
Aspectos Biológicos y Productivos de la Pubertad de la ... · ... no obstante recoger aspectos...
42
Aspectos Biológicos y Productivos de la Pubertad de la Hembra Bovina Dr. Roberto Faure Garcí - [email protected] 1. Resumen 2. Introducción 3. Aspectos biológicos de la pubertad 4. Aspectos productivos 5. A modo de conclusión 6. Bibliografía RESUMEN: Se discuten los principales factores que influyen sobre la aparición de la pubertad y se enriquecen con experiencias propias de trabajo en el tema. Principal atención reciben, por su importancia en las condicio- nes subtropicales, el sistema de alimentación y manejo como variables claves que condicionan este even- to. Se tratan los aspectos neuroendocrinos reguladores y se enfatiza en el mecanismo fisiológico que explica la aparición de la pubertad en una determinada etapa del desarrollo somático, como consecuen- cias de interacciones metabólicas del individuo con el medio, que condiciona la puesta en marcha del generador de pulsos de la GnRH a nivel hipotalámico; dando lugar a la secuencia de eventos neurohor- monales, que posibilitan la aparición del estro y la ovulación. Además se discuten tres aspectos cardinales relacionados con las implicaciones económicas de la apari- ción de la pubertad: el retardo en la aparición de la pubertad, las alternativas biotécnicas para evitar el mismo y la relación de aparición de la pubertad con la eficiencia reproductiva y productiva. Se enfatiza en los aspectos de importancia para las condiciones tropicales y particulares de Cuba y se enriquecen con nuestras experiencias de trabajo en el tema. Se reconoce como principal causa de retardo en la pubertad la inadecuada alimentación y el manejo deficiente, durante la etapa de desarrollo somático de los anima- les. Se muestran diferentes productos y esquemas que pueden ser utilizados para la inducción de la pubertad en los animales reúnan un desarrollo somático y genital mínimo, una vez rectificados los problemas de manejo y alimentación. Palabras claves: pubertad, regulación neuroendocrina, factores influyentes, pubertad retardada; inducción; tratamientos hormonales; eficiencia reproductiva. BIOLOGICAL AND PRODUCTIVE ASPECTS OF THE BOVINE FEMALE PUBERTY ABSTRACT: The main factors that influence on appearance of the puberty are discussed and enrich with our work experiences in the topic. Main attention receives, for its importance in the sub-tropical conditions, the feeding and handling system, that function like key variables that condition this event. The neuroendo- crine regulator aspects are approached and it is emphasized in the physiologic mechanism that explains the appearance of the puberty in a certain stage of the somatic development, as consequences of the individual's metabolic interactions with the environment that conditions the activation of the hypothalamic pulses generator of GnRH; giving place to the sequence of neurohormonal events that make possible the presentation of estrous and ovulation. Also three cardinal aspects related with the economic implications of the appearance of the puberty are discussed: the delay in the appearance of the puberty, the biotechnology alternatives to avoid the same one and the relationship of puberty appearance with the reproductive and productive efficiency. It is em- phasized in the aspects of importance for the tropical conditions and particularly for Cuba, enhance with our work experiences in the topic. It is recognized as main causes in puberty retard the inadequate feeding and the defective handling, during the stage of somatic development of animals. Different products and treatments, which can be used for puberty induction in animals with a minimum somatic and genital devel- opment, once rectified the handling and feeding problems. Key words: puberty, neuroendocrine regulation, related factors, delay puberty; induction; hormonal treat- ments; reproductive efficiency.
Transcript of Aspectos Biológicos y Productivos de la Pubertad de la ... · ... no obstante recoger aspectos...

Aspectos Biológicos y Productivos de la Pubertad de la Hembra Bovina Dr. Roberto Faure Garcí - [email protected]
1. Resumen
2. Introducción
3. Aspectos biológicos de la pubertad
4. Aspectos productivos
5. A modo de conclusión
6. Bibliografía
RESUMEN:
Se discuten los principales factores que influyen sobre la aparición de la pubertad y se enriquecen con
experiencias propias de trabajo en el tema. Principal atención reciben, por su importancia en las condicio-
nes subtropicales, el sistema de alimentación y manejo como variables claves que condicionan este even-
to. Se tratan los aspectos neuroendocrinos reguladores y se enfatiza en el mecanismo fisiológico que
explica la aparición de la pubertad en una determinada etapa del desarrollo somático, como consecuen-
cias de interacciones metabólicas del individuo con el medio, que condiciona la puesta en marcha del
generador de pulsos de la GnRH a nivel hipotalámico; dando lugar a la secuencia de eventos neurohor-
monales, que posibilitan la aparición del estro y la ovulación.
Además se discuten tres aspectos cardinales relacionados con las implicaciones económicas de la apari-
ción de la pubertad: el retardo en la aparición de la pubertad, las alternativas biotécnicas para evitar el
mismo y la relación de aparición de la pubertad con la eficiencia reproductiva y productiva. Se enfatiza en
los aspectos de importancia para las condiciones tropicales y particulares de Cuba y se enriquecen con
nuestras experiencias de trabajo en el tema. Se reconoce como principal causa de retardo en la pubertad
la inadecuada alimentación y el manejo deficiente, durante la etapa de desarrollo somático de los anima-
les.
Se muestran diferentes productos y esquemas que pueden ser utilizados para la inducción de la pubertad
en los animales reúnan un desarrollo somático y genital mínimo, una vez rectificados los problemas de
manejo y alimentación.
Palabras claves: pubertad, regulación neuroendocrina, factores influyentes, pubertad retardada; inducción;
tratamientos hormonales; eficiencia reproductiva.
BIOLOGICAL AND PRODUCTIVE ASPECTS OF THE BOVINE FEMALE PUBERTY
ABSTRACT: The main factors that influence on appearance of the puberty are discussed and enrich with
our work experiences in the topic. Main attention receives, for its importance in the sub-tropical conditions,
the feeding and handling system, that function like key variables that condition this event. The neuroendo-
crine regulator aspects are approached and it is emphasized in the physiologic mechanism that explains
the appearance of the puberty in a certain stage of the somatic development, as consequences of the
individual's metabolic interactions with the environment that conditions the activation of the hypothalamic
pulses generator of GnRH; giving place to the sequence of neurohormonal events that make possible the
presentation of estrous and ovulation.
Also three cardinal aspects related with the economic implications of the appearance of the puberty are
discussed: the delay in the appearance of the puberty, the biotechnology alternatives to avoid the same
one and the relationship of puberty appearance with the reproductive and productive efficiency. It is em-
phasized in the aspects of importance for the tropical conditions and particularly for Cuba, enhance with
our work experiences in the topic. It is recognized as main causes in puberty retard the inadequate feeding
and the defective handling, during the stage of somatic development of animals. Different products and
treatments, which can be used for puberty induction in animals with a minimum somatic and genital devel-
opment, once rectified the handling and feeding problems.
Key words: puberty, neuroendocrine regulation, related factors, delay puberty; induction; hormonal treat-
ments; reproductive efficiency.

1 Introducción
Los años 1960-1978, la época del surgimiento y desarrollo del radioinmunoanálisis (RIA), así como del
descubrimiento y uso de las GnRH. El avance alcanzado en la fisiología de la reproducción, en el trata-
miento de diversos trastornos reproductivos, y la aplicación de métodos para elevar la eficiencia de la
multiplicación de los animales, ha estado condicionado en gran parte por el progreso de las determinacio-
nes de las hormonas proteicas, esteroides y prostaglandinas que participan en la función gonadal.
El hallazgo de un patrón de secreción pulsátil de prácticamente todas las hormonas involucradas en el
control del ciclo de procreación reviste una importancia trascendental, demostrándose que todos los cam-
bios de un estado reproductivo a otro se asocian a un incremento de la frecuencia de pulsos de LH.
Se han abierto nuevos temas interesantes en el campo de la reproducción como son: el líquido folicular
con sus diferentes componentes (inhibinas, complemento, inhibidor de aromatasa, etc.), la acción directa
de los esteroides a nivel ovárico, los péptidos opiodes, la oxcitocina ovárica y su relación con la luteolisis,
aparición de nuevos elementos luteotrópicos, la foliculogénesis, etc. Además son capitales los relaciona-
dos con la transferencia de embriones (microcirugía, transferencia intergenéricas), control de la reproduc-
ción por métodos cibernéticos, evaluación de la fertilidad a nivel de laboratorio (capacitación del esperma-
tozoide y fertilización in vitro del óvulo). La biotecnología tiene en la reproducción un gran campo para su
desarrollo, de esta manera ya se han logrado animales transgénicos así como animales clonados a partir
de células somáticas y recientemente bovinos transgénicos clonados.
Dentro del proceso reproductivo su inicio lo constituye la aparición del fenómeno puberal cuyo retraso
posee una gran implicación zootécnica y económica para la industria pecuaria lo que es un marcapaso de
la eficiencia reproductiva de los rebaños bovinos, el cual debe tenerse en cuenta para los sistemas de
manejo del reemplazo de las vacas. Actualmente se enriquece el concepto del control central de la puber-
tad en la hembra al considerar la interacción de estructuras neurosecretoras neuronales con las células
gliales de la línea astrocítica, con la participación de factores de crecimiento, sus receptores y la Prosta-
glandina E2 lo que en conjunto conducen la estimulación del generador de pulsos de LH por la hipófisis y
por esa vía la aparición de la pubertad.
1.1 Concepto de Pubertad
Desde el punto de vista etimológico, viene del latín pubere, que quiere decir “cubrirse de pelo”; es decir, la
pubertad sería la época en que el vello sexual comienza a desarrollarse (Botella, 1982). Tal acepción fue
válida para la especie humana, pero no para la vacuna. En la actualidad la pubertad ha pasado a significar
el período de la vida en que tiene lugar la maduración sexual y se hace factible la procreación (Sawin,
1971).
Asdell (1946), precisó la maduración sexual, como el tiempo en el cual la primera reproducción se hace
posible, cuya peculiaridad es la liberación de células germinales; mientras que, la madurez sexual no se
alcanza hasta que el animal expresa su potencia reproductiva completa. Por su parte, Bonadonna (1953)
consideró, que la pubertad queda señalada en las hembras por la aparición de las primeras manifestacio-
nes sexuales, lo cual sucede a edades diferentes, según las especies animales.
Donavan y Van der Werff Ten Bosch (1965) conceptuaron esta etapa de la vida, como la fase del desarro-
llo corporal durante la cual, las gónadas secretan hormonas en cantidades suficientes para causar un
crecimiento acelerado de los órganos genitales y la aparición de los caracteres sexuales secundarios.
Este concepto, no obstante recoger aspectos fundamentales de la pubertad, todavía es limitado; pues, el
hecho que se produzcan cantidades suficientes de hormonas y crecimiento de los genitales no necesa-
riamente implican la liberación de gametos. Como señalara Ganong (1971), junto a la función endocrina,
la función gametogénica posibilita la reproducción.
Holy y Martínez (1970) definieron la pubertad como “un período de la vía en el cual se cambia en el orga-
nismo la fase de la tranquilidad sexual por la fase de la función activa, caracterizada por la facultad de
reproducción”. Esta definición es demasiado amplia, ya que existen etapas en la vida animal donde se
cumplen esos requisitos, y no se pueden catalogar de pubescentes (Ej. la vaca con anestro prolongado,
que luego reanuda su actividad cíclica).
Para Thibaut y Lavasseur (1974), la pubertad queda enmarcada en el período de la adolescencia en que,
la hembra o el macho liberan gametos por primera vez; y para Mc Donald (1975) tal período, es el mo-
mento en el cual la hembra se convierte en sexualmente madura y las características sexuales secunda-
rias se hacen notables, quedando vinculado al primer celo.

Según Döcke (1981), la pubertad representa la terminación de un amplio proceso de maduración, el cual
comienza ya prenatal, y conduce hacia la capacidad funcional completa de los órganos sexuales primarios
y secundarios; así como la expresión de las características sexuales somáticas y las formas de conducta
específica del sexo.
Como dijera Sadleir (1969), la pubertad es difícil de definir su inicio coincide con un período de la vida del
individual, en que formularla con términos generales no resulta fácil. Dicho inicio está fuertemente ligado a
un aumento previo de la secreción de LH. Pelletier et al., (1981) señalaron que los mecanismos implica-
dos en la expresión de la pubertad son eventos progresivos, en lugar de abruptos.
Así, podemos decir, que la pubertad “es un proceso gradual que aparece en la etapa de desarrollo somá-
tico de un individuo, donde por primera vez se hace posible la reproducción, y finaliza con la madurez
sexual; este proceso se caracteriza por la presencia de un marcado incremento de la frecuencia de pulsos
de la secreción de gonadotropinas (especialmente LH), desarrollo de los genitales y los caracteres sexua-
les secundarios, pudiendo ser dicho proceso espontáneo o inducido”.
En el caso de la hembra se han utilizado diferentes indicadores para revelar la presencia de la pubertad,
entre los que se destacan la medición de la temperatura rectal cuyos resultados han sido infructuosas
(Shotton et al., 1978); las manifestaciones del primer celo (Glencross, 1984) o la primera ovulación segui-
da de una fase lútea (Hall, et al, 1995 y Prendiville, et al 1995) que es lo más aceptado en el caso de la
hembra ovina. La aparición del primer estro se ha enlazado tanto a la pubertad que a veces queda defini-
da enmarcándose a la fecha del primer ciclo sexual; en tal sentido es bueno aclarar, que la pubertad no
es una fecha sino un proceso que ocurre en un período de tiempo difícil de delimitar su inicio y fin. Es una
continuación de la infancia que se introduce gradualmente en la fase de madurez sexual.
Willi (1944), distinguió tres etapas en el desenvolvimiento y maduración de los órganos reproductivos de
la vaca:
Maduración hipofisaria de 3-6 meses de edad
Maduración de los ovarios de 6-12 meses de edad
Maduración del útero de 3 ó más años
El desarrollo continuo del aparato genital, después de entrada la pubertad, ha sido confirmado por mu-
chos trabajos (Swanson et al., 1972; Okano et al., 1980 a, b).
En nuestro país, Nuñez et al., (1977) informaron que a los 40 meses de edad fue que se alcanzó un ta-
maño estable de la vagina y el útero, concomitante con una intensa actividad ovárica en novillas del cruce
Holstein x Cebú. En este crecimiento y maduración orgánica juega un importante papel el peso y la edad
de los animales.
2 Aspectos Biológicos de la Pubertad
2.1 Factores que influyen sobre la aparición de la pubertad
2.1.1 INTRÍNSECOS
2.1.1.1 Edad y peso
En general, en todas las especies de animales la pubertad se adelanta al desarrollo corporal, esto signifi-
ca que las hembras pueden multiplicarse antes de que sus órganos estén en plenitud de su capacidad
para la producción y la reproducción. La capacidad reproductora aparece cuando el animal alcanza un
peso determinado, este peso es aproximadamente el 45% del peso del adulto en el caso del bovino euro-
peo (Thibault y Levaseeur, 1974; Mc Donald, 1975), el 60% en novillas Brahman y del 64 al 67% en novi-
llas mestizas (González-Stagnaro, 1995). Además, el peso es más importante que la edad, ya que deter-
mina el momento de arribo a la pubertad, aunque siempre es necesaria una edad mínima (Arije y Wilt-
bank, 1974).
Existen otros indicadores estrechamente relacionados con el peso corporal que constituyen expresión del
estado fisiológico y metabólico del animal y de cuya interpretación puede estimarse el potencial reproduc-
tivo a corto plazo. Estos son la Condición Corporal (CC) que estima las reservas energéticas y la Califica-
ción del Tracto Reproductivo (CTR) que es un indicador del desarrollo genital del animal. Para que las
novillas incorporadas a la reproducción tengan un comportamiento reproductivo adecuado es necesario
que además de un peso mínimo, posean una condición corporal mayor de 2.5, preferiblemente entre 3 y
4, y una CTR de 4 o más.

Jung y Kiracofe (1985) señalaron que existía una correlación entre el peso y la primera gestación (r =
0.449) y que a su vez, el peso estaba asociado a la puntuación (score) de la Condición Corporal; en tal
sentido, también se indicó que el estado físico representado por el score influye marcadamente en el
desarrollo folicular a través de la secreción de gonadotropinas (Rhind y Mc Neully, 1986) y puede servir
para predecir la edad de la pubertad en novillas de carne (Brooks et al., 1985).
La edad a la cual las novillas logran su primer ciclo estral varía considerablemente. Desde los clásicos
trabajos de Hamond (1927) sobre reproducción en la especie bovina, se calculó que la edad a la pubertad
de las novillas lecheras fluctuaban entre 3 y 15 meses con promedio de 9 Fahr et al., (1984) observaron
que las novillas alcanzaban la primera ovulación de 9 a 11 meses de edad con peso de 235 a 242 Kg.; en
tanto, Roy et al., (1985) hallaron edades de 6,9 meses para la raza Friesian y 8.4 a 9 meses para la raza
Angus. En Cuba se han publicado valores de 27 a 29 meses como edad de incorporación a los servicios
de IA para el cruce 3/4H x 1/4C (Morales et al., 1977), cruce en el cual se pudo disminuir la edad a los 77
meses cambiando el régimen de manejo (Tarrero et al., 1981). La edad y peso promedio nacional de in-
corporación de las novillas son de 28-32 meses y 268-270 Kg. respectivamente (Betancourt et al., 1986).
En la tabla 1 se muestran los indicadores en algunos rebaños genéticos.
Tabla 1: Edad (meses) y peso (Kg.) de incorporación a los servicios de inseminación artificial y edad al
primer parto en algunos rebaños genéticos (1985).
Incorporación 1er. parto
Razas Edad Peso Edad
Holstein Tropical 20.6 318 31.6
Siboney de Cuba 21.1 311 33.4
3/4H x 1/4C 21.9 304 33.4
Mambí de Cuba 22.7 393 34.2
Cebú 21.8 308 35.7
Charoláis 21.5 316 30.0
Santa Gertrudis 21.0 307 34.6
Fuente: MINAGRI, según Betancourt et al, (1986).
En la actualidad se acepta como más conveniente que las novillas lecheras deben comenzar la pubertad
entre 10 y 15 meses (IAEA, 1984).
Que se haya enfatizado el peso como más importante que la edad, no menosprecia el valor que tiene ésta
en el proceso puberal. Basta recordar la exposición de Willi (1944) relacionada con las etapas de la madu-
ración sexual o los resultados de Burfening (1979), el cual al someter dos grupos de novillas con diferen-
tes edades a un tratamiento inductor de la pubertad, encontró muy baja efectividad en el grupo de menos
edad (11,1 meses) respecto al de mayor edad (13,5 meses). Además Yelich, et al (1995) planteó que el
porciento de grasa corporal no es solo el regulador de la pubertad sino que la edad debe ser un regulador
importante en determinar la pubertad en novillas de carne.
La trascendencia del peso es relativa, porque tiene un límite fuera del cual resulta perjudicial para la re-
producción (Owens, et al, 1993).
2.1.1.2 Genéticos
Desde las investigaciones iniciales de Hammond (1927) se pudo en evidencia el efecto que tiene la raza
sobre la edad en que aparece la pubertad en vacas lecheras, observándose para las razas Jersey,
Guernsey, Friesian y Ayrshire edades promedios de 8, 11, 11 y 13 meses respectivamente.
La influencia genética sobre la edad de aparición de la pubertad se ha demostrado por algunos experi-
mentos que confirman el hecho de que los genes de la madurez sexual temprana son dominantes sobre
los de la madurez sexual tardía (El Amin, 1976). Esto está de acuerdo a lo previamente señalado por
Plasse (1966), de que independientemente del sistema de alimentación, el ganado Bos indicus presenta
la pubertad y el primer parto a edades más avanzadas que el ganado Bos taurus. En este sentido Holy y
Martínez (1970) puntualizaron que en el Cebú la pubertad aparece de 6 a 12 meses más tarde que en el
ganado europeo.

Por otra parte, también se ha observado que el grado de heterosis influye notablemente sobre la edad
(Rivera et al., 1982) y peso a la pubertad (Lee et al., 1984).
Si la causa básica de las diferencias del comportamiento reproductivo entre dos genotipos distintos de
animales reside en el genofondo, la causa inmediata hay que buscarla en el aparato neurofisiológico que
controla el ciclo reproductivo. En conexión con esto, se comunicó que la vaca Brahman posee un cuerpo
lúteo pequeño, tiene deficiencias de la enzima 3 esteroide deshidrogenasa y en consecuencia menos
producción de progesterona (Aguilar y Colina, 1982); además el Brahman tratado con estradiol responde
con un celo poco intenso y presenta una curva de LH de menor dimensión cuando se compara con una
raza europea (Rhodes et al., 1979) y concluyen que existe una diferencia racial inherente al mecanismo
de retroalimentación a nivel del eje hipotálamo - hipófisis entre estos dos tipos de razas.
Por otra parte la influencia del padre en la ganancia de peso de sus hijas influye a través del peso en la
edad de la pubertad (Freetly y Cundiff 1997). Asimismo algunos criterios de selección aplicados como es
el basado en la producción de grasa y proteína disminuyen la velocidad de crecimiento y engorde en novi-
llas; (Coffey, Hickey et al. 2006) lo cual está relacionado directamente a la edad de aparición de la puber-
tad.
2.1.2 EXTRÍNSECOS
2.1.2.1 Nutrición y Manejo
La reproducción de los rumiantes, componente esencial de la producción, solo permite explotar menos de
las dos terceras partes de su potencial, constituyendo las deficiencias nutricionales su principal causa
(Topps, 1977).
El retraso de la pubertad en ganado es el efecto fundamental que produce la subalimentación (Topps,
1977, Patterson, et al, 1992), mientras que un plano nutricional alto la acelera (Busbrik, et al, 1995,
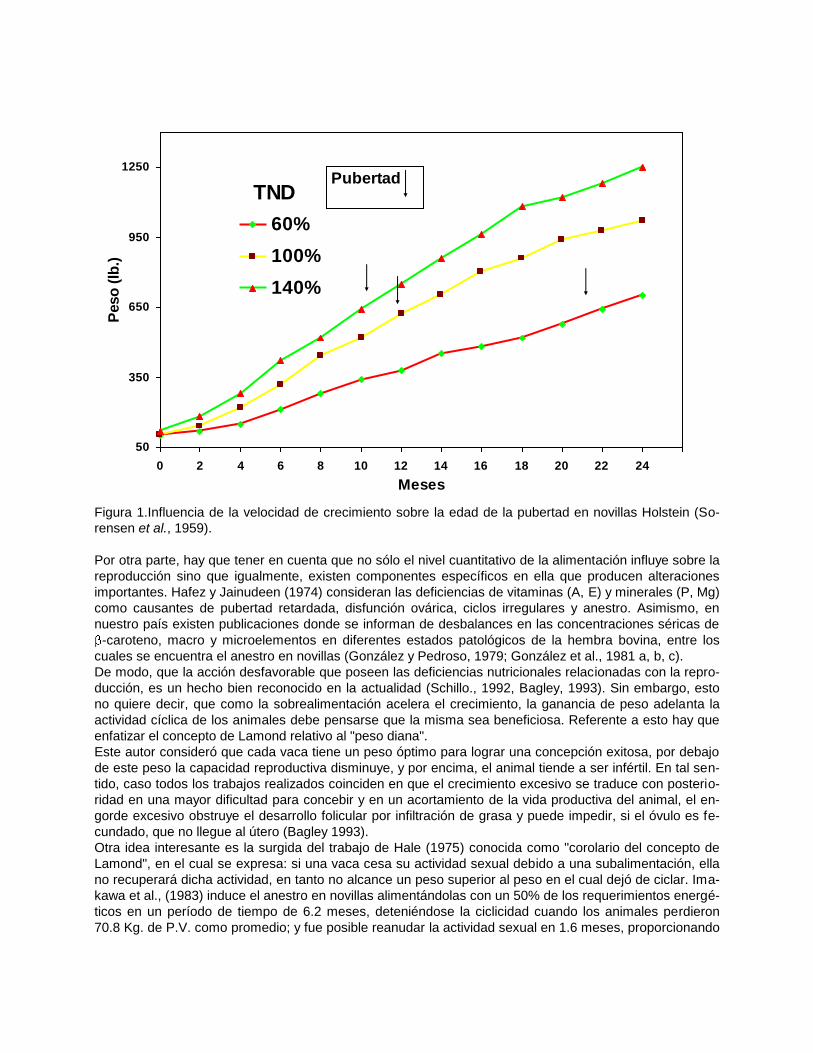
Radcliff, et al, 1997). Este efecto se había demostrado por Sorensen et al., (1959), quienes en un estudio
con novillas lecheras alimentadas en grupos con 140, 100 y 60% de los requerimientos de Morrison, ob-
servaron que el primer celo se presentó a los 8,5; 11,3; 16,3 meses de edad y 261, 261, 243 Kg. de P.V.
respectivamente(figura 1). Perón y Tarrero (1982) en novillas Holstein x Cebú alimentadas con dos dietas
una normal y otra por debajo de las necesidades encontraron una diferencia de 4,7 meses en las edades
al primer celo.
Se ha demostrado que la restricción de alimentos provoca disminución en la expresión del gen de la lepti-
na y en los niveles de leptina circulante, así como de Factor de Crecimiento similar a la e insulina (IGF-I),
de insulina y de LH lo cual da bases para considerar a la leptina como la hormona que indica el estatus
energético al eje reproductivo central en ganado bovino (Amstalden, et al. 2000).
50
350
650
950
1250
0 2 4 6 8 10 12 14 16 18 20 22 24
Meses
Peso
(lb
.)
60%
100%
140%
TNDPubertad
Figura 1.Influencia de la velocidad de crecimiento sobre la edad de la pubertad en novillas Holstein (So-
rensen et al., 1959).
Por otra parte, hay que tener en cuenta que no sólo el nivel cuantitativo de la alimentación influye sobre la
reproducción sino que igualmente, existen componentes específicos en ella que producen alteraciones
importantes. Hafez y Jainudeen (1974) consideran las deficiencias de vitaminas (A, E) y minerales (P, Mg)
como causantes de pubertad retardada, disfunción ovárica, ciclos irregulares y anestro. Asimismo, en
nuestro país existen publicaciones donde se informan de desbalances en las concentraciones séricas de
-caroteno, macro y microelementos en diferentes estados patológicos de la hembra bovina, entre los
cuales se encuentra el anestro en novillas (González y Pedroso, 1979; González et al., 1981 a, b, c).
De modo, que la acción desfavorable que poseen las deficiencias nutricionales relacionadas con la repro-
ducción, es un hecho bien reconocido en la actualidad (Schillo., 1992, Bagley, 1993). Sin embargo, esto
no quiere decir, que como la sobrealimentación acelera el crecimiento, la ganancia de peso adelanta la
actividad cíclica de los animales debe pensarse que la misma sea beneficiosa. Referente a esto hay que
enfatizar el concepto de Lamond relativo al "peso diana".
Este autor consideró que cada vaca tiene un peso óptimo para lograr una concepción exitosa, por debajo
de este peso la capacidad reproductiva disminuye, y por encima, el animal tiende a ser infértil. En tal sen-
tido, caso todos los trabajos realizados coinciden en que el crecimiento excesivo se traduce con posterio-
ridad en una mayor dificultad para concebir y en un acortamiento de la vida productiva del animal, el en-
gorde excesivo obstruye el desarrollo folicular por infiltración de grasa y puede impedir, si el óvulo es fe-
cundado, que no llegue al útero (Bagley 1993).
Otra idea interesante es la surgida del trabajo de Hale (1975) conocida como "corolario del concepto de
Lamond", en el cual se expresa: si una vaca cesa su actividad sexual debido a una subalimentación, ella
no recuperará dicha actividad, en tanto no alcance un peso superior al peso en el cual dejó de ciclar. Ima-
kawa et al., (1983) induce el anestro en novillas alimentándolas con un 50% de los requerimientos energé-
ticos en un período de tiempo de 6.2 meses, deteniéndose la ciclicidad cuando los animales perdieron
70.8 Kg. de P.V. como promedio; y fue posible reanudar la actividad sexual en 1.6 meses, proporcionando

a las novillas una ración que les permitiera una ganancia diaria de 1.4 Kg., lo cual a quedado establecido
como metodología para provocar anestro en novillas (Schoppee et al, 1996).
¿Qué mecanismos tiene la hipoalimentación para producir estas alteraciones reproductivas?
Suzuki et al., (1982), subalimentaron novillas y encontraron un acortamiento de la duración del celo (4.8 h
vs 20.2h), disminución de la libido, reducción del intervalo celo - ovulación (2.0 h vs 14.1h) y disminución
de las concentraciones de progesterona en los dos primeros ciclos experimentales. La depresión de los
niveles periféricos de progesterona ha sido confirmada en otras investigaciones (Donaldson et al., 1970;
Gombe y Hansel al., 1978; Kinder et al., 1982; Alonso et al., 1984, 1985), aunque Spitzer et al., (1978) no
observaron esa relación, no obstante haber detectado que los animales con dietas hipoenergéticas tuvie-
ron el ovario (con CL) un tamaño 57% más pequeño que las dietas adecuadas. La disminución del tama-
ño y peso (Gombe y Hansel, 1973) y la secreción (in vitro) de progesterona del cuerpo lúteo (Kinder et al.,
1982) también fue señalada. Análogamente, las raciones deficientes en lípidos o colesterol conducen a un
menoscabo de la producción de progesterona (Thatcher et al., 1996).
La ingestión deficitaria de nutrientes tiene un efecto pronunciado sobre las concentraciones de gonadotro-
pinas. Según Rombauts et al., (1961), diversos trabajos sugieren no sólo una disminución en la síntesis
de estas hormonas a nivel hipofisario sino también una modificación de su liberación. Sin embargo, en
novillas Beal et al., (1978) midieron cantidades significativas de LH en hipófisis, al parecer constituyentes
de un pool de fácil y mayor liberación al exponer la glándula al GnRH (Beal et al., 1978; Fitzgerald, 1984;
Spicer et al., 1984), comparadas con grupos que recibieron una mejor alimentación. Igualmente, en la
actualidad, otra característica endocrina del bajo consumo de nutrientes es la presencia de una reducción
importante de las frecuencias de pulsos de LH (Kinder et al., 1982 a; Imakawa et al., 1983; Lindsay et al.,
1984; Imakawa et al., 1984, 1985), las cuales son responsables en parte del anestro en novillas inducido
por la subalimentación (Imakawa et al., 1984). También se han indicado cambios en los niveles de FSH,
prolactina y estradiol (Rone, 1984)
De modo, que la subalimentación puede actuar directamente a nivel ovárico o hipofisario, produciendo
alteraciones que conducen a un pobre desarrollo folicular, deficiente producción de hormonas esteroides,
las cuales no pueden promover el celo.
El mecanismo exacto de estos cambios, aún no están esclarecidos. Es posible que la hipoalimentación
(dependiendo si es crónica o aguda, si es energética o proteica, etc.) produzca un aumento en la sensibi-
lidad al mecanismo de retroalimentación negativo de los estrógenos, lo que a su vez implica una disminu-
ción en la liberación hipotalámica de GnRH con el consiguiente cuadro de baja frecuencia en la secreción
pulsátil de gonadotropinas y la falta de estimulación ovárica. En este sentido Schillo (1992) propuso que
metabolitos combustibles oxidables como la glucosa y los ácidos grasos no esterificados y/u hormonas
metabólicas reflejando el estado nutricional influyen en la actividad de las neuronas que controlan la libe-
ración de GnRH.
Manejo: Este es otro aspecto importante a tener en cuenta, cuando se trabajan animales en desarrollo.
Según Web (1972), un buen manejo es necesario desde el nacimiento hasta el momento del primer parto.
De acuerdo a este autor hay tres áreas principales en disciplinas de manejo.
1. Mantenimiento de un óptimo desempeño reproductivo
2. Control de enfermedades y parásitos
3. Mantenimiento de un elevado nivel nutritivo
Factores tales como el medio donde se deben desarrollar las novillas, el traslado frecuente, suministro de
agua, carga por ha de los pastos, frecuencia de baños acaricidas, etc. Son muy influyentes en la aparición
de una pubertad temprana.
Actualmente (Bagley, 1993) el sistema de producción de novillas de reemplazo para lograr partos a los
dos años de edad requiere un plan de manejo integral animal forraje y debe comenzar en la fase de pro-
ducción vaca ternero y en este sentido además propone tener en cuenta:
1. Raza
2. Peso al destete
3. Técnica de manejo pre y post destete (antihelminticos, anabólicos, probióticos y ionofo-
ros)
4. Nutrición
5. Época de monta

2.1.2.2 Climáticos
Temperatura: Dale et al. (1959) comprobaron que las altas temperaturas influyen en el inicio de la puber-
tad. Estudiando novillas de diferentes razas observaron, que a temperatura constante de 26.66°C la pu-
bertad aparecía a los 13.3 meses de edad, mientras que mantenidas a 10°C esta aparecía a los 10 me-
ses.
Otro efecto lo refiere (Tucker, 1982), de que las temperaturas elevadas disminuyen la duración e intensi-
dad del celo y si se mantienen por encima de 28°C (especialmente si el estrés es prolongado) conduce a
una reducción del índice de crecimiento de los bóvidos (Johnson y Ragsdale, 1959; Mendel et al., 1971).
El retardo de la pubertad a altas temperaturas se relaciona con un bajo consumo de alimento y un retraso
del crecimiento en esas condiciones, lo que posiblemente esté vinculado a la elevación de la tasa de cor-
ticoides en sangre que se presenta en el estrés de calor (Gwazdauskas, 1985; Faure et al., 2004) y a la
disminución de la secreción de hormonas tiroideas.
Fotoperíodo: Contrario a los equinos, ovinos y caprinos, la vaca es considerada como una especie que
tiene reproducción no estacionaria. Sin embargo, ciertos aportes científicos llaman a la reflexión sobre
este punto de vista.
Pumm y Kush (1934) y Salisbury (1946) estudiaron el influjo de la estación sobre la edad de parto en novi-
llas primíparas, observando, este último, en el Estado de Iowa, que en el mes de mayo el parto se logró
aproximadamente con 25 meses de edad, mientras que en septiembre fue de aproximadamente 29 me-
ses. Ellos concluyeron, que sobre todo, en las zonas más alejadas del ecuador la influencia del mayor
número de horas diurnas propio de la primavera y el verano, tiende a disminuir la edad de concepción de
las novillas.
Se han publicado muchos trabajos donde aparece una relación entre la edad de la pubertad y la época en
que nacen las terneras (Hansen et al., 1983; Kanwanja, 1984). Fauconneau y Gauthier (1984) trabajando
en el trópico con hembras criollas prepúberes, informaron que la adición de 4 horas de luz diaria incre-
mentaba en un 37.5% el número de animales que ciclaban, en relación a un grupo que sólo recibió luz
natural. Estas observaciones fueron confirmadas por Kamwanja (1984), al notar que el aporte en una
suplementación de luz a un grupo de novillas adelantaba la pubertad en 27 días.
Que el fotoperíodo afecta las edades de aparición de la pubertad y la actividad ovárica en novillas Brah-
man puras y cruzadas ha sido confirmado por Mezzadra, et al, (1993). Más directamente se ha compro-
bado que mayor cantidad de luz diaria adelanta la edad de aparición de la pubertad (Ringuet, et al, 1994).
Aunque, Sergent et al, (1984) no encontraron alteraciones significativas en las variables del pico preovula-
torio de LH, ni en los niveles plasmáticos de progesterona en novillas con mayor tiempo de exposición a la
luz solar, sí parece ser que el fotoperíodo puede modificar la edad de la pubertad modulando la secreción
pulsátil de LH (Louw et al., 1983) o la secreción de prolactina y velocidad de crecimiento (Tucker, 1982;
Schillo et al., 1983a). De tal manera que el manejo del fotoperiodo puede ser usado para incrementar el
crecimiento en novillas y maximizar la adquisición de tejido graso incluyendo parénquima mamario. (Dahl
and Petitclerc, 2003)
Ciclo lunar, Roy et al., (1980) publicaron por primera vez haber encontrado una conexión entre la edad de
aparición de la pubertad en novillas y las fases de la luna. Posteriormente (Roy et al., 1985a) publicaron
otro trabajo relacionado con hembras cruzadas bajo luz continua, donde de 10 novillas, el 50 porciento
presentaron celo 3 a 4 días antes de la luna llena (p < 0.01); destacando además la existencia de una alta
correlación (r = 0.83) entre la edad del primer celo de las hijas respecto a la edad del primer celo de la
madre y la siguiente luna nueva. Sin embargo, Greer (1984) trabajando 556 novillas de carne observó que
la distribución de los primeros celos fue uniforme a lo largo del ciclo lunar. Este es un tema novedoso que
como se ve, para su esclarecimiento se necesitan futuras investigaciones.
En general, al igual que Thatcher et al., (1986), pensamos que cuando se estudian variables ambientales
como es el clima en relación con la fisiología de los animales, debe tenerse en cuenta que con ellas se
entrelazan un conjunto de factores como son: temperatura, humedad, radiaciones, nutrición, manejo, etc.
por lo cual, establecer relaciones de causas efectos no resultan fáciles si no se realizan experimentos
muy bien controlados.

2.1.2.3 Sociales, sexo, enfermedad
Los factores sociales también pueden modificar el comienzo de la maduración sexual en los animales.
Thibaut y Levasseur (1974) refirieron que en ovinos la presencia del sexo opuesto adelanta la pubertad y
contrariamente, la presencia del mismo sexo la retrasa.
En el bovino se sabe que exhibir el toro a una vaca en celo produce aumento de la motilidad uterina y
adelanta el momento de la ovulación (Vandermark y Hays, 1953). Sin embargo no se ha observado una
influencia positiva de la presencia del macho en la edad de aparición de la pubertad en novillas (Wehr-
man, et al, 1996)
Por su lado, Mc Donald (1975) planteó que las hembras de todas las especies pubescen más temprano
que los machos, siendo del orden de 2 a 4 meses en el vacuno. La causa de esta diferencia sexual no
está clara, pero probablemente resida en los centros superiores del SNC.
Hawk et al (1954) estudiaron la influencia de las diarreas y de la consanguinidad sobre el momento de la
aparición del estro en la raza Holstein, encontrando que producían un retraso de la pubertad por su acción
frenadora del crecimiento, por lo cual Salisbury y Vandermark (1961) consideraron que todo factor que
interfiriera en el desarrollo orgánico, alteraba la edad de aparición de los primeros ciclos estrales.
Swift et al., (1983; 1984) dan cuenta de la inducción de anestro en novillas de carne por una anaplasmosis
provocada experimentalmente. Cuyo cuadro se caracterizó (aparte de la fiebre y la anemia) por una dis-
función ovárica con disminución de la producción de progesterona y estradiol, tal vez por un aporte defi-
ciente de oxígeno para la síntesis de esteroides.
Esta última información es valiosa, ya que pudiera explicar en parte algunos casos de anestrías vincula-
dos a los brotes de hemosporidosis en nuestro país. Incluso pudiera ser también que los animales porta-
dores (sanos) de los parásitos sufran daños crónicos en sus órganos reproductivos y como consecuen-
cias aparezcan disturbios de la fertilidad o el celo, similares a los encontrados (abortos) en la vaca gestan-
te (Muñoz y Cisse, 1983).
2.1.2.4 Actividad ovárica y conducta sexual
El feto femenino de la vaca a los 110 días de gestación tiene 2,7 x 106
ovocitos, los cuales se reducen a
68 000 al momento del nacimiento (Thibaut y Levasseur, 1974). Desde el nacimiento hasta los 3 meses
de edad se produce un desarrollo de estos aumentando considerablemente el número de folículos antra-
les (Erickson, 1986; Goff et al., 1984) y este permanece constante hasta el inicio de la pubertad (Forrest
et al., 1984). De modo, que las terneras al nacer poseen una cantidad fija de folículos, que pasan con el
tiempo un proceso de desarrollo, atresia y desaparición; constituyendo la pubertad el momento donde
comienza una intensa actividad ovárica, a partir de donde se hace viable la liberación y fertilización del
óvulo.
Los datos ofrecidos por varios autores demuestran que los ovarios de las novillas evidencian actividad
folicular mucho antes de alcanzar el primer ciclo estral. Morrow (1969) en la raza Holstein, durante la pre-
pubertad detectó folículos palpables de 0.5 a 2 cm. de diámetro en la mayoría de los animales estudiados
y en el 74% la primera ovulación se acompañó de celo silente.
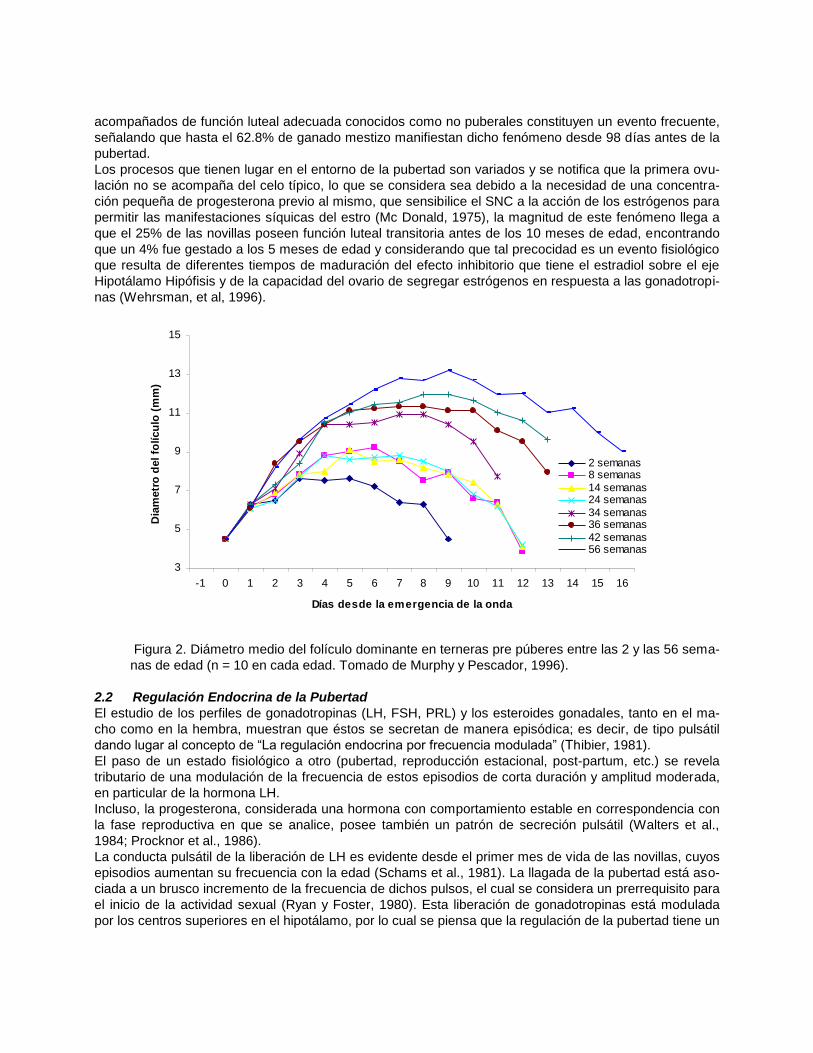
Con el advenimiento de la técnica ecográfica se ha podido determinar la dinámica del desarrollo folicular
en animales pre y peripúberes (Calderón, et al, 1993; Bergfeld, et al, 1994 y Adams y Brogliatti, 1996)
comprobándose que la composición de las ondas foliculares son similares a las presentadas en los ani-
males adultos difiriendo solo en su magnitud y no en su esencia, los mecanismos que controlan el fenó-
meno de emergencia de ondas selección del folículo dominante y regresión folicular están presentes des-
de muy temprano comenzando desde los dos meses de vida en la ternera ( figura 2) .
Dofourt (1975) refiere que aproximadamente el 80% de las novillas Friesan ovularon antes de alcanzar la
pubertad sin mostrar signos estrales, y el 33% presentaron su primer ciclo estral con una duración menor
de 10 días. Por otro lado, Rutter (1984) observó que sólo el 48.3% de los animales por él estudiados for-
maron cuerpos lúteos al llegar a su primer ciclo sexual.
En otro estudio (Morrow et al., 1976) donde se evaluaron las características de 245 ciclos estrales en
novillas peripúberes se encontró, que los signos iniciales de celo aparecieron a los 9.3 meses de edad,
mientras que el primer cuerpo lúteo fue detectado un mes mas tarde. Además, la ocurrencia de celo silen-
te, no típico y típico fue del 7.25 y 68% respectivamente y sólo en el 61% de dichos ciclos aparecieron
cuerpos lúteos normales. González Stagnaro (1993) en su revisión del tema refiere que los celos no
acompañados de función luteal adecuada conocidos como no puberales constituyen un evento frecuente,
señalando que hasta el 62.8% de ganado mestizo manifiestan dicho fenómeno desde 98 días antes de la
pubertad.
Los procesos que tienen lugar en el entorno de la pubertad son variados y se notifica que la primera ovu-
lación no se acompaña del celo típico, lo que se considera sea debido a la necesidad de una concentra-
ción pequeña de progesterona previo al mismo, que sensibilice el SNC a la acción de los estrógenos para
permitir las manifestaciones síquicas del estro (Mc Donald, 1975), la magnitud de este fenómeno llega a
que el 25% de las novillas poseen función luteal transitoria antes de los 10 meses de edad, encontrando
que un 4% fue gestado a los 5 meses de edad y considerando que tal precocidad es un evento fisiológico
que resulta de diferentes tiempos de maduración del efecto inhibitorio que tiene el estradiol sobre el eje
Hipotálamo Hipófisis y de la capacidad del ovario de segregar estrógenos en respuesta a las gonadotropi-
nas (Wehrsman, et al, 1996).
3
5
7
9
11
13
15
-1 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Días desde la emergencia de la onda
Dia
metr
o d
el fo
lícu
lo (
mm
)
2 semanas8 semanas
14 semanas24 semanas
34 semanas36 semanas
42 semanas56 semanas
Figura 2. Diámetro medio del folículo dominante en terneras pre púberes entre las 2 y las 56 sema-
nas de edad (n = 10 en cada edad. Tomado de Murphy y Pescador, 1996).
2.2 Regulación Endocrina de la Pubertad
El estudio de los perfiles de gonadotropinas (LH, FSH, PRL) y los esteroides gonadales, tanto en el ma-
cho como en la hembra, muestran que éstos se secretan de manera episódica; es decir, de tipo pulsátil
dando lugar al concepto de “La regulación endocrina por frecuencia modulada” (Thibier, 1981).
El paso de un estado fisiológico a otro (pubertad, reproducción estacional, post-partum, etc.) se revela
tributario de una modulación de la frecuencia de estos episodios de corta duración y amplitud moderada,
en particular de la hormona LH.
Incluso, la progesterona, considerada una hormona con comportamiento estable en correspondencia con
la fase reproductiva en que se analice, posee también un patrón de secreción pulsátil (Walters et al.,
1984; Procknor et al., 1986).
La conducta pulsátil de la liberación de LH es evidente desde el primer mes de vida de las novillas, cuyos
episodios aumentan su frecuencia con la edad (Schams et al., 1981). La llagada de la pubertad está aso-
ciada a un brusco incremento de la frecuencia de dichos pulsos, el cual se considera un prerrequisito para
el inicio de la actividad sexual (Ryan y Foster, 1980). Esta liberación de gonadotropinas está modulada
por los centros superiores en el hipotálamo, por lo cual se piensa que la regulación de la pubertad tiene un

origen central (Generador de Pulsos de GnRH), donde el eje hipotálamo - hipófisis - ovario juega un papel
fundamental; aunque, aún no está bien esclarecido (Apter, 1997).
2.2.1 Concentraciones de hormonas en sangre
En un estudio (Nakada, et al; 2000) donde se registraron los cambios hormonales del nacimiento a la
pubertad en novillas se observaron cambios en ciclos de 7 a 8 días en la concentración de FSH desde los
10 días a los 9 meses de edad. Las variaciones de las hormonas reproductivas evidenciaron la existencia
de tres momentos críticos, el primero fue del nacimiento a los 7 días donde fueron observados los mayo-
res niveles de hormonas esteroideas los que fueron paulatinamente declinando a la vez que aumentaron
los niveles de las gonadotropinas, el segundo momento fue a las cuatro semanas donde hubo un incre-
mento en los valores de LH, estradiol, testosterona e inhibina y el tercero 5 semanas antes de la primera
ovulación donde hubo incrementos en las concentraciones de estradiol seguido por aumentos de la LH.º
2.2.1.1 Hormona luteinizante (LH)
La LH es una glucoproteína que tiene un peso molecular de 30 000 daltons. Es producida y liberada por la
adenohipófisis bajo el influjo de la GnRH proveniente del hipotálamo.
Entre sus funciones en la hembra se encuentran: estimular la secreción de estrógenos en el folículo, pro-
vocar la ovulación y la formación del cuerpo lúteo, así como activar la producción de progesterona (Nis-
wender et al., 1974). Dichas funciones probablemente se logren mediante la acción intermediaria del
Ampc (teoría del segundo mensajero), como puntualizaron Niswender et al., (1972) o quizás alterando el
metabolismo de los fosfolípidos en la membrana plasmática de las células lúteas (Smith, 1986).
El feto bovino contiene en su sangre concentraciones de LH medibles. A los 90 días de gestación estas
concentraciones son de 3.0 ng/mL, y luego descienden paulatinamente a 1.28, 0.85 y 0.45 ng/mL los días
180, 260 de gestación y séptimo día postnatal respectivamente (Oxender et al., 1971ª.Los niveles de esta
hormona se van incrementando gradualmente con la edad, ganando en magnitud y frecuencia; ya a los
siete meses su cantidad es mayor, que 2 ng y mas tarde que 4.1 ng por mL, para alcanzar la máxima
concentración en el umbral del primer ciclo estral (González-Padilla, 1975a; Johnsson y Obst, 1978; Mc
Leod, et al., 1984).
Al llegar a la pubertad, las novillas pueden tener en su primer pico preovulatorio de LH con magnitudes
máximas que oscilan entre 11 a 52 ng/mL (González-Padilla et al., 1975a); pudiendo llegar hasta 193.7
ng/mL cuando se les suministran cantidades de 2 g de GnRH en forma de pulsos (Mc Leod et al., 1985).
2.2.1.2 Progesterona
Mucho antes del primer estro los niveles periféricos de progesterona pueden ser menores a 0.1 ng/mL de
suero sanguíneo (González.Padilla et al., 1975a) o fluctuar entre 0.68 a 0.82 ng/mL (Shotton et al., 1978).
La característica fundamental que tiene el perfil de la concentración de progesterona durante el período
prepuberal es que aparecen los primeros picos previos al primer ciclo estral.
En este momento, se producen cambios en las concentraciones de la hormona, que están muy relaciona-
dos con los signos del celo, los picos de LH y la edad de la pubertad. Por ejemplo, González-Padilla et al.,
(1975a), informaron que existían dos picos de LH, uno primario y otro que ocurre simultáneo con la puber-
tad; y a la vez observaron, la presencia de dos elevaciones de progesterona. La primera sucedía previa al
pico primario de LH y la segunda ocurría entre ambos picos. Estos autores concluyeron enfatizando que la
progesterona debía tener una función clave en el inicio de la maduración sexual.
Por otra parte, Shotton et al., (1978) indicaron en novillas de la raza Friesian la presencia de un ciclo pre-
puberal de progesterona, similar en concentración y duración a los de una fase lútea normal. Ellos tam-
bién encontraron picos de progesterona hasta 5.5 ng/mL por encima de los niveles básales, 2 a 8 días
antes de la ocurrencia del primer celo. Además, dichos autores correlacionaron la edad de la pubertad
con los niveles básales de progesterona (r = 0.541).
Parece existir una concordancia entre los diferentes autores, respecto a las variaciones de las concentra-
ciones de progesterona que tienen lugar alrededor de la edad de la pubertad, ya que estos resultados han
sido confirmados por otros trabajos (Suzuki y Sato, 1980, Schillo et al., 1983a).

Los ovarios de las novillas prepúberes son susceptibles de ser estimulados por gonadotropinas exógenas
y responder con una asombrosa producción de progesterona y formación de cuerpos lúteos (CL). Así,
Spilman et al. (1973), en terneras de 2 a 5 meses de edad, inyectadas con PMSG y LH cuantificaron valo-
res de progesterona de casi 100 ng/mL a los 8-10 días postratamiento y se obtuvieron como promedio 36
Cls por animal. Se halló también una alta correlación entre los niveles de progesterona y el número de Cls
(r = 0.76). Esa capacidad funcional de CL de ternera ya se había señalado previamente (Spilman et al.,
1972).
Están bien establecidos los valores periféricos de progesterona durante el ciclo estral de las novillas. En
general, son inferiores a 1 ng/mL durante la fase folicular pueden alcanzar durante la fase diestral prome-
dios de 2.3 a 5.2 ng/mL dependiendo de la época del año (Solano et al., 1983), e incluso, ser superiores a
10 ng/mL (Schillo et al., 1983a).
2.2.1.3 Estradiol
El líquido folicular de folículos ováricos colectados de novillas con 3.6 y 9 meses de edad contiene canti-
dades pequeñas, pero apreciables de estradiol, las cuales no difieren entre dichas edades (Forrest et al.,
1984).
En suero, las concentraciones (pg/mL) en novillas pueden oscilar entre 5 a 20 (González-Padilla et al.,
1975a; Suzuki y Sato, 1989) o de 3.99 a 13.60 (Radchenkov et al., 1984) e incluso de 1 a 9 (Glencross,
1984; Moseley et al., 1984). El estradiol circula en muy bajas concentraciones, por lo cual, su determina-
ción no resulta muy fácil; siendo posible que éstas diferencias de magnitudes, como regla, respondan a
diferencias metodológicas.
En el estudio de González-Padilla et al., (1975a) no se encontró asociación entre los niveles de estradiol
(E2) y los picos de LH, quizás debido a la frecuencia del muestreo, pero se observó una ligera correlación
con el nivel de GnRH (r = 0.16) durante el período prepuberal. Sin embargo, Glencross (1984) indicó que
el E2 variaba aleatoriamente entre 1 a 4 pg/mL durante la prepubertad y que existían tres picos de esta
hormona: uno de 6 pg/mL que ocurría ocho días antes del primer celo, otro (9 pg/mL) el día de inicio de la
pubertad y el tercero (4 pg/mL) cuatro días después del celo.
Este autor concluyó destacando que precisamente a los 8 días antes de la pubertad disminuye la sensibi-
lidad del eje hipotálamo - hipófisis a la retroalimentación negativa del E2 dando lugar a la liberación de
gonadotropinas y por tanto la aparición del primer celo.
Si durante el estadio prepuberal como hemos dicho, se mantiene una concentración pobre o una frecuen-
cia disminuida de gonadotropinas, se pudiera pensar que esto se debe a un contenido deficiente en estas
hormonas a nivel hipofisario o que su liberación se encuentra perturbada; motivo por el cual no podrían
reflejarse a nivel periférico. Veremos que esto no es así.
2.2.1.4 Testosterona
No disponemos de información acerca de los niveles de testosterona en relación a la pubertad; pero se
sabe que el ovario de la vaca es la mayor fuente de producción de los andrógenos hallados en sangre
periférica (Mori, 1975; Kanchev et al., 1976).
Las concentraciones de testosterona (pg/mL) durante el celo en las novillas son de 40, siendo de 1809
siete días y 78 un día antes del mismo. En muchos trabajos se informa una asociación entre los niveles
de testosterona y el aumento del estro (Shemesh y Hansel, 1974; Kanchev et al., 1976; Nessan y King,
1981; Sekizawa et al., 1982); aunque se considera que el patrón de la concentración de andrógenos a lo
largo del ciclo sexual es impredecible dada su variabilidad (Spitcer y Echternkamp, 1986).
2.2.2 Eje hipotálamo – hipófisis
El peso de la hipófisis de ternera en el momento del nacimiento es de 0.69g y a partir de este instante se
incrementa en relación directa con la edad hasta los 9 meses, estabilizándose entonces para alcanzar un
peso de 1.8g a los 12 meses (Desjardins y Hafs, 1969). Ese peso responde fundamentalmente al de la
adenohipófisis.
El contenido de LH en la hipófisis fetal varia marcadamente, siendo de 0. 323, 0.474 y 0.535 µg/mg a los
90, 180 y 260 días de la gestación respectivamente, no teniendo relación con las concentraciones en sue-
ro (Oxender et al., 1971 b). Al nacimiento es de 2,4 µg/mg, aumentando hasta los 3 meses (9.1) y luego
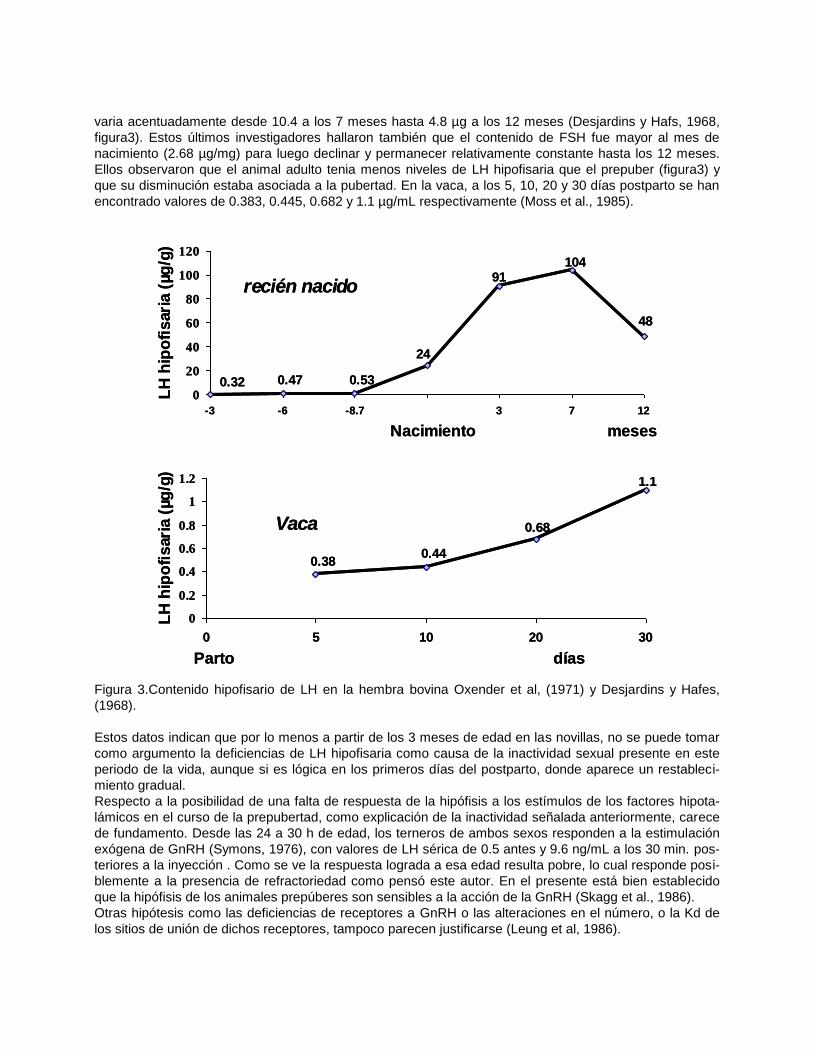
varia acentuadamente desde 10.4 a los 7 meses hasta 4.8 µg a los 12 meses (Desjardins y Hafs, 1968,
figura3). Estos últimos investigadores hallaron también que el contenido de FSH fue mayor al mes de
nacimiento (2.68 µg/mg) para luego declinar y permanecer relativamente constante hasta los 12 meses.
Ellos observaron que el animal adulto tenia menos niveles de LH hipofisaria que el prepuber (figura3) y
que su disminución estaba asociada a la pubertad. En la vaca, a los 5, 10, 20 y 30 días postparto se han
encontrado valores de 0.383, 0.445, 0.682 y 1.1 µg/mL respectivamente (Moss et al., 1985).
104
48
91
0.32 0.47 0.53
24
0
20
40
60
80
100
120
-3 -6 -8.7 3 7 12
Nacimiento meses
LH
hip
ofi
sari
a (
µg
/g)
recién nacido
1.1
0.380.44
0.68
0
0.2
0.4
0.6
0.8
1
1.2
0 5 10 20 30
Parto días
LH
hip
ofi
sari
a (
µg
/g)
Vaca
104
48
91
0.32 0.47 0.53
24
0
20
40
60
80
100
120
-3 -6 -8.7 3 7 12
Nacimiento meses
LH
hip
ofi
sari
a (
µg
/g)
recién nacido
1.1
0.380.44
0.68
0
0.2
0.4
0.6
0.8
1
1.2
0 5 10 20 30
Parto días
LH
hip
ofi
sari
a (
µg
/g)
Vaca
Figura 3.Contenido hipofisario de LH en la hembra bovina Oxender et al, (1971) y Desjardins y Hafes,
(1968).
Estos datos indican que por lo menos a partir de los 3 meses de edad en las novillas, no se puede tomar
como argumento la deficiencias de LH hipofisaria como causa de la inactividad sexual presente en este
periodo de la vida, aunque si es lógica en los primeros días del postparto, donde aparece un restableci-
miento gradual.
Respecto a la posibilidad de una falta de respuesta de la hipófisis a los estímulos de los factores hipota-
lámicos en el curso de la prepubertad, como explicación de la inactividad señalada anteriormente, carece
de fundamento. Desde las 24 a 30 h de edad, los terneros de ambos sexos responden a la estimulación
exógena de GnRH (Symons, 1976), con valores de LH sérica de 0.5 antes y 9.6 ng/mL a los 30 min. pos-
teriores a la inyección . Como se ve la respuesta lograda a esa edad resulta pobre, lo cual responde posi-
blemente a la presencia de refractoriedad como pensó este autor. En el presente está bien establecido
que la hipófisis de los animales prepúberes son sensibles a la acción de la GnRH (Skagg et al., 1986).
Otras hipótesis como las deficiencias de receptores a GnRH o las alteraciones en el número, o la Kd de
los sitios de unión de dichos receptores, tampoco parecen justificarse (Leung et al, 1986).

Queda por pensar, según sugieren Mc Leod et al. (1984) que la baja frecuencia de los episodios de LH,
en novillas prepúberes sea motivada por una inactividad hipotalámica, lo cual resulta lógico al observar
que el suministro exógeno de GnRH induce el aumento de los pulsos de la hormona, por lo tanto debe
existir una pulsación endógena deficiente de este factor hipotalámico. En tal caso hay que analizar los
factores que modulan la secreción de las gonadotropinas y los factores liberadores; el mecanismos de
retroalimentación.
2.2.3 Mecanismo de retroalimentación (feedback)
La regulación de la secreción de hormonas depende de varios mecanismos. La liberación de una hormo-
na en particular puede tener más de un mecanismos de control, donde la hipófisis ocupa un lugar central,
ya que puede recibir influencias tanto por vía hipotalámica como por los receptores periféricos.
Un concepto útil en la modulación de la secreción hormonal es la idea de la retroacción (feedback o ser-
vomecanismo), por lo cual el efecto provocado por una hormona tiende a aumentar o anular la ulterior
secreción de la misma. La situación más sencilla es la establecida entre dos variables (hormonas), según
Rasmussen (1974): Si A = f (B) y B = f (A), existe una relación de retroacción entre las dos. Si la concen-
tración o el efecto de A aumenta cuando aumenta B, la retroacción es positiva, mientras que si A disminu-
ya cuando B aumenta, la retroacción es negativa.
De acuerdo entre quienes se establezca la relación en los órganos del eje SNC - glándula efectora el
mecanismo feedback se clasifica en:
Largo, que es la relación entre órgano efector periférico y eje hipotálamo - hipófisis.
Corto, se enmarca de hipófisis a hipotálamo
Ultracorto, dentro del hipotálamo o la hipófisis
Estas ideas fueron tomadas de Botella (1982) y Piva et al (1983).

Tabla 2. Tipo de control de la secreción endocrina
1 HUMORAL
1.1 Concentración de metabolitos
1.2 Concentración de otra hormona
1.2.1 Circuito largo
1.2.2 Circuito corto
1.2.3 Circuito ultracorto
2 NERVIOSO
2.1 Conexión nerviosa periférica
2.2 Conexión hipotalámica (hipófisis)
3 GENETICO
Según Gorbman y Bern (1962) con ligera modificación
Con respecto a la pubertad, todo parece indicar que el servomecanismo juega un papel fundamental para
su desencadenamiento.
Desde épocas tempranas Kallas (1929) demostró que si se castraba a una compañera de ratas hembras
parabióticas sexualmente inmaduras, la otra experimentaba maduración sexual precoz. Presumiblemente
la castración producía un aumento de la producción de gonadotropinas, con estimulación de las gónadas
en la rata intacta. Luego Byrnes y Meyer (1951) comprobaron que si se inyectaba dosis muy pequeñas de
estradiol al animal castrado, sexualmente inmaduro, parabiótico hembra podía evitarse la pubertad precoz
en su compañera intacta.
Se ha confirmado bien que la castración induce un incremento significativo de gonadotropinas en sangre
(Anderson et al., 1985). Lo cual refleja que las gónadas ejercen una marcada inhibición sobre la liberación
de dicha hormona. Entre los esteroides ováricos, los estrógenos son los más potentes inhibidores (Weick
y Noh, 1984).
De acuerdo a Staigmiller et al (1979) ninguno de los modelos de animales experimentales hasta ahora
poseen su mecanismo de retroacción positivo al estradiol desarrollado al momento del nacimiento. Esa
sensibilidad del eje hipotálamo - hipofisario se desarrolla a partir de los 3 a 5 meses de edad (Staigmiller
et al, 1979; Schillo et al, 1983b). Sin embargo, el control de retroacción negativa ejercido por el estradiol
existe durante todo ese tiempo, siendo en el momento de la pubertad cuando disminuye (Schillo et al,
1982; Day et al, 1984). Glencross (1984) comprobó en novillas, que una disminución de la sensibilidad a
la acción negativa del E2 se lograba ocho días antes del primer ciclo estral mientras que Day, et al (1987)
planteó que esta disminución ocurrió 50 días antes de la pubertad.
¿Qué mecanismo es el que desencadena el inicio de la pubertad?
Los terneros responden con un acentuado desarrollo folicular y ovulación a la estimulación con gonado-
tropinas exógenas (Onuma et al, 1970; Seidel et al, 1971) a pesar de que Spitcer y Echternkamp (1986)
aclaran que en el bovino no se conoce el tiempo en que se obtiene un completo desarrollo de las enzimas
esteroidogénicas, ni de los receptores a gonadotropinas en el folículo ovárico. Además la hipófisis de las
terneras es sensible a la acción gonadotrópica y responden al feedblack positivo del estradiol exceptuan-
do la etapa neonatal temprana (Evans, et al, 1992, 1994), por lo tanto la hipótesis más aceptada es que la
pubertad comienza cuando decrece la sensibilidad a la retroacción negativa del estradiol (Enomoto y
Minowada, 1997).
Quedaría por saber qué es lo que conlleva a esa modificación, para lo cual habría que tener en cuenta la
alimentación, fotoperíodo, ciclo lunar, etc. Todo indica que el estado metabólico del animal es muy impor-
tante así como el fotoperíodo en los animales estacionarios. En la actualidad el mecanismo intrínseco que
conduce a ese proceso está vinculado con interacciones de estructuras neuronales y células gliales de la
línea astrocítica las cuales afectan la función neuronal vía un mecanismo de señalización célula a célula
que involucra a factores de crecimiento y sus receptores entre los cuales se han identificado al EGF (Fac-
tor de crecimiento epidérmico), al TGF (Factor de crecimiento transformante) y al NDF (Factor de dife-
renciación neuras) los que son producidos en los astrocitos hipotalámicos estimulando la liberación de
GnRH a través de un intermediario glial como es la Prostaglandina E2 que activa directamente las neuro-

nas secretoras de GnRH (Ojeda, et al, 1997, Ma y Ojeda, 1997). Otras evidencias indican que la leptina
juega un papel fundamental en este mecanismo, así hay evidencias que la misma estimula a los gonado-
trofos no solo en los periodos puberales y de madurez sino en el juvenil antes de la pubertad haciendo
pensar que modula la sensibilidad de los gonadotrofos hasta la aparición del estímulo de la GnRH y pu-
diera ser el factor que genera la aparición de la pubertad (Tezuka, et al. 2002).
En este sentido además se ha planteado la asociación existente entre la leptina y la secreción tanto de
GnRH como de LH (Zieba, et al. 2004). Sin embargo estos mismos autores notificaron que la administra-
ción crónica de leptina no indujo la pubertad ni alteró el patrón endocrino en novillas alrededor del inicio
del tiempo esperado de la pubertad (Maciel, et al. 2004). Aunque si dejaron aclarado que el tratamiento
con leptina en novillas prepúberes en estado de ayuno evitó la disminución de la frecuencia de pulsos de
LH y modificó la respuesta de la LH mediada por GnRH (Maciel, et al. 2004).
Así podríamos hipotetizar que el inicio de la pubertad transcurriría de la siguiente forma: Disminución de la
sensibilidad del eje hipotálamo - hipófisis a la retroacción negativa del estradiol,(figura 4), determinado por
señales que provienen del estado metabólico del animal probablemente a través de la leptina, aumento de
la frecuencia pulsátil de GnRH y de LH con el consiguiente desarrollo folicular, producción de estrógenos,
aparición del primer celo, descarga preovulatoria de LH y ovulación. Apareciendo de esta forma la activi-
dad cíclica que caracteriza la etapa reproductiva.

-130 días
-60 días
-40 días
-20 días
Primera
Ovulación
Receptores de E2
(Hipotálamo e Hipófisis)
Variable
Retroacción del E2
sobre la secreción de LH
Secreción Pulsátil
de LH
Secreción de E2 y
peso uterino
Figura 4. Modelo del control endocrino de la pubertad en novillas según Kinder et al. (1987).
3 Aspectos Productivos
3.1 Pubertad retardada.
La mayoría de las novillas tienen potencial suficiente para alcanzar la pubertad y cubrirse satisfactoria-
mente al año de edad, si se le proveen una nutrición y manejos adecuados. El costo para hacer esto pue-
de variar grandemente entre razas y entre tipos dentro de una misma raza. Las novillas con capacidad la
pubertad a edades tempranas alcanzan tales propósitos a menor costo que aquellas con edad tardía a la
pubertad (Patterson, et al, 1992).
Se ha reportado la existencia de una regresión entre edad a la pubertad con peso al primer parto, produc-
ción de leche a la primera lactancia así como con el porciento de grasa en la leche, de lo cual se des-
prende que el retraso de la pubertad conduce a una disminución de la eficiencia productiva en la industria
pecuaria. Las diferencias de edad a la pubertad en diferentes regiones geográficas dependen del sistema
de manejo, disponibilidad y calidad del forraje, así como de la adaptación del tipo racial a las condiciones
ambientales específicas. La edad a la pubertad adquiere un valor productivo cuando las novillas se gestan
para parir a los dos años y los sistemas de explotación imponen épocas reproductivas limitadas, de modo
que se postula que las novillas tienen que alcanzar la pubertad a los quince meses al menos con vistas a
lograr el primer parto a los veinticuatro meses, sin embargo alrededor de un 35% no logra este objetivo
por retraso puberal.
El anestro en novillas en nuestro país se corresponde con una maduración sexual tardía o pubertad retar-
dada. La causa fundamental de esta situación reside en una inadecuada alimentación y un manejo defi-
ciente durante la etapa de desarrollo somático de los animales. Los datos obtenidos respecto a la edad y
el peso con los cuales llegan las novillas a incorporarse a los servicios de IA y al primer parto lo demues-
tran.
Hay empresas que incorporan a las hembras con 18 meses de edad y 336 Kg. de PV, lo que es reflejo de
una ganancia en peso por animal de 0.62 Kg. / d. En el Libro de Metodología de Balance Alimentario para
el Ganado Vacuno en Cuba (1975) se indicó que en nuestro país se debe seguir como criterio para gestar
nuestras novillas lecheras un peso aproximado de 230 Kg. y 18 meses de edad. Zamora et al (1985) ma-
nejando bien los pastos y la carga/ha consiguió la primera inseminación en novillas a una edad (16.5 -
17.7 meses) y peso (367 - 376 Kg.) deseables. Estas edades y pesos es posible lograrlos siempre que la
ganancia diaria de peso sea igual o superior a 0.5 Kg. (Metodología b.a., 1975, Santos y Pereiros, 1982;
Zamora et al., 185; Zamora y González, 1986).

3.2 Inducción de la pubertad.
3.2.1 Corrección dietética
La influencia del ambiente sobre la secuencia de eventos que conducen a la pubertad está dictada, princi-
palmente, por el estado nutricional y los efectos vinculados con la tasa de crecimiento y el desarrollo. Un
buen plano nutricional no solo tiene consecuencias con una pubertad a edad temprana sino que influye el
crecimiento y desarrollo mamario en novillas lecheras (Niezen, et al, 1996).
Así en nuestro país se han realizado muchos ensayos en este sentido, haciéndose corrección en el nivel
energético (Simón y Hernández, 1977; Tarrero y Perón, 1983; Alonso et al., 1985) proteico (Pérez y Ramí-
rez, 1986; Zamora y González, 1986) y vitamino - mineral (Perón et al, 1976; Contreras et al, 1986; Gon-
zález et al, 1986) obteniéndose en todos los casos un saldo positivo en el porcentaje de animales que
entran en la pubertad o se gestan o logran el parto a una edad temprana.
El plano energético de la ración parece tener una influencia grande y más rápida que el proteico en la
inducción de la ciclicidad estral (Lam, 1986), encontrándose una correlación inversa entre la edad de pu-
bertad y la ingestión energética (Barash, et al, 1994).
En países con clima tropical existen dos estaciones en el año caracterizadas fundamentalmente por la
cantidad de precipitaciones. Hay que tener en cuenta que la estación de sequía en este medio, constituye
la época más crítica desde el punto de vista de la disponibilidad de alimentos y es en este momento don-
de más se justifica la suplementación si no se dispone de un buen pasto y su tecnología de cultivo. Se ha
demostrado que el suministro de una dieta elevada en concentrados permite un destete temprano y pro-
voca un incremento en la frecuencia de los pulsos de LH (Gasser, Grum et al. 2006), acelera de la madu-
ración ovárica (Gasser, Burke et al. 2006) y que esto se logra por disminución del feedback negativo que
ejerce el estradiol sobre la LH (Gasser, Bridges et al. 2006).
Por otra parte, hay algunos antibióticos que ejercen una profunda influencia en el metabolismo ruminal
como es el caso de la Monensina y otros como la Ivermectina que son capaces de provocar un adelanto
de la pubertad (Purvis y Whitier, 1996). La Monensina produce un cambio en la relación molar de AGV en
el rumen sin alterar la concentración total. Ella aumenta el porciento molar de propionato a expensas de
acetato y butirato. Este ionóforo influye en la reproducción dando un estado nutritivo más favorable, dismi-
nuyendo los requerimientos del animal (Mason y Randel, 1983) y produciendo cambios acentuados en el
sistema endocrino (Randel et al, 1981 y Hardin y Randel, 1983a) lo que provoca una disminución de 21
días en la aparición de la pubertad (Lalman, et al, 1993). En el caso de la Ivermectina solo se sabe que
este efecto es independiente de su acción antiparasitaria ya que no se observó que esto haya sido pro-
ducto de un aumento en la ganancia de peso (Larsen, et al, 1995).
Por otra parte el empleo de anabólicos no esteroideos como la STH no han mostrado que adelanten la
edad de la pubertad (Hall, et al, 1994), aunque si la aplicación de anticuerpos contra el factor liberador de
dicha hormona retrasa la pubertad en novillas (Schoppee, et al, 1996) probablemente por modificación de
la sensibilidad al efecto inhibitorio que tiene el E2 sobre el eje hipotálamo hipófisis (Schoppee, et al, 1995).
3.2.2 Melatonina
La melatonina es el prototipo de una familia de compuestos biológicamente activos, los metoxindoles,
producidos por la glándula pineal de los mamíferos, y considerada como un transductor neuroendocrino
(Wurtman, 1983). Así se le ha prestado una gran atención por su posible función moduladora del inicio de
la pubertad (Yellon et al., 1985; Stanisiewski et al., 1986). En roedores la pinealectomía avanza la puber-
tad (Rolkin, 1971), observándose lo contrario en oveja (Kennaway et al., 1985). En este último trabajo se
concluyó que en ovinos, la pineal juega un papel clave como mediador de la señal hormonal que controla
la pubertad.
En esta especie ha sido posible la inducción de la actividad reproductiva con el suministro oral o parente-
ral de melatonina (Roche et al., 1984; Hanrahan et al., 1985; Márquez et al., 1985). Similares resultados
se indicaron en cabras (Contestabile et al., 1985).
Según Thibaut y Lovasseur (1974), se ha postulado que la secreción de la pineal puede modular la sensi-
bilidad de los receptores esteroideos a nivel del cerebro.
En cuanto al vacuno se informó que el empleo de suero antipineal reportaba buenos resultados en la in-
ducción del celo en novillas anéstricas (Portal et al., 1978 a, b), aunque otros resultados fueron menos
prometedores (Portal et al., 1978c).

Similar a otros mamíferos, la pineal bovina posee un patrón de actividad diurna, siendo la secreción de
melatonina mayor durante la oscuridad (Hedlund et al., 1977; Martín et al., 1983). La melatonina puede
causar supresión de la secreción de LH en vacas castradas (Rhodes et al., 1979); aunque no altera los
niveles basales, ni la secreción de progesterona in vitro inducida por la LH (Battista y Condon, 1986).
Además, la pineal bovina contiene una variedad de péptidos implicados en el proceso productivo, tales
como un tripéptido antigonadotrópico (Orts et al., 1980), arginina - vasopresina (Pavel, 1971) y GnRH
(White et al., 1974).
Stanisiewski et al., (1986), estudiando terneros pinealectomizados en relación con la luz, la concentración
de PRL y testosterona, concluyeron que la pineal estaba involucrada en el inicio de la pubertad en los
toros. Se ha demostrado que a más horas luz más temprana es la aparición de la pubertad posiblemente
a través de cambios en la foliculogénesis, secreción de LH y mayor tasa de crecimiento, estos efectos
pueden estar mediados por la retina y la glándula pineal (Hansen, 1985).
3.2.3 Hormona liberadora de gonadotropinas (GnRH)
El factor liberador de gonadotropinas (LH-RH/FSF-RH, LH-RH,LRF. GnRH) es un decapéptido de PM =
1182 daltons, biosintetizado y liberado por el hipotálamo y conducido por el sistema porta hasta la ade-
nohipófisis, donde produce la descarga de FSH y LH.
El aislamiento de la GnRH se logró simultáneo e independiente en porcino (Schally et al., 1971) y ovino
(Amoss et al., 1971). Determinándose rápidamente su estructura (Matsue et al., 1971a) y síntesis (Matsuo
et al., 1971); Burgus et al., 1972).
En general, después de la administración de GnRH la respuesta de la LH es superior a la de FSH (Barnes
et al., 1980 Bolt, 1981), y en ocasiones dependiendo de la frecuencia y magnitud de los pulsos de GnRH
suministrados (no se han observado cambios en las concentraciones periféricas de FSH (Clarke et al.,
1984a; Clarke y Cummins, 1985a; Rieger y Rawlings, 1985).
La GnRH posee una vida media muy breve, varía entre 4 y 7 min. (Redding et al., 1972; Nott et al, 1973)
lo cual motiva que la respuesta hipofisaria medida por los niveles de LH no dura, comúnmente, más de
seis horas (Mollin et al., 1975; Barnes et al., 1980; González-Menció et al., 1980; Faure et al, 1986a).
Esto se ha tratado de solventar inyectando la GnRH con un medio que la libere lentamente, como son las
cápsulas de gelatina o de Metilcelulosa (Vincent, 1983; Trzel, 1983), o administrándola en inyecciones
repetidas (Mayer et al., 1978) o en forma de infusiones (Chakraborty et al., 1974; Skagg et al., 1986).
Los efectos que se desean obtener con el empleo de GnRH dependen de muchos factores: dosis (Braun
et al., 1985; Rodway y Swift, 1985) y tipos de agonistas aplicados (Braun et al., 1985; McMillan et al.,
1985), forma y vías de administración; sexo (Zolman et al., 1973) y estado reproductivo del animal (Caon-
voy, 1973; Dun et al., 1985), nivel de esteroides endógenos (Convoy, 1973, 1973; Zaied et al., 1980;
Growder et al., 1983; Braun, 1985); existencia y tipo de pool de LH en hipófisis (Madej et al., 1980; Azzani
et al., 1983), así como de un estado refractario de la glándula (Rippel et al., 1974; Mayar et al., 1978), lo
cual se debe en parte a la poca capacidad de los receptores de las células adenohipofisarias de unir la
GnRH después de haberse expuesto a un tratamiento crónico (Mayer et al., 1978; Lofated et al., 1981).
La GnRH se ha utilizado para sincronizar la ovulación (Fernández, 1979) y mejorar el índice de gestación
o realizar la IA a un tiempo fijo (Goldbeck, 1976; Spitzer, 1982). También se emplea para eliminar los
quistes foliculares del ovario con buenos resultados (Bierschwal et al., 1974; Nesler et al., 1978).
Respecto a la inducción de ciclicidad en animales sexualmente inactivos, Conveu (1973) fue el primero en
sugerir que la GnRH podía utilizarse para adelantar la pubertad y el ciclo estral después del parto. Sin
embargo, un gran número de resultados no apoyan tal sugerencia, cuando se emplea la GnRH sola para
inducir la pubertad en bovino (Gil et al., 1981a; McLeod et al., 1985 Skagg et al., 1986: Larson, et al,
1993). Sin embargo el empleo de anticuerpos antiGnRH provoca un retardo marcado de la pubertad en
dependencia de la forma de aplicación (Wattemann y Castree, 1994, Prendiville, et al, 1995)
La falta de estimulación ovárica y la aparición de fases lúteas anormales ha sido común después del tra-
tamiento con GnRH (Rippel et al., 1974a; James et al., 1975; Mawhinney y Roche, 1980; Ball y Lamming,
1983; Dobson, 1983). Aunque algunos investigadores informan haber promovido actividad ovárica con
dicho tratamiento (Secane y Bravo, 1986).

Se admite que un prerequisito para lograr funcionalidad del ovario con el uso del GnRH, es la existencia
de folículos desarrollados en el momento del tratamiento, cuyo tamaño no debe ser menor a 10 mm de
diámetro (Garverick et al., 1980; Zaied et al., 1980).
En la actualidad, para lograr la promoción de crecimiento folicular, ovulación y desarrollo normal del cuer-
po lúteo, existen dos enfoques con el empleo de la GnRH.
a) Suministrar esta hormona de forma pulsátil, tratando de imitar el patrón de secreción fisiológico (Glen-
cress, 1985; Verstermans y Walton, 1985).
b) Realizar un pretratamiento con otras hormonas. El más usual ha sido la progesterona, con muy buenos
resultados (Legan et al., 1985; Hunter et al., 1986); también se utilizó la PMSG, (Nellin et al., 1975).
3.2.4 Gonadotropina del suero de yegua gestante o Gonadotropina coriónica equina (PMSG,
eCG, PMS, Prolan A).
La PMSG es una gonadotropina extrahipofisaria, descubierta por Cole y Hart (1930). Pertenece a las gli-
coproteína, con peso molecular de 68000 - 75000 daltons. Es producida por las copas endometriales (en-
dometrial cups) de la yegua gestante. Aparece en la circulación materna el día 40, alcanza el máximo
entre los 60 y 80 y desciende a los 120 días de gestación (Allen y Morr, 1972).
Dado su gran contenido de ácido siálico tiene una vida media larga (50 - 120h), siendo medible en sangre
hasta 10 días después de su administración (Schams et al., 1978).
Posee tanto actividad FSH como LH, con predominio de la primera, cuyo cociente varía de acuerdo al tipo
de raza y días de gestación (González Menció et al., 1978). Su mecanismo de acción se ejecuta a través
del AMPc (Gosling et al., 1979); promoviendo la formación de cuerpos lúteos secundarios y producción de
progesterona en la yegua. Para Marina y Machado (1976), la PMSG actúa sobre los dos componentes del
desarrollo folicular (masa celular y % de antrum), secreción y multiplicación de las células de la granulosa
y sobre toda la población de folículos; aunque parece que la PMSG no es capaz de recuperar los folículos
atrésicos (Dott et al., 1979).
La PMSG funciona con un inductor de la liberación de LH (Pelletier y Thimonier, 1975). Por inducir creci-
miento folicular, liberación de LH y superovulación ha sido utilizada intensivamente en la especie ovina
(Thimonier y Cognie, 1971) y bovina (Mauleon et al., 1970; Bellows y Short, 1972). Después de la inte-
rrupción de un tratamiento con progesterona la PMSG produce un pico de LH similar al obtenido espontá-
neamente (Pelletier y Thimonier, 1969); aunque dicho pico coincide con el inicio del celo, lo cual es con-
trario al ciclo normal, donde el pico ocurre 8 h más tarde (Schams y Butz, 1972). La liberación de LH res-
ponde a la rápida producción de estradiol causando por el desarrollo folicular (Schams et al., 1978; Wet-
terman et al., 1982).
La PMSG ha sido la hormona más ampliamente empleada en nuestro país para la inducción del celo en
novillas (Vichot et al., 1976; Alba y Revuelta, 1981; Gil et al., 1981a Alcalá et al., 1983; Faure et al., 1983;
Roque, 1984; Martínez y Alcalá, 1986) y vacas (Rysanek y Alba, 1975; Cruz et al., 1983; Gil, 1984; Jimé-
nez et al., 1986). Todos los resultados han sido promisorios, lográndose hasta un 63% más de animales
gestados con una sola aplicación (2000 UI) de PMSG (Gil, 1984), sin embargo la reducción (1/4) de esta
dosis combinada a 50 mg de progesterona previamente condujo a la obtención del 30% de eficiencia en el
tratamiento (Faure et al., 1983).
El tratamiento con PMSG sola o asociada a HCG en animales prepúberes produce aumento de la activi-
dad ovárica y superovulación (Debirian y Baker, 1975; Schurrbush et al., 1980; Paterson y Lindsay, 1981;
Davis y Johnstone, 1985); pero lograr que los animales tratados continúen ciclando no resulta fácil. Estos
animales hacen fase lútea por más de 20 días con valores de P4 superiores a 85 ng/mL (Spilman et al.,
1973; Bonavera et al., 1986), quizás debido a la ausencia del factor luteolítico (Spilman et al., 1973). Esto
pone en evidencia que le inicio del proceso reproductivo tiene una edad límite.
La PMSG se utiliza también en la sincronización del celo en animales cíclicos (Wagner et al., 1973; Ro-
che, 1975) y en el diagnóstico de gestación por pruebas inmunológicas (Zemjanis, 1974).
El empleo práctico de la PMSG se ve afectado por diversos factores: se consideró fundamental la carac-
terística del lote (Betteridge y Mitchell, 1974) no confirmándose posteriormente (Schams et al., 1978;
Newcomb et al., 1979); las proporciones de FSH y LH que contenga la hormona (Steward et al., 1976;
González Mencio et al., 1978), siendo probablemente el más importante la población de folículos antrales
presente en los animales tratados (Saumande, 1978).

Aunque, dando el largo período de vida media que posee la PMSG, merece particular atención la capaci-
dad que tenga la hormona de inducir desensibilización y pérdidas de receptores en las células efectoras
(Gosling et al., 1979).
Un aspecto a tener en cual usar PMSG es la formación de anticuerpos que se produce con inyecciones
repetidas, creando así una refractoriedad ovárica (Willer et al., 1953); aunque no fue observada por
Schams et al., (1978).
La PMSG constituye una hormona muy útil en la práctica veterinaria. Los resultados en cuanto a su-
perovulación mejoran con el uso de PGF2 y la neutralización de la hormona con anticuerpos. Asimismo,
para la inducción del estro, el pretratamiento con progesterona permite disminuir la dosis de PMSG y los
resultados son más favorables (Faure, et al, 1996).
3.2.5 Hormonas esteroides
La vida reproductiva está directamente gobernada por las hormonas gonadales, tanto en el macho como
en la hembra. En el caso de esta última se sabe que el ovario no produce solamente dos hormonas (es-
trógenos y progesterona) como durante muchos años se creyó; sino que en la actualidad se ha compro-
bado que el ovario produce 5 tipos diferentes de hormonas: 3 esteroides (gestágenos, andrógenos, y es-
trógenos) y 2 peptídicas (oxcitocina y relaxina) (Fields, 1984).
Químicamente las hormonas esteroides derivan de un precursor común, teniendo varias fuentes para su
biosíntesis como son: ovario, testículos, glándula adrenal, unidad feto placentaria y algunos tejidos efecto-
res (Botella, 1982). En la sangre circulan libre (biológicamente activos) o unidos a proteínas y su tasa de
depuración metabólica (MCR) ha tenido poca atención, no obstante ser importante. Durante el anestro se
señaló que la MCR se encuentra elevada para la progesterona (Bodford et al., 1972), y los resultados en
novillas ovariectomizadas que recibieron diferentes dosis de la hormona, también surgieren lo mismo
(Alonso et al., 1986 a).
Que los animales posean un patrón de liberación de FSH y LH cíclico (hipotálamo cíclico) o tónico (hipotá-
lamo tónico) depende de si el SNC recibe el influyo de las hormonas esteroides durante el período de
diferenciación (Thibault y Levasseur, 1974). Esto se ha explicado como la pérdida de la sensibilidad del
cerebro a la retroalimentación positiva de los estrógenos, cuando se expone a la acción de los esteroides,
lo cual conduce al comportamiento sexual masculino (Clarine y Searenuz, 1978). Tal es el principio de las
vacas androgenizadas utilizadas como celadoras (Kiser et al., 1977; Hackert y Lin, 1985).
Para ejecutar sus acciones las hormonas esteroides necesitan de la proteína transportadora y un receptor
citosólico en la célula efectora (Botella, 1982); aunque algunas funciones se promueven sin receptores
(Siiteri y Febres, 1983).
Los esteroides penetran en la célula por difusión pasiva ( Jenesen, 1979) y se unen al receptor en el cito-
plasma (Clark, et al., 1977), esta unión induce cambios estructurales al receptor, lo que provoca que el
complejo pase al núcleo por un proceso de translocación (Clark et al., 1977; Kaltonbach y Dunn, 1980). La
translocación depende en parte de las variaciones en los niveles periféricos de la hormona y de la presen-
cia del receptor (Mainwaring, 1979). Se señala que tal proceso depende de la temperatura al inducir un
cambio del receptor a una forma activa que le permite unirse a la cromatina (Jensen, 1979).
Ya en el núcleo el complejo esteroide - receptor promueve la síntesis de RNA-m específicos, los cuales
pasan al citoplasma e inducen la síntesis de proteína (Clark et al., 1977; Jensen, 1979; las cuales son
responsables de estimular las actividades biológicas en las células (Niswender et al., 1974).
3.2.5.1 Estrógenos
La definición de estrógenos no es una definición química, sino biológica. Son derivados alcohólicos e hi-
drocarburos aromáticos (naturales o artificiales) capaces de promover el celo o decamación vaginal de la
rata y cambios proliferativos en el tracto genital de las hembras de todos los vertebrados (Botella, 1982).
El 17-beta-estradiol es el estrógeno natural más potente.
En la hembra las principales funciones de los estrógenos se ejercen en los procesos reproductivos. Ellos
son necesarios para la expresión del estro (Irvin et al., 1978; Cook et al., 1985). A nivel uterino inducen el
desarrollo del tejido endometrial (Henricks y Mayer, 1977; Dorrington, 1979) y la metaplasia del tejido va-
ginal; estimulando la síntesis de proteína y RNA. La gran hiperemia en los órganos reproductivos parece
mediana por la acción de las PGs y no por receptores (Siiteri y Febres, 1983).
La infusión intrauterina de estradiol y PGE-2 en novillas poseen un efecto luteotrópico en la gestación
temprana (Reynolds et al., 1983). Los estrógenos potencian el efecto de la oxcitocina y las PGs durante el
parto y la implantación (Baird, 1972; Kaltenbach y Dunn, 1980).
El estradiol parece ser un regulador importante de la secreción infrafolicular de progesterona (Fortune y
Hansel, 1979), a la vez que promueve la síntesis de receptores a esta última en los órganos efectores
(Siiteri y Febres, 1983).
También los estrógenos a dosis bajas producen liberación de gonadotropinas y a dosis altas las inhiben.
Se ha comprobado que pueden provocar descargas de LH (Faure et al., 1986) y FSH (Reeves et al.,
1974; Peters, 1984). En muchos casos se ha observado un efecto bifásico.
El mecanismo por el cual los estrógenos producen este efecto no está bien aclarado. En bovino existen
receptores a estas hormonas a nivel hipotalámico e hipofisario (Kahwanago et al., 1969; Clarke et al.,
1984).
Quizás actúen en este sentido aumentando la liberación de GnRH, del hipotálamo (Clarke y Cummins,
1985b) o la concentración de receptores de GnRH en la hipófisis (Schoenemann, 1985), o funcionen co-
mo una hormona liberadora (Wildt et al., 1981). Bajo estas acciones es que el estradiol es el responsable
del pico preovulatorio de LH.
Con respecto a la aplicación de los estrógenos, Melampy et al., (1957) indujeron el celo de la vaca con
una dosis < 0,25 mg de benzoato de estradiol (BE2) y Ray (1965) concluyó que la dosis mínima para lo-
grar conducta receptiva era de 0,3 mg. Así, Nancarrow y Radford (1976) con el empleo de 0.5 mg de BE2
provocaron un 100% de presentación de celo en las vacas, lo cual ha sido confirmado por nosotros (Faure
et al., 1982, Faure,1987) en novillas con el uso de 2; 1 y 0.5 mg , aunque los resultados de la tasa de
gestación al primer servicio fueron pobres. Para solucionar estos inconvenientes se ha intentado el uso
combinado con progestágenos. González-Padilla et al., (1975b) con una formulación de 17B-E2 unido a
la progesterona, pudieron estimular en un 75% la aparición de la pubertad en novillas. Similares resulta-
dos se obtuvieron con el implante de progesterona y valerato de estradiol (González-Padilla et al., 1975c;
Burfening, 1979); aunque parece que la edad (menor de 13 meses) es un factor limitante en la inducción
de la pubertad (Burfening, 1979; Spitzer, 1982) y pasada esa edad el pero es el más influyente en los
resultados.
Nosotros hemos demostrado, en novillas anéstricas, que la inyección de Cloprostenol en la submucosa
vulvar en el momento de la inseminación mejora significativamente los resultados obtenidos con un es-
quema de inducción del estro con P4 y E2. (Tabla 3)
Tabla 3. Presentación del celo y tasa de gestación en dos grupos de tratamiento y un control (Gil-
Figueredo y Faure, 1989)
Animales en Estro Tasa de Gestación
Grupo Categoría n n % n %
I. P4 + BE2 + Clop. Vacas 47 35 74d 20 57ª
Novillas 30 25 83ª 17 68ª
Total 77 60 78ª 37 62ª
II. P4 + BE2 Vacas 47 36 76ª 9 25b
Novillas 30 25 83ª 7 28b
Total 77 61 79ª 16 26b
III. Control Vacas 50 14 28b 10 71ª
Novillas 40 10 25b 7 70ª
Total 90 24 27b 17 71ª
Letras diferentes difieren con p<0.05
Los animales recibieron 50 mg de P4, 48 h después 0.5 mg de benzoato de estradiol y en el caso del Gru-
po I, 50 µg de Cloprostenol
Signoret (1967) en la cerda observó una relación lineal entre las dosis de BE2 y el tiempo de duración de
la receptividad sexual lo cual fue confirmado en vacuno (Cook et al., 1985) a pesar de que la dosis mayor

prolongó el celo. En vacas ovariectomizadas no se observó celo con dosis única (0,1 ó 0,25 mg de BE2 o
repetidas (3 x 50 mcg); pero sí se obtuvo un 92% de las vacas con síntomas estrales con inyecciones de
3 x (0.25 ó 0,5 mg), las cuales dieron niveles de E2 en plasma similares a las fases de proestro y estro
(Nessan y King, 1981).
Con otras asociaciones de estrógenos a implantes de progesterona ha sido posible promover conducta
estral y actividad ovárica en novillas acíclicas (Rajamahendran et al., 1981; De Armas et al., 1983). Asi-
mismo se han empleado cipionato de estradiol (Holy y Hrivnak, 1965) y diétilestilbestrol (Gibbons, 1963;
Hernández et al., 1976) El DEB parece no participar en la liberación de gonadotropinas, pero estimula el
nivel de receptores a FSH en ovario; estos receptores a FSH probablemente no dependan de la tasa de
estrógenos, contrario a lo observado para receptores de LH (Moudgal y Vidyashankar, 1984).
Los estrógenos en animales cíclicos, pueden producir variados efectos (luteotrópicos, antiluteotrópico,
luteolítico, antiluteolítico) los cuales dependen de la fase del ciclo estral en que se administren (Lemon,
1975).
Se ha comprobado que ellos unidos con gestágenos pueden disminuir la fertilidad en celos sincronizados
(Wiltbank et al., 1971: Roche, 1974; Hansel et al., 1975), pero unidos a PGF2 la fertilidad resulta normal
(Dailey et al., 1986; Figueredo et al., 1986). Estas acciones deletéreas sobre la fertilización pueden res-
ponder a disturbios en el transporte del óvulo (Humphrey, 1968) o en el transporte y supervivencia del
esperma (Robinson, 1973).
Referente a la promoción de actividad cíclica en los animales los estrógenos pueden seguir dos vías en
sus acciones: desencadenar la liberación de gonadotropinas, las cuales estimulan el crecimiento folicular
con producción de esteroides y conducta estral, o por lo contrario inducen directamente el celo sin cam-
bios ováricos. Estos resultados dependen de factores tales como tipo y dosis de estrógeno, duración de la
administración, el vehículo de inyección, el sinergismo con otras hormonas, raza y estado reproductivo de
los animales tratados.
3.2.5.2 Progestágenos
Son las sustancias capaces de provocar la reacción prográvidad o progestacional en la mucosa uterina
para la anidación del huevo. Su principal representante (la progesterona) fue extraída del cuerpo lúteo y
cristalizada por Butenandt (1934).
El ovario es la mayor fuente de la progesterona hallada en sangre periférica (Kanchev et al., 1976). Ella
está sujeta a un mecanismo de control dependiente de estímulos luteotrópicos y luteolíticos. Está bien
documentado el efecto luteotrópico de la LH en la vaca (Donaldson y Hansel, 1965; Hixon y Hansel, 1979;
Rotchild, 1981). Sin embargo, se ha observado que los pulsos de progesterona son más frecuentes que
los de LH (Hizon et al., 1983; Walters et al., 1984) y durante la segunda mitad de la gestación donde des-
aparecen los pulsos de LH se encontró una relación entre los pulsos de FSH y progesterona (Schallen-
berger et al., 1983); lo cual fue confirmado en la fase lútea de la vas (Walters et al., 1984). Además el
cuerpo lúteo bovino posee receptores a FSH (Manns et al., 1984) y la adición de esta hormona a tejido
lúteo "in vitro" aumenta la síntesis de P4 (Romanoff, 1966). Esto indica que tanto la FSH como la LH se-
rían hormonas luteotrópicas.
Análogamente, se han señalado como agentes luteotrópicos a la prostaciclina (Milvae y Hansel, 1980),
factores angiogénicos (Koss y LeMarie, 1983) y la serotonina (Battista y Condon, 1986).
Las funciones principales de la P4 son la preparación del útero para la nutrición e implantación del huevo,
desarrollo del blastoquiste y mantenimiento del feto hasta el termino (Stabenfeldt, 1974). En la vaca a
partir de los 215 días, la gestación queda sostenida por la P4 procedente de la adrenal después de la ova-
riectomía (Wendorf et al., 1983). La P4 inhibe las contracciones uterinas disminuyendo el potencial de
membrana (Ganong, 1971) y el nivel de receptores estrogénicos (Leavitt et al., 1985).
Los gestágenos controlan la duración del ciclo estral. Walters et al., (1982) propusieron que la P4 actua-
ban como un "brake" bloqueando los eventos endocrinos necesarios para la ovulación.
Suministrada en dosis apropiadas en el ciclo inhibe el piso preovulatorio de LH, correlacionándose negati-
vamente con esta hormona (Peters y Lamming, 1986). En hipotálamo e hipófisis existen receptores de
progesterona (Rajadurai, 1977), la acción inhibitoria puede realizarse disminuyendo la frecuencia de pul-
sos de GnRH y/o interfiriendo el efecto per se del GnRH y del estradiol sobre la hipófisis (Azzazi et al.,
1983).

En la terapia con GnRH, el pretratamiento con P4 promueve una duración del ciclo estral normal (Grobe-
laar et al., 1984; Rutter et al., 1985; Legan et al., 1985). Comprobándose que la progesterona influye di-
rectamente sobre el folículo posibilitando el desarrollo adecuado del cuerpo lúteo (Hunter et al., 1986).
La progesterona controla la receptividad sexual. Dosis altas bloquean el celo, dosis pequeñas lo favore-
cen (Stabenfeldt, 1974; Stabenfeldt et al., 1976). Sin embargo, existen trabajos (Bulman y Lamming,
1979; Hruska et al., 1983; Fernández et al., 1988; Alonso et al., 1986b) donde se señalan valores altos de
P4 en el momento del estro concomitante con buena fertilidad posterior a la IA, lo cual no ha tenido expli-
cación. No es muy probable que tales concentraciones se correspondan a la fracción de P4 libre, (biológi-
camente activa), pues ella no sólo bloquea el celo y la ovulación, sino también administrada el día 1 ó 2
del ciclo produce una luteolisis temprana, ocasionada por una liberación prematura de oxcitocina y PGH
(Peterson, 1985); o en el mismo momento del celo conduce a una persistencia folicular (Lee et al., 1986).
En estas condiciones es difícil obtener una buena fertilidad.
Por esas razones, el aumento de P4 durante el celo puede ser debido a un incremento de la fracción de la
hormona unida a las proteínas transportadoras (albúmina, CBG, SHBG), lo cual proporcionaría una re-
ducción de su MCR y de su actividad fisiológica.
Se sabe que la P4 es un fuerte antagonista de los corticoides (Baxter, 1976) y que se une fuertemente a la
CBG (Siiteri y Febres, 1983), por lo tanto durante el celo de algunos animales donde existe el pico crítico
de E2 (pudiera estimularse significativamente la producción de CBG, resultando en una elevación de la
tasa de P4 unida a la proteína. Esto se apoya en experimentos donde la inmunización contra esteroides
produce un cuadro similar. (Hillier et al., 1974; French y Spenneta, 1981). Otras razones pueden ser la
existencia de hiperalbuninemia o hepatopatías.
Con respecto a la aparición del celo y su inducción, numerosos trabajos han confirmado los picos de pro-
gesterona que se asocian a la pubertad y el primer estro postparto (González-Padilla et al., 1975a; Shot-
tan et al., 1978; Suzuki y Sato, 1980; 1982; Schillo et al., 1983a; Olujohungbe y Glencross, 1985). Lo cual
sugiere que la P4 tiene una función clave en la etapa de transición del estado acíclico al estado cíclico.
Los cambios en el cociente de las concentraciones de entre vana yugular y arteria carótida durante seis
períodos estrales en novillas, sugieren que en el entorno del celo se produce una gran modificación en la
utilización y/o unión de la hormona a nivel de la cabeza (La Voi et al., 1971).}
González-Padilla et al., (1975b), estimularon la pubertad en novillas con 3 dosis de 20 mg de progestero-
na y 17B-E2 (2 mg), lo cual no se logró con el estrógeno sólo. Combinando la P4 con E2 se ha podido
inducir la pubertad en animales prepúberes, dependiendo los resultados de la edad y peso con los cuales
se inician los tratamientos (Short et al., 1976; Burfenigm 1979; Spintzer, Portal et al., (1976) aplicaron un
tratamiento basado sólo en progesterona (30 mg/d x 15) y obtuvieron el 100% de gestación en los celos
inducidos. Igualmente, Sheffield y Ellicott (1982) en ovino lograron con dosis bajas P4 el 37% y el 86% de
celo y actividad ovárica respectivamente. En tanto, Gíl (1984) no recomendó el uso de P4 aislada para la
promoción del celo en hembras bovinas, nosotros sí demostramos la utilidad de aplicar 50 mg en el caso
de las novillas (Faure et al., 1982, 1996; Faure,1987; tabla 4). La aplicación de implantes de norgestomet
a novillas impúberes produjo respuestas variables en la secreción posterior de P4 y E2 (Tanaka, et al,
1995) mientras que se reportó que el progestágeno solo indujo celo en una alta proporción en novillas
prepúberes y estimuló la frecuencia de los picos de LH (Anderson, et al, 1996). Por su parte Hall, et al
(1997) corroboran el efecto inductor del celo que tiene los progestágenos encontrando que no fue alterado
por la ganancia diaria de peso de los animales tratados, pero si era dependiente de la edad a que fueron
tratados los animales concluyendo que la P4 induce la pubertad acelerando la cascada de eventos endo-
crinos y ováricos que se asocian a la pubertad espontánea, la edad y no la ganancia de peso paree ser el
factor crítico en su efecto.

Tabla 4. Porcentajes de novillas con síntomas de estro y gestadas luego del tratamiento con progesterona
y progesterona mas eCG (Faure et al., 1996).
Celo Tasa de Gestación
1er Servicio
Tasa de Gesta-
ción
Total
Tasa de preñez
Grupo n n % n % n % n Dif.
Ensayo I. Progesterona
Trata-
das
140 77 55.0a 38 49.4 50 65.0 36
c 12
Control 240 85 35.4b 47 55.3 57 67.0 24
d 0
Ensayo II. Progesterona + eCG
Trata-
das
51 41 80.0a 27 65.7 40 97.6 78
c 28.4
Control 38 19 50.0b 14 73.7 10 100 50
d 0
a y b difieren con p<0.01; c y d difieren con p<0.05
Los gestágenos combinados con PMSG, GnRH y E2 han sido más ampliamente utilizados con el fin de
inducir la pubertad resultando más eficaz (Faure, et al, 1996).
3.2.5.3 Andrógenos
Son todos aquellos cuerpos esteroides que poseen acción masculinizante; cuyas acciones en la hembra
pueden ser antifeminizantes, seudoprogestógenas y precursoras de estrógenos (Botella, 1982).
La testosterona estimula la síntesis y/o liberación de FSH (Naqvi y Johnson, 1969; Fernández - Limia y
Kuroedov, 1976; Pant, 1977; Thompson et al., 1984) y LH (Restall et al., 1972, 1977 Pant, 1977); o puede
no alterar el nivel de LH (Radford y Wallace, 1971).
Se ha observado tras la aplicación de testosterona previo a una inyección de GnRH, que la respuesta
hipofisaria puede retrasarse (Fernández - Limia et al., 1977) o atenuarse (Thompson et al., 1984). Su
mecanismo de acción en la hembra no esta claro. Puede actuar directamente (Drouin y Labrie, 1976) o
por conversión a 5-alfa-dihidrotestosterona-DHT (según Wilson y Gloyna, 1970); o estrógenos (Baird et
al., 1969). Pant (1977) en ovejas acíclicas logró con una dosis única de PT (2,5-10mg) producir la descar-
ga de gonadotropinas entre 16 y 24 horas posterior a su administración con picos máximos de LH de 31-
65 ng/mL.
La inmunización activa contra andrógenos en ovejas produjo un incremento de la liberación de gonadotro-
pinas (Martensz et al., 1976; Martensz y Scaramuzzi, 1979) y la tasa de ovulación (Scaramuzzi et al.,
1981). En vacas inmunizadas contra androstendiona se presentó quiste folicular, ninfomanía y ovulación
(Walton, 1985). El síndrome de Stein-Leventhal y la atresia folicular se han enlazado con el metabolismo
de los andrógenos (Siiteri y Febres, 1983). Sin embargo, la regresión folicular entraña un proceso comple-
jo, en el cual el flujo sanguíneo y ambiente nutricional parecen ser transcendentes (Hay et al., 1979; Kruio
y Dieleman, 1985).
Está bien definido el efecto que causan los andrógenos en el proceso de sexualización, la diferenciación
del hipotálamo tónico y cíclico (Thibaut y Levasseur, 1974). La administración crónica de andrógenos con-
lleva al virilismo.
El hecho de encontrarse niveles sanguíneos de testosterona concomitante con estradiol sugirió que el
andrógeno sería necesario para la expresión de la conducta sexual. La administración de testosterona
exógena puede estimular la secreción de estrógenos ováricos y adelantar el pico preovulatorio de gonado-
tropinas (Kotwica y Williams, 1982). Así, la inyección de andrógenos estimula la libido de humano (Ban-
croft y Skakkebaek, 1979), la receptividad sexual en monos Rhesus (Herbert y Trimble, 1967) y el estro
en ovejas (Lindasy y Robinson, 1961, 1964). El propionato de testosterona/PT/ en dosis única (1-25mg)
ha inducido la ovulación con formación de cuerpos lúteos (Restall et al., 1977); lográndose desde un 33 a

100% de animales ovulantes. (Faure et al., 1982a, 1987) en novillas lograron inducir el estro con buena
fertilidad, utilizando el PT en dosis única de 50 mg.
3.3 Aparición de la pubertad y su relación con la eficiencia reproductiva y productiva.
Según Brinks (1990) la edad a la pubertad parece ser un buen indicador de la fertilidad y expresión de la
eficiencia reproductiva. La heredabilidad estimada es elevada (0.40) comparada con otros rasgos repro-
ductivos. Debe tenerse en cuenta que la IA o monta de las novillas en su primer celo puberal resulta en un
21% de concepción inferior comparado con el tercer celo, lo que implica que las novillas deben alcanzar la
pubertad como promedio de uno a tres meses antes de ser gestadas (Patterson, et al, 1992). Estos auto-
res han registrado relaciones fenotípicas favorables entre edad a la pubertad y tasa de gestación tempra-
na citándose cifras de -0.75 y -0.40, a la vez que reportan una relación negativa entre edad a la pubertad y
el intervalo parto - celo para el cruce de Hereford x Angus, no así para el cruzado con Brahman. Las novi-
llas más pesadas al destete alcanzan la pubertad más jóvenes pero presentan mayor intervalo parto - celo
(Patterson, et al, 1992a), los que además reportaron que el estudio de las correlaciones existentes entre
edad a la pubertad y peso corporal, en diferentes momentos de la etapa de desarrollo, han evidenciado
una mayor relación del peso corporal de ese parámetro en el momento del destete (-0.52), lo cual ratifica
el nexo existente entre la edad a la pubertad y la curva de crecimiento.
Werre y Brinks (1986) notificaron correlaciones entre edad a la pubertad y capacidad productiva probable
en cuatro lactancias siendo de -0.65 y -0.38 para la primera y segunda respectivamente, lo cual corroboró
los resultados de Laster, et al, (1979) quienes registraron relaciones de -0.88 de edad a la pubertad con la
producción de leche.
Los estudios de aspectos productivos de novillas que llegan temprano a la pubertad indican la necesidad
de evaluar el desarrollo genital y medir el área pélvica de modo que se seleccione el semental apropiado
para evitar dificultades al parto, la heredabilidad del área pélvica es del 61% lo que constituye un indicador
útil para la selección (Motoroga y Gavan 1994). El crecimiento acelerado y el parto muy temprano dismi-
nuyen el desempeño de la primera lactancia por un mecanismo aun no esclarecido (Hoffman, et al, 1996).
4 A modo de conclusión
Como se ha visto, son diversos los factores que modifican de alguna manera la aparición de la pubertad.
De la integración y manejo de ellos por parte del productor pecuario está condicionada la productividad
que pueda alcanzar.
El genofondo del animal, el ambiente, la nutrición, la tecnología y la formación técnica de los trabajadores
de la esfera son aspectos de los cuales pueden obtenerse resultados favorables si se enfocan como un
sistema y se realizan las inversiones necesarias que converjan hacia una mayor eficiencia ganadera.
Si bien es cierto, que en países desarrollados la tendencia a una incorporación a la reproducción muy
temprana de las novillas acarrean perjuicios en la producción de estos animales, en los países del área
subtropical y tropical es muy distinto: por un lado está la época de sequía la que ocasiona un desbalance
nutricional, aparejado a manejo con bajo nivel técnico, existencia de enfermedades propias del clima, y
a su vez, baja selección genética, todo lo cual comprometen marcadamente la industria pecuaria, ha-
ciéndola menos rentable, por lo que sobre estos aspectos es que hay que dirigir la atención, con vistas a
desarrollar una ganadería cada día más competitiva en un mundo cada vez más globalizado.

5 Bibliografía
1. Adams GP. y Brogliatti GM. (1996). La función ovárica en las terneras prepuberes y peripuberes. II
Simposio Int. Reprod. Anim. 31 Oct.- 2 Nov. Carlos Paz. Córdoba, Argentina pp. 13-22.
2. Aguilar A. y Colina C.S. (1982). Estudio morfológico comparativo de los ovarios de la vaca cebú y la
vaca Holstein. VIII Congr. Nac. Buiatría, Veracruz, Ver. México p. 211.
3. Alba, L.O. y Revuelta, J.F. (1981). Efecto de la progesterona y gonadotropina sérica combinada en
novillas anéstricas. Resúmenes II Sem. Científico. CENSA p. 147.
4. Alcalá L., Kreen K. y Martínez G. (1983). Terapéutica hormonal como posible solución del anestro
en novillas. Libro Res. 4ta. Reun. ACPA 12-14 dic. La Habana p. 132.
5. Allen W.R. and Moor R.M. (1972). The origin of the equine endometrial cups. I. Production of PMSG
by fetal trophoblast cells. J. Reprod. Fert. 29: 313.
6. Alonso J.C., Faure R., Gil A., Fernández O. (1986a). Niveles séricos de progesterona en novillas
ovariectomizadas después de la aplicación de progesterona exógena. Informe técnico. Dpto. Re-
prod., CENSA. La Habana.
7. Alonso J.C., Faure R., R. Castellanos. (1984). Niveles de progesterona en novillas ¾ Holstein x ¼
Cebú suplementadas con miel/urea o concentrado. Rev. Salud Anim. 6: 313.
8. Alonso J.C., Libertad García, Faure R., Fernández O. (1986b). Niveles de progesterona sérica du-
rante la inseminación artificial en animales Holstein y su relación con el índice de gestación. 5ta.
Reunión ACPA. La Habana, Cuba.
9. Alonso J.C., R. Castellanos, R. Faure., O. Fernández. (1985). Efecto de la suplementación energé-
tica en novillas con pubertad ratardada sobre la concentración de progesterona sérica y la primera
inseminación artificial. Rev. Salud Anim. 7: 371.
10. Amoss M., R. Burgus, R. Blackwell, W. Vele, R. Fellows and R. Guillermin. (1971). Purification,
amino acid composition and N.terminus of the hypotalamic hormone releasing factor (LRF) of ovine
origin. Biochem. Biophys. Res. Comm. 44: 205.
11. Amstalden, M.,; M. R. Garcia; Williams, S.W.; Stanko, R.L.; Nizielski, S.E.; Morrison, C.D.; Keisler,
D.H. and Williams, G.L. (2000). Leptin Gene Expression, Circulating Leptin, and Luteinizing Hor-
mone Pulsatility Are Acutely Responsive to Short-Term Fasting in Prepubertal Heifers: Relationships
to Circulating Insulin and Insulin-Like Growth Factor I1 Biol Reprod 63: 127.
12. Anderson L.H., McDowell C.M., Day M.L. (1996). Progestin - induced puberty and secretion of lute-
inizing hormone in heifers. Biology of Reprod. 4:1025.
13. Anderson W.J., Forrest D.W., Schulze A.L., Kraemer D.C., Bowen M.J., Harms P.G. (1985). Ovari-
an inhibition of pulsatile luteinizing hormone secretion in prepuberal Holstein Heifers. Domestic
Anim. Endocr. 2: 85.
14. Apter D. (1997). Development of the hypothalamic - pituitary - ovarian axis. Ann N. Y. Acad. Sci.
816:9.
15. Arije G.F., Wiltbank J.N. (1974). Prediction of age and weight at puberty in eef heifers. J.Anim. Sci.
38: 803
16. Asdell S.A. (1946). Patterns of Mammalian Reproduction. Comstock, Ithaca, NY, 437 pp.
17. Azzazi F., G.F. Krause and H. A. Garvericck. (1983). Alteration of the GnRH-induced LH release by
steroids in postpartum dairy catttle. J. Anim. Sci. 57: 1251.
18. Bagley C.P. (1993). Nutritional management of replacement beef heifers: a review. J. Anim. Sci.
71:3151.
19. Baird D.T. (1972). Reprodutive hormones. En: Reproduction in mammls. tomo 3. Hormones in re-
production Austin C.R. and short R.V., Cambridge Univ. Press. Cambridge Univ. Press. Cambridge.
20. Ball P.J.H. and G.E. Lamming. (1983). Diagnosis of ovarian acyclicity in lactating dairy cows and
evaluation of treament with gonadotrophin releasing hormone or a progesterone-releasing intravagi-
nal device. Br. Vet. J. 139:522.
21. Bancroft J. and N.E. Skakkebaek. (1979). Androgens and human sexual behavior. In.
Sex.Hormones and Behavior. Ciba Found. Symp. 62, (N.S.) Exc. Med. Amst. Oxf. N.Y. p. 209.
22. Barash H., Bar-Meir Y., Bruckental Y. (1994). Effects of a low energy diet followed by a compesatory
diet on growth, puberty and milk production in dairy heifers. Livestock Prod. Sci. 39:263.

23. Barnes, M. A., S.T. Bierley, R.D. Halman and D.M. Henricks. (1980). Follicle stimulating hormone
luteinizing hormone and estradiol-17B response in GnRH treated prepubertad Holstein heifers. Biol.
Reprod. 22: 459.
24. Battista, P.J., Condon, W.A. (1986). Serotonin- induced stimulation of progesterone production by
cow luteal cells in vitro. J. of Reprod. and. Fert. 76: 231.
25. Baxter J.D. (1976). Glucocorticoid hormone action. Pharmacol ther. B. 2: 605.
26. Beal W.E., R.E. Short, R.B. Staigmiller, R.A. Bellows, c.c. Kaltenbach and T.G. Dunn. (1978). Influ-
ence of dietary energy intake on bovine putuitary and luteal function. J. Anim. Sci. 46: 181.
27. Bellows R.A, Short R.E. (1972). Superovulation and multiple births in beef cattle. J. Anim. Sci. 34:
66.
28. Bergfeld E.G.M., Kojima F.N., Cupp A.S. et al. (1994). Ovarian follicular development in prepubertal
heifers is influenced by level of dietary energy intake. Biology of Reprod. 51:1051.
29. Betancourt J., García Libertad, Santiago O. (1986). Situación actual de la reproducción en el gana-
do vacuno. Plan Reprod. 5ta. Reun. ACPA. La Habana, Cuba.
30. Betteridge K.F. y Mitchell D. (1974). Embryo transfer in cattle. Experience of twenty four completed
cases. Theriogencology 1: 69.
31. Bierschwal C.J., H.A. Garverick, C.E. Martin, R.S. Younquist, T.C. Cantly and M.D. Brown. (1974).
Clinical response of dairy cows with cystic ovaries to GnRH. J.Anim. Sci. 35: 199.
32. Bonadonnna T. (1953). Fisiopatología de la reproducciòn y de la fecundación artificial de los anima-
les domésticos. Ed. Revol., La Habana, 1967, p. 376.
33. Bonavera G.G., D.T. Tortonese y G.M. Doray. (1986). Influencia de un tratamiento superovulatorio
en la inducción a la pubertad en hembras bovinas. Vet. Argentina 3: 416.
34. Botella LL. J. (1982). Endocrinología de la mujer. Tomo 1. Edic. Revol., La Habana.
35. Braun U. (1985). LH and FSH in cell cultures of the anterior pituitary lobe in relation to stage of oes-
trous cycle, in pregnant cows and in cows with ovarian cysts. Schweizer Archiv für Tierheilkunde
127: 353.
36. Braun U., Schams D., Leidl, W. (1985). Effects of gonadotropin releasing hormone on the release of
luteinizing hormone and follicle-stimulating hormone from bovine anterior pituitary cell cultures. Zen-
tralblatt für Veterinärimedizin, A. 31: 594
37. Brinks JS. (1990). Improving reproductive efficiency (beef cattle). Proceedings of the fourth J.S.
Davies Beef Research Forum, 2nd and 4th April. Adelaide, S.A. (Austria) pp. 1-12.
38. Brooks A.L., Morrow R.E., Youngeuist R.S. (1985). Body composition of beef heifers at puberty.
Theriogenology 24: 235.
39. Bulman D.C. and Lamming G.E. (1979). The use of milk progesterone analysis in the study of oes-
trus detection, herd fertility and embryonic mortality in dairy cws. Brit. Vet. J. 135: 559.
40. Burfening P.J. (1979). Induction of puberty and subsequent reproductive performance. Theriogenol-
ogy 12: 215.
41. Burgus R., R.M. Butcher, M. Amoss, N. Ling, M- Monahan, J. Rivier, R. Fellows, R.B. Blackwel, W.
Vale and R. Guillermin. (1972). Primary structure of the ovine hypothalamic luteinizing hormone re-
leasing factor (LRF). Proc. Ntl. Acad. Sci. 69: 278.
42. Buskirk D.D., Faulkmer D.B., Ireland F.A. (1995). Increased postweaning gain of beef heifers en-
hances fertility and milk production. J. Anim. Sci. 73:937.
43. Butenandt A. (1934). Wien Klin. Wochscht. 47: 936. Citado por Botella (1982).
44. Byrnes W.W. and Meyer R.K. (1951). The inhibition of gonadotrophic hormone secretion by physio-
logical doses of estrogen. Endocrinol. 48: 133.
45. Calderon robles R.C., Villa Godoy A., Lagunes Lagunes J. (1993). Dynamics of folicular develop-
ment at puberty. National Meeting of livestock res. Guadalajara Jal. 27-30 sep. Abstr. p.162. Mexico,
DF.
46. Chakraborty P.K., T.E. Adams, Gladys K. Tarnavsky and J.J. Reeves. (1974). Serum and pituitary
LH concentrations in ewes infused with LH-RH/FSF-RH. J. anim. Sci. 39: 1150.
47. Clark J.H., Peck E.J. and Glasser S.R. (1977). Mechanism of action of sex steroid hormones in the
female. En: Reproduction in domestic animals. Cole H.H. and Cupps P.T., 3a. de. Academic Press.
New York.

48. Clarke I.J. and Cummins J.T. (1985a). GnRH pulse frequency determine LH pulse amplitud by alter-
ing the amount of releasable LH in pituitary glands of ewe. J. Reprod. Fert. 73: 425.
49. Clarke I.J., Cummins J.T. (1984). Direct pituitary effects of estrogen and progesterone on gonado-
tropin secretion in the ovarrectomized ewe. Neuroendocrinol. 39: 267.
50. Clarke I.J., Cummins J.T., Findlay J.K., Burman K.J. Doughton B.W. (1984a). Effects on plasma
luteinizing hormone and follicle-stimulating hrmone of varging the frequency and amplitude of gon-
adotropin releasing hormone pulses in ovariectomized ewes. Neuroendocrinol. 39:267.
51. Clarke J.J.m Wright P.J., Chamley W.A., Burman K. (1984). Differences in the reproductive endo-
crine status of ewes in the early post-partum period and during seasonal anoestrus. Reproduc. Fert.
70: 591.
52. Clarke, I.J; Cummins, J.T. (1985b). Increased gonadotropin-releasing hormone pulse frequency
associated with estrogen-induced luteinizing hormone surges in ovariectomized ewes. Endrocrinol-
ogy 116: 2376.
53. Coffey, M. P., J. Hickey, et al. (2006). Genetic Aspects of Growth of Holstein-Friesian Dairy Cows
from Birth to Maturity. J. Dairy Sci. 89: 322.
54. Cole H.H. and Hart G.H. (1930). The potency of blood serum of mares in progresive stages of preg-
nancy in effecting the sexual maturity of the immature rat. Amer. J. Physiol. 93: 57.
55. Contestabile O:I., Sandoval V.A., Pérez D.E. y Pijoan A.P. (1985). Efecto de la administración de
Melatonina como inductor del estro en cabras durante el anestro estacional. Reunión de investi-
gación pecuaria en México. 1985.
56. Contreras F., R. Portal y Vivian Contreras. (1986). Efecto de la suplementación con yodo en la pre-
sentación de celo en novillas y en la conservación de la salud de los terneros. III Congr. Med. Vet.
Libro Res., res. 43. La Habana.
57. Cook D.K., T.A. Winters, L.A Horstman and R.D. Alleich. (1985). Induction of estrus in ovariecto-
mized cows: Effect of increasing amounts of estradiol benxoate. J. Dairy Sci. 68: 188.
58. Cruz M. de la Barreto G., Capote A. y Valle M. (1983). Producción de gonadotropina sérica (PMSG)
en Cuba y su comparación con PMSG importada. Ciencia y Técnica en la Agricultura 5: 97.
59. Dailey R.A., J.C. Price, K.R. Simmons, E.M. Meisterling, P.A. Quinn and S.P. Washburn. (1986).
Synchronization of estrus in dairy cows with prostaglandin F2 and estradiol benzoate. J. Dairy Sci.
69: 1110.
60. Dahl, G. E. and D. Petitclerc (2003). "Management of photoperiod in the dairy herd for improved
production and health." J. Anim Sci. 81(suppl_3): 11.
61. Dale H.E., Ragsdale A.C. and Cheng C.S. (1959). Effect of constant environmental temperature 50°
and 80°F on appearence of puberty in beef calves. J. Anim. Sci. 18: 1363.
62. Day M.L., K. Imakawa, M. García-Winder, D.D. Zalesky, B.D. Schanbacher, R.J. Kittok and J.E.
Kinder (1984). Endocrine mechanisms of puberty in heifers: estradiol negative feed back regulation
of luteinizing hormone secretion. Biol. Reprod. 31: 332.
63. Day MI, Imakawa K, Wolfe Pl al al. (1987). Endocrine mechanisms of puberty in heifers. Role of
hypothalamo. Pituitary estradiol receptors in the negative feedback of estradiol on luteinizing hor-
mone secretion. Biol Reprod. 37: 1054.
64. Desjardins C. and H.D. Hafs. (1968). Levels of pituitary FSH and LH in heifers from birth through
puberty. J. Anim. Sci. 27: 472.
65. Döcke F. (1981). Veterinärmedizinische Endokrinologie. Veb Gustav Fischer Verlag Jena. p. 464.
66. Donaldson L.E., Bassett J.M. and Thorburn G.D. (1970). Peripheral plasma progesterone concen-
tration of cow during puberty-oestrous cycles, pregnancy and lactation, and the effects of under nu-
trition or exogennos oxitoxin in progesterone concentration. J. Endocrinol. 48:599.
67. Donaldson L.E., W. Hansel, (1965). Prolongation of life span of the bovine corpus luteum by single
injections of bovine luteinizing hormone. J. Dairy Sci. 48: 903.
68. Dorrington J.H. (1979). Pituitary and placental hormone. En: Reproduction in mammals. tomo 7.
Mechanisms of hormone action. Austin C.R. and short R.V. Univ. Press. Cambridge.
69. Dott H.M., Mary F. Hay, D.G. Gran and R.M. Moor. (1979). Effect of exogenous gonadotrophin
(PMSG) on the antral foliche population in the sheep. J. Reprod. Fert. 56: 683.

70. Dun R.T. Jr., M.F. Smith, H.A. Garverick and C.W. Foley. (1985). Effects of 72 hr calf removal
and/or gonadotropin releasing hormone on luteinizing hormone release and ovarian activity in post-
partum beef cows. Theriogenology. 23:767.
71. El amin F.M. (1976). La edad al primer parto. Breve análisis de los trabajos publicados sobre gana-
do europeo y Ceú. Bol Res., Gen. y Reprod. CIDA 2: 1(3): 10, 1978.
72. Enomoto Y., Minowada S. (1997). Endocrinoligical mechanism of puberty in men. Nippon Rinsho
55:2835.
73. Erickson B.H. (1966). Development and senescence of the postnatal bovine ovary. J. Anim. Sci. 25:
800.
74. Evans A.C.O., Adams G.P., Rawlings NC. (1994). Follicular and hormonal development heifers from
2 to 36 WK of age. J. Reprod. Fertil 102: 463.
75. Evans A.C.O., Currie WD, Rawlings NC. (1992). Effect of naloxane on circulating gonadotropin con-
centrations in prepuberal heifers. J. Reprod. Fertil 96: 847.
76. Evidence for an oscillator other than luteinizing hormone controlling the secretion of progesterone in
cattle. Biol Reprod. 29: 1155.
77. Fahr R.D., Jähne M., Schwark H.H. (1984). Relationships between growth and sexual maturity in
female cattle. Wissenchaftliche Zeittschrift der Karl Marx Universität Laipzig, Mathmatisch-
Naturwissenscha ftliche Rihe. Vol. 33: 263.
78. Fauconneau, B., Gauthier, D. (1984). Effect of 4 hours of additional light on growth and onset of
puberty in criollo heifers in the tropics. Rev. Cub. Reprod. Anim. 10:49
79. Faure R, Alonso JC, Tur Zenaida et al. (1982) Ensayo terapéutico con progesterona y benzoato de
estradiol contra el anestro en novilla. Rev. Salud Anim. 4:211
80. Faure R, Alonso JC, Fernández O, González F, Garcia Libertad.(1982a).Inducción del estro median-
te propionato de testosterona en novillas anéstricas. Rev. Salud Anim. 4:201
81. Faure R , Fernández O, Alonso JC, González F, Garcia Libertad.(1986).Respuesta hipofisaria al
benzoato de estradiol en novillas con pubertad retardada. Rev. Salud Anim. 4:281
82. Faure R, , Fernández O, Alonso JC, Garcia Libertad.(1986a).Secreción pulsátil de LH y respuesta
hipofisaria a la inyección de GnRH en novillas con pubertad retardada. Rev. Salud Anim 8:369.
83. Faure R.( 1987). Concentraciones séricas de LH y progesterona, e inducción del estro en novillas
anéstricas. Tesis Dr en Ciencias Vet.La Habana, Cuba.
84. Faure, R., Fernández, O., Alonso, J. C, Gil, A. (1996). Concentraciones de progesterona sérica e
inducción de estro fértil con la inyección de dosis bajas de esta hormona en novillas anestricas.
Rev. Salud Anim. 18: 39.
85. Faure R, Fernández-Limia O, Morales Denis C. (2004). Concentraciones de cortisol serico en novi-
llas Holstein durante las dos estaciones del año en Cuba. www.veterinaria.org
/revistas/redvet/n121204.html.
86. Fernández O., Mendoza A., Faure R., García L. (1988). Niveles de hormona luteinizante, estradiol y
progesterona durante el estro en novillas Holstein en clima subtropical. Rev. Salud Anim. 10: 316
87. Fernández-Limia O. (1979). Varciaciones estacionales y control de las descargas preovulatorias de
hormona luteinizante en ganado bovino. Tesis C.Dr. Ciencia. CENSA. La Habana. Cuba.
88. Fernández-Limia O., R. Faure, L. García. (1986). Valores de progesterona durante el ciclo estral
postparto en vacas Holstein en período seco y de lluvias. Proceedings of Symp. “Nuclear and relat-
ed techniques in animal production and health” IAEA-FAO, Viena 17-21 march page 672.
89. Fields M.J. (1984). Relaxin and oxytocin: Recently discovered ovarian hormones in the cow. Florida
Agric. Res., 3: 44.
90. Fitzgerald J.A. (1984). The effect of castration, estradiol and LHRH on secretion of lambs fed differ-
ent levels of dietary energy. J. Anim. Sci. 59: 460.
91. Forrest D.W., A.L. Schulze, D.C. Kraemer and N.H. McArthur. (1984). Relation ship berween follicu-
lar and plasma estradiol.17 in prepuberal Holstein heifers. J. Anim. Sci. 59 (suppl.1): 313.
92. Fortune, J.E. and W. Hansel. (1979). The effect of 17 -estradiol on progesterone secretion by bo-
vine theca and granulosa cells. Endocrinol 104:6, 1834.

93. Freetly H.C., Cundiff L.V. (1997). Postweanng growth and reproduction characteristics of heifers
sires by bulls of seven breeds and raised on different levels of nutrition. J. Anim. Sci. 75:2841.
94. French L.R. and B. Spenneta. (1981). Effects of antibodies to progesterone on reproduction of
ewes. Theriogenology 16: 407.
95. Ganong W.F. (1971). Manual de fisiología médica 3ra. Ed. El Man. Mod. S.A. México. p.358.
96. Garverick H.A., Elmore R.G., Vaillancourt D.H., and Sharp A.J. (1980). Ovarian response to gonad-
otropin-releasing hormone in postpartum dairy cows. Am. J. Vet. Res. 41: 1582.
97. Gasser, C. L.; Grum, D. E.; Mussard, M. L.; Fluharty, F. L.; Kinder, J. E. and Day, M. L. (2006). In-
duction of precocious puberty in heifers I: Enhanced secretion of luteinizing hormone
10.2527/jas.2005-636. J. Anim Sci. 84(8): 2035.
98. Gasser, C. L.; Burke, C. R.; Mussard, M. L.; Behlke, E. J.; Grum, D. E.; Kinder, J. E. and Day, M. L.
(2006). Induction of precocious puberty in heifers II: Advanced ovarian follicular development
10.2527/jas.2005-637. J. Anim Sci. 84(8): 2042.
99. Gasser, C. L.; Bridges, G. A.; Mussard, M. L.; Grum, D. E.; Kinder, J. E. and Day, M. L. (2006). In-
duction of precocious puberty in heifers III: Hastened reduction of estradiol negative feedback on
secretion of luteinizing hormone 10.2527/jas.2005-638. J. Anim Sci. 84(8): 2050.
100. Gil A., García J., González J., Fernández O., Rizo J.M. (1981). Terapia comparativa entre el factor
liberador gonadotrópico, asociado con estrgenos y la gonadotropina sérica contra el anestro de la
hembra bovina. Rev. Salus Anim. 3: 187.
101. Gil F.A. (1984). Diagnóstico clínico y tratamiento hormonal del anestro en vacas y novillas. Tesis
C.Dr. Cienc. CENSA. La Habana.
102. Gil Figueredo C, Faure R. (1989). Aumento de la tasa de concepción mediante la inyección de Clo-
prostenol en estros inducidos con progesterona y estradiol en vacas y novillas Cebú. Rev. Salud
Anim. 2: 173-78
103. Glencross R.G. (1984). A note on the concentrations of plasma oestradiol-17 and progesterone
around the time of puberty in heifers. Anim. Prod. 39: 137.
104. Glencross R.G. (1985). Induction of follicular growth and oestradiol secretion in cattle by pulsatile
infusion of GnRH during periods of high plasma progesterone concentration. Final report 1984-5 Na-
tional Instituye for Research in Dairying Reading, U.K.: p. 112.
105. Goldbeck V. (1976). Verbesserung der Erstbesa mung sergelomisse bei rinder durch gonadotropin-
releasing-hormone (GnRH) Inaugural-Dissertation zur Erlangung des Grades eines Doctor Medici-
nae Veterinariae durch die tierarztliche Hochschule Hannover. Hannover.
106. Gombe S. and Hansel W. (1973). Plasma luteinizing hormone (LH) and progesterone levels in heif-
er on restricted energy intakes. J. Anim. Sci. 37: 728.
107. González Menció F., Gil A., Fernández O. y Faure R. (1980). Respuesta a la administración de
hormonas liberadoras de gonadotropinas (GnRH) en vacas lecheras con anestro postparto prolon-
gado. Rev. Salud Anim. 2: 151.
108. González Noelia, C. Geerken, R. Pedroso y L. Lavandeira, (1986). Efecto de la suplementación
mineral en la presentación de celo de novillas. Resum. 5ta. Reun. ACPA. La Habana.
109. González Stagnaro C. (1993). Cap. VIII Fisiología reproductiva en vacas mestizas de doble propósi-
to En: Ganadería mestiza de doble propósito. Editor C.González- Stagnaro. 1ra. edic. Astro Deta
S.A. Maracaibo. Venezuela.
110. González, Noelia y Pedroso, R. (1979). Niveles de algunos componentes sanguíneos en novillas y
su relación con la presentación estral. Rev. Cub. Reprod. Anim. 5: 21.
111. González, Noelia; Guerkeen, C.; Pedroso, R. y Lavandeira, L.E. (1981a). Niveles de macroelemen-
tos en el suero sanguíneo de novillas mestizas (HxC) con diferentes estados reproductivos. II Sem.
Científico CENSA. Resúmenes p. 149.
112. González, Noelia; Guerkeen, C.; Pedroso, R. y Lavandeira, L.E. (1981b). Desbalances de minerales
en rebaños de novillas F2 (BsxC) estabuladas durante la estación seca. Resúmenes II Sem.
Científico CENSA p. 151.

113. González, Noelia; Guerkeen, C.; Pedroso, R. y Lavandeira, L.E. (1981c). Niveles de microelemen-
tos en el suero sanguíneo de novillas mestizas (HxC) con diferentes estados reproductivos.
Resúmenes II Sem. Científico CENSA p. 152.
114. González-Menció F., J. Manns and B.D. Murphy. (1978). FSH and LH activity of PMSG from mares
at different stages of gestation. Anim. Reprod. Sci.1: 137.
115. González-Padilla E., G.F. Niswender and J.N. Wiltbank (1875b). Puberty in beef heifers. II Effect of
injections of progesterone and estradiol- 1717 on serum LH, FSH and ovarian activity. J. Anim. Sci.
40: 1105.
116. González-Padilla E., J.N. Wiltbank and G.N. Niswender (1975a). Puberty in Beef Heifers. Y. the
interrelationship b etween pituitary, hypothalamic and ovarian hormones. J. Anim. Sci. 40: 1091.
117. González-Padilla E., R. Ruíz, D. LeFever, A. Fenham and J.N. Wiltbank. (1975c). Pu
. berty in beef heifer. III Induction of fertile estrus J. Anim. Sci. 40: 1110.
118. Gonzalez-Stagnaro C. (1995). Manejo reproductivo en las novillas mestizas de reemplazo. En Mane-
jo de la ganadería mestiza de doble propósito. Ninoska Madrid-Bury y E. Soto Belloso. Univ del Zulia.
Ed Astro Data S.A. Maracaibo, Venezuela.
119. Gosling J.O., Mairead Ryan, Patricia Morgan. (1979). Comls hormone-induced loss of gonadotro-
phin receptors reduce the efficiency of superovulations stimulated by PMSG. Ann. Biol. Anim. Bioch.
Biophys. 19: 1499.
120. Greer R.C. (1984). Effect of daylenght at birth and lunar phase on the occurrenceof first oestrus in
beef heifers. Anim. Prod. 39: 59.
121. Gwazdauskas F.C. (1985). Effects of climate on reproduction in cattle. J. Dairy Sci. 68: 1568.
122. Hafez E.S.E. and M.R. Jainudeen. (1974). Reproductive failure in females. En: Reproduction in farm
animals. E.S.E Hafez (ed).3th. Ed. Lea and Febiger. Philadelphia.
123. Hall J.B., Staigmiller R.B., Bellows R.A., et al. (1995). Body composition and metabolic profiles as-
sociated with puberty in beef heifers. J. Anim. Sci. 73:3409.
124. Hall J.B., Staigmiller R.B., Short R.E. et al (1997). Effect of age and pattern of gain on induction of
puberty with a progestin in beef heifers. J. Anim. Sci. 75:1606.
125. Hall JB, Schillo KK, Fitzgerals BP, et al. (1994). Effects of recombinant bovine somatotropin and
dietary energy intaje on growth secretion of luteinizing hormone, follicular development, and onset of
puberty in beef heifers. J. Anim. Sci. 72: 709.
126. Hammond J. (1927). The physiology of reproduction in the cow, Cambridge, Univ. Press.
127. Hanrahan J.P., Quirke J.F., Roche J.F., (1985). Melatonin and the onser of the breeding season in
ewes. ABA 1985, 53: 5827 abstr.
128. Hansel, W., R.J. Schechter, P.V. Malven, K.R. Simmons D.L. Black, A. J. Hackett and R.R. Saat-
man. (1975). Plasma hormone levels in G.methyl-17-acetoxy progesterone and estradiol benxoate
treated heifers. J. Anim. Sci. 40, 671.
129. Hansen P.J. (1985). Seasonal modulation of puberty and the post-partum anestrus in cattle: a re-
view. Livestock production Sci. 12: 309.
130. Hansen P.J. (1985). Seasonal Modulation of puberty and the postpartum anestrus in cattle: A Re-
view. Livestock Prod. Sci. 12:309.
131. Hansen P.J., Kamwanja L. A., Hauser E.R. (1983). Photoperiod influences age at puberty of heifers
J. Anim. Sci. 57: 985.
132. Hawk H.W., W.J. Tyler and L.E. Casida. (1954). Some factors affecting age at puberty in Holstein.
Friesian heifers. J. Dairy Sci. 37: 252.
133. Hay F. Maru, R.M. Moor, D.G. Gran and H.M. Dott. (1979). Regeneration of atretic sheep ovarian
follicles in vitro. J. Reprod. Fert. 55, 195.
134. Henricks D.M. and Mayer D.T. (1977). Gonadal hormones and uterine factors. En: Reproduction in
domestic animals. Cole H.H. and Cupps. P.T., 3 th. ed. Academic Press. New York.
135. Herbert J. and M.R. Trimble. (1967). Effect of oestradiol and testosterone on the sexual receptivii
and attractiveness of the female rhesus monkey. Nature 216: 165.
136. Hillier G.S., Groom V.G., Boyns R.A., Cameron D.H.E. (1974). Development of polycistic ovaries in
rats actively immunised against T-3-BSA. Nature 250: 433.

137. Hixon J.E., W. Hansel. (1979). Effects ofprostaglandin F2 estradiol and luteinizing hormone in
dispersed cell preparation of bovine corpora lutea. Adv. Exp. Med. Biol. 112: 613.
138. Hoffman P.C., Brehm NM; Price SG. (1996). Effect of accelerated post pubertal growth and early
calving on lactation perfomance of primiparous Holstein heifers. J. of. Dairy Sci. 70: 2024.
139. Hohson H.D. and Ragsdale A.C. (1959). Effects of constant environmental temperatures of 50º for
growth responses of Holstein, Brown Swiss and Jersey calves, Miss. Agr. Exo. Sta. Res. Bull. 705.
140. Hóly L. and Hrivnak J. (1965). Influences of low closes of oestrogen on libido, vaginal cell picture
and crystallization pattern in cervical mucus of spayed cows. Vet. Med. , Praha 10: 119.
141. Holý L. y G. Martínez. (1970). Biología de la reproducción bovina. 1ra. edic. p. 45. Ed. Rev., Inst.
Cub. Libro, La Habana.
142. Hruska K., Fránek M., Bursa J., Kaláb P. (1983). Radioinmmunoassay of progesterone in cow. En-
docrinologia Experimentalis 17: 213.
143. Humphrey K.W. (1968). The effects of oestradiol-3, 17 on tubal transport in the laboratory mouse.
J. Endocr. 42: 17.
144. IAEA. (1984). Laboratory training manual on radioimmunoassay in animal reproduction. Techn. Rep.
Ser. 233 p.73.
145. Imakawa K., Day M.L., García-Winder M., Zalesky D.D., Kittok R.J., Kinder J.E. (1983).Endocrine
changes during restoration of estrous cycles following nutritionally induced anestrus in heifers. In.
Proc. Fifth World Conf. Anim. Prod. Japan. Soc. Zootech. Sci. August. 14-19. p.225.
146. Imakawa K., R.J. Kittok and J. E. Kinder (1984). Luteinizing hormone secretion after withdrawal of
exogenous progestogen in heifers fed three levels of dictary energy. J. Anim. Sci. 58: 151.
147. Imakawa, K. (1985). Influence of dietary energy on endocrine mechanisms controlling reproductibe
functions in the bovine female. Diss. Abstr. Int., B (Sci. and Enginer.) 45: 3731.
148. Jiménez J., A.O. Torres y Alba J.O. (1986). Valoración de un esquema de tratamiento hormonal
para la inducción del celo en vacas aciclícas, usando progesterona y gonadotropina seríca. Resum.
5ta. Reun. ACPA. La Habana.
149. Johsson, I.D. and J.M. Obst. (1978).Pheripheral plasma LH levels inprepuberal heifers. Proc. 10th
Ann. conf. Austr. Soc. Reprod. Biol., Sidney, August 30-31, Sept. 1: 89.
150. Jung. J.K., Kiracofe G.H. (1985). Effect of body weight on conception in beef heifers. In Proc. 3ra.
AAAP Anim. Sci. Congr., May 6-10, 1985, Vol 1, Seoul, Korea; The Organ Comm. p. 469.
151. Kahwanago, I., W.L.R. Heinrichs, W.L. Herrmann. (1969). Isolation of oestradiol “Receptors” form
bovine hypothalamus and anterior pituitary gland. Nature, 223: 313.
152. Kallas H. (1929). Puberte precore par parabiose CR Soc. Biol. 100: 979
153. Kaltenbach c.c. and Dunn T.G. (1980). Endocrinology of reproduction. En: Reproduction in farm
animals. 4ta. ed. Hafez E.S.E. Lea an Febiger, Philadelphia.
154. Kamwanja, L. A. (1984). Regulation of onset of puberty in the bovine female: the rol of season. Diss.
Abst. Int. B. (Sci. and Engin) 45: 1332.
155. Kannaway D.J., Gilmore T.A., Dunstan E.A. (1985). Pinealectomy delays puberty in ewe lambs. J.
Reprod. Fertil.74: 119.
156. Kinder J.E., Imakawa K., Day M.L., Kittot R.J. (1982). Reproductive hormone secretion in beed heif-
ers. I. In beef cattle report. 1982. Univ. Nebraska 19-20.
157. Kinder J.E., Imakawa R., Day M.L., Kittot R.J. (1982a). Reproductive hormone secretion in beed
heifers II. In Beef cattle report, 1982. Univ. Nebraska 21-22.
158. Kinder JE, Day ML, Kittok RJ. (1987). Endocrine regulation of puberty in cows and . ewes
. J. Reprod. Fertil. 34 (Suppl.): 167.
159. Kiser T.F., J.H. Britt and H.D. Richie. (1977). Testosterone treatment of cows for use in detection of
estrus. J. Anim. Sci. 44: 1030.
160. La Voi V.A., M.W. Hull and E.L. Moody, (1971). Progesterone ratios in heifers at estrus. J. Anim.
Sci. 33(1) a 244 abstr.
161. Lalman D.L., Petersen M.K., Asotegui R.P. et al. (1993). The effects of ruminally undergradable
protein, propionic acid, and monensin on puberty and pregnancy in beef heifers. J. Anim. Sci.
71:2843.

162. Lam R.F. (1986), Contribución al estudio de los principales indicadores sanguíneos del metabolis-
mo proteico y enérgetico de hembras bovinas en desarrollo. Tesis C. Dr. Ciencias. ISCAH-MES. La
Habana.
163. Larson C.L., Miller H.L., Marshall D.M., et al. (1993). effects of administering progesterone and
GnRH on age at puberty in crossbred beef heifers. South Dakota beef report No.93-10:40.
164. Larson R.L., Corah L.R., Spire M.F., et al. (1995). Effect of treatment with ivermectin on reproduc-
tive performance of yearling beef heifers. Theriogenology 44:189.
165. Leavitt W.W., Okulics W.C., McCracken J.A., Schramm W., Robidoux W.F. Jr. (1985). Rapid re-
covery of nuclear estrogen receptor and oxitocin in the ovine uterus followiun progesterone with-
drawal. J. of steroid Biochemistry 22: 687.
166. Lee C.N., D.L. Cook, J.R. Parfet, C.A. Smith, and H.A. Garverick, (1986). Administration of proges-
terone at anset of estrus caused the preovullatory follicle to persist. J. Dairy Sci. 69 (Suppl.1) p14
(Abstr).
167. Lee K.S., Na K.J., Kim H.S. (1984). _Studies on growth and puberty in korean native and Korean
native x charolais heiferss. Research reports of the Office of Rural Development, Suweonn (Live-
stock and Veterinary) 26:1
168. Legan S.J., I´Anson H. Fitzgerald B.P., Fitzovich D. (1985). Does the seasonal increase in estradiol
negative feedback prevent luteinizing hormone surges in anestrous ewes by suppresing hypotha-
lamic gonodotropin releasing hormone pulse frequency. Biology Reprod. 33:117.
169. Lemon Meredith. (1975). The effect of oestrogens alone or in association with progestagens on the
formation and regression of the corpus luteum of the cyclic cow. Ann. Biol Anim. Bioch. Biophys. 15:
243.
170. Leung K., V. Padmanablhan, L.J. Spicer, H.A. Tucker, R.E., Short, E.M. Convey. (1986) Relation-
ship between pituitary GnRH bibnding sites and pituitary release of gonadotrophins in post-partum
beef cows. J. Reprod. Fert. 76:53.
171. Lindsay D.R., Pelletier J., Pisselet C., Courot M. (1984). Changes in photoperiodd and nutrition and
their effect on testicular growth of rams. J. Reprod. Fert. 71:351.
172. Louw B.P., Lishman A.W., Nieker A.V. and Lesch S.F. (1983). The influence of season on tonic
luteinizing hormone (LH) and porgesterone levels in cattle in a subtropical en viroment. South Afric.
J. Anim. Sci. 13:253.
173. Ma Y.J., Ojeda S.R. (1997). Neuroendocrine control,of female puberty: glial and neuronal interac-
tions. J. Investig. Dermatol. Symp. Proc. 2:19.
174. Maciel, M. N., D. A. Zieba, et al. (2004). Chronic administration of recombinant ovine leptin in grow-
ing beef heifers: Effects on secretion of LH, metabolic hormones, and timing of puberty. J. Anim Sci.
82(10): 2930.
175. Maciel, M. N., D. A. Zieba, et al. (2004). Leptin Prevents Fasting-Mediated Reductions in Pulsatile
Secretion of Luteinizing Hormone and Enhances Its Gonadotropin-Releasing Hormone-Mediated
Release in Heifers. Biol Reprod 70(1): 229.
176. Madet, A., B. Baqrcikowski, R. Stupnicki, Ewea Kula and Z. Binienda. (1980). Luteinizing hormone
response to injection of synthetic gonodatrophin releasinf hormone of infusion of oestradiool - 17
Beta ibn heifers. Anim. Repdoc. Sci. 3:17.
177. Manns J.G., G.D. Niswender and T. Braden (1984). FSH receptors in the bovine corpus luteum,
Theriogenology 22:321.
178. Márquez B.G.A., Pineda F.M.P., Cardoso A.V.M., De Lucas T.J. y Pijoan A.P. (1985). Inducción de
la actividad sexual en ovejas corriedale mediante administración de melatonina y la variación del fo-
toperíodo. Reunión de Investigación Pecuaria en México. (1985). P 210.
179. Martensz N.D., D.T. Baird, R.J. Scaramuzzi and P.F.A. Van Look. (1976). Androstenedione and the
control of luteinizing hormone in the ewe during aanoestrus. J. Endocr. 69:227.
180. Martín LC, Brinks JS, Bourdon RM et al. (1992). Genetic Effects. on Beef Heifer. Puberty and Sub-
sequent Reproduction. J. Anim Sci. 70: 4006.
181. Matsuo H., A. Arimura, R.M.G. Nair and A. V. Schally. (1971). Synthesis of thew porcine Lh and
FSH releasing hormone by the solid phase method. Bichem Biophys, _Res. Comm, 45:822.

182. Mawhinney S. and Roche J.F. (1980). Induction of ovarian activity in beef suckler cows using pro-
gesterone and oestradiol benzoate or gonadotrophin releasing hormone. Anim. Breed. Abstr.
48:3704 abstr.
183. Mayar M.Q., Gladys K. Ternavsky and J.J. Reeves. (1978). Pitvitary responsive ness following infu-
sion or repeated injections of (D-Leu6, des Gly - Nh210
) LHRH - ethylamide in prepubertal ewes. J.
Anim. Sci. 47:235.
184. Mc Leod B.J., Peters A.R., Hartesing W. , Lamming G.E. (1985). Plasma LH and FSH responses
and ovarian activity in prepubertal heifers treated with repetead injections of low doses of _GnRH for
72 h. J. Reprod. Fert. 74:589
185. Mc Leod B.J., W. Haresign, A.R. Peters and G-.E: Lamming. (1984). Plasma LH and FSH concen-
trations in prepubertal beef heifers before and in response to repeated injections of low doses
GnRH. J. Reprod. Fert. 70:137.
186. Mc Donald L.E. (1975). Veterinary endocrinology and reproduction. 2nd
ed., Lea and Febiger, Phila-
delphia.
187. Mendel V.E., Morrison S.R., Bond T.E. and Lofgreen G.P. (1971). Duration of heat exposure and
performance of beef cattle. J. Anim. Sci. 33:850.
188. Mezzadra C., Homse A., Sampedro D., et al. (1993). Pubertal traits and seasonal variation of the
sexual activity in Brahman, Hereford and crossbred heifers. Theriogenology 40:987.
189. Milvae R.A., W. Hansel (1980). The effects of prostaciclyn (PGI2) and 6-keto-PGF1alfa on bovine
plasma progesterone and LH concentrations. Protaglandins 20:641.
190. Morales, J., Perón, N. Menéndez, A. Iglesias C. (1977). Desarrollo reproductivo en novillas 3/4 cebú
- 1/4 Holstein. Características reproductivas en ganado en reproducción. Rev. Cubana Reprod.
Animal 3:37.
191. Murphy BD y Pescador N. (1996). Biología celular de la foliculogénesis bovina. IIo Simposio Inter-
nacional de Reproducción Animal. Carlos Paz - Cordoba, Argentina.
192. Mori, T. (1975). Steroid hormone formation in biovine ovarian follicles. Endocrinol. Japon. 22:361.
193. Morrow, D.A., L.V. Swanson, H.D. Hafs. (1976). Estrous behavieur and ovarian activity in peripuber-
al heifers,. Theriogenology 6:427
194. Morrow, D.A: (1969). Estrous behavior and ovarian activity in prepubertal and postpubertal dairy
heifers. J. Dairuy Sci,. 52:224.
195. Moseley W.M., Dun T.G., Kaltenbanch C.C:, Short r.E., -Staigmiller R.B. (1984). Nagative fee dback
control of luteinizing hormone secretion in prepubertal beef heifers at 60 and 200 days of age. J.
Anim. Sci, 58:145.
196. Motoroga V, Gavan C. (1994). The puberty in heifers an important element in determination of effi-
ciency in reproduction. Lucrari stiintifice (Romania): 339
197. Moudgal N.R., Vidyashankar N. (1984). _Studies on follicle - stimulating hormone receptor induction
in the intact inmature rat. Hormone receptors in growrth and reproduction New York, USA, Raven
Press 227-236 Vol.9.
198. Muñoz María Cristina y M, Cisse. (1983). Morfopatología del aborto bovino. I.Babesiosis cerebral en
fetos. Rev. Salud Anim. 5:325.
199. Muss, G.E., J.R. Parfet, C.A. Marvin, R.D. Allrich and M.A. Diekman. (1985). Pitvitary concentrations
of gonadotrophins and receptors for GnRH in suckled beef cows at various intervals after calving
J.Anim. Sci. 60:1, 285.
200. Nakada, K.; M. M., T. Nakao, G. Watanabe and K. Taya (2000). Changes in concentrations of
plasma immunoreactive follicle-stimulating hormone, luteinizing hormone, estradiol-17ß, testos-
terone, progesterone, and inhibin in heifers from birth to puberty. Domestic Animal Endocrinology
18: 57.
201. Nancarrow C.D., Radford H.M. (1976). Response in ovariectomized cows to repeated injections of
thyrotrophin releasin hormone (TRH) and to estradiol benzoate (OBD). Theriogenology 6: 642. Ab-
str.
202. Niezen J.H., Grieve D.G., McBride B.W., ewt al. (1996). Effect of plane of nutrition before and after
200 Kg of body weight on mammary development of prepubertal Holstein heifers. J. Dairy Sci.
79:1255.

203. Niswender G.D., Reichert L.E., Midgley A.R. y Nalbandov A.V. (1969). Radioinmunoassay for bovine
and ovine luteinizing hormone. Endocrinology 84:1166.
204. Niswender G.D., T.M. Nett and A.M. Akbar. (1974). The hormones of reproduction. ER: Reproduc-
tion in farm Animal. E.S.E, Hafez. 3th. Ed. Lea and Febiger. Philadelphia.
205. Nuñez, D., Mirva, V. y Montes, I. (1977), Desarrollo reproductivo en novillas 3/4 Cebú - 1/4 Holstein.
Características anatomicas del aparato reproductor. Rev. Cub. Reprod. Anim,. 3:47.
206. Ojeda, S.R., Ma Y.J., Rage F. (1997). The transforming growth factor alpha gene family is involved
in the neuroendocrine control of mammalian puberty. Mol Psychiatry 2:355.
207. Okano A., Terada T. Obata T. (1980a). Histological studies on uterine growth from birth to puberty in
Japanese Black heifers. Japan. J. Anim. Reprod. 26:183.
208. Okano A., Terada T. Obata T. (1980b). Grouth pattern of the genitalia from birth to puberty in Japa-
nese Black heifers. Japan. J. Anim. Reprod. 26:178.
209. Olujohungbe A., Glencross R.G. (1985). Direct addition radioimmunoassay of progesterone in plas-
ma of prepubertal heifers. Final report 1984-5 National Institute for research in Dairying. Reading
UK (1985): 113.
210. Onuma H., J. Hahn and R.H. Foote, (1970). Factors affecting superovulation, fertilization and recov-
ery of superovulated ova in prepubertal cattle. J. Reprod. Fert. 21: 119.
211. Owens F.N., Dubeski P., Hanson C.F (1993). Factors that alter the growth and development of ru-
minants. J. Anim. Sci. 71:3130.
212. Oxender W. D., H.D. Hafs and E.M. Convey. (1971b). Fetal pituitary GH, LH and prolactin. J.Anim.
Sci. 33(1): 260 abstr.
213. Paterson, A.M. and D.R. Lindsay. (1981). Induction of puberty in gitts. 2. The effect of boars on
maintenance of cyclic activity in gilts induced to ovulate with pregnant mare´s serum gonado trophin
and human chorionic gonadotrophin. Anim. Prod. 32:51.
214. Patterson DJ, Corah LR, Brethour JR. et al. (1992a). Evaluation of Reproductive Traits in Bos teurus
and Bos indicus Crossbred Heifers: Relationship of Age at Puberty to Length of the Postpartum In-
terval to Estrus. J. Anim. Sci. 70: 1994.
215. Patterson DJ, Perry RC, Kiracofe GH, et al. (1992). Manegement Considerations in Heifer Devel-
opment and Puberty. J. Anim Sci. 70: 4018.
216. Pavel S. (1971). Evidence for the ependymal origin of arginine vasotocin in the bovine pineal gland.
Endocrinology 89: 613.
217. Pelletier J. and. J. Thimonier (1975). Interactions between ovarian steroids or progestagens and LH
release. Ann. Biol. Anim. Bioch. Biophys. 15: 131.
218. Pelletier J., Carrez. Camous S., Thiery J.C. (1981). Basic. neuroendocrine events before puberty in
cattle, sheep and pigs. J. Reprod. Fert. Supple. 30: 91-102.
219. Pelletier J., Thimonier J. (1969). Etude de la decharge ovulante par dosage radioimmunologique de
la LH plasmatique chez la brebis normale ou traitee par un progestagene. C.R. Acad. Sc. Paris,
268: 573.
220. Pérez Arelys y J. Ramirez. (1986). Respuesta de novillas estabuladas a la suplementación con ha-
rina de soya o pienso en el período de seca. Resum 5ta. Reun. ACPA. La Habana.
221. Perón N., Tarrero R. (1982). Efecto de un regimen de subalimentación durante el crecimiento para
novillas lecheras en la edad y peso a la pubertad. Rev. Cub. Reprod. Anim. 8:33.
222. Perón, N., Martínez G., Iglesias, C. y Morales J.R. (1976). Efecto de la suplementación de minera-
les, vitaminas y proteína en lapresentación del estro en novillas alimentadas a base de caña molida
y miel/urea Rev. Cub. Reprod. Anim. 2: 82.
223. Peter A.R. (1984). Effect of exogenous oestradiol-17 on gonadotrophin Secretion in post-partum
beef cows. J. Reprod. Fert. 72:473.
224. Peter A.R., Lamning, G.E. (1986). The effect of epostane (Win 32739) an inhibitor of 3 hidroxy
steroind de hydrogenase on luteal fanction and gonadotrophin secretion in cows. Anim. Reprod. Sci.
10:91.

225. Peterson, A.J. (1985). Administration of progesterone early in the sheep cycle and the release of
oxitocin and prostaglandin F. In. New Zealand Ministry of Agricul and Fisheries, Agricultural Res.
Div. Ann. Report, 1983/84, Wellington, New Zealand p.60.
226. Piva F., M. Mota y L. Martini. (1983). Regulación de la función hipotalámica e hipofisaria; Mecanis-
mo de feedback largos, cortos y ultracortos. En: Endocrinología. Tomo 1. L.J. De. Groot. Edic. Re-
vol. La Habana.
227. Plasse, D. (1966). El Brahman americano. Revista del Cebú (mayo.agosto). Venezuela.
228. Portal M., Contrera F., Cabrera E., Portal A., Martínez G., Hernández J. y Portal R. (1978b). Induc-
ción artificial del celo en hembras bovinas mediante suero artipineal. Resum II Jorn. Cienc. Vet., Vil-
la Clara. Cuba. p. 25.
229. Portal M., Portal R., Reyes H. y Cabrera E. (1976). Inducciòn artificial del celo y gestación en vacas
F2 rown Swiss-Cebú. II Congreso Nac. Cienc. Vet., La Habana.
230. Portal M., Sierra G., Dulce Portal, F. Contrera, J. Pérez y R. Portal. (1978a). Estudio electroforético
en suero de hembras bovinas tratadas con extractos pineales. Resum. II Jorn. Cienc. Vet., Villa Cla-
ra. Cuba p. 37.
231. Prendiville D.J; EnrightWJ; Crowe MA: et al. (1995). Immunization of prepubertal beef heifers
against gonadotropin-releasing hormone: immune, estrus, ovarian and growth responses. J. Anim.
Sci. 73: 3030.
232. Procknor M., S. Dachir, R.E. Owens, D.E. Little and P.G. Itarms. (1986). Temporal relationship of
the pulsatil fluctuation of luteinizing hormone and progesterone in cattle: A time series cross-
correlation analysis J. Anim. Sci. 62: 191.
233. Purvis H.T., Whittier J.C. (1996). Effects of ionophore feeding and anthelmintic administration on
age and weight at puberty in spring - born beef heifers. J. Anim.. Sci. 74:736.
234. Radchenkov V.P., Butrov E.V., Golenkevich E.K., Sukhikh V.F., Panasenki V.N., Saounov E.G.,
Matveer V.A., Averin V.S., Ismailov S.A. (1984). Hormonal profile, growth and development of the
sexual fuction in heifers. Sal´skok hozyaistvennaya Biologiya 12: 93 (ABA, 1985: 6985 abst.)
235. Radcliff R.P., Vandeltaar M.J., Skidmore A.L. et al (1997). Effects of diet and bovine somatotropin
on heifer growth and mammary development. J. Dairy Sci. 80:1996.
236. Rajadurai. R. (1977). Role of ovarian steroids on estruc control in bovine. Diss. Abstr. Intern. B, 38:
3.
237. Rajamahendran R., P.C. Laque and R.D. Baker. (1981). Serum hormone levels and occurrence of
oestrus following use of an intravaginal devive containing. progesterone and oestradiol 17 in heif-
ers. Anim. Reprod. Sci. 3: 271.
238. Rasmussen H. (1974). Organization and control of endocrine systems. En: Textbook of endocrinol-
ogy. R.H. Williams. Saunders Co. Philadelphia.
239. Redding T.W., A.V. Schally, A. Arimura and H. Marsvo. (1972). Stimulation of release and synthesis
of luteinizing hormone (LH) and follicle stimulating hormone (FSH) in tissue cultures of rat pituitaries
in response to natural and synthetic LH and FSH releasing hormone. Endocrinol. 90: 764.
240. Reeves J.J., T.W. Beck and T.M. Nett. (1974). Serum FSH in anestrous ewes treated with 17 -
estradiol J. Anim. Sci, 38: 374.
241. Restall B.J., Radford H.M., Wallace A.L.C. (1972). Response of lactating ewes to injection of orstra-
diolbenzoate or testosterone propionate. J. Reprod. Fert. 28: 164. Abstr.
242. Reynolds L.P., D.A. Robertson and S.P. Ford. (1983). Effects of intrauterine infusion of oestradiol-
17 and prostaglandin E-2 on luteal function in non-pregnant neifers. J. Reprod. Fert. 69. 103.
243. Rhind S.M. and A.S. McNeilly. (1986). Follicle populations, ovulation rates and plasma profiels of
LH, FSH and prolactin in scottish blackface ewes in high and low levels of body condition. Anim.
Repdioc. Sci. 10:105.
244. Rieger D., Rawlings N.C. (1985). The influence of estradiol 17 and progesterone on serum levels
of LH and FSH in the ewe before and after GnRH treatment. Dom Anim. Endocrinol 2:17.
245. Ringuet H., Pelletier G., Brazeau P. et al. (1994). Long - torm effects of human growth hormone
releasing hormone and photoperiod on hormone release and puberty in dairy heifers. J. Anim. Sci.
72:2709.

246. Rippel R.H., E.S. Johnson and W.F. White. (1974). Effect of consecutive injections of synthetic gon-
adotropin releasing hormone on LH release in the anestrous and ovariectomized ewe. J. Anim. Sci.
39: 907.
247. Rippel, R.H., R.H. Moyer, E.S. Johnson and R.E. Maver. (1974). Response of the ewe to synthetic
gonadotropin releasing hormone. J. Anim. Sci. 38: 3; 605.
248. Rivera J.M., Castillo H.R., J.J.P. Hernández L., J, Pérez. (1982). Crecimiento y reproducción hasta
el primer parto en ganado cebú y cebú x europeo en clima tropical. VIII Cong. Nasc. Buiatría, Vera-
cruz, México p.209.
249. Robinson T.J. (1973). Factors involved in the failure of sperm transport and survival in the female
reproductive tract. J. Reprod. Fert. Suppl. 18: 103.
250. Roche J.F. (1974). Effect of short-term progesterone treatment on aestrous response and fertility in
heifers. J. Reprod. Fert. 40: 433.
251. Roche J.F. (1975). Control of time of ovulation in heifers treated with progesterone and gonadotro-
pin releasing hormone. J. Reprod. Fert. 43 (3): 471.
252. Roche J.F. Hanrahan J.P., Quirke J.F., Ronayne E. (1984). Effect of melatonin on the time of onset
of the breding season in different breeds of sheep. ABA 1986; 54: 1664.
253. Rodes III R.C., R.D. Randel and P.G. Harms, (1978). Reproductive studies of Brahman cattle IV.
Luteinizing hormone levels in ovariectomized Brahman, Brahman x Hereford and Hereford cows fol-
lowing a 20 mg doseof estradiol - 17 . Theriogenology 10: 429.
254. Rodway R.J., Swift, A.D. (1985). A comparison of PMSG and LHRH agonist treatment in the induc-
tion of ovulation in the anoestrous ewe. Anim. Reprod. Sci. 9: 153.
255. Romanoff E.B. 1966. Steroidogenesis in the perfused bovine ovary. J. Reprod. Fert. Suppl. 1: 89.
256. Rombauts P., Courot M., Levasseur M-L, Thibault ch. (1961). Nutrition et puberte. Extrait des an-
nales de la nutrition et de L´alimentation Vol. XV. No. 2.
257. Roque, P.T. (1984). Inducción del celo con progesterona y 500 UI de PMSG en novillas anestricas
mestizas de Holstein. Tesis de Diploma. ISCAH.
258. Roy J.H., Perfitt M.W., Ganderton P., Stobo I.J.F. (1985). The effect of continuous ligt from birth on
the age of puberty of friesian heifers reared on high planes of nutrition. Final report 1984-5 <National
Institute for Research in Dairy Ing. Reading, U.K. 135.
259. Roy J.H.B., Gillies Catherine M., Perfitt M.W. and Stobo I.J.F. (1980). Effect of season of the year
and phase of the noon on puberty and on the occurrence of oestrus and conception in dairy heifers
reared on high planes of nutrition. Anim. <prod. 31:13.
260. Roy J.H.B., Perfitt M.W. , Ganderton P., Stobo J.J.F. (1985). Comparisobn of the age Puberty of
arbeedeen augus x frieseian heifers with that of friesians. Final Report 1984-5 National Institute for
Research in Dairying Reading UK 135.
261. Rutter L.M., T.D. Carrithers and J.G. Manns. (1985). The postpartum induced corpus luteum: Func-
tional differences from thar of cycling cows and the effect of progesterone pretreament Biol Reprod.
33: 560.
262. Rutter, L.M. (1984). Nutrition/reproduction interactions in beef females. Diss. Abst. Intern., B.
(Sci.and Enginee) 44: 2024.
263. Rysanek M. y Alba O. (1975). Tratamiento de la aciclia postpartum en vacas Cebú mal alimentadas
durante el período de sequía con PMSG y HCG. Centro Ser. Cienc. Anim. II, 1-2: 125.
264. Sadleir R.M.F.S. (1969). The ecology of Reproduction in wild and domestic mammals. Methven,
London, 321 pp.
265. Salirbury G.W. (1946). (Inédito) Cornell Univ. Citada por Salisbury y Vandemark. 1961.
266. Salirbury G.W. and N.L. Vandemark, (1961). Physiology of reproduction and artificial insemination of
cattle. Phyladelphia, New York, W.H. Freeman and company.
267. Santos A. y M. Pereiro. (1982). Efecto del tratamiento hormonal en la fertilidad de novillas F1 (Hols-
tein x Cebú) en pastoreo y suplementadas con miel/urea 2% durantela seca. Rev. Cubana Cienc.
Agric. 16: 17.

268. Saumande J. (1978). Relationships between ovarian stimulation by PMSG and steroid secretion,
169-194. En: Sreenan J.M., EEC Symp.: Control of reproduction in the cow, Nijhoff. The hague,
Boston, London.
269. Sawin C.T. (1971). Las hormonas, Fisiología endocrina. 1ra. ed. p.153. Salvat, S.A. Barcelona.
270. Scaramuzzi J., D.T. Baird. (1977). Pulsatile release of luteinizing hormone and the seccretion of
ovarian steroids in sheeep during anestrus. Endocrinol. 101:1801
271. Scaramuzzi R.J., D.T. Baird, N.D. Martenz, K.E. Turnbukll and P.F.A. Van Look. (1981). Ovarian
function in the ewe after active inmunization against testosterone. J. Reprod. Fert. 61:1.
272. Schallenberger E., J. Rampp and D.L. Walters. (1983). Pulsatile progesterone secretion occurs
during midgestation in the cow even though pulsatile LH secretion is abolished. J. Anim. Sci. 57:
371.
273. Schally A.V., A. Arimura, Y. Baha, R.M.G. Noir, H. Matsvo, T.W. Redeling, L. Debeljok and W.F.
white. (1971). Isolation and properties of the FSH and LH realising hormone. Biochm. Biophys. Res.
Comm. 43: 393.
274. Schams D. Buts E.D. (1972). Zeitliche Bezichungen swischen Brunstsymptomen, elektrischn widers
tandsmessungen des vaginalscheims, präovulatorischer Ausschüttung des luteinisierungshormons
und ovulation brim rind. Zuchthyg 7:49.
275. Schams D., Ch. Menzer, E. Schallenberger, B. Hoffmann, J. Hahn and R. Hahn. (1978). Some stud-
ies on pregnant mare serum gonadotrophin (PMSG) and on endocrine responses after application
for superovulation in cattle. En: Sreenan J. M., CEC Semin. Control of reproduction in de cow. Gal-
way. (1977), 122-143.
276. Schams D., Schallenberger E., Gombe S., Karg H. (1981). Endocrine patterns associated with pu-
berty in male and female cattle. J. Reprod. Fert. Suppl. 30, 103-110
277. Schillo K.K., D.J. Diersch ke and E.R. Hauser. (1982). Influences of month of birth and age on pat-
terns of luteinizing hormone secretion inprepubertal heifers. Theriogenology 18: 593.
278. Schillo K.K., Dierschke D.J., Hauser E.R. (1983b). Estrogen induced release of luteinizing hormone
in prepubertal and postpubertal heifers. Theriogenology 19: 727.
279. Schillo K.K., Hansen P.J., Kamwanja L.A., Dierschke D.L., Hauser E.R. (1983a). Influence of sea-
son on sexual development in heifers: Age at puberty as related to growth and serum concentrations
of gonadotropins, prolactin, thyroxine and progesterone. Biol Reprod. 28: 329.
280. Schillo Keith K. (1992). Effects of dietary energy on control of luteinizing hormone secretion in cattle
and sheep. J. Anim. Sci. 70:1271.
281. Schoenemann H.M. (1985). III Studies of pituitary luteinizing hormone releasing hormone (LHRH)
receptor in the cow. Diss. Abstr. Interns. B. (Sci. and Engin) 45: 2459.
282. Schoppec PD; Armstrong JD; Harvey RW; et al. (1995). Endocrine and ovarian response to exoge-
nous estradiol-17 beta in 6-month old heifers previously immunized against growth hormone releas-
ing factor. J. Anim. Sci. 73: 2071.
283. Schoppec PD; Armstrong JD; Harvey RW; et al. (1996). Immunization against growth hormone re-
leasing factor or chronic feed restriction initiated at 3.5 months of age reduces ovarian response to
pulsatile administration of gonado tropin-releasing hormone at 6 months of age and delays onset of
puberty in heifers. Biology Reprod. 55: 87.
284. Seidel G.E., L.L. Larson and R.H. Foote. (1971). Effects of age and gonadotropin treatment on su-
perovulation in the calf. J. Anim. Sci. 33: 617
285. Seift B.L., R.W. Johnson and G.M. Thomas. (1983). Anestrus in cycling beef heifers experimenttally
incoculated with anaplasma marginale. Theriogenology 20: 715.
286. Sergent D., Gauthier D., Vhemineau R. (1984). Effect of exposure to sunlght on some endocrinolog-
ical traits in creole goats and cattle in Guadeloupe. Reproduction des ruminants en zone tripicale.
Reunión internationale, Pointe-á-Pitre, Guadeloupe, 8-10.
287. Sheffield L.G., Ellicott A.R. (1982). Effect of levels of exogenous progesterone on puberty in beef
heifers. Theriogenology 18: 177.
288. Shemesh M. and Hansel W. (1974). Measurement of bovine plasma testosterone by radioimmuno-
assay (RIA) and by rapid competitive protein binding (CPB) assay. J. Anim. Sci. 39: 720.

289. Shorron S.M., J.H.B. Roy and G.S. Pope. (1978). Plasma progesterone concentrations from before
puberty to after parturition in british friesian heifers reared on high planes of nutrition and inseminat-
ed at their first oestrus. Anim Prod. 27: 89.
290. Short R.E. y Bellows R.A. (1971). Relation ships among weihgt gains, age at puberty and reproduc-
tive performance in heifers. J. Anim. Sci. 43: 1254.
291. Short R.E., Howland B.E., Randel R.., Christensen D.S. and Bellows R.A. (1973). Induced LH re-
lease in spayed cows. J. Anim. Sci. 37: 551.
292. Short R.E., R.A. Bellows, J.B. Carr, R.B. Staigmiller and R.D. Randel. (1976). Induced or synchro-
nized puberty in heifers. J. Anim. Sci. 43: 1252.
293. Short R.E., R.D. Randel, R.B. Staigmiller and R.A. Bellows. (1979). Factors affecting estrogen-
induced LH release in the cow. Biol. Reprod. 21: 683.
294. Skagg C.L., B.V. Able and J.S. Stevenson (1986). Pulsatile or continuous infusion of LH-RH and
hormonal concentration in prepubertal beef heifers. J. Anim. Sci. 62: 1034.
295. Smith, M.F. (1986). Recent advances in corpus luteum physiology. J. Dairy Sci. 69: 911.
296. Solano R., Fernández O. y Martínez G. (1983). Dinámica de los niveles de progesterona y LH ca-
racterísticas del ciclo estral en novillas Holstein bajo condiciones climáticas tropicales. Libro Res. III
Sem. Cient. CENSA, La Habana dic. 15-17, p. 200.
297. Sorensen AM, Hansel W, Hough WH, et al.(1959).Causes and prevention of reproductive failures in
dairy cattle. I. Influence of under feeding and overfeedingon growth and development in Holstein
heifers. Cornell Univ Agr Expt Sta Bull :936.
298. Spicer. L.J., K.Sejrsen, H.A. Tucker and J.T. Huber, (1984). Secretion of luteinizing hormona and
follicle. Stimulating hormone from overfeediung dairy heifers. J. Dairy Sci. 67; 1993.
299. Spilman C.H., G.E. Jr. Seidel, L.L. Larson and R.H. Foote. (1972). In vitro progesterone synthesis by
corpora lutea induced in prepuberal cattle. J. Anim. Sci, 34: 1025.
300. Spilman, C.H., G.E. Seidel, J.R., L.L. Larson, G.R. Vukman and R.H. Foote, (1973). Progesterone,
20 -hidroxipreg.4.en.3.one, and luteinizing hormone levels in superovulated prepuberal and post
puberal. Cattle. Biol. Reprod. 9: 116.
301. Spitzer J.C. (1982). Pregnancy rate in peripuberal beef heifers following treatment with syncromate-
B and gonadotropin releasing.hormone. Theriogenology 17. 373.
302. Spitzer, J.C., G.D. Niswender, G.E. Seidel. Jr. and J.N. Wiltbank, 1978. Fertilization and blood levels
of progesterone and LH in beef neifers on a restricted energy diet. J. Anim. Sci. 46: 4; 1071.
303. Stabenfeldt G.H. (1974). Physiologic, pathologic and therapeutic roles of progestins in domestic
animals. J. Amer. Vet. Med. Assoc. 164:311.
304. Stabenfeldt G.H., H. Kindahl, L.E. Edquist. (1976). Radioisotopic techniques for the study of repro-
ductive physiology in domestic animals; 2, Physiological implications. En. Nuc. Tech.Anim. Prod.
Hesalth Proc. Symp.
305. Staigmiller R.B., R.E. Short and R.A. Bellows. (1979). Induction of LH surges with 17 estradiol in
prepuberal beef heifers: An age dependent response. Theriogenology 11: 453.
306. Stanisiewski E.P., N.K. Ames, L.T. Chaplin, C. Blaze, and H.A. Tucker. (1986). Effect of pinealec-
tomy on prolactin and testosterone concentrations in plasma of bull calves exposed to 8 or 16 hours
of light per day. J. Dairy Sci. 69 (Suppl. 1) p. 166 (abstr.)
307. Steward F., Allen W.R. and Moor R.M. (1976). Pregmant marte serungonadotropin;Rtio of follicle
stimulating and luteinizing hormone activities measured by radioreceptor assay. J. Endocri. 71:371.
308. Suzuki O. and Sato M. (1980). (Puberty and serum levels of progesterone and estradiol-17 in beef
heifers) Bull. Natn. Grassld Res. Inst. Japan 16: 96.
309. Suzuki O. Sato M., Kubota Y. (1982). Effects of under feeding on oestrous behavious and ovarian
function in post pubertal beed heifers. Japanese J. Anim. Reprod. 28:205.
310. Swanson L.V., H.D. Hafs and D.A. Morroe. (1972). Ovarian characteristic and serum LH, prolactin,
progesterone and glucocorticoid from first estrus to breeding size in Holstein heifers. J. Anim. Sci.
34: 284.

311. Swift B.L., W.J. Murdoch and R.R. Dahlgren. (1984). Anemia associeted with anestrus in beef heif-
ers inoculated with anaplasma marginale: Endocrinology and ovarian changes. Theriogenology 22:
643.
312. Sygnoret J.P. (1967). Duree du cycle oestrien et de loestrus chez la trvi, action du benzoate does-
tradiol chez la femelle ovariectomisee. Ann. Biol. Anim. Bioch. Biophys 7:1.
313. Symons, A.M. (1976). The LH response of pregnant cows and their calves to synthetic LH-RH. J.
Reprod. Fert. 46: 235.
314. Tanaka Y., Vincent D.L., Ledgerwood K.S. et al.. (1995). Variable progesterone response and es-
tradiol secretion in prepubertal beef heifers following teatment with norgestomet implants. Theri-
ogenology 43:1077.
315. Tarrero R., Pedroso, R. Perón N. y Santiago N. (1981). Influencia de la alimentación en terneras ¾
Holstein x ¼ Cebú durante el crecimiento y su comportamiento reproductivo posterior. Informe pre-
liminar. Resumenes II Seminario Cient. CENSA. La Habana.
316. Tarrero R. y Perón N. (1983). Influencia de tres dietas de miel y urea en el comportamiento repro-
ductivo de las novillas. Libro Res. Y 4ta. Reunión ACPA, 12-14 dic. La Habana p.145.
317. Tezuka, M., M. Irahara, et al. (2002). Effects of leptin on gonadotropin secretion in juvenile female
rat pituitary cells. Eur J Endocrinol 146(2): 261.
318. Thatcher W.W, Smith EJP, de la Sota RL et al. (1996). Desarrollo folicular en el período del post-
parto: anestro, efecto, del balance energético, nutrición y condición corporal sobre el desarrollo foli-
cular y la fertilidad en el postparto de vacas en lactación. II Simposio Int. Reprod. Anim. 31 Oct.- 2
Nov. Carlos Paz. Córdoba, Argentina pp. 23-52.
319. Thibaulth Ch. and Marie - Claire Levasseur. (1974). Reproductive life Cycle. En reproduction in
Farm Animals, de. E.S.E. Hafez, De. Lea and Febiger Philad. 3era. de. p.82.
320. Thibier M. (1981). Hormonologie de la reproduction un now veau concept: La regulation endocrine
par modulation de frecuence. Rec. Med. Vet., 157:15
321. Topps J.H. (1977). The relationship between reproduction andundernutrition in beet cattle. Woel
Reviev Anim. Reprod. 13:2; 43.
322. Tucker H.A. (1982). Seasonality in cattle. Theriogenology 17:53.
323. Vichot, O. Cortada A., Estrada J., Fernández F. (1976). Comportamiento reproductivo de novillas
anéstricas tratadas con progesterona y gonadotropina sérica (PMSG). II Cong. Nac. Cienc. Vet. LA
Habana. p.6.
324. Vincent D.L. (1983). Ovarian and pitvitary responses of seasonally and postpartum anestrous
eswesto gonadotropin releasing hormone. Dissertation Abstracts International, B. (Sciences and
Angineering) 44:1657.
325. Wagner J.F., Veenhuizen E.L., Tankinson L.V> and Rathmacher R.P. (1973).Effect of placental
gonadotrophin on pregnancy rate in the bovine. J. Anim. Sci. 36:1129
326. Walters D.L., D. Schams and E. Schallenberger. (1984). Pulsatile secretion of gonadotrophins,
ovarian steroids and ovarian, oxytocin during the luteal phase of the oestrous cycle in the cow. J.
Reprod. Fert. 71:479.
327. Walters D.L., R.E. Short, E.M. Convey, R.B. Staigmiller, T.G. Dunn and C.L. Kaltenbach. (1982).
Pitvitary and ovarian function in postpartum beef cows. II Endocrine Changes prior to ovulation in
suckled and non sucklet postpartum cows compared to cycling cows. Biol. Reprod. 26:647.
328. Wehrman M.E., Kijima F.N., Sánchez T. (1996). Incidence of precocious puberty in developing beef
heifers. J. Anim. Sci. 74:2462.
329. Werre JF, Brinks JS. (1986). Relationships of age at pubertu with growth and subsequent productivi-
ty in beef heifers. Proc. West sect Am. Soc. Anim. Sci. 37: 300.
330. Wettemann RP, Castree J.W. (1994). Immunization of heifers against gonadotropin releasing hor-
mone delays puberty and causes the cessation of estrous cycles. Anim Reprod. Sci. 36: 49.
331. Yelich J.V., Wettemann R.P., Dolezal H.G. et al. (1995). Effect of growth rate on carcass composi-
tion and lipid partitioning ot puberty and growth hormone, insuline like growth factor Y, insulin and
metabolites before puberty in beef heifers. J. Anim. Sci. 73:2390.

332. Yellon S.M., Bittman E.L., Lehman M.N., Olster D.H., Robinson J.E., Karsch F.J. (1985). Importance
of duration of nocturnal malatonin secretion in determining the reproductive response to inductive
photoperiod in the ew. Biol. of Reprod. 32:523.
333. Zamora A. y Amparo González. (1986). Efecto del nivel de suplementación en la ganancia diaria y
comportamientoreproductivo de novillas Holstein en pasto estrella. Y Primera Réplica 1982-83. Re-
sum. 5ta. Reunión ACPA. La Habana.
334. Zamora A., A. Elias y L. Zarragoitia. (1985). Comportamiento de novillas lecheras en pasto pangola
(Digitaria decumbens Stent) con diferentes cargas. Y Primer año iniciado en la época de seca. Rev.
Cub. Cienc. Agric. 19:247.
335. Zemjanis R. (1974). Pregnancy diagnosis en: Reproduction in farm animal E.S.E. Hafez, 3th. de.
Lea and Febiger. Philadelphia.
336. Zhied A.A., H.A. Garberick, C.J. Bierschwal, R.G. Elmore, R.S. Youngquist and A.J. Sharp. (1980).
Effect of ovarian activity and endogenous reproductive hormones on GnRH - induced ovarian cycles
in postpartum dairy cows. J. Anim. Sci. 50:3, 508.
337. Zieba, D. A., M. Amstalden, et al. (2004). Regulatory Roles of Leptin at the Hypothalamic-
Hypophyseal Axis Before and after Sexual Maturation in Cattle. Biol Reprod 71(3): 804
338. Zolman J.E., Convey L.W., Brittand H.P. y Hafs H.D. (1973). Release of bovine LH by purified por-
cine and synthetic gonadotrophin releasing hormone. Proc. Sco. Expth. Bio. Med. 142:189
Autores:
Dr. Roberto Faure García
Dr. Octavio Fernández Limia
Dirección de Salud y Producción Animal,
Centro Nacional de Sanidad Agropecuaria
La Habana, Cuba
Septiembre de 2006


















