Sall1-dependent signals affect Wnt signaling and ureter ... · KEY WORDS: Sall1, Spalt, Ureter,...
8
3099 RESEARCH ARTICLE INTRODUCTION A crucial step in kidney development is the interaction between the outgrown ureteric bud (ub) and the metanephric mesenchyme (mm). Reciprocal signaling triggers the ub to initiate its first T- branch and induces the mm to condense and proliferate. If these tissues fail to interact properly, renal agenesis or severe hypoplasia will occur. Despite the importance of this interaction, defining the precise molecular events that regulate these processes has been particularly difficult because the mm undergoes rapid apoptosis if it is not properly invaded (Donovan et al., 1999). Here, we describe a mouse model in which the initial invasion of the ureter has been captured and can be studied to identify factors that are crucial for this process. The gene mutated in this mouse model is the transcription factor Sall1 – a multi-zinc-finger transcriptional regulator that is expressed in the mm (Nishinakamura et al., 2001). It is mutated in the autosomal dominant syndrome Townes Brocks (TBS) which is characterized by multi-organ defects (including renal hypoplasia). In this report, we use a Sall1 mouse model that mimics TBS (Sall1- Zn) and demonstrate that these mice form blind ureters with an overlying undifferentiated cap of mm (Kiefer et al., 2003). The ub grows out and invades the mm, but does not form the first T- branch. The unbranched ub and mm remain associated throughout development, thereby allowing the precise dissection of the initial events in ub morphogenesis that depend on Sall1. We show that Sall1 function in the mm triggers downregulation of the stalk- specific marker Wnt9b and the canonical Wnt downstream gene Axin2 in the outgrown ub. Early ub branching is rescued by a reduction in -catenin dose supporting the conclusion that ureter arrest is due to altered Wnt signaling in Sall1-Zn mutants. This theory is further strengthened by findings that ub branching is reduced in Wnt9b transgenic lines and in a stabilized -catenin mutant. These results demonstrate that downregulation of Wnt9b and canonical Wnt signaling in the ureter tip is required to initiate ub branching. MATERIALS AND METHODS Mouse strains Sall1-Zn mice have been described elsewhere (Kiefer et al., 2003). Double heterozygous Sall1-Zn;Hoxb7GFP were mated to Sall1-Zn to create EGFP-labeled mutant Wolffian ducts and ureters (Srinivas et al., 1999). Sprouty1 (Spry1) mice were a gift from J. Licht (Basson et al., 2005). Reduction of -catenin activity was achieved by using a -catenin floxed allele (Ctnnb1fl/+, Jackson Laboratories) that had been first crossed to a -actin-cre transgenic and then to Sall1-Zn to create double heterozygous Sall1-Zn +/– ;Ctnnb1 +/– males. These were mated to Sall1- Zn +/– ;Hoxb7GFPtg/+ females to analyze Sall1-Zn –/– ;Ctnnb1 +/– embryos. The presence of -actin-cre or Hoxb7GFP transgenes had no effect on the phenotypes observed. Activation of -catenin was achieved by crossing Hoxb7creEGFP transgenic mice (Zhao et al., 2004) to a - catenin exon 3 floxed allele (Ctnnb1 ex3fl/ex3fl, Jackson Laboratories) (Harada et al., 1999). Conditional deletion of this exon produces a stabilized -catenin protein in both ureter and Wolffian duct. These studies were performed under the auspices of St Louis University and St Louis VA Medical Center animal care guidelines. Noon on the day of the vaginal plug detection was considered to be E0.5. Overexpression of Wnt9b in the ub Transgenic mice were created to conditionally overexpress Wnt9b (ACTWnt9b). A C-terminally epitope-tagged Wnt9b (Wnt9b-FLU) was inserted into the EcoRI site of a previously described transgenic construct [a gift from Yiping Chen, Tulane University (Chen et al., 2004)]. In this construct, expression of Wnt9b-FLU is prevented by an upstream loxed Development 137, 3099-3106 (2010) doi:10.1242/dev.037812 © 2010. Published by The Company of Biologists Ltd 1 John Cochran Veterans Affairs Medical Center, St Louis, MO 63106, USA. 2 Department of Internal Medicine, Division of Nephrology, St Louis University, St Louis MO 63104, USA. 3 Department of Medicine, Division of Dermatology, Washington University St Louis, St Louis, MO 63110, USA. *Author for correspondence ([email protected]) Accepted 25 June 2010 SUMMARY Development of the metanephric kidney depends on precise control of branching of the ureteric bud. Branching events represent terminal bifurcations that are thought to depend on unique patterns of gene expression in the tip compared with the stalk and are influenced by mesenchymal signals. The metanephric mesenchyme-derived signals that control gene expression at the ureteric bud tip are not well understood. In mouse Sall1 mutants, the ureteric bud grows out and invades the metanephric mesenchyme, but it fails to initiate branching despite tip-specific expression of Ret and Wnt11. The stalk-specific marker Wnt9b and the - catenin downstream target Axin2 are ectopically expressed in the mutant ureteric bud tips, suggesting that upregulated canonical Wnt signaling disrupts ureter branching in this mutant. In support of this hypothesis, ureter arrest is rescued by lowering -catenin levels in the Sall1 mutant and is phenocopied by ectopic expression of a stabilized -catenin in the ureteric bud. Furthermore, transgenic overexpression of Wnt9b in the ureteric bud causes reduced branching in multiple founder lines. These studies indicate that Sall1-dependent signals from the metanephric mesenchyme are required to modulate ureteric bud tip Wnt patterning in order to initiate branching. KEY WORDS: Sall1, Spalt, Ureter, Wnt9b, Kidney development, Ureteric bud, Metanephric mesenchyme, -catenin, Wnt, Tip fate, Mouse Sall1-dependent signals affect Wnt signaling and ureter tip fate to initiate kidney development Susan M. Kiefer 1,2 , Lynn Robbins 1,2 , Kelly M. Stumpff 2 , Congxing Lin 3 , Liang Ma 3 and Michael Rauchman 1,2, * DEVELOPMENT
Transcript of Sall1-dependent signals affect Wnt signaling and ureter ... · KEY WORDS: Sall1, Spalt, Ureter,...

3099RESEARCH ARTICLE
INTRODUCTIONA crucial step in kidney development is the interaction between theoutgrown ureteric bud (ub) and the metanephric mesenchyme(mm). Reciprocal signaling triggers the ub to initiate its first T-branch and induces the mm to condense and proliferate. If thesetissues fail to interact properly, renal agenesis or severe hypoplasiawill occur. Despite the importance of this interaction, defining theprecise molecular events that regulate these processes has beenparticularly difficult because the mm undergoes rapid apoptosis ifit is not properly invaded (Donovan et al., 1999). Here, we describea mouse model in which the initial invasion of the ureter has beencaptured and can be studied to identify factors that are crucial forthis process.
The gene mutated in this mouse model is the transcription factorSall1 – a multi-zinc-finger transcriptional regulator that isexpressed in the mm (Nishinakamura et al., 2001). It is mutated inthe autosomal dominant syndrome Townes Brocks (TBS) which ischaracterized by multi-organ defects (including renal hypoplasia).In this report, we use a Sall1 mouse model that mimics TBS (Sall1-Zn) and demonstrate that these mice form blind ureters with anoverlying undifferentiated cap of mm (Kiefer et al., 2003). The ubgrows out and invades the mm, but does not form the first T-branch. The unbranched ub and mm remain associated throughoutdevelopment, thereby allowing the precise dissection of the initialevents in ub morphogenesis that depend on Sall1. We show that
Sall1 function in the mm triggers downregulation of the stalk-specific marker Wnt9b and the canonical Wnt downstream geneAxin2 in the outgrown ub. Early ub branching is rescued by areduction in -catenin dose supporting the conclusion that ureterarrest is due to altered Wnt signaling in Sall1-Zn mutants. Thistheory is further strengthened by findings that ub branching isreduced in Wnt9b transgenic lines and in a stabilized -cateninmutant. These results demonstrate that downregulation of Wnt9band canonical Wnt signaling in the ureter tip is required to initiateub branching.
MATERIALS AND METHODSMouse strainsSall1-Zn mice have been described elsewhere (Kiefer et al., 2003).Double heterozygous Sall1-Zn;Hoxb7GFP were mated to Sall1-Zn tocreate EGFP-labeled mutant Wolffian ducts and ureters (Srinivas et al.,1999). Sprouty1 (Spry1) mice were a gift from J. Licht (Basson et al.,2005). Reduction of -catenin activity was achieved by using a -cateninfloxed allele (Ctnnb1fl/+, Jackson Laboratories) that had been first crossedto a -actin-cre transgenic and then to Sall1-Zn to create doubleheterozygous Sall1-Zn+/–;Ctnnb1+/– males. These were mated to Sall1-Zn+/–;Hoxb7GFPtg/+ females to analyze Sall1-Zn–/–;Ctnnb1+/–
embryos. The presence of -actin-cre or Hoxb7GFP transgenes had noeffect on the phenotypes observed. Activation of -catenin was achievedby crossing Hoxb7creEGFP transgenic mice (Zhao et al., 2004) to a -catenin exon 3 floxed allele (Ctnnb1 ex3fl/ex3fl, Jackson Laboratories)(Harada et al., 1999). Conditional deletion of this exon produces astabilized -catenin protein in both ureter and Wolffian duct. These studieswere performed under the auspices of St Louis University and St Louis VAMedical Center animal care guidelines. Noon on the day of the vaginalplug detection was considered to be E0.5.
Overexpression of Wnt9b in the ubTransgenic mice were created to conditionally overexpress Wnt9b(ACTWnt9b). A C-terminally epitope-tagged Wnt9b (Wnt9b-FLU) wasinserted into the EcoRI site of a previously described transgenic construct[a gift from Yiping Chen, Tulane University (Chen et al., 2004)]. In thisconstruct, expression of Wnt9b-FLU is prevented by an upstream loxed
Development 137, 3099-3106 (2010) doi:10.1242/dev.037812© 2010. Published by The Company of Biologists Ltd
1John Cochran Veterans Affairs Medical Center, St Louis, MO 63106, USA.2Department of Internal Medicine, Division of Nephrology, St Louis University,St Louis MO 63104, USA. 3Department of Medicine, Division of Dermatology,Washington University St Louis, St Louis, MO 63110, USA.
*Author for correspondence ([email protected])
Accepted 25 June 2010
SUMMARYDevelopment of the metanephric kidney depends on precise control of branching of the ureteric bud. Branching events representterminal bifurcations that are thought to depend on unique patterns of gene expression in the tip compared with the stalk andare influenced by mesenchymal signals. The metanephric mesenchyme-derived signals that control gene expression at the uretericbud tip are not well understood. In mouse Sall1 mutants, the ureteric bud grows out and invades the metanephric mesenchyme,but it fails to initiate branching despite tip-specific expression of Ret and Wnt11. The stalk-specific marker Wnt9b and the -catenin downstream target Axin2 are ectopically expressed in the mutant ureteric bud tips, suggesting that upregulatedcanonical Wnt signaling disrupts ureter branching in this mutant. In support of this hypothesis, ureter arrest is rescued bylowering -catenin levels in the Sall1 mutant and is phenocopied by ectopic expression of a stabilized -catenin in the uretericbud. Furthermore, transgenic overexpression of Wnt9b in the ureteric bud causes reduced branching in multiple founder lines.These studies indicate that Sall1-dependent signals from the metanephric mesenchyme are required to modulate ureteric bud tipWnt patterning in order to initiate branching.
KEY WORDS: Sall1, Spalt, Ureter, Wnt9b, Kidney development, Ureteric bud, Metanephric mesenchyme, -catenin, Wnt, Tip fate, Mouse
Sall1-dependent signals affect Wnt signaling and ureter tipfate to initiate kidney developmentSusan M. Kiefer1,2, Lynn Robbins1,2, Kelly M. Stumpff2, Congxing Lin3, Liang Ma3 and Michael Rauchman1,2,*
DEVELO
PMENT

3100
stop cassette (Novak et al., 2000). Upon exposure to cre recombinase, thestop cassette is removed and transgenic expression is driven by the chicken-actin promoter. Ten independent founder lines were established by theWashington University Mouse Genetics core. Five of these lines werecrossed to Hoxb7creEGFP transgenic mice to overexpress Wnt9b in the ublineage. Kidneys were photographed using a Leica DVC300FX camera anda M165FC fluorescence-enabled microscope. Photographs of all littermateswere scored by at least two individuals blind to the genotype and examinedfor reduced kidney size and abnormal morphology. Statistical significancewas determined by chi square analysis. The ub branch tips were alsocounted and are reported with standard error and the statistical differencedetermined by t-test. Expression of Wnt9b-FLU in E13.5 embryonickidneys was confirmed by western blotting of kidney lysates probed withanti-FLU antibody (12CA5, Roche, Indianapolis, IN, USA) and detectedwith anti-mouse HRP (Cell Signaling Technology, Danvers, MA, USA) asdescribed previously (Kiefer et al., 2002).
Histology, immunohistochemistry and immunofluorescenceUrogenital regions were dissected at E11.0-18.5. Tissues were fixed,sectioned and stained with Hematoxylin and Eosin as described previously(Kiefer et al., 2003). Corresponding unstained slides were used forimmunohistochemistry with anti-uroplakin III (1:10, Fitzgerald Industries,Concord, MA, USA) and anti-smooth muscle actin (1:100, Millipore,Billerica, MA, USA) as described previously (Mahoney et al., 2006).Immunofluorescence was performed on 10 m frozen sections using rabbitanti-Pax2 antibody (1:2000, Covance, Princeton, NJ, USA) and rat anti-laminin B2 chain antibody (1:2000, Chemicon, Temucula, CA, USA).Antibody reactivity was detected using Alexa 488-labeled anti-rabbit(1:400, Invitrogen, Carlsbad, CA, USA) and Cy3-labeled anti-rat (1:2000,Jackson ImmunoResearch, West Grove, PA, USA), and mounted inMowiol (Polysciences, Warrington, PA, USA) as previously described(Kiefer et al., 2002). Sections were incubated without primary antibody tocontrol for non-specific staining of secondary antibodies.
In situ hybridizationWhole-mount in situ hybridization was performed using digoxigenin-labeled antisense riboprobes for Ret (nucleotides 818-1222), Pax2 (563-1248), Six2 (136-656), Cited1 (1-612), Wnt11 (61-674), Wnt9b (486-1076)and Sall1 (421-848) (Kiefer et al., 2008). In situ hybridization wasperformed as described previously (Little et al., 2007) on 20 m frozensections using a digoxigenin-labeled Gdnf riboprobe (nucleotides 122-781).After incubation with digoxigenin-alkaline phosphatase antibody (1:2500),signal was visualized using the alkaline phosphatase substrate BM purple(Roche, Indianapolis, IN, USA).
Quantitative RT-PCRE11.5 wild-type or Sall1Zn mutants were dissected to isolate the uretertips and mesenchyme from each side of the T-stage or unbranched ureter,respectively. Ten tip regions were pooled and cDNA was synthesized asdescribed previously (Kiefer et al., 2008). Samples that lacked reversetranscriptase were prepared as negative controls. Primer sets were designedto amplify Gdnf, Pax2 and Gpc4 specifically using Primer Bank(http://pga.mgh.harvard.edu/primerbank/). cDNA (10 ng) was used forquantitative real-time PCR using SYBR green master mix (AppliedBiosystems, Foster City, CA, USA) and an ABI 7300 quantitative PCRinstrument (Kiefer et al., 2008). Relative levels of mRNA expression werecalculated according to the CT method and normalized to the geneRpl19. Transgenic copy number was estimated using primers at the Cterminus of Wnt9b-FLU (forward GGAGCTCGTGTATACCTGCAA,reverse CCCTTTCGTCTTCAAGAATTCTC) and 10 ng of HindIII cutgenomic DNA. CT values were normalized against a Gapdh-positivecontrol for transgene PCR.
Organ cultureAffi-blue sepharose beads (Biorad, Hercules, CA, USA) were preincubatedwith 10 mg/ml recombinant GDNF (R&D Systems, Minneapolis, MN,USA) or BSA (Sigma, St Louis, MO, USA) for 30 minutes at 37°C.Hoxb7GFP-expressing E11.5 wild-type and Sall1-Zn ubs and theirassociated mm were cultured in Trowell-type organ culture for 48-72 hours
with DMEM containing 10% FBS. Two or three beads were placed on eachfilter near the rudiments and imaged on the day of culture and 48-72 hourslater. At least five individual cultures were tested for each treatment. Forrecombination experiments, metanephric regions were digested with 0.5%Trypsin/0.02 g/l EDTA (Hyclone, Logan, UT, USA) for 5-15 minutes onice to aid separation of the ub and mm. Wild-type E11.0 ubs or Sall1-ZnE12.5 blind ureters were recombined with wild-type E12.5 mm tissue andcultured for 48-72 hours. The ub branching was monitored by imaging thecultures on a Leica MZ16F stereomicroscope equipped with GFPfluorescence and a DFC300FX color camera.
RESULTSSall1-Zn mice exhibit blind ureters that aremorphologically normalSall1-Zn/ homozygous mice exhibit bilateral renal agenesis(Kiefer et al., 2003). It has been suggested that this phenotype is dueto incomplete ureter outgrowth and to mesenchymal apoptosis inSall1-null mice (Nishinakamura et al., 2001). However, upon closerinvestigation, we were able to discern outgrown ureters in at least 25independent E14.5-18.5 Sall1-Zn/ embryos in which there wasno evidence of kidney formation (Fig. 1). These blind ureters hadgrown away from the bladder similar to those in wild-typelittermates, despite no evidence of a morphologically distinctmetanephric mesenchyme. Blind ureters exhibited normalhistological architecture that consisted of urothelium (u) and smoothmuscle (sm) layers, as revealed by Hematoxylin and Eosin staining.By E16.5, the smooth muscle layer should have differentiated andbegun to express smooth muscle actin; this occurred normally in themutant ureters (Fig. 1E,F). Both wild-type and mutant ureters alsoproperly expressed a marker that is exclusive to the differentiatedepithelium of the urinary tract: uroplakin III (Fig. 1G,H). Six1–/–
mutants display a very similar phenotype in which renal agenesis isaccompanied by growth and differentiation of the ureter (Bush et al.,2006), supporting the conclusion that development of the extra-renalureter is independent of the proximal branching ub that forms theintra-renal collecting system. However, unlike Six1 (Nie et al., 2010),Sall1 is not required for proper formation of the smooth muscle layer.Sall1 expression is not preserved in Six1–/– mutant mm and it hasbeen suggested that Sall1 is a direct transcriptional target of Six1regulation (Chai et al., 2006). These findings suggest that Six1 andSall1 function together in a developmental pathway that regulatesinitial branching of the ureter.
Blind ureters arrest before the T-stage andmaintain persistent mesenchymal cellsTo verify the identity of the outgrown ureters and to visualize ureterdevelopment at multiple stages, we crossed Sall1-Zn mice with theHoxb7GFP transgenic line (F. Costantini, Columbia University).Visualization of GFP fluorescence verified the existence of blindureters in Sall1-Zn/ mutant E16.5 embryos (Fig. 2). These uretershad grown at the proper site on either side of the aorta and weremorphologically distinguishable from the surrounding tissue. Thedistal tips appeared dilated and the lengths were often asymmetric.At E11.5, when the wild-type ub has formed its first branch (T-stage),mutant ubs remained unbranched (Fig. 2C,D). Both wild-type andmutant ub emerged from the distal Wolffian duct and invaded themetanephric mesenchyme; however, the mutant ub failed to form theT-stage. This unbranched ub remained viable and continued to growin the absence of proper mesenchymal induction.
Ret was properly expressed in the mutant ub and becamerestricted to the ureter tip by E12.5 (Fig. 3C,D). Analysis of Pax2expression revealed a small cluster of mesenchyme surrounding the
RESEARCH ARTICLE Development 137 (18)
DEVELO
PMENT

ureter tip, suggesting that blind ureter formation is associated witha persistent cap of mesenchyme (Fig. 3E,F). The mutantmesenchyme contained Six2-positive metanephric progenitor cells(Fig. 3G,H). These cells did not progress beyond the progenitorstage because a gene expressed specifically in cap mesenchyme,Cited1, was absent (Fig. 3I,J) (Mugford et al., 2009). Consistentwith this observation, there was no evidence of formation ofpretubular aggregates in the persistent mesenchymal cells and,similar to Sall1-null mice (Nishinakamura et al., 2001), expressionof several other mesenchymal markers, including Eya1 and Wnt4,were reduced compared with wild-type controls (data not shown).Together, these data show that uninduced metanephricmesenchyme persists atop mature blind ureters and demonstratethat Sall1 expression is required for initial T-branch formation.
Upregulation of GDNF signaling is not sufficientto rescue branchingOur data suggest that Sall1-dependent signals from the mm tothe ub are required to initiate branching. It has been proposedthat a failure to maintain Gdnf expression probably accounts forrenal agenesis or for severe hypoplasia in Sall1 mutants (Schedl,
2007). The requirement for proper Gdnf expression in the criticalwindow of initial ub invasion has been shown through analysisof nephronectin (Npnt) and integrin 8 (Itga8) mutant mice(Linton et al., 2007). In these mutants, Gdnf expression appearsto be normal at the time of ub outgrowth, but is dramaticallyreduced at E11.5 when the ub invades the mesenchyme. Agenetic reduction of Spry1 gene expression rescued the kidneydefect in Itga8 mutants confirming that decreased GDNF-Retsignaling was responsible for renal agenesis in these mice.Therefore, we assayed expression of Gdnf in Sall1-Zn/
mutant mesenchyme at E11.5 and found that its expression wassignificantly reduced when measured using in situ hybridizationand quantitative real-time PCR (Fig. 3N, 0.22±0.05 fold change).Mesenchymal Pax2 expression appeared similar to wild-type atthe same time point (Fig. 3K,L).
To test the hypothesis that Gdnf deficiency accounts for ub arrestin the Sall1-Zn mutant, we next performed metanephric organculture with exogenous GDNF and with increased GDNF signalingachieved by reducing the dose of the Spry1 tyrosine kinaseinhibitor in vivo. For organ culture, E11.5 Hoxb7GFP wild-typeand homozygous Sall1-Zn/ metanephric regions were culturedwith beads coated with bovine serum albumin (BSA) orrecombinant GDNF protein. Recapitulating the in vivo phenotypes,mutant explants failed to branch, whereas wild-type explantsunderwent several rounds of branching when incubated with BSA-coated beads (Fig. 3O,P). As previously described, exogenousGDNF caused increased branching and altered patterning of thewild-type ub, confirming that GDNF was active in our culturesystem (Fig. 3Q). Addition of GDNF-coated beads resulted in onlya small single branch with abnormal morphology in eight out of 11mutants (Fig. 3R). Interestingly, mutant ureteric buds did not swellas dramatically as wild-type tissues when treated with exogenousGDNF, suggesting that ampulla formation is abnormal. A similarreduction in Gdnf expression at E11.5 was observed for Grem1-deficient embryos, which arrest prior to ub outgrowth. However, inthis mutant, exogenous GDNF induced ub branching of Grem1–/–
metanephroi in organ culture that was comparable with that seen inthe wild type (Michos et al., 2007). Thus, the failure of GDNF torescue the Sall1-Zn/ explants is best explained by an inabilityof the Sall1-Zn mutant to respond to this branch-promoting factor.
3101RESEARCH ARTICLESall1 affects ub Wnt signaling
Fig. 1. Sall1-Zn mutants exhibit blind ureters that properlydifferentiate into urothelium and smooth muscle layers.(A,B)E18.5 urogenital tracts from wild-type and homozygous mouseembryos demonstrate that Sall1-Zn mutants do not form kidneys (k),but that morphologically distinct ureters are visible. Testes (t) andbladder (bl) are formed. (C,D)Transverse sections at the level indicatedby the arrows are shown in the lower panels. Haemotoxylin and Eosinstaining reveals a proper urothelium (u) and smooth muscle (sm) layerin cross sections of both wild-type and mutant ureters. The mutantexhibits decreased luminal space that might be secondary to the lack ofurine flow. (E-H)Immunohistochemistry with anti-smooth muscle actinand anti-uroplakin III appears similar in wild-type and mutant sections,and confirms the proper differentiation of mutant ureters.
Fig. 2. Blind ureters are caused by failure to initiate the firstbranch. (A-D)Hoxb7GFP-labeled genital tracts from wild-type andmutant mice at E16.5 (A,B) and E11.5 (C,D) reveal that the wild-typeureteric bud (ub) properly forms a T-branch at E11.5 and a branchedkidney at E16.5. Sall1-Zn homozygous mutant ubs properly grow outfrom the Wolffian duct (wd), but fail to branch. Blind ureters persist atE16.5 in the mutant.
DEVELO
PMENT

3102
We next tested whether the deficiency in Gdnf could beovercome by reducing Spry1 dose in vivo. Spry1 normallyfunctions to attenuate GDNF activation of the Ret receptor, andloss of Spry1 in Spry1-null embryos produces multiple ectopic budsbecause of increased Ret signaling (Basson et al., 2005; Basson etal., 2006). Therefore, we postulated that increasing Ret signalingin the presence of low levels of GDNF would allow mutant uretericbuds to branch. In contrast to the rescue observed for Itga8-nullmice, heterozygosity of a Spry1-null allele (J. Licht, NorthwesternUniversity) did not alter the frequency of renal agenesis and blindureter formation for Sall1-Zn mutants in any of theSpry1+/–;Sall1/ mutants examined (n0/5). Together, these resultssuggest that although low levels of Gdnf might contribute to ureterarrest in the Sall1-Zn mutant, reduced signaling through thispathway alone cannot account for this phenotype.
Although Sall1 is not expressed in the ub, it is expressed in thenephric duct at E9-10, immediately prior to ub outgrowth. Thus,Sall1 expression in the duct might be required to pattern the ub andenable it to form an ampulla properly. To test this possibility, weperformed recombination experiments to determine whether Sall1-Zn/ ub is competent to branch when recombined with wild-typemm. As shown in Fig. 3S,T, mutant ubs underwent several roundsof branching morphogenesis (average4.3 ub tips with four out ofsix recombinations exhibiting branching) when recombined withwild-type mesenchyme; this was comparable with that observed inthe wild type } wild type recombination (average4.8 ub tips withfour out of six recombinations exhibiting branching). Wild-typerecombinations were performed prior to the T-stage to verify thatunbranched ubs were capable of branching in organ culture.Together, these results indicate that the Sall1-Zn/ mutant ub iscompetent to respond to mesenchymal signals, and suggest thatSall1-dependent factors in the mm, independently of GDNF, mustbe involved in the regulation of ampulla formation and theinitiation of ureteric bud branching.
Wnt9b is ectopically expressed in mutant uretericbud tipsWe next postulated that Sall1 might be affecting ureter tip fate. Ifmutant ub tips are not properly specified, they could adopt a distalub phenotype and prohibit branching (Caruana et al., 2006; Michaeland Davies, 2004; Schmidt-Ott et al., 2005; Shakya et al., 2005;Sweeney et al., 2008). Tip identity was specified in Sall1-Zn/
mutant ubs because both Ret (Fig. 3D) and Wnt11 (Fig. 4D) wereproperly tip restricted. As the ampulla begins to bifurcate in the wildtype, Wnt11 expression is polarized to delineate the emerging tips.However, in the mutant, Wnt11 remains restricted to the single tip,indicating that bifurcation has not been initiated. Wnt11 expressiondepends on intact GDNF-Ret signaling and is maintained by aWnt11-GDNF feedback loop (Majumdar et al., 2003; Schuchardt etal., 1995; Schuchardt et al., 1996). Therefore, the preservation ofWnt11 expression in the Sall1-Zn/ mutants is consistent with theidea that low GDNF activity is not the principal defect in blind ureterformation.
However, a different paradigm emerged when we analyzedWnt9b. Wnt9b expression is normally restricted to the ub stalk andWnt9b–/– embryos arrest when the ub has formed the T-stage (Carrollet al., 2005). In Sall1-Zn/ mutant ureters, Wnt9b expression wasnot downregulated and continued to be expressed at the ureter tip(Fig. 4F). Wnt9b is thought to signal through the canonical -cateninpathway during kidney development, as Wnt9b–/– embryos can berescued by expression of Wnt1 in the ub (Carroll et al., 2005).Consistent with this interpretation, a -catenin pathway member andtarget gene (Jho et al., 2002), Axin2, was also ectopically expressedin the ub tip in the Sall1-Zn/ mutant (Fig. 4G,H). Upregulation ofGpc4, a gene shown to be modulated by -catenin signaling in thekidney (Schmidt-Ott et al., 2007), also occurred in mutant rudiments(2.3±0.11 fold change). These data suggest that Sall1-dependentsignals in the mm cause the downregulation of canonical Wntsignaling in the ub tip resulting in initiation of branching.
RESEARCH ARTICLE Development 137 (18)
Fig. 3. Mutant cap mesenchyme persists andexpresses low Gdnf, but this does not account forthe blind ureter phenotype. (A-J)Hoxb7GFPexpression (A,B) or whole-mount in situ hybridization(C-J) in wild-type or Sall1-Zn homozygous mutant E12.5mouse tissues. Ret is properly tip restricted in the blindureter. Pax2 and Six2 expression reveal a cap ofmetanephric mesenchyme (mm) atop the blind ureterthat does not express Cited1 (ureter tip is marked by anasterisk in J). (K,L)Anti-Pax2 (green) and anti-laminin(red) staining show that Pax2-positive mm cells encirclethe Pax2-positive ureteric bud (ub) outlined with anti-laminin in both heterozygous and homozygous tissues atE11.5. (M,N)Gdnf expression in the mutant mm isreduced compared with wild-type mm at E11.5.(O-R)Organ culture of Hoxb7GFP;Sall1-Zn+/ orHoxb7GFP;Sall1-Zn/ tissues at E11.5 with the additionof either BSA- or GDNF-coated beads imaged at 0 and48 hours of culture. BSA-treated explants recapitulatethe in vivo phenotypes with normal branching (O, +/)and a blind ub (P, /). GDNF-treatment causes swollenbranch events (Q, +/) or a single branch that does notprogress (R, /). (S,T)Recombination of wild-type E12.5mm with E11.0 ampulla-stage ubs (S, +/) or E12.5 blindureters (T, /) show that the mutant (/) is capable ofmultiple branching events, similar to the control (+/)ampulla, when wild-type mm is present.
DEVELO
PMENT

Reduction in -catenin dosage rescues earlybranchingSince Wnt9b and at least two canonical Wnt-responsive genes wereupregulated in Sall1-Zn/ mutant ureters, we hypothesized thatub branching could be rescued by lowering -catenin activity. Incontrast to reduced Spry1 dose, which had no effect on the Sall1-Zn/ phenotype, lowering the dose of -catenin rescuedbranching in 50% of Sall1-Zn/;Ctnnb1+/– E12.5-13.5 embryos.Seven out of 14 mutant kidneys branched to the T-stage andinitiated at least a second round of branching from each tip,resulting in a statistically significant rescue of the blind ureterphenotype (Fisher’s exact test P<0.005, Fig. 4K,N,O). Sall1-Zn/;Ctnnb1+/+ littermates (21 out of 22) and the remainder ofthe Sall1-Zn/;Ctnnb1+/– kidneys arrested with blind ureters(Fig. 4J,M,O). Sall1-Zn+/+;Ctnnb1+/– control kidneys branched
normally (Fig. 4I,L), verifying that lowered -catenin dose alonehas no effect on kidney branching. This rescue supports the ideathat increased -catenin signaling is a major component of theSall1-Zn/ blind ureter phenotype.
Ectopic Wnt9b or -catenin activity in the ureterinhibits branchingTight control of -catenin is required during nephrogenesis in boththe ureter and mesenchyme (Bridgewater et al., 2008; Carroll et al.,2005; Iglesias et al., 2007; Kuure et al., 2007; Marose et al., 2008;Park et al., 2007). Therefore, we next sought to understand whetherincreased -catenin signaling in the ureter tip could be responsiblefor defective initiation of ub branching. To test this hypothesis, wecreated a new transgenic mouse to conditionally overexpressWnt9b (ACTWnt9b) and used an existing conditionally active
3103RESEARCH ARTICLESall1 affects ub Wnt signaling
Fig. 4. Blind ureters exhibit altered tip fate by ectopic Wnt9bexpression and are rescued by reducing -catenin dose.(A,B)Hoxb7GFP expression at E11.0 (+/+) in mouse ampullas and E12.5Sall1-Zn/ blind ureters. (C,D)Wnt11 is expressed similarly in the wild-type ampulla and mutant blind ureter and is absent from ureteric bud(ub) stalks (outlined). (E,F)Wnt9b expression detected by whole-mountin situ hybridization is upregulated in mutant ub tips (/) at E11.5 (notshown) and E12.5. Wild-type ub tip is outlined. (G,H) In the wild type,Axin2, a canonical Wnt signaling component and -catenin responsivegene, is more strongly expressed in the Wolffian duct and ub stalk, andis downregulated in the ub tip (outlined) similar to Wnt9b. Axin2 isupregulated in the Sall1-Zn/ mutant ub tip. (I-O)Reduction of -catenin dose rescues Sall1Zn/ blind ureter formation. RepresentativeHoxb7GFP expression (I-K) or whole-mount bright-field photos (L-N) areshown and the results are quantitated (O). Two or more rounds ofbranching were seen in 50% (seven out of 14) of E12.5 Sall1-Zn/;Ctnnb1+/– compared with 5% (one out of 22) of Sall1-Zn/;Ctnnb1+/+ kidneys. Blind ureters were observed in 50% ofSall1-Zn/;Ctnnb1+/– and 95% of Sall1-Zn/;Ctnnb1+/+ kidneys.Sall1+/+;Ctnnb1+/– kidneys (100%, 18 out of 18) had branched at leasteight times by E12.5. Statistical significance was calculated usingFisher’s exact test (P<0.0005).
Fig. 5. Ectopic Wnt9b or increased -catenin signaling in theureteric bud (ub) is sufficient to reduce branching. (A-J)Ectopic ubexpression of Wnt9b in E12.5 Hoxb7creEGFPtg/+; ACTWnt9btg/+
transgenic mice causes reduced branching and abnormal morphologyof the ureter. Four different founders are shown [founders 8 (B,J), 7 (D),3 (F) and 10 (H)]. Analysis of Pax2 (G,H) and Six2 expression (I,J) showsnormal mesenchymal cap formation in both wild-type and transgenickidneys. (K-T)GFP expression and whole-mount in situ hybridizationreveal unbranched ubs in Hoxb7creEGFP;Ctnnb1ex3fl/+ at E12.5. Thisconstitutively active -catenin allele would be expected to increase Wntsignaling in the ub tip more significantly than ectopic Wnt9b alone andproduces a more dramatic ub arrest. None of the genes assayed(Wnt9b, Wnt11, Pax2 and Sall1) was significantly altered inHoxb7creEGFP;Ctnnb1ex3fl/+ embryos, suggesting that Sall1 actsupstream of -catenin signaling.
DEVELO
PMENT

3104
dominant allele of -catenin [Ctnnb1ex3fl/+(Harada et al., 1999)].When crossed to Hoxb7creEGFP (Zhao et al., 2004), the progenywould express ectopic Wnt9b or stabilized -catenin throughoutthe Wolffian duct and ureter, including the tip. The resultingembryos were used to test the effects of overexpression ofWnt9b or canonical Wnt signaling in the ureter tip and throughoutthe ub.
For Wnt9b transgenic experiments, E12.5 kidneys wereexamined by two independent individuals who were blind tothe genotype. Thirty-two out of 43 (74%) Hoxb7creEGFPtg/+;ACTWnt9btg/+ kidneys were scored as reduced in size or asexhibiting abnormal branching morphology such as asymmetry(Fig. 5). Nine out of 31 (29%) Hoxb7creEGFPtg/+ littermatesappeared small with normal morphology and acted as controls forvariation in the extent of branching at this developmental stage.These differences were observed for at least two litters from eachof five different transgenic founders and were found to bestatistically significant (Chi Square, P<0.0005). Quantification fortwo independent founders showed a 36% decrease in number of ubbranch tips for Hoxb7creEGFPtg/+;ACTWnt9btg/+ when comparedwith littermate controls (11.2±1.2 versus 7.2±1.0 ub branch tips,t-test, P<0.05, n23 kidneys). Whole-mount in situ hybridizationanalysis of Pax2 and Six2 revealed that normal mesenchymal capswere formed in transgenic ub tips (similar to wild-type littermates)but the number of caps was reduced in proportion to the reducedub tips (Fig. 5G-J). Wnt9b-FLU was detected by western blottingof kidney lysates confirming its expression in transgenics (data notshown). Furthermore, ACTWnt9b founders that were found to havethe most dramatic reduction in branching exhibited the highesttransgene copy number (data not shown). These results show thatectopic Wnt9b expression in the ub is sufficient to alter branchingmorphogenesis.
As the effects of overexpression of Wnt9b alone were less severethan the Sall1-Zn/ blind ureter phenotype, we increased Wntsignaling more dramatically in the ub using a stabilized -cateninallele. A significant number of E11.5-12.5 Hoxb7creEGFP;Ctnnb1ex3fl/+ embryos (35 out of 60) exhibited unbranched uretersthat were remarkably similar to those seen in Sall1-Zn embryos(Fig. 5L). The remaining mutants were able to form the T-branch,but arrested shortly thereafter, and often exhibited abnormal branchpatterns (data not shown). Hoxb7creEGFP;Ctnnb1+/+ littermatesbranched normally at all stages, confirming that the transgene alonehad no deleterious effects. The high frequency of renal agenesis forthis cross has recently been reported, but no detailed analysis wasprovided (Karner et al., 2010). Similarly, inhibition of ureteric budbranching was observed in organ culture when -catenin signalingwas activated using an inhibitor of GSK-3: LiCl (n8/8, data notshown). Gene marker analysis demonstrated the presence of blindureters that properly express tip restricted Wnt11 (Fig. 5O,P).Wnt11 expression was expanded in Hoxb7creEGFP;Ctnnb1ex3fl/+
kidneys compared with Sall1-Zn mutants (compare Fig. 4D withFig. 5P) and this pattern probably indicates sites that could initiateabnormal branching later in development in these embryos. Wnt9bexpression was preserved in ub stalks and was properlydownregulated in wild-type and Ctnnb1ex3/+ mutant ub tips,demonstrating that dominant -catenin has no effect on Wnt9b geneexpression (Fig. 5M,N). Pax2 was expressed in the ureter and theoverlying mm in a very similar pattern to that seen in Sall1-Znembryos (compare Fig. 5R with Fig. 3F). Sall1 expression in themesenchyme was not altered, suggesting that its role in affectingureter branching occurs upstream of -catenin signaling. Overall,the strong correlation between the phenotypic consequences of lossof Sall1 in the mm and increased Wnt9b--catenin signaling in theub suggests that increased -catenin signaling at ub tips is animportant mechanism for the Sall1-Zn blind ureter phenotype.
DISCUSSIONSall1 has been implicated as a regulator of canonical Wntsignaling, but it is not clear whether this is a direct or indirect effect(Bohm et al., 2006; Chihara and Hayashi, 2000; Ma et al., 2006;Onai et al., 2004; Ribeiro et al., 2004; Sato et al., 2004; Uez et al.,2008). As Sall1 is a transcription factor expressed in the mm, it islikely to regulate expression of membrane bound or secretedfactors that would signal to the ub to undergo initial branching.Based on our results, Sall1 would be predicted to repress Wnt9bexpression and -catenin signaling in ureteric bud tip cells. AsSall1 acts as a potent transcriptional repressor through binding toa nucleosome remodeling and deactylase (NuRD) complex, anattractive hypothesis is that Sall1 is necessary to repress a branchinhibitor that is secreted by the mm (Kiefer et al., 2002; Lauberthand Rauchman, 2006). Whereas branch inhibitors normallyfunction to restrict ub outgrowth to a single bud at the properlocation, ectopic expression of these factors could disruptbranching. An alternative possibility is that there is a deficiency inbranch-promoting factors in the Sall1-Zn mutant. In addition toGDNF, FGFs and other growth factors are thought to positivelyregulate ub branching (Costantini, 2006). Further studies areneeded to determine which of these factors might regulate initialbranch formation by the ub via modulation of Wnt9b/-cateninactivity in ub tip cells.
Although our studies point to an important mechanism by whichSall1-dependent signals from the mm regulate the ub, it is likelythat this transcription factor also has cell-autonomous functions inthe mm. Progenitors derived from Sall1 mutant mm can be induced
RESEARCH ARTICLE Development 137 (18)
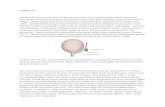
Fig. 6. Model for the effect of mesenchymal Sall1 on ureter stalkexpression of Wnt9b and canonical Wnt signaling. When theureteric bud invades the metanephric mesenchyme (pink),mesenchymal progenitor cells (light gray and blue) coalesce at theureter tip (brown). Wnt9b and -catenin signaling (yellow) aredownregulated in the ub tip, but continue to be expressed in the ubstalk. Signaling from the mesenchyme causes the ureter tips to branch.Reciprocal signals from the ureter to the mesenchyme induce themesenchyme to form condensates that differentiate into renal vesiclesnear the Wnt9b-positive stalk. When Sall1 is not expressed in themesenchyme (dark gray and blue, below the dotted line), Wnt9b isectopically expressed in ureter tips and ureter branching arrests beforethe T-stage.
DEVELO
PMENT

to differentiate into nephron precursors but form smaller coloniesthan wild-type cells, suggesting that Sall1 is required formaintenance or proliferation of metanephric progenitors (Osafuneet al., 2006). Recently, it has been suggested that Sall1 also has arole in modulating compact adhesion of mm cells adjacent to theub tip through direct regulation of the kinesin family gene Kif26(Uchiyama et al., 2010). Identification of Sall1 target genes in thekidney will better elucidate its multiple roles in the mm.
Several key studies have addressed the role of canonical Wntsignaling in the developing kidney using both loss- and gain-of-function approaches (Bridgewater et al., 2008; Carroll et al., 2005;Marose et al., 2008; Park et al., 2007). A general conclusion fromthese studies is that kidney development is extremely sensitive tothe level or duration of -catenin signaling both in the mm and inthe ub. This tight control of -catenin activity appears to be crucialat the ureteric bud tip/cap mesenchyme region in order to balancethe need to induce nephrons yet maintain the pool of progenitorcells (Park et al., 2007). Our results suggest that tight control of -catenin activity in the ub tip cells is also required to initiate normalbranching morphogenesis. Genetic reduction of -catenin (Ctnnb1)levels rescued initial branching in the Sall1-Zn mutant but failedto rescue kidney development fully. This could suggest that Sall1-Wnt signaling regulates early stages of branching but that laterstages use other pathways, as demonstrated by several recentstudies (Jain et al., 2006; Shakya et al., 2005).
We propose a model in which Wnt9b is restricted to ureter stalksto maintain undifferentiated mesenchymal progenitors and topromote subsequent ureter branches in the cortex (Fig. 6). Wnt9bexpression is confined to the ureter stalk to allow induction ofmesenchymal differentiation without affecting ub tip branchingmorphogenesis. In the absence of Sall1, this delicate balance of tipversus stalk Wnt signaling is disrupted, which leads to ubbranching arrest. Our studies reveal a requirement for Sall1 for theinitial interaction between mm and ub that modulates ureter -catenin signaling and branching competence.
AcknowledgementsWe thank Emily Royse for excellent technical assistance with genotyping, andJeannine Basta and Darcy Denner for constructive advice. Jim Kiefer providedassistance with figures and F. Costantini gave valuable criticism of the data. F.Costantini, J. Licht and Feng Chen generously provided mice, and theWashington University St Louis Developmental Biology Morphology coreprovided histology services. This work was supported by an NIH RO1 grant(R01DK067222 to M.R.), a Veterans Affairs Merit Award (M.R.) and anAmerican Heart Association Established Investigator Award (0840117N, M.R.).S.K. is supported by an American Society of Nephrology Gottschalk researchgrant and L.M. is funded by NIH R01ES014482. Deposited in PMC for releaseafter 12 months.
Competing interests statementThe authors declare no competing financial interests.
ReferencesBasson, M. A., Akbulut, S., Watson-Johnson, J., Simon, R., Carroll, T. J.,
Shakya, R., Gross, I., Martin, G. R., Lufkin, T., McMahon, A. P. et al. (2005).Sprouty1 is a critical regulator of GDNF/RET-mediated kidney induction. Dev. Cell8, 229-239.
Basson, M. A., Watson-Johnson, J., Shakya, R., Akbulut, S., Hyink, D.,Costantini, F. D., Wilson, P. D., Mason, I. J. and Licht, J. D. (2006). Branchingmorphogenesis of the ureteric epithelium during kidney development iscoordinated by the opposing functions of GDNF and Sprouty1. Dev. Biol. 299,466-477.
Bohm, J., Sustmann, C., Wilhelm, C. and Kohlhase, J. (2006). SALL4 is directlyactivated by TCF/LEF in the canonical Wnt signaling pathway. Biochem. Biophys.Res. Commun. 348, 898-907.
Bridgewater, D., Cox, B., Cain, J., Lau, A., Athaide, V., Gill, P. S., Kuure, S.,Sainio, K. and Rosenblum, N. D. (2008). Canonical WNT/beta-cateninsignaling is required for ureteric branching. Dev. Biol. 317, 83-94.
Bush, K. T., Vaughn, D. A., Li, X., Rosenfeld, M. G., Rose, D. W., Mendoza, S.A. and Nigam, S. K. (2006). Development and differentiation of the uretericbud into the ureter in the absence of a kidney collecting system. Dev. Biol. 298,571-584.
Carroll, T. J., Park, J. S., Hayashi, S., Majumdar, A. and McMahon, A. P.(2005). Wnt9b plays a central role in the regulation of mesenchymal to epithelialtransitions underlying organogenesis of the mammalian urogenital system. Dev.Cell 9, 283-292.
Caruana, G., Cullen-McEwen, L., Nelson, A. L., Kostoulias, X., Woods, K.,Gardiner, B., Davis, M. J., Taylor, D. F., Teasdale, R. D., Grimmond, S. M. etal. (2006). Spatial gene expression in the T-stage mouse metanephros. GeneExpr. Patterns 6, 807-825.
Chai, L., Yang, J., Di, C., Cui, W., Kawakami, K., Lai, R. and Ma, Y. (2006).Transcriptional activation of the SALL1 by the human SIX1 homeodomain duringkidney development. J. Biol. Chem. 281, 18918-18926.
Chen, Y. X., Krull, C. E. and Reneker, L. W. (2004). Targeted gene expression inthe chicken eye by in ovo electroporation. Mol. Vis. 10, 874-883.
Chihara, T. and Hayashi, S. (2000). Control of tracheal tubulogenesis byWingless signaling. Development 127, 4433-4442.
Costantini, F. (2006). Renal branching morphogenesis: concepts, questions, andrecent advances. Differentiation 74, 402-421.
Donovan, M. J., Natoli, T. A., Sainio, K., Amstutz, A., Jaenisch, R., Sariola, H.and Kreidberg, J. A. (1999). Initial differentiation of the metanephricmesenchyme is independent of WT1 and the ureteric bud. Dev. Genet. 24, 252-262.
Harada, N., Tamai, Y., Ishikawa, T., Sauer, B., Takaku, K., Oshima, M. andTaketo, M. M. (1999). Intestinal polyposis in mice with a dominant stablemutation of the beta-catenin gene. EMBO J. 18, 5931-5942.
Iglesias, D. M., Hueber, P. A., Chu, L., Campbell, R., Patenaude, A. M.,Dziarmaga, A. J., Quinlan, J., Mohamed, O., Dufort, D. and Goodyer, P. R.(2007). Canonical WNT signaling during kidney development. Am. J. Physiol.Renal Physiol. 293, F494-F500.
Jain, S., Encinas, M., Johnson, E. M., Jr and Milbrandt, J. (2006). Critical anddistinct roles for key RET tyrosine docking sites in renal development. Genes Dev.20, 321-333.
Jho, E. H., Zhang, T., Domon, C., Joo, C. K., Freund, J. N. and Costantini, F.(2002). Wnt/beta-catenin/Tcf signaling induces the transcription of Axin2, anegative regulator of the signaling pathway. Mol. Cell. Biol. 22, 1172-1183.
Karner, C. M., Dietrich, M. F., Johnson, E. B., Kappesser, N., Tennert, C.,Percin, F., Wollnik, B., Carroll, T. J. and Herz, J. (2010). Lrp4 regulatesinitiation of ureteric budding and is crucial for kidney formation-a mouse modelfor Cenani-Lenz syndrome. PLoS ONE 5, e10418.
Kiefer, S. M., McDill, B. W., Yang, J. and Rauchman, M. (2002). Murine Sall1represses transcription by recruiting a histone deacetylase complex. J. Biol.Chem. 277, 14869-14876.
Kiefer, S. M., Ohlemiller, K. K., Yang, J., McDill, B. W., Kohlhase, J. andRauchman, M. (2003). Expression of a truncated Sall1 transcriptional repressoris responsible for Townes-Brocks syndrome birth defects. Hum. Mol. Genet. 12,2221-2227.
Kiefer, S. M., Robbins, L., Barina, A., Zhang, Z. and Rauchman, M. (2008).SALL1 truncated protein expression in Townes-Brocks syndrome leads to ectopicexpression of downstream genes. Hum. Mutat. 29, 1133-1140.
Kuure, S., Popsueva, A., Jakobson, M., Sainio, K. and Sariola, H. (2007).Glycogen synthase kinase-3 inactivation and stabilization of beta-catenin inducenephron differentiation in isolated mouse and rat kidney mesenchymes. J. Am.Soc. Nephrol. 18, 1130-1139.
Lauberth, S. M. and Rauchman, M. (2006). A conserved 12-amino acid motif inSall1 recruits the nucleosome remodeling and deacetylase corepressor complex.J. Biol. Chem. 281, 23922-23931.
Linton, J. M., Martin, G. R. and Reichardt, L. F. (2007). The ECM proteinnephronectin promotes kidney development via integrin 81-mediatedstimulation of Gdnf expression. Development 134, 2501-2509.
Little, M. H., Brennan, J., Georgas, K., Davies, J. A., Davidson, D. R., Baldock,R. A., Beverdam, A., Bertram, J. F., Capel, B., Chiu, H. S. et al. (2007). Ahigh-resolution anatomical ontology of the developing murine genitourinarytract. Gene Expr. Patterns 7, 680-699.
Ma, Y., Cui, W., Yang, J., Qu, J., Di, C., Amin, H. M., Lai, R., Ritz, J., Krause, D.S. and Chai, L. (2006). SALL4, a novel oncogene, is constitutively expressed inhuman acute myeloid leukemia (AML) and induces AML in transgenic mice.Blood 108, 2726-2735.
Mahoney, Z. X., Sammut, B., Xavier, R. J., Cunningham, J., Go, G., Brim, K.L., Stappenbeck, T. S., Miner, J. H. and Swat, W. (2006). Discs-large homolog1 regulates smooth muscle orientation in the mouse ureter. Proc. Natl. Acad. Sci.USA 103, 19872-19877.
Majumdar, A., Vainio, S., Kispert, A., McMahon, J. and McMahon, A. P.(2003). Wnt11 and Ret/Gdnf pathways cooperate in regulating uretericbranching during metanephric kidney development. Development 130, 3175-3185.
3105RESEARCH ARTICLESall1 affects ub Wnt signaling
DEVELO
PMENT

3106
Marose, T. D., Merkel, C. E., McMahon, A. P. and Carroll, T. J. (2008). Beta-catenin is necessary to keep cells of ureteric bud/Wolffian duct epithelium in aprecursor state. Dev. Biol. 314, 112-126.
Michael, L. and Davies, J. A. (2004). Pattern and regulation of cell proliferationduring murine ureteric bud development. J. Anat. 204, 241-255.
Michos, O., Goncalves, A., Lopez-Rios, J., Tiecke, E., Naillat, F., Beier, K., Galli,A., Vainio, S. and Zeller, R. (2007). Reduction of BMP4 activity by gremlin 1enables ureteric bud outgrowth and GDNF/WNT11 feedback signalling duringkidney branching morphogenesis. Development 134, 2397-2405.
Mugford, J. W., Yu, J., Kobayashi, A. and McMahon, A. P. (2009). High-resolution gene expression analysis of the developing mouse kidney definesnovel cellular compartments within the nephron progenitor population. Dev.Biol. 333, 312-323.
Nie, X., Sun, J., Gordon, R. E., Cai, C. L. and Xu, P. X. (2010). SIX1 actssynergistically with TBX18 in mediating ureteral smooth muscle formation.Development 137, 755-765.
Nishinakamura, R., Matsumoto, Y., Nakao, K., Nakamura, K., Sato, A.,Copeland, N. G., Gilbert, D. J., Jenkins, N. A., Scully, S., Lacey, D. L. et al.(2001). Murine homolog of SALL1 is essential for ureteric bud invasion in kidneydevelopment. Development 128, 3105-3115.
Novak, A., Guo, C., Yang, W., Nagy, A. and Lobe, C. G. (2000). Z/EG, a doublereporter mouse line that expresses enhanced green fluorescent protein uponCre-mediated excision. Genesis 28, 147-155.
Onai, T., Sasai, N., Matsui, M. and Sasai, Y. (2004). Xenopus XsalF: anteriorneuroectodermal specification by attenuating cellular responsiveness to Wntsignaling. Dev. Cell 7, 95-106.
Osafune, K., Takasato, M., Kispert, A., Asashima, M. and Nishinakamura, R.(2006). Identification of multipotent progenitors in the embryonic mouse kidneyby a novel colony-forming assay. Development 133, 151-161.
Park, J. S., Valerius, M. T. and McMahon, A. P. (2007). Wnt/-catenin signalingregulates nephron induction during mouse kidney development. Development134, 2533-2539.
Ribeiro, C., Neumann, M. and Affolter, M. (2004). Genetic control of cellintercalation during tracheal morphogenesis in Drosophila. Curr. Biol. 14, 2197-2207.
Sato, A., Kishida, S., Tanaka, T., Kikuchi, A., Kodama, T., Asashima, M. andNishinakamura, R. (2004). Sall1, a causative gene for Townes-Brocks
syndrome, enhances the canonical Wnt signaling by localizing toheterochromatin. Biochem. Biophys. Res. Commun. 319, 103-113.
Schedl, A. (2007). Renal abnormalities and their developmental origin. Nat. Rev.Genet. 8, 791-802.
Schmidt-Ott, K. M., Yang, J., Chen, X., Wang, H., Paragas, N., Mori, K., Li, J.Y., Lu, B., Costantini, F., Schiffer, M. et al. (2005). Novel regulators of kidneydevelopment from the tips of the ureteric bud. J. Am. Soc. Nephrol. 16, 1993-2002.
Schmidt-Ott, K. M., Masckauchan, T. N., Chen, X., Hirsh, B. J., Sarkar, A.,Yang, J., Paragas, N., Wallace, V. A., Dufort, D., Pavlidis, P. et al. (2007). -catenin/TCF/Lef controls a differentiation-associated transcriptional program inrenal epithelial progenitors. Development 134, 3177-3190.
Schuchardt, A., D’Agati, V., Larsson-Blomberg, L., Costantini, F. and Pachnis,V. (1995). RET-deficient mice: an animal model for Hirschsprung’s disease andrenal agenesis. J. Intern. Med. 238, 327-332.
Schuchardt, A., D’Agati, V., Pachnis, V. and Costantini, F. (1996). Renalagenesis and hypodysplasia in ret-k- mutant mice result from defects in uretericbud development. Development 122, 1919-1929.
Shakya, R., Watanabe, T. and Costantini, F. (2005). The role of GDNF/Retsignaling in ureteric bud cell fate and branching morphogenesis. Dev. Cell 8, 65-74.
Srinivas, S., Wu, Z., Chen, C. M., D’Agati, V. and Costantini, F. (1999).Dominant effects of RET receptor misexpression and ligand-independent RETsignaling on ureteric bud development. Development 126, 1375-1386.
Sweeney, D., Lindstrom, N. and Davies, J. A. (2008). Developmental plasticityand regenerative capacity in the renal ureteric bud/collecting duct system.Development 135, 2505-2510.
Uchiyama, Y., Sakaguchi, M., Terabayashi, T., Inenaga, T., Inoue, S.,Kobayashi, C., Oshima, N., Kiyonari, H., Nakagata, N., Sato, Y. et al.(2010). Kif26b, a kinesin family gene, regulates adhesion of the embryonickidney mesenchyme. Proc. Natl. Acad. Sci. USA 107, 9240-9245.
Uez, N., Lickert, H., Kohlhase, J., de Angelis, M. H., Kuhn, R., Wurst, W. andFloss, T. (2008). Sall4 isoforms act during proximal-distal and anterior-posterioraxis formation in the mouse embryo. Genesis 46, 463-477.
Zhao, H., Kegg, H., Grady, S., Truong, H. T., Robinson, M. L., Baum, M. andBates, C. M. (2004). Role of fibroblast growth factor receptors 1 and 2 in theureteric bud. Dev. Biol. 276, 403-415.
RESEARCH ARTICLE Development 137 (18)
DEVELO
PMENT