
Le Corps. Le bras Le dos le ventre La tête La jambe Le corps.

Sissejuhatus eukarüootide mitmekesisusse - protistid ja seenedBiodiversiteet ja elustik Eestis ja Euroopas
Veekogude elustikAlgoloogia
Protistide osa konspekt
MegasüstemaatikaKalle Olli1
KokkuvõteTänapäeva megasüstemaatiikas moodustavad loomad(Metazoa), kitsas mõistes taimed (maismaataimed) ja seened euka-rüootide evolutsioonipuu kolm külgharu.Eukarüoote klassifitseeritakse kuude fülogeneetilisse klaadi:
Unikonta ehk ühevibursed. Siia kuuluvad Amoebozoa, Apusozoa, Ophistokonta. VIimane jaotub kaheks, Holozoaja Holomycota, millesse kuuluvad vastavalt ka hulkraksed loomad (Metazoa) ja seened (Opisthosporidia).
Excavata ehk ekskavaadid. Siia kuuluvad Metamonadida (Diplomonada, Retortomonadida, Parabasalia, Oxymo-nadida) ja Discoba (Euglenozoa, Heterolobosea, Jakobida).
Archeplastida ehk arheplastiidsed — primaarsete plastiididega organismid: Glacucophyta (liitvetikad), Rhodophyta(punavetikad) ja Viridiplantae (rohevetikad ja streptofüüdid, sh. maismaataimed).
Rhizaria ehk risaarid on lähedased kromalveolaatidele. Siia kuulub palju amööbe, nagu Cercozoa (Filosa, Endomyxa)ja Retaria (Polycystinea, Acantharia, Foraminifera ehk kambrilised).
Chromalveolata ehk kromalveolaadid — Alveolata ehk alveolaadid (Ciliophora — ripsloomad; Dinophyta — vaguvi-
burlased; Chromerida, Apicomplexa — tippeoslased) ja stramenopiilid (Bigyra, pseudoseened, Ochrophyta).
Harcrobia — pisut ebaselge kuuluvusega; Haptophyta — haptofüüdid ja Cryptophyta — neelvetikad.
Risaarid ja kromalveolaadid on lähedased ja moodsutavad koos SAR-rühma, sõnadest Stramenopiilid–Alveolaadid–Risaarid.1EMU; UT
Sisukord
Megasüstemaatika 2Kes on taimed? . . . . . . . . . . . . . . . . . . . . 3Kes on loomad? . . . . . . . . . . . . . . . . . . . . 3Kes on seened? . . . . . . . . . . . . . . . . . . . . 4
Elu puu ajalooline ja filosoofiline taust 5Dihhotoomia: taimed — loomad . . . . . . . . . . . 5
Aristoteles . . . . . . . . . . . . . . . . . . . . 5Leeuwenhoek . . . . . . . . . . . . . . . . . . . . . 5Linnaeus . . . . . . . . . . . . . . . . . . . . . . . . 6Evolutsioon . . . . . . . . . . . . . . . . . . . . . . 6Ernst Haeckel . . . . . . . . . . . . . . . . . . . . . 7Dihhotoomia: eukarüodoodid — prokarüoodid . . . . 8
Bakterite kontseptsioon . . . . . . . . . . . . . 8
Uus paradigma: eluslooduse kolm domeeni 9
Endosümbioosi teooria 10Mitokondri omastamine . . . . . . . . . . . . . . . .11
Mitokondri sekundaarne kadumine . . . . . . .11
Arhezoa hüpotees . . . . . . . . . . . . . . . .11Mitokondri ehitus . . . . . . . . . . . . . . . .11Vaba hapniku roll eukarüootide mitmekesi-
suse kujunemisel . . . . . . . . . . . .12Plastiidi omastamine . . . . . . . . . . . . . . . . .13
Plastiidide primaarne endosümbioos . . . . . .13Arheplastiidsed — kolm haru primaarsete
plastiididega organisme . . . . . . . .13Sekundaarsed endosümbioosid . . . . . . . . .14Nukleomorfiga organismid . . . . . . . . . . .14Nukleomorf on kadunud . . . . . . . . . . . .14Tertsiaarsed endosümbioosid . . . . . . . . . .14Plastiidi eellane . . . . . . . . . . . . . . . . .15Plastiidi genoomi redutseeritus . . . . . . . . .15Plastiidide värvikirevus . . . . . . . . . . . . .17
Eukarüoodid 18
Kiire lõppsõna 19

MegasüstemaatikaOrganisme võib klassifitseerida mitmel viisil. Ajalooliselton süstemaatika pürginud ’loomuliku klassifikatsiooni’ poo-le, eeldades, et organismid moodustavad loodusliku süstee-mi ja teadlaste asi on seda mõista.Organisme võib klassifitseerida näiteks elupaiga järgi (vees,
maismaal), mikro- ja makroorganismideks, morfoloogilisevõi geneetilise sarnasuse alusel, toitumise järgi (fototroofid,heterotroofid). Nii jagunevad organismid näiteks produtsen-tideks ehk tootjateks, konsumentideks ehk tarbijateks ja re-dutsentideks ehk lagundajateks:
Produtsendid ehk tootjad —autotroofsed organismid kessünteesivad orgaanilist ainet. On ökosüsteemides toiduahe-la esimesteks lülideks.
Konsumendid ehk tarbijad — (Lad.: consumere— tarbi-ma) heterotroofsed organismid, kes otseselt või kaudselt(teiste tarbijate kaudu) toituvad muudest elusatest või sur-nud organismidest. Toiduahelas moodustavad konsumen-did teise lüli ja sellele järgnevad lülid.
Redutsendid ehk lagundajad — hetertotroofsed organis-mid, kes lagundavad surnud orgaanilist ainet. Näiteks bak-terid, seened.
Joonis 1.Molekulaarsetel andmetel põhinev fülogeneetilineklassifikatsioon näitab organismide sugulust. Osaliselt ei langesee kokku intuitiivsete ootustega — krokdillid on lähemassuguluses lindude kui teiste roomajatega; vetikad ei moodustaühtset rühma, siin näiteks rohevetikad (äärmine parempoolne) eirühmitu kokku koldvetikategha (keskmine parempoolne). Üllatavon fototroofsete ja miksotroofsete koldvetikate (siin Dinobryon)ühte rühmitamine heterotroofsetega (siin Spumella).
Joonis 2. Klassifitseerimine üldiste morfoloogiliste tunnustealusel. Krokodillid on koos roomajatega kuna neil on sarnaneehitus, elustiil ja paljud tunnused (soomustega kaetud keha,amfiibne eluviis).
Joonis 3. Klassifitseerimine ökoloogilisete ja funktsionaalsetetunnuste alusesl. Kõnekeelselt mõistetakse taimede allfotoautotroofselt toituvaid organisme. Loomad on kõnekeelseltteistest heterotroofselt toituvad ja vabalt ringiliikuvad organismid.Siin on vetikad rühitatud kokku primaarprodutsentideks, erialdiheterotroofidest. Kõigusoojased krokodillid on koos teisteroomajatega.
Eelistatuim viis organisme klassifitseerida on fülogenee-si1 järgi.
Üldjuhul — fülogeneetiliselt lähedased organismid on
omavahes suuremas sarnasuses kui fülogeneetilisetl kau-
ged organismid.
Sõltuvalt rõhuasetusest on ka teistsugused klassifitseeri-mise viisid mõistlikud. Kõige olulisem on mitte omavahelsegi ajada erinevaid klassifitseerimise aluseid. Näiteks aja-des segi toitumistüübi (autotroof vs heterotroof) ja füloge-neesi on varem või hiljem tulemuseks suuremat sorti sega-dus.Mitmed toitumistüübil või üldisel sarnasuses põhinevad
klassifikatsioonisüsteemid loodi heas usus, et need peegel-davad fülogeneesi. Hiljem, lähtudes ultrastruktuursetest võimolekulaarsetest tunnustest on osutunud aga polü- või para-füleetilisteks.Klassikalised näited on krokodillide arvamine roomajate
hulka, protistide klassifitseerimine vetikateks ja alglooma-deks, või isegi taimede ja loomade eristamine.Krokodillid on fülogeneetiliselt lähemalt seotud lindude-
ga kui roomajatega — sellest hoolimata klassifitseeritakseneid koos roomajatega kuna neil on roomajatega sarnane lä-hedane ning palju morfoloogilisi ja füsioloogilisi sarnasusi(joon. 1, 2).Loomad (Metazoa) ja maismaataimed on selgelt piirit-
letud monofüleetilised rühmad. Ajalooliselt ja kõnekeelselton taime- ja loomariiki eristatud toitumistüübi alusel — vas-tavalt fototroofne ja heterotroofne. Vetikate, sh sinivetikateliigitamine koos taimedega primaarprodutsentide rühma onmõistlik ja kasulik isegi tänapäeva ökoloogiliste teadmistepõhjal (joon. 3). Samas fülogeneesi ja mitme teise teadusha-ru perspektiivis on selline rühmitaine täiesti arulage.
1õpetus organismide sugulussuhetest ja evolutsioonist läb Maa ajaloo.

Kes on taimed?Intuitiivsed tunnused, mis iseloomustavad taimi: nad on ro-helised, fotosünteesivad ja juurtegamaapinnas (joon. 4). Tai-mi isoloomustavad eht, vars ja juuri. Kõigil taimedel ei olekõiki neid tunnuseid. Parasiitsed taimed ei ole ei rohelisedega fotosünteesivad, on putuktoidulisi taimi (joon. 5). Samb-lad ei ole juurtega maapinnas kinni. Fotosünteesiv mere tiguElysia ei ole taim.Vetikad loetakse sageli taimedeks, kuna nad fotosüntee-
sivad. Ometi on vetikad morfoloogiliselt erinevad ja vaidvähesed on fülogeneetiliselt taimedega lähedased. Siniveti-kad ehk tsüanobakterid rühmitatakse sageli koos taimedega,kuna neid ühendab fotosüntees (joon. 6). MIda arvata vabaltringi ujuvast silmviburlaseest Euglena? On see taim, loom,mõlemad või ei kumbki?
Taimi saab defineerida mitmel viisil, mida kõnekeelselt
ka tehakse, põhimides funktsiooni (fotosüntees), öko-
loogiat, morfoloogiat (juur, vars, leht) ja fülogeneesi.
Fülogeneetiliselt onmaismaataimedmonofüleetiline rühm,kuhu kuuluvad samblad, sõnajalad, seemnetaimed — selli-ne definitsioon sobitub kõige paremini taimede olemusega.Funktsioonil (fotosünteesil) põhinev taimede määratlus hõl-mab ka vetikaid ja sõltuvalt käsitlusest ja sinivetikaid.Linnaeus Systema Naturae jagas looduse kolme riiki: La-
pides—kiviriik,Plantae—taimeriik,Animalia—looma-riik. Seened, samblikud, eukarüootsed vetikad ja sinivetikadkuuluside ajalooliselt taimede alla, lähtudes fotosünteesistvõi seente puhul — viljakeha kasvuvormil. Bakterid olidsamuti taimede all, Schizophyta, lähtuvalt rakuseina ole-masolust.Rangelt fülogeneetilisest printsiibist lähtuvalt saab kok-
kuleppeliselt taimedeks lugeda mitmeid eri mahuga mono-füleetilisi rühmi:
• Maismaataimed (eelistatuim määratlus)• Streptophyta—maismaataimed ja mändvetikate haru• Viridiplantae—maismaataimed ja kõik rohevetikad• Archaeplastida—primaarsete plastiididega organismid,Viridiplantae + Rhodophyta +GlaucophytaErinevalt taimedest, mida praegusajal käsitletakse enamas-
ti maismaataimede monofüleetilise taksoni kontekstis, onvetikad puhtalt funktsionaalselt ja ökoloogiliselt määratle-tud termin. Kõige üldisemalt defineeritakse vetikaid kui fo-tosünteetilisi organisme2, mis ei ole maismaataimed. See vä-ga üldine ja negatiivne definitsioon hõlmab ka sinivetikaid,kuigi vetikate all mõeldakse praegu enamasti eukarüootseidorganisme. Ökoloogilises ja funktsionaalses kontekstis pan-
2Fotosüntees kitsamas, oksügeense fotosüntessi mõistes, kus elektron-doonoriks on vesi ja eraldub vaba hapnik. Bakteriklorofüll ja aneroobnefotosüntees ei lähe arvesse.
Joonis 4. Fotosünteesivad maismaataimed kvalifitseeruvad alatitaimedeks. Neil on kõik kõnekeelsete taimede omadused — nadei liigu, nad on fotoautotroofsed.
Joonis 5.Mitte kõigil maismaataimedel ei ole kõiki taimedeiseloomulikke omadusi. Parasiitsed taimed toituvadheterotroofselt (vasakult: harilik seenlillMonotropa hypopitys,soomukas Orobanche haenseleri. Putuktoidulised taimed saavadtoitaineid loomadest, kärbespüünis Dionaea muscipula. LemmelLemna ujub veepinnal, ega ole kinnitunud juurtega pinnasesse.
nakse fotosünteetilised primaarprodutsendid enamasti kok-ku ja ei ole põhjust eristada sinivetikaid eukarüootsetest ve-tikatest.
Kes on loomad?Looma mõiste on väga intuitiivne ja seostub eriti hästi selg-roogsetega — imetajad, linnud, roomajad, kahepaiksed, ka-lad — ja ka selgrootutega — lülijalgsed, limused, etc. —kõik on kindlalt loomad (joon. 7). Sessiilsete loomade jamikroorganismidega on intuitiivsust vähe. Käsnad, korallid,meriroosid on sessiilsed — ei rutta ringi (joon. 8). Seda eba-kindlust kajastab ajalooline taksoni nimetus Anthozoa —
Joonis 6. Vetikad, sh sinivetikad on fotoautotroofsed nagutaimed. Enamik vetikaid ei ole aga fülogeneetiliselt kuigivõrdmaismaataimedega lähedased. Pruunvetikad (vasakult: põisadru(Fucus) ja ränivetikad (Porosira glacialis) kuuluvadstramenopiilide hulka. Sinivetikad (Anabaena) on bakterid jaenamasti ei nimetata neid vetikateks, kuigi ökoloogiliselt täidavadnad sama funktsiooni kui eukarüootsed vetikad.
3MEGASÜSTEMAATIKA

Joonis 7. Käibekeelne ’loomad’ sobitub hästi ’kõrgemateloomade’ kohta, kes liiguvad aktiivselt ja söövad partiklilist toitu.Siia kuulub suurem osaMetazoa taksonist, eriti ilmekalt suuredloomad.
Joonis 8.Mitmed loomad ja heterotroofsed protistid suudavadenergiat saada sümbiontsete vetikate abil (korallid, zooksantellaja zooklorella) või plastiidide abil, mis on omastatud allaneelatudtoidust (kelptokloroplastid) (ripsloom Paramecium bursaria,mere tigu Elysia chlorotica).
õisloomad.Ainuraksete eukarüootide, nagu ripsloomad (Ciliophora)
või silmvibursed (Euglenozoa) liigitamine on olnud sega-dusttekitav, nagu nimedestki selgub (joon. 9). Nagu taimedepuhul on ka loomade määratlemine olnud mitmetähendus-lik, sõltuvalt liigitamise alusest — kas see on ökoloogilinefunktsioon (liikumine, toitumine) või fülogenees.Kaks põhilist klassikalist loomadega seostuvat tunnust on
vaba liikumine ja taju, mis hiljem asendus partiklilise toidutarbimisega, kuna seda on lihtsam tõestada ja mõõta. Sellinelai määratlus paneb paljud protistid loomade alla. Partiklilk-se toidu omastamine on omane lisaks paljudele vetikatele.Fülogeneetiliselt on loomade kõige parem vasteMetazoa.Rääkides loomadest on seega oluline mõista anda, kas
peetakse silmas süstemaatilist fülogeneetilist mõistet või öko-loogilist funktsionaalset. Kumbki ei ole teisest parem või õi-gem, kuid käibekeeles ei pruugi me koheselt aru saada, mil-lest jutt ja tekib segadus. Selgroogsed ja ’kõrgemad loomad’sobituvad hästi mõlema käsitluse alla; ’alamad loomad’ japrotistid võivad tekitada segadust.Süstemaatikamõttes on loomadMetazoa; ökoloogias või-
dakse lugeda loomadeks kõiki heterotroofseid ja fagotroof-seid organisme kui ühte funktsionaalset üksust.
Protistide ’loomne’ pool võetakse ühte ökoloogilise ter-
miniga algloomad; ’taime’ pool ökoloogilise terminiga ve-
tikad.
Joonis 9. Ripsloomad ja amööbid on heterotroofsed, toiduvadbakteritest ja väiksematest protistidest. Kuid ei olefülogeneetiliselt seotud loomdegaMetazoa mõistes. Ripsloomad(vasakult: kingloom Paramecium
kuuluvad Alveolata rühma. Amööbe leidub vägaerinevates eukarüootide harudes. Amööb Saccamoeba
limax) kuulub Amoebozoa hulka.
Kes on seened?Seened ei ole erand — sarnaselt taimedele ja loomadele onka seente puhul käibekeeles segi fülogeneetiline ja ökoloo-giline käsitlus. Sessiilselt eluviisi ja rakuseinte tõttu olid see-ned algselt botaanika all, kuid tõsteti peatselt omaette riikilähtuvalt unikaalsest toitumisest.Seened on kemoorganotroofsed (s.t. heterotroofsed), ena-
masti kas saprotroofsed — toitudes surnud orgaanilisest ai-nest või parasiitsed. Fülogeneetiliselt moodustavad seenedHolomycota rühma, mis on Holozoa sõsar-rühmaks. See-ni (sensu Holomycota) iseloomustab kitiinist, harvem tsel-luloosist või glukaanist rakukest, hõlmab järgmisi rühmi:Nuc-leariidae,Microsporidia,Chytridiomycota,Glomeromyco-ta (varem tuntud kui Zygomycota),Ascomycota ja Basi-diomycota.Funktsionaalselt on seentega sarnased mitmed fülegenee-
tiliselt kaugel olevad rühmad, nagu limaseened ehk lima-kud, munasseened (Oomycetes) ja kiirikseened ehk ank-tinomütseedid (Actinomycetales).Limakud kuuluvad fülogeneetiliselt Amoebozoa klaadi,
neil on keerukas elutsükkel kus vahelduvad ainurakne amöö-bi faas ja hulgatuumaline (plasmodiaalneI) või hulkraknefaas mis moodustab viljakeha.Munasseened (Oomycetes) on tsellulosist rakuseinaga
ja sarnaselt seentegamoodustavad hüüfe, kuid kuuluvad stra-menopiilide hulka. Nende zoospooridel on spetsiifilised kol-meharuliste karvakestega eripikkused viburid, mis on stra-menopiilidemorfoloogiline sünapomorf. Paljud neist onma-janduslikult tähtsad, olles haiguste tekitajad põllukultuuri-del ja kaladel.Kiirikseened ehk anktinomütseedid (Actinomycetales)
on aeroobsed bakterid (Actinobakteria), mis moodustavadvihmaga spoore. Hüüfid moodustavad põhimkuid ja harune-nud võrgustikke mis oli ka algseks põhjuseks nende seen-teks pidamisel.Munasseened, kiirikseened ja limaseened on seega käi-
bekeelsed mõisted, mis viitavad nende organismide seene
4MEGASÜSTEMAATIKA

Joonis 10. Seened on osmotroofsed — omastavad keskkonnastlahustunud madalmolekulaarseid orgaanilisi aineid. Palja silmaganäeme eelkõige pinnasest väljas olevaid viljakehi. Vasakult:harilik kukeseen Cantharellus cibarius, vääveltorik Laetiporussulphureus.
Joonis 11. Limakud toituvad osmotroofselt ja saprotroofselt,moodustades viljakehi (vasakul) ja hulgatuumalisi plasmoodiume(paremal). Füloteneetiliselt kuuluvad Amoebozoa hulka ja onseentest üsna kaugel.
sarnasele eluviisile ja morfoloogiale, kuid fülogeneetiliselton eksitavad.
Elu puu ajalooline ja filosoofilinetaust’Elu Puu’ all tuleb mõista elusorganismide genealoogia kõi-ge üldisemat, suurestruktuurilisemat arhitektuuri.
Dihhotoomia: taimed — loomadAjalooliselt on eristatud eluta ja elusat loodust. Antiikajastkuni kahekümnenda sajandini jaotas inimene elusloodust kah-
Joonis 12. Paljud taimepatogeenid kuuluvad stramenopiilidehulka. Näiteks ebajahukastelised (Peronosporaceae) naguvasakul kurgi lehel haigust põhjustav Pseudoperonosporacubensis, kalade haigusi põhjustav vesihallik (Saprolegnia).Kiirikseened (Actinobakteria) kuuluvad bakterite hulka.
te suurde rühma— taimed ja loomad. Need ajaloolised klas-sifikatsiooni põhialused on kujundanud ka ülikoolide struk-tuuri, kus loodusteadused jagunesid geoteadusteks (eluta loo-dus), zooloogiaks (loomad), botaanikaks (taimed) ja medit-siiniks (inimene). Seejuures defineeriti botaanikat pikka ae-ga kui teadust kõigest elavast mis ei kuulu loomade alla.Selle peegeldusena õpetati ülikoolides ka seeni ja baktereidbotaanika osakondades. Eluslooduse taimseks ja loomsekspooleks jaotamises nähti eluslooduse suurimat hiaatust.
Aristoteles
Sellise kategoriseerimise ajalooline taust ulatub Kreeka filo-soofiani (Aristoteles ja tema õpilane Theophrastus) ja kret-sionismini. Loomade olulisim tunnus oli liikumine ja meel-tega tajumine,mis oli vajalik saagi äratundmisel ja tabamisel.Toitumine ja liikumine on olulised kriteeriumid, kuidas
näiteks laps tajub loomade ja taimede erisust, tekitades se-gadust kui ette satuvad korallid, käsnad, seened ja ka ... mik-roorganismid, nagu protistid.Aristotelese järgi iseloomustab elusorganisme võime toi-
tude ja paljuneda (anima vegetativa). Ainult loomad, sh ini-mene, suudavad tajuda (anima sensitiva). Mõtlemine, järel-damine ja intellekt (anima rationalis) on unikaalne inimes-tele.
LeeuwenhoekMikroorganismid, kuhu kuuluvad arhed, bakterid ja ainurak-sed protistid on ajaloolisse klassifikatsiooni halvasti sobitu-vad. Mikroorganismide klassifitseerimisel on olulised teh-noloogia jametoodikate areng. Eelkõigemikroskoopia areng(nomarski kontrast, elektronmikroskoopia) jamolekulaarsedtunnused.Mikroobide3 tundmaõppimine algas mikroskoobi leiuta-
misega Antonie van Leeuwenhoek’i poolt (joon. 13). Nähtutklassifitseeris Leeuwenhoek kui väikeseid loomi (animalcu-les, joon 14), lähtuvalt nende vabast liikumisest vees.Leeuwenhoek oli esimene, kes detailselt kirjeldas pro- ja
eukarüootsete mikroobide morfoloogiat. Leeuwenhoek nägika ainurakse organelle, pidades neid seede- ja paljunemisor-ganiteks, nagu on loomadel. Arusaam ainuraksete olemusestparanes alles 19 sajandil seoses rakuteooriaga.Kokkuvõttes peeti mikroobe lihtsateks organismideks, kes
paigutati teime ja loomariigi alumistele harudele.
Kuna paljudel mikroobidel on nii taimedele (plastiidid,
pigmendid) kui loomadele (liikuvus, toidu neelamine)
omadusi, kuulusid nad nii botaanilisse kui zooloogilisse
klassifikatsiooni süsteemi.
3mikroobid ehkmikroorganismid on need, keda me palja silmaga ei näe.Nende nägemiseks on vaja mikroskoopi.
5ELU PUU AJALOOLINE JA FILOSOOFILINE TAUST

Joonis 13. Antonie van Leewenhoek (1632–1723) oli Hollandiriidekaupmees, kes avastas lihtsa viisi kuidas suure lahutusegaoptilisi läätsesid toota. Ta disainis ja ehitas oma mikroskoobi.Väidetavalt tegi ta üle 500 läätse ja ehitas kokku üle 200mikroskoobi, millest võimsaimad suurendasid kuni 500x.Mikroskoop ise oli väike, ca 5 cm. Leewenhoek ei avaldanudkunagi oma läätsede tegemise saladust.
Selline klassifikatsioon on segadustekitav ja tänaseni la-hendamata.
LinnaeusCarl Linnaeus (1707–1778) oli Rootsi loodusteadlane, kesavaldas oma põhiteose Systema Naturae, milles andis üle-vaate kõigist tollel ajal tuntud looma- ja taimeliikidest (joon.15). Ta kirjeldas ca 7,700 taime, 6,200 looma ja 500mineraa-li. Linnaeus arvas, et ta on kirjeldanud kogu eluslooduse mit-mekesisuse, kuid reaalselt moodustas see vaid 1–2% praegu-seks tuntud liikidest. Kuigi mikroobide olemasolu oli tol ajaljuba teada, ei käsitlenud Linneaus neid eraldi, vaid nimetasneid ühiselt Chais infusorium. Linnaeus tajus hästi oma tööolulisust ning tavatses väljenduda—Deus creavit, Linnaeusdisposuit — Jumal lõi Linnaeus korrastas.Linnaeuse suurim teene ei ole mitte organismide kirjel-
dused, vaid binaarse nomenklatuuri kasutuselevõtus. Vare-malt olid liikide nimed pikad kirjeldavad fraasid4. Linnaeusnimetas iga liiki perekonna nime ja liigiepiteediga ning lõiühtse hierarhilise nomenklatuuri kõigi elusorganismide ni-metamiseks, mis on kasutusel tänaseni. Linnaeuse põhitöödon Systema Naturae (1735) ja Species Plantarum (1753),mis panid aluse vastavalt loomade ja taimede nimetamisesüsteemidele ehk zooloogilisele ja botaanilisele nomenkla-tuurile.
Nomenklatuuri alused olid juba algselt kahes eri Linna-
euse töös, mis osaliselt seletab botaanilise ja zooloogi-
4Näiteks Physalis angulata — tiivuline füüsal, nimetus oli varasemaltPhysalis annua ramosissima, ramis angulosis glabris, foliis dentato-serratis. Nõme ju?
Joonis 14. Leewenhoek’i mikroskoop ja sellega nähtudmikroobid. Leeuwenhoek oli kirjavhehetuses LondoniKuningliku Seltsiga, kes kuigi algselt skeptiline, oli vaimustusestema tööst ning valis ta 1680 oma liikmeks.
lise nomenklatuuri erinevust ja üksteisest sõltumatust.
Eriti häiriv on see pärand protistide puhul, kus mingi liigi
nimetus võib olla legaalne ühe nomenklatuuri järgi, kuid
illegaalne teise järgi.
Systema Naturae kümnendas köites klassifitseerib Linna-eus kolme peamist klaadi: kivimid, taimed ja loomad:
• Lapides corpora congesta, nec viva, nec sentientia (kivid;tahked kehad, ei elus ega tajuta.
• Vegetabilia corpora organisata& viva, non sentientia (tai-med: organiseeritud keha ja elus, tajuga.
• Animalia corpora organisata, viva et sentientia (loomad:organiseeritud keha, elus ja tajuga, liiguvad ise.
Needmegasüstemaatilised kirjeldused on pärand tolleaeg-sest klassifikatsioonist, aristotellikust filosoofiast ja teoloo-giast. Erinevalt oma kaasaegsetest paigutas Linnaeus inim-ese loomariiki (esimest korda pärast Aristotelese Historiaanimalium). Inimese (Homo sapiens) tüüpeksemplar5 onLin-naeus ise. Iseenda tajumine oli Linnaeuse omane ainult ini-mesele6.
EvolutsioonEvolutsiooni ja kõige elava ühise eellase idee pärineb antiik-ajast. Sokratese eelne Kreeka filosoof Anaximander (610–547 ema) arutles, et inimene põlvneb teistest organismidest,
5Bioloogias on tüüpeksemplar eriline isend, mille järgi on tehtud lii-gikirjeldus. Tüüpeksemplare säilitatakse muuseumites ja kogudes erilisehoolega.
6See ei pea tänapäeval enam paika. Iseenda tajumist omistatakse kamitmetele loomadele, delfiinid, elevandid, võimalik et ka mitmed linnud.
6ELU PUU AJALOOLINE JA FILOSOOFILINE TAUST

Joonis 15. Carl Linnaeus ja tema tähtteoste tiitellehed: SystemaNaturae ja Species plantarum. Systema Naturae esimene trükkilmus 1735, kuid alles kümnes väljaanne (1758) sisaldas loomadebinaarset nomenklatuuri.
et elu tekkis vees ja et elu tekkis elutust materjalist.Lamarck. 19 saj esitles Prantsuse bioloog Jean-Baptiste deLamarck (1744–1820) evolutsiooniteooriat, milles organis-mid pärandasid elu jooksul omandatud tunnuseid järglastele.Liikide järkjärgulist muutumist võeti kui fakti, kuid sellemuutumise mehhanism ja põhus olid ebaselged. Lamarckpostuleeris, et keskkonna järkjärgulinemuutus põhjustasmuu-tusi organismide eluviisis, mis omakorda põhjustas organismi-de endi muutumise. Elu jooksul tekkinud muutuste pärandu-mist uurib tänapäeval epigeneetika.Lamarck postuleeris, et kõigele elusale sisemiselt oma-
se täiustusmiskalduvus tingib evolutsiooni ja organismidejärjest keerukamaks muutumise. Elu iseeneslik tekkimineon pidev ja jätkuv protsess, mis ühtasi seletab lihtsamateorganismide olemasolu.Lamarck’i tööde tuules esitasid Alfred Russel Wallace ja
Charles Darwin 1858 teise evolutsiooniteooria, millel põhi-neb ka praegune arusaam organismide arengust (joon. 16).Erinevalt Lamarck’i täiustusmiskalduvusest, pakkusid Wal-lace ja Darwin välja loodusliku valiku, kui evolutsiooni meh-hanismi. Uus teooria ilmus Darwini raamatus Liikide tekki-mine, 1859 a.Selleks ajaks oli just ilmunud rakuteooria, mis väitis et
kõik organismid koosnevad rakkudest. See toetas evolutsioo-niteooriat, et nii taimed kui loomad, kes mõlemad koosne-vad rakkudest, põlvnevad ühisest rakulisest eellasest ningevolutsiooniteooria on üldistatav kõigi elusorganismide koh-ta.Darwini ideid mõjutas majandusteadlase ja demofgraafi
ThomasRobertMalthus’i (1766–1834)Essee rahvastiku print-siibist (AnEssay on the Principle of Population, 1798).Malt-hus kirjeldas populatsiooni kasvu väliste piirangute puudu-des eksponentsiaalsena, samas kui toidu tootmine oli lineaar-ne. Tulemusena saab populatsiooni eksponentsiaalne kasvolla ainult lühiajaline ja seejärel hakkavad populatsiooni isen-did üksteisega konkureerima piiratud ressursside pärast. Seedeograafiline–sotsiaalteaduslik nägemus oli kesksel kohalDarwini teoorias.Umbes samal ajal näitas Pasteur eksperimentaalselt, et elu
Joonis 16. Charles Darwin (1809–1882) — loodusteadlane jaevolutsiooniteooria autor. Liikide muutumine ei olnud Darwiniajaks enam uus teema. Darwin oli esimene, kes kogus erivaldkondadest massiliselt tõendusmaterjali evolutsiooniteooriakinnituseks. Vasakul: Darwini 1837 a päevik, mis visandab EluPuu põhiolemuse.
Joonis 17. Louis Pasteur (1822–1896) — prantsuse keemik jamikrobioloog. Tema panus on elu iseenesliku tekke, ehkabiogeneesi teooria ümberlükkamises. Enne seda arvati, etmikroobid tekivad iseeneslikult eluta ainest nagu vesi, muda võimuld. Enamgi, ka kõrgamad organismid nagu loomad (vaglad,ussid) tekivad iseeneslikult. Pasteur väitis, et steriilseskeskkonnas ei teki elu iseeneslikult, vaid ainult pärastkontaminatsiooni mitte-steriilse materjaliga.
ei teki eluta ainest (joon. 17). Varem arvati, et elu madala-mad vormid võivad sobivas keskkonnas (vesi, muda, muld)mistahes ajal iseeneslikult tekkida. Pasteur’i eksperimendidnäitasid, et elu iseeneslik tekkimine ei ole võimalik ning etainult elu sigitab elu. Pasteur väitis, et varasemad näited eluiseeneslikust tekkest olid põhjustatud kontaminatsioonist —mikroobid levivad nii vee kui õhu kaudu.Pasteur’i argumendid üldistasid Darwini teooriat ka alge-
lisematele organismidele ja toetasid ideed kõigi organismi-de sugulusest ning põlvnemisest ühest ühisest eellasest.Seega vaid mõne aasta vältel muutus põhiarusaam bioloo-
giast väga kardinaalselt.
Ernst HaeckelErnst Heinrich Philipp August Haeckel (1834–1919) oli sak-sa loodusteadlane, kunstnik ja arstiteadlane (joon. 18). Tema
7ELU PUU AJALOOLINE JA FILOSOOFILINE TAUST

Joonis 18. Ernst Haeckel. Allikas.
elutöö on mikroorganismide, peamiselt mere plankton, vägatäpne joonistamine, kirjeldamine ja nimetamine. Temalt pä-rinevad mõisted ja terminid hõimkond, fülogenees, ökoloo-gia ja .... protistid.Ernst Haeckeli andeka kunstnikukäe all kinnistus teadu-
ses eluslooduse jaotumien kolme riiki: taimed, loomad japrotistid (joon. 19)
Dihhotoomia: eukarüodoodid—prokarüoo-didAntiikajast eluslooduse esmane jaotus taimedeks ja looma-deks muutus prokarüootide avastamisega 20 sajandil. Orga-nismide esmane jaotus prokarüootideks ja eukarüootideksraiuti kivisse 1962 Roger Stanier’i ja C. B. van Niel poolt ar-tikliga The concept of a bacterium [Stanier and Niel, 1962],milles tsiteeritakse tublisti Édouard Chatton’i 1937 ilmunudraamatutTitres et Travaux Scientifiques [Chatton, 1938], kusChatton ühes lõigus märgib prokarüootide ja eukarüootidepõhimõttelist erinevust. Chatton oli esimene kes lähtuvaltrakuehitusest tegi vahe sisse eukarüootidel (defineeriti tõeli-se ‘tuuma’ alusel) ja prokarüootidel organismid millel ei oletuuma). ja võttis 1925 a vastavad terminid kasutusele [Sapp,2005]. Prokarüoodi mõiste suht jäi unustusse, kuni Stainerja Niel selle Cattoni lõigu avastasid ja taas rambivalgusessetõid. Arusaam prokarüootse ja eukarüootse raku ehituslikuerinevuse põhimõttelisusest on üks olulisemaid elusloodusemitmekesisuse mõistmisel.
Bakterite kontseptsioon
Dihhotoomia eukarüoodid — prokarüoodid ei olnud kahe-kümnenda sajandi algusemikrobioloogidele iseenesestmõis-tetav. Põhjuseks see, et prokarüoote (termin mida tollel ajalkasutati väga vähe) defineeriti täielikult ‘negatiivsete tun-nuste alusel’. S.t. prokarüoodid on organismid, millel puudu-vad teatud kindlad tunnused, mis iseloomustavad eukarüoo-
Joonis 19. Ernst Haeckel’i joonis elu puust kujutas tolleaegsetarusaama eluslooduse jaotusest ja evolutsioonist.
te — membraaniga ümbritsetud tuum, rakuskelett, rakusi-sesed membraansüsteemid, jne. Ei olnud palju midagi, misprokarüoote ühendaks. Hästi oli teada prokarüootide suurfüsioloogiline varieeruvus — aeroobsed, anaeroobsed, või-malised kasutama erinevaid energia ja süsinikuallikaid. Pro-karüoodid on füsioloogialt ja metabolismilt palju mitmeke-sisemad kui eukarüoodid.Umbusule vaatamata juurdus arusaam, et nii nagu euka-
rüoodid, nii on ka prokarüoodidmonofüleetiline takson. CarlWoese on tõdenud [Woese, 1998], et tegemist oli ‘uue usu-ga’, mitte ‘uute faktidega’. Suur osa bakterite rakubioloogi-listest ja molekulaarsetest tunnustest saadi Escherichia colija paari teise liigi kohta ja keegi ei esitanud kriitilist kü-simust — kas neid tunnuseid võib ekstrapoleerida kõigileprokarüootidele. Prokarüootide monofüleetilisust võeti vai-kimisi kui midagi iseenesest mõistetavat.
8ELU PUU AJALOOLINE JA FILOSOOFILINE TAUST

Joonis 20. Carl Woese (1928–2012) oli USA Illinoise Ülikoolismikrobioloog. Tuntud eelkõige eluslooduse kolme domeeniparadigma loojana.
Uus paradigma: eluslooduse kolmdomeeni
Üks esimesi tähtsamaidmakromolekule,mille järjestuste ana-lüüs viis suuremate üldistusteni oli ribosomaalse RNA väikesub-ühik (ssu-rRNA). Esiteks, ssu-rRNA sisaldab nii küllaltvarieeruvaid kui ka väga konservatiivseid piirkondi. See või-maldab uurida vastavalt lähedasi ja väga kaugeid evolutsioo-nilisi suhteid organismi(rühma)de vahel. Teiseks, molekulisuurus (ca 1400–1800 aluspaari) on piisavalt suure infor-matsiooni hulgaga esmasteks üldistusteks. Samas on seemu-gavalt väike, nii et üsna peatselt jõuti sekveneerida suuremhulk organisme mis võimaldas analüüse ja üldistusi teha.Carl Woese. Kasutades molekulaarseid meetodeid ja ssu-rRNA andmeid õõnestas Woese 1970ndate lõpus [Woeseand Fox, 1977, Woese et al., 1990] seni valitsenud ettekuju-tust eluslooduse esmasest jaotamisest prokarüootideks ja eu-karüootideks. Uuridesmetanogeensete bakterite ssu-rRNAdtulid nad järeldusele, et metanogeenid on fülogeneetiliseltsama lähedased eukarüootidele kui teistele bakteritele; li-saks on metanogeenide haru evolutsiooniliselt sama sügav(vana) kui mistahes teiste bakterite haru. Arvestades meta-nogeenide metabolismi eripära, leidsid Woese ja Fox [Wo-ese and Fox, 1977] selle olevat ideaalse elu tekkimiseaegsehapnikuvaba ja süsihappegaasirikka keskkonna jaoks ningtegid ettepaneku nimetada metanogeene and nende lähedaltseotud organisme arhebakteriteks ehk arheteks (Archaea,eristamaks neid päris bakteritest ehk eubakteritest ja rõhuta-maks nende vähemalt sama ürgset päritolu.
rRNA fülogeneesi tulemusena valminud ‘Elu Puu’ näitas,
et monoliitseks peetud prokarüootide rühm koosneb
põhimõtteliselt erisugustest organismirühmadest, mis
on omavahel fülogeneetiliselt sama kauged, kui iga rühm
eraldivõetuna on eukarüootidest.
Woese ja Fox [Woese and Fox, 1977] panid ette klassifit-seerida kaasaegsed elusorganismid kolme suurde peamisserühma, mida nimetatakse ka domeenideks (riigist kõrgemakategooria takson): eubakterid, arhebakterid ja euakrüoo-did. Kahte prokarüootset rühma nimetatakse tänapäeval ar-head (ka arhed) ja bakterid, rõhutamaks, et need kaks rühmaei ole omavahel lähedasemas suguluses kui kumbki rühm eu-karüootidega; sünonüümidena kasutatakse ka arhebakteriteja eubakterite nimetusi. Kõik kolm suurjaotust on arenenudühisest eellasest; neil on väga pikk sõltumatu evolutsiooni-line ajalugu ja on võimalik, et nad on lahknenud peatseltpärast esimeste elusorganismide tekkimist. Selle hüpoteesikohaselt ei ole prokarüoodid monofüleetiline rühm, nagutavapäraselt arvati.
Woese põhiväide oli. et prokarüootide kaks rühma, bak-
terid ja arhed, on omavahel sama erinevad kui kumbki
rühm eukarüootidega. Woese esitas kolm domeeni juu-
rimata puuna, ehk ei öelnud midagi selle kohta, millised
kaks domeeni on sõsar-rühmad.
‘Woese Revolutsioon’ muutis põhjalikult nägemust elus-looduse mitmekesisusest. Selle tulemusena levis uus para-digma: organismide esmane jaotus ei ole mitte teljel pro-karüoodid — eukarüoodid, vaid organismid moodustavadesmaselt kolm domeeni, Archeae (arhebakterid), Bacte-ria (eubakterid) ja Eucaryota (eukarüoodid) [Woese et al.,1990]. Seega on meil tegemist kolme, mitte kahe peamiseeluslooduse grupeeringuga. Iseäranis oluline on tähele pan-na, et need grupeeringud on (vähemalt taotluslikult) füloge-neetilised. Selline Elu Puu esmane hierarhia on hüpotees or-ganismide genealoogiast, nende põlvnemisest ühest ühisesteellasest. Sellel ei ole a priori midagi tegemist raku ehitus-tüübi, funktsiooni, füsioloogia või muu sellisega — ainultfülogeneesiga. Kas see hüpotees vastab tegelikkusele (tõe-le), ei saa me kunagi teada — paradoksaalselt ’Tõde’ ei oleteaduse kategooria. Kõikmida usin uurimistöö ja suured avas-tused teha saavad, on kas püstitatud hüpoteese toetada võiümber lükata.Kolme domeeni mudel on tänapäeval üldiselt aktsepteeri-
tud (joon. 21).Arhed ja bakterid — ühised jooned. Mõlemal on proka-rüootne rakuehitus, 70S ribosoomid ja rõngasjas kromosoo-mid (enamikel liikidel üks). Eukarüootidel rakuehitus on eri-nev, neil on membraaniga ümbritsetud tuum paljude lineaar-sete kromosoomidega ja selgelt nähtav rakusisene kompart-
9UUS PARADIGMA: ELUSLOODUSE KOLM DOMEENI

Joonis 21. Klassikaline kolme domeeni skeem, mis näitabeukarüootide, ahede ja bakterite sügavat harunemist.
mentalisatsioon7.Arhed ja eukarüoodid — ühised jooned.Mõlemil on gee-nides intronid. Arhede RNA polümeraas ja transkritsiooni-faktorid on sama tüüpi mis eukarüootidel ja väga erinevadbakterite vastavatest.Arhede unikaalsed tunnused. Arhede membraani lipiididon isoprenoidid, samas kui bakteritel ja eukarüootidel koos-nevad need ester-sidemetega rasvhapetest.Kumb on olulisem, kas geenijärjestus või erinevused ra-
kuehituses? Päris üksmeelt ei ole.Horisontaalne geeniülekanne. Eri viisidel kanduvad gee-nid üle horisontaalselt organismilt organismile, sh eri do-meenide vahel. Seega võib elu puu olla pigem võrgustikutaoline, kui diskreetselt kolme domeeni jaotunud.
Endosümbioosi teooriaEukarüootidele omaste rakuorganellide (tsentrioolid/viburid,mitokondrid, kloroplastid, peroksüsoomid) teket on tihti sa-mastatud eukarüootse organismi tekkega. Endosübmioosi teooriaväidab, et eukarüootse raku mitmed organellid, sealhulgasmitokonder ja plastiid, on endosümbiontset päritolu — ontekkinud sümbiontsest suhtest kahe ainurakse organismi va-hel. S.t. ürgne fagotroofne eukarüootne rakk on all neelanudsaakorganismi, kuid mingil põhjusel seda ei seeditud, vaidsee säilis uues peremeesorganismis kui rakusisene sümbiont(endosümbiont), mis edasise evolutsiooni käigus muutus ra-kuorganelliks.Kõige olulisemad eukarüootsed organellid, mida endosüm-
bioosiga seostatakse, onmitokondrid ja plastiidid (kloroplas-tid). Välistada ei saa mõnede teiste rakuorganellide endo-sümbiontset päritolu. Vaid plastiididel ja mitokondritel onoma genoom, mis on jäänuk nende organellide vabalt elavaeellase genoomist, ning on seega kindlamaks tõendiks nendeendosümbiontsest päritolust.Mitokondri eellaseks on alfa rühma proteobakater, plastii-
di eelaseks on tsüanobakter ehk sinivetikas. Endosümbioosiaeg ulatud ilmselt miljardite aastate taha. Mitokondri endo-sümbioos on ilmselt toimunud evolutsioonis ainult ühel kor-
7Membraanidega ümbritsetud organellid.
ral ja tõenäoliselt on mitokondrid seega oma päritolult mo-nofüleetilised.Primaarne plastiidi endosümbioos on ka toimunud vaid
ühel korral evolutsioonis, koos ühe lisa-erandiga8. Kuid plas-tiididele on omane sekundaarsed ja tertsiaarsed endosüm-bioosid. Primaarne endosümbioos tähistab vabaltelava sini-vetika omastamist ja kujunemist primaarseks plastiidiks (näi-teks rohevetikatel ja taimedel). Sekundaarse endosümbioosikäigus omastati juba eukarüootne vetikas ning see muutusuue peremeesorganismi sees sekundaarseks plastiidiks. See-ga on primaarsete plastiidide otseseks eellaseks vabalt elavsinivetikas. Sekundaarsete plastiidide otseseks eellaseks onvabalt elav eukarüootne vetikas, omakorda kelle plastiidieellaseks on ikkagi vabalt elav sinivetikas.Endosümbioosi teooria esmane versioon seostub üleeel-
mise sajandi lõpul ilmunud Shimperi tööga [Schimper, 1883]ja eelmise sajandi alguseMereschkowsky töödega [Mereschkows-ky, 1905, 1910]. Laiemalt need ideed tollel ajal aga ei le-vinud ja üldisem tuntus (ja tunnustatus) saabus alles LynnMargulis’e töödega [Margulis, 1970, 1981].Endosümbioosi teooria taaselustavas ülevaates [Margu-
lis, 1970] nägi Lynn Margulis eukarüootide tekkimist ast-meliselt toimunud sümbiootilise ühendusena prokarüootserakuga. Selle skeemi kohaselt tekkisid eukarüootidele ise-loomulikud tunnused kas samaaegselt endosümbioosiga võihiljem. Endosümbioosi eelset eukarüootset evolutsiooni eiolnud — eukarüootide evolutsiooni algust saab hakata luge-ma endosümbioosi tekkimise momendist.Teised endosümbioosi teooria pooldajad olid eriarvamu-
sel. Nemad nägid eukarüootsete organismide tekkimise es-mase eeldusena fagotsütoosi võimet. Koos fagotsütoosigaarenesid ka sellised eukarüootsed tunnused nagu rakuskelettja endomembraanne süsteem [Stainer, 1970]. Mehhanistli-kult on endosümbioos palju paremini mõistetav kui rakulon juba olemas tunnused mis võimaldavad fagotsütoosi.Endosümbioosi hüpotees osutus väga edukaks organelli-
de mitmete tunnuste seletamisel (e.g. bakteriaalset tüüpi or-ganelli DNA, tundlikkus antibiootikumidele, organelli ja eu-bakterite nukleotiidse järjestuse sarnasus). Tänapäeval onüldiselt aktsepteeritud, et mitokondri eellane on endosüm-biootiline α - rühma purpurbakter ja plasatiidi eellaseks olivabalt elav sinivetikas ehk tsüanobakter.Nii plastiid kui mitokonder on rakuorganellid. Rakuor-
ganellideks ei muutunud endosümbiondid mitte üleöö, vaidsellega kaasnes pikk evolutsiooniliste sündmuste rida. Mison vahet endosümbiondil ja organellil? Peamine erinevuson genoomi suuruses. Nii plastiidi kui mitokondri genoomon ca. kümnendik eellas-prokarüoodi genoomi suurusest. Jä-relikult üks olulisemaid muutusi, mis endosümbiontidegatoimus oli genoomi vähenemine. Suur osa kaasaegsete eu-karüootsete organismide organellide valkudest kodeeritakseperemeesorganismi tuuma genoomi poolt, vajalikud valgud
8Teadagi ei saa me midagi öelda väljasurnud organismide kohta.
10ENDOSÜMBIOOSI TEOORIA

sünteesitakse peremeesorganismi rakuplasmas ja transpordi-takse seejärel läbi kaksikmembraani organelli. Teatud osavalkudest kodeeritakse ka organelli genoomi poolt.
Mitokondri omastamineMitokonder on kaksikmembraaniga ümbritsetud eukarüoot-se raku organell, milles toimub rida metaboolseid protsesseja kus toimuva aeroobse hingamise käigus toodetakse ATPd.
Mitokondri endosümbioos:
Vabalt elava aeroobselt hingava bakteri omastamineanaeroobse heterotroofse protisti poolt ning selleevolutsioneerumine esmalt endosümbiondiks, seejä-rel püsivaks rakuorganelliks—mitokondriks. Tekibesimene aeroobselt hingav eukarüootne protist.
Endosümbioosi teooria kohaselt muutus bakter esmalt eu-karüootse raku endosümbiondiks ja kaotas suurema osa omaautonoomsusest. Mitokondrite arv rakus võib ulatuda ühestmitmesajani. Mõnedel taksonitel on mitokondrid muundu-nud ja tagavad tavalisest erinevaid, kuid lähedasi biokeemili-si funktsioone.Ilmselt omastati mitokondrid eukarüootide poolt evolut-
sioonis vaid üks kord, unikaalse evolutsioonilise sündmuse-na. Kui see on nii, siis mitokondrid on monofüleetilised.Lähtuvalt mitokondriaalseDNAväiksemast hulgast ja suu-
remast integratsiooniastmestmitokondri ja peremeesraku va-hel võib arvata, et võrreldes kloroplastiga toimusmitokondriomastamine ajaliselt varem.
Mitokondri sekundaarne kadumine
Paljudel juhtudel on mitokonder kadunud sekundaarselt janeid organisme nimetatakse amitokondrilisteks. Amitokond-rilisi protiste iseloomustab anaeroobne metabolism. Näitekskui organism on väga spetsialiseerunud ja asunud elama anae-roobsesse keskkonda. Anaeroobses keskkonnad on näiteksseisva veega veekogude orgaanikarikkad setted, komposti-hunnikud, reoveepuhastusjaamad. Sellistes keskkondades eiole aeroobne hingamine võimalik jamitokondriga ei ole suurtmidagi peale hakata. Amitokondriaalsete hulka kuulub kaparasiite ja soolestikufloora esindajaid. Neid on leitud me-rest ja riimveest.
Arhezoa hüpotees
Amitokondrilistes protistides nähti ülemineku-eukarüoote, alg-seid ‘eelmitokondrilisi organisme’. Sellest lähtuvalt tegi Tho-mas Cavalier-Smith 1983 a. ettepaneku luua eukarüootidealamriik Archezoa [Cavalier-Smith, 1983], millesse pididkuuluma primitiivselt amitokondrilised protistid, s.t. orga-nismid, kellel ei ole evolutsioonilise ajaloo vältel kunagi mi-tokondreid olnud. Enamasti on amitokondrilised protistid
A B CJoonis 22.Morfoloogiliselt eri tüüpi mitokondrid. Mitokondritümbritseb kahemembraaniline kate; välimine on sile jamolekulidele vabalt läbitav, sisemsel on sissesopistised (kristad),see on keeruka ehitusega ja selles toimuvadrespiratsiooniprotsessid. A — kettakujuliste kristadega; B —lamedate kristadega; C — tubulaarsete kristadega.
rakuehituslikult väga lihtsad. Paljudel puuduvad iseloomu-likud eukarüootsed organellid, sageli on ka raku sisememb-raanistik puudulikult arenenud. See asjaolu süvendas veelgiarvamust, et tegu on ‘primitiivsete’ eukarüootidega.Ilmselt põlvnevad kõik tänapäeval elavad amitokondrili-
sed protistid aeroobsetest, mitokondriga eellastest. Näitekskui eukarüoodi tuumas leidub mitokondri genoomi marke-reid, viitab see mitokondri olemasolule eellasel. Seega onamitokondrilisus nende puhul mitte primitiivne, vaid hoopisevolutsiooniliselt arenenud, ehk apomorfne tunnus.
Mitokondri ehitus
Mitokondrit ümbritseb kahemembraaniline kate. Siseminemembraan on tugevalt sisse sopistunud, moodustades kris-tad. Kristad võivad kujult olla (i) ketasjad, (ii) tubullaarsed,(iii) lamedad (joon. 22 ja 23).Peamiselt morfoloogilistele tunnustele tuginedes on mi-
tokondrite päritolu peetud ka polüfüleetiliseks. Molekulaar-sed andmed kinnitavad vastupidist — mitokondrid on mo-nofüleetilised ja järelikult on erinev kristade kuju juba mi-tokondri evolutsiooni tulemus.Mitkonondrite monofüleetili-sus tähendab, et mitokondri endosümbioos on evolutsioonistoimunud vaid üks kord ja kõik kaasaegsete organismidemitokondrid põlvnevad ühes ühisest eellasest.Kui mitokondrid on tõepoolest monofüleetilised, nagu on
praegune konsensuslik arusaam, siis on kristade kuju ükstähelepanuväärne ultrastruktuurne tunnus, mis potentsiaal-selt viitab eukarüootide suurematele evolutsiooniharudele.Kristade kujul tundub olevat fülogeneesi seisukohalt suu-rem tähtsus kui võiks esmapilgul arvata. Järelikult on mi-tokondrite evolutsioon toimunud paralleelselt peremeesor-ganismide evolutsiooniga ja nende ultrastruktuurne mitme-kesisus peaks peegeldama peremeesorganismide mitmeke-sisust. Taksoneid, kus esineks mitut tüüpi mitokondreid, onvähe. Ketasjate kristadega taksonid eralduvad teistest mito-kondrilistest varem. Kuigi ülejäänute omavahelised seosed
11ENDOSÜMBIOOSI TEOORIA

Joonis 23. Kolme peamist tüüpi mitokondrid. Vasakul —ketasjate kristadega; keskel — tubullaarsete kristadega; paremal— lamedate kristadega. Skemaatilist pilti vaata jooniselt 22.
on vähem selged, on põhjust arvata, et tubullaarsete ja lame-date mitokondri kristadega rühmad on monofüleetilised.Mitokondri kristade kuju järgi on provisoorselt pakutud
ka taksonite nimetusi:
Discichristata: Ketasjate kristadega eurkarüoodid. Siia kuu-luvad hõimkonnad Euglenozoa, Percolozoa.
Platychristata: Lamellaarsete kristadega eurkarüoodid. Pu-navetikad, rohevetikad (sealhulgas maismaataimed), loo-mad, seened, samuti hütriidid.
Tubulochristata: Tubullaarsete kristadega eurkarüoodid. Al-veolaadid (e.g. dinoflagellaadid, tsiliaadid), stramenopii-lid (e.g. ränivetikad, pruunvetikad, koldvetikad, etc.)
Vaba hapniku roll eukarüootidemitmekesisuse ku-junemiselVaba hapniku ilmumine Maa elukeskkondadesse on seotudsinivetikate tekkimisega. Sinivetikad, mis fülogeneetiliseltmoodustavadmonofüleetilise gram-negatiivse eubakterite rüh-ma, olidMaa esimesed oksügeensed fotoautotroofid. S.t. omaelutegevuseks kasutasid nad energiaallikana valgusenergiat(fototroofia), süsinikuallikana kasutasid süsihappegaasi (au-totroofia) ja selle protsessi tulemusena eraldus vaba hapnik(O2). Sinivetikate ilmumise aja kohta on andmed küllalt eba-määrased, suurusjärk on kusagil kolme miljardi aasta taga.Sellele viitavad stromanoliitide dateeringud—need on vani-mad fossiilsed säilmed kunagi elanud sinivetikatest (joon. 24).Vaba hapniku varajane akumulatsioon ookeanis oli väga aeg-lane. Seda põhjusel, et algses anaeroobses ookeanis oli paljuredutseerunud ioone, eriti rauda Fe(II). Fe(II) oksüdeerimi-ne F(III)-ks nõudis pikema ajaperioodi vältel kogu vaba hap-niku. Alles vaba hapniku olemasolu keskkonnas võimaldasareoobselt respireerivate bakterite teket.Kui ookeani vees lahustunud Fe(II) oli oksüdeeritud, tek-
kis võimalus vaba hapniku kontsentratsiooni suurenemiseksvees, kust see aegamisi difundeerus kaMaa atmosfääri. Ena-mikule anaeroobsetele organismidele oli vaba hapnik ilm-selt väga toksiline. Sellistes tingimustes on aeroobselt res-
Joonis 24. Stromatoliidid on arvatavalt vanimad fossiilsedsäilmed sinivetikatest. Shark Bay, Autsrtaalia.
pireeriva bakteri omastamine anaeroobse organismi poolt,mis on endosümbioosi ja mitokondri tekkimise eelmänguks,ilmselgelt kasulik kohastumus. Omastatud aeroobne külali-ne kasutas vaba hapniku ja seega alandas peremeesorganis-mi rakusisest vaba hapniku kontsentratsiooni. Suure tõenäo-susega oli peremeesorganismi esmane ‘kasu’ endosümbion-dist kaitse toksilise hapniku vastu. Alles seejärel evolutsio-neerus endosümbiont mitokondriks ja korraldas kogu pere-meesraku anaeroobse metabolismi ümber aeroobseks. Sel-lest arenes tohutu energeetiline hüpe peremeesraku metabo-lismi effektiivsuses, mis omakorda võimaldas suure evolut-sioonilise divergeerumise. Tõenäoliselt oli anaeroobse pe-remeesorganismi peamine energia tootmise viis fermentat-sioon. Kui selleks sai, tänu mitokondri endosümbioosile, ae-roobne respiratsioon, tähendas see kordades effektiivsematenergia tootmise viisi.Atmosfäärsel hapnikul on kaks olulist rolli:
• See andis olulise tõuke aeroobselt respireerivate organismi-de arenguks. Kuna aeroobse respiratiooni kasutegur onoluliselt (keskmiselt ca. 5 korda) suurem fermentatsioonikasutegurist, said sellised organismid olulise eelise. Ae-roobne respiratsioon võimaldab orgaanilise süsiniku ’põ-letamist’ maksimaalselt oksüdeeritud anorgaanilise vor-mini (CO2). Fermenteerivad organismid suutsid omasta-da vaid väikese osa orgaanilises süsinikus olevast kee-milisest energiast; jääkproduktid olid ikka enamasti veelküllalt energiarikkad orgaanilised ained. Orgaanilise C sü-sihappegaasini (või metaanini) lõhustamine nõudis mit-mete eri organismide tegevust, kusjuures igaühe osaks saivaid murdosa algses substraadis sisaldunud keemilisestenergiast. Seega oli iga organismi kasutegur eraldi võe-tuna suhteliselt madal.
• Teiseks, arvestatav hulk hapniku tekitas atmosfääri üle-mistes kihtides, stratosfääris, 25–30 km kõrgusel, osooni-kihi. See andis tõuke elu arenguks maismaal, kus varemoli tugev ultravioletkiigus organismidele hukatuslik.
Eeltoodu on otseses seoses ka fülogeneesi ja süstemaa-tikaga. Vaba hapniku tekkimine keskkonda ja elu tungimi-
12ENDOSÜMBIOOSI TEOORIA

ne maismaale põhjustasid olemasolevate organismide kiireevolutsioonilise radiatsiooni. Samas ilmselt ka küllalt ulatus-liku anaeroobsete organismide väljasuremise. Hapniku aku-muleerumine esmalt veekeskkonda (kuhu sinivetikad sedaeritasid) langeb kokku loomade (Metazoa) kiire radiatsioo-niga — paljude harude tekkimine suhteliselt lühikese aja-vahemiku jooksul. Elu levimine maismaale langeb kokkukõrgemate taimede kiire radiatsiooniga.
Aeroobne respiratsioon kombinatsioonis fagotroofse toi-
tumisega on üks olulisemaid sündmusi elusorganismide
evolutsioonis: see tegi võimalikuks loomsete organismi-
de ning toiduahelatel põhinevate ökosüsteemide tekki-
mise.
NB — oksügeenne fotosüntees ja aeroobne respirat-
sioon kui eukarüootse elu võtmeprotsessid on mõle-
mad sügavalt prokarüootset päritolu. Mis oleksid euka-
rüoodid ilma aeroobse respiratsiooni võimeta?
Valdav osa eukarüootidest kasutab aeroobset respiratsioo-ni energeetilise lähteprotsessina, sealhulgas ka fototroofid.See ei oleks ilmselt mõeldav piisava hulga vaba hapnikuolemasoluta, mis jällegi eeldab suure hulga fototroofide ole-masolu. Enne plastiidide endosümbioosi olid ainsateks va-ba hapniku tootjateks sinivetikad. Praegusel ajal toodetaksesuurem osa hapnikust eukarüootsete organismide poolt.Kaks peamsit protsessi — aeroobne respiratsioon ja ok-
sügeenne fotosüntees, mis teevad eukarüootidest planeediMaa ‘suured ja vägevad’, on mõlemad prokarüootidelt ‘lae-natud’.Kuigi prokarüootide morfoloogiline mitmekesisus on eu-
karüootidega võrreldes täiesti kahvatu, tuleb tunnistada, etmetaboolse paindlikuse ja mitmekesisusega on lood täpseltvastupidi. Nii oksügeenne fotosüntees kui aeroobne respirat-sioon on vaid pisike osa prokarüootide tohutust metabool-sest arsenalist.
Plastiidi omastaminePlastiidide evolutiooni peatükk peaks andma hea ettekuju-tuse vetikate olmusest. Oluliseim on aru saada, et vetikad eiole takson vaid ökoloogiline ja funktsionaalne kategooria.Mis puutub kloroplasti, siis on tõenäoline, et endosüm-
biontide evolutsioon kloroplastiks on toimunud sõltumatultmitmeid kordi. See on oluline erinevust mitokondri endo-sümbioosist.Oluline vahet teha primaarsel ja sekundaarsel endosüm-
bioosil:
Primaarne endosümbioos —vabalt elava sinivetika omas-tamine heterotroofse protisti poolt ning selle evolutsio-
neerumine esmalt endosümbiondiks, seejärel püsivaks ra-kuorganelliks — primaarseks plastiidiks. Tekib esimeneeukarüootne vetikas.
Sekundaarne endosümbioos —juba eukarüootse vetika omas-tamine uue heterotroofse protisti poolt ning selle evolut-sioneerumine esmalt endosümbiondiks, seejärel püsivaksrakuorganelliks — sekundaarseks plastiidiks.
Arvatavalt on primaarne endosümbioos on olnud unikaal-ne sündmus evolutsioonis ja on toimunud vaid üks kord. Se-kundaarsed (ja isegi tertsiaarsed endosümbioosid) on toimu-nud korduvalt ja üksteisest sõltumatult. Mitu korda täpseltja milliste organismidega seoses— selles osas on otsad veellahtised.
Plastiidide primaarne endosümbioosEsmane endosümbioos, ehk kloroplastide eellas-sinivetikaesmane omastamine heterotroofse peremeesraku poolt. Se-da protsessi peetakse unikaalseks, ainult üks kord evolut-siooni vältel toimunuks9. Siit järeldus, et kõik tänapäevalteada olevate prganismide plastiidid pärinevad ühest eella-sest ja kloroplastid on seega monofüleetilised10. Sekundaar-sete plastiididega organismide kohta kehtib see väide kaud-selt — otseselt on nende plastiid eukarüootset päritolu, kuidselle eukarüootset päritolu vetika plastiidi eelane oli ikka-gi vabalt elav sinivetikas. Tertsiaarsete plastiididega orga-nismide puhul, iseenesest mõista, lisandub veel üks vaheas-te.
Arheplastiidsed — kolm haru primaarsete plastii-didega organismePraeguse seisuga on teada kolm primaarsete plastiididegaeukarüootide haru:
1. Rohevetikad (Chlorophyta; sealhulgas kõikmaismaatai-med ehk embrüofüüdid)
2. Punavetikad (Rhodophyta)3. Liitvetikad ehk glaukofüüdid (Glaucophyta).
Primaarsete plastiididega organismidel ümbritseb klo-
roplasti kahemembraaniline kloroplastikate (joon. 25).
Tekkelt on need kaks membraani sinivetika (kui gram-
negatiivse bakteri) sisemine ja välimine rakumembraana.
aVarasema interpretatsiooni kohaselt on kloroplastikatte siseminemembraan tekkelt endosümbiondi rakumembraani, välimine pere-meesorganismi toitevakuooli membraan.
9On üks vähe uuritud erand—Paulinella chromatophora. V.t. joonis 2710terminit ‘mono-, para- ja polüfüleetiline’ võib seega kasutada mitte
ainult organismide, vaid ka (teatud) organellide kohta
13ENDOSÜMBIOOSI TEOORIA

Kui primaarne endosümbioos on evolutsioonis toimunudvaid üks kord, siis järelikult moodustavad puna-, rohe- jaliitvetikad koos ühe monofüleetilise rühma ja plastiidi omas-tas nende kolme ühine eellane. Seetõttu nimetatakse sedakolmest harust koosnevat rühma arheplastiidseteks - Arc-haeplastida ehk ’taimed’ kõige laias mõistes.Arheplastiidsete kolm haru väga erinevate kloroplastide-
ga. Erinev on nii pigmentatsioon kui ka kloroplasti siseehi-tus. Pigmentatsioonilt ja siseehituselt on punavetika (ja kaglaukofüüdi) plastiid suhteliselt sarnane eelas-sinivetika ra-kuga. Peamisteks kaaspigmentideks on fükobiliinid, mis an-navad rakule punaka või sinaka tooni. Tülakoidid — plas-tiidi sisesed membraanid, millel toimub fotosüntees, paik-nevad plastiidis üksikult. Rohevetika plastiid on täiesti eri-nev — fükobiliinid on kadunud, kaaspigmendina on tekki-nud klorofüll b, värvuselt on plastiidid erkrohelised. Tüla-koidid on pakitud kihiliselt lamellideks.Sellest järeldub, et pärast plastiidide primaarset endosüm-
bioosi on toimunud väga kiire plastiidide evolutsioon uueperemeesraku sees. Muutunud on nii plastiidi struktuur kuipigmentatsioon.
Sekundaarsed endosümbioosid
Sekundaarse endosümbioosi puhul on algne heterotroofneeukarüoot neelanud juba primaarsete plastiididega eukarüoot-se vetika (s.t. organismi mis tekkis esmase endosümbioositulemusena), mis seejärel on juba eukarüootset päritolu endo-sümbiont arenenud püsivaks rakuorganelliks — sedapuhkusekundaarseks plastiidiks.Primaarseid ja sekundaarseid endosümbioose on traditsioo-
niliselt eristatud plastiidikatte membraanide arvu järgi.
Sekundaarsete plastiididega organismide kloroplasti ümb-
ritseb üldjuhul neljamembraaniline kloroplastikate: väli-
mine on tekkelt peremeesorganismi toitevakuooli memb-
raan, teine membraan on eukarüootse endosümbiondi
plasmamembraan (neid kahte nimetatakse ka kloroplasti
endoplasmaatiliseks retiikulumiks ehk CER) ja kaks sisemist
on sama interpretatsiooniga kui primaarse endosüm-
bioosi puhul.
Erandid, erandid!. Eugleniididel ja suuremal osal dinofla-gellaatidest (nn. stereotüüpsed dinoflagellaadid, ehk peridi-niini sisaldavad dinoflagellaadid) on plastiidid ümbritsetudkolme membraaniga. Neljanda membraani puudumist on se-letatud ad hoc hüpoteesiga, kus võtmeroll on nende orga-nismide toitumisstrateegial. Nimelt heterotroofne protist eipruugi saaki tervelt neelate, vaid näiteks uuristada spetsiaal-se organelli abil ohvri rakuseina augu, mille kaudu koguraku sisu välja imetakse. Nii puudub nende kloroplastikattesmembraan, mis vastaks tekkelt endosümbiondi rakumemb-
raanile.Sekundaarseid endosümbioose on evolutsioonis iseseis-
valt toimunud mitmeid kordi. Sekundaarsete plastiididegaorganisme saab kloroplasti integreerituse astme järgi jagadaveel kahte rühma: nukleomorfiga organismid ja organismid,kus nukleomorf on kadunud.
Nukleomorfiga organismid
Nukelomorf on sekundaarsete plastiididega vetikate eu-
karüootset päritolu plastiidi tuuma jäänuk.
Nukleomorf on nähatav elektronmikroskoobis — ajaloo-liselt oli see esimesi ja kindlamaid tõendeid plastiidide endo-sümbiootilise päritolu kohta. Plastiidide sekundaarne endo-sümbioos ilmnes esmalt krüptomonaadide (Cryptophyceae)ultrastruktuuri uurides. Krüptomonaadides avastati eukarüoot-set päritolu endosümbiondi tuuma jäänuk. Hiljem avastatisama ka klorarahinofüütidel (Chlorarachinophyceae). Ühel-gi teisel organismirühmal ei ole tänaseni nukleomorfi ole-masolu teada.
Nukleomorf on kadunud
Nukleomorf on kadunud autotroofsetel stramenopiilidel, hap-tofüütidel (Haptophyceae), eugleniididel (Euglenophyceae),dinoflagellaatidel (Dinophyceae) ja parasiitsel tippeoslas-te rühmal (Apicomplexa). Tippeoslastel on funktsioneeru-vad plastiidid kadunud sootuks — alles on vaid klorofüllimittesisaldav plastiidi jäänuk.Stramenopiilide, haptofüütide ja krüptomonaadide plas-
tiidide eelaseks on olnud punavetika rakk. Seda vaatamatapigmentatsiooni erinevustele — stramenopiilidel on kloro-füllid a ja c, puuduvad fükobiliinid; punavetikatete ja krüp-tomonaadide plastiididel on fükobiliinid olemas (ühine ürg-ne tunnus ehk sünplesiomorf sinivetikatega — plastiidideeellasega). Lisaks on krüptomonaadidel ka klorofüll c. Sa-mas, plastiidid välja arvata, ei ole krüptomonaadid, puna-vetikad ja stramenopiilid fülogeneetiliselt lähedalt seotud.Neid ühendab kaudselt ainult sama päritolu plastiid.
Tertsiaarsed endosümbioosid
Nagu eelnevast veel vähe, on plastiidide evolutsioonis kor-duvalt ja iseseisvalt ette tulnud veel tertsiaarset endosüm-bioosi. Sel juhul, pole vist rakse arvata, on endosümbiondiksolnud juba sekundaarsete plastiidiega eukarüootne vetikas.Ei ole küll ühtegi vetikate rühma, kus kõik liigid oleksid
tertsiaarsete plastiididega. Tuntumad näited on dinoflagel-laatide hulgas, kus tertsiaaresid endosümbioose on toimu-nud mitmeid kord iseseisvalt. Näitek dinoflagellaat Peridi-nium foliaceum endosümbiondiks on ränivetikas. Kuna räni-vetikas (nagu kõik teisedki fototroofsed stramenopiilid) on
14ENDOSÜMBIOOSI TEOORIA

Stramenopiilid Haptofüüdid Euglenofüüdid Dinoflagellaadid Apikompleksa
Primaarneendosümbioos
Sekundaarneendosümbioos
TuumSinive tikas
EukarüootEukarüoot
Eukarüoot
Rohevetikad Punavetikad Glaukotsüstofüüdid
Geeni ülekanne
Primaarneplastiid
Nukleomorf kaobPrimaarneplastiid
Sekundaarneplastiid
NukleomorfKlorarahinofüüdidKrüptomonaadid
Joonis 25. Kloroplasti endosümbioosi lihtsustatud skeem. Primaarne endosümbioos on tõenäoliselt toimunud vaid üks kord evolutsioonivältel, sekundaarne aga mitmeid kordi.
Joonis 26. Gymnodinium acidotum endosümbiontsetekrüptomonaadidega. Näide tertsiaarsete plastiididega organismist
juba sekundaarsete plastiididega, siis ongi siin tegemist tert-siaarse endosümbioosiga. Samas, erinevalt eelnevatest, onPeridinium foliaceum endosübiondi tuum veel redutseeru-mata genoomiga. Teine dinoflagellaat,Gymnodinium acido-tum, hoiab endas endosümbiootilisi krüptomonaade (joon. 26).Head näited tertsiaarsest endosümbioosist on fukoksantii-
ni sisaldavad dinoflagellaadid, nagu Karenia brevis, Gym-nodinium galatheanum,Gymnodinium aureolum. Nendel lii-kidel on nii plastiidide pigmentatsioon kui ka ultrastruktuursarnane haptofüütidega (rühm merelisi vetikaid millel onsekundaarsed plastiidid). Kuna nukleomorfi (sedapuhku siishaptofüüdi tuuma jäänukit) ei ole säilinud, siis on tegemist‘tõeliste’ tertsiaarsete plastiididega.
Tertsiaarsete plastiididega protist on ka tsiliaatide hulkakuuluv fototroofne Mesodinium rubrum, mille rakk on täisendosümbiontseid krüptomonaade. Krüptomonaadid ise onjuba sekundaarsete plastiididega.
Plastiidi eellanePlastiidi eellane on vabalt elav sinivetikas ehk tsüanobak-ter. Sinivetikate puhul on iseloomulik, et fotosünteetilisedmembraanid ehk tülakoidid on paigutunud raku perifeerias(seda osa nimetatakse ka kromatoplasmaks); tülakoidid onüksikult (vastandina näiteks rohevetikate ja maismaataime-de plastiididele, kus tülakoidid on mitmekaupa koos lamelli-deks pakitud) ja paiknevad ükdteisest võrdsel kaugusel. Pig-mentatsioonilt on sinivetikatel klorofüll a ja fükobiliinid. Vii-mased on valgust koguvad pigmendid mis annavadki sinive-tikatele nende iseloomuliku värvuse. Fükobiliinid on veeslahustuvad, nad paiknevad spetsiaalsetes pigmendi ja valgukompleksides — fükobilisoomides. Fükobilisoome on kah-te tüüpi: poolketasjad (Joon. 29) ja poolkerajad (Joon. 30).
Fükobilisoomid paiknevad tülakoidide pinnal ja sellest tule-nevalt ei saagi tülakoidid olla lamellideks pakitud — tekibruumiline takistus.
Plastiidi genoomi redutseeritusKõige drastilisem muutus mis plastiididega on toimunud,võrreldes sinivetikast eellasega, on genoomi reduktsioon —genoomi suuruse ja geenide arvu vähenemine. Siin on täiestikohane paralleel mitokondrite genoomi reduktsiooniga. Näi-teks sinivetikas Synechocystis genoom on umbes 3,573 kbja selles on umbes 3200 geeni; punavetikas Porphyra pur-purea plastiidi genoom on ainult 191 kb ja umbes 250 geeni.Rohevetikate jamaismaataimede genoom on veelgi enam re-dutseerunud:Marchantia polymorpha (maksasammal) plas-tiidi genoom on vaid 121 kb ja sisaldab umbes 120 geeni.
Genoomi redutseeritus ja täielik sõltuvus peremeesor-
ganismist on see mis eristab plastiide endosümbionti-
dest.
Eri plastiidide genoomid on ka kaunis ebavõrdse genoomisuurusega. Laias laastus, rohevetikate ja maismaataimedeplastiidid on väiksema genoomiga (veidi üle 100 kb), pu-navetikate ja glaukotsüstofüütide plastiidide genoomid onsuuremad ja kodeerivad ka suuremat hulka valke kui rohe-vetikatel.Plastiidi proteiinid ei ole sugugi lihtsustunud või redut-
seerunud: eri hinnangutel on plastiidides 500–5000 erinevatvalku — igal juhul enam kui mistahes plastiidi geenide arv.Järelikult need valgud, mida ei kodeerita plastiidi genoomis,peavad olema kodeeritud tuuma genoomis ja transporditak-se seejärel plastiidi.Plastiidi genoomi reduktsioon (ja sama kehtib ilmselt ka
mitokondri genoomi kohta) on toimunud kolme protsessikaudu:
Geenide kadumine: Geenid, mille produktid on uutes tin-gimustes ebavajalik või koguni kahjulik, kaovad. Näitekssinivetika paks rakusein ilmselt takistaks ainevahetust or-ganelli ja raku vahel. Rakusein on kadunud ning tõenäoli-selt koos vastavate geenidega. Erandiks on siin glaukofüü-did, mille plastiididel on eellas-sinivetikale iseloomulikrakusein alles (vähemalt osadel liikidel).
15ENDOSÜMBIOOSI TEOORIA
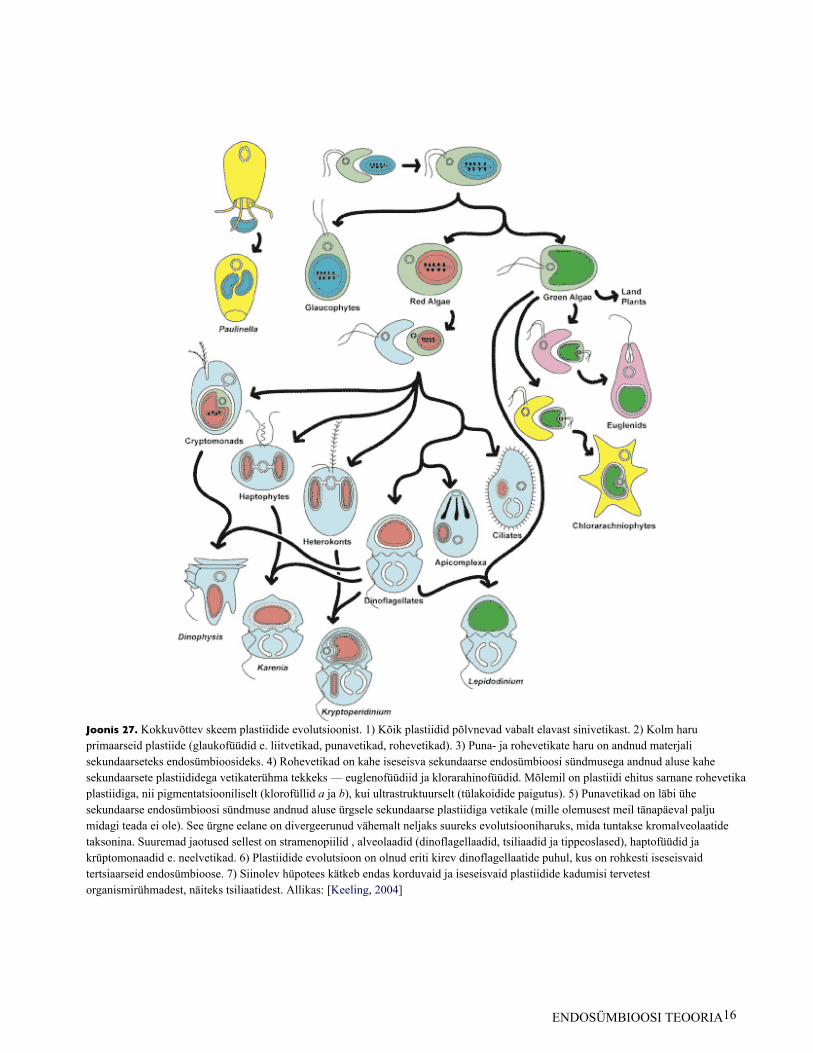
Joonis 27. Kokkuvõttev skeem plastiidide evolutsioonist. 1) Kõik plastiidid põlvnevad vabalt elavast sinivetikast. 2) Kolm haruprimaarseid plastiide (glaukofüüdid e. liitvetikad, punavetikad, rohevetikad). 3) Puna- ja rohevetikate haru on andnud materjalisekundaarseteks endosümbioosideks. 4) Rohevetikad on kahe iseseisva sekundaarse endosümbioosi sündmusega andnud aluse kahesekundaarsete plastiididega vetikaterühma tekkeks — euglenofüüdiid ja klorarahinofüüdid. Mõlemil on plastiidi ehitus sarnane rohevetikaplastiidiga, nii pigmentatsiooniliselt (klorofüllid a ja b), kui ultrastruktuurselt (tülakoidide paigutus). 5) Punavetikad on läbi ühesekundaarse endosümbioosi sündmuse andnud aluse ürgsele sekundaarse plastiidiga vetikale (mille olemusest meil tänapäeval paljumidagi teada ei ole). See ürgne eelane on divergeerunud vähemalt neljaks suureks evolutsiooniharuks, mida tuntakse kromalveolaatidetaksonina. Suuremad jaotused sellest on stramenopiilid , alveolaadid (dinoflagellaadid, tsiliaadid ja tippeoslased), haptofüüdid jakrüptomonaadid e. neelvetikad. 6) Plastiidide evolutsioon on olnud eriti kirev dinoflagellaatide puhul, kus on rohkesti iseseisvaidtertsiaarseid endosümbioose. 7) Siinolev hüpotees kätkeb endas korduvaid ja iseseisvaid plastiidide kadumisi tervetestorganismirühmadest, näiteks tsiliaatidest. Allikas: [Keeling, 2004]
16ENDOSÜMBIOOSI TEOORIA

Joonis 28. Elektronmikroskoobi läbilõige sinivetika rakust.Tülakoidid (fotosünteetilised membraanid) on raku perifeersesosas (nim. kromatoplasma), raku keskosas tülakoide ei ole; sinnaon koondunud DNA (nim. tsentroplasma). Tülakoidid on üksikult(mitte llamellideks pakitud), üksteisest võrdsetel kaugustel.Tülakoidide pinnal on fükobilisoomid.
FÜKOERÜTRIIN
TÜLAKOID
ALLOFÜKOTSÜANIIN
FÜKOTSÜANIIN
Joonis 29. Poolketasja fükobilisoomi skeem. Näidatud onfükobiliproteiinide – fükoerütriini, fükotsüaniini jaallofükotsüaniini paiknevus.
ALLOFÜKOTSÜANIIN
FÜKOTSÜANIIN
FÜKOERÜTRIIN
TÜLAKOID
Joonis 30. Poolkeraja fükobilisoomi skeem. Näidatud onfükobiliproteiinide – fükoerütriini, fükotsüaniini jaallofükotsüaniini paiknevus.
Geenide asendus: Geenid mis on olnud tuuma genoomisalgusest peale ja mis ekspresseeruvad tsütosoolis, võivaduues olukorras kodeerida produkte mis on plastiidi suuna-tud. Sellisel juhul on valgu plastiidi ja tsütosooli versioo-nid väga sarnased olnud, ehk isegi identsed.
Geenide ülekanne: Sellemehhanismi puhul on plastiidi geenesmalt duplitseerunud ja geeni üks koopiatest kandub ületuuma genoomi. Seejärel kaob plastiidi genoomis paiknevduplikaat.
Plastiidide värvikirevus
Maismaataimed on rohelised. Nagu ka rohevetikad, mille-st maismaataimed on evolutsioneerunud. Enamik vetikaidei ole rohelised. Pigem on tüüpiline vetika värvus kollakas-pruun. Miks nii?Lugu selles, et klorofüll a neelab valgust hästi spektri sini-
ses ja punases osas, kuid väga halvasti keskmises rohelisesosas. Mistap klorofüll paistabki roheline. Vees on aga peaalati valgust vähe, alati vähem kui veest väljas, maismaal.Sestap ei saa vetikad enamasti lubada endale luksust lastaroheline spektriosa raisku. Selleks on vetikatel kaaspigmen-did ehk antennpigmendid. Kaaspigmendid neelavad valgustselles spektriosas, kus klorofüll ebaõnnestub. Nii saab ve-tikas kasutada valgust kogu valguse spektri osas. Võis siis— suuremas osas sellest. Kaaspigmendid annavad valgus-energia edasi klorofüllile, mis kannab omakorda energia fo-tosünteesi reaktsioonitsentrile. Seda viimast toimetab ainultklorofüll a.
Klorofüll a on olemas kõigil oksügeenselt fotosünteesi-
vatel organismidela.
aValgust kasutavad energiaallikana ka mõned purpurbakterid.Neil on bakteriklorofüll, mitte klorofüll a ja nende fotosünteesitulemusena ei eraldu hapnik.
Kõik teised fotosünteetilised pigmendid, peale klorofpüllia on kaas- ehk antennpigmendid11. Kui kaaspigmente onrakus palju, varjutavad nad klorofülli ja me näeme vetikaidmuu värvilisena kui roheline.Laias laastus on pigmentide koostis vetikate rühma spet-
siifiline, mis kajastub ka vetikarühmade nimetustes. Näiteksrohevetikad, pruunvetikad, punavetikad, sinivetikad—neednimetused viitavad kõik peamistele pigmentide värvustele.Nagu näha, on pigmente ja värvust kasutatud vetikate klas-sifitseerimisel ja teatud mõttes on see õigustatud siiani:
• Rohevetikatel, eugleniididel ja klorarahinofüütidel on klo-rofüllid a ja b.
11Vetikatel ja ka maismaataimedel on ka muid pigmente kui fotosün-teetilised. Vahel võib sama pigment täita mitut eri rolli, näiteks mitmedkarotenoidid.
17ENDOSÜMBIOOSI TEOORIA

• Punavetikatel ja glaukotsüstofüütidel on klorofüll a ja fü-kobiliinid
• Haptofüütidel, fototroofsetel stramenopiilidel ja enamikuldinoflagellaatidel on klorofüllid a ja c;
• Krüptofüütidel on klorofüllid a ja c ning fükobiliinid.
Esmase fotosünteetilise eukarüoodi tekkimine toimus si-nivetika fagotroofse omastamise kaudu ürgse heterotroofseeukarüoodi poolt. Ilmselt oli esmase eukarüootse vetika pig-mentatsioon ja plastiid väga sarnane sinivetikast eellasega.Praegu on sinivetikale kõige sarnasem, nii pigmentatsiooniltkui ultrastruktuurilt punavetika plastiid.Glaukofüütide plastiid on ka väga sinivetikast eellase sar-
nane, aga glaukofüüdid on umbsopp plastiidide elvolutsioo-nis — nad ei ole teadaolevalt andnud alust ühelegi sekun-daarsele endosümbioosile.Rohevetikate plastiidid on tohutult erinevad — kadunud
on kõik fükobiliinid, juurde on tekkinud klorofüll b, tülakoi-did on pakitult, mitte üksikult. Seega juba arheplastiidseterühmas on plastiidide pigmentatsioon ja ehitus väga erinev.Sekundaarsete plastiididega vetikate pigmentatsioon va-
satab laias laastus nende plastiidi eukarüootse eellase pig-mentatsioonile. Eugleniidid ja klorarahinofüüdid on kaks rüh-ma, mis on oma sekundaarsed plastiidid saanud roheveti-kast ning pigmentatsioonilt on nad samuti erksalt rohelised(klorofüllid a ja b). Krüptomonaadid on oma plastiidi saa-nud punavetikast, ning neil on fükobiliinid, aga ’uuenduse-na’ on tekkinud klorofüll c. Stramenopiilid ja dinoflagellaa-did on samuti oma plastiidid saanud punavetikast, kuid füko-biliinid on kadunud ja lisandunud on korofüll c ja kollakas-pruunikaid kaaspigmente (peridiniin, fukoksantiin).Plastiidide ‘värvikirevus’ viis intuitiivselt mõttele, et kõik
need eri pigmentatsioonid on saadud erineva, vastava pig-mentatsiooniga sinivetika neelamisel. Seega, intuitiivselt võiksplastiide pidada polüfüleetilisteks [Raven, 1970].Tänapäeval on pigem enam tõendeid selle kasuks, et plas-
tiidid on monofüleetilised. Ehk teisisõnu — plastiidi pri-maarne endosümbioos on olnud unikaalne sündmus evolut-sioonis, see on toimunud vaid üks kord.Ainus teadaolev erand on Paulinella chromatophora, mis
on Cercozoa rühma kuuluv kodaamööb ega ole arheplas-tiidsetega kuidagi seotud, kuid kes on fotosünteesija (erine-valt teistest sama perekonna liikidest) ja kelle endosümbiont(või plastiid) on omandatud iseisvalt (joon. 27).
EukarüoodidEukarüootel on rida iseloomulikke tunnuseid,mis prokarüoo-tidel puuduvad. Kõige tuntum on membraaniga ümbritsetudrakutuum. Eukrarüootidele on omane raku kompartmentli-satsioon ja rakuorganellide olemasolu.Mitokondrid ja nende derivaadid. Kõigil eukarüootidelon mitokondrid, või mitokondrite taandarenenud derivaadid
—hüdrogenosoomid või mitosoomid (joon. 31). Hüdrogeno-soomides toimub anaeroobne ATP süntees, samas kui mito-kondrites toimub aeroobne respiratsioon. Mitosoomides eitoimu energia tootmist, küll aga on nad olulised rauda javäävlit sisaldavate proteiinide sünteesil.Plastiidid. Plastiidid on ainult osadel eukarüootidel ja needfotoünteetilised rühmad ei ole tingimata omavahel läheda-selt seotud (joon. 31).Primaarsed plastiidid. Plastiide on erinevat tüüpi. Esiteks—primaarsed plastiidid, mis on tekkinud vabalt elava sinive-tika endosümbioosi tulemusena, s.t. primaarse plastiidi eel-lane oli vabalt elav sinivetikas. Primaarseid plastiide ümbrit-seb kahemembraaniline plastiidikate. Primaarsed plastiididon arheplastiidsetel — glaukofüüdid Glaucophyta, puna-vetikadRhodophyta, rohevetikadChlorophyta ja strepto-füüdid Streptophyta (joon. 31). Pigmentidelt on primaar-sed plastiidid erinevad: glaukofüütide ja punavetikate plas-tiidides on säilinud sinivetikatele omased kaaspigmendid —fükobiliinid. Rohevetikatel ja sterptofüütidel (sh. maismaa-taimedel) on fükobiliinid kadunud, kuid on klorofüll b. Klo-rofüll a on kõigil.Sekundaarsed plastiidid. Sekundaarsed plastiidid on tekki-nud eukarüootse vetika ja eukarüootse heterotroofse protistiendosümbioosi tulemusena, s.t. sekundaarse plastiidi vahe-tu eellane oli priumaarsete plastiididega eukarüootne veti-kas. Sekundaarsed plastiidid on ümbritsetud kolme või nelja-membraanilise kattega. Alveolaatide, stramenopiilide, hap-tofüütide ja neelvetikate sekundaarsed plastiidid on tekkeltpunavetika raku endosümbioosi tulemus ning pigmentideston neil klorofüllid a ja c. Punavetikatele omased kaaspig-mendid, fükobiliinid, on kadunud kõigil, v.a. neelvetikatel.Eugleniidide ja klorarahinofüütide sekundaarsed plastiididon päritolult rohevetikad ja neil on klorofüllid a ja b, naguka rohevetikail endil (joon. 31).Tertsiaarsed plastiidid. Tertsiaarsed plastiidid on tekkinudsekundaarsete plastiididega vetika ja eukarüootse heterotroof-se protisti endosümbioosi tulemusena, s.t. tersiaarsete plas-tiidid vahetu eellane oli juba seunkdaarsete plastiididega eu-karüootne vetikas. Vaguviburlased on evolutsioonis olnuderakordselt andekad plastiide omastama ja just siin on paljutertsiaarsete plastiididega liike. Enamik fototroofseid vagu-viburlasi on punavetikast pärit plastiidiga, kuid mõned liigidon plastiidi saanud ka haptofüüdist, ränivetikast, neelveti-kast või rohevetikast.Osadel eukarüootidel on plastiidid sedavõrd redutseeru-
nud, et fotosünteesivõime on kadunud ja säilinud on vaidmõni üksik metaboolne funktsioon. Näiteks malaaria para-siidi Plasmodium (Apicomplexa) apikoplast ei ole enamfotosünteetiliselt tegev, kuid osaleb rasvhapete sünteesil.Kõik plastiidid, primaarsed, sekundaarsed, tertsiaarsed ja
ka apikoplastid pärinevad lõppkokkuvõttes ühest ühisest eel-lasest — vabaltelavast sinivetikast ja ühest unikaalsest pri-maarse endosümbioosi aktist. Plastiidide lai levik eukarüoo-
18EUKARÜOODID

tide hulgas on mitme iseseisva sekundaarse endosümbioositulemus.On üks tähelepanuväärne erand— Paulinella chromatop-
hora—-Cercozoa suurrühma kuuluv protist, kellel on kak-sikmembraaniga kaetud primaarsed plastiididmis pärinevadtäiesti iseseisvast primaarsest endosümbioosi aktist.Kui plastiididel on sekundaarsed ja tertsiaarsed endosüm-
bioosid, siis kõik mitokondrid põlvnevad ühest ja ainsastprimaarse endosümbioosi aktist.
Kiire lõppsõnaProtistid ei ole takson. Nagu ei ole takson ka ’selgrootud loo-mad’ või ’prokarüoodid’. Kõik need on negatiivsete tunnus-tega defineeritavad rühmad. Protistid on euakrüoodid, kesjäävad üle kui seened. taimed, loomad kui taksonid euka-rüootidest erialdada.Funktsiooni järgi on protistid jaotatavad fototroofseks ja
heterotroofseks pooleks, esimesi nimetame vetikateks, teistalgloomadeks. Ka need ei ole taksonid, vaid funktsionaalsedrühmad. Protistide hulgas on foto-ja heterotroofne energia-hange väga tihedalt läbi põimunud. Nii liigisiseselt kui kõr-gemate taksonite sees. Kuigi on ka valdavalt või ainult foto-ja heterotroofseid taksoneid.Protistid ei võrdu ainuraksetega. Algloomade poolel on
see nii, kuid vetikate hulgas on küllaga hulkrakseid, meetri-tega mõõdetavaid organisme.Protistide evolutsiooni olulised etapid on olnudmitokond-
ri ja plastiidi evolutsioon, mille käigus vabalt elav bakter(alfa proteobakter ja sinivetikas ehk tsüanobakter) muude-ti endosümiondiks ja seejärel rakuorganelliks. NB — euka-rüootide olulisimad funktsionaalsed omadused, aeroobne res-piratsioon ja oksügeense fotosünteesi võime on mõlemadbakteriaalset päritolu.
ViitedT. Cavalier-Smith. Endosymbiotic origin of the mitochond-rial envilope. In W.Šchwemmler and H. E. A. Schenk,editors, Endocytobiology II: intracellular space as an oli-gogenetic ecosystem, pages 265–280. De Gruyter, Berlin,1983.
Edouard Chatton. Titres et travaux scientifiques (1906-1937) de Edouard Chatton ... Impr. E. Sottano, Sète,1938.
P. J. Keeling. Diversity and evolutionary history of plastidsand their hosts. American Journal of Botany, 91:1481–1493, 2004.
L. Margulis. Origin of Eucaryotic Cells. Yale UniversityPress, New Haven, 1970.
L. Margulis. Symbiosis in cell evolution. Freeman, Chicago,1981.
C. Mereschkowsky. Über natur und ursprung der chroma-tophoren im pflanzenreiche. Biol. Zentralbl., 25:593–604,1905.
C. Mereschkowsky. Theorien der zwei plasmaarten alsgrundlage der symbiogenesis, einer neuen lehre der ents-techung der organismen. Biol. Zentralbl., 30:278–303,1910.
P. H. Raven. A multiple origin of plastids and mitochondria.Bot. L. Linn. Soc., 118:81–105, 1970.
J.Šapp. The Prokaryote-Eukaryote Dichotomy: Meaningsand Mythology. Microbiology and Molecular Bio-logy Reviews, 69(2):292–305, June 2005. ISSN1092-2172. doi: 10.1128/MMBR.69.2.292-305.2005.URL http://mmbr.asm.org/cgi/doi/10.1128/MMBR.69.2.292-305.2005.
A. W. F. Schimper. Über die entwicklung der chlorophyll-körner und farbkörner. Bot. Zeit., 41:105–114, 1883.
R. Y. Stainer. Some aspects of the biology of cells andtheir possible evolutionary significance. In H. P. Charlesand B. D. Knight, editors, Organization and control inprokaryotic and eukaryotic cells: 20th Symposium of theSociety for General Microbiology, pages 1–38. Cambrid-ge University Press, London, 1970.
R. Y. Stanier and C. B. Niel. The concept of a bacterium.Archiv für Mikrobiologie, 42(1):17–35, 1962. ISSN 0302-8933, 1432-072X. doi: 10.1007/BF00425185. URL http://link.springer.com/10.1007/BF00425185.
C. R. Woese and G. E. Fox. Phylogenetic structure of theprokaryotic domain: The primary kingdoms. Proceedingsof the National Academy of Sciences, 74(11):5088–5090,November 1977. ISSN 0027-8424, 1091-6490. doi:10.1073/pnas.74.11.5088. URL http://www.pnas.org/cgi/doi/10.1073/pnas.74.11.5088.
C. R. Woese, O. Kandler, and M. L. Wheelis. Toward anatural system of organisms: proposal for the domainsarchaea, bacteria, and eucarya. Proceedings of the Na-tional Academy of Sciences of the USA, 87:4576–4579,1990.
R. C. Woese. Default taxonomy: Ernst mayr’s view of themicrobial world. Proceedings of the National Academyof Sciences of the USA, 95:11043–11046, 1998.
19VIITED
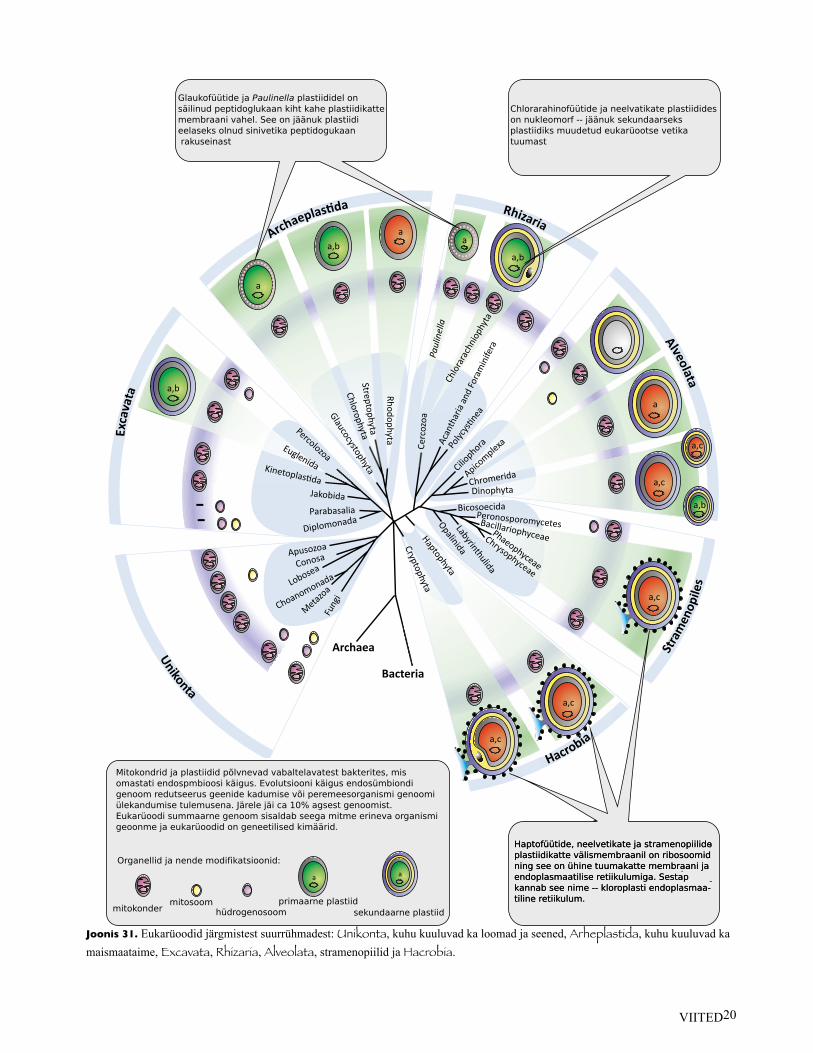
Glaukofüütide ja Paulinella plastiididel onsäilinud peptidoglukaan kiht kahe plastiidikattemembraani vahel. See on jäänuk plastiidi eelaseks olnud sinivetika peptidogukaanrakuseinast
Chlorarahinofüütide ja neelvatikate plastiidides on nukleomorf -- jäänuk sekundaarseksplastiidiks muudetud eukarüootse vetikatuumast
Haptofüütide, neelvetikate ja stramenopiilideplastiidikatte välismembraanil on ribosoomidning see on ühine tuumakatte membraani ja endoplasmaatilise retiikulumiga. Sestap kannab see nime -- kloroplasti endoplasmaa-tiline retiikulum.
Mitokondrid ja plastiidid põlvnevad vabaltelavatest bakterites, misomastati endospmbioosi käigus. Evolutsiooni käigus endosümbiondigenoom redutseerus geenide kadumise või peremeesorganismi genoomiülekandumise tulemusena. Järele jäi ca 10% agsest genoomist. Eukarüoodi summaarne genoom sisaldab seega mitme erineva organismigeoonme ja eukarüoodid on geneetilised kimäärid.
Haptofüütide, neelvetikate ja stramenopiilideplastiidikatte välismembraanil on ribosoomidning see on ühine tuumakatte membraani ja endoplasmaatilise retiikulumiga. Sestap kannab see nime -- kloroplasti endoplasmaa-tiline retiikulum.
Organellid ja nende modifikatsioonid:
mitokondermitosoom
hüdrogenosoomprimaarne plastiid
sekundaarne plastiid
Joonis 31. Eukarüoodid järgmistest suurrühmadest: Unikonta, kuhu kuuluvad ka loomad ja seened, Arheplastida, kuhu kuuluvad kamaismaataime, Excavata, Rhizaria, Alveolata, stramenopiilid ja Hacrobia.
20VIITED


















