
Журнал ЗЛОКАЧЕСТВЕННЫЕ ОПУХОЛИ MALIGNANT TUMOURS № 2-2015 (13) Русскоязычное издание


Л.А. АШРАФЯН, В.И. КИСЕЛЕВ
ОПУХОЛИ РЕПРОДУКТИВНЫХ ОРГАНОВ (ЭТИОЛОГИЯ И
ПАТОГЕНЕЗ)
Москва Компания «Димитрейд График Групп®»
2007

УДК 616 ББК 55.6
А98
Ашрафян Л.А., Киселев В.И. Опухоли репродуктивных органов (этиология и патогенез). М.: Изд-во Димитрейд График Групп, 2007. с. 216, ил. 28
ISBN 5-93620-035-3
Вниманию читателя предлагается монография, в которой авторы попытались рассмотреть механизмы канцерогенеза в органах репродуктивной системы с учетом последних данных, опубликованных в мировой литературе. Особое внимание уделено биохимическим аспектам формирования опухолей в гормон-чувствительных тканях, метаболическим путям женских половых гормонов. Детально рассмотрен метаболический синдром как предвестник канцерогенеза. Акцент на молекулярных факторах малигнизации обусловлен стремлением понять механизмы ранних стадий этих процессов, как с целью доклинической диагностики, так и для своевременной фармакологической коррекции. По существу, авторы попытались обосновать концепцию профилактической онкологии, которая призвана выработать новую стратегию по предупреждению онкологических заболеваний. Насколько это удалось, предстоит решить читателям. Но сегодня уже очевидно, что многие онкологические заболевания являются результатом образа жизни. Если это так, то в наших силах снизить риск их возникновения. Книга рассчитана на широкий круг специалистов, работающих в области гинекологии и онкологии.
Книга создана при поддержке ФГУ «Российский научный центр рентгенорадиологии» Росздрава и Научно-исследовательского
института молекулярной медицины ГОУВПО ММ А им.И.М.Сеченова
У Д К 616
ББК 55.6
© Ашрафян Л.А., Киселев В.И., 2007
© Компания «Димитрейд График Групп®», оригинал-макет, 2007

В медицине нет такой другой области, как онкология, где так демонстративно не вскрывалось бы противоречие между богатством фактического материала и хаосом теоретических представлений об этиологии и сущности опухоли. Обилие фактов, обилие теорий еще не создает подлинной теории.
Давыдовский И.В., 1962
ВВЕДЕНИЕ
Один из самых парадоксальных моментов, касающийся гормонозависимых опухолей у женщин, состоит в том, что пик заболеваемости практически всех опухолей репродуктивных органов приходится на возрастной интервал 55-65 лет. И, несмотря на этот факт, мы (онкологи) продолжаем упорно говорить о гиперэстрогении, как основной причине, приводящей к развитию опухолевого процесса. Сторонники гиперэстрогении могут возразить и аргументировать свою точку зрения реальными успехами антиэстрогенной терапии при раке молочной железы или эффективной гестагеннои терапии, например, при раке эндометрия. В этом случае, на наш взгляд, происходит подмена понятий, когда между гормоноиндуцируемостью и гормонозависимостью ставится знак равенства. Способность опухоли отвечать на гормонотерапию является следствием её генетической памяти, благодаря которой исходная эпителиальная клетка, даже трансформированная в опухолевую, продолжает синтезировать специфические рецепторы. Но этот эффект гормональной чувстви-

ВВЕДЕНИЕ
тельности, к сожалению, недолговечен, что свидетельствует об истощении пула гормоночувствительной популяции, с одной стороны, с другой, подтверждает точку зрения в отношении клеточной гетерогенности опухоли в рамках одного нозологического варианта.
Для того, чтобы понять закономерность сложного процесса или явления, необходимо максимально сузить компоненты анализа, формируя свое понимание наиболее общими и основными факторами, нанизывая впоследствии на основную модель возможные варианты. Исходя из этого, для нас определяющим был тезис о том, что спорадические варианты рака репродуктивных органов — это постменопаузальный рак, а закономерности канцерогенеза, в первую очередь, должны рассматриваться с позиций постменопаузального гомеостаза (обменного, энергетического, гормонального и пр.).
Современные достижения и накопленный за последнее десятилетие опыт в эпидемиологии, клинической онкологии, эндокринологии, молекулярной биологии, вирусологии обозначили ряд фактов, требующих нового осмысления. Нам кажется, что уже сегодня появилась возможность среди множества факторов и причин выделить основные, определяющие начальный этап (первый каскад) канцерогенеза спорадического рака репродуктивных органов.
Наше видение проблемы, безусловно, не является окончательным. Это лишь попытка рассмотреть проблему через призму современных достижений медицинской науки.

ЭПИДЕМИОЛОГИЯ РПКП РЕПРОДУКТИВНЫХ ОРГАНОВ
С конца 70-х годов прошлого столетия отмечена тенденция роста гормонозависимых опухолей репродуктивных органов. Эпидемиологической науке еще предстоит дать объяснение этому феномену. Но уже сегодня совершенно отчетливо обозначается взаимосвязь роста уровня онкологических заболеваний с нарастанием в популяции ждокринно-обменных нарушений. Еще совершенно не
давно многие исследователи считали, что рак в 70-90% вызывается факторами внешней среды (11). Этот вывод, прежде всего, основан на крупных эпидемиологических исследованиях, показывающих связь между многочисленными экзогенными факторами и частотой ряда локализаций злокачественных опухолей (курение — рак легкого, диета богатая жирами — рак толстой кишки, эндометрия, молочной железы и мн. др.). Кроме того, эпидемиологические исследования свидетельствуют о значительном различии в ряде стран в уровне заболеваемости различных типов рака. Эти различия определяются не столько генетическими факторами, сколько своеобразностью уклада жизни, характером диеты. Иллюстрацией к сказанному может служить исследование Johansson (1981), который приводит сравнительные данные, характеризующие мен-струально-генеративный анамнез популяции женщин Швеции, для которых характерна высокая заболеваемость ра-

ГЛАВА 1
ком молочной железы, эндометрия яичников, и популяции женщин Бангладеш с низкой заболеваемостью этими опухолями. У женщин Швеции менархе в среднем наступает в 12,8 лет, а в Бангладеш — в 17 лет. Соответственно менопауза в 51 и 42 года. При этом длительность репродуктивного периода для женщин Швеции составляет 38,2 года, а в Бангладеш — 25 лет при среднем числе родов 1,7 и 8 и длительности лактации 4 и 15 месяцев. Соответственно, число менструальных циклов, не закончившихся беременностью для женской популяции Швеции, составило более 400, а Бангладеш — менее 70.
Поэтому считают, что снижение уровня онкологической заболеваемости можно добиться за счет изменения образа жизни. Подобная точка зрения, вероятно, имеет право на существование.
Злокачественные опухоли репродуктивных органов женщин занимают особое место в клинической онкологии. Именно на модели рака шейки матки впервые четко обозначена этиологическая роль папилломовирусной инфекции, сформирована концепция метаболических нарушений при гормонозависимых злокачественных опухолях. При всем многообразии клинических и морфологических проявлений гинекологического рака можно отметить одну немаловажную особенность — у преимущественного числа больных злокачественный процесс развивается в постменопаузе. Эта особенность рака репродуктивных органов, по сути, обозначает и указывает на патогенетические истоки развития злокачественного процесса. В этом своеобразие гинекологического рака.
Рак вульвы составляет 4% от всех злокачественных опухолей половых путей. Это четвертый по частоте гинекологический рак (2). Большинство раков вульвы развиваются у пожилых женщин, и более чем 5 0 % случаев он

Эпидемиология рока репродуктивных органов
встречается у больных в возрасте от 60 до 79 лет. Лишь в единичных наблюдениях описаны случаи инвазивного плоскоклеточного рака у молодых женщин (12). Увеличение частоты заболеваний среди молодых женщин, возможно, связано с увеличением числа половых партнеров или венерическими инфекциями в популяции.
Более половины случаев (57,6%) рака шейки матки обнаруживаются у женщин в постменопаузальном возрасте, в 13,1% — в перименопаузе (4). Возрастная медиана находится между 50 и 55 годами. К сожалению, в статистику рака шейки матки включаются и все наблюдения с аде-нокарциномой шейки матки, что больше характерно для лиц молодого возраста и несколько искажает истинную статистику плоскоэпителиального рака шейки матки.
Рак эндометрия в структуре женской онкологической заболеваемости занимает четвертое место, составляя 6,5% (Чиссов В.И. с соавт., 2002). За последние три десятилетия во всех экономически развитых странах отмечен неуклонный рост этого заболевания. Почти все авторы отмечают, что 80-90% больных находятся в постменопаузе ( 1 , 2, 3, 5, 6, 7, 9, 10). С увеличением уровня заболевания доля молодых соответственно увеличивается. Однако только этим фактором объяснить сформировавшую тенденцию некоторого омоложения при этом патологическом процессе не совсем верно. Проблема рака эндометрия для пациенток репродуктивного периода должна рассматриваться совершенно в ином ключе и с позиции этиологии и патогенеза, и с позиции лечения, реабилитации и профилактики. В подавляющем проценте наблюдений рак эндометрия у молодых — это продолжение проблемы синдрома поликистозных яичников, в частности, несвоевременная диагностика его и неадекватное лечение.

ГЛАВА 1
Рак яичников составляет четверть всех случаев гинекологического рака, но он является наиболее частой причиной смерти среди всех онкогинекологических больных. В Соединенных Штатах рак яичников составляет 4% от всех раков у женщин, отставая по частоте от рака легкого, молочной железы, кишечника и матки. Среди женщин, умерших от рака, в 6% случаев причиной смерти является рак яичников, а среди умерших от рака женских половых органов почти в половине случаев (13).
Подавляющее большинство случаев рака яичника обнаруживается у женщин постменопаузального возраста и только в 10-15% наблюдений заболевание выявляется в пременопаузальном возрасте (13). Медиана возраста для наиболее часто встречающегося эпителиального типа рака яичников находится между 60 и 65 годами (13). Менее чем в 1 % эпителиальный рак яичников развивается у женщин в возрасте 30 лет или моложе и в большинстве случаев злокачественные опухоли у этих больных представлены герминомами (2, 8, 13). Около 20-30% новообразований яичников, обнаруживаемых у женщин в постменопаузе, являются злокачественными, в то время как у женщин пременопаузального возраста только 7% эпителиальных опухолей яичника являются злокачественными.
Анализ повозрастной заболеваемости гинекологическим раком свидетельствует, что наиболее интенсивно растет заболеваемость раком шейки матки в возрастных группах до 30 лет и 30-39 лет. При раке эндометрия рост заболеваемости имеет место в интервале 40-49 лет. При раке яичников тенденция роста заболеваемости не столь значима в молодом возрасте и равномерно распределена во всех возрастных группах (табл. 1)

Эпидемиология рока репродуктивных органов
Таблица 1 Динамика возрастных показателей заболеваемости
раком репродуктивных органов в России (на 100 000 женского населения) в 1990 и 2002 гг.
Год наблюдения
1990
2002
1990
2002
1990
2002
1990
[ 2002
Возраст, годы
До 30 30-39 40-49 50-59 60-69 70 и
более
Всего
Шейка матки
0,9
2,0
11,5
16,3
25,5
25,2
28,3
27,4
41,6
27,8
36,2
26,1
11,4 I
16,4 |
Эндометрий
0,2
0,2
2,9
3,5
12,6
16,7
36,1
47,9
47,0
62,9
32,0
45,5
18,6
28,5 |
Яичники
U 1,6
5,5
7,1
15,8
19,3
27,3
31,1
36,2
36,6
26,8
32,1
12,9
22,7
Молочная железа
11,7 16,2
62,5
81,2
76,5
103,4
135,7
180,5
189,8
250,6
126,9
179,7
49,8
89,1
Москва — мегаполис, где наиболее четко отлажена система учета онкологической заболеваемости вследствие хорошо отлаженной организационной структуры первичной медико-санитарной помощи (поликлиники, женские консультации, медсанчасти, диспансеры). Иными словами, в рамках Московского здравоохранения имеются все условия для объективного сравнительного анализа.
Анализ заболеваемости раком органов репродуктивной системы за период 1990-2002 гг. по Москве показал, что заболеваемость раком молочной железы увеличилась

ГЛАВА 1
на 67,7% (с 49,8 на 100 000 женского населения в 1990 году до 89,1 в 2002 году) (Рис.1). Аналогичная тенденция имела место и при раке эндометрия: заболеваемость увеличилась на 53,2% (с 18,6 в 1990 году до 28,5 в 2002 году).
При раке яичников отмечено резкое увеличение заболеваемости (на 76%): с 12,9 в 1990 году до 22,7 в 2002 году.
Наметившаяся тенденция к снижению заболеваемости раком шейки матки в 70-80-х годах отмечена медленным и достоверным ростом за период 1990-2002 гг. (с 11,4 до 16,4, что составило 43,9%) преимущественно за счет II-III стадий заболеваний (45%).
Таким образом, повозрастной анализ показывает, что основной пик заболеваемости раком репродуктивных органов приходится на постменопаузу с одновременным увеличением запущенных вариантов злокачественного процесса у пациенток молодого возраста. Иными словами, формирование этиопатогенетической концепции канцерогенеза рака репродуктивных органов у женщин невозможно без учета некоторых сторон гомеостаза пост-менопаузы.

С п и с о к л и т е р а т у р ы к Г л а в е 1
1 Ашрафян Л.А., Стандартизованная диагностика рака эндоме-I fpMH, Дисс. доктора мед. наук, Москва, 1989.
2 Бохман Я.В., Руководство по онкогинекологии, Ленинград, «Ме-I Дядина», 1989. с.463.
3. Вишневская Е.Е., Бохман Я.В., Ошибки в онкогинекологической I Практике, Минск, «Вышейшая школа», 1994, с. 287.
4. Злокачественные новообразования в России в 2002 году (забо-[ЯИаемость и смертность), Под ред. В.И.Чиссова, В.В.Старинского,
Ш.В Петровой. 5. Козаченко В.П., Рак матки, М.: Медицина, 1983, с. 163. 6. Комбинированное и комплексное лечение больных со злокаче-
стиенными опухолями. Под ред. В.И.Чиссова, М.:Медицина, 1989. 7. Руководство по эндокринной гинекологии. Под ред. Вихляевой
f М, M., 1997. 8. Berek J., Haker N., Ovarian and fallopian tubes, Cancer Treatment.
4ih ed. Philadelphia:Saunders, 1995, p. 295. 9. Creasman W.T., Endometrial carcinoma, Medicine Journal,
2001, vol. 2, №7. 10. Fuller A., Seiden M., Young R., Uterine cancer, 2004, London. 11. Higginson J., Muir С Environmental carcinogenesis: Misconceptions
and limitations to cancer control, J. Nat. Cancer Inst.,1979, vol. 63,p. 1291-
12. Iversen Т., Tretli S. Intraepithelian and invasive squamous cell neoplasia of the vulva, Obstet. And Gyn. 1998, 91:969-972.
13. Scully R., Young R., Clement P., Tumors of the ovary, Atlas of Tumor Pathology 3rd end.,Vol. 23.

эндокринологические особенности ПОСТМЕНОПАУЗЫ в Аспекте КАНЦЕРОГЕНЕЗА
Менопауза — это тот период жизни, когда вслед за утратой функции яичников полностью прекращаются менструации и такое состояние стабильно сохраняется в течение не менее года. Постменопауза следует за менопаузой и продолжается в среднем треть жизни женщины. Несмотря на многочисленные и целенаправленные исследования, касающиеся эндокринологии постменопаузы, многие аспекты этой проблемы до настоящего времени не совсем понятны.
Изменения, происходящие в гормональной сфере, как правило, формируются задолго до календарного наступления менопаузы. У большинства женщин за 2-8 лет до прекращения менструации начинает превалировать анову-ляция (89, 91). Изучение яичников человека показало, что подобная ускоренная утрата фолликулов у здоровых женщин начинается с 37-38 лет, когда количество достигает 25000 (60). Этот процесс коррелирует с небольшим, но явным увеличением количества фолликулостимулирующего гормона (ФСГ) и снижением ингибина. Ускоренная утрата фолликулов, вероятно, является вторичной по отношению к росту активности ФСГ. Эти изменения (включая повышение ФСГ) отражают неполноценность стареющих фолли-

Эндокринологические особенности постменопаузы...
кулов и снижение их способности к выработке ингибина, продукта гранулезных клеток, которые обеспечивают противодействие продукции ФСГ гипофизом. Изменения начинаются еще до появления ановуляции. Менструальные циклы удлиняются за счет увеличения фолликулярной фазы, ассоциируемой с повышением уровня ФСГ и снижением ингибина при нормальном содержании эстрадиола и лютеинизирующего гормона (ЛГ) (51, 87). В противоположность прежним представлениям (основанным на сообщении Sherman B.M. et al. (90)) уровень эстрадиола в годы, предшествующие менопаузе, сколько-нибудь заметно не снижается, а остается нормальным вплоть до прекращения роста и развития фолликулов. Если содержание ФСГ и ЛГ увеличивается за 2-3 года до наступления менопаузы, то уровень эстрогена начинает снижаться лишь за несколько месяцев до этого. Уменьшение количества ингибина, приводящее к подъему уровня ФСГ, отражает нарастающую недостаточность фолликулов (64, 80). Падение уровня ингибина начинается в возрасте около 35 лет, но значительно ускоряется после 40 лет. Четко установлено, что относительная неспособность эстрогена подавлять активность ФСГ после наступления менопаузы объясняется утратой ингибина и подтверждает его важную негативную роль в контроле выработки ФСГ.
Вскоре после наступления менопаузы можно однозначно утверждать об отсутствии фолликулов в яичниках (63). В конечном счете, уровень ФСГ повышается в 10-20 раз, Л Г — в 3 раза, достигая максимума через 1-3 года после наступления менопаузы, после чего содержание обоих гонадотропинов начинает постепенно, но незначительно снижаться. Уровень ФСГ выше, чем ЛГ из-за более быстрого удаления последнего из крови (время полувыведе-

ГЛАВА 2
ния ЛГ равно примерно 30 мин., а ФСГ — 4 часа) и, возможно, из-за отсутствия специфического пептида в качестве антагониста ЛГ, каким является ингибин.
После наступления менопаузы уровень андростендио-на в крови уменьшается примерно вдвое (82). Его секреция в постменопаузе осуществляется преимущественно надпочечниками и лишь в незначительной степени — яичниками. Концентрация тестостерона в этот период заметно не падает, причем у большинства (но не у всех) женщин яичники секретируют тестостерон даже больше, чем до менопаузы. После утраты фолликулов и, соответственно, эстрадиола, сохранившаяся строма яичников под воздействием повышенного содержания гонадотропинов начинает продуцировать больше тестостерона. Подавление гонадотропинов агонистами гонадотропин-релизинг гормона (Гн-РГ) или назначение его антагонистов женщинам в постменопаузе приводит к значительному снижению уровня тестостерона в крови, что указывает на его овари-альное происхождение под влиянием гонадотропина (46, 58). Однако общее количество тестостерона, продуцируемое после наступления менопаузы, уменьшается из-за ослабления периферической конверсии андростендиона, главного его источника. Снижение уровня тестостерона и андростендиона в период менопаузы невелико, составляя не более 15% (87). См. табл.2.
Таблица 2 Уровень стероидов в крови (мг/сут)
Гормон
Андростендион
Тестостерон
Эстрадиол
Репродуктивный период
2-3
0,2-0,25
0,35
Постменопауза
0,5-1,0
0,05-0,1
0,045

Эндокринологические особенности постменопоузы.
Уровень эстрадиола в крови после наступления менопаузы равен примерно 10-20 пг/мл (40-75 пмоль/л), и его продукция связана преимущественно с периферической конверсией андростендиона (67,68,82). Уровень эстрона в крови после менопаузы выше, чем содержание эстрадиола, достигая примерно 30-70 пг/мл (110-260 пмоль/л). Суточная выработка эстрогенов в этот период составляет в среднем 45 мг, при этом почти все количество поступает за счет периферической конверсии андростендиона. Соотношение андроген-эстроген после наступления менопаузы резко изменяется в результате более значительного падения уровня эстрогенов, и обычное в этот период развитие легкого гирсутизма отражает существенные сдвиги в гормональном профиле.
В процессе старения происходят определенные гисто-генетические изменения в яичниках. Под влиянием повышенного уровня лютеинизирующего гормона возникает гиперстимуляция тека-ткани яичников, что приводит к усиленному синтезу андростендиона. Дальнейшая его ароматизация происходит экстраовариально, преимущественно в жировой ткани
Выключение яичникового стероидогенеза включает ряд компенсаторных механизмов внеяичниковой продукции эстрогенов. Клиническое значение внегландулярных эстрогенов весьма широко, вариабельно и зависит от многих факторов. Уже сегодня вполне достоверно доказано, что превращение андростендиона в эстрон в процентном отношении коррелирует с весом женщин. Увеличение продукции постменопаузальных эстрогенов по мере нарастания веса связано с участием жировой ткани в процессе ароматизации андрогенов. Следовательно, вес тела положительно коррелирует с уровнем эстрона и

ГЛАВА 2
эстрадиола в крови (65). Следует заметить, что поступление эстрогенов через ароматизацию андрогенов не ограничивается только жировой тканью. Этим свойством обладают почти все ткани. См. рис.2.
Иными словами, в постменопаузе наряду с выключением овуляторной функции яичников, происходит мощное включение ряда компенсаторных механизмов, обеспечивающих достатоный уровень и, вероятно, весьма разнообразный спектр биологически активных продуктов (метаболиты). К сожалению, современная эндокринология еще далека от понимания того клинического эффекта, который сопряжен со взаимодействием постменопаузаль-ных эстрогенов и их метаболитов со специфическими рецепторами (альфа-, бета- и гамма-рецепторы). Вероятность такого взаимодействия очевидна. Дальнейшие исследования будут способствовать более глубокому пониманию этого важного механизма в постменопаузальной эндокринологии.
Варианты гормонального гомеостаза в постменопаузе
Основываясь на концепции Я.В. Бохмана о двух патогенетических вариантах развития рака эндометрия у женщин постменопаузального периода (13), целесообразно предположить, что гормонально-метаболические изменения у женщин в постменопаузе протекают по двум взаимно противоположным направлениям:
1) гипоэстрогения, с характерными для нее проявлениями (урогенитальная атрофия, атеросклероз, остеопороз),
2) гиперэстрогения с высоким уровнем гиперпластических процессов, гормонозависимых опухолей.

Эндокринологические особенности постменопаузы...
Таким образом, при первом варианте постменопаузы в популяции не должны встречаться гормонозависимые опухоли (рак молочной железы, рак эндометрия), в то время как при втором должно быть исключено развитие инволютивно-атрофических процессов. Однако практика показывает о значительном разнообразии сочетаний конституционального типа и заболеваний, возникающих в постменопаузе, что исключает возможность подобного четкого разделения (13, 28, 35, 84).
Оценка уровня гормонов в крови у женщин постменопаузального возраста, как показали исследования, не дает полной информации о возможных вариантах развития патологических процессов, так как для реализации гормонального эффекта необходимо присутствие достаточного уровня цитоплазматических рецепторов (11, 24, 25, 30, 31). Вместе с тем, при назначении заместительной гормональной терапии у женщин пери- и постменопаузального периодов не предусматривается определения уровня рецепторов, (б, 15, 16, 17, 43, 54, 77, 79).
Механизм действия половых гормонов на клетки был изучен благодаря открытию в 1960-1970 гг. рецепторов, специфических белков, посредством которых осуществляется гормональный ответ (11, 30, 31). Благодаря усовершенствованию методов анализа иммуноцитохимии, в 80-х годах было установлено существование только ядерных рецепторов (92).
Дальнейшие исследования показали, что имеются два основных типа рецепторов к стероидным гормонам:
- внутриклеточные ядерные рецепторы, - мембранные рецепторы.
Сродство рецепторов и стероидов определяет биологическую активность гормонов (11, 30,31, 92). При связы-

ГЛАВА 2
вании гормона с рецептором инициируется процесс транскрипции ядерной ДНК с образованием информационной РНК и последующим синтезом рибосомами белков, необходимых для усиленного роста и пролиферации.
Исследованиями последних лет установлено существование двух типов эстрогеновых рецепторов (ЭР-а, ЭР-(3). Оба типа рецепторов кодированы различными генами: ЭР-а находится на хромосоме 6, ЭР-р — на хромосоме 14. Оба подвида рецепторов обладают одинаковым сродством к эстрадиолу. Локализация рецепторов различна в разных органах и тканях — от наличия одного типа рецепторов до их сочетания (6). Так, эстроген-рецепторы (а и Р) обнаружены в мозге, кровеносных сосудах, сердце, костях, молочных железах, яичнике, матке. Только ЭР-р выявлены в легких, почках, мочевом пузыре, кишечнике. В матке и молочных железах доминируют ЭР-а. Результаты исследований рецепции эстрогенов, прогестинов показали, что с возрастом снижается реактивность тканей к действию гормонов, в связи со снижением числа специфических рецепторов в клетках (65).
Начальным этапом действия гормонов на клетки-мишени является его связывание со специфическим белком-рецептором. Если в клетке отсутствуют рецепторы, гормон не способен воздействовать на эту клетку (11, 23, 24, 25, 43). Следовательно, рецептор является необходимым периферическим звеном эндокринной функции, обеспечивающим возможность, интенсивность приема, проведения и реализации гормональных сигналов (30, 31). Все вышесказанное предполагает однозначный вывод для нормальной клетки: нет рецептора гормона — нет действия его на клетку. Клетка, в которой отсутствует рецептор к данному гормону-эффектору, не будет «отвечать»

Эндокринологические особенности постменопоузы
на воздействие этого гормона или его аналогов. Эти понятия и определили понятие ткани-мишени (клетки-мишени), где должны быть рецепторы к гормону, для которого данная клетка, ткань является мишенью (30, 31).
Иногда стероидным гормонам приписывается способность контакта с мембранными рецепторами. По этому вопросу имеется несколько работ. Авторы считают, что первые результаты изучения роли плазматических мембран в реализации гормонального сигнала в норме и при патологии в постменопаузе позволяют предположить участие мембранных компонентов в регуляции поступления тройных гормонов внутрь клеток-мишеней, а также «заинтересованность» этих компонентов в развитии гормонально-зависимых опухолей (33, 42). Однако представленная проблема о роли рецепторов плазматических мембран в реализации гормонального эффекта изучена недостаточно, что предполагает необходимость проведения дальнейших углубленных исследований.
До настоящего времени окончательно не получены данные о динамическом изменении уровня эстроген-рецепторов в неизмененных гормонозависимых тканях (молочной железы, вульвы, эндометрия) в зависимости от продолжительности постменопаузы.
Широкое использование современных технологий в медицине, в частности применение остеоденситометров для определения минеральной плотности костной ткани, позволило не только определять уровень минерализации кости, но и по-другому оценить тактику назначения заместительной гормональной терапии у женщин постмено-паузального периода с учетом возможного онкологического риска.

ГЛАВА 2
По современным представлениям, остеобласты и остеокласты содержат высокоспецифические рецепторы к эстрогенам, и в частности к эстрадиолу, и являются клетками — мишенями для этих гормонов (57, 59). Установлено наличие прогестероновых рецепторов на остеобластах и имеются сообщения об эффективности монотерапии гестеге-нами при снижении минеральной плотности костной ткани.
Исследования по этой проблеме показали, что костная ткань в постменопаузе может служить маркером по отношению к эстрогенам ( 1 , 2, 3, 4, 5).
Работы (73) показывают, что ЗГТ показана женщинам с низкой минеральной плотностью костной ткани, а высокий уровень минеральной плотности костной ткани является фактором риска развития рака молочной железы.
Необходимо отметить, что в отечественной литературе имеются единичные публикации о выявлении у пациенток постменопаузального периода высокой минеральной плотности костной ткани (28), что предполагает проведение дальнейших исследований по этому вопросу.
Среди возрастных эндокринно-обменных изменений, протекающих в постменопаузе проявлениями инсулино-резистентности, особое место занимает внегонадное эс-трогенообразование, которое служит источником дополнительной эстрогенной стимуляции (7, 9).
В последние годы появились доказательства, что нарушенная резистентность к инсулину, и гиперинсулинемия как фактор, усиливают ферментативную активность биосинтеза андрогенов (предшественников эстрогенов) в яичниках (45, 48, 50, 52, 62, 70, 74, 75,85, 86). Ожирение, как известно, встречающиеся при гиперандрогении, способствует снижению тканевой чувствительности к инсулину. Возникает инсулинорезистентность, которая ведет к возникновению компенсаторной гиперинсулинемии. Ин-

Эндокринологические особенности постменопаузы...
сулин опосредованно, через рецепторы различных факторов роста стимулирует яичники на продукцию андроге-нов, подобно ЛГ. Принципиальные механизмы, лежащие в основе гонадотропной активности инсулина, включают прямые эффекты на ферменты стероидогенеза, модуляцию рецепторной активности посредством ЛГ и ФСГ и стимуляцию избыточного синтеза эстрогенов.
Ожирение, как известно, часто сочетается с сахарным диабетом второго типа, артериальной гипертензией, ди-слипидемией, ишемической болезнью сердца. Частота и тяжесть сопряженных с ожирением нарушений зависят не только от степени ожирения (по индексу массы тела — ИМТ), но и особенностей отложения жировой ткани в организме (44, 50, 70, 75, 85). Проведенные исследования показывают, что андроидное ожирение, чаще чем гино-идное, сопровождается в значительной степени метаболическими нарушениями, увеличивает риск развития артериальной гипертонии, сахарного диабета 2 типа, атеро-склеротических заболеваний (26,75,69). Необходимо отметить, что ожирение и инсулинорезистентность с нарушениями в продукции стероидов расцениваются в последнее время как один из факторов, предрасполагающий к развитию рака молочной железы, эндометрия.
Научные исследования последних лет предполагают, что первичным дефектом, лежащим в основе развития ин-сулинорезистентности, является дисфункция эндотели-альных клеток сосудов (45, 76, 78, 85). Именно эти клетки первыми встречаются с реактивными свободными радикалами, с окисленными липопротеинами низкой плотности, с гиперхолестеринемией, с высоким гидростатическим давлением внутри выстилаемых ими сосудов (при артериальной гипертонии), с гипергликемией (при сахарном диабете).

Эндокринологические особенности постменопоузы.
Многочисленными исследованиями установлено, что основными гормонами в постменопаузе являются неклассические эстрогены и эстрон.
Согласно гипотезе, выдвинутой В.М. Дильманом (18, 19), в климактерическом периоде наблюдается повышение порога чувствительности по отношению к действию эстрогенов. Компенсаторное усиление функции яичников в течение определенного периода обеспечивает сохранение механизма обратной связи в этих условиях. В постменопаузе возникает сдвиг в спектре секретируемых гормонов, яичники начинают секретировать так называемые неклассические фенолстероиды. Последние обладают менее выраженным, чем классические эстрогены, ингиби-торным влиянием на гипоталамо-гипофизарную систему. Повышение продукции гонадотропных гормонов обусловливает гиперплазию тека-ткани яичников, что вероятно создает условия для повышенной продукции неклассических фенолстероидов (7, 8, 10).
Известно, что основным эстрогеном в постменопаузе является эстрон, концентрация которого в плазме крови женщин в этой возрастной группе в 3-4 раза выше, чем эстра-диола. Кроме того, эстрон образуется из андростендиона, который большей частью секретируется надпочечниками и в меньшей степени яичниками (20, 2 1 , 22, 32, 34, 37).
С возрастом секреция андростендиона в яичнике снижается и из общего количества циркулирующего в крови, 30% секретируется яичниками, 70% корой надпочечников. В ответ на снижение функциональной активности яичников, по механизму отрицательной обратной связи, повышается секреция ЛГ и ФСГ, снижается концентрация прогестерона и тестостерона. Уровень гонадотропинов достигает максимума через 10-15 лет постменопаузы и на-

ГЛАВА 2
Известно, что при гипергликемии в эндотелиальных клетках активируется фермент протеинкиназа-С, который увеличивает проницаемость сосудистых клеток для белков и нарушает эндотелий — зависимую релаксацию сосудов. При этом, гипергликемия активирует процессы перекисного окисления, продукты которого угнетают сосудорасширяющую функцию эндотелия. При артериальной гипертонии повышенное механическое давление на стенки сосудов приводит к нарушению архитектоники эндотелиальных клеток, повышению их проницаемости для альбумина, усилению секреции сосудосуживающего эн-дотелеина-1, ремоделированию стенки сосудов. Дисли-пидемия, в свою очередь, повышает экспрессию адгезивных молекул на поверхности эндотелиальных клеток, что дает начало формированию атеромы. Все перечисленные состояния, повышая проницаемость эндотелия, экспрессию адгезивных молекул, снижая эндотелий — зависимую релаксацию сосудов, способствуют прогресси-рованию атерогенеза.
Сторонники другой гипотезы считают, что в случае первичного дефекта эндотелиальных клеток трансэндотели-альный транспорт инсулина нарушается. Следовательно, может развиться состояние инсулинорезистентности. В этом случае инсулинорезистентность будет вторичной по отношению к эндотелиопатии (14, 27, 29, 45).
Таким образом, основываясь на вышеизложенных данных, можно сделать вывод, что, несмотря на низкие уровни циркулирующих в крови стероидных гормонов у пост-менопаузальных женщин, имеются все необходимые условия для избыточной продукции эстрогенов и стимуляции органов — мишеней.

ГЛАВА 2
на наш взгляд, способен более четко обозначить показания и противопоказания к этому виду терапии.
Для решения поставленных перед исследованием задач проведено изучение уровня эстроген-рецепторов в ткани молочной железы у 64 пациенток постменопаузаль-ного периода, имеющих картину инволютивных (т.е. соответствующих возрасту) изменений (табл. 3).
Таблица 3 Уровень эстроген-рецепторов (в %) в молочной железе у
пациенток в различные сроки постменопаузы
Продолжительность постмено
паузы
< 5лет (1 гр.)
5-15 лет (Игр.)
> 15 лет (III гр.)
Всего
Число пациен
ток
20
26
18
64
Уровень рецепторов
ЭР-
50,0
65,4
71,4
62,3
ЭР+
22,8
19,2
14,2
18,7
ЭР++
-
3,8
1,9
1,9
ЭР+++
27,2
11,6
12,5
17,1
р<0,005
Как это видно из таблицы, уровень рецепторов достоверно падает с увеличением продолжительности постменопаузы: Э Р + + + — с 27,2% до 12,5%. Следует отметить, что основной пик падения уровня рецепторов в ткани молочной железы наблюдается в течение первых 5 лет постменопаузы — более чем в 70% наблюдений: ЭР(-) 50,0%, ЭР(+) — 22,8%.
В последующем снижение количества рецепторов продолжается, но уже не столь интенсивно. В целом можно констатировать, что основные изменения в рецепторном статусе в эпителии ткани молочной железы на этапе пост-

Эндокринологические особенности постменопаузы...
менопаузы формируются в течение 5 лет с момента выключения функции яичников. Об этом свидетельствует анализ нашего материала: достоверных различий в рецеп-торном статусе между показателями II и III групп практически нет. Немаловажен для нас и тот факт, что у значительной части пациенток (12,5%) ткань молочной железы сохраняет высокий уровень эстроген-рецепторов даже по прошествии 15 лет и более постменопаузы.
Достаточно интересные данные были получены при изучении эстроген-рецепторов в ткани вульвы (табл. 4).
Таблица 4 Уровень эстроген-рецепторов (в %) вульвы у пациенток в
различные сроки постменопаузы
Продолжительность постмено
паузы
< 5лет (I гр.)
5-15 лет (Игр.)
> 15 лет (III гр.)
Всего
Число пациен
ток
20
26
18
64
Уровень рецепторов
ЭР-
45,4
24,1
25,0
31,5
ЭР+
16,2
40,2
33,3
28,6
ЭР++
2,2
4,7
8,4
6,4
Э Р + + + I
36,2
31,0
33,3
33,5
р<0,005
Следует отметить, что к моменту формирования стойкой постменопаузы и далее на протяжении всей постменопаузы число наблюдений, где отмечены высокие концентрации эстроген-рецепторов, стабильны (соответственно: 36,2%, 31,0%, 33,3%). Можно достаточно убедительно констатировать, что нет резких колебаний в динамике концентрации в ткани вульвы эстроген-рецепторов с момента полного выключения эстроген продуцирую-

Эндокринологические особенности постменопаузы...
Как удалось установить, основной пул рецепторов сконцентрирован в базальном слое. По мере увеличения продолжительности постменопаузы отмечено снижение числа наблюдений с Э Р + + + с 36,2% (менопауза < 5 лет), до 27,6% (менопауза 5-15 лет) и 8,3 (менопауза > 15 лет). Следует обратить внимание на такой феномен, что по мере увеличения продолжительности постменопаузы отмечено появление Э Р + + + и в базальном слое, и дерме (менопауза 5-15 лет) в 3,4% наблюдений. Далее, по прошествии 15 лет постменопаузы, в 16,7% наблюдений рецепторы имеются и в дерме, а в 8,3% и в базальном слое, и дерме. Для интерпретации этих данных требуются специальные и целенаправленные исследования.
Следующая ткань-мишень, которая была изучена с позиций установления уровня Э Р + + + в постменопаузе — это атрофичный эндометрий (табл. 6).
Таблица 6 Уровень эстроген-рецепторов (в %) в эндометрии у
пациенток в различные сроки постменопаузы
Продолжительность постмено
паузы
< 5лет (I гр.)
5-15 лет (Игр.)
> 15 лет (III гр.)
Всего
Число пациен
ток
20
26
18
64
Уровень рецепторов
ЭР-
33,3
45,8
55,6
44,9
ЭР+
16,7
25,0
22,2
21,8
ЭР++
13,9
12,5
11,1
11,5
ЭР+++
36,1
16,7
11,1
21,8
р<0,005

ГЛАВА 2
щей функции яичников. Создается впечатление, что состояние рецепторного аппарата в этом случае зависит не столько от уровня эстрадиола, продуцируемого яичниками, сколько, вероятно, генетически детерминировано и в большей степени является отражением программы старения клетки. И вновь необходимо отметить, что практически у трети пациенток ткань вульвы сохраняет высокий уровень концентрации рецепторов (ЭР+++) независимо от продолжительности постменопаузы.
Следует сказать, что, когда речь идет о вульве, как органа-мишени, вероятно, необходимо четко представлять те конкретные зоны, которые ответственны за реализацию гормонального эффекта. Иными словами, в каком из слоев вульвы в клетках сконцентрированы эстроген-рецепторы. Изучение этого вопроса потребовало детализации уровня эстроген-рецепторов в базальном слое, дерме, а также учета возможного сочетания их в различных слоях вульвы (табл. 5).
Таблица 5 Содержание эстроген-рецепторов в
различных слоях вульвы
Продолжительность постмено
паузы
< 5лет (1 гр.)
5-15 лет (Игр.)
> 15 лет (III гр.)
Всего
Число пациен
ток
20
26
18
64
ЭР>+++
Баз. слой
36,2
27,6
8,3
24,0
дерма
-
-
16,7
5,6
Баз. слой
-Ьдерма
-
3,4
8,3
3,9
ЭР++ ЭР+ ЭР-
63,8
69,0
58,4
66,5
» р<0,005

ГЛАВА 2
Как видно из данных таблицы, темпы падения уровня эстроген-рецепторов в эндометрии на протяжении всей постменопаузы в среднем равномерны. Несколько опережающими темпами идет снижение концентрации рецепторов на начальных этапах постменопаузы, далее процесс снижения приобретает более равномерный характер и после 15-летнего периода только в 11,1% наблюдений в эндометрии сохраняется высокая концентрация эстроген-рецепторов. И вновь, даже после длительного периода постменопаузы, у части популяции пациенток в атрофич-ном эндометрии сохраняется достаточно высокий уровень эстроген-рецепторов. Как видим, в представленном материале прослеживается ряд весьма интересных моментов, требующих своего осмысления. Необходимой частью анализа является сравнение наблюдений, где имели место высокие концентрации эстроген-рецепторов в различных тканях-мишенях в популяции пациенток постмено-паузального периода (табл. 7).
Таблица 7 Сравнительные данные уровня ЭР+++ (в %) в тканях-
мишенях у пациенток в различные сроки постменопаузы
Продолжительность постмено
паузы
< 5лет (I гр.)
5-15 лет (Игр.)
> 15 лет (III гр.)
Число пациенток
19
15
10
ЭР+++
Молочная железа
31,8
11,6
12,5
Вульва
36,2
31,0
33,3
Эндоме- ] трий
36,1
16,7
11J
р<0,005

Эндокринологические особенности постменопаузы...
Сравнительный анализ показывает, что имеется определенный синергизм в показателях ЭР+++ в молочной железе и эндометрии. В обеих группах с увеличением продолжительности постменопаузы отмечается синхронное снижение числа пациенток с высокими концентрациями эстроген-рецепторов, в то время как в тканях вульвы количество наблюдений с ЭР+++ остается практически одинаковым. Исходя из этих данных, весьма допустима следующая точка зрения. Вероятно, в циклически функционирующих тканях-мишенях рецепторный аппарат клетки более зависим от яичниковых стероидов, и зависимость эта в последующем в постменопаузе является одной из главных причин, формирующих интенсивность и темпы падения концентрации эстроген-рецепторов в клетке. Там же, где ткани-мишени не столь зависимы от циклической работы яичников, механизм снижения уровня концентрации стероидных рецепторов, скорее всего, обусловлен функциональной дедифференцировкой клетки, что в определенном смысле является отражением реализации программы старения на клеточном уровне. Естественно, это рабочая гипотеза.
Завершая этот фрагмент работы, необходимо отметить, что с началом постменопаузы в клетках тканей-мишеней происходит достаточно интенсивное снижение концентрации эстроген-рецепторов. Наряду с этим, у части пациенток сохраняется наличие высоких концентраций эстроген-рецепторов вне зависимости от продолжительности постменопаузы. Иными словами, если учиты-
ать рецепторный статус тканей-мишеней у пациенток в остменопаузе, то можно вполне четко дифференциро-ать два направления:

ГЛАВА 2
- при первом происходит достаточно интенсивное снижение концентрации эстроген-рецепторов в среднем в течение первых 5-10 лет постменопаузы;
- при втором, вне зависимости от продолжительности постменопаузы, в тканях-мишенях сохраняется высокая концентрация эстроген-рецепторов.

Эндокринологические особенности постменопаузы...
Список литературы к Главе 2
1. Ашрафян Л.А., Титова В.А. и др. Двухфотонная рентгеновская аб-
сорбциометрия в дифференцировании типа постменопаузы. Тезисы
докл. научн. конф. «Лучевая диагностика и лучевая терапия на пороге
третьего тысячелетия», Радиология, 2000, с. 49.
2. Ашрафян Л.А., Харченко Н.В. и др. Изменения минеральной
плотности костной ткани скелета и гормонального статуса у пациенток
после хирургической и лучевой кастрации. Высокие технологии в онко
логии. Материалы V Всероссийского съезда онкологов. Казань. 2000,
ее. 416-17.
3. Ашрафян Л.А., Харченко Н.В. и др. Результаты периферической
денситометрии при популяционном обследовании женщин в постмено
паузе. Вопросы онкологии, 1999, Т. 45, № 1, с. 49-52.
4. Ашрафян Л.А., Харченко Н.В., Акопова Н.Б. и др. Изменение по
казателя минеральной плотности костной ткани в постменопаузе, Ма
териалы VIII Всероссийского съезда рентгенологов и радиологов «Ал
горитмы в лучевой диагностике и программы лучевого и комплексного
лечения больных», Москва, 2001, с. 14.
5. Ашрафян Л.А., Харченко Н.В., Акопова Н.Б., Ивашина СВ. Четы
ре патогенетических варианта изменений в половых органах у женщин
в менопаузе. Климактерий, 2001, № 3, с. 7.
6. Балан В.Е. Качество жизни у женщин с урогенитальными рас
стройствами в климактерии, Тезисы II съезда Всероссийской ассоци
ации акушеров-гинекологов «Гормонозаместительная терапии за и
против», Москва, 1997, с. 10.
7. Берштейн Л.М. Внегонадная продукция эстрогенов, СПб, Наука,
1998.
8. Берштейн Л.М. Выделение классических эстрогенов и суммар
ных феностероидов у больных раком тела матки, Автореф. дисс. канд.
мед. наук, Л, 1967.

Эндокринологические особенности постменопаузы
22. Исследование проблем менопаузы в 90-х годах: Доклад на-
уч.гр. ВОЗ:866, М, Медицина, 1996.
23. Кушлинский Н.Е., Герштейн Е.С. и др. Цитоплазматические ре
цепторы стероидных гормонов в опухолях пищевода, Вестник ОНЦ
АМН России, 1996, №3, с. 42-46.
24. Кушлинский Н.Е., Клименков А.А., Абрамов СБ. и др. Прогресс
в лечении злокачественных новообразований, Материалы конферен
ции, Чимкент, 1991, с. 198-200.
25. Кушлинский Н.Е., Соловьев Ю.Н., Галкина Н.М., Алиев М.Д.,
Березов ТТ., Трапезников Н.Н. Рецепторы эстрадиола 17-бета в цито-
зольной фракции хондросарком и прогноз заболевания, Вестник ОНЦ,
1999, №4, с. 56-59.
26. Липпманн М. Взаимодействие гормонов с рецепторами, М,
1979, с. 213-234.
27. Моисеев B.C., Ивлева А.Я., Кобалава Ж.Д. Гипертония, сахар
ный диабет, атеросклероз — клинические проявления метаболическо
го синдрома X, Вест. Росс. акад. мед. наук, 1995, с. 15-18.
28. Насонов Е.Л. Остеопороз и остеоартроз: взаимоисключающие
или взаимодополняющие болезни? Consilium medicum, 2000. Т 2. № 6.
ее. 248-250.
29. Один В.И. Нарушения жиро-углеводного обмена и надпочечни-
ковые стероиды у лиц с возрастными нарушениями толерантности к
глюкозе и у онкологических больных, Автореф. Дисс. Канд. мед. наук,
Спб, 1994.
30. Рецепторы стероидных гормонов в опухолях человека, под ре
дакцией Бассалык А.С., Москва. Медицина, 1987.
31. Розен В.Б. Физиология гормональной рецепции, Ленинград,
1986, ее. 5-33.
32. Савельева Г.М., Сергеев П.В., Бреусенко В.Г. и др. Рецепторы
эстрогенных плазматических мембран эндометрия при пролифератив-
ных процессах в постменопаузе, Акуш. и гин., 1989, № 12, ее. 60-63.

ГЛАВА 2
9. Берштейн Л.М. Гормональный канцерогенез, СПб, 2000, с. 133-
138.
10. Берштейн Л.М. Эстрогены, старение и возрастная патология,
Усп. герант., 1998, т. 2, с. 90-97.
11. Болье Э.Э. Рецепторы клеточных мембран для лекарств и гор
монов. Междисциплинарный подход, Москва. 1983, ее. 142-165.
12. Борисов С.Н. Изменения минеральной плотности костной ткани
скелета и гормонального статуса у пациенток, перенесших оофорэкто-
мию, дис. к.м.н., М, 1999.
13. Бохман Я.В. Руководство по онкогинекологии, Ленинград. Ме
дицина, 1989, с. 464.
14. Бутрова С.А. Метаболический синдром: патогенез, клиника, ди
агностика, подходы к лечению, Русский мед.журн, 2001, т. 9, № 2.
15. Вихляева Е.М. Постменопаузальный синдром и стратегия заме
стительной гормональной терапии, Акушерство и гинекология, 1997,
№5, ее. 51-56.
16. Гудкова М.А., Кулаков В.И., Крымская М.Л., Соловьева А.Д.
Клинико-физиологическая характеристика я гормонотерапия больных
с типичной формой климактерического синдрома, Акушерство и гине
кология, 1994, №6, с. 49-52.
17. Гуменюк Е.Г., Самородинов Л.А., Цырлина Е.В. Гормональный
статус больных с гиперпластическими изменениями эндометрия и кри
терий выбора метода гормональной терапии дисфункциональных ма
точных кровотечений, Вопросы онкологии, 1999, 45, № 2 с. 147-152.
18. Дильман В.М. Старение, климакс, рак, Л, Медицина, 1968.
19. Дильман В.М. Эндокринологическая онкология, Л, 1983.
20. Зайдиева Я.З. Гормональное лечение женщин в постменопау
зе. Проблемы репродукции, 1996, № 3, ее. 38-43.
21. Зайдиева Я.З. Гормонопрофилактика и коррекция системных
нарушений у женщин в перименопаузе, Автореф. дисс. докт. мед.
наук, М, 1997.

Эндокринологические особенности постменопаузы...
43. Хаит ОБ. Клинико-иммунологические аспекты комплексного
лечебного воздействия при гиперпластических процессах эндо-и мио-
метрия, Автореф. док. мед. наук, Харьков, 1990.
44. Шестакова М.В. Дисфункция эндотелия — причина или след
ствие метаболического синдрома? Русский мед. журн., 2001, т. 9, № 2.
45. Шлянкевич М.А. Вестник академии медицинских наук СССР,
1973, № 4 , ее. 82-89.
46. Andreyko J., Monroe S. Concordant suppression of serum immuno-
reactive LH, J.Clin. Endicrinol.Metab., 1992, 74:399.
47. Barlow D.H. Towards better recognition of urogenital ageing, Swit
zerland, 1995,15-18.
48. Bjorntorp P. Adipose tissue distribution and function, Int.J. Obes,
1991, sep.15(2), p. 67-81.
49. Bone Markers: What,s Their Place? Special Report, The Genesis
Report, 1996, Dx, Nov., USA.
50. Bruning P.F., Bonfrer J.M.G., van Noord P.A.H., Nooijen W.J. Insu
lin resistance and breast cancer risk, Int.J.Cancer, 1992, vol. 52, p. 511-516.
51. Buckler H., Evans A., Mamlora H., Bulgier H. Gonadotropin, steroid
and inhibin levels in women with incipient ovarian failure during anovulatory
and ovulatory «rebound» cycles, J.Clin. Endocrinol. Metab, 1991, 72:116.
52. Campos H., McNamara J.R., Wilson P.W. Differences in low density
lipoprotein subfractions andapolipoproteins in premenopausal and postme
nopausal women, J.Clin. EndocrinolMetab., 1988, 67:30.
53. Colditz G.A., Egan K.M., Stampfer M.J. Hormone replacement thera
py and the risk of breast cancer; results from epidemiologic studies, Am.J.
Obstet.Gynaecol, 1993,168, pp1473-1480.
54. Colditz G.A., Hankinson S.E, Hunter D.J. et al. The use of estrogen
and progestins and risk of breast cancer in postmenopausal woman, N. En
glish J. Med., 1995,1589-93.
55. Coop J. Hormonal and nongormonal interventions for menopausal
symptoms, Maturitas, 1996; No 23,159-168.

ГЛАВА 2
33. Сафроний Д.Ф., Цырлина Е.В. и др. Динамика изменений уров
ня цитоплазматических рецепторов эстрадиола и прогестерона в опу
холевой ткани рака эндометрия в группе больных, чувствительных и
нечувствительных к гормонотерапии тамоксифеном и его сочетании с
ОПК, Тезисы Всесоюзного симпозиума «Скрининг в раннем выявлении
опухолей репродуктивной системы и проведение органосохраняющего
лечения». Кострома, 1991.
34. Серебренникова К.Г. Дисгормональная патология эндометрия
в периоде постменопаузы (патогенез, клиника, диагностика, лечение),
Автореф. дисс. док. мед. наук, СПб, 1992.
35. Серебренникова К.Г., Копасова Т.Л., Кузьмина З.В. Рецепторы
стероидных гормонов в комплексной оценке состояния эндометрия и
молочных желез у больных внутренним эндометриозом, Тезисы докла
дов VI съезда акушеров-гинекологов РСФСР, Новосибирск, 1987,
с. 219-220/
36. Сметник В.П. Климактерические расстройства и принципы
заместительной гормональной терапии, Терапевт. Архив, 1995,
№10, ее. 70-74.
37. Сметник В.П. Лечение и профилактика климактерических рас
стройств, Клиническая фармакология, 1997, № 6(2), ее. 86-91.
38. Сметник В.П. Принципы заместительной гормонопрофилактики
и терапии климактерических расстройств. Проблемы пери- и постмено-
паузального периода, Материалы симпозиума, М, 1996, се. 72-75.
39. Сметник В.П. Системные изменения у женщин в климактерии,
Русский медицинский журнал, 2001, Т. 9, № 9.
40. Сметник В.П. Состояние нейроэндокринной системы при син
дроме истощения яичников и при климактерическом синдроме, Авто
реферат дисс. д.м.н., М, 1980.
41. Сметник В.П., Тумилович Л.Г. Неоперативная гинекология, М,
1997.
42. Таджибаева Ю., Нейштадт Э.Л. Койлоцитоз при краурозе и раке вульвы, Арх. Патологии, 1990 г., т 52, 11, с. 17-20.

Эндокринологические особенности постменопаузы
68. Jadd H., Shamonki I., Frumar A. Origin of serum estradiol in postme
nopausal women, Obstet. Gynecol, 1982, 59:680.
69. Kaaks R., Lukanova A. Effects of weight control and physical activi
ty in cancer prevantion: role of endogenous hormone metabolism, Int. Agen
cy for Research on Cancer, France.
70. Kaplan J. et al. Atherogenic and protective effects of sex steroids on
the monkey cardiovascular system, Post. Med., 1989, pp. 69-73, discussion
pp. 89-90.
71. Kaufman J.M. Pharmacokinetics of oestrogens and hormone repla
cement therapy, Gur. Menopause J., 1997; No 4, p. 14-22.
72. Khan S.A., Rogers M.A.M., Khurana K.K. et al, Estrogen receptor ex
pression in bening breast epithelium and breast cancer risk, J.Nat.Cancer
Inst, 1998, vol. 90, p.37-42.
73. Kuhl H. Pharmacokinetics of oestrogens and progestagens, Maturi-
tas, 1990; No 12, p. 171-197.
74. Lacey J., Swanson C, et al. Obesity as a potential risk factor for ade
nocarcinoma and squamous cell carcinomas of the uterine cervix, Cancer,
2003, Aug. 15; 98(4):814-21.
75. Lange C.A. Richer J.K., Horwitz K.B. Hipothesis: progesterone pri
mes breast cancer cells for cross-talk with proliferative or antiproliferative sig
nals, MoI.Endocrinol., 1999,13, p. 829-836.
76. Lin S.C., Yamate Т., Taguchi Y. et al. Regulation of the gp 80 and gp
130 subunits of the IL-6 receptors by sex steroids in the murine bone mar
row, J. Clin. Invest., 1997,100(8), 1980-90.
77. Longscope C, Bourget C, Flood С The production and aromatiza-
tion of dehydroepiandro-sterone in postmenopausal women, Maturitas,
1982,3, p. 325-332
78. Longscope C, Hunter R., Franz C.Steroid secretion by the post-me-
nopausal ovary, Am.J .Obstet.Gynecol., 1980,138, p. 564-68.
79. Lover R.R, Mazess R.B. Barden H.S. et al. Effects of Tamoxifen on
bone marrow density in postmenopausal women with breast cancer, N.
Engl.J.Med., 1992, 336, p. 852-856.

ГЛАВА 2
56. Cordozo L, Rekers H., Tapp A. Estriol in the treatment of postmeno
pausal urgency: a multycentral stady, Maturitas, 1993; No 18(1), 47-53.
57. Cummings S.R., Black D.M., Rubin S.M. Lifetime risk of hip. Colins
or vertebral fracture and coronary heart disease among white postmenopau
sal women, Arch. Intern. Med., 1989,149, 2445-48.
58. Dowsett M., Cantwell В., Anshumala L. Suppression of postmeno
pausal ovariansteroidogenesis with the luteinizing hormone-releasing hor
mone agonist goserelin, J.Clin. Endocrinol. Metab., 1988, 66:672.
59. Eiken P. Continuous combined Hormone Replacement Therapy for
osteoporosis 10 yeardata on Kliogest//Eur. Menopause J., 1996,3,141-146.
60. Faddy M., Gosden R., Gougeon A. Accelerated disappearance of
ovarian follicles in mid-life: implications for forecasting menopause, Hum.Re-
prod.,1992, 7:1342
61. Finch С The regulation of physiological changes during mammalian
aging, Q. Rev. Biol., 1976, 51(1), p. 49-83.
62. Gamajunova V.B., Bobrov Ju.F. et al Comparative study of blood in
sulin levels in breast and endometrial cancer, Neoplasma, 1997, vol. 44,
p. 123-126.
63. Gosden R. Follicular status at menopause, Hum. Reprod., 1987,
2:617.
64. Нее J., MacNaughton J., Banhag M. Perimenopausal patterns of go
nadotropins, immunoreactive inhibin, estradiol and progesterone, Maturitas,
1993,18:9.
65. Hess G., Roth G.S. Changes in mechanisms of hormone and neuro
transmitter action during aging: current status of the role receptor and postre-
ceptor alteration, Mech. Ageing Dev., 1982, Nov. 20(3), 175-194.
66. Holland J.F., Frei E. Cancer Medicine, Philadelphia: Lea and Febi-
ger, 1973.
67. Jadd H., Jadd G., Lucas W. Endocrin function of the postmenopau
sal ovary; concentration of androgens and estrogens in ovarian and periphe
ral vein blood, J.Clin. Endocrinol. Metab, 1974, 39:1020.

МЕТАБОЛИТЫ ЭСТРПДИОЛП И ПАТОЛОГИЧЕСКИЕ ПРОЦЕССЫ В РЕПРОДУКТИВНОЙ СИСТЕМЕ
Эстрогены: структура, место синтеза, роль в физиологии и патофизиологии (канцерогенезе)
По своей химической природе женские половые гормоны эстрогены (от греч. oistros — страстное влечение) являются С18-стероидами с ароматическим кольцом и ги-дроксильной группой у третьего углеродного атома. Существуют три физиологически важных эстрогена: эстрон (Е1), имеющий кетогруппу в положении С-17; эстрадиол (Е2), обладающий гидроксильными группами в положениях С-3 и С-17, и эстриол (ЕЗ) с гидроксильными группами в положениях С-3, С-16 и С-17 (обозначения Е1, Е2 и ЕЗ даны в соответствии с номерами гидроксильных групп, связанных со стероидным кольцом).
В организме взрослой женщины эстрогены вырабатываются фолликулами яичников, плацентой (во время беременности) и частично корой надпочечников. Яичник се-кретирует вдвое больше эстрадиола, чем эстрона. При этом эстрон обладает лишь десятипроцентной эстроген-ной активностью по отношению к эстрадиолу. Значительное количество эстрона образуется в результате аромати-

ГЛАВА 2
80. MacNaughton J., Banah M., McCloud P. Age related changes in follicle simulating hormone, luteinizing, hormone, estradiol and immunoreactive inhibin in women of reproductive age, Clin. Endocrinol, 1992, 36:339.
81. Mahmud N., Murakami Т., Gyotoku Y. et al. Vulvar dystrophy: a clinical follow-up, Asia Oceania J. Obstet. Gynaecol, 1992, Sep; Vol 18, No3:231-8.
82. Meldrum D., Davidson В., Tataryn I., Changes in circulating steroids with aging in postmenopausal women, Obstet Gynecol. 1981, 57:624.
83. Modan M. et.al. Hyperinsulinemia: a link between hypertension obesity and glucose intolerance, J. Clin.Invest., 1985, 75, p. 809-817.
84. Newcomb P.A., Longnecker M.P., Storer B.E. et al. Long-term hormone replacement therapy and risk of breast cancer in postmenopausal women, Am. J. Epidem, 1995,142, pp. 788-795.
85. Pinzger G., Heim K. et al. Diagnosis and therapy of vulvar dystrophy, Gynacol. Rundsch, 1991; 31 Suppl 2:225-9.
86. Pirisi L, Yasumoto S., Felary M. J. Virol., 1987, Vol. 6 1 , pp. 1061-1066.
87. Rannevik G., Jeppsson S. Longitudinal study of the perimenopausal transition: altered profiles of steroid and pituitary hormones, Maturitas, 1995,21:103.
88. Reid I.R. et al. Determinants of total body and regional bone mineral density in normal postmenopausal women — a key role for fat mass, Journ. of din. endocrin. and metabolism, 1992, 75, 45-51.
89. Richardson S., Senikas V., Nelson J. Follicular depletion during the menopausal transition, J.Clin. Endocr.Metab, 1987, 65:1231.
90. Sherman В., West J., Korenman S. The menopausal transition, J.Clin. Endocrin.Metab. 1976, 42:629.
91. Treloar A., Boynton R., Borhild G., Brown B. Variation of human menstrual cycle through reproductive life, Int. J.Fertil., 1967,12:77.
92. Tsai M-J., Clark J., Schrader W.T. et al. Mechanisms of action of hormones that act as transcription-regulatory factors, Williams textbook of endocrinology, Ed. by J.D. Wilson, Philadelphia, W.B. Saunders Comp, 1998,55-87.

Метаболиты эстрадиола и патологические процессы..
Большинство метаболитов эстрогенов выделяются с мочой в связанной с белками плазмы форме, ~ 20% из них обнаруживаются в желчи. Скорость метаболического клиренса эстрадиола составляет приблизительно 1,0 л/мин, тогда как скорость утилизации эстрона — 1,5 л/мин. Поскольку эти эстрогены метаболизируются по сходным механизмам, это различие отражает, вероятно, более сильное связывание с сывороточными белками эстрадиола, нежели эстрона.
Эстрогены оказывают множественное влияние на женский организм. Основное их предназначение (совместно с прогестероном и гонадотропинами) заключается в контроле развития и функционирования женской репродуктивной системы и обеспечении всего комплекса мероприятий, направленных на подготовку женского организма к беременности, вынашиванию плода и родам, а именно: развитие вторичных половых признаков, появление полового влечения, обеспечение выхода яйцеклетки в половые пути и возможность его оплодотворения после овуляции, структурные изменения в тканях половой системы (пролиферация эпителия слизистой оболочки влагалища), сохранение кислого рН среды, гипертрофия и ритмические сокращения матки, развитие молочных желез, распределение подкожного жира, характерного для женского типа.
Помимо этого, эстрогены участвуют в регуляции метаболизма костной ткани (поддержание прочности и предотвращение резорбции костей), выделительной системы (регуляция водно-солевого обмена), а также в функционировании сердечно-сосудистой (понижение уровня циркулирующих в крови липидов и защита от атеросклероза) и нейроэндо-кринной систем. Отрицательное влияние эстрогенов выражается в стимуляции канцерогенеза в гормон-зависимых

ГЛАВА 3
зации андростендиона в других тканях, преимущественно в жировой. Эстриол — наименее активный из трех указанных эстрогенов (2-4 % от активности эстрадиола) — у небеременных женщин образуется лишь при метаболической деградации эстрона и эстрадиола. Однако он играет важную роль при беременности, и плацента производит значительные количества данного гормона.
Эстрадиол — главный эстроген, синтезируемый в яичниках. В среднем в организме женщины репродуктивного возраста продуцируется несколько сот микрограммов эстрадиола в сутки. Время полужизни данного гормона составляет около трех часов (22). Кроме эстрогенов, фолликул яичника секретирует ряд других половых гормонов, в частности 17 -гидроксипрогестерон и несколько андро-генов, основным из которых является андростендион.
Синтез и секреция эстрогенов регулируются лютеини-зирующим и фолликулостимулирующим гормонами гипофиза. В свою очередь, эстрогены индуцируют овулятор-ный выброс лютеинизирующего гормона.
У женщин отмечаются два пика секреции эстрогенов — во время овуляции и в период максимальной активности желтого тела (30-40 мкг/мл). Во время беременности содержание эстрогенов в плазме крови повышается до 70-80 мкг/мл за счет их биосинтеза в плаценте.
Продукты обмена эстрогенов экскретируются в виде эфиров с серной или глюкуроновой кислотами, синтезируемыми в печени, причем эстриол выделяется преимущественно в виде глюкуронида, а эстрон — в виде эфира с серной кислотой. Главными продуктами являются С-3-глюкозидуронаты эстрона и 2-оксиэстрона, а также эс-тронсульфат, этерифицированный по С-3.

Метаболиты эстродиола и патологические процессы
вании, а не в окислительном фосфорилировании. Электроны NADPH, обладающие высоким энергетическим потенциалом, переносятся на флавопротеин этой цепи; затем они передаются на адренодоксин, белок, содержащий негемовое железо. Адренодоксин переносит электроны на окисленную форму цитохрома Р-450, после чего восстановленная форма Р-450 активирует 0 2 . Система
цитохромов Р-450 играет важную роль в обезвреживании чужеродных (ксенобиотических) соединений, что увеличивает их растворимость и облегчает выведение из организма. Так же, как и в случае с эстрогенами, введение ги-дроксильных групп позволяет присоединить затем к этим молекулам полярные группы (глюкуронат или сульфат), существенно увеличивающие растворимость модифицированных ароматических молекул.
Превращение тестостерона в эстрадиол включает три стадии ферментативного гидроксилирования, в которых (при образовании одной молекулы эстрогена) участвуют три молекулы 02 и три молекулы NADPH. Первое и второе гидроксилирования происходят по С^-метильной
группе; при этом последовательно образуются 19-оксите-стостерон и 19-(альдегид)-производные. Третье гидрок-силирование, осуществляемое по С-2, является скорость-лимитирующей стадией процесса ароматизации и приводит к продукту, который быстро и неферментативно превращается в эстрадиол.
Два типа эстроген индуцированного
канцерогенеза
Влияние эндокринной системы на процессы опухолеоб-разования было обнаружено еще до появления самого

ГЛАВА 3
органах и тканях. Являясь ключевыми индукторами и проводниками внутриклеточных пролиферативных сигналов, эстрогены (главным образом эстрадиол) при определенных условиях способны стимулировать рост доброкачественных и злокачественных опухолей в эпителии молочной железы, эндометрии и шейке матки, эпителии и эндотелии слизистых (гортань, пищевод, прямая кишка).
Биосинтез эстрогенов и прогестерона
Первоначальным источником половых гормонов является холестерин. Холестерин подвергается последовательным реакциям гидроксилирования, окисления и отщепления боковой цепи с образованием прегненолона. Непосредственными предшественниками эстрона и эстра-диола являются андрогены — андростендион и тестостерон (рис. 3), которые подвергаются уникальной реакции ароматизации C-jg-стероидов.
Реакция ароматизации А-кольца с одновременным удалением метильной группы в положении С-19 является ключевой стадией в синтезе эстрогена. Эти превращения катализируются ферментом оромотозой (изоформой цитохрома Р-450) в присутствии NADPH и 02 и происходят в гладком эндоплазматическом ретикулуме. Активность ароматазы находится под гормональным контролем. Таким образом, как первая, так и последняя стадии синтеза эстрогена регулируются.
Ферменты — представители семейства цитохромов Р-450 — представляют собой конечные компоненты цепи переноса электронов, обнаруженной в митохондриях надпочечников и в микросомах печени. Роль данной системы цитохромов состоит прежде всего в гидроксилиро-

Метаболиты эстродиола и патологические процессы...
В пользу этой концепции говорят и классические наблюдения, и представления о факторах риска и гормонально-метаболической предрасположенности к развитию опухолей, и многочисленные эпидемиологические и лабораторные данные.
Как известно, в тканях женской репродуктивной системы ежемесячно происходят колебания митотической клеточной активности, регулируемые ритмическими изменениями уровня половых гормонов. В первой (т.н. пролифе-ративной) фазе менструального цикла секретируемый яичниками эстроген индуцирует пролиферацию клеток эндометрия, гибнущих во время менструации. Аналогично, каждый менструальный цикл эстроген стимулирует пролиферацию внутреннего слоя эпителиальных клеток протоков молочной железы, впоследствии также подвергающихся апоптозу. Всего в течение приблизительно 40-летнего репродуктивного периода в организме женщины осуществляется несколько сотен подобных циклов.
Попадая в клетку, эстроген (Е) активирует эстрогено-вый рецептор (ER), находящийся в цитоплазме в неактивном состоянии. Взаимодействие гормона с рецептором активирует последний и способствует его проникновению в ядро. Попав в ядро, гормон-рецепторный комплекс стимулирует экспрессию т.н. эстроген-зависимых генов, среди которых большая часть прямо или опосредованно контролирует клеточную пролиферацию, а также повышает чувствительность клеток к факторам, активирующим гиперпластические процессы (рис. 4).
Регулярно повторяющиеся циклы эстроген-индуциро-ванного клеточного деления могут двояко влиять на развитие опухолевого процесса в гормон-зависимых тканях. Во-первых, пролиферировать под действием эстро-

ГЛАВА 3
термина «гормон». Это произошло в 1896 г., когда британский хирург Г.Т. Битсон, удалив яичники у больных раком молочной железы, добился регресса опухолей и тем самым доказал зависимость роста последних от субстанций, продуцируемых этими половыми железами. Впоследствии канцерогенный эффект женских половых гормонов эстрогенов (уже после их открытия) был установлен и для других гормон-зависимых органов. В настоящее время участие эстрогенов в развитии неопластических процессов в тканях женской репродуктивной системы (эпителий молочной железы, эндометрий и шейка матки) общепризнано и рассматривается как один из ведущих этиологических факторов их возникновения (25, 27, 30).
Согласно современным представлениям, эстрогены могут быть как промоторами, так и инициаторами многоступенчатого процесса опухолеобразования. В соответствии с этим различают два основных типа гормонального эстроген-индуцированного канцерогенеза: «промотор-ный» (или физиологический) и «генотоксический».
Концепция об участии эстрогенов в промоции роста опухолей, формировавшаяся на протяжении нескольких десятилетий, считается традиционной. В большинстве публикаций, посвященных данной тематике, эстрогены рассматриваются исключительно в качестве кофакторов, усиливающих клеточное деление (стадию промоцию). Такой вариант гормонального канцерогенеза иногда называют физиологическим, поскольку он базируется на естественных, физиологических, гормон-зависимых реакциях, регулирующих рост тканей и закрепленных (в том числе, в процессе естественного отбора) за выполнением именно этих функций.

Метаболиты эстродиоло и патологические процессы...
ганах и тканях (рис. 5). Все эти факторы могут иметь значение при оценке гормонального статуса пациентки и выборе методов антиэстрогенной терапии гормон-зависимых опухолей.
Концепция «генотоксического» эстроген-индуцирован-ного канцерогенеза начала формироваться относительно недавно, приблизительно со второй половины 80-х годов. Однако с каждым годом появляется все больше работ, подтверждающих способность эстрогенов и/или их метаболитов повреждать ДНК: образовывать аддукты, усиливать рас-плетение ее цепей, инициировать разрывы и т.д., что может приводить к другим, более специфическим (пробластомо-генным) изменениям на уровне клеточного генома.
В модельной системе клеток дольково-протокового эпителия молочных желез мышей и человека установлено, что, помимо катехолэстрогенов (4-гидроксиэстрона), генотоксическими свойствами обладает и 16-а-гидрокси-производное эстрадиола (47).
Оказалось, что, кроме того, что метаболиты эстрогенов могут увеличивать доступность хроматина к влиянию истинных канцерогенов, модифицировать метаболическую активацию последних и элиминацию организмом инициированных клеток, действуя по генотоксическому механизму, они способны усиливать образование свободных радикалов, связанное с этим окислительное повреждение клеточных мембран, формировать ковалентные аддукты с ДНК, вызывать гормон-зависимую индукцию ДНК-аддуктов с последующей активацией протоонкоге-нов и ослаблять репарацию повреждений генома. Есть данные, что под влиянием эстрогенов может ослабляться репарация повреждений ДНК, индуцированных и другими соединениями.

ГЛАВА 3
генов могут уже малигнизированные (подвергнувшиеся действию внешнего канцерогена или имеющие набор терминальных мутаций) клетки, пролиферация которых приводит к формированию «опухолевого клона». Однако чаще всего имеет место другая ситуация. Периодически повторяющиеся циклы клеточного деления неизбежно повышают частоту появления новых, спонтанных мутаций (особенно с увеличением возраста женщины, а также при наличии других факторов риска). При условии накопления от 3 до 7 подобных генетических ошибок, не ликвидированных иммунной и/или репаративными клеточными системами, эстроген-зависимая пролиферация трансформированных клеток также приведет к образованию опухоли. При этом инициатор опухолевого процесса, в его традиционном понимании, во втором случае отсутствует.
Говоря об участии эстрогенов в канцерогенезе в качестве «промоторов», необходимо учитывать колоссальные изменения, произошедшие за последние 10-15 лет в понимании молекулярно-генетических механизмов действия эстрогена на гормончувствительные клетки, а именно экспериментально установленные: 1) множественность изоформ (ядерных а-, (5- и неядерных мембранных) эстрогеновых рецепторов, имеющих различные (часто противоположные) биологические функции;
2) участие в проведении гормон-индуцируемых внутриядерных сигналов факторов транскрипции, обладающих различной специфичностью в отношении связывания с молекулой ДНК и экспрессии эстроген-зависимых генов; 3) полиморфизм генов, кодирующих рецепторы эстрогенов, как этиологический фактор опухолеобразования; 4) широкое распределение эстрогеновых рецепторов в ор-

Метаболиты эстрадиола и патологические процессы.
торый затем реагирует с глюкуроновой кислотой и эк-скретируется.
Основной же пул эндогенных эстрогенов утилизируется с помощью реакций гидроксилирования эстрона — продукта превращения эстрадиола, осуществляемых посредством изоферментов, принадлежащих к уже упоминавшейся нами монооксигеназной системе цитохромов Р-450. Изоферменты цитохромов Р-450 (CYP1B1 и CYP1A1) обеспечивают конверсию эстрадиола в два основных его метаболита: 16а-гидроксиэстрон (16а-0НЕ1) и 2-гидроксиэстрон (2-0НЕ1) (рис. 6). Полигидроксипро-изводные эстрона, как мы уже говорили, конъюгируют с глюкуронатом или сульфатом или подвергаются реакции О-метилирования в присутствии S-аденозилметионина, катализируемой относительно неспецифической кате-хол-0-метилтрансферазой. После чего метоксипроиз-водные и их конъюгаты выводятся с мочой и через жел-чевыводящие пути (18).
Показано, что 1ба-гидроксиэстрогены — эстриол и 16а-гидроксиэстрон — проявляют утеротропную активность, сравнимую с таковой у эстрадиола (12, 31). 16а-гидроксиэстрон (1ба-0НЕ1) превращается в эстриол с помощью 17-ОН-стероиддегидрогеназы, который затем связывается с белками плазмы и выводится из организма. 2-0НЕ1 или сразу связывается с сывороточными белками, или предварительно превращается в 2- меток-сиэстрон.
Цитохром Р-450 1А1 (CYP1A1), катализирующий 2-ги-дроксилирование эстрона, является индуцибельным ферментом. В больших количествах CYP1A1 локализован в микросомальнои фракции печени и активируется в ответ на некоторые пищевые ингредиенты, а также на сигарет-

ГЛАВА 3
По мнению некоторых авторов, в случае эстроген-ин-дуцированного канцерогенеза равновероятно вовлечение в процесс обоих его типов — физиологического (где гормоны выступают в качестве митогенов и (или) промоторов) и генотоксического (где они же играют роль инициаторов), и выбор между ними осуществляется в зависимости от условий существования организма и некоторых других факторов ( 1 , 5 ) .
Метаболизм эстрадиола
Первичный метаболизм эстрадиола происходит в печени. Образуясь из андрогенных предшественников — те
стостерона и андростендиона, «классические» эстрогены — эстрадиол и эстрон — подвергаются дальнейшим взаимопревращениям с формированием значительного числа метаболитов.
Как и в процессах синтеза, наибольшее значение в реакциях обмена эстрогенов имеют реакции гидроксилиро-вания. На этот факт было обращено внимание достаточно давно, более 40 лет тому назад (16). При этом часть гидроксилированных производных эстрогенов — на основании особенностей их структуры и свойств — получили название неклассических фенолстероидов. С этого момента увеличение продукции этих соединений стало рассматриваться как свидетельство качественных изменений в стероидогенезе и как фактор развития некоторых злокачественных новообразований, в частности рака тела матки (17).
Небольшая часть эстрадиола сразу подвергается реакции 16 -гидроксилирования с образованием эстриола, ко-

Метаболиты эстрадиола и патологические процессы...
В отличие от 2-ОН-гидроксиэстрона, 16а-0НЕ1 является мощным агонистом эстрадиола. Данный метаболит обладает некоторыми особенностями химической структуры (уникальной взаимной ориентацией 16а-0Н-группы и кето-группы эстрона (15), позволяющими ему образовывать ковалентные связи со специфическими эстрогено-выми рецепторами (45), в результате чего продолжительность эстроген-зависимого пролиферативного сигнала возрастает с нескольких часов до нескольких дней (29, 44, 47) (рис. 7).
Есть мнение, что 1ба-ОНЕ1-опосредованный эффект длится до момента начала деградации связывающих данный метаболит плазменных белков (28). Кроме того, 16а-0НЕ1 может индуцировать канцерогенные эффекты и по генотоксическому механизму (15, 47).
Все вместе это делает 16а-0НЕ1 «агрессивным» метаболитом эстрогена, проявляющим выраженные канцерогенные свойства.
Считается, что подобным образом, одновременно влияя на клеточную пролиферацию и целостность генетического аппарата, процесс опухолеобразования индуцируется в ответ на действие онкогенных вирусов и химических канцерогенов (7).
Следует отметить, что, помимо 2-0НЕ1 и 16а-0НЕ1, в ходе метаболизма эстрадиола образуется еще некоторое количество 4-гидроксиэстрона (4-0НЕ1), который, так же, как 16а-гидроксиэстрон, является агонистом эстрогена с «агрессивными свойствами», хотя и менее выраженным. Однако относительная концентрация 4-0НЕ1 намного ниже, чем концентрация 2-0НЕ1 и 16а-ОНЕ1, и поэтому не оказывает существенного влияния на соотношение метаболитов эстрогенов.

ГЛАВА 3
ный дым. Второй фермент — CYP1B1 — катализирует 1ба-гидроксилирование эстрона. Его активность не повышается при добавлении диетических компонентов, но стимулируется в ответ на ксенобиотические карциногены и пестициды (19).
Принципиальным моментом является тот факт, что 2- и 16а-гидроксипроизводные эстрона обладают противоположными биологическими свойствами.
2-0НЕ1 практически не влияет на клеточную пролифе-ративную активность, в большинстве случаев действуя как слабый агонист эстрадиола (низкоактивный эстроген), а в некоторых опытных моделях даже как антиэстроген (23, 37). Тот же вывод следует из экспериментов in vitro на опухолевых клеточных культурах (42). При этом 2-ги-дроксилирование эстрогенов превращает А-кольцо этих стероидов в катехол, структуру, обнаруживаемую у биологически активных катехоламинов (например, дофамина). Поэтому 2-ОН-эстрогены (наряду с 4-ОН-эстрогенами — еще одной разновидностью гидроксилированных производных эстрогена, не показано на рис.) часто называют катехолэстрогенами и предполагают их участие в регуляции нейро-эндокринных функций организма (21, 40).
Интересно, что у человека на скорость гидроксилиро-вания стероидных гормонов и превращения окси-групп в кето-группы значительно влияет состояние щитовидной железы. Так, при гипертиреозе образование 2-гидрокси-эстрона увеличивается столь значительно, что он оказывается главным метаболитом эстрогенов. При миксидеме, напротив, образование 2-гидроксиэстрона уменьшается. При повышенной тиреоидной активности уменьшается образование эстриола.

ГЛАВА 3
Роль метаболитов эстрогенов в канцерогенезе репродуктивных органов
Общепризнано, что гормональный фактор является первопричиной возникновения опухолей в гормон-зависимых органах и тканях. Наиболее интенсивно роль эстрогенов в канцерогенезе изучалась на модели рака молочной железы (РМЖ). С начала 80-х гг. прошлого века было проведено несколько десятков эпидемиологических рандомизированных исследований, посвященных роли эстрогенов в этиологии данного заболевания. Было показано, что уровень эстрадиола в сыворотке крови, варьируя в различных этнических популяциях, достоверно коррелирует с повышенным риском заболевания РМЖ. Постменопау-зальные женщины с диагнозом РМЖ имели 15%-ное превышение концентрации эстрадиола в сыворотке крови по сравнению со здоровыми пациентками (р = 0.003).
Вместе с тем хорошо известно, что РМЖ может возникать и на фоне нормальной (или близкой к нормальной) концентрации эстрогена в кровотоке. Предполагается, что в таких случаях ведущую роль в стимуляции патологической пролиферации клеток молочной железы играет повышенный локальный уровень эстрогена, особенно при начавшемся опухолевом процессе (известно, что опухолевые клетки гормон-зависимых тканей способны самостоятельно синтезировать эстрогены), а также усиливающийся с возрастом дисбаланс половых гормонов, вызванный нарушением метаболизма эстрогенов.
В последнее время становится все более популярной точка зрения, что нарушение гормонального равновесия играет ведущую роль в процессах опухолеобразования в гормон-зависимых тканях (органах репродуктивной системы). Основанием для подобного вывода послужил уве-

ЛЛетоболиты эстродиоло и патологические процессы...
личивающийся с каждым годом объем информации, касающейся «агрессивных» (канцерогенных) свойств одного из ключевых метаболитов эстрогена — 16а-0Н-ги-дроксиэстрона.
Концепция, предполагающая, что нарушение гормонального равновесия в организме, а именно дисбаланс соотношения 2- и 16а-гидроксипроизводных эстрогена, является основной причиной возникновения злокачественных гормон-зависимых опухолей (РМЖ), начала формироваться достаточно давно, в начале 80-х годов прошлого века. Тогда было впервые установлено значимое 50%-ное (!) увеличение скорости процесса 16а-ги-дроксилирования у больных с РМЖ по сравнению с контрольной группой {43). На сегодняшний день справедливость представлений о связи метаболитов эстрогена с процессами опухолеобразования подкреплена результатами огромного числа лабораторных и клинических исследований (табл. 8).
Таблица 8 Метаболиты эстрогена и канцерогенез (28)
Год
1982
1984
1988
1992
Результаты исследований (основной вывод)
Установлена взаимосвязь между 16а-гидроксилирован-ными метаболитами эстрогенов и РМЖ
Пациентки с РМЖ и раком эндометрия имеют повышенный уровень 16а-гидроксилазной активности
16а-ОНЕ1 способен ковалентно и необратимо взаимодействовать с эстрогеновыми рецепторами
Установлены пролиферативный и генотоксический механизмы 16а-ОНЕ1-индуцированного канцерогенеза (клеточные культуры РМЖ)

Метаболиты эстрадиола и патологические процессы
Так в расширенных клинических исследованиях на большой выборке больных (5 104 женщин от 35 лет и старше; время наблюдения 9,5 лет) с диагнозом РМЖ было показано, что постменопаузальные женщины с прогрессирующими опухолями имели как минимум 15%-ное статистически значимое снижение величины 2-ОНЕ1/16 -ОНЕ1 по сравнению с контрольной группой (32).
В других, более поздних, исследованиях, проведенных на еще более обширной выборке женщин (10 876 чел.; 35-69 лет), наблюдавшихся в течение 5,5 лет, было установлено, что в группе пременопаузальных женщин высокий уровень 2-ОНЕ1/16а-ОНЕ1 коррелировал с пониженным риском возникновения РМЖ (39).
Можно ли рассматривать соотношение 2-ОНЕ1/ 16а-ОНЕ1 как адекватный прогностический фактор РМЖ? Оказалось, что да. В рандомизированных исследованиях, проведенных в конце 90-х гг., было четко показано, что постменопаузальные женщины с диагнозом РМЖ имели пониженное значение данного параметра по сравнению с соответствующим контролем. Более того, было показано, что у пациентов с III и IV стадиями РМЖ определялись более низкие значения 2-ОНЕ1/16а-ОНЕ1, чем у пациентов с I и II стадиями заболевания (24).
В настоящее время считается доказанным, что для поддержания нормального гормонального баланса как у пре-, так и у постменопаузальных женщин необходимо, чтобы концентрация 2-ОНЕ1 превышала концентрацию 16а-ОНЕ1 как минимум в два раза. При понижении данного соотношения статистически значимо возрастает риск возникновения РМЖ (28, 41).
Таким образом, соотношение 2-ОНЕ1 к 16а ОНЕ1 можно считать адекватным универсальным биомар-

ГЛАВА 3
Год
1994
1995
1995
1996
1997
1997
1997
1997
1998
1998
2000
2001
Результаты исследований (основной вывод)
Агенты, повышающие уровень 2-ОНЕ1, подавляют канцерогенез
Соотношение 16а-ОНЕ1/2-ОНЕ1 повышено на поздних стадиях РМЖ (эксп. in vivo на животных и клинич. исследования)
Пестициды (органохлорин) активируют цитохром Р-450, ответственный за синтез 16а-ОНЕ1
Полиморфизм CYP1A1 является причиной пониженного соотношения 2-ОНЕ1/16а-ОНЕ1 в Афро-Американской популяции женщин по сравнению с кавказской популяцией
16а-ОНЕ1 индуцирует канцерогенез в эпителиальных клетках молочной железы по пролиферативному и гено-токсическому механизму (повреждение ДНК), подобно канцерогену 7,12-диметил-(а)6ензатрацен (DMBA)
Бифункциональность (усиление пролиферации и геното-ксический эффект) канцерогенного действия 16а-ОНЕ1
Установлена четкая обратная корреляция между соотношением 2/16 и РМЖ (клинич. исследования)
Постменопаузальные женщины с РМЖ имели 15%-ное статистически значимое снижение величины 2-ОНЕ1/ 16а-ОНЕ1 по сравнению с контрольной группой (клинич. исследования)
Эстрадиол и 16а-ОНЕ1 усиливают аномальную пролиферацию и малигнизацию кератиноцитов. Данный эффект блокируется индол-3-карбинолом
Увеличение соотношения 2-ОНЕ1/16а-ОНЕ1 коррелирует с улучшением статуса больных возвратным респираторным папилломатозом
У пременопаузальных женщин высокий уровень 2-ОНЕ1/ 16а-ОНЕ1 коррелировал с пониженным риском возникновения РМЖ (клинич. исследования)
Соотношение 2/16а— предполагаемый биологический маркер риска опухолей головы и шеи

Метаболиты эстродиола и патологические процессы...
щих» онкогенов. Заставить их проснуться может множество причин — канцерогенных факторов.
Известны десятки зарубежных и отечественных эпидемиологических исследований, проводившихся с целью количественной оценки значимых канцерогенных факторов, приводивших к смерти от всех видов рака. Было установлено, что для некоторых его видов первоочередное значение может приобретать какой-то один или несколько факторов (например, курение для некоторых типов рака легких; воспалительные процессы для опухолей желудка и кишечника; токсические канцерогены, радиационное излучение для лейкозов). По некоторым данным, не менее 30% всех случаев заболеваний раком прямо или косвенно связано с особенностями питания (высокое содержание в пище животных жиров, консервантов и канцерогенов). Большое влияние оказывают также ухудшающаяся с каждым годом экология; рост инфекционных заболеваний (в некоторых случаях провоцирующих предраковые состояния) и отягощенная наследственность.
И, тем не менее, ведущим этиологическим фактором канцерогенеза был и остается возраст пациента. На сегодняшний день устоявшимся считается мнение, что рак (наряду с другими болезнями XXI века — инфарктом и инсультом) — это «болезнь пожилых людей». Согласно статистике, около 60% всех случаев рака приходится на людей старше 65 лет.
Исходя из генетической многостадийной теории происхождения рака, такая точка зрения представляется вполне закономерной. Действительно, есть расчеты, что примерно 1 из 109 нуклеотидов ДНК мутирует при каждом клеточном делении. На сегодняшний день известно около 100 протоонкогенов (нормальных клеточных генов, при

ГЛАВА 3
кером и надежным диагностическим критерием при определении риска и прогноза развития эстроген-зависимых опухолей.
В настоящее время клиницистам стал доступен простой, неинвазивный, достоверный и относительно недорогой иммуно-ферментный ELISA-метод определения 2-0НЕ1 и 1ба-0НЕ1 в моче (26), что в будущем открывает неограниченные перспективы по расширению масштабов профилактики и лечения доклинических форм гормон-зависимых опухолей (особенно среди пациентов, входящих в группы риска по данным заболеваниям), т.е. решению одной из ведущих проблем современной онкологии.
Что такое «канкрофилия» и «метаболический синдром», или «почему гормон-зависимые опухоли у женщин чаще возникают в постменопаузе?
Как известно, у любой болезни есть причина (причины) возникновения — этиология и механизмы развития — патогенез. На сегодняшний день считается, что «рак» — это полиэтиологическое, но монопатогенетическое заболевание. Это означает, что не существует одной-единственной причины для развития рака. Данное заболевание, которое имеет около 100 различных форм проявления, многолико и может развиться под влиянием широкого спектра факторов: химических, физических, биологических (вирусов), гормональных и др., в принципе вызывающих одни и те же изменения в геноме. Другими словами, в каждой клетке заложена потенциальная предрасположенность к злокачественному перерождению в виде «спя-

Метаболиты эстрадиоло и патологические процессы...
этиологических факторов не оказывает такого влияния на вероятность заболеть данным опухолевым заболеванием, как обычное старение.
Все эти рассуждения хорошо укладываются в базисные положения теории «канкрофилии». В основе данной теории лежат представления о том, что вероятность развития рака зависит не только от интенсивности и времени действия внешних канцерогенных факторов, но также и от состояния внутренней среды — определенных гормонально-обменных условий, предрасположенности организма к возникновению злокачественных опухолей.
Впервые представление о синдроме канкрофилии (буквально означающей — «любовь к раку») было выдвинуто в середине 70-х гг. русским ученым-онкоэндокриноло-гом В.М. Дильманом (4).
Это содержательная гипотеза, основанная на многолетних наблюдениях автора, объясняла многие недостающие до сих пор «триггерные» («запускающие») условия и механизмы возникновения рака. Суть ее заключается в следующем: для возникновения и развития рака как болезни в организме вне зависимости от причинных канцерогенных факторов нужны гормонально-метаболические (обменные) условия-сдвиги, а именно: увеличение пула-совокупности пролиферирующих (делящихся) клеток в ткани, снижение активности клеточного иммунитета и макрофагов и угнетение активности систем, ответственных за репарацию повреждений генома. Каждое из этих трех условий может самостоятельно способствовать развитию рака (3).
По мнению В.М. Дильмана, ключевым фактором, необходимым для реализации трех вышеуказанных условий развития канкрофилии, является преимущественное

ГЛАВА 3
неблагоприятных условиях трансформирующихся в индуцирующие злокачественный рост онкогены). Длина кодирующей части одного гена в среднем равна примерно 3000 нуклеотидов, длина кодирующих частей всех 100 онкогенов — 300 000. Это означает, что хотя бы 1 онкоген в 3000 клетках с высокой вероятностью получит мутацию в кодирующей области за одно деление этих клеток. Человек содержит приблизительно 10 1 3 -10 1 4 клеток. Следовательно, в процессе развития организма должны возникать миллиарды клеток, несущих мутантные онкогены (если бы одной мутации было достаточно для развития рака, то еще до рождения наш организм состоял бы из одних опухолей). В результате с возрастом накапливается количество мутаций, необходимое для опухолевой трансформации. Считается, что в среднем существует приблизительная закономерность: у(вероятность развития рака) = (возраст)5.
С другой стороны, периодически возникающие сбои регуляторных механизмов размножения клеток, в норме ликвидирующиеся репаративными клеточными структурами и иммунной системой организма, также со временем изнашиваются, а следовательно, вероятность генетических поломок, приводящих к образованию опухолевых клонов, возрастает.
Особенно актульным данный вывод представляется в отношении т.н. гормон-зависимых опухолей, т.е. опухолей, возникающих в органах и тканях, являющихся мишенями гормонов. К таковым относятся органы женской репродуктивной системы: эндометрий и миометрий матки, яичники, цервикальная зона матки, молочные железы. Известно, что за исключением очень тяжелой наследственной склонности к раку молочной железы ни один из

Метаболиты эстрадиола и патологические процессы
пролиферацию внутреннего слоя эпителиальных клеток протоков молочной железы, впоследствии также подвергающихся апоптозу. Всего в течение приблизительно 40-летнего репродуктивного периода в организме женщины осуществляется несколько сотен подобных циклов.
Регулярно повторяющиеся циклы эстроген-индуциро-ванного клеточного деления могут двояко влиять на развитие опухолевого процесса в гормон-зависимых тканях. Во-первых, пролиферировать под действием эстрогенов могут уже малигнизированные (подвергнувшиеся действию внешнего канцерогена или имеющие набор терминальных мутаций) клетки, пролиферация которых приводит к формированию «опухолевого клона» и впоследствии к наследственному раку. Кроме того, периодически повторяющиеся циклы клеточного деления неизбежно повышают частоту появления новых, спонтанных мутаций.
Очевидно, что вероятность появления «генетических поломок» в органах репродуктивной системы будет неизбежно возрастать с увеличением возраста женщины. Во-первых, просто в силу временного фактора (увеличение времени экспозиции клеток-мишеней с эстрогенами — промоторами пролиферации и индукторами генотокиче-ских поврежедений), а во-вторых, по причине накопления в анамнезе возрастных и системных заболеваний, дополнительно ослабляющих иммунитет, а также продолжительного воздействия других факторов риска онкозаболеваний.
В самом деле, согласно данным статистики, первый пик заболеваемости раком молочной железы (80-100 случаев на 100 000 женщин) приходится на репродуктивный период женщины — от 30 до 45 лет. Второй и третий пики отмечаются соответственно в возрасте 50-60 лет (180 слу-

ГЛАВА 3
использование тканями в качестве энергетического материала насыщенных жирных кислот вместо глюкозы. Подобные сдвиги встречаются при нормальном старении, болезнях компенсации (терминология автора) — атеросклерозе, гипертонической болезни, ожирении, диабете, климаксе, возрастной психической депрессии, а также при хроническом стрессе и переедании (избытке в питании жиров, белков или углеводов). Например, при переедании или нерациональном питании в крови увеличивается уровень инсулина и холестерина — индукторов деления соматических клеток.
Таким образом, канкрофилия — это сумма гормонально-метаболических условий-сдвигов, повышающих вероятность злокачественной трансформации клетки.
В отношении гормон-зависимых опухолей теория кан-крофилии справедлива в наибольшей степени. Ведь увеличение пула-совокупности пролиферирующих (делящихся) клеток (а, следовательно, частота возникновения потенциально онкогенных мутаций) в гормон-зависимых тканях предопределено самим фактом существования последних. В разделе «Два типа эстроген-индуцированного канцерогенеза» мы подробно обсуждали промоторную роль эстрогенов в процессах опухолеобразования. Еще раз напомним, что в тканях женской репродуктивной системы ежемесячно происходят колебания митотической клеточной активности, регулируемые ритмическими изменениями уровня половых гормонов. В первой (т.н. проли-феративной) фазе менструального цикла секретируемыи яичниками эстроген индуцирует пролиферацию клеток эндометрия, гибнущих во время менструации. Аналогично, каждый менструальный цикл эстроген стимулирует


ГЛАВА 3
чаев на 100 000 женщин) и после 65 лет (250 случаев на 100 000 женщин). Кумулятивный риск развития РМЖ у женщины, дожившей до 85 лет, достигает 10% (в данных рассуждениях не учитывается наследственный рак, вероятность которого составляет от 3 до 5% всех случаев данного опухолевого заболевания).
Таким образом, в отличие от большинства других опу-холеобразующих органов и тканей гормон-зависимые ткани представляют собой регулируемую биологическую систему, подвергающуюся на протяжении всего репродуктивного периода женщины периодической стимуляции к клеточному росту. Пока защитные силы каждой отдельной клетки и организма в целом достаточны для того, чтобы противостоять раскачивающим устойчивость данной системы «качелям», ситуация находится под контролем. В случае их ослабления — при неблагоприятных внешних или внутренних условиях или просто по причине старения организма — нарушаются регуляторные механизмы клеточного деления и появляется вероятность возникновения опухолевого процесса. Риск возникновения гормон-зависимых опухолей многократно возрастает при наличии одного или нескольких факторов риска.
И в заключение еще об одном обстоятельстве, непосредственно касающемся метаболических сдвигов в организме, предшествующих канцерогенезу. Как мы отмечали выше, в главе «Роль метаболитов эстрогенов в канцерогенезе репродуктивных органов», в последние годы появляется все больше данных, свидетельствующих о том, что нарушение баланса метаболитов эстрогенов — 2-гидрокси-эстрона и 16а-гидроксиэстрона — играет ведущую роль в процессах опухолеобразования в гормон-зависимых тканях (органах репродуктивной системы) у женщин, находя-





Метаболиты эстродиоло и патологические процессы...
щихся в менопаузе (постменопаузе). Причины данного феномена пока до конца не ясны. Одно из возможных объяснений может выглядеть следующим образом.
Известно, что цитохромы Р-450, относящиеся к ферментам 1-й фазы трансформации и катализирующие ключевые фазы метаболизма эстрогенов, а именно образование их 2- и 16а-гидроксипроизводных, метаболизируют еще и ксенобиотики, в том числе ряд канцерогенов, находящихся в окружающей среде, например полициклические ароматические углеводороды (ПАУ). Показано, что последние способны изменять активность цитохромов Р-450 для эстрогенов таким образом, что происходит сдвиг метаболизма в сторону накопления более «агрессивных» форм эстрогенов (2).
Возможно также, что дисфункция изоферментов цито-хрома Р-450, контролирующих нормальный метаболизм эстрогенов, наступает вследствие нарушения внутренних регуляторных систем организма.
Нельзя исключить и тот факт, что на преимущественное образование «агрессивных» форм эстрогенов у пост-менопаузальных женщин оказывает влияние смещение места синтеза половых гормонов из яичников в жировую ткань, где основным эстрогеновым продуктом является образующийся из андростендиона эстрон.
Фармакологическая коррекция гормонального баланса и профилактики образования гормон-зависимых опухолей
Довольно давно было замечено, что важнейшим фактором, влияющим на процессы канцерогенеза, является диета, и что некоторые пищевые компоненты (преимуще-

ГЛАВА 3
ственно растительного происхождения) способны тормозить образование опухолей в организме. Одними из первых в этом списке традиционно назывались растения семейства крестоцветных: различные виды капусты (бело-,'' краснокачанная, брюссельская, цветная, брокколи), кабачки, брюква, турнепс, горчица. Позднее выяснилось, что антиканцерогенная активность перечисленных овощных культур обусловлена наличием в их составе пищевого индола — индол-3-карбинола (I3C), высвобождающегося в результате гидролиза его предшественника индо-лилметилглюкозинолата — глюкобрассицина (11). Было установлено, что I3C обладает уникальной особенностью значительно снижать риск возникновения опухолей в гормон-зависимых органах и тканях посредством нормализации гормонального баланса (соотношения метаболитов эстрогенов) в организме.
Впервые противоопухолевое действие I3C было продемонстрировано на модели рака молочной железы в 70-80-х гг. В экспериментах in vitro и in vivo на животных было установлено, что I3C обладает выраженной способностью снижать частоту возникновения опухолей молочной железы, индуцированных экзогенными канцерогенами (диметилбензантраценом) (38, 49).
Спустя приблизительно 10 лет был впервые постулирован молекулярный механизм, посредством которого I3C обезвреживает канцерогенные ксенобиотики (б). Оказалось, что пищевые индолы являются активаторами входящих в состав монооксигеназной системы печени индуцибельных изоформ цитохрома Р-450. Напомним, что изоферменты цитохрома Р-450 (функции и механизм действия которых были подробно рассмотрены нами в предыдущих разделах) относятся к ферментам 1-й фазы

Метаболиты эстрадиола и патологические процессы...
трансформации и, помимо того, что метаболизируют (ги-дроксилируют) ксенобиотики, катализируют ключевые фазы метаболизма эстрогенов, а именно образование их 2- и 16а-гидроксипроизводных. В начале 90-х гг. XX века было окончательно доказано, что I3C способен значимо влиять на метаболизм эстрадиола в женском организме, избирательно активируя изоферменты цитохрома Р-450 и повышая таким образом уровень его 2-гидроксилирован-ных производных (9, 36).
С этого момента I3C как противоопухолевый агент становится объектом интенсивного изучения в ведущих научных и медицинских центрах. Проводятся многочисленные лабораторные эксперименты in vitro и in vivo по выявлению его внутриклеточных мишеней, изучению метаболических путей превращения в организме и механизмов противоопухолевой активности (10, 11, 14, 20, 33, 34), а также широкомасштабные клинические исследования по оценке его влияния на концентрации 2-ОН- и 16а-ОН-ме-таболитов эстрогена и установлению эффективных терапевтических доз при назначении I3C в качестве средства противоопухолевой терапии (8, 46, 48, 50).
Подробно основные свойства и механизмы действия I3C будут рассмотрены нами ниже в отдельной главе данной книги. Кратко основные выводы, вытекающие из этих разработок, можно сформулировать следующим образом.
I3C является универсальным корректором патологических пролиферативных процессов в органах женской репродуктивной системы. Универсальность противоопухолевого действия I3C обусловлена способностью данного соединения блокировать все основные пути трансдукции внутриклеточных сигналов, стимулирующих клеточный рост, что позволяет использовать его при адъювантном

ГЛАВА 3
лечении и профилактике не только гормон-чувствительных, но и гормон-нечувствительных (не содержащих эс-трогеновые рецепторы) опухолей, а также индуцировать избирательный апоптоз (программированную клеточную гибель) опухолевых клеток. I3C является поистине уникальным соединением, поскольку способен подавлять развитие опухолевого процесса практически на всех его уровнях: субмолекулярном (активация опухоль-су прес-сорных генов и супрессия генов, стимулирующих онкоге-нез), молекулярном (блокировка пролиферативных сигналов на всем их протяжении — от рецепторов до ядерных белков и ДНК) и надмолекулярном (торможение процессов клеточной миграции и инвазии). Кроме того, выяснилось, что I3C обладает выраженным противовирусным действием, в частности в отношении вируса папилломы человека, вызывающего неопластические изменения в цервикальной области матки и в эпителии гортани.
Однако основное свойство I3C, придающее ему качество блокатора пролиферативных гормон-индуцируемых сигналов, безусловно, состоит в его способности восстанавливать нормальный баланс гидроксипроизводных эстрогена — слабого эстрогенового агониста 2-гидроксиэс-трона и «агрессивного» метаболита — 16а-гидроксиэс-трона. Последний, как мы отмечали выше, способен значительно пролонгировать эстроген-зависимый пролифе-ративный сигнал, а также вызывать опосредованные эстрогенами (и/или продуктами их метаболизма) геноток-сические повреждения наследственного аппарата, стимулируя тем самым гормон-зависимый канцерогенез.
Данный вывод был сделан на основании ряда проведенных за рубежом рандомизированных клинических исследований (8, 35, 50). В ходе этих исследований на ши-

Метаболиты эстродиола и патологические процессы...
рокой выборке пациентов было показано, что после трехмесячного курса регулярного приема I3C в ежедневной дозе 300-400 мг у 85% женщин из основной опытной группы статистически достоверно улучшалось соотношение гидроксиметаболитов эстрогена 2-ОНЕ1/16а-ОНЕ1, причем данный эффект сохранялся на протяжении нескольких месяцев после отмены препарата. При этом долговременный прием I3C не вызывал каких-либо отрицательных побочных эффектов.
На сегодняшний день известна целая группа сывороточных и тканевых маркеров, позволяющих контролировать течение и рецидивирование уже обнаруженных гормон-зависимых опухолей, в частности РМЖ. Вместе с тем маркеров их"доклинической диагностики практически не существует. Соотношение 2- и 16а-гидроксиметаболитов эстрогена является наиболее адекватным и практически единственным реальным критерием, коррелирующим с риском появления новых эстроген-чувствительных опухолей (28). Кроме того, как мы говорили, уровень 2-ОНЕ1/16а-ОНЕ1 может быть и прогностическим фактором течения подобных опухолевых заболеваний. Доводя баланс метаболитов эстрогена до нормы, I3C нормализует метаболические гормональные сдвиги в организме и способствует регрессии уже образовавшихся и снижению риска появления новых гормон-зависимых новообразований.
Из всего вышеизложенного с очевидностью следует, что препараты на основе I3C должны быть высокоэффективны в отношении лечения и профилактики рака репродуктивных органов. Однако все попытки создания таких препаратов, предпринимавшиеся за рубежом, наталкива-

Метаболиты эстродиоло и патологические процессы...
ской, но и социально значимой. Несмотря на усилия традиционной онкологии, стремящейся усовершенствовать существующие методы борьбы с этим грозным недугом, выбор оптимального варианта лечения с минимальными для больного побочными эффектами по-прежнему остается серьезной проблемой. Ситуация дополнительно усугубляется сложностью канцерогенеза РМЖ, в частности его высокой агрессивностью (потенцией к метастазирова-нию) и трудностью ранней диагностики. В России почти половина женщин, страдающих раком груди, обращается к врачу при распространенном (метастатическом) процессе, когда лечение часто оказывается неэффективным. В целом при РМЖ 5-летняя выживаемость больных составляет около 50%, однако при I стадии заболевания она приближается к 95%, а 10-летняя выживаемость при этой же стадии — к 80%. Таким образом, первоочередную значимость на сегодняшний день приобретает разработка новых эффективных методов доклинической диагностики и широкомасштабной профилактики РМЖ.
Есть все основания надеяться, что новый отечественный препарат «Индинол», принимаемый с лечебной и/или профилактической целью, позволит принципиально улучшить ситуацию с заболеваемостью гормон-зависимыми опухолями (в том числе РМЖ) в нашей стране и значительно отодвинуть возрастную границу, при которой наблюдается ее активный рост.

ГЛАВА 3
лись на значительные трудности, связанные с их нестабильностью.
Отечественной научно-производственной компании «МираксФарма» удалось успешно решить эти проблемы и получить стабильное соединение на основе индол-3-кар-бинол — препарат «Индинол». К настоящему моменту Индинол доказал высокую эффективность противоопухолевого действия при лечении доброкачественных опухолевых заболеваний молочной железы, а также неопластических образований эндометрия и шейки матки (в том числе вызванных вирусом папилломы человека) и рецидивирующего респираторного папилломатоза. Ближайшей перспективой компании является создание комплексных лекарственных препаратов для лечения новообразований репродуктивной системы на основе индол-3-карбинола, в состав которых, помимо данного соединения, будут входить другие противоопухолевые вещества растительного и синтетического происхождения. К числу последних принадлежит, в частности, антиэстроген Тамоксифен, широко используемый в настоящее время в качестве средства эндокринной адъювантной терапии при лечении гормон-зависимого РМЖ, но, к сожалению, обладающий рядом значимых побочных эффектов. Доказано, что препарат на основе комплекса (Индинол + Тамоксифен) будет обладать значительно более выраженной противоопухолевой активностью по сравнению с индивидуальным Тамокси-феном (13) и при этом лишен отрицательных побочных эффектов, присущих последнему.
Неутешительная статистика заболеваемости и смертности женской части населения от опухолевых заболеваний свидетельствует о том, что проблема РМЖ в нашей стране становится в последнее время не только медицин-

Метаболиты эстрадиоло и патологические процессы...
12. Clark J.H., Paszko Z., Peck E.J. Nuclear binding and retention of the receptor estrogen complex: relation to the agonistic and antagonistic properties of estriol. Endocrinology, 1977,100, 91-96.
13. Cover СМ., Hsieh J., Cram E.J., Hong C, Riby J.E., Bjeldanes L.F., Firestone G.L. lndole-3-carbinol and tamoxifen cooperate to arrest the cell cycle of MCF-7 human breast cancer cells. Cancer Res, 1999, 59,1244-1251.
14. Dashwood R.H., Fong AT., Arbogast D.N. et al. Anticarcinogenic activity of indole-3-carbinol acid products: ultrasensitive bioassay by trout embryo microinjection. Cancer Res., 1994, 54, 3617-3619.
15. Davis D.L., Telang NT., Osborne M.P., Bradlow H.L Medical hypothesis: bifunctional genetic-hormonal pathways to breast cancer. Environ. Health Perspect., 1997,105, 571-576.
16. Diczfalusy E., Lauritzen С Oestrogene beim Menschen. Berlin: Springer, 1961. p. 257.
17. Dilman V.M., Berstein L.M., Bobrov J.F., Bokhman J.V., Kovaleva L.G., Krylova N. V. Hypothalamo-pituitary hyperactivity and endometrial carcinoma. Amer J. Obstet. and Gynecol., 1968,102, 880-890.
18. Fishman J., Bradlow H.L, Gallagher T.F. Oxidative metabolism of estradiol. J. Biol. Chem., 1960, 235, 3104-3107.
19. Fishman J., Osborne M.P., Telang NT. The role of estrogen in mammary carcinogenesis. Ann. NY Acad. Sci., 1995, 768, 91-100.
20. Ge X., Yannai S., Rennert G et al. 3,3'-diindolylmethane induces apoptosis in human cancer cells. Biochem. Biophys. Res. Commun., 1996, 228,153-158.
21. Gethmann U., Knuppen R. Effect of 2-hydroxyoestrone on lutropin (LH) and follitropin (FSH) secretion in the ovarectomized primed rat. Hoppe-Seyler'sZ. Physiol. Chem., 1976, 357,1011-1013.
22. Ginsburg E.S., Gao X., Shea B.F., Barbieri R.L. Half-life of estradiol in postmenopausal women. Gynecol. Obstet. Invest., 1998, 45, 45-48.
23. Gordon J., Cantrall W., Alberts H. et al. Steroids and lipid metabolism: The hypocholesteremic effect of estrogen metabolites. Steroids, 1964, 4,267-291.
24. Kabat G.C., Chang C.J., Sparano J.A. et al. Urinary estrogen metabolites and breast cancer: a case-control study. Cancer Epidemiol. Biomar-kersPrev., 1997,6, 505-509.

ГЛАВА 3
Список литературы к Главе 3
1. Берштейн Л.М. Гормональный канцерогенез. — СПб.: Наука, 2000.
2. Глушков А.Н. «Общебиологические закономерности и механиз
мы канцерогенеза». Медицина в Кузбассе. 2004, № 1, стр. 3-9.
3. Дильман В.М. Синдром канкрофилии: раковая клетка, факторы
риска развития опухоли и организм // Организация противораковой
борьбы, профилактика злокачественных опухолей. — Л.:
Медицина, 1976.
4. Дильман В.М. Эндокринологическая онкология. — Л.: Медицина,
1983.
5. Дильман В.М. Большие биологические часы (введение в инте
гральную медицину). — М.: Знание, 1986.
6. Bradfield C.A., Bjaeldanes L.F. Structure-activity relationships of die
tary indoles: a proposed mechanism of action as modifiers of xenobiotic me
tabolism. J. Toxicol. Environ. Health, 1987, 21, 311-323.
7. Bradlow H I . Nutrient modulation of female hormone metabolism: Mo
difying breast cancer risk. In: Functional Medicine Approaches to Endocrine
Disturbances of Aging. Vancouver, British Columbia: Institute of Functional
Medicine Proceedings; 2001.
8. Bradlow H I . , Michnovicz J., Halper M. et al. Long-term responses of
women to indole-3-carbinol or a high fiber diet. Cancer Epidemiol. Biomar-
kersPrev., 1994,3,591-595.
9. Bradlow H I . , Michnovicz J., Telang NT., Osborne M.P. Effects of die
tary indole-3-carbinol on esradiol metabolism and spontaneous mammary tu
mors in mice. Carcinogenesis, 1991,12,1571-1574.
10. Bradlow H I . , Sepkovic D.W., Telang NT., Osborne M.P. Multifun
ctional aspects of the action of indole-3-carbinol as an antitumor agent. Ann.
NY Acad. Sci., 1999, 889, 204-213.
11. Chang Y.C., Riby J., Chang G.H. et al. Cytostatic and antiestrogenic
effects of 2-(indol-3-ylmethyl)3,3'-diindolylmethane, a major in vivo product of
dietary indole-3-carbinol. Biochem. Pharmacol. 1999, 58, 825-834.

Метаболиты эстрадиоло и патологические процессы...
37. Michnovicz J.J., Hershcopf R.J., Naganuma H. et al. Increased 2-hy-droxylation of estradiol as a possible mechanism for the antiestrogenic effect of cigarette smoking. N. Engl. J. Med. 1986, 315,1305-1309.
38. Mori M., Tominaga Т., Tamaoki B.I. Steroid metabolism in normal mammary gland and in the dimethylbenzanthracene-induced mammary tumor of rats. Endocrinology, 1978,102,1387-1397.
39. Muti P., Bradlow H I . , Micheli A. et al. Estrogen metabolism and risk of breast cancer: a prospective study of the 2:16alpha-hydroxyestrone ratio in premenopausal and postmenopausal women. Epidemiology, 2000, 11, 635-640.
40. Naftolin F., Morishita H., Davies I.J., Todd R., Ryan K.J., Fishman J. 2-hydroxyestrone induced rise in serum luteinizing hormone in the immature male rat. Biochem. Biophys. Res. Commun., 1975, 64, 905-910.
41. Persson I. The risk of endometrial and breast cancer after estrogen treatment. A review of epidemiological studies. Acta Obstet. Gynecol. Scand. Suppl., 1985,130,59-66.
42. Schneider J., Huh M.M., Bradlow H I . , Fishman J. Antiestrogen action of 2-hydroxyestrone on MCF-7 human breast cancer cells. J. Biol. Chem., 1984,259,4840-4845.
43. Schneider J., Kinne D., Fracchia, Pierce V., Anderson K., Bradlow H.L, Fishman J. Abnormal oxidative metabolism of estradiol in women with breast cancer. Proc. Natl. Acad. Sci. USA, 1982, 79, 3047-3051.
44. Suto A., Bradlow H I . , Wong G.Y. et al. Experimental down-regulation of intermediate biomarkers of carcinogenesis in mouse mammary epithelial cells. Breast Cancer Res. Treat., 1993, 27,193-202.
45. Swaneck G.E., Fishman J. Covalent binding of the endogenous estrogen 16 alpha-hydroxyestrone to estradiol receptor in human breast cancer cells: characterization and intranuclear localization. Proc. Natl. Acad. Sci. USA, 1988, 85, 7831-7835.
46. Telang NT., Katdare M., Bradlow H.L. et al. Inhibition of proliferation and modulation of estradiol metabolism: novel mechanisms for breast cancer prevention by the phytochemical indole-3-carbinol. Proc. Soc. Exp. Biol. Med., 1997, 216,246-252.

ГЛАВА 3
25. Kirschner M.A. In Breast Cancer 3, Advances in Research and Treatment: Current Topics, ed. McGuire, W.(Plenum, New York), 1979, p. 199-226.
26. Klug I., Dressendorfer R., Strasburger С et al. Cortisol and 17-hy-droxyprogesterone levels in saliva of healthy neonates: normative data and relation to body mass index, arterial cord blood pH and time of sampling after birth. Biol. Neonate, 2000, 78, 22-26.
27. Levin M l . & Thomas D.B. In Breast Cancer, Progress in Clinical and Biological Research, eds. Montague AC, Stonesifer GL & Lewison EF (Liss, New York), 1977,12,9-35.
28. Lord R.S., Bongiovanni В., Bralley J.A. Estrogen metabolism and the diet-cancer connection: rationale for assessing the ratio of urinary hydroxyla-ted estrogen metabolites. Altern Med Rev., 2002, 7,12-29.
29. Lustig R., Kendrick-Parker C, Jordan V. Effects of 16 -hydroxyes-trone on MCF-7 cell proliferaton and estrogen receptor regulation in vitro. Endocr. Soc. Proa, 1994, 75, 317.
30. MacMahon В., Cole P. & Brown J. Etiology of human breast cancer: a review. J. Natl. Cancer Inst., 1973, 50, 21-42.
31. Martucci C, Fishman J. Direction of estradiol metabolism as a control of its hormonal action — uterotrophic activity of estradiol metabolites. Endocrinology, 1977,101,1709-1715.
32. Meilahn E.N., De Stavola В., Allen D.S. et al. Do urinary oestrogen metabolites predict breast cancer? Guernsey III cohort follow-up. Br. J. Cancer, 1998, 78,1250-1255.
33. Meng Q., Yuan F., Goldberg I.D. et al. lndole-3-carbinol is a negative regulator of estrogen receptor-alpha signaling in human tumor cells. J. Nutr., 2000, 130,2927-2931.
34. Meng Q., Qi M., Chen D.Z. et al. Supression of breast cancer invasion and migration by indole-3-carbinol: associated with up-regulation of BRCA1 and EOcadherin/catenin complexes. J. Mol. Med., 2000,78,155-165.
35. Michnovicz J.J. Increased estrogen 2-hydroxylation in obese women using oral indole-3-carbinol. Int. J. Obes. Relat. Metab. Disord., 1998, 22, 227-229.
36. Michnovicz J.J., Bradlow H I . Induction of estradiol metabolism by dietary indole-3-carbinol in humans. J. Natl. Cancer Inst. 1990, 82, 947-949.

ГЛАВА 3
47. Telang NT., Suto A., Wong G.Y. et al. Induction by estrogen metabolite 16 alpha-hydroxyestrone of genotoxic damage and aberrant proliferation in mouse mammary epithelial cells. J. Natl. Cancer Inst. 1992, 84, 634-638.
48. Terry P., Wolk A., Persson I., Magnusson C. Brassica vegetables and breast cancer risk. JAMA, 2001, 285, 2975-2977.
49. Wattenberg L.W., Loub W.D. Inhibition of polycyclic aromatic hydrocarbon-induced neoplasia by naturally occurring indoles. Cancer Res., 1978, 38,1410-1413.
50. Wong G.Y., Bradlow L, Sepkovic D. et al. Dose-ranging study of in-dole-3-carbinol for breast cancer prevention. J. Cell. Biochem. Suppl., 1997, 29,111-116.
МЕТАБОЛИЧЕСКИЙ СИНДРОМ КПК МАРКЕР ВНЕГОНАДНОГО СТЕРОИДОГЕНЕЗА В ПОСТМЕНОПАУЗЕ
Отрицательное влияние избыточного веса на здоровье человека известно еще с древних времен. Но по настоящему научное объяснение и целенаправленное исследование этого феномена началось с середины XX столетия, когда вполне четко сформировалось такое понятие, как болезни цивилизации. Однако формирование этого понятия стало возможным вследствие увеличения средней продолжительности жизни и накопления в популяции пожилых людей. Понятно, что при средней продолжительности жизни менее 40 лет влияние избыточной массы тела не могло стать объектом исследования.
Крупные многоцентровые исследования последних лет показали, что ожирение тесно связано с целым рядом заболеваний и синдромов. У женщин ожирение сопровождается высокой частотой ановуляций, нарушениями менструального цикла, бесплодием, гиперпластическими процессами эндометрия, высоким риском развития рака репродуктивных органов (шейка матки, вульва, эндометрий, яичники, молочная железа).
Установлено, что большинство взрослых американцев — 6 1 % , имеют избыточную массу тела и более четверти

Метаболический синдром кои маркер внегонадного...
Еще в 60-е годы прошлого столетия делались попытки объединения некоторых взаимосвязанных метаболических нарушений, ускоряющих развитие атеросклероза и сахарного диабета. В 1988 году G. Reaven, объединив нарушения углеводного обмена, артериальную гипертензию и дислипидемию, обозначил понятием «синдром X», предположив, что общей основой этих нарушений может быть инсулинорезистентность и компенсаторная гиперин-сулинемия. Автор описал синдром X у лиц с нормальной массой тела, однако в дальнейшем было показано, что в большинстве случаев при таких нарушениях имеется ожирение, как правило, по абдоминальному типу. Уже через год, в 1989 году, N. Kaplan описал «смертельный квартет», включив абдоминальное ожирение в число важных составляющих синдрома наряду с артериальной гипертонией, нарушением толерантности к глюкозе и гипертри-глицеридемией. В 1990-е годы метаболические нарушения и заболевания, развивающиеся у лиц с ожирением, объединили рамками метаболического синдрома. До настоящего времени концепция метаболического синдрома продолжает интенсивно разрабатываться, пополняя объем понятия новыми составляющими.
В истории формирования концепции метаболического синдрома особую значимость имеют исследования В.М. Дильмана (8, 9, 10, 11, 12, 13, 14) и Я.В.Бохмана (4, 5, 7, 17). Именно они впервые отметили взаимосвязь метаболического синдрома и рака репродуктивных органов. Фундаментальным исследованием стала работа Я.В. Бох-мана (17), касающаяся двух патогенетических вариантов рака тела матки. I (гормонозависимый) патогенетический вариант наблюдается у 60-70% больных и характеризуется многообразием и глубиной различных метабо-

ГЛАВА 4
— 26% страдают ожирением (ИМТ>30) (20). Многие исследователи отмечают, что сформировалась четкая тенденция омоложения ожирения. В США среди детей 6-17 лет распространенность ожирения возросла с 4% в 1963 г. до 10% и более в 1994 г.
Аналогичные изменения выявлены и в других промы-шленно развитых и развивающихся странах. В России ожирением страдают 54% населения, в Великобритании — 5 1 % , в Германии — 50%. В Китае ожирение отмечено у 15% взрослого населения, что вызывает крайнюю обеспокоенность медиков, учитывая численность населения этой страны (29).
Само по себе ожирение является важной проблемой для любого национального здравоохранения. Но в значительной степени эта проблема возрастает и усугубляется в сочетании с тяжелыми соматическими заболеваниями такими, как диабет 2 типа, артериальная гипертензия, ИБС, атеросклероз, онкологические заболевания. Целый ряд клинических и эпидемиологических исследований выявил и подтвердил, что наиболее значимыми медицинскими последствиями ожирения являются сахарный диабет 2 типа и сердечно-сосудистые заболевания. Было показано, что больные с абдоминальным типом ожирения часто имеют сочетание нескольких факторов риска ИБС. Абдоминальный тип ожирения был признан независимым от степени ожирения фактором риска развития сахарного диабета 2 типа и сердечно-сосудистых заболеваний. В последнее десятилетие многие ученые стали рассматривать различные метаболические нарушения и заболевания, ассоциированные с ожирением, в комплексе и высказывать предположения об общности этих нарушений.

Метаболический синдром как маркер внегонадного
- метаболический сосудистый синдром (Hanefeld М., 1997)
В индустриальных странах среди населения старше 30 лет распространенность метаболического синдрома составляет 10-30%, а у женщин это заболевание резко возрастает в менопаузальном периоде (30).
Как было уже указано, для метаболического синдрома характерно отложение жира в абдоминальной области. Исследования свидетельствуют, что наиболее значимо увеличение не столько подкожной, сколько висцеральной жировой ткани (16). Именно при этом типе ожирения имеют место:
- повышение кортизола - повышение тестостерона и андростендиона у женщин - усиленная трансформация андрогенов в эстрогены
(эстрон) - снижение соматотропного гормона - повышение инсулина - повышение норадреналина - повышение липопротеидов низкой плотности. Таким образом, ожирение, являясь патогенетическим
субстратом продукции эстрона в постменопаузе, становится как бы маркером гиперэстрогении.
С точки зрения нозологического обозначения метаболический синдром имеет весьма условные границы. В рамках сегодняшнего понимания основными симптомами и проявлениями метаболического синдрома являются: абдоминально-висцеральное ожирение, инсулинорезистентность и гиперинсулинемия, дислипидемия, артериальная гипер-тензия, нарушение толерантности к глюкозе (сахарный диабет типа 2), ранний атеросклероз (ИБС), нарушения го-меостаза, гиперурикемия и подагра, микроальбуминурия,

ГЛАВА 4
лических нарушений (ожирение, диабет 2 типа, гипертоническая болезнь). При II патогенетическом варианте указанные эндокринно-обменные нарушения выражены нечетко или совершенно отсутствуют.
В.М. Дильман обосновал гипоталамическую концепцию, обуславливающую многообразный спектр эндо-кринно-обменных нарушений и сформировал понятие канкрофилии, являющейся патогенетической базой при развитии злокачественных опухолей.
Многочисленные клинические и эпидемиологические исследования последних лет свидетельствуют, что увеличение и омоложение ряда опухолей репродуктивных органов у женщин идет параллельно с нарастанием индекса массы тела в популяции (2, 3, б, 15, 18, 22, 23, 24, 25, 27).
Таким образом, с каждым годом значимость метаболического синдрома все более расширяется и науке удается более детально и предметно обозначить патогенетические звенья, ведущие к развитию многих заболеваний, в том числе и онкологических. В медицинской литературе встречаются следующие синонимы метаболического синдрома:
- полиметаболический синдром (Avogaro P., 1965) - метаболический трисиндром (Camus J., 1966) - синдром «изобилия» (Mehnert A., 1968) - синдром X (Reaven G., 1988) - «смертельный квартет» (Kaplan J., 1989) - гормональный метаболический синдром
(Bjorntorp Р., 1991) - метаболический синдром (Hanefeld M., 1991) - синдром инсулинорезистентности (Haffner S., 1992) - смертельный секстет (Enzi G., 1994)

ГЛАВА 4
гиперандрогения. В последние годы ряд исследователей предлагают расширить рамки метаболического синдрома, включив стеатоз печени и синдром обструктивных апноэ во сне. Иными словами, конкретизация клинических и метаболических нарушений при метаболическом синдроме не завершена. С точки зрения клинического осмысления значимость нарушений и заболеваний, объединенных рамками синдрома, заключается в том, что их сочетание в значительной степени ускоряет развитие и прогрессирование социально значимых заболеваний, определяющих сегодня основной вектор причин смертности — сердечно-сосудистые и онкологические заболевания.
Распространенность метаболического синдрома в общей популяции довольно высока и колеблется от 14 до 24% (16, 19, 23). Лидируют в этом отношении высокоразвитые страны Северной Америки и Европы. Исследование распространенности метаболического синдрома, проведенное среди китайского населения в Гонконге, показало, что сочетание сахарного диабета типа 2, артериальной ги-пертензии и дислипидемии имеют 10% населения. Поданным американских исследователей метаболический синдром, согласно критериям Adult Treatment Panel III (ATP), в этой стране имеют в среднем 23,7% населения (т.е. 47 млн. человек) (19). При этом частота метаболического синдрома увеличивается с возрастом: так среди лиц от 20 до 29 лет он имеется у 6,7%, от 60 до 69 — у 43,5%. Среди молодых пациентов с ожирением в возрасте 16-22 лет частота метаболического синдрома составила 35%.
Основным и, пожалуй, наиболее манифестным звеном патогенеза метаболического синдрома, по мнению большинства исследователей, является первичная инсулино-резистентность и компенсаторная гиперинсулинемия. Ин-
Метоболический синдром кок маркер пнегонодного...
сулинорезистентность — это снижение биологического эффекта инсулина. Развивающаяся компенсаторная гиперинсулинемия, с одной стороны, вначале поддерживает углеводный обмен в норме, с другой — индуцирует развитие метаболических, гемодинамических и органных нарушений, приводящих в итоге к возникновению сахарного диабета типа 2 и сердечно-сосудистых заболеваний. В пользу ведущей роли инсулинорезистентности в развитии метаболического синдрома и отдельных его компонентов свидетельствуют результаты популяционного исследования, проведенного в Италии (авторы). Было выявлено, что инсулинорезистентность имелась у 58% обследованных с артериальной гипертензией, у 84% — с гипертриглицери-демией, у 42% — с гиперхолестеринемией, у 66% — с нарушениями к толерантности к глюкозе, у 95% — с метаболическим синдромом.
Большой толчок в развитии абдоминально-висцерального ожирения и инсулинорезистентности привносят гормональные нарушения, возникающие особенно на этапе формирования менопаузального гомеостаза. Развивающийся гормональный дисбаланс способствует накоплению жира в висцеральных депо и развитию и прогресси-рованию инсулинорезистентности. С увеличением общей жировой массы тела резко возрастает тканевой синтез эс-трона и его метаболитов. Причем, нарушение секреции половых гормонов сопряжено со значительным увеличением продукции гонадотропинов и выраженным дефицитом гормона роста. При абдоминальном ожирении нарушается и периферический метаболизм кортикостерои-дов: увеличивается метаболический клиренс кортизола. В адипоцитах сальника вследствие повышения активности 11в-гидроксистероиддегидрогеназы ускоряется превра-

ГЛАВА 4
щение неактивного кортизола в кортизол, который оказывает местное действие, стимулируя адипогенез в висцеральных депо (28).
Жировая ткань является источником синтеза ряда ци-токинов, из которых определенный интерес в рамках онкологической проблематики имеет фактор некроза опу-холей-а и интерлейкин-6. Многие исследователи рассматривают ФНО-а как медиатор инсулинорезистентности при ожирении — отмечены положительная корреляция между экспрессией ФНО-а и величинами ОТ/ОБ и ИМТ, а также снижение экспрессии ФНО-а и его концентрации в крови при уменьшении массы тела (21). Пропорционально нарастанию массы жировой ткани в крови увеличивается концентрация интерлейкина-6. Продукция интерлей-кина-б жировой тканью сальника в 2-3 раза превышает таковую подкожной абдоминальной жировой тканью (26). Следует заметить, что при метаболическом синдроме значимость сальника, как одного из наиболее активных анатомических зон биосинтеза целого спектра ферментов, гормонов, метаболитов, цитокинов, резко возрастает.
В настоящее время многими эпидемиологическими исследованиями установлена корреляция между ожирением, перееданием, гипертонией и т.д., с одной стороны, и частотой рака ряда локализаций, в особенности репродуктивных органов — с другой (15,18, 22, 23, 24, 25, 27). И эта связь в большей степени сопряжена с различными сторонами метаболического синдрома, из которых наиболее важным и определяющим становится гормональная составляющая канцерогенеза.
С позиций наших рассуждений, представляло определенный интерес проследить, имеется ли какая-либо связь или параллель между индексом массы тела (ИМТ) и уров-
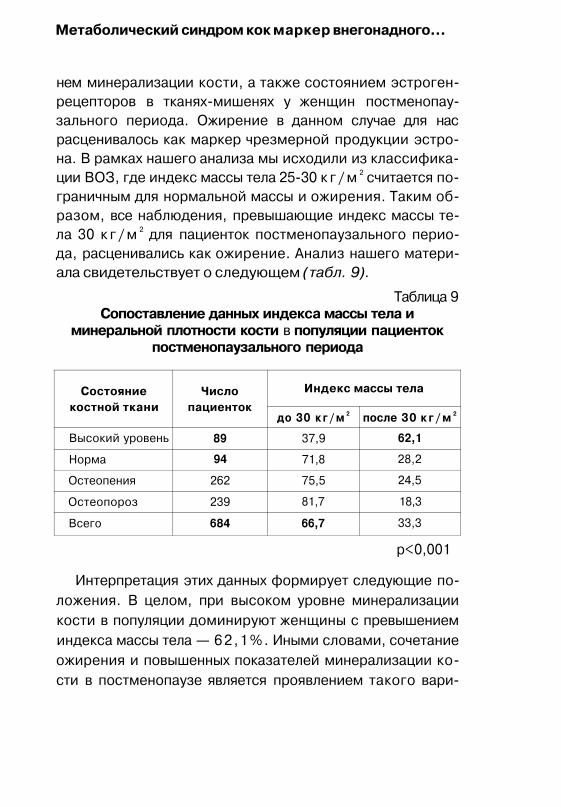
Метаболический синдром кок маркер внегонадного...
нем минерализации кости, а также состоянием эстроген-рецепторов в тканях-мишенях у женщин постменопау-зального периода. Ожирение в данном случае для нас расценивалось как маркер чрезмерной продукции эстро-на. В рамках нашего анализа мы исходили из классификации ВОЗ, где индекс массы тела 25-30 к г / м 2 считается пограничным для нормальной массы и ожирения. Таким образом, все наблюдения, превышающие индекс массы тела 30 к г / м 2 для пациенток постменопаузального периода, расценивались как ожирение. Анализ нашего материала свидетельствует о следующем (табл. 9).
Таблица 9 Сопоставление данных индекса массы тела и
минеральной плотности кости в популяции пациенток постменопаузального периода
Состояние костной ткани
Высокий уровень
Норма
Остеопения
Остеопороз
Всего
Число пациенток
89
94
262
239
684
Индекс массы тела
до 30 к г / м 2
37,9
71,8
75,5
81,7
66,7
после 30 к г / м 2
62,1
28,2
24,5
18,3
33,3
р<0,001
Интерпретация этих данных формирует следующие положения. В целом, при высоком уровне минерализации кости в популяции доминируют женщины с превышением индекса массы тела — 6 2 , 1 % . Иными словами, сочетание ожирения и повышенных показателей минерализации кости в постменопаузе является проявлением такого вари-

Метаболический синдром кок маркер внегонодного...
Однако в эту логическую цепочку не совсем вписываются наблюдения, где имеется сочетание ожирения с остео-пений и остеопорозом и наоборот, высоких показателей минерализации кости и нормального индекса массы тела. Анализируя эти данные, мы в конечном итоге пришли к необходимости учета такого фактора, как уровень эстроген-рецепторов в тканях-мишенях. Именно взаимосвязь эстроген — эстроген-рецептор в постменопаузе может обеспечить протективный эффект на кость равно как и стимулировать гиперпластические процессы в органах-мишенях. Эти рассуждения привели нас к идее изучения уровней эстроген-рецепторов раздельно при I и II варианте постменопаузы. Результаты отражены в таблицах 10, 11, 12.
Таблица 10 Сопоставление данных индекса массы тела и уровня
рецепторов-эстрогенов в молочной железе
Уровень рецепторов
ЭР+++
Э Р *
Число пациенток
11
53
Индекс массы тела
до 30 кг/м 2
27,2
67,9
после 30 к г / м 2
72,8
32,1
р<0,05
Таблица 11 Сопоставление данных индекса массы тела и уровня
рецепторов-эстрогенов вульвы
Уровень рецепторов
ЭР+++
ЭР-*
Число пациенток
21
43
Индекс массы тела
до 30 к г / м 2
47,6
79,1
после 30 к г / м 2
52,4
20,9 |
р<0,05

ГЛАВА 4
анта метаболического синдрома, когда с наибольшей вероятностью можно говорить о гиперэстрогении. Эта точка зрения подкреплена в исследовании Н.Б. Акоповой (2002) на материале больных краурозом вульвы, раком молочной железы, гиперпластическими процессами и раком эндометрия, где установлена кореллятивная взаимосвязь между раком эндометрия и молочной железы, высоким уровнем минерализации кости и ожирением. Наоборот, при явно дистрофических процессах в кости (остеопения, остеопороз) данные индекса массы тела свидетельствуют об отсутствии ожирения у подавляющего числа пациенток: при остеопении в 75,5% наблюдений, при остеопоро-зе в 81,7%. Подобная ситуация отмечена и у большинства пациенток с краурозом вульвы (1). Таким образом, при явно дистрофических процессах в кости (остеопения, остеопороз) данные индекса массы тела свидетельствуют об отсутствии ожирения у преимущественного числа пациенток. Иными словами, при отсутствии проявлений метаболического синдрома как бы отсутствует патогенетическая база избыточной продукции эстрона и состояние может быть обозначено как гипоэстрогенная.
Таким образом, как уже было отмечено, популяция пациенток в постменопаузе может быть условно разделена на две группы:
- I гипоэстрогенный вариант постменопаузы, для которой характерен низкий ИМТ, высокая частота остеопении и остеопороза, относительное превалирование крау-роза вульвы;
- II гиперэстрогенный вариант постменопаузы, где доминируют проявления метаболического синдрома (ожирение), высокая частота гормонозависимых опухолей в органах-мишенях (рис. 8)

Метаболический синдром кок маркер внегонадного.
Вполне очевидно, что вероятная модель варианта постменопаузы должна формироваться не только с учетом возможного гормонального фона, но и с учетом уровня эстроген-рецепторов в органах-мишенях. Опираясь на эти рассуждения, можно предположить следующие варианты гормонального гомеостаза в постменопаузе (рис. 9).
Таким образом, можно говорить о 4 вариантах постменопаузы, которые наиболее полно отражают сочетание гормонального и рецепторного статуса, формирующегося в постменопаузе. Нам представляется, что подобное разделение постменопаузы дает возможность уже сегодня более целенаправленно решать вопросы заместительной гормональной терапии, с одной стороны. С другой, возникает достаточно реальная перспектива патогенетического подхода к профилактике и ранней диагностике гормонозависимых опухолей женской репродуктивной системы.
Возвращаясь к концепции Я.В. Бохмана (1968, 1972) о двух патогенетических вариантах рака эндометрия, надо заметить, что без учета рецепторного статуса действительно в группе больных раком эндометрия дифференцируются две категории пациенток: пациентки с явным превышением индекса массы тела и с сопутствующими признаками метаболического синдрома (гиперэстрогенный тип) и пациентки с нормальным индексом массы тела без явных признаков метаболических нарушений (гипоэстро-генный тип). Однако в каждой из двух категорий пациенток имеются варианты опухоли с высоким и низким рецеп-торным статусом. Иными словами, если, наряду с конституциональным статусом больных учитывать и уровень эстроген-рецепторов, то четко дифференцируются уже че-

ГЛАВА 4
Таблица 12 Сопоставление данных индекса массы тела и уровня
рецепторов-эстрогенов в эндометрии
Уровень рецепторов
ЭР+++ ЭР-*
Число пациенток
14
50
Индекс массы тела
до 30 кг/м 2
21,4
72,0
после 30 кг/м 2
78,6
28,0 |
р<0,05
* ЭР- соответствует данным в диапазоне ЭР++ до ЭР-
Анализируя эту часть работы, можно констатировать ряд моментов, имеющих определяющее значение в трактовке тех несоответствий, о которых упоминалось выше. Как это видно из таблиц, во всех ситуациях, где отмечены высокие уровни эстроген-рецепторов, имеется превалирование повышенного индекса массы тела. Причем, результаты, полученные для молочной железы и эндометрия, весьма схожи: 72,8% и 78,6% соответственно. При вульве эти данные несколько ниже — 52,4%. Таким образом, одно из проявлений метаболического синдрома, выражающегося в ожирении и, соответственно, в гиперэс-трогении, обеспечено достаточно высоким пулом эстроген-рецепторов в тканях-мишенях. Наряду с этим, мы наблюдаем и то, что у определенной части пациенток при высоком уровне эстроген-рецепторов в органах-мишенях индекс массы тела не превышает границы нормальных величин. Наоборот, там, где имеется ожирение, сохраняется процент наблюдений с низким уровнем рецепторов: при молочной железе — 32,1%, вульве — 20,9% и эндометрии — 28,0%.

Метаболический синдром кок маркер внегонадного.
Список литературы к Главе 4
1. Акопова Н.Б. Возможности рентгеновской денситометрии в диагностике инволюционно-атрофических и гиперпластических процессов у женщин в постменопаузальном периоде, Дис. канд. мед. наук, Москва, 2002 г.
2. Берштейн Л.М. Гормональный канцерогенез, СПб, Наука, 2000 3. Берштейн Л.М. Эстрогены, старение и возрастная патология,
Усп. герант., 1998, т. 2, с. 90-97. 4. Бохман Я.В. Об особенностях организма больных раком тела
матки, Вопр. онкологии, 1963, № 9, с. 30-37. 5. Бохман Я.В. Рак тела матки, Кишинев: Штиница, 1972. 6. Бохман Я.В. Руководство по онкогинекологии, Ленинград, Меди
цина, 1989. 7. Бохман Я.В., Данова Л.А. О предрасположенности к раку тела
матки, Совр. проблемы онкологии, Л., 1967, с. 202-207. 8. Дильман В.М. Большие биологические часы, М, Знание, 1981. 9. Дильман В.М. Мутационно-метаболическая модель развития ра
ка и развитие опухолевого процесса, Вопр. онкологии., 1976, № 8, с. 3-16.
10. Дильман В.М. Нейро-эндокринный механизм старения и рак, Вопр. онкологии, 1967, № 10, с. 39-48.
11. Дильман В.М. Старение, климакс и рак., Л, Медицина, 196. 12. Дильман В.М. Четыре модели медицины, 1987. 13. Дильман В.М. Эндокринологическая онкология, Л, Медицина,
1974. 14. Дильман В.М., Бобров Ю.Ф. Гиперхолестеринемия и рак. В кни
ге: Современные проблемы онкологии. Под ред. А.И. Ракова, Л, Медицина, 1966, с. 76-80.
15. Abu-Abid S., Szold A., Klausner J. Obesity and cancer, J.Med., 2002,33(1-4)73-86.
16. Angel A. Pathophysiologic changes in obesity, Canad. Med. Ass. J., 1978, vol. 119, p. 1401-1406.
17. Bokhman J. Two pathogenetic types of endometrial carcinoma, Gynecol. Oncol.1983; 15:10-17.

ГЛАВА 4
тыре варианта рака эндометрия. Наши рассуждения сводятся к тому, что при любом варианте рака репрподук-тивных органов в постменопаузе достаточно четко дифференцируется четыре патогенетических варианта заболевания. В этом смысле заболевание несет на себе отпечаток того варианта постменопаузы, на фоне которого сформировался канцерогенез.
Обобщая эти данные, можно сказать, что одним из ключевых моментов гормонального гомеостаза в постменопаузе становится взаимодействие системы эстроген — эстроген-рецептор, формируя гиперпластическую или ин-волютивную направленность тканей-мишеней.

МОЛЕКУЛЯРНЫЕ ФПКТОРЫ КАНЦЕРОГЕНЕЗА
Как нормальная клетка становится раковой
Исследование молекулярных механизмов пролифера-тивных (гиперпластических) процессов и поиск путей их фармакологической коррекции — одна из самых динамично развивающихся областей современной молекулярной медицины. Понимание базисных основ индукции клеточного роста, особенно в условиях опухолевой трансформации клеток, является неотъемлемой частью грамотного подхода к управлению и мониторированию про-лиферативной активности (2). Ведь раннее выявление в организме очагов патологической пролиферации и направленная регуляция этих процессов открывают неограниченные возможности в профилактике и лечении таких заболеваний женской репродуктивной системы как мастопатии, эндометриоз, фибромиомы, диплазии шейки матки. В настоящее время подробно изучены ключевые механизмы активации сигнальных путей, стимулирующих клетки к патологической пролиферации.
На Рис.10 представлены этапы прогрессии неопластических заболеваний, начиная с хронических воспалительных изменений в нормальной ткани, которые впоследствии вызывают активацию пролиферативных процессов, развитие локальных опухолей и переход последних в ме-

ГЛАВА 4
18. Engeland A., Tretli S., Bjorge Т. Height, body mass index, and ovarian cancer, J. Natl. Cancer Inst.,2003, Aug 20;95(16):1244-8.
19. Ford E., Giles W., Dietz W. Prevalence of the Memetabolic syndrome among US Adults, JAMA, 2002; 287:356-9.
20. Harlan W., Landis J., Regal K. Secular trends in body mass in the United States, Am.J.Epidemiol, 1988,128:1065.
21. Hauner H., Bender M., Hube F. Plasma concentration of TNFa and its soluble receptors in obese subjects, Int. J. Obes. Relat. Metab. Disord., 1998,22:1239-43.
22. Jonsson F., Wolk A., Pedersen N. et al. Obesity and hormon-depen-dent tumors, Int J Cancer, 2003, Sept. 10; 106(4):594-9.
23. Kaaks R., Lukanova A. Effects of weight control and physical activity in cancer prevantion: role of endogenous hormone metabolism, Int. Agency for Research on Cancer, France.
24. Kaaks R., Lukanova A., Kurzer M. Obesity, endogenous hormones and endometrial cancer risk: synthetic review, Cancer Epidemiol. Biomarkers Prev, 2002, Dec; 11(12):1531-43.
25. Lapinska-Szumczyk S., Emerich Janusz. Obesity, hypertension and diabetes melitus in patients with endometrial cancer, /Ginecol. Pol., 2003, Apr; 74(4):274-81.
26. Papanicolaou D. lnterleukin-6: the endocrine cytokine, J.Clin. Endocrinol. Metab, 2000; 85:1331-33.
27. Parker E., Folsom A. International weight loss and incidence\of obesity-related cancer: the Iowa Women,s Health Study, Int.J.Obes.Relat.Me-tab.Disord., 2003, Dec;27(12):1447-52.
28. Rask E., Olson Т., Soderberg S. Tissue-specific dysregulation of Cortisol metabolism in human obesity, J Clin. Endocrinol. Metab., 2001; 86:1418-21.
29. Seidell J. The worldwide epidemic of obesity. 8th International congress on obesity, London, 1999; 661-8.
30. Woods S., Porte D. The central nervous system, pancreatic hormones, feeding and obesity.- In: Advencer in metabolic disorder/ Ed. R. Levin, R. Luft. New-York, 1978, vol. 4, p. 283-312.

ГЛАВА 5
ков. Все эти белки повышают чувствительность клеток молочной железы к факторам, индуцирующим гиперпластические процессы. В случае, если в организме женщины имеется повышенное содержание одного из производных эстрогенов, а именно 16а-гидроксиэстрона, эти процессы многократно усиливаются (6). Именно поэтому повышенное содержание 16а-гидроксиэстрона рассматривается в настоящее время как фактор риска развития рака молочной железы.
Однако, как уже упоминалось, существуют эстроген-независимые варианты развития опухолей. Какие же механизмы «работают» в этом случае?
Цитокиновый путь регуляции связан с фактором некроза опухоли (TNF-a). В больших концентрациях этот цито-кин активирует про-апоптотические (рецептор-опосредованные) сигнальные каскады, то есть останавливает процессы клеточного деления и вызывает физиологическую гибель клеток. Однако в малых дозах — действует как фактор выживания и пролиферации. При этом стимулируется активность циклооксигеназы-2 (СОХ-2) — основного фермента, участвующего в биосинтезе простагландинов (PGE2). Последние являются активаторами ядерного фактора NF-kB, который включает экспрессию генов эпидер-мального фактора роста (EGFR), фактора роста керати-ноцитов (KGF), циклин-зависимых киназ (CDK), фактора роста эндотелия сосудов (VEGF), инсулино-подобного фактора роста (IGF). Все эти белки, как уже упоминалось, стимулируют клеточное деление.
Наконец, третий путь, через который происходит активация клеточного деления в молочных железах — это сигнальные каскады, стимулируемые ростовыми факторами. Для опухолей молочной железы это, чаще всего, эпидер-мальный фактор роста (EGF), который также способен ак-

Молекулярные факторы канцерогенеза
тивировать ядерный фактор транскрипции NF-kB и через него стимулировать клеточное деление.
Таким образом, в настоящее время мы достаточно хорошо осведомлены о патогенетических механизмах развития опухолей молочной железы. Очевидно, что, блокируя основные каналы поступления сигналов, стимулирующих пролиферацию, мы можем рассчитывать на успех в профилактике и лечении этой патологии. Другими словами, фармакологическая коррекция должна осуществляться на всех этапах и по отношению ко всем сигналь- ] ным каскадам, которые выполняют ключевые патофизиологические функции.
С п и с о к литературы к Главе 5
1. Боженко В.К. Молекулярные маркеры рака молочной железы. В сб. «Клиническая маммология» (под ред. В.П. Харченко, Н.И. Рожковой) (вып. 1), «Фирма СТРОМ», М., 2005.
2. Киселев В.И., Ляшенко А.А. Молекулярные механизмы регуляции гиперпластических процессов, Изд-во «Димитрейд График Групп», 2005, монография; 346 стр.
3. Baud V., Karin M., Signal transduction by tumor necrosis factor and its relatives, Trends Cell Biol, 2001,11, 372-377.
4. Bukovsky A., Caudle M.R., Cekanova M. et al. Placental expression of estrogen receptor beta and its hormone binding variant — comparison with estrogen receptor alpha and a role for estrogen receptors in asymmetric division and differentiation of estrogen-dependent cells, Reproductive Biology and Endocrinology, 2003,1:36.
5. Cao Y. & Karin M. NF-kappaB in mammary gland development and breast cancer, J. of Mammary Gland Biology and Neoplasia, 2003, 8, 215-23.
6. demons M., Goss P. Estrogen and risk of breast cancer. N. Engl. J. Med., 2001, 344 (4), 276-285.

ИНФЕКЦИОННЫЕ ПГЕНТЫ И КАНЦЕРОГЕНЕЗ
Инфекционные болезни — одна из великих и трагических страниц человеческой цивилизации. До последнего времени инфекционные и паразитарные болезни являются ведущими причинами смертности. Ежегодно в мире умирает 50 млн. человек. Почти 17 млн. — от инфекций и паразитов. Для сравнения: сердечно-сосудистые болезни уносят около 10 млн. человек.
Наш мир наполнен миром микроорганизмов. В процессе длительной эволюции возникли различные формы симбиоза, т.е. биологического сосуществования различных организмов. Между микроорганизмами и высшими организмами сформировалась сложная внутренняя постоянная связь, необходимая, но в то же время изменчивая в ходе эволюции. И.В. Давыдовский рассматривал инфекционную болезнь как своеобразный процесс нарушения адаптации (И.В. Давыдовский, 1962). Это совершенно справедливо: в эру антибиотиков мы наблюдаем интенсивный процесс селекции возбудителей. Ответом на массовое, зачастую неразумное использование препаратов становится приобретение возбудителями новых свойств, увеличивающих их патогенность и вирулентность. Классические примеры того — пенициллиноустойчивые пневмококки и гонококки, полирезистентные штаммы возбудителей тропической малярии, туберкулеза, токсигенные

ГЛАВА 5
7. Fishman J., Osborne M.P., Telang NT. The role of estrogen in mammary carcinogenesis, Ann. NY Acad. ScL, 1995, 768, 91-100.
8. Ford Ch. HJ. Impact of Molecular Biology on Cancer Treatment: I Therapeutic Targets. Kuwait Medical Journal, 2003,35 (4), 253-262.
9. Frasor J., Danes J.M., Komm B. et al, Profiling of estrogen up- and down-regulated gene expression in human breast cancers cells: insights into gene networks and pathways underlying estrogenic control of proliferation and cell phenotype, Endocrinology, 2003,144,4562-4574.
10. Ghosh S. & Karin M. Missing pieces in the NF-B puzzle, Cell, 2002, 109 81s-96s.
11. Gomperts B.D., Kramer I.M., Tatham PER. Signal Transduction. Elsevier Academic Press, USA, Orlando, Florida, 2003, p. 257-392.
12. Gustafsson J.A., Warner M. Estrogen receptor in the breast: role in estrogen responsiveness and development of breast cancer, J. Steroid Biochem. Mol. Biol., 74, 2000, 245-248.
13. Lacroix M., Toillon R-A, Leclercq G. Stable 'portrait' of breast tumors during progression: data from biology, pathology and genetics. Endocrine-Related Cancer, 2004,11, 497-522.
14. Mosselman S., Polman J., Dijkema R. ER beta: identification and characterization of a novel human estrogen receptor, FEBS Lett, 1996, 392(1), 49-53.
15. Roger P., Sahla M.E., Makela S. et al. Decreased Expresion of Estrogen Receptor Protein in Proliferative Preinvasive Mammary, Tumors Cancer Research 61, 2001, 2537-2541
16. Ross J.S., Linette G.P., Stec J. et al. Breast cancer biomarkers and molecule medicine. Expert Rev. Mol. Diagn., 2003, 3(5), 573-585
17. White R., Parker M.G., Molecular mechanism of steroid hormone action, Endocrine-Related Cancer, 1998, 5,1-14
18. Yarden Y., Sliwkowski M. Untangling the ErbB signaling network, Nat. Rev. Mol. Cell. Biol., 2001, 2,127-137. V

Инфекционные агенты и канцерогенез
остатических систем, можно предположить, что результатом этого могут стать нарушения во многих системах, в том числе в системе взаимодействия макро-микроорганизм. Система урогенитального синуса является весьма важной экосистемой, где даже минимальные сдвиги приводят к обострению разнообразного спектра инфекции.
Воспалительные заболевания женских половых органов занимают первое место (55-70%) в структуре гинекологической заболеваемости. Значительную долю в них занимают инфекции вульвы и влагалища. В постменопаузе проблема эта приобретает дополнительное звучание и в определенном смысле усложняется. Общеизвестно, что все воспалительные процессы гениталий делятся на неспецифические и вызванные инфекцией, передающейся половым путем. В последнее десятилетие под влиянием антибиотиков второго и третьего поколений (полусинтетические пенициллины, цефалоспорины, аминогликози-ды) в корне изменился спектр микроорганизмов, доминирующих в этиологии воспалительных процессов вульвы и влагалища. Все чаще стали встречаться микробные ассоциации, обуславливающие более тяжелое течение процесса или же протекающие в виде хронической, невыраженной клинически заболеванием. С биологической точки зрения влагалище необходимо рассматривать в качестве очень важной экологической ниши, которая характеризуется рядом параметров: концентрация определенных биологически активных веществ (ферменты, микроэлементы, гормоны, белки, аминокислоты, простагландины), рН, окислительно-востановительный потенциал, макроорганизма и что для нас особенно важно, функциональной активностью яичников температура, ассоциация микроорганизмов и пр. Эти идея высказана ещё в 1988 году

ГЛАВА 6
стрептококки группы А, вызывающие синдром токсического шока. В последние годы широкое распространение получил термин «новые инфекции». Сегодня можно высказать три вероятных пути обозначения возбудителя, как нового. Первый — выделение известных ранее микроорганизмов или вариантов известных, вызывающих «новую» болезнь из группы клинически сходных заболеваний, прежде трактовавшихся, как одна нозологическая форма. Пример — грипп, гепатиты, диареи. Второй — появление неизвестной болезни. Сравнительно недавно открыты геморрагические лихорадки Лаоса, Эбола, вирусы иммунодефицита человека, др. инфекции. Третий путь — выявление возбудителей болезней, ранее считавшихся неинфекционными. Это самая многочисленная группа среди так называемых «новых инфекций». В неё входят, к примеру, первичная карцинома печени (вирус гепатита С), гастриты, язва и рак желудка или В-клеточная лимфома желудка (Helicobacter pylory), рак шейки матки (вирус папилломы человека), саркома Калоши (вирус герпеса 8 типа). По материалам, опубликовавнным ВОЗ, до 84% некоторых форм рака этиологически связаны с вирусами, паразитами или бактериями (Cancer Pr, 1996). Согласно оценкам экспертов ВОЗ, до 1,5 млн. новых случаев рака можно избежать, используя методы профилактики инфекционных и паразитарных болезней. Особое место среди «новых инфекций» занимает сочетание инфекции, когда в результате одновременного или последовательного воздействия разными инфекционными агентами клинические проявления болезни претерпевают значительные изменения.
Рассматривая постменопаузу, как период резкой и весьма разнообразной качественной перестройки многих гоме-

Инфекционные агенты и канцерогенез
тельной тканью. Резко возрастает частота микротравм, трещин и мацераций кожи. В связи с истончением эпидермиса создаются реальные условия, когда микротравмы и трещины могут достигать базального слоя. Условно патогенная и патогенная инфекция в этих условиях способна вегетировать на уровне всех слоев эпидермиса, вызывая длительно текущий, хронический воспалительный процесс, со всеми вытекающими для воспаления признаками. Кроме того, развитие инволютивно-атрофических процессов в зоне больших и малых половых губ, а также снижение тонуса мышц промежности, обеспечивающих сужения вульварного кольца, нивелируют роль такого важного биологического барьера, как сомкнутое состояние половой щели. В итоге, с некоторой относительностью можно говорить о формировании единой биологической среды, включающей в себя вульву, влагалище и шейку матки.
Вульвовагинальная зона является наиболее ярким примером конфликта в относительно стабильной экосистеме при формировании менопаузы. Естественно, что с той или иной интенсивностью подобные конфликты имеют место в любых эстрогензависимых зонах (эндометрий, яичники, протоки молочных желез и др.). В предыдущих главах мы акцентировали наше внимание на резкое увеличение пула онкологической заболеваемости органов репродуктивной системы именно в постменопаузе. Несомненно, что одним из важных составляющих в этиопатогенетиче-ской цепи канцерогенеза при этом становится инфекционный фактор.
В рамках наших рассуждений изучена микрофлора (ПЦР-диагностика) вульвы, влагалища и шейки матки (цервикальный канал) у 76 женщин постменопаузального периода (Табл. 13).

ГЛАВА 6
В. Larsen. Состав микрофлоры определяется разнообразием взаимоотношений видов микроорганизмов, состоянием иммунной системы.
Дефицит эстрогенов в постменопаузе включает цепь разнообразных событий, конечным звеном которых становятся ряд хронических патологических состояний, а у ряда пациенток — формируется злокачественная опухоль. Следует сказать, что изменения, происходящие в вульвовагинальной области в постменопаузе, достаточно хорошо изучены и описаны (7, 8, 9, 10, 11, 12, 13, 15, 16, 19, 20, 22, 23, 28, 30, 31, 34). В основе этих изменений, как было отмечено, лежит эстрогенная недостаточность. Именно эстрадиол способствует дифференциации вуль-вовагинального эпителия, формирует соответствующую экосистему этой зоны, оказывает благоприятное влияние на ангиогенез и способствует формированию хорошо развитой сосудистой системы. Под влиянием эстрадиола поверхностные слизистой вульвы и влагалища в значительном количестве накапливают гликоген. Лактобактерии и ацидогенные коринебактерии используют этот богатый запас гликогена, продуцируя органические кислоты, которые и поддерживают кислую реакцию среды (рН 3,8-4,2). Именно благодаря кислой реакции подавляется избыточный рост патогенов. Отсутствие эстрадиола приводит к исчезновению гликогенсодержащих поверхностных клеток, что приводит к ощелачиванию среды и резкому изменению микробиоценоза. Формируется агрессивная микрофлора из условнопатогенных, анаэробных бактерий и вирусов. Наряду с изменениями поверхностного эпителия редуцируется богатая сосудистая система, многослойный дермальный эпителий вульвы, влагалища, шейки матки резко истончается и замещается соедини-

Инфекционные агенты и канцерогенез
вается в постменопаузе. Обращает на себя внимание разнообразный спектр различных инфекционных агентов. При этом относительно чаще диагностируется стафилококк, грибы кандида, гарднерелла и вирус простого герпеса 2 се-ротипа, а для цервикального канала и серотипы вируса папилломы человека высокого риска. Кроме того, результаты исследования свидетельствуют о достаточной близости инфекционного спектра влагалища и цервикального канала, что вполне объяснимо анатомической близостью этих зон, а также единой плоскоэпителиальной выстилкой.
Определенный интерес представлял вопрос и о частоте сочетаний различных инфекций, косвенно характеризующих глубину биоцинотических и местных иммунных нарушений (см. табл. 14).
Таблица 14 Частота инфекционных ассоциаций (в %) в
цервикальном канале, влагалище и с поверхности вульвы у пациенток постменопаузального периода
Анатомическая зона
Цервикальный канал
Влагалище
Вульва
Не обнаружено
32,4
36,8
15,9
Число инфекционных ассоциаций
2
8,2
7,9
43,4
3
26,6
14,5
28,9
более 3
32,8
40,8
11,8
Как видно из таблицы, во всех анатомических зонах отмечена достаточно высокая частота сочетаний различных инфекционных агентов. При этом опять же близки показатели для влагалища и цервикального канала.
В рамках данной проблемы изучен микробиологический фон яичников у пациенток постменопаузального пе-

ГЛАВА 6
Таблица 13 Частота выявления инфекционных агентов (в %) в
цервикальном канале, влагалище и с поверхности вульвы у пациенток постменопаузального периода
Инфекционный агент
Стафилококк
Кандида
Трихомонада
Гарднерелла
Гонококк
Микоплазма
Уреаплазма
Хламидия Кишечная палочка Цитомегалови-рус Вирус простого герпеса II
Вирус папилломы человека н.р.
Вирус папилломы человека в.р.
Символ, обозначе
ние
Stph
Cal
Tva
Gva
Ngo
Mho
Ure
Ctr
Esc
HCMV
HSVII
HPV Lr.
HPV H.r.
Церви-кальный
канал
16,4
20,8
8,2
10,8
2,6
3,9
5,8
6,6
2,2
1,3
10,8
2,4
10,4
Влагалище
18,4
22,4
7,9
13,2
2,6
3,9
6,6
6,6
3,9
1,3
9,2
3,9
6,6
Вульва
10,5
7,9
5,3
23,7
3,9
2,6
2,6
6,5
10,5
2,6
10,5
1,3 I
3,9
Р<0,5
Следует отметить, что представленные данные статистически недостоверны, вместе с тем, для нас важным было получить общее представление по этому вопросу и выяснение той тенденции в формировании микробиоценоза цер-викального канала, влагалища и вульвы, которая склады-

Инфекционные агенты и канцерогенез
плоскоклеточных раков, мы попытались дополнить этот раздел онкогинекологии целенаправленными исследованиями, которые обобщены в отдельной монографии (2).
Был изучен максимально возможный спектр ВПЧ в образцах опухоли у 80 больных раком вульвы (см. табл. 15). При этом амплификацию выделенной ДНК проводили, используя праймеры, позволяющие идентифицировать следующие типы ВПЧ: 6, 11, 42, 53, 54, 66, относящиеся к низкой степени риска, и 16, 18, 31, 33, 35, 39, 45, 51, 52, 55, 58, 59, 68, 73, 83, относящиеся к серотипам высокой степени риска.
Таблица 15 Частота (абс. число/ %) ДНК вируса папилломы человека
при инвазивном раке вульвы
Метод исследов.
ПЦР-диаг-ностика
Число больных
80
Не обнаружено
19
23,8
Серотипы
6
24*
30,0
11
24*
30,0
16
21
26,3
18
14(3*)
17,5
55
2*
2,5
59
3*
3,8
83
4(2*)
5,0
Р<0,05 * число серотипов, сочетающихся с другими серотипами
Для исключения возможных ложноположительных результатов, которые могут иметь место при ПЦР-диагно-стике, параллельно проведено исследование с помощью технологии Murex-Digene Capture II ДНК HPV, при которой также дифференцировались две группы пациенток: пациентки с серотипами низкого риска и пациентки с серотипами высокого риска. Подобный подход методом «двойного контроля» был крайне необходим, дабы исключить возможный минимальный процент ложнополо-

ГЛАВА 6
риода. Из всего спектра инфекционных агентов установлена лишь герпетическая инфекция: вирус простого герпеса 2 серотипа — 32% наблюдений, цитомегаловирус — 4 % , вирус Эпштейн-Барра — 12%. Полученные результаты исследования не позволяют в настоящее время однозначно высказаться о наиболее вероятных путях миграции герпетической инфекции в яичниковую ткань. Можно лишь в некоторой мере предположить, что инфецирова-ние происходит по принципу восходящей инфекции, о чем имеются единичные сведения (29, 32).
Таким образом, можно совершенно однозначно говорить о серьезных нарушениях в биоценозе генитального тракта. Обозначая роль инфекционного фактора в канцерогенезе, многие исследователи обычно подразумевают два момента: инфекция, как кофактор канцерогенеза, и инфекция, как основной этиологический фактор канцерогенеза. Как ни пародоксально, но сегодня, гораздо более убедительно можно говорить об этиологических факторах канцерогенеза, чем указать четкое место инфекционного агента в качестве кофактора канцерогенеза. Любые рассуждения в этом плане — это скорее попытка дополнить воображением и некоторой логикой недостающие звенья в патогенетической цепочке событий.
Папилломовирусная инфекция при гинекологическом раке
Накопившиеся в онкогинекологии за последние два десятилетия факты свидетельствуют о доминирующей роли папилломовирусной инфекции в этиологии (генезе) плоскоклеточного рака (рак шейки матки). Развивая идею причастности папилломовирусной инфекции к индуцированию


ГЛАВА 6
жительных заключений. Результаты же, полученные технологией Murex-Digene Hybrid Capture II, относятся к высокоспецифичным тестам.
На основе проведенных исследований, включающих два независимых варианта методик по идентификации ДНК вируса папилломы человека в группе 80 больных раком вульвы, установлено его присутствие по результатам ПЦР-диагностики, с корректированными технологией Murex-Digene Hybrid Capture II, в 76,2% наблюдений.
С наибольшей частотой при раке вульвы установлено наличие б и 11 серотипов. Причем, как правило, эти два серотипа выявлялись одновременно. Вероятно, подобная дупликация серотипов связана с крайне близкой идентичностью и последовательностью нуклеотидов в геноме па-пилломовируса человека у 6 и 11 серотипов. Эти данные, используя метод молекулярной гибридизации (dot blot technique), были получены и в исследованиях Gregory P. Sutton et al. (35). Ассоциация серотипов не ограничилась только 6 и 11 серотипами. На материале наших исследований проявилась ассоциация 18 серотипа с 55 (1 наблюдение), с 59 (1 наблюдение), с 83 (1 наблюдение), 55 серотипа с 59 (1 наблюдение), 59 и 83 серотипа (1 наблюдение). Наряду с этим, не отмечено ни одного наблюдения ассоциации какого-либо серотипа с 16 серотипом. Как правило, 16 серотип выявлялся изолированно. Доля пациенток, у которых выявлены серотипы 16 и 18, трактуемые (на модели рака шейки матки) как серотипы высокого онкологического риска, на нашем материале установлены в 43,8% наблюдений.
Вирусный канцерогенез можно условно разделить на несколько этапов (3, 4):
1. первичное инфицирование вирусом;



Рис. 16. Частота определения серотипов высокого и низкого риска ВПЧ (в %) в образцах серозного и муцинозного рака яичников с использованием метода ПЦР

Рис. 15. Частота определения серотипов высокого и низкого риска ВПЧ (в %) в образцах эпителиального рака яичников с использованием метода ПЦР

Инфекционные агенты и канцерогенез
2. персистенция генома вируса папиллом в эписо-мальной форме и возможность продукции вирусных частиц с последующей вторичной инфекцией;
3. интеграция вирусной ДНК в клеточный геном без видимой специфичности сайта интеграции; на этапах 2 и 3 начинают проявляться функции Е6 и Е7, нарушающих контроль деления клеток;
4. индукция мутаций в клеточной ДНК, вызывающая нестабильность клеточного генома;
5. селекция клона клеток с мутантной ДНК, содержащих интегрированную ДНК вирусов папилломы;
6. активное размножение данного клона клеток и рост опухоли.
В рамках вирусного канцерогенеза наиболее значим именно третий этап, определяющий новый качественный сдвиг во взаимоотношениях вируса и клетки хозяина.
Интеграция ДНК ВПЧ в геном клеток сопровождается двумя молекулярными событиями:
1. Встраивание вирусной ДНК в хромосому всегда сопровождается нарушением структуры гена Е2, который является репрессором Е7.
2. при интеграции ДНК-вируса синтез белка Е2 прекращается вследствие нарушения структуры соответствующего гена и активируется синтез белка Е7.
Таким образом, факт интеграции вирусной ДНК может быть зарегистрирован или изучением структуры гена Е2 (эти методы в стадии разработок) или измерением уровня синтеза онкобелка Е7. Из этого следует, что наличие он-кобелка Е7 в пробах из опухоли может рассматриваться как однозначное свидетельство вирус-ассоциированного злокачественного процесса. Неоспоримым достоинством Е7, как маркера, является то, что в норме этот белок в тка-

ГЛАВА б
нях не синтезируется. Его происхождение полностью связано с жизненным циклом интегративной формы ВПЧ-ин-фекции. Так же преимуществом метода является то, что берется материал непосредственно из органа-мишени. В нашей работе иммуноферментный анализ выполнен в 34 наблюдениях рака вульвы (28 больных с наличием ВПЧ в опухоли и б больных без ПЦР данных ВПЧ инфекции) — см. табл. 16. Из 28 больных с вирусассоциированным раком вульвы у 20 имели место 16 и 18 серотипы. В семи наблюдениях установлены 6/11 серотипы и в одном наблюдении 83 серотип. Во всех наблюдениях вирусассоцииро-ванного рака определялся онкобелок Е7.
Таблица 16 Уровень белка Е7 в образцах опухоли рака вульвы
Уровень белка Е7 (ng/ml)
Не обнаружен
До 1,0
1,1-5,0
5,1 — 10,0
Более 10,1
Всего
Число больных
15
5
5
2
7
34
Без ВПЧ
6
б
Серотипы ВПЧ
6/11
7
7
16
2
4
2
7
15
18
1
3
1
-
5
83
1
1
Положительные результаты получены только в тех пробах, где имелись серотипы 16 и 18. Значения онкобелка Е7 были весьма вариабельны и колебались в диапазоне 0,06-12 ng/ml. Из 6 больных с вируснезависимым опухолевым процессом онкобелок Е7 выявлен в 1 наблюдении (1,8 ng/ml), что требует отдельного изучения. Значимость изучения синтеза онкобелка Е7 в вирусиндуцируемых
Инфекционные агенты и канцерогенез
опухолях гораздо более обширна с диагностической, лечебной и прогностической точек зрения. Специфичность теста на белок Е7, вероятно, для отдельных серотипов различается, что не позволяет сегодня говорить о широком его использовании в ранней диагностике рака вульвы, учитывая спектр серотипов ВПЧ, встречающийся при этой локализации злокачественного процесса. В данном же фрагменте исследования мы преследовали одну цель — привнести еще одно доказательство причастности ВПЧ в канцерогенезе рака вульвы.
Следующая локализация, не относящаяся к традиционной модели плоскоклеточного рака — рак яичников. Интерес к этой локализации определялся тем, что в группе эпителиальных раков яичника отсутствует вариант плоскоэпителиального процесса. Следовательно, учитывая тропность папилломовирусной инфекции к плоскому эпителию, как бы автоматически снимался вопрос о причастности этой инфекции в генезе эпителиального рака яичников. Вместе с тем, в литературе имеются сведения о наличии ВПЧ в образцах неопластической трансформации эпителия яичников и эта частота варьирует от 32% до 52% (5, 14, 24, 36, 37, 38). Предметом изучения в настоящем разделе явились образцы ткани яичников 67 больных эпителиальным РЯ и 25 пациентов контрольной группы без признаков злокачественной патологии яичников (б). Из таблицы /7видно, что при ДНК-диагностике ПВИ в 51 (76,1%) из 67 наблюдений ЗЭОЯ ВПЧ был положительным (р<0,001). И только в 2 из 25 случаев (8,0%) нормальных тканей яичников зарегистрированы позитивные результаты (р<0,001). Причем, частота определения ВПЧ в образцах серозного (п=52) и муцинозного (п=15) РЯ имела практически одинаковые значения — 75,0% и

Инфекционные агенты и канцерогенез
матки, рак вульвы и рак влагалища продемонстрирована некая условность градации ПВИ в соответствии с ее онкологическим потенциалом, которая вероятно обусловлена локализацией опухолевого процесса. Поэтому в данной работе нами была проведена идентификация ДНК максимально широкого спектра типов ВПЧ как высокого онкологического риска, так и относящихся к низкой степени риска.
Установлено, что с наибольшей частотой эпителиальный РЯ ассоциируется с ВПЧ-18 (29,9%), ВПЧ-52 (29,9%), ВПЧ-55 (26,9%), ВПЧ-83 (17,9%), ВПЧ-39 (16,4%) серотипами. ПВИ 73, 31, 51, 45, 33, 66, 68 и 16 типов определялась в 11,9%; 10,4%; 9,0%; 6,0%; 4,5%; 3,0%; 3,0% и 3,0% наблюдений, соответственно, (рис.15). Наряду с этим, ни в одном образце эпителиального РЯ не отмечено присутствие ВПЧ-35, ВПЧ-58 и ВПЧ-59 типов и ВПЧ т. н. низкого онкологического риска.
Как правило, в большинстве случаев (53,0%) определялось сочетание двух и более серотипов ВПЧ высокого онкологического риска. Изолированная форма инфекции отмечена в 24 образцах (47,0%), из которых в 9 (37,5%) наблюдениях был выявлен 18 тип ВПЧ, в 6 (25,0%) наблюдениях — 52 серотип, по 3 (12,5%) наблюдений — 39 и 55 тип и по 1 наблюдению — 31, 51 и 31 серотипы. Не отмечено изолированных вариантов при детекции 16, 33, 45,68, 73 и 83 типов ПВИ.
Следует особо остановиться на случаях определения 16 и 18 серотипов ВПЧ в овариальных пробах. Так, доля больных эпителиальным РЯ положительных по этому признаку в нашем исследовании составила лишь 32,9%. Аналогичные данные были получены в исследовании опухолевой ткани яичников методом ПЦР-диагностики HJ.

ГЛАВА 6
80,0% соответственно (р>0,05). То обстоятельство, что ПВИ при различных вариантах РЯ выявляется статистически достоверно выше, чем в нормальных тканях, позволяет предположить ее причастность к возникновению и развитию ЗЭОЯ.
Таблица 17 Частота определения ВПЧ (абс. число/%) в образцах
опухолевой и нормальной тканях яичников с использованием метода ПЦР
Группа исследования
Эпителиальный рак яичников
в том числе: Серозный рак яичников
Муцинозный рак яичников
Контрольная группа
Число больных
67 100%
52 100%
15 100%
25 100%
ВПЧ-позитивные образцы
5 1 * 7 6 , 1 %
3 9 * * 75,0%
1 2 * * 80,0%
2* 8,0%
ВПЧ-негативные образцы
16 23,9%
13 25,0%
3 20,0%
23 92,0%
*р<0,001;**р>0,05
В дальнейшем наши взгляды были устремлены в направлении изучения гипотетически значимых серотипов ПВИ в онкогенезе РЯ. Надо сказать, что в мировой литературе существуют только сведения о детекции ВПЧ 16 и 18 типов в опухолевой ткани яичников (5, 36, 37). Более того, на моделях канцерогенеза злокачественных новообразований генитального тракта, включая рак шейки

ГЛАВА 6
Yang et al. (37). В то же время, результаты некоторых исследовательских работ, представленных в мировой периодической литературе с использованием методик иммуно-гистохимического анализа и гибридизации in situ, показали присутствие ПВИ 16 и 18 серотипов приблизительно в половине случаев ( 1 , 36). По всей вероятности, противоречивость полученных результатов может быть обусловлена использованием методов детекции вирусных инфекций, которые в сравнении с ПЦР-диагностикой обладают меньшей чувствительностью и специфичностью.
Значительный научный интерес представляло изучение вопроса о своеобразии ассоциации различных серотипов ПВИ при серозном и муцинозном вариантах ЗЭОЯ (рис.16). Проводя анализ полученных нами данных, выяснилось, что муцинозный РЯ с наибольшей частотой сочетается с ВПЧ 18 (80,0%) серотипа (р<0,001), а при серозном РЯ в большинстве овариальных проб выявлено присутствие ВПЧ 52 (36,5%) и 55 (32,7%) типов (р<0,01). Разница статистически достоверна.
Однако поскольку верификация диагноза ЗЭОЯ включает в себя проведение пункции брюшной полости через задний свод влагалища, что может увеличивать вероятность распространения вирусной инфекции, и как следствие приводить к ложноположительным результатам, мы сочли возможным проанализировать частоту выявления спектра серотипов ВПЧ в группах пациентов с наличием пункции на диагностическом этапе и без нее. Нами не было отмечено существенных различий частоты детекции серотипов ВПЧ в исследуемых группах пациентов (табл. 18). Это дает нам право аргументировано утверждать об истинности статуса ВПЧ-инфекции в опухолевой ткани яичников.

Инфекционные агенты и канцерогенез
Таблица 18 Частота определения серотипов ВПЧ (абс. число/%) в образцах опухолевой ткани яичников с учетом пункции
брюшной полости
Группа исследования
без пункции
с пункцией
Число больных
30 100,0%
21 100,0%
Частота серотипов ВПЧ
18
13 43,3%
7 33,3%
39
7 23,3%
4 19,0%
52
12 40,0%
8 38,1%
55
11 36,7%
7 33,3%
83
9 30,0%
3 14,3%
р>0,05
Подводя промежуточный итог из всей совокупности имеющихся на этот момент данных, мы хотели бы подчер-
ь два важных обстоятельства. Во-первых, ВПЧ, в частности 18, 52, 55, 83 и 39 его серотипы, по всей вероятности, обладают высокой тропностью в отношении эпителия
чиков. Во-вторых, принимая во внимание тот факт, что при сравнении спектра ПВИ в пробах муцинозного и серозного РЯ нами обнаружено достоверное различие в частоте детекции ВПЧ 18, 52 и 55 типов, по-видимому, указывает на важность дифференцированного подхода с позиций значимости того или иного серотипа для различных •лриантов эпителиального РЯ.
Между тем присутствие ПВИ в ткани не тождественно м участию в процессе опухолевого перерождения клетки. Такое заключение необходимо осмыслить в первую Ьмередь с позиции хронологии течения инфекционного коцесса, тогда станет понятным, какие из множества событий и определяют потенциальную готовность клетки к Ь/юкачественной трансформации.
Накопленные сведения показывают, что во взаимоотношении с клеткой хозяина ВПЧ может существовать в двух

ГЛАВА 6
состояниях — эписомальном и интегративном. Эписомаль-ная форма является инфекционной единицей, заражающей клетки. Персистенция генома ВПЧ во внехромасо-мальной форме обеспечивает продукцию вирусных частиц с последующей вторичной инфекцией. В определенных случаях происходит интеграция вирусного генома в генетический аппарат клетки хозяина, которая на молекулярном уровне сопровождается следующими событиями. Так, встраивание вирусной ДНК в хромосому всегда заканчивается полным разрушением генов Е 1 — Е 2. Последние отвечают за регуляцию транскрипции вирусных генов Е 6 и Е 7, что в свою очередь приводит к их неконтролируемой экспрессии и активному синтезу онкопротеинов Е 6 и Е 7. Не останавливаясь подробно на описании функциональных особенностей и механизмов реализации канцерогенного эффекта вирусных протеинов Е б и Е 7, которые были рассмотрены в главе, посвященной обзору литературных источников, и хорошо известны широкому кругу специалистов, подчеркнем лишь то положение, что процесс неопластической трансформации характеризуется интеграцией вирусной ДНК, выражающейся в продукции большого количества онкобелков Е б и Е 7 в исследуемой ткани.
В нашей работе анализ 85 овариальных проб (61 образец тканей ЗЭОЯ и 24 — нормальных тканей яичников) был проведен методом ИФА с использованием широкой панели моноклональных антител к различным детерминантам белка Е 7 для ВПЧ-18 серотипа (табл. 19).
Прежде всего, обратим внимание на итоги результатов по ВПЧ-18 позитивным образцам. Из 20 проб, забранных у больных эпителиальным РЯ I — IV стадий, продукция белка отмечена в 14 (70%) образцах преимущественно с ранними стадиями опухолевого процесса (р<0,0001).
Инфекционные агенты и канцерогенез
Таблица 19 Уровень онкопротеина Е 7 ВПЧ-18 в образцах
эпителиального рака яичников
Группа исследования
ВПЧ-18 позитивные пробы
ВПЧ-18 негативные пробы
I
Контрольная группа
Общее число
20 100%
41 100%
24 100%
Уровень онкобелка Е 7 (нг/мл)
отсутствует
6 30,0%
38* 92,7%
24* 100%
Д о 1
13
-
-
1,1-5,0
1
-
-
5,1-10,0
-
-
-
более 10,0
-
3
-
всего
14 70,0%
3* 7,3%
0 * 0 %
р<0,0001;*р>0,05
Показатель уровня ВПЧ-18 Е 7 протеина у пациенток варьировал в пределах пороговых значений и составил 0,3 — 1,2 нг/мл. Отсутствие вирусного протеина зафиксировано только в 6 (30%) пробах больных распространенным РЯ. Можно предположить, что синтез онкопротеина Е 7, начало которого в клиническом эквиваленте проецируется на стадию предрака, вероятно, в течение развития опухолевого процесса имеет тенденцию к снижению. Принимая во внимание незначительное число Е 7-по-зитивных наблюдений, нами не предусматривалось изучение зависимости продукции вирусного протеина от стадии распространения опухолевого процесса. Тем не менее, дальнейшая разработка этого направления позволит определить практическую ценность онкопротеина Е 7 в

Инфекционные агенты и канцерогенез
Герпесвирусная инфекция при злокачественных эпителиальных опухолях яичников
В настоящее время результаты немногочисленных эпидемиологических и экспериментальных научных работ свидетельствуют о том, что среди прочих вирусных агентов определенным онкогенным потенциалом могут обладать вирусы герпеса (вирус Эпштейн-Барр (ВЭБ), вирус простого герпеса 1 и 2 типов (ВПГ) и цитомегаловирус (ЦМВ)). По нашему мнению целенаправленный поиск кофакторов в первую очередь необходимо рассматривать с точки зрения дальнейшего изучения особенностей ПВИ при эпителиальном РЯ.
Определение обозначенных герпесвирусов мы провели в 67 образцах опухолевой и 25 образцах нормальной ткани яичников (рис.17).
Как нам удалось установить, частота определения ВЭБ-инфекции в основной группе составила 38,8% наблюдений и была статистически достоверно выше по сравнению с контрольной группой, где составила 12% наблюдений (р<0,01). Кроме того, в контрольной группе отмечалось повышение частоты детекции ВПГ 2 типа (32,0%), которая была в четыре раза выше, чем у больных ЗЭОЯ (7,5%) (р<0,001). Значения частоты ЦМВ-инфек-ции в основной и контрольной группе были практически равные — 3,0% и 4,0% наблюдений соответственно (р>0,05).
Таким образом, полученные нами результаты свидетельствуют о том, что с наибольшей частотой в опухолевой ткани определяется ВЭБ-инфекция, которая, как правило, носит сочетанный характер. Так, ассоциация ВЭБ с

ГЛАВА 6
качестве диагностического биомаркера ВПЧ-ассоцииро-ванных неопластических процессов яичников.
В группе ВПЧ-18 негативных образцов, онкобелок Е 7 был выявлен в 3 (7,3%) из 41 наблюдений (р<0,0001). При этом уровень белка имел критическое значение — более 20 нг/мл. Безусловно, этот эпизод нуждается в дополнительном изучении, однако в литературных источниках имеется описание феномена «элиминации» или «hit and run transformation» вируса в возникшей опухоли. Впервые на это явление обратил свое пристальное внимание Л.А. Зильбер, который в эксперименте на крысах продемонстрировал отсутствие вирусного агента в опухоли, индуцируемой только одним штаммом вируса куриной саркомы Рауса (Л.А. Зильбер и И.Н. Крюкова, 1958). Говоря о вирусе папилломы, следует отметить, что аналогичная модель была описана К.Т. Smith (1988), где в культивируемые клетки фибробластов мышей вводили линейную ДНК (т.е. интегрированную форму) бычьего вируса папилломы 4 серотипа и только в 5 (15%) из 21 трансформированных клеточных линий была идентифицирована вирусная ДНК, что возможно имеет место и в нашем случае.
Изучение уровня онкобелка Е 7 ВПЧ 18 серотипа в 24 образцах нормальных тканей яичников показало отсутствие синтеза последнего во всех пробах, в том числе и ВПЧ-позитивных (р<0,0001).
Разумеется, продукция вирусного белка Е 7 в тканях ЗЭОЯ, происхождение которого полностью связано с жизненным циклом интегративной формы ПВИ, наряду с его отсутствием в нормальной ткани яичников в определенном смысле может быть еще одним аргументом в пользу ВПЧ-ассоциированного опухолевого процесса.

ГЛАВА 6
ВПЧ отмечена в 28,4% наблюдений, а изолированное течение инфекционного процесса — в 10,4%. При этом чаще других установлено сочетание ВЭБ-инфекции с 52 (17,9%), 55 (13,4%) и 18 (6,0%) серотипами ВПЧ, т.е. типами высокого онкологического риска.
Если взглянуть на сведения, представленные в таблице 20, характеризующие частоту ВЭБ при различных вариантах эпителиального РЯ, становится очевидным, что для серозного РЯ свойственно увеличение частоты определения ВЭБ — этот показатель был положительным у 24 (46,2%) пациенток. В группе больных муцинозным РЯ только у 2 (13,3%) женщин был отмечен позитивный результат. Разница является достоверной (р<0,01).
В целом, представленные нами материалы о частоте определения ВЭБ в опухолевой ткани яичников наряду с доказанной моноклональностью генома вируса в опухолевой клетке дают основание предполагать, что ВЭБ-ин-фекция может выступать в качестве кофактора, приводящего к возникновению и развитию, по крайней мере, некоторой части ЗЭОЯ.
Таблица 20 Частота ВЭБ (абс. число/%) в образцах серозного и
муцинозного вариантов эпителиального РЯ с использованием метода ПЦР
Группа исследования
[ Серозный рак ! яичников
Муцинозный рак яичников
Число больных
52 100%
15 100%
ВЭБ-позитив-ные образцы
24* 46,2%
2* 13,3%
ВЭБ-негатив-ные образцы
28 53,8%
13 86,7% |
*р<0,01

Инфекционные агенты и канцерогенез
До настоящего времени роль ВЭБ-инфекции в контексте вирусного канцерогенеза обсуждалась в основном с позиций нарушения локального противоопухолевого иммунного ответа при инфицировании вирусом В-лимфоци-тов. Однако в работах целого ряда исследователей продемонстрировано, что ВЭБ также способен инфицировать и эпителиальные клетки, в частности носо-, ротоглотки и желудка (17, 2 1 , 27). Нам не встретились литературные данные об инфицировании ВЭБ эпителиальных клеток яичника.
Произведенные нами исследования показали, что ВЭБ-инфекция в опухолевой ткани яичников определяется в 38,8% наблюдений. Однако необходимо учитывать особенности применявшегося в работе метода ПЦР, а именно, то, что метод не отражает, в каком состоянии находились обнаруженные фрагменты ДНК. Мы не исключаем возможности, что это могли быть обрывки молекул, заносимые, например, лимфоцитами и макрофагами из других участков.
Вместе с этим следует добавить, что ВЭБ диагностирован приблизительно в половине случаев (46,2%) серозного РЯ, в то время как при муцинозном РЯ этот показатель не превышал 13,3% наблюдений. Объяснением этой закономерности может быть предположение о том, что ВЭБ-инфекция обладает тропностью именно к ПЭЯ, являющегося гистогенетическим источником серозной аде-нокарциномы.
Общеизвестно, что онкогенный потенциал ВЭБ обусловлен модифицирующим влиянием вирусного онкопротеи-на LMP1 на рецепторы эпидермального фактора роста в эпителиальных клетках, вероятно, путем взаимодействия его с клеточным протеином TRAFs (рецептор-ассоцииро-ванные факторы ФНО) (18, 25, 33). Некоторые исследо-

ГЛАВА 6
ватели также высказывают точку зрения о способности онкопротеина LMP1 ингибировать р53-зависимый механизм репарации ДНК в эпителиальных клетках (26). В создавшихся условиях синергическое действие ВЭБ и ВПЧ может приводить к более выраженным нарушениям генетического гомеостаза клетки. Так, по нашим данным ассоциация ВЭБ и ПВИ высокого онкологического риска отмечалась в 28,4% от всех наблюдений ЗЭОЯ.
Тем не менее, нельзя не сказать и о том, что у 7 (10,4%) больных серозным РЯ наблюдался изолированный тип или ВЭБ-ассоциированный эпителиальный РЯ. Обнаружение таких вариантов ЗЭОЯ создает предпосылки для проведения комплекса дальнейших исследований в рамках отдельной диссертационной работы, направленных на изучение как патогенеза, так и клинико-морфологических особенностей новообразований яичников, в возникновении которых принимает участие ВЭБ.
Таким образом, представленные данные, а также многочисленные факты, накопленные в онкологии, свидетельствуют о существенной роли инфекционного фактора в канцерогенезе. Наука находится в самом начале осознания инфекционной природы большинства онкологических заболеваний. Очевидно, что некоторые представления могут быть спорными. Но сегодня важно не цепляться за старые представления, плутать в кругу ложных понятий, сформированных многие десятилетия назад на примитивной исследовательско-технической базе. Многие отрасли медицины (генетика, вирусология, молекулярная биология, клиническая медицина) за последние годы накопили множество новых данных. Сегодня имеет смысл интегрировать этот фактический материал и обозначить наиболее верные и оптимальные пути продвижения в проблеме.

Инфекционные агенты и канцерогенез
С п и с о к литературы к Главе 6
1. Антонеева И.И. Экспрессия антигенов вирусов папилломы человека, простого герпеса и фенотип рецепторов эстрогенов и прогестерона в малигнизированных эпителиальных опухолях яичников, Авто-реф. дисс. канд. мед. наук, СПб, 2003, с. 24.
2. Ашрафян Л.А., Харченко Н.В., Киселев В.И. и др. Рак вульвы: эт-иопатологическая концепция, 2006, Москва.
3. Киселев Ф.Л. Вирус-ассоциированные опухоли человека: Рак шейки матки и вирус папилломы, Биохимия, 2000, № 1, с. 79-91.
4. Киселев Ф.Л. Вирусы папилломы человека как этиологический фактор рака шейки матки: значение для практики здравоохранения, Вопросы вирусологии, 1997, № 6, с. 248-251.
5. Максимов С.Я. Факторы риска возникновения злокачественных новообразований органов репродуктивной системы женщин, Вопр. онкологии, 2003, т. 49, с. 496-501.
6. Мухтаруллина СВ. Вирусная и бактериальная инфекция при злокачественных эпителиальных опухолях яичников, Дисс. канд. мед. наук, Москва, 2005.
7. Петрова А.С., Агамова К.А., Ермолаева А.Г. Роль и место цитологической диагностики в клинической практике, Клин, лабораторная диагностика, 1996, № 4, с.4-7.
8. Погосянц Е.Е. Генетика злокачественных новообразований, Проблемы мед. Генетики, М. Медицина, 1970, с. 267-282.
9. Попков С.А. ЗГТ в коррекции функциональных и метаболических нарушений у женщин с патологией сердца в климактерическом периоде, Дисс. докт. мед. наук, М., 1997
10. Розен В.Б., Смирнов А.И. Рецепторы и стероидные гормоны, М, 1981, с. 268-308.
11. Сафроний Д.Ф., Цырлина Е.В., Максимов С.Я., Обручков А.С. Динамика изменений уровня цитоплазматических рецепторов эстра-диола и прогестерона в опухолевой ткани рака эндометрия в группе больных чувствительных и нечувствительных к гормонотерапии та-моксифеном и его сочетании с ОПК, Тезисы Всесоюзного симпозиума

ГЛАВА 6
«Скрининг в раннем выявлении опухолей репродуктивной системы и проведение органосохраняющего лечения», Кострома, 1991.
12. Серов В.П., Прилепская В.Н., Пшеничникова Т.Я. Практическое руководство по гинекологической эндокринологии, М, 1995, с. 168-198.
13. Сметник В.П. Клинические аспекты климактерических расстройств, Материалы конференции, М, 1994, с. 4-8.
14. Antilla M., Syrjanen S., Saarikoski S. Failure to demonstrate HPV DNA in epithelial ovarian cancer by general primer PCR, Gyn. Oncol.,1999, Vol. 72, p. 337-341
15. Becagli L, Scrimin F., De Salva D. et al. Hormone receptors in dystrophic and neoplastic vulvar disease, Preliminary consideration, Clin. exp. Obstet. Gynecol, 1983, Vol. 23,145-49.
16. Crodstein F. et. al. Postmenopausal hormone therapy and mortality, N. Engl.J. Med, 1997, 336, P. 1769-1775.
17. Dadmarz R., Sgagias M., Rosenberg S. CD4+T lymphocytes infiltrating human breast cancer recognise autulogous tumor in an MHC-class-ll restracted fashion, Cancer Immunol. Immunother, 1995, Vol. 40(1), p. 1-9
18. di Maio D., Lai C, Klein 0. Viricrin transformation: the interaction between viral transforming protein and cellular signal transduction pathway, Annu. Rev. Microbiol, 1998-Vol. 52, p. 397-421.
19. Engeland A., Tretli S., Bjorge T. Height, body mass index, and ovarian cancer J. Natl. Cancer Inst., 2003, Aug 20; 95(16):1244-8.
20. Enzi G. et al. Bone mineral density and body composition in underweight and normal elderly subjests, Osteopor.lnt., 2000-11(12), p. 1043-50.
21. Epstein M. Historical background; Burkit's Lymphoma and Epstein-Barr virus, IARC Sci Publ, 1985, Vol. 60, p. 17-27.
22. Eriksen E.F., Colvard D.S., Berg N.J. et al. Evidence of estrogen receptor in normal human osteoblast-like cells, Science, 1988, 241-84.
23. Hebert-Croteau N.A. A meta — analysis of hormone replacement therapy and colon cancer in women, Cancer Epidemiol. Biomark.and Prevent, 1998,7, p. 653-660.
24. Lai C, Hsueh S., Lin С et al. HPV in benigh and malignant ovarian and endometrial tissues, Int. Gyn. Pathol., 1992,11, p. 210-215

Инфекционные агенты и канцерогенез
25. Li Н-Р., Chang Y-S. Epstein-Barr virus latent membran protein, J.Biomed.ScL, 2003, Vol.10(5), p. 490-504.
26. Liu M., Chang Y., Chen S. Epstein-Barr Virus latent membran protein 1 represses p53-mediated DNA repair and transcriptional activity, Oncogen, 2005, Vol. 24(16), p. 2635-2646.
27. Lopez V., Young L, Murray P. Epstein-Barr virus-associated cancer: Aetiology and Treatment. Herpes, 2003, Vol. 10(3), p. 78-82.
28. Lover R.R.,Mazess R.B. Barden H.S.et.aLEffects of Tamoxifen on bone marrow density in postmenopausal women with breast cancer, N. Engl.J.Med, 1992, 336, p. 852-856.
29. Manolitsas Т., Lanham S., Hitchcock A., Watson R. Synchronous ovarian and cervical squamous intraepithelial neoplasia: an analysis of HPV status, Gynec. Oncol., 1998, Vol. 70, p. 428-431.
30. Newcomb P.A., Longnecker M.P., Storer B.E. et al. Long-term hormone replacement therapy and risk of breast cancer in postmenopausal women, Am. J." Epidem, 1995,142, p. 788-795.
31. Omsjo I.H., Wright P.B., Bormer O.P. Estrogen and progesterone receptors in normal and malignant vulvar tissue, Ginecol Obstet Invest 1984, 17(6); 281-3.
32. Pins M., Young R., Crum С Cervacal squamous cell carcinoma in situ with intraepithelial extension to upper genital tract and invasion of tube and ovaries: report of case with human papilloma virus analysis, Int.J.Gynecol. Pathol, 1997, Vol.16, p.272-278.
33. Raab-Traub N. Epstein-Barr virus, lymphoproliferative diseases and nosopharyngeal carcinoma, Oxford University Press, Oxford, UK, 1999, p. 180-206.
34. Smith K.T., Campo M.S., «Hit and run» transformation of mouse C127 cells by bovin papillomovirus type 4: The viral DNA is reguired for the initiation but not for the maintenance of the transformed phenotype, Virology, 1988;164:39-47.
35. Sutton G.P., Stehman F.B. et al. Human Papillomavirus DNA in lesion of the famale Genital Tract: Evidence for Type 6/11 in Squamous Carcinoma of the Vulva, Obstet. and Gynec, Vol. 70, № 4,1987, P. 564-568.

ГЛАВА 6
36. Wu Q-J., Guo M., Li Т., Detection of papillomavirus-16 in ovarian malignancy, Brit. J.Cancer, 2003, Vol. 89, p. 672-675.
37. Yang H., Liu V., Tsang P. Comparison of HPV DNA in giynecologi-cal cancer, Tumor Biol, 2003, Vol. 24(6), p. 310-316.
38. Zimma K., Poreba E., Kedzia W. HPV in upper genital tract carcinoma of women, Eur.J.Gynecol. Oncol., 1997,Vol. 18, p. 415-417.

РАННИЙ PRK И ПРЕДРПК ШЕЙКИ МПТКИ, ОБУСЛОВЛЕННЫЕ ВИРУСАМИ ПАПИЛЛОМЫ ЧЕЛОВЕКА
Рак шейки матки является одной из наиболее распространенных форм новообразований у женщин. Ежегодно в мире регистрируется около 500 тыс. новых случаев заболевания. По данным ВОЗ за 2001 г. во всем мире эта локализация злокачественных опухолей у женщин занимает 2 место, уступая только раку молочной железы (19). По данным НИИ онкологии им. Герцена в 2000 г. заболеваемость и смертность от рака шейки матки среди женщин составила 12342 и 6288, соответственно. Выявляемость заболевания в зависимости от стадии составила:
1-2 стадия — 57,8%, 3 стадия — 29,2 %, 4 стадия — 10,4%. А летальность на первом году жизни с момента поста
новки диагноза — 20,2%. Следует более подробно остановиться на определе
ниях предрак и ранний рак. Итак, к предраку шейки матки относятся диспластические процессы шейки матки всех степеней выраженности. Дисплазия, по определению экспертов ВОЗ, представляет собой атипию эпителия шейки матки с нарушением слоистости, но без вовлечения в про-

ГЛАВА 7
цесс поверхностного слоя и стромы. Термин «дисплазия» был впервые предложен в 1953 г. J.W. Reagan и соавт. Они предложили деление на легкую, умеренную и тяжелую дисплазию. Условно дисплазия определена чисто гистологическими терминами: учитывались две главные особенности: клеточная атипия и степень дифференцировки (6).
В 1968 г. R.M. Richait ввел термин «цервикальная ин-траэпителиальная неоплазия» (cervical intraepithelial neoplasia — CIN). Его целью было показать, что это — различные стадии одного и того же процесса, прогрессирующего во времени через Са in situ (карцинома in situ) в ин-вазивный рак.
- CIN I соответствует легкой дисплазии, - CIN II — умеренной. - Поскольку трудно цитологически и гистологически
различить тяжелую дисплазию и CIS, они были объединены в CIN III.
Эта терминология принята Всемирной организацией здравоохранения (ВОЗ).
Несмотря на это, в конце 80-х годов S.L. Woodhouse и соавт. была предпринята новая попытка классифицировать неопластические процессы шейки матки. В результате были представлены следующие категории:
- SEA — squamous epithelial abnormal: доброкачественные изменеия плоского эпителия.
- ASCUS — atypical squamous cells of determined significance: атипические клетки плоского эпителия неопределенного происхождения.
- LSIL — low — grade Intraepithelial Lesion: поражения эпителия легкой степени, что соответствует CIN 1,
- HSIL — high — grade Intraepitelial Lesion: поражения эпителия тяжелой степени, что соответствует CIN 2 — 3.

Роннии рои и предрек шейки мотки, обусловленные...
Тем не менее, предпринятые усилия показали, что ошибки в дифференциальной диагностике между HSIL и LSIL очень велики. Трудность усугубляется тем, что часто HSIL имеет компонент LSIL, клетки которого могут доминировать и маскировать HSIL. Поэтому ведение подобных больных, основанное лишь на цитологических данных, может иметь неудовлетворительные результаты. Аналогичные результаты были получены А. Е. Smith и соавт., которыми было проведено изучение 100 «атипичных» мазков с целью их отнесения к отрицательным, ASCUS или плоскоклеточному внутриэпителиальному поражению (SIL). Исследование проводилось двумя независимыми группами цитологов. Процент совпадения ответов составил 40-60, что свидетельствует о недостаточной точности результатов на основе применения существующих классификаций. К сожалению, ASCUS интерпретируется в разных лабораториях по-разному. В связи с этим высказывается мысль о необходимости ревизии классификационной системы цервико-вагинальной цитологии, использовании последней в качестве инструмента скрининга, а не как диагностического теста; разработке новых, более эффективных и объективных путей диагностики (4).
Поскольку рак шейки матки имеет ортодоксальное течение, то прогрессия дисплазии в преинвазивный рак (Са in situ) приводит к формированию микроинвазивного, а затем и инвазивного рака. Именно первые две формы и подразумевают понятие раннего рака.
• Под преинвазивным раком понимают патологию покровного эпителия шейки матки, во всей толще которого имеются гистологические признаки рака, утрата слоистости и полярности, но отсутствует инвазия в подлежащую строму. Развитие инвазивного рака

ГЛАВА 7
происходит в 40% случаев, в 35% процесс подвергается обратному развитию (6). Тем не менее, во всех случаях следует рассматривать Са in situ как промежуточное звено в развитии инвазивного рака. Это разные стадии одного неопластического процесса, несмотря даже на то, что преинвазивный рак может исчезать или держаться сравнительно долгий срок. Са in situ чаще всего наблюдается во влагалищной части шейки матки в зоне стыка эпителиальных слоев: многослойного плоского и призматического, реже в начальных отделах цервикального канала. Могут иметь место очаговые, изолированные и мульти-центрические поражения. Чрезвычайно важно подчеркнуть, что с возникновением преинвазивного рака на поверхности слизистой оболочки шейки поражаются и железы. До 40 лет Са in situ чаще локализуется в экзоцервиксе, в 40 — 45 лет с одинаковой долей вероятности в экзо — и эндоцервиксе, а после 45 лет — в цервикальном канале. Подобная закономерность в различном возрасте связана с определенными сдвигами многослойного плоского и призматического эпителия и перемещением в связи с этим зоны их стыка, в области которого чаще всего и развивается Са in situ (Яковлева И.А., Кукутэ Б.Г.).
• Микроинвазивному раку (1-ая стадия) соответствует поражение эпителия шейки матки с глубиной инвазии до 3 мм и длиной поражения до 7 мм. При этом отсутствуют мультицентрические очаги роста и раковые эмболы в кровеносных и лимфатических сосудах стромы. Развитие инвазивного рака на фоне Са in situ выражается в пролиферативном процессе, при котором наблюдается утолщение пласта атипичных

Роннии рок и предрек шейки мотки, обусловленные...
клеток, погружение их в подлежащую ткань в виде массивных, чаще округлой формы тяжей, оттесняющих перед собой базальную мембрану, которая не прослеживается отчетливо на всем протяжении. Выделяют и другой процесс, который называют «спорной инвазией». При этом отмечается нечеткость границ анаплазированного эпителия, разобщенность эпителиальных клеток внутри комплексов, наклонность их к погружению в подлежащую ткань. Однако отшнуровки от основного пласта не наблюдается.
В России за год диагностируется более 11 тыс. новых случаев инвазивного рака. У значительной части этих пациенток опухоль выявляется на поздних стадиях, когда эффективность всех современных методов лечения резко снижается.
Каждый такой случай — это поражение, которое терпит современная медицина. Это признание неспособности вовремя диагностировать и лечить болезнь, а значит признание собственной беспомощности перед ней. Средний возраст больных раком шейки матки составляет 57 лет — эта категория относительно молодых, социально активных женщин, что делает проблему раннего выявления неопластических процессов шейки матки социально значимой. Сохраняющаяся высокая частота этого заболевания в большей степени связана с недостаточной обращаемостью женщин и в меньшей, но достаточно значительной степени — с трудностями лабораторной диагностики и неудачами лечения. Поэтому основным направлением в борьбе с раком шейки матки является более широкое внедрение скрининговых программ в группах риска, направленных на раннее выявление заболевания и профилактические мероприятия. К последним следует отне-

Ронний рок и предрек шейки мотки, обусловленные...
Распространение инфекции происходит преимущественно половым путем, достигая максимума инфицирования до 30 лет. Более 50% сексуально активного населения мира в течение жизни инфицируется вирусом папилломы человека, и это является «первичным» событием в патогенезе РШМ. Большинство случаев инфицирования заканчивается спонтанным выздоровлением. Однако в некоторых случаях развивается персистирующая инфекция, способная запускать механизмы клеточной трансформации эпителиальных клеток. Так при CIN 1 наблюдается активная репликация вируса и его бессимптомное выделение. Превращение CIN 1 в инвазивный рак происходит с большой частотой и, как правило, сопровождается интеграцией вирусной ДНК в геном клетки хозяина (10, 18, 23, 24).
ВПЧ высокой степени онкогенного риска (преимущественно 16 и 18 типов) обнаруживают в 50 — 80% образцов умеренной и тяжелой дисплазии плоского эпителия шейки матки и в 90% инвазивного рака. Важным обстоятельством является то, что клинические проявления ВПЧ-инфекции не обнаруживаются достаточно длительное время; однако последствия персистенции вируса от этого не становятся более легкими. Развитие легкой и умеренной дисплазии в тяжелую происходит соответственно в 10 и 20 % случаев. Такая статистика не является фатальной: переход одной стадии атипии эпителия в другую происходит достаточно долго, что дает большие возможности в плане диагностики и лечения. Кроме того, немаловажным является тот факт, что носительство ВПЧ не является пожизненным: по данным ВОЗ (2001 г.), при отсутствии отягощающих факторов в течение 3 лет плоскоклеточные внутриэпителиальные поражения низкой степени тяжести, содержащие ВПЧ, подвергаются регрессии в 50-62%

ГЛАВА 7
emu лабораторные исследования, направленные на вы-явление вирусов папилломы человека «высокого онко-генного риска», измерение уровня метаболитов эс-традиола и коррекцию этого параметра с помощью Индинола.
Роль В П Ч - и н ф е к ц и и в развитии
неопластических процессов шейки матки
В середине 70-х годов впервые было высказано предположение о возможном участии вирусов папилломы человека в патогенезе рака шейки матки и начаты исследования по идентификации вируса в биопсийном материале. Было установлено, что в злокачественных опухолях шейки матки выявляются наиболее часто два типа вирусов ВПЧ 16 и ВПЧ 18. А в доброкачественных поражениях присутствуют в основном вирусы 6 и 11 типов (13, 27). При дальнейшем изучении роли ВПЧ в неопластических процессах все серотипы были разделены на три группы:
• Низкая степень онкогенного риска: 6,11,42,43,44 • Средняя степень онкогенного риска: 31,33,51,52,58 • Высокая степень онкогенного риска: 16,18,45,56 В настоящее время в ряде исследований показано, что
ДНК ВПЧ обнаруживается в среднем в 98,9% и 94,7% образцов остроконечных кондилом и цервикального рака, соответственно. В популяции у здоровых женщин ВПЧ обнаруживаются в 35% случаев, причем 6, 11, 16 и 18 типы — только в 21,6%. Интересным для дальнейшего изучения оказался и тот факт, что большая часть ДНК ВПЧ 16 в клетках остроконечных кондилом находится в эписо-мальной форме, а в образцах цервикального рака — интегрирована в клеточный геном (26).

Ранний рок и предрек шейки мотки, обусловленные...
са и формы существования его ДНК (эписомальная или интегрированная в геном);
- Опухолевая трансформация возникает с большой вероятностью при взаимодействии ВПЧ с другими канцерогенными или инфекционными агентами (табакокурение, гормональный статус, генетическая предрасположенность и т.д.);
- Иммунологические механизмы защиты могут влиять на течение инфекционного процесса;
- По итогам 4-летних наблюдений ВПЧ-инфекция в 15-20% случаев заканчивается той или иной онкологической патологией генитальной сферы.
Механизмы канцерогенного действия ВПЧ
Для того, чтобы понять механизм трансформирующего действия ВПЧ, необходимо хотя бы кратко остановиться на устройстве вирусного генома.
Генетический аппарат вируса представлен кольцевой двухцепочечной ДНК. Одна из нитей ДНК содержит 9 открытых рамок считывания, которые потенциально кодируют до 10 протеинов, и регуляторный участок генома. Другая нить ДНК некодирующая. Открытые рамки считывания генома разделены на ранний (early, E) и поздний (late, L) участки. Ранний фрагмент включает гены Е1 — Е7, кодирующие синтез белков, ответственных за различные функции в процессе репликации вируса и трансформации клеток. Гены Е1 и Е2 ответственны за репликацию вируса, а также участвуют в регуляции транскрипции вирусного генома. Белок, кодируемый геном Е1, отвечает за поддержание персистенции вирусного генома в эписомальной форме. Белок Е2 подавляет экспрессию ранних генов.

ГЛАВА 7
наблюдений. По сведениям ученых Калифорнийского университета у 70% молодых ВПЧ-инфицированных женщин ДНК ВПЧ перестает определяться на протяжении 24 мес. Скорость элиминации значительно снижается при инфицировании несколькими видами ВПЧ, а также — при наличии в анамнезе кондилом вульвы. В то же время, среди женщин, у которых по меньшей степени 3 раза были получены положительные определения ВПЧ, риск тяжелых онкогенных поражений эпителия возрастал в 14 раз (15,22,28,31).
Многолетние наблюдения за женщинами, инфицированными ВПЧ, позволили прийти к следующим выводам (В.И. Киселев):
- ВПЧ-является заболеванием, передающимся половым путем, и должна считаться фактором риска возникновения РШМ;
- ВПЧ является причиной предопухолевых состояний (дисплазий), что подтверждается иммунологическими и иммуногистохимическими данными, а также результатами с использованием методов молекулярной биологии (полимеразной цепной реакции);
- Трансформированные вирусом клетки эпителия шейки матки с высокой вероятностью приводят к возникновению цервикальных карцином, если не предпринимаются своевременные усилия по лечению предраковых состояний;
- Генитальная ВПЧ-инфекция может длительное время существовать в латентном состоянии, причем это явление не зависит от пола пациента;
- Прогнозы по поводу онкогенной трансформации при инфицировании вирусом, вероятно, зависят от типа виру-

Ронний рок и предрек шейки мотки, обусловленные...
2. При интеграции ДНК-вируса синтез белка Е2 прекращается вследствие нарушения структуры соответствующего гена и активируется синтез белка Е7.
Таким образом, факт интеграции вирусной ДНК может быть зарегистрирован или изучением структуры гена Е2 (эти методы разрабатываются нами, однако в настоящее время они слишком трудоемки для клинических лабораторий) или измерением уровня синтеза онкобелка Е7.
По нашему мнению, наличие онкобелка Е7 в цервикаль-ных пробах может рассматриваться как однозначное свидетельство начавшегося процесса малигнизации эпителиальных клеток, содержащих интегрированную копию генома ВПЧ. Неоспоримым достоинством Е7 как онкомарке-ра является и то, что в норме этот белок в тканях не синтезируется. Его происхождение полностью связано с жизненным циклом интегративной формы ВПЧ-инфекции. Так же преимуществом метода является то, что берется материал непосредственно из органа-мишени (5).
В 2002 г. началась исследовательская работа на базе НИИ молекулярной медицины ММА им. И.М. Сеченова, Всероссийского научного центра молекулярной диагностики и лечения (ВНЦМДЛ), Российского научного центра рентгенорадиологии и Центра акушерства гинекологии и перинатологии РАМН, целью которой является оценка эффективности Е7 в диагностике неопластических процессов шейки матки.
Онкобелок Е7 в цервикальном материале определяется методом иммуноферментного анализа. С этой целью созданы и опробованы специальные тест-системы. Полученные нами результаты очень обнадеживающие: Е7 определялся в 8 1 % дисплазий и 75% всех цервикальных раков. Остальные 25% злокачественных поражений шей-

ГЛАВА 7
Протеин Е4 участвует в экспрессии вирусных частиц, а Е5 — Е7 обладают трансформирующим потенциалом. Поздний фрагмент генома состоит из генов L1 — L2, кодирующих структурные белки вириона (9, 17).
В процессе интеграции в клеточный геном разрыв кольцевой молекулы вирусной ДНК может происходить в любом месте, однако наиболее часто происходит в области Е1 \Е2. Исчезновение супрессорной функции белка Е2 обуславливает суперэксперессию вирусных онкогенов Еб — Е7, трансформирующее действие которых способствует прогрессии неоплазий.
Как уже говорилось ранее, в последние годы было показано, что в инфекционном процессе ВПЧ различают две стадии (11, 12, 16, 30):
1. Стадия репродуктивной инфекции, когда вирусная ДНК находится в свободном (эписомальном) состоянии,
2. Стадия интегративной инфекции, когда ДНК вируса встраивается в геном инфицированных клеток.
Первая стадия является обратимой и у многих инфицированных наступает ремиссия, о чем мы уже упоминали выше. Стадия интегративной инфекции является первым шагом к опухолевому перерождению клетки и очень часто заканчивается развитием карциномы. В исследованиях Кузнецовой Ю.Н. с соавт. 2003 г. установлено, что эти стадии вирусной инфекции соответствуют двум различным клиническим формам заболевания — транзитор-ная и латентная, соответственно.
Интеграция ДНК ВПЧ в геном клеток сопровождается двумя молекулярными событиями:
1. Встраивание вирусной ДНК в хромосому всегда сопровождается нарушением структуры гена Е2, который является репрессором Е7.

ГЛАВА 7
ки матки носили ВПЧ-независимый характер, и Е7 в них не определялся.
Измерение уровня онкобелка Е7 в цервикальных пробах позволяет ответить на следующие вопросы:
1. Определить стадию развития вирусной инфекции. Повышенный синтез онкобелка Е7 указывает на интегра-тивную фазу инфекционного процесса, при которой очень мала вероятность спонтанной ремиссии;
2. Уровень Е7 достоверно коррелирует с агрессивностью зарождающегося опухолевого процесса и может рассматриваться как неблагоприятный прогностический признак;
3. Онкобелок Е7 является мощным иммуносупрес-сивным агентом, существенно снижая эффективность таких иммунокоррегирующих методов лечения, как интер-феронотерапия, использование индукторов интерферона (циклоферон) и иммуномодуляторов. Поэтому при выявлении с помощью лабораторных методов высокого уровня Е7, необходимо включить в комплексную терапию «Индинол» как препарат, избирательно ингибирую-щий синтез Е7 в эпителиальных клетках, инфицированных ВПЧ (3).
Иммуносупрессорные функции онкобелка Е7
Из клинической практики известно, что один из способов терапии ВПЧ-ассоциированных заболеваний — это вагинальные суппозитории, содержащие интерферон 2. Известно также, что примерно в 50% случаев даже длительное назначение интерферона не приводит к клиническим улучшениям.

Ронний рок и предрек шейки мотки, обусловленные...
Феномен резистентности ВПЧ-инфицированных клеток к действию интерферона исследовался недавно более детально. Оказалось, что это явление напрямую зависело от уровня синтеза онкобелка Е7. Авторы установили, что белок Е7 нейтрализует противовирусную и противоопухолевую активность IFN-a2 за счет того, что он способен избирательно блокировать большинство генов, индуцируемых интерфероном, сводя на нет все усилия интер-феронотерапии (8, 20, 21).
Другая группа авторов обнаружила способность онкобелка Е7 внутриклеточно инактивировать IRF (interferon regulatory factor — фактор регуляции активности интерферона). IRF является внутриклеточным фактором транскрипции, который активируется в клетках при действии на них IFN-a и IFN-y. IRF «включает» транскрипцию генов, которые кодируют синтез противовирусных белков (20).
Представленные данные свидетельствуют о том, что онкобелок Е7 не только выполняет функции основного фактора малигнизации, но и определяет устойчивость инфицированных клеток к интерферонам. Из этого следует важный вывод для клинической практики:
1. Местная интерферонотерапия остается важным методом лечения ВПЧ-ассоциированных заболеваний;
2. Измерение содержания белка Е7 ВПЧ является необходимым условием для назначения терапии и прогнозирования ее эффективности;
3. При высоком содержании Е7 необходимы мероприятия по его снижению, что сделает опухолевые клетки более восприимчивыми к интерферону.
Постоянный синтез онкобелков необходим для поддержания опухолевого фенотипа клеток. Очевидно, что появление в организме клеток, несущих чужеродные ан-

Роннии рок и предрек шейки мотки, обусловленные.
вать с ним), приводящий к нежелательным последствиям.
Второй метаболит (2-ОН), обладая умеренными функция
ми, нормализует клеточный рост (7, 25, 29).
Давно было отмечено, что тканевые изменения в цер-
викальном канале, вызванные ВПЧ, локализованы глав
ным образом в эстроген-чувствительных зонах. Более
того, было установлено, что там, где наблюдается актив
ная экспрессия белков ВПЧ, отмечен высокий уровень
синтеза (1ба-ОН), сравнимый с таковым в раковых клет
ках молочной железы. Следует подчеркнуть, что в норме
эпителиальные клетки шейки матки не способны обеспе
чивать превращение эстрадиола в 16-гидроксистерон.
Таким образом, активная репродукция ВПЧ индуцирует
образование агрессивного метаболита в инфицирован
ных клетках.
Инфицированные кератиноциты человека проявляют
пролиферативную активность in vitro, но при этом не име
ют опухолевого фенотипа при микроскопическом иссле
довании. Добавление в культуральную среду экзогенного
1ба-0Н превращает клетки в типично раковые. На основа
нии этого становится понятным, что инфицирование эпи
телиальных клеток ВПЧ — необходимое, но недостаточ
ное событие для ракового перерождения. Для формиро
вания необратимой неоплазии необходима:
1. активная экспрессия генов Е6 и Е7 вируса,
2. индукция метаболических механизмов конверсии
эстрадиола в 16а-ОН,
3. индукция множественных повреждений хромосом
ной ДНК в инфицированной клетке, которая завершает
процесс перерождения.

ГЛАВА 7
тигены (Е6 или Е7), должно индуцировать соответствующие иммунные реакции. Однако больные с цервикальны-ми карциномами имеют очень низкий уровень Т-клеточно-го иммунитета в отношении Еб и Е7 (14).
При вакцинации этими белками также не наблюдается выраженной реакции, несмотря на хорошие характеристики иммунного статуса. Эти наблюдения, а также эксперименты на животных показывают, что онкобелок Е7, синтезирующийся в эпителиальных клетках, способен «ускользать» или даже су премировать иммунные реакции в отношении себя.
Роль гормональных факторов в развитии рака шейки матки
Как уже упоминалось, эстрогены играют исключительно важную роль в развитии неопластических процессов в так называемых эстроген-чувствительных тканях. К числу последних относится и эпителий шейки матки. Эстрадиол — один из наиболее активных женских половых гормонов — обладает высоким сродством к эстрогеновым рецепторам и, взаимодействуя с ними, оказывает существенное влияние на метаболическую и пролиферативную активность клеток эпителия шейки матки.
Ферментативная система цитохромов Р-450 обеспечивает конверсию эстрадиола в два основных метаболита: 16сх-гидроксистерон (16сх-0Н) и 2-гидроксистерон (2-ОН) (рис.18).
Первый из них (16а-0Н) относится к категории «агрессивных гормонов», вызывающих длительный эффект (есть данные, что 16а-0Н способен образовывать ковалентные связи с рецептором, то есть необратимо взаимодейство-

ГЛАВА 7
Ключевая роль 16а-ОН-стерона в раковом перерождении клеток, инфицированных ВПЧ, подтверждается прямыми исследованиями на клетках эпителия шейки и цер-викальной карциномы.
Из наших данных следует, что один из основных путей малигнизации клеток, инфицированных ВПЧ, заключается в том, что вирус модифицирует клеточный метаболизм таким образом, что клетка приобретает способность превращать эстрадиол преимущественно в 16а-ОН, который является прямым активатором экспрессии гена Е7, ответственного за опухолевую трансформацию клеток. Таким образом, формируется порочный круг, при котором вирус через образование агрессивной формы эстрадиола создает благоприятные условия для развития опухоли, стимулируя синтез онкобелка Е7. В свою очередь онкобе-лок, с одной стороны, активирует механизмы патологической пролиферации клеток, а с другой — блокирует механизмы развития иммунологической защиты (Рис. 19).
Из этой модели очевидно, что один из способов профилактики и лечения опухолей, обусловленных ВПЧ, может быть нацелен на разрыв этого патогенетического механизма. Это предположение легло в основу наших экспериментальных исследований, посвященных способности Индинола регулировать метаболизм эстрадиола в эпителиальных клетках, инфицированных ВПЧ. Как следует из данных, представленных в Таблице 21, Индинол подавляет образование 1ба-гидроксистерона (16а-0Н) в клетках CaSki, содержащих копии гена Е7 ВПЧ, и трансформированных клетках цервикального эпителия. В этих условиях должен снижаться уровень экспрессии гена Е7, что и было подтверждено нашими исследованиями (1,2).

Ранний рок и предрек шейки мотки, обусловленные...
Таблица 21 Изучение особенностей метаболизма эстрадиола в эпителиальных клетках, инфицированных ВПЧ, под
воздействием Индинола
Тип клеток
CaSki ВПЧ +
С 33-А ВПЧ -
Цервикальный эпителий
Цервикальный эпителий из
I трансформированных зон
% 16а гидроксиэстрона на мкг клеточного белка
Эстрадиол
16 ±0,1
0,08 ± 0,03
0,7 ± 0
12,6 ±0,5
Эстрадиол + ИНДИНОЛ
2,2 ±0,5
0,08 ± 0,08
0,5 ±0,1
2,1 ±0,3

ГЛАВА 7
С п и с о к литературы к Главе 7
1. Киселев В.И., Ашрафян Л.А., Бударина СО. и др. Этиологиче
ская роль вируса папилломы человека в развитии рака шейки матки:
генетические и патогенетические механизмы, возможности терапии и
профилактики, Гинекология, 2004, том 6, № 4 стр. 174-179.
2. Киселев В.И., Киселев О.И., Северин Е.С. Исследование спе
цифической активности индол-3 карбинола в отношении клеток, инфи
цированных вирусом папилломы человека, Вопросы биологической,
медицинской и фармацевтической химии, 2003, 4, стр. 28-32.
3. Киселев В.И., Киселев О.И. Этиологическая роль вируса па
пилломы человека в развитии рака шейки матки: генетические и пато
генетические механизмы, Цитокины и воспаление, 2003, том 2, № 4,
стр. 31-38.
4. Киселев В.И., Ляшенко А.А. Молекулярные механизмы регуля
ции гиперпластических процессов, монография, 2005, 346 стр.
5. Свешников П.Г., Шемчукова О.Б. и др. Ранняя диагостика ра
ка шейки матки, ассоциированного с вирусами папилломы человека.
Диагностическая система и моноклональные антитела для определе
ния белка Е7, Молекулярная медицина, 2004 № 2, стр. 32-38.
6. Хмельницкий O.K., Цитологическая и гистологическая диагно
стика заболеваний шейки матки, 2000.
7. Auborn К., Abramson A. et al Estrogen metabolism and laryngeal
papillomatosis a pilot study on dietary prevention, Anticancer research, 1998,
18,4574-4589.
8. Barnard P., Payne E. & McMillan N.A. The human papillomavirus
E7 protein is able to inhibit the antiviral and anti-growth functions of interf
eron-. Virology, 2000, 277, 411-419.
9. Bedell M.A., Jones K.H., Laimins LA. The E6-E7 region of hu man
papillomavirus type 18 is sufficient for transformation of NIH 3T3 and rat-1
cells. J Virol 1987; 61:3635-40.
10. Campo M.S., Moar M.H., Jarrett WFH, Laird H.M. A new papilloma
virus associated with alimentary cancer in cattle. Nature 1980; 286:180-2.

Ранний рои и предрек шейки мотки, обусловленные...
11. Durst M., Сгосе СМ., Gissmann L, Schwarz E., Huebner K. Papillomavirus sequences integrate near cellular onco genes in some cervical carcinomas. Proc Nati Acad Sci USA 1987; 84:1070-1076.
12. Dyson N.. Howley P.M., Munger K., Harlow E. The human papilloma virus-16 E7 oncoprotein is able to bind to the retinoblastoma gene product. Sciece 1986; 243:934-7.
13. Gross G., Joblonska S., Pfister H. Genital pappiloma- virus infections, 1989.
14. Gtannini S i . et al. Cytokine expression in squamous intraepilheial lesions of the uterine cervix: impications for the generation of local immunosuppression. Clin.Exp.lmmunoL, 1998,113,183-189.
15. Howley P.M., Schlegel R. The human papillomaviruses. As overview. Am J Med 1988; 85:155-158.
16. Lehn H., Villa L L , Marziona F., Hilgarth M., Hillemans H.G., Sauer G. Physical state an<J biological activity of human papillomavirus genomes in precanrous lesions of the female genital tract. J Gen Virol 1988; 69:187-96.
17. Matlashewski G., Schneider J., Banes L, Jones N., Murray A., Crawford L. Human papillomavirus type 16 DNA cooperation with activated ras in transforming primary cells. EMBO J 1987; 6:1741-6.
18. McCance D.J. Human papillomavirus (HPV) infection in the aetiology of cervical cancer. Cancer Sury 1988; 7:499-506.
19. Muir C, Waterhouse J., Mack Т., Powell J., Whelan S., ets. Cancer incidence in five continents. Lyon, France:lnternational Agency for on Cancer, 1987, Vol. 5. Publ. n. 88.
20. Nees M. et al. Papillomavirus type16 oncogenes downregulate expression of interieron-responsive genes and upregulate proiferation-associ-ated and NF-kB-responsive genes in cervical keratinocytes. J. Vrd., 2001, 75, 4283-4296
21. Park J.S. et at Inactivation of interferon regulatory factor-1 tumor suppressor protein by HPV E7 oncoprotein. Implication for the E7-medated immune evasion macharism in cervical carcinogenesis. J. Biol. Chem, 2000, 275, 6764-6769.
22. Reeves W.G., Rawls W.E., Brinton L.A. Epidemiology of genital papillomavirus and cervical cancer. Rev Infect Dis 1989; 11:426-39.

ГЛАВА 7
23. Schneider A., Olterdorf Т., Schneider V., Gissmann L. Distribution pattern of human virus 16 genome in cervical neoplasia by molecular in situ hybridization of tissue sections. Int J Cancer 1987; 39:717-21.
24. Schwarz E., Freese U.K., Gissmann L, Mayer W., Roggenbuck В., Stremlau A., zur Hausen H. Structure and transcription of human papillomavirus sequences in cervical carcinoma cell. Nature 1985; 314:111-4.
25. Swaneck G.E. and Fishman J. Covalent binding of the endogenous estrogen 16a-hydroxyestrone to estradiol receptor in human breast cancer cells: Characterization and intranuclear localization (cell proliferation/estra-diol metabolism/MCF-7 eels/estrogen receptor), Proc. Natl. Acad. Sci. USA, 1988, Vol. 85, pp. 7831-7835.
26. Syrjanen K., Mantyjarvi R., Saarikoski S. et al Factors associated with progression of cervical human papillomavirus (HPV) infections into carcinoma in suti during a long-term prospective follow-up. Br.J.Obstet-Gynoe-col, 1988,95:1096-1102.
27. Syrjanen K. Human papillomavirus (HPV) infections of the female genital tracts and their associations with intraepithelial neoplasia and squ-amoces cell carcinoma. Pathol.Ann. 1986, 21:53-89.
28. Syrjanen K.L. Epidemiology of human papillomavirus (HPV) infections and their association with genital squamous cell cancer. APMIS 1989, 97:957-70.
29. Taylor A., Gaston K., Webster K. Oestrogen and progesterone increase the levels of apoptosis induced by the human papillomavirus type 16 E2 and E7 proteins, J. Gen. Virol, 2001, Vol. 82, 201-213.
30. Werness B.A., Levine A.J., Howley P.M. Association of human papillomavirus types 16 and E6 proteins with p53. Science 1990; 248:76-9.
31. zur Hausen H. Intracellular surveillance of persisting viral infections: human genital cancer results from deficient cellular control of papillomavirus gene expression. Lancet 1986; 2:489-91.

ОБОСНОВАННЫЕ НАПРАВЛЕНИЯ ТЕРАПИИ И ПРОФИЛАКТИКИ РАКА РЕПРОДУКТИВНЫХ ОРГАНОВ
Данные, представленные выше, свидетельствуют о том, что опухолевая клетка обладает многими системами для восприятия пролиферативных сигналов. В том случае, если одна из них блокируется приемом лекарственных препаратов, например, назначением антиэстрогенов при терапии эстроген-зависимых опухолей, то немедленно включается другая. При таких способностях опухолей к выживанию даже трудно вычленить, какой именно стимул является ключевым: блокада одного из них приводит к моментальному включению других сигнальных каскадов, с возможностью использовать компоненты одного пути в пользу другого. Уникальная адаптационная способность опухолевых клеток диктует необходимость разработки новых подходов к этой проблеме.
На многочисленных онкологических международных форумах в последние годы намечены основные тенденции в борьбе с этим видом патологий. Коротко их можно обозначить в виде следующих тезисов:
1. В последние годы стала очевидной необходимость комплексного (или, как принято называть в иностранной литературе, мультифокального) подхода к терапии опухо-

ГЛАВА 8
лей. Блокаду пролиферативной активности клеток-мишеней целесообразно проводить на уровне мембран, цитоплазмы, ядра. При этом в качестве внеклеточных способов коррекции рассматриваются, в числе прочих, блокада лиганд-рецепторных взаимодействий — ростовых факторов, гормональных, а также управление экспрессией «нужных» форм цитохрома Р-450 через взаимодействие с Ah-рецептором. Цитоплазматическими мишенями могут быть киназы, как рецепторные, так и свободные, а также множество других участников каскадной передачи от мембраны к ядру. Внутриядерная коррекция подразумевает блокаду активаторов транскрипции, модуляцию активности ядерных рецепторов, защиту от изменений ну-клеотидов (например, метилирование) ДНК. В настоящее время на рынке имеется множество блокаторов пролиферации опухолевых клеток с самой различной направленностью. Клиницистам предстоит большая работа по выработке оптимальных доз и комбинаций лекарственных препаратов в лечении опухолевых заболеваний, которая, будем надеяться, закончится успехом.
2. Усилить акцент на разработке новых прогностических маркеров, позволяющих регистрировать начинающиеся в организме процессы малигнизации. Специалистам хорошо известно, что чем раньше поставлен диагноз, тем выше эффективность проводимых лечебных мероприятий. Необходимо широкое внедрение скрининго-вых программ, позволяющих выявлять начальные стадии заболевания и существенно снижать смертность.
3. Сконцентрировать усилия на профилактике онкологических заболеваний. Эпидемиологические исследования в области онкологии убедительно свидетельствуют о том, что рак во многом является результатом образа жизни.

Обоснованные направления терапии и профилактики...
В последнее время потенциальным источником противоопухолевой и антиопухолевой защиты рассматривается диета. Эпидемиологические исследования показывают, что в странах, где традиционно потребляется большое количество овощей, риск возникновения рака гораздо меньше. Многочисленные исследования последних лет посвящены фитонутриентам — веществам растительного происхождения: терапевтические концентрации некоторых из них оказывают поразительный эффект, связанный с избирательным ингибированием роста опухолевых клеток, индукцией их апоптоза.
В литературе широко представлены и подробно изучены молекулярные механизмы блокады пролиферативной активности фитонутриентов, которые проявляют свое действие на уровне мембран, цитоплазмы и ядра клеток-мишеней, модулируя сигнальные каскады как на белковом, так и транскрипционном уровне. Механизмы избирательной активности смеси фитонутриентов вмешиваются в гормонозависимую (эстроген- и андроген-зависимую) пролиферацию, блокаду сигнальных путей, индуцируемых ростовыми факторами, осуществляют нормализацию защиты естественных анти-оксидантных систем клетки, метаболическую коррекцию цитохромной системы (цито-хромов Р-450) оксигенации эстрогенов и канцерогенов, вмешиваются в клеточный цикл (78, 84).
Роль диеты в индукции противоопухолевой защиты вызывает небывалый интерес и в настоящее время интенсивно изучается. Этому вопросу посвящены многочисленные эпидемиологические исследования, на основе которых Всемирная организация здравоохранения констатирует ряд показательных выводов: диета является определяю-

ГЛАВА 8
щим фактором в развитии и профилактике опухолей. Масштабные эпидемиологические исследования показали, что традиционные привычки в питании вносят значительный вклад в показатели смертности от опухолей в различных странах. Приведенная на Рис. 20 диаграмма отражает общую тенденцию по всем опухолевым заболеваниям, с тем лишь отличием, что доля смертей на определенное количество населения (обычно на 100 тысяч) варьирует в разных странах. Однако общая тенденция сохраняется, и в странах, где традиционным считается употребление рыбы, сырых овощей и фруктов (Юго-Восточная Азия, страны Средиземноморья), уровни заболеваемости гораздо ниже по сравнению с таковыми в Европе и Америке. Современные технологии приготовления пищи, стремление к увеличению ее сроков годности, добавление консервантов и искусственных красителей, различных стабилизаторов и усилителей вкуса вносят отрицательное влияние на здоровье человека. Изучение заболеваемости эмигрантов также подтверждает эти выводы: смена традиционной диеты влечет развитие тех же заболеваний, которыми страдает местное население. Эти показатели отодвигают на второй план значение генетической предрасположенности к развитию опухолей и ставят диету в один ряд с другими влиятельными факторами — окружающая среда, курение, иные привычки, относящиеся к стилю жизни. Можно сказать, что рак, среди прочего, — это заболевание стиля жизни. По крайней мере, статистика эти выводы не опровергает.
Все эти показатели стимулируют изучение биологических особенностей основных компонентов пищи, составляющих основу традиций питания той или иной страны.
Обоснованные направления терапии и профилактики...
Катехины зеленого чая. Эпигаллокатехин-3-галлат
Второй после воды, чай является одним из наиболее популярных напитков, и его частое употребление напрямую связывают с поддержанием нормальной работы отдельных органов и систем — сердечно-сосудистой, нервной, мочевыделительной, а также улучшением общего эмоционального состояния организма. Экстракт зеленых листьев различных сортов кустарника Camellia sinensis, именуемое чайным деревом, позволяет получить множество видов чая, отличающихся способом приготовления, содержанием катехинов, кофеинов, других полифеноль-ных компонентов.
Имеются поразительные данные эпидемиологических исследований о противоопухолевых эффектах отдельных чайных компонентов. Так, например, по данным итальянских исследователей, ежедневное употребление зеленого чая способствует защите от рака ротовой полости, гортани, глотки. По другим данным, полученным из Шанхая, частое употребление чая ассоциируется со снижением заболеваемости раком пищевода, даже среди курящих и употребляющих алкоголь (102). Защитный эффект зеленого чая подтвержден в Турции, Швеции, Японии: ежедневное употребление этого напитка местными жителями способствует защите от рака желудка. Употребление японскими женщинами более 10 чашек чая в день (около 2 литров) снижает риск возникновения и метастазирования рака молочной железы. При этом аналогичных данных не получено при употреблении черного чая. Таким образом, зеленый чай обладает сильнейшим превентивным эффектом на многие солидные опухоли, и изучение активностей

ГЛАВА 8
чайных компонентов постепенно становится в один ряд с масштабными исследованиями, посвященными изучению механизмов действия традиционных противоопухолевых средств.
Наибольший интерес в последнее время вызывают полифенолы зеленого чая, в основном, катехины, обладающие целым спектром защитных эффектов на органы и ткани. Водный экстракт зеленого чая содержит эпигалло-катехин-3-галлат (epigallocatechin-3-gallate, EGCG), эпи-галлокатехин (epigallocatechin, EGC), эпикатехин-3-гал-лат (epicatechin-gallate, ECG) и эпикатехин (epicatechin, ЕС) (Рис.21). Все они обладают сильнейшей антиокси-дантной активностью, по своей силе сравнимой с таковой у токоферола-альфа (витамина Е). Важнейшим дополнением является мощный антипролиферативный потенциал: клетками-мишенями являются все клетки, где наблюдается стимуляция экспрессии тех компонентов, от которых зависит рост и количество данного типа клеток. Можно сказать, что чайные катехины обладают избирательной активностью по отношению к пролиферирую-щим, активно делящимся клеткам. Поэтому уровень избирательной активности в отношении гиперпластических процессов, вне зависимости от стимулов, у чайных кате-хинов чрезвычайно широк.
Сложным процессам ингибирования опухолевого роста катехинами есть многочисленные доказательства, как in vitro, так и in vivo. Общим механизмом антиопухолевой активности катехинов считался антиоксидантный эффект. Однако, благодаря интенсивным изучениям этих веществ, стало известно, что катехины обладают комплексным воздействием на многочисленные пролиферативные пути в опухолевых клетках, среди которых наибольший вклад

Обоснованные направлении терапии и профилактики...
в стимуляцию роста, развития и метастазирования опухолей вносят рецептор-опосредованная стимуляция ростовыми факторами, как следствие этого — активация цито-плазматических и рецепторных тирозин-киназ, активация транскрипционных факторов, развитие провоспалитель-ной реакции, стимуляция неоангиогенеза. Модуляция вышеуказанных процессов катехинами характеризует их с позиций мощных противовоспалительных, анти-пролифе-ративных, анти-ангиогенных природных компонентов.
Одним из интереснейших представителей чайных кате-хинов является эпигаллокатехин-3-галлат (epigalloca-techin-3-gallate, EGCG). Максимально возможное употребление чая в течение дня приводит к увеличению концентрации EGCG до 326 нг/мл в плазме. Время полувыведения EGCG составляет 5-5,5 часов, что намного выше, чем у других катехинов. Абсолютно нетоксичен; механизмам его противоопухолевой активности посвящено множество исследований.
Модуляция внутриклеточных сигнальных каскадов
В основе антипролиферативной активности EGCG лежит, в частности, его способность блокировать развитие воспалительных реакций. Цитокины IL-8, TNF-a, активируемый ими NF-kB являются основными факторами, вовлеченными в развитие воспаления. Оно имеет место при всех заболеваниях с пролиферативным и (или) аутоиммунным компонентом, среди которых наиболее трудноизлечимыми являются атеросклероз, псориаз, полиартриты, эндометриоз, мастопатии, а также при доброкачественных и злокачественных новообразованиях. Показа-

ГЛАВА 8
но, что EGCG ингибирует TNF-опосредованную активацию NF-kB (49, 96). Кроме того, NF-kB ингибируется также через блокаду другого пути, индуцируемого IL-1 (99). Инактивация обоих сигнальных путей достигается благодаря уникальной возможности EGCG вмешиваться на любом уровне провоспалительных сигнальных каскадов. Показано, что в клетках-мишенях IL-1 и TNF-aEGCG является ингибитором IRAK-киназы, вовлеченной в IL-1-индуци-рованный сигнальный каскад, блокатором фосфорилиро-вания субъединицы рб5 NF-kB. В отсутствие NF-kB нарушается экспрессия IL-8 — основного хемоаттрактанта и стимулятора миграции моноядерных лейкоцитов (лимфоцитов) и нейтрофилов в зону воспаления. Кроме того, IL-8 также стимулирует экспрессию фактора роста эндотелия сосудов (VEGF).
Ингибирование металлопротеиназ
Матриксные металлопротеиназы (ММР) очень активно экспрессируются в опухолевых клетках, а также в клетках эндотелия (50). Они вовлечены в деградацию клеточного матрикса и миграцию эндотелиальных клеток. Основная их функция в опухолевых клетках — разрушение базаль-ной мембраны и облегчение клеточной инвазии. ММР активируются параллельно на про-воспалительном фоне, совместно с IL-1 и TNF-a. В экспериментах установлено, что инвазивный потенциал опухолевых клеток ингибируется блокаторами ММР — как синтетическими, так и тканевыми (59). Эти препараты сейчас активно используются в клинических испытаниях в качестве противоопухолевых средств. Однако они обладают нежелательными побочными эффектами: их употребление сопровождается, в част-

Обоснованные направления терапии и профилактики.
ности, мышечной и костной болью. Недавно было показано, что EGCG эффективно ингибирует активность некоторых представителей металлопротеиназ, — ММР-2 и ММР-9 (Gabrisa et al., 2001). Интересно, что подобная активность достигается при такой концентрации EGCG, которая наблюдается у людей, употребляющих даже небольшое количество чашек зеленого чая ежедневно. При этом, согласно экспериментам in vivo, степень опухолевой инвазии и размер опухоли уменьшались примерно на 50% (34).
Поскольку EGCG образует комплексы с металлами, ин-гибирование ММР может быть объяснено его способностью хелатировать цинк, являющийся необходимым для проявления ферментативной активности металлопротеиназ (64). Кроме того, в других исследованиях показано, что добавление цинка в присутствии EGCG абсолютно подавляло активность ММР, фактически усиливая это инги-бирование (34) (Рис. 22).
Инактивация ММР может быть использована и при лечении заболеваний неопухолевого генеза, но которые также сопровождаются не менее сильной пролиферативной реакцией. Классическим примером является артрит. Воспаление, индуцируемое IL-ip, сопровождается усиленной экспрессией и активностью ММР: всё направлено на разрушение и потерю хрящевой ткани, что ассоциируется с хроническим воспалением при данном заболевании. Показано, что EGCG не только абсолютно нетоксичен для хрящевой ткани, он ингибирует экспрессию воспалительных медиаторов в суставах при артрите. EGCG блокирует IL-ip-индуцируемую активность матриксных протеиназ ММР-1 и ММР-13 в хрящевой ткани, активацию NF-kB и АР-1, обеспечивая тем самым стабильную молекулярную основу для анти-воспалительных процессов и заживления (2).

Рис. 18. Метаболизм эстрадиола

ГЛАВА 8
Ингибирование урокиназы
Урокиназа — это гидролаза, также вовлеченная в инвазию и метастатический потенциал опухолевых клеток (46). Как и металлопротеиназы, урокиназа усиливает деградацию внеклеточного матрикса. В экспериментах регистрировалась повышенная экспрессия урокиназы в опухолевых клетках молочной железы, яичников, простаты, где отмечена ее ключевая роль в развитии метастазирую-щего рака (36). Ингибирование активности плазминоген-активатора урокиназного типа (иРА) значительно редуцировало объем и размер опухолевой массы, а в некоторых случаях наступала полная ремиссия опухолей in vivo (47). EGCG является непосредственным блокатором урокиназы, изменяя ее ферментную активность и, как следствие, способность к деградации внеклеточного матрикса. Компьютерное моделирование комплекса EGCG с урокина-зой позволило идентифицировать аминокислотные остатки, которые ответственны за ее ферментативную активность: комплексообразование EGCG* с каталитической триадой — гистидином 57, серином 195 и аргинином 35 в молекуле урокиназы полностью нивелирует ее активность: фермент перестает распознавать субстрат (48). Однако uPA-ингибирующая активность EGCG наблюдается только в концентрациях, намного превышающих таковую в крови при обычном употреблении чая (14).
Интересные данные получены на клетках фибросарко-мы человека НТ 1080. EGCG индуцировал не только супрессию промоторной активности иРА, но и дестабилизацию мРНК иРА. Отмечено также ингибирование фосфо-рилирования митоген-активируемых протеинкиназ (МАРК) — Erk1/2 и р38 МАРК (56). Благодаря данным, полученным авторами, можно сделать вывод, что EGCG

Рис. 19. Роль эстрогенов в канцерогенезе эпителиальных клеток шейки матки, инфицированных ВПЧ
Рис. 20. Эпидемиологические показатели заболеваемости раком молочной железы в мире (по данным ВОЗ за 2002 год). Цифры отражают статистику смертей в разных странах на 100 тыс. населения

ОН
Рис. 21. Химические формулы чайных катехинов
Рис. 22. Спектр механизмов противоопухолевого действия EGCG

Рис. 23. Сравнительный эффект чайных катехинов (EGCG и ЕС) на апоптоз опухолевых клеток НТ29 (карцинома прямой кишки) на мышиных моделях. Исследовались объем опухоли (А) и опухолевая масса (Б)
Рис. 24. Ингибирование ангиогенеза и миграции эндотелиальных клеток в зависимости от различных концентраций EGCG. Диаграмма отражает результаты трех экспериментов in vivo
(количество живых клеток в зависимости от дозы EGCG)

Обоснованные направления терапии и профилактики...
регулирует экспрессию uPA, как минимум, за счет двух механизмов: путем подавления промоторнои активности и нарушения структурной стабильности мРНК uPA.
EGCG и циклооксигеназа-2
В опухолевых клетках наблюдается усиленная экспрессия циклооксигеназы-2 (СОХ-2) — фермента, играющего ключевую роль в синтезе простагландинов (PGE2). Простагландины, в свою очередь, стимулируют экспрессию VEGF — ростового фактора, вовлеченного в специфические митогенные и хемотаксические эффекты в клетках эндотелия сосудов. Снижение рецидива опухолей наблюдается при приеме нестероидных противовоспалительных препаратов — ингибиторов СОХ-2. Вначале этот эффект был отмечен при раке прямой кишки, впоследствии — на всех солидных опухолях, где регистрируется повышенная активность СОХ-2. После приема EGCG наблюдалось снижение уровня PGE2, что характеризует EGCG как специфический СОХ-2 — ингибитор (6). Эти результаты дополняют исследования Hong и соавторов, которыми показано, что чайные полифенолы вмешиваются в СОХ-2-зависимый метаболизм арахидоновой кислоты, что ассоциировалось со снижением инцидентов развития рака прямой кишки (43). В то же время доказано отсутствие какого-либо угнетающего эффекта на промотор-ную область гена СОХ-2 в ряде опухолевых линий карциномы прямой кишки человека (69). Интересно, что СОХ-2-стимулирующая активность под влиянием EGCG обнаружена в макрофагах (73). Очевидно, что проявление разных эффектов EGCG на экспрессию СОХ-2 зависит от дозы катехина, типа клеток-мишеней, способов введения, каких-то других дополнительных факторов.

Рис. 25. Механизм EGCG-индуцированного апоптоза

Обоснованные направлении терапии и профилактики...
том числе при довольно низких концентрациях (61). При этом отмечено, что при концентрации, которая устанавливается в крови человека после выпитой чашки чая (0,1 цд/ml EGCG), наблюдается эффект довольно сильного синергизма с традиционными противоопухолевыми препаратами.
Блокада неоангиогенеза
Опухолевый неоангиогенез характеризуется усиленной экспрессией VEGF и его рецепторов, что способствует улучшению трофики новообразований. Появление новых сосудов в опухоли — классический пример патологического неоангиогенеза. Существенный вклад в противоопухолевую стратегию вносит блокада ангио-генеза. Блокирование VEGF или его рецепторов (VEGF-В, VEGF-C и VEGF-D) почти полностью прекращает про-лиферативную активность в эндотелии сосудов. Это означает, что антиангиогенные факторы вносят существенную коррекцию в пролиферативную реакцию. Показано, что EGCG значительно снижает уровень VEGF и плотность его рецепторов in vitro (91). Механизм инги-бирования VEGF и его рецепторов может быть объяснен следующими фактами. Блокада указанных факторов происходит, в частности, на уровне активаторов транскрипции. В промоторной области VEGF имеются несколько сайтов связывания с фактором транскрипции АР-1, который является мишенью для EGCG. Препятствие для взаимодействия с ДНК является определяющим в EGCG-индуцированной блокаде транскрипции VEGF и его рецепторов.

ГЛАВА 8
Регуляция клеточного цикла
Клеточный цикл млекопитающих, способы управления его регуляцией в последнее время довольно интенсивно изучаются. Особенно, если речь идет о методах и подходах к лечению опухолевых заболеваний: анормальное клеточное деление — основная характеристика активно делящихся опухолевых клеток и восстановление (угнетение) скорости их деления — одна из важных составляющих противоопухолевой терапии. В процесс клеточного деления вовлечено множество комплексов, факторов транскрипции, адапторных белков, онкогенов.
Ключевую роль в процессе деления клеток выполняют циклины — «положительные» регуляторы клеточного цикла, а также циклин-зависимые киназы (CDK) — целое семейство киназ с антиапоптотическими свойствами. Проли-феративные процессы сопровождаются комплексообра-зованием циклинов с CDK в зависимости от стадий клеточного цикла. Так, например, ранняя Gl-фаза регулируется «циклин D — CDK4/6» комплексом, в S-фазу вовлечены «циклин Е — CDK2» и «циклин А — CDK2», остальные фазы управляются «циклин В — CDK1/2» комплексами (100).
Опухолевые клетки нуждаются в гиперэкспрессии указанных компонентов для активации синтеза ДНК необходимых генов (S-фаза), а также митоза (М-фаза клеточного цикла). Поэтому в опухолях циклины и циклин-зависимые киназы обладают повышенной экспрессией, в то время как CDK-ингибиторы, такие как р21, р27, р1б, р15 практически отсутствуют.
Показано, что EGCG ингибирует CDK2 и CDK4, и при этом усиливает экспрессию p 2 1 w a f VCIP1 и р 2 7 к , Р 1 (60), в

Обоснованные направления терапии и профилактики...
неза с последующим вовлечением в апоптоз самих опухолевых клеток (7).
Еще один транскрипционный фактор, Ets-1, вовлечен в регуляцию неоангиогенеза. Он обладает совместным влиянием на экспрессию матриксных металлопротеиназ и VEGF, поскольку также имеет сайты связывания в промо-торных регионах указанных генов (41). Влияет ли EGCG и на этот фактор транскрипции — пока неизвестно. Однако этот факт сам по себе интересен и еще раз доказывает наличие определенной сочетанной регуляции в опухолях, направленной на реализацию программы выживания опухолевых клеток любым способом. Этим, в частности, можно объяснить феномен «ускользания» опухолей от монотерапии. Следовательно, наиболее предпочтительным выбором при лечении опухолей является комплексный подход, сочетающий влияние на все звенья опухолевой программы выживания, а не на конкретный сигнальный путь.
He-опухолевый неоангиогенез наблюдается, в частности, при псориазе. При этом заболевании также отмечена усиленная экспрессия VEGF и его рецепторов, что способствует улучшению трофики псориатической бляшки, дальнейшему ее увеличению и развитию локального воспаления. Блокирование VEGF или его рецепторов VEGF-B, VEGF-C и VEGF-D (например, моноклональными антителами) тормозит развитие воспаления, прекращает локальную пролиферативную активность. Это означает, что антиангиогенные факторы вносят существенную коррекцию в воспалительную реакцию. EGCG является антиан-гиогенным фактором, ингибирующим как белковую активность VEGF и рецепторов, так и уровень их транскрипции. Вовлечение ангиогенеза в развитие псориаза доказа-

ГЛАВА 8
В экспериментах in vivo также установлено блокирующее влияние EGCG на развитие сосудистой сети. Сравнительная антиангиогенная активность EGCG и ЕС исследовалась на животных моделях (51). Мышам подкожно вводились опухолевые клетки НТ29 (карцинома прямой кишки). В данном эксперименте демонстрировалась четкая зависимость концентрации и времени воздействия EGCG на ингибирование роста сосудов и индукцию апоптоза в опухолевых клетках (Рис. 23). Животные получали ежедневную инъекцию EGCG в количестве 1,5 мг в день интра-перитонально (ЕС использовался в качестве отрицательного контроля).
По сравнению с контролем, наблюдалось 30%-ное ингибирование плотности сосудистой сети, 58%-ное замедление опухолевого роста, замедление скорости опухолевой пролиферации на 27%, торможение метастазирова-ния. Аналогичные данные с эффектом EGCG на ангиоге-нез-зависимую дифференцировку эпителиальных клеток получены Singh и соавт. (87), (Рис. 24).
EGCG ингибирует активность эпидермального фактора роста и индуцирует апоптоз эндотелиальных клеток, что поддерживает гипотезу, высказанную в работе Shaahen с соавт., о том, что VEFG является фактором выживания опухолевого эндотелия (86). Блокирование VEGF на транскрипционном и белковом уровне ведет к апоптозу эндотелиальных клеток внутри опухолей, что не только ингибирует неоангиогенез, но и коммитирует сами опухолевые клетки к апоптозу (Рис. 25).
На другой модельной системе (человеческая не-Ходж-кинская лимфома) на животных показано, что включение EGCG в ежедневную диету также сопровождалось апоп-тозом эндотелиальных клеток, блокированием ангиоге-

Обоснованные направления терапии и профилактики...
сию, оксидативныи стресс в кератиноцитах. Совместное применение UV-облучения и EGCG при псориазе обеспечивает, с одной стороны, индукцию излишней пролиферации, сопровождающей заживление под влиянием облучения, а с другой — снимает излишнюю воспалительную реакцию под влиянием EGCG. Двойной механизм EGCG на выживание кератиноцитов связан, с одной стороны, со стимуляцией фосфорилирования Akt и Erk — киназ, препятствующих UV-индуцированному апоптозу, а с другой
— смещению баланса Bax/Bcl в сторону увеличения Bel. Это нормализует время жизни кератиноцитов (с 9 у больных псориазом до 27 дней в норме), способствует их диф-ференцировке (21).
Иммуномодулирующий аспект также вносит немаловажный вклад в дерматологический контроль пролифе-ративной активности. Сбой в системе ТМ/ТТгё-лимфо-цитов, дисбаланс про- и антивоспалительных цитокинов способствуют, как и в любом другом органе, смещению иммунологических ответов с соответствующими последствиями. Как правило, воспаление и аутоиммунные реакции сопровождаются и развиваются на фоне превышения активности Thl-подтипа лимфоцитов, развитие опухолей характеризуется снижением Thl-ответа, аллергий
— усилением Th2. При сильном ультрафиолетовом облучении наблюдается иммуносупрессия, как системная, так и местная. Увеличивается спектр ТИ2-цитокинов, наиболее активным из них является интерлейкин-10 (IL-10). EGCG угнетает экспрессию IL-10 в кератиноцитах, снимает локальное UV-индуцированное воспаление, блокирует синтез циклобутановых пиримидиновых ди-меров — факторов риска UV-ассоциированного рака

ГЛАВА 8
но с открытием генного полиморфизма по отношению к VEGF. У пациентов с серьезной формой псориаза отмечено нарушение системной регуляции VEGF из-за полиморфных вариантов рецепторов к VEGF в активированных кератиноцитах кожи (30, 104). Блокирование ангио-генеза катехинами приводило к стойкой и длительной ремиссии, сопровождаемой очевидными улучшениями состояния кожи, а также общим эмоциональным статусом пациентов.
Еще одно доказательство важности анти-ангиогенной терапии при псориазе получено при исследовании ретино-идов. Жирорастворимые витамины традиционно используются для лечения псориаза. Недавно выяснен механизм их анти-псориатического действия. Ретиноиды становятся биологически активными в результате активации ими одного из двух путей: взаимодействия со своими ядерными рецепторами (рецепторами ретиноевой кислоты), либо взаимодействуя с ядерным фактором транскрипции АР-1. Интересно, что в промоторной области гена VEGF имеются четыре АР-1-связывающих сайта; взаимодействие ре-тиноидов с АР-1 блокирует экспрессию VEGF, т.е. ретиноиды обладают анти-АР-1 и, следовательно, анти-VEGF-aK-тивностью.
Фотозащитный эффект EGCG
При определенных уровнях воздействия ультрафиолет становится сильным митогеном, что повышает риск развития рака кожи. Было показано, что употребление зеленого чая, либо его поверхностная аппликация на кожу оказывает защитный противоопухолевый эффект, предотвращает UV-индуцированное воспаление, иммуносупрес-

Обоснованные направления терапии и профилактики
Термогенез
Механизм индукции термогенеза — еще один интересный аспект, благодаря которому EGCG и некоторые другие фитонутриенты обладают свойством т.н. «сжигателей жира». На основе экстрактов зеленого чая на рынке имеются препараты, позволяющие снизить вес. EGCG ингиби-рует липазы желудка и панкреаса, при этом стимулируя термогенез (17). Наблюдаемые в процессе клинических испытаний эффекты были настолько сильные, что речь идет не просто о коррекции, а о лечении ожирения: через 3 месяца приема AR25 (Exolise™, экстракт зеленого чая) у добровольцев вес снизился на 4,6%, объем талии уменьшился на 4,8% (17). Интересные данные получены при изучении совместного влияния катехинов зеленого чая, кофеина и танина. Показано, что от комбинированного приема «катехин+кофеин» наблюдалось наиболее значимое снижение веса, в то время как от остальных комбинаций компонентов результаты были менее значительными. Т.е. кофеин и катехины обладают определенным синергизмом и препятствуют ожирению (106). Уровень три-глицеридов, жирных кислот, липопротеидов низкой плотности заметно снижается после употребления EGCG. Кроме того, уменьшается сывороточный уровень адипоцито-кина лептина — одного из косвенных показателей степени ожирения организма (85).
Чтобы замкнуть круг, следует, очевидно, связать влияние ожирения на риск развития рака. В рамках данного изложения авторы сочли необходимым порекомендовать читателю недавние полновесные исследования на эту интересную тему: Crew & Neugut, 2004 (27); Furberg et al., 2004 (33); Calle & Kaaks, 2004 (13); Lin et al., 2004 (62), a также более ранние работы.

ГЛАВА 8
кожи. В проведенных клинических испытаниях установлено, что после применения EGCG исчезали кожные эритемы, замедлялось фотостарение, а также снижался риск меланомного и не-меланомного рака кожи (54). В других исследованиях также подтверждался защитный эффект EGCG от UV-облучения за счет разных механизмов: предотвращения перекисного окисления липидов и разрушения оксидантных систем клетки и восстановления антиоксидантных ферментов (95), ингибирования транскрипционной активации NF-kB в кератиноцитах (1), блокады АР-1 (55). По мнению Elmets и соавт., 30-минутной аппликации экстракта зеленого чая достаточно для мощной превентивной защиты от UV-облучения (Elmets etal., 2001).
Анти-фотоканцерогенная активность EGCG тестировалась in vivo на мышах SKH-1 (безволосые). Накожная аппликация EGCG в концентрации примерно 1 м г / с м 2 наносилась в виде крема. Велись наблюдения за размером опухоли, частотой и множественностью опухолевых разрастаний, инцидентами возникновения новых опухолей. EGCG препятствовал онкогенной трансформации папиллом, индуцированной UV-облучением (68). Механизм предотвращения фотоканцерогенеза, по данным других авторов, связан также со значительным ингибированием метилирования ДНК, вызываемого ультрафиолетом (52, 53). При этом долговременная аппликация на коже не вызывала каких-либо признаков токсичности, как локальных, так и системных. Не отмечалось никакого повреждения кожи, потери массы тела, плотности костной ткани; при этом наблюдалось значительное снижение количества белой жировой ткани у животных.

Обоснованные направления терапии и профилактики...
слабореактивных свободных радикалов, которые не обладают столь сильным повреждающим эффектом.
В нормальных физиологических условиях небольшие количества кислорода, потребляемого митохондрией, постоянно конвертируются в супероксид анионы, перекись водорода, гидроксильные радикалы. В небольших количествах они выполняют важную роль в регуляции многих клеточных функций и действуют как вторичные мессен-джеры на активаторы транскрипции, такие как NF-kB (29). Избыточная продукция этих радикалов является фактором повреждения, и на этот счет в клетке имеется естественная анти-оксидантная система. Она представлена ферментами, среди которых наибольшее значение имеют Мп2+- и Си2+-зависимые супероксид-дисмутазы (SOD, ЕС 1.15.1.1), глютатион пероксидаза (GPX, ЕС 1.11.1.9), глютатион-редуктаза (GR, ЕС 1.8.1.7) и каталаза (CAT, EC 1.11.1.6). SOD конвертирует супероксид-анионы в перекись водорода, которая затем трансформируется в воду другими ферментами, GPX и CAT (98).
На активность антиоксидантных ферментов оказывают влияние также и возрастные изменения, которые выражаются в снижении их экспрессии и уровня в крови и тканях. Наибольший вклад в снижение активности естественных анти-оксидантных систем клетки вносят нарушения респираторной функции митохондрий, снижение митохон-дриального потенциала, выход цитохрома с, стимуляция про-апоптотических белков теплового шока (HSP10, HSP60), дисбаланс в системе белков Bax/Bcl, активация каспаз-3 и -9, других апоптогенных факторов. Таким образом, гиперпродукция свободных радикалов является также и характеристикой старения, что проявляется токсичностью на разных уровнях — органном, клеточном,

ГЛАВА 8
Антиоксидантная активность
Свойство EGCG и других чайных катехинов защищать от оксидативного стресса является наиболее изучаемым. Это, по сути, самое первое свойство, благодаря которому EGCG стал рассматриваться как заслуживающее пристального внимания вещество. Антиоксидантная активность EGCG обнаруживается во всех здоровых клетках, в то время как в опухолевых — наоборот, фиксируются про-оксидантные эффекты, вызывающие EGCG-индуци-рованное повреждение ДНК и апоптоз опухолей. Чем обусловлена такая «интеллектуальная» подстройка и различия в типе клеток — пока не совсем ясно.
Продукция свободных радикалов, приводящая к окислению липидов клеточных мембран, является одним из мощных стимулов, повреждающих клетки и ткани. Усиле-ние оксидативного стресса, в свою очередь, приводит к нарушению естественных антиоксидантных систем. Для предотвращения или коррекции оксидативного стресса применяются различные антиоксиданты, обладающие разной силой защитного ответа. В ряду известных натуральных и синтетических антиоксидантов известны какао, транс-ресвератрол, компоненты зеленого чая, оливкового масла, витамины Е (а-токоферол) и С, некоторые минералы (селен). Кроме того, существует также множество искусственно созданных веществ с аналогичными свойствами — гидроксианизол (butylated hydroxyanisole, ВНА), гидрокситолюэн (butylated hydroxytoluene, BHT), пропил-галлат (PG) (97). В основе механизма антиокси-дантного действия всех этих веществ лежит ингибирова-ние окисления липидов формированием относительно

Обоснованные направления терапии и профилактики...
Другие антипролиферативные эффекты EGCG
EGCG оказывает ингибирующее действие на развитие гипертрофии гладкомышечных клеток, предотвращая, таким образом, развитие атеросклеротических бляшек, утолщения стенок сосудов, гипертензии. Молекулярной основой ингибирования является прямая блокада ангио-тензин-И-индуцируемой активности JNK-киназы (c-Jun N-terminal kinase) в гладкомышечных клетках. Для сравнения, другие катехины зеленого чая не обладают аналогичными эффектами (105).
Антифибротические свойства EGCG подтверждены в экспериментах in vitro. Воспалительные изменения в печени диктуют развитие фибротических процессов. На культуре клеток LI90 EGCG ингибировал PDGF-BB — индуцируемую клеточную пролиферацию, а также мРНК коллагенов альфа-l и IV типов (83). В этой работе показано, что EGCG снижает способность к аутофосфорилированию рецептора к PDGF-BB, что прерывает пролиферативный каскад и блокирует фибротические образования в печеночных клетках.
Исследовался эффект EGCG на рост, пролиферацию, апоптоз и теломеразную активность при цервикальном канцерогенезе. Изучались различные дозы EGCG (от 0 до 100 мМ) на культуре первичных эндоцервикальных клеток человека различных линий, в том числе зараженных вирусом папилломатоза (HPV18). Показано, что антиканцерогенный эффект EGCG опосредуется за счет индукции апоп-тоза и снижении активности теломеразы в клетках дис-пластического эпителия (103). Цервикальные повреждения, вызванные HPV, нивелировались после употребления 200 мг/день EGCG у добровольцев, либо принимающих

ГЛАВА 8
генном. Стимуляция естественных антиоксидантных систем, привнесение экзогенных антиоксидантов — ключевой момент лечения и профилактики воспаления, канцерогенеза, раннего старения.
Антиоксидантная активность EGCG была исследована на культуре гепатоцитов крыс. Гепатоциты инкубировались с канцерогенами — беномилом и 1-нитропеном. Воздействие EGCG на клетки мониторировалось измерением малонилальдегида — маркера перекисного окисления липидов, а также изменением активности глютатиона (GSH), глютатион-пероксидазы, глютатион-редуктазы. По сравнению с контролем, концентрация антиоксидантных ферментов увеличилась более чем на 200%, а малонилальдегида уменьшилась на 23% (79).
Антиоксидантные свойства EGCG изучались на животных моделях с почечной недостаточностью, вызванной метилгуанидином (MG). Это уремический токсин, продуцируемый из креатинина под влиянием гидроксильного радикала. Мышам вводился креатинин интраперитонеаль-но. Добавление мышам в питьевую воду EGCG за 30 минут до и после инъекции креатинина способствовало ингиби-рованию продукции MG за счет инактивации гидроксильного радикала (71).
Похожие данные получены при исследовании нейродеге-неративных заболеваний — болезни Альцгеймера, Паркин-сона, хореи Гентингтона, боковом амиотрофическом склерозе. EGCG выступал в качестве монотерапии и обнаруживал свойства скевенджера свободных радикалов, хелатора металлов (железа и меди), стимулятора защитных анти-ок-сидантных ферментов, чья экспрессия нарушается в процессе ишемии-реперфузии нейронов, токсическом повреждении, окислении, других повреждающих стимулов (66).

Обоснованные направления терапии и профилактики.
вещество обладает анти-аллергическими свойствами, подавляя экспрессию высокоафинного рецептора к IgE. Известно, что IgE является одним из наиболее ключевых факторов в развитии аллергических реакций. Анти-ал-лергические свойства EGCG основаны также на блокаде ERK1/2 — сигнального пути, что также ингибирует экспрессию рецептора к IgE.
Таким образом, EGCG является не просто противовоспалительным веществом: влияние на активность Th1- и ТЬ2-подтипа лимфоцитов обеспечивает восстановление Th1/Th2 баланса — свойство, которым не обладает ни один современный иммуномодулирующий препарат, основное назначение которых — лишь активация Т-клеточно-го ответа (т.е. стимуляция ТМ-звена иммунной системы).
Получены интересные данные об анти-кариесном действии чая: EGCG полностью блокирует глюкозилтрансфе-разу стрептококков, живущих в ротовой полости (40). Кроме того, EGCG ингибирует продукцию токсических метаболитов Porphyromonas gingivalis — вирулентного микроорганизма, вызывающего пародонтоз. Наблюдалось снижение продукции пропионовой и других кислот, разрушающих зубную эмаль, в концентрации 1-2 мг/мл (82). Makimura и соавторы обнаружили полное ингибиро-вание ферментной активности коллагеназы чайными ка-техинами у пациентов с острыми гингивитами (65).
На фоне употребления зеленого чая снижался риск развития катаракты: ослабевало помутнение хрусталика за счет снижения преципитации белков, стимулируемое EGCG (92). Сочетание антиангиогенной и антипролифе-ративной активностей по отношению к эпителиальным клеткам вносит дополнительную ясность в объяснение эффектов EGCG при данной патологии.

ГЛАВА 8
катехин в виде капсул в течение 8-12 недель, либо в виде крема местно. Эффект от крема отмечен у 74% пациентов, от приема капсулы — у 50%, а в группе, где использовались оба вида приема EGCG — 75% (3). Эти данные характеризуют EGCG как мощный противовирусный препарат, с направленной активностью на эндотелиальные клетки, к которым HPV имеет высокую тропность.
Модуляция цитохромной системы EGCG исследована in vivo на мышах Swiss-Webster. EGCG вовлечен в метаболизм и синтез эстрадиола — одного из основных эстрогенов с сильной пролиферативной активностью. При добавлении EGCG в различных концентрациях (12.5, 25 или 50 мг/кг/день) были измерены активность ароматазы (CYP19), а также цитохромов Р-450 CYP3A, CYP1A. Под влиянием EGCG активность ароматазы была ингибирована на 56%, активность CYP3A возросла на 31-47%, a CYP1A1 — оставалась неизменной. Эти данные говорят о наличии антипролиферативной активности EGCG на тканях-мишенях ароматазы и эстрадиола (опухоли эндометрия, миоме-трия, опухоли молочной железы), что объясняет блокаду гиперпластических процессов и противоопухолевые свойства EGCG в тканях репродуктивной системы (38).
Употребление зеленого чая пожилыми женщинами (65-76 лет) ассоциировалось с увеличением плотности и минерализации костной ткани, несмотря на гипоэстрогенемию (42). Но этот эффект достигается, очевидно, не за счет ка-техинов, а за счет фитоэстрогенов чая, которые стимулируют остеобластогенез: EGCG ингибирует щелочную фосфатазу — маркер созревания остеобластов, и никак не влияет на минерализацию кости (101).
Антипролиферативные активности EGCG распространяются и на ТИ2-подтип лимфоцитов. Показано, что это

Обоснованные направления терапии и профилактики...
низации Здравоохранения ежегодно регистрируется около 570 тысяч новых случаев. Во всем мире он занимает второе, в России — первое место в ряду смертности от опухолевых заболеваний.
Как было отмечено ранее, одним из необходимых факторов, которые используют как здоровые, так и опухолевые клетки, является эстроген. Взаимодействие эстрогена с рецептором (ER) инициирует проникновение образовавшегося комплекса в ядро, где эстрогеновый рецептор выполняет роль активатора транскрипции, стимулируя экспрессию множества эстроген-стимулируемых генов. Действуя в паракринной и аутокринной манере, вновь синтезируемые белки стимулируют рост самой клетки, а также вовлечены во многие другие антиапоптотические функции. В числе таких генов — изоформы цитохрома Р-450 (CYP450), а именно — CYP1B1. Субстратом для данной формы цитохрома является сам эстроген, в результате чего образуется его гидроксиметаболит, окисленный в 16-м положении, а именно — 16-альфа-гидрок-сиэстрон (16ссОНЕ1) (63). Эта форма эстрогена обладает наиболее сильной пролиферативной активностью по сравнению как с нативным эстрогеном, так и его любыми другими метаболитами. Именно эта форма эстрогена несет мощный пролиферативный и онкогенный потенциал и способна активировать эстрогенный рецептор (31, 74).
Модуляция цитохромной системы индол-3-карбинолом состоит в индукции другой изоформы CYP450, а именно — CYP1A1 (5). Данная изоформа гидроксилирует эстроген во 2-м положении, с образованием 2-гидроксиэстрона (20НЕ1). Полученный метаболит является антагонистом рецептора эстрогена и блокирует его активацию самим эстрогеном, а также его опасными метаболитами (9). Так

ГЛАВА 8
Таким образом, EGCG вовлечен практически во все процессы, которыми характеризуется гиперпластический рост. В данном разделе описан один из ярких примеров влияния природных фитонутриентов на предотвращение и блокаду опухолевого роста. Среди многих других не менее интересным представляется другой природный компонент, который способен вмешиваться в гормональный метаболизм и корректировать рост-стимулирующие влияния гормонов (эстрогенов) на клетках. Речь идет об ин-дол-3-карбиноле.
Механизмы противоопухолевой активности индол-3-карбинола
Индол-3-карбинол (I3C) — компонент овощей семейства крестоцветных (в основном, это все виды капусты: брокколи, цветная, белокочанная, брюссельская) (78). За более чем 10-летнюю историю исследований по I3C показано, что это вещество обладает .высокой избирательной противоопухолевой активностью, влияющей на многие типы клеток, включая клетки молочной железы. Изначально считалось, что I3C оказывает влияние только на те клетки, в которых эстрогены являются факторами, провоцирующими пролиферацию. Однако позже было установлено, что I3C вызывает апоптоз опухолевых клеток вне зависимости от наличия эстрогеновых рецепторов.
Анти-эстрогенный эффект
Наиболее типичный пример эстрогенной стимуляции — пролиферативные заболевания молочной железы. Наиболее опасным является рак: по данным Всемирной орга-

Обоснованные направления терапии и профилактики...
как эпидермальный фактор роста (EGF), фибробластиче-ский фактор роста (FGF), в том числе фактор роста кера-тиноцитов (FGF-7 или KGF), инсулино-подобный фактор роста (IGF-I). В отсутствие какого-либо препятствия, ростовые факторы активируют свои рецепторы, обладающие киназной активностью. Лиганд-индуцируемая димериза-ция рецепторов активирует каскад цитоплазматических киназ (МАРК, Akt), протоонкогенов (fos, ras), активаторов транскрипции (NF-kB), что приводит к синтезу лиганд-индуцируемых генов, среди которых — ростовые факторы и рецепторы к ним, цитоплазматические киназы, факторы для роста новых сосудов, белки теплового шока, другие необходимые для опухолевого метаболизма белки. I3C, проникая в клетку, блокирует множество сигнальных путей, препятствуя фосфорилированию тирозиновых остатков киназ, что мешает каскадной передаче пролифе-ративных сигналов с поверхности к ядру клетки (20). Кроме того, он ингибирует NF-kB, который является основным фактором активации транскрипции множества генов, вовлеченных в пролиферацию и воспаление (12).
Накапливаются данные по ингибированию I3C специфических опухолевых генов. MUC1 — трансмембранный гликопротеин, экспрессирующийся в большинстве карцином. Уровень экспрессии MUC1 ассоциируется со степенью агрессивности опухоли; MUCI-антиген используется в качестве маркера для мониторирования пациентов с опухолями молочной железы. Было показано, что I3C ингибирует экспрессию MUC1 как в эстроген-зависимых (MCF-7), так и эстроген-независимых (MDA-MB-468) клетках, в доза-зависимой и время-зависимой манере, на белковом и транскрипционном уровне (57).

ГЛАВА 8
прерывается синтез эстроген-зависимых генов, и опухолевая клетка не способна получить эстроген-зависимую сти- | муляцию. В этом суть анти-эстрогенного эффекта I3C.
Применение I3C как противоопухолевого средства в отношении рака молочной железы было начато в 1995 году (9). Было показано, что уровень CYP1A1 регулировался в зависимости от концентрации I3C: ферментная активность CYP450 увеличивалась линейно от 10 до 150 | пмоль/мг белка при дозах от 1 до 500 ммоль I3C на одно животное. В сравнении с другими диетическими индолами, I3C проявлял наиболее сильную СУР1А1-стимули-рующую активность, сопровождающуюся повышением 2-гидроксиэстрона. При этом наблюдалась статистически значимая регрессия эстроген-зависимого роста опухолевой ткани по сравнению с таковой в контрольной группе животных.
Нужно отметить, что восприимчивость к I3C довольно вариабельна: индивидуальный ответ проявляется благодаря генетическим вариациям и зависит от индуцибельно-сти CYP1A1. Этот эффект проанализирован в популяциях европейских и африкано-американских женщин: у первых соотношение 2/16а почти в 2 раза выше (2.25±1.42) (89).
Модуляция сигнальных путей ростовых факторов
Антипролиферативная активность I3C распространяется как на эстроген-зависимые, так и эстроген-независимые клетки, где пролиферативные каскады осуществляются без участия эстрогена (т.е. не имеющих эстрогено-вых рецепторов). Напомним, что рост таких клеток чаще всего зависит от активности ростовых факторов, таких

Обоснованные направления терапии и профилактики.
Гены
Вах
CDK6
Akt
Р 2 7 , р 2 1 , р16
HSP27
EGF, EGFR, TGFb, FGF
NF-kB и NF-kB-ин-дуцибель-ные гены
Функция в опухолевой клетке
Цитоплазматический белок, маркер апоптоза. Транслокация в митохондрию сопровождается выходом цитохрома с с последующим вовлечением каспаз 3 и 9 и развитием апоптоза
Циклин-зависимая киназа. Стимулирует рост опухолевых клеток, наиболее активна в Gl-фазе клеточного цикла. Эстроген-независима
Киназа, один из участников EGF-индуцируемого сигнального каскада. Стимулирует пролиферацию опухолевых клеток
CDK-ингибиторы, опухолевые су-прессоры
Белок теплового шока, выполняющий функцию стабилизации структуры белков, участвующих в пролиферации клеток. Экспрессия многократно повышена. Ингибитор апоптоза. Ассоциирован с агрессивной формой рака молочной железы. Вовлечен в мета-стазирование
Ростовые факторы и их рецепторы. Стимулируют пролиферацию опухолей
Ядерный фактор активации транскрипции генов, индуцируемых ростовыми факторами. Вовлечен в пролиферацию и выживание опухолевых клеток
Эффект I3C
Усиление транскрипции
Блокада активности
Блокада активности, вызванная нарушением фосфорилиро-вания
Усиление транскрипции
Блокада активности
Ингибирование транскрипции
Ингибирование транскрипции

ГЛАВА 8
Еще один механизм блокады опухолевого роста ин-дол-3-карбинолом связан с ингибированием активности циклооксигеназы-2 (СОХ-2) — фермента, вовлеченного в синтез простагландинов (PGE2). СОХ-2 активируется в ответ на провоспалительные цитокины — фактор некроза опухоли-альфа (TNF-ct) и его «партнера» — интерлейки-на-1 (IL-1). Взаимодействие цитокинов со своими рецепторами приводит, в частности, к активации СОХ-2, синтезу PGE2 с последующей транскрипцией фактора роста эндотелия сосудов (VEGF) — основного компонента опухолевого неоангиогенеза.
Таким образом, I3C регулирует баланс про- и антиапоп-тотических факторов как на уровне транскрипции генов, так и посредством блокады активности зрелых белков. Сведения о влиянии I3C на регуляцию активности ключевых внутриклеточных пролиферативных сигналов суммированы в Таблице 22.
Таблица 22 Регуляция I3C транскрипции генов в опухолевой клетке
Гены
CYP1A1
[ BRCA1
Функция в опухолевой клетке
Гидроксилирование эстриола во 2 положении с образованием 2-ОНЕ1 производного, обладающего антипролиферативной активностью
Специфический опухолевый су-прессор в клетках молочной железы. Осуществляет блокаду транскрипции генов, индуцируемых ERa
Эффект I3C
Усиление транскрипции через активацию Ah-pe-цептора
Усиление транскрипции

Обоснованные направления терапии и профилактики...
модисплазия — вот неполный перечень заболеваний, которые ассоциируются с этой инфекцией. Папилломатозом старадают как взрослые, так и дети: случаи папилломато-за гортани у новорожденных — явление не такое уж и редкое. Первые признаки могут появиться уже в 6-месячном возрасте, и если не предпринимать ургентных мер, быстровырастающие гроздеобразные папилломы могут привести к асфиксии. Совершенно очевидно, что хирургическое лечение эффективно только до следующего рецидива, который наступает очень быстро. Больные папилломатозом гортани переносят до 200 операций ежегодно.
Распространение заболеваемости цервикальным раком в развивающихся странах сродни эпидемии. Все сексуально активное население так или иначе находится в группе риска. Ежегодно регистрируется более 500 тысяч новых случаев в мире. Рак шейки матки является первым опухолевым заболеванием, для которого однозначно идентифицирован этиологический фактор. Эпидемиологические и лабораторные исследования установили, что основной причиной является вирус папилломатоза человека (ВПЧ), обнаруженный в 100% случаев.
Примерно 30 типов вируса папилломатоза инфицируют генитальный тракт, а четыре из них (ВПЧ 16, 18,31 и 45 типа) вносят значительный (до 80%) вклад в эту статистику.
Лечение цервикального рака в настоящее время довольно сложное, и далеко не всегда проходит удачно. Диагностические критерии отсутствовали до недавнего времени: трудность заключается в том, что 90% случаев ге-нитального папилломатоза заканчивается спонтанной ремиссией, а 10% случаев — малигнизацией. В случае ремиссии никаких клинических проявлений обычно нет, но вирус остается в организме, что сохраняет риск заражения партнеров и развития неоплазий.

ГЛАВА 8
I3C обладает мощным антипролиферативным эффектом, позволяющим ингибировать воспаление, рост и пролиферацию опухолевых клеток, в том числе зараженных вирусом папилломатоза, а также рост новых сосудов. Кроме того, отмечен прямой апоптотический эффект I3C на опухолевые клетки: I3C усиливает экспрессию естественных опухолевых супрессоров, р21 и р53, которые, в свою очередь, являются ингибиторами циклин-зависимой киназы (CDK) — основного стимулятора опухолевого клеточного цикла (11, 26).
Противовирусный потенциал I3C в отношении вируса папилломатоза человека
Относительно недавно установлен еще один необычный эффект I3C: он вызывает апоптоз клеток, зараженных вирусом папилломатоза человека, в том числе теми типами вируса, которые обладают онкогенным потенциалом (23, 88). С учетом трудностей, сопровождающих терапию папилломатоза слизистых, данное свойство делает I3C незаменимым противовирусным препаратом с параллельной стимуляцией Т-клеточного звена иммунитета.
Медико-социальные аспекты папилломатоза
Вирус папилломатоза человека (HPV) имеет очень широкую распространенность; согласно данным ВОЗ, носителями HPV являются 23% населения земного шара, по другим данным — все 60%. Кожные папилломы, остроконечные кондиломы, рак агоненитальной области (шейки матки, вульвы, влагалища), ювенильный папилломатоз гортани (веррукозный ларингит), бородавчатая эпидер-

Обоснованные направления терапии и профилактики.,
тельством в какой-то конкретный сигнальный путь. Еб и Е7 также вмешиваются в регуляцию тех путей, принципиально необходимых клетке для выживания. Один из них — прямая регуляция активности ядерных рецепторов — тироидных (TR), андрогеновых (AR), эстрогеновых (ER). Еб и Е7 обладают способностью образовывать с ними комплексы и активировать их транскрипционный потенциал. Функциональная роль этих белков зависит от клеточного типа, особенностей промоторного региона, а также самого ядерного рецептора. К примеру, регуляция ER происходит через АР-1 и АР-2 сайты — т.е. те участки промотора эстрогенового рецептора, через которые осуществляется основная регуляторная роль и стимуляция транскрипции эстроген-зависимых генов. Вторая причина — подавление транскрипции TGF-p, выполняющего в некоторых типах клеток функции опухолевого супрессора. Показано, что HPV 16 Е7 значительно подавляет промо-торную активность одной из изоформ TGFp — TGFp-2 в клетках NIH/3T3 (дикий тип, имеющий опухолевые су-прессоры р53 и pRb). Необходимым условием блокирования TGFp-промоторной активности является взаимодействие Е7 с pRb.
Супрессоры опухолевого роста р53 и pRb вовлечены также в регуляцию экспрессии про- и анти-ангиогенных факторов, и их блокада в HPV-клетках, очевидно, должна нарушать регуляторные механизмы ангиогенеза. Вопрос об альтерациях онкобелков в процесс неоангиогенеза исследовался в следующем эксперименте. Кератиноциты человека были инфицированы рекомбинантным ретрови-русом, экспрессирующим Еб и Е7 (93). RT-PCR, ELISA и Вестерн-блот показали, что в клетках, экспрессирующих HPV 16 Еб и Е7, экспрессионный уровень двух ингибито-

ГЛАВА 8
HPV-онкомаркеры
В последнее время идентифицированы антигены, являющиеся специфическими для HPV-ассоциированных инфекций. Наибольшее диагностическое и клиническое значение имеют два белка, онкобелки — маркеры Е6 и Е7. Во многих случаях вирус папилломатоза интегрируется в геном зараженных клеток хозяина, при этом наблюдается повышение экспрессии этих антигенов на клеточной поверхности. Они довольно иммуногенны, хорошо распознаются Т-лимфоцитами организма и являются, таким образом, специфическими Т-антигенами. Онкобелки проявляют свой эффект через формирование комплексов со специальными белками — р53 и pRb1), которые обычно выполняют в клетке противоопухолевую функцию. В связанном с онкобелками состоянии они не способны выполнять эти функции, что вызывает снижение внутриклеточных противоопухолевых систем защиты. Инактивация р53 и Rb вызывает нарушения в клеточном цикле клетки, что способствует дальнейшей «генетической нестабильности» и развитию дополнительных генетических альтераций. Необходимо отметить, что, в зависимости от активности вируса, блокада противоопухолевых свойств этих белков происходит с разной силой: у вирусов с высоким потенциалом развития опухолей комплексы «Е6-р53» и «E7-Rb» более стабильны.
Молекулярные механизмы иммортальности клетки, зараженной вирусом, определены благодаря изучению регулирующих активностей онкобелков Е6 и Е7. Как обычно бывает, воздействие онкогена не ограничивается вмеша-
Ретинобластома — связывающий белок, опухолевый супрессор

ГЛАВА 8
ров ангиогенеза, тромбоспондина-1 и маспина, был даже ниже, чем в контроле. При этом наблюдалась гиперэкспрессия основных стимуляторов ангиогенеза — VEGF и IL-8. В HPV-зараженных кератиноцитах отмечались продукты альтернативного сплайсинга VEGF — изоформы VEGF12v VEGF145» VEGF165 и VEGF18g. Стимулирование опухолевого вирус-индуцированного ангиогенеза — еще один механизм, объясняющий иммортальность HPV-зараженных клеток. В дополнение к этому можно отметить Е6-стимулированную активацию МАРК-сигнальных антиапоп-тотических каскадов (16), индукцию теломеразы в кератиноцитах белком Еб HPV 16 типа (75, 76), повышение активности СОХ-2 в клетках с цервикальной дисплазией (37, 56), экспрессии матриксных металлопротеиназ (90), уро-киназы (иРА) (81, 94), катастрофическое снижение соотношения Bax/Bcl в HPV-инфицированных клетках (70), многих других антиапоптотических стимулов.
Иммуномодулирующий эффект I3C
Относительно недавно появились новые исследования, касающиеся одной из наиболее важных составляющих активностей I3C. Блокада пролиферативных стимулов — не единственная роль I3C в HPV-инфицированной клетке. Сочетания противоопухолевых и противовирусных эффектов I3C исследовали Chatterji с соавторами (18). Оказалось, что в промоторной области рецептора к интерфе-рону-гамма (IFN-yR1) имеются сайты связывания для I3C. Попадание в клетку I3C запускает, кроме всего прочего, транскрипцию IFN-yR1, что, в свою очередь, стимулирует восприимчивость опухолевых клеток к интерферону (IFN-y). Этот эффект был проверен авторами на клетках

Обоснованные направлении терапии и профилактики...
опухоли молочной железы. Показано, что комбинационное использование I3C и IFN-y активирует STAT 1-белки усилением фосфорилирования тирозина Tyr7ov Кроме того, синергизм этих компонентов вызывает прекращение клеточного цикла в G1 фазе и стимулирует экспрессию р21, ингибитора клеточного цикла. Эти интереснейшие данные вносят дополнительный вклад в объяснение необычной полифункциональности индол-3-карбинола и также объясняют его противовирусный эффект: интерфе-роны 2 типа, к которым относится IFN-y, являются сильнейшими факторами с мощной противовирусной активностью.
Связывание интерферонов с рецептором индуцирует экспрессию ряда новых генов в клетке-мишени. Это ну-клеазы, синтазы и протеинкиназы, которые оказывают влияние на белковый синтез и ответственны за проявление антивирусной активности. В частности, вновь синтезированные протеинкиназы инактивируют фактор инициации elF2a и таким образом ингибируют синтез вирусных белков. Экспрессия другого интерферон-индуцируе-мого фермента — 2',5'-полиаденилат синтазы, способствует активации эндонуклеаз, которые подвергают деградации вирусную мРНК.
Влияние на плотность кости
Ингибирование активности эстрогенов индол-3-карби-нолом теоретически должно приводить к остеопорозу, т.е. усилению стимуляции остеокластогенеза. Однако наблюдения in vivo опровергают эти опасения: показано, что одна из форм индол-3-карбинола, DIM, усиливает активность щелочной фосфатазы, маркера остеобластогенеза, в 4 раза. Повышение тРНК щелочной фосфатазы проис-

ГЛАВА 8
ходит за счет активации TGF-(5 — ростового фактора, индуцируемого индол-3-карбинолом. Последнее свойство очень ценно, так как TGF-J3— это как раз тот цитокин, который модулирует (трансформирует) клеточный рост: стимулирование его экспрессии в опухолях способствует замедлению роста опухолевой клетки. Рекомбинантные изоформы TGF-p вовлечены в дифференцировку многих типов опухолевых клеток in vitro и in vivo, выступая, таким образом, в качестве опухолевых супрессоров.
Механизм апоптоза опухолевых клеток под влиянием I3C
К настоящему времени определены ключевые механизмы и факторы, вовлеченные в развитие апоптотиче-ского процесса клеток-мишеней. Нарушение баланса ан-ти- и проапоптотических факторов сдвигает равновесие в пользу какого-то из них, в случае преобладания последних регистрируется гибель клетки. Одними из важнейших в этом процессе рассматриваются белки семейства Вс1-2, среди которых — Вах, Bcl-2, BCI- X L > некоторые другие. Вах индуцирует апоптоз, в то время как Bcl-2, Bcl- X L — ингибируют его развитие. Показано, что отношение Вах:Вс1-2 является наиболее важным показателем апоптоза опухолевых клеток, индуцируемого противоопухолевыми препаратами (67). Кроме того, комплексный анализ апоптоза подразумевает изучение внутриклеточной локализации белка Вах. Транслокация цитозольного Вах в митохондрию индуцирует выход цитохрома с, который активирует каспазы-3 и -9, являющиеся ключевыми в последовательности апоптотических процессов (8, 32).

Обоснованные направления терапии и профилактики...
Под влиянием I3C в опухолевых клетках молочной желе-зы происходит модуляция экспрессии генов, вовлеченных в апоптоз. Так, на клеточных моделях MCF10A и СА1а, являющимися нормальными и опухолевыми эпителиальными клетками молочной железы, показано, что I3C значительно блокирует экспрессию Bcl-2 в опухолевых СА1а клетках, в то время как в здоровых MCF10A — нет (78, 84). В обоих типах клеток регистрировалась транслокация Вах в митохондрию, однако потеря митохондриального потенциала, выход цитохрома с и индукция апоптоза наблюдались только в опухолевых клетках. Как видно из этих наблюдений, транслокации Вах в митохондрию недостаточно для индукции апоптоза; необходимы дополнительные механизмы, вовлекающие митохондриальную деполяризацию с последующей активацией про-апоптотических каспаз. Все эти события в опухолевой клетке индуцирует I3C.
Еще два сигнальных пути, а именно — Akt (митоген-ак-тивируемая протеинкиназа) и NF-kB, вовлечены во взаимодействие с белками семейства Bcl-2 и являются важнейшими регуляторами программы выживания клетки.
Сигнальные пути, в которые вовлечена киназа Akt, ведут к анти-апоптотическим событиям в нормальных и опухолевых клетках. В норме они активируются в ответ на стимулы от ростовых факторов, обладающих также и ми-тогенными активностями, и приводят к росту клеток-мишеней, а в случае опухолевой трансформации — пролиферации клеток. В регуляции активностей факторов, вовлеченных в каскад индуцируемых митогенами реакций, Akt играет важную роль. Она осуществляет анти-апопто-тические функции за счет многих механизмов. В частности, ингибирует конформационные изменения белка Вах, тем самым предотвращая его транслокацию в митохон-

ГЛАВА 8
дрию, что, в свою очередь, предотвращает потерю потенциала внутренней митохондриальной мембраны, активацию цитохрома с и развитие апоптоза (45).
I3C оказывает терапевтическое воздействие на эпителиальные клетки, одновременно являющиеся тканями-мишенями эстрогенов. Сигнальными молекулами, индуцирующими запуск пролиферативного механизма в этих клетках, являются, среди прочих, эндотелиальный фактор роста (EGF), его рецепторы (EGFR) и эстрогены. EGF, взаимодействуя со своими рецепторами, индуцирует сигнальный каскад с вовлечением специфических цитоплазматических белков (РОК, Akt). Активация этого пути в клетке ведет к стимуляции ее пролиферации. В свою очередь, взаимодействие эстрогенов со своими рецепторами приводит к фос-форилированию ассоциированного с рецептором белка теплового шока HSP27, выполняющего анти-апоптотические функции в клетке (15, 35). HSP27 экспрессируется в опухолях млекопитающих и его экспрессия многократно повышается после стимуляции эстрогенами (22, 77). Таким образом, существуют две возможности выживания опухолевой эстроген-зависимой эпителиальной клетки.
Уникальность действия I3C заключается в том, что он вмешивается в оба эти пути, нарушая внутриклеточные каскады и препятствуя клеточной пролиферативной активности. С одной стороны, он обладает антиэстрогенным эффектом, стимулируя образование анти-пролиферативного 2-гидроксиэстрона, а с другой — препятствует фосфори-лированию цитоплазматических белков — участников каскадной передачи, индуцируемой EGF. Блокирование анти-апоптотического пути в эпителиальной клетке — суть антиопухолевого эффекта индол-3-карбинола на тех опухолях, на которых повышена экспрессия рецепторов к EGF.

Обоснованные направления тсропии и профилактики...
Сравнение активностей тамоксифена и индол-3 к а р б и н о л а
В последние 30 лет самым популярным препаратом, назначаемым при лечении пролиферативных заболеваний молочной железы, был и остается тамоксифен. Тем не менее, в литературе накапливаются данные о том, что длительный прием тамоксифена сопровождается рядом нежелательных побочных эффектов.
Успех лечения от тамоксифена определяется, в основном, наличием эстрогеновых рецепторов на клетках карцином молочной железы. При этом примерно у половины пациентов с эстроген-зависимой формой опухоли происходит довольно быстрое улучшение, тогда как у остальных заболевание прогрессирует и развивается тамоксифен-устойчивый фенотип, что ставит под сомнение и делает небезопасным дальнейшее применение этого препарата: показано, что длительный прием тамоксифена зачастую приводит к развитию опухолей в здоровых тканях, и на этот счет имеется множество публикаций (10, 24, 28, 39, 58, 72). В частности, тамоксифен ассоциирован с риском развития опухолей эндометрия (19,44). Показано, что на фоне приема тамоксифена частота опухолей эндометриаль-ного происхождения выросла в 2-3 раза и составляет 80 случаев на 10000 (4). По мнению этих авторов, механизм опухолевой индукции связывают с блокадой тамоксифе-ном гена р53 — естественного супрессора опухолей. У здоровых субъектов р53 «посылает» сигналы к прекращению клеточного цикла, и, если в цикл вовлечена атипичная клетка, вызывает ее деструкцию. И если подобная активность тамоксифена, несомненно, полезна для усиления чувствительности раковой клетки к химиотерапии, в здо-

ГЛАВА 8
ровой клетке этот феномен провоцирует нарушение устойчивости к развитию тамоксифен-зависимого рака.
Таким образом, индол-3-карбинол отчасти похож ната-моксифен по тем показателям, которые направлены на гибель опухолевых клеток. Он также прерывает клеточный цикл. Однако его анти-опухолевая активность выше и безопаснее, чем у тамоксифена. Сравнительный анализ антиопухолевой активности показал, что тамоксифен вызывает 60%-ную гибель клеток, тогда как индол-3-карбинол — 90% (25). Важным отличием между ним является тот факт, что в ER-негативных опухолях индол-3-карбинол прекращает синтез ДНК вновь образованных клеток, в то время как тамоксифен аналогичным эффектом не обладает. Индол-3-карбинол восстанавливает р21, р53 и другие опухолевые супрессоры, которые работают в процессе синтеза новых клеток. Тамоксифен же вызывает в них мутации (80). В отличие от тамоксифена, индол-3-карбинол вызывает апоптоз клеток, вызванных химическими канцерогенами, в том числе и диоксином. Остается добавить, что индол-3-карбинол малотоксичен. Согласно клиническим испытаниям, в дозе, многократно превышающей терапевтическую, которая составляет 400 мг/день, индол-3-карбинол не вызывает никаких побочных эффектов.
Таким образом, можно вычленить ряд преимуществ ин-дол-3-карбинола перед тамоксифеном. Во-первых, ин-дол-3-карбинол не имеет побочных эффектов, в то время как тамоксифен вызывает ряд последствий, которые превращаются в самостоятельные заболевания. Во-вторых, лечение индол-3-карбинолом основного заболевания (например, опухолей молочной железы) сопровождается параллельной защитой органов и тканей от возникновения опухолей любого происхождения, будь то результат ме-

Обоснованные направления терапии и профилактики...
тастазирования, либо химическая индукция веществами из пищи или окружающей среды. Кроме того, спектр применения индол-3-карбинола гораздо выше, чем тамокси-фена: последний проявляет избирательную активность исключительно на эстроген-зависимых опухолях, в то время как индол-3-карбинол вызывает апоптоз опухолевых клеток вне зависимости от того, чем вызван их рост — эстрогеном, либо эпидермальным фактором роста, иными стимулами. Отсюда вытекает еще одно важное свойство данного вещества: его противоопухолевая активность распространяется не только на клетки молочной железы, но и на другие ткани, где работает EGF и другие ростовые факторы — простата, желудок, пищевод, мочевой пузырь, щитовидная железа и т.д. Очевидно, что лечение индол-3-карбинолом опухолей вышеназванных тканей — вопрос самого ближайшего будущего.
Сравнительный анализ активностей тамоксифена и ин-дол-3-карбинола приведен в Таблице 23.
Представленные данные неопровержимо доказывают преимущество индол-3-карбинола в терапии пролифера-тивных заболеваний молочной железы, к которым, вообще говоря, относится не только рак. Не меньшей проблемой перед маммологами стоит мастопатия. И если узелковую форму можно лечить хирургически, то диффузный вариант такому лечению не поддается. Поэтому и в этом случае объектом для терапии выбора может стать индол-3-карбинол.
Вместе с тем молекулярные механизмы, благодаря которым I3C вызывает избирательную гибель именно опухолевых клеток, еще малоизучены. Как видно из накопленных к настоящему времени данных, мы имеем лишь констатацию противоопухолевой активности I3C и пока не

ГЛАВА 8
Таблица 23 Сравнение активностей индол-3 карбинола и
тамоксифена
Индол 3 карбинол
Усиливает конверсию эстрона в «слабый» эстроген, 20НЕ1, обеспечивая профилактический и терапевтический эффект на эстроген-зависимые опухоли
Проявляет эффекты как на эстроген-зависимых, так и эстроген-независимых клетках
Ингибирует рост и вызывает апоптоз MCF7 клеток опухоли молочной железы на 90%
Защищает клетки от токсинов, диоксина (профилактика опухолей)
Восстанавливает р21 — супрес-сор опухолей
Вмешивается в пути, индуцируемые другими пролифератив-ными сигналами (с участием ростовых факторов, цитоплаз-матических киназ, активаторов транскрипции)
Вызывает апоптоз всех эстроген-зависимых, эстроген-независимых опухолевых клеток, блокирует и предупреждает рост опухолей других органов и тканей
Обеспечивает антиоксидантную защиту
Тамоксифен
Смещает равновесие 20НЕ1/16аОНЕ1 в пользу 16аОНЕ1
Работает только на эстроген-позитивных клетках опухоли молочной железы
Ингибирует рост и вызывает апоптоз MCF7 клеток опухоли молочной железы на 60%
Не обладает защитными свойствами против химически-инду-цированных опухолей
Вызывает мутации р21, р53
Не блокирует пролиферацию опухолевых клеток, чей рост не индуцируется эстрогеном
Индуцирует рост опухолей в эстроген-зависимых тканях (эндометрий)
Не обладает анти-оксидантной функцией

Обоснованные направления терапии и профилактики..
можем понять, как это вещество «отличает» опухолевые клетки от здоровых. Скорее всего, I3C лишь использует механизмы, которыми отличаются опухоли, постоянно живущие в условиях метаболического стресса. Эти°меха-низмы направлены на регуляцию пролиферативной активности опухолевой клетки, ее выживание в условиях стресса, поддержание относительно постоянного гомеостаза. А это, в свою очередь, достигается усилением транскрипционного контроля белковых факторов, ответственных за эти процессы. Так, например, в клетках молочной железы повышена экспрессия ростовых факторов и их мембран-но-связанных рецепторов (EGF, EGFR), факторов транскрипции (субъединиц NF-kB, SP-1, других активаторов транскрипции), ядерных рецепторов (в частности, рецепторов к эстрогенам), а также Ah-рецептора. I3C использует свой потенциал воздействия на сигнальные пути, индуцируемые указанными факторами, которых несравнимо больше в опухолевой, чем в здоровой клетке. Это, в свою очередь, приводит к кажущейся «избирательности» действия I3C, что на самом деле можно объяснить большей вероятностью взаимодействия. В пользу этого рассуждения говорит также тот факт, что потенциальными мишенями I3C являются, вообще говоря, все солидные опухоли, рост которых напрямую и не зависит от эстрогенов, и пролиферация которых проходит в эстроген-независимой манере (например, эстроген-независимые опухоли молочной железы, опухоли простаты). При этом никакой избирательности между разными типами опухолей I3C не проявляет. Просто там больше потенциальных объектов для взаимодействия. Другими словами, опухоль сама «предоставляет» множество объектов для проявления противоопухолевых активностей I3C.

ГЛАВА 8
Не менее интересным является свойство I3C предотвращать развитие опухолей, особенно когда речь идет о химически-индуцированном канцерогенезе. Вредное влияние окружающей среды вносит свой вклад в повышение риска мутагенности, нарушение метаболизма, сбоя защитных систем клетки, что немедленно сказывается на статистике смертей от опухолей в индустриально развитых странах. Мутагены, онкогены, циклические углеводороды, яды проникают в организм и накапливаются в тканях. Своеобразными «токсикологическими» воротами клетки служит Ah-рецептор, лигандами которого являются ариловые углеводороды. Долгое время не было известно ни одного природного лиганда для этого рецептора. Сейчас установлено, что один из них — индол-3-кар-бинол. Он служит селективным модулятором Ah-рецепто-ра, изменяет его функции в плане регуляции экспрессии эстроген-зависимых генов и обладает сильным превентивным эффектом от активации Ah его химическими лигандами, такими, как, например, диоксин.

Обоснованные направления терапии и профилактики...
С п и с о к литературы к Главе 8
1. Afaq F., Adhami V.M., Ahmad N., Mukhtar H. Inhibition of ultraviolet B-mediated activation of nuclear factor kappaB in normal human epidermal keratinocytes by green tea Constituent (-)-epigallocatechin-3-gallate. Oncogene, 2003, Feb 20, 22(7), 1035-44.
2. Ahmed S., Wang N., Lalonde M., Goldberg V.M., Haqqi T.M. Green tea polyphenol epigallocatechin-3-gallate (EGCG) differentially inhibits inter-leukin-1 beta-induced expression of matrix metalloproteinase-1 and -13 in human chondrocytes. J Pharmacol Exp Ther. 2004, Feb, 308(2), 767-73.
3. Ahn W.S., Yoo J., Huh S.W., Kim C.K., Lee J.M., Namkoong S.E., Bae S.M., Lee IP. Protective effects of green tea extracts (polyphenon E and EGCG) on human cervical lesions. Eur J Cancer Prev, 2003, Oct, 12(5), 383-90.
4. Altaras M.M., Aviram Ft., Beyth Y. Tamoxifen-associated endometrial carcinoma in postmenopausal breast cancer patients. Gynecol Oncol, 1991, 41,3,270.
5. Auborn K.J., Fan S., Rosen E.M., Goodwin L, Chandraskaren A., Williams D.E., Chen D., Carter Т.Н. lndole-3-carbinol is a negative regulator of estrogen. J Nutr, 2003, Jul, 133(7 Suppl), 2470S-2475S.
6. August D.A., Landau J., Caputo D., Hong J., Lee M.J., Yang C.S. Ingestion of green tea rapidly decreases prostaglandin E2 levels in rectal mucosa in humans. Cancer Epidemiol, 1999, 8, 709-713.
7. Bertolini F., Fusetti L, Rabascio C, Cineri S., Martinellil G., Pruneri G. Inhibition of angiogenesis and induction of endothelial and tumor cell apoptosis by green tea in animal models of human high-grade non-Hodgkin's lymphoma. Leukemia, 2000,14,1477-1482.
8. Blanc C, Deveraux Q.L., Krajewski S., Janicke R.U., Porter A.G., Reed J.C., Jaggi R., Marti A. Caspase-3 is essential for procaspase-9 processing and cisplatin-induced apoptosis of MCF-7 breast cancer cells. Cancer Res, 2000, Aug 15, 60(16), 4386-90.
9. Bradlow H I . , Telang NT., Sepkovic D.W., Osborne M.P. 2-hydroxy-estrone, the 'good' estrogen. J Endocrinol, 1996, Sep, 150 Suppl, S259-65.
10. Braithwaite R.S., Chlebowski R.T., Lau J., George S., Hess R., Col N.F. Meta-analysis of vascular and neoplastic events associated with tamoxifen. J Gen Intern Med, 2003, Nov, 18(11), 937-47.

ГЛАВА 8
11. Brandi G., Paiardini M., Cervasi В., Fiorucci C, Filippone P., De Marco C, Zaffaroni N.. Magnani M. A new indole-3-carbinol tetrameric derivative inhibits cyclin-dependent kinase 6 expression, and induces G1 cell cycle arrest in both estrogen-dependent and estrogen-independent breast cancer cell lines. Cancer Res, 2003, Jul 15, 63(14), 4028-36.
12. Brignall M.S. Prevention and treatment of cancer with indole-3-carbi-nol. Altern Med Rev, 2001, Dec, 6(6), 580-9.
13. Calle E.E, Kaaks R. Overweight, obesity and cancer, epidemiological evidence and proposed mechanisms. Nat Rev Cancer, 2004, Aug, 4(8), 579-91.
14. Cao Y., Cao R. Angiogenesis inhibited by drinking tea. Nature, 1999, 398,381.
15. Cardoso F., Di Leo A., Larsimont D., Gancberg D., Rouas G., Dolci S., Ferreira F., Paesmans M., Piccart M. Evaluation of HER2, p53, bcl-2, to-poisomerase ll-alpha, heat shock proteins 27 and 70 in primary breast cancer and metastatic ipsilateral axillary lymph nodes. Ann Oncol, 2001, May, 12(5), 615-20.
16. Chakrabarti O., Veeraraghavalu K., Tergaonkar V., Liu Y., Androphy E.J., Stanley M.A., Krishna S. Human papillomavirus type 16 E6 amino acid 83 variants enhance E6-mediated МАРК signaling and differentially regulate tumorigenesis by notch signaling and oncogenic Ras. J Virol, 2004, Jun, 78(11), 5934-45.
17. Chantre P., Lairon D. Recent findings of green tea extract AR25 (Exolise) and its activity for the treatment of obesity. Phytomedicine, 2002, Jan, 9(1), 3-8.
18. Chatterji LL, Riby J.E., Taniguchi Т., Bjeldanes E.L., Bjeldanes LF, Firestone GL lndole-3-carbinol stimulates transcription of the interferon gamma receptor 1 gene and augments interferon responsiveness in human breast cancer cells. Carcinogenesis, 2004, Feb 26.
19. Chen P., Yang C.C., Chen Y.J., Wang P.H. Tamoxifen-induced endometrial cancer. Eur J Gynaecol Oncol, 2003, 24(2), 135-137.
20. Chinni S.R., Sarkar F.H. Akt inactivation is a key event in indole-3-carbinol-induced apoptosis in PC-3 cells. Clin Cancer Res, 2002, Apr, 8(4), 1228-36.
21. Chung J.H., Han J.H., Hwang E.J., Seo J.Y., Cho K.H., Kim K.H., Youn J.I., Eun H.C. Dual mechanisms of green tea extract (EGCG)-induced

Обоснованные направления терапии и профилактики...
cell survival in human epidermal keratinocytes. FASEB J, 2003, Oct, 17(13), 1913-5.
22. Ciocca D.R., Luque E.H. Immunological evidence for the identity between the hsp27 estrogen-regulated heat shock protein and the p29 estrogen receptor-associated protein in breast and endometrial cancer. Breast Cancer Res Treat, 1991, 20(1), 33-42.
23. Coll D.A., Rosen C.A., Auborn K„ Potsic W.P., Bradlow H i . Treatment of recurrent respiratory papillomatosis with indole-3-carbinol. Am J Otolaryngol, 1997, Jul-Aug, 18(4), 283-5.
24. Couillard S., Gutman M., Labrie C, Belanger A., Candas В., Labrie F. Comparison of the effects of the antiestrogens EM-800 and tamoxifen on the growth of human breast ZR-75-1 cancer xenografts in nude mice. Cancer Res, 1998, Jan 1, 58(1), 60-4.
25. Cover СМ., Hsieh S.J., Cram E.J., Hong C, Riby J.E., Bjeldanes L.F., Firestone G.L. lndole-3-carbinol and tamoxifen cooperate to arrest the cell cycle of MCF-7 human breast cancer cells. Cancer Res, 1999, Mar 15, 59(6), 1244-51.
26. Cram E.J., Liu B.D., Bjeldanes L.F., Firestone G.L. lndole-3-carbinol inhibits CDK6 expression in human MCF-7 breast cancer cells by disrupting Sp1 transcription factor interactions with a composite element in the CDK6 gene promoter. J Biol Chem, 2001, Jun, 22, 276(25), 22332-40.
27. Crew K.D., Neugut A.I. Epidemiology of upper gastrointestinal malignancies. Semin Oncol, 2004, Aug, 31(4), 450-64.
28. Curtis R.E., Freedman D.M., Sherman M.E., Fraumeni J.F. Jr. Risk of malignant mixed mullerian tumors after tamoxifen therapy for breast cancer. J Natl Cancer Inst, 2004, 96,1, 70-74.
29. Dalton T.P., Shertzer H.G., Puga A. Regulation of gene expression by reactive oxygen. Annu Rev Pharmacol Toxicol, 1999, 39, 67-101.
30. Detmar M. Evidence for vascular endothelial growth factor (VEGF) as a modifier gene in psoriasis. J Invest Dermatol, 2004, Jan, 122(1), XIV-XV.
31. Firestone G.L., Bjeldanes L.F. lndole-3-carbinol and З-З'-diindolyl-methane antiproliferative signaling pathways control cell-cycle gene transcription in human breast cancer cells by regulating promoter-Sp1 transcription factor interactions. J Nutr, 2003, Jul, 133(7 Suppl), 2448S-2455S.

Обоснованные направления терапии и профилактики...
42. Hegarty V.M., May H.M., Khaw K.T. Tea drinking and bone mineral density in older women. Am J Clin Nutr, 2000, 71,1003-1007.
43. Hong J., Smith T.J., Но СТ., August D.A., Yang C.S. Effects of purified green and black tea polyphenols on cyclooxygenase- and lipoxyge-nase-dependent metabolism of arachidonic acid in human colon mucosa and colon tumor tissues. Biochem Pharmacol, 2001, 62,1175-1183.
44. Houghton J.P, loffe O.B., Silverberg S.G., McGrady B, McClugga-ge W.G. Metastatic breast lobular carcinoma involving tamoxifen-associated endometrial polyps: report of two cases and review of tamoxifen-associated polypoid uterine lesions. Mod Pathol, 2003, Apr, 16(4), 395-8.
45. Howells L.M., Gallacher-Horley В., Houghton C.E., Manson M.M., Hudson E.A. lndole-3-carbinol inhibits protein kinase B/Akt and induces apo-ptosis in the human breast tumor cell line MDA MB468 but not in the nontu-morigenic HBL100 line. Mol Cancer Ther, 2002, Nov, 1(13), 1161-72.
46. Huang S., New L, Pan Z., Han J., Nemerow G.R. Urokinase plasminogen activator/urokinase-specific surface receptor expression and matrix invasion by breast cancer cells requires constitutive p38alpha mitogen-acti-vated protein kinase activity. J Biol Chem, 2000, 275,12266-12272.
47. Jankun J., Keck R.W., Skrzypczak-Jankun E., Swiercz R. Inhibitors of urokinase reduce size of prostate cancer xenografts in severe combined immunodeficient mice. Cancer Res, 1997a, 57, 559-563.
48. Jankun J., Selman S.H., Swiercz R., Skrzypczak-Jankun E. Why drinking green tea could prevent cancer. Nature, 1997b, 387, 561.
49. Jeong W.S., Kim I.W., Hu R., Kong A.N. Modulatory properties of various natural chemopreventive agents on the activation of NF-kappaB signaling pathway. Pharm Res, 2004, Apr, 21(4), 661-70.
50. Jung Y.D., Ellis LM. Inhibition of tumour invasion and angiogenesis by epigallocatechin gallate (EGCG), a major component of green tea. Int J Exp Pathol, 2001, Dec, 82(6), 309-16.
51. Jung Y.D., Kim M.S., Shin B.A., Chay K.O., Ahn B.W., Liu W., Buca-na CD., Gallick G.E., Ellis LM. EGCG, a major component of green tea, inhibits tumour growth by inhibiting VEGF induction in human colon carcinoma cells. Br J Cancer, 2001, Mar, 23, 84(6), 844-50.

ГЛАВА 8
32. Friedrich К., Wieder Т., Von Haefen С, Radetzki S., Janicke R., Schulze-Osthoff K., Dorken В., Daniel P.T. Overexpression of caspase-3 restores sensitivity for drug-induced apoptosis in breast cancer cell lines with acquired drug resistance. Oncogene, 2001, May, 17, 20(22), 2749-60.
33. Furberg A.S., Veierod M.B., Wilsgaard Т., Bernstein L, Thune I. Serum high-density lipoprotein cholesterol, metabolic profile, and breast cancer risk. J Natl Cancer Inst, 2004, Aug, 4, 96(15), 1152-60.
34. Garbisa S., Sartor L, Biggin S, Salvato В., Benelli R., Albini A. Tumor gelatinases and invasion inhibited by the green tea flavanol epigalloca-techin-3-gallate. Cancer, 2001, 91, 822-832.
35. Garrido C, Schmitt E., Cande C, Vahsen N., Parcellier A., Kroemer G. HSP27 and HSP70: potentially oncogenic apoptosis inhibitors. Cell Cycle, 2003, Nov-Dec, 2(6), 579-84.
36. Gohji K., Nakajima M., Boyd D, Dinney СР., Bucana CD., Kitaza-na S., Kamidono S., Fidler I.J. Orgap-site dependence for the production of urokinase-type plasminogen activator and metastasis by human renal cell carcinoma cells. Am J Pathol, 1997, Dec, 151(6), 1655-61.
37. Golijanin D., Tan J.Y, Kazior A., Cohen E.G., Russo P., Dalbagni G., Auborn K.J., Subbaramaiah K., Dannenberg A.J. Cyclooxygenase-2 and microsomal prostaglandin E synthase-1 are overexpressed in squamous cell carcinoma of the penis. Clin Cancer Res, 2004, Feb, 1,10(3), 1024-31.
38. Goodin M.G., Rosengren R.J. Epigallocatechin gallate modulates CYP450 isoforms in the female Swiss-Webster mouse.Toxicol Sci, 2003, Dec, 76(2), 262-70.
39. Gottardis M.M, Jordan V.C Development of tamoxifen-stimulated growth of MCF-7 tumors in athymic mice after long-term antiestrogen administration. Cancer Res, 1988, 48, 5183-5187.
40. Hamada S., Ooshima Т., Fijiwara Т., Minami Т., Kimura S. Development of preventive measures based on the aetiology of dental caries-a review. Microb Ecol Health Dis, 1996, 9, 349-357.
41. Hashiya N., Jo N., Aoki M., Matsumoto K., Nakamura Т., Sato Y., Ogata N., Ogihara Т., Kaneda Y., Morishita R. In vivo evidence of angiogen-esis induced by transcription factor Ets-1, Ets-1 is located upstream of angio-genesis cascade. Circulation, 2004, Jun, 22,109(24), 3035-41.

Обоснованные направления терапии и профилактики...
62. Lin J., Zhang S.M., Cook N.R., Rexrode K.M., Lee I.M., Buring J.E. Body mass index and risk of colorectal cancer in women (United States). Cancer Causes Control, 2004, Aug, 15(6), 581-9.
63. Lord R.S., Bongiovanni В., Bralley J.A. Estrogen metabolism and the diet-cancer connection, rationale for assessing the ratio of urinary hydroxyla-ted estrogen metabolites. Altern Med Rev, 2002 Apr, 7(2), 112-29.
64. Maeda-Yamamoto M., Kawahara H., Tahara N., Tsuju K., Hara Y., Isemura M. Effects of tea polyphenols on the invasion and matrix metallopro-teinases activities of human fibrosarcoma HT1080 cells. J Agric Food Chem, 1999,47,2350-2354.
65. Makimura M., Hirasawa M., Kobayashi K., Indo J., Sakanaka S., Ta-guchi Т., Otake S. Inhibitory effect of tea catechins on collagenase activity. J Periodontal, 1993, Jul, 64(7), 630-6.
66. Mandel S., Weinreb O., Amit Т., Youdim M.B. Cell signaling pathways in the neuroprotective actions of the green tea polyphenol (-)-epigalloca-techin-3-gallate, implications for neurodegenerative diseases. J Neurochem, 2004, Mar, 88(6), 1555-69.
67. Martinez-Arribas F., Nunez-Villar M.J., Lucas A.R., Sanchez J., Tej-erina A., Schneider J. Immunofluorometric study of Bcl-2 and Bax expression in clinical fresh tumor samples from breast cancer patients. Anticancer Res, 2003, Jan-Feb, 23(1 B), 565-8.
68. Mittal A., Piyathilake C, Hara Y., Katiyar S.K. Exceptionally high protection of photocarcinogenesis by topical application of (-)-epigallocatechin-3-gallate in hydrophilic cream in SKH-1 hairless mouse model, relationship to inhibition of UVB-induced global DNA hypomethylation. Neoplasia, 2003, 5, 6, 555-65.
69. Mutoh M., Takahashi M., Fukuda K., Matsushima-Hibiya Y., Mutoh H., Sugimura Т., Wakabayashi K. Suppression of cyclooxygenase-2 promoter-dependent transcriptional activity in colon cancer cells by chemopreventi-ve agents with a resorcin-type structure. Carcinogenesis, 2000, 21, 959-963.
70. Nair P., Nair K.M., Jayaprakash P.G., Pillai M.R. Decreased programmed cell death in the uterine cervix associated with high risk human papillomavirus infection. Pathol Oncol Res, 1999, 5, 2, 95-103.
71. Nakagawa Т., Yokozawa Т., Sano M., Takeuchi S., Kim M., Minamo-to S. Activity of (-)-Epigallocatechin 3-O-Gallate against Oxidative Stress in

ГЛАВА 8
52. Katiyar S.K., Bergamo B.M., Vyalil P.K., Elmets C.A. Green tea polyphenols, DNA photodamage and photoimmunology. J Photochem Photo-biolB, 2001, Dec, 31, 65(2-3), 109-14.
53. Katiyar S.K., Mukhtar H. Green tea polyphenol (-)-epigallocatechin-3-gallate treatment to mouse skin prevents UVB-induced infiltration of leukocytes, depletion of antigen-presenting cells, and oxidative stress. J Leukoc Biol, 2001, May, 69(5), 719-26.
54. Katiyar S.K. Skin photoprotection by green tea, antioxidant and immunomodulatory effects. Curr Drug Targets Immune Endocr Metabol Disord, 2003, Sep, 3(3), 234-42.
55. Kim J., Hwang J.S., Cho Y.K., Han Y., Jeon Y.J., Yang K.H. Protective effects of (-)-epigallocatechin-3-gallate on UVA- and UVB-induced skin damage. Skin Pharmacol Appl Skin Physiol, 2001, Jan-Feb, 14(1), 11-9.
56. Kim M.H., Jung M A , Hwang Y.S., Jeong M., Kim S.M., Ahn S.J., Shin B.A., Ahn B.W., Jung YD. Regulation of urokinase plasminogen activator by epigallocatechin-3-gallate in human fibrosarcoma cells. Eur J Pharmacol, 2004, Mar 8, 487(1-3), 1-6.
57. Lee I.J., Han F., Baek J., Hisatsune A., Kim K.C. Inhibition of MUC1 expression by indole-3-carbinol. Int J Cancer, 2004, May 10,109(6), 810-6.
58. Legha S.S. Tamoxifen in the treatment of breast cancer. Ann Intern Med, 1988,109,219-228.
59. Li H., Lindenmeyer F., Grenet C, Opolon P., Menashi S., Soria C, Yeh P., Perricaudet M., Lu H. AdTIMP-2 inhibits tumor growth, angiogenesis, and metastasis, and prolongs survival in mice. Hum Gene Ther, 2001, Mar 20,12(5), 515-26.
60. Liang Y.C., Huang Y., Tsai S., Lin-Shiau S., Chen C, Lin J. Suppression of inducible cyclooxygenase and inducible nitric oxide synthase by api-genin and related flavonoids in mouse macrophages. Carcinogenesis, 1999a, 20, 1945-1952.
61. Liang Y.C., Lin-Shiau S.Y., Chen C.F., Lin J.K. Inhibition of cyclin-de-pendent kinases 2 and 4 activities as well as induction of Cdk inhibitors p21 and p27 during growth arrest of human breast carcinoma cells by epigalloca-techin- 3-gallate. J Cell Biochem, 1999b, 75,1-12.

О б о с н о в а н н ы е н а п р а в л е н и я т е р а п и и и п р о ф и л а к т и к и . . .
tamoxifen associated endometrial carcinomas. Anticancer Res, 1998, Nov-Dec, 18(6B), 4661-5.
81. Riethdorf L, Riethdorf S., Petersen S., Bauer M., Herbst H., Janic-ke F., Loning T. Urokinase gene expression indicates early invasive growth in squamous cell lesions of the uterine cervix. J Pathol, 1999, Oct, 189(2), 245-50.
82. Sakanaka S., Okada Y. Inhibitory effects of green tea polyphenols on the production of a virulence factor of the periodontal-disease-causing anaerobic bacterium Porphyromonas gingivalis. J Agric Food Chem, 2004, 52(6), 1688-92.
83. Sakata R., Ueno Т., Nakamura Т., Sakamoto M., Torimura Т., Sata M. Green tea polyphenol epigallocatechin-3-gallate inhibits platelet-derived growth factor-induced proliferation of human hepatic stellate cell line LI90. J Hepatol, 2004, Jan, 40(1), 52-9.
84. Sarkar F.H., Rahman K.M., Li Y. Bax translocation to mitochondria is an important event in inducing apoptotic cell death by indole-3-carbinol (I3C) treatment of breast cancer cells. J Nutr, 2003,133, 7, 2434-2439.
85. Sayama K., Lin S., Zheng G., Oguni I. Effects of green tea on growth, food utilization and lipid metabolism in mice. In Vivo, 2000, Jul-Aug, 14(4), 481-4.
86. Shaheen R.M., Davis D.W., Liu W, Zebrowski B.K., Wilson M.R., Bucana CD., McConkey D.J., McMahon G., Ellis LM. Antiangiogenic therapy targeting the tyrosine kinase receptor for vascular endothelial growth factor receptor inhibits the growth of colon cancer liver metastasis and induces tumor and endothelial cell apoptosis. Cancer Res, 1999, Nov 1, 59(21), 5412-6.
87. Singh A.K, Seth P., Anthony P., Husain M.M., Madhavan S., Mukhtar H, Maheshwari R.K. Green tea constituent epigallocatechin-3-gal-late inhibits angiogenic differentiation of human endothelial cells. Arch Biochem Biophys, 2002, May 1, 401(1), 29-37.
88. Stanley M. Chapter 17, Genital human papillomavirus infections-current and prospective therapies. J Natl Cancer Inst Monogr, 2003, 31,117-24.
89. Taioli E., Garte S.J., Trachman J., Garbers S., Sepkovic D.W., Osborne M.P., Mehl S., Bradlow H I . Ethnic differences in estrogen metabolism in healthy women. J Natl Cancer Inst, 1996, May 1, 88(9), 617.

Г Л А В А 8
Rats with Adenine-lnduced Renal Failure. J Agric Food Chem, 2004, Apr 7, 52(7), 2103-2107.
72. Osborne C.K., Coronado-Heinsohn E.B., Hilsenbeck S.G., McCue B.L., Wakeling A.E., McClelland R.A., Manning D.L, Nicholson R.I. Comparison of the effects of a pure steroidal antiestrogen with those of tamoxifen in a model of human breast cancer. J Natl Cancer Inst, 1995, May 17, 87(10), 746-50.
73. Park J.W, Choi Y.J., Suh S.I., Kwon Т.К. Involvement of ERK and protein tyrosine phosphatase signaling pathways in EGCG-induced cycloox-ygenase-2 expression in Raw 264.7 cell. Biochem Biophys Res Commun, 2001,286,721-725.
74. Parkin D.R., Malejka-Giganti D. Differences in the hepatic P450-de-pendent metabolism of estrogen and tamoxifen in response to treatment of rats with 3,3'-diindolylmethane and its parent compound indole-3-carbinol. Cancer Detect Prev, 2004, 28(1), 72-9.
75. Peitsaro P., Ruutu M., Syrjanen S., Johansson B. Divergent expression changes of telomerase and E6/E7 mRNA, following integration of human papillomavirus type 33 in cultured epithelial cells. Scand J Infect Dis, 2004, 36(4), 302-4.
76. Plug-DeMaggio A.W., Sundsvold Т., Wurscher M.A, Koop J.I., Klin-gelhutz A.J., McDougall J.K. Telomere erosion and chromosomal instability in cells expressing the HPV oncogene 16E6. Oncogene, 2004, Apr 29, 23(20), 3561-71.
77. Porter W., Wang F., Duan R., Qin C, Castro-Rivera E., Safe S. Transcriptional activation of heat shock protein 27 gene expression by 17b-estradiol and modulation by antiestrogens and aryl hydrocarbon receptor agonists, estrogenic activity of IC1164,384. J Mol Endocrinol, 2000, 26, 31-42.
78. Rahman K.M., Aranha O., Sarkar F.H. lndole-3-carbinol (I3C) induces apoptosis in tumorigenic but not in nontumorigenic breast epithelial cells. Nutr Cancer, 2003, 45, 1,101-112.
79. Ramirez-Mares M.V., de Mejia E.G. Comparative study of the antioxidant effect of ardisin and epigallocatechin gallate in rat hepatocytes exposed to benomyl and 1-nitropyrene. Food Chem Toxicol, 2003, Nov, 41(11), 1527-35.
80. Ramondetta L.M., Palazzo J.P., Dunton C.J., Kovatich A.J., Carlson J.A. A comparative analysis of Ki-67, p53, and p21 WAF1CIP1 expression in

Обоснованные направления терапии и профилактики...
100. Wilkinson M.G., Millar J.B.A. Control of the eukaryotic cell cycle by MAP kinase signaling pathways. FASEB J, 2000,14, 2147- 2157.
101. Yamaguchi M., Jie Z. Effect of polyphenols on calcium content and alkaline phosphatase activity in rat femoral tissues in vitro. Biol Pharm Bull, 2001, Dec, 24(12), 1437-9.
102. Yang C.S., Landau J.M. Effects of tea consumption on nutrition and health. J Nutr, 2000, Oct, 130(10), 2409-12.
103. Yokoyama M., Noguchi M., Nakao Y., Pater A., Iwasaka T. The tea polyphenol, (-)-epigallocatechin gallate effects on growth, apoptosis, and telo-merase activity in cervical cell lines. Gynecol Oncol, 2004, Jan, 92(1), 197-204.
104. Young H.S., Summers A.M., Bhushan M., Brenchley P.E., Griffiths C.E. Single-nucleotide polymorphisms of vascular endothelial growth factor in psoriasis of early onset. J Invest Dermatol, 2004, Jan, 122(1), 209-15.
105. Zheng G., Sayama K., Okubo Т., Juneja L.R., Oguni I. Anti-obesity effects of three major components of green tea, catechins, caffeine and the-anine, in mice. In Vivo, 2004, Jan-Feb, 18(1), 55-62.
106. Zheng Y, Song H.J, Kim C.H., Kim H.S, Kim E.G., Sachinidis A, Ahn H.Y. Inhibitory effect of epigallocatechin 3-O-gallate on vascular smooth muscle cell hypertrophy induced by angiotensin II. J Cardiovasc Pharmacol, 2004, Feb 43(2), 200-8.

ГЛАВА 8
90. Talvensaari A., Apaja-Sarkkinen M., Hoyhtya M., Westerlund A., Pu-istola U., Turpeenniemi Т. Matrix metalloproteinase 2 immunoreactive protein appears early in cervical epithelial dedifferentiation. Gynecol Oncol, 1999, Mar, 72(3), 306-11.
91. Tang F.Y., Nguyen N., Meydani M. Green tea catechins inhibit VEGF-induced angiogenesis in vitro through suppression of VE-cadherin phosphorylation and inactivation of Akt molecule. Int J Cancer, 2003, Oct 10, 106(6), 871-8.
92. Tarani A., Negri E., LaVecchia C. Food and nutrient intake and risk of cataract. Ann Epidemiol, 1996, 6, 41-46.
93. Toussaint-Smith E., Donner D.B., Roman A. Expression of human papillomavirus type 16 E6 and E7 oncoproteins in primary foreskin keratino-cytes is sufficient to alter the expression of angiogenic factors. Oncogene, 2004, Apr 15, 23(17), 2988-95.
94. Turner M.A., Palefsky J.M. Urokinase plasminogen activator expression by primary and HPV 16-transformed keratinocytes. Clin Exp Metastasis, 1995, Jul, 13(4), 260-8.
95. Vayalil P.K., Elmets С A., Katiyar S.K. Treatment of green tea polyphenols in hydrophilic cream prevents UVB-induced oxidation of lipids and proteins, depletion of antioxidant enzymes and phosphorylation of МАРК proteins in SKH-1 hairless mouse skin. Carcinogenesis, 2003, May, 24(5), 927-36.
96. Vayalil P.K., Katiyar S.K. Treatment of epigallocatechin-3-gallate inhibits matrix metalloproteinases-2 and -9 via inhibition of activation of mito-gen-activated protein kinases, c-jun and NF-kappaB in human prostate carcinoma DU-145 cells. Prostate, 2004, Apr 1, 59(1), 33-42.
97. Vitaglione P., Fogliano V. Use of antioxidants to minimize the human health risk associated to mutagenic/carcinogenic heterocyclic amines in food. J Chromatogr В Analyt Technol Biomed Life Sci, 2004, Mar 25,802(1), 189-99.
98. Wei Y.H., Lee H.C. Oxidative stress, mitochondrial DNA mutation, and impairment of antioxidant enzymes in aging. Exp Biol Med (Maywood), 2002, Oct, 227(9), 671-82.
99. Wheeler D.S., Catravas J.D., Odoms K., Denenberg A., Malhotra V., Wong H.R. Epigallocatechin-3-gallate, a green tea-derived polyphenol, inhibits IL-1 beta-dependent proinflammatory signal transduction in cultured respiratory epithelial cells. J Nutr, 2004, May, 134(5), 1039-44.

ЗПКЛЮЧЕНИЕ
В этиологии в течение долгого времени доминировал принцип монокаузальности. Этому способствовали открытия в микробиологии и вирусологии, которые наложили свой отпечаток в трактовке причин и неинфекционных заболеваний. Вот уже более века в онкологии предпринимаются попытки сформировать некую, четко очерченную (обозначенную) причину развития опухолевого процесса. Этот этап в онкологической науке, безусловно, важен, так как позволил выделить из множества причин наиболее значимые и близкие к вероятным. Уже сегодня ясно, что причины, ведущие к развитию злокачественного процесса, многофакторны и разнообразны при разных локализациях.
Человеческая природа такова, что при попытке разгадать какую-либо научную тайну, в первую очередь, формируются гипотезы, в основе которых лежит принцип универсальной модели [теория хронического раздражения (Virchow R., 1839); вирусная теория (Borrel A., 1903; Мечников И.И., 1909; Rous P.,1911,1936); мутационная теория (Bovery Th., 1914); теория химического канцерогенеза (Yamagiwa К., Ichikawa К., 1918)]. В истории онкологии они сыграли неоценимую роль, дав мощный стимул научного поиска для многих поколений исследователей. Чуть позже стало ясно, как трудно в один универсальный механизм вместить все разнообразие вариантов, характеризующих канцерогенез.

ЗАКЛЮЧЕНИЕ
На смену универсальным теориям пришли теории, объединяющие самые разнообразные этиологические факторы [регенарационно-мутационная теория (Фишер-Вазельс В., 1936); теория наследственной передачи рака, генетические факторы и факторы окружающей среды (Bittner J., 1936, Loeb L, 1940); теория комплексной этиологии рака (Southman С, 1963); клонально-селекционная теория рака (PrehnR., 1964); вирусо-генетическая теория (Зильбер Л.А., 1945,1963); полиэтиологическая теория (Н.Н. Петров, 1947)].
И сегодня следует признать, что универсальной этиологической концепции, объединяющей весь спектр злокачественных опухолей, нет. С учетом знаний и пониманий, к которым пришла онкология к началу XXI века, в проблеме канцерогенеза следует выделить три основных последовательных этапа:
- этап трансформации (превращение нормальных клеток в опухолевые, наследственно передающееся последующим популяциям клеток);
- этап некотролируемого размножения трансформированных клеток;
- этап прогрессии (индивидуальное развитие опухоли путем устойчивых необратимых качественных и количественных изменений одного или нескольких признаков [FouldsL.,1969]).
Условия и исходные факторы, определяющие развитие каждого этапа, различны и разнообразны, что изначально исключает универсальность и монокаузальность канцерогенеза. Эта точка зрения разделяется не всеми исследователями, часть которых вкладывают в понятие канцерогенеза лишь этап трансформации — превращение нормальной клетки в опухолевую. На первый взгляд,

ЗАКЛЮЧЕНИЕ
подобный подход к трактовке канцерогенеза весьма близок к четкому обозначению причинно-следственных связей, и, вследствие этого, понятен широкому кругу специалистов. Согласиться с тем, что для сформированной опухолевой клетки необходим еще ряд условий, без которых нет продолжения злокачественного прогресса, достаточно сложно и обременительно, так как требует учета целого ряда факторов, причем, весьма разнообразных даже в рамках одной нозологии.
Приступая к данной работе, авторы изначально определили свою позицию по данному вопросу. Итак, канцерогенез — биологический процесс, в основе которого многоэтапное^ и необратимость. Опираясь на этот постулат, мы постарались обозначить основные моменты, определяющие современное понимание некоторых звеньев канцерогенеза рака репродуктивных органов. Вполне понятно, что сегодня нет возможности представить исчерпывающую картину по всем трем направлениям (этапам) канцерогенеза. Вместе с тем, при всем многообразии фактов в экспериментальной и клинической онкологии проблема рака репродуктивных органов несколько выделяется вследствие ряда весьма общих моментов, формирующих некий принцип закономерности. Благодаря некоторой схожести ряда параметров, характеризующих общий (биологический) фон онкологического больного, раньше, чем при других локализациях, выделены факторы и группы риска именно для больных раком репродуктивных органов. Тем самым исследователи изначально обозначили общую патогенетическую базу, предопределяющую высокий риск канцерогенеза некоторых органов-мишеней у женщин. Развивая эту точку зрения, мы достаточно четко и аргументированно можем сказать,

ЗАКЛЮЧЕНИЕ
что наиболее значимыми факторами являются метаболический синдром и постменопауза.
При метаболическом синдроме интенсивно развиваются многчисленные пути внеяичникового (и яичникового — тека-ткань) стероидогенеза, обеспечивая высокий уровень продукции агрессивных метаболитов (16аОН). Комплекс метаболит-рецептор достаточно прочен, что и обеспечивает пролонгированный пролиферативный эффект.
Постменопауза — период жизни женщины, когда происходит весьма качественная и резкая перестройка в различных звеньях гомеостаза (энергетическом, обменном, гормональном, иммунологическом и т.д.). Немаловажно и то, что в постменопаузе идет резкое увеличение общего объема соединительной ткани (как в норме, так и в процессе репарации) и снижение эпителиального компонента многих органов и тканей. В подобной ситуации основные компоненты пролиферации могут находиться в состоянии напряжения или даже усиливаться (как эффект в рамках компенсаторного ответа). Кроме того, во всех гормоноза-висимых органах имеет место резкое изменение микробиоценоза с усилением патологического влияния вирусного и условнопатогенного компонентов.
Действительно, онкологическая наука и практика в течение обозримой истории были нацелены (в диагностическом и лечебном плане) на опухолевый процесс, который четко обозначен и клинически и морфологически. Математическое выражение этого тезиса означает, что первичная опухоль достигла уровня, как минимум, 109 опухолевых клеток, а интересы клинической онкологии вообще ограничены рамками диапазона 109-101 3 опухолевых клеток. Следует признать, что, несмотря на значительные успехи, достигнутые в диагностическом и лечебном разде-

ЗАКЛЮЧЕНИЕ
лах онкологической науки, средние показатели смертности практически не изменились. Иными словами, лечебные пособия, какими бы современными они ни были, не способны существенно изменить отдаленные результаты излечения опухолевого процесса в рамках диапазона 109-10 1 3 опухолевых клеток. Из этого вытекает необходимость переориентации усилий с клинической онкологии на доклинический или ранний клинический этап злокачественного процесса. Попытка осмыслить эту задачу стала основной причиной появления этой книги.
Таким образом, начало и развитие злокачественной опухоли — это многофакторный и многоэтапный процесс, формирующийся в течение длительного времени. Не понимая и не представляя себе пусковых механизмов этого процесса, мы не можем строить сколько-нибудь эффективные программы профилактики и современной диагностики. Сегодня единственно возможным путем развития онкологии как науки является сдвиг интересов в сторону молекулярных основ онкогенеза.
Вот основные моменты, проясняющие в некоторой степени феномен роста онкологической заболеваемости ряда локализации у женщин после 50 лет. Представленная в книге точка зрения, ряд положений, трактовка некоторых результатов отражают взгляды авторов и могут стать предметом дискуссий. Мы будем искренне благодарны читателям за критические замечания и конструктивные предложения.

СОДЕРЖАНИЕ
Введение стр. 3
Глава 1. Эпидемиология рака репродуктивных
органов стр. 5
- список литературы к Главе 1
Глава 2.
Эндокринологические особенности постменопаузы в аспекте канцерогенеза стр. 12
- Варианты гормонального гомеостаза в постменопаузе
- Эстроген-рецепторы (ER) в некоторых тканях-мишенях (молочная железа, вульва, эндометрий) у пациенток постменопаузального периода
- список литературы к Главе 2
Глава 3.
Метаболиты эстрадиола и патологические
процессы в репродуктивной системе стр. 41
- Эстрогены: структура, место синтеза, роль в
физиологии и патофизиологии (канцерогенезе)
- Биосинтез эстрогенов и прогестерона
- Два типа эстроген-индуцированного канцерогенеза
- Метаболизм эстрадиола
- Роль метаболитов эстрогенов в канцерогенезе
репродуктивных органов
- Что такое «канкрофилия» и «метаболический
синдром», или «почему гормон-зависимые опухоли
у женщин чаще возникают в постменопаузе?
- Фармакологическая коррекция гормонального
баланса и профилактика образования гормон-
зависимых опухолей
- список литературы к Главе 3

СОДЕРЖАНИЕ
Глава 4. Метаболический синдром как маркер внегонадного стероидогенеза в постменопаузе стр. 77
- список литературы к Главе 4
Глава 5. Молекулярные факторы канцерогенеза стр. 93
- Как нормальная клетка становится раковой - Молекулярные механизмы активации клеточного
деления - Как клетка воспринимает сигналы, стимулирующие её
деление - Молекулярные механизмы развития опухолей
молочной железы - список литературы к Главе 5
Глава 6. Инфекционные агенты и канцерогенез стр. 103
- Папилломовирусная инфекция при гинекологическом раке
- Герпесвирусная инфекция при злокачественных эпителиальных опухолях яичников
- список литературы к Главе б
Глава 7. Ранний рак и предрак шейки матки, обусловленные вирусами папилломы человека стр. 131
- Роль ВПЧ - инфекции в развитии неопластических процессов шейки матки
- Механизмы канцерогенного действия ВПЧ - Иммуносупрессорные функции онкобелка Е7 - Роль гормональных факторов в развитии рака
шейки матки - список литературы к Главе 7

СОДЕРЖАНИЕ
Глава 8.
Обоснованные направления терапии и профилактики
рака репродуктивных органов стр. 151
- Катехины зеленого чая. Эпигаллокатехин-3-галлат
- Модуляция внутриклеточных сигнальных каскадов
- Ингибирование металлопротеиназ
- Ингибирование урокиназы
- EGCG и циклооксигеназа-2
- Регуляция клеточного цикла
- Блокада неоангиогенеза
- Фотозащитный эффект EGCG
- Термогенез
- Антиоксидантная активность
- Другие антипролиферативные эффекты EGCG
- Механизмы противоопухолевой активности
индол-3-карбинола
- Анти-эстрогенный эффект
- Модуляция сигнальных путей ростовых факторов
- Противовирусный потенциал I3C в отношении вируса
папилломатоза человека
- Медико-социальные аспекты папилломатоза
- HPV-онкомаркеры
- Иммуномодулирующий эффект I3C
- Влияние на плотность кости
- Механизм апоптоза опухолевых клеток под
влиянием I3C
- Сравнение активностей тамоксифена и
индол-3-карбинола
- список литературы к Главе 8
Заключение стр. 208

ЛА АШРАФЯН, В.И. КИСЕЛЕВ
ОПУХОЛИ РЕПРОДУКТИВНЫХ ОРГАНОВ (ЭТИОЛОГИЯ И ПАТОГЕНЕЗ)
Художники: Толмачев О.В., Щетилина А.А. Верстка: Коломеец Д.С, Галицкий М.А.
Технический редактор: Дьячкова Э.Г. Корректоры: Филановская B.C., Иванова Н.Н.
В оформлении обложки использована репродукция картины А. Модильяни «Лежащая обнаженная» (1917-1918)
Подписано в печать 06.03.2007 Формат 60x88/16
Печать офсетная. Бумага офсетная №1. Гарнитура TextBookC. Печ.л. 13,5 + 1,75 вкл.
Тираж 1000 экз. Заказ 4703
Издательство «Димитрейд График Групп» Тел.: (495) 558-80-58, (495) 558-80-05,
E-mail: [email protected] Отпечатано с готовых диапозитивов в ФГУП «ПИК ВИНИТИ»
140010, г. Люберцы, Октябрьский пр-т, 403
![ОПУХОЛИ БРЮШНОЙ ПОЛОСТИ · 2017-09-19 · >eybg^b\b^mZevgh]h bkihevah\Zgby ©=WHL:J F_^bZª 298 Опухоли брюшной полости Ежедневное](https://static.fdocuments.net/doc/165x107/5ee1d3d1ad6a402d666c90c1/-2017-09-19-eybgbbmzevghh.jpg)

















