TEMA. CITOLOGIA. CÉLULA EUCARIOTA. ENVOLTURAS CELULARES

of 12
-
Upload
mgonate4814 -
Category
Documents
-
view
586 -
download
0
Transcript of TEMA. CITOLOGIA. CÉLULA EUCARIOTA. ENVOLTURAS CELULARES
Biologa 2 Bach.
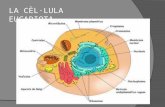
CLULA EUCARIOTA ENVOLTURAS CELULARESLas envolturas celulares que presentan las clulas eucariotas son la membrana plasmtica y la pared celular vegetal. Las clulas eucariotas animales presentan adems, por fuera de la membrana plasmtica una matriz extracelular.
1. La membrana plasmtica.La membrana plasmtica (tambin denominada citoplasmtica, celular o plasmalema) es una envoltura continua que delimita a todas las clulas (procariotas y eucariotas) y que est en contacto con el medio exterior celular por una de sus caras y con el hialoplasma celular por la otra. Al microscopio electrnico se observa cmo una envuelta de unos 75 de espesor (en realidad se aprecian dos bandas oscuras separadas por un espacio claro) que en el caso de las clulas animales es la nica estructura que separa las clulas de en medio ambiente que las rodea. Es una cubierta con dos caractersticas: es una estructura dinmica y asimtrica. Dinmica. Las molculas que la constituyen se pueden desplazar lateralmente, lo que permite que la membrana pueda autorrepararse, si sufre una ruptura, o fusionarse con cualquiera otra membrana. Gracias a eso pueden ocurrir los procesos de endocitosis y de exocitosis. Asimtrica. El conjunto de los oligosacridos que se unen a los lpidos y protenas de la membrana formando el glucoclix, slo se encuentra en la cara externa de la membrana de las clulas (estos oligosacridos son receptores de membrana y estn relacionados con las funciones de reconocimiento celular). La cara interna de la membrana no tiene glicoclix, sino una serie de protenas perifricas, muchas de las cuales constituyen puntos de anclaje para lo citoesqueleto.
1.1 Estructura y composicin de la membranaEstructura. Se puede generalizar la arquitectura de la membrana asumiendo el modelo terico elaborado por Singer & Nicholson (1972) denominado modelo del mosaico fluido. Segn este la membrana plasmtica est constituida por una doble capa de lpidos a la cual se asocian molculas proteicas, que se observan en las dos caras de la superficie de esta doble capa, total o parcialmente englobadas en ella (mosaico). Tanto los lpidos como las protenas se pueden mover (fluido). Este modelo aparece en todas las membranas que hay en la clula, ya que muchos orgnulos son de naturaleza membranosa (retculo, mitocondrias, lisosomas, etc) por eso tambin se habla de membrana unitaria. Composicin. A pesar de las mltiples posibilidades y funciones que se pueden encontrar en las membranas biolgicas la composicin qumica siempre es: lpidos, protenas, y glcidos.
LPIDOS. Son las ms abundantes, constituyen la mitad de la masa total de la membrana.Diferenciamos fosfolpidos y colesterol. FOSFOLPIDOS Una doble capa de fosfolpidos (tanto esfingolpidos cmo fosfoglicridos) conforma el entramado bsico de este modelo. Forman una bicapa, en las que las regiones polares de losCitologa. Envolturas celulares
1
Biologa 2 Bach.
fosfolpidos se disponen orientadas hacia las dos superficies de la misma (interior y exterior celular), mientras las regiones apolares se enfrentan entre s en el interior. Esta disposicin de los fosfolpidos no es rgida sino que es fluida, es decir, las molculas se pueden mover lateralmente dentro de la misma capa (a ms insaturacin, mayor fluidez); rotar; o incluso cambiar de una capa a la otra (movimiento conocido cmo flip-flop). COLESTEROL Intercaladas en la bicapa lipdica aparecen molculas de colesterol interaccionando con las cadenas hidrfobas de los fosfolpidos. Esta disposicin limita la movilidad de los fosfolpidos, reduciendo su fluidez y dndole estabilidad a la membrana.
PROTENASSon caractersticas de cada especie, e incluso de cada tipo de clula. Su presencia determina en gran medida las especializaciones funcionales de la membrana. En su mayora son de tipo globular, por lo que la solubilidad de las mismas depende de la presencia de cadenas laterales polares en la superficie de la molcula: la zona de la protena con los radicales polares queda fuera de la bicapa, mientras la zona que tiene grupos lipfilos establecer contacto con la zona hidrfoba de la membrana. De este modo aparecen protenas: -Perifricas o extrnsecas. Situadas en el exterior de la membrana, pudiendo estar unidas a los lpidos o a otras protenas transmembrana. Son solubles en soluciones acuosas y se aslan fcilmente por estar dbilmente unidas a los lpidos. Integrales o intrnsecas. Con cierta anfipata ya que tienen una parte incluida en la bicapa lipdica. Pueden incluso atravesar la membrana de parte a parte denominndose entonces protenas transmembrana. Estn ntimamente asociadas a los
Citologa. Envolturas celulares
2
Biologa 2 Bach.
lpidos, representan el 70% del total y son muy difciles de separar. Las protenas de la membrana tambin pueden rotar y desplazarse en el plano de la bicapa, y as su concentracin puede modificarse segn las necesidades de la clula, simplemente arrastrando las protenas hacia un mismo punto.
GLCIDOSEn la cara externa de la membrana aparecen multitud de glcidos (oligosacridos) unidos a los propios lpidos formando glucolpidos o tambin a las protenas (glucoprotenas). Esta capa as definida es el denominado glucoclix o cubierta celular. En las clulas animales este glucoclix se encuentra conectado o inmerso en una matriz extracelular (formada por colgeno, elastina y otras protenas), que resulta de la secrecin de la misma clula. Cuando se observa la membrana en el microscopio electrnico se aprecian tres capas: una central clara, correspondiente a la bicapa lipdica, y dos capas externas ms oscuras que se corresponderan con las protenas que sobresalen en ambas superficies. A la disposicin de los componentes proteicos y lipdicos de la membrana se refiere el nombre del modelo: forman una especie de mosaico. La capacidad de movimiento de los lpidos y las protenas dentro de la membrana es la causa por la que el modelo propuesto por Singer y Nicholson reciba tambin el nombre de fluido.
1.2 Propiedades de la membranaEste modelo as descrito determina una arquitectura molecular que explica varias propiedades observadas en las membranas celulares. Asimetra. Los medios que separa la membrana son de naturaleza muy distinta, y las funciones que lleva a cabo en ambos lados deben ser tambin diferentes. Las molculas que asoman a la cara externa da membrana (glucoclix) son distintas de las que aparecen en la cara interna (protenas perifricas). Esta asimetra incluye a todos los componentes de la membrana, tanto lpidos, como protenas y por supuesto a los glcidos. Fluidez. La mayora de las funciones que desenvuelve la membrana de una clula precisan de una alta flexibilidad y dinamismo (la fusin entre membranas, la redistribucin de los componentes, etc.). En realidad la membrana casi tiene un comportamiento de fluido pues sus componentes tienen un alto grado de movilidad, especialmente los fosfolpidos y tambin las protenas. Esta fluidez sin embargo est condicionada por varios factores, tanto qumicos como fsicos: - La mayor o menor presencia de colesterol (disminuye la movilidad de los FL). - El grado de insaturacin de los cidos grasos de los fosfolpidos de la bicapa (a mayor insaturacin, mayor capacidad de movimiento). - La longitud de los cidos grasos (se estos son saturados, menor movilidad). - La temperatura (a menor temperatura, menor fluidez). Permeabilidad. Debido a su interior hidrfobo, las membranas son muy impermeables a los iones y a la mayor parte de las molculas polares. En muchos casos estas molculas atravesarn a membrana si existen protenas que lo posibiliten. Se dice por esto que la membrana plasmtica tiene permeabilidad selectiva
Citologa. Envolturas celulares
3
Biologa 2 Bach.
1.3 Funciones de la membranaLa membrana plasmtica de la clula es una estructura altamente diferenciada y con funciones indispensables que se relacionan con alguna de las subestructuras presentes en su composicin. No obstante, estas funciones no son posibles fuera de la estructura de la membrana, que funciona como un todo. Resumen de las funciones de la membrana asociadas a subestructuras.
Adems la membrana realiza las siguientes funciones: Confiere individualidad a la clula. Separa el interior muy organizado del entorno inestable e desorganizado. Esta separacin no implica en cambio un aislamiento, sino un intercambio controlado y sobre todo selectivo de substancias con el exterior.Citologa. Envolturas celulares
4
Biologa 2 Bach.
Interaccin con el entorno. La funcin de relacin de la clula con su medio se realiza a travs de su membrana, percibiendo estmulos del exterior e incluso generando respuestas a ellos. Tambin recibe e transmite seales (transduccin) y es la responsable del reconocimiento celular.
1.3.1 Mecanismos de transporte a travs de la membranaLa membrana plasmtica controla la entrada y salida de substancias de la clula, mediante distintos tipos de transporte que dependen fundamentalmente de la naturaleza y del tamao de estas. Estos tipos de transporte se realizan bsicamente de dos formas: sin deformacin de la membrana y con deformacin de la misma.
A.
Sin deformacin de la membrana.
Las molculas pequeas o los iones pueden atravesar la membrana sin necesidad de que se produzca una alteracin en su estructura. Este proceso puede ser por transporte pasivo (no requiere energa) o activo (requiere energa).
Transporte pasivo Las molculas simplemente difunden (atraviesan) a travs de la membrana siguiendo su propia tendencia a equilibrar concentraciones a ambos los lados de la membrana, es decir, se mueven a favor de gradiente de concentracin. Se trata por lo tanto de un transporte que no requiere gasto de energa. Hay dos modalidades: Difusin simple. Algunas molculas de pequeo tamao y sin carga, como O2, CO2, difunden rpidamente a travs de la bicapa, a favor de su gradiente de concentracin (es decir, del medio ms concentrado al menos concentrado). Tambin substancias liposolubles como hormonas esteroideas, ter y cloroformo. Incluso pueden pasar molculas dbilmente polares y de baja masa molecular como etanol y urea. Se piensa que tiene lugar gracias a los huecos que aparecen entre los fosfolpidos motivados por el continuo movimiento de estos.
Citologa. Envolturas celulares
5
Biologa 2 Bach.
Difusin facilitada. Las molculas polares que no pueden cruzar libremente la bicapa utilizan protenas transmembrana. Estas pueden actuar como: - Protenas de canal. Tienen un orificio o canal interno por el que pasan iones como Na+, K+, Ca2+ e Cl-. - Protenas transportadoras, carriers (ou permeasas). Transportan glcidos pequeos, aminocidos y nuclesidos. Se unen especficamente a determinadas molculas y, mediante un cambio en la conformacin espacial de la protena, esa molcula pode acceder al otro lado de la membrana. Transporte activo Se realiza en contra de un gradiente electroqumico (diferencia de carga elctrica y de concentracin), yendo por lo tanto en contra de la propia tendencia de las molculas a difundir hacia el lado menos concentrado. Para conseguir esto la membrana dispone de protenas especficas para esas substancias, tambin llamadas permeasas, capaces de transportar molculas en contra de gradiente. Por supuesto que este trabajo se hace a costa de un gasto energtico que normalmente es suministrado por el ATP (por la hidrlisis del mismo). Es el caso de las bombas de iones, encargadas de crear un desequilibrio elctrico entre ambas caras de la membrana plasmtica. As, la bomba de Na+-K+ (imprescindible para mantener el potencial de membrana en las clulas animales; por este mecanismo el interior de la clula es - y el exterior es +). Bombea activamente 3 iones de Na+ hacia el exterior de la clula, mientras introduce 2 iones de K+ en el citoplasma; as la concentracin de Na+ es mayor en el exterior celular y la de K+ es mayor en el interior celular. Se trata de un mecanismo extremadamente importante en los fenmenos de transmisin nerviosa o en la contraccin muscular, incluso para el mantenimiento de la presin osmtica en las clulas animales (estas gastan ms del 30 % de su ATP en mantener la bomba de Na+-K+ y las nerviosas el 70%). Tambin es importante el transporte activo en las clulas que precisan almacenar nutrientes en el citoplasma a pesar de estar ms concentrados que en el exterior, tal como sucede con la glucosa.
B. Con deformacin de la membrana.Las clulas animales tambin pueden captar o expulsar macromolculas como polisacridos, o incluso otras clulas o partes de ellas. Es un mecanismo menos especfico que precisa de vesculas membranosas, encargadas de transportar en su interior dichas molculas disueltas. Estas vesculas conservan la asimetra de la membrana plasmtica. La entrada de materia a la clula mediante estos mecanismos es la endocitosis, mientras que la salida de materia de la clula es la exocitosis. La transcitosis es un sistema de transporte que combina estos dos procesos.
Citologa. Envolturas celulares
6
Biologa 2 Bach.
Por otra parte, la continua prdida de fragmentos membranosos debido a la entrada de substancias por endocitosis se ve compensada por la salida de otras mediante exocitosis. Se consigue de este modo la renovacin continua de la membrana plasmtica. Endocitosis La clula introduce molculas de gran tamao, partculas slidas o incluso fluidos que contienen macromolculas o partculas de pequeo tamao. Estas se adhieren a la superficie de la membrana y desencadenan la formacin de una depresin, que va progresando hasta envolverlas completamente y formar una vescula. - Fagocitosis. Cuando las vesculas tienen un gran tamao por envolver partculas grandes, como restos celulares o microorganismos, se habla de fagocitosis (phagos = el que come). La clula emite una especie de prolongaciones denominadas pseudpodos que rodean a la substancia. La gran vescula que se forma recibe el nombre de vacuola fagoctica, fagocito o fagosoma y mide varios micrmetros. Las enzimas de los lisosomas se encargan despus de descomponer los materiales fagocitados (se produce la unin de este fagosoma con uno o varios lisosomas formando un fagolisosoma). Se da en algunos protozoos (modo de alimentacin) y glbulos blancos (defensa contra agentes patgenos). - Pinocitosis. Si las vesculas son menores y contienen fluidos, el proceso se denomina pinocitosis (pinein = beber). Consiste en la toma indiscriminada de lquido extracelular. Una depresin en canal de la membrana atrapa en su extremo una gota de lquido que se libera hacia el interior del citoplasma en forma de una pequea vescula de dimetro inferior a un micrn, denominada pinocito. De esta forma se incorporan a la clula las molculas pequeas o micropartculas presentes en la gota. - Existe un tercer tipo de endocitosis: la endocitosis mediada por receptor. A diferencia de la pinocitosis, que es relativamente lenta, este es un proceso rpido y altamente especfico. Los receptores de membrana protenas transmembrana- toman en este caso especficamente del lquido extracelular una determinada substancia, denominada ligando, que es una molcula proteica o una partcula pequea, para la que existe un receptor determinado en la membrana que las reconoce especficamente. Estos receptores se pueden acumular en determinadas zonas de la superficie de la membrana, denominadas depresiones revestidas con clatrina (esta es una protena que forma una estructura de recubrimiento) o Exocitosis Es un proceso inverso a la endocitosis. Determinadas vesculas internas producidas por la propia clula (normalmente por el complejo de Golgi, o el retculo endoplasmtico) pueden emigrar hasta la cara interior de la membrana, fusionarse con ella y finalmente liberar su contenido al exterior. Este proceso produce un aumento de la superficie de la membrana plasmtica. Estas molculas segregadas suelen cumplir una funcin fuera de la clula ya que muchas veces son substancias sintetizadas, como polisacridos o protenas. A veces son molculas de desecho.
Citologa. Envolturas celulares
7
Biologa 2 Bach.
o Transcitosis Constituye un sistema de transporte a travs del citoplasma, segn el cual vesculas formadas por endocitosis no quedan en el interior de la clula, sino que se dirigen al extremo opuesto de esta y liberan su contenido al exterior, por exocitosis. Es un mecanismo que se da, por ejemplo, en las clulas de los endotelios de los capilares sanguneos, en los que determinadas substancias tienen que atravesar esa pared desde la luz del vaso al exterior de este o viceversa.
1.3.2. Transduccin de sealesLas clulas son capaces de recibir y transmitir seales gracias a los receptores especficos (normalmente glcidos o protenas de membrana) de estmulos externos. Estos pueden ser fsicos o qumicos, y son capaces de desencadenar una respuesta en el interior de la clula. Esta funcin est especialmente presente en los mecanismos de coordinacin entre las clulas de organismo pluricelular. Aunque algunas de los mensajeros qumicos que viajan por el interior de estos organismos pueden atravesar fcilmente la membrana de las clulas diana, otros muchos mensajeros deben unirse la algn receptor de la membrana, pues no pueden pasar al interior del citoplasma. Estos primeros mensajeros o ligandos hacen que la unin con el receptor induzca una segunda seal intracelular (segundo mensajero) que modificar el metabolismo de la clula. Tal es el caso del AMPc, que se fabrica en la clula cuando a nivel de la membrana tiene lugar a unin entre el ligando y su receptor.
1.3.3 Reconocimiento celularLa complejidad de los oligosacridos en el glucoclix, as como el hecho de encontrarse en la superficie exterior celular, sugiere que estos azcares son los principales marcadores de identidad que actan nos diversos procesos de reconocimiento celular. En los procesos infecciosos las bacterias deben adherirse a la superficie del tejido; esta unin se hace a nivel de los azcares de la membrana de la clula hospedadora. De ah que las infecciones tengan un alto grado de especificidad entre la bacteria y el tejido que va a ser infectado. Las clulas que pertenecen a un mismo tejido se reconocen entre s, y son capaces de adherirse y provocar respuestas directas en las vecinas. Este mecanismo se denomina histocompatibilidad. Incluso las clulas de distintos individuos, aunque sean de la misma especie, presentan molculas propias en la superficie de sus membranas. Por esto, cuando se transplantan clulas de un organismo a otro, este receptor puede generar defensas (anticuerpos) especficos contra las clulas extraas.
Citologa. Envolturas celulares
8
Biologa 2 Bach.
Los espermatozoides, por ejemplo, distinguen las ovoclulas de la propia especie gracias al glucoclix de ambas clulas sexuales.
1.4. Diferenciaciones de la membrana plasmticaEn la membrana plasmtica podemos encontrar distintos tipos de especializaciones segn la funcin que desempee la clula. Algunas estn destinadas a aumentar la superficie celular. Las ms importantes son: - Microvellosidades. Aparecen en la membrana de las clulas epiteliales del intestino para aumentar la superficie de absorcin de los nutrientes. - Invaginaciones. En las clulas que tapizan el tbulo contorneado de las nefronas. Son profundos entrantes que aumentan la superficie celular para que puedan ser transferidos a la sangre todos los metabolitos que no deben ser eliminados en la orina. En otros casos las modificaciones son estructuras de unin entre clulas, esenciales para estabilizar la formacin y el funcionamiento de los tejidos. Las ms importantes son: Uniones ntimas o de oclusin. Son uniones que no dejan espacio intercelular y, por lo tanto, no permiten el paso de substancias a travs de las capas celulares. Estn formadas por protenas transmembrana (claudinas y ocludinas) que se disponen formando filas que sueldan las membranas plasmticas entre s. Estn, adems, reforzadas por protenas filamentosas intracelulares. Abundantes en el epitelio intestinal donde permiten la absorcin de los nutrientes. Uniones adherentes. Son uniones que dejan un espacio por el que pueden pasar substancias, entre clulas que pertenecen a un tejido. Pueden ser: desmosomas y hemidesmosomas. Los desmosomas forman como botones a distintos niveles entre las membranas plasmticas de clulas contiguas. La unin se produce gracias a protenas transmembrana (cadherinas) que se fijan, en la cara interna de las membranas, en una placa (placa desmosmica) que tiene forma de disco. Esta placa tambin se une a los filamentos intermedios del citoesqueleto por una red de filamentos de queratina. Se encuentran principalmente entre las clulas de los tejidos epiteliales.
Los hemidesmosomas se localizan en la parte basal de las clulas epiteliales y las conectan con las clulas del tejido conectivo subyacente. Su estructura es la de medio desmosoma, de ah su nombre.
Citologa. Envolturas celulares
9
Biologa 2 Bach.
Uniones de comunicacin o tipo gap. Son uniones que no dejan espacio intercelular, pero s dejan un pequeo espacio de comunicacin entre los citoplasmas de las dos clulas por medio de canales proteicos y, por lo tanto permiten el paso de molculas entre ellas. Estn constituidas por dos conexiones. Cada conexin es una especie de tubo fino de contorno hexagonal formado por seis protenas transmembrana (llamadas conexinas) que dejan un canal en el centro. Esta conexin est alineada con la de la clula adyacente. Las clulas slo quedan separadas por el espacio que corresponde a las porciones de las conexinas que sobresalen de las membranas plasmticas.Este tipo de unin interviene en la transmisin del impulso nervioso entre las neuronas. La membrana de las clulas vegetales tambin presenta diferenciaciones que permiten la comunicacin y el intercambio de substancias entre las clulas. Son los plasmodesmos y las punteaduras
1.5. La matriz extracelularConstituye el medio ambiente de las clulas animales. Es un entramado de molculas, protenas y glcidos dispuestos en el espacio intercelular, que son sintetizadas y secretadas por las propias clulas. Tiene mltiples funciones: aporta propiedades mecnicas a los tejidos, mantiene la forma celular, permite la adhesin de las clulas para formar tejidos, permite la comunicacin intercelular, forma sendas por las que se mueven las clulas, modulan la diferenciacin celular y la fisiologa celular, secuestran factores de crecimiento, etctera. La cantidad, la composicin y la disposicin de la matriz extracelular depende del tipo de tejido considerado. Hay algunos como el epitelial y el nervioso que carecen o tienen muy poca matriz extracelular, mientras que en otros como el tejido conectivo es el elemento ms importante en volumen.
2. La pared celular vegetalEs una cubierta inerte, rgida e insoluble que les confiere forma, proteccin, rigidez e inmovilidad a las clulas, mantiene su balance osmtico, contribuye la que las plantas se mantengan erguidas y, une, conecta y permite los intercambios entre las clulas adyacentes. Es de naturaleza principalmente celulsica (microfibrillas unidas por puentes de hidrgeno), aunque presenta una matriz en la que hay agua, sales minerales, hemicelulosa y pectina (substancia con gran capacidad de retener agua). Todas estas substancias las produce y segrega la clula (exocitosis)
2.1 Composicin y estructuraLa pared celular de las plantas (la de las algas es muy parecida) est compuesta bsicamente de celulosa y matriz. En todas las clulas en crecimiento y en determinadas clulas maduras consta de dos capas: la lmina media y la pared primaria. En muchas clulas presenta una tercera capa, la pared secundaria, que se desarrolla cuando las clulas concluyeron su crecimiento y que, adems, puede ser reforzada con substancias como lignina, suberina y cutina.
Citologa. Envolturas celulares
10
Biologa 2 Bach.
Lmina media: Es delgada y flexible. Es una estructura que se sita entre las paredes primarias de clulas vecinas, actuando como un cemento que une ntimamente las paredes primarias adyacentes. Tiene una estructura simple y una consistencia gelatinosa debido a su contenido en pectinas (macromolculas constituidas por sales clcicas y magnsicas, con una base estructural que es el cido galacturnico). Cuando las clulas de las partes viejas del tejido conductor de las plantas mueren, las lminas medias se impregnan de lignina, lo que aumenta la resistencia mecnica del tejido. Durante la divisin celular (en la citocinesis), cuando las vesculas procedentes del aparato de Golgi se alinean para constituir la placa celular (fragmoplasto), se vierten en su interior los precursores que van a dar origen a la lmina media. A ambos los lados de ella se van depositando las paredes primarias de las clulas hijas. Pared primaria: consiste en una cubierta delgada, flexible y elstica, que delimita externamente la clula vegetal a modo de caja. Qumicamente est formada por un retculo microfibrilar y una matriz amorfa. El retculo microfibrilar lo constituyen largas y finas molculas de celulosa que se agrupan, alineadas y en paralelo, para dar fibrillas elementales de celulosa. Estas se entrelazan para dar microfibrillas de celulosa. Estas ltimas permiten el crecimiento de la clula por su facilidad de extensin. Las microfibrillas se enrollan helicoidalmente entre s, como los hilos de un cable, originando las macrofibrillas. Estas se agrupan siguiendo el mismo patrn y forman las fibras celulsicas. De esta manera se forman cables de celulosa (fibras) que, a igualdad de grosor, alcanzan una resistencia similar a la del acero. Entre las fibras de celulosa se sita una matriz amorfa de sustancias no celulsicas formada por agua, molculas ramificadas de otros polisacridos, substancias pcticas, monosacridos libres y protenas. En las paredes de clulas muertas suele depositarse lignina aglutinando las microfibrillas de celulosa, lo que constituye la madera. La lignina es un material amorfo, de color marrn oscura, que proporciona resistencia tanto mecnica cmo qumica. En funcin de las misiones que desempee la clula en la planta, pueden encontrarse en su pared primaria, impregnadas o depositadas sobre ella, otras substancias, como sales minerales, gomas, ceras o compuestos pcticos. Un ejemplo es la capa de cutina, que tiene por misin evitar la prdida de agua a travs de las paredes exteriores. La suberina, substancia impermeable depositada en lminas sobre la pared, da origen al sber o corteza. A medida que se desarrolla la clula, la pared celular primaria se estira y crece gracias al depsito de nuevas fibrillas de celulosa hasta alcanzar el su tamao definitivo. Las molculas de celulosa llegan hasta la pared en vesculas procedentes del aparato de Golgi. Pared secundaria: en muchas clulas vegetales slo existe una pared primaria, pero en otras se forma internamente una pared suplementaria de mayor grosor que tiene como misin otorgar una resistencia mecnica ms elevada al tejido. La formacin de esta pared se produce cuando la pared primaria ya no aumenta de tamao, al finalizar el crecimiento celular.Citologa. Envolturas celulares
11
Biologa 2 Bach.
Este tipo de paredes adquiere gran importancia en las clulas de los tejidos conductores o de resistencia mecnica. En ellos, las clulas mueren tras formar la pared secundaria (por la dificultad de intercambio de substancias con el exterior) que puede llegar a ocupar toda la luz celular. La composicin qumica de la pared secundaria es similar a la de la pared primaria, aunque es ms rica en celulosa, tiene menos agua y hemicelulosa y carece de pectinas. La pared secundaria presenta una estructura laminar con 3 capas diferentes, aunque en ocasiones el n de capas puede llegar a veinte (hojas de loe). La diferencia en relacin con la pared primaria estriba en la disposicin en hlice y en la mayor densidad que presentan las microfibrillas de celulosa, lo que impide el crecimiento celular.
Es muy habitual la presencia de lignina, como continuacin del depsito iniciado en la lmina media y en la pared primaria. En las clulas de los tejidos conductores y de sostn, la pared secundaria no llega a extenderse sobre la totalidad de la pared primaria y se deposita en forma de anillos, tiras helicoidales, retculos o refuerzos irregulares. Estructura de la pared celular secundaria
Plasmodesmos y punteaduras.El intercambio de agua y solutos entre las clulas vecinas se facilita por la existencia de los plasmodesmos y las punteaduras. Al formarse la lmina media, se mantiene la comunicacin entre las clulas hijas a travs de orificios que atraviesan finas comunicaciones citoplasmticas, a modo de cordones, llamados plasmodesmos. Aparecen en nmero variable y segn crecen las clulas se van formando nuevos plasmodesmos. Pueden estar distribuidos por toda la superficie o formar grupos en ciertas zonas de la pared primaria, constituyendo los llamados campos primarios de punteaduras. Las punteaduras son cavidades o depresiones de la pared secundaria que se corresponden entre dos clulas vecinas. Se distinguen dos tipos de punteaduras: las simples y las que tienen borde o aureola. Punteaduras simples: no existen restos de pared secundaria en la punteadura. Punteaduras con borde: son aquellas en las que la pared secundaria se arquea sobre la cavidad de la punteadura, constituyendo un borde.
Citologa. Envolturas celulares
12
![UNIDAD 8: LA CÉLULA EUCARIOTA: ENVOLTURAS … · Citosol o hialoplasma. ... PASIVO: a favor de gradiente de [] o de carga. No gasto de E. Es una difusión. ... INCLUSIONES CITOPLASMÁTICAS](https://static.fdocuments.net/doc/165x107/5bb6f87c09d3f20c668d0166/unidad-8-la-celula-eucariota-envolturas-citosol-o-hialoplasma-pasivo.jpg)