
Regulation of Lipid Synthesis in Soybeans byTwo … · Thin Layer Chromatography. ... homogenate...
6
Plant Physiol. (1977) 60, 730-735 Regulation of Lipid Synthesis in Soybeans by Two Benzoic Acid Herbicides' Received for publication February 8, 1977 and in revised form August 2, 1977 RAAD K. MUSLIH2 AND DEAN L. LINSCOTr Department of Agronomy, Cornell University, Ithaca, New York 14853 ABSTRACT The effects of 3-nitro-2,5-dichlorobenzoic acid (dinoben) and 3- amino-2,4-dlchlorobenzolc acid (chloramben) on lipid formation and on the incorporation of various substrates into lipids by intact seeds and subceflular fractions of germinating soybean (Glycine max [L.] Merr. 'Amsoy') were studied. Dinoben (20 lg/ml) inhibited synthesis of total lipils 67%, neutral lipids 73%, glycolpids 51%, and pbospholipids 39% in germinating seeds. When polar lipids were analyzed further, inhibition of individel lipid dcass was also observed. Chloramben (20 pg/ml) stimulated total lpid synthesis 25%. With the exception of the mitochondrial fraction where malonate thiokinase was absent, dinoben inhibited up to 99% the incorporation of acetate and malonate into lpids, but did not inhibit acetyl-CoA and malonyl-CoA incorporation. Chloramben stimulated the incorporation of aU substrates tested into lipids by all fractions except the mitochodrial fraction when malonate was the substrate. When dinoben and chloramben were used in combi- nations, chioramben did not reverse the inhibitory effect of dinoben. It is concluded that the dinoben inhibitory effect specific and is assodated with the acetate and malonate thiokinase systems. The chioramben effect b stimulatory to either acetyl-CoA carboxylase or fatty add synthetase or both. Recently, research on various aspects of soybean metabolism has increased substantially. However, little is known about specific metabolic processes in soybean during germination. In particular, lipid synthesis during germination of soybean has not been studied in great detail. Quantitative and qualitative alteration of membrane lipid composition results in changes in chloroplast and mitochondrial functions and membrane permeability capacity (1, 9). Thus, chemical inhibition of lipid synthesis could ultimately result in loss of photosynthetic and respiratory activity, plant necrosis, and changes in the nutritional balance of seedlings. Mann and Pu (8) have shown that 3-nitro-2,5-dichlorobenzoic acid (di- noben) inhibits 2-14C-malonate incorporation into lipids by excised hypocotyl of hemp sesbania (Sesbania exaltata [Raf.] 'Cory'). The reduced form, 3-amino-2,4-dichlorobenzoic acid (chloramben) stimulated 2-14C-malonate incorporation into lip- ids by the same tissue (8). Both dinoben and chloramben are selective herbicides applied to soil for control of weeds in the germination-early growth stage. Thus, knowledge of their effects on lipid synthesis could bring better understanding of the reason(s) for selectivity. I Cooperative investigations of Cornell University and the Agricul- tural Research Service, United States Department of Agriculture. Department of Agronomy Journal Series 216. 2 Present address: College of Agriculture, Baghdad University, Bagh- dad, Iraq. In this study we report the effect of dinoben and chloramben on incorporation of labeled acetate and other substrates into lipids and the enzyme systems involved. MATERIALS AND METHODS Seed Germination. After 3 hr imbibition, soybean seeds (Glycine max [L.] Merr. 'Amsoy') were germinated at 25 C in small test tubes (one seed/tube) in 0.5 ml of deionized H20 containing 0.25 ,uCi of 1-14C-acetate and the proper concentra- tion of chloramben or dinoben. After 24 hr (unless otherwise indicated), the germinated seeds were transferred to 95 C deionized H20 for 2 min and then washed three times with 100 ml of water. Extraction of Total Lipids. Ten germinated seeds (about 4 g) were homogenized for 3 min with 10 volumes (ml/g) of chloro- form-methanol (2:1, v/v). The homogenate was left overnight at room temperature. The fine suspension was then filtered and the tissue residue was further extracted with 10 ml of the same solvent system. Removal of Nonlipid Material. Sephadex G-253 column chro- matography was used as described by Wells and Dittmer (22) and modified by Wuthier (23). Silicic Acid Column Chromatography. Silicic acid column chromatography was used for the initial separation of neutral lipids from phospholipids and glycolipids by a stepwise elution. Heat-activated Hi-Flosil 100-200 mesh was treated and packed in a chromatography tube (2.5 x 30 cm) as described (12). The solvent systems described by Rouser et al. (17) were used (Table I). Diethylaminoethyl (DEAE)-Celiulose Column Chromatogra- phy. The acetate form of DEAE-cellulose powder with an exchange capacity of 0.9 ± 0.1 meq/g was prepared according to the method of Rouser et al. (16). The mixture of phospholip- ids to be fractionated (fraction 4 from the silicic acid column) was transferred quantitatively to the column as a solution in pure methanol. The solvent systems reported by Hitchcock and Nichols (5) were used (Table I). Thin Layer Chromatography. TLC was used to confirm the fractionation results of silicic acid chromatography and DEAE column chromatography qualitatively and to identify the labeled lipid classes. Aluminum TLC sheets precoated with Silica Gel F245, 0.25 mm thick, and plastic Polygram Sil G/UV254, 0.25- mm-thick sheets were used. Chromatography was performed at room temperature in chambers lined with filter paper. The solvent systems used for one-dimensional and two-dimensional TLC for fractions specified are shown in Table I. Spots were made visible by charring (15) and Rhodamine 6G (14) for total lipids. The molybdenum blue reagent (3) was used to detect 3 Mention of a trade name or company designation here and else- where in the paper is for identification purposes and does not imply endorsement by the U.S. Department of Agriculture over products of a similar nature. 730 www.plantphysiol.org on August 28, 2018 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.
Transcript of Regulation of Lipid Synthesis in Soybeans byTwo … · Thin Layer Chromatography. ... homogenate...

Plant Physiol. (1977) 60, 730-735
Regulation of Lipid Synthesis in Soybeans by Two Benzoic AcidHerbicides'
Received for publication February 8, 1977 and in revised form August 2, 1977
RAAD K. MUSLIH2 AND DEAN L. LINSCOTrDepartment ofAgronomy, Cornell University, Ithaca, New York 14853
ABSTRACT
The effects of 3-nitro-2,5-dichlorobenzoic acid (dinoben) and 3-amino-2,4-dlchlorobenzolc acid (chloramben) on lipid formation andon the incorporation of various substrates into lipids by intact seeds andsubceflular fractions of germinating soybean (Glycine max [L.] Merr.'Amsoy') were studied. Dinoben (20 lg/ml) inhibited synthesis of totallipils 67%, neutral lipids 73%, glycolpids 51%, and pbospholipids39% in germinating seeds. When polar lipids were analyzed further,inhibition of individel lipid dcass was also observed. Chloramben (20pg/ml) stimulated total lpid synthesis 25%. With the exception of themitochondrial fraction where malonate thiokinase was absent, dinobeninhibited up to 99% the incorporation of acetate and malonate intolpids, but did not inhibit acetyl-CoA and malonyl-CoA incorporation.Chloramben stimulated the incorporation of aU substrates tested intolipids by all fractions except the mitochodrial fraction when malonatewas the substrate. When dinoben and chloramben were used in combi-nations, chioramben did not reverse the inhibitory effect of dinoben.
It is concluded that the dinoben inhibitory effect specific and isassodated with the acetate and malonate thiokinase systems. Thechioramben effect b stimulatory to either acetyl-CoA carboxylase orfatty add synthetase or both.
Recently, research on various aspects of soybean metabolismhas increased substantially. However, little is known aboutspecific metabolic processes in soybean during germination. Inparticular, lipid synthesis during germination of soybean hasnot been studied in great detail.
Quantitative and qualitative alteration of membrane lipidcomposition results in changes in chloroplast and mitochondrialfunctions and membrane permeability capacity (1, 9). Thus,chemical inhibition of lipid synthesis could ultimately result inloss of photosynthetic and respiratory activity, plant necrosis,and changes in the nutritional balance of seedlings. Mann andPu (8) have shown that 3-nitro-2,5-dichlorobenzoic acid (di-noben) inhibits 2-14C-malonate incorporation into lipids byexcised hypocotyl of hemp sesbania (Sesbania exaltata [Raf.]'Cory'). The reduced form, 3-amino-2,4-dichlorobenzoic acid(chloramben) stimulated 2-14C-malonate incorporation into lip-ids by the same tissue (8). Both dinoben and chloramben areselective herbicides applied to soil for control of weeds in thegermination-early growth stage. Thus, knowledge of their effectson lipid synthesis could bring better understanding of thereason(s) for selectivity.
I Cooperative investigations of Cornell University and the Agricul-tural Research Service, United States Department of Agriculture.Department of Agronomy Journal Series 216.
2 Present address: College of Agriculture, Baghdad University, Bagh-dad, Iraq.
In this study we report the effect of dinoben and chlorambenon incorporation of labeled acetate and other substrates intolipids and the enzyme systems involved.
MATERIALS AND METHODS
Seed Germination. After 3 hr imbibition, soybean seeds(Glycine max [L.] Merr. 'Amsoy') were germinated at 25 C insmall test tubes (one seed/tube) in 0.5 ml of deionized H20containing 0.25 ,uCi of 1-14C-acetate and the proper concentra-tion of chloramben or dinoben. After 24 hr (unless otherwiseindicated), the germinated seeds were transferred to 95 Cdeionized H20 for 2 min and then washed three times with 100ml of water.
Extraction of Total Lipids. Ten germinated seeds (about 4 g)were homogenized for 3 min with 10 volumes (ml/g) of chloro-form-methanol (2:1, v/v). The homogenate was left overnightat room temperature. The fine suspension was then filtered andthe tissue residue was further extracted with 10 ml of the samesolvent system.Removal of Nonlipid Material. Sephadex G-253 column chro-
matography was used as described by Wells and Dittmer (22)and modified by Wuthier (23).
Silicic Acid Column Chromatography. Silicic acid columnchromatography was used for the initial separation of neutrallipids from phospholipids and glycolipids by a stepwise elution.Heat-activated Hi-Flosil 100-200 mesh was treated and packedin a chromatography tube (2.5 x 30 cm) as described (12). Thesolvent systems described by Rouser et al. (17) were used(Table I).
Diethylaminoethyl (DEAE)-Celiulose Column Chromatogra-phy. The acetate form of DEAE-cellulose powder with anexchange capacity of 0.9 ± 0.1 meq/g was prepared accordingto the method of Rouser et al. (16). The mixture of phospholip-ids to be fractionated (fraction 4 from the silicic acid column)was transferred quantitatively to the column as a solution inpure methanol. The solvent systems reported by Hitchcock andNichols (5) were used (Table I).Thin Layer Chromatography. TLC was used to confirm the
fractionation results of silicic acid chromatography and DEAEcolumn chromatography qualitatively and to identify the labeledlipid classes. Aluminum TLC sheets precoated with Silica GelF245, 0.25 mm thick, and plastic Polygram Sil G/UV254, 0.25-mm-thick sheets were used. Chromatography was performed atroom temperature in chambers lined with filter paper. Thesolvent systems used for one-dimensional and two-dimensionalTLC for fractions specified are shown in Table I. Spots weremade visible by charring (15) and Rhodamine 6G (14) for totallipids. The molybdenum blue reagent (3) was used to detect
3 Mention of a trade name or company designation here and else-where in the paper is for identification purposes and does not implyendorsement by the U.S. Department of Agriculture over products of asimilar nature.
730 www.plantphysiol.orgon August 28, 2018 - Published by Downloaded from
Copyright © 1977 American Society of Plant Biologists. All rights reserved.

HERBICIDAL REGULATION OF LIPID SYNTHESIS
Table 1. Chromatography coluans, solvents and sequences for fractionationof germinating soybean seed lipids and their identificationby TLC.
Column and Solvent systems for Column Lipidfraction columns and TLC eluate class
volume
(v/v) El
Sephadex Folch (FLP). Two-dimensional TLC, 50x-chloroform-methanol-14 N NH40H,65:35:5; y-chloroform-acetone- Total lipidsmethanol-acetic acid-water, 100:40:20:20:10.
Silicic Acid
1 Chloroform. 200 Neutral lipids2 Chloroform-acetone 1:0. TLC, 140 MGMG, ESG, SC.
chloroform-_ethanol-7 N NI40H, 80:20:2.
3 Acetone. TLC, chloroform-ethanol- 600 DGDG, SL, CE14 N 1N1H40H, 65:25:4.
4 Methanol. Two-dimensional TLC x- 200 Phospholipids -
chloroform-methanol-water, 65:25:4; PA, PI, PE, PC,y-diisobutyl ketone-acetic acid- PS, PGwater, 80:50:10.
DEAE-Cellulose
1 Chloroform-methanol 95:5. TLC, 300 PC, LPC, U1chloroform-methanol-water, 65:25:4.
2 Chloroform-methanol -6:4. TLC, chlor- 300 PE, LPE, U2oform-methanol-water, 65:25:4.
3 Chloroform-methanol 2:1 saturated 300 PI, PG, PS, PAwith 14 N NH40H. TLC, as fraction4 of silicic acid column separation.
phospholipids and ninhydrin (18) to detect compounds contain-ing free amino groups. The orcinol test was used to detectglycolipids (20).
Gas-Liquid Chromatography. GLC was performed on an
Aerograph HY-FI model 600 gas chromatograph equipped witha flame ionization detector. The chromatograph was furtherequipped with a matrix temperature programmer and a columneffluent stream splitter. The carrier gas was nitrogen. Fatty acidmethyl esters were injected into a coiled borosilicate glasscolumn (300 cm x 4 mm) packed with ethylene succinatestructure polymers treated with methyl silicon on 100-200 meshGas-chrom Q. Oven temperature was 180 C and injector anddetector temperatures were 205 C.The "4C-fatty acid methyl esters from the GLC column
effluent were trapped in borosilicate glass tubes (4 x 0.5 cm)packed with 60-80 mesh HMDS-treated Chromosorb P whichwas coated with Dow-Coming DC 550 silicone oil, 5% byweight at room temperature. The entire contents of the tubeswere expressed into a counting vial containing liquid scintillator.
Isotope Assay. "4C-Labeled compounds were assayed withBeckman LS-100 C liquid scintillation spectrometer. The liquidscintillator fluid was prepared by adding 5 g of Nuclear-ChicagoPremix-M (95% PPO and 5% dimethyl POPOP) to a liter of95% toluene-5% n-propyl alcohol (v/v). Counting efficiency(62%) was determined by the channels ratio method. Quantita-tive determination of "4C-labeled compounds from TLC plateswas accomplished by scraping off the identified zones andassaying as described above. Radioactive regions were alsodetected by autoradiography by exposing no-screen x-ray filmto the developed thin layer sheets for 2 weeks. Liquid samplesfrom column chromatography were concentrated to minimalvolume and assayed as described above.
Protein Determination. Protein was determined by themethod of Lowry et al. (6). BSA was used as a standard.
Preparation of Homogenates and Cell Fractions. Seeds were
germinated between moist sterilized towel papers in an incuba-tor at 25 C for 24 hr. Germinated seeds appearing to be normalwere selected and washed with deionized H20. The washedseeds (50 g) were homogenized at 4 C in a mortar to a smoothpaste in a medium (2 ml/g) containing 0.05 M K-phosphatebuffer and 0.3 M sucrose (pH 7.2). The homogenate was
strained through four layers of cheesecloth to remove cellulardebris. The crude homogenate was centrifuged at 21,600g for
30 min at 4 C to obtain mainly mitochondrial materials. Thehomogenate separated into a top lipid layer, a middle clearlayer of crude extracts, and a pellet. The three layers wereseparated by decanting the supernatant liquid through severallayers of cheesecloth to remove the lipid layer which wasdiscarded. The pellet was washed with 10 ml of the homogeniz-ing medium and recentrifuged. The pellet was resuspended inhomogenizing medium and designated 21,600g P. The extractswere combined and centrifuged at 198,000g in a Spinco Ti-50rotor for 60 min at 4 C. The precipitate was resuspended in 5ml of the homogenizing medium and designated 198,000g P.The remaining supernatant was designated 198,000g S.
Assay Medium. Cell-free fractions were incubated in thefollowing media except where stated otherwise: ATP, 2 iumol;NADH, 0.5 ,umol; NADPH, 0.2 ,umol; glucose-6-P, 4 ,umol;glucose-6-P dehydrogenase, 0.5 unit; GSH, 8 umol; soybeanacyl carrier protein, 0.5 mg of protein; CoA, 0.2 j.mol; MnCl2,2 itmol; KHCO3, 30 ,umol; cell fraction preparations, 3 mg ofprotein; 0.2 jACi of the appropriate labeled substrate; and theappropriate concentration of dinoben or chloramben in a totalvolume of 2 ml. The mixture was incubated in test tubes at 37C for 2 hr. The reaction was stopped with 0.2 ml of 40% KOH.The lipids were extracted twice with 3 ml of chloroform-metha-nol (2:1, v/v). The nonlipid layer was removed and the lipidlayer was separated from nonlipid material by Sephadex columnchromatography as above. An aliquot was transmethylated with15% boron trifluoride-methanol for fatty acid analysis. Themethyl esters were analyzed by GLC. Two additional aliquotswere taken: one from the lipid layer to determine the labelincorporation into total lipids, and the other from the nonlipidlayer for comparison studies.
Chemicals and Substrates. BSA, CoA, and GSH were ob-tained from Nutritional Biochemicals Corp. Glucose-6-P, ATP,NADH, NADP, NADPH, and glucose-6-P dehydrogenase wereobtained from Calbiochem. 1-14C-Malonate and 2-14C-malonatewere obtained from ICN Isotope and Nuclear Division. 1_14C_malonyl-CoA and 2-14C-malonyl-CoA were prepared as de-scribed (21); ACP was isolated and purified from germinatingsoybean seeds as described (19). Solutions of dinoben andchloramben technical grade were prepared as described (8).
Data. All data are averages of four to six replications.
RESULTS AND DISCUSSION
Preliminary Experiments. Fractionation of total lipids bycolumn chromatography combined with quantitative and quali-tative TLC of unlabeled soybean seeds, germinated for 24 hr at25 C, revealed the presence of eight phosphorous-containinglipids. The four major components were: PC,4 PE, PI, and PG.The minor components were: PS, PA, and the remaining twowere unidentified. MGDG and DGDG were the major compo-nents of the glycolipid fraction. Another component gave apositive test with orcinol but was not identified. Sulfolipids,sterol glucosides and cerebrosides were also present, but noattempt was made to fractionate and identify their components.The distribution of radioactivity in the newly synthesized
lipid classes and individual lipid classes was determined in seedsgerminated under the same conditions with labeled acetate(Table II). The onset of lipid synthesis from 1-14(C-acetateshowed a lag period of 9 hr after treatment (Fig. 1). Themajority of the radioactive fatty acids in germinating seed werepresent in phospholipids (68%). The second highest radioactive
4 Abbreviations: CE: cerebrosides; DGDG: digalactosyl diglycerides;ESG: esterified sterol glucosides; LPC: lysophosphatidyl choline; LPE:lysophosphatidyl ethanolamine; MGDG: monogalactosyl diglycerides;PA: phosphatidic acid; PC: phosphatidyl choline; PE: phosphatidylethanolamine; PG: phosphatidyl glycerol; PI: phosphatidyl inositol; PS:phosphatidyl serine; SG: sterol glucoside; SL: sulfolipids; U: unknown.
731Plant Physiol. Vol. 60, 1977
www.plantphysiol.orgon August 28, 2018 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.

MUSLIH AND LINSCOTP
Table II. Distribution of C in lipid fractions of soybean.The seeds were germinate4 for 24 hr at 25C in 0.5 ml
water containing 0.25 bCi 1-1 C-acetate.
14Lipid Class C Distributionby Class1 <16 16:0 18:0 18:1 18:2 18:3 >18
2 ~~~~~~~~%
Neutral lipids 25.6 8 38 34 15 3 2 0Phospholipids 68.3
PC 8.2 n 60 37 1 0 0 2PE 12.7 0 55 45 0 0 0 tr
PG,PI,PA 44.8 0 58 39 0 0 0 3Glycolipids 6.1 0 91 9 0 0 0 tr
MGDG 3.9DGDG 1.2
Based on 1.48 X 105 dpo/g total lipid.
20
IS0x
0.i-I
0
0
&
5 10 15 20INCUBATION, HOURS
25 30
FIG. 1. Time course of lipid synthesis initiation in germinating seedsas measured by 1-_4C-acetate incorporation. Each of 10 seeds wasgerminated for 24 hr at 25 C in 0.5 ml of water containing 0.25 ,uCi of1-_4C-acetate.
fraction was the neutral lipids (26%). Sulfolipids and glycolipidswere found also to be highly labeled. The distribution patternof newly synthesized fatty acids was different from that ofendogenous fatty acids in mature seeds. Mature soybean seedscontain about 65% unsaturated fatty acids as their food reserve(10). In germinating seeds, only 18% of the newly synthesizedfatty acids were unsaturated and the bulk of radioactivity wasfound in palmitic and stearic acids (Table II). Our results aresimilar to those of Harwood and Stumpf (4) for lipid synthesisin pea seeds germinating in the presence of 14C-acetate. How-ever, in safflower and castor bean they detected labeled oleicand linoleic acids during the early stages of germination (4).The localization of the different enzyme activities associated
with the fatty acid-synthesizing system in germinating soybeanseeds was tested by using various cell-free fractions and sub-strates (Table III). The mitochondrial fraction incorporatedacetate, acetyl-CoA, and malonyl-CoA but not malonate. Thesefindings indicate that the mitochondrial fraction lacks malonatethiokinase. The soluble fraction incorporated all four substratesmore efficiently than the mitochondrial fraction. Malonyl-CoAwas the most efficient substrate, which indicates that the solublefraction was richer in fatty acid synthetase than the mitochon-drial fraction. The microsomal fraction incorporated acetate,acetyl-CoA, and malonyl-CoA, but utilized malonate verypoorly. Again, malonyl-CoA was a more efficient substratethan either acetate or acetyl-CoA. These results indicate thatacetate thiokinase was associated with all three fractions. Acetyl-CoA carboxylase was also detected in all three fractions. Malo-nate thiokinase was found to be associated with the solublefraction. Clearly, all three cell fractions tested were activelyparticipating in lipid synthesis during the initial stages of soybeangermination. In developing soybean seeds most of the activitynecessary to incorporate acetate into lipids is associated withthe soluble fraction (13). Therefore, there seem to be moresites of lipid synthesis in the germinating seed than in thedeveloping seed.
Cofactor requirements were determined for substrate incor-poration into lipids by mitochondrial, soluble, and microsomalfractions. All systems responded markedly to limitations ofCoA, ATP, and NADPH (see assay medium for concentra-tions). NADH did not have the same effect as NADPH,although its absence in the incubation medium had sufficienteffect on the cell systems for it to be routinely included. Theabsence of MnCl2 caused a significant decrease in acetate andmalonate incorporation into lipids, while HC03 was found tobe essential for acetyl-CoA incorporation. The addition of ACPproduced a much higher stimulation with the soluble fractionthan was observed with the mitochondrial and microsomalfractions. These results indicate that in soybeans the cofactorrequirements for substrate incorporation into lipids do not differsubstantially from other plants (2, 7, 11).
Herbicide Effects on Lipid Synthesis. Dinoben inhibited theincorporation of 1-_4C-acetate into lipids during the first 24 hrof germination of intact seed (Fig. 2). Inhibition increased withincreased concentration of dinoben up to 67% at 20 ,ug/ml, themaximum for the experiment. Dinoben inhibited not only totallipids, but also lipid classes and their individual components(Table IV). At 20 ,ug of dinoben/ml the incorporation of 1-'4C-acetate into the neutral lipid fraction was inhibited up to 73%while incorporation into the glycolipid and phospholipid frac-
Table III. Incorporation of 1- C labeled substrates intolipids by three subcellular preparations of germinatingsoybean seeds.
Fractionmitochondrial soluble microsomal
Substrate (21,600 & P) (198,000 g S) (198,000 j P)
nmol incorporated mg protein hr
1- C-acetate 1.0 1.8 1.2
1- C-acetyl-CoA 0.9 1.7 2.3
2- C-malonate trace 2.1 0.2
2- C-malonyl-CoA 0.4 4.5 4.6
z0
I-5z
4
I-Co
* Dinobeno Dinoben incubated for 30 minutes*ChlorombenoChloromben incuboted for 30 minutes
Chloromben12 16 C0
pg/ml
FIG. 2. Effect of dinoben and chloramben, incubated 0 and 30 minbefore addition-of labeled acetate, on 1-'4C-acetate incorporation intototal lipids by germinating seeds. Percentage calculated from a base of1.43 and 1.44 x 105 dpm/g of lipid, respectively, for dinoben andchloramben treatments. Seeds were germinated for 24 hr at 25 C in 0.5ml of solution containing 0.25 ,uCi of 1-'4C-acetate.
732 Plant Physiol. Vol. 60, 1977
www.plantphysiol.orgon August 28, 2018 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.

HERBICIDAL REGULATION OF LIPID SYNTHESIS
Table IV. Inhibition of acetate incorporation into lipids bydinoben.
Soybean seeds were germinated at 25C for 24 hrin 0.5 ml water containing 0.25 uCi 1-14C-acetate.
Lipid dpm/mg lipid %class Control +20 ug/ml dinoben inhibition
Neutral lipids 36.0 9.7 73Glycolipids 9.9 4.8 51MGDG 5.6 2.3 58DGDG 1.6 1.1 32
Phospholipids 102.2 63.3 39PC 13.5 5.4 60PE 17.5 7.9 55PI,PG,PA 63.7 42.9 33
tions was inhibited by 51 and 39%, respectively. When polarlipid fractions were analyzed further, inhibitions of individuallipid classes were observed as high as 60% for phosphatidylcho-line at 20 ,ug/ml concentration. Cerebrosides of the DGDGfraction, sulfolipids of DGDG and MGDG fractions, and sterolglycosides detected in the MGDG fraction were not testedindividually. The PE fraction contained a significant amount ofLPE. LPC was also detected in small amounts in the PCfraction. Since all end products of lipid syn'thesis tested wereinhibited to various extents, it is likely that the inhibition site isassociated with fatty acid synthesis enzymes and not withcomplex lipid synthesis enzymes. This is probably related to thelimited amounts of fatty acids being synthesized rather than theactual incorporation of these acids into neutral and complexlipids. The evident decrease in the free fatty acids fractionsupports this suggestion.Chloramben stimulated the incorporation of labeled acetate
into lipids (Fig. 2). At 20 ,ug/ml the stimulation was as high as25% of control. No attempt was made to fractionate total lipidsinto lipid classes.
Figure 3a shows the inhibition pattern of incorporation oflabeled acetate into lipids by dinoben in the mitochondrial,soluble, and microsomal fractions. Inhibition of acetate incor-poration in the microsomal fraction by dinoben reached 99%(highest) of control at 20 ,ug/ml. Lipid synthesis from acetate inthe soluble fraction was inhibited up to 92% of control at 50jig/ml of dinoben. Inhibition was only 65% (lowest) in themitochondrial fraction at 50 jig/ml of dinoben. This inhibitionlevel was low in comparison with the crude preparation whichwas inhibited by 81% of control at the same concentration.The pattern and levels of inhibition of lipid synthesis from
acetate by dinoben were much different with the subcellularfractions than with intact seeds. Dinoben was a very effectiveinhibitor at low concentrations with cell-free fractions whichmay indicate that dinoben is not freely transported intercellu-larly in germinating soybean seeds.The effect of chloramben on 1-_4C-acetate incorporation into
lipids by cell-free fractions of germinating soybean seeds isshown in Figure 3b. At concentrations of up to 6 ,ig/ml, thesoluble fraction was the only system being stimulated. Atconcentrations over 6 ,ig/ml the incorporation of acetate intolipids was increased in all systems. The highest stimulation was43% with the soluble fraction at 50 ,ig/ml chloramben.When dinoben (20 ,ug/ml) and chloramben (0, 5, 10, 15, 20,
and 50 jig/ml) were tested in combination for effect on 1-'4C-acetate incorporation into lipids, no chloramben-induced differ-ences in incorporation were detected at any concentration.Chloramben did not reverse or modify the inhibitory effect ofdinoben.These results with subcellular fractions indicate that dinoben
and chloramben may affect different enzyme systems of fattyacid synthesis.
Herbicide Effects on Enzyme Systems. Up to 100 jig/ml
dinoben had no significant effect on the incorporation of 1-'4C-acetyl-CoA into lipid by the cell-free fractions. Chloramben,however, at similar concentrations, and as low as 2 ug/ml,stimulated the incorporation of acetyl-CoA into lipids in allenzyme preparations tested (Fig. 4a). The highest response was
Z 4
m
I-
a0- A
z0
4-j
(nII
0
.44I,
l0l-4
20- wo 1.0
0 o_09_°0 CHLORAMBN
4-8112 ~ ~ t 1100'Fg/mI
FIG. 3. Incorporation of labeled acetate into lipids by subcellularfractions of germinating seeds as affected by (a) dinoben and (b)chloramben. The fractions were incubated at 37 C for 2 hr in 2 ml ofsolution containing 0.2 iLCi of 1-'4C-acetate. Percentages were calcu-lated from a base for (a) of 1.92, 2.83, 2.01 x 104 dpm (3 mg ofprotein hr)-' and for (b) of 1.52, 2.41, 2.23 x 10" dpm (3 mg ofprotein hr)-', respectively, for mitochondrial, soluble, and microsomalfractions.
J-ox
I-'
n
z0
I--i
U)
pg/mIFIG. 4. Chloramben effect on the incorporation of (a) 1-14C-acetyl-
CoA, and (b) 2-"4C-malonyl-CoA, into lipids by subcellular fractions ofgerminating seeds. Percentages calculated from a base of (a) 0.5, 1.3,1.49 dpm (3 mg of protein hr)-' and (b) 2.25, 2.55, 2.50 x 104 dpm (3mg of protein hr)-', respectively, for mitochondrial, soluble, andmicrosomal fractions. Fractions were incubated 2 hr at 37 C.
733Plant Physiol. Vol. 60, 1977
www.plantphysiol.orgon August 28, 2018 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.
MUSLIH AND LINSCOTP
z
0
1
z
0
I;-
(n
jg/ml
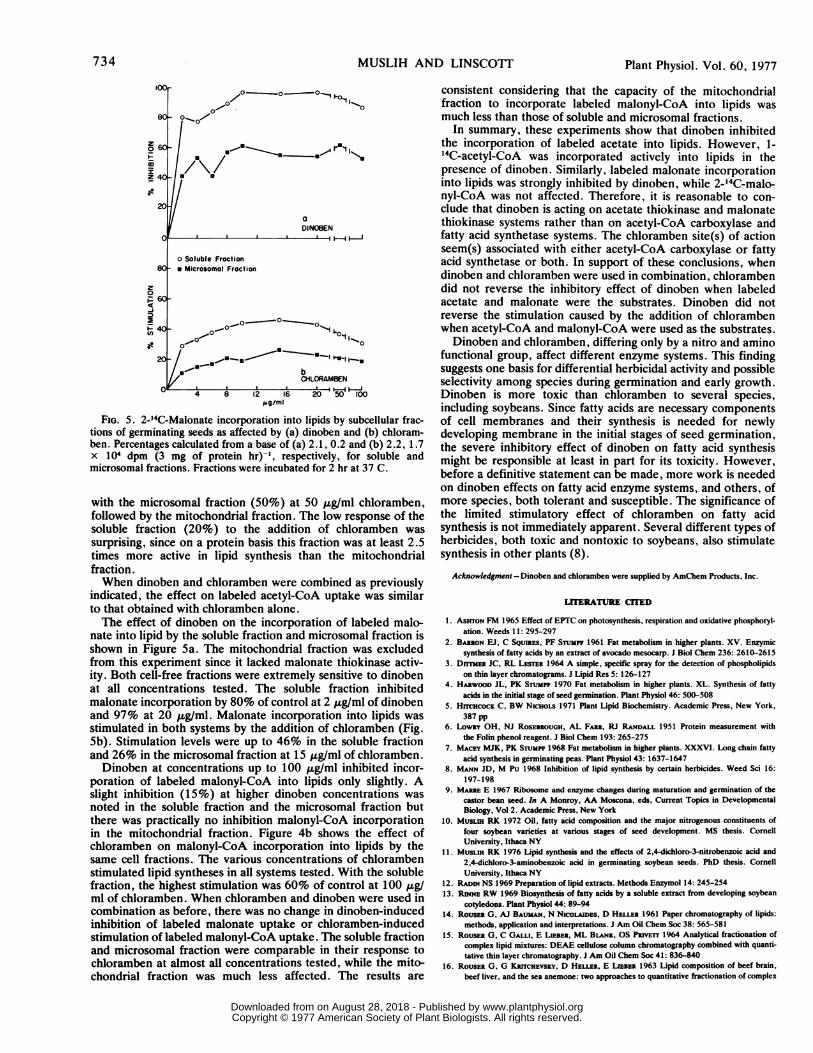
FIG. 5. 2-14C-Malonate incorporation into lipids by subcellular frac-tions of germinating seeds as affected by (a) dinoben and (b) chloram-ben. Percentages calculated from a base of (a) 2.1, 0.2 and (b) 2.2, 1.7x 104 dpm (3 mg of protein hr)-', respectively, for soluble andmicrosomal fractions. Fractions were incubated for 2 hr at 37 C.
with the microsomal fraction (50%) at 50 ,ug/ml chloramben,followed by the mitochondrial fraction. The low response of thesoluble fraction (20%) to the addition of chloramben was
surprising, since on a protein basis this fraction was at least 2.5times more active in lipid synthesis than the mitochondrialfraction.When dinoben and chloramben were combined as previously
indicated, the effect on labeled acetyl-CoA uptake was similarto that obtained with chloramben alone.The effect of dinoben on the incorporation of labeled malo-
nate into lipid by the soluble fraction and microsomal fraction isshown in Figure 5a. The mitochondrial fraction was excludedfrom this experiment since it lacked malonate thiokinase activ-ity. Both cell-free fractions were extremely sensitive to dinobenat all concentrations tested. The soluble fraction inhibitedmalonate incorporation by 80% of control at 2 ug/ml of dinobenand 97% at 20,ug/ml. Malonate incorporation into lipids wasstimulated in both systems by the addition of chloramben (Fig.Sb). Stimulation levels were up to 46% in the soluble fractionand 26% in the microsomal fraction at 15,ug/ml of chloramben.Dinoben at concentrations up to 100,ug/ml inhibited incor-
poration of labeled malonyl-CoA into lipids only slightly. Aslight inhibition (15%) at higher dinoben concentrations was
noted in the soluble fraction and the microsomal fraction butthere was practically no inhibition malonyl-CoA incorporationin the mitochondrial fraction. Figure 4b shows the effect ofchloramben on malonyl-CoA incorporation into lipids by thesame cell fractions. The various concentrations of chlorambenstimulated lipid syntheses in all systems tested. With the solublefraction, the highest stimulation was 60% of control at 100,ug/ml of chloramben. When chloramben and dinoben were used incombination as before, there was no change in dinoben-inducedinhibition of labeled malonate uptake or chloramben-inducedstimulation of labeled malonyl-CoA uptake. The soluble fractionand microsomal fraction were comparable in their response tochloramben at almost all concentrations tested, while the mito-chondrial fraction was much less affected. The results are
consistent considering that the capacity of the mitochondrialfraction to incorporate labeled malonyl-CoA into lipids wasmuch less than those of soluble and microsomal fractions.
In summary, these experiments show that dinoben inhibitedthe incorporation of labeled acetate into lipids. However, 1-'4C-acetyl-CoA was incorporated actively into lipids in thepresence of dinoben. Similarly, labeled malonate incorporationinto lipids was strongly inhibited by dinoben, while 2-'4C-malo-nyl-CoA was not affected. Therefore, it is reasonable to con-clude that dinoben is acting on acetate thiokinase and malonatethiokinase systems rather than on acetyl-CoA carboxylase andfatty acid synthetase systems. The chloramben site(s) of actionseem(s) associated with either acetyl-CoA carboxylase or fattyacid synthetase or both. In support of these conclusions, whendinoben and chloramben were used in combination, chlorambendid not reverse the inhibitory effect of dinoben when labeledacetate and malonate were the substrates. Dinoben did notreverse the stimulation caused by the addition of chlorambenwhen acetyl-CoA and malonyl-CoA were used as the substrates.Dinoben and chloramben, differing only by a nitro and amino
functional group, affect different enzyme systems. This findingsuggests one basis for differential herbicidal activity and possibleselectivity among species during germination and early growth.Dinoben is more toxic than chloramben to several species,including soybeans. Since fatty acids are necessary componentsof cell membranes and their synthesis is needed for newlydeveloping membrane in the initial stages of seed germination,the severe inhibitory effect of dinoben on fatty acid synthesismight be responsible at least in part for its toxicity. However,before a definitive statement can be made, more work is neededon dinoben effects on fatty acid enzyme systems, and others, ofmore species, both tolerant and susceptible. The significance ofthe limited stimulatory effect of chloramben on fatty acidsynthesis is not immediately apparent. Several different types ofherbicides, both toxic and nontoxic to soybeans, also stimulatesynthesis in other plants (8).
Acknowledgment -Dinoben and chloramben were supplied by AmChem Products, Inc.
LIMRATURE CITED
1. ASHrON FM 1965 Effect of EPTC on photosynthesis, respiration and oxidative phosphoryl-ation.Weeds 11: 295-297
2. BARON EJ, C SQuass, PF STrUvs 1961 Fat metabolism in higher plants. XV. Enzymicsynthesis of fatty acids by an extract of avocado mesocarp. J Biol Chem 236: 2610-2615
3. DrrmEaJC, RL LEssE 1964 A simple, specific spray for the detection of phospholipidson thin layer chromatograms. J Lipid Res 5:126-127
4. HARwooD JL, PK STusm 1970 Fat metabolism in higher plants. XL. Synthesis of fattyacids in the initial stage of seed germination. Plant Physiol 46: 500-508
5. HImcocK C, BW NxCHoLs 1971 Plant Lipid Biochemistry. Academic Press, New York,387 pp
6. Loway OH, NJ ROSEBROUGH, AL FAi, RJ RANDALL 1951 Protein measurement withthe Folin phenol reagent. J Biol Chem 193: 265-275
7. MACsY MJK, PK STuMrm 1968 Fat metabolism in higher plants. XXXVI. Long chain fattyacid synthesis in germinating peas. Plant Physiol 43: 1637-1647
8. MAmN JD, M Pu 1968 Inhibition of lipid synthesis by certain herbicides. Weed Sci 16:197-198
9. MARE E 1967 Ribosome and enzyme changes during maturation and germination of thecastor bean seed. In A Monroy, AA Moscona, eds, Current Topics in DevelopmentalBiology, Vol 2. Academic Press, New York
10. MuSLI RK 1972 Oil, fatty acid composition and the major nitrogenous constituents offour soybean varieties at various stages of seed development. MS thesis. CornellUniversity, Ithaca NY
11. MusLns RK 1976 Lipid synthesis and the effects of 2,4-dichloro-3-nitrobenzoic acid and2,4-dichloro-3-aminobenzoic acid in germinating soybean seeds. PhD thesis. CornellUniversity, Ithaca NY
12. RADIN NS 1969 Preparation of lipid extracts. Methods Enzymol 14: 245-25413. RnNP RW 1969 Biosynthesis of fatty acids by asoluble extract from developing soybean
cotyledons. Plant Physiol 44: 89-9414. RousesG, AJ BAUMAN, N NCoLomDEs, D HELsEa 1961 Paper chromatography of lipids:
methods, application and interpretations.J Am Oil Chem Soc 38: 565-58115. Rouses G, C GALL, E LiBEa, ML BLANK, OS Pavwm 1964 Analytical fractionation of
complex lipid mixtures: DEAE cellulose column chromatography combined with quanti-tative thin layer chromatography.J Am Oil Chem Soc 41: 836-840
16. ROUSER G,G KRrCHEVSsY, D HELLER, E LisasB 1963 Lipid composition of beef brain,beef liver, and the sea anemone: two approaches to quantitative fractionation of complex
734 Plant Physiol. Vol. 60, 1977
www.plantphysiol.orgon August 28, 2018 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 60, 1977 HERBICIDAL REGULATION OF LIPID SYNTHESIS 735
lipid mixtures. J Am Oil Chem Soc 40: 425-454 573-58117. ROUSER G, G KEITCHEVSKY, G SIMON GJ NELSON 1967 Quantitative analysis of brain 20. SVENNERHOLM L 1956 The quantitative estimation of cerebrosides in nervous tissue. J
and spinach leaf lipids employing silicic acid column chromatography and acetone for Neurochem 1: 42-46elution of glycolipids. Lipids 2: 37-40 21. TRANs FG, RO BAODY 1960 The synthesis of malonyl-'4C-coenzyme A. J Am Oil Chem
18. SHttKl RF PENTERSON, M BARcLAY 1962 Separation of phosphatidyl ethanolamine, Soc 82: 2972-2973phosphatidyl serine, and other phospholipids by thin-layer chromatography. J Lipid Res 22. Wells MA, JC DrrrIER 1963 The use of Sephadex for the removal of nonlipid contaminants3: 467-470 from lipid extracts. Biochemistry 2: 1259-1263
19. SIMONi RD. RS CRDDLE, PK SrunmpF 1967 Fat metabolism in higher plants. XXXI. 23. WUTHmR RE 1966 Purification of lipids from nonlipid contaminants on Sephadex beadPurification and properties of plant and bacterial acyl carrier proteins. J Biol Chem 242: columns. J Lipid Res 7: 558-567
www.plantphysiol.orgon August 28, 2018 - Published by Downloaded from Copyright © 1977 American Society of Plant Biologists. All rights reserved.


















