
Principles of energetics and stability in human locomotion · Principles of energetics and...
18
Handbook of Humanoid Robotics 1 Principles of energetics and stability in human locomotion Jeremy D Wong 1 , J Maxwell Donelan 1 People are skilled walkers and runners. We move with economy, agility and speed, and can do so even while we travel through our rough and variable world. Present- day humanoid robots are certainly much less capable than humans at accomplishing the same locomotor tasks (1). One potential path to improving the design and control of robots is to draw inspiration and guidance from biology. That is, we may be able to build more capable robots if we better understand how people move. One argument against this possibility is that humans and robots are comprised of fundamentally different components. Where robots are built using metals, encoders, wires, computers, magnetic motors, and batteries, humans have evolved to use bone, sensory cells, nerves, brains, muscles and food. While humans and robots have quite different components, these components are assembled into a common system-level design. Both humans and humanoid robots employ a skeletal system comprised of relatively rigid segments connected by relatively low friction joints. Actuators power movements by applying forces between segments. Sensors make measurements that help estimate the configuration of the segments, the forces acting on them, where they are with respect to the outside world, and the state of the world itself. Control systems interpret the sensor signals and implement the policy that commands the actuators to move the body. And power systems supply energy to the working actuators, sensors, and computers. Given that there are both similarities and differences, what types of insights into robot design should we expect to gain from studying humans? Here it is helpful to consider the overlap between robots and humans at three different levels of analysis (2). The highest level concerns their objectives, the middle level focuses on the principles employed to meet these objectives, and the lowest level concerns the physical implementation of these principles. Restricting our high-level analysis to locomotion, we suspect that humanoid robots either already share the same objectives as their human counterparts, or we might learn how to improve their design by studying objectives of human locomotion not currently embodied in robots. In contrast, and as we noted earlier, humans and robots use strikingly different hardware leaving perhaps little insight to be gained into 1 Dept. of Biomedical Physiology and Kinesiology, Simon Fraser University, Burnaby BC, V5A 1S6 improving robot design by mimicking component level physiology. Given these shared objectives and different underlying hardware, the most fruitful insights into robot locomotion are most likely reached with an analysis focused on the middle-level principles of human locomotion. Our goal in this chapter is to identify and explain central principles underlying why people walk and run the way they do. We consider principles that span biomechanics, energetics and control. Despite our breadth of scope, or perhaps even because of it, we don’t intend this chapter to be an exhaustive review. Instead, we focus on principles of energetics and stability that we suspect are most relevant for designing and controlling new humanoid robots, or aiding in the understanding of existing ones. But human locomotion is not just about energetics and stability. Stability, for example, is only one aspect of locomotor robustness, and principles of energetics and stability don’t at all explain how humans achieve their locomotor agility. We nevertheless narrowly focus on energetics and stability as that is where our expertise lies. We endeavor to emphasize principles that are grounded in both theory and experimental evidence, as these principles have the strongest foundation. While there are many non-human animal studies that have greatly informed our understanding of human locomotion, here we focus on biped theory and human experiments to keep the chapter length manageable. In writing reviews such as these, one has to make choices about the intended readership: we have aimed this chapter at roboticists that haven’t had much experience with the physiological sciences. An important caveat: the principles of human locomotion that we are about to describe should be viewed as a guide more than a formula. It is unlikely that the solutions favoured in our human physiology are the only solutions, and alternatives may in fact be better for a robot, especially given its hardware differences. But guidance is perhaps more necessary than one might initially think because it is only possible for a roboticist to evaluate a very small part of their very large design and control spaces within realistic amounts of time. Studying evolution’s solutions helps constrain the huge numbers of choices available to a roboticist to
Transcript of Principles of energetics and stability in human locomotion · Principles of energetics and...

HandbookofHumanoidRobotics 1
Principles of energetics and stability in human locomotion JeremyDWong1,JMaxwellDonelan1Peopleareskilledwalkersandrunners.Wemovewitheconomy,agilityandspeed,andcandosoevenwhilewetravel through our rough and variableworld. Present-day humanoid robots are certainly much less capablethanhumansataccomplishingthesamelocomotortasks(1). One potential path to improving the design andcontrol of robots is to draw inspiration and guidancefrom biology. That is, we may be able to build morecapable robots if we better understand how peoplemove. One argument against this possibility is thathumans and robots are comprised of fundamentallydifferent components. Where robots are built usingmetals, encoders, wires, computers, magnetic motors,andbatteries,humanshaveevolvedtousebone,sensorycells,nerves,brains,musclesandfood.While humans and robots have quite differentcomponents, these components are assembled into acommon system-level design. Both humans andhumanoidrobotsemployaskeletalsystemcomprisedofrelatively rigid segments connected by relatively lowfrictionjoints.Actuatorspowermovementsbyapplyingforcesbetweensegments.Sensorsmakemeasurementsthathelpestimatetheconfigurationofthesegments,theforcesactingonthem,wheretheyarewithrespecttotheoutsideworld,andthestateoftheworlditself.Controlsystemsinterpretthesensorsignalsandimplementthepolicy that commands theactuators tomove thebody.And power systems supply energy to the workingactuators,sensors,andcomputers.Given that there are both similarities and differences,whattypesofinsightsintorobotdesignshouldweexpectto gain from studying humans? Here it is helpful toconsider the overlap between robots and humans atthree different levels of analysis (2). The highest levelconcernstheirobjectives,themiddlelevelfocusesontheprinciples employed tomeet these objectives, and thelowest level concerns the physical implementation ofthese principles. Restricting our high-level analysis tolocomotion, we suspect that humanoid robots eitheralready share the same objectives as their humancounterparts, orwemight learn how to improve theirdesignbystudyingobjectivesofhumanlocomotionnotcurrently embodied in robots. In contrast, and as wenotedearlier,humansandrobotsusestrikinglydifferenthardwareleavingperhapslittleinsighttobegainedinto1Dept.ofBiomedicalPhysiologyandKinesiology,SimonFraserUniversity,BurnabyBC,V5A1S6
improvingrobotdesignbymimickingcomponent levelphysiology.Giventhesesharedobjectivesanddifferentunderlying hardware, the most fruitful insights intorobot locomotion are most likely reached with ananalysisfocusedonthemiddle-levelprinciplesofhumanlocomotion.Ourgoalinthischapteristoidentifyandexplaincentralprinciplesunderlyingwhypeoplewalkandrunthewaytheydo.Weconsiderprinciplesthatspanbiomechanics,energeticsandcontrol.Despiteourbreadthofscope,orperhapsevenbecauseofit,wedon’tintendthischapterto be an exhaustive review. Instead, we focus onprinciplesofenergeticsandstabilitythatwesuspectaremost relevant for designing and controlling newhumanoid robots, or aiding in the understanding ofexisting ones. But human locomotion is not just aboutenergeticsandstability.Stability,forexample,isonlyoneaspect of locomotor robustness, and principles ofenergeticsandstabilitydon’tatallexplainhowhumansachieve their locomotor agility. We neverthelessnarrowly focus on energetics and stability as that iswhere our expertise lies. We endeavor to emphasizeprinciples that are grounded in both theory andexperimental evidence, as these principles have thestrongestfoundation.Whiletherearemanynon-humananimal studies that have greatly informed ourunderstandingofhuman locomotion,herewe focusonbiped theory and human experiments to keep thechapter lengthmanageable. Inwriting reviews suchasthese, one has to make choices about the intendedreadership: we have aimed this chapter at roboticiststhathaven’thadmuchexperiencewiththephysiologicalsciences.An important caveat: the principles of humanlocomotion that we are about to describe should beviewedasaguidemorethanaformula.Itisunlikelythatthesolutionsfavouredinourhumanphysiologyaretheonlysolutions,andalternativesmayinfactbebetterfora robot, especially given its hardware differences. Butguidance is perhaps more necessary than one mightinitiallythinkbecauseitisonlypossibleforaroboticisttoevaluateaverysmallpartof theirvery largedesignand control spaces within realistic amounts of time.Studyingevolution’ssolutionshelpsconstrainthehugenumbers of choices available to a roboticist to

HandbookofHumanoidRobotics 2
combinationsthatmightactuallywork.Afterall,nature’sexemplarsaretheonlyideasthatwehavethataren’tourown, and unlike our own ideas, we know Nature’ssolutionswork(3).BipedallocomotionisadefiningtraitofhumansTherewasa timewhenourancestorsweren’tbipedal.Theseanimals—thecommonancestorbetweenhumansand apes— primarily moved around by climbingbetweentreesandwhen theymovedoverground, theyemployed all four limbs. The clearest evidence of theoriginofbipedalismcomesfromhominidfossilsthatareabout4millionyearsold,althoughitpossiblyoriginatedmillions of years sooner (4). This bipedalism plays acentral role in our human origins—evidence suggeststhatitonlyevolvedoncewithinwhatwenowrecognizeasmany hominid lineages, and it occurredmillions ofyears before our ancestors acquired distinctly humantraitslikelargebrains,longlife-spans,language,tooluse,andfire(5,6).Thesemillionsofyearsofnaturalselectionhave adapted the human locomotor system in manyways, including straightening leg posture, lengtheningthe legs, increasingenergystoragecapacity in the feet,andchangingtherelativesizesofmusclegroups(7,8).Not only has human locomotion been shaped byevolution,butwealsotuneitoveralifetimeofpractice.The typical human baby takes its first steps around 1yearofage,andquicklyrampsuptotakinganaverageofabout14,000stepsperdaywhilecoveringaround5kmof ground (9). At first, walking is at the boundary ofinfants’capabilities—theyfallmorethan100timesperdayonaverage(9).Butskillrapidlyimproves—walkingkinematicsresembletheadultformbyaboutfouryearsofage,andfallsbecomeveryrare,eveninolderadults(10).Thefrequencyofwalkingismaintainedovermuchofthelifespan,withadultsaveragingabout6,000-10,000steps per day (11). These daily step counts are nottypicallyaccomplishedwithafewperiodsoflongwalks,but instead are dominated by a large number of veryshortdurationbouts(11-13).Wearemovingaround—ratherthanstanding,sitting,orlyingdown—forabout8hourseachday.Andthismovementdemandsaboutone-thirdofourdailyenergyrequirements,andabout95%of the energy we use above that required to simplysustainlife(14).Whiletheamountofwalkingdeclinesinolderadults(15),wehaveloggedabout300milliontotalstepsbythetimewetakeourlastone.Acentralobjective forhuman locomotion is tominimizeenergeticcostAn important performance metric for walking andrunning is therateatwhich thebody, includingall theactive muscles, consumes chemical energy. This isreferred toasmetabolicenergyexpenditure,orsimply
energetic cost. While it is not technically possible tomeasure the energy consumed by each individualmuscle,wecanmeasurethetotalenergyconsumptioninafewdifferentways(16).Themostcommonmethod—termed indirect calorimetry for historical reasons—measurestheairwebreatheinandouttodeterminethevolumeofoxygenconsumedbyourbody.Ourbodiesusethisoxygentoliberatetheenergystoredinthechemicalbondsoffatandothermolecules,inthesamewaythatacampfire uses oxygen to release the energy stored inwood.Thereisafixedrelationshipbetweenthenumberof oxygenmolecules required tobreak eachbond, andtheamountofenergyreleasedwitheachbreak.So,wecansimplycountthedifferencebetweenthenumberofoxygenmoleculesthatgointothebodyandthenumberthat leave to determine the body’s chemical energyconsumption. It is a little more complicated thanconveyed in the previous few sentences because thebodyhasdifferentstoredfueltypes—fats,carbohydratesand proteins—and the energy liberated per oxygenmolecule depends on the fuel type. Fortunately, theamountofcarbondioxidereleaseddiffers foreachfueltype, so we can increase our accuracy in estimatingenergetic cost by also measuring carbon dioxideproduction(17).Nevertheless,anaccurateruleofthumbisthat21Joulesofchemicalenergyisliberatedforeachmilliliter of oxygen consumed when the bodymetabolizesatypicalmixofitsfuels(17).The energy required forwalking and runningdependsuponhowthetaskisaccomplished.Atfasterspeeds,forexample,walkingor runningdemandsahigher rateofenergeticcost(measuredinenergy(J)perunittime(s)andperunitmass(kg))thanwhenmovingmoreslowly(Figure 1). For walking, the rate of energetic costincreasesmonotonicallyand isroughlyproportional tothe square of speed (18, 19). For running, it increaseslinearly with speed (20). Rather than express theenergetic cost as energyusedper unit time, it is oftenpreferredtoexpressitasenergyusedperunitdistance.This is probably a more relevant metric for humanbehaviourbecauseweusuallyhavetogetfromoneplacetoanotherratherthansimplywalkorrunforacertainperiod of time. In walking, the energetic cost per unitdistance—termed cost of transport—is bowl-shapedwith the minimum cost at an intermediate walkingspeed.Thiscostbowlisquiteflataroundtheminimum—forchanges inspeedof10%weexpect thatpeople,onaverage, pay an energetic penalty of only about 1.5%(21). The minimum has a dimensionless value ofapproximately 0.3, when the measured energy isnormalized for the distance travelled and the bodyweightoftheindividual.Thiseconomyisaboutequaltothatmeasured for a similar-size quadrupedmoving atthesamespeed(22)andaround10timeslowerthanthat

HandbookofHumanoidRobotics 3
estimated for Honda’s Asimo humanoid robot (23). Inrunning,thecostoftransportisnearlyindependentof
speed(19)(Figure1).Thissurprisingresultmeansthatwhile therateofenergyexpenditure ismuchhigheratfastspeeds,itisnearlyexactlybalancedbyshortertotaltimethemuscleshavetobeactivelyconsumingenergy—the sameamountof energy isneeded to cover a givendistancenomatterifitisrunfastorslow.Importantly,energeticcostdoesn’tjustdependonspeed.Itisstronglyaffected by step frequency, stepwidth, degree of kneeflexion,and injury, toname justa few(24-26). Indeed,almostany change tohowapersonwalksor runswillchangethewaymusclesareusedandthushaveaneffectontheresultingenergeticcost.People prefer to walk and run in ways that minimizetheirenergeticcost(Figure2).Forexample,wetendtomoveataspeedthatminimizesourcostoftransport(27-30).Andatagivenspeed,weselectthebestgait—beitwalking or running—to approximately minimizeenergetic cost (31-33). People also consistently selectthe appropriate step frequency, step length and stepwidththatkeepscostminimized(25,34,35).Thisisnottosaythattherearen’tothergoalsduringlocomotion.Aparticularspeedmayberequiredifoneistryingtocatchabus,forexample,whilestabilityisnecessarytoavoidfalls.Yet,withintheseconstraints,energyminimizationappears to be paramount. Indeed, there are no knownelective changes that we can make to our locomotionbiomechanicsthatwilldecreaseourenergyexpenditurerelativetotheirpreferredgait(36).Our ability to find economical gaits reflects processesoperating on different timescales. On the longesttimescale, evolution appears to have favouredmorphologiesthatmadeourancestorsmoreeconomical(37).Movementcomprisesasubstantialfractionofourdaily energy needs (38, 39), and adaptions for moreeconomical movements would have meant less timerequiredtosearchforfood,leavingmoretimeavailableforotherfactorsthataffectevolutionaryfitnesssuchashiding from predators, reproduction, and raisingoffspring(40).Overshortertimescales,oureconomicalgait might also be the product of the tuning of ourneuromuscular physiology over developmental time.Indeed,ourgaitbiomechanicsmatureoverthecourseofchildhood (12), and are mirrored by systematicdecreasesinthecostoftransport(41).Ontheshortesttimescales, our nervous systems employ real-timeoptimization to find energetically optimal controlstrategies.Wethinktheclearestevidenceofthiscomesfrom our own studies that have used exoskeletons toreshapeenergycostasafunctionofaperson’sgait(42).After the exoskeletons present the wearer with newrelationships betweenwalkingmovements and energyexpenditure, people rapidly adjust theirmovements toconvergeonthenewenergeticallyoptimalgait,anddo
walk
0 1 2 3 4 5Speed (m/s)
0.2
0.4
0.6
0.8
1
1.2
1.4
Cos
t of t
rans
port
0 1 2 3 4 5Speed (m/s)
0
5
10
15
20
25M
etab
olic
pow
er (J
/s/k
g)
run
Figure1.Metabolicdataforwalkingandrunning.(A)Metabolicpower of walking and running. For running, the metabolicpowerislinearlyrelatedtospeed(blue).Themetabolicpowerofwalkingincreasesnon-linearlywithwalkingspeed.(B)Thecostof transportof runningandwalking. Costof transport ismade dimensionless by dividing energetic cost (J) by thedistance travelled andbodyweight. For running it decreasesrapidly below speeds of 2 m/s, and remains roughlyindependent of speed thereafter. The cost of transport forwalkinghasaminimumnearthepreferredwalkingspeed(~1.5m/s),andexceedsthecostofrunningat~2.3m/s.Thewalkingdata and slow runningdata (< 3m/s, filled circles) are datafrom unpublished experiments at University of California,Berkeley.Thehigh-speedrunningdata(opensquares)arefromMayhewetal.(60).Errorbarsdenotestandarddeviations.Thepolynomialfitlinesareforillustrativepurposesonly.

HandbookofHumanoidRobotics 4
soforremarkablysmallsavings.Thenervoussystemisbothrapidandfrugalinitsdesiretominimizeenergeticcost.Walking and running employ distinct underlyingmechanismsAlmost no net mechanical work is required whenwalking or running over level ground at a constantspeed.Thisisbecauseifthegroundishard—likepackeddirt rather than sand—the legs need to perform verylittlemechanicalworkontheenvironment.Andthebody
doesn’tneed toperformnetmechanicalworkon itselfbecauseitsaveragepotentialandkineticenergyarenotchanging. This is not to say that the gravitational andpotential energy of the body don’t fluctuate within astridecycle—theydo(Figure3).But,thesefluctuationsneed not be entirely due to muscle and other tissuedissipating energy by performing negative mechanicalwork on the body, and thenmuscle restoring it usingmetabolically expensive positive mechanical work.Instead, in both walking and running, these energyfluctuations could be due to passive mechanisms that
1.5 2 2.5 3 3.5
Frequency (Hz)
0.25
0.3
0.35
0.4
0.45
0.5
0.55C
ost
of
tra
nsp
ort
0 0.5 1 1.5 2
Speed (m/s)
0.25
0.3
0.35
0.4
0.45
0.5
0.55
Co
st
of
tra
nsp
ort
1.5 2 2.5 3 3.5
Frequency (Hz)
0.25
0.3
0.35
0.4
0.45
0.5
0.55
Co
st
of
tra
nsp
ort
0 0.1 0.2 0.3 0.4
Step width (m)
0.25
0.3
0.35
0.4
0.45
0.5
0.55
Co
st
of
tra
nsp
ort
Figure2.Humansprefertomoveinwaysthatminimizeenergeticcost.Thegreylinesineachfiguredenotetheenergeticcostatthepreferred(A)speedofwalking(21),(B)stepfrequencyofwalking(24),(C)stepwidthofwalking(25),and(D)stepfrequencyofrunning(59).Errorbarsdenotestandarddeviations.Thepolynomialfitlinesareforillustrativepurposesonly.
HandbookofHumanoidRobotics 5
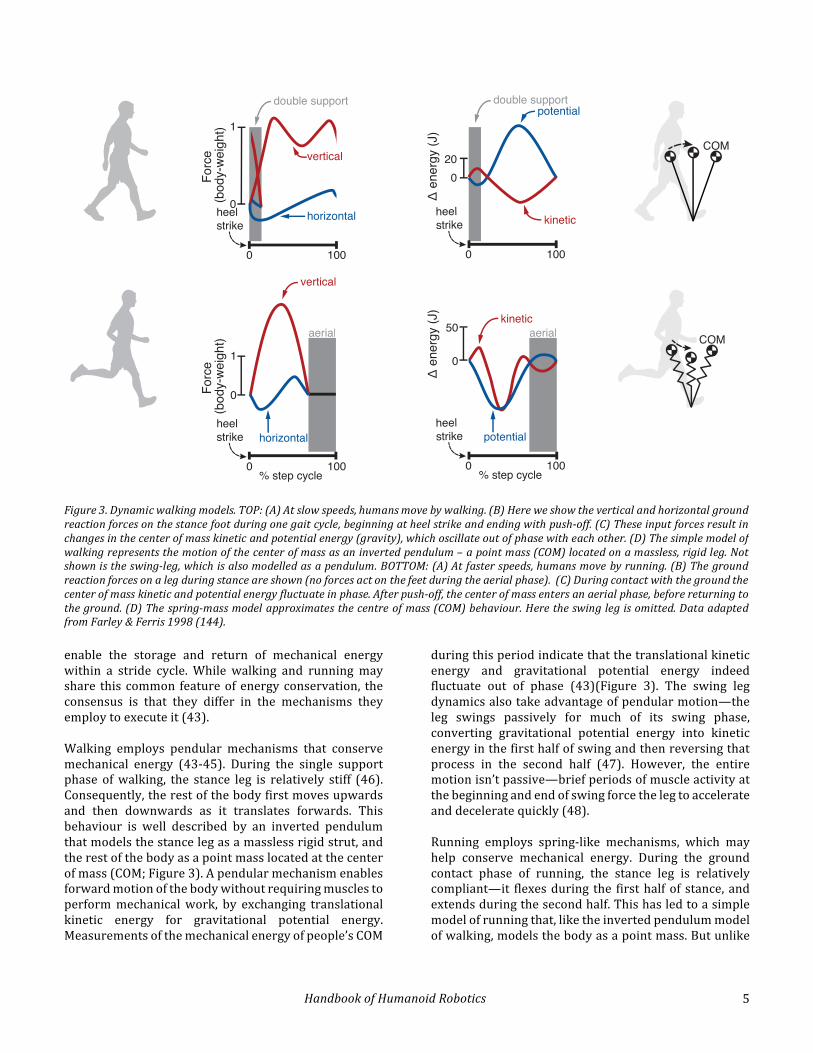
enable the storage and return of mechanical energywithin a stride cycle.While walking and running mayshare this common featureofenergyconservation, theconsensus is that they differ in the mechanisms theyemploytoexecuteit(43).Walking employs pendular mechanisms that conservemechanical energy (43-45). During the single supportphase ofwalking, the stance leg is relatively stiff (46).Consequently,therestofthebodyfirstmovesupwardsand then downwards as it translates forwards. Thisbehaviour is well described by an inverted pendulumthatmodelsthestancelegasamasslessrigidstrut,andtherestofthebodyasapointmasslocatedatthecenterofmass(COM;Figure3).Apendularmechanismenablesforwardmotionofthebodywithoutrequiringmusclestoperformmechanicalwork, by exchanging translationalkinetic energy for gravitational potential energy.Measurementsofthemechanicalenergyofpeople’sCOM
duringthisperiodindicatethatthetranslationalkineticenergy and gravitational potential energy indeedfluctuate out of phase (43)(Figure 3). The swing legdynamicsalsotakeadvantageofpendularmotion—theleg swings passively for much of its swing phase,converting gravitational potential energy into kineticenergyinthefirsthalfofswingandthenreversingthatprocess in the second half (47). However, the entiremotionisn’tpassive—briefperiodsofmuscleactivityatthebeginningandendofswingforcethelegtoaccelerateanddeceleratequickly(48).Running employs spring-like mechanisms, which mayhelp conserve mechanical energy. During the groundcontact phase of running, the stance leg is relativelycompliant—it flexesduring the firsthalfof stance,andextendsduringthesecondhalf.Thishasledtoasimplemodelofrunningthat,liketheinvertedpendulummodelofwalking,modelsthebodyasapointmass.Butunlike
kinetic
potential
potential
kinetic
vertical
horizontal
horizontal
aerial
heelstrike
heelstrike
0 1000 100
heelstrike
heelstrike
aerial
double support double support
20
50
0
0
Δ e
nerg
y (J
)
0 100% step cycle
0 100% step cycle
Δ e
nerg
y (J
)
0
1
1
0
Forc
e (b
ody-
weig
ht)
Forc
e (b
ody-
weig
ht)
verticalCOM
COM
Figure3.Dynamicwalkingmodels.TOP:(A)Atslowspeeds,humansmovebywalking.(B)Hereweshowtheverticalandhorizontalgroundreactionforcesonthestancefootduringonegaitcycle,beginningatheelstrikeandendingwithpush-off.(C)Theseinputforcesresultinchangesinthecenterofmasskineticandpotentialenergy(gravity),whichoscillateoutofphasewitheachother.(D)Thesimplemodelofwalkingrepresentsthemotionofthecenterofmassasaninvertedpendulum–apointmass(COM)locatedonamassless,rigidleg.Notshownistheswing-leg,whichisalsomodelledasapendulum.BOTTOM:(A)Atfasterspeeds,humansmovebyrunning.(B)Thegroundreactionforcesonalegduringstanceareshown(noforcesactonthefeetduringtheaerialphase).(C)Duringcontactwiththegroundthecenterofmasskineticandpotentialenergyfluctuateinphase.Afterpush-off,thecenterofmassentersanaerialphase,beforereturningtotheground.(D)Thespring-massmodelapproximatesthecentreofmass(COM)behaviour.Heretheswinglegisomitted.DataadaptedfromFarley&Ferris1998(144).

HandbookofHumanoidRobotics 6
walking, the leg is approximated as a massless springrather than a rigid strut (Figure 3). This mechanism,which is sometimes called a spring-mass model andsometimes called a spring-loaded inverted pendulum,conserves mechanical energy by storing it as elasticpotentialenergyduringthefirsthalfofstancephaseandthenreturningitduringthesecondhalf.Indeed,humanexperimentsdemonstrate that theCOM isat its lowestduringmidstance in running,butat itshighestduringmidstanceinwalking(46).Furthermore,measurementsof the mechanical energy of the body’s COM duringstanceindicatethatthetranslationalkineticenergyandgravitational potential energy fluctuate in phase witheach other, but out of phasewith the elastic potentialenergyoftheleg(18,49,50).Perhapsmostremarkably,human experiments demonstrate that the entire legbehaves like a single spring during ground contact,delivering force to the ground in proportion to itscompression.Thislegstiffnessisremarkablylinear,andexhibitsverylittlehysteresis(51).The inverted pendulum and spring-mass modeldescribed above are more conceptual and descriptivethan predictive. They do describe the measured COMtrajectoryduringwalkingandrunning,andhaveplayedan important role in conceptualizing the utility ofmechanical energy conservation in moving the bodyforwardeconomically.Theyalsodemonstratethatthereare multiple candidate mechanisms for useful energyconservation.However,humansandhumanlocomotiondon’tstrictlyconformtothesemodels.Forexample,ourbodiesaren’tpointmassesandourlegsaren’tmassless.Andduringwalking, our legs aremore compliant thanimpliedbyastrut-likeinvertedpendulum(46).Indeed,modelswithspringylegscanreproducehumangroundreaction forces and COM trajectories not only forrunning,butalso forwalking(52).Andinrunning,ourspring-like leg behaviour may be less related toconservationofelasticenergyinstructuresliketendons,andmore related to howour nervous system controlsour muscles and joints. Importantly, the originalinverted-pendulumandspring-massmodelsdon’tmakepredictionsaboutourlocomotorbehaviour,suchasourenergetic cost, our preferred biomechanics, and ourstability. Despite this major explanatory shortcoming,these models have nevertheless been central to ourunderstanding of locomotion primarily by influencingthe direction ofmore predictivemodels. For example,the idea of pendular walking has been taken to theunder-actuated extreme in the seminal work of TadMcGeer (1990)(53). Inspired by the original work ofCavagnaandMcMahon,McGeershowedthatsimulatedandphysicalmodelsofstraight-leggedandkneedrobotsarecapableofgeneratingdynamicwalkinggaits in theabsence of any actuation, save for that provided by
gravity.TheoriginalMcGeermodels,andextensionsofthem that we collectively refer to as dynamicwalkingmodels, do make predictions about why walkingrequiresenergyandcontrol,andhowmuchenergyandcontrol it requires under different circumstances. Thespring-mass model has similarly spawned usefulpredictivemodels thatharness spring-massmechanics(54-57).Wefurtherdescribemanyofthesewalkingandrunningmodelswithinthischapter,anddosowhentheybecome useful for explaining locomotion’s underlyingprinciples.WewalkatslowspeedsandrunatfastspeedsbecauseitismosteconomicaltodosoThewayswe normallywalk and run are not the onlywaysofusingour legs to get around. For example,wemight skip, hold our centre ofmass (COM) completelylevel,orlandwithstifflegswhenwerun,butweseldomdoso.Tounderstandwhywewalkandrunthewaythatwe do, Srinivasan and Ruina (2005)(58) developed asimplemodelofbipedallocomotionthatcouldperformvirtually any type of gait. They used numericaloptimization to discover which types of gait wouldminimize muscle mechanical work at each speed.Unbiased by an expectation of what the optimal gaitsmay be, the optimization algorithm converged on theclassic gaits of inverted-pendulum walking at slowspeedsandspring-likerunningatfastspeeds.Notethatin the case of running, this behaviour was observeddespitethefactthatthebipeddidnotstoreandreturnelasticenergy,indicatingthatthespring-likebehaviourof the human leg has energetic benefits beyond theconservationofmechanicalenergy.Theseoptimizationresults suggest that we walk the way we do at slowspeeds,andrun thewaywedoat fast speeds,becausethey minimize energetic cost. Experimentally, it isimpossible to compare all possible gaits. Because it ispossibletorunatallwalkingspeeds,andwalkatmanyrunning speeds, a logical comparison is between thesetwo gaits at the same speeds. Measurements of theenergetic costofwalkingandrunning (Figure1) showthatitismoreexpensivetorunthanwalkatslowspeeds,whileatfastspeedsitismoreexpensivetowalkthanrun(21,24,25,59,60).And,peopleprefertoswitchbetweenthe two gaits at about the speed where it becomescheaper to use the other gait (32). A second logicalcomparison is to gaits where the center of masstrajectoryiskeptflatandmovingataconstantspeed,likethe center of mass motion in a rolling wheel. This issimilartothegaitsmadefamousbytheunlikelyduoofGrouchoMarx, the actor, andAsimo, the robot. Such agaitcanbeusedatbothwalkingandrunningspeeds,butin either case, there is a very large energetic penaltyrelative to typical walking and running gaits (61, 62).These results demonstrate that it is energetically

HandbookofHumanoidRobotics 7
disadvantageous to eliminate the mechanical energyfluctuationsthatnormallyoccurwithinagaitcycle.EnergeticcostofwalkingisdominatedbytransitioncostsandlegswingcostsOurmovements arepoweredbymuscles.Ourmusclescan behave like motors, generating forces whileshortening by converting stored chemical energy intopositivemechanicalwork that liftsandacceleratesourbodies.Thischemicalenergyoriginatesfromthefoodweconsume, and isoften referred to asmetabolic energy.Whenperformingpositivework,themuscle’smaximumefficiency – the ratio between mechanical workperformed to the chemical energy consumed – isapproximately25%(20).Thatis,forevery1Jofpositivemechanical work performed by muscle, it requires atleast 4 J of chemical energy. This is the maximumefficiency—it can be much less than 25% under lessfavourableconditions.Behavinglikeamotorisperhapsthemost intuitiverole formuscle,particularly ifone isusedtothinkingaboutrobotactuators.Butmusclehastwo othermechanical roles in humanmovement (55).First,itcanbehavelikeabrake,generatingforceswhileitislengtheningandconvertingmechanicalenergythatwasstoredinthemotionofourbodiesintoheat.Unlikemagneticmotors that act as generatorswhen braking,musclesconsume, rather theproduce, chemicalenergywhen performing negative mechanical work. Theenergeticefficiencyofsuchnegativeworkconditionsisapproximately-120%.Thatis,whenamuscletakes1Jofmechanicalenergyfromthebodyandturnsitintoheatthroughbraking, itconsumes0.83Jofchemicalenergytodoso(20,63).Thefinalmechanicalroleofamuscleisthatofastrutoracable,whereitgeneratesforceswhilekeeping its length constant. This active muscle stillconsumes chemical energyeven though itsmechanicalworkiszero,resultinginanefficiencyof0%.Aswewilldescribeindetaillater,abenefitofthisroleisthatitcanallow tendons and other elastic tissues to store andreturnelasticenergywhileavoidinghavingtoperformcostly positivemechanicalwork. Thus,muscle force ismost costly during shortening contractions whendelivering positive mechanical work, and less costlyunder isometric or negative work conditions (55, 64).Butnomatterhow themuscle isbeingused, anactivemusclealwaysconsumeschemicalenergy.Since walking uses pendular mechanisms to conservemechanicalenergy,whydoesitexactanenergeticcost?The answer appears to be that there are energeticconsequences to behaving like an inverted pendulum.During single support, when the body is swingingupwardlikeapendulum,thelegisbehavinglikearigidstrut.Whenalegisn’tchanginglength,thenthereisnocomponentofthecenterofmassvelocitythatactsalong
theleg—insteaditisdirectedperpendiculartotheleg.Ateachnewstep,thebodymusttransitionfromatrailingleg inverted pendulum to a leading leg invertedpendulum(53,65,66).Thus,thecenterofmassvelocitymust be redirected from being perpendicular to thetrailing leg to being perpendicular to the leading leg.People accomplish this redirection partially byperforming positive mechanical work by pushing offwith the trailing leg, and partially by performingnegativemechanicalworkbytheleadingleg.Theperiodoverwhichtheredirectionoccurs iscalledthestep-to-step transition (67). This period is modeled as aninstantaneous event inmost dynamicwalkingmodels,andtheleadinglegnegativemechanicalworkismodeledasaninelasticcollision.Butinhumans,thestep-to-steptransition is stretched out over the double supportperiod and sometimes longer (68). Experimentsdesigned to tease apart the work at the step-to-steptransitionfromothercontributorstotheenergeticcostof walking have demonstrated that transition work isresponsible forabout two-thirdsof thecostofwalkingundernormalconditions(66,69).In principle, COM velocity redirection could beaccomplished entirely with leading leg negativemechanical work. But to prevent slowing down, thisnegativeworkhastobereplacedbyanequalamountofpositivework.As longas it is in therightamount, thispositiveworkcouldbeperformedatanytimeduringthegait cycle. But, there is an energetic advantage toperformingitduringpush-off—ithelpsredirecttheCOMvelocity so that less negative work is required of theleading leg, and thus less positivework is required toreplace it.Models analyzing the sequencingofpositiveworksuggestthattheenergylostatthetransitionwithan appropriately timedpush-off canbe as little as onequarterofthatlostifthereplacementworkisperformedwith other strategies (65). In linewith this prediction,humansdoperformmostofthepositiveworkinwalkingduringthetrailinglegpushoff,andmostofthenegativeworkduringtheleadinglegcollision(68).Andifpush-off is impaired, the negative mechanical work of theleadinglegincreases,thereforerequiringmorepositiveworkduringtherestofthegaitcycleandincreasingtheenergeticcostofmovement(70,71,72).Oneapplicationfor this theory is gait assistance for amputees, byreplacingthe lostpush-offworkintheamputatedlimbwithanactuatedprosthetic.However,theeffectofaddedpush-offworkhasproducedresultsthatareachallengeto explain with existing theory. In a recent study theaddition of positive push-off work with an ankleprosthetic reduced energetic cost but not by reducingnegativework by the leading leg. Instead, the positiveworkseemstohelpreducethecostsofswingingtheleg(73).atthistime,itisunclearwhethertheseresultsare

HandbookofHumanoidRobotics 8
causedbyparticularsofamputeegait,limitedtimespentlearning to use the actuated prosthetic, or ratherrepresentashortcomingincurrenttheory.People actively swing their legs to reduce the step-to-steptransitioncost.Weretransitioncosttheonlycosttowalking,itwouldfavourwalkingwithveryshortsteps.This isbecause,at theextremestep lengthofzero, thewalkingmotionbecomesthatofawheelandnoworkisrequired to redirect the COM velocity. For a givenwalking speed, short step lengths require high stepfrequencies.Thiscanbeaccomplishedwithperiodichiptorques to entrain the leg to swing with frequencieshigherthanitsnaturalfrequency.Butactivelyswingingthe leg is not free—it is estimated to contribute aboutone-thirdofthetotalcostofwalking(36).Experimentsonhumanparticipantsindicatethatthecostoflegswingincreases steeply with increases in swing frequency,whenamplitudeiskeptfixed.Thispatternisoppositetothe step-to-step transition cost, placing the two intension—people should avoid long steps to avoid hightransitionworkandavoidshortstepstoavoidhighlegswingcost.Indeed,minimizingthesumofthetwocostsacross a range of walking speeds well predicts thepreferred relationship between walking speed, steplength and step frequency (65). And summing theirindependentenergetic costsyieldsavalue close to thetotalmeasuredenergeticcost—transitionandlegswingcostsaretheprimarycontributorstowalking’senergeticcostinhealthyadults(36).Energetic cost of running is dominated by the cost ofproducingisometricforceAswiththependularmechanismsinwalking,thereareenergeticconsequencesforthespring-likebehaviourofthestancelegduringrunning.Anatomically,theprimaryspringswithin our legs are our tendons. These tissuesarequiteeffectiveatstoringandreturningenergy,anddo so without any energetic cost (74). However, theydon’tindependentlycrosshumanlegjointsanddirectlyattachbonetobone. Instead, theyattach inserieswithmuscleasamuscle-tendonunit.Consequently,musclesmust generate force to allow the tendons to lengthenunder load and storemechanical energy. As describedearlier, this muscle force generation comes with anenergeticcost.Themuscleneednotperformexpensivemechanicalwork,however,ifenergycanberecycledinapassivetendon. Instead, itcangenerate forceandkeepitslengthconstant.Thisstillexactsanenergeticcost,butlessso than if themusclehad toperformnewpositivemechanical work (75). It is technically difficult tomeasure muscle and tendon lengths and forces inrunning humans. The datawe do have in humans andother running animals demonstrate that importantmuscles in running generate large forceswhile indeed
keepingtheir lengthconstant(76-78).Thisenablesthein-series tendons to lengthen and then shorten, firststoringmechanicalenergyandthenreturningitintotherunning gait cycle. Moreover, muscles benefit fromoperatingatthelengthforwhichtheirforceismaximal.Atthislength,thefewestnumberofmusclefibresneedto be active to generate a given force. At shorter andlonger lengths, the same total muscle force requiresmore active muscle fibres, and active muscle fibresconsume chemical energy to generate force. Thusproducing isometric force at or near the muscle’soptimum fibre length further improves runningeconomy(77-79).Theenergeticcostofisometricforcegenerationdependsonboththemagnitudeofforceandthetimeavailabletogeneratetheforce.Iftheaverageforceiskeptconstant,it is also more expensive to generate a short burst offorcefollowedbyalongperiodofinactivity,ratherthantogeneratealongburstfollowedbyashortperiodofnoforce (80). This time dependence is attributed to anumberoffactorsthatincludeanaddedcosttoturningamuscle on and off, and a greater rate of consumingchemical energy in faster muscle fibres (80-83). Thedependence onmagnitude of force and burst durationare collectively referred to as the cost of generatingforce, and they explain much of the energetic cost ofhumanrunningatdifferentspeeds(19).Thatis,runningat faster speeds results in a higher rate of energyconsumption due, in part, to the shorter durations ofgroundcontact,resultinginshorter,andmoreexpensive,burstsofmuscleactivity.Howmuchdoesitcosttoswingthelegsduringrunning?While spring-massmodels of running often ignore thedynamicsoflegswing,orassumeamasslessleg,runninghas been studied experimentally in guinea fowl whoconveniently havemuscles partitioned into those usedfor legswingandthoseused for legstance.Bloodflowestimatesintheseanimalssuggestswingingthelegcostsroughly 25% of the total energy during running (84).This is in line with estimates from human runningexperimentswhere leg swing is assistedwith externalsprings(85).Giventhatbothlegswingcostandthecostof generating force appear to penalize shorter stepperiods,whydon’tpeopleadoptlongerstepperiodsforagivenrunningspeed?Theanswerappearstobethat,likewhenwalking,peopleprefertominimizeenergeticcost during running, and step periods that are longerthanpreferredcomewithanenergeticpenalty(35,59).Butunlikewalking,wherewehaveagoodmechanisticunderstandingofthedeterminantsofenergeticcost,thesourceofthislongstepperiodpenaltyisunknown.Onecandidate explanation is that longer steps require

HandbookofHumanoidRobotics 9
increased leg compliance and thus perhaps increasedmuscleforcestosupportamorebentlegposture(86).ThefootcanactlikeaspringandawheelHumanfeetarecomplex.Theycontainonequarterofthebones inthehumanbody,morethan100muscles,andanastonishing33joints(87).Ourfootdesignisalmostcertainly not optimal—our ancestors’ feet were likelyprehensileandevolutionhasadaptedthismorphologytobeusedasourprimary interfacewith theground.Thefunction of the foot depends on whether a person iswalkingorrunning.Inwalking, itappearsthatthefoothelpstominimizestep-to-steptransitionwork.Thiswasstudied first in dynamic walkingmodels by using feetshapedlikecirculararcs(53).Itwasfoundthattheworkrequiredofeachtransitiondecreaseswith increases inthe radius of curvatureof the feet. This is because thepoint of application of the ground reaction forcetranslatesalongthefoot,reducingtheangleoverwhichthe COM needs to be redirected. Indeed, for footcurvaturesequaltoleglength,andfootlengthsequaltosteplength,theredirectionanglegoestozerorequiringnotransitionworkandtheCOMtravelsinastraightlinewithout change in speed, like the hub of a wheel.Measurementsonhumansindicatethatthemotionofthepointofgroundreactionforceapplicationalongthefootof awalkingperson iswell predictedby a circular arcshape, and that the arc has a radius of 0.3 times leglength. Why is this value optimal? Dynamic walkingmodels show that larger curvatures than 0.3with ourforward-facing and human-length feet have increasedenergy losses during step-to-step transitions. Theselosses are due to the unfavourable position of the legmass relative to thepointof collisionwith theground.Whilelongerfeet(atthesamecurvature)candecreaseredirectioncostfurther,longfeethaveadditionalcosts.Theyareheavierandthushavegreaterswingcost,andthe added mass makes them less maneuverable.Experiments that varied artificial foot shape whilemeasuringenergeticcosthavedemonstratedthatarcedfeet with curvature and lengths similar to human feetprovidemeasurableenergeticsavingsoverpointcontactwiththeground(88).Several elastic storage mechanisms distributed acrossthe entire leg allow humans to store energy duringrunning in amanner similar to the carbon fiber lowerlimbs of modern Paralympians. As the foot hits thegroundandinitiatesastanceperiodforaparticularleg,thefootitselfactsasanelasticenergymechanismasitstores energy during collision with the ground andreleasesitduringpush-off.Theforcesthatasinglefootappliestothegroundarelarge—oftenmorethanthreetimes body weight at fast running speeds (89).Measurementsofthestrainenergystoredandreleased
under compression estimate that the ligaments andothersofttissueinthehumanfeetstoreandreturn17Jperstep(90).Ananalogousstorageandreturncycleactsabout the ankle joint, as forces on the forefoot duringcollision dorsiflex the ankle and store energy in theAchillestendon.Thismechanismisthoughttostoreandreturn35Jperstep,slightlymorethandoublethatofthefoot (90).Taken together, this representsa substantialfractionofthe100Jincrementinthemechanicalenergyofthebodyduringstance.Giventhelargecontributionofthese elastic storage mechanisms, variations in theireffectivenessbetweenindividualscouldhavesignificantconsequences on individual running economy. Forexample,allelsebeingequal,theamountofstrainenergythat is stored in the Achilles tendon is inverselyproportionaltoitsmomentarm,andmeasuredmomentarms turn out to be a strong predictor of runningeconomy(91,92).Another central objective for human locomotion is toremainstableWhilewerefineourgaitstominimizeenergeticcost,itisparamountthatthesegaitsarestable.Afterall,ifyouaretoo unstable to get to where you want to go, howimportantisitthatyourattemptedgaitminimizedcost?It is reasonable to assume that one of the strongestevolutionary pressures on our ancestors’ locomotionwas placed on adaptations that stabilized gait—fallingduringlocomotionwouldhaveincreasedvulnerabilitytoinjuryorpredation,anddecreasedthechanceofcatchingpreywhenhunting.Whiletoddlersfallfrequentlywhenlearning how to walk, we almost never fall duringlocomotionashealthyadults(10).Butthischangesaswecontinue to age—older adults are typically less stable,walkingisamajorcauseoffalling,andtheconsequencesforfallingaresevere.Hipfracturesareamongthemostseriousconsequences,withmorethanonemillion fallsper year causing hip fractures in older adults, 90% ofwhichhappenduringwalking(93).Twentypercentofallhip fractures lead to deathwithin1 year (93). Clearly,stability has been important in human evolution anddevelopmentandcontinuestobeimportantinourdailylives.Researcherstypicallyusethreetypesofexperimentstostudygaitstability.First, theymeasureactual falls thattakeplaceintherealword.Thisnaturalexperimentisinmanywaysthemostdesirable,butthedataarethemostdifficulttoacquiregiventheinfrequencyoffallsandtheiroccurrence outside of the well-controlled labenvironment (93, 94). More common is the use ofexperimental paradigms that perturb walking andrunning to understand the causes of falling. In suchexperiments,researcherscanprobestabilityusingbothphysical and sensory perturbations. That is, they can

HandbookofHumanoidRobotics 10
apply external forces to the system, or they can altersensory information to test the neural systemsunderlying thecontrolof stability.Onecomplicationofthese experiments is that it can be difficult to ensurepeopledon’t adopt adifferent stabilizing strategy thanthey would have used outside of the experimentalparadigm.Peopletendtoonlyrespondnaturallytothefirst unanticipated perturbation, and then adopt adifferent and more stable strategy for subsequentperturbations (95). Finally, rather than use anexperimentalperturbation,researchershaveattemptedto analyze normal walking data to determinecharacteristics that may correlate with instability orfalls,suchasstepwidthvariability.But,thiscorrelationalapproachishardtovalidategivenhowinfrequentlyfallsoccur.Moreover,sincetheseobservedcharacteristicsofmovementareaffectedbyfeedback,theymaynotevenreflect a cause of instability. For example, is stepvariability indicative of decreased stability or ofincreasedactivecontrolviafootplacement?PhysiologicalfeedbackcontrolisrelativelyslowOnewaytostabilizeasystemiswithfeedbackcontrol,but feedback delays in human locomotion areremarkablylong.Physiologicalfeedbackcontrolinvolvessensing changes to the body’s state, and then makingadjustments to the control of muscle based on thissensoryfeedback.Oneofthefastestneuralresponseinhumansisamono-synapticstretchreflex(Figure4).Thisis the fastest feedback response because it uses thefastestnerveandmusclefibresandthesignalonlyhastopass through a single synapse. However, nerves andmuscles are quite slow—the maximum conductionvelocityalongahumannervefibreisonlyabout65m/s(96,97),andittakesabout65msfromthetimeasignalarrives at amuscle until it can generate its peak force(98). Consequently, the shortest total time betweenstimulus and response is remarkably long—about 120ms from stimulus to peak force for the muscles thatextendthehumanankle(98).Thisisaboutequaltothetotaltimethatthefootspendsincontactwiththegroundduring sprinting (89, 98). At these speeds, a stimulusoccurringatthebeginningofgroundcontactmaybeableto contribute to the response at the end of groundcontact, but it would be impossible to implementmultiplecyclesofclosed-loopfeedbackcontrol(99).Thelatencies are even more extreme in other feedbackpathways—signals from our touch receptors travelslower than from stretch receptors, and signals frompainandtemperaturereceptorstravelslowerstill(100).Therearemajoradditionaldelaysifcommandsrequirecomputationbythebrain,duetoboththeaddeddistanceover which the signal is conducted and the additionalsynapses involved in the computation.Taken together,thesedelaysmakeitdifficultforournervoussystemto
userapidfeedbackcontroltostabilizefastlocomotioninresponsetounanticipatedperturbations(101,102).
Figure4.DelaysNeuromusculardelaysinbiologicallocomotion.Here we depict the delays that result from a single-synapticreflex, which includes 6 principle delays: 1) transduction ofsensorystimuliintoaneuralsignal;2)transmissionoftheneuralsignal along a nerve returning to the spinal cord; 3) synaptictransmission of the neural signal from one neuron, across asynapse,toanother;4)transmissionoftheneuralsignaltowardthemuscle;5)transductionoftheneuralsignalarrivingattheneuromuscular junction intomuscle activity; 6) the change inmuscleactivationthatproducesachange in forcedeliveredbythemuscle.ReflexeshelpstabilizeslowlocomotionWalkingorslowrunningmayallowtimeforreflexestocontribute to gait stability throughout the gait cycle.These reflex responses used to be thought of as fixedmotor commands that were tailored to specific andsudden sensed events. For example, if cutaneousreceptors inourskindetectasharpobjectunderneathone’s foot, this signal is relayed tomotorneurons thatinitiateflexionoftheinjuredlegtoremoveitfromharmandsimultaneousextensionoftheotherlegsothatitcanbearthebody’sweight(103).Wenowknowthatthesereflexes are functionally tuned for the behaviouralcontext (104, 105). This means that the same signalsfrom cutaneous receptors on the foot elicit differentmuscularresponses,dependingonwhetherthepersoniswalking,standingorlyingdown(106).Moreover,themagnitude,timing,andevendirectionofresponsesaretunedwithindifferentphasesofthegaitcycle.Instanceforexample,sensorsthatdetectanincreaseinloadingofthe stance leg can signal the muscles of that leg toincrease output force (107). Similarly, a suddenextensionofthestanceanklebeyonditsnormalangulardeflection can trigger feedback inhibition of extensor

HandbookofHumanoidRobotics 11
muscles (108). To stabilize transitions between stanceand swing, sensors that measure hip joint angle cantrigger when a step should begin (109). To measurewhen a swing phase has ended, cutaneous receptorssignalheel contact (110).And to stabilizewalking gaitusingswinglegretraction,reflexesinthehamstringareupregulatedattheendofswing(111).Inprinciple,asetof reflexes could stabilize an entire gait cycle byintegratingsignalsfrommanydifferentafferentsensorypathways.Physics-based models of walking demonstrate thatlocomotionispossibleevenwhendrivenexclusivelybythese kinds of reflexes (112, 113). In the absence ofdescending commands, active stability relies solely onthecomparativelysimplenatureofreflexestoadaptgait.These models are stable in the face of considerablechanges in surface height and sloped terrain, withoutrequiringanyadjustmentinreflexgainsforthedifferentsurfaces. Of course, a model containing a very largenumber of reflexes might achieve stability by bruteforce—each unique state in a gait cycle could have areflex-driven muscle command, and each of thesecommandscouldevenbeoptimizedforstability.Inthatcase the resultingmodelwould achieve stable gait butwould also no longer reflect simple or necessaryproperties of biological gait. Since the models notedabovehaveused a relatively small numberof reflexes,they do serve as impressive tests of the benefits ofreflexesforstabilizinggait.MusclepropertieshelpstabilizehumanlocomotionRatherthanrelyentirelyonneuralfeedbackcontrol,ourgaitsalsotakeadvantageofbiomechanicalmechanismsforself-stabilization.Onesuchmechanismconcernsthevisco-elastic properties of activemuscle—they rapidlygenerate more force when stretched. This zero-delayintrinsicmuscleresponsetoperturbationsisreferredtoasapreflex,as itoccursbeforeeven the fastestneuralreflex (114). Some simulations suggest that preflexesendow human movement with additional intrinsicstability. In simulations of jumping, for example,whenthecommandstomusclewereunderopen-loopcontrolthe jumps were remarkably stable to initial-stateperturbations, with the muscle forces adjustingappropriately(115).Incontrast,modelsthatwereunderopen-loopcontrolof jointtorquesfailedinresponsetoeven small perturbations. In simulations of hopping,which is often viewed as zero-speed running,unpredictable changes to surface stiffness werestabilized by the passive mechanical properties ofmuscle—leg stiffnesswas adapted in a stablemannerwithoutanychangeintheneuralcommand(116).Thesesimulations accurately predict the self-stabilizingbehavior of humans when hopping during the same
unexpected changes to surface stiffness—researchersmeasuredappropriateadjustments to legstiffness thatwere too rapid to be neural in origin (117). And, inresponse to anticipated changes to surface stiffness,researchers found increases in thecommandedmuscleactivity that preceded ground contact, resulting in afeedforwardadjustmenttolegstiffness(118-120).Thatis, the nervous system doesn’t benefit from preflexivemechanismsonlywhentheyhappentoarise,butinsteadactivelyadoptscontrolstrategiestopurposefullybenefitfromtheirrapidresponses.PassivedynamicsalsohelpstabilizehumanlocomotionPassivedynamicsreferstomotionthat isgeneratedbythebodywithouttheuseofactivecontrolbythenervoussystem. Whenever a system is perturbed, there is apassive dynamic response. Remarkably, this responsesometimesactstorejecttheperturbation,endowingthesystemwithpassivedynamicstability.Inwalking,thisisperhapsmost clearly seen in dynamicwalkingmodelsthat are constrained to two-dimensional sagittal planemotion. For some set of speeds, these models arepassively stable—they do not require a controller toactively adjust the limbs to prevent falling over whengivenasmallpush.Thedynamicsaresuch thatapushthatincreasestheenergyofthewalkercausesittotakea longer step. The longer step elicits greater negativeworkbytheleadinglegatheel-strikethanthenominalcyclicgait,thusdissipatingaportionoftheenergyaddedfrom the push (53, 121). In this way, the passivedynamicscontinuestoadjuststeplengthwitheachnewstep, monotonically reducing the added energy of thepush,andcausingthewalkertoconvergetoitsoriginalgaitcycle.Aperturbationthatslowsthewalkerhastheoppositeeffect: it acts todecreasestep length, therebyreducingdissipationandallowing thewalker to regainenergy and return to its original gait cycle. That thesewalkers are passively stable to small perturbationsdoesn’tmean that they are stable to all perturbations.Likerealsystems,ifthepushishardenough,itwillfallover.Still,theregionsofstabilityarelargeenoughthatpassivedynamicstability isofpractical importance forhumangaits(122).Human walking appears to harness passive dynamicstability to assist with the control of locomotion. It isexperimentally challenging to assign responsibility forstability to either active control or passive dynamicsbecause,unlikeinpassivedynamicwalkingmodels,theyare both always at work in human locomotion. Oneexperimentaltrickmentionedpreviouslyistostudytheshortintervalafteraperturbation,withinwhichneuralcontrolistooslowtohaveaneffect.Studiesthatemploythisapproachdemonstratethatpassivedynamicsplaysan important role for the adjustment of leg stiffness

HandbookofHumanoidRobotics 12
duringrunning(116,117).BaubyandKuo(2000)(123)employedadifferentapproach—theycontaminatedthesensoryfeedbackofparticipantswalkingovergroundbyhavingthemclosetheireyes.Theauthorsreasonedthatthiswouldnegatively impact the stabilityof aspectsofgaitunder active feedback control,while leaving thosethat benefit from passive dynamic stability relativelyunaffected.Aspredictedbydynamicwalkingmodels,theresearchers found that variability in step length wasrelativelyunaffectedbytheaddedsensorynoise—theseadjustments appear to be primarily driven by passivedynamics. In a related experiment, O’Connor and Kuo(2009)(124) used virtual reality to apply visualperturbations and again found that step length wasinsensitive to thesensorydisturbances. Itappears thatpeople harness passive dynamic stability to helpstabilizetheirmotioninthesagittalplane.The stability of human walking can’t be accomplishedpurelybypassivedynamics.Three-dimensionaldynamicwalking models demonstrate that passive dynamicadjustments to step length that compensate forperturbations to system energy don’t apply to stepwidth—lateral motion in bipedal walking is passivelyunstable(125).Consequently,virtuallyanypushtothesewalkerswill cause them to topple over sideways. Thissuggeststhatlateralmotioninhumanwalkingrequiresactivecontrol(125).Walkingusesactivecontrolof footplacementtoachievestabilityIn principle, passively unstable lateral motion can bestabilizedwithactivecontrolinanumberofways(125).One approach is to lean back toward the midline,opposite the tipping direction. Another approach is torotate or pinwheel the arms to control the body’srotation, since the total angular momentum of thesystem(thebodyandthearms)isconstant,thusmovingthe body towards the middle(126, 127). Anotherapproachistogenerateankletorquetorotatethebodyabout theankles.A final approach is toadjust the footplacementfortheswingleg,therebymovingthecenterof mass back between the base of support. Modelssuggest that this foot placement strategy achieves thelargest region of stability (125), and is the mosteconomicalchoice,primarilybecausethecontrolofsteplocation takes place over the entire swing phase, thusreducingenergeticcostsrelatedtogeneratingrapidandlarge forces. It also reduces the computational cost ofstability,sincethelongdurationoverwhichtheresponseis executed leaves ample opportunity for feedbackcorrectionstocompensateforanyimprecision.Human experiments support the model predictions—peopleappeartouseactivecontroloffootplacementto
stabilize lateral motion. The previously mentionedexperimentsthattestedforactivecontrolbyperturbingsensoryfeedbackmaynothaveobservedlargeeffectsonstep length, butdid findmajor increases in stepwidthvariability(123,128).Thissuggeststhattheside-to-sideplacement of the feet is under active control thatemployssensoryfeedback.Ratherthanmakingitmoredifficult for people to stabilize walking, it is insteadpossibletoassistactivecontrolbyusingspringstoapplystabilizing lateral forces (25,129).Thisaddedstabilitydecreasedstepwidthvariabilityinbothyoungandolderparticipants,indicatingareducedneedforactivecontrolof footplacement.A further indication that lateral footplacement is under active control is that decreases instep width variability were paralleled by decreases inenergetic cost (25, 129). In healthy young people,stabilizationappearstobethethirdmaincontributortoenergetic cost (after step-to-step transition and swingleg costs), accounting for 5-10% of the total energeticcost(25,130).Andtheelevatedenergeticcostofwalkinginolderadultsappears tobeentirelyexplainedby theadditional active control that they require to stabilizetheirlateralmotion(129).Converging evidence supports the idea that stabilizingbehaviours occur within just one or two steps after aperturbation.Dynamicwalkingmodelshavebeenusedtoinvestigatethecontrollabilityofwalking–theabilityof a system to achieve a target state, typically a targetvelocity(131). Itturnsoutthat if thesystemcanreachthetargetatall—say,toregainasteady-statespeedafterbeing pushed a little bit—it can almost always do sowithin two steps. What could have been a verycomputationally expensive task to plan many futuresteps therefore collapses into a very manageable one.The effects of lateral perturbations on normal humanwalkingarealsotypicallyeliminatedbythesecondstepaftertheperturbation(132,133).Whenplanningfuturefoot placements, people typically look only two stepsahead (134), and when planning steps over obstaclespeopleusemostoftheireyemovementstoexplorethenext two step lengths of visual space(135). Thus thenumberofstepsthatachievecontrollabilityaspredictedbymodelsmatchesthenumberofstepsthatpeopleusetobothstabilizetheirgait,andgathervisualinformationtoplantheirupcomingsteps.Running stability is partially determined by swing-legretractionTo study the stability of running, researchers haveextendedspring-massmodelsofthestancelegtoincludetheswing leg.Thereareavarietyof thesemodels,butthey generally have a stance leg that behaves like aspring, and a swing leg that repositions for the nextstance phase. Some are fully dynamic models that

HandbookofHumanoidRobotics 13
accomplishrepositioningusingahipspring(56),whileothersarequasi-dynamicandassumethattheswinglegcanbemadetoreachaspecifiedstateatgroundcontact.Unlikewalkingmodels,thedynamicrunningmodelsarepassively unstable in the sagittal plane at realisticrunningspeedsandcadences(56,136).McGeer’selegantanalysisofdynamicrunningmodelsdemonstrates thatthey can be readily stabilized with once-per-stepfeedbackcontrol,similartotheuseoffootplacementtostabilize walking (56). The feasibility of intermittentfeedback control such as this is important given therelatively lengthydelays inhumannervous systems atfast running speeds. Also useful in the face of longfeedback delays is feedforward control, which canstabilize these sorts of models with a mechanismreferred to as swing leg retraction (137). In swing legretraction, a burst of hip torque actively reverses theforward direction of the swing leg prior to groundcontact. The swing leg isoftenmodelledasmovingatconstant velocity during the flight phase. Since the legretracts without feedback, the backwards angle sweptbefore contact is defined by flight time. Longer flighttimes than the nominal trajectorymake the leg swingfurtherbackward, so that theangleof the legwith theground is more vertical when the foot contacts theground. When the model leaves the ground after thiscontact phase its exit velocity is more parallel to theground.Thusthecenterofmassachievesalowerheightonthenextstride,therebyprovidingnegativefeedbackaboutthenominalrunningtrajectory.Swing leg retraction offers several benefits, includingproviding asymptotic stability, peak disturbancerejection, and improved foot placement stability (138-140).Inthisarea,thereismuchmoretheoreticalworkthan experimental tests (141). But the few controlledexperiments designed to testwhether humans employswing leg retraction as a stabilization mechanismsupport it. For example, when runners are presentedwith changes to surface height, retraction angle atground contact varies in proportion to the change inheight,andinthedirectionpredictedbytheory(142).RunnersadjustlegstiffnesstoremainstableModels of running suggest that stable running isachievablewithoutchanging legstiffness.Asdescribedearlier, variouscontrol schemescanstabilizepassivelyunstablerunninggaitseveninmodelswithaconstantlegstiffness(56,137).Furthermore,modelswithaconstantlegstiffnesscanrunacrossawiderangeofspeeds(143).Yet experiments show that people do adjust their legstiffnessinresponsetoperturbations,andcandosooverquite a large range (120). And these leg stiffnessadjustments result in a conservation of runningkinematics, preserving characteristics of gait such as
contactduration,stepfrequency,andCOMmotion(144).For example, when transitioning from a stiff groundsurfacetoacompliantone,peopleincreaselegstiffnessin such a way that the COM motion is essentiallyunchanged.Itisanopenquestionastowhythenervoussystemappearstoconservekinematics.Onepossibilityis that maintaining a nominal motion of the center ofmassreflectsafeedbackcontrolstrategybythenervoussystem to preserve the motion itself (120). Anotherpossibilityisthatreflexcontrolstrategiesthatstabilizegait operate about this nominal movement (144). Yetanotherpossibilityisthataknownnominalmotionofthecenterofmasscouldallowsensorysystemssuchasthevestibular and visual systems to operate effectively. Afurther,and, inouropinion,unlikely,possibility is thatthis adaptation is made strictly to minimize energeticcost,sinceitseemsunlikelythattheminimumcostgaitonbothstiffandcompliantgaitswouldsharethesamekinematicsofthecenterofmass.ConcludingRemarksOur goal in this chapter was to identify and explaincentralprinciplesunderlyingwhypeoplewalkandrunthe way they do. We focused on principles becausehumans and robots are comprised of very differenthardware, but both may benefit from using similaralgorithmstosolvethecommonproblemoflocomotion.Inbrief,peoplequicklylearnhowtostabilizethemselvesearly in development and rarely fall as adults. Thisstability isespeciallyremarkablebecausephysiologicalfeedback delays are relatively long. Humans overcomethese long delays using a combination of preflexes,passive stability, feedforward strategies, intermittentfeedback control and, when time allows, continuousfeedback control.While people need to be sufficientlystable to walk and run, they don’t need to maximizestability for most situations. This leaves considerableflexibility inhowpeopleachievetheir locomotorgoals.The solution for most circumstances is to minimizeenergeticcost.Thisemphasisonenergyoptimizationisevidentinouranatomy,throughtheuseofmechanismslike energy storage and return in tendons, and in ourcontrol,throughtheuseofcontrolpoliciesthatconservemechanical energy. The energetics and stabilitymechanisms employed by humans have arisen overevolutionary,developmentalandreal-timelearningtimescales. That is, adaptations for economicalmovementsand rapid stabilization are evident in human anatomy,and people learn over time to leverage their inherentbodydynamicstoimproveeconomyandstability.Althoughwearefarfromexpertsontheengineeringofhumanoid robots, it seems to us that many of theprinciplesofhumanlocomotionthatwehavedescribedhere are implementable in robots. Indeed, some

HandbookofHumanoidRobotics 14
roboticists are already actively employing similarapproaches (145-147). Importantly, the principles wedescribehereshouldbeusedasinspiration,ratherthanbeblindlycopied,assomeofthemmaybeaconsequenceofhumanlimitationswithwhichrobotsneednotsuffer.Forexample,humanadaptationstocompensateforlongfeedbackdelaysmaynotbecharacteristics thatshouldbeimplementedinhumanoidrobotsunlesstheserobotsalsosufferfromlongdelays.Ontheotherhand,thefactthathumancontrolissorelativelygoodinthepresenceof these delays suggests that such delays are not inthemselves an obstacle to control, and that highfrequencycontrolisnotanecessaryfeatureofstableandeconomicalmovement.Wesuspectthatthereisnothingparticularlyspecialabouthumanhardwarethatcan’tbematchedorexceededbyengineeredcomponents.Iftrue,human locomotion capabilities arenot only achievableby humanoid robots, but possibly exceedable. Suchrobots, we suspect, will employ some of the humanlocomotionprinciplesthatwehavedescribedheretobebothstableandeconomical.AcknowledgementsThis work was supported by a Michael SmithPostdoctoralFellowship(JDW),andanNSERCDiscoveryGrant(JMD).References1. AtkesonCG,BabuBPW,BanerjeeN,BerensonD,
BoveCP,CuiX,DeDonatoM,DuR,FengS,FranklinP,GennertM,GraffJP,HeP,JaegerA,KimJ,KnoedlerK,LiL,LiuC,LongX,PadirT,PolidoF,TigheGG,XinjilefuX.In:2015IEEE-RAS15thInternationalConferenceonHumanoidRobots(Humanoids).IEEE,2015,p.623–630.
2. MarrD.TheMITPress,1982.
3. VogelS.WWNorton&Company,1998.
4. WardCV.AmJPhysAnthropol119:185–215,2002.
5. NicholsJ.1998.
6. WranghamRW.2009.
7. LovejoyCO.GaitPosture21:113–124,2005.
8. BrambleDM,LiebermanDE.Nature432:345–352,2004.
9. AdolphKE,ColeWG,KomatiM,GarciaguirreJS,BadalyD,LingemanJM,ChanGLY,SotskyRB.PsycholSci23:1387–1394,2012.
10.O'LoughlinJL,RobitailleY,BoivinJF,SuissaS.AmJEpidemiol137:342–354,1993.
11.OrendurffMS,SchoenJA,BernatzGC,SegalAD,KluteGK.JRehabilResDev45:1077–1089,2008.
12.SutherlandDH,OlshenR,CooperL,WooSL.JBoneJointSurgAm62:336–353,1980.
13.SutherlandDH.GaitPosture14:61–70,2001.
14.LevineJA.PublicHealthNutrition8:1123–1132,2005.
15.Tudor-LockeC,CraigCL,AoyagiY,BellRC,CroteauKA,DeBourdeaudhuijI,EwaldB,GardnerAW,HatanoY,LutesLD,MatsudoSM,Ramirez-MarreroFA,RogersLQ,RoweDA,SchmidtMD,TullyMA,BlairSN.IntJBehavNutrPhysAct8:80,2011.
16.BrooksGA,FaheyTD,WhiteTP,BaldwinKM.MountainView,1996.
17.BrockwayJM.HumNutrClinNutr41:463–471,1987.
18.CavagnaGA,HeglundNC,TaylorCR.AmJPhysiol233:R243–61,1977.
19.KramR,TaylorCR.Nature346:265–267,1990.
20.MargariaR,CerretelliP,AghemoP,SassiG.JApplPhysiol18:367–370,1963.
21.BrowningRC,KramR.ObesRes13:891–899,2005.
22.TuckerVA.AmSci63:413–419,1975.
23.CollinsS,RuinaA,TedrakeR,WisseM.Science307:1082–1085,2005.
24.UmbergerBR,MartinPE.JExpBiol210:3255–3265,2007.
25.DonelanJM,KramR,KuoAD.ProcBiolSci268:1985–1992,2001.
26.WatersRL,MulroyS.GaitPosture9:207–231,1999.
27.MolenNH,RozendalRH,BoonW.ProcKNedAkadWetC75:305–314,1972.

HandbookofHumanoidRobotics 15
28.ElftmanH.JBoneJointSurgAm48:363–377,1966.
29.AtzlerE,HerbstR.PflügersArch215:291–328,1927.
30.RalstonHJ.IntZAngewPhysiol17:277–283,1958.
31.MinettiAE,ArdigòLP,SaibeneF.ActaPhysiolScand150:315–323,1994.
32.HreljacA.MedSciSportsExerc25:1158–1162,1993.
33.HreljacA.JBiomech28:669–677,1995.
34.BertramJE,RuinaA.JTheorBiol209:445–453,2001.
35.CavanaghPR,WilliamsKR.MedSciSportsExerc14:30–35,1982.
36.KuoAD,DonelanJM.PhysTher90:157–174,2010.
37.AlexanderRM.JBiomech26Suppl1:1–6,1993.
38.LevineJA.BestPractResClinEndocrinolMetab16:679–702,2002.
39.SeethapathiN,SrinivasanM.BiolLett11:20150486,2015.
40.GarlandTJr.AmericanNaturalist:571–587,1983.
41.DeJaegerD,WillemsPA,HeglundNC.PflügersArch441:538–543,2001.
42.SelingerJC,O'ConnorSM,WongJD,DonelanJM.CurrBiol25:2452–2456,2015.
43.CavagnaGA,ThysH,ZamboniA.JPhysiol(Lond)262:639–657,1976.
44.MinettiAE,AlexanderRM.JTheorBiol186:467–476,1997.
45.AlexanderRM.JExpBiol160:55–69,1991.
46.LeeCR,FarleyCT.JExpBiol201:2935–2944,1998.
47.MochonS,McMahonTA.JBiomech13:49–57,1980.
48.BasmajianJV.OrthopClinNorthAm7:1027–1029,1976.
49.RobertsTJ,AziziE.JExpBiol214:353–361,2011.
50.HeglundNC,FedakMA,TaylorCR,CavagnaGA.JExpBiol97:57–66,1982.
51.FerrisDP,LouieM,FarleyCT.ProcBiolSci265:989–994,1998.
52.GeyerH,SeyfarthA,BlickhanR.ProcBiolSci273:2861–2867,2006.
53.McGeerT.IntJRobRes9:62–82,1990.
54.GeyerH,SeyfarthA,BlickhanR.JTheorBiol232:315–328,2005.
55.DickinsonMH,FarleyCT,FullRJ,KoehlMA,KramR,LehmanS.Science288:100–106,2000.
56.McGeerT.ProcBiolSci240:107–134,1990.
57.RaibertM,ChepponisM.IEEERobotAutomat2:70–82,1986.
58.SrinivasanM,RuinaA.Nature439:72–75,2006.
59.HunterI,SmithGA.EurJApplPhysiol100:653–661,2007.
60.MayhewJL.BrJSportsMed11:116–121,1977.
61.OrtegaJD,FarleyCT.JApplPhysiol99:2099–2107,2005.
62.GordonKE,FerrisDP,KuoAD.ArchPhysMedRehabil90:136–144,2009.
63.FennWO.AmJPhysiol93:433–462,1930.
64.FennWO.58:373–395,1924.
65.KuoAD.JBiomechEng123:264–269,2001.
66.DonelanJM,KramR,KuoAD.JExpBiol205:3717–3727,2002.
67.RuinaA,BertramJEA,SrinivasanM.JTheorBiol237:170–192,2005.
68.DonelanJM,KramR,KuoAD.JBiomech35:117–124,2002.
69.KuoAD,DonelanJM,RuinaA.ExercSportSciRev33:88–97,2005.

HandbookofHumanoidRobotics 16
70.SooCH,DonelanJM.JExpBiol213:4265–4271,2010.
71.WezenbergD,CuttiAG,BrunoA,HoudijkH.JRehabilResDev51:1579–1590,2014.
72.WatersRL,PerryJ,AntonelliD,HislopH.JBoneJointSurgAm58:42–46,1976.
73.CaputoJM,CollinsSH.ScientificReports4:7213,2014.
74.MaganarisCN,PaulJP.JBiomech35:1639–1646,2002.
75.BiewenerAA,RobertsTJ.ExercSportSciRev28:99,2000.
76.RobertsTJ,MarshRL,WeyandPG,TaylorCR.Science275:1113–1115,1997.
77.LichtwarkGA,BougouliasK,WilsonAM.JBiomech40:157–164,2007.
78.LaiA,LichtwarkGA,SchacheAG,LinY-C,BrownNAT,PandyMG.JApplPhysiol118:1266–1275,2015.
79.HilberK,SunYB,IrvingM.JPhysiol(Lond)531:771–780,2001.
80.HoganMC,InghamE,KurdakSS.AmJPhysiol274:E397–402,1998.
81.RallJA.ExercSportSciRev13:33–74,1985.
82.ChasiotisD,BergströmM,HultmanE.JApplPhysiol63:167–174,1987.
83.BergströmM,HultmanE.JApplPhysiol65:1500–1505,1988.
84.MarshRL,EllerbyDJ,CarrJA,HenryHT,BuchananCI.Science303:80–83,2004.
85.ModicaJR,KramR.JApplPhysiol98:2126–2131,2005.
86.FarleyCT,GonzálezO.JBiomech29:181–186,1996.
87.PlatzerW.5ed.Thieme,2004.
88.AdamczykPG,CollinsSH,KuoAD.JExpBiol209:
3953–3963,2006.
89.WeyandPG,SternlightDB,BellizziMJ,WrightS.JApplPhysiol89:1991–1999,2000.
90.KerRF,BennettMB,BibbySR,KesterRC,AlexanderRM.Nature325:147–149,1987.
91.LeeSSM,PiazzaSJ.JExpBiol212:3700–3707,2009.
92.ScholzMN,BobbertMF,vanSoestAJ,ClarkJR,vanHeerdenJ.JExpBiol211:3266–3271,2008.
93.RobinovitchSN,EvansSL,MinnsJ,LaingAC,KannusP,CriptonPA,DerlerS,BirgeSJ,PlantD,CameronID,KielDP,HowlandJ,KhanK,LauritzenJB.In:OsteoporosInt.Springer-Verlag,2009,p.1977–1988.
94.SchwickertL,BeckerC,LindemannU,MaréchalC,BourkeA,ChiariL,HelbostadJL,ZijlstraW,AminianK,ToddC,BandinelliS,KlenkJ,FARSEEINGConsortiumandtheFARSEEINGMetaDatabaseConsensusGroup.ZGerontolGeriatr46:706–719,2013.
95.BohmS,MademliL,MersmannF,ArampatzisA.SportsMed45:1759–1777,2015.
96.BurkeD,GandeviaSC,McKeonB.JPhysiol(Lond)339:535–552,1983.
97.MarsdenCD,MertonPA,MortonHB.JPhysiol(Lond)259:531–560,1976.
98.DietzV,SchmidtbleicherD,NothJ.JNeurophysiol42:1212–1222,1979.
99.GrillnerS.ActaPhysiolScand86:92–108,1972.
100.CraigAD.NatRevNeurosci3:655–666,2002.
101.HoganN,SternadD.BiolCybern106:727–739,2012.
102.MoreHL,HutchinsonJR,CollinsDF,WeberDJ,AungSKH,DonelanJM.ProcBiolSci277:3563–3568,2010.
103.RossiA,DecchiB.JPhysiol(Lond)481(Pt2):521–532,1994.
104.ProchazkaA.ProgNeurobiol33:281–307,1989.

HandbookofHumanoidRobotics 17
105.ProchazkaA,ClaracF,LoebGE,RothwellJC,WolpawJR.ExpBrainRes130:417–432,2000.
106.ZehrEP,SteinRB.ProgNeurobiol58:185–205,1999.
107.DietzV,ColomboG.MovementDisorders13:255–261,1998.
108.SinkjaerT,AndersenJB,LadouceurM,LO,NielsenJB.JPhysiol(Lond)523Pt3:817–827,2000.
109.DietzV,HarkemaSJ.JApplPhysiol96:1954–1960,2004.
110.DietzV,GollhoferA,KleiberM,TrippelM.ExpBrainRes89:229–231,1992.
111.VandeCrommertHW,FaistM,BergerW,DuysensJ.BrainRes734:341–344,1996.
112.GeyerH,HerrH.IEEETransNeuralSystRehabilEng18:263–273,2010.
113.SongS,GeyerH.JPhysiol(Lond)593:3493–3511,2015.
114.BrownIE,LoebGE.In:BiomechanicsandNeuralControlofPostureandMovement.SpringerNewYork,2000,p.148–163.
115.vanSoestAJ,BobbertMF.BiolCybern69:195–204,1993.
116.vanderKrogtMM,deGraafWW,FarleyCT,MoritzCT,RichardCasiusLJ,BobbertMF.JApplPhysiol107:801–808,2009.
117.MoritzCT,FarleyCT.JApplPhysiol97:1313–1322,2004.
118.MoritzCT,FarleyCT.JBiomech39:1030–1038,2006.
119.MoritzCT,FarleyCT.JExpBiol208:939–949,2005.
120.FerrisDP,LiangK,FarleyCT.JBiomech32:787–794,1999.
121.GarciaM,ChatterjeeA,RuinaA,ColemanM.JBiomechEng120:281–288,1998.
122.SchwabAL,WisseM.ProcASMEDesEng6:531–
539,2001.
123.BaubyCE,KuoAD.JBiomech33:1433–1440,2000.
124.O'ConnorSM,KuoAD.JNeurophysiol102:1411–1419,2009.
125.KuoAD.IntJRobRes18:917–930,1999.
126.LibbyT,MooreTY,Chang-SiuE,LiD,CohenDJ,JusufiA,FullRJ.Nature481:181–184,2012.
127.PijnappelsM,KingmaI,WezenbergD,ReurinkG,vanDieënJH.ExpBrainRes201:689–699,2010.
128.O'ConnorSM,XuHZ,KuoAD.GaitPosture36:102–107,2012.
129.DeanJC,AlexanderNB,KuoAD.IEEETransBiomedEng54:1919–1926,2007.
130.IjmkerT,HoudijkH,LamothCJC,BeekPJ,vanderWoudeLHV.JBiomech46:2109–2114,2013.
131.ZaytsevP,HasaneiniSJ,RuinaA.In:IEEEIntConfRobotAutom.IEEE,2015,p.6295–6300.
132.HofAL,VermerrisSM,GjaltemaWA.JExpBiol213:2655–2664,2010.
133.TangPF,WoollacottMH,ChongRK.ExpBrainRes119:141–152,1998.
134.PatlaAE,VickersJN.NeuroReport8:3661–3665,1997.
135.MatthisJ,BartonS,FajenB.JVis13:121–121,2013.
136.RaibertM.1sted.TheMITPress,1987.
137.SeyfarthA,GeyerH,HerrH.JExpBiol206:2547–2555,2003.
138.HaberlandM,KarssenJ,KimS.In:IEEEIntConfRobotAutom.2011.
139.HaberlandM,KarssenJGD,KimS,WisseM.In:2011IEEE/RSJInternationalConferenceonIntelligentRobotsandSystems(IROS2011).IEEE,2011,p.3957–3962.
140.KarssenJGD,HaberlandM,WisseM,KimS.

HandbookofHumanoidRobotics 18
Robotica33:2137–2155,2015.
141.DaleyMA.CurrBiol18:R1064–R1066,2008.
142.GrimmerS,ErnstM,GüntherM,BlickhanR.JExpBiol211:2989–3000,2008.
143.McMahonTA,ChengGC.JBiomech23:65–78,1990.
144.FarleyCT,FerrisDP.ExercSportSciRev26:253–285,1998.
145.RaibertM,BlankespoorK,NelsonG,PlayterR.In:Proc17IFACWorldCongress.2008.
146.HurstJW,ChestnuttJE,RizziAA.In:IEEEIntConfRobotAutom.2007,p.1863–1868.
147.BhounsuleP,CortellJ,RuinaA.2012.


















