
PHOSPHORIC ACID* Glucose-l-phosphoric acid was first isolated
14
THE ISOLATION AND SYNTHESIS OF GLUCOSE-l- PHOSPHORIC ACID* BY CARL F. CORI, SIDNEY P. COLOWICK, AND GERTY T. CORI (From the Department of Pharmacology, Washington University School of Medicine, St. Louis) (Received for publication, August 19, 1937) Glucose-l-phosphoric acid was first isolated from minced and washed frog muscle which had been incubated for 3 hour in phos- phate buffer containing small amounts of adenylic acid (2). Under these conditions part of the glucose-l-phosphoric acid (l-ester) was converted to hexose-6-phosphoric acid (8ester), so that fractional crystallization of the brucine salts had to be resorted to as a means of separating the two esters. This rather tedious step in the preparation became unnecessary when it was found that the l-ester was the main reaction product in a dialyzed muscle extract incubated with glycogen, inorganic phosphate, and adenylic acid* (3). Glucose-l-phosphoric acid has also been pre- pared synthetically (1). In the present paper are recorded experi- mental data concerning the preparation, synthesis, and properties of the l-ester. EXPERIMENTAL Preparation of Muscle Extract-The hind limbs of a rabbit are perfused with Ringer’s solution until most of the blood is re- moved. The muscles are excised and chilled in an ice-cold mortar in a cold room before being passedtwice through a meat grinder; then they are mixed with an equal weight of ice-cold distilled water, triturated with a pestle, and extracted for 2 hours at 0” * A preliminary account was read before the meeting of the American Society of Biological Chemists at Memphis (1). This work was supported by a grant from a fluid research fund of Washington University School of Medicine. 1 The purpose of the dialysis is the removal of magnesium ions which accelerate the enzymic conversion of the l- to the g-ester. The dialysis also removes all but traces of organic phosphate compounds. 465 by guest on April 5, 2019 http://www.jbc.org/ Downloaded from
Transcript of PHOSPHORIC ACID* Glucose-l-phosphoric acid was first isolated

THE ISOLATION AND SYNTHESIS OF GLUCOSE-l- PHOSPHORIC ACID*
BY CARL F. CORI, SIDNEY P. COLOWICK, AND GERTY T. CORI
(From the Department of Pharmacology, Washington University School of Medicine, St. Louis)
(Received for publication, August 19, 1937)
Glucose-l-phosphoric acid was first isolated from minced and washed frog muscle which had been incubated for 3 hour in phos- phate buffer containing small amounts of adenylic acid (2). Under these conditions part of the glucose-l-phosphoric acid (l-ester) was converted to hexose-6-phosphoric acid (8ester), so that fractional crystallization of the brucine salts had to be resorted to as a means of separating the two esters. This rather tedious step in the preparation became unnecessary when it was found that the l-ester was the main reaction product in a dialyzed muscle extract incubated with glycogen, inorganic phosphate, and adenylic acid* (3). Glucose-l-phosphoric acid has also been pre- pared synthetically (1). In the present paper are recorded experi- mental data concerning the preparation, synthesis, and properties of the l-ester.
EXPERIMENTAL
Preparation of Muscle Extract-The hind limbs of a rabbit are perfused with Ringer’s solution until most of the blood is re- moved. The muscles are excised and chilled in an ice-cold mortar in a cold room before being passed twice through a meat grinder; then they are mixed with an equal weight of ice-cold distilled water, triturated with a pestle, and extracted for 2 hours at 0”
* A preliminary account was read before the meeting of the American Society of Biological Chemists at Memphis (1). This work was supported by a grant from a fluid research fund of Washington University School of Medicine.
1 The purpose of the dialysis is the removal of magnesium ions which accelerate the enzymic conversion of the l- to the g-ester. The dialysis also removes all but traces of organic phosphate compounds.
465
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

466 Glucose-l-Phosphoric Acid
under occasional stirring. After sharp centrifugation the residue is extracted a second time with the same volume of distilled water. The combined extracts are dialyzed in collodion membranes for 5 hours against running tap water of about 8” and for 12 hours against distilled water in the cold room. A globulin precipitate settles out which is removed by centrifugation or filtration.*
Small amounts of denatured protein continue to be deposited while the extract is standing in the refrigerator preserved with toluene. After an initial decrease, the phosphorylizing activity of the extract remains fairly constant, as shown in the following example in which the activity is expressed in arbitrary units: 10 days, 4.7; 14 days, 4.0; 16 days, 3.7; 19 days, 3.6; 23 days, 3.5; 28 days, 3.5.
Preparation of l-Ester from Muscle Extract-It is advisable to test the activity of the extract on a small scale before proceeding with the preparation of the l-ester. In this test 6 cc. of extract are mixed with 3 cc. of ~/3 phosphate buffer of pH 7 containing 50 mg. of glycogen and 0.2 mg. of adenylic acid. The mixture is incubated for 30 minutes at 25c3 and is deproteinized by addition of 6 cc. of 2.5 per cent HgC12 in 0.5 N HCl. After centrifugation, removal of the excess Hg as HgS4 and aeration, the solution is made faintly alkaline to phenolphthalein by addition of Ba(OH)*. This should be done without delay because of the instability of the l-ester in acid solution. The procedure previously described for the determination of hexosemonophosphate (4) is then fol- lowed. The reducing power before hydrolysis corresponds to the amount of g-ester formed (which should be less than 10 per cent of the total esterification in an extract suitable for the above prepara- tion), while the increase in reducing power (and in inorganic P) after 10 minutes hydrolysis in N HzSO4 corresponds to the amount of l-ester formed.5
* The specific conductivity of such an extract is close to 2 X 10W4, the pH is about 6 as measured with a glass electrode, and the total nitrogen content is 1.8 to 2.0 mg. per cc.
3 An increase in the length of time of incubation and in temperature favors the formation of g-ester.
4 If a considerable amount of glycogen is left, it prevents the flocculation
of HgS, a condition which can be remedied by addition of solid HgCl,. 6The amount of glycogen which disappears corresponds closely to the
sum of the l- and g-esters formed. In an extract with good activity 30 to 40
mg. of glycogen disappear in the above test in 30 minutes.
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Cori, Colowick, and Cori 467
About 0.5 gm. of the barium salt of glucose-l-phosphate can be prepared by means of 100 cc. of extract to which are added 10 cc. of M phosphate buffer of pH 7, 750 mg. of glycogen,‘j and 3 mg. of adenylic acid. After $ hour of incubation at 25” the proteins are precipitated by addition of 70 to 90 cc. of 2.5 per cent HgClz in 0.5 N HCl and the solution is filtered on a Buchner funnel. The filtrate is treated with HZS, filtered, and aerated. It is neutralized as soon as possible by rubbing successive portions with solid Ba(GH)2 in a mortar, the final adjustment to a faint pink (phenolphthalein) being made by addition of a saturated solution of Ba(CIH)2.
The solution is centrifuged. Since the barium phosphate pre- cipitate includes a considerable amount of l-ester, it is dissolved in the smallest required amount of N HCI, diluted to about 50 cc., and again treated with Ba(OH)2 until faintly alkaline to phenol- phthalein. The combined supernatant fluids are mixed with 1.5 volumes of 95 per cent alcohol and the solution is placed in the refrigerator.7 A flocculent precipitate forms which is allowed to settle so that most of the supernatant liquid can be siphoned off.
The precipitate of barium glucose-l-phosphate is centrifuged and drained; it includes small amounts of barium phosphate and carbonate, which are insoluble in water. The precipitate is ex- tracted twice with water, the insoluble residue is discarded, and the supernatant liquid is precipitated with 1.5 volumes of alcohol. This is repeated until a material almost completely soluble in water is obtained. Generally three to four solutions and pre- cipitations are required. The final precipitate is washed with a mixture consisting of 60 per cent ethanol and 40 per cent ether and with dry ether. The product is dried in vacua and ground in an agate mortar. The barium salt of glucose-l-phosphate so obtained is a white, non-hygroscopic, stable powder, easily soluble in water; it contains 3Hz0 which cannot be removed completely at 115” in vacua;* it is free of inorganic phosphate but may contain
6 Glycogen prepared by the method of Somogyi (5) was used. 7 1 volume of alcohol is sufficient if the solution is kept standing over-
night in the refrigerator, while 2 volumes are required if the solution is kept for only a short time before centrifugation.
* Drying at higher temperatures leads to decomposition, making it impossible to carry out a direct determination of the water content. One
preparation which had been kept for 4 months in a desiccator over CaCll lost 2HzO.
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

468 Glucose-l-Phosphoric Acid
small amounts of the barium salt of hexose-6-phosphate. A complete separation of the l- and 6-esters as the brucine salts is made possible by the fact that upon cooling a hot methanol solution the former ester crystallizes much more rapidly than the latter.
The l-ester, since it does not possess a free aldehyde group, does not reduce alkaline copper or iodine solutions. It is almost com- pletely (97 to 99 per cent) hydrolyzed to glucose and inorganic phosphate when heated for 10 minutes in N HzS04 in the boiling water bath.g A phosphatase, prepared from intestinal mucosa, also splits the ester to glucose and inorganic phosphate. The [oL]~ of the sugar obtained after the action of the phosphatase for 18 hours at pH 7 and 25” (which resulted in a hydrolysis of 96.5 per cent of the ester added) was +52.5’. In an experiment with acid hydrolysis the [a], of the sugar formed was +52.2”. The sugars were completely fermentable by yeast and they gave a typical glucosazone.
The following is an analysis of a preparation obtained by the action of minced frog muscle and purified as the brucine salt.
CsH1105POaBa.3Hz0 Calculated Found
C H
16.05 3.56
Pregl technique
P Ba Aldose
6.90 6.94, 6.9i Gravimetric 30.6 30.4, 30.7 Method of King (6) 40.1 39.8, 40.2 Hypoiodite titration
after acid hydroly- sis
[(Y]: = +75.5” (for anhydrous Ba salt in water; concentration 1.26%) [al: = +118” (for the free acid; concentration 1.01%)
Synthesis of 1-Ester-Phosphorylation of a- and p-2,3,4, 6-tetraacetyl glucose with POCI, in dry pyridine was first tried, but was not successful. The action of silver phosphate on a-l- bromo-tetraacetyl glucose dissolved invarious organic solvents was studied next. The expected product of the reaction, tri-(tetra- acetyl glucose-1)-phosphoric acid, was obtained in crystalline form in theoretical yield in the following way.
9 We have not been able to prepare a sample of l-ester containing 100 per cent of easily hydrolyzable P. It seems possible that the 1 to 3 per cent of difficultly hydrolyzable P is not originally present but is formed during hydrolysis.
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Cori, Colowick, and Cori
A solution of 10 gm. of dry acetobromoglucose in 30 cc. of ben- zene (dried by refluxing over sodium for 1 hour and distilled at 79.1-79.4”) is shaken with 3.5 gm. (or slightly more than the theoretical amount) of freshly prepared silver phosphatelO until the latter is finely divided. The mixture is refluxed for 1 hour with occasional shaking, with a calcium chloride tube attached to the condenser. The silver salts are centrifuged off and arewashed once with dry benzene. The combined benzene solutions, if not per- fectly clear, are centrifuged again; they are concentrated in vacua
at about 50” to a thick syrup. Upon further evacuation the syrup forms a foamy mass which crystallizes completely when left over- night in a high vacuum over CaC12. The yield is 8.9 gm. The substance is easily soluble in organic solvents (alcohol, ether, chloro- form, carbon tetrachloride) but only sparingly soluble in water.”
The preparation of glucose-l-phosphoric acid from the inter- mediate compound requires deacetylation and the splitting off of two sugar residues from the molecule. Several methods of de- acetylation (with barium hydroxide, sodium and barium methyl- ate, and ammonia in methanol) were tried, but they all caused some decomposition resulting in the formation of phosphorylized prod- ucts difficult to hydrolyze in acid and having a low specific rotation from which glucose-l-phosphoric acid could not be sep- ara,ted. The following procedure gave the desired product in pure form in a yield of 25 to 30 per cent of the theory.
To a 5 per cent solution of the intermediate substance in meth- anol is added the required amount of 5 N HCl to make a 0.2 N
solution which is kept for 16 hours at 250.12 The solution is made
lo 1.19 gm. of Na2HP0, and 4.27 gm. of AgNOa are dissolved in 100 cc. of water each and the solution of the former is poured into the latter with
vigorous stirring. The precipitate is collected on a Ruchner funnel, washed with water, absolute alcohol, and dry ether, and dried in air. All these
operations are carried out in subdued light. 11 Three different preparations showed a P content of 2.74, 2.76, 2.73,
average 2.74 per cent; calculated for tri-(tetraacetyl glucose-1)-phosphoric acid (molecular weight lo%), 2.84 per cent. [cz]: = +122’ (0.97 per cent in methanol). It is assumed that this is the OL form.
12 During that time some inorga.nic P is formed, amounting to about 10
to 15 per cent of the total organic P present, and the optical rotation of the solution shows a decrease to about 4/10 of the original value. After short periods of incubation (4 to 5 hours) the glucose-l-phosphate contains an admixture of the impurities mentioned in the preceding paragraph.
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

470 Glucose-l-Phosphoric Acid
just alkaline to phenolphthalein by addition of a saturated solu- t,ion of Ba(OH)2 and cooled for several hours in the refrigerator. The precipitate is centrifuged off, drained, and extracted with several portions of water. The insoluble residue and the methyl alcoholic mother liquor, although they include some l-ester, are discarded. The combined aqueous extracts are precipitated with 1 to 1.5 volumes of ethanol and this is repeated, as described in the preceding section, until a precipitate almost completely soluble in water is obtained. The final product, after washing with the alcohol-ether mixture and with ether, is dried in the vacuum desiccator; it contains 3Hz0 and appears to be identical with the natural product as regards chemical and physiological prop- erties.
In three different synthetic preparations of the barium salt the P content was 6.78, 6.95, 6.87, average 6.87 per cent. The glucose content (after acid hydrolysis) was 37.6,37.5,37.9, average 37.7 per cent. Calculated for P and glucose, 6.90 and 40.1 per cent, respectively. [(Y]:~ = +76”, +74”, +75”, average +75” (for the anhydrous Ba salt in water). One preparation, after removal of Ba with the equivalent amount of HvS04, gave an [(Y]: value for the free acid of +120”. The expected rotation due to the different molecular weights of the barium salt and the free acid is 75” X 395/260 = +114”.
The interaction of acetobromoglucose and silver phosphate per- mits the entrance of the phosphoric acid group only in position (1) of the glucose molecule. Walden inversion, though common in this type of reaction, does not seem to occur in the present case. The acetobromoglucose has a pyranoid structure. The synthetic product is therefore assumed to be a-glucopyranose-l-phosphoric acid.13 The natural product, which is formed by the disruption of the a-glucosidic linkages in the glycogen molecule through the entrance of the phosphoric acid group in position (l), would also
13 An attempt has been made to prepare the corresponding p compound by interaction of p-1-chloro-tetraacetyl glucose and silver phosphate. In one case in which Ba(OH)2 was used as the deacetylating agent, the barium
salt of the phosphate ester was 99 per cent hydrolyzable during 10 minutes of heating in N H&404 and had an [a], = +9.7”. However, a repetition of the experiment with the method of deacetylation described above gave a
product with an [al, of $69”.
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Cori, Colowick, and Cori 471
be expected to have an ar-pyranoid structure. Further work seems desirable in order to substantiate this structure.
Hydrolysis in Acid-The rate of hydrolysis of the natural and synthetic esters was determined in 0.25 N HCl at 37” f 0.01”. The free ester was prepared by adding the exact equivalent of HzS04 to an aqueous solution of the barium salt. After removal of BaS04 by centrifugation, the ester solution was diluted one-half with 0.5 N HCI and distributed in small individual tubes which were stoppered and almost completely submerged in the bath. The inorganic phosphate and glucose formed during hydrolysis
TABLE I
Hydrolysis of Natural and Synthetic Glucose-l-Phosphoric Acid in 0.65 N
HC1 at $7”
K was calculated by means of the formula for a unimolecular reaction.
Natural ester (0.193 mg. P per co.) Synthetic ester (0.329 mg. P per cc.)
Time
min.
60
120 180 240
300 360
P split off
per cent
16.2 29.2
40.3 51.0
59.2
K x 103
1.28 1.23
1.23 1.43
1.17
Average . . . .I 1.27 / 1.30 / 1 1.30
P split off
per cent 17.5
30.3 42.0 51.8
59.6 66.2
KX 103
1.38 1.21
1.32 1.35
1.27 1.30
GlUCOS0 split off
per cent
17.4
30.4 43.4 51.9
60.6 66.2
-i-
_-
KX 10’
1.39
1.25 1.50 1.18
1.44 1.12
were determined by the Fiske and Subbarow (7)14 and the Shaffer and Somogyi (8) methods, respectively.
As may be seen in Table I, there is no significant difference in the rate of hydrolysis of the natural and synthetic esters. The values for K remain constant (within the limits of error of the
1’The acid concentration during the development of the color in the
Fiske-Subbarow method is 0.5 N H,SOr. During the 10 or more minutes elapsing until the calorimetric readings are made, a small but measurable amount of the l-ester is being split, for which a correction should be applied if maximal accuracy is required. This correction is made on the basis of a determination of the velocity constant of the reaction at the prevailing room temperature. The values for inorganic phosphate in Table I have been so corrected.
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

472 Glucose-l-Phosphoric Acid
methods used) up to 66 per cent of hydrolysis, showing that these preparatjions do not contain appreciable quantities of other phos- phate esters. The sugar phosphates hitherto described are more resistant to acid hydrolysis than the l-cster.ls The glucose-g- phosphoric acid, in particular, with which the l-ester is most likely to be contaminated, is not measurably hydrolyzed under the con- ditions specified in Table I. Consequently, this ester, if present, will cause a falling off of the value of K as the hydrolysis of the l-ester progresses.
Robison (9) found that the rate of hydrolysis of glucose- and mannose-6-phosphoric acid did not rise with an increase in H ion concentration; almost identical values were obtained for the autol- ysis of the free esters and for their hydrolysis in 0.1 N and 1.0 N sulfuric acid. Robison explained these results by assuming that these aldose esters can exist in aqueous solution in two forms, perhaps as furanose and pyranose derivatives, and that the ad- dition of acid favored the formation of the more stable pyranoid form. As further evidence for the existence of dual forms Rob- ison cites the fact that a rise in optical rotation occurs when the esters are liberated from their salts. Glucose-l-phosphoric acid does not behave in this way; its rate of hydrolysis rises witch an increase in H ion concentration and the specific rotation of the free ester corresponds closely to that calculated from its barium salt.
Dissociation Constants-The apparent first and second dissocia- tion constants of orthophosphoric acid and of natural and syn- thetic glucose-l-phosphoric acid were determined by electro- metric titration, with a glass electrode assembly which gave readings to 0.5 millivolt. The pK’, was calculated by means of Van Slyke’s (10) formula for strong acids,
[H’l (B + fH+l) K’ = C - (B + [H+l)
the pK’2 by means of the Henderson-Hasselbalch equation, pK’, = pH - log B/(C - B).
A 0.1 M solution of the free ester was prepared by treating the
15 The only naturally occurring organic phosphate more easily hydro- lyzed in acid is phosphocreatine.
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from
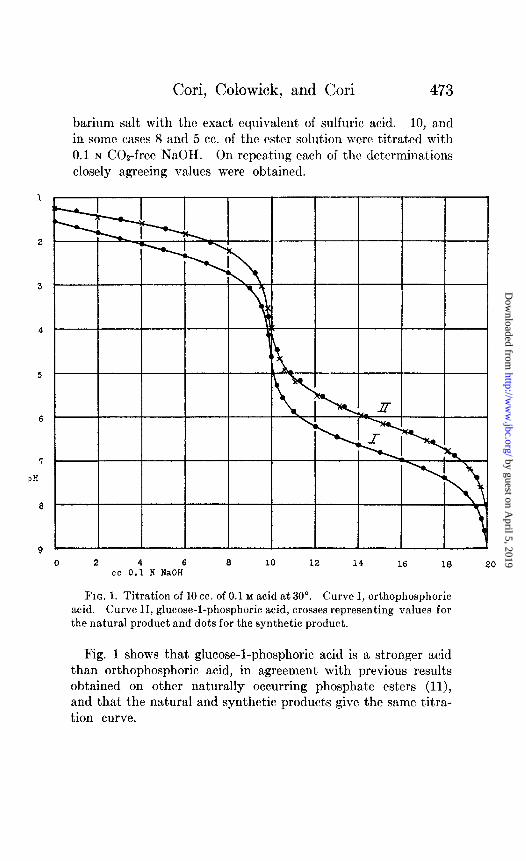
Cori, Colowick, and Cori 473
barium salt with the exact equivalent of sulfuric acid. 10, and in some cases 8 and 5 cc. of the ester solution were titrated with 0.1 N COz-free NaOH. On repeating each of tho dctcrminations closely agreeing values were obtained.
0 2 4 6 8 10 12 14 16 18 cc 0.1 N NaOH
FIG. 1. Titration of 10 cc. of 0.1 M acid at 30”. Curve I, orthophosphoric acid. Curve II, glucose-l-phosphoric acid, crosses representing values for
the natural product and dots for the synthetic product.
Fig. 1 shows that glucose-l-phosphoric acid is a stronger acid than orthophosphoric acid, in agreement with previous results obtained on other naturally occurring phosphate esters (ll), and that the natural and synthetic products give the same titra- tion curve.
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

474 Glucose-l-Phosphoric Acid
The following average values were obtained (for an initial con- centration of the acid of 0.1 M and a temperature of 30”).
pK, Orthophosphoric acid 2.03 Glucose-l-phosphoric acid (natural). 1.11
‘I “ (synthetic) 1.10 Hexose-6-phosphoric acid (11) 0.97
*KS
6.8; 6.13
6.13 6.11
Biological Behavior-It has been shown previously (3) that natural glucose-l-phosphoric acid is converted to glucose-6- phosphoric acid and then to the equilibrium or Embden ester by an enzyme system present in muscle and that magnesium ions accelerate this conversion. The kinetics of this reaction will be dealt with in a later paper; at present we merely wish to show that the biological behavior of the synthetic product is the same as that of the natural product.
In the following experiment Embden ester (which consists of an equilibrium mixture of glucose-, fructose-, and mannose-6- phosphate) was isolated as the barium salt after incubation of a dialyzed rabbit muscle extract with synthetic glucose-l-phos- phoric acid.
To 35 cc. of a dialyzed muscle extract (prepared as described earlier in this paper) was added the equivalent of 70 mg. of the an- hydrous barium salt of the l-ester, the barium having been re- moved by Na2SOd. After incubation for 3 hours at 25”, during which time almost all of the added l-ester had disappeared, the solution was deproteinized with HgC12. The barium salt was isolated and purified by repeated solutions in water and precipi- tations with alcohol. The final product, after drying in a high vacuum, weighed 41.9 mg., corresponding to 60 per cent of the l-ester added.
[cy]: = +34” (for the free acid in water, concentration 0.58 per cent). Hydrolysis in N HCI at 100” gave the following values for the velocity constant (K X lo”): 30 minutes 1.82, 60 minutes 0.63, 120 minutes 0.60, 180 minutes 0.33, 300 minutes 0.24. The final value agrees with that given by Lohmann (12). The falling off in the value of K is due to the fact that the aldose components of the Embden ester are much more difficult to hydrolyze in acid than the fructose component. A determination of the latter by
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Cori, Colowick, and Cori 475
means of the Seliwanoff reaction as modified by Roe (13) showed that 17 per cent of the total esterified hexose was fructose. This is lower than the value reported by Lohmann and the initial rate of acid hydrolysis is correspondingly lower; it seems obvious, however, that a constant composition of the isolated Embden ester is not to be expected.
The accelerating effect of magnesium ions on the conversion of synthetic l-ester to 6-ester in a dialyzed muscle extract is shown in Table II.
In a non-dialyzed or in a dialyzed and reactivated muscle extract glucose-l-phosphoric acid is fermented to lactic acid.
TABLE II
Eflect of Magnesium Ions on Conversion of I- to B-Ester
A solution of synthetic glucose-l-phosphoric acid (as the sodium salt),
containing 0.714 mg. of easily hydrolyzable P per cc., was mixed with an equal volume of a dialyzed and IO-fold diluted rabbit muscle extract.
The temperature was 23”.
No magnesium added
Period of incubation Conversion
0 0.357 0 15 0.335 6.2
30 0.318 10.9 45 0.297 16.8
0.2 mg. magnesium added per ee.
Easily hydrolya- able P Conversion
“8. per cc. per cent
0.357 0 0 313 12.3
0.246 31.1 0.139 61.2
The following experiment with synthetic l-ester may serve as an example. To 2 cc. of frog muscle extract, dialyzed for a short time, were added 1 cc. of ~/3 phosphate buffer pH 7.2, 1 mg. of adenosinetriphosphate, and 1 cc. containing 1.2 mg. (as hexose) of synthetic magnesium glucose-l-phosphate. The mixture was incu- bated for 1 hour at 29” and deproteinized by addition of HgC12. The results in mg. of lactic acid per 4 cc. of mixture were as follows: initial 0.74, after 1 hour without substrate 0.74, after 1 hour with substrate 1.56. ((1.56 - 0.74) X 100) /1.2 = 68 per cent of added glucose-l-phosphoric acid converted to lactic acid.
The good agreement in a number of chemical and biological properties justifies the conclusion that the synthetic ester is
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

476 Glucose-l-Phosphoric Acid
identical with the ester isolated from muscle extract as the first product of glycogen degradation.
SUMMARY
1. The isolation of glucose-l-phosphoric acid, produced by the action of dialyzed muscle extract on glycogen, and the synthesis of this compound by the interaction of acetobromoglucose and silver phosphate have been described. An elementary analysis of the barium salt agrees with the composition CsHII05P04- Ba. 3Hz0.
2. The ester is non-reducing. After 10 minutes in N sulfuric acid at loo”, 97 to 99 per cent of the ester is hydrolyzed to reducing sugar and inorganic phosphate, which permits its determination in the presence of other phosphate esters. The sugar formed by acid hydrolysis or by the action of an intestinal phosphatase has a specific rotation corresponding to that of glucose.
3. The natural and synthetic products agree with respect to the following values: (a) [01]i5 = +75” (for the anhydrous Ba salt in water), [cv]:~ = +120” (for the free acid); (b) the velocity con- stant for hydrolysis in 0.25 N HCl at 37” = 1.30 X 1O-3 (for both glucose and inorganic P formed) ; (c) apparent dissociation con- stants, pK: = 1.1, pKG = 6.13. A considerationof thestructural possibilities suggests that the compound is cu-glucopyranose-l- phosphoric acid.
4. The behavior of the natural and synthetic glucose-l-phos- phoric acid, when added to muscle extract, is the same in regard to conversion to hexose-6-phosphoric acid (Embden ester) which was isolated as the barium salt, in regard to the effect of mag- nesiurn ions in accelerating this conversion and in regard to fer- mentation to lactic acid.
The authors wish to thank Dr. S. A. Thayer of the Department of Biochemistry, St. Louis University, for the C and H analyses.
BIBLIOGRAPHY
1. Cori, C. F., Cori, G. T., and Colowick, S. P., Proc. A.m. Sot. Biol. Chew,., 8, xix (1937) (J. Biol. Chem., 119 (1937)).
2. Cori, C. F., and Cori, G. T., Proc. Sot. Exp. Biol. and Med., 34, 702 (1936).
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Cori, Colowick, and Cori
3. Cori, G. T., and Cori, C. F., Proc. Sot. Exp. Biol. and Med., 36, 119
(1937). 4. Cori, G. T., and Cori, C. F., J. Biol. Chem., 94,561 (193132). 5. Somogyi, M., J. Biol. Chem., 104,245 (1934).
6. King, E. J., Biochem. J., 26,586 (1932). 7. Fiske, C. H., and Subbarow, Y., J. Biol. Chem., 66,375 (1925).
8. Shaffer, P. A., and Somogyi, M., J. Biol. Chem., 100,695 (1933). 9. Robison, R., Biochem. J., 26,219l (1932).
10. Van Slyke, D. D., J. Biol. Chem., 62,525 (1922).
11. Meyerhof, O., and Lohmann, K., Biochem. Z., 166,113 (1927). 12. Lohmann, K., Biochem. Z., 194,306 (1928). 13. Roe, J. H., .I. Biol. Chem., 107,15 (1934).
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

CoriCarl F. Cori, Sidney P. Colowick and Gerty T.
GLUCOSE-1-PHOSPHORIC ACIDTHE ISOLATION AND SYNTHESIS OF
1937, 121:465-477.J. Biol. Chem.
http://www.jbc.org/content/121/2/465.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/121/2/465.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on April 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from


















