
Original Article - Diabetesdiabetes.diabetesjournals.org/content/diabetes/54/8/2294.full.pdf ·...
11
Original Article Mechanisms of Compensatory -Cell Growth in Insulin-Resistant Rats Roles of Akt Kinase Thomas L. Jetton, James Lausier, Kyla LaRock, Winifred E. Trotman, Brooke Larmie, Aida Habibovic, Mina Peshavaria, and Jack L. Leahy The physiological mechanisms underlying the compen- satory growth of -cell mass in insulin-resistant states are poorly understood. Using the insulin-resistant Zucker fatty (fa/fa) (ZF) rat and the corresponding Zucker lean control (ZLC) rat, we investigated the factors contributing to the age-/obesity-related en- hancement of -cell mass. A 3.8-fold -cell mass in- crease was observed in ZF rats as early as 5 weeks of age, an age that precedes severe insulin resistance by several weeks. Closer investigation showed that ZF rat pups were not born with heightened -cell mass but developed a modest increase over ZLC rats by 20 days that preceded weight gain or hyperinsulinemia that first developed at 24 days of age. In these ZF pups, an augmented survival potential of -cells of ZF pups was observed by enhanced activated (phospho-) Akt, phos- pho-BAD, and Bcl-2 immunoreactivity in the postwean- ing period. However, increased -cell proliferation in the ZF rats was only detected at 31 days of age, a period preceding massive -cell growth. During this phase, we also detected an increase in the numbers of small -cell clusters among ducts and acini, increased duct pancre- atic/duodenal homeobox-1 (PDX-1) immunoreactivity, and an increase in islet number in the ZF rats suggesting duct- and acini-mediated heightened -cell neogenesis. Interestingly, in young ZF rats, specific cells associated with ducts, acini, and islets exhibited an increased frequency of PDX-1 /phospho-Akt staining, indicating a potential role for Akt in -cell differentiation. Thus, several adaptive mechanisms account for the compensa- tory growth of -cells in ZF rats, a combination of enhanced survival and neogenesis with a transient rise in proliferation before 5 weeks of age, with Akt serving as a potential mediator in these processes. Diabetes 54: 2294 –2304, 2005 T he pancreatic islet -cell regulates cellular fuel metabolism and glucose homeostasis through its secretion of insulin. Numerous studies in ro- dents and humans have shown a tremendous capacity for -cell expansion in response to physiological and pathophysiological changes in tissue insulin require- ments, i.e., insulin resistance (1–3). Over the last several years, much has been learned about the molecular regula- tion of -cell development from mice with targeted genetic mutations of transcription factors and cell signaling inter- mediates. These studies have identified a key role for the insulin/IGF-1 signaling pathway in the adaptive response to insulin resistance, particularly the intermediates insulin receptor substrate-2 (IRS-2) (4 –9), Akt/protein kinase B (PKB) (10 –13), and the forkhead transcription factor Foxo1 (14) as putative regulators of the gene encoding pancreatic-duodenal homeobox-1 (PDX-1) (15). Although the details are not known about how the -cell mass is regulated in response to progressive insulin resistance, a recent study implicates PDX-1 with a key role (16). Al- though the relative contribution of hyperplasia of existing -cells versus new cell development (neogenesis) from pancreatic epithelial precursors of death from apoptosis and hypertrophy of -cells is not clear, recent evidence underscores the importance of -cell proliferation as a principal mechanism of -cell growth and mass mainte- nance (16 –18). We previously investigated -cell regeneration following a partial pancreatectomy in normally insulin-sensitive Sprague-Dawley rats (19). We observed that the growth factor–activated kinase Akt, which has been implicated in regulating duct-derived neogenesis and -cell prolifera- tion, cell size, differentiation, and survival (10–13,20–24), was activated in a subset of IRS-2 /PDX-1 epithelial cells in the common pancreatic duct epithelium (19). This suggested a possible regulatory mechanism whereby en- hanced signaling through IRS-2 and Akt may promote duct-derived -cell differentiation during islet regenera- tion. Whether these proteins play a similar role in the -cell growth response to insulin resistance is not known. The Zucker fatty (ZF) (fa/fa) rat is a model of hyperlip- idemia, insulin resistance, and obesity because of a mutated From the Division of Endocrinology, Diabetes, and Metabolism, University of Vermont, Burlington, Vermont. Address correspondence and reprint requests to T.L. Jetton, University of Vermont College of Medicine, Department of Medicine, Given C331, Burling- ton, VT 05405. E-mail: [email protected]. Received for publication 15 October 2004 and accepted in revised form 4 May 2005. IRS-2, insulin receptor substrate-2; PDX-1, pancreatic/duodenal homeobox-1; PKB, protein kinase B. © 2005 by the American Diabetes Association. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 2294 DIABETES, VOL. 54, AUGUST 2005
-
Upload
truongkhanh -
Category
Documents
-
view
218 -
download
0
Transcript of Original Article - Diabetesdiabetes.diabetesjournals.org/content/diabetes/54/8/2294.full.pdf ·...

Original Article
Mechanisms of Compensatory �-Cell Growth inInsulin-Resistant RatsRoles of Akt KinaseThomas L. Jetton, James Lausier, Kyla LaRock, Winifred E. Trotman, Brooke Larmie, Aida Habibovic,
Mina Peshavaria, and Jack L. Leahy
The physiological mechanisms underlying the compen-satory growth of �-cell mass in insulin-resistant statesare poorly understood. Using the insulin-resistantZucker fatty (fa/fa) (ZF) rat and the correspondingZucker lean control (ZLC) rat, we investigated thefactors contributing to the age-/obesity-related en-hancement of �-cell mass. A 3.8-fold �-cell mass in-crease was observed in ZF rats as early as 5 weeks ofage, an age that precedes severe insulin resistance byseveral weeks. Closer investigation showed that ZF ratpups were not born with heightened �-cell mass butdeveloped a modest increase over ZLC rats by 20 daysthat preceded weight gain or hyperinsulinemia that firstdeveloped at 24 days of age. In these ZF pups, anaugmented survival potential of �-cells of ZF pups wasobserved by enhanced activated (phospho-) Akt, phos-pho-BAD, and Bcl-2 immunoreactivity in the postwean-ing period. However, increased �-cell proliferation inthe ZF rats was only detected at 31 days of age, a periodpreceding massive �-cell growth. During this phase, wealso detected an increase in the numbers of small �-cellclusters among ducts and acini, increased duct pancre-atic/duodenal homeobox-1 (PDX-1) immunoreactivity,and an increase in islet number in the ZF rats suggestingduct- and acini-mediated heightened �-cell neogenesis.Interestingly, in young ZF rats, specific cells associatedwith ducts, acini, and islets exhibited an increasedfrequency of PDX-1�/phospho-Akt� staining, indicatinga potential role for Akt in �-cell differentiation. Thus,several adaptive mechanisms account for the compensa-tory growth of �-cells in ZF rats, a combination ofenhanced survival and neogenesis with a transient risein proliferation before 5 weeks of age, with Akt servingas a potential mediator in these processes. Diabetes 54:2294–2304, 2005
The pancreatic islet �-cell regulates cellular fuelmetabolism and glucose homeostasis through itssecretion of insulin. Numerous studies in ro-dents and humans have shown a tremendous
capacity for �-cell expansion in response to physiologicaland pathophysiological changes in tissue insulin require-ments, i.e., insulin resistance (1–3). Over the last severalyears, much has been learned about the molecular regula-tion of �-cell development from mice with targeted geneticmutations of transcription factors and cell signaling inter-mediates. These studies have identified a key role for theinsulin/IGF-1 signaling pathway in the adaptive responseto insulin resistance, particularly the intermediates insulinreceptor substrate-2 (IRS-2) (4–9), Akt/protein kinase B(PKB) (10–13), and the forkhead transcription factorFoxo1 (14) as putative regulators of the gene encodingpancreatic-duodenal homeobox-1 (PDX-1) (15). Althoughthe details are not known about how the �-cell mass isregulated in response to progressive insulin resistance, arecent study implicates PDX-1 with a key role (16). Al-though the relative contribution of hyperplasia of existing�-cells versus new cell development (neogenesis) frompancreatic epithelial precursors of death from apoptosisand hypertrophy of �-cells is not clear, recent evidenceunderscores the importance of �-cell proliferation as aprincipal mechanism of �-cell growth and mass mainte-nance (16–18).
We previously investigated �-cell regeneration followinga partial pancreatectomy in normally insulin-sensitiveSprague-Dawley rats (19). We observed that the growthfactor–activated kinase Akt, which has been implicated inregulating duct-derived neogenesis and �-cell prolifera-tion, cell size, differentiation, and survival (10–13,20–24),was activated in a subset of IRS-2�/PDX-1� epithelial cellsin the common pancreatic duct epithelium (19). Thissuggested a possible regulatory mechanism whereby en-hanced signaling through IRS-2 and Akt may promoteduct-derived �-cell differentiation during islet regenera-tion. Whether these proteins play a similar role in the�-cell growth response to insulin resistance is not known.
The Zucker fatty (ZF) (fa/fa) rat is a model of hyperlip-idemia, insulin resistance, and obesity because of a mutated
From the Division of Endocrinology, Diabetes, and Metabolism, University ofVermont, Burlington, Vermont.
Address correspondence and reprint requests to T.L. Jetton, University ofVermont College of Medicine, Department of Medicine, Given C331, Burling-ton, VT 05405. E-mail: [email protected].
Received for publication 15 October 2004 and accepted in revised form 4May 2005.
IRS-2, insulin receptor substrate-2; PDX-1, pancreatic/duodenal homeobox-1;PKB, protein kinase B.
© 2005 by the American Diabetes Association.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked “advertisement” in accordance
with 18 U.S.C. Section 1734 solely to indicate this fact.
2294 DIABETES, VOL. 54, AUGUST 2005

leptin receptor gene (25–30). These rats physiologicallyadapt to the insulin resistance through increased �-cellmass and enhanced �-cell function such that hyperglyce-mia is largely prevented. As a result, this model has beenused extensively to investigate the biochemical and mo-lecular bases for �-cell compensation. A study of adult ZFrats has reported a striking increase of �-cell mass inresponse to their insulin resistance attributable to en-hanced �-cell hypertrophy and neogenesis (1). This studydid not address the mechanisms that initiate or mediatethese �-cell mass–augmenting processes. Furthermore,the temporal relationship between the growth of �-cellsand the onset of fully established insulin resistance inthese rats has never been determined.
The present study has investigated the time of onset andorigin of the cells that mediate �-cell expansion in ZF rats.We have also screened for potential signaling intermedi-ates. Importantly, we observed that �-cell mass expansionpreceded the onset of obesity and marked insulin resis-tance. Our results have uncovered potential regulatorymechanisms whereby Akt/PKB signaling promotes en-hanced �-cell mass in an animal that develops severeinsulin resistance in adulthood.
RESEARCH DESIGN AND METHODS
Tissue processing. Adult male ZF and ZLC (�/fa or �/�) rats at 5, 10, and13 weeks of age (n � 5 for each group) were obtained from Harlan and housedin the UVM Animal Facility at least 5 days before use. Young male and female10-, 20-, 28-, and 31-day fa/fa and �/� rats were obtained from litters derivedfrom pregnant �/fa mothers (also acquired from Harlan) that had been matedto �/fa studs (see genotyping details below). The guidelines set forth by theUVM IACUC Committee were strictly adhered to for these studies. Six hoursbefore they were killed, adult rats were given 100 mg/kg BrdU by intraperi-toneal injection. Body weight and nonfasting blood glucose concentrationwere measured in nonanesthetized rats before euthanasia by exsanguinationfollowing intraperitoneal sodium pentobarbital injection. Pancreata wererapidly excised, cleared of fat and lymph nodes, blotted, and weighed beforeimmersion fixing overnight in 4.0% paraformaldehyde in 0.1 mol/l PBS at 4°C.After washing in several changes of PBS, tissues were dehydrated in ascend-ing ethanols and embedded in paraffin.
For the detection of labile signaling intermediates (phospho-Akt andphospho-BAD) in relation to the transcription factor PDX-1, pancreata from25-day-old rats (in the postweaning period) (n � 4 each group) were rapidlyexcised, cut into smaller pieces, and paraformaldehyde fixed for 1 h. Theywere immediately washed, then equilibrated in 30% sucrose/PBS, embedded inOCT medium (Miles Scientific), and sectioned at 5 �m in a cryostat.Zucker pup genotyping. Tail genomic DNA was isolated from Zucker pupsusing Qiagen’s DNeasy Tissue Kit and following manufacturer’s instructions(Qiagen). The introduction of an MspI site by the fa mutation was detected byPCR using a previously described protocol (29). Briefly, PCR primers spanningthe fa mutation were designed to amplify a 1.8-kb fragment. PCR wasperformed in 20 �l containing 100 ng of genomic DNA mixed with 1X Taq
buffer (HotStarTaq DNA polymerase buffer; Qiagen), 500 nmol/l of primers(forward: 5�-GTTTGCGTATGGAAGTCACAG-3�, reverse: 5�-ACCAGCAGAGATGTATCCGAG-3�), 250 �mol/l dNTPs, and 1 unit of HotStarTaq DNA Poly-merase (Qiagen). PCR amplication protocol began with one cycle of 92°C for15 min, followed by 35 cycles at 92°C for 30 s, 54°C for 1 min, 68°C for 5 min,and finished with an extension step at 72°C for 10 min. A total of 20 �l of PCRproduct was digested with MspI for 1 h at 37°C and analyzed on a 1% agarosegel. MspI restriction enzyme digestion of the 1.8-kb product from wild-typeZucker pups produced fragments of 1,100 and 700 bp, whereas the fa mutationproduced fragments of 950, 700, and 130 bp.Plasma glucose and insulin measurements. Fed glucose levels weredetermined in whole blood using a glucose meter (Freestyle; TheraSense) at9:00 A.M., and the plasma was subsequently analyzed for insulin employing arat insulin standard-range enzyme immunoassay kit (Alpco Diagnostics).�-Cell mass measurements. Five-micrometer paraffin sections throughoutthe entire pancreas were mounted as ribbons on microscope slides tofacilitate section counting. At 200-�m intervals in the young rats or 400-�mintervals in the adult rats, sections were rehydrated, blocked, and immuno-stained with guinea pig anti-insulin IgG (Linco Research) followed by donkey
anti–guinea pig IgG-alkaline phosphatase (Jackson Immunoresearch). Follow-ing development with Vector Red substrate (Vector Laboratories), sectionswere counterstained with hematoxylin, cleared, and mounted in Permount(Fisher Scientific).
The proportion of islet �-cell surface area versus surface area of the wholepancreas was determined planimetrically by digitally imaging 14–20 (youngrats) or 50–60 (adult rats) nonoverlapping �3.63-mm2 fields per pancreas ona Zeiss Universal microscope coupled to a Spot RT color charge-coupleddevice camera (Diagnostics Instruments). Single and small clusters of �-cellswere presumed to be neogenic and were below the threshold for accuratesurface area measurements; hence, we measured these cells in differentassays (see below). Therefore, only islets �5 cell diameters were included forislet �-cell surface area measurements. Image files were analyzed using NIHImage version 1.62 with the “Area-Measure” tool, then tabulated (in pixelvalues) and entered into Microsoft Excel for statistical analyses (significancedetermined by Student’s t test where P � 0.05 was considered significant).This planimetric method also allowed us to estimate relative islet numbersand compare islet sizes. The possibility of measuring large islets twice wasminimized due to the wide sampling interval between sections. �-Cell masswas estimated for each animal by determining the average �-cell surface areaper animal multiplied by their pancreatic weight.�-Cell and duct proliferation. At least two slides per adult pancreas wereimmunohistochemically stained for BrdU and insulin. Hydrated tissue sec-tions were subjected to pretreatment with 0.125% trypsin solution (Zymed) for10 min at 37°C or a 10-min boil in 10 mmol/l citrate buffer, pH 6.0, followed bydenaturation in 2N HCl for 20 min. After blocking, sections were incubatedovernight in mouse anti–BrdU-biotin (1:20; Zymed), washed, and then incu-bated in Z-avidin–horseradish peroxidase (1:300; Zymed Laboratories) for 1 h.In other experiments examining young ZF �-cell and duct proliferation, slideswere immunostained for the proliferation marker Ki-67 and insulin. Thisalternative strategy was used due to inconsistencies in BrdU labeling in theseyoung rats that are known to occur in certain situations (31), includingparaformaldehyde fixation (32). Since BrdU labeling marks the “S” phase ofthe cell cycle and Ki-67 marks a cycling cell, these two markers are notinterchangeable but are related. Therefore, in young Zucker pups, the valuesfor proliferation were expressed as Ki-67� �-cells (%) and were much higherthan the values for BrdU incorporation. Hydrated tissue sections weresubjected to pretreatment as above and then were incubated overnight inmouse anti–Ki-67 (1:500; Transduction Laboratories), washed, and then incu-bated in donkey anti–mouse-IgG-biotin (Jackson Immunoresearch) followedby streptavidin–horseradish peroxidase (1:300; Zymed). Following develop-ment in DAB/H2O2, sections were then stained for insulin as described aboveand finally counterstained with hematoxylin. The number of Ki-67� nuclei per1,000–1,500 islet �-cells or per 300 common duct cells was counted for eachanimal.�-Cell size measurements. Islet �-cell size was determined by doubleimmunofluorescence labeling and confocal imaging of insulin and eitherGLUT2 (Alpha Diagnostics; young rats and 5-week groups) or pan-cadherin(Zymed) (10-week group) to mark �-cell surface boundaries. A nuclearcounterstain was also used in an effort to maximize the number of cellsoptically sectioned through the middle of the cell. Only those �-cells with aclearly defined nucleus were measured. Pseudocolored images were mergedusing Adobe Photoshop, and �-cell boundaries were outlined and the surfacearea measured (�m2) using NIH Image.Islet number and size measurements. Using the primary data obtained forthe �-cell mass measurements, the relative islet number for each animal(calculated as average number per microscopic field in 400-�m intervals) wasdetermined. With this same primary data set, we also determined the relativesizes, or cross-sectional surface area of individual islets, by converting pixelvalues to micrometers squared and tabulating for each animal.Analysis of neogenesis. Single and small clusters of cells too small to beconsidered definitive islets and thus not accounted for in the islet massmeasurements for the 10-, 20-, 28-, and 31-day and 10-week-old rats weretallied in each group (n � 4–5 each group). Using the same insulin-stainedsections that were used for islet �-cell mass measurements, the total numberof �-cells as singles and clusters as doublets, triplets, quadruplets, and up tofive cell diameters (very small islets) were counted for each animal. Single andclustered �-cells, regardless of their location, show no signs of replication andtherefore were considered neogenic and may aggregate to form new islets.Multiple-labeling immunofluorescence. Hydrated sections of pancreaswere blocked in 5% normal donkey serum � 1% BSA and then incubatedovernight at 4°C in an antibody mixtures of sheep anti-amylase (Biogenesis),guinea pig anti-insulin or C-peptide (Linco), rabbit or guinea pig anti-glucagon(Linco), sheep anti-somatostatin (Cortex), and rabbit or guinea pig anti–pancreatic polypeptide (Linco). For staining of PDX-1, a rabbit antiserum wasused at 1:1,000 (gift from Dr. Chris Wright, Vanderbilt University). Staining for
T.L. JETTON AND ASSOCIATES
DIABETES, VOL. 54, AUGUST 2005 2295

PDX-1 required antigen retrieval as detailed previously. Labeling of phospho-Akt and phospho-BAD required rapidly fixed and processed frozen sectionsthat were stained with antiserum specific for activated Akt using a mousemonoclonal anti–phospho-S473Akt (Cell Signaling Technology) or a sheepanti–phospho-S473Akt (Upstate), and mouse anti–phospho-BAD (Cell Signal-ing Technology). Polyclonal antibody to Bcl-2 was obtained from Santa CruzBiotechnology.
Following washing, sections were incubated in a secondary antibodymixture that contained “ML-” grade donkey anti–species-specific IgG conju-gated to CY2 (1:300), CY3 (1:2000), or CY5 (1:500; all from Jackson Immu-noResearch), counterstained in 0.5 �g/ml Hoecsht 33258, and mounted inAqua Polymount (Polysciences). Alternate sections were stained with nonim-mune serum from the appropriate species. Controls for secondary antibodyspecificity were performed as previously detailed (19). Images were acquiredusing the aforementioned color charge-coupled device camera for conven-tional epifluorescence or by confocal microscopy.Immunoblot analyses. Purified islets were lysed in ice-cold buffer consistingof 50 mmol/l HEPES (pH 7.5), 1% (vol/vol) Nonidet P-40, 2 mmol/l sodiumorthovanadate, 100 mmol/l sodium fluoride, 10 mmol/l sodium pyrophosphate,4 mmol/l EDTA, 1 mmol/l PMSF, and protease inhibitors. The lysate wassonicated, spun, and the protein concentration determined. Protein wasseparated by SDS-PAGE, transferred to polyvinylidine fluoride membranes,and probed with the relevant antibodies followed by enhanced chemilumines-cence detection. Antibodies used to probe for total protein, and activated(phosphorylated) forms of Akt and BAD were obtained from Cell Signaling.Bcl-2 antibody was purchased from Santa Cruz Biotechnology. These were thesame antibodies used for the immunostaining experiments.Analysis of apoptosis for �-cells in situ and in vitro. For determination ofthe apoptotic �-cells in young rats (10–35 days), a modified TUNEL stainingprotocol was used to fluorescently mark cells with fragmented DNA strands(33). A Dead-End TUNEL Kit (Promega) was used with the initial steps of theprotocol followed by the manufacturer’s suggestions. Modification of the kit(TdT-based end labeling with biotin-UTP as the labeled nucleotide) includedincubation with streptavidin-CY3 (1:2,000, Jackson ImmunoResearch) in thefinal labeling step. Pancreas or islet sections were immunofluorescentlystained for insulin (CY2) and finally counterstained with 0.5 �g/ml Hoechst.Verification of TUNEL labeling included staining adjacent slides with rabbitanti-cleaved caspase 3 (R&D Systems) or propidium iodide to label apoptoticnuclei. TUNEL labeling of short-term cultured islets was accomplished byisolating islets from 4.5-week-old rats (ZLC, n � 3; ZF, n � 6; mean islets perrat, 15) using standard methods and incubating in RPMI-1640 � 10% fetalbovine serum at 37°C for 1 h before fixing and processing for frozen sections.Confocal imaging. Sections were imaged using a Bio-Rad MRC 1024 scan-ning confocal head coupled to an upright Olympus BX50 microscope with anAr-Kr laser utilizing the 488-, 568-, and 647-nm excitation lines (UVM Micros-copy Imaging Facility). Images were acquired with the Bio-Rad LaserSharpsoftware merged and formatted on a Power Macintosh running AdobePhotoshop.Quantitative imaging for Bcl-2 and PDX-1. Semiquantitative comparisonsof Bcl-2 immunofluorescence intensity for islet �-cells from young pups wereaccomplished by batch staining and sampling by confocal microscopy forBcl-2 (CY3), insulin (CY5), and a nuclear counterstain (YO-PRO1; MolecularProbes). The approach allowed us to measure the relative levels of immuno-reactive Bcl-2 within individual �-cells without potential signal from thenon–�-cells, an inherent pitfall in whole-islet analyses. Samples were imagedwith the confocal microscope using a 60� PlanApo objective lens (N.A. � 1.4)and the 568-, 488-, and 647-nm excitation lines, respectively, of an ArKr laser.For each field, the microscope was focused to maximize the number of cellsoptically sectioned through the middle of the nucleus. All confocal imagingparameters were identical for each imaged field with four nonoverlappingfields of �170 � 170 �m captured for each animal analyed. Grayscale images(512 � 512 pixels) were transferred to a Power Macintosh running NIH Imagefor analyses. For each field studied, the Area-Measure tool was used todetermine the mean pixel intensity (range 0–255 grayscale levels) of Bcl-2immunofluorescence in randomly picked 20-pixel areas of the perinuclearregion of �-cells. The final corrected intensity values were calculated for eachcell after subtraction of the field background fluorescence values.
Using an imaging strategy similar to method above, semiquantitativecomparisons of duct nuclear PDX-1 immunofluorescence intensity wereobtained from sections of the common pancreatic duct epithelium from10-week-old ZF and ZLC rats (n � 5 per group).
RESULTS
�-Cell mass of adult ZF rats. These studies were initi-ated to pinpoint both the age at which the increased �-cell
mass occurs in ZF rats and to study the underlyingmechanisms. We first quantified �-cell mass of ZF and ZLCrats at several time points from 5 to 13 weeks of age (Fig.1A). Unexpectedly, an �3.5-fold increase was observed inthe ZF rats at 5 weeks of age (P � 0.008), which is wellbefore marked insulin resistance that occurs at �7–10weeks (25,34,35). Very similar and proportional �-cellmass differences between the ZF and ZLC rats were seenat 10 and 13 weeks of age, which suggested that thecompensatory �-cell expansion was completed before 5weeks of age and maintained into adulthood. Examiningthe morphological bases for the increased �-cell mass in 5-and 10-week-old ZF rats revealed enhancement in the sizeof islets (Fig. 1B and C, respectively). These data initiallysuggested that the mass augmentation effect was princi-pally due to �-cell hyperplasia. Surprisingly, however, the�-cell proliferation rate of ZF rats was not increased at 5,
FIG. 1. �-Cell mass enhancement in adult ZF rats is due to increasedislet number and size. A: �-Cell mass was increased significantly in theZF rat (■ ). *P < 0.01 vs. ZLC (�) at all time points examined (n � 5 foreach group). Five-week-old (B) and 10-week-old (C) rats; the ZF groupexhibited more islets of a larger size than those of ZLC rats. 1K � 1,000pixels surface area or �8,500 �m2.
�-CELL MASS COMPENSATION IN THE ZUCKER RAT
2296 DIABETES, VOL. 54, AUGUST 2005

10, or 13 weeks (Fig. 2A). We also quantified the relativenumber of islets by assessing their prevalence in sectionsat 400-�m intervals: 5-week-old ZF rats had an 18% in-crease, and 10-week-old ZF rats exhibited a �40% increase(P 0.01) in the islet number over age-matched ZLC rats(Fig. 1B and C). In addition, no detectable differences in�-cell apoptosis as judged by TUNEL, Hoechst nuclearstaining, and cleaved caspase-3 immunohistochemistrywere found between the ZF and ZLC rats (data not shown).There was a 33–38% increase in the size (by surface area)of individual �-cells of 5- and 10-week-old ZF rats versusthe lean controls, respectively (Fig. 2B; P 0.001 for bothtime points), but this degree of hypertrophy alone couldaccount for only a portion of the increased �-cell mass.Since there was also an increase in the number of islets inZF rats, we reasoned that another source for the �-cellmass increase might be accelerated islet neogenesis. Ac-cordingly, we quantified single �-cells and small �-cellclusters and observed significant increases in 10-week-oldZF rats (Fig. 2C), thus suggesting the possibility of in-creased �-cell neogenesis. Importantly, these cells nevershowed signs of proliferation (data not shown). Thus,these scattered �-cells probably differentiate directly orindirectly (postmitotically) from a non–� precursor celltype. Increased ductal proliferation and PDX-1 expressionin the adult has been correlated with enhanced neogenicpotential (rev. in 2). Although we did not detect significantdifferences in proliferation among the epithelium of thecommon or main ducts in 10-week-old ZF rats (data notshown), we found an �50% increase in nuclear PDX-1immunofluorescence intensity in the common duct of ZFrats (Fig. 2D).
Collectively, these results suggested that the increased�-cell mass in adult ZF rats occurs from both �-cellhypertrophy and enhanced neogenesis but not increasedproliferation. However, the most dramatic observationwas that the mass augmentation was essentially com-pleted by 5 weeks of age. As such, resolving the underlyingmechanisms for this differential growth response requiredus to study younger rats.�-Cell mass studies of ZF pups. Before this study, �-cellmass measurements and growth parameters of ZF rats 5weeks of age had not been reported. We investigated 10-,20-, 28-, and 31-day-old ZLC and ZF pups and observed asubtle increase in �-cell mass of the ZF pups at 20 days ofage (1.4-fold over lean littermate controls; P � 0.029) (Fig.3A). This early �-cell growth preceded any differences inbody weight or insulinemia compared with the ZLC pups(Fig. 3B and C and Table 1). Also, blood glucose levelswere not different between the ZF and ZLC pups at anytime point (Fig. 3D).
The �-cell mass increase in the ZF pups at 20 and 28days old did not appear to be the result of enhanced �-cellproliferation (Fig. 3E). Interestingly, however, at 31 daysof age, ZF pups exhibited a twofold enhancement (P �0.004) in �-cell proliferation over the ZLC pups (Fig. 3E).There were no differences among the two groups in thesize of �-cells, nor were there detectable differences in�-cell apoptosis (as determined by a sensitive TUNELfluorescence assay, activated [cleaved] caspase-3 immuno-staining, and propidium iodide/Hoechst staining), in 10-,20-, 28-, or 31-day-old pups (data not shown). In fact,
FIG. 2. �-Cell hypertrophy and enhanced �-cell recruitment throughneogenesis, but not hyperplasia, contribute to �-cell mass maintenancein adult ZF rats. A: No differences in �-cell proliferation were observedbetween the adult animal groups. �, ZLC rats; f, ZF rats. B: Individual�-cell size had increased �30–40% (*P < 0.0001) in the 5- and10-week-old ZF rats. C: A tally of presumed nascent �-cells in 10-week-old rats, single insulin� cells, doublets, etc., up to five cell diametersper pancreas (section intervals every 200 �m), were grouped andcharted. Significant increases were observed in all �-cell cluster sizegroups from ZF rat pancreata (*P < 0.05; **P < 0.01). D: Semiquanti-tative analysis of nuclear PDX-1 staining in the common pancreaticduct epithelium revealed an �50% increase (*P � 0.014) in 10-week-old ZF rats (n � 5 per group).
T.L. JETTON AND ASSOCIATES
DIABETES, VOL. 54, AUGUST 2005 2297

apoptotic cells were quite rare in all animals examined(e.g., 1 in 1,000 �-cells, or 0.1%). Since a wave ofapoptosis in the pancreas has been reported to occur inSprague-Dawley rat pups during the 3rd week of life,presumably to serve a remodeling function (36), we alsostudied apoptosis in 15-day-old rats. Again, we found toolow frequencies of apoptosis in these rats to be useful forcomparison.Survival potential is enhanced in islets from young
ZF rats. The rarity of TUNEL� �-cells in tissue sectionsmade cell survival potential determinations between thegroups untenable, possibly due to efficient apoptotic clear-ance mechanisms in Zucker rat pancreata. To circumventthis problem, we performed TUNEL staining on frozensections of isolated islets from 4.5-week-old rats (ZLC, n �3; ZF, n � 6) that were then cultured for 1 h beforefixation. Accordingly, we found that islets from ZF ratsexhibited a significantly reduced fraction (P � 0.009) ofTUNEL� �-cells compared with the ZLC islets (Fig. 3F).
In the absence of finding any detectable differences inislet �-cell death in situ, we predicted that the early �-cellmass expansion in young ZF rats was initially because of
enhanced neogenesis. This was supported by our findingof significant increases in the number of small �-cellclusters at 20–28 days of age that, unexpectedly, tran-siently wane at 31 days (Fig. 4A). Since no differenceswere seen in the numbers of clusters between the groupsat 10 days of age before the �-cell mass increase (Fig. 4A),there appears to be a temporal correlation with theincreased �-cell mass and increased prevalence of �-cellclusters from 20 days of age onward. These �-cells consis-tently lacked any indications of proliferation (not shown).Upon closer analysis, these clusters were composed ofinsulin�/PDX-1� cells that were associated with bothducts and acini (Figs. 4B and C), and may serve as thesource of new islets. This was followed by a transient risein proliferation at 31 days that preceded a substantial�-cell mass increase at 35 days of age.Enhanced Akt/PKB signaling in ducts of ZF pups. Ourprior study of �-cell regeneration following a 60% pancre-atectomy in Sprague-Dawley rats implicated Akt/PKB sig-naling in duct-related �-cell neogenesis (19). We examinedthis possibility in 25-day-old ZF and ZLC pups, an age thatimmediately follows the normal weaning period. We de-
FIG. 3. �-Cell growth characteristics inyoung Zucker rats. A: �-Cell mass in-creased significantly by 20 days of age inthe ZF rat (*P < 0.03; **P < 1 � 10�5;***P < 0.04). �, ZLC rats; ■ , ZF rats. B:Body mass increased significantly (*P <0.001) in the ZF rats by 28 days, as didhyperinsulinemia (P < 0.01) (C). D: Nodifferences in blood glucose levels wereobserved between ZF and ZLC rats. E:No differences in �-cell proliferationwere observed between the groups inyoung Zucker rats except at 31 days,when there was a twofold increase(*P � 0.004). F: TUNEL analysis of�-cells from short-term–cultured isletsfrom 31-day-old rats revealed a signifi-cant reduction (*P � 0.009) in labelingin the ZF group (ZLC, n � 3; ZF, n � 6).
�-CELL MASS COMPENSATION IN THE ZUCKER RAT
2298 DIABETES, VOL. 54, AUGUST 2005

tected activated (phospho-) Akt/PKB staining in noncon-tiguous cells of the common duct epithelium in both thesurface lining and in the evaginations (Fig. 5A and B).While rare in the ZLC pups, these phospho-Akt� cells wereconsiderably more frequent in the ducts of ZF pups.Further characterization of these cells revealed that theywere Ki-67 negative and did not express any of theprincipal islet hormones. In contrast, they routinely exhib-ited very high levels of nuclear PDX-1 (Fig. 5A and B). Wealso observed insulin�/PDX-1� cells in small ducts pre-sumed to be nascent �-cells that displayed little or nodetectable phospho-Akt staining (Fig. 5C). Cells with thesame staining profiles were also observed in the centroaci-nar region (Fig. 5D) of both groups of 25-day-old rats. Thislatter observation supports the current hypotheses ofacinar cell transdifferentiation (37–39) and centroacinarcells (40) as a potential source of �-cells. Collectively,these findings suggested a developmental sequencewhereby activation of Akt in exocrine precursors initiatesor mediates early events in �-cell differentiation, but itsactivity is then rapidly suppressed during the progressionto fully mature �-cells.Enhanced Akt/PKB signaling and survival in islet
�-cells of ZF pups. In the 25-day-old ZF pups, wedetected moderate phospho-Akt immunoreactivity in islet�-cells, whereas in the ZLC rat �-cells, it was generallyquite low (compare Fig. 6A with C). In contrast, �-cell totalAkt immunostaining (not shown) and immunoblot analysis(Fig. 6I) revealed no differences among islets of ZF andZLC pups. Immunoblot analyses of phospho-Akt in isletsfrom young ZLC and ZF rats failed to show consistentdifferences between the groups, possibly due to non–�-cellAkt activation in islets or inadequate stabilization of isletphosphatases during islet isolation. On the other hand,using a rapid fixation and processing method for pancreascryosections (19), we can reproducibly detect phospho-Akt and other labile phospho-intermediates in �-cell insitu. Since Akt is an established survival factor for manycells types including �-cells (12,21,23,24), we next exam-ined the expression patterns of the genes encoding tworelated �-cell proteins that function in the mitochondrialregulation of apoptosis, BAD, and Bcl-2, a proapoptoticfactor and a survival factor, respectively. BAD activity isinhibited upon phosphorylation and is an establishedtarget of Akt/PKB (41). In 4-week-old rats, we found nodifferences in BAD or Bcl-2 mRNA expression between ZFand ZLC rats by RT-PCR (data not shown), nor did we findconsistent differences in their respective protein levels byimmunoblot (Fig. 6I). In contrast, immunostaining forphospho-BAD showed that it was markedly elevated inislets of 25-day-old ZF pups compared with the ZLCs (Fig.6B and D), suggesting functional inhibition of this protein.Although we found no significant differences in Bcl-2immunoreactivity in �-cells of ZF and ZLC pups at 20 daysof age (not shown), at 28 days of age, Bcl-2 immunofluo-rescence intensity was clearly increased (Fig. 6E–H).Using semiquantitative confocal analysis to measure cyto-plasmic Bcl-2 immunofluorescence at the level of individ-ual �-cells, the signal was 2.2-fold enhanced in ZF rats(Fig. 6J) (P � 0.046). This disparity between the whole-islet immunoblot analysis and the �-cell immunofluores-cence measurements of Bcl-2 may be due to non–�-cell
TA
BLE
1T
rendsin
increasedbody
weight
gainand
progressivehyperinsulinem
iain
ZFrat
pups
20day
22day
24day
26day
28day
Body
Weight(g)
Blood
Insulin(ng/m
l)
Body
Weight(g)
Blood
Insulin(ng/m
l)
Body
Weight(g)
Blood
Insulin(ng/m
l)
Body
Weight(g)
Blood
Insulin(ng/m
l)
Body
Weight(g)
Blood
Insulin(ng/m
l)
ZLC34.4
1.1
0.25
0.0540.8
1.06
0.34
0.0449.0
1.4
0.42
0.0557.9
1.6
0.56
0.2964.6
1.1
0.38
0.03ZF
37.4
1.70.26
0.05
44.3
2.10.32
0.03
54.8
2.7*0.75
0.11†
67.4
3.2†0.62
0.18
75.9
2.4‡3.24
1.53*
Values
representm
easurements
fromthe
28-daygroup
comprised
of6
ZFand
11ZLC
rats*.Body
weight
andblood
insulinw
erem
easuredfrom
fedrats
atthe
same
time
eachm
orning.*P
0.05;
†P
0.01;
‡P
0.001.
T.L. JETTON AND ASSOCIATES
DIABETES, VOL. 54, AUGUST 2005 2299

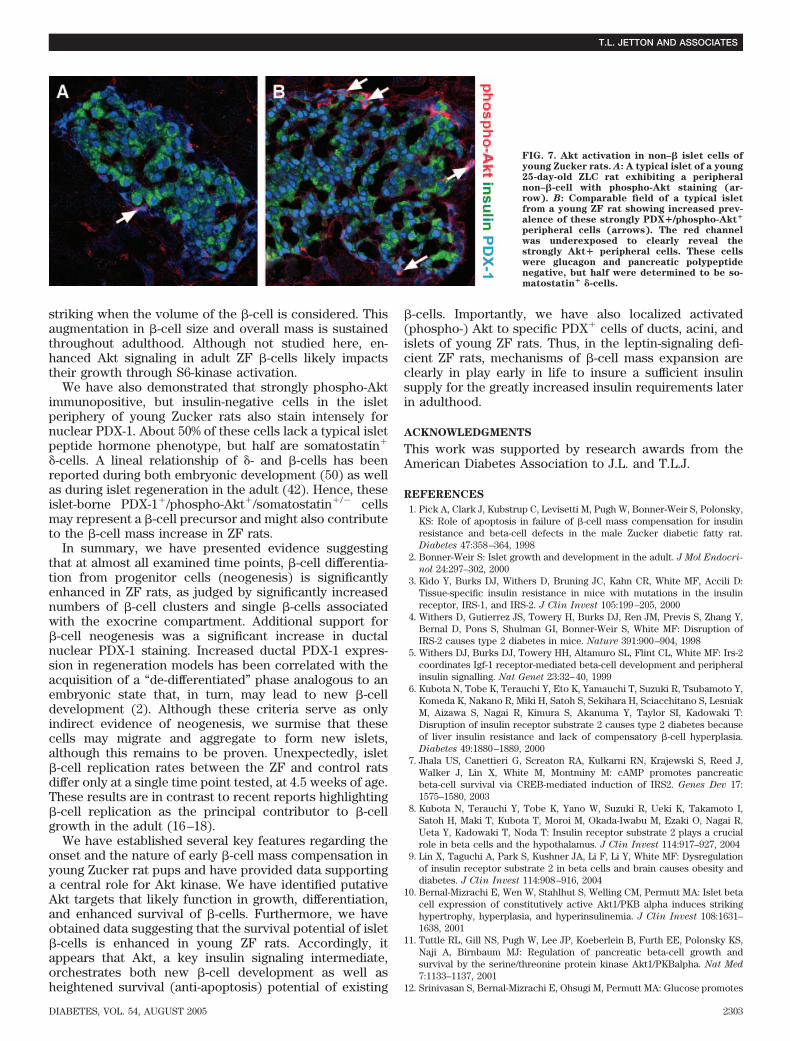
Bcl-2 protein in the whole-islet extracts. Based on theseobservations of detecting increased phospho-Akt, Bcl-2,and phospho-BAD immunoreactivity in islet �-cells ofyoung ZF rats, we propose that activated Akt in �-cellsmay be targeted to mitochondrial BAD, thus inhibiting itsfunction and thereby promoting �-cell survival.Enhanced Akt/PKB signaling in non–�-cells of ZF
islets. A closer examination of islets revealed an addi-tional cell type–specific staining pattern of phospho-Akt inthe ZF pups at the postweaning age. In addition to the�-cells, we noted an increased prevalence of intenselystained, peripheral cells in the ZF rats compared with theZLCs (Figs. 6C and 7) that were glucagon and pancreaticpolypeptide immunonegative, but stained strongly forPDX-1. About half of these cells lacked a classical islet cellhormonal phenotype, whereas the remainder was found tobe somatostatin� �-cells (data not shown). Since it hasbeen proposed that somatostatin�/PDX-1� cells may serveas �-cell progenitors during islet compensatory growth(42), this observation may relate to new �-cell develop-ment within the islet periphery of young ZF rats.
DISCUSSION
The ZF model has been widely used to study the stepwisefunctional (2,25,35) and morphological (1,43) adaptationsin islet �-cells that occur in response to insulin resistanceand obesity. In the current study, we have uncovered anovel mechanism for the �-cell expansion in ZF rats thatincorporates enhanced neogenesis, a transient rise inproliferation, and increased �-cell survival potential andprovides evidence of potential roles for Akt/PKB kinase inmediating these responses.
At the outset of these experiments, we had anticipatedthat the �-cell mass of ZF rats would grow in concordancewith their progressive insulin resistance and peak whensevere insulin resistance is established. We were unable toestablish the dominant mechanisms of �-cell mass expan-sion in adult ZF rats due to the fact that a steady-stateincrease in mass was already established by the time ofadulthood. In fact, we found that the �-cell mass of ZF ratswas substantially elevated by 5 weeks of age. Althoughseveral studies on ZF rats have considered the 10- to12-week-old age period when insulin resistance is fully
FIG. 4. �-Cell neogenesis is enhanced in youngZF rats. A: A �-cell neogenesis assay: a tally ofpresumed nascent �-cells not accounted for inthe �-cell mass measurements, whereby singleinsulin� cells, doublets, etc., up to 15 cells percluster (intervals every 100 �m), were groupedand charted. Significant increases in very smallclusters were observed in the 20- and 28-day-oldZF rat pups (*P < 0.03; **P < 0.01). �, ZLC rats;■ , ZF rats. B: Representative fields from a 28-day-old ZLC and a ZF (C) rat pancreas stainedfor insulin (green), PDX-1 (red), and amylase(blue). *Acini-associated �-cells are prevalentin the ZF rat. �-Cells are also associated withsmall intercalated ducts (arrow points to lu-men).
�-CELL MASS COMPENSATION IN THE ZUCKER RAT
2300 DIABETES, VOL. 54, AUGUST 2005

established, mild insulin resistance in skeletal muscle hasbeen detected as early as 30 days of age (44). This earlyperiod of mild insulin resistance temporally correlateswell with our finding of increased �-cell mass and prolif-eration. Our investigation of earlier time points has shownthat ZF pups are not born with enhanced �-cell mass;instead, a significant increase was observed just beforeweaning (at 20 days), which clearly precedes majorchanges in energy metabolism after 23 days that result inhyperphagia, hyperinsulinemia, and significant weight gain(29). Thus, our results demonstrate an apparent temporaldisparity between the �-cell expansion and ensuing meta-bolic alterations that merits further study.
We thus focused on ZF rat pups before 5 weeks of ageand have demonstrated that the key insulin signalingmolecule, Akt, is activated in ducts, acini, and �-cells.Downstream of IRS-2/PI3-kinase signaling, Akt, and inparticular, the Akt-2 isoform (13) appears to be a crucialelement in compensatory �-cell expansion (10,11). Acti-vated (phospho-) Akt staining levels in the duct epitheliumwere dramatically enhanced in ZF rats in the postweaningperiod. Furthermore, the most intense PDX-1� cells con-sistently costained for phospho-Akt. This observation issignificant, since studies of mouse models harboring mu-tations of genes encoding key insulin signaling intermedi-ates indicate that the link between the insulin signalingpathway and �-cell mass compensation to insulin resis-tance is through the stimulation of pdx-1 expression vianuclear exclusion of the forkhead protein transcriptionfactor Foxo1 in �-cells, an established target of Akt(14,15). Thus, the phospho-Akt�/PDX-1� cells we have
identified in ducts and centroacinar regions of young ZFrats may represent a novel insulin/IGF-responsive progen-itor for �-cell neogenesis, although this remains to beproven.
In addition to the exocrine duct epithelium and cen-troacinar region, we also observed increased phospho-Aktimmunoreactivity in islet �-cells of ZF rats. In �-cells, Akthas been proposed to function in mediating proliferation(10,22,24,45), size (10,11), and survival (11,12,21,23,24).Enhanced �-cell proliferation in ZF rats was limited to arestricted time (31 days of age) that preceded the onset ofthe largest �-cell mass increase (�3.8-fold at 35 days ofage). Since Akt has been ascribed with roles associatedwith enhanced �-cell proliferation in vivo (10), futurestudies are necessary to determine whether it may beserving such a role in the young ZF rat.
The Bcl-2 family member BAD, a well-established sub-strate of Akt that is inhibited upon phosphorylation,functions as a proapoptotic factor by antagonizing Bcl-2activity, a related prosurvival protein (46). Both BAD andBcl-2 are expressed in �-cells and have been implicated inthe coordinate control of �-cell apoptosis/survival, respec-tively (46). We detected both increased phospho-BAD andBcl-2 staining in �-cells of ZF pups by immunostaining thatis consistent with the notion of an enhanced survivalpotential of �-cells of ZF rats. Elevated Bcl-2 stainingmight also represent increased compartmentalization ofthe protein to cellular membranes. We speculate fromthese findings in young rats that activated Akt in �-cellsmay be targeted to mitochondrial BAD, thus inhibiting itsfunction and promoting �-cell survival. We failed to detect
FIG. 5. Akt/PKB activation in ducts and aciniof young Zucker rats. A: In the main lining andevaginations (B) of the common pancreaticduct of 25-day-old ZF rats, epithelial cells (ar-rows) with strong nuclear PDX-1 staining(blue) also exhibit intense phospho-Akt stain-ing (red). These cells were rare in the ZLCrats. Lumen is marked with an asterisk (*). C:A section through a small intercalated duct(dotted line) showing a PDX-1�/phospho-Akt�/insulin- cell (3), and PDX-1�/phospho-Akt- cells (�), which are insulin� (green). D:An acinus (dotted line) with a central regioncomposed of PDX-1�/phospho-Akt� cells (3),a glucagon� cell (blue), and a PDX�/phospho-Akt–negative cell (�) that is probably insulinexpressing.
T.L. JETTON AND ASSOCIATES
DIABETES, VOL. 54, AUGUST 2005 2301

decreased apoptosis in the ZF rat pancreas based on insitu apoptosis assays, but the rarity of these profiles(0.1% in all rats and time points), probably from theirrapid engulfment from phagocytes or neighboring �-cells(47), made this approach ineffective to demonstratechanges in apoptosis. However, we quantified apoptosis in�-cells from 4.5-week-old ZF and ZLC pups using anotherstrategy. Hence, we observed increased survival potentialin isolated, short-term cultured islets from young ZF ratsbased on decreased TUNEL staining. Relatedly, a recentstudy of pdx-1�/� haploinsufficient mice has ascribedPDX-1 with a pro-survival role in islets (48). Although themechanisms involved are unresolved, a similar role forPDX-1 could be operating in the pancreas of young ZF rats,although this has yet to be explored.
Akt/PKB has been proposed to regulate �-cell size(10,11) and may also be playing such a role in the �-cellsof ZF rats. Transgenic overexpression of Akt in mouse�-cells results not only in a several-fold enhancement in�-cell mass, but also increased �-cell size, as well as theirheightened survival potential (11). A global knockout ofthe gene encoding for the ribosomal protein p70S6-kinase 1,a downstream target in the Akt-mTOR pathway, results inglucose intolerance and atrophied �-cells (49). Collec-tively, Akt in �-cells may control size through regulation ofp70S6-kinase activation, hence, increasing protein transla-tion and cellular growth. We have determined that �-cellsize, as measured by surface area, is enhanced 33% in5-week-old ZF rats with over a 3.5-fold enhancement in�-cell mass. The magnitude of this growth is actually more
FIG. 6. Increased phospho-Akt, phospho-BAD, and Bcl-2 in islets from young ZF rats. A and B: A typical 25-day-old ZLC control rat islet showingvery low levels of phospho-Akt (red) and phospho-BAD (blue). Islet boundaries are indicated with dotted lines. C and D: Comparable field of arepresentative islet from a young ZF rat exhibiting markedly increased phospho-Akt and phospho-BAD immunostaining, respectively. E and F: Arepresentative field of an islet from a 28-day-old ZLC rat stained for Bcl-2 (red) and insulin (green). �-Cell boundaries are indicated with dottedlines. G and H: A representative field of an islet from a 28-day-old ZF rat stained for Bcl-2 and insulin. Note that punctate �-cell Bcl-2immunoreactivity is increased in the ZF islet. I: Representative immunoblots of whole-islet extracts from 30-day-old rats showing no markedchanges in total protein levels of Akt, BAD, or Bcl-2. J: Semiquantitative Bcl-2 immunofluorescence analysis revealed a more than twofoldincrease in the �-cell signal in 28-day-old ZF rats (*P < 0.05).
�-CELL MASS COMPENSATION IN THE ZUCKER RAT
2302 DIABETES, VOL. 54, AUGUST 2005
striking when the volume of the �-cell is considered. Thisaugmentation in �-cell size and overall mass is sustainedthroughout adulthood. Although not studied here, en-hanced Akt signaling in adult ZF �-cells likely impactstheir growth through S6-kinase activation.
We have also demonstrated that strongly phospho-Aktimmunopositive, but insulin-negative cells in the isletperiphery of young Zucker rats also stain intensely fornuclear PDX-1. About 50% of these cells lack a typical isletpeptide hormone phenotype, but half are somatostatin�
�-cells. A lineal relationship of �- and �-cells has beenreported during both embryonic development (50) as wellas during islet regeneration in the adult (42). Hence, theseislet-borne PDX-1�/phospho-Akt�/somatostatin�/� cellsmay represent a �-cell precursor and might also contributeto the �-cell mass increase in ZF rats.
In summary, we have presented evidence suggestingthat at almost all examined time points, �-cell differentia-tion from progenitor cells (neogenesis) is significantlyenhanced in ZF rats, as judged by significantly increasednumbers of �-cell clusters and single �-cells associatedwith the exocrine compartment. Additional support for�-cell neogenesis was a significant increase in ductalnuclear PDX-1 staining. Increased ductal PDX-1 expres-sion in regeneration models has been correlated with theacquisition of a “de-differentiated” phase analogous to anembryonic state that, in turn, may lead to new �-celldevelopment (2). Although these criteria serve as onlyindirect evidence of neogenesis, we surmise that thesecells may migrate and aggregate to form new islets,although this remains to be proven. Unexpectedly, islet�-cell replication rates between the ZF and control ratsdiffer only at a single time point tested, at 4.5 weeks of age.These results are in contrast to recent reports highlighting�-cell replication as the principal contributor to �-cellgrowth in the adult (16–18).
We have established several key features regarding theonset and the nature of early �-cell mass compensation inyoung Zucker rat pups and have provided data supportinga central role for Akt kinase. We have identified putativeAkt targets that likely function in growth, differentiation,and enhanced survival of �-cells. Furthermore, we haveobtained data suggesting that the survival potential of islet�-cells is enhanced in young ZF rats. Accordingly, itappears that Akt, a key insulin signaling intermediate,orchestrates both new �-cell development as well asheightened survival (anti-apoptosis) potential of existing
�-cells. Importantly, we have also localized activated(phospho-) Akt to specific PDX� cells of ducts, acini, andislets of young ZF rats. Thus, in the leptin-signaling defi-cient ZF rats, mechanisms of �-cell mass expansion areclearly in play early in life to insure a sufficient insulinsupply for the greatly increased insulin requirements laterin adulthood.
ACKNOWLEDGMENTS
This work was supported by research awards from theAmerican Diabetes Association to J.L. and T.L.J.
REFERENCES
1. Pick A, Clark J, Kubstrup C, Levisetti M, Pugh W, Bonner-Weir S, Polonsky,KS: Role of apoptosis in failure of �-cell mass compensation for insulinresistance and beta-cell defects in the male Zucker diabetic fatty rat.Diabetes 47:358–364, 1998
2. Bonner-Weir S: Islet growth and development in the adult. J Mol Endocri-
nol 24:297–302, 20003. Kido Y, Burks DJ, Withers D, Bruning JC, Kahn CR, White MF, Accili D:
Tissue-specific insulin resistance in mice with mutations in the insulinreceptor, IRS-1, and IRS-2. J Clin Invest 105:199–205, 2000
4. Withers D, Gutierrez JS, Towery H, Burks DJ, Ren JM, Previs S, Zhang Y,Bernal D, Pons S, Shulman GI, Bonner-Weir S, White MF: Disruption ofIRS-2 causes type 2 diabetes in mice. Nature 391:900–904, 1998
5. Withers DJ, Burks DJ, Towery HH, Altamuro SL, Flint CL, White MF: Irs-2coordinates Igf-1 receptor-mediated beta-cell development and peripheralinsulin signalling. Nat Genet 23:32–40, 1999
6. Kubota N, Tobe K, Terauchi Y, Eto K, Yamauchi T, Suzuki R, Tsubamoto Y,Komeda K, Nakano R, Miki H, Satoh S, Sekihara H, Sciacchitano S, LesniakM, Aizawa S, Nagai R, Kimura S, Akanuma Y, Taylor SI, Kadowaki T:Disruption of insulin receptor substrate 2 causes type 2 diabetes becauseof liver insulin resistance and lack of compensatory �-cell hyperplasia.Diabetes 49:1880–1889, 2000
7. Jhala US, Canettieri G, Screaton RA, Kulkarni RN, Krajewski S, Reed J,Walker J, Lin X, White M, Montminy M: cAMP promotes pancreaticbeta-cell survival via CREB-mediated induction of IRS2. Genes Dev 17:1575–1580, 2003
8. Kubota N, Terauchi Y, Tobe K, Yano W, Suzuki R, Ueki K, Takamoto I,Satoh H, Maki T, Kubota T, Moroi M, Okada-Iwabu M, Ezaki O, Nagai R,Ueta Y, Kadowaki T, Noda T: Insulin receptor substrate 2 plays a crucialrole in beta cells and the hypothalamus. J Clin Invest 114:917–927, 2004
9. Lin X, Taguchi A, Park S, Kushner JA, Li F, Li Y, White MF: Dysregulationof insulin receptor substrate 2 in beta cells and brain causes obesity anddiabetes. J Clin Invest 114:908–916, 2004
10. Bernal-Mizrachi E, Wen W, Stahlhut S, Welling CM, Permutt MA: Islet betacell expression of constitutively active Akt1/PKB alpha induces strikinghypertrophy, hyperplasia, and hyperinsulinemia. J Clin Invest 108:1631–1638, 2001
11. Tuttle RL, Gill NS, Pugh W, Lee JP, Koeberlein B, Furth EE, Polonsky KS,Naji A, Birnbaum MJ: Regulation of pancreatic beta-cell growth andsurvival by the serine/threonine protein kinase Akt1/PKBalpha. Nat Med
7:1133–1137, 200112. Srinivasan S, Bernal-Mizrachi E, Ohsugi M, Permutt MA: Glucose promotes
FIG. 7. Akt activation in non–� islet cells ofyoung Zucker rats. A: A typical islet of a young25-day-old ZLC rat exhibiting a peripheralnon–�-cell with phospho-Akt staining (ar-row). B: Comparable field of a typical isletfrom a young ZF rat showing increased prev-alence of these strongly PDX�/phospho-Akt�
peripheral cells (arrows). The red channelwas underexposed to clearly reveal thestrongly Akt� peripheral cells. These cellswere glucagon and pancreatic polypeptidenegative, but half were determined to be so-matostatin� �-cells.
T.L. JETTON AND ASSOCIATES
DIABETES, VOL. 54, AUGUST 2005 2303

pancreatic islet beta-cell survival through a PI 3-kinase/Akt-signalingpathway. Am J Physiol Endocrinol Metab 283:E784–E793, 2002
13. Garofalo RS, Orena SJ, Rafidi K, Torchia AJ, Stock JL, Hildebrandt AL,Coskran T, Black SC, Brees DJ, Wicks JR, McNeish JD, Coleman KG:Severe diabetes, age-dependent loss of adipose tissue, and mild growthdeficiency in mice lacking Akt2/PKB beta. J Clin Invest 112:197–208, 2003
14. Nakae J, Biggs WH, Kitamura T, Cavenee WK, Wright CV, Arden KC, AcciliD: Regulation of insulin action and pancreatic beta-cell function bymutated alleles of the gene encoding forkhead transcription factor Foxo1.Nat Genet 32:245–253, 2002
15. Kitamura T, Nakae J, Kitamura Y, Kido Y, Biggs WH, Wright CV, White MF,Arden KC, Accili D: The forkhead transcription factor Foxo1 links insulinsignaling to Pdx1 regulation of pancreatic beta cell growth. J Clin Invest
110:1839–1847, 200216. Kulkarni RN, Jhala US, Winnay JN, Krajewski S, Montminy M, Kahn CR:
PDX-1 haploinsufficiency limits the compensatory islet hyperplasia thatoccurs in response to insulin resistance. J Clin Invest 114:828–836, 2004
17. Dor Y, Brown J, Martinez OI, Melton DA: Adult pancreatic beta-cells areformed by self-duplication rather than stem-cell differentiation. Nature
429:41–46, 200418. Georgia S, Bhushan A: Beta cell replication is the primary mechanism for
maintaining postnatal beta cell mass. J Clin Invest 114:963–968, 200419. Jetton TL, Liu YQ, Trotman WE, Nevin PW, Sun XJ, Leahy JL: Enhanced
expression of insulin receptor substrate-2 and activation of protein kinaseB/Akt in regenerating pancreatic duct epithelium of 60%-partial pancrea-tectomy rats. Diabetologia 44:2056–2065, 2001
20. Accili D: A kinase in the life of the beta cell. J Clin Invest 108:1575–1576,2001
21. Dickson LM, Rhodes CJ: Pancreatic beta-cell growth and survival in theonset of type 2 diabetes: a role for protein kinase B in the Akt? Am J
Physiol Endocrinol Metab 287:E192–E198, 200422. Holst LS, Mulder H, Manganiello V, Sundler F, Ahren B, Holm C, Degerman
E: Protein kinase B is expressed in pancreatic beta cells and activatedupon stimulation with insulin-like growth factor I. Biochem Biophys Res
Commun 250:181–186, 199823. Maeda H, Rajesh KG, Maeda H, Suzuki R, Sasaguri S: Epidermal growth
factor and insulin inhibit cell death in pancreatic beta cells by activation ofPI3-kinase/AKT signaling pathway under oxidative stress. Transplant Proc
36:1163–1165, 200424. Wang Q, Li L, Xu E, Wong V, Rhodes C, Brubaker PL: Glucagon-like
peptide-1 regulates proliferation and apoptosis via activation of proteinkinase B in pancreatic INS-1 beta cells. Diabetologia 47:478–487, 2004
25. Bray GA: The Zucker-fatty rat: a review. Fed Proc 36:148–153, 197726. Cleary MP, Phillips FC, Morton RA: Liver, serum and adipose tissue fatty
acid composition in suckling Zucker rats. Lipids 29:753–758, 199427. Takaya K, Ogawa Y, Isse N, Okazaki T, Satoh N, Masuzaki H, Tamura N,
Hosoda K, Nakao K: Molecular cloning of rat leptin receptor isoformcomplementary DNAs: identification of a missense mutation in Zuckerfatty (fa/fa) rats. Biochem Biophys Res Commun 225:75–83, 1996
28. Phillips MS, Liu Q, Hammond HA, Dugan V, Hey PJ, Caskey CT, Hess JF:Leptin receptor missense mutation in the fatty Zucker rat. Nat Genet
13:18–19, 199629. Truett GE, Walker JA, Harris RB: A developmental switch affecting growth
of fatty rats. Am J Physiol Regul Integr Comp Physiol 279:R1956–R1963,2000
30. Williams G, Bing C, Cai XJ, Harrold JA, King PJ, Liu XH: The hypothalamusand the control of energy homeostasis: different circuits, different pur-poses. Physiol Behav 74:683–701, 2001
31. Gould E, Gross CG: Neurogenesis in adult mammals: some progress andproblems. J Neurosci 22:619–623, 2002
32. Ezaki T: Antigen retrieval on formaldehyde-fixed paraffin sections: its
potential drawbacks and optimization for double immunostaining. Micron
31:639–649, 200033. Jetton TL, Liang Y, Cincotta AH: Systemic treatment with sympatholytic
dopamine agonists improves aberrant beta-cell hyperplasia and GLUT2,glucokinase, and insulin immunoreactive levels in ob/ob mice. Metabolism
50:1377–1384, 200134. York D, Steinke J, Bray GA: Hyperinsulinemia and insulin resistance in
genetically obese rats. Metabolism 21:277–284, 197235. Argiles JM: The obese Zucker rat: a choice for fat metabolism 1968–1988:
twenty years of research on the insights of the Zucker mutation. Prog
Lipid Res 28:53–66, 198936. Scaglia L, Cahill CJ, Finegood DT, Bonner-Weir S: Apoptosis participates in
the remodeling of the endocrine pancreas in the neonatal rat. Endocrinol-
ogy 138:1736–1741, 199737. Lipsett M, Finegood DT: �-cell neogenesis during prolonged hyperglycemia
in rats. Diabetes 51:1834–1841, 200238. Song KH, Ko SH, Ahn YB, Yoo SJ, Chin HM, Kaneto H, Yoon KH, Cha BY,
Lee KW, Son HY: In vitro transdifferentiation of adult pancreatic acinarcells into insulin-expressing cells. Biochem Biophys Res Commun 316:1094–1100, 2004
39. De Haro-Hernandez R, Cabrera-Munoz L, Mendez JD: Regeneration ofbeta-cells and neogenesis from small ducts or acinar cells promoterecovery of endocrine pancreatic function in alloxan-treated rats. Arch
Med Res 35:114–120, 200440. Nagasao J, Yoshioka K, Amasaki H, Mutoh K: Centroacinar and interca-
lated duct cells as potential precursors of pancreatic endocrine cells in ratstreated with streptozotocin. Ann Anat 185:211–216, 2003
41. Datta SR, Brunet A, Greenberg ME: Cellular survival: a play in three Akts.Genes Dev 13:2905–2927, 1999
42. Guz Y, Nasir I, Teitelman G: Regeneration of pancreatic beta cells fromintra-islet precursor cells in an experimental model of diabetes. Endocri-
nology 142:4956–4968, 200143. Shino A, Matsuo T, Iwatsuka H, Suzuoki Z: Structural changes of pancre-
atic islets in genetically obese rats. Diabetologia 9:413–421, 197344. Penicaud L, Ferre P, Terretaz J, Kinebanyan MF, Leturque A, Dore E,
Girard J, Jeanrenaud B, Picon L: Development of obesity in Zucker rats:early insulin resistance in muscles but normal sensitivity in white adiposetissue. Diabetes 36:626–631, 1987
45. Trumper K, Trumper A, Trusheim H, Arnold R, Goke B, Horsch D:Integrative mitogenic role of protein kinase B/Akt in beta-cells. Ann N Y
Acad Sci 921:242–250, 200046. Federici M, Hribal M, Perego L, Ranalli M, Caradonna Z, Perego C, Usellini
L, Nano R, Bonini P, Bertuzzi F, Marlier LN, Davalli AM, Carandente O,Pontiroli AE, Melino G, Marchetti P, Lauro R, Sesti G, Folli F: High glucosecauses apoptosis in cultured human pancreatic islets of Langerhans: apotential role for regulation of specific Bcl family genes toward anapoptotic cell death program. Diabetes 50:1290–1301, 2001
47. Trudeau JD, Dutz JP, Arany E, Hill DJ, Fieldus WE, Finegood DT: Neonatalbeta-cell apoptosis: a trigger for autoimmune diabetes? Diabetes 49:1–7,2000
48. Johnson JD, Ahmed NT, Luciani DS, Han Z, Tran H, Fujita J, Misler S,Edlund H, Polonsky KS: Increased islet apoptosis in Pdx1�/� mice. J Clin
Invest 111:1147–1160, 200349. Pende M, Kozma SC, Jaquet M, Oorschot V, Burcelin R, Le Marchand-
Brustel Y, Klumperman J, Thorens B, Thomas G: Hypoinsulinaemia,glucose intolerance and diminished beta-cell size in S6K1-deficient mice.Nature 408:994–997, 2000
50. Guz Y, Montminy MR, Stein R, Leonard J, Gamer LW, Wright CV, TeitelmanG: Expression of murine STF-1, a putative insulin gene transcription factor,in beta cells of pancreas, duodenal epithelium and pancreatic exocrine andendocrine progenitors during ontogeny. Development 121:11–18, 1995
�-CELL MASS COMPENSATION IN THE ZUCKER RAT
2304 DIABETES, VOL. 54, AUGUST 2005


















