
Makalah Fosforilasi Oksidatif (1)
41
HEME SPIN RENDAH SITOKROM OKSIDASE C SEBAGAI PENDORONG PROSES POMPA PROTON Oleh: Anne Carolina (20504011) Eka Wulandari (20504012)
Transcript of Makalah Fosforilasi Oksidatif (1)

HEME SPIN RENDAH SITOKROM OKSIDASE C SEBAGAI
PENDORONG PROSES POMPA PROTON
Oleh:
Anne Carolina (20504011)
Eka Wulandari (20504012)
PROGRAM MAGISTER KIMIA
BIDANG KHUSUS BIOKIMIA
INSTITUT TEKNOLOGI BANDUNG
2005

ABSTRAK
Sitokrom c oksidase mitokondria berperan penting dalam respirasi seluler aerob, mereduksi oksigen menghasilkan air, dalam proses kopling dengan pompa hidrogen melewati membran mitokondria dalam. Residu aspartat, Asp-51, yang 0berlokasi dekat permukaan enzim, mengalami perubahan struktur dalam Sinar-X. Hal ini mengindikasikan bahwa residu ini berperan dalam proses pemompaan proton. Meskipun bukti keterlibatan mekanistik dan fungsional dari residu ini dalam proses pemompaan proton belum jelas diperoleh, mutasi Asp-52 Asn dari enzim hati memperlihatkan fungsi pompa proton tanpa mempengaruhi aktivitas reduksi dioksigen. Struktur Sinar-X (pada resolusi 1.8/1.9 Å dalam bentuk teroksidasi dan reduksi penuh) menunjukkan bahwa muatan positif total yang dibentuk selama oksidasi mendorong transport proton aktif dari ruang mitokondria ke Asp-51 melewati enzim lewat saluran air dan jaringan ikatan hidrogen, yang berlokasi di tandem. Selain itu reduksi enzim menginduksi pengeluaran proton dari aspartat ke mitokondria bagian luar. Ikatan peptida dalam jaringan ikatan hidrogen menginhibisi transfer balik proton melalui jaringan. Perubahan redoks dalam kapasitas saluran air, diinduksi oleh gugus hidroksi farnesil etil dari heme spin-rendah. Hal ini mengindikasikan bahwa fungsi saluran air sama efektifnya dengan daerah pengumpul proton. Hasil infrared mengindikasikan bahwa konformasi Asp-51 dikontrol hanya oleh bentuk oksidasi heme spin rendah. Hasil ini mengindikasikan bahwa heme spin rendah mendorong proses pompa proton.

BAB I
PENDAHULUAN
Sitokrom c oksidase yang berlokasi di bagian dalam membran mitokondria
merupakan enzim kunci pada rantai respirasi organisme aerob. Enzim ini
berfungsi dalam katalisis transfer elektron dari sitokrom c ke oksigen molekular,
sehingga terjadi reduksi menjadi air. Reaksi ini terjadi melalui proses transport,
yaitu melalui pemompaan empat proton melewati membran. Sitokrom c oksidase
memiliki empat kofaktor reaksi redoks yang aktif, yaitu dua atom Cu pembentuk,
dinamakan CuA, suatu heme a spin rendah, serta pusat binuklir heme a3 dan Cu B.
Cu A menerima elektron dari sitokrom c dan elektron ditransfer melalui heme a ke
pusat binuklir tempat terjadinya reduksi oksigen.
Kesetimbangan elektron dicapai dari sitokrom c di luar membran dalam
mitokondria (ruang inter membran) melalui sisi Cu A dan heme spin rendah (heme
a) ke sisi reduksi O2. Proton yang digunakan untuk pembentukan air dari O2,
ditransfer dari membran dalam mitokondria (ruang matriks) melalui dua jaringan
ikatan hidrogen yang disebut jalur K dan D. Selain dalam proses pompa proton,
transfer proton dan elektron ke sisi reduksi O2 menghasilkan perpindahan muatan
positif ke ruang intermembran, yang menghasilkan gaya proton motive yang
diperlukan untuk menggerakkan ATP sintase.

Mutasi residu asam amino di dalam jalur D dapat menyebabkan penurunan
dalam efesiensi pompa proton dan penurunan aktivitas reduksi O2. Pengamatan
menunjukkan bahwa proses yang menyertai pompa proton terjadi melalui jalur
proton untuk membentuk air. Struktur Sinar-X memperlihatkan bahwa sitokrom c
oksidase dari hati sapi dalam bentuk teroksidasi dan tereduksi penuh pada resolusi
2.3 dan 2.35 Å, menunjukkan adanya pergerakan Asp-51 dari subunit I (subunit
paling besar yang mengandung heme a dan sisi reduksi O2) dari interior protein
menuju permukaan inter membran melalui reduksi enzim. Dalam bentuk
teroksidasi, Asp-51 berkontak dengan ruang matriks melalui jaringan ikatan
hidrogen sehingga molekul air dapat melewati ruang dalam matriks (jalur H).
Struktur ini memberikan gambaran bahwa proses pompa proton terjadi pada Asp-
51. Akan tetapi usulan mengenai hal ini belum banyak diterima, karena enzim
bakteri dan tumbuhan tidak mempunyai residu analog Asp-51 (penomoran dalam
sapi) dan juga karena mutasi jalur D menyebabkan penurunan efisiensi pompa
proton dan aktivitas reduksi O2. selain itu tidak ada mekanisme untuk penggerak
pompa proton pada Asp-51 yang menjelaskan. Mutasi Asp-51 memberikan satu
alternatif untuk menjelaskan fungsi Asp-51. Dalam penelitian ini akan dijelaskan
peranan jalur H melalui sisi mutagenesis terarah, struktur Sinar-X dan
spektroskopi infra merah.

BAB II
TINJAUAN PUSTAKA
Semua tahap-tahap enzimatis pada degradasi oksidatif karbohidrat, lemak
dan asam amino didalam sel aerobik menyatu menjadi tahap akhir respirasi sel.
Disini terjadi pengaliran electron dari senyawa organik menuju oksigen,
menghasilkan energi untuk membuat ATP dari ADP dan fosfat. Rantai respirasi
terdiri dari serangkaian protein dengan gugus prostetik yang terikat kuat dan
mampu menerima dan memberikan elektron.
Pada sel eukariot, hampir semua dehidrogenase spesifik yang diperlukan
pada oksidasi piruvat dan bahan bakar lain melalui siklus asam sitrat terletak pada
bagian sebelah dalam mitokondria yaitu matriks. Molekul pemindah electron dari
rantai respirasi dan molekul enzim yang melakukan sintesa ATP dari ADP dan
fosfat terbenam dalam membran sebelah dalam. Bahan bakar siklus asam sitrat
seperti piruvat, harus dipindahkan dari sitosol (tempat dilakukannya sintesis
molekul-molekul tersebut) melalui membran mitokondria ke dalam bagian
matriks disebelah dalam, sebagai tempat aktivitas dehidrogenase.
Demikian pula, ADP yang dibentuk dari ATP selama aktivitas yang
memerlukan energi dari sitosol harus dipindahkan ke dalam matriks mitokondria,
untuk mengikat fosfat kembali, menjadi ATP. ATP baru yang dibentuk harus
dikembalikan ke sitosol.

Jadi membran mitokondria sebelah dalam, merupakan struktur komplek
yang mengandung molekul pembawa electron, sejumlah enzim dan beberapa
system transport membran.
Gambar Proses Fosforilasi Oksidatif dalam Mitokondria
Beberapa jenis gugus pembawa elektron, semuanya berikatan dengan protein.
Golongannya antara lain Nikotinamida adenin dinukleotida (NAD) yang aktif
dengan dehidrogenase; flavin mononukleotida (FMN) pada NADH
dehidrogenase;ubiquinon atau koenzim Q, suatu senyawa kuinon isoprenoid yang
larut dalam lemak, yang berfungsi dalam bentuk ikatannya dengan satu atau lebih
protein; dua jenis protein yang mengandung pusat besi-sulfur (Fe-S) sitokrom;
dan tembaga pada sitokrom aa3.
Senyawa sitokrom adalah protein mengandung besi pemindah elektron dan
berwarna merah atau coklat, yang bekerja secara berurutan untuk mengangkut
elektron dari ubikuinon ke molekul oksigen. Golongan ini merupakan protein
heme, dengan besi yang berada pada kompleks porfirin-besi, atau heme, yang
serupa dengan pada hemoglobin.Terdapat tiga kelas sitokrom, a, b, dan c yang

berbeda dalam spektra absorbsi sinarnya. Setiap jenis sitokrom dalam keadaaan
tereduksinya atau ferro memiliki tiga pita absorbsi yang jelas pada kisaran sinar
tampak.
Sitokrom pada rantai respirasi disusun dalam urutan b c1 aa3. Sitokrom b
berada dalam dua bentuk, menerima elektron dari ubikuinon dan
memindahkannya ke sitokrom c1 yang selanjutnya memberikan elektron yang
diterima ke sitokrom c. Setiap sitokrom berada dalam bentuk feri [Fe(III)]
menerima satu elektron menjadi bentuk fero [Fe(II)]. Pembawa elektron terakhir
adalah sitokrom aa3 atau oksidase sitokrom yang dapat memberikan elektron
langsung ke oksigen untuk menyempurnakan proses transport elektron.
Sitokrom aa3 berbeda dengan sitokrom lain. Protein ini mengandung dua molekul
heme A yang terikat kuat, yang berbeda dari protoheme oada hemoglobin, dalam
cincin porfirinnya, yang memiliki rantai sisi hidrokarbon yang panjang. Lebih
jauh lagi sitokrom aa3 juga mengandung dua atom tembaga yang esensial. Setelah
komponen sitokrom a menerima elektron dari sitokrom c dab tereduksi menjadi
bentuk Fe(II), molekul ini memberikan elektronnya ke sitokrom a3. Sitokrom a3
tereduksi lalu memberikan elektron kepada molekul oksigen. Unsur yang
berpartisipasi dengan kedua gugus heme didalam proses ini adalah kedua atom
tembaga yang terikat, yang mengalami perubahan redoks kupro-kupri [Cu(I)-
Cu(II)] dalam fungsinya. Ini adalah suatu tahap yang kompleks dan penting di
dalam transport elektron, karena ke empat elektron harus diberikan hampir
bersamaan kepada O2 untuk menghasilkan dua H2O, dengan mengambil empat H+

dari medium cair. Dari semua anggota rantai transport elektron, hanya sitokrom
aa3 yang dapat bereaksi langsung dengan oksigen.

BAB III
METODOLOGI PENELITIAN
3.1 Mutagenesis Terarah (Site-directed Mutagenesis)
Jumlah sitokrom c pada mitokodria ditentukan menggunakan western blot.
Sejumlah sampel dijalankan pada on 4–12% Mes NuPage gels (Novex) selama 42
menit pada 200V dan 40C. Kemudian electrotransfer basah ke membran
nitroselulosa menggunakan sistem Novex dengan 5% metanol pada buffer transfer
selama 1 jam pada 30V. Efisiensi transfer dipertegas dengan noda transfer awal
pada gel dan membran menggunakan EasyStain (Novex) and Ponceau S.
Membran residu noda dicuci menggunakan Tris-buffer saline dengan
0.2% Tween- 20 (TBS-T),yang diblok semalam menggunakan 1.5% susu bubuk
pada TBS-T. Sitokrom c ditandai menggunakan imunoglobulin antibodi sitokrom
c oksidasi dari tikus dengan pengenceran 1 : 2000 dengan TBS-T dengan 1,5%
selama 1 hari. Inkubasi selanjutnya menggunakan secondary anti tikus Ig
berikatan dengan horseradish peroxidase (Amersham) pada pengenceran 1:2000
selama 40 menit, pitanya divisualisasikan menggunakan reagen luminol dan film
sinar-x.

3.2 Pemurnian Sitokrom c Oksidase dari Hati Sapi dan Kristalisasi
Penyiapan pemurnian enzim dengan rekristalisasi berulang adalah
pengkondisian kristalisasi enzim yang sangat penting. Kristal dalam bentuk
reduksi penuh dan bentuk reduksi ikatan-CO dimulai dengan merendam bentuk
teroksidasi penuh dengan medium yang mengandung askorbat dan sejumlah
katalitik sitokrom c sebagai sistem pereduksi dan polietilen glikol (PEG 4000)
untuk menstabilkan kristal dibawah atmosfer N2 dan CO.
Difraksi sinar-X dilakukan dengan cara menempatkan kristal pada kapiler
yang sesuai dengan medium perendaman.
Keadaan teroksidasi dan keadaan pengikatan ligan dari enzim dipertegas
dengan spektra absorpsi darikristal ini pada kondisi medium yang sama.
Bentuk azida disiapkan dengan merendam kristal pada keadaan teroksidasi penuh
ke dalam buffer yang mengandung azida dan jumlah PEG 4000 yang sesuai pada
keadaan aerobik. Data intensitas diambil menggunakan radiasi synchroton pada
1.0 Å Photon Factory, Tsukuba, Japan, dengan modified Weissenberg camera
untuk makromolekul.

BAB IV
HASIL DAN PEMBAHASAN
4.1 Mutagenesis Asp-51 dalam Subunit I Hati Sapi Sitokrom c Oksidase
Subunit I Asp-51 digantikan oleh Asn melalui metode pembentukan
hibrid. Vektor ekspresi dibuat untuk memproduksi, di dalam sitosol sel HeLa,
yang mengkode subunit I dari enzim sapi dengan signal targeting mitokondria
pada terminal N- dan ekor heksahistidin pada terminal C-, untuk penentuan
kuantitatif subunit I yang terekspresi. Struktur Sinar-X menunjukkan bahwa
terminal C subunit I mempunyai ruang yang cukup untuk menerima tag (ekor)
His tanpa mempengaruhi konformasi enzim.
Efisiensi produksi enzim hibrid sapi/manusia dievaluasi dengan antibodi
yang spesifik. Seperti yang ditunjukkan oleh analisis western blot dari SDS
mitokondria hati sapi dan Sel HeLa. Antibodi Subunit I sapi menunjukkan pita
yang jelas tapi tidak pada Subunit I manusia (Fig. 1A, garis 1 dan 2). Antibodi
untuk protein manusia hanya bereaksi dengan Subunit I manusia tapi tidak
dengan Subunit I sapi (Fig 1A, garis 3 dan 4).

Gambar 1A. Pengaruh mutasi sub unit I Asp-51 3 Asn (Asp51Asn) pada fungsi sitokrom c oksidase. Mitokondria dari hati sapi (jalur 1 dan 3) dan sel HeLa (jalur 2 dan 4) diperlakukan 4% SDS 3,5M urea dan difraksinasi menggunakan SDS PAGE. 70 gram sampel protein dimasukkan kecuali untuk jalur 1 (20gram protein). Antibodi untuk subunit I sapi digunakan untuk jalur 1 dan 2, antobodi sub unit 1 untuk manusia digunakan jalur 3 dan 4.
Dodesil maltosida melarutkan sitokrom c okidase dari membran
mitokondria tanpa mendenaturasi protein dan menghasilkan pita 210 kDa dari
Sitokrom c oksidase dalam blue native PAGE. Antibodi sapi bereaksi dengan
fraksi 210 kDa dari mitokondria yang larut dalam dodesil maltosida. Fraksi ini
diisolasi dari sel HeLa transfektan gen sapi wild-type (Fig 1B, garis 2).
Gambar 1B. Hasil gel elektroforesis di fraksinasi dengan 1,4% dodesil maltosidesolubilized metokondria dari tiruan-transfektan sel HeLa (jalur 1,4 dan 6) dan dari sel HeLa menempatkan wild type (jalur 2 dan 5) dan sub unit I gen mutan Asp51Asn (jalur 3 dan 7) dari sitokrom c oksidase. Pada jalur 1,2, dan 3,180,90 dan 90 gram protein diberikan secara berturut-turut. Pada jalur 4-7 ditambahkan 70 gram protein. Anyibodi spesifik sub unit I untuk sapi digunakan jalur 1-3 atau manusia pada jalur 4-7.

Fraksi yang berhubungan diambil dari suatu sel transfektan oleh vektor
yang tidak membawa gen subunit I (mock-transfected cell line) tidak
menghasilkan pita yang analog (Fig.1B, garis 1). Disamping itu antibodi manusia
bereaksi dengan fraksi dari mock-transfected cell line (Fig 1B, garis 4 dan 6).
Kemudian hasil menunjukkan bahwa subunit I sapi tersusun oleh subunit
manusia. Keberadaan pita yang lemah dari fraksi 210 kDa mitondria yang larut
dari garis cell transfected gen sapi wild-type (Fig. 1B, garis 5) menunjukkan
bahwa subunit I manusia tidak sempurna digantikan oleh subunit I manusia dalam
cell transfected. Jumlah residu dari nonhibrid enzim ditentukan secara kuantitatif
dengan membandingkan intensitas pita dari pita 210 kDa dari mock-transfected
dan gen wild-type transfected cell seperti dijelaskan dalam metode yaitu menjadi
sekitar 20% (Fig. 1B, garis 4 dan 5). Pita yang ditunjukkan dalam Fig. 1B yaitu
pita blue native PAGE, tidak sejelas pita hasil SDS PAGE. Spektrum sinar
tampak dari mitokondria yang larut dalam dodesil maltosida dan mitokondria
tereduksi penuh dari gen wild-type sel transfektan menunjukkan suatu - pita pada
640 nm, yang karakteristik bagi sitokrom c oksidase natif. Tidak ada komponen
mitokondria yang mempunyai absorbansi signifikan pada 604 nm selain sitokrom
c oksidase.
Selanjutnya, preparasi enzim hibrid yang larut memperlihatkan aktivitas
enzim spesifik (laju oksidasi ferositokrom c per molekul enzim) yang sama tinggi
dengan enzim preparasi yang larut dari sel transfektan tiruan. Spektrum dan
aktivitas enzim dari enzim hibrid memberikan bukti kuat bahwa enzim hibrid

membawa konformasi natif. Sel transfektan dengan gen subunit I yang termutasi
Asp51Asn memperlihatkan efisiensi produksi enzim hibrid yang sama, seperti
halnya sel transfektan gen wild-type pada garis diatas (Fig. 1B, garis 2 dan 3).
Spektrum absorpsi dan aktivitas transfer elektron dari preparasi yang bersifat larut
juga sama identik dengan enzim hibrid wild-type. Hasil ini menunjukkan bahwa
mutasi Asp51Asn tidak mengganggu struktur 3D enzim.
Aktivitas transfer elektron dan pompa proton dari enzim hibrid dalam
mitokondria ditentukan dengan mengukur laju oksidasi sitokrom c dan
pengeluaran elektron oleh preparasi mitoplast dengan keberadaan beberapa reagen
pemblok aktivitas komponen mitokondria lain, termasuk valimisin dan KCl untuk
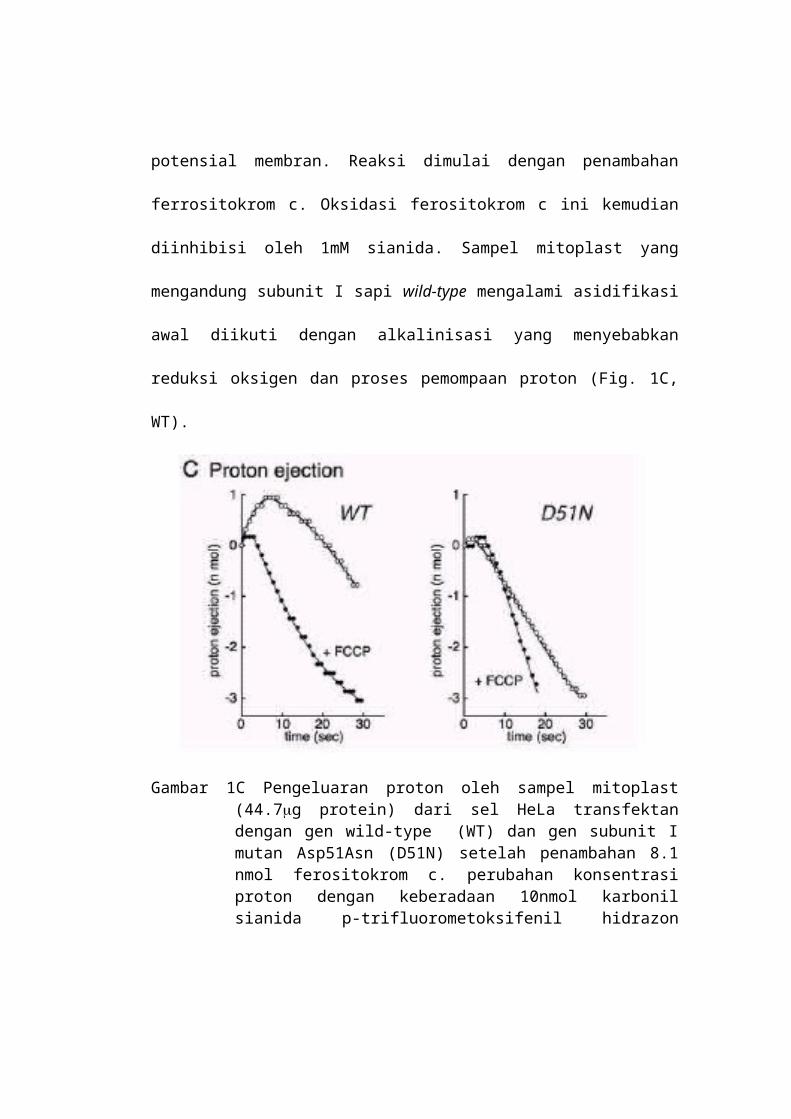
eliminasi potensial membran. Reaksi dimulai dengan penambahan ferrositokrom
c. Oksidasi ferositokrom c ini kemudian diinhibisi oleh 1mM sianida. Sampel
mitoplast yang mengandung subunit I sapi wild-type mengalami asidifikasi awal
diikuti dengan alkalinisasi yang menyebabkan reduksi oksigen dan proses
pemompaan proton (Fig. 1C, WT).

Gambar 1C Pengeluaran proton oleh sampel mitoplast (44.7g protein) dari sel HeLa transfektan dengan gen wild-type (WT) dan gen subunit I mutan Asp51Asn (D51N) setelah penambahan 8.1 nmol ferositokrom c. perubahan konsentrasi proton dengan keberadaan 10nmol karbonil sianida p-trifluorometoksifenil hidrazon (FCCP) per mg protein mitoplast yang ditunjukkan dengan +FCCP
Kurva yang sama diperoleh dari sampel sel transfektan tiruan (tidak
ditunjukkan). Karbonil sianida p-trifluorometoksi fenilhidrazon (FCCP)
memindahkan asidifikasi awal (Gambar 1C, WT + FCCP). Adaptasi awal
alkalinisasi dengan keberadaan FCCP menunjukkan bahwa alkalinisasi karena
reduksi O2 diabaikan dalam 4 detik pertama. Kemudian kecepatan pengeluaran
proton dapat ditentukan dari fase linear dalam 4 detik pertama setelah inisiasi
reaksi. Perkiraan kuantitatif proton dan elektron yang ditransfer dalam 5 detik
pertama adalah 0.96 nmol proton (Gambar 1C, WT) dan 1.17 nmol ekuivalen
electron. Hal ini berarti perbandingan H+/ e- adalah sekitar 0.82. Tiga pengukuran
lain menggunakan sampel mitoplast, termasuk subunit I wild-type sapi dari garis

sel yang sama, memberikan perbandingan 0.83, 0.58, dan 0.79. Harga ini sama
dengan hasil dari sampel mitoplast yang dibuat dari garis sel trasnfektan tiruan
(0.85 dan 0.82). Selain itu harga yang sama juga ditunjukkan oleh enzim mamalia
lain. Hasil ini juga menunjukkan bahwa konformasi natif enzim ada pada enzim
hibrid. Preparasi mitoplast termasuk juga mutan Asp51Asn dari subunit I sapi
tidak menunjukkan adanya asidifikasi awal (Gambar. 1C, D51N). Asidifikasi juga
tidak terlihat dengan keberadaan FCCP (Fig.1C, D51N + FCCP). Mutan mitoplast
memperlihatkan oksidasi ferrositokrom yang sensitif terhadap sianida pada laju
yang lebih cepat sekitar 70% dari wild-type. Kandungan residu enzim manusia
(sekitar 20%) terlalu rendah untuk mendeteksi proses pompa proton pada kondisi
percobaan ini. Hasil ini dipertegas oleh data yang diperoleh dari garis sel
transfektan gen wild-type dan tiga garis sel transfektan gen mutan Asp51Asn
yang berbeda.
Hipotesis endosimbiosis yang diterima secara luas yaitu bahwa asal
muasal organel menyebutkan gen asal ditransfer ke inti selama evolusi. Subunit I
dan sitokrom b yang memiliki 12 dan 8 transmembran -heliks, dikode oleh DNA
mitokondria oleh semua organisme eukariot. Pemindahan yang terlihat berhasil
dari subunit I ke mitokondria adalah bahwa daerah hidrofobik merupakan alat
yang mencegah subunit I dab sitokrom b mengalami transfer dari genom
mitokondria selama evolusi. Pembentukan sistem ekspresi subunit I tidak
menggunakan metode pembentukan hibrid seperti digunakan disini karena

percobaan untuk pengangkutan apositokrom b (yang lebih kecil dari subunit I) ke
mitokondria tidaklah berhasil.
4.2 Struktur Sinar-X dari Jalur H
Struktur sinar-X dari sitokrom c oksidase bentuk teroksidasi dan tereduksi
penuh pada resolusi 1.8 dan 1.9 Å memperlihatkan bahwa perubahan
konformasional yang besar dari Subunit I Asp-51 terjadi di dekat permukaan sisi
intermembran (Gambar 2A).
Gambar 2A Perubahan konformasional redoks berpasangan dalam Asp-51. penggambaran Spektroskopi jaringan ikatan hidrogen dalam bentuk teroksidsi dan tereduksi penuh (struktur biru)pada resolui 1.8 dan 1.9 Å, dilihat dari sisi intermembran. Dua histidin terikat ke Fea (besi heme a), tidak ditunjukkan.

Perubahan konformosional ini meliputi penyusunan ulang interaksi ikatan
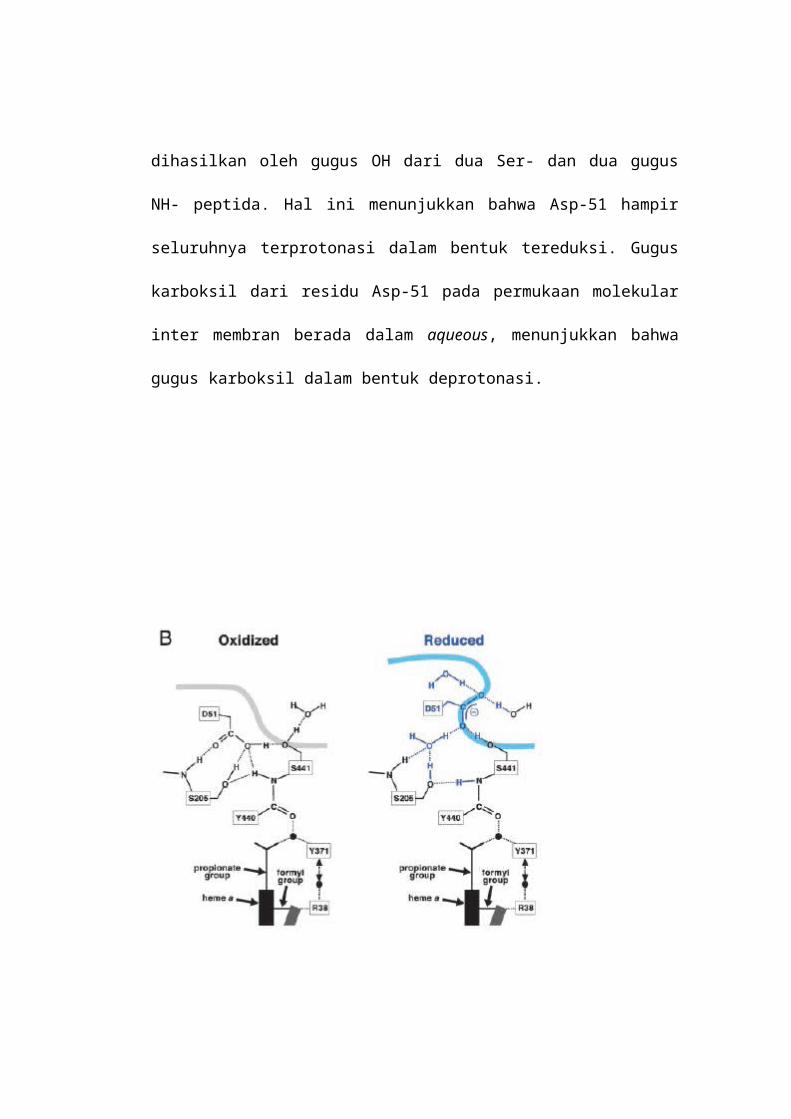
hidrogen (Gambar 2B). Harga pKa dari gugus karbonil dipengaruhi oleh
lingkungannya. Sebagai contoh, pKa asam asetat adalah 4.8 dalam air dan 9.5
dalam methanol. Oleh karenanya lingkungan polar yang non aqueous dari Asp-51
dalam bentuk teroksidasi dihasilkan oleh gugus OH dari dua Ser- dan dua gugus
NH- peptida. Hal ini menunjukkan bahwa Asp-51 hampir seluruhnya terprotonasi
dalam bentuk tereduksi. Gugus karboksil dari residu Asp-51 pada permukaan
molekular inter membran berada dalam aqueous, menunjukkan bahwa gugus
karboksil dalam bentuk deprotonasi.

Gambar 2B.Struktur Ikatan Hidrogen Asp-51 pada keadaan Teroksidasi (kiri) dan keadaan Tereduksi (kanan). Garis tebal menunjukkan permukaan molecular dimana molekul air berada pada ruang intermembran dapat diakses.Perubahan konformasi diinduksi oleh reduksi dari enzim yang diperlihatkan dengan struktur berwarna biru disebelah kanan. Bola berwarna biru (A) dan hitam (B) menggambarkan molekul air. Garis putus-putus menunjukkan ikatan hidrogen. Panah dua menunjukkan kemungkinan pergerakan molekul air dari Arg-38 ke Tyr-71.
Gugus karbonil dari ikatan peptida antara Tyr-440 dan Ser-441
dihubungkan dengan Arg-38 oleh jaringan ikatan hydrogen yang terdiri dari Tyr-
371 dan molekul penarik air kedua (Fig. 2). Molewkul air yang terikat dengan
ikaan hidrohen ke Arg- 38 berlokasi sekitar 4 Å dari Tyr-371. Jarak ini terlalu
jauh untuk membentuk ikatan hydrogen. Molekul air ini kemudian dapat
mendekati Tyr-371 untuk membentuk ikatan hydrogen setelah berpindah dari
Arg-38, yang diindikasikan oleh tanda panah bertitik. Air yang terikat antara tyr-

371 dan gugus karbonil peptida juga terhubung dengan hydrogen ke gugus
propionat dari heme a.
Transfer proton dimungkinkan terjadi melalui ikatan peptida. Ketika
proton ditambahkan ke gugus kmarbonil peptida, akan terbentuk asam imidat [-
C(OH)=N+H-]. Jika gugus penerima proton berlokasi dekat bagian =N+H-, proron
akan diambil untuk membentuk enol dari peptida [-C(OH)=N-]. Stabilitas yang
lebih besar ada pada bentuk keto (-CO-NH-) dibandingkan bentuk enol [-
C(OH)=N-]. Perubahan konformasi dari bentuk ketoenol mengindikasikan
pembalikan ke bentuk keto. Peptida tidak memberikan arah yang karakteristik
transfer protonmelalui peptida yang menghalangi transfer proton dari sisi
intermembran.
Asp-51 dalam bentuk teroksidasi dihubungkan dengan ikatan hydrogen ke
Ser-441 pada permukaanmolekul. Pada tempat itu bentuk reduksi dari jaringan
ikatan hydrogen termasuk air yang terikat antara Asp-51 dan Ser-205
menghubungkan Asp-51 dengan jaringan ikatan hydrogen terbentang ke Arg-38
(Fig 2B). Kemudian Asp-51 dapat dilalui melalui ikatan hydrogen ke kedua sisi
molekul bentuk oksidasi. Transfer proton kebalikan dihalangi oleh ikatan peptida.
Disamping itu struktur sebelumnya pada resolusi 2.3/2.35Å menunjukkan bahwa
Asp-51 dalam bentuk teroksidasi dihubungkan ke ruang matriks oleh jaringan
ikatan hydrogen dan terkubur di dalam permukaan inter membran. Sementara itu
bentuk tereduksi enzim, terdesosiasi dari jaringan ikatan hydrogen dan terpapar ke
ruang intermembran. Model terbaru ini menunjukan bahwa proses pompa proton
didorong oleh perubahan pKadari Asp-51 dan transfer proton tak berarah melalui
ikatan peptida berbeda halnya dengan usulan yang dijelaskan sebelumnya.
Besi heme a yang terikat ke enam nitrogen, (dua dari histidin dan empat
dari forfirin) keduanya dalam bentuk teroksidasi. Dalam bentuk tereduksi , dua
muatan positif Fe2+ dinetralkan oleh dua muatan negatif forfirin. Dalam bentuk
teroksidasi, Fe3+ mempunyai satu muatan positif yang belum ternetralkan.
Relokasi muatan positif ini melalui system electron -forfirin dapat menyebabkan
deprotonasi gugus bersifat asamyang berlokasi dekat heme a. struktur terbaru
Sinar-X menunjukkan bahwa struktur heme a gugus formil adalah coplanar
dengan cincin forfirin dan dapat berkonjugasi dengan system electron -forfirin
dalam kedua bentuk oksidasi. Hal ini konsisten denganbuktin resonansi Raman
yaitu pengaruh bentuk oksidasi besi pada karbonil formil ditunjukkan oleh
pergeseran vibrasional tarikan C-O dari 1610-1650 cm-1dari oksidasi heme a.
Karenanya perubahan dalam bentuk oksidasi heme a dapat diharapkan
mempunyai pengaruh elektrostatik yang signifikan terhadap Arg-38 yang terikat
dengan ikatan hydrogen ke gugus formil, walaupun tidak teradi perubahan
kopnformasional yang disebabkan oleh system Arg-38-formil (Fig 2A). resolusi
terbaru dari struktur Sinar-X tidak cukup untuk meneliti perubahan
konformasional yang diinduksi oleh pengaruh elektrostatik Fea. gugus propionat
heme a yang terhubung dengan ikatan hydrogen ke air yang terperangkap (Fig 2),
dapat memicu transfer proton melewati jaringan ikatan hydrogen, juga oleh
pengaruh elektrostatik dari oksidasi Fea.

Arg-38 yang terhubung dengan ikatan hydrogen ke gugus formil heme a,
dimana molekul air dalam ruang matriks dapat masukmelalui saluran air. Saluran
di dekat ujung formil, secara skematik diperlihatkan dengan garis putus dalam
gambar 3A. ruang yang dapat dilalui air pada saluran, ditentukan oleh perhitungan
permukaan molekul, menunjukkan bahwa saluran memilki 4 lubang, yang
masing-masing cukup besar untuk mengandungsatu sampai tiga molekul air (fg
3B dan ruang bertitik merah dalam Fig 3A. gugus OH dari gugus hidrofarnesiletil
dari heme a terhubung dengan ikatan hydrogen ke Ser-382 dekat saluran airdalam
bentuk oksidasi ( Fig. 3A, struktur merah). Selama reduksi enzim, ikatan
hydrogen OH…Ser-382 diputus , memungkinakn gugus OH dari gugus
hidroksifarnesiletil berotasi 120 dengan pergerakan dari rantai hidrokarbon dekat
gugus OH [-CH(OH)-CH2-CH2-CH=C(CH3)-] dan memungkinkan rotasi 110 dari
gugus OH Ser-382 (Fig.3A, struktur biru), dipasangkan dengan perubahan
konformasional dalam pergantian heliks-X yang termasuk Ser-382, Leu-381 dan
Val-380 (Fig.3A, struktur asam amino merah dan biru dlam Heliks-X). perubahan
kondformasiional ini menyebabkan terbentuknya lubang baru antara gugus
hidroksi farnesiletil dan Heliks-X (Fig 3A, ruang bertitik biru dengan ruang yang
tidak ertitik merah dan Fig. 3B, oval biru). Perubahan konformasional diatur
olehbentuk oksidasi heme a karena ikatan hydrogen OH…Ser-382 yang berlokasi
dekat system electron- forfirin heme a. posisi dan ukuran empat lubang yang
teramati dalam bentuk teroksidasi tidak dipengaruhi secara signifikan selama
reduksi (Fig.3 ruang bertitik biru dan merah). Perubahan konformasional redoks

yang dipasngkan menunjukkan perubahan kapasitas air dalam saluran, yang
sepertinya memberikan kontribusi terhadap kumpulan proton efektif dari ruang
matriks ke Arg-38.
Gambar 3A Struktur Sinar-X dari saluran air jalur H. perubahan konformasional redoks berpasangan dari saluran air. Bagian atas saluran ditunjukkan. Merah dan biru menunjukkan struktur dalam bentuk teroksidasi dan tereduksi. Permukaan bertitik merah dan biru menunjukkan lubang yang dideteksi dalam bentuk teroksidasi dan tereduksi. Garis putus-putus menunjukkan jalur pengubung air ke lubang. Garis bertitik menunjukkan ikatan hidrogen. Bola kecil menunjukkan posisi molekul air yang terikat. His-61 terikat ke Fea dari sisi yang berlawanan dengan heme tidak ditunjukkan.

Gambar 3B Representasi skematik dari perubahan konformasional redoks berpasangan dalam saluran air. Daerah yang diberi kotak disebut daerah A. Bola hitam dan biru menunjukkan molekul air yang terikat. Struktur yang teramati hanya pada bentuk tereduksi yaitu oleh warna biru.
4.3 Analisis FTIR Dari Perubahan Konformasi Reaksi Redoks Yang Berpasangan
FTIR digunakan untuk mengidentifikasi sisi logam redoks-aktif yang
mengatur konformasi Asp-51. Berbeda dengan spektrum FTIR dari keadaan
teroksidasi, enzim keadaan tereduksi pada H2O, Asp-51 memberikan puncak pada
1,738 cm-1 dan 1,585 cm-1, yang memberikan bukti adanya uluran COO model

dari COOH dan COO. Perbedaan spektra pada redoks dengan adanya sianida dan
CO yang juga memberikan pita yang identik pada 1,738 cm-1 dan 1,585 cm-1.
sianida menstabilkan heme a3 pada keadaan teroksidasi. Jadi enzim tereduksi
dengan adanya sianida yang dimiliki oleh heme a3, CuA dan CuB pada keadaan
tereduksi dan heme a3 pada keadaan sianida terikat yang berikatan. Sehingga
spektrum yang berbeda antara enzim teroksidasi pada sianida berikatan dan enzim
tereduksi yang mengikat sianida menghasilkan penjumlahan perbedaan spektra
redoks yang diinduksi oleh heme a, CuA dan CuB. Dengan kata lain, Co
menstabilkan CuB dan heme a3 pada keadaan tereduksi memberikan spektra redoks
yang berbeda yang diinduksi oleh heme a, CuA. Karena itu hasil ini
mengindikasikan bahwa keadaan terprotonasi dari Asp-51 dikendalikan oleh heme
a dan CuA.
Pada titrasi reduksi enzim sapi ini terdapat sianida, dengan penurunan
intensitas dua pita adalah proporsional pada ekuivalen elektron yang ditambahkan.
Tiga elektron ekuivalen dibutuhkan untuk memenuhi eliminasi dari dua pita pada
spektrum yang berbeda. Hasil ini mengindikasikan bahwa spektra infra
merahberubah karena single elektron ekuivalen.
Titrasi redukrif enzim sianida terikat, dimonitor menggunakan keadaan
oksidasi dari tempat logam redoks aktif, heme a, CuA dan CuB. Sehingga jika dua
atau tiga elekron dibutuhkan untuk perubahan spektra infra merah, maja perubahn
spektra tidak akan terlihat dibawah satu atau dua elektron secara ekuivalen.
Karena itu COOH dari Asp-51 dipisahkan selama reduksi hanya satu tempat
logam, selain heme a atau CuA.
Elektron equivalen dari sitokrom c ditransfer dari CuA ke heme a kemudian
dari heme a ke tempat reduksi O2.jadi heme a dapat memberikan tanda sebagai
tempat kontrol logam pada konformasi Asp-51.
4.4 Mekanisme Proton-Pumping
Mekanisme keseluruhan berdasarkan hasil kesimpulan adalah : ketika
heme a berada pada keadaan teroksidasi, Arg-38 terprotonasi pertama walaupun
dibawah muatan positif dari heme a, karena molekul air pada ruang matriks
accessible ke Arg-38 melalui terowongan air.Asp-51 terkubur didalam protein dan
terprotonasi. Selama heme a mengalami reduksi, dimana kehilangan muatan
positif di heme a, Asp-51 keluar ke ruang intermembran dan kapasitas terowongan
air meningkat. Karena itu molekul air akan bergerak ke ruang matriks, ketika
proton pada Asp-51 dilepaskan ke ruang intermembran. Selama heme a
mengalami oksidasi, Asp-51 bergerak ke belakang ke interior protein dan
memperlihatkan muatan positif pada heme a menurunkan afinitas dari formil
oksigen untuk membagi proton dengan Arg-38. penurunan aktivitas promosi
proton transfer dari Arg-38 ke Asp-51. gugus propionat ikatan hidrogen pada air
juga mempercepat proton transfer sepanjang jaringan ikatan hidrogen.

Gambar 4. Mekanisme Proton pumping yang diusulkan. Besi, porfirin dan gugus samping formil dari heme a diperlihatkandengan Fea, Pr dan OCHO.COOH pada Pr menunjukkan satu gugus propionat dari heme a.
Tanda kurung ([ ]1_and [ ]0) menunjukkan jumlah muatan dari enam koordinat heme Warna kotak yang lebih hitam menunjukkan keadaan struktur yang stabil dan keadaan intermediet. Panah tebal menunjukan pengaruh elektrostatik dari jumlah muatan positif heme a dan proton transfer selama heme a mengalami oksidasi. Garis putus-putus menunjukkan ikatan hidrogen yang menghubungkan Arg-38 dengan Asp-51,termasuk ikatan peptida yang menghalangi reverse transfer dari sisi intermembran.
Hasil Arg-38 yang terprotonasi mengekstrak proton dari molekul air pada
terowongan air sebelum kapasitas terowongan air menurun menjadi pemaksaan
keluar OH-. Keikutsertaan gugus formil heme a dan molekul air disekitar proton
pumping sebelumnya telah dianalisa menggunakan resonansi raman.

KESIMPULAN
1. Mutasi Asp-51 Asn dari enzim hati memperlihatkan fungsi pompa proton
tanpa mempengaruhi aktivitas reduksi dioksigen
2. Struktur Sinar-X (pada resolusi 1.8/1.9 Å dalam bentuk teroksidasi dan reduksi
penuh) menunjukkan bahwa muatan positif total yang dibentuk selama
oksidasi mendorong transport proton aktif dari ruang mitokondria ke Asp-51
melewati enzim lewat saluran air dan jaringan ikatan hidrogen, yang berlokasi
di tandem.
3. Hasil infrared mengindikasikan bahwa konformasi Asp-51 dikontrol hanya oleh
bentuk oksidasi heme spin rendah. Hasil ini mengindikasikan bahwa heme
spin rendah mendorong proses pompa proton.

DAFTAR PUSTAKA
Lehninger, L. 1991.Dasar-dasar Biokimia.Alih bahasa : Dr Maggy Thenawidjaja. Edisi kedua.Penerbit Erlangga. Jakarta.
Michel H. 1998. The Mechanism of proton pumping by cytochrome c oxidase.PNAS 95, 12819-12824.
Tsukihara, T., Shimokata, K., Katayama, Y., Shimada H, Muromoto K, Aoyama H, Mochizuki M, Shinzawa K, Yamashita E, Yao M, Ishimura Y, Yoshikawa S. 2003.The Low Spin Heme of Cytochrome c oxidase as the driving element of the proton pumping process.PNAS 100.15304-15309.
Yoshikawa, S., Shinzawa-Itoh, K., Nakashima, R., Yaono, R., Yamashita, E., Inoue, N., Yao, M., Fei, M. J., Libeu, C. P., Mizushima, T., et al. (1998)
Science 280, 1723–1729.


















