
Jurnal Cumi Cumi
8
Squid gelatin hydrolysates with antihypertensive, anticancer and antioxidant activity A. Alemán a , E. Pérez-Santín a , S. Bordenave-Juchereau b , I. Arnaudin b , M.C. Gómez-Guillén a, ⁎, P. Montero a a Instituto de Ciencia y Tecnología de Alimentos y Nutrición (ICTAN, CSIC) (formerly Instituto del Frío), C/ José Antonio Novais, 10, 28040, Madrid, Spain b Laboratoire Littoral ENvironnement et Societés (LIENSS), Université de La Rochelle, CNR UMR 6250 UFR Sciences, Avenue M. Crepeau 17042, La Rochelle, France a b s t r a c t a r t i c l e i n f o Article history: Received 22 January 2011 Accepted 6 March 2011 Keywords: Bioactive peptides Gelatin hydrolysates Antioxidant ACE inhibition Anticancer Gela tin obtained fro m gi ant squ id (Dosidicus gigas) inner and out er tuni cs was hydr olyz ed by seve n comme rcia l proteases (Protam ex, Trypsin, Neutrase, Savinase, NS37005, Esperase and Alcalase ) to produce bioactive hydrolysates. The Alcalase hydrolysate was the most potent angiotensin-converting enzyme (ACE) inhibitor (IC 50 =0.34 mg/mL ) while theEsperas e hydr oly sateshowed thehighestcytoto xiceffecton canc er cells, wit h IC 50 valu es of 0.13and 0.10 mg/m L forMCF-7 (human breastcarcino ma)and U87(glioma ) celllines,respect ivel y. The radical scavenging capacity of gelatin increased approximately 3-fold for Protamex, Neutrase and NS37005 hydrolysates and between 7 and 10-fold for Trypsin, Savinase, Esperase and Alcalase hydrolysates. Trypsin, Savinase, Esperase and Alcalase hydrolysates had a metal chelating capacity above 80% whereas Protamex, Neut rase and NS3700 5 hydro lysa tes regi ster ed less than 25%. The antio xida nt activi ty measur ed by FRAP (fer ric ion reducing power) was largely unaffected by the enzyme used, increasing approximately 2-fold for all hydrolysates. The most active hydrolysates (Alcalase and Esperase) were comprised mostly of peptides with mole cula r weights rang ing from500 to 1400 Da, howe ver, a clea r relatio nshi p between bioa ctiv e properties and molecular weight distribution of all the hydrolysates was not fully established. © 2011 Elsevier Ltd. All rights reserved. 1. Introduction Hypertension is a multifactorial process and the main cause of illness in industr ialized countries. The angioten sin-I converting enzyme (ACE) plays an important role in the regulation of blood pressure and hypertension because it catalyses the conversion of inactive angiotensin-I into angiotensin-II, a potent vasoconstrictor (Goodf rien d, Elliott, & Catt, 1996) and inac tivat es brad ykin in, a potent vasodilator (Witherow, Helmy, Webb, Fox, & Newby, 2001 ). Consequently synthe tic ACE inhibitors such as Captopril and Enalapril are often used to treathypertension and other card io-r elate d dise ases . However as these synthetic ACE inhibitors can cause adverse side effects including cough, taste disturbance, rashes and angioedema (Messerli, 1999), there is a growing interest in natural inhibitors. Fish prote in hydro lysa tes produ ced using commerci ally available proteases have been shown to possess combined ACE-inhibitory and antioxidant capacities ( Je, Lee, Lee, & Ahn , 2009; Samaran ayaka, Kitts, & Li-Chan, 2010), and it has been postulated that these two activities could contribute to their antihypertensive effect in vivo (Hou, Chen, & Lin, 2003). On th e other ha nd, ca nc er is on e of the la rg est si nglecaus es of deat h in both men and women ( Kaufman & Earnshaw, 2000). Frequently, resistance to anticancer drugs has been observed ( Lind et al., 2001). The ref ore , theresea rchanddevelo pme nt of moreeff ect iveand les s toxic drugs by the pharma ceuti cal indust ry has becomenecessar y, prompti ng a growing interest in the identi cation and characterization of natural antitumor agents. The elimination of cancer in the early stages is an int egr al part of che mop rev ent ion , and measur ing the cyt otox ic properties of a given compound against cancer cells provides useful insight into its chemoprotective potential. Antioxidants have also been proposed as potentially suitable candidates for the prevention and treatment of diseases associated with active oxygen species, especially applicable to cancer diseases ( Leng, Liu, & Chen, 2005 ). Fish or squid skin gelatins have been reported to give rise to biologically active peptides with high ACE-inhibitor y and antioxidant act ivity, the lat ter due to its radical scavengin g cap acity, metal chelating effects and reducing power or lipid peroxidation inhibition (Zhao, Li, Liu, Dong, Zhao, & Zeng, 2007; Kim, Byun, Park, & Shahidi, 2001; Mendis, Rajapakse, Byun, & Kim, 2005; Mendis, Rajapakse, & Kim, 2005; Lin & Li, 2006; Giménez, Alemán, Montero, & Gómez- Guillén, 2009; Alemán, Giménez, Montero, & Gómez-Guillén, 2011 ). However, no reference has been made in previous reports of any anti proli fera tive /cyt otoxic effect of gela tin derived pept ides or hydrolysates on cancer cells. The biological properties of peptides are to a large extent in uenc ed by their molec ular weight and confo rmat ional structur e, which are in turn , greatly affected by processing conditions . In contrast, the amino acid composition of the hydrolysates resulting from the hydrolysis with different types of enzymes is very similar to that of the parent gelatins, as reported by Kim, Kim, Byun, Nam, Joo, and Shahidi (2001) with Alaska pollack Food Research International 44 (2011) 1044 –1051 ⁎ Corresponding author. Tel.: +34 91 5492300; fax: +34 91 5493627. E-mail address: [email protected] (M.C. Gómez-Guillén). 0963-9969/$ – see front matter © 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.foodres.2011.03.010 Contents lists available at ScienceDirect Food Research International j ournal h o me p a g e : www. els ev i e r. c o m/lo c a t e / food r e s
-
Upload
putri-rahmawati -
Category
Documents
-
view
233 -
download
0
Transcript of Jurnal Cumi Cumi

8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 18
Squid gelatin hydrolysates with antihypertensive anticancer and antioxidant activity
A Alemaacuten a E Peacuterez-Santiacuten a S Bordenave-Juchereau b I Arnaudin b MC Goacutemez-Guilleacuten a P Montero a
a Instituto de Ciencia y Tecnologiacutea de Alimentos y Nutricioacuten (ICTAN CSIC) (formerly Instituto del Friacuteo) C Joseacute Antonio Novais 10 28040 Madrid Spainb Laboratoire Littoral ENvironnement et Societeacutes (LIENSS) Universiteacute de La Rochelle CNR UMR 6250 UFR Sciences Avenue M Crepeau 17042 La Rochelle France
a b s t r a c ta r t i c l e i n f o
Article history
Received 22 January 2011Accepted 6 March 2011
Keywords
Bioactive peptidesGelatin hydrolysatesAntioxidantACE inhibitionAnticancer
Gelatin obtained from giant squid (Dosidicus gigas) inner and outer tunics was hydrolyzed by seven commercialproteases (Protamex Trypsin Neutrase Savinase NS37005 Esperase and Alcalase) to produce bioactivehydrolysates The Alcalase hydrolysate was the most potent angiotensin-converting enzyme (ACE) inhibitor(IC50=034 mgmL) while theEsperase hydrolysateshowed thehighestcytotoxiceffecton cancer cells with IC50values of 013and 010 mgmL forMCF-7 (human breast carcinoma)and U87(glioma) celllinesrespectively Theradical scavenging capacity of gelatin increased approximately 3-fold for Protamex Neutrase and NS37005hydrolysates and between 7 and 10-fold for Trypsin Savinase Esperase and Alcalase hydrolysates TrypsinSavinase Esperase and Alcalase hydrolysates had a metal chelating capacity above 80 whereas ProtamexNeutrase and NS37005 hydrolysates registered less than 25 The antioxidant activity measured by FRAP (ferricion reducing power) was largely unaffected by the enzyme used increasing approximately 2-fold for allhydrolysates The most active hydrolysates (Alcalase and Esperase) were comprised mostly of peptides withmolecular weights ranging from500 to 1400 Da however a clear relationship between bioactive properties andmolecular weight distribution of all the hydrolysates was not fully established
copy 2011 Elsevier Ltd All rights reserved
1 Introduction
Hypertension is a multifactorial process and the main cause of illness in industrialized countries The angiotensin-I convertingenzyme (ACE) plays an important role in the regulation of bloodpressure and hypertension because it catalyses the conversion of inactive angiotensin-I into angiotensin-II a potent vasoconstrictor(Goodfriend Elliott amp Catt 1996) and inactivates bradykinin apotent vasodilator (Witherow Helmy Webb Fox amp Newby 2001)Consequently synthetic ACE inhibitors such as Captopril and Enalaprilare oftenused to treathypertension and othercardio-related diseasesHowever as these synthetic ACE inhibitors can cause adverse sideeffects including cough taste disturbance rashes and angioedema(Messerli 1999) there is a growing interest in natural inhibitorsFish protein hydrolysates produced using commercially availableproteases have been shown to possess combined ACE-inhibitory andantioxidant capacities ( Je Lee Lee amp Ahn 2009 Samaranayaka Kittsamp Li-Chan 2010) and it has been postulated that these two activitiescould contribute to their antihypertensive effect in vivo (Hou Chen ampLin 2003)
On the other hand cancer is one of the largest single causes of deathin both men and women (Kaufman amp Earnshaw 2000) Frequentlyresistance to anticancer drugs has been observed (Lind et al 2001)
Therefore theresearchand developmentof more effectiveand less toxicdrugsby the pharmaceutical industry has becomenecessary promptinga growing interest in the identi1047297cation and characterization of naturalantitumor agents The elimination of cancer in the early stages is anintegral part of chemoprevention and measuring the cytotoxicproperties of a given compound against cancer cells provides usefulinsight into its chemoprotective potential Antioxidants have also beenproposed as potentially suitable candidates for the prevention andtreatment of diseases associated with active oxygen species especiallyapplicable to cancer diseases (Leng Liu amp Chen 2005)
Fish or squid skin gelatins have been reported to give rise tobiologically active peptides with high ACE-inhibitory and antioxidantactivity the latter due to its radical scavenging capacity metalchelating effects and reducing power or lipid peroxidation inhibition(Zhao Li Liu Dong Zhao amp Zeng 2007 Kim Byun Park amp Shahidi2001 Mendis Rajapakse Byun amp Kim 2005 Mendis Rajapakse ampKim 2005 Lin amp Li 2006 Gimeacutenez Alemaacuten Montero amp Goacutemez-Guilleacuten 2009 Alemaacuten Gimeacutenez Montero amp Goacutemez-Guilleacuten 2011)However no reference has been made in previous reports of anyantiproliferativecytotoxic effect of gelatin derived peptides orhydrolysates on cancer cells The biological properties of peptidesare to a large extent in1047298uenced by their molecular weight andconformational structure which are in turn greatly affected byprocessing conditions In contrast the amino acid composition of thehydrolysates resulting from the hydrolysis with different types of enzymes is very similar to that of the parent gelatins as reportedby Kim Kim Byun Nam Joo and Shahidi (2001) with Alaska pollack
Food Research International 44 (2011) 1044ndash1051
Corresponding author Tel +34 91 5492300 fax +34 91 5493627E-mail address cgomezictancsices (MC Goacutemez-Guilleacuten)
0963-9969$ ndash see front matter copy 2011 Elsevier Ltd All rights reserved
doi101016jfoodres201103010
Contents lists available at ScienceDirect
Food Research International
j o u r n a l h o m e p a g e w w w e l s ev i e r c o m l o c a t e f o o d r e s
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 28
skin gelatin or Goacutemez-Guilleacuten et al (2010) with tuna and giant squidskin gelatins
A number of commercial proteases have been used for theproduction of antioxidant gelatin hydrolysates including trypsinchymotrypsin pepsin Properase E Pronase collagenase bromelainand papain (Kim Kim et al 2001 Mendis Rajapakse Byun et al2005 Mendis Rajapakse amp Kim 2005 Linamp Li2006YangHo Chu ampChow 2008) Alcalase which is a commercial protease from a
microbial source (Bacillus licheniformis) has been shown to be one of the most ef 1047297cient in the hydrolysis of 1047297sh protein (Gueacuterard DufosseacuteDe La Broise amp Binet 2001) This enzyme preparation has also beenreported to produce high levels of hydrolysis in skin gelatin fromAlaska pollack squid Todarodes paci 1047297cus or giant squid Dosidicus gigas
(Kim Byun et al 2001 Nam You amp Kim 2008 Gimeacutenez et al 2009)In a recent work a peptide sequence Gly-Pro-X-Gly-X-X-Gly-Phe-X-Gly-Pro-X-Gly-X-Ser with the X positions occupied by either Hyp orLeu and a molecular mass of 1411 Da together with a compound witha molecular mass of 952 Da associated with the carbohydrate fucosewere identi1047297ed as being responsible for most of the high anti-radicalandACE-inhibitoryactivityin an Alcalasehydrolysate fromgiant squidgelatin (Alemaacuten Gimeacutenez Peacuterez-Santiacuten Goacutemez-Guilleacuten amp Montero2011) Further work with giant squid gelatin revealed that Alcalasealso produced hydrolysates with much higher radical scavengingand ferric reducing capacities than those of other single enzymessuch as collagenase trypsin or pepsin (Alemaacuten Gimeacutenez Monteroet al 2011)
The aim of this study was to explore the potential health bene1047297tswhich could be derived from giant squid tunic gelatin by investigatingthe ACE-inhibitory anticancer and antioxidant properties (radicalscavenging metal chelating and reducing power) of hydrolysateshaving different molecular weight distributions obtained by usingseveral commercial proteases
2 Materials and methods
21 Materials
Gelatin from inner and outer giant squid (D gigas) tunics wasextracted using the method described by Gimeacutenez et al (2009)Commercial proteases (Protamex Trypsin Neutrase 08 L Savinase16 L NS37005 Esperase 08 L and Alcalase 24 L) were kindly suppliedby Novozymes (Bagsvaerd Denmark) NaOH HCl NaCl glycerol andpotassium persulfate were from Panreac (Barcelona Spain) All otherreagents were from Sigma-Aldrich Inc (St Louis MO USA)
22 Preparation of gelatin hydrolysates
The squid gelatin (25 wv) was dissolved in water and subjectedto enzymatic hydrolysis for 3 h under optimal temperature and pHconditions The optimal condition for each protease was as followsProtamex (pH 65 60 degC) Trypsin (pH 8 55 degC) Neutrase 08 L (pH
65 55 degC) Savinase 16 L (pH 95 55 degC) NS37005 (pH 7 55 degC)Esperase 08 L (pH 85 60 degC) and Alcalase 24 L (pH 8 50 degC) with anenzymendashsubstrate ratio of 120 (ww) The pH of the reaction waskept constant by adding a 1 N NaOH solution to the reaction mediumusing a pH-stat (TIM 856 Radiometer Analytical Villeurbanne CedexFrance) The enzymes were inactivated by heating at 90 degC for 10 minand the samples were centrifuged at 3000 g for 15 min The super-natants comprised the hydrolysates and were lyophilized and storedat minus80 degC for further assays
23 Molecular weight distribution
The gelatin hydrolysates were analyzed by Tricine SDS-PAGEaccording to Schagger and Von Jagow (1987) using a 5 stacking
gel and a 165 resolving gel The hydrolysates were dissolved at
a concentration of 10 mgmL in the loading buffer (50 mM TrisndashHCl4 SDS 12 glycerol 2 mercaptoethanol and 001 SERVA Blue G)heat-denatured at 90 degC for 10 min and run in a Mini Protean II unit(Bio-Rad Laboratories Hercules CA USA) Protein bands were stainedwith Coomassie Brilliant Blue R250 The approximate molecular weight(MW) of the hydrolysates was determined using a low molecularweight marker kit (Amersham Pharmacia Biotech Uppsala Sweden)consisting of triosephosphate isomerase (266 kDa) myoglobin
(169 kDa) α-lactalbumin (144 kDa) aprotinin (65 kDa) oxidizedinsulin β chain (35 kDa) and bacitracin (14 kDa)Hydrolysate molecular weight distribution was also evaluated
by size-exclusion HPLC (model SPE-MA10AVP Shimadzu Kyoto Japan) on a superdex peptide PC 3230 column (GE Healthcare Bio-Sciences Barcelona Spain) with a fractionation range between 7000and 100 Da The injection volume was 10 μ L and the 1047298ow rate was01 mLmin using Milli-Q water in the mobile phase Vitamin B12(1355 Da) and hippuryl-L -histidyl-L -leucine (HHL 502 Da) bothfrom Sigma-Aldrich Inc (St Louis MO USA) were used as molecularweight standards
24 ACE-inhibition
Reversed-phase high performance liquid chromatography (RP-HPLC) was used to determine ACE inhibition applying the methodused by Wu Aluko and Muir (2002) with slight modi1047297cations Thetotal reaction volume was 225 μ L made up of 50 μ L of 5 mM HHL160 μ L of 25 mUmL of ACE and 20 μ L of sample at six differentconcentrations (from 01 to 15 mgmL 1047297nal concentration) preparedwith 100 mM potassium-phosphate buffer containing 300 mM NaClpH 83 The mixture was incubated at 37 degC for 120 min and thereaction was quenched by adding 100 μ L of 01 M HCl The releasedhippuric acid (HA) was quanti1047297ed by RP-HPLC (model SPE-MA10AVPShimadzu Kyoto Japan) on a C18 column (Tracel excel 120 ODSA5 μ m 25times046 Teknokroma Barcelona Spain) The injected volumewas 50 μ L and the 1047298ow rate was 08 mLmin using an acetonitrilegradient of 20 to 60 in 01 tri1047298uoroacetic acid (TFA) for 26 minThe HA and HHL were monitored at 228 nm and the retention times
were 830 and 1570 min respectively All determinations werecarried out at least in triplicate The IC50 value of ACE inhibitoryactivity was de1047297ned as the concentration of hydrolysate (mgmL)required to reduce the HA peak by 50
25 Cytotoxic and antiproliferative activities
The study of cytotoxic and antiproliferative activities of squidgelatin hydrolysates was performed according to the methodologyproposed by Picot et al (2006) MCF-7 human breast carcinomaand U87 glioma cell lines were grown at 37 degC in a 5 CO 2 95 airhumidi1047297ed atmosphere in DMEM-Hams F12 medium (11 vvGibco) The medium was supplemented with 5 heat inactivatedFetal Calf Serum (FCS Dutscher) to which penicillin (100 UmL) and
streptomycin (100 μ gmL) had been added (Pan biotech GmbH) Thecells grown in 1047298asks were trypsinized centrifuged at 800 rpm for5 min and dissolved in fresh culture medium at 104 cells90 μ LSubsequently a 90 μ L volume of suspension cells (10000 cells) wasadded to each well of a 96-well microplate and incubated at 37 degC for24 h In order to perform cytotoxicity screening of the hydrolysateshydrolysate stock solution at a concentration of 10 mgmL wasprepared in PBS 01 M (pH 74) and diluted 10-fold in cell culturemedium containing the cells The microplate was then incubated at37 degC for24 48 and72 h changing the culture mediumevery 24 h andadding the hydrolysate at a 1047297nal concentration of 1 mgmL At the endof every incubation period 15 μ L of 5 mgmL tetrazolium salt (MTT)solution was added to each well and the plate was incubated for3 h To stop succinate-tetrazolium reductase activity and solubilize
formazan crystals 200 μ L of dimethyl sulfoxide (DMSO) was then
1045 A A lemaacuten e t al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 38
added to each well and kept at 37 degC for 1 h Absorbance was read ona plate reader (VERSAmax Molecular Devices Saint Gregoire France)at 570 nm The data were analyzed to calculate the percentageof viability inhibition induced by the presence of hydrolysate incell culture medium determined by the equation Viability inhibition()=100minus(AHtimes100AC) where AH and AC are the absorbenciesmeasured for cell viability in culture medium containing onlyhydrolysate or PBS respectively
Following the same procedure the effect of addition of hydro-lysates at the same time as the cancer cells at the beginning of theexperiment was also evaluated For this evaluation the initialconditions of the culture medium and the added hydrolysates weremaintained for 72 h
The viability inhibition of the Esperase and Alcalase hydrolysateswas also measured using six different concentrations ranging from005 to 15 mgmL in order to determine the IC50 All experimentswere performed at least in triplicate
26 Antioxidant activities
Different assays (ABTS chelating ability and FRAP) were per-formed to study the antioxidant properties of the squid gelatin andthe corresponding hydrolysates at a concentration of 10 mgmL TheABTSradical [22prime-azino-bis-(3-ethylbenzothiazoline-6-sulfonic acid)]scavenging assay was carried out according to the method describedby Re Pellegrini Proteggente Pannala Yang and Rice-Evans (1999)The stock solution of ABTS radical consisted of 7 mM ABTS inpotassium persulfate 245 mM kept in the dark at room temperaturefor 16 h An aliquot of stock solution was diluted with Milli-Q waterin order to give the working solution of ABTS radical an absorbanceat 734 nm of 070plusmn002 A 20 μ L aliquot sample was mixed with980 μ L of ABTS reagent and then left to stand in the dark at 30 degCfor 10 min Absorbance values were read at 734 nm using a UV-1601spectrophotometer (Model CPS-240 Shimadzu Japan) Resultswere expressed as mg Vitamin C Equivalent Antioxidant Capacity(VCEAC)g of sample (gelatin or hydrolysate) based on a standardcurve of vitamin C
The Fe2+ chelating activity of the hydrolysates at a concentrationof 10 mgmL was measured by the method introduced by ChungChang Chao Lin andChou (2002) Brie1047298y a test sample of 800 μ Lwasmixed with 10 μ L of 2 mM FeCl2 and 20 μ L of 5 mM ferrozine Themixture was vortexed and kept at room temperature for 10 min priorto measuring the absorbance at 562 nm using a UV-1601 spectro-photometer (Model CPS-240 Shimadzu Japan) Chelating abilityexpressed as a percentage was calculated as follows Chelatingability ()=[1minus(Abs sample Abs control)]times 100 The control wasa mixture composed of 800 μ L of Milli-Q water 10 μ L of 2 mM FeCl2and 20 μ L of 5 mM ferrozine
FRAP a measure of the reducing power of samples was performedaccording to the method described by Benzie and Strain (1996) withsome modi1047297cations A 60 μ L of dissolved sample (at a concentration of
10 mgmL)was incubated at 37 degCwith 60μ L of Milli-Q waterand 900 μ L of FRAP reagent containing 10 mM of TPTZ (246-tripyridyl-s-triazine)and 20 mMof FeCl3 Absorbance valueswerereadat595 nmafter 30 minusing a UV-1601 spectrophotometer (Model CPS-240 Shimadzu Japan)Results were expressed as μ mol Fe2+ equivalentsg of sample (gelatin orhydrolysate) based on a standard curve of FeSO4middot7H2O All determina-tions were carried out at least in triplicate
27 Statistical analysis
Statistical tests were performed using the SPSSreg computerprogramme (SPSS Statistical Software Inc Chicago IL USA) Datawere expressed as meanplusmn standard deviation of the mean (n= 3 orn=24) Tukeys test was used to determine the level of signi1047297cance
at ple005
3 Results and discussion
31 Molecular weight distribution
Analysis of the overall molecular weight distribution of thedifferent hydrolysates (Fig 1) was carried out by Tricine SDS-PAGEThe Trypsin and NS37005 hydrolysates showed the highest quantityof peptide bands in the MW range between 26 and 35 kDa An
appreciable amount of protein with MW even higher than 26 kDawas also present in the Trypsin hydrolysates denoting the lowestef 1047297ciency in protein breakdown All the hydrolysates were character-ized by presenting an intense band at MWle14 kDa Howeverno peptide bands of more than 14 kDa could be observed in theAlcalase hydrolysates and hardly any in the Neutrase hydrolysateand Esperase hydrolysate indicating a considerably high degree of proteolysis Hydrolysates composed of a wide range of polydisperseproducts with varying degrees of hydrolysis and mostly small peptidescannot usually be properly detected under analytical electrophoresisconditions (Khantaphant amp Benjakul 2008 Gimeacutenez et al 2009)Consequently in order to know the differences in the molecularweight distribution of the smallest peptides (MWle14 kDa) size-exclusion HPLC (SE-HPLC) was performed in a fractionation rangebetween 7000 and 100 Da (Fig 2) The varying MW pro1047297lesrevealed considerable differences in the degree of protein breakdowndepending on the enzyme used In accordance with the electrophoreticpro1047297le the Alcalase Neutrase and Esperase hydrolysates showedhigher levels of peptides with MW below 1400 Da whereas in Trypsinand NS37005 hydrolysate peptides with MW above 1400 Da werepredominant
32 ACE-inhibitory capacity
The amount of hydrolysate required to inhibit 50 of the ACEactivity (IC50) is shown in Table 1 Unlike the parent squid gelatin allthe hydrolysates presented ACE inhibitory capacity although it wasmarkedly differentdepending on theenzyme used It wasnot possibleto determine the IC50 value in the NS37005 hydrolysate whereas the
Alcalase hydrolysate showed the most potent effect with an IC50
of 034 mgmL followed by the Neutrase hydrolysate (063 mgmL)(ple005) As revealed by SDS-PAGE and SE-HPLC the peptides inthe Alcalase and Neutrase hydrolysates were mostly between ~1400and 500 Da In another study Zhao et al (2007) found the highestACE-inhibitory capacity for a sea cucumber gelatin hydrolysate in thefraction with MWb1 kDa Nevertheless there are other studies where
Fig 1 Electrophoretic pro1047297les of gelatin hydrolysates at a concentration of 10 mgmL (Tricine SDS-PAGE) (m) molecular weight markers (a) Protamex (b) Trypsin
(c) Neutrase (d) Esperase (e) NS37005 (f) Savinase and (g) Alcalase
1046 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 48
very small peptides did not appear to increase the activity of thewhole hydrolysate but rather decreased it (Theodore amp Kristinsson2007)
Bougatef et al (2008) reported IC50 values in the concentrationrange of 12ndash74 mgmL for sardinelle by-product hydrolysatesprepared with different proteases while the IC50 of a marine shrimphydrolysate digested by a crude protease was 097 mgmL (Hai-lunXiu-Lan Cai-Yun Yu-Zhong amp Bai-Cheing 2006) Three peptides Val-Tyr-Ala-Pro Val-Ile-Ile-Phe and Met-Ala-Trp with IC50 values of 61 87and 1632 μ M respectively had been reported from cuttle1047297sh muscleprotein hydrolysates (Balti Nedjar-Arroume Bougatef Guillochon amp
Nasri 2010) Although values of IC50 for ACE-inhibitory capacity of skingelatin whole hydrolysates had been previously reported by Kim Kimet al (2001) and Zhao et al (2007) comparisons with IC50 valuesin thepresent study were not fully possible due to the different methodologyemployed for measuring activity
Proline or aromatic amino acids existing at theC-terminal are suitablefor peptide binding to ACE (Cheung Wang Miguel Emily amp Dvid 1980)Alcalase has broad speci1047297city hydrolyzing most peptide bonds preferen-tially those containing aromatic amino acid residues (Doucet OtterGauthier amp Foegeding 2003) suggesting that this enzyme preparationshould generate peptides with the most favorable amino acids at the C-
terminal position Some hydrolysates prepared withAlcalase also exerteda strong in vitro and in vivo ACE-inhibitory activity (Cheng et al 2008)Other studies demonstrated that Pro at the antepenultimate position inthe peptide sequence also enhanced binding to the ACE enzyme(Contreras Carroacuten Montero Ramos amp Recio 2009) Most commercialinhibitors (CaptoprilandEnalapril) also bear Proresidue intheirstructureThe reasonably high ACE-inhibitory capacity (IC50=034ndash106 mgmL)achieved with most of the hydrolysates tested may be favored by theabundanceof Proresidues(89residues1000 residues)intheparentsquidgelatin (Alemaacuten Gimeacutenez Montero et al 2011)
33 Cytotoxic and antiproliferative effects against cancer cells
Cytotoxic activity against cancer cells was considered to be the
effect produced when the hydrolysate was added to the culture
0 10 20 30 40 50 min0
250
500
750
mAUTrypsin
50 min0 10 20 30 400
250
500
750
mAUNeutrase
mAU
50 min0 10 20 30 400
250
500
750Esperase
mAU
0 10 20 30 40 50 min0
250
500
750NS37005
0 10 20 30 40 50 min0
250
500
750
mAUSavinase
0 10 20 30 40 50 min0
250
500
750
mAUAlcalase
0 10 20 30 40 50 min0
250
500
750
mAUProtamex
gt 1375 Da lt 500 Da1375-500 Da
Fig 2 Molecular weight distribution of gelatin hydrolysates at a concentration of 10 mgmL by SE-HPLC The absorbance was monitored at 215 nm
Table 1
ACE-inhibitory capacity of squid gelatin hydrolysates
IC50
Protamex 106 plusmn 001e
Trypsin 091 plusmn 003d
Neutrase 063 plusmn 001b
Esperase 089 plusmn 003cd
NS37005 ndSavinase 082 plusmn 008c
Alcalase 034 plusmn 002a
nd not determinedDifferent letters (a b chellip) indicate signi1047297cant differences(ple005) IC50 concentration (mgmL) required for inhibiting
50 of ACE activity
1047 A A lemaacuten e t al F ood Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 58
medium after the cells had been growing for 24 h antiproliferativeactivity was considered to be the effect produced when thehydrolysates were incorporated before cell growth started
Asexpectedsquidgelatin didnot show cytotoxicor antiproliferativeeffect on cancer cells However the hydrolysates prepared from squidgelatin affected the viability of both cell lines differently depending onthe enzyme used (Fig 3A B) Cell viability was determined after 24 48and 72 h with a change of culture medium and theaddition of 1 mgmL
concentration of the hydrolysate on each occasion Only EsperaseAlcalase and Savinase hydrolysates intensi1047297ed their effect on bothcancer cell lines after 48 and 72 h (ple005) The Esperase hydrolysatehad the highest (ple005) cytotoxic effect on both cancer cells (966plusmn05 and 912plusmn27 of viability inhibition on MCF-7 and U87 cellsrespectively after 72 h) It is worth noting that the same results wereobtained for a concentration of 03 mgmL for both cell lines Alcalasehydrolysate also showed relatively high cytotoxic activity (678plusmn56and 839plusmn48 viability inhibition on both cell lines respectively)
Jang Jo Kang and Lee (2008) puri1047297ed four peptides from beef proteinand caused a 20 viability inhibition on MCF-7 at 04 mgmL
Regarding antiproliferative activity again Esperase hydrolysatefollowed by Alcalase hydrolysate exhibited the highest (ple005)
activity on both cell lines (Fig 4) Picot et al (2006) evaluated theantiproliferative activity of 18 1047297sh protein hydrolysates and foundthe highest effect on MCF-7 cell lines to be around 40 for a codhydrolysate obtained with Protamex and Alcalase At the sameconcentration (1 mgmL) the Alcalase squid gelatin hydrolysatecaused a similar effect on MCF-7 cells (4164plusmn53 of growthinhibition) however the Esperase hydrolysate had a much stronger(ple005) effect (9141plusmn 18 of inhibition) and remained similar for
a concentration reduction to 05 mgmLThe IC50 was determined for both MCF-7 and U87 cells in the case
of hydrolysates that had produced the strongest cytotoxic effects(Esperase and Alcalase hydrolysates) at 24 48 and 72 h (Table 2)Both hydrolysates showed low IC50 values for both cell linesespecially the Esperase hydrolysate A pepsin hydrolysate fromalgae protein waste showed an IC50 of 174 mgmL against AGC celllines (Sheih Fang Wu amp Lin 2010) To our knowledge the IC50 valuesfor cytotoxiceffect (013 and 010 mgmL forMCF-7 andU87 cell linesrespectively) of the Esperase hydrolysate were the lowest found forwhole food protein hydrolysates although similar IC50 values havebeen reported for isolated peptides The nonapeptide X-Met-Leu-Pro-Ser-Tyr-Ser-Pro-Tyr obtained by hydrolysis of defatted soy protein
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
v i a b i l i t y i n h i b i t i o n ( )
24h 48h 72h
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
V i a b i l i t y i n h i b i t i o n ( )
24h 48h 72h
A
B
a x
c x
a b y
b x a
b x
b x
a y
a x
a x
b x
b y c
y
b x b
y d x
d x
e y e
z
e x
d y f z
a x a
b y
b y
a x
b y c
y
a x b
y b y
c x d
y e z
a x
a b y
b y
a x
b y c
z b x
c y d
z
Fig 3 Effect of gelatin hydrolysates on the viability of cancer cells for 24 48 and 72 h in cell culture medium containing a concentration of 1 mgmL of hydrolysate MCF-7 cell line(A) U87cellline (B) Differentletters (ab chellip) withinthe same time periodindicatesigni1047297cantdifferences (ple005)Different letters (xy z) withinthe same hydrolysate indicatesigni1047297cant differences (ple005)
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
G r o w t h i n h i b i t i o n ( )
U87 MCF-7
bd
c c
c
d
e
f
b ebc
aa
e
Fig 4 Effect of gelatin hydrolysates on growth of MCF-7 and U87human cells for 72 h in cell culture mediumcontaining a concentration of 1 mgmL of hydrolysate Different letters
(a b chellip) within the same cell cultures indicate signi1047297cant differences (ple005)
1048 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 68
hadanIC50 of 016 mgmL on a mouse monocyte macrophage cell line(Kim et al 2000) A peptide fraction from algae protein waste exertedan IC50 value of 007 mgmL against AGS cells (Sheih et al 2010) Thecytotoxic effect of Esperase hydrolysate on MCF-7 cells increasedafter 48 and 72 h but to a lesser extent than on U87 cells After 24 hthe IC50 for MCF-7 cells (015 mgmL) was lower than that for U87cells (031 mgmL) however after 72 h the Esperase hydrolysate wasmore cytotoxic against U87 cells (IC50=010 mgmL) than againstMCF-7 cells (IC50=013 mgmL)
The Esperase and Alcalase hydrolysates had most of their peptidesin the range of 1400 to 500 Da Sheih et al (2010) fractioned a pepsinhydrolysate from algae protein waste and found that the mosteffective peptide fraction against AGS cells had a molecular weightlower than 6500 Da and most of the peptides (N90) were locatedbetween the range of 2000 and 200 Da Kannan Hettiarachchy
Johnson and Nannapaneni (2008) observed the highest antiprolifera-tive activity in human colon and liver cancer cells in the lowermolecular weight fraction (b5 kDa) of an Alcalase hydrolysate fromrice bran However molecular weight cannot be considered the mostimportant factor in1047298uencing anticancer activity since the Neutrasehydrolysate which has relatively low cytotoxic and antiproliferativeactivities is also made up mostly of peptides in the range of 1400ndash
500 Da Thus the antiproliferativecytotoxic effect against MCF-7and U87 cells should be linked for the most part to the presenceof speci1047297c peptides that exert a direct cytotoxic effect on cancer
cells as previously reported for valorphin a hemoglobin-derivedpeptide (Blishchenko et al 2005) Nevertheless the hypothesis of abinding competition between peptides and FCS growth factor on cellmembrane receptors cannot be excluded either Three peptides froma 1047297sh source have been described as having antitumor activity (i) a4409 Da anchovy hydrophobic peptide able to induce apoptosis inhuman U937 lymphoma cells by increasing caspase-3 and caspase-8 activity (Lee Kim Lee Kim amp Lee 2004) (ii) tilapia hepcidin TH2-3that manifested signi1047297cant antitumor activity in human 1047297brosarcomacells (Chen Lin amp Lin 2009) and (iii) epinecidin-1 a peptide from1047297sh (Epinephelus coioides) which had an antitumor effect similar tolytic peptides in human 1047297brosarcoma cells (Lin et al 2009) Howevernone of these peptides originate from 1047297sh gelatincollagen
34 Antioxidant activity
The radical scavenging capacity of squid gelatin exhibited a 3-foldincrease when it was hydrolyzed with Protamex Neutrase orNS37005 whereas such an increase was between 7 and 10-foldwhen the enzymes used were Trypsin Savinase Esperase or Alcalase(Fig 5A) However radical scavenging capacity could not be wellcorrelated with the molecular weight distribution of the wholehydrolysate since Trypsin hydrolysate was the one with the highestnumber of larger peptides This 1047297nding suggested that the radicalscavenging activity of protein hydrolysates is dependent not onlyon the peptide size but also to a greater extent on their sequencesas a result of the speci1047297city of the enzyme used
The metal chelating ability of a given compound may also serve
as a useful indicator of its potential antioxidant activity As depicted
in Fig 5B the squid gelatin did not show any Fe2+ chelating ability at10 mgmL However when gelatin was hydrolyzed with ProtamexNeutrase or NS37005 enzymes the resulting hydrolysates did exert alow chelating capacity (less than 25) In contrast the TrypsinSavinase or Esperase hydrolysates at the same concentration caused~80 of Fe2+ chelation which was near to 100 with the Alcalasehydrolysate Dong Zeng Wang Liu Zhao and Huicheng(2008) founda metal chelation activity of 9297 with an Alcalase hydrolysate fromsilver carp at 5 mgmL
Hydroxyl radicals can be formed from superoxide anion andhydrogen peroxide in the presence of transition metal ions such asFe2+ andCu2+ Thus chelating metal ions may inhibit theformation of hydroxyl radicals Presumably peptide cleavage leads to enhancedmetal ion binding because carboxyl and amino groups in the sidechains of the acidic (Glx and Asx) and basic (Lys His and Arg) aminoacids are thought to play an important role in chelating metal ions(Saiga Tanabe amp Nishimura 2003)
The molecular weight of peptidesis also believed to play a key rolein chelating metals Klompong Benjakul Kantachote and Shaidi(2007)and Dong et al (2008) pointed out that metal-chelating abilityand molecular weight had a linear relationship and that metal-chelating ability increased with decreasing molecular weight Never-theless Megiacuteas et al (2007) reported that if peptide length is tooshort chelation is unstable In our study hydrolysate with peptidesmostly between ~1400 and 500 Da (Alcalase hydrolysate) exerted
the highest metal chelating ability However it is worth noting theconsiderably high metal chelating activity in the Trypsin hydrolysateas compared to other hydrolysates with higher proportions of smallerpeptides (Neutraseor Protamex) denoting onceagainthe importanceof the presence of speci1047297c peptide sequences rather than the wholehydrolysate molecular weight distribution
The behavior for Fe2+ chelation and ABTS+ scavenging followed asimilar trend suggesting that the two antioxidant mechanisms mightbe related Zhu Chen Tang and Xiong (2008) also found a similartrend between Cu2+ chelating capacity and ABTS radical scavengingfor an Alcalase hydrolysate from zein protein
Another method for measuring antioxidant potential is theferric reducing antioxidant power (FRAP) assay Reducing power isa measurement that provides an estimation of a compounds ability
to reduce ferric iron (III) to ferrous iron (II) and is determinedusing a redox-linked colorimetric reaction Samples with higherreducing power are more able to donate electrons or hydrogenFree radicals form stable substances by accepting donated electronsand thus the free radical chain reaction could be interruptedThe reducing power of squid gelatin and hydrolysates is shownin Fig 5C The ability of the hydrolysates to reduce Fe3+ to Fe2+
was approximately twice than that of the gelatin with all thehydrolysates showing a similar (pN005) capacity independently of the enzyme used and consequently of molecular weight distribu-tion too In several studies involving protein hydrolysates someresearchers reported that the reducing capacity improved as thedegree of protein hydrolysis increased (Thiansilakul Benjakulamp Shahidi 2007) while some researchers reported the opposite
(Klompong et al 2007)
Table 2
Cytotoxic activity of Esperase and Alcalase squid gelatin hydrolysates
IC50
MCF-7 cells U87 cells
24 h 48 h 72 h 24 h 48 h 72 h
Esperase 015 plusmn001a 014plusmn001a 013plusmn001a 031plusmn001c 013plusmn001b 010plusmn001a
Alcalase 118 plusmn002c 104plusmn002b 081plusmn003a 122plusmn003c 098plusmn003b 085plusmn002a
Different letters (a b) within the same hydrolysate and the same cell line indicate signi1047297cant differences (ple005) IC50 concentration (mgmL) required to induce 50 cells death
1049 A A lemaacuten e t al F ood Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 78
Alcalase Esperase Savinase and Trypsin signi1047297cantly improvedthe antioxidant activity of the resulting hydrolysates via the enhance-ment of antiradical activity and metal chelation Reducing powerwas also increased by the action of these enzymes but the degreeof enhancement was much lower Consistent with our results thehydrolysis of snapper gelatin with proteases from pyloric caecacaused a 55 to 65-fold increase in ABTS radical scavenging capacitywhereas ferric reducing power did not increase more than 2-fold(Khantaphant amp Benjakul 2008) Free radical quenchingwas reportedto be the main antioxidative mechanism of some peptides derivedfrom squid gelatin hydrolysates (Mendis Rajapakse amp Kim 2005)
4 Conclusions
The Esperase hydrolysate exerted a potent cytotoxic and anti-proliferative effects on the two cancer cell lines studied withsigni1047297cant antioxidant activity especially with respect to metalchelating and antiradical activities The Alcalase hydrolysate was themost potent ACE-inhibitor and metal chelator while at the same timeexerting strong radical scavenging and cytotoxic activity againstcancer cells These results suggest that squid gelatin hydrolysatesprepared with Esperase or Alcalase enzymes could be utilized as newmaterials in the development of functional foods with potentialanticancer antihypertensive and antioxidant capacities
Acknowledgments
Funding for this research was provided by (i) the Interministerial
Scienti1047297c and Technical Commission of Spains Ministry of Science
and Innovation under project AGL2008-02135ALI and (ii) Xunta deGalicia Program Aacutengeles Alvarintildeo (European Social Fund)
References
AlemaacutenA Gimeacutenez BMonteroP amp Goacutemez-GuilleacutenM C(2011) Antioxidantactivityof several marine skin gelatins LWT Food Science and Technology 44 407minus413
Alemaacuten A Gimeacutenez B Peacuterez-Santiacuten E Goacutemez-Guilleacuten M C amp Montero P (2011)Contribution of Leu and Hyp residues to antioxidant and ACE-inhibitory activitiesof peptide sequences isolated from squid gelatin hydrolysates Food Chemistry 125334minus341
Balti R Nedjar-Arroume N Bougatef A Guillochon D amp Nasri M (2010) Threenovel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttle1047297sh(Sepia of 1047297cinalis) using digestive proteases Food Research International 43(4)1136minus1143
BenzieI F Famp StrainJ J(1996) Theferric reducingabilityof plasma(FRAP) asa measureof ldquoantioxidant powerrdquo The FRAP assay Analytical Biochemistry 239(1) 70minus76
Blishchenko E Y Sazonova O V Kalinina O A Moiseeva E V Vass A A KarelinA A et al (2005) Antitumor effect of valorphin in vitro and in vivo Combinedaction with cytostatic drugs Cancer Biology amp Therapy 4(1) 118minus124
Bougatef A Nedjar-Arroume N Ravallec-Pleacute R Leroy Y Guillochon D Barkia Aet al (2008) Angiotensin I-converting enzyme (ACE) inhibitory activities of sardinelle (Sardinella aurita) by-products protein hydrolysates obtained bytreatment with microbial and visceral 1047297sh serine-proteases Food Chemistry 111350minus356
Chen J Y Lin W J amp Lin T L (2009) A 1047297sh antimicrobial peptide tilapia hepcidinTH2-3 showspotent antitumor activity againsthuman 1047297brosarcomacells Peptides
30 1636minus1642ChengFYWanTCLiuYTLaiKMLinLCampSakataR(2008)Astudyof invivo
antihypertensive properties of enzymatic hydrolysate from chicken leg boneprotein Animal Science Journal 79 614minus619
Cheung H S Wang F L Miguel A O Emily F S amp Dvid W C (1980) Binding of peptide substrates and inhibitors of angiotensin-I-converting enzyme The Journalof Biological Chemistry 255 401minus407
Chung Y C Chang C T Chao W W Lin C F amp Chou S T (2002) Antioxidativeactivity and safety of the 50 ethanolic extract from red bean fermented by Bacillus
subtilis IMR-NK1 Journal of Agricultural and Food Chemistry 50 2454minus2458
0
20
40
60
80
100
C h e l a t i n g a b i l i t y ( )
0
5
10
15
20
micro m o l e s F e E q g
A
a
b
c
d
bb
d d
B
bc
d
b
d
c
de
a
C
a
bbbbb bb
0
10
20
30
40
50
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
V C E A C ( m g g )
Fig 5 Antioxidant activity of gelatin hydrolysates at a concentration of 10 mgmL ABTS radical scavenging capacity (A) Fe chelating ability (B) and Fe reducing power (FRAP) (C)Different letters (a b chellip) indicate signi1047297cant differences (ple005)
1050 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 88
Contreras M Carroacuten R Montero M J Ramos M amp Recio I (2009) Novel casein-derived peptides with antihypertensive activity International Dairy Journal 19566minus573
Dong S Zeng M Wang D Liu Z Zhao Y amp Huicheng Y (2008) Antioxidantand biochemical properties of protein hydrolysates prepared from silver carp(Hypophthalmichthys molitrix) Food Chemistry 107 1485minus1493
Doucet D Otter E O Gauthier S F amp Foegeding E A (2003) Enzyme-inducedgelation of extensively hydrolyzed whey protein by Alcalase Peptides identi1047297ca-tion and determination of enzyme speci1047297city Journal of Agricultural and FoodChemistry 51 6300minus6308
Gimeacutenez B Alemaacuten A Montero P amp Goacutemez-Guilleacuten M C (2009) Antioxidant and
functional properties of gelatin hydrolysates obtained from skin of sole and squidFood Chemistry 114(3) 976minus983Goacutemez-Guilleacuten MC Loacutepez-Caballero ME Alemaacuten ALoacutepezde LaceyA Gimeacutenez B
amp Montero P (2010) Antioxidant and antimicrobial peptide fractions from squidandtunaskin gelatin In E LeBihan(Ed) Seaby-productsas real materialNew waysof application (pp 89minus115) Kerala India Transworld Research Network
Goodfriend T L Elliott M E amp Catt K J (1996) Drug therapy ndash Angiotensin receptorsand their antagonists The New England Journal of Medicine 334(25) 1649minus1654
Gueacuterard F Dufosseacute L De La Broise D amp Binet A (2001) Enzymatic hydrolysis of proteins from yellow1047297n tuna (Thunnus albacares) wastes using Alcalase Journal of Molecular Catalysis B Enzymatic 11 1051minus1059
Hai-Lun H Xiu-Lan C Sai-Yun S Yu-Zhonga Z amp Bai-Chenga Z (2006) Analysisof novel angiotensin-I-converting enzyme inhibitory peptides from protease-hydrolyzed marine shrimp Acetes chinensis Journal of Peptide Science 12 726minus733
Hou W C Chen H J amp Lin Y H (2003) Antioxidant peptides with angiotensinconverting enzymeinhibitory activitiesand applications for angiotensin convertingenzyme puri1047297cation Journal of Agricultural and Food Chemistry 51 1706minus1709
Jang A Jo C Kang K S amp Lee M (2008) Antimicrobial and human cancer cellcytotoxic effect of synthetic angiotensin-converting enzyme (ACE) inhibitory
peptides Food Chemistry 107 327minus336 Je J -Y Lee K -H Lee M H amp Ahn C -B (2009) Antioxidant and antihypertensive
protein hydrolysates produced from tuna liver by enzymatic hydrolysis FoodResearch International 42(9) 1266minus1272
Kannan A Hettiarachchy N Johnson M G amp Nannapaneni R (2008) Human colonand liver cancer cell proliferation inhibition by peptide hydrolysates derived fromheat-stabilized defatted rice bran Journal of Agriculture and Food Chemistry 56 11643minus11647
Kaufman S H amp Earnshaw W C (2000) Induction of apoptosis by cancerchemotherapy Cell Research 256 42minus49
Khantaphant S amp Benjakul S (2008) Comparative study on the proteases from 1047297shpyloric caeca and the use for production of gelatin hydrolysate with antioxidativeactivity Comparative Biochemistry and Physiology 151 410minus419
Kim S K Byun H G Park P J amp Shahidi F (2001) Angiotensin I converting enzymeinhibitory peptides puri1047297ed form bovine skin gelatin hydrolysate Journal of
Agricultural and Food Chemistry 49 2992minus2997Kim S K Kim Y T Byun H G Nam K S Joo D S amp Shahidi F (2001) Isolation and
characterization of antioxidative peptides from gelatin hydrolysate of Alaskapollack skin Journal of Agricultural and Food Chemistry 49 1984minus1989
Kim S E Kim H H Kim J Y Kang Y I Wooa H J amp Lee H J (2000) Anticanceractivity of hydrophobic peptides from soy proteins Biofactor 21 63minus67
Klompong V Benjakul S Kantachote D amp Shahidi F (2007) Antioxidative activityand functional properties of protein hydrolysate of yellow stripe trevally(Selaroides leptolepis) as in1047298uenced by the degree of hydrolysis and enzyme typeFood Chemistry 102 1317minus1327
LeeY G Kim J Y Lee KW Kim K H amp Lee H J (2004) Peptides from anchovy sauceinduce apoptosisin a human lymphoma cell(U937) throughthe increaseof caspase-3and -8 activities Annals of the New York Academy of Sciences 1010 399minus404
Leng B Liu X D amp Chen Q X (2005) Inhibitory effects of anticancer peptide fromMercenaria on the BGC-823 cells and several enzymes FEBS 579 1187minus1190
Lin W J Chien Y L Pan C Y Lin T L Chen J Y Chiu S J amp Hui C F (2009)Epinecidin-1 an antimicrobial peptide from 1047297sh (Epinephelus coioides) which
hasan antitumoreffect like lytic peptides in human 1047297brosarcoma cells Peptides 30283minus290
Lin Lamp LiB (2006) Radical scavengingproperties of protein hydrolysates from jumbo1047298ying squid (Dosidicus eschrichitii Steenstrup) skin gelatin Journal of the Science of Food and Agriculture 86 2290minus2295
Lind D S Hochwald S N Malaty J Rekkas S Hebig P Mishra G et al (2001)Nuclear factor-kB is upregulated in colorectal cancer Surgery 130 363minus369
Megiacuteas C Pedroche J Yust M M Giroacuten-Calle J Alaiz M Millaacuten F amp Vioque J(2007) Af 1047297nity puri1047297cation of copper chelating peptides from chickpea proteinhydrolysates Journal of Agricultural and Food Chemistry 55 3949minus3954
Mendis E Rajapakse N Byun H G amp Kim S K (2005) Investigation of jumbo squid
(Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects LifeScience 77 2166minus2178Mendis E Rajapakse N amp Kim S K (2005) Antioxidant properties of a radical-
scavenging peptide puri1047297ed from enzymatically prepared 1047297sh skin gelatinhydrolysate Journal of Agricultural and Food Chemistry 53(3) 581minus587
Messerli F H (1999) Hypertension and sudden cardiac death American Journal of Hypertension 12 181minus188
Nam K A You S G amp Kim S M (2008) Molecular and physical characteristicsof squid (Toradores paci 1047297cus) skin collagens and biological properties of theirenzymatic hydrolysates Journal of Food Science 73(4) 249minus255
PicotL Bordenave S Didelot S Fruitier-Arnaudin I Sannier F ThorkelssonG et al(2006) Antiproliferative activity of 1047297sh protein hydrolysates on human breastcancer cell lines Process Biochemistry 41 1217minus1222
Re R Pellegrini N Proteggente A Pannala A Yang M amp Rice-Evans C (1999)Antioxidant activity applying an improved ABTSradical cation decolorization assayFree Radical Biology amp Medicine 26 1231minus1237
Saiga A Tanabe S amp Nishimura T (2003) Antioxidant activity of peptides obtainedfrom porcine myo1047297brillar proteins by protease treatment Journal of Agriculturaland Food Chemistry 51(12) 3661minus3667
Samaranayaka A G P Kitts D D amp Li-Chan E C Y (2010) Antioxidative andangiotensin-I-converting enzyme inhibitory potential of a paci1047297c hake (Merluccius
productus) 1047297sh protein hydrolysate subjected to simulated gastrointestinaldigestion and caco-2 cell permeation Journal of Agricultural and Food Chemistry58(3) 1535minus1542
Schagger H amp Von Jagow G (1987) Tricine-sodium dodecyl sulphate polyacrylamidegel electrophoresis for the separation of proteins in the range from 1 to 100 kDa
Analytical Biochemistry 166 368minus379Sheih I Fang T J Wu T K amp LinP H (2010) Anticancer and antioxidant activities of
the peptide fraction from algae protein waste Journal of Agricultural and FoodChemistry 58 1202minus1207
Theodore A E amp Kristinsson H G (2007) Angiotensin converting enzyme inhibitionof 1047297sh protein hydrolysates prepared from alkaline-aided channel cat1047297sh proteinisolate Journal of the Science of Food and Agriculture 87 2353minus2357
Thiansilakul Y Benjakul S amp Shahidi F (2007) Compositions functional propertiesand antioxidative activity of protein hydrolysates prepared from round scad(Decapterus maruadsi) Food Chemistry 103(4) 1385minus1394
Witherow F N Helmy A Webb D J Fox K A A amp Newby D E (2001) Bradykinincontributes to the vasodilator effects of chronic angiotensin converting enzymeinhibition in patients with heart failure Circulation 104(18) 2177minus2181
Wu J Aluko R E amp Muir A D (2002) Improved method for direct high-performanceliquid chromatography assay of angiotensin-converting enzyme-catalyzed reactions
Journal of Chromatography A 950 125minus130Yang J L Ho H Y Chu Y J amp Chow C J (2008) Characteristic and antioxidant
activity of retorted gelatin hydrolysates from cobia (Rachycetron canadum) skinFood Chemistry 110 128minus136
Zhao Y Li B Liu Z Dong S Zhao X amp Zeng M (2007) Antihypertensive effect andpuri1047297cation of an ACE inhibitory peptide from sea cucumber gelatin hydrolysateProcess Biochemistry 42 1586minus1591
Zhu LChenJ Tang Xamp XiongY (2008) Reducingradical scavenging andchelationproperties of in vitro digests of Alcalase-treated zein hydrolysate Journal of
Agricultural and Food Chemistry 56 2714minus2721
1051 A A lemaacuten e t al Food Research International 44 (2011) 1044ndash1051

8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 28
skin gelatin or Goacutemez-Guilleacuten et al (2010) with tuna and giant squidskin gelatins
A number of commercial proteases have been used for theproduction of antioxidant gelatin hydrolysates including trypsinchymotrypsin pepsin Properase E Pronase collagenase bromelainand papain (Kim Kim et al 2001 Mendis Rajapakse Byun et al2005 Mendis Rajapakse amp Kim 2005 Linamp Li2006YangHo Chu ampChow 2008) Alcalase which is a commercial protease from a
microbial source (Bacillus licheniformis) has been shown to be one of the most ef 1047297cient in the hydrolysis of 1047297sh protein (Gueacuterard DufosseacuteDe La Broise amp Binet 2001) This enzyme preparation has also beenreported to produce high levels of hydrolysis in skin gelatin fromAlaska pollack squid Todarodes paci 1047297cus or giant squid Dosidicus gigas
(Kim Byun et al 2001 Nam You amp Kim 2008 Gimeacutenez et al 2009)In a recent work a peptide sequence Gly-Pro-X-Gly-X-X-Gly-Phe-X-Gly-Pro-X-Gly-X-Ser with the X positions occupied by either Hyp orLeu and a molecular mass of 1411 Da together with a compound witha molecular mass of 952 Da associated with the carbohydrate fucosewere identi1047297ed as being responsible for most of the high anti-radicalandACE-inhibitoryactivityin an Alcalasehydrolysate fromgiant squidgelatin (Alemaacuten Gimeacutenez Peacuterez-Santiacuten Goacutemez-Guilleacuten amp Montero2011) Further work with giant squid gelatin revealed that Alcalasealso produced hydrolysates with much higher radical scavengingand ferric reducing capacities than those of other single enzymessuch as collagenase trypsin or pepsin (Alemaacuten Gimeacutenez Monteroet al 2011)
The aim of this study was to explore the potential health bene1047297tswhich could be derived from giant squid tunic gelatin by investigatingthe ACE-inhibitory anticancer and antioxidant properties (radicalscavenging metal chelating and reducing power) of hydrolysateshaving different molecular weight distributions obtained by usingseveral commercial proteases
2 Materials and methods
21 Materials
Gelatin from inner and outer giant squid (D gigas) tunics wasextracted using the method described by Gimeacutenez et al (2009)Commercial proteases (Protamex Trypsin Neutrase 08 L Savinase16 L NS37005 Esperase 08 L and Alcalase 24 L) were kindly suppliedby Novozymes (Bagsvaerd Denmark) NaOH HCl NaCl glycerol andpotassium persulfate were from Panreac (Barcelona Spain) All otherreagents were from Sigma-Aldrich Inc (St Louis MO USA)
22 Preparation of gelatin hydrolysates
The squid gelatin (25 wv) was dissolved in water and subjectedto enzymatic hydrolysis for 3 h under optimal temperature and pHconditions The optimal condition for each protease was as followsProtamex (pH 65 60 degC) Trypsin (pH 8 55 degC) Neutrase 08 L (pH
65 55 degC) Savinase 16 L (pH 95 55 degC) NS37005 (pH 7 55 degC)Esperase 08 L (pH 85 60 degC) and Alcalase 24 L (pH 8 50 degC) with anenzymendashsubstrate ratio of 120 (ww) The pH of the reaction waskept constant by adding a 1 N NaOH solution to the reaction mediumusing a pH-stat (TIM 856 Radiometer Analytical Villeurbanne CedexFrance) The enzymes were inactivated by heating at 90 degC for 10 minand the samples were centrifuged at 3000 g for 15 min The super-natants comprised the hydrolysates and were lyophilized and storedat minus80 degC for further assays
23 Molecular weight distribution
The gelatin hydrolysates were analyzed by Tricine SDS-PAGEaccording to Schagger and Von Jagow (1987) using a 5 stacking
gel and a 165 resolving gel The hydrolysates were dissolved at
a concentration of 10 mgmL in the loading buffer (50 mM TrisndashHCl4 SDS 12 glycerol 2 mercaptoethanol and 001 SERVA Blue G)heat-denatured at 90 degC for 10 min and run in a Mini Protean II unit(Bio-Rad Laboratories Hercules CA USA) Protein bands were stainedwith Coomassie Brilliant Blue R250 The approximate molecular weight(MW) of the hydrolysates was determined using a low molecularweight marker kit (Amersham Pharmacia Biotech Uppsala Sweden)consisting of triosephosphate isomerase (266 kDa) myoglobin
(169 kDa) α-lactalbumin (144 kDa) aprotinin (65 kDa) oxidizedinsulin β chain (35 kDa) and bacitracin (14 kDa)Hydrolysate molecular weight distribution was also evaluated
by size-exclusion HPLC (model SPE-MA10AVP Shimadzu Kyoto Japan) on a superdex peptide PC 3230 column (GE Healthcare Bio-Sciences Barcelona Spain) with a fractionation range between 7000and 100 Da The injection volume was 10 μ L and the 1047298ow rate was01 mLmin using Milli-Q water in the mobile phase Vitamin B12(1355 Da) and hippuryl-L -histidyl-L -leucine (HHL 502 Da) bothfrom Sigma-Aldrich Inc (St Louis MO USA) were used as molecularweight standards
24 ACE-inhibition
Reversed-phase high performance liquid chromatography (RP-HPLC) was used to determine ACE inhibition applying the methodused by Wu Aluko and Muir (2002) with slight modi1047297cations Thetotal reaction volume was 225 μ L made up of 50 μ L of 5 mM HHL160 μ L of 25 mUmL of ACE and 20 μ L of sample at six differentconcentrations (from 01 to 15 mgmL 1047297nal concentration) preparedwith 100 mM potassium-phosphate buffer containing 300 mM NaClpH 83 The mixture was incubated at 37 degC for 120 min and thereaction was quenched by adding 100 μ L of 01 M HCl The releasedhippuric acid (HA) was quanti1047297ed by RP-HPLC (model SPE-MA10AVPShimadzu Kyoto Japan) on a C18 column (Tracel excel 120 ODSA5 μ m 25times046 Teknokroma Barcelona Spain) The injected volumewas 50 μ L and the 1047298ow rate was 08 mLmin using an acetonitrilegradient of 20 to 60 in 01 tri1047298uoroacetic acid (TFA) for 26 minThe HA and HHL were monitored at 228 nm and the retention times
were 830 and 1570 min respectively All determinations werecarried out at least in triplicate The IC50 value of ACE inhibitoryactivity was de1047297ned as the concentration of hydrolysate (mgmL)required to reduce the HA peak by 50
25 Cytotoxic and antiproliferative activities
The study of cytotoxic and antiproliferative activities of squidgelatin hydrolysates was performed according to the methodologyproposed by Picot et al (2006) MCF-7 human breast carcinomaand U87 glioma cell lines were grown at 37 degC in a 5 CO 2 95 airhumidi1047297ed atmosphere in DMEM-Hams F12 medium (11 vvGibco) The medium was supplemented with 5 heat inactivatedFetal Calf Serum (FCS Dutscher) to which penicillin (100 UmL) and
streptomycin (100 μ gmL) had been added (Pan biotech GmbH) Thecells grown in 1047298asks were trypsinized centrifuged at 800 rpm for5 min and dissolved in fresh culture medium at 104 cells90 μ LSubsequently a 90 μ L volume of suspension cells (10000 cells) wasadded to each well of a 96-well microplate and incubated at 37 degC for24 h In order to perform cytotoxicity screening of the hydrolysateshydrolysate stock solution at a concentration of 10 mgmL wasprepared in PBS 01 M (pH 74) and diluted 10-fold in cell culturemedium containing the cells The microplate was then incubated at37 degC for24 48 and72 h changing the culture mediumevery 24 h andadding the hydrolysate at a 1047297nal concentration of 1 mgmL At the endof every incubation period 15 μ L of 5 mgmL tetrazolium salt (MTT)solution was added to each well and the plate was incubated for3 h To stop succinate-tetrazolium reductase activity and solubilize
formazan crystals 200 μ L of dimethyl sulfoxide (DMSO) was then
1045 A A lemaacuten e t al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 38
added to each well and kept at 37 degC for 1 h Absorbance was read ona plate reader (VERSAmax Molecular Devices Saint Gregoire France)at 570 nm The data were analyzed to calculate the percentageof viability inhibition induced by the presence of hydrolysate incell culture medium determined by the equation Viability inhibition()=100minus(AHtimes100AC) where AH and AC are the absorbenciesmeasured for cell viability in culture medium containing onlyhydrolysate or PBS respectively
Following the same procedure the effect of addition of hydro-lysates at the same time as the cancer cells at the beginning of theexperiment was also evaluated For this evaluation the initialconditions of the culture medium and the added hydrolysates weremaintained for 72 h
The viability inhibition of the Esperase and Alcalase hydrolysateswas also measured using six different concentrations ranging from005 to 15 mgmL in order to determine the IC50 All experimentswere performed at least in triplicate
26 Antioxidant activities
Different assays (ABTS chelating ability and FRAP) were per-formed to study the antioxidant properties of the squid gelatin andthe corresponding hydrolysates at a concentration of 10 mgmL TheABTSradical [22prime-azino-bis-(3-ethylbenzothiazoline-6-sulfonic acid)]scavenging assay was carried out according to the method describedby Re Pellegrini Proteggente Pannala Yang and Rice-Evans (1999)The stock solution of ABTS radical consisted of 7 mM ABTS inpotassium persulfate 245 mM kept in the dark at room temperaturefor 16 h An aliquot of stock solution was diluted with Milli-Q waterin order to give the working solution of ABTS radical an absorbanceat 734 nm of 070plusmn002 A 20 μ L aliquot sample was mixed with980 μ L of ABTS reagent and then left to stand in the dark at 30 degCfor 10 min Absorbance values were read at 734 nm using a UV-1601spectrophotometer (Model CPS-240 Shimadzu Japan) Resultswere expressed as mg Vitamin C Equivalent Antioxidant Capacity(VCEAC)g of sample (gelatin or hydrolysate) based on a standardcurve of vitamin C
The Fe2+ chelating activity of the hydrolysates at a concentrationof 10 mgmL was measured by the method introduced by ChungChang Chao Lin andChou (2002) Brie1047298y a test sample of 800 μ Lwasmixed with 10 μ L of 2 mM FeCl2 and 20 μ L of 5 mM ferrozine Themixture was vortexed and kept at room temperature for 10 min priorto measuring the absorbance at 562 nm using a UV-1601 spectro-photometer (Model CPS-240 Shimadzu Japan) Chelating abilityexpressed as a percentage was calculated as follows Chelatingability ()=[1minus(Abs sample Abs control)]times 100 The control wasa mixture composed of 800 μ L of Milli-Q water 10 μ L of 2 mM FeCl2and 20 μ L of 5 mM ferrozine
FRAP a measure of the reducing power of samples was performedaccording to the method described by Benzie and Strain (1996) withsome modi1047297cations A 60 μ L of dissolved sample (at a concentration of
10 mgmL)was incubated at 37 degCwith 60μ L of Milli-Q waterand 900 μ L of FRAP reagent containing 10 mM of TPTZ (246-tripyridyl-s-triazine)and 20 mMof FeCl3 Absorbance valueswerereadat595 nmafter 30 minusing a UV-1601 spectrophotometer (Model CPS-240 Shimadzu Japan)Results were expressed as μ mol Fe2+ equivalentsg of sample (gelatin orhydrolysate) based on a standard curve of FeSO4middot7H2O All determina-tions were carried out at least in triplicate
27 Statistical analysis
Statistical tests were performed using the SPSSreg computerprogramme (SPSS Statistical Software Inc Chicago IL USA) Datawere expressed as meanplusmn standard deviation of the mean (n= 3 orn=24) Tukeys test was used to determine the level of signi1047297cance
at ple005
3 Results and discussion
31 Molecular weight distribution
Analysis of the overall molecular weight distribution of thedifferent hydrolysates (Fig 1) was carried out by Tricine SDS-PAGEThe Trypsin and NS37005 hydrolysates showed the highest quantityof peptide bands in the MW range between 26 and 35 kDa An
appreciable amount of protein with MW even higher than 26 kDawas also present in the Trypsin hydrolysates denoting the lowestef 1047297ciency in protein breakdown All the hydrolysates were character-ized by presenting an intense band at MWle14 kDa Howeverno peptide bands of more than 14 kDa could be observed in theAlcalase hydrolysates and hardly any in the Neutrase hydrolysateand Esperase hydrolysate indicating a considerably high degree of proteolysis Hydrolysates composed of a wide range of polydisperseproducts with varying degrees of hydrolysis and mostly small peptidescannot usually be properly detected under analytical electrophoresisconditions (Khantaphant amp Benjakul 2008 Gimeacutenez et al 2009)Consequently in order to know the differences in the molecularweight distribution of the smallest peptides (MWle14 kDa) size-exclusion HPLC (SE-HPLC) was performed in a fractionation rangebetween 7000 and 100 Da (Fig 2) The varying MW pro1047297lesrevealed considerable differences in the degree of protein breakdowndepending on the enzyme used In accordance with the electrophoreticpro1047297le the Alcalase Neutrase and Esperase hydrolysates showedhigher levels of peptides with MW below 1400 Da whereas in Trypsinand NS37005 hydrolysate peptides with MW above 1400 Da werepredominant
32 ACE-inhibitory capacity
The amount of hydrolysate required to inhibit 50 of the ACEactivity (IC50) is shown in Table 1 Unlike the parent squid gelatin allthe hydrolysates presented ACE inhibitory capacity although it wasmarkedly differentdepending on theenzyme used It wasnot possibleto determine the IC50 value in the NS37005 hydrolysate whereas the
Alcalase hydrolysate showed the most potent effect with an IC50
of 034 mgmL followed by the Neutrase hydrolysate (063 mgmL)(ple005) As revealed by SDS-PAGE and SE-HPLC the peptides inthe Alcalase and Neutrase hydrolysates were mostly between ~1400and 500 Da In another study Zhao et al (2007) found the highestACE-inhibitory capacity for a sea cucumber gelatin hydrolysate in thefraction with MWb1 kDa Nevertheless there are other studies where
Fig 1 Electrophoretic pro1047297les of gelatin hydrolysates at a concentration of 10 mgmL (Tricine SDS-PAGE) (m) molecular weight markers (a) Protamex (b) Trypsin
(c) Neutrase (d) Esperase (e) NS37005 (f) Savinase and (g) Alcalase
1046 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 48
very small peptides did not appear to increase the activity of thewhole hydrolysate but rather decreased it (Theodore amp Kristinsson2007)
Bougatef et al (2008) reported IC50 values in the concentrationrange of 12ndash74 mgmL for sardinelle by-product hydrolysatesprepared with different proteases while the IC50 of a marine shrimphydrolysate digested by a crude protease was 097 mgmL (Hai-lunXiu-Lan Cai-Yun Yu-Zhong amp Bai-Cheing 2006) Three peptides Val-Tyr-Ala-Pro Val-Ile-Ile-Phe and Met-Ala-Trp with IC50 values of 61 87and 1632 μ M respectively had been reported from cuttle1047297sh muscleprotein hydrolysates (Balti Nedjar-Arroume Bougatef Guillochon amp
Nasri 2010) Although values of IC50 for ACE-inhibitory capacity of skingelatin whole hydrolysates had been previously reported by Kim Kimet al (2001) and Zhao et al (2007) comparisons with IC50 valuesin thepresent study were not fully possible due to the different methodologyemployed for measuring activity
Proline or aromatic amino acids existing at theC-terminal are suitablefor peptide binding to ACE (Cheung Wang Miguel Emily amp Dvid 1980)Alcalase has broad speci1047297city hydrolyzing most peptide bonds preferen-tially those containing aromatic amino acid residues (Doucet OtterGauthier amp Foegeding 2003) suggesting that this enzyme preparationshould generate peptides with the most favorable amino acids at the C-
terminal position Some hydrolysates prepared withAlcalase also exerteda strong in vitro and in vivo ACE-inhibitory activity (Cheng et al 2008)Other studies demonstrated that Pro at the antepenultimate position inthe peptide sequence also enhanced binding to the ACE enzyme(Contreras Carroacuten Montero Ramos amp Recio 2009) Most commercialinhibitors (CaptoprilandEnalapril) also bear Proresidue intheirstructureThe reasonably high ACE-inhibitory capacity (IC50=034ndash106 mgmL)achieved with most of the hydrolysates tested may be favored by theabundanceof Proresidues(89residues1000 residues)intheparentsquidgelatin (Alemaacuten Gimeacutenez Montero et al 2011)
33 Cytotoxic and antiproliferative effects against cancer cells
Cytotoxic activity against cancer cells was considered to be the
effect produced when the hydrolysate was added to the culture
0 10 20 30 40 50 min0
250
500
750
mAUTrypsin
50 min0 10 20 30 400
250
500
750
mAUNeutrase
mAU
50 min0 10 20 30 400
250
500
750Esperase
mAU
0 10 20 30 40 50 min0
250
500
750NS37005
0 10 20 30 40 50 min0
250
500
750
mAUSavinase
0 10 20 30 40 50 min0
250
500
750
mAUAlcalase
0 10 20 30 40 50 min0
250
500
750
mAUProtamex
gt 1375 Da lt 500 Da1375-500 Da
Fig 2 Molecular weight distribution of gelatin hydrolysates at a concentration of 10 mgmL by SE-HPLC The absorbance was monitored at 215 nm
Table 1
ACE-inhibitory capacity of squid gelatin hydrolysates
IC50
Protamex 106 plusmn 001e
Trypsin 091 plusmn 003d
Neutrase 063 plusmn 001b
Esperase 089 plusmn 003cd
NS37005 ndSavinase 082 plusmn 008c
Alcalase 034 plusmn 002a
nd not determinedDifferent letters (a b chellip) indicate signi1047297cant differences(ple005) IC50 concentration (mgmL) required for inhibiting
50 of ACE activity
1047 A A lemaacuten e t al F ood Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 58
medium after the cells had been growing for 24 h antiproliferativeactivity was considered to be the effect produced when thehydrolysates were incorporated before cell growth started
Asexpectedsquidgelatin didnot show cytotoxicor antiproliferativeeffect on cancer cells However the hydrolysates prepared from squidgelatin affected the viability of both cell lines differently depending onthe enzyme used (Fig 3A B) Cell viability was determined after 24 48and 72 h with a change of culture medium and theaddition of 1 mgmL
concentration of the hydrolysate on each occasion Only EsperaseAlcalase and Savinase hydrolysates intensi1047297ed their effect on bothcancer cell lines after 48 and 72 h (ple005) The Esperase hydrolysatehad the highest (ple005) cytotoxic effect on both cancer cells (966plusmn05 and 912plusmn27 of viability inhibition on MCF-7 and U87 cellsrespectively after 72 h) It is worth noting that the same results wereobtained for a concentration of 03 mgmL for both cell lines Alcalasehydrolysate also showed relatively high cytotoxic activity (678plusmn56and 839plusmn48 viability inhibition on both cell lines respectively)
Jang Jo Kang and Lee (2008) puri1047297ed four peptides from beef proteinand caused a 20 viability inhibition on MCF-7 at 04 mgmL
Regarding antiproliferative activity again Esperase hydrolysatefollowed by Alcalase hydrolysate exhibited the highest (ple005)
activity on both cell lines (Fig 4) Picot et al (2006) evaluated theantiproliferative activity of 18 1047297sh protein hydrolysates and foundthe highest effect on MCF-7 cell lines to be around 40 for a codhydrolysate obtained with Protamex and Alcalase At the sameconcentration (1 mgmL) the Alcalase squid gelatin hydrolysatecaused a similar effect on MCF-7 cells (4164plusmn53 of growthinhibition) however the Esperase hydrolysate had a much stronger(ple005) effect (9141plusmn 18 of inhibition) and remained similar for
a concentration reduction to 05 mgmLThe IC50 was determined for both MCF-7 and U87 cells in the case
of hydrolysates that had produced the strongest cytotoxic effects(Esperase and Alcalase hydrolysates) at 24 48 and 72 h (Table 2)Both hydrolysates showed low IC50 values for both cell linesespecially the Esperase hydrolysate A pepsin hydrolysate fromalgae protein waste showed an IC50 of 174 mgmL against AGC celllines (Sheih Fang Wu amp Lin 2010) To our knowledge the IC50 valuesfor cytotoxiceffect (013 and 010 mgmL forMCF-7 andU87 cell linesrespectively) of the Esperase hydrolysate were the lowest found forwhole food protein hydrolysates although similar IC50 values havebeen reported for isolated peptides The nonapeptide X-Met-Leu-Pro-Ser-Tyr-Ser-Pro-Tyr obtained by hydrolysis of defatted soy protein
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
v i a b i l i t y i n h i b i t i o n ( )
24h 48h 72h
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
V i a b i l i t y i n h i b i t i o n ( )
24h 48h 72h
A
B
a x
c x
a b y
b x a
b x
b x
a y
a x
a x
b x
b y c
y
b x b
y d x
d x
e y e
z
e x
d y f z
a x a
b y
b y
a x
b y c
y
a x b
y b y
c x d
y e z
a x
a b y
b y
a x
b y c
z b x
c y d
z
Fig 3 Effect of gelatin hydrolysates on the viability of cancer cells for 24 48 and 72 h in cell culture medium containing a concentration of 1 mgmL of hydrolysate MCF-7 cell line(A) U87cellline (B) Differentletters (ab chellip) withinthe same time periodindicatesigni1047297cantdifferences (ple005)Different letters (xy z) withinthe same hydrolysate indicatesigni1047297cant differences (ple005)
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
G r o w t h i n h i b i t i o n ( )
U87 MCF-7
bd
c c
c
d
e
f
b ebc
aa
e
Fig 4 Effect of gelatin hydrolysates on growth of MCF-7 and U87human cells for 72 h in cell culture mediumcontaining a concentration of 1 mgmL of hydrolysate Different letters
(a b chellip) within the same cell cultures indicate signi1047297cant differences (ple005)
1048 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 68
hadanIC50 of 016 mgmL on a mouse monocyte macrophage cell line(Kim et al 2000) A peptide fraction from algae protein waste exertedan IC50 value of 007 mgmL against AGS cells (Sheih et al 2010) Thecytotoxic effect of Esperase hydrolysate on MCF-7 cells increasedafter 48 and 72 h but to a lesser extent than on U87 cells After 24 hthe IC50 for MCF-7 cells (015 mgmL) was lower than that for U87cells (031 mgmL) however after 72 h the Esperase hydrolysate wasmore cytotoxic against U87 cells (IC50=010 mgmL) than againstMCF-7 cells (IC50=013 mgmL)
The Esperase and Alcalase hydrolysates had most of their peptidesin the range of 1400 to 500 Da Sheih et al (2010) fractioned a pepsinhydrolysate from algae protein waste and found that the mosteffective peptide fraction against AGS cells had a molecular weightlower than 6500 Da and most of the peptides (N90) were locatedbetween the range of 2000 and 200 Da Kannan Hettiarachchy
Johnson and Nannapaneni (2008) observed the highest antiprolifera-tive activity in human colon and liver cancer cells in the lowermolecular weight fraction (b5 kDa) of an Alcalase hydrolysate fromrice bran However molecular weight cannot be considered the mostimportant factor in1047298uencing anticancer activity since the Neutrasehydrolysate which has relatively low cytotoxic and antiproliferativeactivities is also made up mostly of peptides in the range of 1400ndash
500 Da Thus the antiproliferativecytotoxic effect against MCF-7and U87 cells should be linked for the most part to the presenceof speci1047297c peptides that exert a direct cytotoxic effect on cancer
cells as previously reported for valorphin a hemoglobin-derivedpeptide (Blishchenko et al 2005) Nevertheless the hypothesis of abinding competition between peptides and FCS growth factor on cellmembrane receptors cannot be excluded either Three peptides froma 1047297sh source have been described as having antitumor activity (i) a4409 Da anchovy hydrophobic peptide able to induce apoptosis inhuman U937 lymphoma cells by increasing caspase-3 and caspase-8 activity (Lee Kim Lee Kim amp Lee 2004) (ii) tilapia hepcidin TH2-3that manifested signi1047297cant antitumor activity in human 1047297brosarcomacells (Chen Lin amp Lin 2009) and (iii) epinecidin-1 a peptide from1047297sh (Epinephelus coioides) which had an antitumor effect similar tolytic peptides in human 1047297brosarcoma cells (Lin et al 2009) Howevernone of these peptides originate from 1047297sh gelatincollagen
34 Antioxidant activity
The radical scavenging capacity of squid gelatin exhibited a 3-foldincrease when it was hydrolyzed with Protamex Neutrase orNS37005 whereas such an increase was between 7 and 10-foldwhen the enzymes used were Trypsin Savinase Esperase or Alcalase(Fig 5A) However radical scavenging capacity could not be wellcorrelated with the molecular weight distribution of the wholehydrolysate since Trypsin hydrolysate was the one with the highestnumber of larger peptides This 1047297nding suggested that the radicalscavenging activity of protein hydrolysates is dependent not onlyon the peptide size but also to a greater extent on their sequencesas a result of the speci1047297city of the enzyme used
The metal chelating ability of a given compound may also serve
as a useful indicator of its potential antioxidant activity As depicted
in Fig 5B the squid gelatin did not show any Fe2+ chelating ability at10 mgmL However when gelatin was hydrolyzed with ProtamexNeutrase or NS37005 enzymes the resulting hydrolysates did exert alow chelating capacity (less than 25) In contrast the TrypsinSavinase or Esperase hydrolysates at the same concentration caused~80 of Fe2+ chelation which was near to 100 with the Alcalasehydrolysate Dong Zeng Wang Liu Zhao and Huicheng(2008) founda metal chelation activity of 9297 with an Alcalase hydrolysate fromsilver carp at 5 mgmL
Hydroxyl radicals can be formed from superoxide anion andhydrogen peroxide in the presence of transition metal ions such asFe2+ andCu2+ Thus chelating metal ions may inhibit theformation of hydroxyl radicals Presumably peptide cleavage leads to enhancedmetal ion binding because carboxyl and amino groups in the sidechains of the acidic (Glx and Asx) and basic (Lys His and Arg) aminoacids are thought to play an important role in chelating metal ions(Saiga Tanabe amp Nishimura 2003)
The molecular weight of peptidesis also believed to play a key rolein chelating metals Klompong Benjakul Kantachote and Shaidi(2007)and Dong et al (2008) pointed out that metal-chelating abilityand molecular weight had a linear relationship and that metal-chelating ability increased with decreasing molecular weight Never-theless Megiacuteas et al (2007) reported that if peptide length is tooshort chelation is unstable In our study hydrolysate with peptidesmostly between ~1400 and 500 Da (Alcalase hydrolysate) exerted
the highest metal chelating ability However it is worth noting theconsiderably high metal chelating activity in the Trypsin hydrolysateas compared to other hydrolysates with higher proportions of smallerpeptides (Neutraseor Protamex) denoting onceagainthe importanceof the presence of speci1047297c peptide sequences rather than the wholehydrolysate molecular weight distribution
The behavior for Fe2+ chelation and ABTS+ scavenging followed asimilar trend suggesting that the two antioxidant mechanisms mightbe related Zhu Chen Tang and Xiong (2008) also found a similartrend between Cu2+ chelating capacity and ABTS radical scavengingfor an Alcalase hydrolysate from zein protein
Another method for measuring antioxidant potential is theferric reducing antioxidant power (FRAP) assay Reducing power isa measurement that provides an estimation of a compounds ability
to reduce ferric iron (III) to ferrous iron (II) and is determinedusing a redox-linked colorimetric reaction Samples with higherreducing power are more able to donate electrons or hydrogenFree radicals form stable substances by accepting donated electronsand thus the free radical chain reaction could be interruptedThe reducing power of squid gelatin and hydrolysates is shownin Fig 5C The ability of the hydrolysates to reduce Fe3+ to Fe2+
was approximately twice than that of the gelatin with all thehydrolysates showing a similar (pN005) capacity independently of the enzyme used and consequently of molecular weight distribu-tion too In several studies involving protein hydrolysates someresearchers reported that the reducing capacity improved as thedegree of protein hydrolysis increased (Thiansilakul Benjakulamp Shahidi 2007) while some researchers reported the opposite
(Klompong et al 2007)
Table 2
Cytotoxic activity of Esperase and Alcalase squid gelatin hydrolysates
IC50
MCF-7 cells U87 cells
24 h 48 h 72 h 24 h 48 h 72 h
Esperase 015 plusmn001a 014plusmn001a 013plusmn001a 031plusmn001c 013plusmn001b 010plusmn001a
Alcalase 118 plusmn002c 104plusmn002b 081plusmn003a 122plusmn003c 098plusmn003b 085plusmn002a
Different letters (a b) within the same hydrolysate and the same cell line indicate signi1047297cant differences (ple005) IC50 concentration (mgmL) required to induce 50 cells death
1049 A A lemaacuten e t al F ood Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 78
Alcalase Esperase Savinase and Trypsin signi1047297cantly improvedthe antioxidant activity of the resulting hydrolysates via the enhance-ment of antiradical activity and metal chelation Reducing powerwas also increased by the action of these enzymes but the degreeof enhancement was much lower Consistent with our results thehydrolysis of snapper gelatin with proteases from pyloric caecacaused a 55 to 65-fold increase in ABTS radical scavenging capacitywhereas ferric reducing power did not increase more than 2-fold(Khantaphant amp Benjakul 2008) Free radical quenchingwas reportedto be the main antioxidative mechanism of some peptides derivedfrom squid gelatin hydrolysates (Mendis Rajapakse amp Kim 2005)
4 Conclusions
The Esperase hydrolysate exerted a potent cytotoxic and anti-proliferative effects on the two cancer cell lines studied withsigni1047297cant antioxidant activity especially with respect to metalchelating and antiradical activities The Alcalase hydrolysate was themost potent ACE-inhibitor and metal chelator while at the same timeexerting strong radical scavenging and cytotoxic activity againstcancer cells These results suggest that squid gelatin hydrolysatesprepared with Esperase or Alcalase enzymes could be utilized as newmaterials in the development of functional foods with potentialanticancer antihypertensive and antioxidant capacities
Acknowledgments
Funding for this research was provided by (i) the Interministerial
Scienti1047297c and Technical Commission of Spains Ministry of Science
and Innovation under project AGL2008-02135ALI and (ii) Xunta deGalicia Program Aacutengeles Alvarintildeo (European Social Fund)
References
AlemaacutenA Gimeacutenez BMonteroP amp Goacutemez-GuilleacutenM C(2011) Antioxidantactivityof several marine skin gelatins LWT Food Science and Technology 44 407minus413
Alemaacuten A Gimeacutenez B Peacuterez-Santiacuten E Goacutemez-Guilleacuten M C amp Montero P (2011)Contribution of Leu and Hyp residues to antioxidant and ACE-inhibitory activitiesof peptide sequences isolated from squid gelatin hydrolysates Food Chemistry 125334minus341
Balti R Nedjar-Arroume N Bougatef A Guillochon D amp Nasri M (2010) Threenovel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttle1047297sh(Sepia of 1047297cinalis) using digestive proteases Food Research International 43(4)1136minus1143
BenzieI F Famp StrainJ J(1996) Theferric reducingabilityof plasma(FRAP) asa measureof ldquoantioxidant powerrdquo The FRAP assay Analytical Biochemistry 239(1) 70minus76
Blishchenko E Y Sazonova O V Kalinina O A Moiseeva E V Vass A A KarelinA A et al (2005) Antitumor effect of valorphin in vitro and in vivo Combinedaction with cytostatic drugs Cancer Biology amp Therapy 4(1) 118minus124
Bougatef A Nedjar-Arroume N Ravallec-Pleacute R Leroy Y Guillochon D Barkia Aet al (2008) Angiotensin I-converting enzyme (ACE) inhibitory activities of sardinelle (Sardinella aurita) by-products protein hydrolysates obtained bytreatment with microbial and visceral 1047297sh serine-proteases Food Chemistry 111350minus356
Chen J Y Lin W J amp Lin T L (2009) A 1047297sh antimicrobial peptide tilapia hepcidinTH2-3 showspotent antitumor activity againsthuman 1047297brosarcomacells Peptides
30 1636minus1642ChengFYWanTCLiuYTLaiKMLinLCampSakataR(2008)Astudyof invivo
antihypertensive properties of enzymatic hydrolysate from chicken leg boneprotein Animal Science Journal 79 614minus619
Cheung H S Wang F L Miguel A O Emily F S amp Dvid W C (1980) Binding of peptide substrates and inhibitors of angiotensin-I-converting enzyme The Journalof Biological Chemistry 255 401minus407
Chung Y C Chang C T Chao W W Lin C F amp Chou S T (2002) Antioxidativeactivity and safety of the 50 ethanolic extract from red bean fermented by Bacillus
subtilis IMR-NK1 Journal of Agricultural and Food Chemistry 50 2454minus2458
0
20
40
60
80
100
C h e l a t i n g a b i l i t y ( )
0
5
10
15
20
micro m o l e s F e E q g
A
a
b
c
d
bb
d d
B
bc
d
b
d
c
de
a
C
a
bbbbb bb
0
10
20
30
40
50
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
V C E A C ( m g g )
Fig 5 Antioxidant activity of gelatin hydrolysates at a concentration of 10 mgmL ABTS radical scavenging capacity (A) Fe chelating ability (B) and Fe reducing power (FRAP) (C)Different letters (a b chellip) indicate signi1047297cant differences (ple005)
1050 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 88
Contreras M Carroacuten R Montero M J Ramos M amp Recio I (2009) Novel casein-derived peptides with antihypertensive activity International Dairy Journal 19566minus573
Dong S Zeng M Wang D Liu Z Zhao Y amp Huicheng Y (2008) Antioxidantand biochemical properties of protein hydrolysates prepared from silver carp(Hypophthalmichthys molitrix) Food Chemistry 107 1485minus1493
Doucet D Otter E O Gauthier S F amp Foegeding E A (2003) Enzyme-inducedgelation of extensively hydrolyzed whey protein by Alcalase Peptides identi1047297ca-tion and determination of enzyme speci1047297city Journal of Agricultural and FoodChemistry 51 6300minus6308
Gimeacutenez B Alemaacuten A Montero P amp Goacutemez-Guilleacuten M C (2009) Antioxidant and
functional properties of gelatin hydrolysates obtained from skin of sole and squidFood Chemistry 114(3) 976minus983Goacutemez-Guilleacuten MC Loacutepez-Caballero ME Alemaacuten ALoacutepezde LaceyA Gimeacutenez B
amp Montero P (2010) Antioxidant and antimicrobial peptide fractions from squidandtunaskin gelatin In E LeBihan(Ed) Seaby-productsas real materialNew waysof application (pp 89minus115) Kerala India Transworld Research Network
Goodfriend T L Elliott M E amp Catt K J (1996) Drug therapy ndash Angiotensin receptorsand their antagonists The New England Journal of Medicine 334(25) 1649minus1654
Gueacuterard F Dufosseacute L De La Broise D amp Binet A (2001) Enzymatic hydrolysis of proteins from yellow1047297n tuna (Thunnus albacares) wastes using Alcalase Journal of Molecular Catalysis B Enzymatic 11 1051minus1059
Hai-Lun H Xiu-Lan C Sai-Yun S Yu-Zhonga Z amp Bai-Chenga Z (2006) Analysisof novel angiotensin-I-converting enzyme inhibitory peptides from protease-hydrolyzed marine shrimp Acetes chinensis Journal of Peptide Science 12 726minus733
Hou W C Chen H J amp Lin Y H (2003) Antioxidant peptides with angiotensinconverting enzymeinhibitory activitiesand applications for angiotensin convertingenzyme puri1047297cation Journal of Agricultural and Food Chemistry 51 1706minus1709
Jang A Jo C Kang K S amp Lee M (2008) Antimicrobial and human cancer cellcytotoxic effect of synthetic angiotensin-converting enzyme (ACE) inhibitory
peptides Food Chemistry 107 327minus336 Je J -Y Lee K -H Lee M H amp Ahn C -B (2009) Antioxidant and antihypertensive
protein hydrolysates produced from tuna liver by enzymatic hydrolysis FoodResearch International 42(9) 1266minus1272
Kannan A Hettiarachchy N Johnson M G amp Nannapaneni R (2008) Human colonand liver cancer cell proliferation inhibition by peptide hydrolysates derived fromheat-stabilized defatted rice bran Journal of Agriculture and Food Chemistry 56 11643minus11647
Kaufman S H amp Earnshaw W C (2000) Induction of apoptosis by cancerchemotherapy Cell Research 256 42minus49
Khantaphant S amp Benjakul S (2008) Comparative study on the proteases from 1047297shpyloric caeca and the use for production of gelatin hydrolysate with antioxidativeactivity Comparative Biochemistry and Physiology 151 410minus419
Kim S K Byun H G Park P J amp Shahidi F (2001) Angiotensin I converting enzymeinhibitory peptides puri1047297ed form bovine skin gelatin hydrolysate Journal of
Agricultural and Food Chemistry 49 2992minus2997Kim S K Kim Y T Byun H G Nam K S Joo D S amp Shahidi F (2001) Isolation and
characterization of antioxidative peptides from gelatin hydrolysate of Alaskapollack skin Journal of Agricultural and Food Chemistry 49 1984minus1989
Kim S E Kim H H Kim J Y Kang Y I Wooa H J amp Lee H J (2000) Anticanceractivity of hydrophobic peptides from soy proteins Biofactor 21 63minus67
Klompong V Benjakul S Kantachote D amp Shahidi F (2007) Antioxidative activityand functional properties of protein hydrolysate of yellow stripe trevally(Selaroides leptolepis) as in1047298uenced by the degree of hydrolysis and enzyme typeFood Chemistry 102 1317minus1327
LeeY G Kim J Y Lee KW Kim K H amp Lee H J (2004) Peptides from anchovy sauceinduce apoptosisin a human lymphoma cell(U937) throughthe increaseof caspase-3and -8 activities Annals of the New York Academy of Sciences 1010 399minus404
Leng B Liu X D amp Chen Q X (2005) Inhibitory effects of anticancer peptide fromMercenaria on the BGC-823 cells and several enzymes FEBS 579 1187minus1190
Lin W J Chien Y L Pan C Y Lin T L Chen J Y Chiu S J amp Hui C F (2009)Epinecidin-1 an antimicrobial peptide from 1047297sh (Epinephelus coioides) which
hasan antitumoreffect like lytic peptides in human 1047297brosarcoma cells Peptides 30283minus290
Lin Lamp LiB (2006) Radical scavengingproperties of protein hydrolysates from jumbo1047298ying squid (Dosidicus eschrichitii Steenstrup) skin gelatin Journal of the Science of Food and Agriculture 86 2290minus2295
Lind D S Hochwald S N Malaty J Rekkas S Hebig P Mishra G et al (2001)Nuclear factor-kB is upregulated in colorectal cancer Surgery 130 363minus369
Megiacuteas C Pedroche J Yust M M Giroacuten-Calle J Alaiz M Millaacuten F amp Vioque J(2007) Af 1047297nity puri1047297cation of copper chelating peptides from chickpea proteinhydrolysates Journal of Agricultural and Food Chemistry 55 3949minus3954
Mendis E Rajapakse N Byun H G amp Kim S K (2005) Investigation of jumbo squid
(Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects LifeScience 77 2166minus2178Mendis E Rajapakse N amp Kim S K (2005) Antioxidant properties of a radical-
scavenging peptide puri1047297ed from enzymatically prepared 1047297sh skin gelatinhydrolysate Journal of Agricultural and Food Chemistry 53(3) 581minus587
Messerli F H (1999) Hypertension and sudden cardiac death American Journal of Hypertension 12 181minus188
Nam K A You S G amp Kim S M (2008) Molecular and physical characteristicsof squid (Toradores paci 1047297cus) skin collagens and biological properties of theirenzymatic hydrolysates Journal of Food Science 73(4) 249minus255
PicotL Bordenave S Didelot S Fruitier-Arnaudin I Sannier F ThorkelssonG et al(2006) Antiproliferative activity of 1047297sh protein hydrolysates on human breastcancer cell lines Process Biochemistry 41 1217minus1222
Re R Pellegrini N Proteggente A Pannala A Yang M amp Rice-Evans C (1999)Antioxidant activity applying an improved ABTSradical cation decolorization assayFree Radical Biology amp Medicine 26 1231minus1237
Saiga A Tanabe S amp Nishimura T (2003) Antioxidant activity of peptides obtainedfrom porcine myo1047297brillar proteins by protease treatment Journal of Agriculturaland Food Chemistry 51(12) 3661minus3667
Samaranayaka A G P Kitts D D amp Li-Chan E C Y (2010) Antioxidative andangiotensin-I-converting enzyme inhibitory potential of a paci1047297c hake (Merluccius
productus) 1047297sh protein hydrolysate subjected to simulated gastrointestinaldigestion and caco-2 cell permeation Journal of Agricultural and Food Chemistry58(3) 1535minus1542
Schagger H amp Von Jagow G (1987) Tricine-sodium dodecyl sulphate polyacrylamidegel electrophoresis for the separation of proteins in the range from 1 to 100 kDa
Analytical Biochemistry 166 368minus379Sheih I Fang T J Wu T K amp LinP H (2010) Anticancer and antioxidant activities of
the peptide fraction from algae protein waste Journal of Agricultural and FoodChemistry 58 1202minus1207
Theodore A E amp Kristinsson H G (2007) Angiotensin converting enzyme inhibitionof 1047297sh protein hydrolysates prepared from alkaline-aided channel cat1047297sh proteinisolate Journal of the Science of Food and Agriculture 87 2353minus2357
Thiansilakul Y Benjakul S amp Shahidi F (2007) Compositions functional propertiesand antioxidative activity of protein hydrolysates prepared from round scad(Decapterus maruadsi) Food Chemistry 103(4) 1385minus1394
Witherow F N Helmy A Webb D J Fox K A A amp Newby D E (2001) Bradykinincontributes to the vasodilator effects of chronic angiotensin converting enzymeinhibition in patients with heart failure Circulation 104(18) 2177minus2181
Wu J Aluko R E amp Muir A D (2002) Improved method for direct high-performanceliquid chromatography assay of angiotensin-converting enzyme-catalyzed reactions
Journal of Chromatography A 950 125minus130Yang J L Ho H Y Chu Y J amp Chow C J (2008) Characteristic and antioxidant
activity of retorted gelatin hydrolysates from cobia (Rachycetron canadum) skinFood Chemistry 110 128minus136
Zhao Y Li B Liu Z Dong S Zhao X amp Zeng M (2007) Antihypertensive effect andpuri1047297cation of an ACE inhibitory peptide from sea cucumber gelatin hydrolysateProcess Biochemistry 42 1586minus1591
Zhu LChenJ Tang Xamp XiongY (2008) Reducingradical scavenging andchelationproperties of in vitro digests of Alcalase-treated zein hydrolysate Journal of
Agricultural and Food Chemistry 56 2714minus2721
1051 A A lemaacuten e t al Food Research International 44 (2011) 1044ndash1051

8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 38
added to each well and kept at 37 degC for 1 h Absorbance was read ona plate reader (VERSAmax Molecular Devices Saint Gregoire France)at 570 nm The data were analyzed to calculate the percentageof viability inhibition induced by the presence of hydrolysate incell culture medium determined by the equation Viability inhibition()=100minus(AHtimes100AC) where AH and AC are the absorbenciesmeasured for cell viability in culture medium containing onlyhydrolysate or PBS respectively
Following the same procedure the effect of addition of hydro-lysates at the same time as the cancer cells at the beginning of theexperiment was also evaluated For this evaluation the initialconditions of the culture medium and the added hydrolysates weremaintained for 72 h
The viability inhibition of the Esperase and Alcalase hydrolysateswas also measured using six different concentrations ranging from005 to 15 mgmL in order to determine the IC50 All experimentswere performed at least in triplicate
26 Antioxidant activities
Different assays (ABTS chelating ability and FRAP) were per-formed to study the antioxidant properties of the squid gelatin andthe corresponding hydrolysates at a concentration of 10 mgmL TheABTSradical [22prime-azino-bis-(3-ethylbenzothiazoline-6-sulfonic acid)]scavenging assay was carried out according to the method describedby Re Pellegrini Proteggente Pannala Yang and Rice-Evans (1999)The stock solution of ABTS radical consisted of 7 mM ABTS inpotassium persulfate 245 mM kept in the dark at room temperaturefor 16 h An aliquot of stock solution was diluted with Milli-Q waterin order to give the working solution of ABTS radical an absorbanceat 734 nm of 070plusmn002 A 20 μ L aliquot sample was mixed with980 μ L of ABTS reagent and then left to stand in the dark at 30 degCfor 10 min Absorbance values were read at 734 nm using a UV-1601spectrophotometer (Model CPS-240 Shimadzu Japan) Resultswere expressed as mg Vitamin C Equivalent Antioxidant Capacity(VCEAC)g of sample (gelatin or hydrolysate) based on a standardcurve of vitamin C
The Fe2+ chelating activity of the hydrolysates at a concentrationof 10 mgmL was measured by the method introduced by ChungChang Chao Lin andChou (2002) Brie1047298y a test sample of 800 μ Lwasmixed with 10 μ L of 2 mM FeCl2 and 20 μ L of 5 mM ferrozine Themixture was vortexed and kept at room temperature for 10 min priorto measuring the absorbance at 562 nm using a UV-1601 spectro-photometer (Model CPS-240 Shimadzu Japan) Chelating abilityexpressed as a percentage was calculated as follows Chelatingability ()=[1minus(Abs sample Abs control)]times 100 The control wasa mixture composed of 800 μ L of Milli-Q water 10 μ L of 2 mM FeCl2and 20 μ L of 5 mM ferrozine
FRAP a measure of the reducing power of samples was performedaccording to the method described by Benzie and Strain (1996) withsome modi1047297cations A 60 μ L of dissolved sample (at a concentration of
10 mgmL)was incubated at 37 degCwith 60μ L of Milli-Q waterand 900 μ L of FRAP reagent containing 10 mM of TPTZ (246-tripyridyl-s-triazine)and 20 mMof FeCl3 Absorbance valueswerereadat595 nmafter 30 minusing a UV-1601 spectrophotometer (Model CPS-240 Shimadzu Japan)Results were expressed as μ mol Fe2+ equivalentsg of sample (gelatin orhydrolysate) based on a standard curve of FeSO4middot7H2O All determina-tions were carried out at least in triplicate
27 Statistical analysis
Statistical tests were performed using the SPSSreg computerprogramme (SPSS Statistical Software Inc Chicago IL USA) Datawere expressed as meanplusmn standard deviation of the mean (n= 3 orn=24) Tukeys test was used to determine the level of signi1047297cance
at ple005
3 Results and discussion
31 Molecular weight distribution
Analysis of the overall molecular weight distribution of thedifferent hydrolysates (Fig 1) was carried out by Tricine SDS-PAGEThe Trypsin and NS37005 hydrolysates showed the highest quantityof peptide bands in the MW range between 26 and 35 kDa An
appreciable amount of protein with MW even higher than 26 kDawas also present in the Trypsin hydrolysates denoting the lowestef 1047297ciency in protein breakdown All the hydrolysates were character-ized by presenting an intense band at MWle14 kDa Howeverno peptide bands of more than 14 kDa could be observed in theAlcalase hydrolysates and hardly any in the Neutrase hydrolysateand Esperase hydrolysate indicating a considerably high degree of proteolysis Hydrolysates composed of a wide range of polydisperseproducts with varying degrees of hydrolysis and mostly small peptidescannot usually be properly detected under analytical electrophoresisconditions (Khantaphant amp Benjakul 2008 Gimeacutenez et al 2009)Consequently in order to know the differences in the molecularweight distribution of the smallest peptides (MWle14 kDa) size-exclusion HPLC (SE-HPLC) was performed in a fractionation rangebetween 7000 and 100 Da (Fig 2) The varying MW pro1047297lesrevealed considerable differences in the degree of protein breakdowndepending on the enzyme used In accordance with the electrophoreticpro1047297le the Alcalase Neutrase and Esperase hydrolysates showedhigher levels of peptides with MW below 1400 Da whereas in Trypsinand NS37005 hydrolysate peptides with MW above 1400 Da werepredominant
32 ACE-inhibitory capacity
The amount of hydrolysate required to inhibit 50 of the ACEactivity (IC50) is shown in Table 1 Unlike the parent squid gelatin allthe hydrolysates presented ACE inhibitory capacity although it wasmarkedly differentdepending on theenzyme used It wasnot possibleto determine the IC50 value in the NS37005 hydrolysate whereas the
Alcalase hydrolysate showed the most potent effect with an IC50
of 034 mgmL followed by the Neutrase hydrolysate (063 mgmL)(ple005) As revealed by SDS-PAGE and SE-HPLC the peptides inthe Alcalase and Neutrase hydrolysates were mostly between ~1400and 500 Da In another study Zhao et al (2007) found the highestACE-inhibitory capacity for a sea cucumber gelatin hydrolysate in thefraction with MWb1 kDa Nevertheless there are other studies where
Fig 1 Electrophoretic pro1047297les of gelatin hydrolysates at a concentration of 10 mgmL (Tricine SDS-PAGE) (m) molecular weight markers (a) Protamex (b) Trypsin
(c) Neutrase (d) Esperase (e) NS37005 (f) Savinase and (g) Alcalase
1046 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 48
very small peptides did not appear to increase the activity of thewhole hydrolysate but rather decreased it (Theodore amp Kristinsson2007)
Bougatef et al (2008) reported IC50 values in the concentrationrange of 12ndash74 mgmL for sardinelle by-product hydrolysatesprepared with different proteases while the IC50 of a marine shrimphydrolysate digested by a crude protease was 097 mgmL (Hai-lunXiu-Lan Cai-Yun Yu-Zhong amp Bai-Cheing 2006) Three peptides Val-Tyr-Ala-Pro Val-Ile-Ile-Phe and Met-Ala-Trp with IC50 values of 61 87and 1632 μ M respectively had been reported from cuttle1047297sh muscleprotein hydrolysates (Balti Nedjar-Arroume Bougatef Guillochon amp
Nasri 2010) Although values of IC50 for ACE-inhibitory capacity of skingelatin whole hydrolysates had been previously reported by Kim Kimet al (2001) and Zhao et al (2007) comparisons with IC50 valuesin thepresent study were not fully possible due to the different methodologyemployed for measuring activity
Proline or aromatic amino acids existing at theC-terminal are suitablefor peptide binding to ACE (Cheung Wang Miguel Emily amp Dvid 1980)Alcalase has broad speci1047297city hydrolyzing most peptide bonds preferen-tially those containing aromatic amino acid residues (Doucet OtterGauthier amp Foegeding 2003) suggesting that this enzyme preparationshould generate peptides with the most favorable amino acids at the C-
terminal position Some hydrolysates prepared withAlcalase also exerteda strong in vitro and in vivo ACE-inhibitory activity (Cheng et al 2008)Other studies demonstrated that Pro at the antepenultimate position inthe peptide sequence also enhanced binding to the ACE enzyme(Contreras Carroacuten Montero Ramos amp Recio 2009) Most commercialinhibitors (CaptoprilandEnalapril) also bear Proresidue intheirstructureThe reasonably high ACE-inhibitory capacity (IC50=034ndash106 mgmL)achieved with most of the hydrolysates tested may be favored by theabundanceof Proresidues(89residues1000 residues)intheparentsquidgelatin (Alemaacuten Gimeacutenez Montero et al 2011)
33 Cytotoxic and antiproliferative effects against cancer cells
Cytotoxic activity against cancer cells was considered to be the
effect produced when the hydrolysate was added to the culture
0 10 20 30 40 50 min0
250
500
750
mAUTrypsin
50 min0 10 20 30 400
250
500
750
mAUNeutrase
mAU
50 min0 10 20 30 400
250
500
750Esperase
mAU
0 10 20 30 40 50 min0
250
500
750NS37005
0 10 20 30 40 50 min0
250
500
750
mAUSavinase
0 10 20 30 40 50 min0
250
500
750
mAUAlcalase
0 10 20 30 40 50 min0
250
500
750
mAUProtamex
gt 1375 Da lt 500 Da1375-500 Da
Fig 2 Molecular weight distribution of gelatin hydrolysates at a concentration of 10 mgmL by SE-HPLC The absorbance was monitored at 215 nm
Table 1
ACE-inhibitory capacity of squid gelatin hydrolysates
IC50
Protamex 106 plusmn 001e
Trypsin 091 plusmn 003d
Neutrase 063 plusmn 001b
Esperase 089 plusmn 003cd
NS37005 ndSavinase 082 plusmn 008c
Alcalase 034 plusmn 002a
nd not determinedDifferent letters (a b chellip) indicate signi1047297cant differences(ple005) IC50 concentration (mgmL) required for inhibiting
50 of ACE activity
1047 A A lemaacuten e t al F ood Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 58
medium after the cells had been growing for 24 h antiproliferativeactivity was considered to be the effect produced when thehydrolysates were incorporated before cell growth started
Asexpectedsquidgelatin didnot show cytotoxicor antiproliferativeeffect on cancer cells However the hydrolysates prepared from squidgelatin affected the viability of both cell lines differently depending onthe enzyme used (Fig 3A B) Cell viability was determined after 24 48and 72 h with a change of culture medium and theaddition of 1 mgmL
concentration of the hydrolysate on each occasion Only EsperaseAlcalase and Savinase hydrolysates intensi1047297ed their effect on bothcancer cell lines after 48 and 72 h (ple005) The Esperase hydrolysatehad the highest (ple005) cytotoxic effect on both cancer cells (966plusmn05 and 912plusmn27 of viability inhibition on MCF-7 and U87 cellsrespectively after 72 h) It is worth noting that the same results wereobtained for a concentration of 03 mgmL for both cell lines Alcalasehydrolysate also showed relatively high cytotoxic activity (678plusmn56and 839plusmn48 viability inhibition on both cell lines respectively)
Jang Jo Kang and Lee (2008) puri1047297ed four peptides from beef proteinand caused a 20 viability inhibition on MCF-7 at 04 mgmL
Regarding antiproliferative activity again Esperase hydrolysatefollowed by Alcalase hydrolysate exhibited the highest (ple005)
activity on both cell lines (Fig 4) Picot et al (2006) evaluated theantiproliferative activity of 18 1047297sh protein hydrolysates and foundthe highest effect on MCF-7 cell lines to be around 40 for a codhydrolysate obtained with Protamex and Alcalase At the sameconcentration (1 mgmL) the Alcalase squid gelatin hydrolysatecaused a similar effect on MCF-7 cells (4164plusmn53 of growthinhibition) however the Esperase hydrolysate had a much stronger(ple005) effect (9141plusmn 18 of inhibition) and remained similar for
a concentration reduction to 05 mgmLThe IC50 was determined for both MCF-7 and U87 cells in the case
of hydrolysates that had produced the strongest cytotoxic effects(Esperase and Alcalase hydrolysates) at 24 48 and 72 h (Table 2)Both hydrolysates showed low IC50 values for both cell linesespecially the Esperase hydrolysate A pepsin hydrolysate fromalgae protein waste showed an IC50 of 174 mgmL against AGC celllines (Sheih Fang Wu amp Lin 2010) To our knowledge the IC50 valuesfor cytotoxiceffect (013 and 010 mgmL forMCF-7 andU87 cell linesrespectively) of the Esperase hydrolysate were the lowest found forwhole food protein hydrolysates although similar IC50 values havebeen reported for isolated peptides The nonapeptide X-Met-Leu-Pro-Ser-Tyr-Ser-Pro-Tyr obtained by hydrolysis of defatted soy protein
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
v i a b i l i t y i n h i b i t i o n ( )
24h 48h 72h
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
V i a b i l i t y i n h i b i t i o n ( )
24h 48h 72h
A
B
a x
c x
a b y
b x a
b x
b x
a y
a x
a x
b x
b y c
y
b x b
y d x
d x
e y e
z
e x
d y f z
a x a
b y
b y
a x
b y c
y
a x b
y b y
c x d
y e z
a x
a b y
b y
a x
b y c
z b x
c y d
z
Fig 3 Effect of gelatin hydrolysates on the viability of cancer cells for 24 48 and 72 h in cell culture medium containing a concentration of 1 mgmL of hydrolysate MCF-7 cell line(A) U87cellline (B) Differentletters (ab chellip) withinthe same time periodindicatesigni1047297cantdifferences (ple005)Different letters (xy z) withinthe same hydrolysate indicatesigni1047297cant differences (ple005)
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
G r o w t h i n h i b i t i o n ( )
U87 MCF-7
bd
c c
c
d
e
f
b ebc
aa
e
Fig 4 Effect of gelatin hydrolysates on growth of MCF-7 and U87human cells for 72 h in cell culture mediumcontaining a concentration of 1 mgmL of hydrolysate Different letters
(a b chellip) within the same cell cultures indicate signi1047297cant differences (ple005)
1048 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 68
hadanIC50 of 016 mgmL on a mouse monocyte macrophage cell line(Kim et al 2000) A peptide fraction from algae protein waste exertedan IC50 value of 007 mgmL against AGS cells (Sheih et al 2010) Thecytotoxic effect of Esperase hydrolysate on MCF-7 cells increasedafter 48 and 72 h but to a lesser extent than on U87 cells After 24 hthe IC50 for MCF-7 cells (015 mgmL) was lower than that for U87cells (031 mgmL) however after 72 h the Esperase hydrolysate wasmore cytotoxic against U87 cells (IC50=010 mgmL) than againstMCF-7 cells (IC50=013 mgmL)
The Esperase and Alcalase hydrolysates had most of their peptidesin the range of 1400 to 500 Da Sheih et al (2010) fractioned a pepsinhydrolysate from algae protein waste and found that the mosteffective peptide fraction against AGS cells had a molecular weightlower than 6500 Da and most of the peptides (N90) were locatedbetween the range of 2000 and 200 Da Kannan Hettiarachchy
Johnson and Nannapaneni (2008) observed the highest antiprolifera-tive activity in human colon and liver cancer cells in the lowermolecular weight fraction (b5 kDa) of an Alcalase hydrolysate fromrice bran However molecular weight cannot be considered the mostimportant factor in1047298uencing anticancer activity since the Neutrasehydrolysate which has relatively low cytotoxic and antiproliferativeactivities is also made up mostly of peptides in the range of 1400ndash
500 Da Thus the antiproliferativecytotoxic effect against MCF-7and U87 cells should be linked for the most part to the presenceof speci1047297c peptides that exert a direct cytotoxic effect on cancer
cells as previously reported for valorphin a hemoglobin-derivedpeptide (Blishchenko et al 2005) Nevertheless the hypothesis of abinding competition between peptides and FCS growth factor on cellmembrane receptors cannot be excluded either Three peptides froma 1047297sh source have been described as having antitumor activity (i) a4409 Da anchovy hydrophobic peptide able to induce apoptosis inhuman U937 lymphoma cells by increasing caspase-3 and caspase-8 activity (Lee Kim Lee Kim amp Lee 2004) (ii) tilapia hepcidin TH2-3that manifested signi1047297cant antitumor activity in human 1047297brosarcomacells (Chen Lin amp Lin 2009) and (iii) epinecidin-1 a peptide from1047297sh (Epinephelus coioides) which had an antitumor effect similar tolytic peptides in human 1047297brosarcoma cells (Lin et al 2009) Howevernone of these peptides originate from 1047297sh gelatincollagen
34 Antioxidant activity
The radical scavenging capacity of squid gelatin exhibited a 3-foldincrease when it was hydrolyzed with Protamex Neutrase orNS37005 whereas such an increase was between 7 and 10-foldwhen the enzymes used were Trypsin Savinase Esperase or Alcalase(Fig 5A) However radical scavenging capacity could not be wellcorrelated with the molecular weight distribution of the wholehydrolysate since Trypsin hydrolysate was the one with the highestnumber of larger peptides This 1047297nding suggested that the radicalscavenging activity of protein hydrolysates is dependent not onlyon the peptide size but also to a greater extent on their sequencesas a result of the speci1047297city of the enzyme used
The metal chelating ability of a given compound may also serve
as a useful indicator of its potential antioxidant activity As depicted
in Fig 5B the squid gelatin did not show any Fe2+ chelating ability at10 mgmL However when gelatin was hydrolyzed with ProtamexNeutrase or NS37005 enzymes the resulting hydrolysates did exert alow chelating capacity (less than 25) In contrast the TrypsinSavinase or Esperase hydrolysates at the same concentration caused~80 of Fe2+ chelation which was near to 100 with the Alcalasehydrolysate Dong Zeng Wang Liu Zhao and Huicheng(2008) founda metal chelation activity of 9297 with an Alcalase hydrolysate fromsilver carp at 5 mgmL
Hydroxyl radicals can be formed from superoxide anion andhydrogen peroxide in the presence of transition metal ions such asFe2+ andCu2+ Thus chelating metal ions may inhibit theformation of hydroxyl radicals Presumably peptide cleavage leads to enhancedmetal ion binding because carboxyl and amino groups in the sidechains of the acidic (Glx and Asx) and basic (Lys His and Arg) aminoacids are thought to play an important role in chelating metal ions(Saiga Tanabe amp Nishimura 2003)
The molecular weight of peptidesis also believed to play a key rolein chelating metals Klompong Benjakul Kantachote and Shaidi(2007)and Dong et al (2008) pointed out that metal-chelating abilityand molecular weight had a linear relationship and that metal-chelating ability increased with decreasing molecular weight Never-theless Megiacuteas et al (2007) reported that if peptide length is tooshort chelation is unstable In our study hydrolysate with peptidesmostly between ~1400 and 500 Da (Alcalase hydrolysate) exerted
the highest metal chelating ability However it is worth noting theconsiderably high metal chelating activity in the Trypsin hydrolysateas compared to other hydrolysates with higher proportions of smallerpeptides (Neutraseor Protamex) denoting onceagainthe importanceof the presence of speci1047297c peptide sequences rather than the wholehydrolysate molecular weight distribution
The behavior for Fe2+ chelation and ABTS+ scavenging followed asimilar trend suggesting that the two antioxidant mechanisms mightbe related Zhu Chen Tang and Xiong (2008) also found a similartrend between Cu2+ chelating capacity and ABTS radical scavengingfor an Alcalase hydrolysate from zein protein
Another method for measuring antioxidant potential is theferric reducing antioxidant power (FRAP) assay Reducing power isa measurement that provides an estimation of a compounds ability
to reduce ferric iron (III) to ferrous iron (II) and is determinedusing a redox-linked colorimetric reaction Samples with higherreducing power are more able to donate electrons or hydrogenFree radicals form stable substances by accepting donated electronsand thus the free radical chain reaction could be interruptedThe reducing power of squid gelatin and hydrolysates is shownin Fig 5C The ability of the hydrolysates to reduce Fe3+ to Fe2+
was approximately twice than that of the gelatin with all thehydrolysates showing a similar (pN005) capacity independently of the enzyme used and consequently of molecular weight distribu-tion too In several studies involving protein hydrolysates someresearchers reported that the reducing capacity improved as thedegree of protein hydrolysis increased (Thiansilakul Benjakulamp Shahidi 2007) while some researchers reported the opposite
(Klompong et al 2007)
Table 2
Cytotoxic activity of Esperase and Alcalase squid gelatin hydrolysates
IC50
MCF-7 cells U87 cells
24 h 48 h 72 h 24 h 48 h 72 h
Esperase 015 plusmn001a 014plusmn001a 013plusmn001a 031plusmn001c 013plusmn001b 010plusmn001a
Alcalase 118 plusmn002c 104plusmn002b 081plusmn003a 122plusmn003c 098plusmn003b 085plusmn002a
Different letters (a b) within the same hydrolysate and the same cell line indicate signi1047297cant differences (ple005) IC50 concentration (mgmL) required to induce 50 cells death
1049 A A lemaacuten e t al F ood Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 78
Alcalase Esperase Savinase and Trypsin signi1047297cantly improvedthe antioxidant activity of the resulting hydrolysates via the enhance-ment of antiradical activity and metal chelation Reducing powerwas also increased by the action of these enzymes but the degreeof enhancement was much lower Consistent with our results thehydrolysis of snapper gelatin with proteases from pyloric caecacaused a 55 to 65-fold increase in ABTS radical scavenging capacitywhereas ferric reducing power did not increase more than 2-fold(Khantaphant amp Benjakul 2008) Free radical quenchingwas reportedto be the main antioxidative mechanism of some peptides derivedfrom squid gelatin hydrolysates (Mendis Rajapakse amp Kim 2005)
4 Conclusions
The Esperase hydrolysate exerted a potent cytotoxic and anti-proliferative effects on the two cancer cell lines studied withsigni1047297cant antioxidant activity especially with respect to metalchelating and antiradical activities The Alcalase hydrolysate was themost potent ACE-inhibitor and metal chelator while at the same timeexerting strong radical scavenging and cytotoxic activity againstcancer cells These results suggest that squid gelatin hydrolysatesprepared with Esperase or Alcalase enzymes could be utilized as newmaterials in the development of functional foods with potentialanticancer antihypertensive and antioxidant capacities
Acknowledgments
Funding for this research was provided by (i) the Interministerial
Scienti1047297c and Technical Commission of Spains Ministry of Science
and Innovation under project AGL2008-02135ALI and (ii) Xunta deGalicia Program Aacutengeles Alvarintildeo (European Social Fund)
References
AlemaacutenA Gimeacutenez BMonteroP amp Goacutemez-GuilleacutenM C(2011) Antioxidantactivityof several marine skin gelatins LWT Food Science and Technology 44 407minus413
Alemaacuten A Gimeacutenez B Peacuterez-Santiacuten E Goacutemez-Guilleacuten M C amp Montero P (2011)Contribution of Leu and Hyp residues to antioxidant and ACE-inhibitory activitiesof peptide sequences isolated from squid gelatin hydrolysates Food Chemistry 125334minus341
Balti R Nedjar-Arroume N Bougatef A Guillochon D amp Nasri M (2010) Threenovel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttle1047297sh(Sepia of 1047297cinalis) using digestive proteases Food Research International 43(4)1136minus1143
BenzieI F Famp StrainJ J(1996) Theferric reducingabilityof plasma(FRAP) asa measureof ldquoantioxidant powerrdquo The FRAP assay Analytical Biochemistry 239(1) 70minus76
Blishchenko E Y Sazonova O V Kalinina O A Moiseeva E V Vass A A KarelinA A et al (2005) Antitumor effect of valorphin in vitro and in vivo Combinedaction with cytostatic drugs Cancer Biology amp Therapy 4(1) 118minus124
Bougatef A Nedjar-Arroume N Ravallec-Pleacute R Leroy Y Guillochon D Barkia Aet al (2008) Angiotensin I-converting enzyme (ACE) inhibitory activities of sardinelle (Sardinella aurita) by-products protein hydrolysates obtained bytreatment with microbial and visceral 1047297sh serine-proteases Food Chemistry 111350minus356
Chen J Y Lin W J amp Lin T L (2009) A 1047297sh antimicrobial peptide tilapia hepcidinTH2-3 showspotent antitumor activity againsthuman 1047297brosarcomacells Peptides
30 1636minus1642ChengFYWanTCLiuYTLaiKMLinLCampSakataR(2008)Astudyof invivo
antihypertensive properties of enzymatic hydrolysate from chicken leg boneprotein Animal Science Journal 79 614minus619
Cheung H S Wang F L Miguel A O Emily F S amp Dvid W C (1980) Binding of peptide substrates and inhibitors of angiotensin-I-converting enzyme The Journalof Biological Chemistry 255 401minus407
Chung Y C Chang C T Chao W W Lin C F amp Chou S T (2002) Antioxidativeactivity and safety of the 50 ethanolic extract from red bean fermented by Bacillus
subtilis IMR-NK1 Journal of Agricultural and Food Chemistry 50 2454minus2458
0
20
40
60
80
100
C h e l a t i n g a b i l i t y ( )
0
5
10
15
20
micro m o l e s F e E q g
A
a
b
c
d
bb
d d
B
bc
d
b
d
c
de
a
C
a
bbbbb bb
0
10
20
30
40
50
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
V C E A C ( m g g )
Fig 5 Antioxidant activity of gelatin hydrolysates at a concentration of 10 mgmL ABTS radical scavenging capacity (A) Fe chelating ability (B) and Fe reducing power (FRAP) (C)Different letters (a b chellip) indicate signi1047297cant differences (ple005)
1050 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 88
Contreras M Carroacuten R Montero M J Ramos M amp Recio I (2009) Novel casein-derived peptides with antihypertensive activity International Dairy Journal 19566minus573
Dong S Zeng M Wang D Liu Z Zhao Y amp Huicheng Y (2008) Antioxidantand biochemical properties of protein hydrolysates prepared from silver carp(Hypophthalmichthys molitrix) Food Chemistry 107 1485minus1493
Doucet D Otter E O Gauthier S F amp Foegeding E A (2003) Enzyme-inducedgelation of extensively hydrolyzed whey protein by Alcalase Peptides identi1047297ca-tion and determination of enzyme speci1047297city Journal of Agricultural and FoodChemistry 51 6300minus6308
Gimeacutenez B Alemaacuten A Montero P amp Goacutemez-Guilleacuten M C (2009) Antioxidant and
functional properties of gelatin hydrolysates obtained from skin of sole and squidFood Chemistry 114(3) 976minus983Goacutemez-Guilleacuten MC Loacutepez-Caballero ME Alemaacuten ALoacutepezde LaceyA Gimeacutenez B
amp Montero P (2010) Antioxidant and antimicrobial peptide fractions from squidandtunaskin gelatin In E LeBihan(Ed) Seaby-productsas real materialNew waysof application (pp 89minus115) Kerala India Transworld Research Network
Goodfriend T L Elliott M E amp Catt K J (1996) Drug therapy ndash Angiotensin receptorsand their antagonists The New England Journal of Medicine 334(25) 1649minus1654
Gueacuterard F Dufosseacute L De La Broise D amp Binet A (2001) Enzymatic hydrolysis of proteins from yellow1047297n tuna (Thunnus albacares) wastes using Alcalase Journal of Molecular Catalysis B Enzymatic 11 1051minus1059
Hai-Lun H Xiu-Lan C Sai-Yun S Yu-Zhonga Z amp Bai-Chenga Z (2006) Analysisof novel angiotensin-I-converting enzyme inhibitory peptides from protease-hydrolyzed marine shrimp Acetes chinensis Journal of Peptide Science 12 726minus733
Hou W C Chen H J amp Lin Y H (2003) Antioxidant peptides with angiotensinconverting enzymeinhibitory activitiesand applications for angiotensin convertingenzyme puri1047297cation Journal of Agricultural and Food Chemistry 51 1706minus1709
Jang A Jo C Kang K S amp Lee M (2008) Antimicrobial and human cancer cellcytotoxic effect of synthetic angiotensin-converting enzyme (ACE) inhibitory
peptides Food Chemistry 107 327minus336 Je J -Y Lee K -H Lee M H amp Ahn C -B (2009) Antioxidant and antihypertensive
protein hydrolysates produced from tuna liver by enzymatic hydrolysis FoodResearch International 42(9) 1266minus1272
Kannan A Hettiarachchy N Johnson M G amp Nannapaneni R (2008) Human colonand liver cancer cell proliferation inhibition by peptide hydrolysates derived fromheat-stabilized defatted rice bran Journal of Agriculture and Food Chemistry 56 11643minus11647
Kaufman S H amp Earnshaw W C (2000) Induction of apoptosis by cancerchemotherapy Cell Research 256 42minus49
Khantaphant S amp Benjakul S (2008) Comparative study on the proteases from 1047297shpyloric caeca and the use for production of gelatin hydrolysate with antioxidativeactivity Comparative Biochemistry and Physiology 151 410minus419
Kim S K Byun H G Park P J amp Shahidi F (2001) Angiotensin I converting enzymeinhibitory peptides puri1047297ed form bovine skin gelatin hydrolysate Journal of
Agricultural and Food Chemistry 49 2992minus2997Kim S K Kim Y T Byun H G Nam K S Joo D S amp Shahidi F (2001) Isolation and
characterization of antioxidative peptides from gelatin hydrolysate of Alaskapollack skin Journal of Agricultural and Food Chemistry 49 1984minus1989
Kim S E Kim H H Kim J Y Kang Y I Wooa H J amp Lee H J (2000) Anticanceractivity of hydrophobic peptides from soy proteins Biofactor 21 63minus67
Klompong V Benjakul S Kantachote D amp Shahidi F (2007) Antioxidative activityand functional properties of protein hydrolysate of yellow stripe trevally(Selaroides leptolepis) as in1047298uenced by the degree of hydrolysis and enzyme typeFood Chemistry 102 1317minus1327
LeeY G Kim J Y Lee KW Kim K H amp Lee H J (2004) Peptides from anchovy sauceinduce apoptosisin a human lymphoma cell(U937) throughthe increaseof caspase-3and -8 activities Annals of the New York Academy of Sciences 1010 399minus404
Leng B Liu X D amp Chen Q X (2005) Inhibitory effects of anticancer peptide fromMercenaria on the BGC-823 cells and several enzymes FEBS 579 1187minus1190
Lin W J Chien Y L Pan C Y Lin T L Chen J Y Chiu S J amp Hui C F (2009)Epinecidin-1 an antimicrobial peptide from 1047297sh (Epinephelus coioides) which
hasan antitumoreffect like lytic peptides in human 1047297brosarcoma cells Peptides 30283minus290
Lin Lamp LiB (2006) Radical scavengingproperties of protein hydrolysates from jumbo1047298ying squid (Dosidicus eschrichitii Steenstrup) skin gelatin Journal of the Science of Food and Agriculture 86 2290minus2295
Lind D S Hochwald S N Malaty J Rekkas S Hebig P Mishra G et al (2001)Nuclear factor-kB is upregulated in colorectal cancer Surgery 130 363minus369
Megiacuteas C Pedroche J Yust M M Giroacuten-Calle J Alaiz M Millaacuten F amp Vioque J(2007) Af 1047297nity puri1047297cation of copper chelating peptides from chickpea proteinhydrolysates Journal of Agricultural and Food Chemistry 55 3949minus3954
Mendis E Rajapakse N Byun H G amp Kim S K (2005) Investigation of jumbo squid
(Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects LifeScience 77 2166minus2178Mendis E Rajapakse N amp Kim S K (2005) Antioxidant properties of a radical-
scavenging peptide puri1047297ed from enzymatically prepared 1047297sh skin gelatinhydrolysate Journal of Agricultural and Food Chemistry 53(3) 581minus587
Messerli F H (1999) Hypertension and sudden cardiac death American Journal of Hypertension 12 181minus188
Nam K A You S G amp Kim S M (2008) Molecular and physical characteristicsof squid (Toradores paci 1047297cus) skin collagens and biological properties of theirenzymatic hydrolysates Journal of Food Science 73(4) 249minus255
PicotL Bordenave S Didelot S Fruitier-Arnaudin I Sannier F ThorkelssonG et al(2006) Antiproliferative activity of 1047297sh protein hydrolysates on human breastcancer cell lines Process Biochemistry 41 1217minus1222
Re R Pellegrini N Proteggente A Pannala A Yang M amp Rice-Evans C (1999)Antioxidant activity applying an improved ABTSradical cation decolorization assayFree Radical Biology amp Medicine 26 1231minus1237
Saiga A Tanabe S amp Nishimura T (2003) Antioxidant activity of peptides obtainedfrom porcine myo1047297brillar proteins by protease treatment Journal of Agriculturaland Food Chemistry 51(12) 3661minus3667
Samaranayaka A G P Kitts D D amp Li-Chan E C Y (2010) Antioxidative andangiotensin-I-converting enzyme inhibitory potential of a paci1047297c hake (Merluccius
productus) 1047297sh protein hydrolysate subjected to simulated gastrointestinaldigestion and caco-2 cell permeation Journal of Agricultural and Food Chemistry58(3) 1535minus1542
Schagger H amp Von Jagow G (1987) Tricine-sodium dodecyl sulphate polyacrylamidegel electrophoresis for the separation of proteins in the range from 1 to 100 kDa
Analytical Biochemistry 166 368minus379Sheih I Fang T J Wu T K amp LinP H (2010) Anticancer and antioxidant activities of
the peptide fraction from algae protein waste Journal of Agricultural and FoodChemistry 58 1202minus1207
Theodore A E amp Kristinsson H G (2007) Angiotensin converting enzyme inhibitionof 1047297sh protein hydrolysates prepared from alkaline-aided channel cat1047297sh proteinisolate Journal of the Science of Food and Agriculture 87 2353minus2357
Thiansilakul Y Benjakul S amp Shahidi F (2007) Compositions functional propertiesand antioxidative activity of protein hydrolysates prepared from round scad(Decapterus maruadsi) Food Chemistry 103(4) 1385minus1394
Witherow F N Helmy A Webb D J Fox K A A amp Newby D E (2001) Bradykinincontributes to the vasodilator effects of chronic angiotensin converting enzymeinhibition in patients with heart failure Circulation 104(18) 2177minus2181
Wu J Aluko R E amp Muir A D (2002) Improved method for direct high-performanceliquid chromatography assay of angiotensin-converting enzyme-catalyzed reactions
Journal of Chromatography A 950 125minus130Yang J L Ho H Y Chu Y J amp Chow C J (2008) Characteristic and antioxidant
activity of retorted gelatin hydrolysates from cobia (Rachycetron canadum) skinFood Chemistry 110 128minus136
Zhao Y Li B Liu Z Dong S Zhao X amp Zeng M (2007) Antihypertensive effect andpuri1047297cation of an ACE inhibitory peptide from sea cucumber gelatin hydrolysateProcess Biochemistry 42 1586minus1591
Zhu LChenJ Tang Xamp XiongY (2008) Reducingradical scavenging andchelationproperties of in vitro digests of Alcalase-treated zein hydrolysate Journal of
Agricultural and Food Chemistry 56 2714minus2721
1051 A A lemaacuten e t al Food Research International 44 (2011) 1044ndash1051
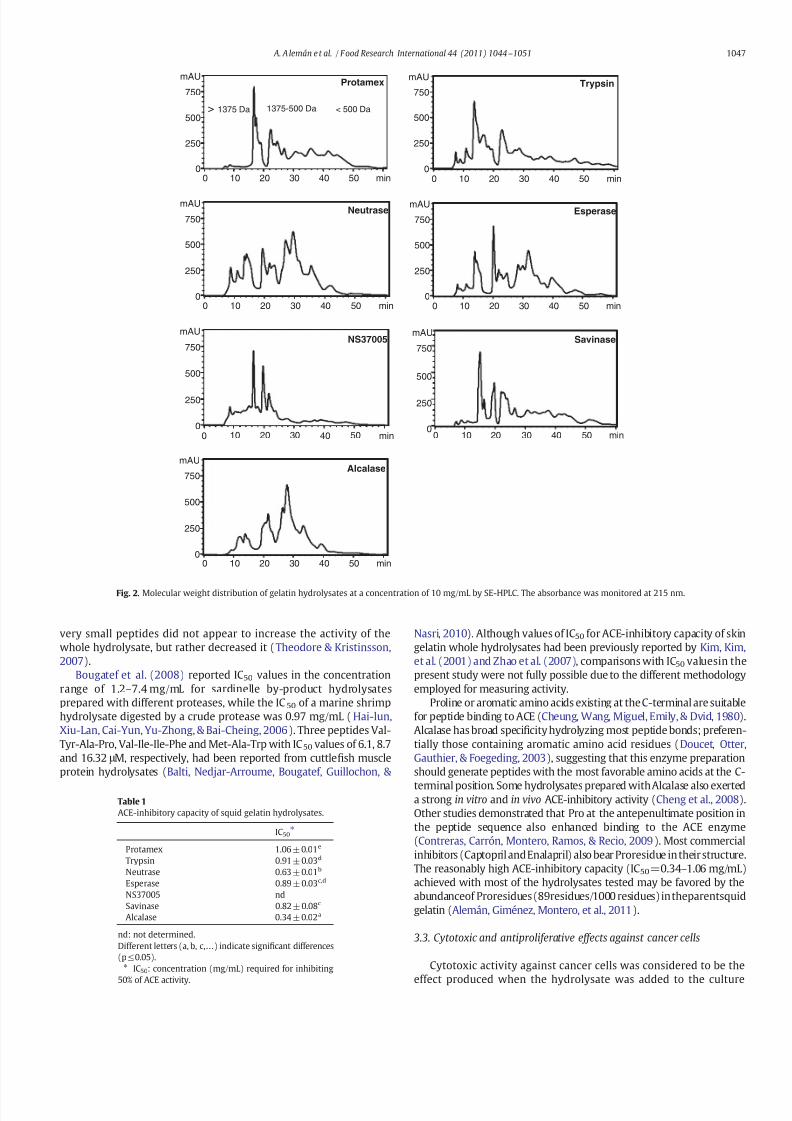
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 48
very small peptides did not appear to increase the activity of thewhole hydrolysate but rather decreased it (Theodore amp Kristinsson2007)
Bougatef et al (2008) reported IC50 values in the concentrationrange of 12ndash74 mgmL for sardinelle by-product hydrolysatesprepared with different proteases while the IC50 of a marine shrimphydrolysate digested by a crude protease was 097 mgmL (Hai-lunXiu-Lan Cai-Yun Yu-Zhong amp Bai-Cheing 2006) Three peptides Val-Tyr-Ala-Pro Val-Ile-Ile-Phe and Met-Ala-Trp with IC50 values of 61 87and 1632 μ M respectively had been reported from cuttle1047297sh muscleprotein hydrolysates (Balti Nedjar-Arroume Bougatef Guillochon amp
Nasri 2010) Although values of IC50 for ACE-inhibitory capacity of skingelatin whole hydrolysates had been previously reported by Kim Kimet al (2001) and Zhao et al (2007) comparisons with IC50 valuesin thepresent study were not fully possible due to the different methodologyemployed for measuring activity
Proline or aromatic amino acids existing at theC-terminal are suitablefor peptide binding to ACE (Cheung Wang Miguel Emily amp Dvid 1980)Alcalase has broad speci1047297city hydrolyzing most peptide bonds preferen-tially those containing aromatic amino acid residues (Doucet OtterGauthier amp Foegeding 2003) suggesting that this enzyme preparationshould generate peptides with the most favorable amino acids at the C-
terminal position Some hydrolysates prepared withAlcalase also exerteda strong in vitro and in vivo ACE-inhibitory activity (Cheng et al 2008)Other studies demonstrated that Pro at the antepenultimate position inthe peptide sequence also enhanced binding to the ACE enzyme(Contreras Carroacuten Montero Ramos amp Recio 2009) Most commercialinhibitors (CaptoprilandEnalapril) also bear Proresidue intheirstructureThe reasonably high ACE-inhibitory capacity (IC50=034ndash106 mgmL)achieved with most of the hydrolysates tested may be favored by theabundanceof Proresidues(89residues1000 residues)intheparentsquidgelatin (Alemaacuten Gimeacutenez Montero et al 2011)
33 Cytotoxic and antiproliferative effects against cancer cells
Cytotoxic activity against cancer cells was considered to be the
effect produced when the hydrolysate was added to the culture
0 10 20 30 40 50 min0
250
500
750
mAUTrypsin
50 min0 10 20 30 400
250
500
750
mAUNeutrase
mAU
50 min0 10 20 30 400
250
500
750Esperase
mAU
0 10 20 30 40 50 min0
250
500
750NS37005
0 10 20 30 40 50 min0
250
500
750
mAUSavinase
0 10 20 30 40 50 min0
250
500
750
mAUAlcalase
0 10 20 30 40 50 min0
250
500
750
mAUProtamex
gt 1375 Da lt 500 Da1375-500 Da
Fig 2 Molecular weight distribution of gelatin hydrolysates at a concentration of 10 mgmL by SE-HPLC The absorbance was monitored at 215 nm
Table 1
ACE-inhibitory capacity of squid gelatin hydrolysates
IC50
Protamex 106 plusmn 001e
Trypsin 091 plusmn 003d
Neutrase 063 plusmn 001b
Esperase 089 plusmn 003cd
NS37005 ndSavinase 082 plusmn 008c
Alcalase 034 plusmn 002a
nd not determinedDifferent letters (a b chellip) indicate signi1047297cant differences(ple005) IC50 concentration (mgmL) required for inhibiting
50 of ACE activity
1047 A A lemaacuten e t al F ood Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 58
medium after the cells had been growing for 24 h antiproliferativeactivity was considered to be the effect produced when thehydrolysates were incorporated before cell growth started
Asexpectedsquidgelatin didnot show cytotoxicor antiproliferativeeffect on cancer cells However the hydrolysates prepared from squidgelatin affected the viability of both cell lines differently depending onthe enzyme used (Fig 3A B) Cell viability was determined after 24 48and 72 h with a change of culture medium and theaddition of 1 mgmL
concentration of the hydrolysate on each occasion Only EsperaseAlcalase and Savinase hydrolysates intensi1047297ed their effect on bothcancer cell lines after 48 and 72 h (ple005) The Esperase hydrolysatehad the highest (ple005) cytotoxic effect on both cancer cells (966plusmn05 and 912plusmn27 of viability inhibition on MCF-7 and U87 cellsrespectively after 72 h) It is worth noting that the same results wereobtained for a concentration of 03 mgmL for both cell lines Alcalasehydrolysate also showed relatively high cytotoxic activity (678plusmn56and 839plusmn48 viability inhibition on both cell lines respectively)
Jang Jo Kang and Lee (2008) puri1047297ed four peptides from beef proteinand caused a 20 viability inhibition on MCF-7 at 04 mgmL
Regarding antiproliferative activity again Esperase hydrolysatefollowed by Alcalase hydrolysate exhibited the highest (ple005)
activity on both cell lines (Fig 4) Picot et al (2006) evaluated theantiproliferative activity of 18 1047297sh protein hydrolysates and foundthe highest effect on MCF-7 cell lines to be around 40 for a codhydrolysate obtained with Protamex and Alcalase At the sameconcentration (1 mgmL) the Alcalase squid gelatin hydrolysatecaused a similar effect on MCF-7 cells (4164plusmn53 of growthinhibition) however the Esperase hydrolysate had a much stronger(ple005) effect (9141plusmn 18 of inhibition) and remained similar for
a concentration reduction to 05 mgmLThe IC50 was determined for both MCF-7 and U87 cells in the case
of hydrolysates that had produced the strongest cytotoxic effects(Esperase and Alcalase hydrolysates) at 24 48 and 72 h (Table 2)Both hydrolysates showed low IC50 values for both cell linesespecially the Esperase hydrolysate A pepsin hydrolysate fromalgae protein waste showed an IC50 of 174 mgmL against AGC celllines (Sheih Fang Wu amp Lin 2010) To our knowledge the IC50 valuesfor cytotoxiceffect (013 and 010 mgmL forMCF-7 andU87 cell linesrespectively) of the Esperase hydrolysate were the lowest found forwhole food protein hydrolysates although similar IC50 values havebeen reported for isolated peptides The nonapeptide X-Met-Leu-Pro-Ser-Tyr-Ser-Pro-Tyr obtained by hydrolysis of defatted soy protein
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
v i a b i l i t y i n h i b i t i o n ( )
24h 48h 72h
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
V i a b i l i t y i n h i b i t i o n ( )
24h 48h 72h
A
B
a x
c x
a b y
b x a
b x
b x
a y
a x
a x
b x
b y c
y
b x b
y d x
d x
e y e
z
e x
d y f z
a x a
b y
b y
a x
b y c
y
a x b
y b y
c x d
y e z
a x
a b y
b y
a x
b y c
z b x
c y d
z
Fig 3 Effect of gelatin hydrolysates on the viability of cancer cells for 24 48 and 72 h in cell culture medium containing a concentration of 1 mgmL of hydrolysate MCF-7 cell line(A) U87cellline (B) Differentletters (ab chellip) withinthe same time periodindicatesigni1047297cantdifferences (ple005)Different letters (xy z) withinthe same hydrolysate indicatesigni1047297cant differences (ple005)
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
G r o w t h i n h i b i t i o n ( )
U87 MCF-7
bd
c c
c
d
e
f
b ebc
aa
e
Fig 4 Effect of gelatin hydrolysates on growth of MCF-7 and U87human cells for 72 h in cell culture mediumcontaining a concentration of 1 mgmL of hydrolysate Different letters
(a b chellip) within the same cell cultures indicate signi1047297cant differences (ple005)
1048 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 68
hadanIC50 of 016 mgmL on a mouse monocyte macrophage cell line(Kim et al 2000) A peptide fraction from algae protein waste exertedan IC50 value of 007 mgmL against AGS cells (Sheih et al 2010) Thecytotoxic effect of Esperase hydrolysate on MCF-7 cells increasedafter 48 and 72 h but to a lesser extent than on U87 cells After 24 hthe IC50 for MCF-7 cells (015 mgmL) was lower than that for U87cells (031 mgmL) however after 72 h the Esperase hydrolysate wasmore cytotoxic against U87 cells (IC50=010 mgmL) than againstMCF-7 cells (IC50=013 mgmL)
The Esperase and Alcalase hydrolysates had most of their peptidesin the range of 1400 to 500 Da Sheih et al (2010) fractioned a pepsinhydrolysate from algae protein waste and found that the mosteffective peptide fraction against AGS cells had a molecular weightlower than 6500 Da and most of the peptides (N90) were locatedbetween the range of 2000 and 200 Da Kannan Hettiarachchy
Johnson and Nannapaneni (2008) observed the highest antiprolifera-tive activity in human colon and liver cancer cells in the lowermolecular weight fraction (b5 kDa) of an Alcalase hydrolysate fromrice bran However molecular weight cannot be considered the mostimportant factor in1047298uencing anticancer activity since the Neutrasehydrolysate which has relatively low cytotoxic and antiproliferativeactivities is also made up mostly of peptides in the range of 1400ndash
500 Da Thus the antiproliferativecytotoxic effect against MCF-7and U87 cells should be linked for the most part to the presenceof speci1047297c peptides that exert a direct cytotoxic effect on cancer
cells as previously reported for valorphin a hemoglobin-derivedpeptide (Blishchenko et al 2005) Nevertheless the hypothesis of abinding competition between peptides and FCS growth factor on cellmembrane receptors cannot be excluded either Three peptides froma 1047297sh source have been described as having antitumor activity (i) a4409 Da anchovy hydrophobic peptide able to induce apoptosis inhuman U937 lymphoma cells by increasing caspase-3 and caspase-8 activity (Lee Kim Lee Kim amp Lee 2004) (ii) tilapia hepcidin TH2-3that manifested signi1047297cant antitumor activity in human 1047297brosarcomacells (Chen Lin amp Lin 2009) and (iii) epinecidin-1 a peptide from1047297sh (Epinephelus coioides) which had an antitumor effect similar tolytic peptides in human 1047297brosarcoma cells (Lin et al 2009) Howevernone of these peptides originate from 1047297sh gelatincollagen
34 Antioxidant activity
The radical scavenging capacity of squid gelatin exhibited a 3-foldincrease when it was hydrolyzed with Protamex Neutrase orNS37005 whereas such an increase was between 7 and 10-foldwhen the enzymes used were Trypsin Savinase Esperase or Alcalase(Fig 5A) However radical scavenging capacity could not be wellcorrelated with the molecular weight distribution of the wholehydrolysate since Trypsin hydrolysate was the one with the highestnumber of larger peptides This 1047297nding suggested that the radicalscavenging activity of protein hydrolysates is dependent not onlyon the peptide size but also to a greater extent on their sequencesas a result of the speci1047297city of the enzyme used
The metal chelating ability of a given compound may also serve
as a useful indicator of its potential antioxidant activity As depicted
in Fig 5B the squid gelatin did not show any Fe2+ chelating ability at10 mgmL However when gelatin was hydrolyzed with ProtamexNeutrase or NS37005 enzymes the resulting hydrolysates did exert alow chelating capacity (less than 25) In contrast the TrypsinSavinase or Esperase hydrolysates at the same concentration caused~80 of Fe2+ chelation which was near to 100 with the Alcalasehydrolysate Dong Zeng Wang Liu Zhao and Huicheng(2008) founda metal chelation activity of 9297 with an Alcalase hydrolysate fromsilver carp at 5 mgmL
Hydroxyl radicals can be formed from superoxide anion andhydrogen peroxide in the presence of transition metal ions such asFe2+ andCu2+ Thus chelating metal ions may inhibit theformation of hydroxyl radicals Presumably peptide cleavage leads to enhancedmetal ion binding because carboxyl and amino groups in the sidechains of the acidic (Glx and Asx) and basic (Lys His and Arg) aminoacids are thought to play an important role in chelating metal ions(Saiga Tanabe amp Nishimura 2003)
The molecular weight of peptidesis also believed to play a key rolein chelating metals Klompong Benjakul Kantachote and Shaidi(2007)and Dong et al (2008) pointed out that metal-chelating abilityand molecular weight had a linear relationship and that metal-chelating ability increased with decreasing molecular weight Never-theless Megiacuteas et al (2007) reported that if peptide length is tooshort chelation is unstable In our study hydrolysate with peptidesmostly between ~1400 and 500 Da (Alcalase hydrolysate) exerted
the highest metal chelating ability However it is worth noting theconsiderably high metal chelating activity in the Trypsin hydrolysateas compared to other hydrolysates with higher proportions of smallerpeptides (Neutraseor Protamex) denoting onceagainthe importanceof the presence of speci1047297c peptide sequences rather than the wholehydrolysate molecular weight distribution
The behavior for Fe2+ chelation and ABTS+ scavenging followed asimilar trend suggesting that the two antioxidant mechanisms mightbe related Zhu Chen Tang and Xiong (2008) also found a similartrend between Cu2+ chelating capacity and ABTS radical scavengingfor an Alcalase hydrolysate from zein protein
Another method for measuring antioxidant potential is theferric reducing antioxidant power (FRAP) assay Reducing power isa measurement that provides an estimation of a compounds ability
to reduce ferric iron (III) to ferrous iron (II) and is determinedusing a redox-linked colorimetric reaction Samples with higherreducing power are more able to donate electrons or hydrogenFree radicals form stable substances by accepting donated electronsand thus the free radical chain reaction could be interruptedThe reducing power of squid gelatin and hydrolysates is shownin Fig 5C The ability of the hydrolysates to reduce Fe3+ to Fe2+
was approximately twice than that of the gelatin with all thehydrolysates showing a similar (pN005) capacity independently of the enzyme used and consequently of molecular weight distribu-tion too In several studies involving protein hydrolysates someresearchers reported that the reducing capacity improved as thedegree of protein hydrolysis increased (Thiansilakul Benjakulamp Shahidi 2007) while some researchers reported the opposite
(Klompong et al 2007)
Table 2
Cytotoxic activity of Esperase and Alcalase squid gelatin hydrolysates
IC50
MCF-7 cells U87 cells
24 h 48 h 72 h 24 h 48 h 72 h
Esperase 015 plusmn001a 014plusmn001a 013plusmn001a 031plusmn001c 013plusmn001b 010plusmn001a
Alcalase 118 plusmn002c 104plusmn002b 081plusmn003a 122plusmn003c 098plusmn003b 085plusmn002a
Different letters (a b) within the same hydrolysate and the same cell line indicate signi1047297cant differences (ple005) IC50 concentration (mgmL) required to induce 50 cells death
1049 A A lemaacuten e t al F ood Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 78
Alcalase Esperase Savinase and Trypsin signi1047297cantly improvedthe antioxidant activity of the resulting hydrolysates via the enhance-ment of antiradical activity and metal chelation Reducing powerwas also increased by the action of these enzymes but the degreeof enhancement was much lower Consistent with our results thehydrolysis of snapper gelatin with proteases from pyloric caecacaused a 55 to 65-fold increase in ABTS radical scavenging capacitywhereas ferric reducing power did not increase more than 2-fold(Khantaphant amp Benjakul 2008) Free radical quenchingwas reportedto be the main antioxidative mechanism of some peptides derivedfrom squid gelatin hydrolysates (Mendis Rajapakse amp Kim 2005)
4 Conclusions
The Esperase hydrolysate exerted a potent cytotoxic and anti-proliferative effects on the two cancer cell lines studied withsigni1047297cant antioxidant activity especially with respect to metalchelating and antiradical activities The Alcalase hydrolysate was themost potent ACE-inhibitor and metal chelator while at the same timeexerting strong radical scavenging and cytotoxic activity againstcancer cells These results suggest that squid gelatin hydrolysatesprepared with Esperase or Alcalase enzymes could be utilized as newmaterials in the development of functional foods with potentialanticancer antihypertensive and antioxidant capacities
Acknowledgments
Funding for this research was provided by (i) the Interministerial
Scienti1047297c and Technical Commission of Spains Ministry of Science
and Innovation under project AGL2008-02135ALI and (ii) Xunta deGalicia Program Aacutengeles Alvarintildeo (European Social Fund)
References
AlemaacutenA Gimeacutenez BMonteroP amp Goacutemez-GuilleacutenM C(2011) Antioxidantactivityof several marine skin gelatins LWT Food Science and Technology 44 407minus413
Alemaacuten A Gimeacutenez B Peacuterez-Santiacuten E Goacutemez-Guilleacuten M C amp Montero P (2011)Contribution of Leu and Hyp residues to antioxidant and ACE-inhibitory activitiesof peptide sequences isolated from squid gelatin hydrolysates Food Chemistry 125334minus341
Balti R Nedjar-Arroume N Bougatef A Guillochon D amp Nasri M (2010) Threenovel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttle1047297sh(Sepia of 1047297cinalis) using digestive proteases Food Research International 43(4)1136minus1143
BenzieI F Famp StrainJ J(1996) Theferric reducingabilityof plasma(FRAP) asa measureof ldquoantioxidant powerrdquo The FRAP assay Analytical Biochemistry 239(1) 70minus76
Blishchenko E Y Sazonova O V Kalinina O A Moiseeva E V Vass A A KarelinA A et al (2005) Antitumor effect of valorphin in vitro and in vivo Combinedaction with cytostatic drugs Cancer Biology amp Therapy 4(1) 118minus124
Bougatef A Nedjar-Arroume N Ravallec-Pleacute R Leroy Y Guillochon D Barkia Aet al (2008) Angiotensin I-converting enzyme (ACE) inhibitory activities of sardinelle (Sardinella aurita) by-products protein hydrolysates obtained bytreatment with microbial and visceral 1047297sh serine-proteases Food Chemistry 111350minus356
Chen J Y Lin W J amp Lin T L (2009) A 1047297sh antimicrobial peptide tilapia hepcidinTH2-3 showspotent antitumor activity againsthuman 1047297brosarcomacells Peptides
30 1636minus1642ChengFYWanTCLiuYTLaiKMLinLCampSakataR(2008)Astudyof invivo
antihypertensive properties of enzymatic hydrolysate from chicken leg boneprotein Animal Science Journal 79 614minus619
Cheung H S Wang F L Miguel A O Emily F S amp Dvid W C (1980) Binding of peptide substrates and inhibitors of angiotensin-I-converting enzyme The Journalof Biological Chemistry 255 401minus407
Chung Y C Chang C T Chao W W Lin C F amp Chou S T (2002) Antioxidativeactivity and safety of the 50 ethanolic extract from red bean fermented by Bacillus
subtilis IMR-NK1 Journal of Agricultural and Food Chemistry 50 2454minus2458
0
20
40
60
80
100
C h e l a t i n g a b i l i t y ( )
0
5
10
15
20
micro m o l e s F e E q g
A
a
b
c
d
bb
d d
B
bc
d
b
d
c
de
a
C
a
bbbbb bb
0
10
20
30
40
50
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
V C E A C ( m g g )
Fig 5 Antioxidant activity of gelatin hydrolysates at a concentration of 10 mgmL ABTS radical scavenging capacity (A) Fe chelating ability (B) and Fe reducing power (FRAP) (C)Different letters (a b chellip) indicate signi1047297cant differences (ple005)
1050 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 88
Contreras M Carroacuten R Montero M J Ramos M amp Recio I (2009) Novel casein-derived peptides with antihypertensive activity International Dairy Journal 19566minus573
Dong S Zeng M Wang D Liu Z Zhao Y amp Huicheng Y (2008) Antioxidantand biochemical properties of protein hydrolysates prepared from silver carp(Hypophthalmichthys molitrix) Food Chemistry 107 1485minus1493
Doucet D Otter E O Gauthier S F amp Foegeding E A (2003) Enzyme-inducedgelation of extensively hydrolyzed whey protein by Alcalase Peptides identi1047297ca-tion and determination of enzyme speci1047297city Journal of Agricultural and FoodChemistry 51 6300minus6308
Gimeacutenez B Alemaacuten A Montero P amp Goacutemez-Guilleacuten M C (2009) Antioxidant and
functional properties of gelatin hydrolysates obtained from skin of sole and squidFood Chemistry 114(3) 976minus983Goacutemez-Guilleacuten MC Loacutepez-Caballero ME Alemaacuten ALoacutepezde LaceyA Gimeacutenez B
amp Montero P (2010) Antioxidant and antimicrobial peptide fractions from squidandtunaskin gelatin In E LeBihan(Ed) Seaby-productsas real materialNew waysof application (pp 89minus115) Kerala India Transworld Research Network
Goodfriend T L Elliott M E amp Catt K J (1996) Drug therapy ndash Angiotensin receptorsand their antagonists The New England Journal of Medicine 334(25) 1649minus1654
Gueacuterard F Dufosseacute L De La Broise D amp Binet A (2001) Enzymatic hydrolysis of proteins from yellow1047297n tuna (Thunnus albacares) wastes using Alcalase Journal of Molecular Catalysis B Enzymatic 11 1051minus1059
Hai-Lun H Xiu-Lan C Sai-Yun S Yu-Zhonga Z amp Bai-Chenga Z (2006) Analysisof novel angiotensin-I-converting enzyme inhibitory peptides from protease-hydrolyzed marine shrimp Acetes chinensis Journal of Peptide Science 12 726minus733
Hou W C Chen H J amp Lin Y H (2003) Antioxidant peptides with angiotensinconverting enzymeinhibitory activitiesand applications for angiotensin convertingenzyme puri1047297cation Journal of Agricultural and Food Chemistry 51 1706minus1709
Jang A Jo C Kang K S amp Lee M (2008) Antimicrobial and human cancer cellcytotoxic effect of synthetic angiotensin-converting enzyme (ACE) inhibitory
peptides Food Chemistry 107 327minus336 Je J -Y Lee K -H Lee M H amp Ahn C -B (2009) Antioxidant and antihypertensive
protein hydrolysates produced from tuna liver by enzymatic hydrolysis FoodResearch International 42(9) 1266minus1272
Kannan A Hettiarachchy N Johnson M G amp Nannapaneni R (2008) Human colonand liver cancer cell proliferation inhibition by peptide hydrolysates derived fromheat-stabilized defatted rice bran Journal of Agriculture and Food Chemistry 56 11643minus11647
Kaufman S H amp Earnshaw W C (2000) Induction of apoptosis by cancerchemotherapy Cell Research 256 42minus49
Khantaphant S amp Benjakul S (2008) Comparative study on the proteases from 1047297shpyloric caeca and the use for production of gelatin hydrolysate with antioxidativeactivity Comparative Biochemistry and Physiology 151 410minus419
Kim S K Byun H G Park P J amp Shahidi F (2001) Angiotensin I converting enzymeinhibitory peptides puri1047297ed form bovine skin gelatin hydrolysate Journal of
Agricultural and Food Chemistry 49 2992minus2997Kim S K Kim Y T Byun H G Nam K S Joo D S amp Shahidi F (2001) Isolation and
characterization of antioxidative peptides from gelatin hydrolysate of Alaskapollack skin Journal of Agricultural and Food Chemistry 49 1984minus1989
Kim S E Kim H H Kim J Y Kang Y I Wooa H J amp Lee H J (2000) Anticanceractivity of hydrophobic peptides from soy proteins Biofactor 21 63minus67
Klompong V Benjakul S Kantachote D amp Shahidi F (2007) Antioxidative activityand functional properties of protein hydrolysate of yellow stripe trevally(Selaroides leptolepis) as in1047298uenced by the degree of hydrolysis and enzyme typeFood Chemistry 102 1317minus1327
LeeY G Kim J Y Lee KW Kim K H amp Lee H J (2004) Peptides from anchovy sauceinduce apoptosisin a human lymphoma cell(U937) throughthe increaseof caspase-3and -8 activities Annals of the New York Academy of Sciences 1010 399minus404
Leng B Liu X D amp Chen Q X (2005) Inhibitory effects of anticancer peptide fromMercenaria on the BGC-823 cells and several enzymes FEBS 579 1187minus1190
Lin W J Chien Y L Pan C Y Lin T L Chen J Y Chiu S J amp Hui C F (2009)Epinecidin-1 an antimicrobial peptide from 1047297sh (Epinephelus coioides) which
hasan antitumoreffect like lytic peptides in human 1047297brosarcoma cells Peptides 30283minus290
Lin Lamp LiB (2006) Radical scavengingproperties of protein hydrolysates from jumbo1047298ying squid (Dosidicus eschrichitii Steenstrup) skin gelatin Journal of the Science of Food and Agriculture 86 2290minus2295
Lind D S Hochwald S N Malaty J Rekkas S Hebig P Mishra G et al (2001)Nuclear factor-kB is upregulated in colorectal cancer Surgery 130 363minus369
Megiacuteas C Pedroche J Yust M M Giroacuten-Calle J Alaiz M Millaacuten F amp Vioque J(2007) Af 1047297nity puri1047297cation of copper chelating peptides from chickpea proteinhydrolysates Journal of Agricultural and Food Chemistry 55 3949minus3954
Mendis E Rajapakse N Byun H G amp Kim S K (2005) Investigation of jumbo squid
(Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects LifeScience 77 2166minus2178Mendis E Rajapakse N amp Kim S K (2005) Antioxidant properties of a radical-
scavenging peptide puri1047297ed from enzymatically prepared 1047297sh skin gelatinhydrolysate Journal of Agricultural and Food Chemistry 53(3) 581minus587
Messerli F H (1999) Hypertension and sudden cardiac death American Journal of Hypertension 12 181minus188
Nam K A You S G amp Kim S M (2008) Molecular and physical characteristicsof squid (Toradores paci 1047297cus) skin collagens and biological properties of theirenzymatic hydrolysates Journal of Food Science 73(4) 249minus255
PicotL Bordenave S Didelot S Fruitier-Arnaudin I Sannier F ThorkelssonG et al(2006) Antiproliferative activity of 1047297sh protein hydrolysates on human breastcancer cell lines Process Biochemistry 41 1217minus1222
Re R Pellegrini N Proteggente A Pannala A Yang M amp Rice-Evans C (1999)Antioxidant activity applying an improved ABTSradical cation decolorization assayFree Radical Biology amp Medicine 26 1231minus1237
Saiga A Tanabe S amp Nishimura T (2003) Antioxidant activity of peptides obtainedfrom porcine myo1047297brillar proteins by protease treatment Journal of Agriculturaland Food Chemistry 51(12) 3661minus3667
Samaranayaka A G P Kitts D D amp Li-Chan E C Y (2010) Antioxidative andangiotensin-I-converting enzyme inhibitory potential of a paci1047297c hake (Merluccius
productus) 1047297sh protein hydrolysate subjected to simulated gastrointestinaldigestion and caco-2 cell permeation Journal of Agricultural and Food Chemistry58(3) 1535minus1542
Schagger H amp Von Jagow G (1987) Tricine-sodium dodecyl sulphate polyacrylamidegel electrophoresis for the separation of proteins in the range from 1 to 100 kDa
Analytical Biochemistry 166 368minus379Sheih I Fang T J Wu T K amp LinP H (2010) Anticancer and antioxidant activities of
the peptide fraction from algae protein waste Journal of Agricultural and FoodChemistry 58 1202minus1207
Theodore A E amp Kristinsson H G (2007) Angiotensin converting enzyme inhibitionof 1047297sh protein hydrolysates prepared from alkaline-aided channel cat1047297sh proteinisolate Journal of the Science of Food and Agriculture 87 2353minus2357
Thiansilakul Y Benjakul S amp Shahidi F (2007) Compositions functional propertiesand antioxidative activity of protein hydrolysates prepared from round scad(Decapterus maruadsi) Food Chemistry 103(4) 1385minus1394
Witherow F N Helmy A Webb D J Fox K A A amp Newby D E (2001) Bradykinincontributes to the vasodilator effects of chronic angiotensin converting enzymeinhibition in patients with heart failure Circulation 104(18) 2177minus2181
Wu J Aluko R E amp Muir A D (2002) Improved method for direct high-performanceliquid chromatography assay of angiotensin-converting enzyme-catalyzed reactions
Journal of Chromatography A 950 125minus130Yang J L Ho H Y Chu Y J amp Chow C J (2008) Characteristic and antioxidant
activity of retorted gelatin hydrolysates from cobia (Rachycetron canadum) skinFood Chemistry 110 128minus136
Zhao Y Li B Liu Z Dong S Zhao X amp Zeng M (2007) Antihypertensive effect andpuri1047297cation of an ACE inhibitory peptide from sea cucumber gelatin hydrolysateProcess Biochemistry 42 1586minus1591
Zhu LChenJ Tang Xamp XiongY (2008) Reducingradical scavenging andchelationproperties of in vitro digests of Alcalase-treated zein hydrolysate Journal of
Agricultural and Food Chemistry 56 2714minus2721
1051 A A lemaacuten e t al Food Research International 44 (2011) 1044ndash1051

8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 58
medium after the cells had been growing for 24 h antiproliferativeactivity was considered to be the effect produced when thehydrolysates were incorporated before cell growth started
Asexpectedsquidgelatin didnot show cytotoxicor antiproliferativeeffect on cancer cells However the hydrolysates prepared from squidgelatin affected the viability of both cell lines differently depending onthe enzyme used (Fig 3A B) Cell viability was determined after 24 48and 72 h with a change of culture medium and theaddition of 1 mgmL
concentration of the hydrolysate on each occasion Only EsperaseAlcalase and Savinase hydrolysates intensi1047297ed their effect on bothcancer cell lines after 48 and 72 h (ple005) The Esperase hydrolysatehad the highest (ple005) cytotoxic effect on both cancer cells (966plusmn05 and 912plusmn27 of viability inhibition on MCF-7 and U87 cellsrespectively after 72 h) It is worth noting that the same results wereobtained for a concentration of 03 mgmL for both cell lines Alcalasehydrolysate also showed relatively high cytotoxic activity (678plusmn56and 839plusmn48 viability inhibition on both cell lines respectively)
Jang Jo Kang and Lee (2008) puri1047297ed four peptides from beef proteinand caused a 20 viability inhibition on MCF-7 at 04 mgmL
Regarding antiproliferative activity again Esperase hydrolysatefollowed by Alcalase hydrolysate exhibited the highest (ple005)
activity on both cell lines (Fig 4) Picot et al (2006) evaluated theantiproliferative activity of 18 1047297sh protein hydrolysates and foundthe highest effect on MCF-7 cell lines to be around 40 for a codhydrolysate obtained with Protamex and Alcalase At the sameconcentration (1 mgmL) the Alcalase squid gelatin hydrolysatecaused a similar effect on MCF-7 cells (4164plusmn53 of growthinhibition) however the Esperase hydrolysate had a much stronger(ple005) effect (9141plusmn 18 of inhibition) and remained similar for
a concentration reduction to 05 mgmLThe IC50 was determined for both MCF-7 and U87 cells in the case
of hydrolysates that had produced the strongest cytotoxic effects(Esperase and Alcalase hydrolysates) at 24 48 and 72 h (Table 2)Both hydrolysates showed low IC50 values for both cell linesespecially the Esperase hydrolysate A pepsin hydrolysate fromalgae protein waste showed an IC50 of 174 mgmL against AGC celllines (Sheih Fang Wu amp Lin 2010) To our knowledge the IC50 valuesfor cytotoxiceffect (013 and 010 mgmL forMCF-7 andU87 cell linesrespectively) of the Esperase hydrolysate were the lowest found forwhole food protein hydrolysates although similar IC50 values havebeen reported for isolated peptides The nonapeptide X-Met-Leu-Pro-Ser-Tyr-Ser-Pro-Tyr obtained by hydrolysis of defatted soy protein
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
v i a b i l i t y i n h i b i t i o n ( )
24h 48h 72h
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
V i a b i l i t y i n h i b i t i o n ( )
24h 48h 72h
A
B
a x
c x
a b y
b x a
b x
b x
a y
a x
a x
b x
b y c
y
b x b
y d x
d x
e y e
z
e x
d y f z
a x a
b y
b y
a x
b y c
y
a x b
y b y
c x d
y e z
a x
a b y
b y
a x
b y c
z b x
c y d
z
Fig 3 Effect of gelatin hydrolysates on the viability of cancer cells for 24 48 and 72 h in cell culture medium containing a concentration of 1 mgmL of hydrolysate MCF-7 cell line(A) U87cellline (B) Differentletters (ab chellip) withinthe same time periodindicatesigni1047297cantdifferences (ple005)Different letters (xy z) withinthe same hydrolysate indicatesigni1047297cant differences (ple005)
0
20
40
60
80
100
Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
G r o w t h i n h i b i t i o n ( )
U87 MCF-7
bd
c c
c
d
e
f
b ebc
aa
e
Fig 4 Effect of gelatin hydrolysates on growth of MCF-7 and U87human cells for 72 h in cell culture mediumcontaining a concentration of 1 mgmL of hydrolysate Different letters
(a b chellip) within the same cell cultures indicate signi1047297cant differences (ple005)
1048 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 68
hadanIC50 of 016 mgmL on a mouse monocyte macrophage cell line(Kim et al 2000) A peptide fraction from algae protein waste exertedan IC50 value of 007 mgmL against AGS cells (Sheih et al 2010) Thecytotoxic effect of Esperase hydrolysate on MCF-7 cells increasedafter 48 and 72 h but to a lesser extent than on U87 cells After 24 hthe IC50 for MCF-7 cells (015 mgmL) was lower than that for U87cells (031 mgmL) however after 72 h the Esperase hydrolysate wasmore cytotoxic against U87 cells (IC50=010 mgmL) than againstMCF-7 cells (IC50=013 mgmL)
The Esperase and Alcalase hydrolysates had most of their peptidesin the range of 1400 to 500 Da Sheih et al (2010) fractioned a pepsinhydrolysate from algae protein waste and found that the mosteffective peptide fraction against AGS cells had a molecular weightlower than 6500 Da and most of the peptides (N90) were locatedbetween the range of 2000 and 200 Da Kannan Hettiarachchy
Johnson and Nannapaneni (2008) observed the highest antiprolifera-tive activity in human colon and liver cancer cells in the lowermolecular weight fraction (b5 kDa) of an Alcalase hydrolysate fromrice bran However molecular weight cannot be considered the mostimportant factor in1047298uencing anticancer activity since the Neutrasehydrolysate which has relatively low cytotoxic and antiproliferativeactivities is also made up mostly of peptides in the range of 1400ndash
500 Da Thus the antiproliferativecytotoxic effect against MCF-7and U87 cells should be linked for the most part to the presenceof speci1047297c peptides that exert a direct cytotoxic effect on cancer
cells as previously reported for valorphin a hemoglobin-derivedpeptide (Blishchenko et al 2005) Nevertheless the hypothesis of abinding competition between peptides and FCS growth factor on cellmembrane receptors cannot be excluded either Three peptides froma 1047297sh source have been described as having antitumor activity (i) a4409 Da anchovy hydrophobic peptide able to induce apoptosis inhuman U937 lymphoma cells by increasing caspase-3 and caspase-8 activity (Lee Kim Lee Kim amp Lee 2004) (ii) tilapia hepcidin TH2-3that manifested signi1047297cant antitumor activity in human 1047297brosarcomacells (Chen Lin amp Lin 2009) and (iii) epinecidin-1 a peptide from1047297sh (Epinephelus coioides) which had an antitumor effect similar tolytic peptides in human 1047297brosarcoma cells (Lin et al 2009) Howevernone of these peptides originate from 1047297sh gelatincollagen
34 Antioxidant activity
The radical scavenging capacity of squid gelatin exhibited a 3-foldincrease when it was hydrolyzed with Protamex Neutrase orNS37005 whereas such an increase was between 7 and 10-foldwhen the enzymes used were Trypsin Savinase Esperase or Alcalase(Fig 5A) However radical scavenging capacity could not be wellcorrelated with the molecular weight distribution of the wholehydrolysate since Trypsin hydrolysate was the one with the highestnumber of larger peptides This 1047297nding suggested that the radicalscavenging activity of protein hydrolysates is dependent not onlyon the peptide size but also to a greater extent on their sequencesas a result of the speci1047297city of the enzyme used
The metal chelating ability of a given compound may also serve
as a useful indicator of its potential antioxidant activity As depicted
in Fig 5B the squid gelatin did not show any Fe2+ chelating ability at10 mgmL However when gelatin was hydrolyzed with ProtamexNeutrase or NS37005 enzymes the resulting hydrolysates did exert alow chelating capacity (less than 25) In contrast the TrypsinSavinase or Esperase hydrolysates at the same concentration caused~80 of Fe2+ chelation which was near to 100 with the Alcalasehydrolysate Dong Zeng Wang Liu Zhao and Huicheng(2008) founda metal chelation activity of 9297 with an Alcalase hydrolysate fromsilver carp at 5 mgmL
Hydroxyl radicals can be formed from superoxide anion andhydrogen peroxide in the presence of transition metal ions such asFe2+ andCu2+ Thus chelating metal ions may inhibit theformation of hydroxyl radicals Presumably peptide cleavage leads to enhancedmetal ion binding because carboxyl and amino groups in the sidechains of the acidic (Glx and Asx) and basic (Lys His and Arg) aminoacids are thought to play an important role in chelating metal ions(Saiga Tanabe amp Nishimura 2003)
The molecular weight of peptidesis also believed to play a key rolein chelating metals Klompong Benjakul Kantachote and Shaidi(2007)and Dong et al (2008) pointed out that metal-chelating abilityand molecular weight had a linear relationship and that metal-chelating ability increased with decreasing molecular weight Never-theless Megiacuteas et al (2007) reported that if peptide length is tooshort chelation is unstable In our study hydrolysate with peptidesmostly between ~1400 and 500 Da (Alcalase hydrolysate) exerted
the highest metal chelating ability However it is worth noting theconsiderably high metal chelating activity in the Trypsin hydrolysateas compared to other hydrolysates with higher proportions of smallerpeptides (Neutraseor Protamex) denoting onceagainthe importanceof the presence of speci1047297c peptide sequences rather than the wholehydrolysate molecular weight distribution
The behavior for Fe2+ chelation and ABTS+ scavenging followed asimilar trend suggesting that the two antioxidant mechanisms mightbe related Zhu Chen Tang and Xiong (2008) also found a similartrend between Cu2+ chelating capacity and ABTS radical scavengingfor an Alcalase hydrolysate from zein protein
Another method for measuring antioxidant potential is theferric reducing antioxidant power (FRAP) assay Reducing power isa measurement that provides an estimation of a compounds ability
to reduce ferric iron (III) to ferrous iron (II) and is determinedusing a redox-linked colorimetric reaction Samples with higherreducing power are more able to donate electrons or hydrogenFree radicals form stable substances by accepting donated electronsand thus the free radical chain reaction could be interruptedThe reducing power of squid gelatin and hydrolysates is shownin Fig 5C The ability of the hydrolysates to reduce Fe3+ to Fe2+
was approximately twice than that of the gelatin with all thehydrolysates showing a similar (pN005) capacity independently of the enzyme used and consequently of molecular weight distribu-tion too In several studies involving protein hydrolysates someresearchers reported that the reducing capacity improved as thedegree of protein hydrolysis increased (Thiansilakul Benjakulamp Shahidi 2007) while some researchers reported the opposite
(Klompong et al 2007)
Table 2
Cytotoxic activity of Esperase and Alcalase squid gelatin hydrolysates
IC50
MCF-7 cells U87 cells
24 h 48 h 72 h 24 h 48 h 72 h
Esperase 015 plusmn001a 014plusmn001a 013plusmn001a 031plusmn001c 013plusmn001b 010plusmn001a
Alcalase 118 plusmn002c 104plusmn002b 081plusmn003a 122plusmn003c 098plusmn003b 085plusmn002a
Different letters (a b) within the same hydrolysate and the same cell line indicate signi1047297cant differences (ple005) IC50 concentration (mgmL) required to induce 50 cells death
1049 A A lemaacuten e t al F ood Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 78
Alcalase Esperase Savinase and Trypsin signi1047297cantly improvedthe antioxidant activity of the resulting hydrolysates via the enhance-ment of antiradical activity and metal chelation Reducing powerwas also increased by the action of these enzymes but the degreeof enhancement was much lower Consistent with our results thehydrolysis of snapper gelatin with proteases from pyloric caecacaused a 55 to 65-fold increase in ABTS radical scavenging capacitywhereas ferric reducing power did not increase more than 2-fold(Khantaphant amp Benjakul 2008) Free radical quenchingwas reportedto be the main antioxidative mechanism of some peptides derivedfrom squid gelatin hydrolysates (Mendis Rajapakse amp Kim 2005)
4 Conclusions
The Esperase hydrolysate exerted a potent cytotoxic and anti-proliferative effects on the two cancer cell lines studied withsigni1047297cant antioxidant activity especially with respect to metalchelating and antiradical activities The Alcalase hydrolysate was themost potent ACE-inhibitor and metal chelator while at the same timeexerting strong radical scavenging and cytotoxic activity againstcancer cells These results suggest that squid gelatin hydrolysatesprepared with Esperase or Alcalase enzymes could be utilized as newmaterials in the development of functional foods with potentialanticancer antihypertensive and antioxidant capacities
Acknowledgments
Funding for this research was provided by (i) the Interministerial
Scienti1047297c and Technical Commission of Spains Ministry of Science
and Innovation under project AGL2008-02135ALI and (ii) Xunta deGalicia Program Aacutengeles Alvarintildeo (European Social Fund)
References
AlemaacutenA Gimeacutenez BMonteroP amp Goacutemez-GuilleacutenM C(2011) Antioxidantactivityof several marine skin gelatins LWT Food Science and Technology 44 407minus413
Alemaacuten A Gimeacutenez B Peacuterez-Santiacuten E Goacutemez-Guilleacuten M C amp Montero P (2011)Contribution of Leu and Hyp residues to antioxidant and ACE-inhibitory activitiesof peptide sequences isolated from squid gelatin hydrolysates Food Chemistry 125334minus341
Balti R Nedjar-Arroume N Bougatef A Guillochon D amp Nasri M (2010) Threenovel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttle1047297sh(Sepia of 1047297cinalis) using digestive proteases Food Research International 43(4)1136minus1143
BenzieI F Famp StrainJ J(1996) Theferric reducingabilityof plasma(FRAP) asa measureof ldquoantioxidant powerrdquo The FRAP assay Analytical Biochemistry 239(1) 70minus76
Blishchenko E Y Sazonova O V Kalinina O A Moiseeva E V Vass A A KarelinA A et al (2005) Antitumor effect of valorphin in vitro and in vivo Combinedaction with cytostatic drugs Cancer Biology amp Therapy 4(1) 118minus124
Bougatef A Nedjar-Arroume N Ravallec-Pleacute R Leroy Y Guillochon D Barkia Aet al (2008) Angiotensin I-converting enzyme (ACE) inhibitory activities of sardinelle (Sardinella aurita) by-products protein hydrolysates obtained bytreatment with microbial and visceral 1047297sh serine-proteases Food Chemistry 111350minus356
Chen J Y Lin W J amp Lin T L (2009) A 1047297sh antimicrobial peptide tilapia hepcidinTH2-3 showspotent antitumor activity againsthuman 1047297brosarcomacells Peptides
30 1636minus1642ChengFYWanTCLiuYTLaiKMLinLCampSakataR(2008)Astudyof invivo
antihypertensive properties of enzymatic hydrolysate from chicken leg boneprotein Animal Science Journal 79 614minus619
Cheung H S Wang F L Miguel A O Emily F S amp Dvid W C (1980) Binding of peptide substrates and inhibitors of angiotensin-I-converting enzyme The Journalof Biological Chemistry 255 401minus407
Chung Y C Chang C T Chao W W Lin C F amp Chou S T (2002) Antioxidativeactivity and safety of the 50 ethanolic extract from red bean fermented by Bacillus
subtilis IMR-NK1 Journal of Agricultural and Food Chemistry 50 2454minus2458
0
20
40
60
80
100
C h e l a t i n g a b i l i t y ( )
0
5
10
15
20
micro m o l e s F e E q g
A
a
b
c
d
bb
d d
B
bc
d
b
d
c
de
a
C
a
bbbbb bb
0
10
20
30
40
50
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
V C E A C ( m g g )
Fig 5 Antioxidant activity of gelatin hydrolysates at a concentration of 10 mgmL ABTS radical scavenging capacity (A) Fe chelating ability (B) and Fe reducing power (FRAP) (C)Different letters (a b chellip) indicate signi1047297cant differences (ple005)
1050 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 88
Contreras M Carroacuten R Montero M J Ramos M amp Recio I (2009) Novel casein-derived peptides with antihypertensive activity International Dairy Journal 19566minus573
Dong S Zeng M Wang D Liu Z Zhao Y amp Huicheng Y (2008) Antioxidantand biochemical properties of protein hydrolysates prepared from silver carp(Hypophthalmichthys molitrix) Food Chemistry 107 1485minus1493
Doucet D Otter E O Gauthier S F amp Foegeding E A (2003) Enzyme-inducedgelation of extensively hydrolyzed whey protein by Alcalase Peptides identi1047297ca-tion and determination of enzyme speci1047297city Journal of Agricultural and FoodChemistry 51 6300minus6308
Gimeacutenez B Alemaacuten A Montero P amp Goacutemez-Guilleacuten M C (2009) Antioxidant and
functional properties of gelatin hydrolysates obtained from skin of sole and squidFood Chemistry 114(3) 976minus983Goacutemez-Guilleacuten MC Loacutepez-Caballero ME Alemaacuten ALoacutepezde LaceyA Gimeacutenez B
amp Montero P (2010) Antioxidant and antimicrobial peptide fractions from squidandtunaskin gelatin In E LeBihan(Ed) Seaby-productsas real materialNew waysof application (pp 89minus115) Kerala India Transworld Research Network
Goodfriend T L Elliott M E amp Catt K J (1996) Drug therapy ndash Angiotensin receptorsand their antagonists The New England Journal of Medicine 334(25) 1649minus1654
Gueacuterard F Dufosseacute L De La Broise D amp Binet A (2001) Enzymatic hydrolysis of proteins from yellow1047297n tuna (Thunnus albacares) wastes using Alcalase Journal of Molecular Catalysis B Enzymatic 11 1051minus1059
Hai-Lun H Xiu-Lan C Sai-Yun S Yu-Zhonga Z amp Bai-Chenga Z (2006) Analysisof novel angiotensin-I-converting enzyme inhibitory peptides from protease-hydrolyzed marine shrimp Acetes chinensis Journal of Peptide Science 12 726minus733
Hou W C Chen H J amp Lin Y H (2003) Antioxidant peptides with angiotensinconverting enzymeinhibitory activitiesand applications for angiotensin convertingenzyme puri1047297cation Journal of Agricultural and Food Chemistry 51 1706minus1709
Jang A Jo C Kang K S amp Lee M (2008) Antimicrobial and human cancer cellcytotoxic effect of synthetic angiotensin-converting enzyme (ACE) inhibitory
peptides Food Chemistry 107 327minus336 Je J -Y Lee K -H Lee M H amp Ahn C -B (2009) Antioxidant and antihypertensive
protein hydrolysates produced from tuna liver by enzymatic hydrolysis FoodResearch International 42(9) 1266minus1272
Kannan A Hettiarachchy N Johnson M G amp Nannapaneni R (2008) Human colonand liver cancer cell proliferation inhibition by peptide hydrolysates derived fromheat-stabilized defatted rice bran Journal of Agriculture and Food Chemistry 56 11643minus11647
Kaufman S H amp Earnshaw W C (2000) Induction of apoptosis by cancerchemotherapy Cell Research 256 42minus49
Khantaphant S amp Benjakul S (2008) Comparative study on the proteases from 1047297shpyloric caeca and the use for production of gelatin hydrolysate with antioxidativeactivity Comparative Biochemistry and Physiology 151 410minus419
Kim S K Byun H G Park P J amp Shahidi F (2001) Angiotensin I converting enzymeinhibitory peptides puri1047297ed form bovine skin gelatin hydrolysate Journal of
Agricultural and Food Chemistry 49 2992minus2997Kim S K Kim Y T Byun H G Nam K S Joo D S amp Shahidi F (2001) Isolation and
characterization of antioxidative peptides from gelatin hydrolysate of Alaskapollack skin Journal of Agricultural and Food Chemistry 49 1984minus1989
Kim S E Kim H H Kim J Y Kang Y I Wooa H J amp Lee H J (2000) Anticanceractivity of hydrophobic peptides from soy proteins Biofactor 21 63minus67
Klompong V Benjakul S Kantachote D amp Shahidi F (2007) Antioxidative activityand functional properties of protein hydrolysate of yellow stripe trevally(Selaroides leptolepis) as in1047298uenced by the degree of hydrolysis and enzyme typeFood Chemistry 102 1317minus1327
LeeY G Kim J Y Lee KW Kim K H amp Lee H J (2004) Peptides from anchovy sauceinduce apoptosisin a human lymphoma cell(U937) throughthe increaseof caspase-3and -8 activities Annals of the New York Academy of Sciences 1010 399minus404
Leng B Liu X D amp Chen Q X (2005) Inhibitory effects of anticancer peptide fromMercenaria on the BGC-823 cells and several enzymes FEBS 579 1187minus1190
Lin W J Chien Y L Pan C Y Lin T L Chen J Y Chiu S J amp Hui C F (2009)Epinecidin-1 an antimicrobial peptide from 1047297sh (Epinephelus coioides) which
hasan antitumoreffect like lytic peptides in human 1047297brosarcoma cells Peptides 30283minus290
Lin Lamp LiB (2006) Radical scavengingproperties of protein hydrolysates from jumbo1047298ying squid (Dosidicus eschrichitii Steenstrup) skin gelatin Journal of the Science of Food and Agriculture 86 2290minus2295
Lind D S Hochwald S N Malaty J Rekkas S Hebig P Mishra G et al (2001)Nuclear factor-kB is upregulated in colorectal cancer Surgery 130 363minus369
Megiacuteas C Pedroche J Yust M M Giroacuten-Calle J Alaiz M Millaacuten F amp Vioque J(2007) Af 1047297nity puri1047297cation of copper chelating peptides from chickpea proteinhydrolysates Journal of Agricultural and Food Chemistry 55 3949minus3954
Mendis E Rajapakse N Byun H G amp Kim S K (2005) Investigation of jumbo squid
(Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects LifeScience 77 2166minus2178Mendis E Rajapakse N amp Kim S K (2005) Antioxidant properties of a radical-
scavenging peptide puri1047297ed from enzymatically prepared 1047297sh skin gelatinhydrolysate Journal of Agricultural and Food Chemistry 53(3) 581minus587
Messerli F H (1999) Hypertension and sudden cardiac death American Journal of Hypertension 12 181minus188
Nam K A You S G amp Kim S M (2008) Molecular and physical characteristicsof squid (Toradores paci 1047297cus) skin collagens and biological properties of theirenzymatic hydrolysates Journal of Food Science 73(4) 249minus255
PicotL Bordenave S Didelot S Fruitier-Arnaudin I Sannier F ThorkelssonG et al(2006) Antiproliferative activity of 1047297sh protein hydrolysates on human breastcancer cell lines Process Biochemistry 41 1217minus1222
Re R Pellegrini N Proteggente A Pannala A Yang M amp Rice-Evans C (1999)Antioxidant activity applying an improved ABTSradical cation decolorization assayFree Radical Biology amp Medicine 26 1231minus1237
Saiga A Tanabe S amp Nishimura T (2003) Antioxidant activity of peptides obtainedfrom porcine myo1047297brillar proteins by protease treatment Journal of Agriculturaland Food Chemistry 51(12) 3661minus3667
Samaranayaka A G P Kitts D D amp Li-Chan E C Y (2010) Antioxidative andangiotensin-I-converting enzyme inhibitory potential of a paci1047297c hake (Merluccius
productus) 1047297sh protein hydrolysate subjected to simulated gastrointestinaldigestion and caco-2 cell permeation Journal of Agricultural and Food Chemistry58(3) 1535minus1542
Schagger H amp Von Jagow G (1987) Tricine-sodium dodecyl sulphate polyacrylamidegel electrophoresis for the separation of proteins in the range from 1 to 100 kDa
Analytical Biochemistry 166 368minus379Sheih I Fang T J Wu T K amp LinP H (2010) Anticancer and antioxidant activities of
the peptide fraction from algae protein waste Journal of Agricultural and FoodChemistry 58 1202minus1207
Theodore A E amp Kristinsson H G (2007) Angiotensin converting enzyme inhibitionof 1047297sh protein hydrolysates prepared from alkaline-aided channel cat1047297sh proteinisolate Journal of the Science of Food and Agriculture 87 2353minus2357
Thiansilakul Y Benjakul S amp Shahidi F (2007) Compositions functional propertiesand antioxidative activity of protein hydrolysates prepared from round scad(Decapterus maruadsi) Food Chemistry 103(4) 1385minus1394
Witherow F N Helmy A Webb D J Fox K A A amp Newby D E (2001) Bradykinincontributes to the vasodilator effects of chronic angiotensin converting enzymeinhibition in patients with heart failure Circulation 104(18) 2177minus2181
Wu J Aluko R E amp Muir A D (2002) Improved method for direct high-performanceliquid chromatography assay of angiotensin-converting enzyme-catalyzed reactions
Journal of Chromatography A 950 125minus130Yang J L Ho H Y Chu Y J amp Chow C J (2008) Characteristic and antioxidant
activity of retorted gelatin hydrolysates from cobia (Rachycetron canadum) skinFood Chemistry 110 128minus136
Zhao Y Li B Liu Z Dong S Zhao X amp Zeng M (2007) Antihypertensive effect andpuri1047297cation of an ACE inhibitory peptide from sea cucumber gelatin hydrolysateProcess Biochemistry 42 1586minus1591
Zhu LChenJ Tang Xamp XiongY (2008) Reducingradical scavenging andchelationproperties of in vitro digests of Alcalase-treated zein hydrolysate Journal of
Agricultural and Food Chemistry 56 2714minus2721
1051 A A lemaacuten e t al Food Research International 44 (2011) 1044ndash1051

8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 68
hadanIC50 of 016 mgmL on a mouse monocyte macrophage cell line(Kim et al 2000) A peptide fraction from algae protein waste exertedan IC50 value of 007 mgmL against AGS cells (Sheih et al 2010) Thecytotoxic effect of Esperase hydrolysate on MCF-7 cells increasedafter 48 and 72 h but to a lesser extent than on U87 cells After 24 hthe IC50 for MCF-7 cells (015 mgmL) was lower than that for U87cells (031 mgmL) however after 72 h the Esperase hydrolysate wasmore cytotoxic against U87 cells (IC50=010 mgmL) than againstMCF-7 cells (IC50=013 mgmL)
The Esperase and Alcalase hydrolysates had most of their peptidesin the range of 1400 to 500 Da Sheih et al (2010) fractioned a pepsinhydrolysate from algae protein waste and found that the mosteffective peptide fraction against AGS cells had a molecular weightlower than 6500 Da and most of the peptides (N90) were locatedbetween the range of 2000 and 200 Da Kannan Hettiarachchy
Johnson and Nannapaneni (2008) observed the highest antiprolifera-tive activity in human colon and liver cancer cells in the lowermolecular weight fraction (b5 kDa) of an Alcalase hydrolysate fromrice bran However molecular weight cannot be considered the mostimportant factor in1047298uencing anticancer activity since the Neutrasehydrolysate which has relatively low cytotoxic and antiproliferativeactivities is also made up mostly of peptides in the range of 1400ndash
500 Da Thus the antiproliferativecytotoxic effect against MCF-7and U87 cells should be linked for the most part to the presenceof speci1047297c peptides that exert a direct cytotoxic effect on cancer
cells as previously reported for valorphin a hemoglobin-derivedpeptide (Blishchenko et al 2005) Nevertheless the hypothesis of abinding competition between peptides and FCS growth factor on cellmembrane receptors cannot be excluded either Three peptides froma 1047297sh source have been described as having antitumor activity (i) a4409 Da anchovy hydrophobic peptide able to induce apoptosis inhuman U937 lymphoma cells by increasing caspase-3 and caspase-8 activity (Lee Kim Lee Kim amp Lee 2004) (ii) tilapia hepcidin TH2-3that manifested signi1047297cant antitumor activity in human 1047297brosarcomacells (Chen Lin amp Lin 2009) and (iii) epinecidin-1 a peptide from1047297sh (Epinephelus coioides) which had an antitumor effect similar tolytic peptides in human 1047297brosarcoma cells (Lin et al 2009) Howevernone of these peptides originate from 1047297sh gelatincollagen
34 Antioxidant activity
The radical scavenging capacity of squid gelatin exhibited a 3-foldincrease when it was hydrolyzed with Protamex Neutrase orNS37005 whereas such an increase was between 7 and 10-foldwhen the enzymes used were Trypsin Savinase Esperase or Alcalase(Fig 5A) However radical scavenging capacity could not be wellcorrelated with the molecular weight distribution of the wholehydrolysate since Trypsin hydrolysate was the one with the highestnumber of larger peptides This 1047297nding suggested that the radicalscavenging activity of protein hydrolysates is dependent not onlyon the peptide size but also to a greater extent on their sequencesas a result of the speci1047297city of the enzyme used
The metal chelating ability of a given compound may also serve
as a useful indicator of its potential antioxidant activity As depicted
in Fig 5B the squid gelatin did not show any Fe2+ chelating ability at10 mgmL However when gelatin was hydrolyzed with ProtamexNeutrase or NS37005 enzymes the resulting hydrolysates did exert alow chelating capacity (less than 25) In contrast the TrypsinSavinase or Esperase hydrolysates at the same concentration caused~80 of Fe2+ chelation which was near to 100 with the Alcalasehydrolysate Dong Zeng Wang Liu Zhao and Huicheng(2008) founda metal chelation activity of 9297 with an Alcalase hydrolysate fromsilver carp at 5 mgmL
Hydroxyl radicals can be formed from superoxide anion andhydrogen peroxide in the presence of transition metal ions such asFe2+ andCu2+ Thus chelating metal ions may inhibit theformation of hydroxyl radicals Presumably peptide cleavage leads to enhancedmetal ion binding because carboxyl and amino groups in the sidechains of the acidic (Glx and Asx) and basic (Lys His and Arg) aminoacids are thought to play an important role in chelating metal ions(Saiga Tanabe amp Nishimura 2003)
The molecular weight of peptidesis also believed to play a key rolein chelating metals Klompong Benjakul Kantachote and Shaidi(2007)and Dong et al (2008) pointed out that metal-chelating abilityand molecular weight had a linear relationship and that metal-chelating ability increased with decreasing molecular weight Never-theless Megiacuteas et al (2007) reported that if peptide length is tooshort chelation is unstable In our study hydrolysate with peptidesmostly between ~1400 and 500 Da (Alcalase hydrolysate) exerted
the highest metal chelating ability However it is worth noting theconsiderably high metal chelating activity in the Trypsin hydrolysateas compared to other hydrolysates with higher proportions of smallerpeptides (Neutraseor Protamex) denoting onceagainthe importanceof the presence of speci1047297c peptide sequences rather than the wholehydrolysate molecular weight distribution
The behavior for Fe2+ chelation and ABTS+ scavenging followed asimilar trend suggesting that the two antioxidant mechanisms mightbe related Zhu Chen Tang and Xiong (2008) also found a similartrend between Cu2+ chelating capacity and ABTS radical scavengingfor an Alcalase hydrolysate from zein protein
Another method for measuring antioxidant potential is theferric reducing antioxidant power (FRAP) assay Reducing power isa measurement that provides an estimation of a compounds ability
to reduce ferric iron (III) to ferrous iron (II) and is determinedusing a redox-linked colorimetric reaction Samples with higherreducing power are more able to donate electrons or hydrogenFree radicals form stable substances by accepting donated electronsand thus the free radical chain reaction could be interruptedThe reducing power of squid gelatin and hydrolysates is shownin Fig 5C The ability of the hydrolysates to reduce Fe3+ to Fe2+
was approximately twice than that of the gelatin with all thehydrolysates showing a similar (pN005) capacity independently of the enzyme used and consequently of molecular weight distribu-tion too In several studies involving protein hydrolysates someresearchers reported that the reducing capacity improved as thedegree of protein hydrolysis increased (Thiansilakul Benjakulamp Shahidi 2007) while some researchers reported the opposite
(Klompong et al 2007)
Table 2
Cytotoxic activity of Esperase and Alcalase squid gelatin hydrolysates
IC50
MCF-7 cells U87 cells
24 h 48 h 72 h 24 h 48 h 72 h
Esperase 015 plusmn001a 014plusmn001a 013plusmn001a 031plusmn001c 013plusmn001b 010plusmn001a
Alcalase 118 plusmn002c 104plusmn002b 081plusmn003a 122plusmn003c 098plusmn003b 085plusmn002a
Different letters (a b) within the same hydrolysate and the same cell line indicate signi1047297cant differences (ple005) IC50 concentration (mgmL) required to induce 50 cells death
1049 A A lemaacuten e t al F ood Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 78
Alcalase Esperase Savinase and Trypsin signi1047297cantly improvedthe antioxidant activity of the resulting hydrolysates via the enhance-ment of antiradical activity and metal chelation Reducing powerwas also increased by the action of these enzymes but the degreeof enhancement was much lower Consistent with our results thehydrolysis of snapper gelatin with proteases from pyloric caecacaused a 55 to 65-fold increase in ABTS radical scavenging capacitywhereas ferric reducing power did not increase more than 2-fold(Khantaphant amp Benjakul 2008) Free radical quenchingwas reportedto be the main antioxidative mechanism of some peptides derivedfrom squid gelatin hydrolysates (Mendis Rajapakse amp Kim 2005)
4 Conclusions
The Esperase hydrolysate exerted a potent cytotoxic and anti-proliferative effects on the two cancer cell lines studied withsigni1047297cant antioxidant activity especially with respect to metalchelating and antiradical activities The Alcalase hydrolysate was themost potent ACE-inhibitor and metal chelator while at the same timeexerting strong radical scavenging and cytotoxic activity againstcancer cells These results suggest that squid gelatin hydrolysatesprepared with Esperase or Alcalase enzymes could be utilized as newmaterials in the development of functional foods with potentialanticancer antihypertensive and antioxidant capacities
Acknowledgments
Funding for this research was provided by (i) the Interministerial
Scienti1047297c and Technical Commission of Spains Ministry of Science
and Innovation under project AGL2008-02135ALI and (ii) Xunta deGalicia Program Aacutengeles Alvarintildeo (European Social Fund)
References
AlemaacutenA Gimeacutenez BMonteroP amp Goacutemez-GuilleacutenM C(2011) Antioxidantactivityof several marine skin gelatins LWT Food Science and Technology 44 407minus413
Alemaacuten A Gimeacutenez B Peacuterez-Santiacuten E Goacutemez-Guilleacuten M C amp Montero P (2011)Contribution of Leu and Hyp residues to antioxidant and ACE-inhibitory activitiesof peptide sequences isolated from squid gelatin hydrolysates Food Chemistry 125334minus341
Balti R Nedjar-Arroume N Bougatef A Guillochon D amp Nasri M (2010) Threenovel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttle1047297sh(Sepia of 1047297cinalis) using digestive proteases Food Research International 43(4)1136minus1143
BenzieI F Famp StrainJ J(1996) Theferric reducingabilityof plasma(FRAP) asa measureof ldquoantioxidant powerrdquo The FRAP assay Analytical Biochemistry 239(1) 70minus76
Blishchenko E Y Sazonova O V Kalinina O A Moiseeva E V Vass A A KarelinA A et al (2005) Antitumor effect of valorphin in vitro and in vivo Combinedaction with cytostatic drugs Cancer Biology amp Therapy 4(1) 118minus124
Bougatef A Nedjar-Arroume N Ravallec-Pleacute R Leroy Y Guillochon D Barkia Aet al (2008) Angiotensin I-converting enzyme (ACE) inhibitory activities of sardinelle (Sardinella aurita) by-products protein hydrolysates obtained bytreatment with microbial and visceral 1047297sh serine-proteases Food Chemistry 111350minus356
Chen J Y Lin W J amp Lin T L (2009) A 1047297sh antimicrobial peptide tilapia hepcidinTH2-3 showspotent antitumor activity againsthuman 1047297brosarcomacells Peptides
30 1636minus1642ChengFYWanTCLiuYTLaiKMLinLCampSakataR(2008)Astudyof invivo
antihypertensive properties of enzymatic hydrolysate from chicken leg boneprotein Animal Science Journal 79 614minus619
Cheung H S Wang F L Miguel A O Emily F S amp Dvid W C (1980) Binding of peptide substrates and inhibitors of angiotensin-I-converting enzyme The Journalof Biological Chemistry 255 401minus407
Chung Y C Chang C T Chao W W Lin C F amp Chou S T (2002) Antioxidativeactivity and safety of the 50 ethanolic extract from red bean fermented by Bacillus
subtilis IMR-NK1 Journal of Agricultural and Food Chemistry 50 2454minus2458
0
20
40
60
80
100
C h e l a t i n g a b i l i t y ( )
0
5
10
15
20
micro m o l e s F e E q g
A
a
b
c
d
bb
d d
B
bc
d
b
d
c
de
a
C
a
bbbbb bb
0
10
20
30
40
50
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
V C E A C ( m g g )
Fig 5 Antioxidant activity of gelatin hydrolysates at a concentration of 10 mgmL ABTS radical scavenging capacity (A) Fe chelating ability (B) and Fe reducing power (FRAP) (C)Different letters (a b chellip) indicate signi1047297cant differences (ple005)
1050 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 88
Contreras M Carroacuten R Montero M J Ramos M amp Recio I (2009) Novel casein-derived peptides with antihypertensive activity International Dairy Journal 19566minus573
Dong S Zeng M Wang D Liu Z Zhao Y amp Huicheng Y (2008) Antioxidantand biochemical properties of protein hydrolysates prepared from silver carp(Hypophthalmichthys molitrix) Food Chemistry 107 1485minus1493
Doucet D Otter E O Gauthier S F amp Foegeding E A (2003) Enzyme-inducedgelation of extensively hydrolyzed whey protein by Alcalase Peptides identi1047297ca-tion and determination of enzyme speci1047297city Journal of Agricultural and FoodChemistry 51 6300minus6308
Gimeacutenez B Alemaacuten A Montero P amp Goacutemez-Guilleacuten M C (2009) Antioxidant and
functional properties of gelatin hydrolysates obtained from skin of sole and squidFood Chemistry 114(3) 976minus983Goacutemez-Guilleacuten MC Loacutepez-Caballero ME Alemaacuten ALoacutepezde LaceyA Gimeacutenez B
amp Montero P (2010) Antioxidant and antimicrobial peptide fractions from squidandtunaskin gelatin In E LeBihan(Ed) Seaby-productsas real materialNew waysof application (pp 89minus115) Kerala India Transworld Research Network
Goodfriend T L Elliott M E amp Catt K J (1996) Drug therapy ndash Angiotensin receptorsand their antagonists The New England Journal of Medicine 334(25) 1649minus1654
Gueacuterard F Dufosseacute L De La Broise D amp Binet A (2001) Enzymatic hydrolysis of proteins from yellow1047297n tuna (Thunnus albacares) wastes using Alcalase Journal of Molecular Catalysis B Enzymatic 11 1051minus1059
Hai-Lun H Xiu-Lan C Sai-Yun S Yu-Zhonga Z amp Bai-Chenga Z (2006) Analysisof novel angiotensin-I-converting enzyme inhibitory peptides from protease-hydrolyzed marine shrimp Acetes chinensis Journal of Peptide Science 12 726minus733
Hou W C Chen H J amp Lin Y H (2003) Antioxidant peptides with angiotensinconverting enzymeinhibitory activitiesand applications for angiotensin convertingenzyme puri1047297cation Journal of Agricultural and Food Chemistry 51 1706minus1709
Jang A Jo C Kang K S amp Lee M (2008) Antimicrobial and human cancer cellcytotoxic effect of synthetic angiotensin-converting enzyme (ACE) inhibitory
peptides Food Chemistry 107 327minus336 Je J -Y Lee K -H Lee M H amp Ahn C -B (2009) Antioxidant and antihypertensive
protein hydrolysates produced from tuna liver by enzymatic hydrolysis FoodResearch International 42(9) 1266minus1272
Kannan A Hettiarachchy N Johnson M G amp Nannapaneni R (2008) Human colonand liver cancer cell proliferation inhibition by peptide hydrolysates derived fromheat-stabilized defatted rice bran Journal of Agriculture and Food Chemistry 56 11643minus11647
Kaufman S H amp Earnshaw W C (2000) Induction of apoptosis by cancerchemotherapy Cell Research 256 42minus49
Khantaphant S amp Benjakul S (2008) Comparative study on the proteases from 1047297shpyloric caeca and the use for production of gelatin hydrolysate with antioxidativeactivity Comparative Biochemistry and Physiology 151 410minus419
Kim S K Byun H G Park P J amp Shahidi F (2001) Angiotensin I converting enzymeinhibitory peptides puri1047297ed form bovine skin gelatin hydrolysate Journal of
Agricultural and Food Chemistry 49 2992minus2997Kim S K Kim Y T Byun H G Nam K S Joo D S amp Shahidi F (2001) Isolation and
characterization of antioxidative peptides from gelatin hydrolysate of Alaskapollack skin Journal of Agricultural and Food Chemistry 49 1984minus1989
Kim S E Kim H H Kim J Y Kang Y I Wooa H J amp Lee H J (2000) Anticanceractivity of hydrophobic peptides from soy proteins Biofactor 21 63minus67
Klompong V Benjakul S Kantachote D amp Shahidi F (2007) Antioxidative activityand functional properties of protein hydrolysate of yellow stripe trevally(Selaroides leptolepis) as in1047298uenced by the degree of hydrolysis and enzyme typeFood Chemistry 102 1317minus1327
LeeY G Kim J Y Lee KW Kim K H amp Lee H J (2004) Peptides from anchovy sauceinduce apoptosisin a human lymphoma cell(U937) throughthe increaseof caspase-3and -8 activities Annals of the New York Academy of Sciences 1010 399minus404
Leng B Liu X D amp Chen Q X (2005) Inhibitory effects of anticancer peptide fromMercenaria on the BGC-823 cells and several enzymes FEBS 579 1187minus1190
Lin W J Chien Y L Pan C Y Lin T L Chen J Y Chiu S J amp Hui C F (2009)Epinecidin-1 an antimicrobial peptide from 1047297sh (Epinephelus coioides) which
hasan antitumoreffect like lytic peptides in human 1047297brosarcoma cells Peptides 30283minus290
Lin Lamp LiB (2006) Radical scavengingproperties of protein hydrolysates from jumbo1047298ying squid (Dosidicus eschrichitii Steenstrup) skin gelatin Journal of the Science of Food and Agriculture 86 2290minus2295
Lind D S Hochwald S N Malaty J Rekkas S Hebig P Mishra G et al (2001)Nuclear factor-kB is upregulated in colorectal cancer Surgery 130 363minus369
Megiacuteas C Pedroche J Yust M M Giroacuten-Calle J Alaiz M Millaacuten F amp Vioque J(2007) Af 1047297nity puri1047297cation of copper chelating peptides from chickpea proteinhydrolysates Journal of Agricultural and Food Chemistry 55 3949minus3954
Mendis E Rajapakse N Byun H G amp Kim S K (2005) Investigation of jumbo squid
(Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects LifeScience 77 2166minus2178Mendis E Rajapakse N amp Kim S K (2005) Antioxidant properties of a radical-
scavenging peptide puri1047297ed from enzymatically prepared 1047297sh skin gelatinhydrolysate Journal of Agricultural and Food Chemistry 53(3) 581minus587
Messerli F H (1999) Hypertension and sudden cardiac death American Journal of Hypertension 12 181minus188
Nam K A You S G amp Kim S M (2008) Molecular and physical characteristicsof squid (Toradores paci 1047297cus) skin collagens and biological properties of theirenzymatic hydrolysates Journal of Food Science 73(4) 249minus255
PicotL Bordenave S Didelot S Fruitier-Arnaudin I Sannier F ThorkelssonG et al(2006) Antiproliferative activity of 1047297sh protein hydrolysates on human breastcancer cell lines Process Biochemistry 41 1217minus1222
Re R Pellegrini N Proteggente A Pannala A Yang M amp Rice-Evans C (1999)Antioxidant activity applying an improved ABTSradical cation decolorization assayFree Radical Biology amp Medicine 26 1231minus1237
Saiga A Tanabe S amp Nishimura T (2003) Antioxidant activity of peptides obtainedfrom porcine myo1047297brillar proteins by protease treatment Journal of Agriculturaland Food Chemistry 51(12) 3661minus3667
Samaranayaka A G P Kitts D D amp Li-Chan E C Y (2010) Antioxidative andangiotensin-I-converting enzyme inhibitory potential of a paci1047297c hake (Merluccius
productus) 1047297sh protein hydrolysate subjected to simulated gastrointestinaldigestion and caco-2 cell permeation Journal of Agricultural and Food Chemistry58(3) 1535minus1542
Schagger H amp Von Jagow G (1987) Tricine-sodium dodecyl sulphate polyacrylamidegel electrophoresis for the separation of proteins in the range from 1 to 100 kDa
Analytical Biochemistry 166 368minus379Sheih I Fang T J Wu T K amp LinP H (2010) Anticancer and antioxidant activities of
the peptide fraction from algae protein waste Journal of Agricultural and FoodChemistry 58 1202minus1207
Theodore A E amp Kristinsson H G (2007) Angiotensin converting enzyme inhibitionof 1047297sh protein hydrolysates prepared from alkaline-aided channel cat1047297sh proteinisolate Journal of the Science of Food and Agriculture 87 2353minus2357
Thiansilakul Y Benjakul S amp Shahidi F (2007) Compositions functional propertiesand antioxidative activity of protein hydrolysates prepared from round scad(Decapterus maruadsi) Food Chemistry 103(4) 1385minus1394
Witherow F N Helmy A Webb D J Fox K A A amp Newby D E (2001) Bradykinincontributes to the vasodilator effects of chronic angiotensin converting enzymeinhibition in patients with heart failure Circulation 104(18) 2177minus2181
Wu J Aluko R E amp Muir A D (2002) Improved method for direct high-performanceliquid chromatography assay of angiotensin-converting enzyme-catalyzed reactions
Journal of Chromatography A 950 125minus130Yang J L Ho H Y Chu Y J amp Chow C J (2008) Characteristic and antioxidant
activity of retorted gelatin hydrolysates from cobia (Rachycetron canadum) skinFood Chemistry 110 128minus136
Zhao Y Li B Liu Z Dong S Zhao X amp Zeng M (2007) Antihypertensive effect andpuri1047297cation of an ACE inhibitory peptide from sea cucumber gelatin hydrolysateProcess Biochemistry 42 1586minus1591
Zhu LChenJ Tang Xamp XiongY (2008) Reducingradical scavenging andchelationproperties of in vitro digests of Alcalase-treated zein hydrolysate Journal of
Agricultural and Food Chemistry 56 2714minus2721
1051 A A lemaacuten e t al Food Research International 44 (2011) 1044ndash1051
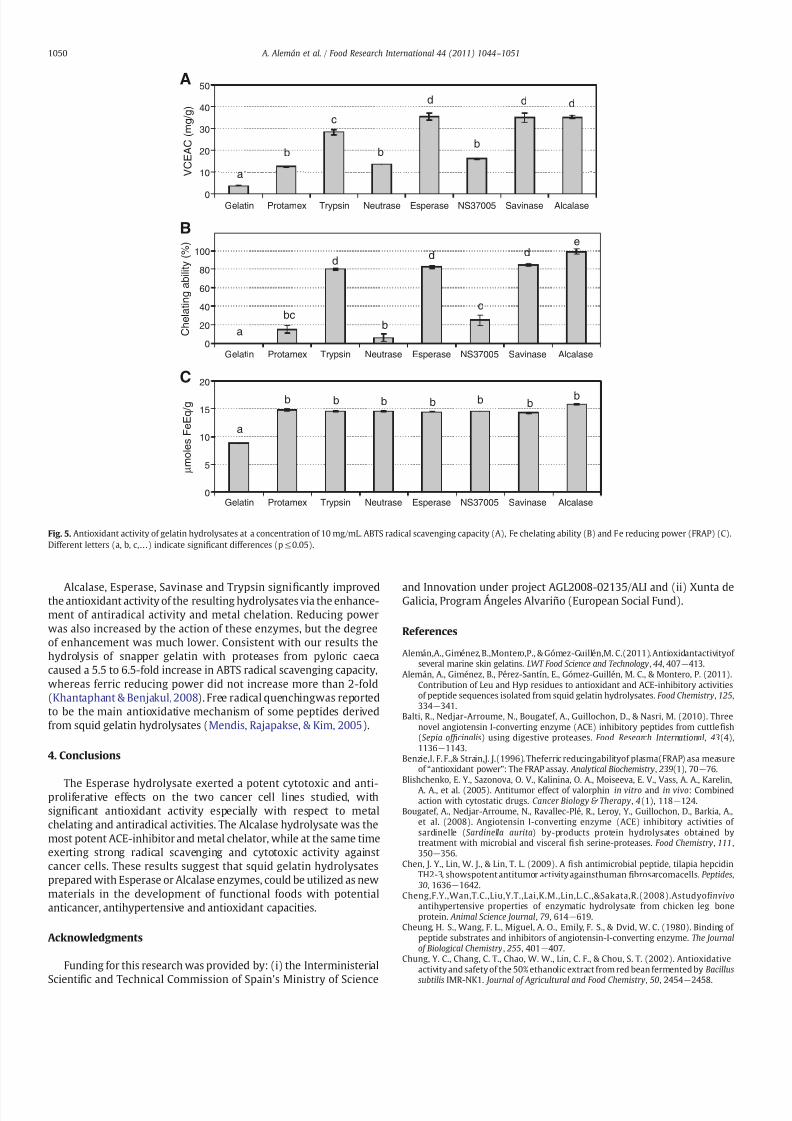
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 78
Alcalase Esperase Savinase and Trypsin signi1047297cantly improvedthe antioxidant activity of the resulting hydrolysates via the enhance-ment of antiradical activity and metal chelation Reducing powerwas also increased by the action of these enzymes but the degreeof enhancement was much lower Consistent with our results thehydrolysis of snapper gelatin with proteases from pyloric caecacaused a 55 to 65-fold increase in ABTS radical scavenging capacitywhereas ferric reducing power did not increase more than 2-fold(Khantaphant amp Benjakul 2008) Free radical quenchingwas reportedto be the main antioxidative mechanism of some peptides derivedfrom squid gelatin hydrolysates (Mendis Rajapakse amp Kim 2005)
4 Conclusions
The Esperase hydrolysate exerted a potent cytotoxic and anti-proliferative effects on the two cancer cell lines studied withsigni1047297cant antioxidant activity especially with respect to metalchelating and antiradical activities The Alcalase hydrolysate was themost potent ACE-inhibitor and metal chelator while at the same timeexerting strong radical scavenging and cytotoxic activity againstcancer cells These results suggest that squid gelatin hydrolysatesprepared with Esperase or Alcalase enzymes could be utilized as newmaterials in the development of functional foods with potentialanticancer antihypertensive and antioxidant capacities
Acknowledgments
Funding for this research was provided by (i) the Interministerial
Scienti1047297c and Technical Commission of Spains Ministry of Science
and Innovation under project AGL2008-02135ALI and (ii) Xunta deGalicia Program Aacutengeles Alvarintildeo (European Social Fund)
References
AlemaacutenA Gimeacutenez BMonteroP amp Goacutemez-GuilleacutenM C(2011) Antioxidantactivityof several marine skin gelatins LWT Food Science and Technology 44 407minus413
Alemaacuten A Gimeacutenez B Peacuterez-Santiacuten E Goacutemez-Guilleacuten M C amp Montero P (2011)Contribution of Leu and Hyp residues to antioxidant and ACE-inhibitory activitiesof peptide sequences isolated from squid gelatin hydrolysates Food Chemistry 125334minus341
Balti R Nedjar-Arroume N Bougatef A Guillochon D amp Nasri M (2010) Threenovel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttle1047297sh(Sepia of 1047297cinalis) using digestive proteases Food Research International 43(4)1136minus1143
BenzieI F Famp StrainJ J(1996) Theferric reducingabilityof plasma(FRAP) asa measureof ldquoantioxidant powerrdquo The FRAP assay Analytical Biochemistry 239(1) 70minus76
Blishchenko E Y Sazonova O V Kalinina O A Moiseeva E V Vass A A KarelinA A et al (2005) Antitumor effect of valorphin in vitro and in vivo Combinedaction with cytostatic drugs Cancer Biology amp Therapy 4(1) 118minus124
Bougatef A Nedjar-Arroume N Ravallec-Pleacute R Leroy Y Guillochon D Barkia Aet al (2008) Angiotensin I-converting enzyme (ACE) inhibitory activities of sardinelle (Sardinella aurita) by-products protein hydrolysates obtained bytreatment with microbial and visceral 1047297sh serine-proteases Food Chemistry 111350minus356
Chen J Y Lin W J amp Lin T L (2009) A 1047297sh antimicrobial peptide tilapia hepcidinTH2-3 showspotent antitumor activity againsthuman 1047297brosarcomacells Peptides
30 1636minus1642ChengFYWanTCLiuYTLaiKMLinLCampSakataR(2008)Astudyof invivo
antihypertensive properties of enzymatic hydrolysate from chicken leg boneprotein Animal Science Journal 79 614minus619
Cheung H S Wang F L Miguel A O Emily F S amp Dvid W C (1980) Binding of peptide substrates and inhibitors of angiotensin-I-converting enzyme The Journalof Biological Chemistry 255 401minus407
Chung Y C Chang C T Chao W W Lin C F amp Chou S T (2002) Antioxidativeactivity and safety of the 50 ethanolic extract from red bean fermented by Bacillus
subtilis IMR-NK1 Journal of Agricultural and Food Chemistry 50 2454minus2458
0
20
40
60
80
100
C h e l a t i n g a b i l i t y ( )
0
5
10
15
20
micro m o l e s F e E q g
A
a
b
c
d
bb
d d
B
bc
d
b
d
c
de
a
C
a
bbbbb bb
0
10
20
30
40
50
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
Gelatin Protamex Trypsin Neutrase Esperase NS37005 Savinase Alcalase
V C E A C ( m g g )
Fig 5 Antioxidant activity of gelatin hydrolysates at a concentration of 10 mgmL ABTS radical scavenging capacity (A) Fe chelating ability (B) and Fe reducing power (FRAP) (C)Different letters (a b chellip) indicate signi1047297cant differences (ple005)
1050 A Alemaacuten et al Food Research International 44 (2011) 1044ndash1051
8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 88
Contreras M Carroacuten R Montero M J Ramos M amp Recio I (2009) Novel casein-derived peptides with antihypertensive activity International Dairy Journal 19566minus573
Dong S Zeng M Wang D Liu Z Zhao Y amp Huicheng Y (2008) Antioxidantand biochemical properties of protein hydrolysates prepared from silver carp(Hypophthalmichthys molitrix) Food Chemistry 107 1485minus1493
Doucet D Otter E O Gauthier S F amp Foegeding E A (2003) Enzyme-inducedgelation of extensively hydrolyzed whey protein by Alcalase Peptides identi1047297ca-tion and determination of enzyme speci1047297city Journal of Agricultural and FoodChemistry 51 6300minus6308
Gimeacutenez B Alemaacuten A Montero P amp Goacutemez-Guilleacuten M C (2009) Antioxidant and
functional properties of gelatin hydrolysates obtained from skin of sole and squidFood Chemistry 114(3) 976minus983Goacutemez-Guilleacuten MC Loacutepez-Caballero ME Alemaacuten ALoacutepezde LaceyA Gimeacutenez B
amp Montero P (2010) Antioxidant and antimicrobial peptide fractions from squidandtunaskin gelatin In E LeBihan(Ed) Seaby-productsas real materialNew waysof application (pp 89minus115) Kerala India Transworld Research Network
Goodfriend T L Elliott M E amp Catt K J (1996) Drug therapy ndash Angiotensin receptorsand their antagonists The New England Journal of Medicine 334(25) 1649minus1654
Gueacuterard F Dufosseacute L De La Broise D amp Binet A (2001) Enzymatic hydrolysis of proteins from yellow1047297n tuna (Thunnus albacares) wastes using Alcalase Journal of Molecular Catalysis B Enzymatic 11 1051minus1059
Hai-Lun H Xiu-Lan C Sai-Yun S Yu-Zhonga Z amp Bai-Chenga Z (2006) Analysisof novel angiotensin-I-converting enzyme inhibitory peptides from protease-hydrolyzed marine shrimp Acetes chinensis Journal of Peptide Science 12 726minus733
Hou W C Chen H J amp Lin Y H (2003) Antioxidant peptides with angiotensinconverting enzymeinhibitory activitiesand applications for angiotensin convertingenzyme puri1047297cation Journal of Agricultural and Food Chemistry 51 1706minus1709
Jang A Jo C Kang K S amp Lee M (2008) Antimicrobial and human cancer cellcytotoxic effect of synthetic angiotensin-converting enzyme (ACE) inhibitory
peptides Food Chemistry 107 327minus336 Je J -Y Lee K -H Lee M H amp Ahn C -B (2009) Antioxidant and antihypertensive
protein hydrolysates produced from tuna liver by enzymatic hydrolysis FoodResearch International 42(9) 1266minus1272
Kannan A Hettiarachchy N Johnson M G amp Nannapaneni R (2008) Human colonand liver cancer cell proliferation inhibition by peptide hydrolysates derived fromheat-stabilized defatted rice bran Journal of Agriculture and Food Chemistry 56 11643minus11647
Kaufman S H amp Earnshaw W C (2000) Induction of apoptosis by cancerchemotherapy Cell Research 256 42minus49
Khantaphant S amp Benjakul S (2008) Comparative study on the proteases from 1047297shpyloric caeca and the use for production of gelatin hydrolysate with antioxidativeactivity Comparative Biochemistry and Physiology 151 410minus419
Kim S K Byun H G Park P J amp Shahidi F (2001) Angiotensin I converting enzymeinhibitory peptides puri1047297ed form bovine skin gelatin hydrolysate Journal of
Agricultural and Food Chemistry 49 2992minus2997Kim S K Kim Y T Byun H G Nam K S Joo D S amp Shahidi F (2001) Isolation and
characterization of antioxidative peptides from gelatin hydrolysate of Alaskapollack skin Journal of Agricultural and Food Chemistry 49 1984minus1989
Kim S E Kim H H Kim J Y Kang Y I Wooa H J amp Lee H J (2000) Anticanceractivity of hydrophobic peptides from soy proteins Biofactor 21 63minus67
Klompong V Benjakul S Kantachote D amp Shahidi F (2007) Antioxidative activityand functional properties of protein hydrolysate of yellow stripe trevally(Selaroides leptolepis) as in1047298uenced by the degree of hydrolysis and enzyme typeFood Chemistry 102 1317minus1327
LeeY G Kim J Y Lee KW Kim K H amp Lee H J (2004) Peptides from anchovy sauceinduce apoptosisin a human lymphoma cell(U937) throughthe increaseof caspase-3and -8 activities Annals of the New York Academy of Sciences 1010 399minus404
Leng B Liu X D amp Chen Q X (2005) Inhibitory effects of anticancer peptide fromMercenaria on the BGC-823 cells and several enzymes FEBS 579 1187minus1190
Lin W J Chien Y L Pan C Y Lin T L Chen J Y Chiu S J amp Hui C F (2009)Epinecidin-1 an antimicrobial peptide from 1047297sh (Epinephelus coioides) which
hasan antitumoreffect like lytic peptides in human 1047297brosarcoma cells Peptides 30283minus290
Lin Lamp LiB (2006) Radical scavengingproperties of protein hydrolysates from jumbo1047298ying squid (Dosidicus eschrichitii Steenstrup) skin gelatin Journal of the Science of Food and Agriculture 86 2290minus2295
Lind D S Hochwald S N Malaty J Rekkas S Hebig P Mishra G et al (2001)Nuclear factor-kB is upregulated in colorectal cancer Surgery 130 363minus369
Megiacuteas C Pedroche J Yust M M Giroacuten-Calle J Alaiz M Millaacuten F amp Vioque J(2007) Af 1047297nity puri1047297cation of copper chelating peptides from chickpea proteinhydrolysates Journal of Agricultural and Food Chemistry 55 3949minus3954
Mendis E Rajapakse N Byun H G amp Kim S K (2005) Investigation of jumbo squid
(Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects LifeScience 77 2166minus2178Mendis E Rajapakse N amp Kim S K (2005) Antioxidant properties of a radical-
scavenging peptide puri1047297ed from enzymatically prepared 1047297sh skin gelatinhydrolysate Journal of Agricultural and Food Chemistry 53(3) 581minus587
Messerli F H (1999) Hypertension and sudden cardiac death American Journal of Hypertension 12 181minus188
Nam K A You S G amp Kim S M (2008) Molecular and physical characteristicsof squid (Toradores paci 1047297cus) skin collagens and biological properties of theirenzymatic hydrolysates Journal of Food Science 73(4) 249minus255
PicotL Bordenave S Didelot S Fruitier-Arnaudin I Sannier F ThorkelssonG et al(2006) Antiproliferative activity of 1047297sh protein hydrolysates on human breastcancer cell lines Process Biochemistry 41 1217minus1222
Re R Pellegrini N Proteggente A Pannala A Yang M amp Rice-Evans C (1999)Antioxidant activity applying an improved ABTSradical cation decolorization assayFree Radical Biology amp Medicine 26 1231minus1237
Saiga A Tanabe S amp Nishimura T (2003) Antioxidant activity of peptides obtainedfrom porcine myo1047297brillar proteins by protease treatment Journal of Agriculturaland Food Chemistry 51(12) 3661minus3667
Samaranayaka A G P Kitts D D amp Li-Chan E C Y (2010) Antioxidative andangiotensin-I-converting enzyme inhibitory potential of a paci1047297c hake (Merluccius
productus) 1047297sh protein hydrolysate subjected to simulated gastrointestinaldigestion and caco-2 cell permeation Journal of Agricultural and Food Chemistry58(3) 1535minus1542
Schagger H amp Von Jagow G (1987) Tricine-sodium dodecyl sulphate polyacrylamidegel electrophoresis for the separation of proteins in the range from 1 to 100 kDa
Analytical Biochemistry 166 368minus379Sheih I Fang T J Wu T K amp LinP H (2010) Anticancer and antioxidant activities of
the peptide fraction from algae protein waste Journal of Agricultural and FoodChemistry 58 1202minus1207
Theodore A E amp Kristinsson H G (2007) Angiotensin converting enzyme inhibitionof 1047297sh protein hydrolysates prepared from alkaline-aided channel cat1047297sh proteinisolate Journal of the Science of Food and Agriculture 87 2353minus2357
Thiansilakul Y Benjakul S amp Shahidi F (2007) Compositions functional propertiesand antioxidative activity of protein hydrolysates prepared from round scad(Decapterus maruadsi) Food Chemistry 103(4) 1385minus1394
Witherow F N Helmy A Webb D J Fox K A A amp Newby D E (2001) Bradykinincontributes to the vasodilator effects of chronic angiotensin converting enzymeinhibition in patients with heart failure Circulation 104(18) 2177minus2181
Wu J Aluko R E amp Muir A D (2002) Improved method for direct high-performanceliquid chromatography assay of angiotensin-converting enzyme-catalyzed reactions
Journal of Chromatography A 950 125minus130Yang J L Ho H Y Chu Y J amp Chow C J (2008) Characteristic and antioxidant
activity of retorted gelatin hydrolysates from cobia (Rachycetron canadum) skinFood Chemistry 110 128minus136
Zhao Y Li B Liu Z Dong S Zhao X amp Zeng M (2007) Antihypertensive effect andpuri1047297cation of an ACE inhibitory peptide from sea cucumber gelatin hydrolysateProcess Biochemistry 42 1586minus1591
Zhu LChenJ Tang Xamp XiongY (2008) Reducingradical scavenging andchelationproperties of in vitro digests of Alcalase-treated zein hydrolysate Journal of
Agricultural and Food Chemistry 56 2714minus2721
1051 A A lemaacuten e t al Food Research International 44 (2011) 1044ndash1051

8102019 Jurnal Cumi Cumi
httpslidepdfcomreaderfulljurnal-cumi-cumi 88
Contreras M Carroacuten R Montero M J Ramos M amp Recio I (2009) Novel casein-derived peptides with antihypertensive activity International Dairy Journal 19566minus573
Dong S Zeng M Wang D Liu Z Zhao Y amp Huicheng Y (2008) Antioxidantand biochemical properties of protein hydrolysates prepared from silver carp(Hypophthalmichthys molitrix) Food Chemistry 107 1485minus1493
Doucet D Otter E O Gauthier S F amp Foegeding E A (2003) Enzyme-inducedgelation of extensively hydrolyzed whey protein by Alcalase Peptides identi1047297ca-tion and determination of enzyme speci1047297city Journal of Agricultural and FoodChemistry 51 6300minus6308
Gimeacutenez B Alemaacuten A Montero P amp Goacutemez-Guilleacuten M C (2009) Antioxidant and
functional properties of gelatin hydrolysates obtained from skin of sole and squidFood Chemistry 114(3) 976minus983Goacutemez-Guilleacuten MC Loacutepez-Caballero ME Alemaacuten ALoacutepezde LaceyA Gimeacutenez B
amp Montero P (2010) Antioxidant and antimicrobial peptide fractions from squidandtunaskin gelatin In E LeBihan(Ed) Seaby-productsas real materialNew waysof application (pp 89minus115) Kerala India Transworld Research Network
Goodfriend T L Elliott M E amp Catt K J (1996) Drug therapy ndash Angiotensin receptorsand their antagonists The New England Journal of Medicine 334(25) 1649minus1654
Gueacuterard F Dufosseacute L De La Broise D amp Binet A (2001) Enzymatic hydrolysis of proteins from yellow1047297n tuna (Thunnus albacares) wastes using Alcalase Journal of Molecular Catalysis B Enzymatic 11 1051minus1059
Hai-Lun H Xiu-Lan C Sai-Yun S Yu-Zhonga Z amp Bai-Chenga Z (2006) Analysisof novel angiotensin-I-converting enzyme inhibitory peptides from protease-hydrolyzed marine shrimp Acetes chinensis Journal of Peptide Science 12 726minus733
Hou W C Chen H J amp Lin Y H (2003) Antioxidant peptides with angiotensinconverting enzymeinhibitory activitiesand applications for angiotensin convertingenzyme puri1047297cation Journal of Agricultural and Food Chemistry 51 1706minus1709
Jang A Jo C Kang K S amp Lee M (2008) Antimicrobial and human cancer cellcytotoxic effect of synthetic angiotensin-converting enzyme (ACE) inhibitory
peptides Food Chemistry 107 327minus336 Je J -Y Lee K -H Lee M H amp Ahn C -B (2009) Antioxidant and antihypertensive
protein hydrolysates produced from tuna liver by enzymatic hydrolysis FoodResearch International 42(9) 1266minus1272
Kannan A Hettiarachchy N Johnson M G amp Nannapaneni R (2008) Human colonand liver cancer cell proliferation inhibition by peptide hydrolysates derived fromheat-stabilized defatted rice bran Journal of Agriculture and Food Chemistry 56 11643minus11647
Kaufman S H amp Earnshaw W C (2000) Induction of apoptosis by cancerchemotherapy Cell Research 256 42minus49
Khantaphant S amp Benjakul S (2008) Comparative study on the proteases from 1047297shpyloric caeca and the use for production of gelatin hydrolysate with antioxidativeactivity Comparative Biochemistry and Physiology 151 410minus419
Kim S K Byun H G Park P J amp Shahidi F (2001) Angiotensin I converting enzymeinhibitory peptides puri1047297ed form bovine skin gelatin hydrolysate Journal of
Agricultural and Food Chemistry 49 2992minus2997Kim S K Kim Y T Byun H G Nam K S Joo D S amp Shahidi F (2001) Isolation and
characterization of antioxidative peptides from gelatin hydrolysate of Alaskapollack skin Journal of Agricultural and Food Chemistry 49 1984minus1989
Kim S E Kim H H Kim J Y Kang Y I Wooa H J amp Lee H J (2000) Anticanceractivity of hydrophobic peptides from soy proteins Biofactor 21 63minus67
Klompong V Benjakul S Kantachote D amp Shahidi F (2007) Antioxidative activityand functional properties of protein hydrolysate of yellow stripe trevally(Selaroides leptolepis) as in1047298uenced by the degree of hydrolysis and enzyme typeFood Chemistry 102 1317minus1327
LeeY G Kim J Y Lee KW Kim K H amp Lee H J (2004) Peptides from anchovy sauceinduce apoptosisin a human lymphoma cell(U937) throughthe increaseof caspase-3and -8 activities Annals of the New York Academy of Sciences 1010 399minus404
Leng B Liu X D amp Chen Q X (2005) Inhibitory effects of anticancer peptide fromMercenaria on the BGC-823 cells and several enzymes FEBS 579 1187minus1190
Lin W J Chien Y L Pan C Y Lin T L Chen J Y Chiu S J amp Hui C F (2009)Epinecidin-1 an antimicrobial peptide from 1047297sh (Epinephelus coioides) which
hasan antitumoreffect like lytic peptides in human 1047297brosarcoma cells Peptides 30283minus290
Lin Lamp LiB (2006) Radical scavengingproperties of protein hydrolysates from jumbo1047298ying squid (Dosidicus eschrichitii Steenstrup) skin gelatin Journal of the Science of Food and Agriculture 86 2290minus2295
Lind D S Hochwald S N Malaty J Rekkas S Hebig P Mishra G et al (2001)Nuclear factor-kB is upregulated in colorectal cancer Surgery 130 363minus369
Megiacuteas C Pedroche J Yust M M Giroacuten-Calle J Alaiz M Millaacuten F amp Vioque J(2007) Af 1047297nity puri1047297cation of copper chelating peptides from chickpea proteinhydrolysates Journal of Agricultural and Food Chemistry 55 3949minus3954
Mendis E Rajapakse N Byun H G amp Kim S K (2005) Investigation of jumbo squid
(Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects LifeScience 77 2166minus2178Mendis E Rajapakse N amp Kim S K (2005) Antioxidant properties of a radical-
scavenging peptide puri1047297ed from enzymatically prepared 1047297sh skin gelatinhydrolysate Journal of Agricultural and Food Chemistry 53(3) 581minus587
Messerli F H (1999) Hypertension and sudden cardiac death American Journal of Hypertension 12 181minus188
Nam K A You S G amp Kim S M (2008) Molecular and physical characteristicsof squid (Toradores paci 1047297cus) skin collagens and biological properties of theirenzymatic hydrolysates Journal of Food Science 73(4) 249minus255
PicotL Bordenave S Didelot S Fruitier-Arnaudin I Sannier F ThorkelssonG et al(2006) Antiproliferative activity of 1047297sh protein hydrolysates on human breastcancer cell lines Process Biochemistry 41 1217minus1222
Re R Pellegrini N Proteggente A Pannala A Yang M amp Rice-Evans C (1999)Antioxidant activity applying an improved ABTSradical cation decolorization assayFree Radical Biology amp Medicine 26 1231minus1237
Saiga A Tanabe S amp Nishimura T (2003) Antioxidant activity of peptides obtainedfrom porcine myo1047297brillar proteins by protease treatment Journal of Agriculturaland Food Chemistry 51(12) 3661minus3667
Samaranayaka A G P Kitts D D amp Li-Chan E C Y (2010) Antioxidative andangiotensin-I-converting enzyme inhibitory potential of a paci1047297c hake (Merluccius
productus) 1047297sh protein hydrolysate subjected to simulated gastrointestinaldigestion and caco-2 cell permeation Journal of Agricultural and Food Chemistry58(3) 1535minus1542
Schagger H amp Von Jagow G (1987) Tricine-sodium dodecyl sulphate polyacrylamidegel electrophoresis for the separation of proteins in the range from 1 to 100 kDa
Analytical Biochemistry 166 368minus379Sheih I Fang T J Wu T K amp LinP H (2010) Anticancer and antioxidant activities of
the peptide fraction from algae protein waste Journal of Agricultural and FoodChemistry 58 1202minus1207
Theodore A E amp Kristinsson H G (2007) Angiotensin converting enzyme inhibitionof 1047297sh protein hydrolysates prepared from alkaline-aided channel cat1047297sh proteinisolate Journal of the Science of Food and Agriculture 87 2353minus2357
Thiansilakul Y Benjakul S amp Shahidi F (2007) Compositions functional propertiesand antioxidative activity of protein hydrolysates prepared from round scad(Decapterus maruadsi) Food Chemistry 103(4) 1385minus1394
Witherow F N Helmy A Webb D J Fox K A A amp Newby D E (2001) Bradykinincontributes to the vasodilator effects of chronic angiotensin converting enzymeinhibition in patients with heart failure Circulation 104(18) 2177minus2181
Wu J Aluko R E amp Muir A D (2002) Improved method for direct high-performanceliquid chromatography assay of angiotensin-converting enzyme-catalyzed reactions
Journal of Chromatography A 950 125minus130Yang J L Ho H Y Chu Y J amp Chow C J (2008) Characteristic and antioxidant
activity of retorted gelatin hydrolysates from cobia (Rachycetron canadum) skinFood Chemistry 110 128minus136
Zhao Y Li B Liu Z Dong S Zhao X amp Zeng M (2007) Antihypertensive effect andpuri1047297cation of an ACE inhibitory peptide from sea cucumber gelatin hydrolysateProcess Biochemistry 42 1586minus1591
Zhu LChenJ Tang Xamp XiongY (2008) Reducingradical scavenging andchelationproperties of in vitro digests of Alcalase-treated zein hydrolysate Journal of
Agricultural and Food Chemistry 56 2714minus2721
1051 A A lemaacuten e t al Food Research International 44 (2011) 1044ndash1051


















