INTRA- AND EXTRACELLULAR DISTRIBUTION OF AN ENDOGENOUS...
12
J. Cell Sci. 84, 201-212 (1986) 201 Printed in Great Britain © The Company of Biologists Limited 1986 INTRA- AND EXTRACELLULAR DISTRIBUTION OF AN ENDOGENOUS LECTIN DURING ERYTHROPOIESIS F. LYNNE HARRISON AND JAMES W. CATT Department of Cell Biology, AFRC Institute of Animal Physiology, Babraham, Cambridge CB2 4AT, UK SUMMARY An endogenous /3-galactoside-specific lectin has previously been isolated from rabbit bone marrow. The quantification of extracted lectin now indicates that approximately 75 % of the lectin is intracellular in marrow. Indirect immunofluorescence studies show the extracellular lectin is associated with the erythroblast cell surface and is also found in some acellular areas of the marrow stroma. At enucleation, lectin surrounds the extruded nucleus while some residual lectin is observed in the cytoplasm of circulating reticulocytes and erythrocytes. INTRODUCTION An endogenous /3-galactoside-specific lectin has previously been identified in rabbit bone marrow tissue and preliminary observations suggested that the lectin might mediate inter-erythroblast associations during maturation; the lectin was therefore termed erythroid developmental agglutinin, EDA (Harrison & Chesterton, 1980a). Subsequently a lectin indistinguishable from that isolated from the marrow has been identified in a number of adult and neonatal rabbit tissues (Harrison et al. 1984). Moreover, endogenous /3-galactoside-specific lectins have been identified in a number of other vertebrate species and are probably ubiquitous in animal tissues. On the basis of their structural similarity and saccharide specificity we have proposed that these lectins be collectively termed 'galaptins' (Harrison & Chesterton, 19806) and now refer to the lectin in marrow as bone-marrow galaptin. We report here an investigation of the intra-/extracellular distribution of the marrow galaptin using biochemical techniques and also the confirmation and extension of these studies by the immunocytochemical localization of lectin in both gently disaggregated marrow cells and resin-embedded whole marrow. A prelimi- nary report of some of this work has been published previously (Harrison & Catt, 1985). MATERIALS AND METHODS Materials New Zealand White rabbits (2-0-2-5 kg) were made anaemic by five daily injections of phenyl- hydrazine (2-5 % in water, pH7-4, O'SSmlkg" 1 ) and marrow was extracted from the femur, tibia Key words: lectin, /J-galactoside, erythropoiesis.
Transcript of INTRA- AND EXTRACELLULAR DISTRIBUTION OF AN ENDOGENOUS...

J. Cell Sci. 84, 201-212 (1986) 201Printed in Great Britain © The Company of Biologists Limited 1986
INTRA- AND EXTRACELLULAR DISTRIBUTION
OF AN ENDOGENOUS LECTIN DURING
ERYTHROPOIESIS
F. LYNNE HARRISON AND JAMES W. CATTDepartment of Cell Biology, AFRC Institute of Animal Physiology, Babraham,Cambridge CB2 4AT, UK
SUMMARY
An endogenous /3-galactoside-specific lectin has previously been isolated from rabbit bonemarrow. The quantification of extracted lectin now indicates that approximately 75 % of the lectinis intracellular in marrow. Indirect immunofluorescence studies show the extracellular lectin isassociated with the erythroblast cell surface and is also found in some acellular areas of the marrowstroma. At enucleation, lectin surrounds the extruded nucleus while some residual lectin isobserved in the cytoplasm of circulating reticulocytes and erythrocytes.
INTRODUCTION
An endogenous /3-galactoside-specific lectin has previously been identified inrabbit bone marrow tissue and preliminary observations suggested that the lectinmight mediate inter-erythroblast associations during maturation; the lectin wastherefore termed erythroid developmental agglutinin, EDA (Harrison & Chesterton,1980a). Subsequently a lectin indistinguishable from that isolated from the marrowhas been identified in a number of adult and neonatal rabbit tissues (Harrison et al.1984). Moreover, endogenous /3-galactoside-specific lectins have been identified in anumber of other vertebrate species and are probably ubiquitous in animal tissues. Onthe basis of their structural similarity and saccharide specificity we have proposedthat these lectins be collectively termed 'galaptins' (Harrison & Chesterton, 19806)and now refer to the lectin in marrow as bone-marrow galaptin.
We report here an investigation of the intra-/extracellular distribution of themarrow galaptin using biochemical techniques and also the confirmation andextension of these studies by the immunocytochemical localization of lectin in bothgently disaggregated marrow cells and resin-embedded whole marrow. A prelimi-nary report of some of this work has been published previously (Harrison & Catt,1985).
MATERIALS AND METHODS
MaterialsNew Zealand White rabbits (2-0-2-5 kg) were made anaemic by five daily injections of phenyl-
hydrazine (2-5 % in water, pH7-4, O'SSmlkg"1) and marrow was extracted from the femur, tibia
Key words: lectin, /J-galactoside, erythropoiesis.

202 F. L. Harrison andjf. W. Catt
and humerus 48 h after the last injection. The antiserum used had been raised in a sheep,immunized with bone-marrow galaptin purified from lactose washes of marrow cells. The anti-bodies' specificity for the isoforms of bone-marrow galaptin has previously been determined(FitzGerald et al. 1984). Anti-galaptin (anti-bmg) and preimmune IgG (immunoglobulin G)fractions were prepared from sera by ammonium sulphate precipitation (50%) and subsequentdialysis against 0-l M-NaCl.
Analytical techniquesLactose extracts were analysed by sodium dodecyl sulphate (SDS)-polyacrylamide gel electro-
phoresis (11 % T, 2-6% C), isoelectricfocusing on agarose gels and Western blotting, using anti-bmg followed by peroxidase-linked rabbit anti-sheep IgG as described (FitzGerald et al. 1984).The lectin content of each lactose extraction was determined within 24 h by single radial immuno-diffusion (SRID) assays. Duplicate samples were assayed in triplicate and compared with standarddilutions of purified bone-marrow galaptin on the same diffusion plate. The reliability of the assayin crude lactose extracts was established by adding a known amount of purified galaptin to eachsample and checking that the expected increase in apparent lectin concentration was observed.Experimental details have been reported (Harrison et al. 1984).
Immunofluorescence studiesCytocentrifuge slides, prepared from normal and anaemic marrow, gently disaggregated in
phosphate-buffered saline (PBS; 6-5 mM-phosphate, pH7-4, O15M-NaCl), and from blood cellscollected into PBS and concentrated by centrifugation (1500g'for 5 min), were dried under vacuum(over P2O5) overnight. In anaemic blood 92-98% of the cells were reticulocytes, judged by thepresence of intracellular RNA precipitates after staining with Cresyl Blue ( 1 % , w/v, BrilliantCresyl Blue in PBS, mixed 1:1 (v/v) with blood sample and viewed immediately). The cyto-centrifuge slides were fixed for 10 min in methanol and rehydrated in PBS for 15 min immediatelybefore staining. All immunological reagents were spun for 2 min in a Beckman Microfuge (approx.9000#) just before use and all procedures were performed at room temperature. Cells, covered witha drop of preimmune or anti-bmg IgG, 0-1 mgml"1 in 5 % (w/v) bovine serum albumin (BSA) inPBS, were incubated for 3 h in a wet box and then washed for 3 h in a minimum of 500 ml PBS,with stirring, keeping slides treated with preimmune and anti-bmg IgG separate. Cells were thencovered with a drop of fluorescein isothiocyanate(FITC)-derivatized rabbit anti-sheep IgG(FIRaSh, Miles Laboratories Ltd), diluted 80 times in rabbit serum + 001 % (v/v) Nonidet P40(NP40), incubated for 30 min and then washed for 1 h in PBS. Slides were mounted in 80%glycerol in PBS containing Citifluor mountant, which retards fading of fluorescence duringillumination (obtainable from Chemistry Department, City University, London, UK) and viewedon a Leitz Dialux microscope with epifluorescent illumination. Results were recorded on KodakXP 1 or Kodacolour 400 film and printed on Ilford Multigrade II paper for black and white prints.
To prepare sections, bone marrow was chopped into approximately 5 mm cubes and fixed in 8 %formaldehyde for 1 h. Fixation, embedding in JB-4 resin (Polysciences Ltd) and the cutting ofsections was carried out as previously described (Catt & Harrison, 1985a), except that the high fatcontent of the marrow necessitated a prolonged dehydration of 30 min at 0°C in each of a gradedacetone/water series and overnight dehydration in 100 % acetone at 0°C. Sections were rehydratedfor 30 min in 5 % (w/v) BSA in Tris-saline (lOmM-Tris, pH7-2) and incubated for 2-3 h withanti-bmg IgG, 0"2mgml~' in 5 % BSA (w/v) in Tris-saline, which had been preincubated for30 min before the addition of 0-01 % (v/v) NP40. Slides were spray-rinsed for 1 min and thensoaked in 600ml Tris-saline for 30-45 min before incubation with FIRaSh, 0-1 mgml"1 in 2-5 %(w/v) BSA/50% (V/V) rabbit serum in Tris-saline, which had been preincubated for 30 minbefore the addition of 0-01 % (v/v) NP40. Slides were then spray-rinsed and soaked in Tris-salinefor 4 h before mounting and viewing as described above. Control incubations were performed with0 ' 2 m g m r ' preimmune IgG and with FIRaSh alone to assess non-specific binding. The stainingspecificity of anti-bmg IgG was checked by preincubating both the antibody and the section withexcess purified lectin (0-2mgml~'). Results were recorded on Kodak Ektachrome (daylight) 400ASA and plates were prepared by copying the colour slides on a Bowen's Illumitran 3, through aWratten no. 63 filter, onto Ilford Pan F to improve the distinction on black and white printsbetween the green fluorescence and the red background caused by the Evan's Blue.

An endogenous lectin in erythropoiesis 203
Extraction of the lectin from bone marrowAll procedures were carried out on ice or at 4°C. Extracellular lectin was isolated by gently
disaggregating bone marrow in 10 ml LM/EDTA/NaCl per rabbit (0-3M-lactose in lmll/J-mercaptoethanol,2mM-EDTA (pH7-4), 0-15 M-NaCl) containing 1 mM-phenylmethylsulphonylfluoride. Cells were pelleted at 1000g for 5min and washed again with 10 ml LM/EDTA/NaCl.Pooled washes were centrifuged at 26 500 £ for 15min and the supernatants recentrifuged at122500# for 2h. Intracellular lectin was extracted by lysis of washed bone marrow cells,resuspended in a minimum of LM/EDTA/NaCl and ground in a pestle and mortar with coarsesilver sand under liquid nitrogen. After thawing, the supernatant was diluted to 10 ml and the sandremoved by centrifugation at 800 £ for 1 min then washed in 10 ml LM/EDTA/NaCl. Lysate andwash were pooled and centrifuged as for the cell washes above. Centrifugation pellets wereextracted with either 0-1 % SDS (w/v) or 0-1 % (v/v) Triton X-100 in LM/EDTA/NaCl. Theextent of cell lysis during each procedure was estimated by comparison of the haemoglobin contentof each extraction with the total tissue haemoglobin, determined by the maximal absorption ofsupernatants at 380-440 nm (approx. 418 nm) relative to that of a rabbit haemoglobin standard.
For the quantitative determination of intra- and extracellular lectin alternative homogenizationconditions were used. Marrow was extracted, pooled and mixed by gentle stirring. Two 1-gsamples were weighed and one was washed with LM/EDTA/NaCl as described above. Wholetissue and washed bone marrow cells were then pulverized in an anvil press precooled with 'cardice'and liquid nitrogen and then the frozen wafer of tissue was homogenized in 5 ml LM/EDTA/NaClin an Ultra-Turrax homogenizer for four 15-s periods. Approximately 1 ml volumes of blood wereeither allowed to clot or collected into 5 ml LM/EDTA/NaCl and weighed. Lectin was extractedfrom whole blood (clot) or from cells collected into LM/EDTA/NaCl and washed once, as for themarrow cells. Haemoglobin concentrations were determined spectrophotometrically.
RESULTS
Biochemical studiesExtracellular lectin, purified from lactose washes of marrow cells, as previously
described (Harrison et al. 1984), was identical in molecular weight, isoelectric point,lectin activity and immunological reactivity to intracellular lectin purified fromlysates of lactose-washed cells. Since minor isoforms of bone-marrow galaptin havepreviously been identified in crude lactose washes of bone marrow and in prep-arations of galaptin after purification (FitzGerald et al. 1984), intracellular lectinisoforms were compared with extracellular lectin by isoelectricfocusing followed byWestern blotting. Haemoglobin determinations indicated that lysis of 20-23 %marrow cells occurred during lactose washing, thereby releasing a proportion of theinternal lectin into the washes. Nevertheless, staining the nitrocellulose blots withanti-bmg (Fig. 1) suggested that the isoform of pi 5-6 was predominant both intra-and extracellularly in each of four rabbits investigated. Two other isoforms, at pi 5-2and 5-9, were also detectable in each lactose extract. Biochemical studies thereforeindicated no major difference between the internal and external lectin.
To quantify intra- and extracellular lectin, single radial immunodiffusion assayswere carried out on the crude lactose washes and cell lysates of normal and anaemicblood and marrow. Comparative experiments showed that grinding with sand andliquid nitrogen to lyse the marrow cells depleted the extracts of lectin, but adequatelysis was not achieved by osmotic shock or freeze—thawing procedures. To extractcytoplasmic lectin washed marrow cells were therefore pulverized in a precooledanvil press and subsequently homogenized as described in Materials and Methods.

204 F. L. Harrison andjf. W. Catt
Fig. 1. Western blot of an isoelectricfocusing gel of intra- and extracellular galaptin fromthe bone marrow of four anaemic rabbits. The three isoforms of bone marrow galaptin,pi 5-2, 5-65 and 5-95, are similarly abundant in all preparations. 1, Bone marrow galaptinpurified from lactose washes of marrow cells (1 fig): 2,4,6,8, intracellular lectin from ap-proximately 2 mg marrow;3,5,7,9, extracellular lectin from approximately 2 mg marrow.
Insoluble material, pelleted on centrifugation of crude lactose extracts, was extractedwith SDS or Triton X-100 and subjected to SDS-polyacrylamide gel electro-phoresis. Only trace amounts of galaptin were detected, indicating efficient release ofthe lectin by these extraction procedures. To assess the relative distribution of thelectin, we compared the galaptin content of washed cells with that of wholeundissociated marrow. Circulating erythrocytes from normal rabbits and reticulo-cytes from anaemic rabbits were collected in duplicate samples, and lectin levels inwhole blood and lactose-washed cells were then determined as for the marrowsamples. Results are presented in Tables 1 and 2.
No significant lysis of circulating blood cells occurred during lactose-washingprocedures but there was inevitably some lysis of bone marrow cells on dis-aggregation and washing. Histological staining of cytocentrifuge slides indicated thatthe relative proportions of different cell types in the marrow remained constantduring washing procedures, suggesting that there was not significant differential lysisof particular cell types. We therefore used haemoglobin as an intracellular markerand took the % haemoglobin lost from duplicate marrow samples during dissociationand washing as an estimate of the total cell lysis. The extent of lysis varied between 18and 36 % for normal marrow samples and 25 and 56 % for anaemic marrow. This

An endogenous lectin in erythropoiesis 205
unavoidable lysis resulted in a loss of intracellular lectin from the washed cells. Toaccount for this we expressed the lectin concentrations in both lactose extracts interms of /ig lectin mg"1 haemoglobin. This procedure permits direct comparisonof the experimentally determined lectin concentrations in washed cells and undis-sociated tissue, and also corrects for any differences between duplicate tissue samplesand any losses from either sample during preparation procedures. To compare intra-and extracellular lectin concentrations a least-squares linear regression of total lectinin undissociated marrow (/igmg"1 haemoglobin) on intracellular lectin in washedcells (^gmg"1 haemoglobin) was performed. In all cases intercepts were not sig-nificantly different from zero, so slopes through the origin were estimated (Table 3).In circulating erythrocytes and reticulocytes there were no significant differencesbetween total and intracellular lectin concentrations (i.e. slope = 1), indicating thatthere is very little extracellular lectin in serum or bound to the cell surface. However,in normal and anaemic marrow significant differences were observed (i.e. slopes,which are equivalent to mean ratios of total: intracellular lectin concentration, were1-25 and 1-34, respectively). These differences indicate a partially extracellulardistribution of the galaptin. In terms of the total lectin content, in normal marrowapproximately 25 % and in anaemic marrow approximately 20 % of the total lectin isextracellular. In fact, the intercepts and slopes for regression of total and intra-
Table 1. Determination of total and intracellular galaptin in circulating blood cells
Status
Normal
Anaemic
mgHb/g
Total
66-7 ±1-8
69-0 ±3-069-6 ±1-9
74-7 ±5-0
26-4 ±0-6
21-9± 1-3
22-5 ±0
39-8 ±0
20-6 ±0
tissue
Washed cells
73-8 ±0-2
69-7 ±5-277-2 ±5-8
67-9 ±5-7
26-611-0
22-4 ±1-3
19-5 ±0-8
31-8±0-9
17-7 ±0-4
/^ggalaptin/g blood
Total
131-9 ±7-6102-0 + 11-5135-6 ±5-5195-6 ±10-4213-0+ 11-2218-5 ±8-0230-3 ±10-3
35-4±5-458-1 ±8-924-1 ±6-235-8 ±6-522-6 ±5-346-415-3
105-118-3122-813-141-819-945-713-7
Washed cells
182-91095-015-5
141-419-6198-016-6197-914-2199-5110-0205-517-1
39-014-753-511-130-213-741-815-2
n.d.49-012-575-018-092-215-332-219-231-518-0
/ig galaptin/mg Hb
Total
1-991-452-002-813-062-923-08
4-342-211-101-641-02-072-643-082-022-21
Washed cells
2-481-252-022-562-562-943-03
1-472-011-351-86
2-692-352-901-821-77
Haemoglobin (Hb) was determined spectrophotometrically; data represent the average of two determi-nations. Galaptin concentrations were determined by SRID assay. Paired data represent mean values oftwo independent assays, performed in triplicate, i standard deviation. In anaemic animals circulatingblood cells were 92-98 % reticulocytes. n.d., not determined.

206 F. L. Harrison andjf. W. Catt
cellular lectin were not significantly different for normal and anaemic marrow. Thecombined slope through the origin (1-311 ± 0-063) indicates that 76 ± 4% of totallectin is intracellular in both normal and anaemic animals. This suggests thatanaemia induced by phenylhydrazine treatment does not cause any major redistri-bution of the lectin. From these data the quantitites of intracellular lectin per
Table 2. Determination of total and intracellular galaptin in bone marrow
Status
Normal
Anaemic
mgHb/g
Total
7-5 ±0-2
ll-8±0-4
7-0 ±1-0
10-2 ±0-2
8-3 ±0-4
8-8 ±0-7
l l - 5 ± 0
6-7 ± 0
7-6 ± 0
tissue
Washed cells
4-8 ±0-2
6-7 ±0-1
5-4 ±0-5
5-6±0-2
6-8 ±0-3
8-8 ±0-4
7-4±0-l
5-3 ±0-2
5-1 ± 0 1
fig galaptin/g blood
Total
1101 ± 5-5l l l -2± l -9133-6 ±9-0104-3 + 6-393-3 ±9-477-6 ±7-0
116-5 ±7-473-4±l-0
137-5 ±3-5115-3 ±10-5115-0 ±3-7106-7 ±4-3124-2 ±14-7162-9 ±7-6176-9 ±6-7138-1 ±0-3151-6±9-6116-4 ±5-4
Washed cells
43-4±5-056-0 ±2-069-9 ±12-451-8±9-845-7 ±3-841-913-766-8 ±0-535-9 ±0-4
76-8 ±6-271-2±5-l64-9 ±15-865-812-644-917-386-815-585-8113-064-5110-694-3114-277-611-4
fig galaptin/mg Hb
Total
14-6414-8011-318-84
13-2911-0611-447-21
16-4813-8213-0521-1110-9114-2926-2120-4919-8515-25
Washed cells
9-1711-8310-457-778-497-79
11-936-45
11-2410-587-367-466-08
11-7416-4212-3518-5515-27
Haemoglobin (Hb) was determined spectrophotometrically; data represent the average of two determi-nations. Galaptin concentrations were determined by SRID assay. Paired data represent mean values oftwo independent assays, performed in triplicate, i standard deviation. The average cellular constitutionof normal marrow was 20—30 % erythroblast, 30—55 % reticulpcyte and 20—40 % myeloid, and of anaemicmarrow, 60-70% erythroblast, 10—15% reticulocyte and 20-30% myeloid.
Table 3. Comparison of total and intracellular lectin concentration in circulatingblood cells and in bone marrow, in normal and anaemic rabbits
Tissue Intercept Slope through origin
Circulating erythrocytes (8)Circulating reticulocytes (10)Normal marrow (8)Anaemic marrow (10)
0-2010-550-0510-484-2913-345-8813-11
1-023610-04720-997610-05181-250810-08421-337710-0904
Data were compared by least-squares linear regression of total lectin concentration in intracellu-lar lectin concentration, (n), number of experimental values. Lectin concentrations determinedby SRID assay were corrected for accidental lysis by expressing the data as fig galaptin/mghaemoglobin (see Tables 1 and 2, and Results). Results are given 1 S.E. Degrees of freedom usedin standard errors of parameters = n—2 for intercepts and n— 1 for slopes through the origin.
An endogenous lectin in erythropoiesis
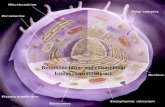
Fig. 2. Indirect immunofluorescent staining of semi-thin section of resin-embeddednormal bone marrow. Bar, 20/fln. After fluorescent staining (A) the section was counter-stained with Toluidine Blue to enable identification of cell type (B);/, fat cell; arrowhead,eosinophil; ey, erythrocytes; e, predominantly erythroid area; m, predominantly myeloidarea. Fluorescent staining (A) shows the galaptin is restricted to certain areas of themarrow, most of which can be identified as erythroid, e.g. e. Predominantly myeloidareas do not contain the lectin. A Y-shaped area of connective tissue (arrow), in A andB, clearly contains galaptin. Fluorescent staining of eosinophils and erythrocytes isartifactual, as discussed in the text.
erythroblast and per circulating cell can be estimated to be 80—170fg/erythroblast,50 fg/reticulocyte and 30fg/erythrocyte, which indicates that there are of the orderof 4x 106 molecules within an erythroblast, decreasing to 1-5X106 molecules withincirculating erythrocytes.
Immunocytochemistry
Sections of resin-embedded bone marrow show the complex multicellular organ-ization of the tissue (Fig. 2B). Large fat cells (/) are prominent and megakaryocytes,eosinophils and neutrophils can also be readily identified. Erythroid (e) and myeloidcells (m) differentiate in clusters within the marrow matrix. Eosinophils (arrowhead)and erythrocytes (ey) seen on sections were fluorescent in all incubations, even when

208 F. L. Harrison andjf. W. Catt
incubated with FIRaSh alone. This non-specific fluorescence has been reported foreosinophils (Zucker-Franklin et al. 1981), which are autofluorescent and also bindthe fluorochrome, FITC. The fluorescence of erythrocytes results from the fixationprocedure, since circulating erythrocytes fixed with 8 % formaldehyde exhibit a highdegree of non-specific staining on cytocentrifuge slides whereas cytocentrifugederythrocytes fixed in methanol show virtually no fluorescence in control incubations.All other cell types on sections showed only a very diffuse background staining incontrol incubations. The pattern of fluorescent staining seen with anti-bmg IgG(Fig. 2A) indicates that the galaptin is restricted to particular areas of the marrow,some of which can be morphologically identified as erythroid, though at the light-microscopic level it is not possible to classify each cell; in particular, early myeloidand erythroid blast cells cannot be distinguished. Erythroblasts at later stages ofdifferentiation appeared to contain the lectin but recognizable myeloid cells did not.Unfortunately, cell surface lectin could not be clearly distinguished in the sectionsbecause of the high intracellular fluorescence and the close packing of the cells withinthe marrow matrix; however, lectin appeared to be extracellular in a Y-shaped area ofcellular material, clearly stained by anti-bmg IgG (Fig. 2A, arrow) and by ToluidineBlue (Fig. 2B, arrow).
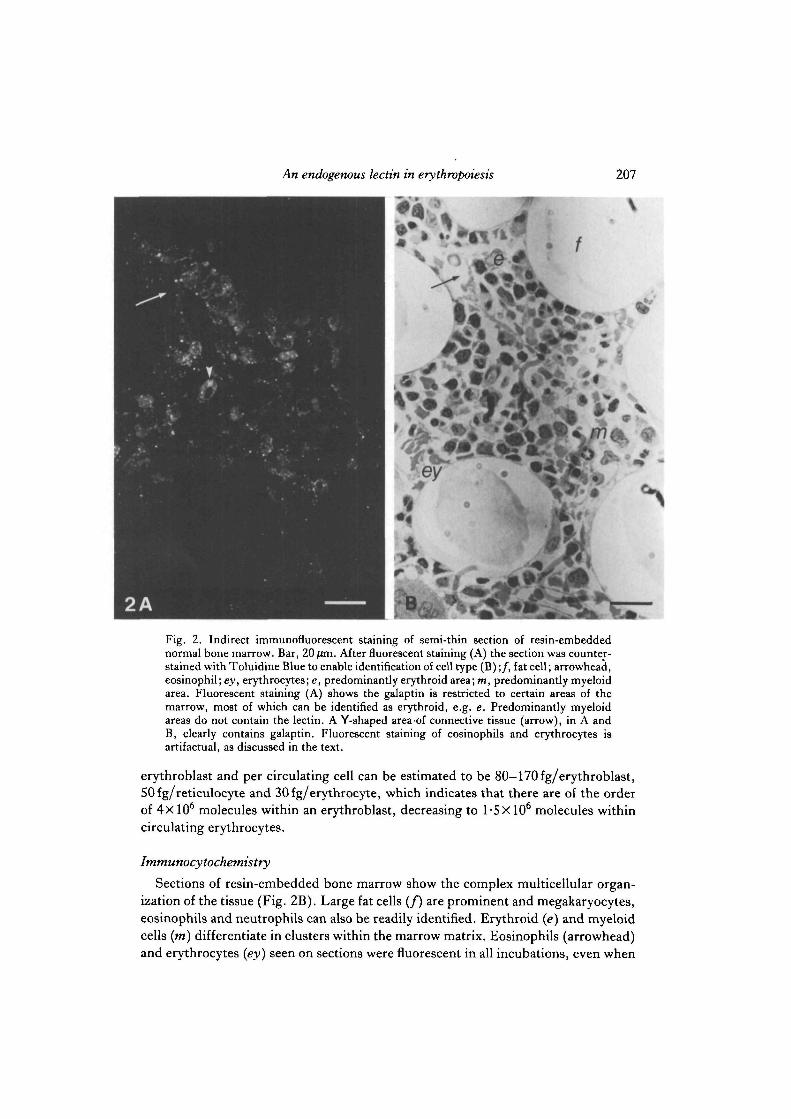
To identify positively those cells showing intracellular lectin cytocentrifuge slidesof gently disaggregated marrow were prepared, and the cells were permeabilized bymethanol fixation. Fig. 3A,B shows that the galaptin is intracellular in erythroblastsbut not in myeloid cells (m). Cells treated with preimmune IgG showed nofluorescent staining (Fig. 3C,D). Occasionally erythroblastic islands were seen,containing a central macrophage surrounded by erythroblasts, and staining showedlectin within and possibly between erythroblasts but not in the cytoplasm of themacrophage nurse cell (n) (Fig. 3E,F). The fluorescent staining becameconcentrated around the nucleus in maturing erythroblasts until, at enucleation, aring of fluorescence surrounded the extruded nucleus and the reticulocyte cytoplasmshowed only weak diffuse staining (Fig. 4A,B). Light-microscopic techniquescannot distinguish whether the lectin is on the external surface or within the smallamount of cytoplasm surrounding the extruded nucleus. Circulating reticulocytesand erythrocytes showed very faint diffuse intracellular staining when cytocentrifugeslides were prepared and stained as described above (data not shown).
DISCUSSION
In spite of their relative abundance the precise functions of lectins in situ remainuncertain. A role in intercellular recognition and adhesion has been proposed forlectins in several other systems, but contradictory evidence has also been presented.A setback has been the discovery that the majority of the lectin is located intra-cellularly. In aggregating slime moulds the extracellular lectin has been estimated tobe as little as 0-2% (Springer et al. 1980) or even 0-02% (Bartlesef al. 1982) of thetotal cellular lectin, and Erdos & Whitaker (1983) were unable to detect any immuno-cytochemically reactive lectin on the cell surface of Dictyostelium discoideum. In the
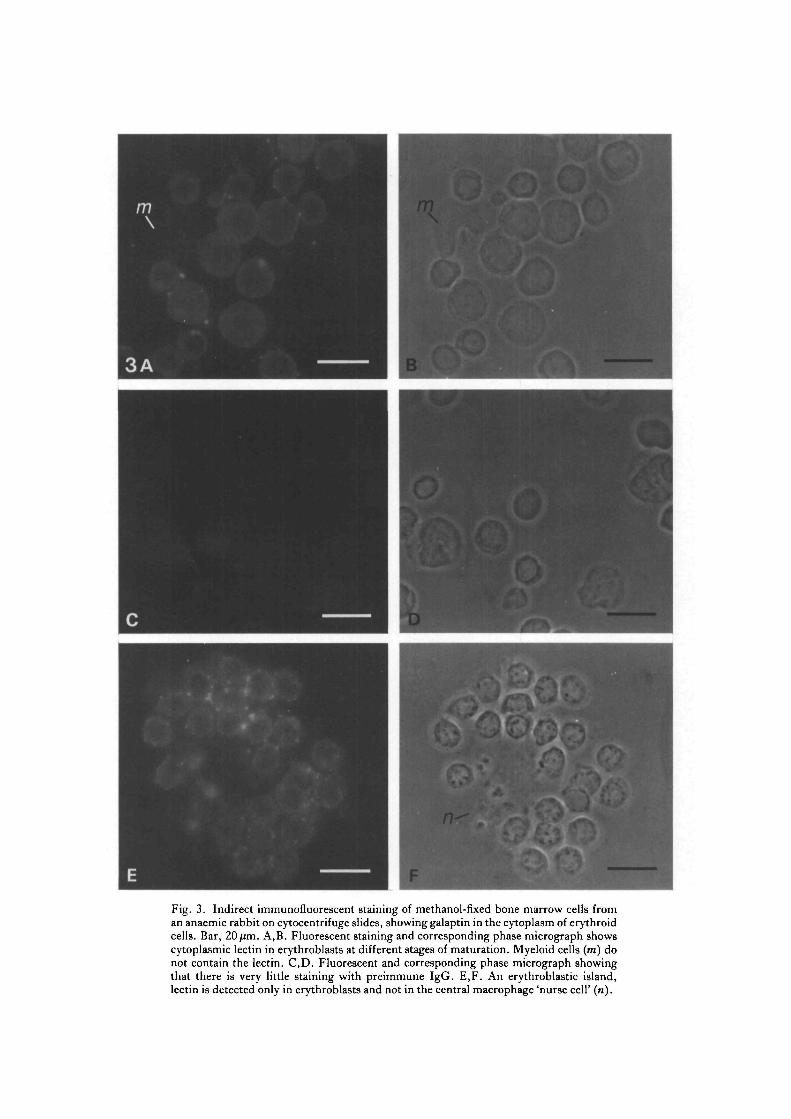
Fig. 3. Indirect immunofluorescent staining of methanol-fixed bone marrow cells froman anaemic rabbit on cytocentrif uge slides, showing galaptin in the cytoplasm of erythroidcells. Bar, 20/im. A,B. Fluorescent staining and corresponding phase micrograph showscytoplasmic lectin in erythroblasts at different stages of maturation. Myeloid cells (m) donot contain the lectin. C,D. Fluorescent and corresponding phase micrograph showingthat there is very little staining with preimmune IgG. E,F. An erythroblastic island,lectin is detected only in erythroblasts and not in the central macrophage 'nurse cell' (n).
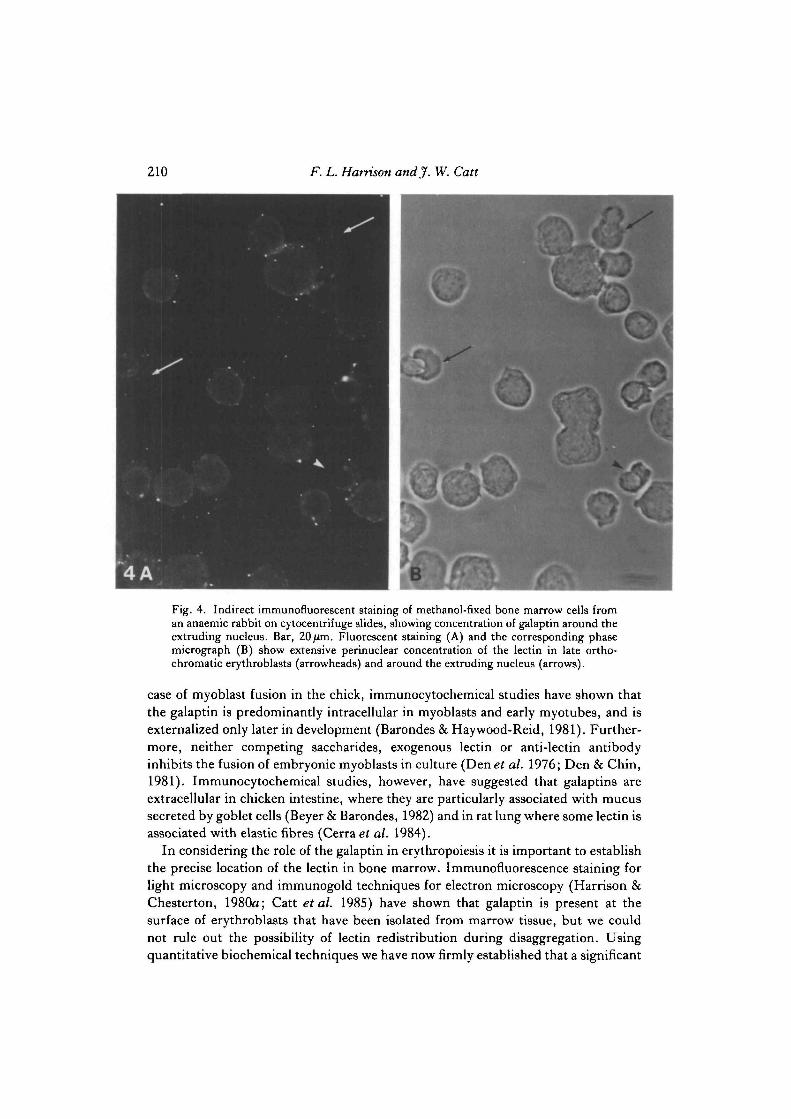
210 F. L. Harrison andjf. W. Catt
Fig. 4. Indirect immunofluorescent staining of methanol-fixed bone marrow cells froman anaemic rabbit on cytocentrifuge slides, showing concentration of galaptin around theextruding nucleus. Bar, 20/im. Fluorescent staining (A) and the corresponding phasemicrograph (B) show extensive perinuclear concentration of the lectin in late ortho-chromatic erythroblasts (arrowheads) and around the extruding nucleus (arrows).
case of myoblast fusion in the chick, immunocytochemical studies have shown thatthe galaptin is predominantly intracellular in myoblasts and early myotubes, and isexternalized only later in development (Barondes & Haywood-Reid, 1981). Further-more, neither competing saccharides, exogenous lectin or anti-lectin antibodyinhibits the fusion of embryonic myoblasts in culture (Dene* al. 1976; Den & Chin,1981). Immunocytochemical studies, however, have suggested that galaptins areextracellular in chicken intestine, where they are particularly associated with mucussecreted by goblet cells (Beyer & Barondes, 1982) and in rat lung where some lectin isassociated with elastic fibres (Cerraef al. 1984).
In considering the role of the galaptin in erythropoiesis it is important to establishthe precise location of the lectin in bone marrow. Immunofluorescence staining forlight microscopy and immunogold techniques for electron microscopy (Harrison &Chesterton, 1980a; Catt et al. 1985) have shown that galaptin is present at thesurface of erythroblasts that have been isolated from marrow tissue, but we couldnot rule out the possibility of lectin redistribution during disaggregation. Usingquantitative biochemical techniques we have now firmly established that a significant

An endogenous lectin in erythropoiesis 211
proportion of the galaptin, approximately 25 %, is indeed extracellular in rabbit bonemarrow. The origin of the extracellular lectin has not been determined but it appearsto be identical to that found within erythroblasts. Using resin-embedded sections toobserve the overall distribution of the lectin in marrow we found that galaptin isassociated with discrete areas of cells, probably the sites of erythroid differentiationand also with some acellular areas of extracellular matrix. This distribution is similarto that observed in other rabbit tissues where immunocytochemistry suggests thatthe lectin is both intra- and extracellular, and is particularly prominent surroundingdifferentiating cells, for example in intestinal crypts and hair follicles in the skin(Catt & Harrison, 1985a). Increasing evidence suggests that matrix components playan important role in the regulation of cellular differentiation in marrow (Zuckerman& Wicha, 1983; Spooncer et al. 1983). The galaptin is presumably not involvedin the induction of erythropoiesis, as erythroblasts themselves seem to be majorproducers of the lectin, but the lectin may play a role in the maintenance andregulation of the very high rates of erythrocyte production (about 150 millionerythrocytes min"1 in human marrow; Dexter, 1984), perhaps by mediating cell-cell or cell-matrix associations during maturation. We are now investigating thesepossibilities using bone marrow cultures where differentiation can be observedand experimentally manipulated. At enucleation, when the maturing erythroblastis released from its close association with the nurse cell, a ring of fluorescencesurrounds the extruded nucleus, while residual lectin in the reticulocyte remainsintracellular. Though electron-microscopic studies are required to confirm that theendogenous lectin is actually associated with the plasma membrane surrounding theextruded nucleus, such a differential distribution of the lectin at enucleation wouldbe of obvious relevance both to the phagocytosis of extruded nuclei and to the releaseof reticulocytes into the circulation.
We are grateful to Sally Martin and Sue Carleton for technical assistance, to Dr John Patersonfor his invaluable assistance with the statistical treatment of the data and to Peter Taylor forphotographic advice. J. W. Catt is a Research Fellow supported by the Cancer ResearchCampaign.
REFERENCESBARONDES, S. H. & HAYWOOD-REID, P. L. (1981). Externalization of an endogenous chicken
muscle lectin with in vivo development. J. CellBiol. 91, 568-572.BARTLES, J. R., FRAZIER, W. A. & ROSEN, S. D. (1982). Slime mould lectins. In Int. Rev. Cytol
(ed. G. H. Bourne & J. F. Danielli), vol. 75, pp. 61-93. New York: Academic Press.BEYER, E. C. & BARONDES, S. H. (1982). Secretion of an endogenous lectin by chicken intestinal
goblet cells. J. CellBiol. 92, 28-33.CATT, J. W. & HARRISON, F. L. (1985a). Selective association of an endogenous lectin with
connective tissues. J. Cell Sci. 73, 347-359.CATT, J. W., PEACOCK, M. A. & HARRISON, F. L. (1985). Surface localization of an endogenous
lectin in rabbit bone marrow. Histochem.J. 17, 189-199.CERRA, R. F., HAYWOOD-REID, P. L. & BARONDES, S. H. (1984). Endogenous mammalian lectin
localised extracellularly in lung elastic fibres. J. CellBiol. 98, 1580-1589.DEN, H. & CHIN, J. H. (1981). Endogenous lectin from chick embryo skeletal muscle is not
involved in myotube formation in vitro. J. Biol. Chem. 256, 8069-8073.

212 F. L. Harrison andj. W. Catt
DEN, H., MALJNZAK, D. A. & ROSENBERG, A. (1976). Lack of evidence for the involvement of a /3-D-galactosyl-specific lectin in the fusion of chick myoblasts. Btochem. Btophys. Res. Commun. 69,621-627.
DEXTER, T. M. (1982). Stromal cell associated haemopoiesis. J. cell. Phys. Suppl. J, 89-94.DEXTER, T. M. (1984). The message in the medium. Nature, Land. 309, 746-747.ERDOS, G. W. & WHTTAKER, D. (1983). Failure to detect immunocytochemically reactive
endogenous lectin on the cell surface of Dictyostelium discoideum.J. CellBiol. 97, 993-1000.FITZGERALD, J. W., CATT, J. W. & HARRISON, F. L. (1984). Isoforms of an endogenous lectin in
rabbit bone marrow. Eur.J. Btochem. 140, 137-141.HARRISON, F. L. & CATT, J. W. (1985). An endogenous /3-galactoside specific lectin is both intra-
and extra-cellular during erythropoiesis. Biochem. Soc. Trans. \3, 1116-1117.HARRISON, F. L. & CHESTERTON, C. J. (1980a). Erythroid developmental agglutinin is a protein
lectin mediating specific cell-cell adhesion between differentiating rabbit erythroblasts. Nature,Lond. 286, 502-504.
HARRISON, F. L. & CHESTERTON, C. J. (19806). Factors mediating cell-cell recognition andadhesion. Galaptins, a new class of bridging molecules. FEBS Letts 122, 157-165.
HARRISON, F. L., FITZGERALD, J. E. & CATT, J. W. (1984). Endogenous /S-galactoside-specificlectins in rabbit tissues. J . Cell Set. 72, 147-162.
SPOONCER, E., GALLAGHER, J. T., KRIZSA, F. & DEXTER, T. M. (1983). Regulation of
haemopoiesis in long-term bone marrow cultures. IV. J. CellBiol. 96, 510-514.SPRINGER, W. R., HAYWOOD, P. L. & BARONDES, S. H. (1980). Endogenous cell surface lectin in
Dictyostelium: quantitation elution by sugar, and elicitation by divalent immunoglobulin. J . CellBiol. 87, 682-690.
ZUCKER-FRANKLTN, D., GREAVES, M. F., GROSSI, C. E. & MARMONT, A. M. (eds) (1981). Atlas
of Blood Cells, Function and Pathology, vol. 1. Philadelphia, USA: Lea & Febiger.ZuCKEKMAN, K. S. & WICHA, M. S. (1983). Extracellular matrix production by the adherent cells
of long-term bone marrow cultures. Blood 61, 540-547.
(Received 30 October 1985 -Accepted, in revised form, 7 April 1986)