
Innhold - Universitetsforlaget · INNHOLD 9 Restitusjon er en læringsprosess . . . . . . . . . . ....
49
Forord . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21 Innledning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 Et fugleperspektiv . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 Hvordan undersøkes nervesystemets bygning og funksjon? . . . . . . . . . . . . . . . . . . . . . 24 Dyreeksperimenter har i særlig grad bidratt til økt kunnskap . . . . . . . . . . . . . 25 Dyreeksperimenter og etikk . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25 Alle metoder har feilkilder . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26 Vitenskapelige «sannheter» må stadig revideres . . . . . . . . . . . . . . . . . . . . . . . 26 D EL 1 H OVEDTREKK I NERVESYSTEMETS BYGNING OG FUNKSJON K APITTEL 1 Nervesystemets bygningselementer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29 Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29 Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29 Nervecellen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30 Nevroner har lange utløpere . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30 Dendritter er utstyrt med spinae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30 De fleste nevroner er multipolare . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32 Kommunikasjon mellom nerveceller foregår ved synapser . . . . . . . . . . . . . . . . . . 33 To hovedtyper av nevroner: projeksjonsnevroner og internevroner . . . . . . . . . . . . 35 Mange aksoner er ekstra isolert for å øke ledningshastigheten . . . . . . . . . . . . . . . . 37 Grå og hvit substans . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37 Nervecellene ligger ordnet i kjerner og ganglier . . . . . . . . . . . . . . . . . . . . . . . . . . . 37 Aksoner danner baner og nerver . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38 Sammenkobling av nevroner og signalveier . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39 Divergens, konvergens og summasjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39 Parallelle signalveier og resiproke forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . 41 Innhold
Transcript of Innhold - Universitetsforlaget · INNHOLD 9 Restitusjon er en læringsprosess . . . . . . . . . . ....

Forord
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
Innledning
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
Et fugleperspektiv . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23Hvordan undersøkes nervesystemets bygning og funksjon? . . . . . . . . . . . . . . . . . . . . . 24
Dyreeksperimenter har i særlig grad bidratt til økt kunnskap . . . . . . . . . . . . . 25Dyreeksperimenter og etikk . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25Alle metoder har feilkilder . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26Vitenskapelige «sannheter» må stadig revideres . . . . . . . . . . . . . . . . . . . . . . . 26
D
EL
1 H
OVEDTREKK
I
NERVESYSTEMETS
BYGNING
OG
FUNKSJON
K
APITTEL
1 Nervesystemets bygningselementer
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29Nervecellen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
Nevroner har lange utløpere . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30Dendritter er utstyrt med spinae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30De fleste nevroner er multipolare . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32Kommunikasjon mellom nerveceller foregår ved synapser . . . . . . . . . . . . . . . . . . 33To hovedtyper av nevroner: projeksjonsnevroner og internevroner . . . . . . . . . . . . 35Mange aksoner er ekstra isolert for å øke ledningshastigheten . . . . . . . . . . . . . . . . 37Grå og hvit substans . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37Nervecellene ligger ordnet i kjerner og ganglier . . . . . . . . . . . . . . . . . . . . . . . . . . . 37Aksoner danner baner og nerver . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
Sammenkobling av nevroner og signalveier . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39Divergens, konvergens og summasjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39Parallelle signalveier og resiproke forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
Innhold
0000 UFb SentrNerv 070401M.book Page 5 Wednesday, July 4, 2007 12:11 PM

6 INNHOLD
Forbindelser mellom de to sidene av sentralnervesystemet . . . . . . . . . . . . . . . . . 41Enkeltnevroner inngår i nevrale nettverk . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
Celleskjelettet og aksonal transport . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43Celleskjelettets komponenter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
Gliaceller . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45Astroglia står i kontakt med kapillærer, cerebrospinalvæsken og nevroner . . . . . 46Astroglia og kontroll av nervecellenes miljø (homøostase) . . . . . . . . . . . . . . . . . 47Myelinskjeder dannes av oligodendroglia og schwannceller . . . . . . . . . . . . . . . . 48Ranvierske innsnøringer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49Umyeliniserte aksoner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50Mikroglia er fagocytter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50Sentralnervesystemets reaksjon på skade og betennelse . . . . . . . . . . . . . . . . . . . . 51Perifere nerver er bygd for å beskytte aksonene . . . . . . . . . . . . . . . . . . . . . . . . . . 52
K
APITTEL
2 Nervecellenes funksjonelle egenskaper
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55Grunnlaget for nervecellers eksitabilitet og impulsledning . . . . . . . . . . . . . . . . . . . . . 56
Ionekanalene bestemmer cellemembranens permeabilitet . . . . . . . . . . . . . . . . . . 56Membranpotensialet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57Også anioner er skjevfordelt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58Transmitterstyrte ionekanaler . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 59Spenningsstyrte ionekanaler . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60Ionekanalenes struktur . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61Endringer av membranpotensialet – depolarisering og hyperpolarisering . . . . . . 61Aksjonspotensialet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62Hvor oppstår aksjonspotensialet? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63Frekvenskoding og impulsmønster: hvordan nervecellene varierer informasjonen de formidler . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63Refraktærperioden . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64Ledning av aksjonspotensialet i aksoner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 65Ledningshastighet i myeliniserte og umyeliniserte aksoner . . . . . . . . . . . . . . . . . 66
Synapsenes virkemåte . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 66Sammendrag og viktige begreper . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 67Frisetting av nevrotransmittere . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68Nevrotransmittere frisettes i kvanter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70Inaktivering av nevrotransmittere . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71Mekanismer for EPSP og IPSP . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71Langsomme synaptiske virkninger modulerer effekten av raske . . . . . . . . . . . . . 72Summasjon og integrasjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 73Synapsers plassering har funksjonell betydning . . . . . . . . . . . . . . . . . . . . . . . . . . 73Aksoaksoniske synapser formidler presynaptisk kontroll av transmitterfrisetting 75Hvorfor trengs det inhibitoriske synapser? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 75Synaptisk plastisitet – bruksavhengige endringer av struktur og funksjon . . . . . . 76Betingelser for synaptisk plastisitet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 77Former for synaptisk plastisitet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 77
Nevrotransmittere . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 78Hva skal til for å klassifisere et stoff som nevrotransmitter? . . . . . . . . . . . . . . . . 79Samlokalisering av nevrotransmittere . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81Transmitterreseptorer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82Syntese av nevrotransmittere . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83
0000 UFb SentrNerv 070401M.book Page 6 Wednesday, July 4, 2007 12:11 PM

INNHOLD 7
Eksitatoriske transmitteraminosyrer: glutamat og aspartat . . . . . . . . . . . . . . . . . . . 83Glutamatsyntese . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 84Glutamatreseptorer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 85Glutamattransportører . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 85Inhibitoriske transmitteraminosyrer: GABA og glysin . . . . . . . . . . . . . . . . . . . . . . 87GABA-reseptorer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87Acetylkolin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88Acetylkolinreseptorer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 89Biogene aminer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 90Virkninger av biogene aminer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 90Monoaminene: noradrenalin, dopamin og serotonin . . . . . . . . . . . . . . . . . . . . . . . 91Syntese av biogene aminer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91Monoaminreseptorer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91Monoamintransportører og monoaminoksidase . . . . . . . . . . . . . . . . . . . . . . . . . . . 92Monoaminer, depresjon, sårbarhet og plastisitet . . . . . . . . . . . . . . . . . . . . . . . . . . . 92Modulerende transmittere: fellestrekk og spesialiseringer . . . . . . . . . . . . . . . . . . . 93ATP og adenosin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 94NO . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 94Nevropeptider . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 95Generelt om medikamenters virkninger på nervesystemet . . . . . . . . . . . . . . . . . . . 96
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 97
K
APITTEL
3 Nervesystemets forskjellige deler
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 99
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 99Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 99Medulla spinalis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 100
Spinalnerver forbinder ryggmargen med kroppen . . . . . . . . . . . . . . . . . . . . . . . . . 101Ryggmargen er delt inn i segmenter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 102Ryggmargen består av grå og hvit substans . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 102Hovedtyper av nevroner i ryggmargen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 103Efferente fibre fra ryggmargen styrer muskler og kjertler . . . . . . . . . . . . . . . . . . . 104Sensoriske celler i ryggmargen påvirkes fra dorsalroten og sender signaler til hjernen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 105Internevroner sørger for samarbeid mellom cellegrupper i ryggmargen . . . . . . . . 106Ryggmargens grå substans kan deles inn i soner – Rexeds laminae . . . . . . . . . . . . 106Spinalnervene deler seg i grener . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 107Ryggmargen består av enheter som styres fra hjernen . . . . . . . . . . . . . . . . . . . . . . 107
Hjernestammen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 107Hjernestammen inneholder 3. og 4. ventrikkel . . . . . . . . . . . . . . . . . . . . . . . . . . . . 107Hjernenervene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 109Retikulærsubstansen strekker seg gjennom sentrale deler av hjernestammen . . . . 109Medulla oblongata . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 110Pons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 111Medulla oblongata og pons sett bakfra . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 112Mesencephalon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 112Diencephalon omfatter thalamus og hypothalamus . . . . . . . . . . . . . . . . . . . . . . . . 113
Cerebrum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 116Storhjernens overflate er sterkt foldet og danner gyri og sulci . . . . . . . . . . . . . . . . 117Storhjernen deles inn i fire lapper . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 117Cortex cerebri er bygd opp av seks cellelag . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 119Hjernebarken inndeles i areae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 119
0000 UFb SentrNerv 070401M.book Page 7 Wednesday, July 4, 2007 12:11 PM

8 INNHOLD
Basalgangliene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 120Cerebellum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 121Sentralnervesystemets hinner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 122Ventrikkelsystemet og cerebrospinalvæsken . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 124
Ventriklenes beliggenhet og form . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 124Cerebrospinalvæsken dannes av årefletninger i ventriklene . . . . . . . . . . . . . . . . . 125Cerebrospinalvæskens sammensetning og funksjon . . . . . . . . . . . . . . . . . . . . . . . 126Cerebrospinalvæskens sirkulasjon og avløp . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 127
Sentralnervesystemets blodforsyning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 128Regulering av cerebral sirkulasjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 128Blod–hjerne-barrieren . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 130Blod–hjerne-barrieren mangler enkelte steder i hjernen . . . . . . . . . . . . . . . . . . . . 131Hjernen forsynes med blod fra arteria carotis interna og arteria vertebralis . . . . . 133Forbindelser mellom grener av arteria carotis og arteria vertebralis . . . . . . . . . . . 135Ryggmargen mottar blod fra mange nivåer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 135Hjernens venøse blod samles i sinuser og forlater skallen i vena jugularis interna 135
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 137
K
APITTEL
4 Nervesystemets utvikling, aldring og restitusjon etter skader
. . . . . . . . . 139
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 139Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 139Nervesystemets utvikling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 140
Kort oversikt over viktige faser i nervesystemets utvikling . . . . . . . . . . . . . . . . . 140Sentralnervesystemet anlegges som et rør . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 140Tidlig utvikling av den kraniale enden av nevralrøret . . . . . . . . . . . . . . . . . . . . . . 141Tidlige faser i utviklingen av nevroepitelet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 144Videre utvikling av medulla spinalis og hjernestammen . . . . . . . . . . . . . . . . . . . 146Videre utvikling av diencephalon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 147Videre utvikling av telencephalon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 147Utviklingen av hjernebarken . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 149Myelinisering . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 151Hvordan etableres nervesystemets spesifikke forbindelser? . . . . . . . . . . . . . . . . . 152Aksonets vekstkonus påvirkes av N-CAM og andre molekyler . . . . . . . . . . . . . . 154Nervevekstfaktorer – nevrotrofiner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 154Programmert celledød og konkurranse om vekstfaktorer . . . . . . . . . . . . . . . . . . . 155Eliminering av aksongrener . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 155Nydannelse og eliminasjon av synapser fra nyfødt til voksen alder . . . . . . . . . . . 156Etablering av topografiske kart . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 157Nervesystemets utvikling etter fødselen er bruksavhengig . . . . . . . . . . . . . . . . . . 157Sensitive (kritiske) perioder . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 159
Nervesystemet og aldring . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 161Hvor mange nevroner mister vi? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 161Distribuerte nettverk, bruksavhengig plastisitet og aldring . . . . . . . . . . . . . . . . . . 161Tap av perifere sanseorganer med alderen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 162Biologiske aldersforandringer i hjernen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 162Demens og nevrodegenerative sykdommer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 165Ligger felles molekylære mekanismer bak nevrodegenerative sykdommer? . . . . 167
Restitusjon etter skader av sentralnervesystemet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 167To faser i restitusjonsprosessen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 168Mekanismer for restitusjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 168Studier av restitusjon etter hjerneslag hos mennesker . . . . . . . . . . . . . . . . . . . . . . 170
0000 UFb SentrNerv 070401M.book Page 8 Wednesday, July 4, 2007 12:11 PM

INNHOLD 9
Restitusjon er en læringsprosess . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 171Restitusjon etter skader i tidlig barnealder . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 173
D
EL
2 S
ENSORISKE
SYSTEMER
K
APITTEL
5 Generelt om sanseorganer
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 177
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 177Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 177Sensoriske enheter og reseptoriske felt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 178Transduksjon: oversettelse av stimulus til aksjonspotensialer . . . . . . . . . . . . . . . . . . . 178
Reseptorpotensial . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 178Transduksjonsmekanismer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 179
Reseptorers egenskaper . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 180Adekvat stimulus, sansemodalitet og sansekvalitet . . . . . . . . . . . . . . . . . . . . . . . . 180Adaptasjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 181Dynamisk og statisk følsomhet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 181
Klassifisering av reseptorer på grunnlag av deres egenskaper . . . . . . . . . . . . . . . . . . . 181Adekvate stimulus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 181Hvor signalene kommer fra . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 182Fullstendig beskrivelse av en reseptor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 182
Reseptorer og sanseopplevelse . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 182Hjernen mottar ikke «sann» informasjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 183Sentral analyse . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 183
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 183
K
APITTEL
6 Det somatosensoriske systemet
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 185
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 185Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 185Hudens følesans. Eksteroseptorer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 186
Frie og innkapslede reseptorer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 187Nociseptorer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 187Termoreseptorer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 188Mekanoreseptorene i huden . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 189Hva informerer lavterskelmekanoreseptorene i huden om? . . . . . . . . . . . . . . . . . . 191Reseptoriske felt i huden . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 191Diskriminerende følesans . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 192Lateral inhibisjon: Inhibitoriske internevroner bedrer den diskriminerende følesansen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 192
Proprioseptorer. Dyp sensibilitet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 193Muskelspolenes bygning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 194Muskelspolens funksjonelle egenskaper . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 196Virkninger av
γ
-fibrene på muskelspolens egenskaper . . . . . . . . . . . . . . . . . . . . . . 198Muskelspoler hos mennesker og
α
–
γ
-koaktivering . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 199Senespolen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 199Hvorfor senespoler er mer følsomme for kontraksjon enn for passivt strekk . . . . . 200Leddenes innervasjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 201Nociseptorer i ledd . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 202Leddsans (kinestesi) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 202Proprioseptorer, balanse og viljestyrte bevegelser . . . . . . . . . . . . . . . . . . . . . . . . . 203
0000 UFb SentrNerv 070401M.book Page 9 Wednesday, July 4, 2007 12:11 PM

10 INNHOLD
De sensoriske fibrene og dorsalrøttene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 204Klassifisering av dorsalrotsfibre etter tykkelse og ledningshastighet . . . . . . . . . . 204Tykkelse av nervefibre og sanseopplevelse . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 206Segmental innervasjon. Dermatomer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 207Dorsalrotsfibrenes endeområder i ryggmargen . . . . . . . . . . . . . . . . . . . . . . . . . . . 208Sensoriske fibre og nevrotransmittere . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 209Nevropeptider i spinalganglieceller og smerte . . . . . . . . . . . . . . . . . . . . . . . . . . . 210Sensoriske fibre er ledd i refleksbuer. Ryggmargens internevroner . . . . . . . . . . . 210
Sentrale somatosensoriske baner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 211Sammendrag og sentrale begreper . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 211Thalamus – omkoblingsstasjon for sensoriske signaler . . . . . . . . . . . . . . . . . . . . 212Bakstrengene og lemniscus medialis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 213Thalamokortikal bane: til SI og SII . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 214Enkeltnevroners egenskaper i bakstrengs-lemniscus-medialissystemet . . . . . . . . 214Bakstrengs-lemniscus-medialissystemets oppgaver . . . . . . . . . . . . . . . . . . . . . . . 215Klinisk undersøkelse av bakstrengs-lemniscus-medialissystemet . . . . . . . . . . . . 216Tractus spinothalamicus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 216Koblinger mellom dorsalrotsfibre og spinotalamiske nevroner . . . . . . . . . . . . . . 217Endeområder for tractus spinothalamicus og videre formidling til korteks . . . . . 217Spinotalamiske nevroners egenskaper . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 218Tractus spinothalamicus’ oppgaver – homøostatisk overvåkning? . . . . . . . . . . . . 219Signaltrafikken i de sensoriske banene kontrolleres fra høyere nivåer . . . . . . . . . 221
De somatosensoriske barkområdene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 221Det primære somatosensoriske området, SI . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 222Videre behandling av sanseinformasjon utenfor SI . . . . . . . . . . . . . . . . . . . . . . . . 223Hvor i korteks behandles informasjon fra nociseptorer? . . . . . . . . . . . . . . . . . . . . 224
Smerte . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 225Nociseptorer og smerte er ikke det samme . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 225Kronisk smerte er forskjellig fra akutt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 225Patologisk smerte . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 226Sentral kontroll av smerte og ledning fra nociseptorer . . . . . . . . . . . . . . . . . . . . . 228Oppstigende forbindelser med virkning på smerteoppfattelse . . . . . . . . . . . . . . . 230Placebo og nocebo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 231Nyere synspunkter på smerte og smertebehandling . . . . . . . . . . . . . . . . . . . . . . . 232
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 233
K
APITTEL
7 Synssystemet
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 237
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 237Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 237Øyeeplet og øyets brytende medier . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 238
Øyet har likhetstrekk med et fotografisk kamera . . . . . . . . . . . . . . . . . . . . . . . . . 238Synsfeltet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 238Akkomodasjon og øyets fjernpunkt og nærpunkt . . . . . . . . . . . . . . . . . . . . . . . . . 238
Retina . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 241Retina er lagdelt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 241Fotoreseptorene og synspigment . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 242Stavene og tappene har forskjellige egenskaper . . . . . . . . . . . . . . . . . . . . . . . . . . 242Signaloverføring i retina . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 243Gangliecellenes reseptoriske felter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 244Gangliecellene overdriver forskjeller . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 246Synsskarphet og størrelsen på reseptoriske felt – macula lutea . . . . . . . . . . . . . . 247
0000 UFb SentrNerv 070401M.book Page 10 Wednesday, July 4, 2007 12:11 PM

INNHOLD 11
Forskjeller mellom sentrale og perifere deler av retina . . . . . . . . . . . . . . . . . . . . . 248Hovedtyper av ganglieceller . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 249
Synsbanenes organisasjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 250Aksonene fra M- og P-celler ender i forskjellige lag av corpus geniculatum laterale 250Synsinntrykk fra synsfeltet på én side kommer til motsatt sides hemisfære . . . . . . 252Synsbanene er retinotopisk ordnet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 252Sentrale deler av synsfeltet er overrepresentert i synsbanene og i korteks . . . . . . . 255
Visuell korteks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 256Egenskaper til nevroner i area striata . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 256Videre behandling av synsinformasjon: dorsal og ventral kanal ut av area striata . . 258Fargesyn . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 259Integrasjon av synsinformasjon og bevisst oppfattelse . . . . . . . . . . . . . . . . . . . . . . 261Utvikling av normalt syn krever riktig bruk . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 263
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 263
K
APITTEL
8 Hørselssystemet
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 265
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 265Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 265Hørselsorganet (cochlea) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 266
Cochlea er en del av labyrinten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 266Hvordan lydsvingningene overføres til sansecellene i cochlea . . . . . . . . . . . . . . . . 268Cortis organ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 269De indre hårcellene og mekanoelektrisk transduksjon . . . . . . . . . . . . . . . . . . . . . . 270De ytre hårcellene forsterker lyden . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 271Forskjellige frekvenser registreres på ulike deler av basilarmembranen. Tonotopisk lokalisasjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 271
Hørselsbanene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 272Nervus cochlearis og cochleariskjernene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 273Videre ledning fra cochleariskjernene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 273Lokalisering av lyd . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 274Nedstigende kontroll av impulstrafikken i hørselsbanene . . . . . . . . . . . . . . . . . . . 274Auditive reflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 276
Auditiv korteks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 277Kjerneområde og belteområder . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 277Egenskaper til nevroner i AI . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 277Videre behandling utenfor AI . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 278
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 278
K
APITTEL
9 Likevektssansen og balanse
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 279
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 279Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 279Likevektsapparatets bygning og virkemåte . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 280
Den vestibulære delen av labyrinten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 280Rotasjon av hodet stimulerer buegangene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 280Sacculus og utriculus registrerer hodets stilling og lineær akselerasjon . . . . . . . . . 283
Likevektskjernene og deres forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 283Vestibulariskjernene og de primærafferente fibrene . . . . . . . . . . . . . . . . . . . . . . . . 283Vestibulariskjernene mottar også afferenter fra andre steder enn labyrinten . . . . . 283Vestibulariskjernenes efferente forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 284
0000 UFb SentrNerv 070401M.book Page 11 Wednesday, July 4, 2007 12:11 PM

12 INNHOLD
Vestibulære reflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 285Labyrintreflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 286Vestibulo-okulære reflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 286Integrasjon av vestibulær og annen sanseinformasjon . . . . . . . . . . . . . . . . . . . . . 286
Reseptorer og posturale reflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 286Posturale reflekser er under sentral kontroll . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 287Ulike reseptortypers betydning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 287Nakke- og labyrintreflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 289
Kortikal behandling av vestibulære signaler . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 289Flere områder integrerer vestibulære signaler med andre sansemodaliteter . . . . . 289Distribuerte nettverk . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 289
Balanse og kroppsbilde . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 290Kroppsbilde og indre modeller . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 290Kroppsbildet forutsetter integrasjon av sanseinformasjon og bevegelsesintensjon 291Ustøhet, svimmelhet og vertigo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 292
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 293
K
APITTEL
10 Luktesansen og smakssansen
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 295
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 295Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 295Det olfaktoriske systemet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 296
Reseptorer for luktesansen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 296Luktreseptorene uttrykker et enormt repertoar av reseptorproteiner . . . . . . . . . . . 297Sentral ledning av luktinformasjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 297Endeområder for fibrene i tractus olfactorius . . . . . . . . . . . . . . . . . . . . . . . . . . . . 297Videre behandling utenfor den primære luktebarken . . . . . . . . . . . . . . . . . . . . . . 299Lukteinntrykk og atferd . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 299
Smakssansen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 299Reseptorene for smak og smakskvaliteter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 300Smaksstoffene virker via ionekanaler og G-proteinkoblede reseptorer . . . . . . . . 301Smakscellene er spesifikke . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 302Ledningsveier fra smaksreseptorer til hjernebarken . . . . . . . . . . . . . . . . . . . . . . . 302Den primære smaksbarken . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 303Videre kortikal behandling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 303
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 304
D
EL
3 M
OTORISKE
SYSTEMER
K
APITTEL
11 Perifere motoriske nevroner og reflekser
. . . . . . . . . . . . . . . . . . . . . . . . . . . . 309
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 309Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 309Motonevroner og muskler . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 310
Motonevronene er ordnet i søyler . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 311Motonevronene er funksjonelt forskjellige . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 312Den motoriske endeplaten og nevromuskulær transmisjon . . . . . . . . . . . . . . . . . . 312Muskelcellens kontraksjonskraft reguleres av motonevronet . . . . . . . . . . . . . . . . 314Fibertyper: muskelceller spesialisert for ulike typer arbeid . . . . . . . . . . . . . . . . . 315Fibertyper hos mennesker . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 315Motoriske enheter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 316Motoriske enheter og fibertyper . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 316
0000 UFb SentrNerv 070401M.book Page 12 Wednesday, July 4, 2007 12:11 PM

INNHOLD 13
Gradering av muskelkraft: rekruttering og frekvens . . . . . . . . . . . . . . . . . . . . . . . . 317Rekruttering ved ulike kraftbehov . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 317
Reflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 318Noen reflekser «forsvinner» under utviklingen . . . . . . . . . . . . . . . . . . . . . . . . . . . 319Refleksbuen og grunnleggende egenskaper ved reflekser . . . . . . . . . . . . . . . . . . . 319Fleksjonsrefleksen (tilbaketrekningsrefleksen) . . . . . . . . . . . . . . . . . . . . . . . . . . . . 320Strekkereflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 321Den monosynaptiske strekkerefleksen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 322Resiprok inhibisjon – hemming av antagonistene ved strekkereflekser . . . . . . . . . 323Langlatensstrekkereflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 323Strekkerefleksenes oppgaver . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 324Hudreseptorer og presisjonsgrep . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 326Sentral modulering av reflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 326
Muskeltonus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 326Hva bestemmer muskeltonus? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 327Individuelle forskjeller i muskeltonus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 328Sykelige forandringer av muskeltonus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 329
Skader av perifere motoriske nevroner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 330Perifere aksoner kan vokse ut igjen etter skade . . . . . . . . . . . . . . . . . . . . . . . . . . . 331Forhold som påvirker regenerasjon etter nerveskade . . . . . . . . . . . . . . . . . . . . . . . 331
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 332
K
APITTEL
12 Sentrale motoriske baner – kontroll av automatiske og viljestyrte bevegelser
. . . . . . . . . . . . . . . . . . . 333
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 333Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 333Pyramidebanen (tractus corticospinalis) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 334
Pyramidebanen kommer fra MI og områdene rundt . . . . . . . . . . . . . . . . . . . . . . . . 334Pyramidebanens forløp . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 335Ledningshastighet og endeområder i ryggmargen . . . . . . . . . . . . . . . . . . . . . . . . . 337Virkninger i ryggmargen – monosynaptisk kobling til motonevroner . . . . . . . . . . 338Utbredelse og virkninger av kortikospinale fibre i ryggmargen . . . . . . . . . . . . . . . 338Pyramidebanen virker særlig på bøyemuskler . . . . . . . . . . . . . . . . . . . . . . . . . . . . 339Pyramidebanen kontrollerer spinale refleksbuer . . . . . . . . . . . . . . . . . . . . . . . . . . . 340
Nedstigende baner til ryggmargen fra hjernestammen – indirekte kortikospinale signalveier . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 341
Kortiko-retikulospinale baner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 341Tektospinale baner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 342Vestibulospinale baner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 342Monoaminerge baner fra hjernestammen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 343
Kontroll av automatiske bevegelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 344Nettverk i hjernestammen og ryggmargen koordineres fra hjernebarken . . . . . . . . 344Postural kontroll . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 344Utvikling av postural kontroll . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 345Kontroll av gangbevegelser – rytmegeneratorer . . . . . . . . . . . . . . . . . . . . . . . . . . . 345Sentral kontroll av gangbevegelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 346
Motoriske barkområder og kontroll av viljestyrte bevegelser . . . . . . . . . . . . . . . . . . . . 346Motoriske nettverk og hierarkisk organisering av motoriske områder . . . . . . . . . . 346De «mest motoriske» barkområdene: MI, SMA og PMA . . . . . . . . . . . . . . . . . . . 347Det primære motoriske barkområdets forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . 348Funksjonell organisering av motorisk korteks . . . . . . . . . . . . . . . . . . . . . . . . . . . . 349
0000 UFb SentrNerv 070401M.book Page 13 Wednesday, July 4, 2007 12:11 PM

14 INNHOLD
Den supplementærmotoriske area (SMA) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 350Den premotoriske area, PMA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 351Motoriske barkområder og motorisk læring . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 352
Symptomer ved avbrytelse av sentrale motoriske baner . . . . . . . . . . . . . . . . . . . . . . . 352«Negative» og «positive» symptomer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 353Plantarrefleksen og andre reflekser som endres ved skade av sentrale nevroner . . 355Spastisitet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 355
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 357
K
APITTEL
13 Basalgangliene
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 359
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 359Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 359Basalganglienes bygning og forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 360
Striatum er basalganglienes mottagende del . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 362Projeksjonsnevroner i striatum sender aksoner til globus pallidus og substantia nigra . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 363Basalgangliene virker på premotorisk nettverk i thalamus og hjernestammen . . . 364Signalveier fra globus pallidus og substantia nigra til thalamus og videre til hjernebarken . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 365Forbindelser fra basalgangliene til hjernestammen . . . . . . . . . . . . . . . . . . . . . . . . 366Basalgangliene er organisert i funksjonelt forskjellige kretser . . . . . . . . . . . . . . . 366Transmittere og synaptiske virkninger av nevroner i sløyfen korteks–basalgangliene–korteks: disinhibisjon . . . . . . . . . . . . . . . . . . . . . . . . . . . 367Funksjonell betydning av disinhibisjon i premotoriske nevrongrupper . . . . . . . . 368Nucleus subthalamicus har stor innflytelse på aktiviteten i pallidum og nigra . . . 368Virkninger av dopamin i striatum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 369Hva aktiverer dopaminerge nevroner? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 370
Ventrale striatopallidum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 371Basalganglienes oppgaver . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 373Sykdommer som rammer basalgangliene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 373Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 377
K
APITTEL
14 Cerebellum
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 379
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 379Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 379Cerebellums inndeling og afferente forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 380
Cerebellum består av tre funksjonelt forskjellige deler . . . . . . . . . . . . . . . . . . . . . 380Afferente forbindelser fra labyrinten og likevektskjernene . . . . . . . . . . . . . . . . . . 382Afferente forbindelser fra medulla spinalis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 382Afferente forbindelser fra cortex cerebri . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 384Intermediærsonen er «møtested» for signaler fra medulla spinalis og hjernebarken 386
Lillehjernebarken, klatrefibrene og mosfibrene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 386Lillehjernebarken består av tre lag . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 387Afferenter til cerebellarkorteks er av to slag: mosfibre og klatrefibre . . . . . . . . . 387Mosfibrene og klatrefibrene formidler ulike typer av informasjon . . . . . . . . . . . . 389
Cerebellums efferente forbindelser. Cerebellarkjernene . . . . . . . . . . . . . . . . . . . . . . . 390Nevronene i cerebellarkjernene er spontantaktive . . . . . . . . . . . . . . . . . . . . . . . . 391Endeområder for de efferente forbindelsene fra cerebellarkjernene . . . . . . . . . . . 392
Cerebellums funksjon og symptomer ved skader . . . . . . . . . . . . . . . . . . . . . . . . . . . . 393Det «flokkulonodulære syndrom» og «lobus anterior-syndromet» . . . . . . . . . . . . 394
0000 UFb SentrNerv 070401M.book Page 14 Wednesday, July 4, 2007 12:11 PM

INNHOLD 15
Cerebellarhemisfærene og det «neocerebellare» syndromet . . . . . . . . . . . . . . . . . . 394Tidsteorien – utfører cerebellum én grunnleggende operasjon som brukes til flere oppgaver? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 396Cerebellum og læring . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 397Cerebellum og kognitive funksjoner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 397
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 398
K
APITTEL
15 Kontroll av øyebevegelser
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 399
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 399Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 399Øyemusklene og bevegelser av øyet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 400
Horisontalbevegelser, vertikalbevegelser og rotasjonsbevegelser . . . . . . . . . . . . . 400De ytre øyemusklene og deres virkninger . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 400Øyebevegelsene foregår konjugert . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 402
Sentral kontroll av øyebevegelsene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 402Typer av konjugerte øyebevegelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 402Sentre i hjernestammen for øyebevegelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 403Cerebellum er viktig for både sakkader og følgebevegelser . . . . . . . . . . . . . . . . . . 405Kortikal kontroll av øyebevegelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 406
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 408
D
EL
4 H
JERNESTAMMEN
OG
HJERNENERVENE
K
APITTEL
16 Retikulærsubstansen
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 411
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 411Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 411Retikulærsubstansens bygning og forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 412
Medial efferent og lateral afferent del . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 412Retikulærsubstansen er bygget for integrasjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . 412Rafekjernene og locus coeruleus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 413Mulige overordnede oppgaver for rafekjernene og locus coeruleus . . . . . . . . . . . . 413Retikulærsubstansens efferente forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 415Retikulærsubstansen mottar alle typer sensorisk informasjon . . . . . . . . . . . . . . . . 417Afferenter til retikulærsubstansen fra hjernebarken og subkortikale kjerner . . . . . 418
Funksjonelle forhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 419Aktiveringssystemet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 419Virkning på skjelettmuskler . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 419Kontroll av åndedrett og sirkulasjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 420
Bevissthet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 421En forutsetning for bevissthet: retikulærsubstansens virkning på hjernebarken . . . 421Signalveier og transmittere som er ansvarlige for aktivering av korteks . . . . . . . . 425Kontroll av sanseinformasjon og fokusering av oppmerksomheten . . . . . . . . . . . . 426
Søvn . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 426Søvn og læring . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 426Nevrongrupper og transmittere som kontrollerer søvnen . . . . . . . . . . . . . . . . . . . . 427Drømmer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 428
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 429
0000 UFb SentrNerv 070401M.book Page 15 Wednesday, July 4, 2007 12:11 PM

16 INNHOLD
K
APITTEL
17 Hjernenervene
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 431
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 431Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 431Generelle trekk i hjernenervenes organisasjon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 432Hjernenervene kan inneholde fire typer fibre . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 433
Prinsipper for beliggenhet av hjernenervekjernene . . . . . . . . . . . . . . . . . . . . . . . . 433Nærmere om beliggenheten av hjernenervekjernene . . . . . . . . . . . . . . . . . . . . . . 433Hjernestammereflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 434Hjernenervene er koblet til sensoriske og motoriske baner . . . . . . . . . . . . . . . . . . 435
Nervus hypoglossus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 436Nervus accessorius . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 438Nervus vagus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 438
Visceral-efferente (parasympatiske) vagusfibre . . . . . . . . . . . . . . . . . . . . . . . . . . 438Visceral-afferente vagusfibre . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 439Viscerale reflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 440Somatisk-efferente vagusfibre . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 440
Nervus glossopharyngeus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 442Sinusnerven og baroreseptorer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 442Nervus glossopharyngeus’ kjerner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 442
Nervus vestibulocochlearis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 442Nervus facialis og nervus intermedius . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 442
Nervus facialis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 443Sentral og perifer facialislammelse . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 443«Ekte» mimikk er ikke avhengig av pyramidebanen . . . . . . . . . . . . . . . . . . . . . . 444Nervus facialis og reflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 444Tåre- og spyttsekresjonen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 445
Nervus trigeminus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 445Den sensoriske trigeminuskjernen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 446Sentral ledning av signaler fra trigeminuskjernene . . . . . . . . . . . . . . . . . . . . . . . . 446Trigeminus og reflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 446Den motoriske trigeminuskjernen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 446
Nervus abducens, nervus trochlearis og nervus oculomotorius . . . . . . . . . . . . . . . . . . 448Nervus abducens . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 449Nervus trochlearis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 449Nervus oculomotorius . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 449Lysrefleksen og akkomodasjonsrefleksen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 451
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 451
D
EL
5 D
ET
AUTONOME
NERVESYSTEMET
OG
KONTROLL
AV
KROPPENS
INDRE
MILJØ
K
APITTEL
18 Perifere deler av det autonome nervesystemet
. . . . . . . . . . . . . . . . . . . . . . . . 455
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 455Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 455Generelle trekk ved organiseringen av det autonome nervesystemet . . . . . . . . . . . . . 456
To efferente nevroner er koblet etter hverandre . . . . . . . . . . . . . . . . . . . . . . . . . . 456Postganglionære fibre danner ikke typiske synapser . . . . . . . . . . . . . . . . . . . . . . . 456Noen organer styres mer presist enn andre . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 456Autonome ganglier og fletninger . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 457Forskjeller mellom det sympatiske og det parasympatiske systemet . . . . . . . . . . 458
0000 UFb SentrNerv 070401M.book Page 16 Wednesday, July 4, 2007 12:11 PM

INNHOLD 17
Perifere deler av det sympatiske systemet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 459Preganglionære fibre og den sympatiske grensestrengen . . . . . . . . . . . . . . . . . . . . 459Postganglionære sympatiske fibre . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 462Sympatisk innervasjon av hodet og ekstremitetene . . . . . . . . . . . . . . . . . . . . . . . . 462Sympatisk innervasjon av innvollsorganer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 464
Perifere deler av det parasympatiske nervesystemet . . . . . . . . . . . . . . . . . . . . . . . . . . . 467Hjernenervene inneholder preganglionære parasympatiske fibre . . . . . . . . . . . . . . 467Den sakrale delen av det parasympatiske systemet forsyner kjønnsorganene, blæren og endetarmen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 467
Det enteriske nervesystemet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 468Nevrontyper og nevrotransmittere . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 468
Det autonome nervesystemets oppgaver . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 469Sympatikus og parasympatikus: forsvar og vedlikehold . . . . . . . . . . . . . . . . . . . . 469Mer nyansert syn på arbeidsdelingen mellom sympatikus og parasympatikus . . . 469Sympatikus’ virkninger på sirkulasjonsorganene og lungene . . . . . . . . . . . . . . . . . 469Regulering av blodtrykk og fordeling av blodstrøm . . . . . . . . . . . . . . . . . . . . . . . . 470Virkning av sympatikus i huden . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 470Sympatikus’ virkninger på organer i bukhulen, kjønnsorganene og øyet . . . . . . . . 471Funksjonsspesifikk sympatisk kontroll . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 471Virkninger av de parasympatiske nervefibrene . . . . . . . . . . . . . . . . . . . . . . . . . . . . 472
Nevrotransmittere i det autonome nervesystemet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 473Non-kolinerg og non-adrenerg transmisjon i det autonome nervesystemet . . . . . . 473Presynaptiske reseptorer regulerer frisettingen av transmittere fra postganglionære nervefibre . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 475Sensibilisering . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 475
Sensorisk innervasjon av innvollsorganer og viscerale reflekser . . . . . . . . . . . . . . . . . 475Viscerale reseptorer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 476Sentral ledning av sensoriske signaler fra innvollene . . . . . . . . . . . . . . . . . . . . . . . 476Viscerale reflekser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 477Reflekser utløst fra reseptorer i lungene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 477Tømmingsrefleksen for urinblæren . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 477Overordnet kontroll av vannlatingen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 478Viscerale smerter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 479Hva slags stimuli fremkaller smerte fra innvoller? . . . . . . . . . . . . . . . . . . . . . . . . . 480Referert smerte . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 481
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 483
K
APITTEL
19 Overordnede deler av det autonome nervesystemet: Hypothalamus
. . . 485
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 485Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 485«Sentre» i hjernestammen for koordinering av atferd . . . . . . . . . . . . . . . . . . . . . . . . . . 486Hypothalamus’ bygning og forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 486
Inndeling i kjerner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 487Hypothalamus inneholder mange nevrotransmittere . . . . . . . . . . . . . . . . . . . . . . . 488Hypothalamus’ afferente forbindelser og andre former for påvirkning . . . . . . . . . 488Hypothalamus’ efferente forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 489
Hypothalamus og det endokrine systemet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 491Hypofysens forlapp danner flere hormoner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 491Hypothalamus’ relasjon til hypofysens baklapp . . . . . . . . . . . . . . . . . . . . . . . . . . . 491Hypothalamus’ virkning på hypofysens forlapp. Det hypofysære portåresystemet 493
Funksjonelle forhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 495
0000 UFb SentrNerv 070401M.book Page 17 Wednesday, July 4, 2007 12:11 PM

18 INNHOLD
Hypotalamiske «sentre» . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 495Temperaturregulering . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 495Hypothalamus og søvn/våkenhet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 496Hypothalamus og sirkadiane rytmer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 497Regulering av fordøyelse og matinntak . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 498Hypothalamus, seksualfunksjon og kjønnsforskjeller . . . . . . . . . . . . . . . . . . . . . . 499
Hypothalamus og immunsystemet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 500Immunsystemets virkning på sentralnervesystemet . . . . . . . . . . . . . . . . . . . . . . . 500
Hypothalamus og psykiske funksjoner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 501Psykosomatiske relasjoner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 501Stress . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 502Hypothalamus og emosjoner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 502
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 505
D
EL
6 C
ORTEX
CEREBRI
OG
LIMBISKE
STRUKTURER
–
TANKER
OG
FØLELSER
K
APITTEL
20 Limbiske strukturer – emosjoner og hukommelse
. . . . . . . . . . . . . . . . . . . . 509
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 509Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 509
«Det limbiske system» – et hensiktsmessig begrep? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 510
Amygdala og emosjoner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 510Amygdalas afferente forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 510Amygdalas efferente forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 512Amygdalas oppgaver . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 513Amygdala og atferd . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 513Amygdala og læring . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 515
Kortikale områder av betydning for autonome funksjoner og for emosjoner . . . . . . . 516Autonome funksjoner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 516Emosjoner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 517
Nevrongrupper i basale deler av hemisfæren. «The basal forebrain» . . . . . . . . . . . . . 518Kolinerge nevroner som sender aksoner til hjernebarken . . . . . . . . . . . . . . . . . . . 518
Hippocampusformasjonen, læring og hukommelse . . . . . . . . . . . . . . . . . . . . . . . . . . . 520Hippocampusformasjonens afferente forbindelser . . . . . . . . . . . . . . . . . . . . . . . . 521Modulerende forbindelser øker plastisiteten i hippocampus . . . . . . . . . . . . . . . . . 522Hippocampusformasjonens efferente forbindelser . . . . . . . . . . . . . . . . . . . . . . . . 522Ulike former for læring og hukommelse . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 524Mediale deler av temporallappen er nødvendig for deklarativ hukommelse . . . . 526Permanente hukommelsesspor ligger i mange deler av hjernen . . . . . . . . . . . . . . 527Hva er hippocampus’ selvstendige bidrag til læring og hukommelse? . . . . . . . . . 527«Hjernens kunnskapssystemer» . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 528
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 529
K
APITTEL
21 Cortex cerebri og høyere mentale funksjoner
. . . . . . . . . . . . . . . . . . . . . . . . 531
Innhold . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 531Oversikt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 531Hjernebarkens oppbygning og synaptiske organisasjon . . . . . . . . . . . . . . . . . . . . . . . 532
Neokorteks’ lagdeling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 532Nevronene er ordnet i søyler . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 533
0000 UFb SentrNerv 070401M.book Page 18 Wednesday, July 4, 2007 12:11 PM

INNHOLD 19
Afferente fibre fra forskjellige steder ender i ulike laminae . . . . . . . . . . . . . . . . . . 534Hjernebarken kan inndeles i cytoarkitektoniske areae . . . . . . . . . . . . . . . . . . . . . . 535Projeksjonsnevroner og internevroner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 535Rask eksitasjon og inhibisjon i korteks: glutamat og GABA . . . . . . . . . . . . . . . . . 536Modulerende synaptiske virkninger i hjernebarken . . . . . . . . . . . . . . . . . . . . . . . . 537Intrakortikal impulstrafikk og informasjonsbehandling . . . . . . . . . . . . . . . . . . . . . 537Kortikale nevroner oppdager sammentreff . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 538Horisontal integrasjon og plastisitet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 538
Hjernebarkens forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 538Hovedtrekk . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 539Spesifikke talamokortikale forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 539De intralaminære thalamuskjernene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 540Kortikotalamiske forbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 541Direkte kortikale afferenter fra nevrongrupper med modulerende transmittere . . . 542Hjernebarkens assosiasjonsforbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 542Foroverrettede og tilbakerettede kortikokortikale forbindelser . . . . . . . . . . . . . . . 543Hjernebarkens kommissuralforbindelser . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 544
Oppgaver knyttet til hjernebarkens assosiasjonsområder . . . . . . . . . . . . . . . . . . . . . . . 544Assosiasjonsområder . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 544Integrasjon av sanseinformasjon i assosiasjonsområdene må læres . . . . . . . . . . . . 545Parietale assosiasjonsområder . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 546Frontallappens assosiasjonsområder . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 548Symptomer ved skader av prefrontalkorteks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 549Temporallappens assosiasjonsområder . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 549Symptomer ved skader av temporallappen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 550Hvordan integreres informasjon i hjernebarken? . . . . . . . . . . . . . . . . . . . . . . . . . . 551
Språkområder i hjernebarken . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 553Er det spesifikke systemer for språk i hjernen? . . . . . . . . . . . . . . . . . . . . . . . . . . . 554
Arbeidsdeling mellom hemisfærene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 554Er det anatomiske forskjeller mellom høyre og venstre hemisfære? . . . . . . . . . . . 554Kommissuralforbindelsenes funksjon. Corpus callosum . . . . . . . . . . . . . . . . . . . . 555Lateralisering og dominans . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 556Lateralisering av språk . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 557Lateralisering av håndmotorikk . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 557Lateralisering og emosjoner . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 558Lateralisering og kognitive kjønnsforskjeller . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 558Psykologiske kjønnsforskjeller og biologi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 558Arv eller miljø? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 559
Noter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 560
Litteratur
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 561
Stikkord
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 601
0000 UFb SentrNerv 070401M.book Page 19 Wednesday, July 4, 2007 12:11 PM

0000 UFb SentrNerv 070401M.book Page 20 Wednesday, July 4, 2007 12:11 PM

Denne boken er i første rekke beregnet for medisin-,fysioterapi- og psykologistudenter, det vil si gruppersom trenger kjennskap til nervesystemet som basis forsenere klinisk virksomhet. Denne fjerde utgaven hargjennomgått en omfattende revisjon og fornyelse. Vedsiden av oppdatering og omskriving der ny kunnskaphar gjort det nødvendig, har bokens layout og illustra-sjonsmateriale gjennomgått store forandringer. Sikte-målet har vært å lette tilgjengeligheten av stoffet, blantannet ved bruk av farger gjennom hele boken. Samtligefigurer er omtegnet, og en del nye er kommet til.
En lærebok er nødvendigvis basert på en hardhendtog ganske subjektiv utvelgelse av stoff. Den samledestoffmengden i denne boken går likevel ut over hvasom er realistisk å tilegne seg i en grunnutdanning.Dette er gjort for å dekke behov for senere fordypning,og fordi de forskjellige profesjonsutdanninger harfokus på ulike sider av nervesystemets virksomhet. Forå gjøre det lettere å velge det som er mest relevant, erutdypende stoff markert med farget bakgrunn, slik atdet ikke skal forstyrre lesingen av hovedteksten.
Jeg har forsøkt å skrive slik at boken kan lesesmed utbytte uten forkunnskaper om nervesystemet.Mitt mål er det samme som min far Alf Brodal haddeda han skrev forløperen til denne boken for snart 60år siden: å stimulere til forståelse fremfor innpren-ting av isolerte fakta. Samtidig ønsker jeg å formidleen viss ydmykhet i forhold til vår tross alt svært be-grensede innsikt i menneskehjernen. Nervesystemet eroverveldende komplisert, og det er fortsatt langt igjen til vi
har svar på mange sentrale spørsmål. Selv om man i engrunnutdanning bare kan nå et stykke på vei i forhold tilhva som kunne være ønskelig å oppnå av faglig fordyp-ning, er ingen tjent med en ukritisk tilegnelse av overfla-diske kunnskaper som gir en falsk tro på egen kompe-tanse. Jeg har derfor forsøkt også å formidle noen avpremissene for det som presenteres, og om begrensnin-gene i vår viten, selv om dette kan gå ut over den umiddel-bare tilgjengeligheten.
Den store utfordringen – for studenten som for fors-keren – er å forstå hvordan nervesystemet løser sineoppgaver. Dette krever en
integrert
tilnærming, hvordet trekkes inn kunnskaper fra alle spesialfelt innenfornevrobiologien, og også fra psykologisk og kliniskforskning. Denne boken er et forsøk på en slik integrertfremstilling. Kanskje fordi mitt eget forskningsfelt ernervesystemets koblingsmønstre, vil jeg likevel frem-heve at presise forestillinger om nervesystemets byg-ning er en forutsetning for å kunne nyttiggjøre segkunnskap fra andre felter. Slik anatomisk kunnskap erogså nødvendig for å kunne tolke symptomer som opp-trer ved sykelige forandringer i hjernen. Kunnskap omhjernens kompliserte anatomi alene er likevel ikke noemål i seg selv, og forsøk på innlæring av mange anato-miske detaljer går ofte på bekostning av forståelse ogsammenheng. Bare der de bidrar til forståelse av funk-sjon eller av viktige sykdomsytringer, er derfor anato-miske trekk beskrevet nærmere.
I arbeidet med bokens fire utgaver har jeg fått rådog hjelp av en rekke kolleger, som jeg er svært takk-
Forord
0000 UFb SentrNerv 070401M.book Page 21 Wednesday, July 4, 2007 12:11 PM

22 FORORD
nemlig for. Jan Bjålie, Niels Christian Danbolt, Sig-bjørn Fossum, Paul Heggelund, Jan Jansen, HaraldKryvi, Kirsten Osen, Ole Petter Ottersen, JonStorm-Mathisen og Eric Rinvik har alle gitt konstruk-tive kommentarer. Mange forbedringer er foreslått avstudenter. Jeg er svært takknemlig for oppmuntrendekommentarer og konstruktiv kritikk fra brukerne avboken. Jeg vil også takke Gunnar Lothe og CarinaKnudsen, som har ansvar for det fotografiske arbeidet.
Professor Dag Gundersen har vært til stor hjelp som år-våken språkkonsulent, og bidratt til språklig konsistensog klarhet, og ikke minst korrekt fornorskning av fag-uttrykk. Til sist er det en glede å takke seniorredaktørMariann Bakken og Universitetsforlaget for et inspire-rende og tillitsfullt samarbeid.
Oslo, mai 2007
Per Brodal
0000 UFb SentrNerv 070401M.book Page 22 Wednesday, July 4, 2007 12:11 PM

Hva er sentralnervesystemets viktigste oppgaver? Etslikt spørsmål er ikke lett å besvare, og i alle fall ikkekort – hjernen står for alt det vi forbinder med å væremenneske. På et overordnet nivå kan vi si at hjernenskaper vår virkelighet: den velger, sorterer og tolkerden overveldende mengden av informasjon vi kan tainn fra våre omgivelser og vår egen kropp, og den sty-rer vår atferd i henhold til sin tolkning av virkeligheten.Det gjelder atferd i vid forstand: et aspekt er kontrollog vedlikehold av kroppen selv og dens indre miljø, etannet er vår interaksjon med andre mennesker gjen-nom handlinger og språk. Et tredje er vår indre, subjek-tive mentale virkelighet, som andre bare delvis kantrenge inn i. I tidlig barnealder må hjernen skape ordenog forutsigbarhet slik at vi lærer å forholde oss til ossselv og våre omgivelser på en hensiktsmessig måte.
Nervesystemets viktigste grunnelement er
nerve-cellen
, som er spesialisert for hurtig ledning av signa-ler over lange avstander. Nervecellene kan videre over-føre signaler seg imellom på en presis måte – de kankommunisere. Til sammen danner milliarder av nerve-celler i vår hjerne kompliserte og velordnede nettverkfor
kommunikasjon
og
informasjonsbehandling
.Nervesystemet mottar enorme mengder med informa-sjon fra våre omgivelser og fra kroppen. Blant alt somkommer inn, siler nervesystemet ut det vesentligste,
sørger for lagring av det som kan trenges siden, og sen-der eventuelt ut kommandoer hvis det trengs et svar påinformasjonen. I noen tilfeller kan svaret kommemeget raskt, med minimal behandlingstid av informa-sjonen og uten bevisst medvirkning, som en
refleks
. Iandre tilfeller kan svaret trenge lang behandlingstid ogsamspill mellom mange deler av nervesystemet, ogvære et resultat av
bevisst tankevirksomhet
. Denoverordnede oppgaven for nervesystemet er uansett åsørge for at organismen som helhet til enhver tid tilpas-ser seg best mulig til omgivelsene.
Nervesystemet er utstyrt med
sanseorganer
,
re-septorer
, som kan fange opp forskjellige typer av san-seinformasjon eller
stimuli
. Uansett hvilken type sti-mulus det gjelder (hvilken form for energi),«oversetter» sanseorganet stimulusenergien til det språ-ket som nervesystemet forstår, nemlig
nerveimpulser
.Dette er ørsmå elektriske utladninger som forplanterseg raskt langs nervetrådene. På den måten ledes signa-ler hurtig fra reseptorene til de delene av nervesystemethvor informasjonsbehandlingen foregår.
Når nervesystemet skal frembringe en ytre reak-sjon på en stimulus, må dette skje ved påvirkning av
ef-fektorer
(utførere), som kan være enten muskler ellerkjertelceller. Det betyr at svaret,
responsen
, kommer iform av
bevegelse
eller
sekresjon
. Sekretet fra kjertel-cellene kan for eksempel bidra til fordøyelsen, eller detkan være et signalstoff som bringes videre med blodettil andre celler. Muskelkontraksjon kan selvfølgelig giseg mange uttrykk, fra kommunikasjon gjennom tale,
Innledning
Et fugleperspektiv
0000 UFb SentrNerv 070401M.book Page 23 Wednesday, July 4, 2007 12:11 PM

24 INNLEDNING
minespill og kroppsholdning, til forflytning av krop-pen, åndedrett og endring av blodtrykket. Uansett harnervesystemet bare muskler og kjertelceller å spille påfor å uttrykke sin «vilje».
På anatomisk grunnlag kan vi dele inn nervesyste-met i
sentralnervesystemet
(CNS), som utgjøres avhjernen og ryggmargen, og
det perifere nervesyste-met
(PNS), som utgjøres av de delene som sørger forkommunikasjon mellom sentralnervesystemet på denene siden og sanseorganer og effektorer på den andre.Nervesystemet kan også inndeles, riktignok utenskarpe skiller, mellom de delene som fortrinnsvis regu-lerer virksomheten til innvollsorganer og kroppensindre miljø, og de delene som styrer vår mer ellermindre bevisste tilpasning til omgivelsene. Den førstedelen kalles det
autonome
eller
vegetative nervesys-temet
, den andre delen kalles gjerne det
somatiske
.Man bruker også betegnelsen
det cerebrospinale ner-vesystemet
om den siste delen, som vi kan si er kjen-netegnet ved å motta informasjon fra sanser som regist-rerer forholdene i våre omgivelser (som syn, hørsel,sanseorganer i huden) ved å kontrollere våre viljestyrtemuskler (tverrstripet skjelettmuskulatur). Det auto-nome nervesystemet derimot har som oppgave å kon-trollere aktiviteten av ikke-viljestyrt muskulatur (glattemuskelceller og hjertemuskelceller) og kjertelceller.Det autonome nervesystemet kan inndeles i to delersom stort sett har motsatte virkninger: det
sympatiskesystemet
, som særlig ivaretar mobilisering av krop-pens ressurser ved økte krav og påkjenninger, og det
parasympatiske systemet
, som i større grad ivaretardet daglige vedlikehold.
Hos virveldyr med en liten og (relativt sett) enkelhjerne – for eksempel en frosk – er det stort sett fastekoblinger mellom sansning og atferd. Det vil si at enstimulus, for eksempel et lite objekt i synsfeltet, utløserbestemte målrettede bevegelser. Det er få nevronerkoblet mellom sanseorganet og effektoren, med tilsva-rende små muligheter for å modifisere responsen på etsanseinntrykk. Det meste av dyrets atferd er derfor in-stinktiv og automatisk, og endres lite ved læring. Hospattedyr med en liten hjerne i forhold til kroppsvekten(f.eks. rotter) brukes en stor del av det de har av hjerne-bark, til bearbeiding av sanseinformasjon og til direktekontroll av bevegelser. Hos noen pattedyrarter – mesthos primatene – er hjernens vekt i løpet av noen milli-oner år økt sterkt i forhold til kroppsvekten. Mest mar-kert er dette hos mennesket, hvor hjernens relative vekter omtrent dobbelt så stor som hos menneskeapene(f.eks. sjimpansen). Hos mennesket er de faste koblin-gene mellom sansning og atferd for en stor del løst opp(med unntak av en del livsviktige reflekser). Samme
sanseinntrykk kan utløse meget forskjellige reaksjoner,avhengig av sammenhengen det opptrer i og forhisto-rien til individet. Det betyr at vi har mulighet til å velgeblant et større repertoar av reaksjoner, og responsenkan endres på grunnlag av erfaring. En slik fleksibilitetforutsetter imidlertid at mange nevroner er koblet innmellom sanseorganet og effektoren – det trengs «data-kraft» for å koble stimulus sammen med andre innkom-mende sansestimuli og tidligere erfaringer.
De høyere integrasjons- og assosiasjonsproses-sene, det vil si det vi gjerne kaller
mentale prosesser
,finner sted først og fremst i
hjernebarken
. Det er daogså først og fremst økningen i dennes størrelse somforklarer tilpasningsevnen og det enorme læringspo-tensialet hos mennesket. Ja, vår hjerne har ikke baregitt oss en enestående evne til å tilpasse oss skiftendemiljøer, den har også gitt oss muligheten til å endrevårt miljø så det passer til oss. I dette ligger enormemuligheter, men også farer, ved at vi som kjent frem-bringer endringer som på kort sikt er fordelaktige, menpå lang sikt truer vår egen eksistens som art.
Et utvalg av metoder for utforskning av nervesystemetblir omtalt i kapitler hvor metodene har vært brukt til åfremskaffe kunnskapen som formidles. Her skal vibare ta for oss noen generelle trekk ved nevrobiologiskforskning. Mange veier har vært fulgt for å klarleggenervesystemets bygning og funksjon, fra de aller enk-leste observasjoner av hjernens makroskopiske utse-ende til bestemmelse av enkeltmolekylers funksjon. Ide senere årene har det skjedd en voldsom utvikling imetoder, slik at man i dag kan angripe problemer somman før ikke hadde mulighet til å gjøre noe med. Dethar også vært en enorm økning i antallet forskere på defleste felt innenfor nevrobiologien, fra det molekylæretil atferdsstudier. Kunnskapsmengden har da også økteksponentielt. Dette har utvilsomt gitt vesentlig ny inn-sikt, men har også gjort det vanskelig for den enkelteforsker å ha tilstrekkelig oversikt til å sette sine funninn i en større sammenheng. Det øker også faren for atforskeren trekker for vidtrekkende slutninger på basisav funn gjort i en kunstig eksperimentell situasjon. Enreduksjonistisk tilnærming er nødvendig for å få sikreresultater, men er samtidig et problem hvis forskerneikke er seg bevisst begrensningene dette legger på mu-ligheten til å trekke slutninger om helheten.
Hvordan undersøkes nerve-systemets bygning og funksjon?
0000 UFb SentrNerv 070401M.book Page 24 Wednesday, July 4, 2007 12:11 PM

INNLEDNING 25
Skjematisk kan vi gruppere metoder til utforskningav nervesystemet i slike som befatter seg med oppbyg-ningen, strukturen, og slike som brukes for å fastslåstrukturenes egenskaper. Grensene er imidlertid fly-tende, og typisk for moderne nevrobiologisk forskninger at anatomiske, fysiologiske, biokjemiske og andremetoder kombineres. Videre har moderne teknikkerfor databasert bildefremstilling gitt muligheter til å un-dersøke både struktur og dynamiske forhold, somblodstrøm og utbredelse av kjemiske stoffer, i den le-vende menneskehjernen. I økende grad anvendes celle-biologiske metoder under studiet av nervesystemet,deriblant ikke minst genteknologiske metoder, og maner blitt klar over at nerveceller ikke er så spesielle i for-hold til andre celler som man trodde tidligere.
Dyreeksperimenter har i særlig grad bidratt til økt kunnskapDet vi i dag vet om nervesystemet, baserer seg i en vissutstrekning på observasjoner hos mennesket, men ilangt større grad på undersøkelser av dyr. Hos mennes-ket er man oftest begrenset til å sammenholde kliniskeobservasjoner etter tilfeldige skader med undersøkelseav hjernen etter døden (postmortalt), selv om modernebildedannende teknikker har åpnet nye muligheter.Ved undersøkelse av menneskemateriale er sjelden totilfeller like, og ofte er skadene så store at slutningerom sammenheng mellom skadested og funksjon blirsvært usikre.
Hos dyr kan man derimot legge de eksperimentelleforholdene til rette, og man kan gjenta eksperimentenefor å kunne trekke pålitelige slutninger. Man kan ogsåundersøke egenskapene til elementer av nervesystemetdirekte, for eksempel kan aktivitet i veldefinerte nev-rongrupper sammenholdes med dyrets atferd. Man kanogså isolere deler av nervesystemet og studere detalene, for eksempel skiver av hjernevev som kan hol-des levende i flere timer, slik at man kan registrere fraceller som siden kan undersøkes mikroskopisk. Under-søkelser hos virvelløse dyr med enkle nervesystemerhar gjort det mulig å klarlegge fundamentale mekanis-mer for nervecellers kommunikasjon.
Når man kommer til spørsmål om funksjoner somer spesielle for de mest kompliserte hjerner, som men-neskets, er man imidlertid henvist til å gjøre eksperi-menter på høyere pattedyr som har en velutviklet hjer-nebark. Særlig katt og ape brukes i dennesammenhengen. Selv på basis av slike eksperimentermå slutninger om menneskets nervesystem trekkesmed forsiktighet. For selv om nervesystemet er bygdetter de samme prinsippene, er det selv ved sammen-ligning av aper og mennesker betydelige forskjeller i
utviklingsgraden for deler av hjernen. Slike anato-miske forskjeller tyder sterkt på at det også finnesfunksjonelle forskjeller. Derfor må studiet av mennes-ket, slik som det gjøres i klinisk nevrologi, psykiatri ogpsykologi, til slutt ha det avgjørende ordet. Men fordiklinikere som regel ikke kan eksperimentere, må devurdere om funn hos pasienter eller frivillige forsøks-personer lar seg forklare ut fra hva man vet fra ekspe-rimentelle studier på dyr. Hvis dette ikke er mulig, rei-ser kliniske observasjoner nye problemstillinger somofte kan angripes i dyreeksperimenter. Dypere sett erimidlertid de metodene som anvendes ved studier avmennesket, de samme som brukes ved utforskningenav nervesystemet hos dyr.
Dyreeksperimenter og etikkEksperimenter på dyr blir ofte kritisert på et etiskgrunnlag. Spørsmålet om det er akseptabelt å bruke dyrtil forskningsformål, kan imidlertid ikke løsrives fradet mer fundamentale spørsmålet om mennesket harrett til å råde over andre skapningers liv, enten det gjel-der til matforsyning eller til overtakelse av deres livs-rom. Det gjelder også retten til å endre dyrs egenskaperved systematisk avl eller genteknologiske manipula-sjoner. Anvendelse av dyr i nevrobiologisk forskninger en forutsetning for å oppnå dypere innsikt i den men-neskelige hjernen – dvs. i det biologiske substratet forvåre tanker, følelser og handlinger. Selv om cellekultu-rer og datasimuleringer kan erstatte noen dyreeksperi-menter, er vi likevel i overskuelig fremtid avhengig avdyreeksperimenter. Hvis for eksempel databaserte mo-deller av hjernebarkens virkemåte skal bli mer enngjetninger, kreves mer detaljerte og ikke minst merkvantitative data om hjernens koblingsmønstre og nev-ronenes egenskaper, som bare kan skaffes ved dyre-eksperimenter.
Økt kunnskap er imidlertid også nødvendig hvis viønsker å bedre behandlingsmulighetene for sykdommersom rammer nervesystemet. Disse sykdommene med-fører ofte betydelig grad av funksjonshemning og li-delse, og inntil i dag har vi bare i liten grad vært i standtil å behandle dem effektivt. Moderne metoder innenfornevrobiologien gir håp for fremtiden, men igjen er manavhengig av muligheten til å teste hypoteser ved blantannet dyreeksperimenter for å komme videre.
Selv om vi mener det er etisk forsvarlig å bruke dyri nevrobiologisk forskning, er det selvsagt grenser forhvor mye lidelse vi kan tillate oss å påføre dyr i men-neskets interesse. Det finnes imidlertid i dag strengeregler for hvem som kan drive med eksperimenter pådyr, og hvordan forsøkene skal gjennomføres for å re-dusere ubehag og smerte til et minimum, for eksempel
0000 UFb SentrNerv 070401M.book Page 25 Wednesday, July 4, 2007 12:11 PM

26 INNLEDNING
ved at alle inngrep gjøres i narkose. Større internasjo-nale tidsskrifter som publiserer forskningsrapporterinnenfor nevrobiologi, forlanger erklæring om at slikeregler har vært fulgt før en artikkel godtas for offentlig-gjørelse.
Alle metoder har feilkilderSelv om vi ikke systematisk kommer til å ta for ossfeilkilder ved metodene som nevnes i denne boken, erdet viktig å huske på at feilkilder finnes ved alle meto-der. En mulighet for feil ligger i forhastede slutningerom forholdene hos mennesker ut fra funn hos dyr.Mer generelt gjelder at alle eksperimentelle metodergår ut på, mer eller mindre, å isolere strukturer ellerprosesser slik at de kan observeres alene. Dette gjør atmange fenomener studeres ute av sin naturlige sam-menheng, og slutninger om hvordan de forskjelligedelene virker sammen i en intakt organisme, blir der-for ofte spekulative.
Også anatomiske metoder – som man kanskjeskulle tro gav entydige og sikre resultater – har sinefeilkilder. Det er for eksempel trukket mange feilslut-ninger om forbindelser mellom cellegrupper. Detfører i sin tur til feilslutninger når det gjelder tolkningav fysiologiske og kliniske data. Ved undersøkelserav mennesker er det selvfølgelig også betydelige feil-kilder, ikke minst av psykologisk art, som blant annet
gjør at man ikke alltid kan basere seg på pasientenseller forsøkspersonens opplysninger. Dette er ikkeminst et problem ved forsøk på å korrelere blod-strømsendringer i hjernen med kognitive prosesserog atferd.
Vitenskapelige «sannheter» må stadig revideresAt metodene som anvendes, har sine feilkilder, og attolkninger av observasjoner og funn ikke alltid er hold-bare, viser seg blant annet ved at mange oppfatningerom nervesystemet stadig er blitt revidert. Ofte skjerdette når det tas i bruk nye teknikker som belyser for-holdene fra en annen vinkel. Som i all vitenskap kanikke konklusjonene som trekkes, betraktes som ende-lige sannheter, men heller som mer eller mindre sann-synlige og foreløpige tolkninger. Forskning går i storutstrekning ut på å stille naturen spørsmål. Hvor for-ståelige og gode svarene er, avhenger av hvor presistspørsmålet er stilt, og hvor relevant det er for proble-met man ønsker å belyse. Selv om det for den enkeltekan være vanskelig å leve etter det, ligger det også iforskningens vesen at konklusjoner og tolkninger måvurderes fordomsfritt ut fra holdbarheten av de grunn-leggende observasjonene og argumentasjonens logiskestyrke. Hvem som presenterer konklusjonene, skalvære likegyldig.
0000 UFb SentrNerv 070401M.book Page 26 Wednesday, July 4, 2007 12:11 PM

Denne første delen av boken omhandler generelle forhold ved nervesystemets bygningog virkemåte, og er et nødvendig grunnlag for de senere kapitlene. Kapittel 1, Nerve-vevets bygningselementer, omhandler hvordan nevroner og glia er bygd, og noengrunnleggende trekk av hvordan nervecellene er sammenkoblet. Kapittel 2, Nerve-cellenes funksjonelle egenskaper, gir grunnlag for å forstå hvordan nerveceller kommu-niserer. Kapittel 3, Nervesystemets forskjellige deler, gir en oversikt over sentralnerve-systemets bygning med stikkordmessig beskrivelse av oppgaver for de forskjelligedelene. I kapittel 4, Nervesystemets utvikling, aldring og plastisitet, behandler vi nerve-systemets utvikling i fosterlivet og etter fødselen, med særlig vekt på hvordan dets kom-pliserte mønstre av forbindelser oppstår. Vi ser dessuten på endringer som skjer i nerve-systemet ved aldring, og på nervesystemets evne til tilpasning og plastisitet, både veddagligdagse endrede krav til ytelse og når man «kommer seg» etter en hjerneskade.
Hovedtrekk i nervesystemets bygning og funksjon DEL 1
0000 UFb SentrNerv 070401M.book Page 27 Wednesday, July 4, 2007 12:11 PM

0000 UFb SentrNerv 070401M.book Page 28 Wednesday, July 4, 2007 12:11 PM

NERVECELLEN S. 30SAMMENKOBLING AV NEVRONER OG SIGNALVEIER S. 39CELLESKJELETTET OG AKSONAL TRANSPORT S. 43GLIACELLER S. 45
Nervesystemet er bygd opp av nerveceller, nevro-ner, og gliaceller. Det er nervecellene som er an-svarlige for de funksjonene som særpreger nerve-systemet, mens gliacellene har en hjelpendefunksjon i forhold til nervecellene. Nevronene harsom oftest mange utløpere. Flere kortere dendrit-ter utgjør en mottagende del, mens et akson ledernerveimpulsen fra nevronet videre til andre nevro-ner eller muskelceller. Nervecellene er karakterisert
ved sin høyt utviklede evne til å reagere på stimulimed en elektrisk utladning, en nerveimpuls, ogved evnen til å lede nerveimpulsen hurtig i akso-net over betydelige avstander. På den måten brin-ges signaler i løpet av millisekunder fra ett sted tilet annet, enten innenfor sentralnervesystemet ellermellom dette og kroppens organer. Når nerveim-pulsen når frem til kontaktstedet med andre nevro-ner – synapsen – frisettes en nevrotransmitter sompåvirker neste nevron. Nevroner kan grupperes iprojeksjonsnevroner, som formidler signaler overlange avstander, og internevroner, som sørger forlokalt samarbeid mellom nevroner innenfor engruppe. Grupper av nevroner er koblet sammen ikompliserte nettverk. Slike nettverk ivaretar defleste av nervesystemets oppgaver.
Gliacellene har flere viktige oppgaver. De ernødvendige for å opprettholde et konstant miljørundt nervecellene (homøostase), og for repara-sjonsprosesser etter skader. De er også ansvar-lige for isolasjon av nervecelleutløpere slik atderes ledningshastighet økes. Gliaceller spillerogså en viktig rolle under nervesystemets utviklingi fosterlivet.
Nervesystemets bygnings-elementer KAP 1
Innhold
Oversikt
0000 UFb SentrNerv 070401M.book Page 29 Wednesday, July 4, 2007 12:11 PM

30 DEL 1 HOVEDTREKK I NERVESYSTEMETS BYGNING OG FUNKSJON
Nevroner har lange utløpereEn nervecelle (et nevron, figur 1.1) har som andre cel-ler et cellelegeme med kjerne, omgitt av cytoplasmamed organeller (figurene 1.2 og 1.3). Cellelegemet kal-les også soma (kropp) eller perikaryon. Fra cellelege-met går det ut lange utløpere. Utløperne kan varieremye i antall og lengde, men er av to hovedtyper. Denene typen, dendritter, har de fleste nevroner mange av,og hver dendritt forgrener seg videre (figur 1.1). På denmåten danner dendrittene en stor overflate – som et tre– som kan motta signaler fra andre nerveceller. Denandre typen utløper, aksonet, er spesialbygd for å ledesignaler (nerveimpulser) over lange avstander. I dealler fleste tilfeller leder aksonet impulser fra celle-legemet og utover. Hver celle har bare ett akson, menaksonet forgrener seg som regel sterkt, slik at et nevronkan påvirke opp til flere tusen andre. En aksongren kal-les også en kollateral eller aksonkollateral (figur 1.1).Betegnelsen nervefiber brukes synonymt med akson.
Dendritter er utstyrt med spinae
Skal man studere nervevevet nærmere, må man bruketynne snitt som kan undersøkes i mikroskop. Forskjel-lige fargemetoder kan brukes, slik at deler av eller helenevronet tydelig kan skilles fra sine omgivelser (figur1.4). Nevronene kan ha svært forskjellig form, bådenår det gjelder størrelse på cellelegemet (figur 1.2), ogantall, lengde og forgrening av dendrittene (figurene
21.5 og 21.6). Dendritt-treets størrelse forteller omhvor mange andre nerveceller en nervecelle blir kon-taktet av, det vil si dens mottagende overflate. Jo størredendritt-tre, desto mer informasjon kan nervecellen ta
Nervecellen
Cellelegeme (soma)
Dendritter
Spina
Akson
Nerveterminaler
Aksonkollateral
FIGUR 1.1 Nevron. Tegning som viser de forskjelligedelene av en typisk nervecelle (forenklet).
Nervecellene inneholder mye mitokondrier og ru endoplasmatisk retikulumKjernen i nerveceller sett i mikroskopiske snitt er karakterisert ved sin størrelse, og ved at den er lys(det vil si at kromatinet er ekstendert, som tegn på at mye av genomet stadig avleses). Den inneholderogså en tydelig nucleolus (figurene 1.2 og 1.3). Utseendet er så karakteristisk at nerveceller lett kanskilles fra andre celler (f.eks. gliaceller), selv om bare kjernen er tydelig farget. I cytoplasma finnes mito-kondrier, ofte i store mengder, som uttrykk for at nervecellene er metabolsk svært aktive med stortbehov for aerob ATP-syntese, som bare kan skje i mitokondriene (figur 1.3). Nervecellene er heltavhengige av aerob ATP-produksjon, det vil si tilførsel av oksygen, og kan ikke som mange andre cellergreie seg lenge med anaerob ATP-produksjon. Glukose er substrat for ATP-produksjonen som skjer imitokondriene i nerveceller – de kan ikke, som for eksempel muskelceller, forbrenne fett. Nervecelleneinneholder også påfallende store mengder av frie ribosomer og ru (granulert) endoplasmatisk retikulum(rER), hvor det foregår syntese av proteiner. Store klumper med rER sees lysmikroskopisk i cytoplasmai litt større nerveceller (figurene 1.2, 1.3 og 1.30), og ble kalt tigroid- eller Nissl-korn lenge før man visstehva de egentlig representerer. Årsaken til nervecellenes svært store produksjon av proteiner er sann-synligvis først og fremst at de har en så enorm overflate dekket av cellemembran. Selv om hovedbe-standdelen i cellemembranen i nervecellen – som i andre celler – er lipid, finnes det mange proteiner imembranen som stadig må fornyes.
0000 UFb SentrNerv 070401M.book Page 30 Wednesday, July 4, 2007 12:11 PM

KAPITTEL 1 NERVESYSTEMETS BYGNINGSELEMENTER 31
Dendritt
Dendritt
Cellekjerne
Nucleolus
rER (Nissl-korn)
A
B
FIGUR 1.2 Nevronets cellelegeme (soma).Bildene viser to nerveceller fra ryggmargen– en stor og en liten. Merk den store, lysekjernen med tydelig nucleolus. Fargemeto-den som er brukt (tionin), fremstiller barecellelegemet og de tykkeste delene avdendrittene. Verken aksoner eller tynneredendritter er farget. De mørke klumpene icytoplasma (Nissl-korn) er ru endoplasma-tisk retikulum. Bildet er tatt i lysmikroskopav et snitt (dvs. en tynn skive av vev) somer ca. 20 μm (20/1000 mm). Forstørrelsenca. 800 ganger. Diameteren på cellelege-met til den største nervecellen er ca. 40μm.
FIGUR 1.3 Nevronets ultrastruktur. Bildertatt i elektronmikroskop av snitt gjennomcellelegemet til A: en liten nervecelle(øverst) og B: deler av en større nederst.Kjernene (N) inneholder en tydelig nucleo-lus (Nu) og er lyse pga. ekstendert kroma-tin. I cytoplasma sees både ru endoplas-matisk retikulum (rER) og golgiapparat (G)– dette er organeller som er ansvarlige forproteinsyntese. Flere mitokondrier (m)sees også, som tegn på at nervecellenehar høyt energibehov. Det sees flerenerveterminaler (B) som danner synapsermed cellens soma og første del av endendritt (D). I øverste venstre hjørne avdet nederste bildet sees en nerveterminalsom danner synapse med en tynndendritt (D). Gliautløpere (g) «smyger» seginntil nevronenes soma og utløpere. Detsees også noen tynne aksoner (A) omgittav myelin (My). Forstørret ca. 9000(øverst) og 15 000 ganger (nederst).
0000 UFb SentrNerv 070401M.book Page 31 Wednesday, July 4, 2007 12:11 PM
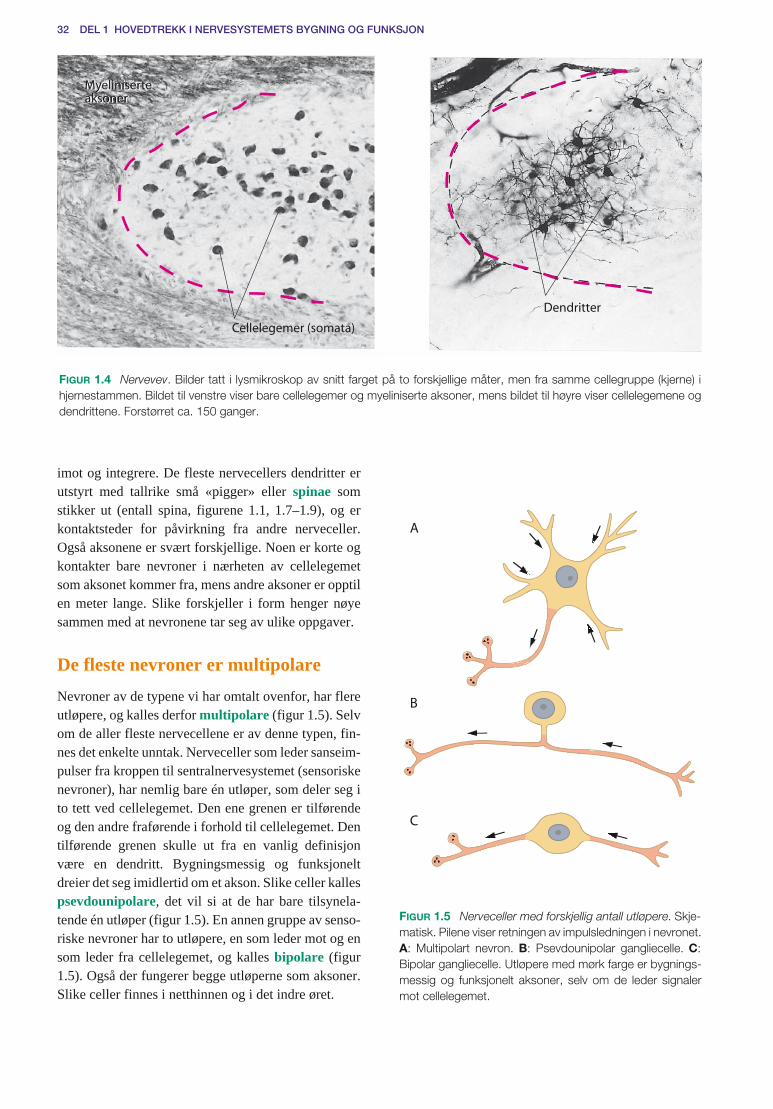
32 DEL 1 HOVEDTREKK I NERVESYSTEMETS BYGNING OG FUNKSJON
imot og integrere. De fleste nervecellers dendritter erutstyrt med tallrike små «pigger» eller spinae somstikker ut (entall spina, figurene 1.1, 1.7–1.9), og erkontaktsteder for påvirkning fra andre nerveceller.Også aksonene er svært forskjellige. Noen er korte ogkontakter bare nevroner i nærheten av cellelegemetsom aksonet kommer fra, mens andre aksoner er opptilen meter lange. Slike forskjeller i form henger nøyesammen med at nevronene tar seg av ulike oppgaver.
De fleste nevroner er multipolare
Nevroner av de typene vi har omtalt ovenfor, har flereutløpere, og kalles derfor multipolare (figur 1.5). Selvom de aller fleste nervecellene er av denne typen, fin-nes det enkelte unntak. Nerveceller som leder sanseim-pulser fra kroppen til sentralnervesystemet (sensoriskenevroner), har nemlig bare én utløper, som deler seg ito tett ved cellelegemet. Den ene grenen er tilførendeog den andre fraførende i forhold til cellelegemet. Dentilførende grenen skulle ut fra en vanlig definisjonvære en dendritt. Bygningsmessig og funksjoneltdreier det seg imidlertid om et akson. Slike celler kallespsevdounipolare, det vil si at de har bare tilsynela-tende én utløper (figur 1.5). En annen gruppe av senso-riske nevroner har to utløpere, en som leder mot og ensom leder fra cellelegemet, og kalles bipolare (figur1.5). Også der fungerer begge utløperne som aksoner.Slike celler finnes i netthinnen og i det indre øret.
Cellelegemer (somata)
Dendritter
FIGUR 1.4 Nervevev. Bilder tatt i lysmikroskop av snitt farget på to forskjellige måter, men fra samme cellegruppe (kjerne) ihjernestammen. Bildet til venstre viser bare cellelegemer og myeliniserte aksoner, mens bildet til høyre viser cellelegemene ogdendrittene. Forstørret ca. 150 ganger.
A
C
B
FIGUR 1.5 Nerveceller med forskjellig antall utløpere. Skje-matisk. Pilene viser retningen av impulsledningen i nevronet.A: Multipolart nevron. B: Psevdounipolar gangliecelle. C:Bipolar gangliecelle. Utløpere med mørk farge er bygnings-messig og funksjonelt aksoner, selv om de leder signalermot cellelegemet.
0000 UFb SentrNerv 070401M.book Page 32 Wednesday, July 4, 2007 12:11 PM

KAPITTEL 1 NERVESYSTEMETS BYGNINGSELEMENTER 33
Kommunikasjon mellom nerveceller foregår ved synapser
Endeforgreningene av et akson har små kølleformedeoppsvulminger som kalles nerveterminaler eller bou-toner (bouton, fransk = knapp) (figurene 1.1, 1.3 og1.6). I noen tilfeller kan nerveterminalen bare være enoppsvulming på aksonet, som så fortsetter videre (figur1.9A). Uansett legger nerveterminalen seg tett inntil
membranen til en annen nervecelle, oftest på endendritt eller på cellelegemet (soma). Et slikt kontakt-sted kalles en synapse (figurene 1.6 og 1.7). I det peri-fere nervesystemet dannes det også synapser mellomaksoner og muskelceller (figurene 11.4 og 11.5). Det erved synapsen at informasjonen overføres fra én nerve-celle til en annen. Dette skjer ikke ved at nerveimpul-sen (som er en elektrisk impuls) overføres direkte, menved frisetting av signalmolekyler som så påvirker
Glia
Akson
Nerveterminal
Synaptiske vesikler
Postsynaptisk membran
Presynaptisk membran
Postsynaptisk nevron
Synaptisk spalte
A B
Synapse
Akson
Dendritt
Cellelegeme (soma)
Nerveterminaler
Presynaptisk nevron
Postsynaptisk nevron
Postsynaptisk fortetning (PSD)
FIGUR 1.6 Synapse. A:Oversikt over nevron medterminaler på postsynaptisknevron B: Skjematisk teg-ning basert på elektron-mikroskopiske bilder somviser de viktigste elemen-tene i en synapse. Sammen-lign med figurene 1.3 og 1.7.
g g
g
g
g
A B
FIGUR 1.7 Aksoner, nerveterminaler og synapser. A: Kontakter mellom nerveterminaler (B) og dendritter (D) danner akso-dendrittiske synapser. Det sees også synapser mellom nerveterminaler og spinae (Sp). Synaptiske vesikler, pre- og postsynap-tisk membran med fortetninger og synaptisk spalte er kjennetegn på en synapse. Merk at den synaptiske spalten er litt videreenn spalten mellom membranene utenfor synapsen. Gliautløpere (G) fyller ut mellom nervecellenes forskjellige utløpere. Leggspesielt merke til hvordan glia omgir synapsene. B: Bunt av umyeliniserte aksoner (A) og en synapse på en spina. De små run-dingene inne i aksonene er mikrotubuli (aksonal transport). Elektronmikroskopiske bilder fra lillehjernebarken. A: Forstørret ca.15 000 B: 30 000 ganger.
0000 UFb SentrNerv 070401M.book Page 33 Wednesday, July 4, 2007 12:11 PM

34 DEL 1 HOVEDTREKK I NERVESYSTEMETS BYGNING OG FUNKSJON
cellen. Stoffet som frisettes, kalles en nevrotransmit-ter eller transmittersubstans (ofte brukes bare trans-mitter). Nevrotransmitteren er lagret i nervetermina-lene i små blærer som kalles synaptiske vesikler(figurene 1.6 og 1.7). Synapsen og nevrotransmitternesvirkemåte blir nærmere omtalt i kapittel 2. Her skal vita for oss synapsens bygning litt nærmere.
Mellom nerveterminalen og den andre nervecellener det en smal spalte på ca. 20 nanometer (dvs. 2/100 000 mm). Denne synaptiske spalten er det selv-følgelig ikke mulig å observere i lysmikroskopet, ogførst med elektronmikroskopet kunne man på 1950-tal-let med sikkerhet fastslå at nervecellene virkelig er se-parate enheter. I elektronmikroskopet ser man også atmembranen på hver side av den synaptiske spalten erfortettet og tykkere enn membranene utenfor synapsen(figurene 1.3 og 1.7). Dette skyldes stor tetthet av pro-teiner, noe som er av betydning for signaloverføringenved synapsen. Nerveterminalens membran, som liggerinn mot spalten, kalles presynaptisk, mens membra-nen til cellen som kontaktes, er postsynaptisk (figur1.6). I den postsynaptiske membranen finnes det spesi-elle mottakermolekyler, reseptorer (proteiner), somnevrotransmitteren binder seg til. Dette fremkaller enkortvarig endring i den postsynaptiske cellen, og et sig-nal er overført.
Den postsynaptiske fortetningen (engelsk: post-synaptic density – PSD; figur 1.6) har forbindelser tilcelleskjelettet, blant annet med aktinfilamenter. Detteer sannsynligvis viktig for å forankre reseptorene imembranen og holde dem samlet der frisettingen avnevrotransmitteren foregår. I tillegg finnes det protei-ner som binder seg til tilsvarende proteiner i den presy-naptiske membranen så nerveterminalen holdes påplass. Blant slike proteiner er kadheriner, som også
Aksodendrittiske synapser på spinae
Aksodendrittisk synapse på dendittskaftet
Aksosomatisk synapse
Aksoaksonisk synapse
Aksosomatisk synapse nær aksonet
FIGUR 1.8 Synapser forskjellige steder på nevronet.Synapsens plassering har betydning for dens virkning pådet postsynaptiske nevronet. For eksempel har en synapsenær aksonets initiale segment større innflytelse enn en somsitter distalt på en dendritt. I sistnevnte tilfelle må mangesynapser virke sammen for å endre det postsynaptiske nev-ronets aktivitet.
Presynaptisk nerveterminal
Postsynaptisk nerveterminal
Postsynaptisk nevron
Nerveterminaler Terminaler «en passage»
Dendritt
Akson Glia
A B
Spina
FIGUR 1.9 A: Aksodendrittiske synapser. Flertallet sitter på spinae. B: Aksoaksonisk synapse. Den presynaptiske nervetermi-nalen kan påvirke (oftest hemme) frisettingen av nevrotransmitter fra den postsynaptiske nerveterminalen.
0000 UFb SentrNerv 070401M.book Page 34 Wednesday, July 4, 2007 12:11 PM

KAPITTEL 1 NERVESYSTEMETS BYGNINGSELEMENTER 35
finnes ved andre typer av cellekontakter (som adhe-renskontakter mellom epitelceller). Mange proteiner iden postsynaptiske fortetningen har regulerende virk-ning på synapsens funksjon – for eksempel ved å på-virke reseptorenes egenskaper. Ved læring er den post-synaptiske fortetningen ett av de stedene hvor det skjervarige endringer.
Hvor på nevronet en synapse er plassert, har betyd-ning for virkningen (figur 1.8). Generelt kan vi si at jonærmere begynnelsen av aksonet synapsen sitter, destosterkere er påvirkningen. Synapser som sitter langt utpå dendrittene, har en tilsvarende svak virkning, ogmange må som regel virke sammen for å ha avgjørendeinnflytelse på nevronets funksjonstilstand. Synapsersom sitter på cellelegemet, kalles aksosomatiske,mens synapser på dendrittene kalles aksodendrittiske(figurene 1.3, 1.7 og 1.8). Der hvor dendrittene er ut-styrt med spinae, dannes det alltid én eller et par sy-napser på hver spina (figurene 1.7–1.9). Spinaes funk-sjonelle rolle er ikke fullt forstått, men det er blantannet holdepunkter for at de kan endre både antall ogform i forbindelse med læring. Nerveterminaler kanogså danne synapse med et annet akson (oftest nær en
nerveterminal), og synapsen kalles da aksoaksonisk(figurene 1.8 og 1.9). Dette er en måte å kontrollere ak-tiviteten i bare denne nerveterminalen på, uten å på-virke alle de andre nerveterminalene som hører tilsamme akson (til forskjell fra aksodendrittiske ogaksosomatiske synapser). Aksoaksoniske synapserøker altså presisjonen i signalformidlingen i sentral-nervesystemet.
Det er langt flere synapser på dendrittene enn påsoma, fordi dendrittover flaten er så mye større. Hvernervecelle har som regel et stort antall (mange tusen)synapser på sin overflate, og det er summen av påvirk-ningene fra alle disse som bestemmer nervecellens ak-tivitetsnivå.
To hovedtyper av nevroner: projeksjonsnevroner og internevronerNoen nevroner skal påvirke celler som ligger langtborte, og må derfor ha et tilsvarende langt akson. Slikenevroner kalles projeksjonsnevroner (figur 1.10). Eteksempel på projeksjonsnevroner er slike som bringersignaler fra ryggmargen og ut til musklene. Andre pro-
Kollateral
Projeksjonsnevron
Dendritt
Dendritt
Akson
Akson
Internevron
FIGUR 1.10 To hovedtyper av nevroner. Et projeksjonsnevron sender sitt akson til nev-roner som ligger langt unna, mens et internevrons akson kontakter nevroner som ligger inærheten (innenfor samme gruppe av nerveceller). Tegning av nerveceller som erimpregnert med sølvsalter (golgimetoden). Fra hjernestammen.
FIGUR 1.11 Projeksjonsnevronmed kollateral. Ved å sende avgrener (kollateraler) underveiskan et projeksjonsnevron påvirkecellegrupper (kjerner) som liggerpå forskjellige steder.
0000 UFb SentrNerv 070401M.book Page 35 Wednesday, July 4, 2007 12:11 PM

36 DEL 1 HOVEDTREKK I NERVESYSTEMETS BYGNING OG FUNKSJON
jeksjonsnevroner finnes blant annet i hjernebarken (fi-gur 21.5). Noen av disse sender aksoner til grupper avnevroner i andre deler av hjernebarken, mens andresender sine aksoner til hjernestammen og ryggmargen.Under sitt forløp sender som regel projeksjonsnevro-nets akson ut mindre sidegrener, kollateraler (figu-rene 1.1, 1.11 og 21.5). På den måten kan ett projek-sjonsnevron bringe informasjon til andre nerveceller iflere deler av nervesystemet.
Den andre hovedtypen av nevroner har et kort,sterkt forgrenet akson, og kalles internevron (figurene1.10 og 20.6). Assosiasjonsnevron er navn på internev-roner. Navnet henspiller på at internevronene er inn-skutt mellom nerveceller som ligger nær hverandre. Detjener også til å spre signaler til mange nevroner i en
gruppe. Ved at slike internevroner kan «slås av og på»,øker antallet mulige koblinger mellom nervecellene ien gruppe enormt.
Etter hvert er vi blitt klar over at det ikke alltid eret skarpt skille mellom projeksjonsnevroner og inter-nevroner. Mange nevroner som før ble ansett for å habare lokale grener, har vist seg også å ha en langgren, slik at de både virker på celler i nærheten og påceller langt borte fra dem. Det gjelder for eksempelmange av nevronene i ryggmargen. Tilsvarende hartypiske projeksjonsnevroner i hjernebarken kollate-raler som går av før aksonet forlater cellegruppennevronet ligger i (figur 21.6). Slike nerveceller fun-gerer altså både som internevroner og som projek-sjonsnevroner.
Internevroners oppgaverFigur 1.12 viser hvordan internevroner kan være innskutt i en signalvei. En impuls i de innkommendeaksonene til venstre aktiverer nervecellen a, som så aktiverer b, som så aktiverer c. Man kunne kanskjetenkt seg at det ville gjøre samme nytten med den enklere løsningen vist nederst på figur 1.12, hvorsignalet går direkte fra A til C. Internevronet fører jo til forsinkelse av impulstrafikken, noe som ofte ikkeer noen fordel. Den vesentligste fordelen med ordningen er imidlertid at den gir muligheter for størrevariasjon i respons som svar på stimuli. Nevronet c er ikke «diktatorisk» styrt av a, fordi internevronetb også påvirkes av nevroner i andre deler av sentralnervesystemet, slik at signaltrafikken kan økes ellerhemmes. Mange «har et ord med i laget» om hva som skal formidles videre fra c. Samme stimulus til akan på den måten gi forskjellig resultat under forskjellige omstendigheter. Dette forklarer blant annethvorfor smertestimuli kan oppfattes svært forskjellig i forskjellige situasjoner – nevrongrupper i fleredeler av sentralnervesystemet virker på internevronene som er innskutt i signalveiene som leder smer-tesignaler til bevissthet.
Et annet eksempel på internevroners oppgaver er vist på figur 1.13. Her virker nevronet A tilbakepå seg selv og hemmer, «bremser» – via internevronet B – sin egen aktivitet. Dette hindrer at impuls-frekvensen i nevronet A blir for høy, eller sørger for at det bare sender noen få impulser før det tar enpause. En slik ordning finnes blant annet for motoriske nevroner som styrer muskelkontraksjon (figur11.14).
AC
a cb A
B +
–
+
FIGUR 1.12 Innskutte nevroner i en signalvei. Denenkleste og raskeste signalveien går direkte fra A til C.Signalveien fra a til c er avbrutt av b. Dette gjør at hvasom påvirker c ikke bestemmes diktatorisk av a, menavhenger av hva som påvirker b til enhver tid. Hvis vi ten-ker oss dette som en signalvei for smerte, kan tilstandeni b være påvirket av signaler som øker ved angst.
FIGUR 1.13 Internevron som sørger for negativ tilbake-kobling. Nevronet A aktiverer internevronet B, som i sintur hemmer A. Dette kan være viktig for å begrense akti-viteten i A, fordi for høy aktivitet kan være skadelig fornevronene. Det kan også bidra til å hindre for store sving-ninger i aktivitet, ev. ukontrollerte rytmiske utladninger.
0000 UFb SentrNerv 070401M.book Page 36 Wednesday, July 4, 2007 12:11 PM
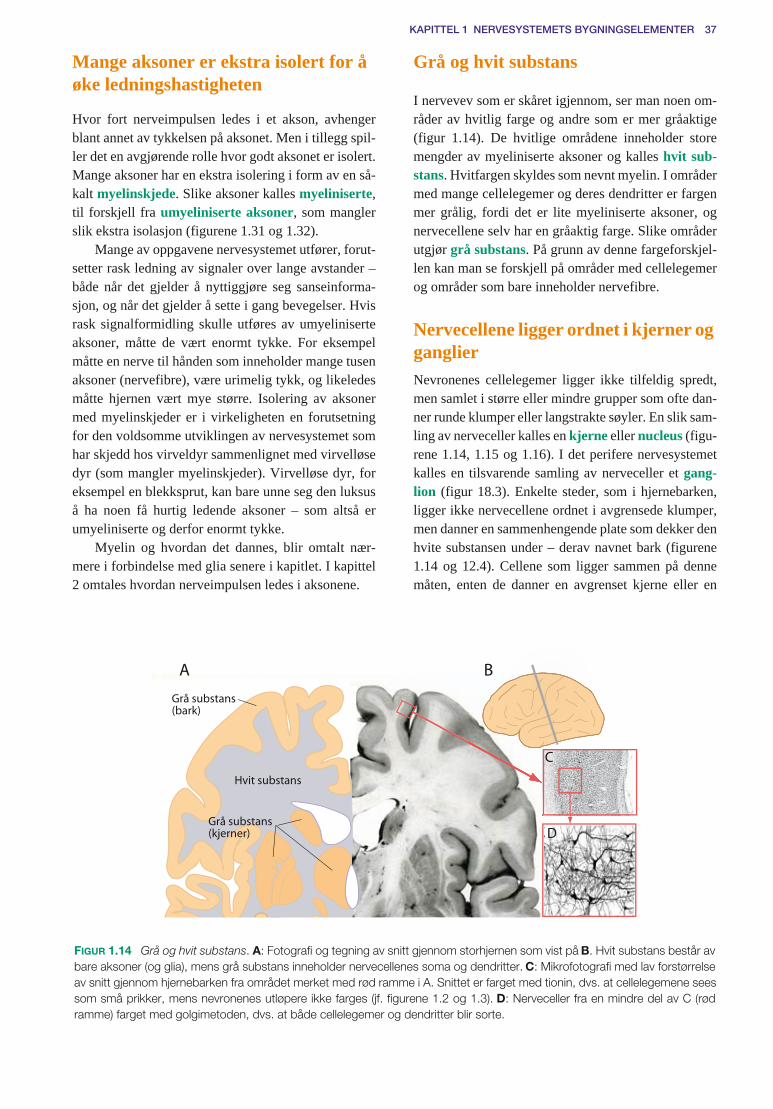
KAPITTEL 1 NERVESYSTEMETS BYGNINGSELEMENTER 37
Mange aksoner er ekstra isolert for å øke ledningshastigheten
Hvor fort nerveimpulsen ledes i et akson, avhengerblant annet av tykkelsen på aksonet. Men i tillegg spil-ler det en avgjørende rolle hvor godt aksonet er isolert.Mange aksoner har en ekstra isolering i form av en så-kalt myelinskjede. Slike aksoner kalles myeliniserte,til forskjell fra umyeliniserte aksoner, som manglerslik ekstra isolasjon (figurene 1.31 og 1.32).
Mange av oppgavene nervesystemet utfører, forut-setter rask ledning av signaler over lange avstander –både når det gjelder å nyttiggjøre seg sanseinforma-sjon, og når det gjelder å sette i gang bevegelser. Hvisrask signalformidling skulle utføres av umyeliniserteaksoner, måtte de vært enormt tykke. For eksempelmåtte en nerve til hånden som inneholder mange tusenaksoner (nervefibre), være urimelig tykk, og likeledesmåtte hjernen vært mye større. Isolering av aksonermed myelinskjeder er i virkeligheten en forutsetningfor den voldsomme utviklingen av nervesystemet somhar skjedd hos virveldyr sammenlignet med virvelløsedyr (som mangler myelinskjeder). Virvelløse dyr, foreksempel en blekksprut, kan bare unne seg den luksuså ha noen få hurtig ledende aksoner – som altså erumyeliniserte og derfor enormt tykke.
Myelin og hvordan det dannes, blir omtalt nær-mere i forbindelse med glia senere i kapitlet. I kapittel2 omtales hvordan nerveimpulsen ledes i aksonene.
Grå og hvit substans
I nervevev som er skåret igjennom, ser man noen om-råder av hvitlig farge og andre som er mer gråaktige(figur 1.14). De hvitlige områdene inneholder storemengder av myeliniserte aksoner og kalles hvit sub-stans. Hvitfargen skyldes som nevnt myelin. I områdermed mange cellelegemer og deres dendritter er fargenmer grålig, fordi det er lite myeliniserte aksoner, ognervecellene selv har en gråaktig farge. Slike områderutgjør grå substans. På grunn av denne fargeforskjel-len kan man se forskjell på områder med cellelegemerog områder som bare inneholder nervefibre.
Nervecellene ligger ordnet i kjerner og ganglierNevronenes cellelegemer ligger ikke tilfeldig spredt,men samlet i større eller mindre grupper som ofte dan-ner runde klumper eller langstrakte søyler. En slik sam-ling av nerveceller kalles en kjerne eller nucleus (figu-rene 1.14, 1.15 og 1.16). I det perifere nervesystemetkalles en tilsvarende samling av nerveceller et gang-lion (figur 18.3). Enkelte steder, som i hjernebarken,ligger ikke nervecellene ordnet i avgrensede klumper,men danner en sammenhengende plate som dekker denhvite substansen under – derav navnet bark (figurene1.14 og 12.4). Cellene som ligger sammen på dennemåten, enten de danner en avgrenset kjerne eller en
Grå substans (bark)
Grå substans (kjerner)
Hvit substans
A B
C
D
FIGUR 1.14 Grå og hvit substans. A: Fotografi og tegning av snitt gjennom storhjernen som vist på B. Hvit substans består avbare aksoner (og glia), mens grå substans inneholder nervecellenes soma og dendritter. C: Mikrofotografi med lav forstørrelseav snitt gjennom hjernebarken fra området merket med rød ramme i A. Snittet er farget med tionin, dvs. at cellelegemene seessom små prikker, mens nevronenes utløpere ikke farges (jf. figurene 1.2 og 1.3). D: Nerveceller fra en mindre del av C (rødramme) farget med golgimetoden, dvs. at både cellelegemer og dendritter blir sorte.
0000 UFb SentrNerv 070401M.book Page 37 Wednesday, July 4, 2007 12:11 PM

38 DEL 1 HOVEDTREKK I NERVESYSTEMETS BYGNING OG FUNKSJON
mindre del av hjernebarken, har felles oppgaver ogdanner derfor en slags funksjonell enhet. Dette skyl-des at nevronene i kjernen har fellestrekk når det gjel-der hvilken informasjon de mottar, og hvor de sendersine beskjeder. Informasjon til nevronene i kjernenkommer som nerveimpulser i tilførende – afferente –aksoner, som så danner synapser på nevronene. Infor-masjonen kjernen sender ut, formidles igjen som ner-veimpulser i nevronenes aksoner til andre kjerner. Ak-soner som går ut fra en kjerne, kalles fraførende ellerefferente. Vi bruker betegnelsene afferent og efferentogså om aksoner som leder henholdsvis inn til og ut frasentralnervesystemet (dvs. sensoriske aksoner somleder signaler fra sanseorganer, er afferente, mens demotoriske aksonene som går til musklene, er efferente).
Aksoner danner baner og nerver
Aksonene fra cellene i en kjerne (eller et ganglion) skalofte til samme sted, og går derfor samlet ut for å ende ién eller flere andre kjerner (slike nevroner hører til grup-pen projeksjonsnevroner omtalt ovenfor). En slik buntav aksoner (nervefibre) som går fra en kjerne til en anneni sentralnervesystemet, kalles en bane, eller på latintractus (figurene 1.15 og 1.16). I det perifere nervesys-temet kalles en tilsvarende bunt av aksoner for en nerve,latin nervus (figurene 1.15, 1.16, 11.1 og 18.3). Vi bru-ker også betegnelsen perifer nerve for å understreke atdet dreier seg om nervefibre i det perifere nervesystemet.Baner danner hvit substans i sentralnervesystemet. Påsamme måte er perifere nerver hvitlige av farge.
Bane (tractus)
Kjerne (nucleus)HJERNESTAMMEN FIGUR 1.15 Kjerne og bane. Bildet tilhøyre er et mikrofotografi (forstørretca. 75 ganger) av et snitt gjennomnedre del av hjernestammen (medullaoblongata), farget slik at cellelegemerog myeliniserte aksoner er mørke.Figuren til venstre viser hvordan kjer-nene på fotografiet egentlig er søylersom ligger på langs av hjerne-stammen. (Den største av kjernenepå bildet sender sine aksoner tiltungemusklene.) Banen som i foto-grafiet er skåret på tvers, går også pålangs av hjernestammen. (Dennespesielle banen formidler signaler fralikevektsorganet til kjerner som styrerøynenes bevegelser.)
Bane (tractus)
Kjerne (nucleus)
Nerve (nervus)
Kjerne
Muskel
HJERNESTAMMEN
RYGGMARGEN
FIGUR 1.16 Kjerner, bane og nerve.Banen på tegningen består av aksonersom går fra en kjerne i hjernestammentil en kjerne i ryggmargen. Aksonenefra denne kjernen forlater sentralnerve-systemet og danner en nerve som gårtil en muskel.
0000 UFb SentrNerv 070401M.book Page 38 Wednesday, July 4, 2007 12:11 PM

KAPITTEL 1 NERVESYSTEMETS BYGNINGSELEMENTER 39
For å forstå oppgavene til en gruppe av nevroner (foreksempel en kjerne i ryggmargen) må vi vite noe om«hvem» (dvs. hvilke andre nevrongrupper) som på-virker den. Vi må også vite «hvem» cellegruppenselv kan påvirke – dvs. hvor den sender sine aksoner.Det betyr at vi må vite noe om organiseringen av ba-nene i sentralnervesystemet – det vi kaller sentralner-vesystemets forbindelses- eller koblingsmønstre. Ide senere kapitlene blir mange enkeltbaner omtalt(baner som formidler sanseimpulser, som kontrolle-rer bevegelser osv.). Her skal vi se på enkelte gene-relle prinsipper for hvordan nervesystemets forbin-delser er organisert.
Divergens, konvergens og summasjon
Et grunnleggende trekk ved sentralnervesystemet er athvert nevron påvirker mange andre (opptil flere tusen)– dvs. at informasjonen spres. Figur 1.17 viser sterktforenklet hvordan sanseimpulser (for eksempel fra fin-gertuppen) ledes inn til ryggmargen i et sensorisk nev-ron (en psevdounipolar gangliecelle, jf. figur 1.5). Im-pulsen fra det sensoriske nevronet spres til mangenevroner (røde) i ryggmargen, mens hver av disse vir-ker på mange andre (blå). Resultatet er altså at signalerfra ett nevron overføres til mange andre. Dette kallesdivergens av forbindelser. Det er altså ikke slik at in-formasjonen bare følger enkle kjeder av nevroner kob-let etter hverandre.
Et annet like viktig prinsipp – konvergens av for-bindelser – er vist på figur 1.18. Det betyr at ett nevronså godt som alltid påvirkes synaptisk fra mange andrenevroner. I eksemplet styrer den motoriske nervecellenkontraksjon av muskelceller, men det kunne vært encelle hvor som helst i sentralnervesystemet. Den moto-riske nervecellen påvirkes både av sanseimpulser (hergjennom to psevdounipolare ganglieceller) og fra nev-roner i sentralnervesystemet (for eksempel nervecelleri hjernebarken som igangsetter viljestyrte bevegelser).Vi ser altså at den motoriske nervecellens aktivitet – og
Sensorisk nevron
Nevroner i ryggmargen
Huden
FIGUR 1.17 Divergens av forbindel-ser. Figuren viser (forenklet) hvordanaksonkollateralene fra en sensorisknervecelle (psevdounipolar ganglie-celle) kontakter mange nerveceller iryggmargen (røde), og at hver av demi sin tur kontakter mange andre. Der-ved spres signalet fra én nervecelle tilsvært mange andre.
Sammenkobling av nevroner og signalveier
Muskelcelle
Motorisk nevron
Sensoriske nevroner med signaler fra kroppen
Nevroner med signaler fra andre deler av sentralnervesystemet
FIGUR 1.18 Konvergens av forbindelser. Som eksempel erher brukt et motorisk nevron i ryggmargen som sender sittakson til tverrstripede muskelceller. Nevronet påvirkes avmange andre, som danner synapser med det fra forskjelligedeler av kroppen (sensoriske nevroner), og fra andre delerav sentralnervesystemet. Det er summen av alle signalenesom konvergerer på det motoriske nevronet, som bestem-mer hvor sterkt det skal påvirke muskelcellene.
0000 UFb SentrNerv 070401M.book Page 39 Wednesday, July 4, 2007 12:11 PM
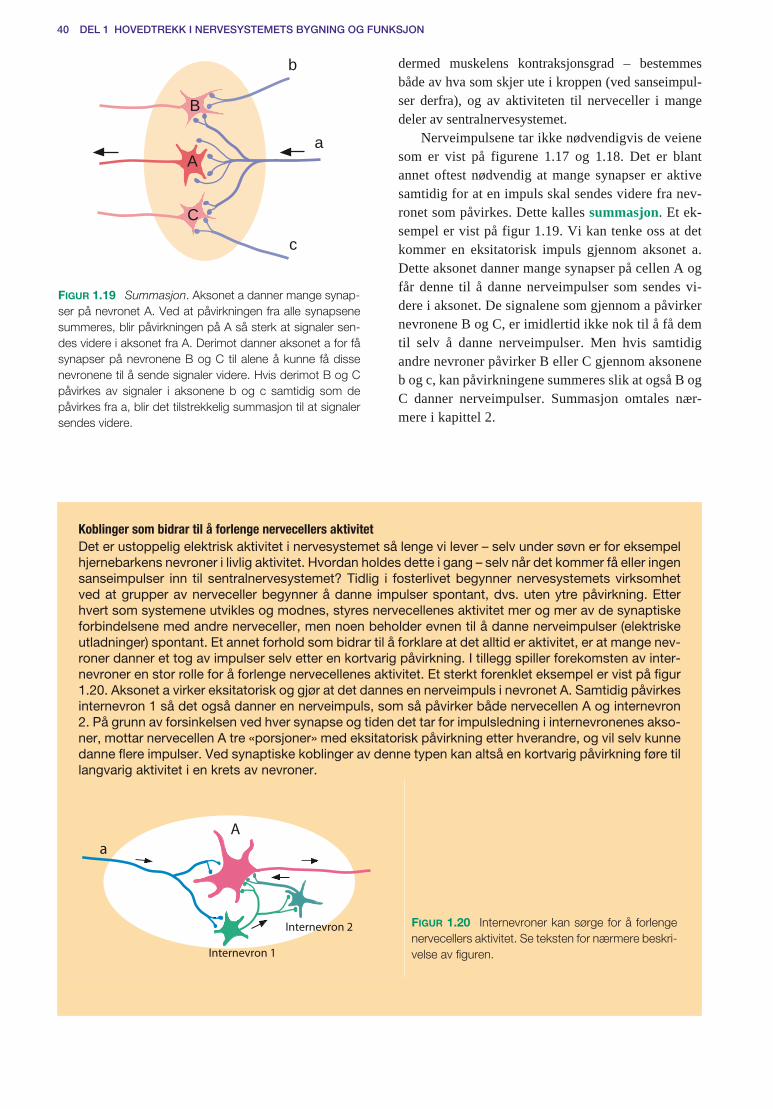
40 DEL 1 HOVEDTREKK I NERVESYSTEMETS BYGNING OG FUNKSJON
dermed muskelens kontraksjonsgrad – bestemmesbåde av hva som skjer ute i kroppen (ved sanseimpul-ser derfra), og av aktiviteten til nerveceller i mangedeler av sentralnervesystemet.
Nerveimpulsene tar ikke nødvendigvis de veienesom er vist på figurene 1.17 og 1.18. Det er blantannet oftest nødvendig at mange synapser er aktivesamtidig for at en impuls skal sendes videre fra nev-ronet som påvirkes. Dette kalles summasjon. Et ek-sempel er vist på figur 1.19. Vi kan tenke oss at detkommer en eksitatorisk impuls gjennom aksonet a.Dette aksonet danner mange synapser på cellen A ogfår denne til å danne nerveimpulser som sendes vi-dere i aksonet. De signalene som gjennom a påvirkernevronene B og C, er imidlertid ikke nok til å få demtil selv å danne nerveimpulser. Men hvis samtidigandre nevroner påvirker B eller C gjennom aksoneneb og c, kan påvirkningene summeres slik at også B ogC danner nerveimpulser. Summasjon omtales nær-mere i kapittel 2.
B
A
C
a
c
b
FIGUR 1.19 Summasjon. Aksonet a danner mange synap-ser på nevronet A. Ved at påvirkningen fra alle synapsenesummeres, blir påvirkningen på A så sterk at signaler sen-des videre i aksonet fra A. Derimot danner aksonet a for fåsynapser på nevronene B og C til alene å kunne få dissenevronene til å sende signaler videre. Hvis derimot B og Cpåvirkes av signaler i aksonene b og c samtidig som depåvirkes fra a, blir det tilstrekkelig summasjon til at signalersendes videre.
Koblinger som bidrar til å forlenge nervecellers aktivitetDet er ustoppelig elektrisk aktivitet i nervesystemet så lenge vi lever – selv under søvn er for eksempelhjernebarkens nevroner i livlig aktivitet. Hvordan holdes dette i gang – selv når det kommer få eller ingensanseimpulser inn til sentralnervesystemet? Tidlig i fosterlivet begynner nervesystemets virksomhetved at grupper av nerveceller begynner å danne impulser spontant, dvs. uten ytre påvirkning. Etterhvert som systemene utvikles og modnes, styres nervecellenes aktivitet mer og mer av de synaptiskeforbindelsene med andre nerveceller, men noen beholder evnen til å danne nerveimpulser (elektriskeutladninger) spontant. Et annet forhold som bidrar til å forklare at det alltid er aktivitet, er at mange nev-roner danner et tog av impulser selv etter en kortvarig påvirkning. I tillegg spiller forekomsten av inter-nevroner en stor rolle for å forlenge nervecellenes aktivitet. Et sterkt forenklet eksempel er vist på figur1.20. Aksonet a virker eksitatorisk og gjør at det dannes en nerveimpuls i nevronet A. Samtidig påvirkesinternevron 1 så det også danner en nerveimpuls, som så påvirker både nervecellen A og internevron2. På grunn av forsinkelsen ved hver synapse og tiden det tar for impulsledning i internevronenes akso-ner, mottar nervecellen A tre «porsjoner» med eksitatorisk påvirkning etter hverandre, og vil selv kunnedanne flere impulser. Ved synaptiske koblinger av denne typen kan altså en kortvarig påvirkning føre tillangvarig aktivitet i en krets av nevroner.
Aa
Internevron 1
Internevron 2 FIGUR 1.20 Internevroner kan sørge for å forlengenervecellers aktivitet. Se teksten for nærmere beskri-velse av figuren.
0000 UFb SentrNerv 070401M.book Page 40 Wednesday, July 4, 2007 12:11 PM

KAPITTEL 1 NERVESYSTEMETS BYGNINGSELEMENTER 41
Parallelle signalveier og resiproke forbindelser
Figur 1.21 illustrerer skjematisk noen prinsipper forhvordan nevrongrupper (kjerner) som ligger et stykkefra hverandre, er sammenkoblet. Det kan for eksempelgjelde kobling mellom forskjellige deler av hjernebar-ken (jf. figur 1.23) eller mellom hjernebarken og kjer-ner i ryggmargen (figur 1.22A). Figur 1.21A viser enordning som synes å gjelde generelt, nemlig det vi kal-ler parallelle signalveier. Det er én direkte vei mellomkjernene N1 og N2, og i tillegg en indirekte vei, synap-tisk avbrutt i andre kjerner. Det betyr at informasjonenN2 mottar, er mer variert og mangfoldig enn om den di-rekte veien formidlet all informasjon. Signaltrafikken iden indirekte veien er jo ikke bare avhengig av det somgår ut fra N1, men også av hvilken påvirkning de inn-skutte kjernene n1 og n2 mottar fra andre deler av ner-vesystemet. Det store antallet parallelle signalveier eren viktig forklaring på den menneskelige hjernesenorme fleksibilitet og kapasitet for informasjonsbe-handling (sammenlignet med dyr med enklere hjerner).
Ved skader i nervesystemet kan slike parallelleveier være av stor betydning for gjenvinning av funk-sjon. Hvis den direkte veien mellom N1 og N2 erbrutt, kan den indirekte, hvis den er bevart, etter hvertta over en del av oppgavene som ble ivaretatt av dendirekte veien. Dette er sannsynligvis noe av grunnla-get for den bedringen som ofte kommer i månedeneetter et hjerneslag.
Et annet viktig forhold er at det ofte er toveisfor-bindelser mellom cellegrupper – det kalles også resip-roke forbindelser (figur 1.21B). Hensikten kan blant
annet være å informere om resultatet av påvirkningen– dvs. tilbakekobling eller «feedback». Dette er sann-synligvis avgjørende for informasjonsbehandlingen,som skjer i et samarbeid mellom nevrongrupper medtoveisforbindelser. I tillegg er toveisforbindelser vik-tige for å hindre ukontrollert aktivitet av nerveceller.Hvis for eksempel påvirkningen den ene veien blir forsterk, sendes beskjed tilbake om å dempe aktiviteten.Derved sikres stabilitet i nervesystemets funksjon.Mange symptomer som opptrer ved nevrologiske syk-dommer, skyldes overaktivitet av nerveceller, og kantil dels forklares ved svikt i slike tilbakekoblings-mekanismer.
Forbindelser mellom de to sidene av sentralnervesystemetEndelig er det viktig at de fleste kjerner har forbindel-ser med begge halvdeler av nervesystemet, de har bila-terale forbindelser (figur 1.22A). Noen slike baner gårkrysset og ukrysset i lik mengde, mens andre baner,som vesentlig går til motsatt – kontralateral – side,også sender noen fibre til samme – ipsilateral – side(se for eksempel figur 1.22). Slike bilaterale forbindel-ser er av betydning for bedring etter ensidige skader isentralnervesystemet. At det må være et utstrakt sam-arbeid mellom de to halvdelene av nervesystemet, blirenda tydeligere når man ser på de store mengdene avdirekte forbindelser mellom tilsvarende deler (for ek-sempel mellom sensoriske barkområder i de to hjerne-halvdelene). Dette kalles kommissuralforbindelser(figur 1.22B), og finnes på alle nivåer av sentralnerve-systemet. Den kraftigste, som forbinder de to halvde-lene av storhjernen med hverandre (corpus callosum,
N1
N2
n1
n2
Parallelle signalveier Resiproke (feedback) forbindelser
A B
FIGUR 1.21 Koblingsmønstre i sentralnervesystemet.Parallelle og resiproke forbindelser.
Kommissuralforbindelser
Bilaterale forbindelser
A B
FIGUR 1.22 Koblingsmønstre i sentralnervesystemet. Bila-terale (tosidige) forbindelser og kommissuralforbindelser.
0000 UFb SentrNerv 070401M.book Page 41 Wednesday, July 4, 2007 12:11 PM

42 DEL 1 HOVEDTREKK I NERVESYSTEMETS BYGNING OG FUNKSJON
figurene 3.26 og 3.27), inneholder rundt 200 millioneraksoner hos mennesket. Dette sikrer at de to hjerne-halvdelene hele tiden samarbeider.
Enkeltnevroner inngår i nevrale nettverkDen enkelte nervecellens ytelse gir bare mening sett isammenheng med de tusenvis av andre nerveceller dener forbundet med. Det er heller ikke slik at én enkeltgruppe (kjerne) av nerveceller alene er ansvarlig for aten oppgave utføres. De fleste av nervesystemets opp-gaver ivaretas ikke av enkelte sentraler eller «sentre»,men ved at mange nevrongrupper samarbeider (ut-veksler informasjon). Slike samarbeidende nevron-grupper kan ligge langt fra hverandre i hjernen. For ek-sempel samarbeider cellegrupper i flere deler avhjernebarken med lillehjernen og de såkalte basalgang-liene om kontroll av bevegelser. Vi bruker betegnelsen
distribuert system om nevronene som er ansvarligefor utførelsen av en bestemt oppgave. Et distribuertsystem er et komplisert nettverk av romlig atskiltenevrongrupper som er forbundet med hverandre – of-test med resiproke (toveis) forbindelser. Figur 1.23 giret meget forenklet eksempel på et nettverk av innbyr-des forbundne nevrongrupper i hjernebarken. Signal-trafikken kan ta mange veier innenfor nettverket, oghver nevrongruppe kan inngå i flere andre nettverk(spesialisert for andre oppgaver). At det er mange inn-ganger til et slikt distribuert system, gjør at det kan ak-tiveres på mange måter. Et eksempel er hvordan etminne eller en assosiasjon kan fremkalles på forskjel-lige måter (en lukt, en lyd, en tanke osv.). Samtidig erdet mange utganger – det vil si at systemet kan virke påmange andre deler av sentralnervesystemet. Et eksem-pel kan være hvordan et bestemt minne som aktiveres,kan fremkalle helt forskjellige handlinger avhengig avsituasjonen minnet fremkalles i.
Toveis forbindelser mellom spesialiserte nevrongrupper
Afferente forbindelser med ulike typer informasjon
Nevrongruppe som er spesialisert for integrasjon av visse typer informasjon
FIGUR 1.23 Distribuerte nettverk i hjernebarken. Tre områder (grupper av nevroner) i hjernebarken er innbyrdes forbundet medtoveisforbindelser (pilene angir retningen av impulstrafikken). Den samlede aktiviteten i områdene er ansvarlig for nettverketsytelse (for eksempel opplevelse av smerte). Figuren illustrerer også at det er mange andre nevrongrupper som virker inn pånettverket og påvirker dets ytelse, mens nettverket i sin tur påvirker flere andre deler av hjernebarken (nettverk som tar seg avandre oppgaver). Sterkt forenklet skjema.
Skader av nevrale nettverkEt viktig kjennetegn ved distribuerte systemer i hjernen er at en begrenset skade av nettverket kanredusere ytelsen, men sjelden eliminerer den fullstendig. Kanskje vil skaden bare merkes ved ekstrastore krav til ytelse – for eksempel når det gjelder bevegelsershastighet, korttidshukommelsens kapa-sitet eller annet. Ved ytterligere reduksjon i antall nevroner som deltar i nettverket, kan det likevel skjeat funksjonen ikke lenger kan ivaretas. Da opptrer brått tydelige symptomer, til tross for at sykdomspro-sessen er gradvis og kanskje har pågått i mange år. Dette sees ved degenerative hjernesykdommerhvor antallet nerveceller i deler av hjernen gradvis reduseres ved celledød. Ved Parkinsons sykdom, foreksempel, opptrer tydelige symptomer først når mer enn to tredjedeler av nervecellene i en bestemtkjerne (substantia nigra) er døde.
0000 UFb SentrNerv 070401M.book Page 42 Wednesday, July 4, 2007 12:11 PM
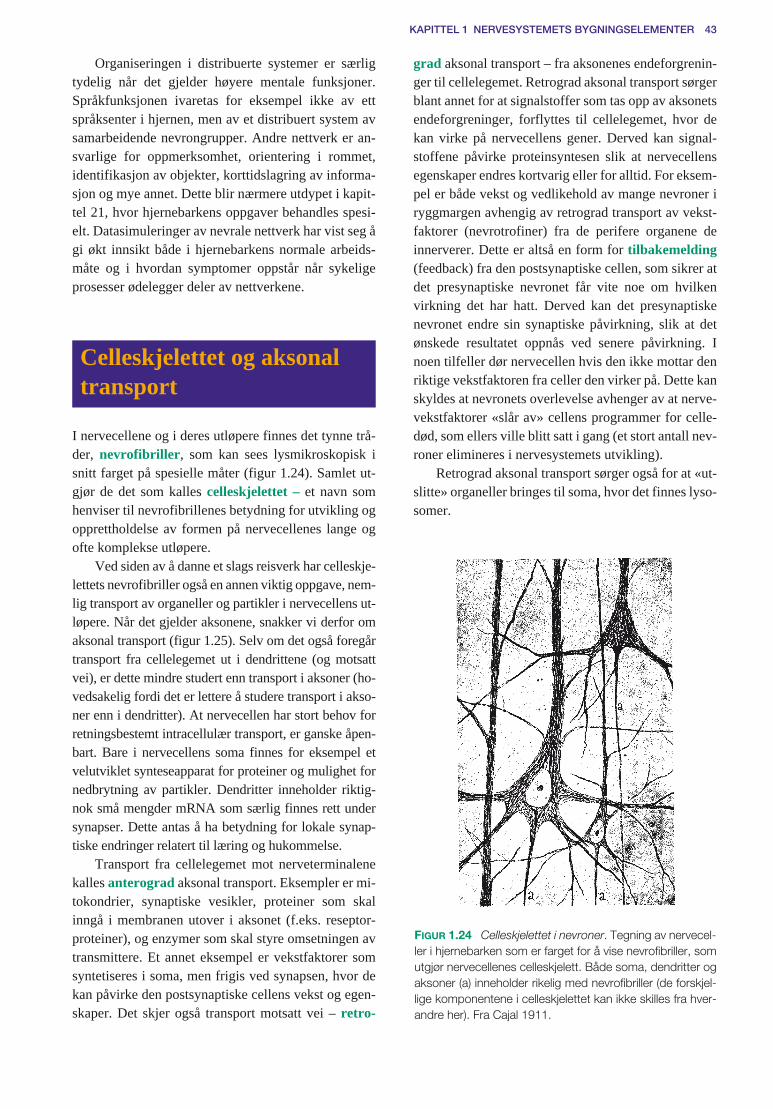
KAPITTEL 1 NERVESYSTEMETS BYGNINGSELEMENTER 43
Organiseringen i distribuerte systemer er særligtydelig når det gjelder høyere mentale funksjoner.Språkfunksjonen ivaretas for eksempel ikke av ettspråksenter i hjernen, men av et distribuert system avsamarbeidende nevrongrupper. Andre nettverk er an-svarlige for oppmerksomhet, orientering i rommet,identifikasjon av objekter, korttidslagring av informa-sjon og mye annet. Dette blir nærmere utdypet i kapit-tel 21, hvor hjernebarkens oppgaver behandles spesi-elt. Datasimuleringer av nevrale nettverk har vist seg ågi økt innsikt både i hjernebarkens normale arbeids-måte og i hvordan symptomer oppstår når sykeligeprosesser ødelegger deler av nettverkene.
I nervecellene og i deres utløpere finnes det tynne trå-der, nevrofibriller, som kan sees lysmikroskopisk isnitt farget på spesielle måter (figur 1.24). Samlet ut-gjør de det som kalles celleskjelettet – et navn somhenviser til nevrofibrillenes betydning for utvikling ogopprettholdelse av formen på nervecellenes lange ogofte komplekse utløpere.
Ved siden av å danne et slags reisverk har celleskje-lettets nevrofibriller også en annen viktig oppgave, nem-lig transport av organeller og partikler i nervecellens ut-løpere. Når det gjelder aksonene, snakker vi derfor omaksonal transport (figur 1.25). Selv om det også foregårtransport fra cellelegemet ut i dendrittene (og motsattvei), er dette mindre studert enn transport i aksoner (ho-vedsakelig fordi det er lettere å studere transport i akso-ner enn i dendritter). At nervecellen har stort behov forretningsbestemt intracellulær transport, er ganske åpen-bart. Bare i nervecellens soma finnes for eksempel etvelutviklet synteseapparat for proteiner og mulighet fornedbrytning av partikler. Dendritter inneholder riktig-nok små mengder mRNA som særlig finnes rett undersynapser. Dette antas å ha betydning for lokale synap-tiske endringer relatert til læring og hukommelse.
Transport fra cellelegemet mot nerveterminalenekalles anterograd aksonal transport. Eksempler er mi-tokondrier, synaptiske vesikler, proteiner som skalinngå i membranen utover i aksonet (f.eks. reseptor-proteiner), og enzymer som skal styre omsetningen avtransmittere. Et annet eksempel er vekstfaktorer somsyntetiseres i soma, men frigis ved synapsen, hvor dekan påvirke den postsynaptiske cellens vekst og egen-skaper. Det skjer også transport motsatt vei – retro-
grad aksonal transport – fra aksonenes endeforgrenin-ger til cellelegemet. Retrograd aksonal transport sørgerblant annet for at signalstoffer som tas opp av aksonetsendeforgreninger, forflyttes til cellelegemet, hvor dekan virke på nervecellens gener. Derved kan signal-stoffene påvirke proteinsyntesen slik at nervecellensegenskaper endres kortvarig eller for alltid. For eksem-pel er både vekst og vedlikehold av mange nevroner iryggmargen avhengig av retrograd transport av vekst-faktorer (nevrotrofiner) fra de perifere organene deinnerverer. Dette er altså en form for tilbakemelding(feedback) fra den postsynaptiske cellen, som sikrer atdet presynaptiske nevronet får vite noe om hvilkenvirkning det har hatt. Derved kan det presynaptiskenevronet endre sin synaptiske påvirkning, slik at detønskede resultatet oppnås ved senere påvirkning. Inoen tilfeller dør nervecellen hvis den ikke mottar denriktige vekstfaktoren fra celler den virker på. Dette kanskyldes at nevronets overlevelse avhenger av at nerve-vekstfaktorer «slår av» cellens programmer for celle-død, som ellers ville blitt satt i gang (et stort antall nev-roner elimineres i nervesystemets utvikling).
Retrograd aksonal transport sørger også for at «ut-slitte» organeller bringes til soma, hvor det finnes lyso-somer.
Celleskjelettet og aksonal transport
FIGUR 1.24 Celleskjelettet i nevroner. Tegning av nervecel-ler i hjernebarken som er farget for å vise nevrofibriller, somutgjør nervecellenes celleskjelett. Både soma, dendritter ogaksoner (a) inneholder rikelig med nevrofibriller (de forskjel-lige komponentene i celleskjelettet kan ikke skilles fra hver-andre her). Fra Cajal 1911.
0000 UFb SentrNerv 070401M.book Page 43 Wednesday, July 4, 2007 12:11 PM

44 DEL 1 HOVEDTREKK I NERVESYSTEMETS BYGNING OG FUNKSJON
Celleskjelettets komponenter
Elektronmikroskopisk og biokjemisk har man analy-sert nærmere hva som danner celleskjelettet. Det dreierseg om proteiner som danner tynne tråder og faller i trehovedgrupper:
1 aktinfilamenter (mikrofilamenter) og assosierteproteiner (ca. 5 nm tykke)
2 mikrotubuli (tynne rør) og assosierte proteiner (ca.20 nm tykke)
3 nevrofilamenter, som er en undergruppe av såkalteintermediære filamenter (ca. 10 nm tykke).
Aktin (mikrofilamenter) finnes blant annet i aksonet.Særlig når aksonet vokser ut i fosterlivet, er aktin viktigfor bevegelsene av aksonets vekstkonus (figur 4.14),som danner en oppsvulming i tuppen av aksonet. Påoppsvulmingen dannes det hele tiden små utløpere som«prøver seg» i forskjellige retninger. Sannsynligvis «le-ter» de etter bestemte molekyler som angir riktig ret-ning for aksonets vekst (aktin finnes i mange celler somhar evne til bevegelse, spesielt rikelig i muskelceller).
Men aktinet er sannsynligvis også av betydning for åopprettholde aksonets form når det først er vokst ut.
Mikrotubuli og mikrotubuliassosierte protei-ner (MAP) finnes i alle slags utløpere, og særlig rikeligi aksoner (figurene 1.7 og 1.31), og spiller en rolle bådefor deres utvekst og opprettholdelse av formen. I til-legg er mikrotubuli nødvendige for transport av orga-neller, proteiner og andre partikler i nervecellens utlø-pere – dvs. for aksonal transport og transport idendrittene (som også går begge veier). Dette er blantannet vist ved at stoffer som ødelegger mikrotubuli(f.eks. kolkisin), hindrer aksonal transport.
Nevrofilamentene vet vi mindre om med hensyntil funksjon. En viktig oppgave er sannsynligvis å opp-rettholde diameteren på tykke myeliniserte aksoner –omtrent som et slags innvendig reisverk (nevrofila-menter utgjør omtrent 10 % av proteinene i aksoner).Det er også påfallende at det ved flere sykdommer inervesystemet hvor nerveceller dør (degenerative syk-dommer), opptrer forandringer i nevrofilamentene.Dette gjelder for eksempel Alzheimers sykdom, hvordet opptrer uorganiserte ansamlinger (som en floke avtråder) av nevrofilamenter i hjernebarken.
Hjernebarken Cerebellum
Hjernestammen Hjernestammen
Injisert sporstoff Injisert sporstoff
Sporstoff i aksonale forgreninger
Sporstoff i cellelegemer
ANTEROGRAD TRANSPORT RETROGRAD TRANSPORT
FIGUR 1.25 Aksonal transport. Mikrofotografiene viser bruk av aksonal transport for å kartlegge forbindelser mellom kjerner isentralnervesystemet. Brøkdelen av en mikroliter av et sporstoff (enzymet pepperrotperoksidase – HRP) injiseres henholdsvis ihjernebarken og lillehjernen. Enzymet tas opp av cellelegemer og nerveterminaler ved endocytose. Blærer med enzym trans-porteres så anterograd (fra hjernebarken til en kjerne i hjernestammen) og retrograd (fra lillehjernebarken til hjernestammen).Tilstedeværelse av HRP vises ved et svart reaksjonsprodukt.
0000 UFb SentrNerv 070401M.book Page 44 Wednesday, July 4, 2007 12:11 PM

KAPITTEL 1 NERVESYSTEMETS BYGNINGSELEMENTER 45
Selv om gliacellene ikke deltar i formidlingen av pre-sis informasjon i nervesystemet, og dermed ikke i ner-vesystemets spesifikke oppgaver, er de av avgjørendebetydning for at nervecellene skal kunne fungere nor-malt. Navnet henspiller på at man fra gammelt avantok at de hadde en slags «lim»-funksjon. Antalletgliaceller i hjernen er faktisk langt høyere enn antalletnevroner. Alle gliacellene har kortere eller lengre utlø-pere (figur 1.26). De fleste oppgavene som man i dagmener gliacellene har, ble foreslått på grunnlag av ob-
servasjoner av deres struktur og beliggenhet for hun-dre år siden. Likevel er det først i de senere årene atman har fått metoder som har gjort det mulig å studeregliacellenes egenskaper direkte. Slike studier har vistat gliacellenes oppgaver er mer mangeartede enn vitrodde, og det mangler fortsatt mye på en full forstå-else av deres oppgaver.
Det er tre hovedtyper av gliaceller, astroglia, oli-godendroglia og mikroglia, som atskiller seg bådeved sin bygning og ved hvilke oppgaver de har. Deulike gliacelletypene skilles også fra hverandre im-muncytokjemisk, fordi de inneholder ulike proteinerknyttet til celleskjelettet.
Mer om aksonal transport og maskineriet som driver denVed å injisere radioaktivt merkede stoffer som opptas av nervecellene, har man kunnet vise at akson-transporten faller i to faser, en rask og en langsom. Den raske går med en hastighet på opptil en halvmeter eller mer per dag, mens den langsomme kan variere mellom 1 og 3 mm per dag. Den raske trans-porten gjelder særlig organeller og vesikler (blærer omgitt av cellemembran), mens den langsommesærlig gjelder enzymer og komponenter av celleskjelettet. Som nevnt er mikrotubuli av spesiell betyd-ning for den aksonale transporten. Hvert mikrotubulusrør er satt sammen av mindre byggesteiner i formav proteinet tubulin. Sammensetningen av tallrike tubulinmolekyler til rør skjer ved hjelp av mikrotubu-liassosierte proteiner (MAP). Disse sørger også for at mikrotubuli forankres til cellemembranen og andredeler av celleskjelettet, blant annet til nevrofilamenter. To typer av MAP som bare finnes i nerveceller –MAP2 og tau – er av spesiell betydning for avstivning av mikrotubuli. Egne typer av MAP sørger for hen-holdsvis anterograd og retrograd transport. Man kaller gjerne disse proteinene «motorer» for aksonaltransport. De er ATP-aser, det vil si at de spalter ATP, og energien fra denne spaltingen gir endring avproteinets form (bevegelse). Derved omdannes altså kjemisk energi til mekanisk. Det som transporte-res, for eksempel vesikler og mitokondrier, forflyttes langs mikrotubuli ved midlertidig binding til prote-iner som stikker ut og beveger seg, det vil si at de «går» langs mikrotubuli. Samme mikrotubulus kantransportere begge veier – det avhenger bare av hvilken «motor» partiklene binder seg til. Proteiner,som alle hører til kinesin-superfamilien, er ansvarlig for anterograd transport – det vil si at de sørger forat partikler beveges langs mikrotubuli fra soma. Forskjellige varianter av kinesin ser ut til å transporterehver sin type «last» – for eksempel er det én type som transporterer mitokondrier, mens en annen trans-porterer forstadier av synaptiske vesikler. Dynein, som er et mer komplekst protein enn kinesin, erhovedansvarlig for retrograd transport, men sannsynligvis deltar også enkelte kinesiner.
Eksperimenter med injeksjon av sporstoffer som transporteres i aksonet og senere kan påvises isnitt av vevet, brukes for å kartlegge forløpet av nervecellenes aksoner, det vil si koblingsmønstrene inervesystemet (figur 1.25).
Gliaceller
Spesielle typer av gliacellerI tillegg til de tre hovedtypene finnes det spesialiserte typer av glia i enkelte deler av nervesystemet.Overflaten av hjernens hulrom (ventrikkelsystemet) er kledd av et lag av sylindriske ependymceller (figur1.27). I retina finnes såkalte müllerske celler, mens Bergman-glia finnes i lillehjernen. I hypofysens bak-lapp er pituicytter en spesialisert form for glia. Olfaktoriske skjedeceller (olfactory ensheathing cells) eren særform av schwannceller som finnes i luktenerven. De er av særlig interesse fordi de ved implan-tasjon i sentralnervesystemet kan stimulere til regenerasjon av aksoner.
0000 UFb SentrNerv 070401M.book Page 45 Wednesday, July 4, 2007 12:11 PM
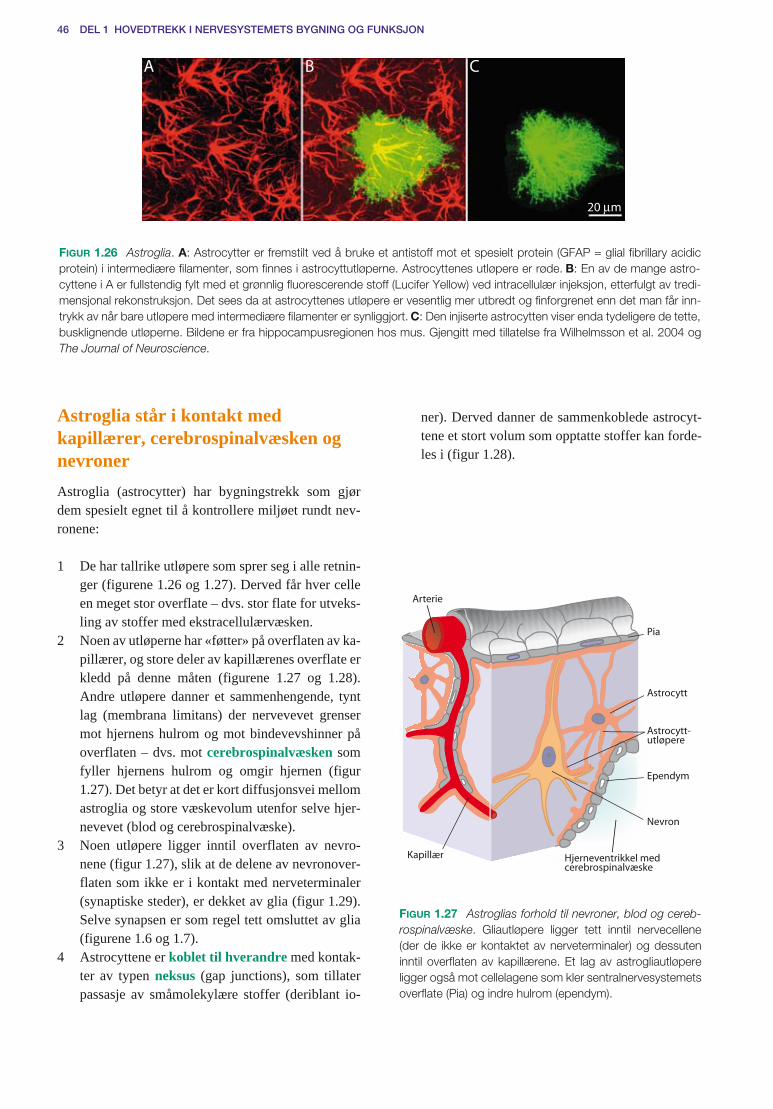
46 DEL 1 HOVEDTREKK I NERVESYSTEMETS BYGNING OG FUNKSJON
Astroglia står i kontakt med kapillærer, cerebrospinalvæsken og nevroner
Astroglia (astrocytter) har bygningstrekk som gjørdem spesielt egnet til å kontrollere miljøet rundt nev-ronene:
1 De har tallrike utløpere som sprer seg i alle retnin-ger (figurene 1.26 og 1.27). Derved får hver celleen meget stor overflate – dvs. stor flate for utveks-ling av stoffer med ekstracellulærvæsken.
2 Noen av utløperne har «føtter» på overflaten av ka-pillærer, og store deler av kapillærenes overflate erkledd på denne måten (figurene 1.27 og 1.28).Andre utløpere danner et sammenhengende, tyntlag (membrana limitans) der nervevevet grensermot hjernens hulrom og mot bindevevshinner påoverflaten – dvs. mot cerebrospinalvæsken somfyller hjernens hulrom og omgir hjernen (figur1.27). Det betyr at det er kort diffusjonsvei mellomastroglia og store væskevolum utenfor selve hjer-nevevet (blod og cerebrospinalvæske).
3 Noen utløpere ligger inntil overflaten av nevro-nene (figur 1.27), slik at de delene av nevronover-flaten som ikke er i kontakt med nerveterminaler(synaptiske steder), er dekket av glia (figur 1.29).Selve synapsen er som regel tett omsluttet av glia(figurene 1.6 og 1.7).
4 Astrocyttene er koblet til hverandre med kontak-ter av typen neksus (gap junctions), som tillaterpassasje av småmolekylære stoffer (deriblant io-
ner). Derved danner de sammenkoblede astrocyt-tene et stort volum som opptatte stoffer kan forde-les i (figur 1.28).
20 μm
A B C
FIGUR 1.26 Astroglia. A: Astrocytter er fremstilt ved å bruke et antistoff mot et spesielt protein (GFAP = glial fibrillary acidicprotein) i intermediære filamenter, som finnes i astrocyttutløperne. Astrocyttenes utløpere er røde. B: En av de mange astro-cyttene i A er fullstendig fylt med et grønnlig fluorescerende stoff (Lucifer Yellow) ved intracellulær injeksjon, etterfulgt av tredi-mensjonal rekonstruksjon. Det sees da at astrocyttenes utløpere er vesentlig mer utbredt og finforgrenet enn det man får inn-trykk av når bare utløpere med intermediære filamenter er synliggjort. C: Den injiserte astrocytten viser enda tydeligere de tette,busklignende utløperne. Bildene er fra hippocampusregionen hos mus. Gjengitt med tillatelse fra Wilhelmsson et al. 2004 ogThe Journal of Neuroscience.
Pia
Astrocytt
Nevron
Ependym
Kapillær
Astrocytt-utløpere
Arterie
Hjerneventrikkel med cerebrospinalvæske
FIGUR 1.27 Astroglias forhold til nevroner, blod og cereb-rospinalvæske. Gliautløpere ligger tett inntil nervecellene(der de ikke er kontaktet av nerveterminaler) og dessuteninntil overflaten av kapillærene. Et lag av astrogliautløpereligger også mot cellelagene som kler sentralnervesystemetsoverflate (Pia) og indre hulrom (ependym).
0000 UFb SentrNerv 070401M.book Page 46 Wednesday, July 4, 2007 12:11 PM

KAPITTEL 1 NERVESYSTEMETS BYGNINGSELEMENTER 47
Astroglia og kontroll av nervecellenes miljø (homøostase)
Den intime kontakten med både kapillærer, cerebrospi-nalvæsken og nevroner gir gode muligheter for utveks-ling av stoffer både mellom gliacellene og nevroneneog mellom gliacellene og blod-/cerebrospinalvæsken(figurene 1.27 og 1.28). At det trengs svært effektivemekanismer for slik utveksling, skyldes blant annet atnervevevet inneholder lite ekstracellulærvæske (rundt20 % av totalvolumet). Elektronmikroskopiske bilderviser at det bare finnes en meget smal spalte mellom decellulære elementene (figurene 1.3 og 1.7). Det betyr atselv små tilførsler av stoffer til ekstracellulærvæskenkan gi store konsentrasjonsendringer. Dette kan haflere uheldige følger. Én uheldig virkning kan være øk-ning av det osmotiske trykket i vevsvæsken. Videre erdet nødvendig for nervecellenes elektriske egenskaperat ionekonsentrasjoner holdes innenfor snevre grenser(dette behandles nærmere i kapittel 2). Til sist – menikke minst – er det nødvendig at nevrotransmittere ikkefår hope seg opp ekstracellulært – de skal bare virkesvært kort etter frisetting ved synapsen, og økte meng-der kan til og med skade nervecellene (nevrotoksisitet).Astroglia har da også reseptorer for flere nevrotrans-mittere, deriblant glutamat, som er den eksitatoriskenevrotransmitteren det finnes mest av i hjernen.
Når det gjelder kontroll av ioner, er særlig konsent-rasjonen av kaliumioner viktig. Nervecellenes eksitabi-litet (dvs. hvor lett de danner nerveimpulser) er nemligspesielt påvirkelig av den ekstracellulære konsentrasjo-nen av K+-ioner. Nervecellene slipper ut K+ når de dan-ner nerveimpulser, slik at langvarig eller høyfrekventaktivitet kan føre til økt K+-konsentrasjon. Økningenkan raskt bli stor, for som nevnt er det ekstracellulærerommet svært begrenset i nervevevet. Astroglia spillerogså en rolle for å kontrollere pH i hjernens ekstracellu-lærvæske ved å ta opp CO2.
De vanligste nevrotransmitterne fjernes fra ekstra-cellulærvæsken i og rundt synapsen ved cellulært opp-
Astroglia kommuniserer med elektriske signalerSelv om glia – til forskjell fra nevroner – ikke formidler presise signaler over lange avstander, kan dedanne kortvarige elektriske strømmer ved åpning av membrankanaler for Ca2+. Fordi astrogliaceller erelektrisk koblet til hverandre, kan et slikt «kalsiumsignal» bre seg forholdsvis raskt blant mange nevro-ner. Man antar derfor at astroglia ved sin påvirkning av ekstracellulær kalsiumkonsentrasjon blant annetkan bidra til å samordne aktiviteten i mange synapser. Frisetting av nevrotransmitteren er nemlig avhen-gig av konsentrasjonen av Ca2+ rundt nerveterminalene (figur 2.8).
Taurin Glutamat K+ H2O
KapillærNeksus (Gap junction)
Cerebrospinalvæske
Nevron
Glia
FIGUR 1.28 Astroglias rolle i nervevevets homøostase.Astrogliacellene er koblet til hverandre med neksus, og dan-ner derved et stort sammenhengende væskevolum somopptatte stoffer kan fordeles i. Astroglia står i kontakt mednevroner, blodet og cerebrospinalvæsken og kan utvekslestoffer med dem. Enkelte viktige stoffer som behandles avglia, er angitt på figuren (transporten går ikke nødvendigvisalltid i pilenes retning). Overskudd av vann, K+ og aminosy-ren taurin kan fordeles til blod og cerebrospinalvæske(strømmen kan også gå den motsatte veien ved behov).Nevrotransmitteren glutamat behandles imidlertid annerle-des. Glutamat omdannes etter opptak i glia til glutamin (avenzymet glutaminsyntetase), derved mister det sin transmit-terfunksjon og blir nøytralt i forhold til nevronene. Glutaminkan derfor gå tilbake til ekstracellulærvæsken og gjenopp-tas i nevronene for nydannelse av glutamat. Siden nervecel-lene trenger store mengder glutamat, er en slik «gjenbruk»en økonomisk måte å sikre tilførselen på. Nest etter gluta-mat er taurin den aminosyren som finnes i høyest konsent-rasjon i nervevevet (dvs. det bidrar vesentlig til vevets osmo-laritet). Taurin har sannsynligvis ingen transmitterfunksjon,men transporten av taurin inn i og ut av astroglia kan væreen mekanisme for regulering av nervecellenes volum. Basertpå E. Nagelhus 1998.
0000 UFb SentrNerv 070401M.book Page 47 Wednesday, July 4, 2007 12:11 PM

48 DEL 1 HOVEDTREKK I NERVESYSTEMETS BYGNING OG FUNKSJON
tak. Opptaket foregår ved hjelp av egne transportpro-teiner i membranen til gliaceller og nevroner. Det erpåvist spesifikke transportører for en rekke forskjelligenevrotransmittere (transportører omtales nærmere i ka-pittel 2). Figur 1.29 gir et inntrykk av gliacellenes ka-pasitet når det gjelder å fjerne nevrotransmitteren glu-tamat fra ekstracellulærvæsken.
Astroglia spiller trolig en sentral rolle også når detgjelder hjernens vannbalanse. Utveksling av småmo-lekylære stoffer (som aminosyren taurin, som det erstore mengder av i nervevevet) kan bidra til å reguleredet osmotiske trykket i ekstracellulærvæsken. I tillegger astroglias membran utstyrt med vannkanaler –akvaporiner – som øker kapasiteten for vannpassasjesterkt (akvaporiner ble først påvist i nyrenes rørsystem,hvor vanntransport er en hovedoppgave). At akvapori-nene er viktige for kontrollen av hjernens vannbalanse,støttes av at de finnes i høyest konsentrasjon på gliaut-løpere som ligger mot kapillærer og mot pia (dvs.cerebrospinalvæsken, se figurene 1.27 og 1.28).
Laget av astrocyttutløpere rundt hjernekapillæreneer også viktig for kapillærenes evne til å hindre stofferi å passere fra blodet til nervevevet (se kapittel 3:Blod–hjerne-barrieren).
Til sist spiller gliacellene viktige roller under ner-vesystemets utvikling, blant annet ved å danne «veier»for nervecellenes vandring og for utvekst av aksonene.
Myelinskjeder dannes av oligodendroglia og schwannceller
Myelinskjeden danner en isolerende sylindrisk hylserundt tykke aksoner og reduserer strømtapet til vevs-væsken under ledning av nerveimpulsen. Dette bidrartil at ledningshastigheten i myeliniserte aksoner er myehøyere enn i de tynne umyeliniserte aksonene. De tyk-keste myeliniserte aksonene leder med rundt 120 m/s,mens umyeliniserte oftest leder med under 1 m/s. I sent-ralnervesystemet er det oligodendroglia som dannermyelinet, mens såkalte schwannceller danner myelin idet perifere nervesystemet.
Myelinskjeden består av tallrike lag av cellemem-bran (figurene 1.30 og 1.31). I de tykkeste myelinskje-dene kan det være mer enn 30 slike myelinlameller.Lamellene er dannet ved at gliaceller har «viklet» segrundt aksonet. Samtidig er cytoplasma i cellen klemtvekk, slik at lagene av cellemembran ligger tett inntilhverandre. Myelinskjedene består derfor overveiendeav lipider, og dette forklarer den hvitlige fargen. Sam-let betegnes materialet som omgir aksonet for myelin.
Cellemembranene som danner myelinlamellene,har en unik sammensetning av lipider og proteiner.Blant annet er det et høyt innhold av kolesterol og gly-kolipider. De siste ser ut til å være avgjørende forblant annet myelinets isolerende egenskaper. Proteiner
FIGUR 1.29 Astrogliautløpere i nervevevet. A: En glutamattransportør er her påvist med immuncytokjemisk teknikk. I dettetynne snittet (ca. 1 μm) fra ryggmargens forhorn er de mørke områdene astrogliautløpere som har høye konsentrasjoner avglutamattransportøren i sin cellemembran. Bildet illustrerer både astroglias evne til å ta opp glutamat fra ekstracellulærvæskenog astroglias enorme overflate mot resten av bestanddelene i nervevevet. Konturen av cellelegemer og dendritter er ujevn fordigliautløpere ligger utenpå nerveterminalene. En ekstra stor nerveterminal er markert med tynn pil. Merk også at vevet inneholdermange kapillærer (*). Cellelegemet til en astrogliacelle er vist med fet pil. B: Til sammenligning vises bildet av et snitt fra sammeområde av ryggmargen, hvor bare cellelegemene er farget (tioninfarging). Verken tynne dendritter, aksoner eller gliautløpere kanskilles fra hverandre i bildet. En gliacelle er merket med fet pil. Bildet A er stilt til rådighet av Jon Storm-Mathisen og Niels Chris-tian Danbolt, Avdeling for anatomi, Institutt for medisinske basalfag, Universitetet i Oslo.
0000 UFb SentrNerv 070401M.book Page 48 Wednesday, July 4, 2007 12:11 PM

KAPITTEL 1 NERVESYSTEMETS BYGNINGSELEMENTER 49
med slektskap til immunglobuliner er integrert i mem-branen og binder yttersidene av lamellene tett sammen.Dette bidrar til å gjøre myelinskjeden kompakt ogugjennomtrengelig. Et annet membranprotein – mye-linbasisk protein (MBP) – forsegler cytoplasmasi-dene i lamellen slik at ikke cytoplasma (med dårlig iso-leringsevne) opptar plass i myelinskjeden. Mus med enmutasjon i BMP-genet danner defekt myelin og får ut-talte bevegelsesforstyrrelser.
Myeliniseringen av aksoner begynner i fosterlivet,men mange nervebaner hos mennesket er ikke ferdigmyelinisert før rundt 2-årsalderen (se kapittel 4: «Mye-linisering»). Først når myeliniseringen er fullført, opp-når nerveforbindelsene sin fulle funksjon.
Ranvierske innsnøringer
I lengdesnitt av aksoner kan man se at myelinskjedener avbrutt med jevne mellomrom (figur 1.30A). Hver
avbrytelse ser ut som en innsnøring av aksonet, og kal-les fra gammelt av en ranviersk innsnøring (eller ran-viersk knute). De ranvierske innsnøringene oppstårved at gliacellene ligger etter hverandre langs aksonetog danner hvert sitt stykke (segment) av myelinskjeden(figur 1.30A).
Elektronmikroskopisk kan man se at aksonet erhelt «nakent», det vil si udekket av myelin, ved inn-snøringen. Her kan ioner vandre gjennom akson-membranen, til forskjell fra der hvor myelinskjedenfinnes. Dette har avgjørende betydning for hvordannerveimpulsen ledes i myeliniserte aksoner, som vikommer nærmere inn på i kapittel 2. Her skal barenevnes at nerveimpulsen «hopper» uhyre raskt frainnsnøring til innsnøring, mens det ved innsnørin-gen blir en liten forsinkelse. Lengden av et segment,det vil si avstanden mellom to ranvierske innsnørin-ger, kan være 0,5 mm eller mer i det perifere nerve-systemet.
Akson
Akson
Myelinlameller
MyelinlamellerAksonets initiale segment Ranviersk innsnøring
Schwannceller
Umyeliniserte aksoner
A B C
FIGUR 1.30 Myeliniserte og umyeliniserte aksoner. A: Skjematisk fremstilling av hvordan myelinskjeden består av oligodendrog-lia eller schwannceller som ligger etter hverandre langs aksonet. Merk den ranvierske innsnøringen mellom to myelinsegmenter.Nerveimpulsen oppstår vanligvis i aksonets initiale segment, og «hopper» så fra innsnøring til innsnøring langs aksonet. B: Viserhvordan myelinskjeden dannes ved at en schwanncelle «vikler» seg rundt aksonet. C: Umyeliniserte aksoner i det perifere ner-vesystemet ligger omgitt av schwannceller, men uten at de danner myelin.
Forskjeller mellom oligodendroglia og schwanncellerSelv om struktur og funksjon av myelinet de danner, er det samme, er ikke oligodendroglia ogschwannceller identiske. En forskjell består i at en enkelt oligodendrogliacelle vanligvis sender flereutløpere som hver bidrar med ett myelinsegment rundt forskjellige aksoner, mens en schwanncellebare danner ett myelinsegment for ett akson. En annen interessant forskjell gjelder deres påvirkning avaksonenes muligheter til å vokse ut igjen etter skade. I det perifere nervesystemet kan et avkuttetakson vokse ut igjen (regenerere) hvis det finnes levedyktige schwannceller på skadestedet. Slikutvekst skjer ikke hvis aksoner skades i sentralnervesystemet, fordi oligodendroglia produserer vekst-hemmende stoffer.
0000 UFb SentrNerv 070401M.book Page 49 Wednesday, July 4, 2007 12:11 PM
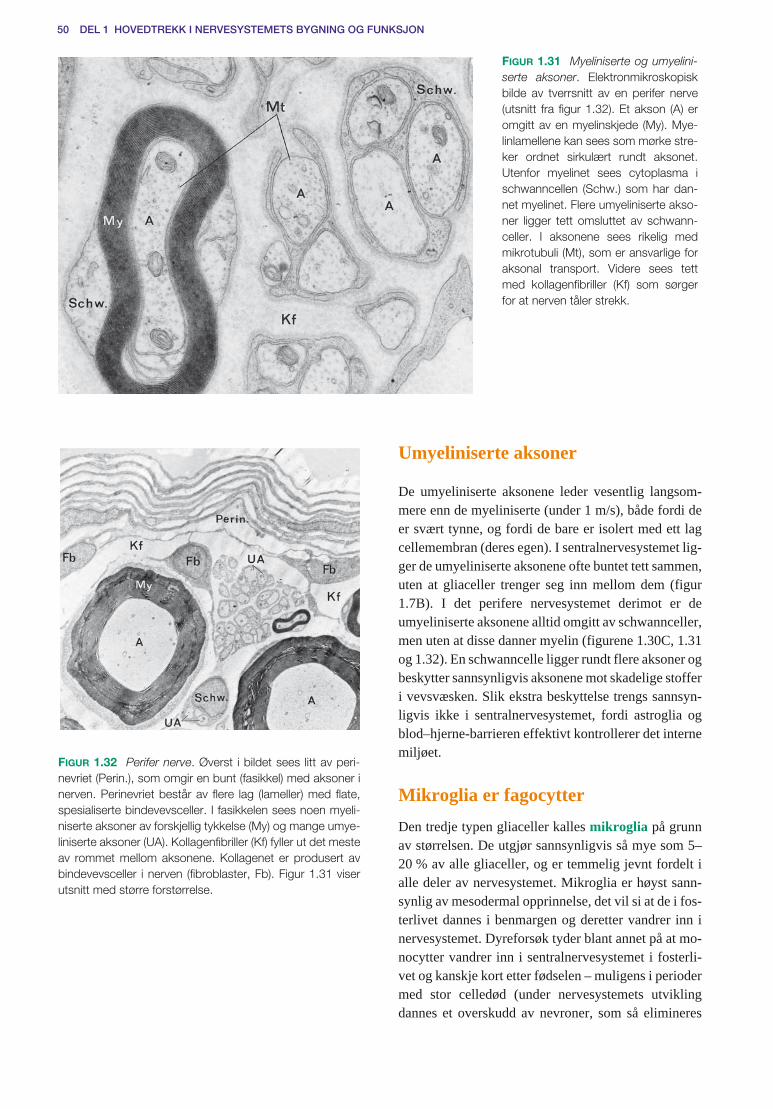
50 DEL 1 HOVEDTREKK I NERVESYSTEMETS BYGNING OG FUNKSJON
Umyeliniserte aksoner
De umyeliniserte aksonene leder vesentlig langsom-mere enn de myeliniserte (under 1 m/s), både fordi deer svært tynne, og fordi de bare er isolert med ett lagcellemembran (deres egen). I sentralnervesystemet lig-ger de umyeliniserte aksonene ofte buntet tett sammen,uten at gliaceller trenger seg inn mellom dem (figur1.7B). I det perifere nervesystemet derimot er deumyeliniserte aksonene alltid omgitt av schwannceller,men uten at disse danner myelin (figurene 1.30C, 1.31og 1.32). En schwanncelle ligger rundt flere aksoner ogbeskytter sannsynligvis aksonene mot skadelige stofferi vevsvæsken. Slik ekstra beskyttelse trengs sannsyn-ligvis ikke i sentralnervesystemet, fordi astroglia ogblod–hjerne-barrieren effektivt kontrollerer det internemiljøet.
Mikroglia er fagocytter
Den tredje typen gliaceller kalles mikroglia på grunnav størrelsen. De utgjør sannsynligvis så mye som 5–20 % av alle gliaceller, og er temmelig jevnt fordelt ialle deler av nervesystemet. Mikroglia er høyst sann-synlig av mesodermal opprinnelse, det vil si at de i fos-terlivet dannes i benmargen og deretter vandrer inn inervesystemet. Dyreforsøk tyder blant annet på at mo-nocytter vandrer inn i sentralnervesystemet i fosterli-vet og kanskje kort etter fødselen – muligens i periodermed stor celledød (under nervesystemets utviklingdannes et overskudd av nevroner, som så elimineres
FIGUR 1.31 Myeliniserte og umyelini-serte aksoner. Elektronmikroskopiskbilde av tverrsnitt av en perifer nerve(utsnitt fra figur 1.32). Et akson (A) eromgitt av en myelinskjede (My). Mye-linlamellene kan sees som mørke stre-ker ordnet sirkulært rundt aksonet.Utenfor myelinet sees cytoplasma ischwanncellen (Schw.) som har dan-net myelinet. Flere umyeliniserte akso-ner ligger tett omsluttet av schwann-celler. I aksonene sees rikelig medmikrotubuli (Mt), som er ansvarlige foraksonal transport. Videre sees tettmed kollagenfibriller (Kf) som sørgerfor at nerven tåler strekk.
FIGUR 1.32 Perifer nerve. Øverst i bildet sees litt av peri-nevriet (Perin.), som omgir en bunt (fasikkel) med aksoner inerven. Perinevriet består av flere lag (lameller) med flate,spesialiserte bindevevsceller. I fasikkelen sees noen myeli-niserte aksoner av forskjellig tykkelse (My) og mange umye-liniserte aksoner (UA). Kollagenfibriller (Kf) fyller ut det mesteav rommet mellom aksonene. Kollagenet er produsert avbindevevsceller i nerven (fibroblaster, Fb). Figur 1.31 viserutsnitt med større forstørrelse.
0000 UFb SentrNerv 070401M.book Page 50 Wednesday, July 4, 2007 12:11 PM

KAPITTEL 1 NERVESYSTEMETS BYGNINGSELEMENTER 51
etter hvert). Etter innvandringen gjennomgår monocyt-tene forandringer – blant annet med utvikling av utlø-pere – slik at de blir til mikroglia, med de særtrekkdenne celletypen har i det voksne sentralnervesyste-met. De beholder imidlertid monocyttenes evne tilfagocytose (dvs. de kan «spise» rester av døde cellerog fremmed materiale). Etter innvandringen ser det uttil at mikrogliacellene er stabile i antall, og at de ikkesenere blir fornyet fra blodbanen (av celler fra benmar-gen). Dette er undersøkt hos dyr hvor benmargen erskiftet ut etter kraftig bestråling, slik at man kan iden-tifisere alle celler som stammer fra benmargen etter ut-skiftingen.
Ved skade av nervevevet øker antallet celler somviser uttalt fagocytose. Økningen i antall slike celler(makrofager) etter skader kan skyldes ny innvandringav monocytter fra blodbanen, men også aktivering avmikroglia. Innvandring av monocytter i slike tilfellerforutsetter sannsynligvis at blod–hjerne-barrieren erskadet (dvs. hjernens kapillærer slipper igjennom be-standdeler i blodet de normalt holder ute).
Sentralnervesystemets reaksjon på skade og betennelse
Vevsskade fører til en betennelsesreaksjon hvor inn-vandring og aktivering av immunsystemets celler spil-ler en sentral rolle. Hensikten med innvandringen er ådrepe mikroorganismer, fjerne rester av dødt vev ogbidra til reparasjon. Likevel arter reaksjonen seg an-nerledes i sentralnervesystemet enn i andre av krop-pens vev og organer. Særlig er det påfallende at inn-vandring av nøytrofile granulocytter ofte mangler, ogselv aktivering av mikroglia og innvandrende mono-cytter til makrofager går langsomt og kan ta flere da-ger. I det hele tatt er immunreaksjoner svakere i sent-ralnervesystemet enn ellers i kroppen. Dette hengersannsynligvis sammen med at sentralnervesystemetmangler lymfedrenasje. Derved holdes ikke immun-systemet løpende informert om tilstanden i vevet, tilforskjell fra nesten alle andre organer. Normalt «pat-ruljeres» nervesystemet bare av et lite antall monocyt-ter og T-lymfocytter fra blodbanen. Muligens utgjør
Multippel skleroseEnkelte sykdommer som rammer nervevev, fører til at myelinskjedene ødelegges. Den vanligste blantslike demyeliniserende sykdommer i sentralnervesystemet er multippel sklerose (MS). (Demyeliniseringav aksoner i perifere nerver omtales nedenfor.) Man kjenner ikke årsaken til multippel sklerose, mensymptomene fremkalles av en betennelsestilstand i hvit substans. Betennelsen rammer «flekkvis»(plakk) på tilsynelatende tilfeldige steder. Sykdommen har hos de fleste pasientene et svingende forløp,hvor perioder med stor sykdomsaktivitet veksler med perioder med lav aktivitet og tilbakegang avsymptomer. Når betennelsesprosessen rammer et område, forsvinner myelinskjedene, mens i alle fallde fleste aksonene forblir intakte. Symptomene som opptrer, forklares vanligvis med at tapet av myelinsvekker eller blokkerer impulsledningen i aksonene. (Det er likevel ikke alltid en klar sammenheng mel-lom graden av demyelinisering og pasientens symptomer.) Ofte begynner sykdommen med at syns-nerven angripes, med delvis blindhet som det første pasienten merker. Dette er som regel forbigående,men etter hvert rammes hos de fleste pasienter andre nervebaner, blant annet slike som styrer beve-gelser, med lammelser til følge. Når sykdomsaktiviteten avtar, kan det skje i alle fall en delvis nydan-nelse av myelin der hvor aksonene fortsatt er uskadd (remyelinisering), og symptomene bedres ellerforsvinner helt for kortere eller lengre tid. Ved gjentatte episoder med betennelse ødelegges etter hvertogså mange aksoner. Dette forklarer at hos de fleste pasientene blir symptomene etter hvert vedva-rende og gradvis mer invalidiserende. Vi vet ikke om aksonene ødelegges som følge av myelintapet,eller om de angripes direkte av immunsystemet, men sannsynligvis spiller begge mekanismene en rolle.
MS regnes blant de såkalte autoimmune sykdommene, hvor immunsystemet angriper kroppensegne celler – i dette tilfellet altså særlig oligodendroglia. Det drives intens forskning for å klargjøre hvor-dan dette egentlig foregår. Både T-lymfocytter, mikroglia og endotelceller i hjernens kapillærer er invol-vert i kontrollen av betennelsesprosessen ved MS. En rekke cytokiner er sannsynligvis også involvert,men man har ikke kunnet utpeke én som er hovedansvarlig. Tumornekrosefaktor (TNFα) ble for eksem-pel antatt å være direkte årsak til demyelinisering, og enkelte dyreforsøk tydet på at stoffer som hem-met TNFα, kunne hindre tap av myelin. Men sammenhengen mellom TNFα og demyelinisering er likeveltvilsom, fordi transgene mus som overproduserer TNFα, får massiv betennelse i hjernen uten tap avmyelin. Beta(β)-interferon, et annet cytokin, brukes terapeutisk og synes å redusere hyppigheten avsykdomsepisoder. De cellulære virkningene av β-interferon er imidlertid komplekse, med en blandingav stimulerende og hemmende effekter på blant annet mikroglia. Man kan derfor ikke foreløpig forklarehvordan det virker ved MS.
0000 UFb SentrNerv 070401M.book Page 51 Wednesday, July 4, 2007 12:11 PM

52 DEL 1 HOVEDTREKK I NERVESYSTEMETS BYGNING OG FUNKSJON
disse forholdene en tilpasning for å hindre at nevro-nene skades av potente stoffer som frigis fra nøytrofilegranulocytter og aktiverte makrofager. Blant annet fårødem – som ellers er en sentral komponent ved beten-nelsesreaksjoner – lett fatale konsekvenser i sentral-nervesystemet, som har minimale utvidelsesmulighe-ter inne i skallen. Likevel kan det som nevnt opptreimmunreaksjoner i sentralnervesystemet, til dels medmeget alvorlige konsekvenser (som ved multippelsklerose).
Astroglias rolle ved skade er sannsynligvis i førsterekke å øke sine normale oppgaver når det gjelder åholde miljøet rundt nervecellene konstant. Ved vevs-ødeleggelse – enten det skyldes blødning, direkte knu-sing eller stopp i sirkulasjonen – øker lekkasjen avioner og nevrotransmittere til ekstracellulærvæsken,og astroglia øker sitt opptak. Fordi stoffene som tasopp, er osmotisk aktive, kan dette føre til oppsvulmingav astroglia allerede de første sekundene til minutteneetter en skade (hvis skaden er så pass alvorlig at astrog-lias normale kapasitet overskrides). Dette kan altsåbidra til volumøkning av hjernen (hjerneødem), som eren fryktet komplikasjon ved hjerneskader. På lengresikt danner astroglia et slags arrvev der hvor nevronerer gått til grunne.
I kapittel 4 kommer vi inn på mer langsiktige pro-sesser i nervesystemet som bidrar til at mye av taptfunksjon ofte kan gjenvinnes etter vevsødeleggelse ihjernen.
Perifere nerver er bygd for å beskytte aksoneneNervevevet er i frisk tilstand temmelig bløtt og gelé-aktig. Det har ingen mekanisk styrke i seg selv. Nårdet gjelder sentralnervesystemet, sørger beliggen-heten i skallen og virvelkanalen samt «innpakning» ibindevevshinner for den nødvendige mekaniske be-
skyttelsen (figurene 1.27, 3.1 og 3.6). Ute i kroppenkan imidlertid buntene av aksoner, dvs. nervene, bliutsatt for ganske store mekaniske påkjenninger der deofte ligger ganske overflatisk og også utsettes forstrekk ved bevegelser av kroppen (spesielt armer ogben). Aksoner tåler bare å forlenges litt før deres led-ningsevne forstyrres, eller de til og med ryker. For åhindre dette inneholder nervene et flettverk av storemengder av bindevevstråder (kollagenfibriller, figur1.31) som er spesielt egnet til å tåle strekkpåkjennin-ger. Dette forklarer hvorfor nervene blir så mye tyk-kere idet de trer ut fra skallen eller virvelkanalen –innenfor trenger de ikke å inneholde kollagen, og gjørdet da heller ikke.
Bindevevselementene i en perifer nerve er ordnet iforskjellige lag. Ytterst er det et kraftig lag av fast fib-ret bindevev som kalles epinevriet. Innenfor liggeraksonene ordnet i fasikler (bunter), omgitt av etganske spesielt bindevev, perinevriet (figur 1.32).Inne i fasiklene finnes det mer sparsomt med bindevev,endonevriet, som inneholder kapillærer. Perinevrietbestår, foruten av langsgående kollagenfibre, av flerelag med avflatede celler som danner lameller i en slagshylse. Mellom disse cellene finnes det cellekontakter(okkludenskontakter eller «tight junctions»), slik at pe-rinevriet representerer en passasjehindring mellomaksonene og vevet utenfor. I tillegg er kapillærene i en-donevriet ekstra tette, slik at de hindrer mange stoffer iblodbanen i å virke på aksonene. Det er altså en blod–nerve-barriere, tilsvarende den som finnes i hjernen(blod–hjerne-barrieren).
Både perinevriet og blod–nerve-barrieren bidrar tilat miljøet rundt aksonene holdes stabilt, slik at deresledningsevne ikke forstyrres. Beskyttelsen av akso-nene i det perifere nervesystemet er likevel ikke så ef-fektiv som i sentralnervesystemet, noe som kanskjekan forklare at perifere nerver relativt ofte rammes avsykdommer.
Aids og sentralnervesystemetSykdommen aids (Acquired Immune Deficiency Syndrome), som fremkalles av hiv (humant immun-sviktvirus), rammer blant annet også nervesystemet. Til tross for at hiv ikke infiserer nerveceller, opptrerdet likevel betydelig nevrondød, uten at det påvises andre infeksjoner i sentralnervesystemet. Pasien-tene kan få forstyrrelser både av bevegelser, sansning og mentale funksjoner. Viruset kommer sann-synligvis inn i sentralnervesystemet ved infiserte T-lymfocytter. Som nevnt «patruljeres» sentralnerve-systemet av et lite antall T-lymfocytter og monocytter. Mikroglia, som så infiseres av hiv, skillersannsynligvis ut toksiske stoffer som dreper nevroner – muligens ved å binde seg til den såkalteNMDA-reseptoren (se «NMDA-reseptoren: formidler både av læring og av celleskade» i kapittel 2, og«Celleskader ved hjerneiskemi» i kapittel 4). Mikroglia har nemlig det såkalte CD4-antigenet – som bin-der hiv – på overflaten.
0000 UFb SentrNerv 070401M.book Page 52 Wednesday, July 4, 2007 12:11 PM

KAPITTEL 1 NERVESYSTEMETS BYGNINGSELEMENTER 53
Sykdommer som rammer perifere nerverNevropati er en fellesbetegnelse for sykdommer som rammer perifere nerver. Nevropatier kan hamange ulike årsaker, men uansett oppheves ledningsevnen til aksonene permanent eller midlertidig.Derved opptrer det symptomer i form av lammelser og føletap (ved noen former opptrer bare lammel-ser eller bare føletap, og ved noen nevropatier kan det opptre plagsomme smerter, til tross for tap avaksoner). Nevropatier kan blant annet opptre ved metabolske sykdommer som sukkersyke og vedvisse typer av forgiftninger. Mange tilfeller skyldes at immunsystemet angriper aksonene eller myelin-skjedene. Dette kan noen ganger opptre etter en infeksjonssykdom eller hos kreftpasienter, sannsyn-ligvis fordi det er dannet antistoffer som «tar feil» og reagerer med antigener knyttet til myelinet elleraksonmembranen. Fordi aksoner uttrykker forskjellige antigener avhengig av forhold som hvor i krop-pen de befinner seg, om de er motoriske eller sensoriske, og om de er tykke eller tynne, kan man forståhvorfor nevropatier ofte rammer bare enkelte nerver eller typer av aksoner. Hvis for eksempel baremotoriske aksoner er skadet, ytrer sykdommen seg bare ved lammelser, mens hvis bare sensoriskeaksoner er skadet, blir det forstyrrelser bare av hudens følesans og av leddsans. Nevropatier kan ogsåramme undergrupper av sensoriske aksoner – for eksempel bare tynne fibre som formidler smerte- ogtemperatursans, eller slike som leder fra ledd og muskler, mens de som leder fra huden, er spart (enpasient med en slik nevropati er beskrevet i kapittel 6: «Kliniske eksempler på tap av somatosensoriskinformasjon»). Nevropatier kan også skyldes genetisk betinget feil i samspillet mellom schwanncellerog aksoner. En stor gruppe nevropatier hos mennesker er arvelige, blant dem den såkalte Char-cot-Marie-Tooths sykdom (også kalt peroneal muskelatrofi). Sykdommen begynner gjerne før 20-års-alderen og fører til langsomt økende lammelser og føletap som begynner distalt i bena. Symptomeneskyldes at aksonene mister myelin og degenererer. De fleste pasientene med denne sykdommen haren endring (dobling) av et gen som koder for et spesielt myelinprotein (PMP-22). Dyremodeller medoverekspresjon av PMP-22 viser at doblingen av dette genet alene fører til defekt myelinisering ogsymptomer som svarer til dem man ser hos mennesker med Charcot-Marie-Tooths sykdom.
0000 UFb SentrNerv 070401M.book Page 53 Wednesday, July 4, 2007 12:11 PM


















