
Fluctuation of Lower Limb Movement in the MRI Bore ...
6
Fluctuation of Lower Limb Movement in the MRI Bore: Different Contributions of the Cortical and Subcortical Locomotor Regions Akira TOYOMURA, *, **, # Koichi YOKOSAWA, **, *** Shinya KURIKI *** Abstract Bipedal locomotion requires an automatic process for rhythmic motor generation, as well as volitional and precise motor control for purposeful action. The neural substrates for these two distinct locomotion controls in humans are still unclear. This functional magnetic resonance imaging (MRI) study investigated the neural activity of the locomotor-related brain regions along with participants’ lower limb behavior. Participants lay face-up in the MRI bore and moved their legs up and down at the knees under two conditions. Under the non-obstacle condition, they moved their legs while watching a point-of-view video that showed walking straight along a corridor at a constant speed. On the other hand, under the obstacle condition, the point-of-view video showed walking while avoiding obstacles in the passage. Two white markers were pasted on the soles of the participants’ feet and images were captured by a video camera throughout the experiment to track the participants’ lower limb movements. The horizontal fluctuation of lower limb movement was extracted offline. We assumed that the horizontal fluctuation reflected participants’ volitional behavior of lower limb movement. The results showed that the obstacle condition elicited greater fluctu- ation of lower limb movement compared to the non-obstacle condition, and elicited brain activation in wider areas including the parietal and occipital lobes compared to the non-obstacle condition. The involvement of the parietal and occipital lobes in voli- tional control is consistent with findings of previous animal studies. In a correlation analysis between the signal change extract- ed from the locomotor-related cortical/subcortical regions and the horizontal fluctuation of the feet, activities in the cortical mo- tor areas (primary motor cortex, premotor cortex, and supplementary motor area) showed weak but significant correlation with the fluctuation of lower limb movement. Activities in the cerebellar and mesencephalic locomotor regions showed no significant correlation, but almost constant activation irrespective of fluctuation of the lower limbs. These results suggest that the parietal, occipital cortices, and cortical motor areas are involved in volitional control. Keywords: lower limb movement, volitional control, cortical and subcortical locomotor regions, functional magnetic resonance imaging. Adv Biomed Eng. 6: pp. 15–20, 2017. 1. Introduction Human locomotion is fundamental to generating goal-directed behavior to support various life activities [1]. Although the be- havioral and mechanical aspects of bipedal locomotion have been widely investigated in previous studies, the neural mechanisms of human bipedal locomotor function is not sufficiently understood. The motor generation/regulation functions of locomotion are known to recruit various locomotion-related regions and net- works in the brain, including the cortical and subcortical areas [2, 3]. Regions in the deep brain, such as the basal ganglia and brain stem, also play important roles in locomotion [4, 5]. Therefore, appropriate measurement methods of activities from deep brain regions are necessary for understanding the neural mechanisms of locomotion. Magnetic resonance imaging (MRI) is one of the non-invasive methods that can be used for this purpose. However, a crucial shortcoming of this method is that volunteers usually lie on the bed during scanning, which makes it difficult for them to perform locomotion control. Some recent studies have reported neural correlates of locomotion and postural control using tasks involving mental imagery [5–7]. Another study has shown that slight lower limb movement in the MRI system also induces ac- tivity in the locomotor-related brain areas [8]. However, detailed mechanisms of locomotion control in the brain in humans remain unclear. Bipedal locomotion has two distinct aspects from the view- point of consciousness [1]. One is an automatic process of rhyth- mic motor generation, which primarily allows movement from point to point. The other is volitional control driven by cognitive processes. This control allows precise lower limb control and achievement of purposeful movement, and is therefore important for the selection of routes to avoid obstacles. Emotion also works as a trigger to initiate locomotion in struggle and escape behav- iors. Ordinarily, individuals move in accordance with the auto- matic process, and at the same time intentionally control their locomotion behavior using volitional control. The neural mechanisms of these controls have been pro- posed based on animal studies and study of impaired locomotion in humans [1]. The automatic process is assumed to be supported primarily by the neural system in subcortical regions and the cer- This study was presented at the Symposium on Biomedical Engi- neering 2016, Asahikawa, September, 2016. Received on July 24, 2016; revised on November 5, 2016; accept- ed on December 26, 2016. * Graduate School of Health Sciences, Gunma University, Mae- bashi, Japan. ** Research and Education Center for Brain Science, Hokkaido Uni- versity, Sapporo, Japan. *** Faculty of Health Sciences, Hokkaido University, Sapporo, Japan. # 3–39–22 Showa-machi, Maebashi, Gunma 371–8514, Japan. E-mail: [email protected] Original Paper Advanced Biomedical Engineering 6: 15–20, 2017. DOI:10.14326/abe.6.15
Transcript of Fluctuation of Lower Limb Movement in the MRI Bore ...

Fluctuation of Lower Limb Movement in the MRI Bore: Different Contributions of the Cortical and Subcortical Locomotor Regions
Akira TOYOMURA,*, **, # Koichi YOKOSAWA,**, *** Shinya KURIKI***
Abstract Bipedal locomotion requires an automatic process for rhythmic motor generation, as well as volitional and precise motor control for purposeful action. The neural substrates for these two distinct locomotion controls in humans are still unclear. This functional magnetic resonance imaging (MRI) study investigated the neural activity of the locomotor-related brain regions along with participants’ lower limb behavior. Participants lay face-up in the MRI bore and moved their legs up and down at the knees under two conditions. Under the non-obstacle condition, they moved their legs while watching a point-of-view video that showed walking straight along a corridor at a constant speed. On the other hand, under the obstacle condition, the point-of-view video showed walking while avoiding obstacles in the passage. Two white markers were pasted on the soles of the participants’ feet and images were captured by a video camera throughout the experiment to track the participants’ lower limb movements. The horizontal �uctuation of lower limb movement was extracted of�ine. We assumed that the horizontal �uctuation re�ected participants’ volitional behavior of lower limb movement. The results showed that the obstacle condition elicited greater �uctu-ation of lower limb movement compared to the non-obstacle condition, and elicited brain activation in wider areas including the parietal and occipital lobes compared to the non-obstacle condition. The involvement of the parietal and occipital lobes in voli-tional control is consistent with �ndings of previous animal studies. In a correlation analysis between the signal change extract-ed from the locomotor-related cortical/subcortical regions and the horizontal �uctuation of the feet, activities in the cortical mo-tor areas (primary motor cortex, premotor cortex, and supplementary motor area) showed weak but signi�cant correlation with the �uctuation of lower limb movement. Activities in the cerebellar and mesencephalic locomotor regions showed no signi�cant correlation, but almost constant activation irrespective of �uctuation of the lower limbs. These results suggest that the parietal, occipital cortices, and cortical motor areas are involved in volitional control.
Keywords: lower limb movement, volitional control, cortical and subcortical locomotor regions, functional magnetic resonance imaging.
Adv Biomed Eng. 6: pp. 15–20, 2017.
1. Introduction
Human locomotion is fundamental to generating goal-directed behavior to support various life activities [1]. Although the be-havioral and mechanical aspects of bipedal locomotion have been widely investigated in previous studies, the neural mechanisms of human bipedal locomotor function is not suf�ciently understood. The motor generation/regulation functions of locomotion are known to recruit various locomotion-related regions and net-works in the brain, including the cortical and subcortical areas [2, 3]. Regions in the deep brain, such as the basal ganglia and brain stem, also play important roles in locomotion [4, 5]. Therefore, appropriate measurement methods of activities from deep brain regions are necessary for understanding the neural mechanisms of
locomotion. Magnetic resonance imaging (MRI) is one of the non-invasive methods that can be used for this purpose. However, a crucial shortcoming of this method is that volunteers usually lie on the bed during scanning, which makes it dif�cult for them to perform locomotion control. Some recent studies have reported neural correlates of locomotion and postural control using tasks involving mental imagery [5–7]. Another study has shown that slight lower limb movement in the MRI system also induces ac-tivity in the locomotor-related brain areas [8]. However, detailed mechanisms of locomotion control in the brain in humans remain unclear.
Bipedal locomotion has two distinct aspects from the view-point of consciousness [1]. One is an automatic process of rhyth-mic motor generation, which primarily allows movement from point to point. The other is volitional control driven by cognitive processes. This control allows precise lower limb control and achievement of purposeful movement, and is therefore important for the selection of routes to avoid obstacles. Emotion also works as a trigger to initiate locomotion in struggle and escape behav-iors. Ordinarily, individuals move in accordance with the auto-matic process, and at the same time intentionally control their locomotion behavior using volitional control.
The neural mechanisms of these controls have been pro-posed based on animal studies and study of impaired locomotion in humans [1]. The automatic process is assumed to be supported primarily by the neural system in subcortical regions and the cer-
This study was presented at the Symposium on Biomedical Engi-neering 2016, Asahikawa, September, 2016. Received on July 24, 2016; revised on November 5, 2016; accept-ed on December 26, 2016.
* Graduate School of Health Sciences, Gunma University, Mae-bashi, Japan.
** Research and Education Center for Brain Science, Hokkaido Uni-versity, Sapporo, Japan.
*** Faculty of Health Sciences, Hokkaido University, Sapporo, Japan.
# 3–39–22 Showa-machi, Maebashi, Gunma 371–8514, Japan. E-mail: [email protected]
Original PaperAdvanced Biomedical Engineering6: 15–20, 2017.
DOI:10.14326/abe.6.15

ebellum, and volitional control by the cortical areas. In emotional reactions, the limbic system is crucial for triggering locomotion. However, because of insuf�cient neuroscience studies on human locomotion, the neural substrates of these mechanisms are still unclear. As preliminary studies, we previously investigated the brain activity under the two controls [9, 10], but the relations be-tween neural activities and participants’ behavior remained un-clear.
This study therefore aimed to investigate the neural cor-relates of automatic and volitional controls of lower limb move-ment in human subjects. We instructed the participants to move their legs inside the MRI bore, and measured their brain activities and lower limb behaviors. One condition involved watching a video depicting walking straight along a corridor at a constant speed, and another walking while avoiding obstacles in the pas-sage. We assumed that the latter condition would induce relative-ly larger �uctuation of lower limb movement than the former con-dition, and investigated the correlation between brain activities in the locomotor-related cortical/subcortical regions and horizontal �uctuation of the feet. This design may be able to illustrate the two distinct lower limb control processes, and thus allows under-standing of the neural mechanism of locomotion in greater detail.
2. Methods
2.1 ParticipantsSixteen right-handed men (mean age 23.1 years, SD = 3.9) partic-ipated in this study. Data analysis described below revealed that the head movement of one participant during the experiment was large (> 3 mm). We excluded this participant from data analysis. All participants signed an informed consent form approved by the Committee of Medical Ethics, Graduate School of Medicine, Hokkaido University.
2.2 Experimental procedureThe participants were instructed to move their legs up and down at the knees inside the MRI bore. During the movement, a point-of-view video of 5 s (Fig. 1) depicting passage along a corridor at a constant speed was presented on a screen, and they were re-quired to look the video in a mirror mounted on the head coil while moving their legs. In addition, to further control the speed of their leg movements, a metronome sound (100 ms length with 10 ms rise and fall times, ISI of 500 ms) was presented through a headset. A 900-Hz pure tone was the signal to start and stop the movement, and a 450-Hz pure tone was the signal for the individ-ual steps. The walking speed of the point-of-view video was ap-
proximately the same as the speed depicted by the metronome sound. Prior to the experiment, the participants thoroughly prac-ticed their tasks.
A sparse image acquisition protocol [11] with a 7-s delay was used to minimize movement-related artifacts. The partici-pants were instructed to perform movement tasks during the non-scanning periods and to stop before the next scan. The video was presented 5 s during the 7-s non-scanning period. To mini-mize head movement of the participants induced by the tasks, their heads and bodies were �xed to the MRI bed using several belts. Cushions were placed in the space between the headset and the head coil. A triangular cushion was placed under the partici-pant’s knees, and soft blankets were put under the ankles. Two white markers were pasted on the soles of participants’ feet and images were captured by a video camera throughout the experi-ment to track their lower limb movement (Fig. 2).
There were two task conditions and one resting condition in this experiment. Under the non-obstacle condition, the presented video depicted walking straight along a corridor at a constant speed. Under the obstacle condition, the video depicted walking while avoiding obstacles in the passage. Six videos were prepared for each condition (total 12 videos), one of which was randomly presented during each task period. The supplementary �les show examples of videos for the obstacle and non-obstacle conditions. Regardless of the condition, we instructed the participants to syn-chronize their leg movements with the rhythm that was presented through the headset while they were watching the video. During the resting condition, they did not move their legs and merely looked at a �xed point on the screen.
Four experimental runs were performed for each participant, and each run had eight blocks: two were assigned to a task under obstacle condition, two for a task under non-obstacle condition, and the remaining four under resting condition. A single task
Fig. 1 Snapshot of the point-of-view video. (a) Non-obstacle condi-tion and (b) Obstacle condition.
Fig. 2 Experimental setup and the �ow of analysis. The white markers were pasted on the soles of the feet and were captured by a video camera (left). The horizontal movement was extracted of�ine (middle) and was used for correlation analysis with brain activity (right).
Advanced Biomedical Engineering. Vol. 6, 2017.(16)

block produced three MR image volumes under the same condi-tion. Therefore, 24 volumes (4 runs × 2 blocks × 3 volumes) were acquired from each participant for each of the obstacle and non-obstacle conditions.
A whole-body 1.5 T Signa EchoSpeed scanner (General Electric, Inc.) was used to acquire MR images with BOLD con-trast in this experiment. Twenty parallel axial slices (thickness, 5 mm; gap, 2 mm) were acquired using echo planar imaging with a 7-s delay (64 × 64 matrix; �eld-of-view, 24 × 24 cm; TR, 3 s; TE, 40 ms; �ip angle, 90°). T1-weighted images (corresponding to 20 axial slices; thickness, 5 mm; gap, 2 mm; 256 × 256 matrix; �eld-of-view, 24 × 24 cm; TR, 500 ms; TE, 14 ms) were acquired as an anatomical reference for the functional images. The experi-mental setup as well as the MRI acquisition/analysis methods described here were partially the same as those used in our previ-ous study [8].
2.3 AnalysisThe functional data from each participant were corrected for head movement, and a mean realigned image was created for each par-ticipant using the Statistical Parametric Mapping toolbox (SPM8; Wellcome Department of Cognitive Neurology, London, UK). The mean image for each participant was then coregistered to the high resolution T1-weighted image. The data sets were spatially normalized into the standard space de�ned by the Montreal Neu-rological Institute (MNI) template, and were spatially smoothed using an 8-mm isotropic Gaussian kernel to compensate for in-ter-individual anatomical variance. One participant was excluded from analysis due to a large amount of head motion (> 3 mm). All analyses described below were performed on the remaining par-ticipants.
For the general linear model approach, �rst-level statistical analysis on each participant’s time series data was used to model each condition independently by convolving with a �nite impulse response function. Motion parameters of the head estimated from the realignment process were added to the design matrix. A high-pass �lter was used to remove low-frequency noise. Group effects were then assessed by treating participants as random effects us-ing the individual contrast images. Statistical parametric maps were generated for each contrast of the t statistic on a vox-el-by-voxel basis.
To identify relations between the participants’ lower limb be-havior and brain activity, activation peaks in the locomotor-relat-ed areas were identi�ed in the following analyses. First, we ana-lyzed areas where the obstacle condition showed greater activation than the non-obstacle condition. Activation was found in the pre-motor cortex (Brodmann area 6/middle frontal gyrus: [26 −2 58] in MNI coordinates) in this contrast. Next, data of the two condi-tions were conjoined to localize the locomotor regions that showed activation in the primary motor cortex (Brodmann area 4: [−12 −34 74]), supplementary motor area (SMA: [8 −18 68]), and cerebellum ([−10 −44 −24]). The mesencephalon and dien-cephalon are known to be involved in locomotor control [1], but they did not show signi�cant activation in these analyses. We used the MNI coordinates [−5 −25 −13] for the mesencephalon based on Jahn et al (2008), who reported locomotor regions in the brain-stem [5], and with reference to the anatomical maps by Nieuwen-huys et al. [12]. For the diencephalon, we used the MNI coordi-nates [6 −1 −12] for the diencephalon based on Baroncini et
al [13]; this location is in the hypothalamus. We extracted the time course of the MR signal within a spherical volume of 2 mm in radius centered on these coordinates one by one from each par-ticipant’s data using the MarsBaR toolbox (http://marsbar.sourceforge.net/) in SPM8. Scanner drift was eliminated from the raw signal using �rst-order approximation by MATLAB. To cor-rect for the different baselines in each participant, the brain activ-ity was evaluated as signal change from the signal recorded under resting condition in each participant’s data. It was calculated as the percent change (%) from the level when the participant was at rest, for each region of interest (ROI) for each task period (the scan after the non-scanning period). If the correlation was evalu-ated based on averaged data of an individual participant, the �uc-tuation of lower limb movement was canceled out. We then ana-lyzed the correlation between the calculated percent signal change and the lower limb movement for each period (non-scanning pe-riod before the scan) extracted by the following step.
Video images captured during the experiment were dis-played full-screen on a 24.1-inch monitor, and the movement of the markers was extracted at pixel level. The movement at pixel level was then converted to the actual length based on a compari-son between the size of the video image and the actual size of the MRI system. The standard deviation of the horizontal movement of the markers was calculated for each task period (non-scanning period, 7s) and used as the degree of �uctuation. The data for the left and right feet were averaged. Through analyses of the imag-ing and behavioral data, 48 sets of data related to �uctuations of lower limbs and brain activities were acquired from each partici-pant. As the participants showed large variability in lower limb behavior, statistical evaluation based on regression coef�cients obtained from individual participants was dif�cult. Thus, we pooled the data acquired from all the participants and used it in the subsequent correlation analysis.
Correlation analysis was then performed in relation to lower limb �uctuation for the cortical motor areas of the primary motor cortex, premotor cortex, and SMA, as well as the cerebellum and subcortical areas of the mesencephalon and diencephalon. To ad-just multiple comparisons according to Bonferroni correction, each calculated p value was multiplied by six, and then the signif-icance was tested for each ROI.
3. Results
The non-obstacle condition caused 1.62 ± 1.22 (mean ± SD) cm of horizontal �uctuation in the lower limb movement, and the ob-stacle condition caused 2.91 ± 1.91 cm of �uctuation (Fig. 3). The �uctuation during obstacle condition was signi�cantly larger than that during non-obstacle condition (p < .001).
Comparison of brain activity between the two conditions showed that the obstacle condition elicited stronger activation in wide brain areas including the precuneus and middle occipital gy-rus in the parietal and occipital lobes compared to non-obstacle condition [Fig. 4 (a) and Table 1]. The cingulate gyrus also showed signi�cant activation. Among the cortical motor areas, the premotor cortex (middle frontal gyrus, BA6) was activated in this comparison. The non-obstacle condition induced greater acti-vation only in the right insula ([36 12 10]) compared to obstacle condition, except for peaks detected on the ventricle, callosum and white matter.
Figure 4 (b) shows the result of conjoined analysis to local-
Akira TOYOMURA, et al: Fluctuation of Lower Limb Movement in MRI (17)
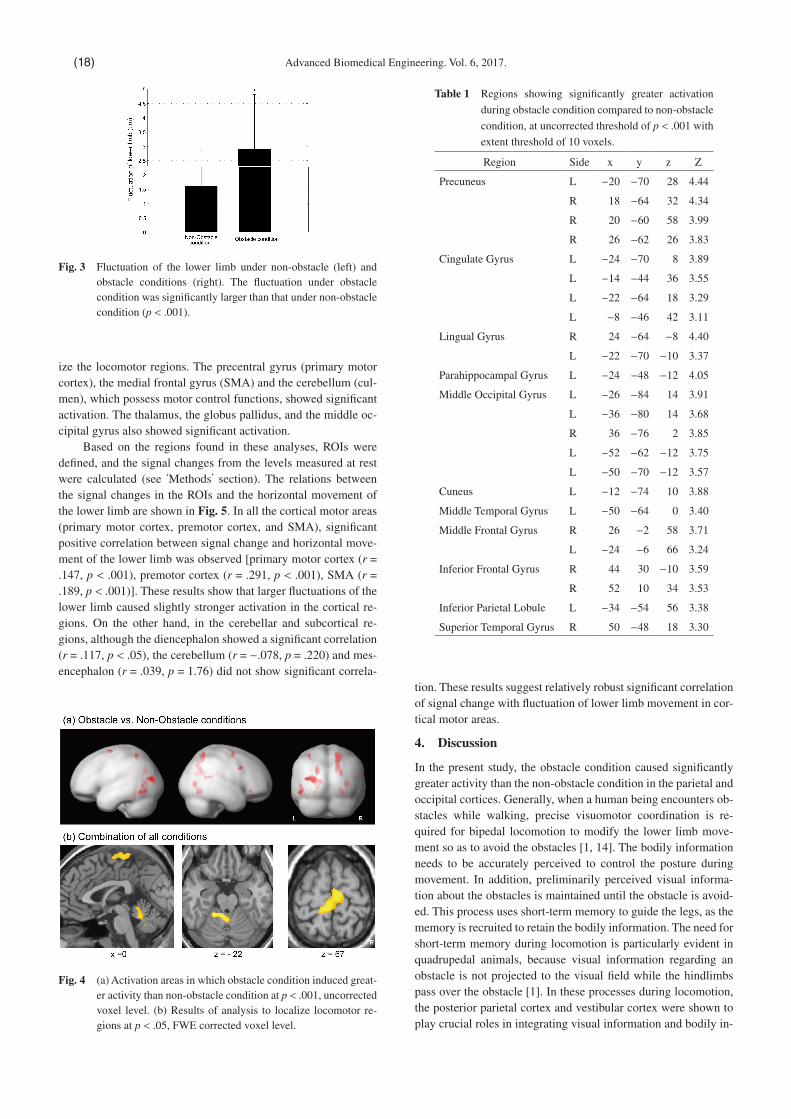
ize the locomotor regions. The precentral gyrus (primary motor cortex), the medial frontal gyrus (SMA) and the cerebellum (cul-men), which possess motor control functions, showed signi�cant activation. The thalamus, the globus pallidus, and the middle oc-cipital gyrus also showed signi�cant activation.
Based on the regions found in these analyses, ROIs were de�ned, and the signal changes from the levels measured at rest were calculated (see ‘Methods’ section). The relations between the signal changes in the ROIs and the horizontal movement of the lower limb are shown in Fig. 5. In all the cortical motor areas (primary motor cortex, premotor cortex, and SMA), signi�cant positive correlation between signal change and horizontal move-ment of the lower limb was observed [primary motor cortex (r = .147, p < .001), premotor cortex (r = .291, p < .001), SMA (r = .189, p < .001)]. These results show that larger �uctuations of the lower limb caused slightly stronger activation in the cortical re-gions. On the other hand, in the cerebellar and subcortical re-gions, although the diencephalon showed a signi�cant correlation (r = .117, p < .05), the cerebellum (r = −.078, p = .220) and mes-encephalon (r = .039, p = 1.76) did not show signi�cant correla-
tion. These results suggest relatively robust signi�cant correlation of signal change with �uctuation of lower limb movement in cor-tical motor areas.
4. Discussion
In the present study, the obstacle condition caused signi�cantly greater activity than the non-obstacle condition in the parietal and occipital cortices. Generally, when a human being encounters ob-stacles while walking, precise visuomotor coordination is re-quired for bipedal locomotion to modify the lower limb move-ment so as to avoid the obstacles [1, 14]. The bodily information needs to be accurately perceived to control the posture during movement. In addition, preliminarily perceived visual informa-tion about the obstacles is maintained until the obstacle is avoid-ed. This process uses short-term memory to guide the legs, as the memory is recruited to retain the bodily information. The need for short-term memory during locomotion is particularly evident in quadrupedal animals, because visual information regarding an obstacle is not projected to the visual �eld while the hindlimbs pass over the obstacle [1]. In these processes during locomotion, the posterior parietal cortex and vestibular cortex were shown to play crucial roles in integrating visual information and bodily in-
Table 1 Regions showing signi�cantly greater activation
during obstacle condition compared to non-obstacle
condition, at uncorrected threshold of p < .001 with
extent threshold of 10 voxels.
Region Side x y z Z
Precuneus L −20 −70 28 4.44
R 18 −64 32 4.34
R 20 −60 58 3.99
R 26 −62 26 3.83
Cingulate Gyrus L −24 −70 8 3.89
L −14 −44 36 3.55
L −22 −64 18 3.29
L −8 −46 42 3.11
Lingual Gyrus R 24 −64 −8 4.40
L −22 −70 −10 3.37
Parahippocampal Gyrus L −24 −48 −12 4.05
Middle Occipital Gyrus L −26 −84 14 3.91
L −36 −80 14 3.68
R 36 −76 2 3.85
L −52 −62 −12 3.75
L −50 −70 −12 3.57
Cuneus L −12 −74 10 3.88
Middle Temporal Gyrus L −50 −64 0 3.40
Middle Frontal Gyrus R 26 −2 58 3.71
L −24 −6 66 3.24
Inferior Frontal Gyrus R 44 30 −10 3.59
R 52 10 34 3.53
Inferior Parietal Lobule L −34 −54 56 3.38
Superior Temporal Gyrus R 50 −48 18 3.30
Fig. 3 Fluctuation of the lower limb under non-obstacle (left) and obstacle conditions (right). The �uctuation under obstacle condition was signi�cantly larger than that under non-obstacle condition (p < .001).
Fig. 4 (a) Activation areas in which obstacle condition induced great-er activity than non-obstacle condition at p < .001, uncorrected voxel level. (b) Results of analysis to localize locomotor re-gions at p < .05, FWE corrected voxel level.
Advanced Biomedical Engineering. Vol. 6, 2017.(18)

formation in animal studies [15, 16].The precuneus, which is located in the posterior parietal cor-
tex, is known to be involved in human visuospatial information processing [17]. This region was strongly and symmetrically acti-vated in the contrast between the obstacle and non-obstacle con-ditions in this study ([−20 −70 28] and [18 −64 32]). The middle occipital gyrus in the occipital lobe is known to be involved in visual processing, and it showed signi�cant activation in this study. Along with these areas related to visual and spatial process-ing, the posterior cingulate cortex was also seen to be signi�cant-ly activated. This area is known to play an important role in the regulation of the focus of attention [18], which is thought to be crucial to visuospatial processing as well. For these reasons, the areas that were signi�cantly activated in the obstacle condition were considered to be related to cognitive processes involved in volitional control.
This study observed whether a relation exists between the activities in the cortical/subcortical locomotor regions and the volitional lower limb behavior experimentally induced by moving images depicting the avoidance of obstacles. The signi�cant in-crease in horizontal �uctuations in the obstacle condition (Fig. 3) was speculated to be caused by an increase in volitional control to avoid the obstacles. Therefore, we assumed that the positive cor-relation of brain activity with the amount of �uctuation re�ected the effect of volitional control. The results indicated that the cor-tical and subcortical regions may have different contributions.
The activities in the cortical motor areas (primary motor cortex, premotor cortex, and SMA) showed weak but signi�cant correla-tion with horizontal �uctuation, suggesting that the cortical motor areas are involved in volitional control. On the other hand, the cerebellum and mesencephalon did not show signi�cant correla-tion but showed almost constant activation irrespective of �uctu-ation of lower limb movement. These suggest that the cerebellar and mesencephalic locomotor regions may not be involved in vo-litional control, but may be rather involved in the automatic pro-cess of locomotion. However, the whole-brain analysis did not show signi�cant activation in the mesencephalon, which might have affected the results of the correlation analysis in this area.
Finally, some possible limitations of this study should be ac-knowledged. First, owing to the large variability in lower limb behavior among the participants, we conducted the correlation analysis by pooling the �uctuation and brain activity data ac-quired from all the participants (2.3 Analysis). This variability in behavior may be caused by the method of instructing participants to perform the experimental task. More elaborate experimental procedures should be explored. Second, in this experiment, the participants lay face up and performed tasks involving lower limb movement. This movement was not actual gait but was a model of gait movement, because real gait movement requires postural control with antigravity functions and is more complex. Recently, MR-compatible devices have been proposed to investigate the neural mechanism of locomotion [19–22]. Among these devices, those proposed by Martinez et al. [19] and Toyomura et al. [22] are relatively close to simulating real locomotor movement. Fur-ther study using these systems is necessary to clarify the functions of the cortical and subcortical locomotor regions, as well as the differences between automatic process and volitional control of human locomotion.
5. Conclusion
Contributions of the cortical and subcortical locomotor regions to the automatic and volitional controls of locomotion were investi-gated using MRI. The results showed that the obstacle condition elicited larger �uctuation of lower limb movement compared to the non-obstacle condition, and elicited brain activation over wid-er areas including the parietal and occipital lobes compared to the non-obstacle condition. Correlation analysis showed signi�cant but weak correlation between activities in the cortical motor areas and �uctuation of lower limb movement. The activities in the cer-ebellar and mesencephalic locomotor regions did not show signif-icant correlation. These results suggest that the parietal, occipital cortices, and cortical motor areas are involved in volitional con-trol, while the cerebellar and mesencephalic locomotor regions may not be involved.
Acknowledgement
The authors would like to thank Prof. Kaoru Takakusaki for pro-viding helpful suggestions on this study. We are also grateful to Mr. Shuhei Bannae and Mr. Yojiro So for providing technical as-sistance. This study was supported by JSPS KAKENHI Grant Numbers 21700232, 24680025, 24650138 and 16K00366 to AT.
Disclosing Con�ict of Interest
We have no con�icts of interest relationship with any companies or commercial organizations.
Fig. 5 Correlation analyses between brain activity in each ROI and horizontal movement of the lower limb.
Akira TOYOMURA, et al: Fluctuation of Lower Limb Movement in MRI (19)

Supplemental Material
Supplementary material to this article can be found online at https://www.jstage.jst.go.jp/article/abe/6/0/6_6_15/_article/supplement
References
1 Takakusaki K: Neurophysiology of gait: from the spinal cord to the frontal lobe. Mov Disord. 28(11), pp. 1483–1491, 2013.
2 la Fougere C, Zwergal A, Rominger A, Forster S, Fesl G, Di-eterich M, Brandt T, Strupp M, Bartenstein P, Jahn K: Real versus imagined locomotion: a [18F]-FDG PET-fMRI comparison. Neuroimage. 50(4), pp. 1589–1598, 2010.
3 Hanakawa T, Katsumi Y, Fukuyama H, Honda M, Hayashi T, Kimura J, Shibasaki H: Mechanisms underlying gait disturbance in Parkinson’s disease: a single photon emission computed to-mography study. Brain. 122(Pt 7), pp. 1271–1282, 1999.
4 Iseki K, Fukuyama H, Oishi N, Tomimoto H, Otsuka Y, Nankaku M, Benninger D, Hallett M, Hanakawa T: Freezing of gait and white matter changes: a tract-based spatial statistics study. J Clin Mov Disord. 2, p. 1, 2015.
5 Jahn K, Deutschlander A, Stephan T, Kalla R, Wiesmann M, Strupp M, Brandt T: Imaging human supraspinal locomotor cen-ters in brainstem and cerebellum. Neuroimage. 39(2), pp. 786–792, 2008.
6 Iseki K, Hanakawa T, Shinozaki J, Nankaku M, Fukuyama H: Neural mechanisms involved in mental imagery and observation of gait. Neuroimage. 41(3), pp. 1021–1031, 2008.
7 Zwergal A, Linn J, Xiong G, Brandt T, Strupp M, Jahn K: Aging of human supraspinal locomotor and postural control in fMRI. Neurobiol Aging. 33(6), pp. 1073–1084, 2012.
8 Toyomura A, Shibata M, Kuriki S: Self-paced and externally trig-gered rhythmical lower limb movements: a functional MRI study. Neurosci Lett. 516(1), pp. 39–44, 2012.
9 Toyomura A, Kuriki S: Involvement of deep brain regions for locomotion control: an MRI study. IEICE Technical Report. MBE2012-21, pp. 43–46, 2012.
10 Toyomura A, Kuriki S: Study on neural substrates of bipedal lo-comotion control. Proceedings of The 27th Symposium on Bio-logical and Physiological Engineering. 2B3-05, 2012.
11 Hall DA, Haggard MP, Akeroyd MA, Palmer AR, Summer�eld AQ, Elliott MR, Gurney EM, Bowtell RW: “Sparse” temporal sampling in auditory fMRI, Hum Brain Mapp. 7(3), pp. 213–223, 1999.
12 Nieuwenhuys R, Voogd J, Huijzen CV: ‘The human central ner-vous system’ Springer, 4th edn. 2008.
13 Baroncini M, Jissendi P, Balland E, Besson P, Pruvo JP, Francke JP, Dewailly D, Blond S, Prevot V: MRI atlas of the human hypo-thalamus. Neuroimage. 59(1), pp. 168–180, 2012.
14 Georgopoulos AP, Grillner S: Visuomotor coordination in reach-ing and locomotion. Science. 245(4923), pp. 1209–1210, 1989.
15 Lajoie K, Andujar JE, Pearson K, Drew T: Neurons in area 5 of the posterior parietal cortex in the cat contribute to interlimb co-ordination during visually guided locomotion: a role in working memory. J Neurophysiol. 103(4), pp. 2234–2254, 2010.
16 Marigold DS, Drew T: Contribution of cells in the posterior pari-etal cortex to the planning of visually guided locomotion in the cat: effects of temporary visual interruption. J Neurophysiol. 105(5), pp. 2457–2470, 2011.
17 Cavanna AE, Trimble MR: The precuneus: a review of its func-tional anatomy and behavioural correlates. Brain. 129(Pt 3), pp. 564–583, 2006.
18 Leech R, Sharp DJ: The role of the posterior cingulate cortex in cognition and disease. Brain. 137(Pt 1), pp. 12–32, 2014.
19 Martinez M, Villagra F, Loayza F, Vidorreta M, Arrondo G, Luis E, Diaz J, Echeverria M, Fernandez-Seara MA, Pastor MA: MRI-compatible device for examining brain activation related to stepping. IEEE Trans Med Imaging. 33(5), pp. 1044–1053, 2014.
20 Mehta JP, Verber MD, Wieser JA, Schmit BD, Schindler-Ivens SM: A novel technique for examining human brain activity asso-ciated with pedaling using fMRI. J Neurosci Methods. 179 (2), pp. 230–239, 2009.
21 Newton JM, Dong Y, Hidler J, Plummer-D’Amato P, Marehbian J, Albistegui-Dubois RM, Woods RP, Dobkin BH: Reliable as-sessment of lower limb motor representations with fMRI: use of a novel MR compatible device for real-time monitoring of ankle, knee and hip torques. Neuroimage. 43(1), pp. 136–146, 2008.
22 Toyomura A, Yokosawa K, Shimojo A, Tetsunoshin F, Kuriki S: Evaluation of brain activity of lower limb movement using MRI-compatible equipment. Transactions of the Japanese Soci-ety for Medical and Biogical Engineering. 54(Suppl. 1), p. 174, 2016.
Akira TOYOMURA
Akira TOYOMURA is an Associate Professor at the
Graduate School of Health Sciences, Gunma Uni-
versity, Japan. He received the Ph.D. degree from
the Graduate School of Engineering, Hokkaido
University in 2005. Before joining Gunma Univer-
sity, he has been a research fellow at Hokkaido
University and an assistant professor at Tokyo Denki University. His
research interests are in human brain functions of motor control, in-
cluding lower limb movement, speech production, and stuttering.
Koichi YOKOSAWA
Koichi YOKOSAWA was born in Sendai, Japan, in
1962. He received B.S. and M.S. degrees in phys-
ics from the Faculty of Science, Hokkaido Univer-
sity in 1984 and 1986 respectively, and a Ph.D.
degree in systems and information engineering
from the Faculty of Engineering, Hokkaido Uni-
versity in 1998. He joined the Central Research Laboratory, Hitachi,
Ltd. in 1986, and was engaged in medical instrument research and de-
velopment. He transferred to Hokkaido University in 2007 where he
holds a professorship and is now engaged mainly in studying brain
function using magnetoencephalography.
Shinya KURIKI
Shinya KURIKI is a Professor Emeritus of Research
Institute for Electronic Science, Hokkaido Univer-
sity, Japan. He received the Ph.D. (Dr. Eng.) de-
gree in electrical engineering from Hokkaido Uni-
versity in 1976. He was a Professor at Hokkaido
University from 1991 and Tokyo Denki University
from 2009. He also currently belongs to the Faculty of Health Sciences,
Hokkaido University. His research interests are in biomagnetism, mea-
surement and instrumentation of biological magnetic �eld, and higher
brain functions.
Advanced Biomedical Engineering. Vol. 6, 2017.(20)


















