
Enzimas
31
ENZIMAS Prof.: Angel Roco Videla Introducción Las enzimas son catalizadores biológicos, la cuales pueden no sólo aumentar la velocidad en factores que van desde 10 6 a 10 12 , sino que son altamente especifica. Actúan sólo en determinadas moléculas, llamadas sustrato (esto es, reactivos). Se ha calculado que una célula promedio puede contener unas 3000 enzimas diferentes, cada una de las cuales cataliza una reacción específica. Prof.: Angel Roco Videla
-
Upload
enfermeriauv07 -
Category
Business
-
view
48.123 -
download
0
Transcript of Enzimas

1
ENZIMAS
Prof.: Angel Roco Videla
Introducción
Las enzimas son catalizadores biológicos, la cuales pueden no sólo aumentar la velocidad en factores que van desde 106 a 1012 , sino que son altamente especifica. Actúan sólo en determinadas moléculas, llamadas sustrato (esto es, reactivos). Se ha calculado que una célula promedio puede contener unas 3000 enzimas diferentes, cada una de las cuales cataliza una reacción específica.
Prof.: Angel Roco Videla

2
Prof.: Angel Roco Videla
Las enzimas no operan independientemente, existe un control interno que activa ciertas enzimas cuando se requieren, y las desactiva cuando se ha formado suficiente producto.
Una teoría sobre la especificidad de las enzimas es la llamada teoría de la “llave cerradura”. Una enzima es generalmente una molécula grande de proteína que contiene uno o más sitios activos donde se lleva a cabo las reacciones con los sustratos, el resto de la molécula mantiene la integridad tridimensional de la red.
Prof.: Angel Roco Videla
ENZIMA
PRODUCTOS
MODELO LLAVE CERRADURA

3
Prof.: Angel Roco Videla
Sustrato
Enzima Complejo enzima-sustrato
Productos
Enzima
Prof.: Angel Roco Videla
De acuerdo con la Hipótesis el sitio activo tiene una estructura rígida , similar a una cerradura. Una molécula de sustrato tiene la estructura complementaria que causa que embonen, y funciona como una llave.
Aunque parece perfecta la hipótesis existen varias discrepancias, por ejemplo se sabe que sustratos de diferentes tamaños embonan con el mismo tipo de enzima. Hoy se sabe que el sitio activo de una enzima tiene gran flexibilidad , durante una reacción cambia para acomodarse a la molécula de sustrato.

4
Prof.: Angel Roco Videla
Propiedades de las enzimas
Las enzimas son muy específicas para los reactantes osustratos, sobre los cuales actúan. Una enzima puede reaccionar con un grupo de sustancias de estructuras estrechamente relacionadas, mientras que otra sólo puede actuar sobre un compuesto molecular único. Muchas enzimas presentan estereoespecificidad, lo cual significa, que actúan sobre un solo estereoisómero del sutrato.
Prof.: Angel Roco Videla
Tal vez el aspecto general más importante de la especificidad de las enzimas es la especificidad de reacción, esto es, la falta de formación de subproductos de desecho. La especificidad de reacción se refleja en los rendimientos excepcionales de producto, los cuales son prácticamente del 100%. La eficiencia de las enzimas no solo ahorra energía para la célula viva, sino que también evita la formación de subproductos del metabolismo potencialmente tóxicos.

5
Prof.: Angel Roco Videla
Algunas enzimas funcionan como puntos de control en el metabolismo el metabolismo puede ser controlado a través de:
* alteraciones en las concentraciones de enzimas
* Alteraciones en las concentraciones de sustratos y de inhibidores de enzimas
* Con la modulación de los niveles de actividad de algunas enzimas.
Prof.: Angel Roco Videla
Las enzimas más comunes son proteínas globulares y poseen la capacidad para unirse específicamente a una o varías moléculas.
Las moléculas de sustrato se unen, en una hendidura un poco hidrofóbica que se conoce como el sitio activo, el cual con frecuencia está situado entre dos dominios o subunidades de una proteína o en una indentación, algo parecido una bolsa de un extremo de un barril.

6
Prof.: Angel Roco Videla
Las enzimas que intervienen en la regulación del metabolismo son por lo general de mayor complejidad que las enzimas que no son reguladoras.
Las enzimas son moléculas oligoméricas que tienen sitios de unión diferentes para sustratos y para moduladores, (compuestos que actúan como señales reguladoras).
Clasificación de las enzimas
La mayor parte de las enzimas tienen un nombre que se forma adicionando el fijo "-asa" al nombre del sustrato sobre el cual actúan o a un término que describe las reacciones que catalizan.
Prof.: Angel Roco Videla
Por ejemplo, la ureasa tiene la urea como sutrato. La alcoholdeshidrogenasa cataliza la remoción de hidrógenos de los alcoholes.
A pesar de ello, algunas enzimas, como tripsina o la quimotripsina, aún se reconocen por sus nombres históricos.
La Unión Internacional de Bioquímica asigna un número a cada enzima y clasifica a las enzimas en seis grupos importantes de acuerdo a la clase general de reacciones químicas que catalizan:

7
Prof.: Angel Roco Videla
1.- Las óxidorreductasas catalizan reacciones de oxidación-reducción. La mayor parte de estas enzimas se conocen como deshidrogenasas, pero algunas de ellas reciben el nombre de oxidasas, peroxidasa, oxigenasas, o reductasas.
Ejemplo
Enzima : Lactato deshidrogenasa
Descripción de la reacción :Oxidación de alcohol secundario. Lactato a piruvato ( una cetona)
Coenzima que participa: NAD+ ( Nicotínamida adenina dinucleótido)
Prof.: Angel Roco Videla
C
C
CH3
HOH
O
O-
NAD++
C
C
CH3
O
O
O-
+ +NADH H+
L-lactato
piruvato

8
Prof.: Angel Roco Videla
Ejemplo
Enzima :Alanina transaminasa ( Alanina aminotransferasa)
Descripción de la reacción :Transferencia de un grupo amino
Coenzima que participa: Fosfato de piridoxal
2.- Las transferasas catalizan reacciones de transferencia de grupos. Muchas de ellas requieren de la existencia de coenzimas. Estas enzimas o sus coenzimas son sustituidas en forma covalente por una porción de la molécula del sustrato.
Prof.: Angel Roco Videla
C
C
CH3
NH3+
O
O-
H +C
C
CH2
O
O-
O
CH2
C
O
O-
L-alaninaα-cetoglutarato
C
C
CH3
O
O
O-
piruvato
CH
C
CH2
O
O-
CH2
C
O
O-
NH3+
+
L-glutamato

9
Prof.: Angel Roco Videla
Ejemplo
Enzima :tripsina
Descripción de la reacción :Hidrólisis del enlaces peptídicos Lis-Y o de Arg-Y donde Y es igual o diferente a Pro,
Coenzima que participa: Ninguna
3.- Las hidrolasas catalizan la hidrólisis. Son una clase especial de transferasas, el agua les sirve como un receptor del grupo transferido.
Prof.: Angel Roco Videla
NH
CH
C
CH2
CH2
CH2
CH2
NH3+
NH
CH
CO
-
RO
-+ OH2
Residuo de lisina dentro de una cadena peptídica
NH
CH
C
CH2
CH2
CH2
CH2
NH3+
O
-
O-
NH3+
CH
C
O
-
R
Fragmento de polipéptido con
lisina C-terminal.
+Fragmento de
polipéptido nuevo N-terminal

10
Prof.: Angel Roco Videla
Ejemplo
Enzima :piruvato descarboxilasa
Descripción de la reacción :Descarboxilación de piruvato
Coenzima que participa: Tiamina pirofosfato
4.- Las liasas catalizan las reacciones de eliminación no hidrolítica, no oxidante, o la lisis de un sustrato en reacciones que generan un enlace doble. En la dirección inversa, una liasa cataliza la adición de un sutrato a un doble enlace de un segundo sustrato. Una liasa que cataliza una reacción de adición en las células se denomina con frecuencia sintasa
Prof.: Angel Roco Videla
C
C
CH3
O
O
O-
piruvato
H++
CH
CH3
OCO2
Acetaldehido
+

11
Prof.: Angel Roco Videla
Ejemplo
Enzima :Alanina racemasa
Descripción de la reacción :Interconversión de isómeros D y L de alanina
Coenzima que participa: Fosfato de piridoxal
5.- Las isomerasas catalizan reacciones de isomerización. Debido a que estas reacciones tienen un solo sustrato y un producto, son por lo regular reacciones enzimáticas más sencillas
Prof.: Angel Roco Videla
C
C
CH3
HNH3+
O
O-
C
C
CH3
H NH3+
O
O-
L-Alanina D-Alanina

12
Prof.: Angel Roco Videla
Ejemplo
Enzima :Glutamina sintetasa
Descripción de la reacción :Síntesis de L-glutamina, dependiente de ATP
Coenzima que participa: ATP
6.- Las ligasas catalizan la ligadura o unión de dos sustratos en reacciones sintéticas que requieren el ingreso de la energía química potencial de un nucleósidotrifosfato, como el ATP. A las ligasas se les da el nombre de sintetasas
Prof.: Angel Roco Videla
C
O
O-
C
CH2
CH2
CO O-
NH3+
HATP+ + NH4
+
C
O
O-
C
CH2
CH2
CO NH2
NH3+
H ADP+ + Pi

13
Prof.: Angel Roco Videla
La ecuación de velocidad enzimática
Los datos cinéticos reflejan las relaciones entre la cantidad de producto P de la reacción formada en la unidad de tiempo, ∆[P]/∆t, y las condiciones experimentales bajo las cuales la reacción tiene lugar.
La base de la mayor parte de las mediciones cinéticas es la observación de que la rapidez o velocidad (v) de una reacción varía directamente con la concentración de cada sustrato o de cada catalizador. Esta afirmación se escribe en una ecuación que se llama ecuación de la velocidad.
Prof.: Angel Roco Videla
Por ejemplo, la velocidad de ecuación para la conversión no enzimática de S→P una reacción de isomerización se escribe como:
∆[P]/∆t = v = k [S]
El símbolo k es la constante de velocidad. Las constantes de velocidad indican la rapidez o la eficiencia de una reacción. Una gráfica de velocidad en comparación a [S]para esta reacción podría ser una línea recta que aumenta linealmente con la concentración del sustrato.

14
Prof.: Angel Roco Videla
El orden cinético general de una reacción es la suma de los exponentes en la ecuación de velocidad y nos dice cuantas moléculas están reaccionando en la etapa más lenta de la reacción.
Un reacción de primer orden es aquella en la cual un solo componente está reaccionando. La constante de velocidad para una reacción de primer orden se expresa en unidades de la recíproca del tiempo ( s-1)
Prof.: Angel Roco Videla
[E]
v
Reacción de primer orden

15
Prof.: Angel Roco Videla
Para una reacción complicada como la reacción
de S1 + S2 → P1 + P2
La velocidad se determina por las concentraciones de ambos sustratos.-
Si ambos sustratos están a concentraciones similares la ecuación de velocidad es :
v = k [S1]1 [S2]1
Esta reacción es de primer orden con respecto a cada reactante y de segundo orden general.-
Prof.: Angel Roco Videla
Las reacciones de segundo orden también se denominabimolecular porque dos moléculas reaccionan formando los productos. Las constantes de velocidad para las reacciones de orden tienen las unidades M-1 s-1. Con frecuencia se estudian reacciones con más de dos reactantes, pero debido a que los cálculos se pueden volver muy complejos, se escogen condiciones de modo que las reacciones se efectúen en varias etapas, y que cada etapa sea de primer o de segundo orden.-

16
Prof.: Angel Roco Videla
Si la concentración de uno de los reactantes es tan alta que permanece prácticamente constante durante el curso de la reacción se dice que la reacción es de pseudo primer orden.
v= k [S1]1 [S2]o = k’ [S1]1
Las condiciones de psuedo primer-orden se utilizan en análisis que se llaman ensayos enzimáticos que determinan las concentraciones de las enzimas.
Prof.: Angel Roco Videla
En los ensayos enzimáticos, todos los reactantes, con excepción de la enzima, existen en exceso para asegurar que las condiciones de la reacción sean de pseudo - primer orden . El ensayo se lleva a cabo a una temperatura constante lo suficiente elevada para producir una actividad alta, pero no para desnaturalizar a la enzima.
Baja estas condiciones, hay suficientes moléculas de sustrato para convertir todas las moléculas de E en ES ( saturación).

17
Prof.: Angel Roco Videla
En los ensayos, por lo regular las enzimas están en concentraciones muy bajas en comparación con los sustratos. Sin embargo, en el interior de las células, no están necesariamente saturadas. La concentración de una enzima en una muestra de prueba se puede determinar con facilidad por comparación de su actividad con una curva de referencia
Prof.: Angel Roco Videla
Los momentos iniciales de una reacción bimolecular se puede escribir como:
E + S ES E + Pk1
k-1
kcat
La velocidad medida durante este corto periodo se denomina velocidad inicial vo

18
Prof.: Angel Roco Videla
Las velocidades iniciales se obtienen a partir de curvas de progreso, gráficas de la concentración del producto contra el tiempo. La velocidad es la tangente o pendiente ∆[P]/ ∆t en el origen de una curva de progreso.
[P]
Tiempo
2E
E
Prof.: Angel Roco Videla
La ecuación de Michaelis-Menten
Cuando se comenzaron a analizar las variaciones en las concentraciones del sustrato con el fin de derivar las ecuaciones de velocidad de los procesos enzimáticos, se hicieron dos observaciones importantes.
1º.- a concentraciones elevadas de sustrato, E esta saturada con S, y la velocidad de reacción es independiente de la concentración de sustrato. El valor de vo para una solución de E que está saturada con S se llama velocidad máxima vmáx.

19
Prof.: Angel Roco Videla
2.- A concentraciones bajas de sustrato , la reacción es de primer orden con respecto al sustrato (segundo orden general : de primer orden con respecto a S y de primer orden con respecto a E). La forma de la curva de vo en comparación a [S] es una hipérbola rectangular.
vmáx
vo
0
Primer orden con respecto a S
Orden cero con respecto a S
[S]
Prof.: Angel Roco Videla
La ecuación de velocidad para una hipérbola de este tipo es
y = ax
b + x
Donde a es la asíntota de la curva ( el valor de y a un valor infinito de x) y b es el punto sobre el eje de la x que corresponde a un valor de y al infinito /2. Podemos obtener la ecuación de velocidad para la reacción bimolecular:
E + S ES E + P

20
Prof.: Angel Roco Videla
Por sustitución de cuatro términos de la cinética enzimática en la ecuación general para una hipérbola rectangular tenemos y = vo , x = [S], a = vmáx. El cuarto termino b en la ecuación general es la constante de Michaelis Km definida como la concentración de sustrato cuando vo es igual a la mitad vmáx.
vmáx
vo =
0[S]
vmáx2
Prof.: Angel Roco Videla
La ecuación completa de velocidad:
y = vmáx[S]
km + [S]
Esta se denomina ecuación de Michaelis-Menten.

21
Prof.: Angel Roco Videla
Constante de velocidad y la eficiencia catalítica
Las constantes cinéticas de las reacciones enzimáticas Kmy kcat se pueden utilizar para medir la eficiencia catalítica de las enzimas.-
Km es una medida de la estabilidad del complejo ES
kcat es la constante de velocidad de primer orden para la conversión de ES a E + P. Es una medida de la actividad catalítica de una enzima y nos dice cuantas reacciones por segundo puede catalizar una molécula de enzima.
Prof.: Angel Roco Videla
La relación kcat / Km es una constante de velocidad de segundo orden aparente para la formación de E + P a partir de E + S cuando la reacción general está limitada por el encuentro de S con E. Esta constante se aproxima a 108 a 109 M-1s-1. , que es la mayor velocidad a la cual dos solutos sin carga se pueden aproximar entre si por difusión.
Los valores absolutos de estas constantes indican la potencia de cada enzima como catalizador.
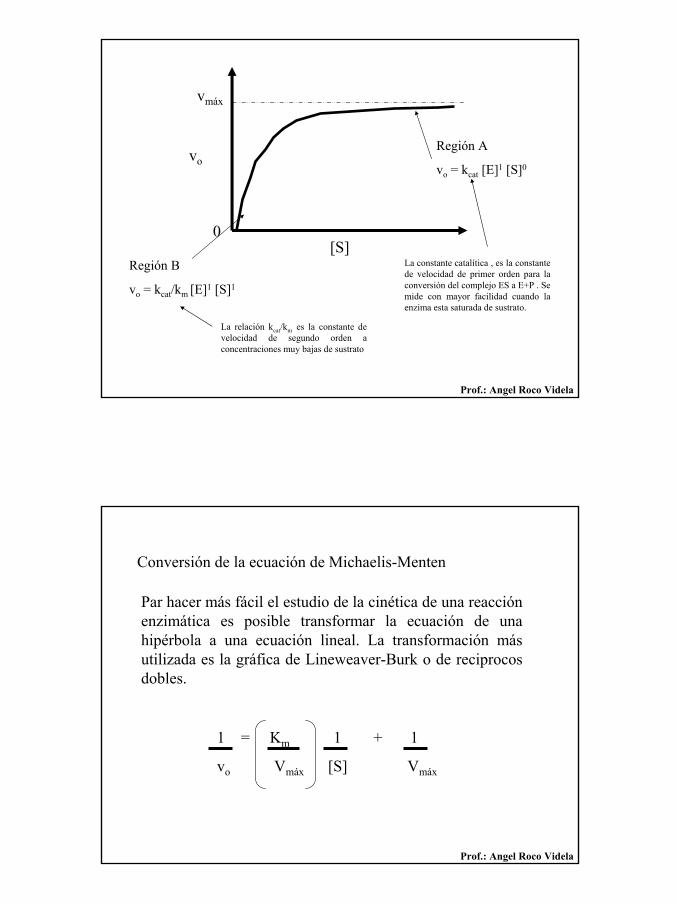
22
Prof.: Angel Roco Videla
vmáx
vo
0[S]
Región A
vo = kcat [E]1 [S]0
Región B
vo = kcat/km [E]1 [S]1
La constante catalítica , es la constante de velocidad de primer orden para la conversión del complejo ES a E+P . Se mide con mayor facilidad cuando la enzima esta saturada de sustrato.
La relación kcat/km es la constante de velocidad de segundo orden a concentraciones muy bajas de sustrato
Prof.: Angel Roco Videla
Conversión de la ecuación de Michaelis-Menten
Par hacer más fácil el estudio de la cinética de una reacción enzimática es posible transformar la ecuación de una hipérbola a una ecuación lineal. La transformación más utilizada es la gráfica de Lineweaver-Burk o de reciprocosdobles.
1 = Km 1 + 1
vo Vmáx [S] Vmáx

23
Prof.: Angel Roco Videla
Se usan los valores de 1/vo contra 1/[S] para la gráfica. El valor absoluto de 1/km se obtiene de la intercepción de la línea en el eje de la x y el valor de 1/Vmáx se obtiene del intercepto en el eje de las y.
Los valores de kcat se pueden obtener a partir de mediciones de Vmáx sólo cuando se conoce la concentración absoluta de la enzima. Los valores de km se pueden determinar aún cuando las enzimas no se hayan purificado, mientras una enzima en una preparación impura pueda catalizar la reacción que se observa.
Prof.: Angel Roco Videla
1/vo
1/[S]
1/vmáx
-1/km
ECUACIÓN DE
LINEWEAVER-BURK

24
FACTORES QUE AFECTAN LA ACTIVIDAD ENZIMÁTICA
Y EL EFECTO DE LOS INHIBIDORES ENZIMÁTICOS
Prof: Angel Roco Videla
Prof: Angel Roco Videla
Efecto del pH sobre la actividad enzimática
Aunque los perfiles de las curvas de actividad en función del pH de muchas enzimas son acampanadas, pueden variar considerablemente de forma.

25
Prof: Angel Roco Videla
La relación entre pH y la actividad de cualquier enzima depende del comportamiento ácido-base de la enzima y del sustrato.
La forma de la curva de actividad-pH varía con la concentración del sustrato, ya que el valor de KM de muchas enzimas varía con el pH.
La relación pH actividad de una enzima puede constituir un factor en el control intracelular de su actividad, esto debido a que el pH optimo de una enzima no es necesariamente idéntico al pH de su entorno intracelular normal.
Prof: Angel Roco Videla
Efecto de la temperatura sobre la actividad enzimática
Aunque las reacciones catalizadas por las enzimas parecen, con frecuencia tener una temperatura optima, el pick que se observa al representar la actividad versus temperatura se produce porque las enzimas se desnaturalizan por la acción del calor y se inactivan cuando la temperatura sobrepasa cierto punto.
La aparente temperatura optima es el resultado de dos procesos
1.- el incremento habitual de la velocidad de reacción con la temperatura.
2.- el incremento en la velocidad de desnaturalización térmica de la enzima al sobrepasar una temperatura critica.

26
Prof: Angel Roco Videla
La mayoría de las enzimas se inactivan a temperaturas entre los 55 y 60ºC. Aunque existen enzimas capaces de seguir activas hasta los 85ºC como las enzimas de las bacterias termofílicas.
Prof: Angel Roco Videla
Algunas enzimas, como la ribonucleasa, pierden actividad por calefacción, pero la recuperan por enfriamiento, lo cual indica que su cadena polipeptídica desplegada vuelve rápidamente a adoptar su conformación nativa.

27
Prof: Angel Roco Videla
Inhibición de las enzimas
Inhibición competitiva
La característica de la inhibición competitiva es que el inhibidor puede combinarse con la enzima libre de tal modo que compite con el sustrato normal para unirse al centro activo. Un inhibidor competitivo reacciona reversiblemente con la enzima para formar un complejo enzima-inhibidor (EI) análogo al complejo enzima-sustrato:
E + I EI
Prof: Angel Roco Videla
Kj[E][I]
[EI]=
La molecula del inhibidor no resulta químicamente alterada por la enzima. Es posible definir entonces la constante de inhibición como Kj. Y esta sería comparable con Ks que es la constante de disociación del complejo enzimas sustrato.
La inhibición competitiva se reconoce experimentalmente, debido a que el porcentaje de inhibición para una concentración de inhibidor constante disminuye al incrementar la concentración del sustrato.

28
Gráficamente este tipo de inhibición se caracteriza por proporcionar una familia de líneas rectas cuyo punto de intersección común se haya sobre el eje de 1/vº.
La presencia de un inhibidor competitivo incrementa KM aparente de la enzima por el sustrato, provocando que se necesite concentraciones mayores de sustrato para que se alcance la velocidad máxima.
Prof: Angel Roco Videla
Prof: Angel Roco Videla
Inhibición acompetitiva
El inhibidor no se combina con la enzima liebre ni afecta a su reacción con el sustrato normal, sin embargo se combina con el complejo ES para formar un complejo inactivo ESI, el cual no experimenta posteriores transformaciones.
ES + I ESI Kj[ES][I]
[ESI]=

29
El grado de inhibición puede aumentar cuando la concentración de sustrato se ve aumentada. Esta inhibición se puede reconocer gráficamente al representar 1/vº frente a 1/[S] para concentraciones constantes de inhibidor. Lo característico de este tipo de inhibición es que pendiente permanece constante al aumentar a concentración del inhibidor pero la Vmax decrece.
Prof: Angel Roco Videla
Inhibición no competitiva
Un inhibidor no competitivo puede combinarse con la enzima libre o bien con el complejo ES, interfiriendo con la acción de ambos. Los inhibidores no competitivos se unen a un centro de la enzima distinto del centro activo, a menudo para deformar a la enzima, de modo que no pueda formarse en complejo ES a su velocidad normal y que una vez formado no se descomponga a su velocidad habitual para liberarlos productos. Sus efectos no se anulan al aumentar la concentración de sustrato.
Prof: Angel Roco Videla

30
Prof: Angel Roco Videla
En la inhibición no competitiva la reacción con en inhibidor produce dos formas inactivas, EI y ESI:
ES + I ESI
E + I EI Kj[E][I]
[EI]=
Kj[ES][I]
[ESI]=
Prof: Angel Roco Videla
Esta inhibición se reconoce gráficamente en el hecho que las líneas no comparten un punto de intersección común. El valor de la intersección es mayor para la enzima inhibida que para las no inhibidas, lo que indica que la Vmax decrece en presencia del inhibidor y no puede restablecerse su valor aunque se aumente la concentración de sustrato.

31
Prof: Angel Roco Videla
Inhibición irreversible: modificación de la enzima
Este tipo de moléculas son capaces de unirse covalentemente con la enzima y modifican de modo permanentemente el grupo funcional, necesario para la catálisis. Este tipo de inhibición no puede ser tratado por los principios de Michaelis-Menten, que suponía la formación de complejos reversibles.
Este tipo de inhibición es muy útil para identificarlos grupos funcionales esenciales para el proceso de catálisis
TAREA
¿QUE SON LAS ENZIMAS REGULADAS ALOSTERICAMENTE (ENZIMAS ALOSTERICAS) Y LAS ENZIMAS MODULADAS COVALENTEMENTE?


















