
Differential Gene Expression - University of Minnesota...
46
Differential Gene Expression IBS 8102 – Cell, Molecular, and Developmental Biology January 22, 2008
Transcript of Differential Gene Expression - University of Minnesota...

Differential Gene Expression
IBS 8102 – Cell, Molecular, and Developmental Biology
January 22, 2008

Differential Gene Expression
Chromatin structure
Gene anatomy
Gene sequences
Control of gene transcription
enhancers
transcription factors
RNA processing
methylation
imprinting
Epigenetic processes (modification of structure of DNA rather than sequence)
epigenetics

Chromatin Structure
~140 bp
~60 bp
chromatin
Histone lysines are protonated = net positive charge
DNA PO 4 deprotonated = net negative charge
electrostatic attraction ensures tight binding DNA inaccessible to polymerases, etc

Nucleosome
DNA bound to an octameric complex composed of two each of histones H2A, H2B, H3, and H4. From Luger et al, 1997. Nature 389:251260.
Tails
Core
Histone H1 – associates nucleosomes into higherorder folded complex

Chromatin Acetylation
3. Histone methylation (CH 3 ) – transcriptional repression
Transcriptional Regulation:
1. “Packing” prevents access
acetylated histone (histone acetyltransferase; HAT) destabilizes = transcription allowed
deacetylated histones (histone deacetylases; HDAC) stabilizes = transcription repressed
2. Acetylation ( ) state of histones controls DNA binding CH 3 C O
e.g. acetylation of K14 and K9 lysines on tail of histone H3 correlated with transcriptional competence
methylation of K9 lysine on H3 associated with heterochromatin
Multiple histone regulatory modifications may occur at once; work together to change the behavior of the chromatin dynamic systematic reproducible
= Histone Code

remains tightly condensed throughout most of the cell cycle
replicates later
John H. Frenster
Chromatin Configuration
Heterochromatin Euchromatin “active” chromatin

Anatomy of the Gene
5’ 3’
core promoter region
transcription initiation site
+1 TATAT
upstream downstream
transcribed region
Transcription produces heterogenous nuclear RNA (hnRNA)
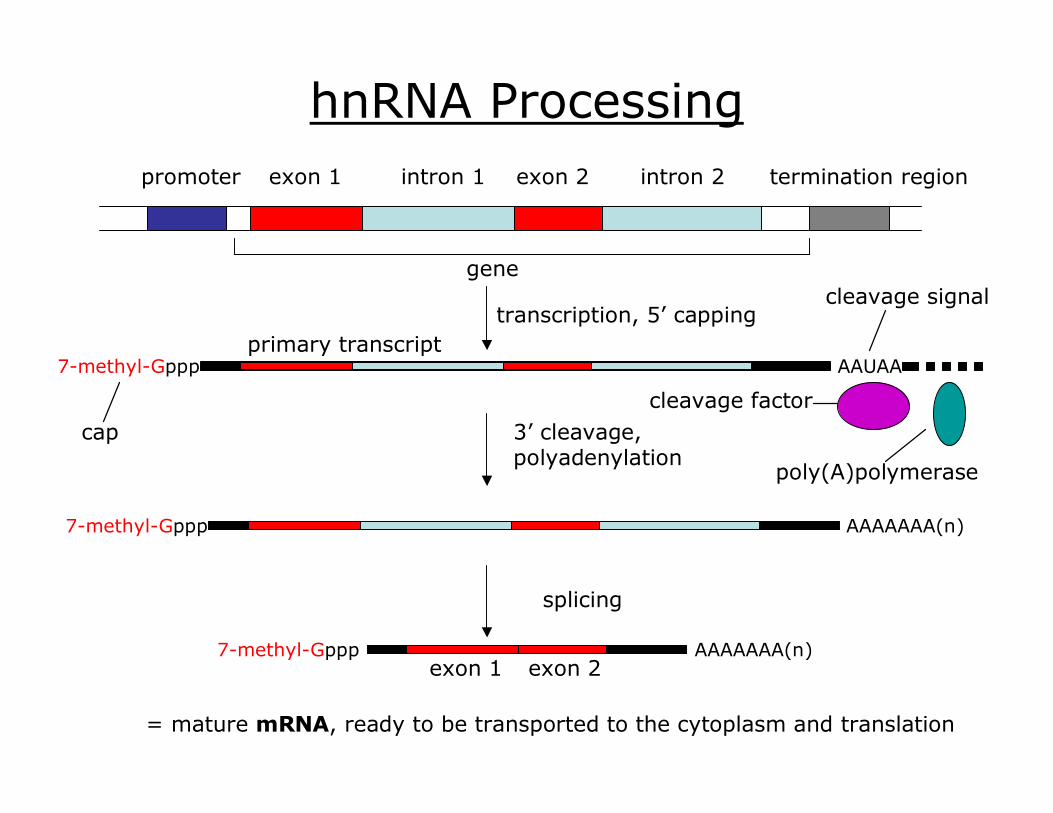
promoter exon 1 intron 1 exon 2 intron 2 termination region
gene
hnRNA Processing
splicing
exon 1 exon 2 7methylGppp AAAAAAA(n)
= mature mRNA, ready to be transported to the cytoplasm and translation
7methylGppp
cap 3’ cleavage, polyadenylation
AAAAAAA(n) 7methylGppp
cleavage signal
poly(A)polymerase
cleavage factor
AAUAA primary transcript
transcription, 5’ capping

Human βglobin Sequence
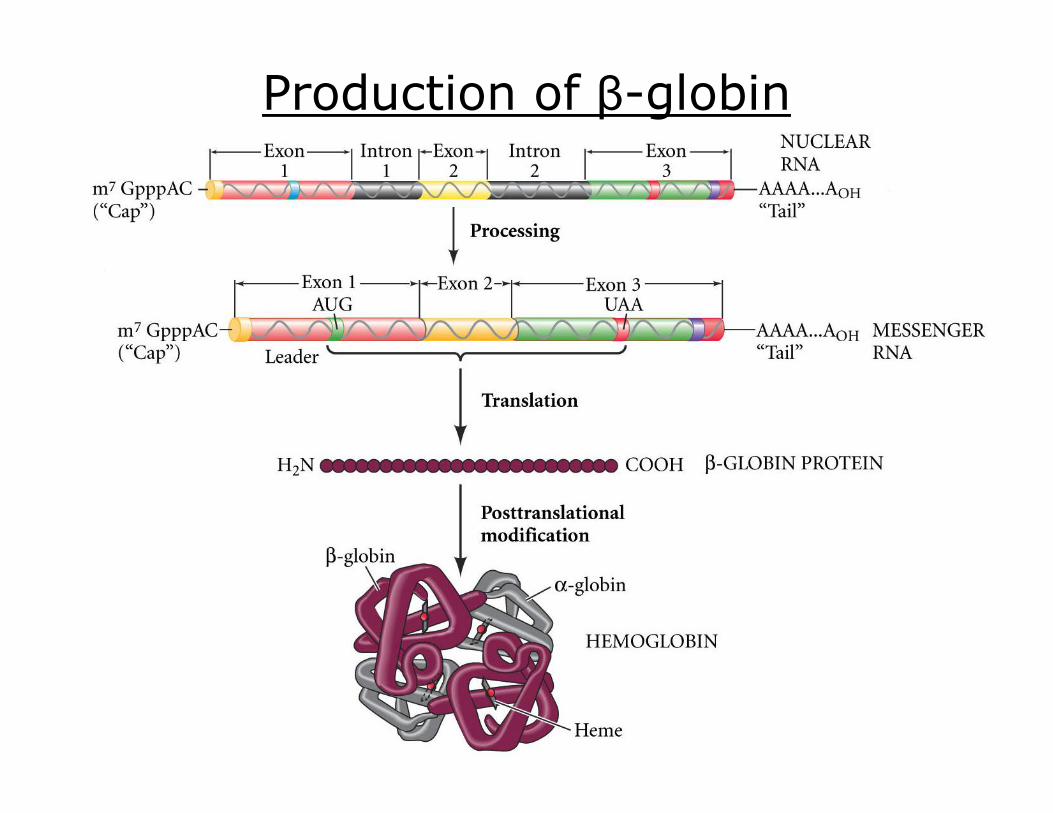
Production of βglobin
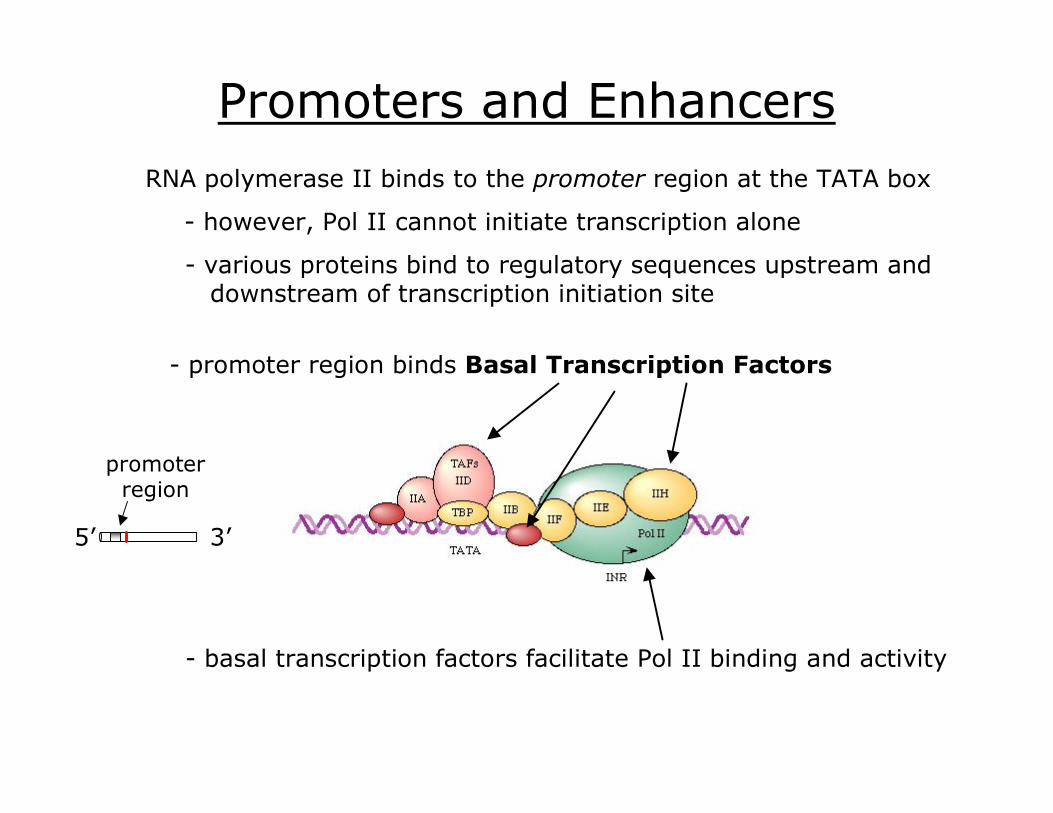
Promoters and Enhancers RNA polymerase II binds to the promoter region at the TATA box
various proteins bind to regulatory sequences upstream and downstream of transcription initiation site
promoter region binds Basal Transcription Factors
basal transcription factors facilitate Pol II binding and activity
promoter region
5’ 3’
however, Pol II cannot initiate transcription alone

Eukaryotic Transcription Initiation Complex
Basal (General) Transcription Factors: small nuclear proteins bind to promoter region constitutive, ubiquitous bind sequentially binding of individual TFs mediated by small proteins TBPassociated factors (TAFs) mediator complex (~25 proteins)
(AKA Preinitiation Complex)
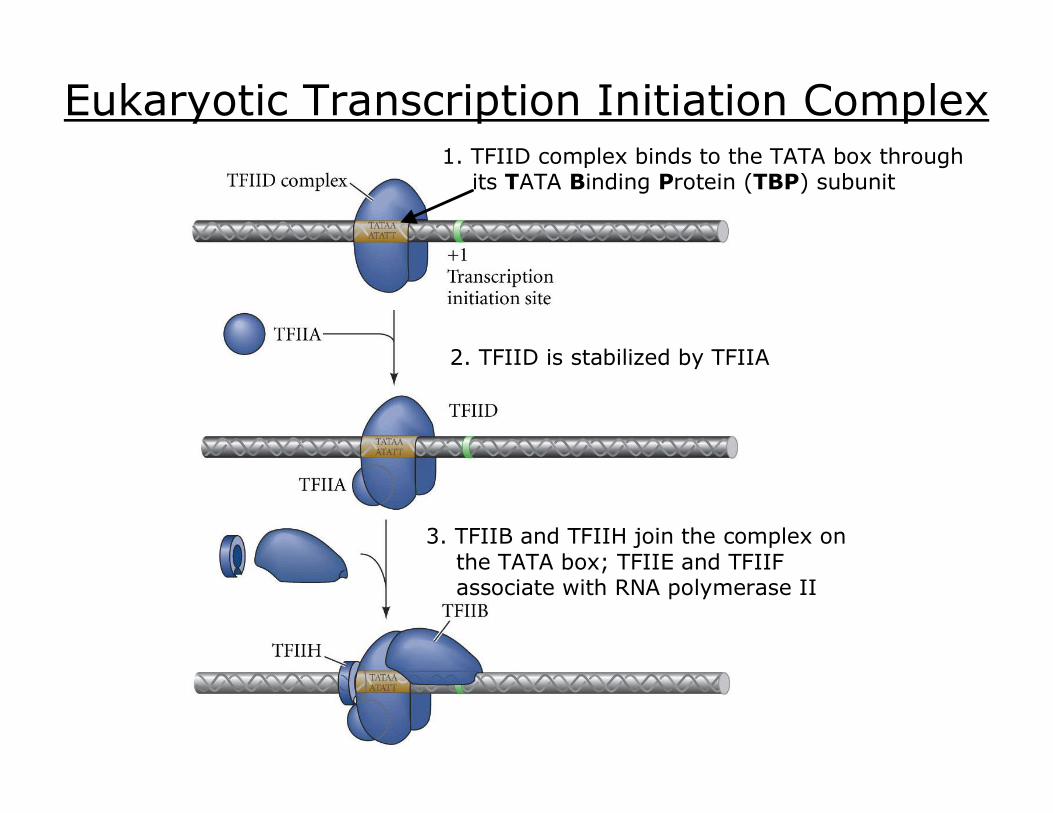
Eukaryotic Transcription Initiation Complex 1. TFIID complex binds to the TATA box through its TATA Binding Protein (TBP) subunit
2. TFIID is stabilized by TFIIA
3. TFIIB and TFIIH join the complex on the TATA box; TFIIE and TFIIF associate with RNA polymerase II

Eukaryotic Transcription Initiation Complex
5. The CTD is phosphorylated by TFIIH and is released by TFIID; RNA polymerase II can now transcribe mRNA
4. RNA polymerase II is positioned by TFIIB, and its carboxyterminal domain (CTD) is bound by TFIID
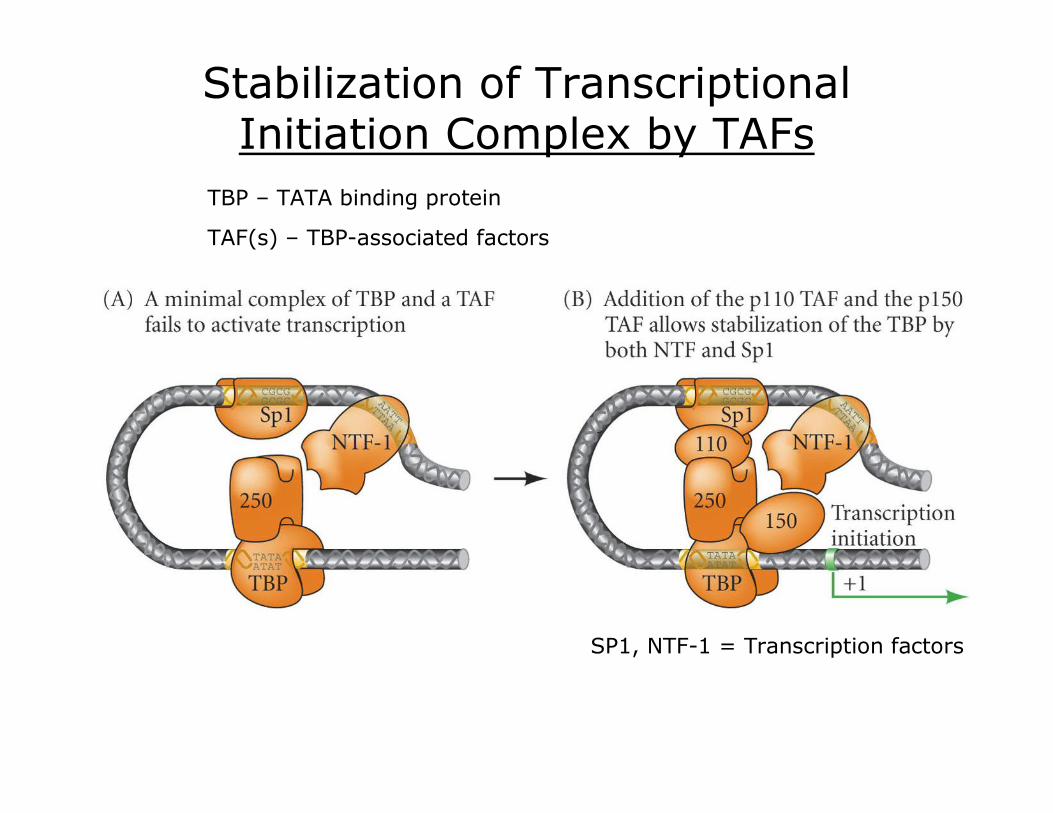
Stabilization of Transcriptional Initiation Complex by TAFs
TBP – TATA binding protein
TAF(s) – TBPassociated factors
SP1, NTF1 = Transcription factors
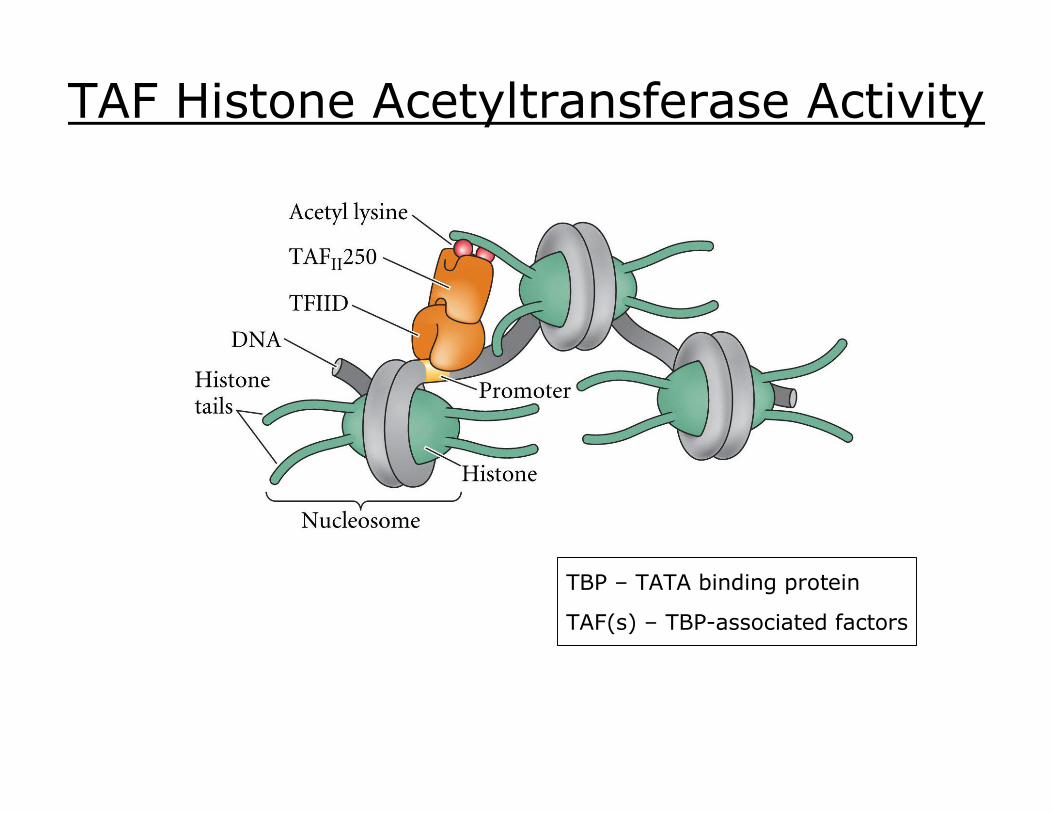
TAF Histone Acetyltransferase Activity
TBP – TATA binding protein
TAF(s) – TBPassociated factors

Enhancers
Function bind specific regulatory proteins (i.e. specific Transcription Factors) TFs generally interact with mediator complex proteins – recruit RNA polII
Enhancers: DNA sequences that regulate gene expression by affecting the transcription initiation complex on the promoter.
Location highly variable with respect to the transcribed portion of the gene. upstream (5’), downstream (3’), or within transcribed region close proximity or as many as 10 6 bp away
Enhancers are cisacting regulatory elements
cis – (same or same side); elements that reside on the same DNA strand; e.g. DNA sequences
trans – (other side); elements that originate from another DNA strand, e.g. regulatory proteins

Enhancers
Enhancers differ from promoters:
3) can work in reverse orientation
2) can work at a distance
1) need a promoter to work

Enhancer Generalizations 1. Most gene transcription requires enhancers.
2. Enhancers are the major determinants of differential transcription in cell types and through developmental stages.
3. There can be multiple signals (e.g. multiple enhancer sites) for a given gene, and each enhancer can be bound by more than one transcription factor (not at the same time).
4. Transcription is regulated by the interaction of transcription factors bound to enhancers and the transcription initiation complex assembled at the promoter.
5. Enhancers are combinatorial. Various DNA sequences regulate temporal and spatial gene expression; these can be mixed and matched.
6. Enhancers are modular. A gene can have several enhancer elements, each of which may turn it on in different sets of cells.
7. Enhancers generally activate transcription by remodeling chromatin to expose the promoter, or by facilitating the binding of RNA polymerase to the promoter by stabilizing TAFs.
8. Enhancers can also inhibit transcription (aka Silencers).

Transcription Factors
Proteins that bind to enhancer or promoter regions activate or repress transcription
Most bind to specific DNA sequences (e.g. enhancers)
Transcription factors are grouped together in families, based on structural similarities families share common framework in DNA binding sites slight differences in binding sites cause differences in recognition
Arabidopsis currently 41 TF families listed eukaryotes > 75
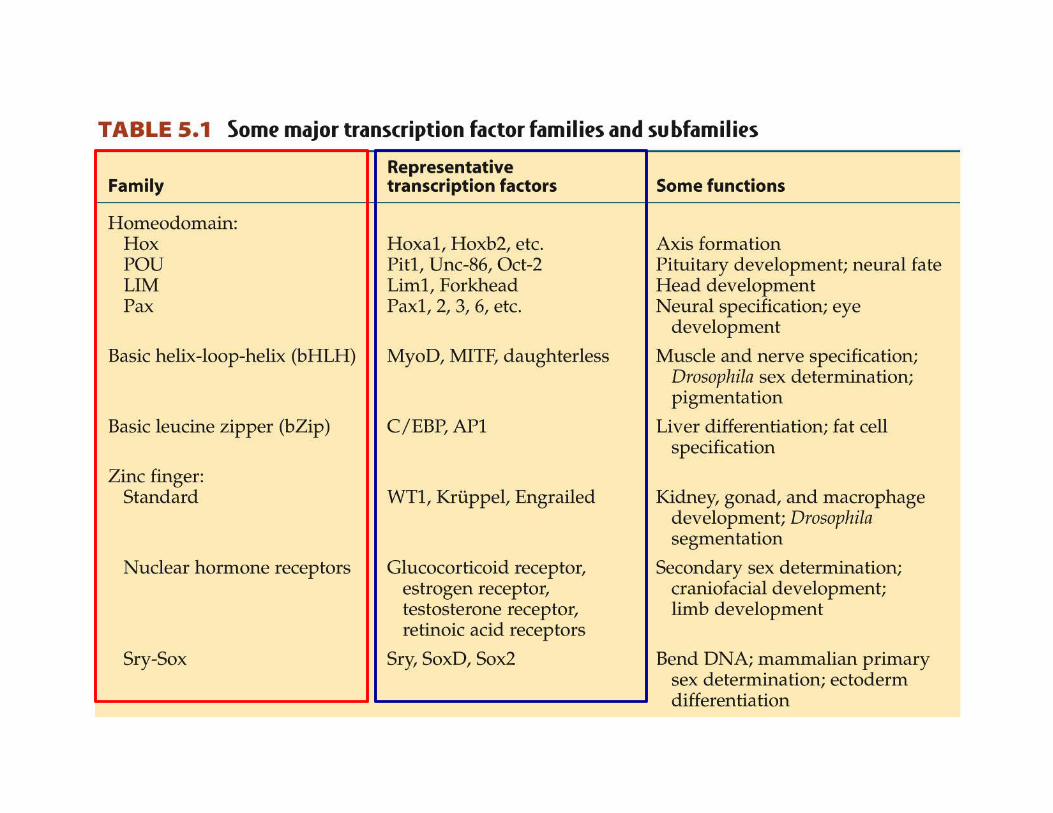

estrogen receptor zinc finger
TF/DNA Interactions
helixloophelix
leucine zipper
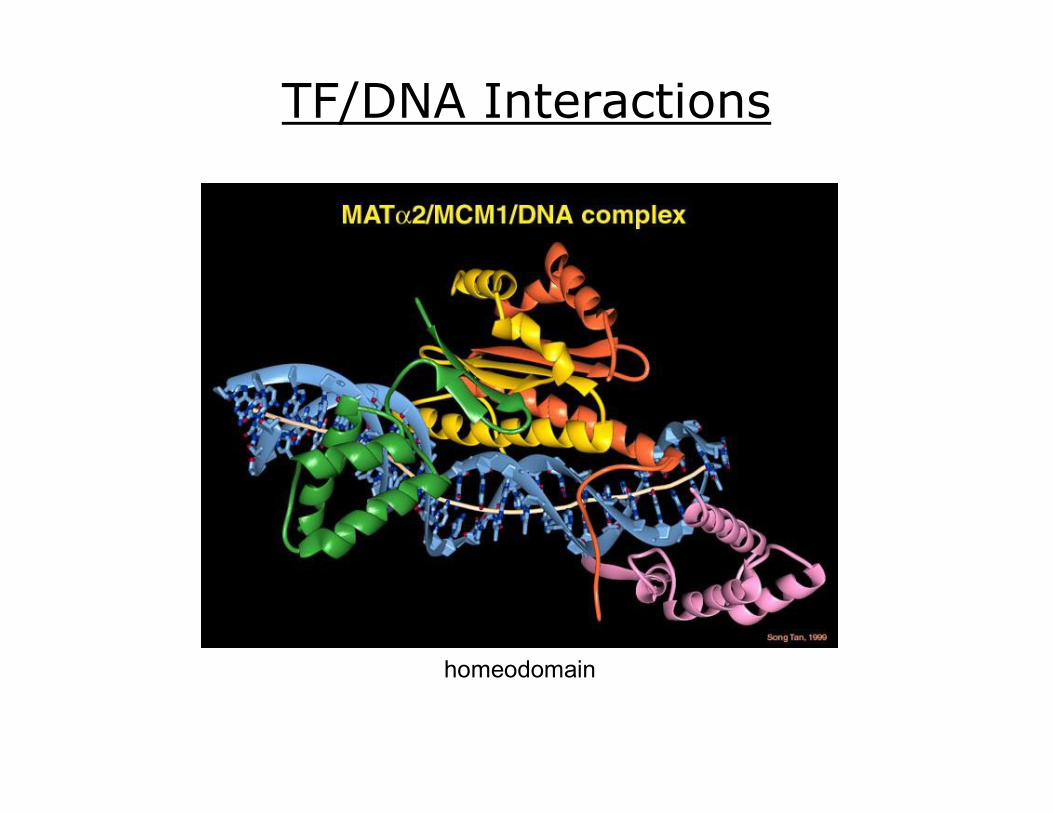
homeodomain
TF/DNA Interactions

Tbox Transcription Factor

NFkB/Rel Transcription Factor

Transcription Factor Domains
Three major domains:
1. DNAbinding recognizes particular DNA sequence
transcription factor engrailed

Transcription Factor Domains
Three major domains:
1. DNAbinding recognizes particular DNA sequence
2. Transactivation – activates or represses transcription often involved with transcription coregulator proteins involved in binding RNA polymerase II; e.g. TFIIB, TFIIE often involved with enzymes that modify histones

Transcription Factor Domains
3. proteinprotein interaction domain promotes dimerization allows it to be modulated by TAFs or other transcription factors
TBP – TATA binding protein
TAF(s) – TBPassociated factors
Three major domains:
1. DNAbinding recognizes particular DNA sequence
2. Transactivation – activates or represses transcription often involved with transcription coregulator proteins involved in binding RNA polymerase II; e.g. TFIIB, TFIIE often involved with enzymes that modify histones
NOTE – alternate categories combine 2 and 3; also add “signal sensing domain” for some TFs

Transcription Factor Domains Transcription factor MITF (microphthalmia) active in ear and pigmentforming cells of eye and skin; osteoclasts pigmentcellspecific tyrosinases expressed MITF mutation = deafness, multicolored irises, white forelock
Structural Family basic helixloophelix leucine zipper
Functional protein is a homodimer dimerization common among TFs
The transactivating domain is located near the amino terminal when bound to a promoter or enhancer, the protein is able to bind TAF p300/CBP TAF p300/CBP is a histone acetyltransferase

Functional Classification of TFs I. Constitutively active – e.g. SP1, NF1 CCAAT
II.B.3.a. Resident nuclear factors – reside in the nucleus regardless of activation state; e.g. CRDEB, AP1, Mef2
II.B.3.b. Latent cytoplasmic factors – inactive form reside in the cytoplasm, but when activated are translocated into the nucleus; e.g. STAT, R0SMAD, NFkB, Notch, TUBBY, NFAT
II.B3. Cell membrane receptordependent – second messenger signaling cascades resulting in the phosphorylation of the transcription factor
II. Conditionally active – require activation
II.A. Developmental (cell specific) expression tightly controlled, but once expressed requires no additional activation; e.g. GATA, HNF, PIT1, MyoD, MNyf5, Hox, Winged Helix
II.B1. Extracellular liganddependent – nuclear receptors
II.B2. Intracellular liganddependent activated by small intracellular molecules; e.g. SREBP, p53, orphan nuclear receptors
II.B. Signal dependent requires external signal for activation
A. H. Brivanlou et al., Science 295, 813 818 (2002)
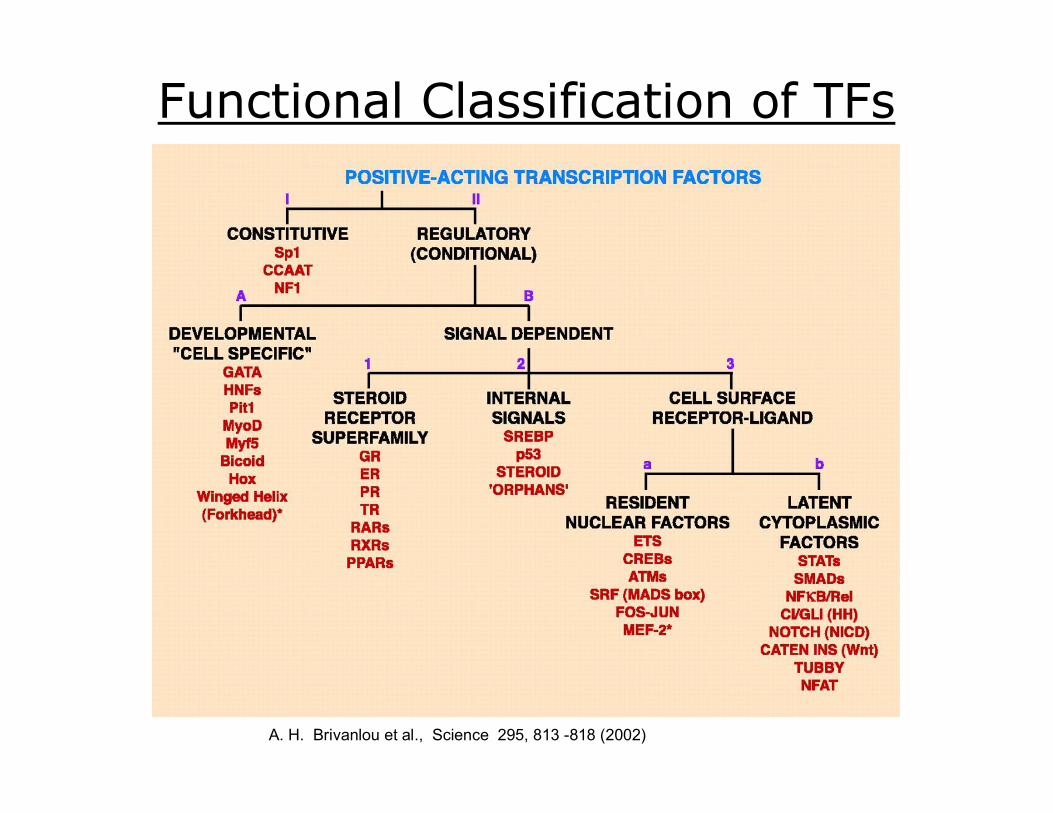
A. H. Brivanlou et al., Science 295, 813 818 (2002)
Functional Classification of TFs
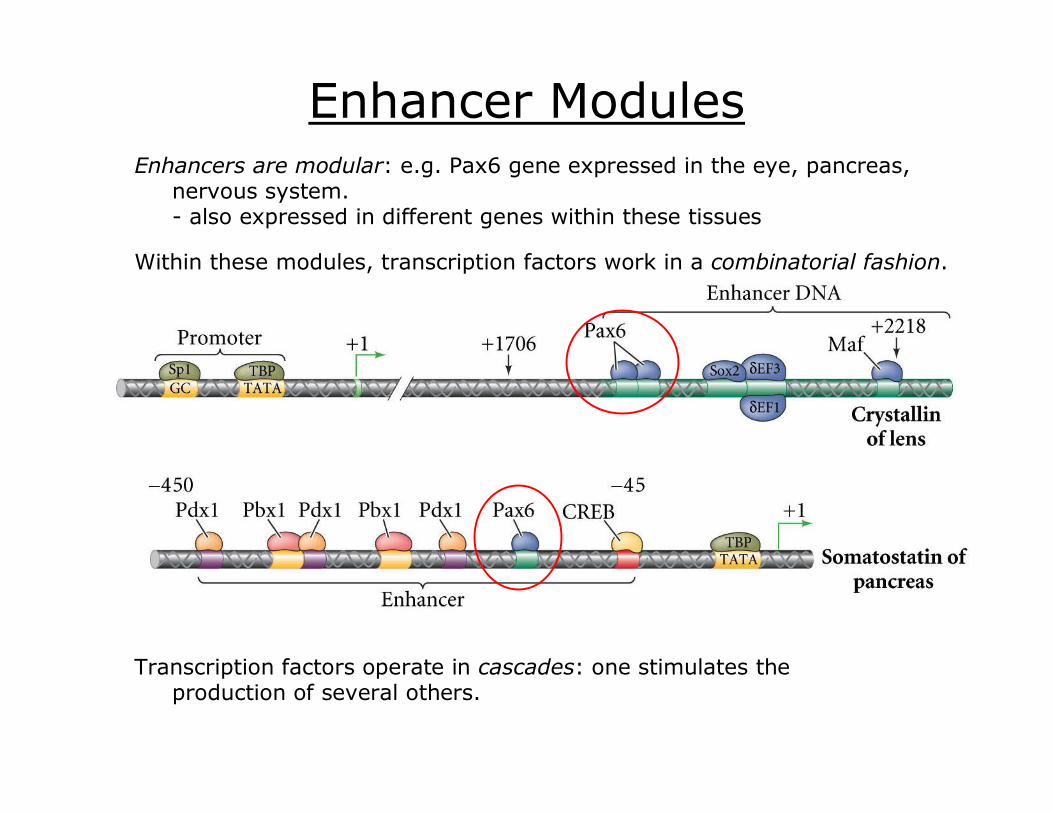
Enhancer Modules Enhancers are modular: e.g. Pax6 gene expressed in the eye, pancreas,
nervous system. also expressed in different genes within these tissues
Within these modules, transcription factors work in a combinatorial fashion.
Transcription factors operate in cascades: one stimulates the production of several others.
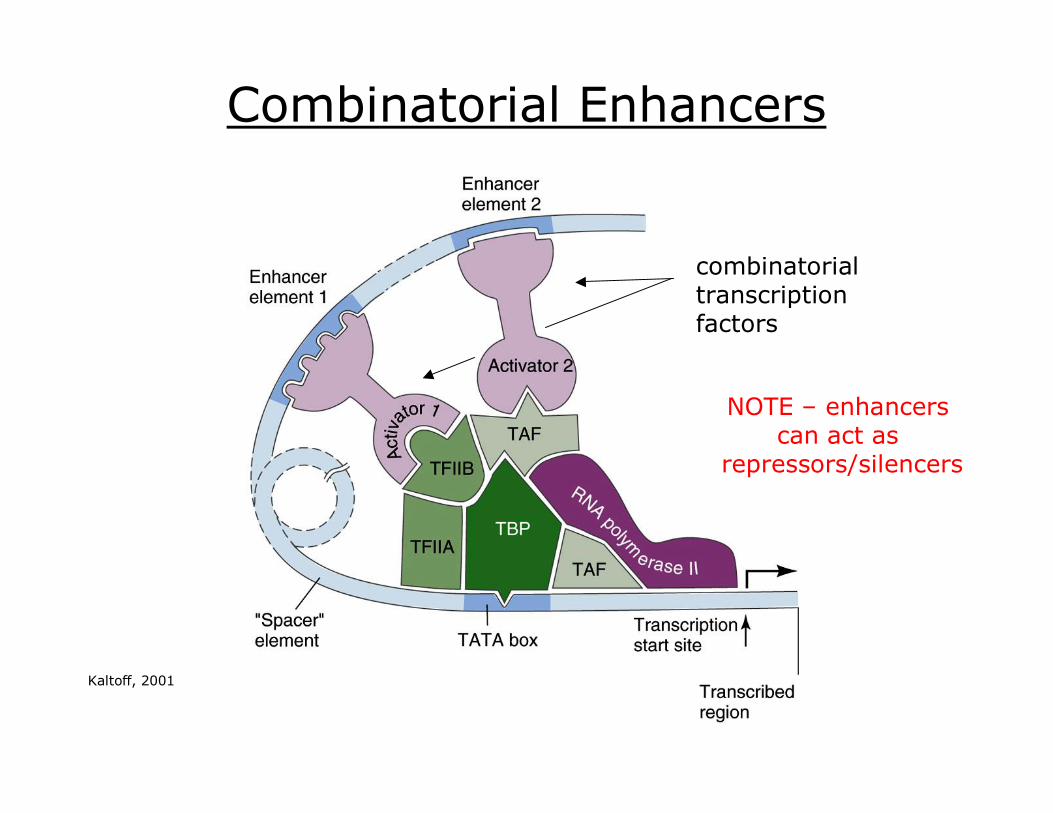
Combinatorial Enhancers
combinatorial transcription factors
NOTE – enhancers can act as
repressors/silencers
Kaltoff, 2001
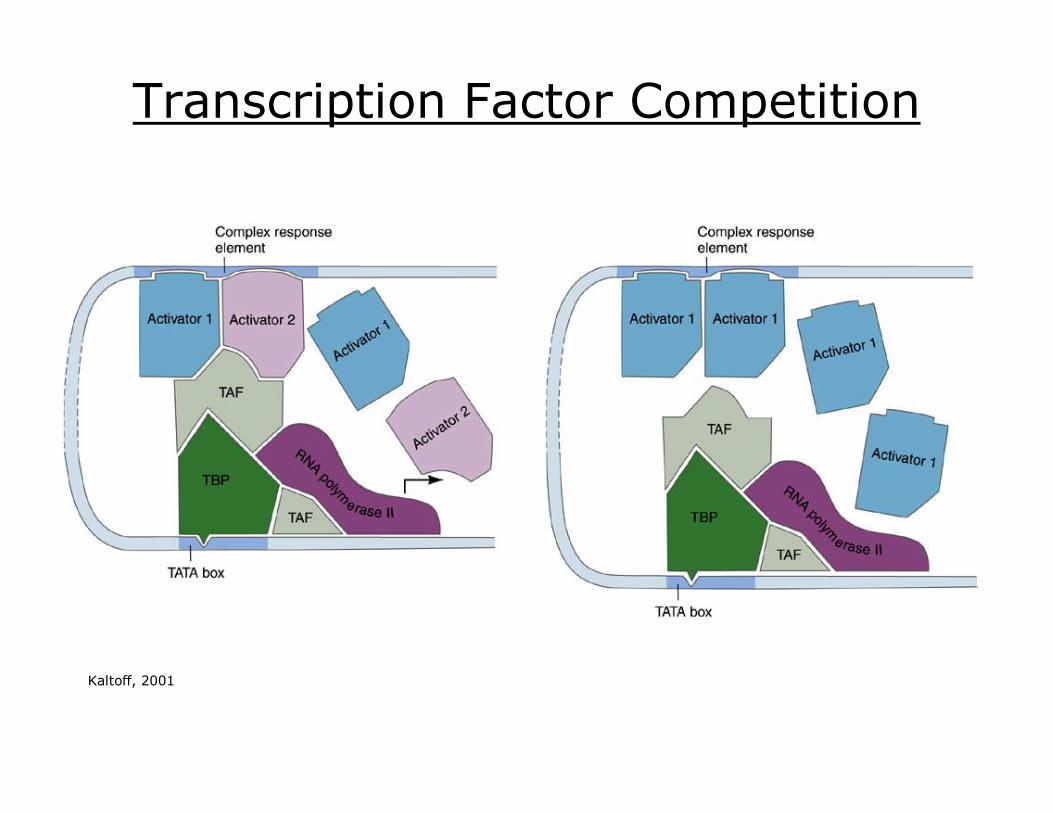
Transcription Factor Competition
Kaltoff, 2001

Epigenetic Mechanisms
Epigenetics
epi – above, etc.
control of gene expression not involving sequence
DNA methylation
imprinting
siRNA
alternative RNA splicing
histone methylation
translational control
“Development is an epigenetic, not a genetic, process.”
Mechanisms

Histone Modification
Acetylation/deacetylation – acetylation destabilizes nucleosome targets: lysines on histone “tails”
methylated histones recruit DNA methylation enzymes further stabilizes nucleosome
Methylation – stabilized nucleosome (transcription repressed) hypermethylation of H3 lysine 9 induces ectopic heterochromatin
demethylated by specific demethylating enzymes
Ubiquitination vertebrates, yeast; core histones H2A, H2B, H3 destabilizes nucleosome
Phosphorylation histone serine phosphorylation correlated with gene activation
Others?
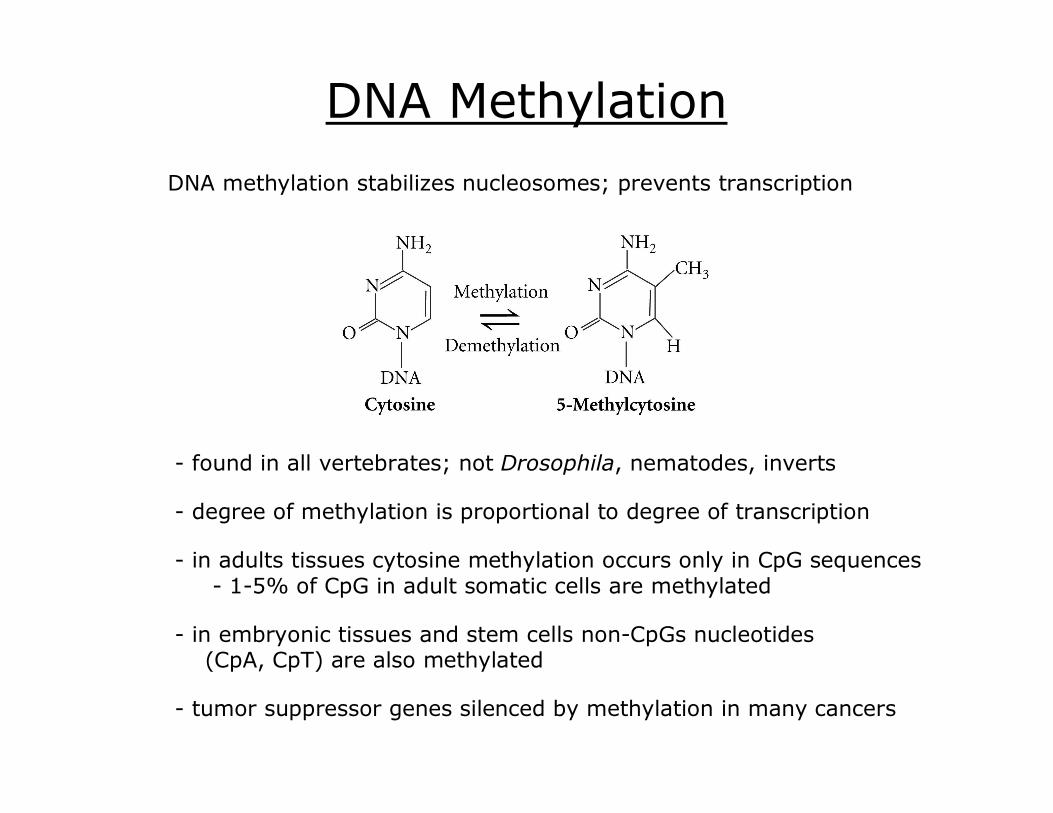
DNA Methylation
found in all vertebrates; not Drosophila, nematodes, inverts
degree of methylation is proportional to degree of transcription
DNA methylation stabilizes nucleosomes; prevents transcription
in adults tissues cytosine methylation occurs only in CpG sequences 15% of CpG in adult somatic cells are methylated
in embryonic tissues and stem cells nonCpGs nucleotides (CpA, CpT) are also methylated
tumor suppressor genes silenced by methylation in many cancers

Embryonic DNA Methylation
methylation correlates inversely with expression
Methylation patterns are established and maintained throughout cell division by DNA (cytosine5)methyltransferases
Methylation patterns differ in stage and tissuespecific manner
humans: DNMT3a, DNMT3b act as de novo methyltransferases DNMT1 maintains pattern during DNA replication
methylation pattern changes during development

Genomic Imprinting Special case of DNA methylation:
alleles from maternal and paternal genome are differentially methylated
methylation patterns can be distinguished based on resulting phenotypes
Approximately 80 imprinted genes in humans; many involved with early developmental processes
No mammalian parthenogenesis because of imprinting; in mice:
in insects, imprinting silences paternal genome produces males (functionally haploid)
Imprinting seen in mammals, insects, flowering plants
androgenetic (two male pronuclei) embryos showed poor embryonic development, but normal placental development
gynogenetic (two female pronuclei) embryos showed normal embryonic development, but poor placental development
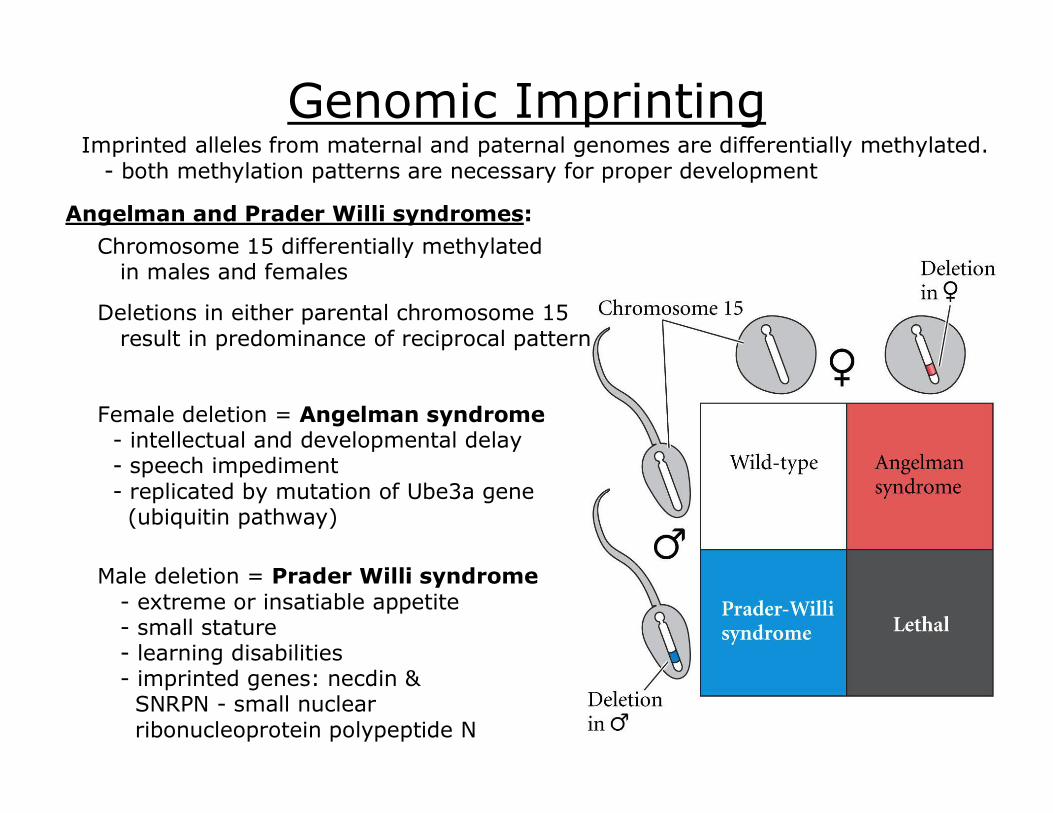
Genomic Imprinting Imprinted alleles from maternal and paternal genomes are differentially methylated. both methylation patterns are necessary for proper development
Chromosome 15 differentially methylated in males and females
Deletions in either parental chromosome 15 result in predominance of reciprocal pattern
Female deletion = Angelman syndrome intellectual and developmental delay speech impediment replicated by mutation of Ube3a gene (ubiquitin pathway)
Male deletion = Prader Willi syndrome extreme or insatiable appetite small stature learning disabilities imprinted genes: necdin & SNRPN small nuclear ribonucleoprotein polypeptide N
Angelman and Prader Willi syndromes:

Dosage Compensation
Mammals Inactivation of a single X chromosome in mammalian XX cells
Barr bodies
Mammals, Drosophila, nematodes – XX = female, XY = male; diploid XXs have double the X gene products as diploid XYs
Drosophila – transcription rate of male X is doubled
C. elegans – both Xs partially repressed ( = hermaphrodite)
XX cell early embryo – both active later embryo – only one active

X Chromosome Inactivation Lyon hypothesis:
1. Both X chromosomes active in very early female development.
2. One X is inactivated in each cell
3. Inactivation is random
4. The process is irreversible. All progeny cells will retain the same inactivation pattern
calico cat: heterozygous for coat color genes contained on X chromosome X chromosome inactivation
early late

Differential RNA Processing Nuclear RNA selection more genes transcribed than processed into mRNA unprocessed genes degraded in nucleus
Alternate splicing ~75% of all human genes may be alternatively spliced (splice isoforms; splice variants)
= one gene – one protein family nRNA splicing mediated through spliceosomes
Spliceosomes – constructed for each new hnRNA small nuclear RNAs (snRNA) splicing factor proteins
Introns contain recognition sequences GU at 5’ end splice site, AG at 3’ end splice site (typical) also variable length polypyrimidinepolypyrimidine tract (PPT) recruit spliceosome factors to site
Cellspecific splicing factors differ in ability to recognize intron sequences exon in one cells type might be recognized as an intron in another = alternative splicing
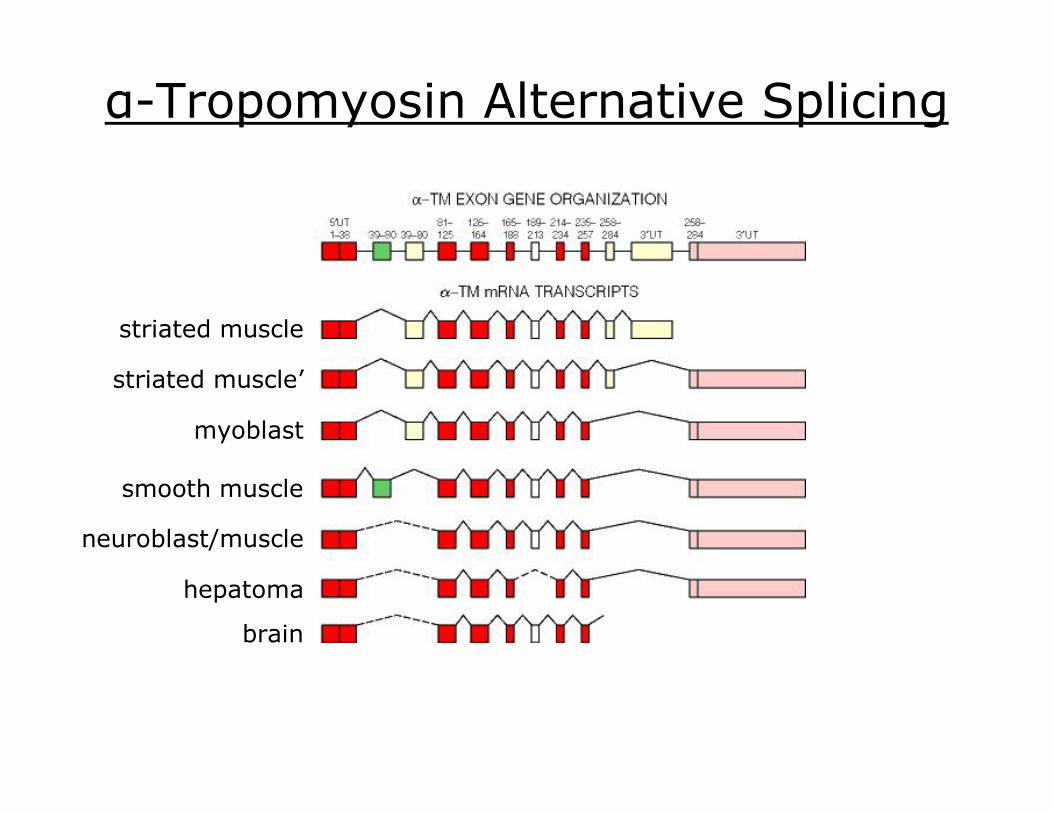
αTropomyosin Alternative Splicing
striated muscle
striated muscle’
myoblast
smooth muscle
neuroblast/muscle
hepatoma
brain

Translational Control
Rhoads, R. E. et al. Mechanism and regulation of translation in C. elegans (January 28, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.63.1, http://www.wormbook.org.
Differential mRNA longevity stability dependent on length of poly(A) tail poly(A) length dependent on 3’UTR
Selective inhibition of message translation (e.g. stored maternal messages) messages lacking 5’ cap or 3’ poly(A) tail are not translated (may be degraded)
mRNAs circularize mediated by proteins bound to 5’ cap and poly(A) tail no 5’ cap = no protein binding
proximity of 5’ and 3’ ends important for ribosomal recognition, initiation
poly(A) binding protein
eukaryotic initiation factors
ribosomal 40 S subunit
mRNA
5’ methylated guanosine cap stored mRNAs often lack methylated cap
at fertilization, methyltransferase completes cap
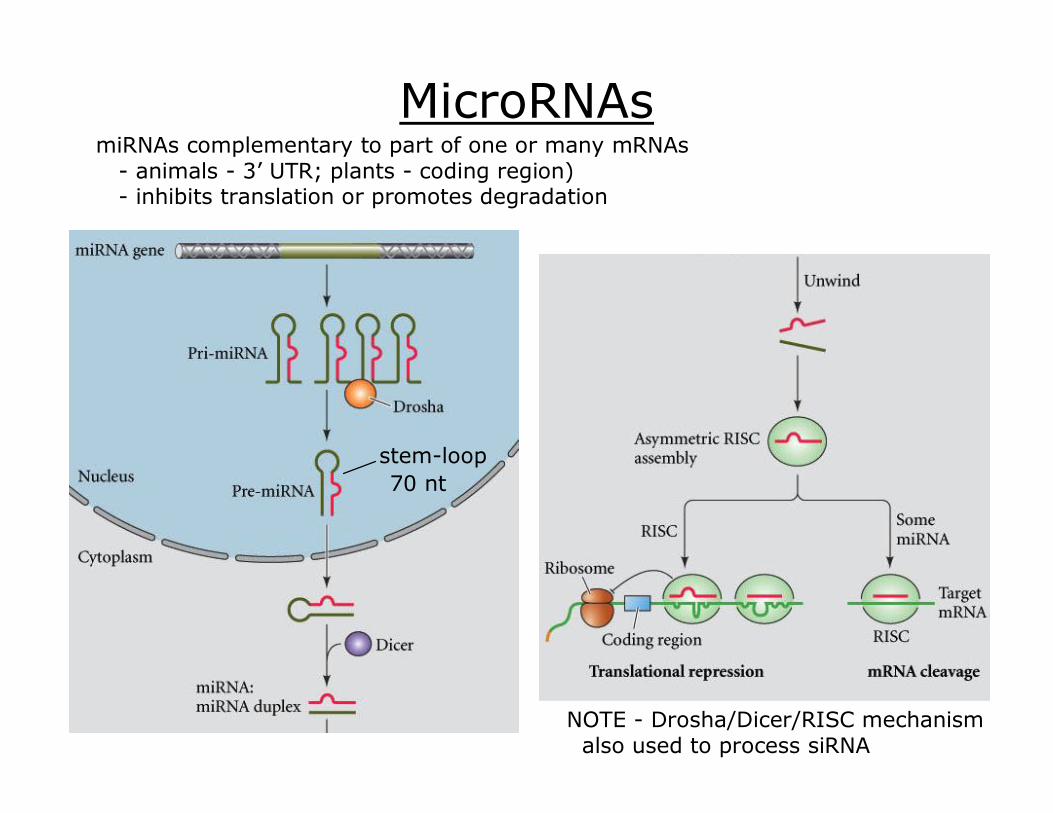
MicroRNAs
NOTE Drosha/Dicer/RISC mechanism also used to process siRNA
stemloop 70 nt
miRNAs complementary to part of one or many mRNAs animals 3’ UTR; plants coding region) inhibits translation or promotes degradation


















