
Differential Expression ofProteinKinaseCIsozyme Messenger...
7
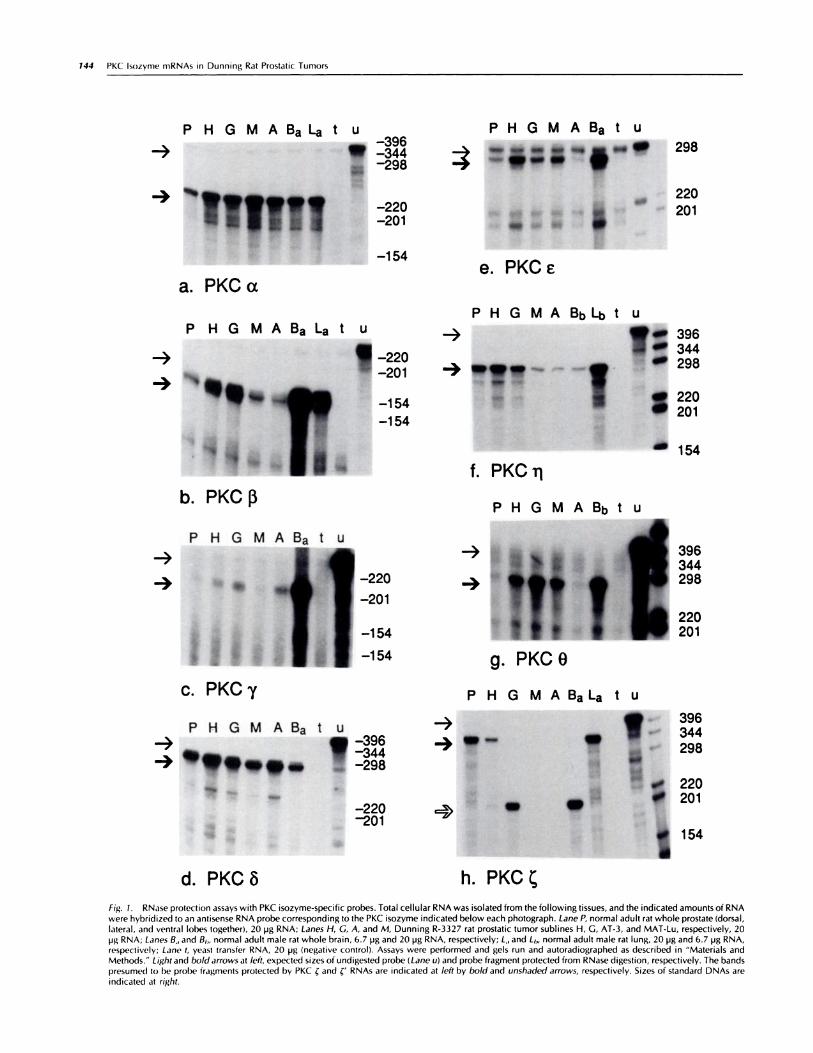
Vol. 5, 143-149, February 1994 Cell Growth & Differentiation 143 Differential Expression of Protein Kinase C Isozyme Messenger RNAs in Dunning R-3327 Rat Prostatic Tumors C. Thomas Powell,2 William R. Fair, and Warren D. W. Heston Urologic Oncology Research Laboratory, Memorial Sloan-Kettering Cancer Center, New York, New York 10021 Abstract Adenocarcinoma of the prostate is the second leading cause of cancer deaths in men. The protein kinase C (PKC) family of signal transducing kinases has been implicated in neoplastic transformation and progression in other tissues, and some evidence suggests roles for PKC in prostate growth and neoplasia. We have detected expression of eight specific PKC isozyme mRNAs (a, fi, 7, 8, #{128}, ii, 0, and ) in normal rat whole prostate and found some of these to be differentially expressed in certain Dunning R-3327 rat prostatic adenocarcinoma sublines. PKC mRNA was detected in normal prostate and Dunning H tumor, whereas an alternatively spliced form of PKC RNA was found in Dunning G tumor and normal brain. Both forms of PKC RNA were markedly reduced in the androgen insensitive, highly metastatic Dunning AT-3, MAT-Lu, and MAT-LyLu tumors. We have cloned and report the sequence of the novel portion of the alternatively spliced form of PKC RNA, which is polyadenylated and present in cytoplasm. Introduction The prostate in aging males is highly prone to benign and malignant proliferative changes (1 ). A greater understanding of the biology of normal and neoplastic prostate growth should suggest novel targets for therapy of proliferative dis- eases ofthe prostate. The PKC3 family of signal transducing, serine/threonine kinases has been implicated in regulation of normal and malignant growth in several cell types (for reviews, see Refs. 2 and 3), but scant evidence is available about PKC in the prostate. The available data support a role for PKC in normal and/or neoplastic prostate growth. First, the tumor promoter and PKC activator/down-regulator 1 2-O-tetradecanoylphorbol-i 3-acetate stimulated growth of a normal human prostatic epithelial cell line in medium containing 3.75% fetal bovine serum (4). i2-O- Tetradecanoylphorbol-i 3-acetate appears to activate most of the PKC isozymes, but not PKC (5). Second, although Received 1 0/7/93; accepted 1 1/1 5/93. 1 These studies were supported in part by NIH Grants CA39203 and DK/CA 47650. C. I. P. receives partial support from American Cancer Society In- stitutional Grant 1 1 4-0 to Memorial Sloan-Kettering Cancer Center. 2 To whom requests for reprints should be addressed, at Memorial Sloan- Kettering Cancer Center, 1 275 York Avenue, Box 516, New York, NY 10021. 3 The abbreviations used are: PKC, protein kinase C; cDNA, complementary DNA; 5’RACE, 5’ rapid amplification ofcDNA ends; PCR, polymerase chain reaction. total PKC activity appears to be low in normal rat (6, 7) and human (8) prostates, elevated PKC activity has been found in a Dunning AT-3 rat prostatic tumor subline (6) and in regressed prostates from castrated rats (7). The PKC activity assays used in those reported experiments appear to detect primarily the classical PKC isozymes a, f3, and ‘y. Finally, several peptide growth factors that have been reported to stimulate prostatic cell growth are known to activate PKC, among other signal transduction pathways (see, for example, Ref. 9). We have chosen to study PKC in the prostate at the level of individual isozymes because diverse data suggest that dif- ferent PKC isozymes may have different functions (for re- views, see Refs. 2, 3, and 10). As a first step in determining roles of PKC isozymes in prostate cancer, we have used highly specific RNase protection assays to identify which isozymes are expressed in normal rat prostate and prostatic tumor sublines. We report here that at least eight ofthe nine known PKC isozyme mRNAs (not counting alternatively spliced forms) are expressed in normal rat prostate, and some of these, including an alternatively spliced form of PKC RNA, aredifferentiallyexpressed in certain Dunning R-3327 rat prostatic tumor subhines. In addition, we have cloned the novel 5’ end of the alternatively spliced PKC RNA, which is expressed in the Dunning R-3327 G subhine and normal rat brain. Results Distribution of PKC mRNAs in Dunning Tumors. We have used RNase protection assays to identify which PKC isozyme mRNAs are expressed in normal rat prostates and in a group of Dunning rat prostatic tumors with different characteristics (summarized in Table 2; see “Materials and Methods”). RNase protection was preferred for initial studies because of its unsurpassed specificity, high sensitivity, and reproduc- ibility (1 1). We examined expression of eight of the nine known PKC isozyme mRNAs (not counting alternatively spliced forms) and found all eight (a, 3, y, 8, #{128}, ,, 0, and in normal rat whole prostate (ventral, dorsal, and lateral lobes together; Fig. 1 , a-h, Lane P). Expression of PKC A was not examined in this study. Of the classical isozymes (a, f3, .y), which appear to be expressed most highly in the brain, PKC a mRNA is expressed at a moderate level, PKC A (mix- ture of l and Il) mRNA at a low level, and PKC y mRNA at a barely detectable level in normal rat whole prostate relative to rat brain (Fig. 1 , a-c, Lanes Pand Ba; Table 1). PKC a mRNA levels are approximately 4-fold higher in all of the Dunning tumors examined than in normal prostate (Fig. 1 a; Table i ). It is not clear whether this is entirely a difference between normal and tumor tissue or whether it reflects lobe specific differences in expression of PKC a, since the Dun- ning sublines were derived from a tumor of the dorsal pros- tate lobe (12). PKC f3 mRNA levels are more than 4-fold higher in Dun- ning H and G tumors than in normal prostate and MAT-Lu
Transcript of Differential Expression ofProteinKinaseCIsozyme Messenger...

Vol. 5, 143-149, February 1994 Cell Growth & Differentiation 143
Differential Expression of Protein Kinase C IsozymeMessenger RNAs in Dunning R-3327 RatProstatic Tumors�
C. Thomas Powell,2 William R. Fair, andWarren D. W. Heston
Urologic Oncology Research Laboratory, Memorial Sloan-Kettering
Cancer Center, New York, New York 10021
Abstract
Adenocarcinoma of the prostate is the second leadingcause of cancer deaths in men. The protein kinase C(PKC) family of signal transducing kinases has beenimplicated in neoplastic transformation and progressionin other tissues, and some evidence suggests roles forPKC in prostate growth and neoplasia. We have detectedexpression of eight specific PKC isozyme mRNAs (a, fi,7, 8, #{128},ii, 0, and �) in normal rat whole prostate andfound some of these to be differentially expressed incertain Dunning R-3327 rat prostatic adenocarcinomasublines. PKC � mRNA was detected in normal prostateand Dunning H tumor, whereas an alternatively splicedform of PKC � RNA was found in Dunning G tumor andnormal brain. Both forms of PKC � RNA were markedlyreduced in the androgen insensitive, highly metastaticDunning AT-3, MAT-Lu, and MAT-LyLu tumors. We havecloned and report the sequence of the novel portion ofthe alternatively spliced form of PKC � RNA, which ispolyadenylated and present in cytoplasm.
IntroductionThe prostate in aging males is highly prone to benign andmalignant proliferative changes (1 ). A greater understandingof the biology of normal and neoplastic prostate growthshould suggest novel targets for therapy of proliferative dis-eases ofthe prostate. The PKC3 family of signal transducing,serine/threonine kinases has been implicated in regulationof normal and malignant growth in several cell types (forreviews, see Refs. 2 and 3), but scant evidence is availableabout PKC in the prostate. The available data support a rolefor PKC in normal and/or neoplastic prostate growth. First,the tumor promoter and PKC activator/down-regulator1 2-O-tetradecanoylphorbol-i 3-acetate stimulated growthof a normal human prostatic epithelial cell line inmedium containing 3.75% fetal bovine serum (4). i2-O-Tetradecanoylphorbol-i 3-acetate appears to activate mostof the PKC isozymes, but not PKC � (5). Second, although
Received 10/7/93; accepted 1 1/1 5/93.
1 These studies were supported in part by NIH Grants CA39203 and DK/CA
47650. C. I. P. receives partial support from American Cancer Society In-stitutional Grant 1 1 4-0 to Memorial Sloan-Kettering Cancer Center.2 To whom requests for reprints should be addressed, at Memorial Sloan-Kettering Cancer Center, 1 275 York Avenue, Box 516, New York, NY 10021.3 The abbreviations used are: PKC, protein kinase C; cDNA, complementaryDNA; 5’RACE, 5’ rapid amplification ofcDNA ends; PCR, polymerase chainreaction.
total PKC activity appears to be low in normal rat (6, 7) andhuman (8) prostates, elevated PKC activity has been foundin a Dunning AT-3 rat prostatic tumor subline (6) and inregressed prostates from castrated rats (7). The PKC activityassays used in those reported experiments appear to detectprimarily the classical PKC isozymes a, f3, and ‘y. Finally,several peptide growth factors that have been reported tostimulate prostatic cell growth are known to activate PKC,among other signal transduction pathways (see, for example,Ref. 9).
We have chosen to study PKC in the prostate at the levelof individual isozymes because diverse data suggest that dif-ferent PKC isozymes may have different functions (for re-views, see Refs. 2, 3, and 10). As a first step in determiningroles of PKC isozymes in prostate cancer, we have usedhighly specific RNase protection assays to identify whichisozymes are expressed in normal rat prostate and prostatictumor sublines. We report here that at least eight ofthe nineknown PKC isozyme mRNAs (not counting alternativelyspliced forms) are expressed in normal rat prostate, and someof these, including an alternatively spliced form of PKC �RNA, aredifferentiallyexpressed in certain Dunning R-3327rat prostatic tumor subhines. In addition, we have cloned thenovel 5’ end of the alternatively spliced PKC � RNA, whichis expressed in the Dunning R-3327 G subhine and normalrat brain.
Results
Distribution of PKC mRNAs in Dunning Tumors. We haveused RNase protection assays to identify which PKC isozymemRNAs are expressed in normal rat prostates and in a groupof Dunning rat prostatic tumors with different characteristics(summarized in Table 2; see “Materials and Methods”).RNase protection was preferred for initial studies because ofits unsurpassed specificity, high sensitivity, and reproduc-ibility (1 1). We examined expression of eight of the nineknown PKC isozyme mRNAs (not counting alternativelyspliced forms) and found all eight (a, �3, y, 8, #{128},,�, 0, and �in normal rat whole prostate (ventral, dorsal, and laterallobes together; Fig. 1 , a-h, Lane P). Expression of PKC A wasnot examined in this study. Of the classical isozymes (a, f3,.y), which appear to be expressed most highly in the brain,PKC a mRNA is expressed at a moderate level, PKC A (mix-ture of �l and �Il) mRNA at a low level, and PKC y mRNAat a barely detectable level in normal rat whole prostaterelative to rat brain (Fig. 1 , a-c, Lanes Pand Ba; Table 1). PKCa mRNA levels are approximately 4-fold higher in all of theDunning tumors examined than in normal prostate (Fig. 1 a;Table i ). It is not clear whether this is entirely a differencebetween normal and tumor tissue or whether it reflects lobespecific differences in expression of PKC a, since the Dun-ning sublines were derived from a tumor of the dorsal pros-tate lobe (12).
PKC f3 mRNA levels are more than 4-fold higher in Dun-ning H and G tumors than in normal prostate and MAT-Lu
�,,� 0’ �
#{149}*��
e. PKCe
P H G M A Ba La t U-396
, -344
! -298
-, �t1i!I.! :�
� -154
a. PKCx
P H G M A Ba La t U
.-� �_220-201
.4 � -154-154
b. PKC�
P H G M A Ba t U
-3
C. PKCy
P H G M A Ba t U
-3-,
*“!� �
d. PKC#{246}
I 1144
f. PKCi
P H G M A Bb t U
z,ttt,g. PKCO
P H G M A BaLa t U
-3
-,,�
9 . .�
Ih. PKC�
396344298
220201
396344298
220201
I 54
-396-344-298
-220-201
144 PKC Isozyme mRNAs in Dunning Rat Prostatic Tumors
P H G M A Ba t U
:� � 298
- 220
201
P H G M A BbLi� t U
.-� #{149},396, . 344
-3 !7J #{149}-� �t. : � 298a 220-, 201
m 154
Fig. 1. RNase protection assays with PKC isozyme-specific probes. Total cellular RNA was isolated from the following tissues, and the indicated amounts of RNA
were hybridized to an antisense RNA probe corresponding to the PKC isozyme indicated below each photograph. Lane P, normal adult rat whole prostate (dorsal,lateral, and ventral lobes together), 20 pg RNA; Lanes H, C, A, and M, Dunning R-3327 rat prostatic tumor sublines H, G, AT-3, and MAT-Lu, respectively, 20pg RNA; Lanes B,, and B,,, normal adult male rat whole brain, 6.7 pg and 20 �ig RNA, respectively; L,, and Lb, normal adult male rat lung, 20 pg and 6.7 pg RNA,respectively; Lane t, yeast transfer RNA, 20 pg (negative control). Assays were performed and gels run and autoradiographed as described in “Materials andMethods.” Lightand boldarrows at left, expected sizes of undigested probe (Lane u( and probe fragment protected from RNase digestion, respectively. The bandspresumed to l)e probe fragments protected by PKC � and �‘ RNAs are indicated at left by bold and unshaded arrows, respectively. Sizes of standard DNAs are
indicated at right.

a. 5’
b.
C.
d.
3&�0 4�3 684
3’ :143nt � 202nt
5’rt12�t
3’
I 3’
C’
-L_,,_z � I
anchor 304nt primer 2
Fig. 2. Schematic representations of rat brain PKC � cDNA sequences (a, C)
and antisense PKC � RNA probe (b). a, a PKC �cDNA sequence with completecoding region, published byOno etal.(i5). c, a published partial PKC �cDNAsequence without a complete coding region (1 4). Regions of sequence iden-
tity among the cDNAs and RNA probe are indicated by dashedlines betweenthe sequences. nt, nucleotides.
Cell Growth & Differentiation 145
Table 1 PKC isoz yme mRNA levels in normal prostate an d Dunning prostatic tumors relative to wh ole braina
IsozymeTissue
P H G M A B L
a 12 45 43 54 42 100 37
f3 3.2 13 14 4.8 2.0 100 23
0.5 1.4 2.4 0.7 3.2 100
6 60 106 80 57 77 100
C 5.5 18 9.1 12 4.0 100
�1 647 830 566 115 68 100 3075
0 5.0 38 75 36 3.4 100
‘ 25 7.6 <0.5 <0.5 <0.5 2.9 33�, <0.5 <0.5 21 <0.5 <0.5 100 <0.5
a Labels of tissues are as in Fig. 1 . Protected fragments in RNase protection assays were quantified as described in AMaterials and Methods.” Signal intensities
per pg RNA obtained from each tissue are shown relative to the value obtained from brain (set at 100) for the same isozyme. All PKC � signals, including brain,are relative to the PKC �‘ signal in brain.
and AT-3 tumors (Fig. 1 b; Table 1). These differences do notcorrelate with tumor growth rate, since the Dunning G tumorgrows 5 times faster than the H tumor, whereas the MAT-Luand AT-3 tumors grow even faster. Although the antisensePKC f3 RNA probe used here will not distinguish between the�3l and �3ll alternatively spliced forms, a separate experimentwith a probe that spans the site of alternative splicing sug-gested that f3II is the predominantform in normal rat prostateand all of the Dunning tumors shown in Fig. 1b (data notshown).
RNAs for the novel PKC isozymes 6, e, rj, and 0 are ex-pressed at low to moderate levelsin normal ratprostate rela-tive to brain (8, #{128},0) and lung (�; Fig. 1 , d-g; Table i). PKC6 mRNA levels are similar among normal whole prostate andthe Dunning tumors (Fig. 1 d), whereas PKC #{128}levels aresomewhat variable (Fig. 1 e). PKC ij mRNA levels are about6- to i 0-fold lower in androgen insensitive, highly metastaticMAT-Lu and AT-3 tumors relative to normal prostate andandrogen sensitive, low metastatic H and G tumors (Fig. i 1).PKC 0 mRNA levels in all of the Dunning tumors examinedexcept the AT-3 subline are at least 7-fold higher than innormal prostate. It should be noted that the level of expres-sion of PKC 0 is very low even in brain relative to skeletalmuscle (1 3).
We chose a fragment of PKC � cDNA that spans a potentialsite of alternative splicing, suggested by comparison of twopublished PKC � cDNA sequences (i4, iS). The antisensePKC � probe used in these studies is complementary tonucleotides 340 to 683 of a sequence published by Ono et
a!.(15), containing a complete protein coding region (Fig.2a). Another rat PKC � sequence containing an incompletecoding region was published earlier by the same group (Ref.1 4; Fig. 2c). The first 28 nucleotides of the earlier sequence(unshaded portion, Fig. 2c) differ from the later, completesequence, but the remainder of the two sequences is iden-tical, starting with nucleotide 483 ofthe complete sequence(Fig. 2, a and c). Thus, RNA with the originally publishedsequence, which we refer to as �‘, would be expected toprotect only 20i nucleotides of the antisense PKC � probe.This was the size protected by the Dunning G tumor RNAand the predominant size protected by normal rat brain RNA(Fig. 1h). PKC � mRNA levels are at least 50-fold lower inMAT-Lu and AT-3 tumors relativeto normal prostate (Fig.ih;Table i). A much longer exposure of the PKC � RNase pro-tection gel revealed very faint 201 nucleotide bands in theMAT-Lu and AT-3 lanes (data not shown), suggesting verylow expression of PKC �‘ RNA in these tumors. A similar
reduction in PKC �/�‘ RNA levels is seen in the androgeninsensitive, highly metastatic MAT-LyLu tumor and in thecorresponding MAT-LyLu cell line (data not shown).
Since the MAT-Lu, AT-3, and MAT-LyLu tumors all lackdetectable androgen receptors, we examined the effect ofandrogen removal on PKC � expression in normal rat pros-tate and in an androgen responsive human prostatic tumorcell line (LNCaP) that expresses PKC � mRNA. In rat ventralprostate, castration results in a decrease in the level of PKC
C mRNA per mg wet tissue that parallels a decrease in theamount of total cellular RNA per mg wet tissue, to a maxi-mum decrease of about 5-fold by 6 days postcastration (datanot shown). This suggests that either PKC � mRNA expressionis androgen stimulated in androgen dependent rat ventralprostate cells, or the level of PKC � mRNA is lower in an-drogen independent than in androgen dependent rat ventralprostate cells. On the other hand, LNCaP cells grown insteroid-free medium express as much PKC � mRNA as LN-CaP cells grown in medium supplemented with 10 hiM tes-tosterone (data not shown), indicating that PKC � expressionis not dependent on androgen in these cells.
PKC �‘ RNA. The published partial PKC � (i’)cDNA Se-quence (1 4) was reported to have been derived from an un-spliced PKC � RNA, since it diverges from the complete PKC� sequence at AG, a potential splice acceptor site (i 6). Wehave found by RNase protection that the PKC �‘ RNA isenriched in polyadenylated selected rat brain RNA to the

I 2345678
4
- 220
� � � �. -. 201
Fig. 3. PKC �/C RNase protection assay of Dunning G cell line and rat brainRNAs. The indicated amounts of RNA samples were hybridized to the anti-
sense PKC C RNA probe and processed as described in “Materials and Meth-ods.” Lane 1, Dunning G cell line total cellular RNA, 20 pg; Lane 2, DunningG cell line cytoplasmic RNA, 20 pg; Lane 3, Dunning G cell line nuclear RNA,
20 pg; Lane 4, rat brain total cellular RNA, 2 pg; Lane 5, rat brain polyade-nylated selected RNA, 2 pg; Lane 6, yeast transfer RNA control; Lane 7,
undigested probe; Lane 8, standard DNAs, sizes of which are indicated atright. Light arrow, size of undigested probe; boldand unshadedarrows, sizes
of probe fragments protected by PKC � and �‘ RNAs, respectively.
146 PKC Isozyme mRNAs in Dunning Rat Prostatic Tumors
. -396
.L �344
M 298
0�
same degree as PKC � RNA (Fig. 3, Lanes 4 and 5), and thatcytoplasmic RNA from the Dunning G cell line containsslightly more PKC �‘ RNA than does C cell nuclear RNA (Fig.3, Lanes 2 and 3). These data suggest that the �‘ form isindeed a messenger RNA.
5’RACE of PKC rand CRNAs. We have used 5’RACE toclone the remainder of the divergent 5’ end of PKC �‘ RNAfrom normal rat brain and the Dunning G cell line. PCRamplification using an ohigonucleotide primer complemen-tary to nucleotides Si 7 to 498 of the complete rat PKC �sequence (i 5) and the 5’RACE System Anchor primer (see“Materials and Methods”) yielded the products shown in Fig.4A. Rat brain yielded two major fragments, visualized on a2% agarose gel: an upper band of about 570 base pairs anda wide lower band ranging from about 420 to 340 base pairsin length (Fig. 4A, Lane 1 ). The G cell line, on the other hand,yielded only one major band, centered at about 410 basepairs in length (Fig. 4A, Lane 2). We cloned and sequencedseveral PCR products from both brain and G cell line thatcontained, from 5’ to 3’, Anchor primer, (dGMP)16_3�, 304nucleotides of novel sequence, the 28 nucleotides at the 5’end of the published partial PKC � (c’) sequence (14), andnucleotides 483 to Si 7 ofthe complete rat PKC � sequence(Ref. 1 5; Figs. 2dand 4B). Presumably these sequences cor-respond to the wide lower band in Fig. 4A, Lane 1, and the410-base pair band in Fig. 4A, Lane 2. In addition, the se-quences of some of the products derived from brain RNA,but none ofthe products from G cell RNA, proved to be the5’ end of PKC � mRNA. This is in agreement with the RNaseprotection assay that showed a small amount of PKC � RNA
in rat brain relative to PKC �‘ RNA, but no detectable PKC� RNA in Dunning G tumor (Fig. 1 h, Lanes Ba and G).
The sequences of PKC �‘ clones from rat brain and Dun-ning G cell line are identical to each other and contain trans-lation termination codons in all three reading frames (Fig.4B). Thus, even if the 5’RACE products do not contain theentire 5’ end of PKC �‘ RNA, the novel sequence cannotcontain an open reading frame. When the novel PKC �‘ RNAsequence is combined with the PKC � mRNA sequencedownstream of the splice site, the first ATG codon headingan open reading frame is 1 0 nucleotides downstream of theend of the coding sequence for the cysteine-rich domain of
PKC � (1 5). Thus, if a protein were to be translated from PKC
C RNA, it would consist of the kinase domain of PKC �without most of its regulatory domain and would have apredicted molecular weight, without posttranslational modi-uications, of 46,600.
DiscussionThe large number of PKC isozyme mRNAs found in rat pros-tate suggests that PKC signaling pathways have importantroles in this organ. Among other tissues studied, only thebrain (13, 17, 18) and retina (19) are known to express asmany isozymes as we have found in the prostate. Althoughthe levels of most of the novel PKC RNAs are lower in themetastatic Dunning tumors, particularly the AT-3 tumor,than in normal prostate, our results are not necessarily in-consistent with the previous report (4) of 3- to 4-fold higherPKC activity in Dunning AT-3 tumors relative to normal ratprostate. In those studies, PKC activity was determined usingan in vitro assay in which the substrate proteins were his-tones, which are phosphorylated much more efficiently bythe classical PKC isozymes a, �3, and ‘y than by the otherisozymes tested to date (& #{128},i’s, �). Thus, although the levelsof PKC #{128},.#{239}#{236},0, and � RNAs are much lower in the DunningAT-3 tumor relative to normal prostate, the difference in PKCactivity reported by Battistone et a!. (6) might be accountedfor by the 3.5- and 6-fold higher levels of PKC a and y mR-NAs, respectively, found in the AT-3 tumor relative to nor-mal whole prostate (Table 1 ). Also, there may be differencesin translation efficiencies of PKC RNAs between tumor andnormal prostate.
The low levels of PKC i� mRNA in the more aggressiveDunning tumors are in agreement with a recent report thatPKC ‘q expression is higher in differentiated versus prohifer-ating skin, lung, and gut epithehial cells (20), suggesting apossible role for this isozyme in differentiation or growthinhibition. The consistent loss of PKC � expression in thehighly metastatic, androgen insensitive Dunning tumors sug-gests a possible involvement of this isozyme in regulation oftumor aggression. Our observations are consistent with re-suIts of a previous study in keratinocytes, in which Northernblot analysis showed lower PKC � mRNA levels in benignneoplastic keratinocyte cell lines 308 and SP-1 than in nor-mal cultured primary keratinocytes (21).
It is unclear whether the lack of PKC � mRNA expressionin the MAT-Lu, AT-3, and MAT-LyLu tumors is due to theabsence ofdetectable androgen receptors in these cell lines.Although the level of PKC � mRNA appears to be androgensensitive in normal rat ventral prostate, the persistence ofPKC � mRNA expression in LNCaP prostatic tumor cells aftersteroid withdrawal suggests that the reduction in PKC � ex-pression in the metastatic Dunning tumors may not be dueto inability ofthese cells to respond to androgen. This ques-tion could be further addressed by examination of PKC �
expression in MAT-LyLu cells transfected with androgen re-ceptor cDNA and expressing functional androgen receptor.
It is not clear at this time whether a protein is translatedfrom PKC �‘ RNA, although it seems unlikely, owing to thelong (553 nucleotides) 5’ untranslated sequence and ab-sence of consensus sequence around the ATG codons of theopen reading frame (22). Also, such a protein would mostlikely be highly unstable. Polyclonal antibodies againstCOOH-terminah PKC � peptides are available but must beused with caution to distinguish between PKC �‘ and pro-teolytic fragments of PKC �, or cross-reacting proteins. Nev-ertheless, the presence of an alternatively spliced form of

A123
B
-3
1 TGACGT CAGCTC CTCCGT ATCCAT GCCGCG TTTTA� TGTGTG CCTCAG CTGCCG GCTGTh
61 �CTTCC CAGGGG AGCCGG GTACCA CCCGGG CCTGGA GACA�Q...AGGAGG CAGAGA TGI�AG
12 1 AGGCGG GGAGCA GGACAG TCAACC TTCTAT Th�ATQ CCTGCT CTCCAG CAGIM CCCACC
181 CTGCCT ATTGTC GATCCG GAGACC CACCCA GCCCAT TGTCGG TCCGGA GGCCAG GGGACG
241 CAGCAT CGGCTC
Eco RI
CTTA�A GGGACG GAAGAT GCCTCC TTGCGC GCCCAC CTTCGG �AGC
3 0 1 AIMAG AATTCC GTGCT�.�AGAGG
361 CCAGAA GATGGA GGA 375
CAGCAG AGAAAG CCGAG/T CCATCT ACCGCC GTGGAG
Fig. 4. PKC �/�‘ 5’RACE. A, 2% agarose minigel of5’RACE products. Lane 1, rat brain RNAtemplate; Lane 2, Dunning G cell linetemplate. Lightand bold arrowsat left, products shown by sequencing to be PKC � and �‘ 5’ end cDNAs, respectively. B, nucleotide sequence of rat PKC �‘ 5’RACE product, beginning after(dGMP),,_,0. Methionine codons are typed in bo/d;termination codons are underlined. I, presumed splice site; italicized sequence, beginning with AG upstreamof the splice site, is common to PKC � and �‘ RNAs. Sequence from the EcoRl site to the 3’ end matches partial PKC � sequence of Ono et al. (14). GenBank
accession number L23321.
HI-S .�Hl-M �HlF--AT-3
. . H�-AT-2OrIgInal ..� N� �MAT-LuR3327 :: AT-iTumor ‘#{176}4�G MAT-LyLu
Fig. 5. Derivation of Dunning R-3327 rat prostatic tumor sublines, from Ref.23. - - - -, multiple serial passages; A, castration-induced selection.
Cell Growth & Differentiation 147
1018
517, 506
396344298
220, 201
154, 134
PKC � RNA in normal rat brain and in one subhine of Dunningrat prostatic tumors is intriguing.
Materials and Methods
Dunning R-3327 Tumors and Cell Lines. The DunningR-3327 H, G, MAT-Lu, MAT-LyLu, and AT-3 rat prostatictumor sub�ines were derived from a spontaneous tumor ofthe dorsal prostate in a 22-month-old Copenhagen rat (1 2),following serial in vivo passaging as described (Ref. 23; Fig.5; Table 2). Tumor sublines were passaged in our laboratoryby inoculating 25- to 30-mg pieces s.c. in the flanks of adultmale Copenhagen rats (Harlan Sprague-Dawley, Indianapo-his, IN). When tumor volumes reached about 2 cm3, tumorswere harvested under sodium pentobarbital anesthesia, im-mediately frozen in liquid nitrogen, and stored at -80#{176}C.Normal brain and lung tissues were harvested from rats not
bearing tumors. The Dunning G and MAT-LyLu cell lineswere established from the corresponding tumors (23) and
generously provided by Dr. J. Isaacs. Cells were maintainedin RPMI 1 640 medium supplemented with 2 mxii-glutamine, 1 0% fetal calf serum (HyClone Laboratories,Logan, UT), 250 nM dexamethasone (Sigma Chemical Co.,St. Louis, MO), 1 00 units/mI potassium penicillin G, and 100units/mI streptomycin sulfate and incubated at 37#{176}Cin ahumidified, 5% CO2 incubator.
RNA Isolation. Total cellular RNA was isolated from nor-mal rat tissues and Dunning R-3327 tumor subhines by ho-mogenization in guanidinium thiocyanate and centrifuga-tion through cesium chloride (24). For isolation ofcytoplasmic and nuclear RNA, monolayer cultures of Dun-ning G cells were washed with ice-cold 4.3 m�i Na2HPO4,1.4 ms� KH2PO4, 2.7 m� KCI, 137 mxi NaCI, scraped intophosphate-buffered saline, pelleted by centrifugation, andresuspended in ice-cold lysis buffer [50 msi Tris-Ch (pH 8.0),1 00 ms� NaCI, 5 m� MgCI2, 0.5% (v/v) Nonidet P-40, pre-pared with diethylpyrocarbonate treated water and filtersterilized]. Lysates were incubated for 5 mm on ice, and thennuclei were pelleted by centrifugation at 2500 X gfor 5 mm.The supernatants were treated with Proteinase K, and cyto-plasmic RNA was isolated by phenol-chloroform extractionas described (25).
RNase Protection Assays. For generation of antisenseRNA probes, cDNA clones for rat PKC �3, y, 6, #{128},and �,allgenerously provided by Dr. Y. Nishizuka (14, 26, 27), wereutilized. Restriction fragments of these cDNAs, encompass-ing the regions shown in Table 3, were subcloned intopBluescript KS(-) or SK(-) vectors (Stratagene, La Jolla, CA),oriented so that transcription from the T3 promoter of eachvectorwould yield a short piece ofvector sequence followed
by antisense PKC RNA. The PKC � cDNA fragment, nucleo-tides 340 to 684 of a PKC � cDNA containing a completeprotein coding region (Ref. 15; Fig. 2, a and b), spans apotential site of alternative splicing. Nucleotides 483 to 684of this fragment are identical to a region near the 5’ end ofa partial PKC � sequence published by the same group (Ref.14; Fig. 2c), whereas the sequence upstream of nucleotide483 differs from the upstream sequence ofthe partial cDNA.
Fragments of DNA for rat PKC a, �, and 0, shown in Table3, were synthesized and amplified from rat brain (a, 0) orlung (�) RNA by reverse transcription-polymerase chain re-action (28), using Superscript RNase H- reverse transcriptase(GIBCO-BRL, Gaithersburg, MD), Taq DNA polymerase(Promega, Madison, WI), and oligonucheotide primers syn-thesized in the Sloan-Kettering Institute MicrochemistryCore Facility. Amplified products were ligated into the TAcloning vector pCR II (Invitrogen, San Diego, CA), andclones were isolated with cDNA inserts oriented so that tran-scription from the SP6 promoter of pCR II yielded antisensePKC RNA. The identities and orientations of the PCR gen-erated clones were confirmed by sequencing the doublestranded DNA, using the dideoxy chain termination method
148 PKC Isozyme mRNAs in Dunning Rat Prostatic Tumors
Table 2 In vivo chara cteristics of Dunning R-332 7 rat prostatic tumor sublines used in this study’
TumorAndrogen
.responsive
Androgen receptorcOntend’
Doubling time (days) Metastatic ability
H Yes 290 21 Low
G Yes 1450 4.0 LowMAT-Lu No ND� 2.7 High (lung)MAT-LyLu No ND 1 .7 High (lymph node, lung)AT-3 No ND 1 .8 High llymph node, lung)
.1 Data from Ref. 23.I, FmoI of receptor/i 08 cells.,ND, not detectable.
Table 3 Regions of PKC cDNA s used for synthesis of RNA probes
lsozyme Fragmenta Ref.Fragment
lengthbComplete
probec
a 193-431 26 238 35813 585-831 27 156d 227
.y 2182-2383 26 202 242
6 959-1252 14 294 371
#{128} 1186-1469 14 284 303
TI 408-705 18 298 418
0 458-753 13 296 416
340-684 15 345, 202’ 416
a Nucleotide numbers according to published sequences.b Length, in nucleotides of PKC cDNA fragment, equals the length of RNA
probe expected to be protected from RNase digestion by the correspondingPKC RNA.C Length in nucleotides of the undigested RNA probe, including vector se-quences on one or both ends.d Length of PKC �3 fragment after digestion with BssHIl at nucleotide 676.,. All 345 nucleotides match full length PKC � cDNA sequences (15);only nucleotides 483 to 684 match the alternatively spliced PKC � cDNA
sequence (14).
(29) with Sequenase (U.S. Biochemical, Cleveland, OH) andfollowi ng the manufacturer’s instructions.
Prior to synthesis of antisense RNA probes, each of theabove clones was cleaved at a restriction site in the vectorpolylinker downstream of the 3’ end ofthe antisense strand.An exception was the PKC 13subclone, which was cleavedat a BssHll site at nucleotide 676 of the PKC �3 cDNA, 156nucleotides from the 5’ end of the antisense strand. Tran-scription of the digested DNA with T3 or 5P6 RNA polym-erase (1 i) and equimolar amounts (35 pmol) of [a-32PICTP(3000 Ci/mmol) and unlabeled CTP yielded high specificactivity antisense RNA probes. Several nucleotides at the 5’end and, in most cases, the 3’ end of each probe consistedof pBluescript sequences that should not be protected fromRNase digestion by eukaryotic RNA, thus providing an in-ternal control for each assay.
Total cellular RNA samples were hybridized to the anti-sense RNA probes, and RNase protection was carried out asdescribed (1 1), using 1 x 10� cpm of probe in each 30 hi1reaction mixture. RNA fragments protected from RNase di-gestion were visualized by electrophoresis on S%
polyacrylamide-urea gelsfollowed by autoradiography. Pro-tected fragments were quantified by scanning the gels witha Betascope Model 603 blot analyzer (Betagen Corp.,Waltham, MA) or the autoradiographs with an UltroScan XLlaser densitometer (Pharmacia LKB, Piscataway, NJ). Al-though a control probefor RNA quality/loading was not usedin the RNase protection assays, the assay was repeated withRNA isolated from two different tumor or tissue samples withsimilar results.
5’ Rapid Amplification of cDNA Ends. Single strandedcDNA was synthesized from 1 .5 �g of rat brain total cellularRNA or Dunning G cell line cytoplasmic RNA, using anoligonucleotide primer complementary to nucleotides 690to 7iOofcomplete PKC �cDNA(15)and Superscript reversetranscriptase (GIBCO-BRL). The mRNA template was de-graded, and the cDNA was purified and tailed with dCMPnucleotides using a 5’RACE System (Ref. 30; GIBCO-BRL)and following the manufacturer’s instructions. The tailedcDNA was amplified using the 5’RACE System AnchorPrimer and an oligonucleotide primer complementary tonucleotides 498 to Si 7 of the complete rat PKC � cDNA.Amplification reactions contained about one-twenty-fifth ofthe product from a given cDNA synthesis reaction in a finalvolume ofSO hi1containing 7.7 m� Tris-CI (pH 8.8), i 6.6 mt�iNH4SO4, 1 .63 mM MgCl2, 200 hiM deoxyribonucleotides,300 ns� of each primer, 200 hig/�1 bovine serum albumin,and 1 unit ofTaq DNA polymerase (Promega). Amplificationwas performed in a Perkin-Elmer Model 480 DNA thermalcycler using a “hot start” protocol (31) and 38 cycles of de-naturation at 94#{176}Cfor 30 s, annealing at 54#{176}Cfor 1 mm, andextension at 72#{176}Cfor 2 mm. One-tenth ofeach reaction wasanalyzed by separation on a 2% agarose minigel, followedby ethidium bromide staining and photography of the UVlight illuminated gels using an Eagle Eye still video system(Stratagene). Amplified products were ligated into the TAcloning vector pCR II (Invitrogen), and clones were se-quenced as described above.
Acknowledgments
We thank Anthony Palattella and Robert Huryk for expert technical assis-tance.
References
1 . Carter, H. B., and Coffey, D. S. The prostate: an increasing medical prob-lem. Prostate, 16: 187-197, 1990.
2. Nishizuka, Y. Studies and prospectives of the protein kinase C family forcellular regulation. Cancer (Phila.), 63: 1892-1903, 1989.
3. Ohno, S., Akita, Y., Hata, A., Osada, S-I., Kubo, K., Konno, Y., Akimoto,K., Mizuno, K., Saido, I., Kuroki, I., and Suzuki, K. Structural and functionaldiversities of a family of signal transducing protein kinases, protein kinase C
family; two distinct classes of PKC, conventional cPKC and novel nPKC. Adv.EnzymeRegul., 31:287-303, 1991.
4. Lechner,J. F., and Kaighn, M. E. Nutrition ofprostatecells. Prog. Clin. Biol.Res., 37:217-232, 1980.
5. Nakanishi, H., and Exton, J. H. Purification and characterization of the �isoform of protein kinase C from bovine kidney. J. Biol. Chem., 267: 16347-16354, 1992.
6. Battistone, M. J., Padilla, G. M., and Petrow, V. 1-Dehydromelengestrolacetate inhibits the growth and protein kinase C activity of androgen-independent Dunning rat prostatic tumors. Cancer Chemother. Pharmacol.,31:407-411, 1993.
7. Durham, I. P., Butcher, F. R., Teoh, T. S., Miller, R., and Mawhinney, M.

Cell Growth & Differentiation 149
G. Characterization and androgenic regulation ofsoluble protein kinases andprotein phosphorylation in rat ventral prostate gland. Arch. Biochem. Bio-phys., 244:692-700, 1986.
8. Kase, D. J., Kirschenbaum, A., Li, H. C., and Droller, M. J. Protein kinaseC in the human prostate. J. Urol., 139: 220A, 1988.
9. McKeehan, W. 1. Growth factor receptors and prostate cell growth. CancerSurv., 11: 165-175, 1991.
1 0. Clemens, M. J., Trayner, I., and Menaya, J. The role of protein kinase Cisozymes in the regulation ofcell proliferation and differentiation. J. Cell Sci.,103:881-887, 1992.
1 1 . Melton, D. A., Krieg, P. A., Rebagliati, M. R., Maniatis, T., Zinn, K., andGreen, M. R. Efficient in vitro synthesis of biologically active RNA and RNAhybridization probes from plasmids containing a bacteriophage 5P6 pro-moter. Nucleic Acids Res., 12: 7035-7056, 1984.
1 2. Dunning, W. F. Prostate cancer in the rat. NatI. Cancer Inst. Monogr., 12:351-369, 1963.
1 3. Osada, S., Mizuno, K., Saido, I. C., Suzuki, K., Kuroki, I., and Ohno, S.A new member of the protein kinase C family, nPKC 0, predominantly ex-pressed in skeletal muscle. Mol. Cell. Biol., 12: 3930-3938, 1992.
14. Ono, Y., Fujii, I., Ogita, K., Kikkawa, U., Igarashi, K., and Nishizuka, Y.The structure, expression and properties of additional members ofthe proteinkinase C family. J. Biol. Chem., 263: 6927-6932, 1988.
15. Ono, Y., Fujii, I., Ogita, K., Kikkawa, U., Igarashi, K., and Nishizuka, Y.Protein kinase C � subspecies from rat brain: its structure, expression and
properties. Proc. NatI. Acad. Sci. USA, 86: 3099-3103, 1989.
16. Smith, C. W. J., Patton, J. G., and Nadal-Ginard, B. Alternative splicingin the control of gene expression. Annu. Rev. Genet., 23: 527-577, 1989.
1 7. Wetsel, W. C., Khan, W. A., Merchenthaler, I., Rivera, H., Halpern, A.E., Phung, H. M., Negro-Vilar, A., and Hannun, Y. A. Tissue and cellulardistribution of the extended family of protein kinase C isoenzymes. J. CellBiol., 117:121-133, 1992.
1 8. Osada, S., Mizuno, K., Saido, I. C., Akita, Y., Suzuki, K., Kuroki, I., andOhno, S. A phorbol ester receptor/protein kinase, nPKC �,a new member ofthe protein kinase C family predominantly expressed in lung and skin. I. Biol.Chem., 265:22434-22440, 1990.
1 9. Fujisawa, N., Ogita, K., Saito, N., and Nishizuka, Y. Expression of proteinkinase C subspecies in rat retina. FEBS Left., 309: 409-41 2, 1992.
20. Osada, S., Hashimoto, Y., Nomura, S., Kohno, Y., Chida, K., Tajima, 0.,Kubo, K., Akimoto, K., Koizumi, H., Kitamura, Y., Suzuki, K., Ohno, S., and
Kuroki, I. Predominant expression of nPKC i�, a Ca2�-independent isoformof protein kinase C in epithelial tissues, in association with epithelial differ-entiation. Cell Growth & Differ., 4: 167-175, 1993.
21 . Dlugosz, A. A., Knopf, J. L., and Yuspa, S. H. mRNA encoding proteinkinase C a, d, e and � is expressed in normal and neoplastic mouse kera-tinocytes in vitro. Clin. Res., 39: 485A, 1991.
22. Kozak, M. An analysis of vertebrate mRNA sequences: intimations oftranslational control. J, Cell Biol., 1 15: 887-903, 1991.
23. Isaacs, J. I., Isaacs, W. B., Feitz, W. F. J., and Scheres, J. Establishmentand characterization ofseven Dunning rat prostatic cancer cell lines and theiruse in developing methods for predicting metastatic abilities of prostatic can-cers. Prostate, 9: 261-281, 1986.
24. Chirgwin, J. J., Przybyla, A. E., MacDonald, R. I., and Rutter, W. J. Iso-lation of biologically active ribonucleic acid from sources enriched in ribo-nuclease. Biochemistry, 18: 5294-5299, 1979.
25. Berger, S. L. Isolation ofcytoplasmic RNA: ribonucleoside-vanadyl com-plexes. Methods Enzymol., 152: 227-233, 1987.
26. Ono, Y., Fujii, T., Igarashi, K., Kikkawa, U., Ogita, K., and Nishizuka, Y.Nucleotide sequences of cDNAs for a and y subspecies of rat brain proteinkinase C. Nucleic Acids Res., 16: 5199-5200, 1988.
27. Ono, Y., Kurokawa, I., Fujii, I., Kawahara, K., Igarashi, K., Kikkawa, U.,Ogita, K., and Nishizuka, Y. Two types of complementary DNAs of rat brain
protein kinase C. FEBS Left., 206: 347-352, 1986.
28. Kawasaki, E. S. Amplification of RNA. In: M. A. Innis, D. H. Gelfand,J. J. Sninsky, and I. J. White (eds.), PCR Protocols: A Guide to Methods and
Applications, pp. 21-27. San Diego: Academic Press, 1990.
29. Sanger, F., Nicklen, S., and Coulson, A. R. DNA sequencing with chain-terminating inhibitors. Proc. NatI. Acad. Sci. USA, 74: 5463-5467, 1977.
30. Frohman, M. A., Dush, M. K., and Martin, G. R. Rapid production offull-length cDNAs from rare transcripts: amplification using a single gene-specific oligonucleotide primer. Proc. NatI. Acad. Sci. USA, 85:8998-9002,1988.
31 . Saiki, R. K. Amplification ofgenomic DNA. In:M. A. Innis, D. H. Gelfand,J, J. Sninsky, and I. I. White (eds.), PCR Protocols: A Guide to Methods andApplications, pp. 1 3-20. San Diego: Academic Press, 1990.


















