





Novel Treatment Strategies for Unconjugated Hyperbilirubinemia

Cellular uptake of unconjugated TAT peptide involves clathrin-
dependent endocytosis and heparan sulfate receptors.
Jean Philippe Richard1,3, Kamran Melikov2,3, Hilary Brooks1, Paul Prevot1,
Bernard Lebleu1 and Leonid V. Chernomordik2
1UMR 5124 CNRS, Université Montpellier 2, place Eugène Bataillon, 34095 Montpellier
cedex 5, France; 2Section on Membrane Biology, Laboratory of Cellular and Molecular
Biophysics, NICHD, National Institutes of Health, Bldg.10/Rm.10D05, 10 Center Drive,
Bethesda, MD 20892-1855;
3These authors contributed equally to this work.
Running title: Endocytic pathway and receptors in TAT uptake Address correspondence to: Kamran Melikov, Ph.D. LCMB, NICHD, NIH, 10 Center Dr., Bldg. 10, Rm. 10D05, Bethesda, Maryland 20892, USA TEL: (301) 4029010 FAX: (301) 4802412 Email: [email protected]
JBC Papers in Press. Published on February 1, 2005 as Manuscript M401604200 by guest on February 2, 2019
http://ww
w.jbc.org/
Dow
nloaded from

SUMMARY
Delivery of macromolecules mediated by protein transduction domains (PTDs)
attracts a lot of interest due to its therapeutic and biotechnological potential. A major
reevaluation of the mechanism of PTD-mediated internalization and the role of
endocytosis in this mechanism has been recently initiated. Here we demonstrate that
entry of TAT peptide (one of the most widely used PTDs) into different primary cells is
ATP- and temperature- dependent indicating the involvement of endocytosis. Specific
inhibitors of clathrin-dependent endocytosis partially inhibit TAT peptide uptake
implicating this pathway in TAT peptide entry. In contrast, the caveolin-dependent
pathway is not essential for the uptake of unconjugated TAT peptide as evidenced by
the efficient internalization of TAT in the presence of the known inhibitors of
raft/caveolin-dependent pathway and for cells lacking or deficient in caveolin-1
expression. While a significant part of TAT peptide uptake involves heparan sulfate
receptors, efficient internalization of peptide is observed even in their absence,
indicating involvement of other receptors. Our results suggest that unconjugated
peptide might follow endocytic pathways different from those utilized by TAT peptide
conjugated to different proteins.
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

INTRODUCTION
Recent advances in identification of new molecular therapy targets and disease-
relevant proteins, accelerated by the completion of the human genome project,
emphasized an importance of high molecular weight information-rich biomolecules,
such as peptides, proteins, antisense DNA and small interfering RNA, for molecular
therapy. However, delivery of proteins and nucleic acids into cells is greatly hampered
by the low permeability of the cell plasma membrane to polar molecules. Not
surprisingly, the discovery that a number of cationic peptides, known as “protein
transduction domains” (PTDs) can facilitate cytoplasmic and nuclear delivery of a
conjugated cargo has attracted a lot of interest (1-3). Up to date a wide range of cargo
molecules, including low molecular weight drugs (4), oligonucleotides (5), peptides (6)
and even full-length proteins (7-10) have been successfully delivered into cells using
PTD’s and, most importantly, functional activity of the delivered cargo has been
observed (7-10).
Despite significant progress in the cytoplasmic and nuclear delivery of various
cargo molecules using PTD’s, the underlying mechanisms remain under active debate.
Until recently it was widely assumed that the internalization of cationic PTD’s is an
energy- and receptor- independent process based on direct transport through the lipid
bilayer (11-15). On the other hand, there have been indications that uptake of full length
TAT protein, from which one of the most commonly used PTD’s referred to as TAT
peptide is derived, occurs via endocytosis and depends on cell surface heparan sulfate
receptors (16). Moreover the validity of some of the important data, supporting a direct
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

transport model for synthetic TAT peptide, has been questioned in several recent
studies (17-19). It has been shown that apparent ATP and temperature independence of
the cellular uptake of the TAT peptide, as well as its fast nuclear accumulation, results
from experimental artifacts due to cell fixation and incomplete removal of cell bound
peptide (18,19). Recently the uptake of TAT and (Arg)9 peptides has been attributed to a
vesiculation process inhibited by cellular ATP depletion and by low temperature (19).
While several very recent papers provide additional evidence for the involvement of
endocytic pathways in the transduction of PTD’s alone as well as PTD’s conjugated to a
cargo (20-23), the alternative mechanism of direct translocation has been also argued for
in the recent literature (24,25).
In the present work we further clarify the mechanisms by which TAT peptide
enters living cells. As in the case of the stable cell lines used in our earlier work, TAT
entry into several different primary cells is found to be ATP- and temperature-
dependent indicating the involvement of endocytosis. Judging from the effects of
specific inhibitors, unconjugated TAT peptide enters cells mainly by a clathrin-
dependent endocytic pathway, while raft/caveolin-dependent pathway is not required
for internalization. While a significant part of TAT peptide uptake involves heparan
sulfate receptors, these receptors are not a pre-requisite for TAT entry.
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

EXPERIMENTAL PROCEDURES
Peptide Synthesis and Labeling. Synthesis of TAT peptide with sequence Gly-
Arg-Lys-Lys-Arg-Arg-Gln-Arg-Arg-Arg-Pro-Pro was carried out by solid phase on a
Pioneer peptide synthesizer (Applied Biosystems) following the Fmoc chemistry
protocol. A cysteine was added to the C-terminal end of the peptide to provide a
sulfhydryl group for ligation to a fluorochrome. Peptides were purified by preparative
HPLC and characterized by analytical HPLC and matrix-assisted laser desorption
ionization time-of-flight (MALDI-TOF) analysis (data not shown). Labelling with the
fluorochromes was performed by conjugation with a 10 molar excess of the
fluorochrome-maleimide derivatives (Molecular Probes) in 50 mM Tris-HCl buffer pH
7.2 for 4 h in the dark. Labelled peptides were purified by semi-preparative HPLC,
freeze-dried, and resuspended in deionized water. Peptides were stored frozen at -20°C
until further use.
Cells and Cell Cultures. HeLa and HepG2 cells were cultured as exponentially
growing subconfluent monolayers on 90-mm plates in RPMI 1640 medium (Invitrogen)
supplemented with 10% (v/v) fetal calf serum and 2mM glutamine. Wild type CHO
cells (CHO-K1 cell line) and mutants lacking all glycosaminoglycans (pgsA-745 cell line)
or heparan sulfate (pgsD-677 cell line) were cultured in 75cm2 flasks in Vitacell Ham's
F12K medium (ATCC) supplemented with 10% (v/v) fetal bovine serum and 2mM
glutamine. HUVEC (primary human umbilical vein endothelial cells) cells were
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

cultured as exponentially growing subconfluent monolayers on 90-mm plates in
endothelial cell basal medium (EBM) (Clonetics) supplemented with 2% (v/v) fetal calf
serum and 2 mM glutamine, Hydrocortisone 1mg/ml, Gentamicin 50µg/ml,
Amphotericin B 50pg/ml, Bovine brain extract 0.75µg/ml and recombinant human
epidermal growth factor 10µg/ml. Peripheral blood, obtained from healthy donors, was
collected in heparinized tubes. The peripheral blood mononuclear cells (PBMC) were
separated by Ficoll-Hypaque (Sigma) as described previously (26). Flow cytometry. In
the case of HeLa, HepG2 and human macrophages exponentially growing cells were
dissociated with a nonenzymatic cell dissociation medium (Sigma). 5x105 cells were
plated and cultured overnight on 30-mm dishes. The culture medium was discarded,
and cells were washed with NaCl/Pi (pH 7.3). NaCl/Pi was discarded, and cell
monolayers were preincubated with Opti-MEM during 30 min. Subsequently, the cell
monolayers were incubated, as described below, with peptides and drugs dissolved in
Opti-MEM. Following incubation, cells were washed with NaCl/Pi and incubated 10
min with 0.1% trypsin (except HUVEC which were treated with 0.05% trypsin) to
detach them and to remove surface- bound material. In some experiments membrane-
bound peptides were removed by 15 min incubation at 4°C with 0.1% pronase and
1mM EDTA. After the incubation, one volume of serum was added to stop the trypsin
(or pronase) treatment and 10 volumes of NaCl/Pi were added to detach completely the
cells. The cell suspension was centrifuged at 800g, washed with NaCl/Pi, centrifuged
again, and resuspended in 500µl of NaCl/Pi.
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

In case of wild type and mutant CHO cells, exponentially growing cells were
dissociated with a nonenzymatic cell dissociation medium and resuspended in serum-
free Ham’s F12K medium. Subsequently, 5x105 cells were incubated, as described in the
figure legends, with 200µl of TAT peptide dissolved in F12K medium. Following
incubation, cell suspension was centrifuged at 800g, washed twice with 2ml of cold
NaCl/Pi and incubated 10 min with 200µl of 0.1% trypsin to remove surface-bound
peptide. Finally cells were washed once with 2ml F12K medium supplemented with
10% fetal calf serum and once with 2ml of cold NaCl/Pi and resuspended in 500µl of
NaCl/Pi, containing 10µM propidium iodide (Molecular Probes) to exclude dead cells.
Fluorescence analysis was performed with a FACScalibur fluorescence-activated
cell sorter (BD Biosciences). A minimum of 30,000 events per sample was analyzed,
except for HUVEC cells for which 10,000 events were analyzed. To quantify effects of
various treatments on cellular uptake, median of cell fluorescence distribution in
experiment was normalized to the cell fluorescence distribution median in untreated
control. Each experiment was performed at least twice in duplicate or triplicate. Mean
and S.D. of total number of analyzed samples (from 3 to 14) are indicated on Figures.
Significance of the effects of various treatments as compared to untreated control was
evaluated by paired t-test at the 95% confidence level.
RESULTS
Uptake of TAT peptide by primary cells is ATP- and temperature- dependent.
While established cell lines are convenient and useful experimental models, many
important biotechnological and biomedical applications involve primary cells. We
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

studied the effects of ATP depletion and low temperature on TAT peptide uptake by
human PBMC macrophages and HUVEC endothelial cells. The primary cells were
incubated with AlexaFluor488-tagged TAT peptide for 30 min and the amount of
internalized peptide was evaluated using FACS. As shown on Fig. 1, TAT uptake was
greatly inhibited at 4°C in both tested cell types. When cells were pre-incubated with
sodium azide and deoxyglucose to deplete the cellular ATP pool, a smaller but still
significant inhibition of TAT uptake was observed (Fig. 1). Temperature- and ATP-
dependence of TAT uptake by the primary cells suggest that entry by endocytosis is a
general mechanism of TAT internalization valid for primary cells rather than a
mechanism characteristic only for established cell lines.
Heparan sulfate receptors involved in TAT peptide internalization. Earlier work
indicated that the uptake of either full length TAT protein (16) or TAT peptide
conjugated to streptavidin (20) depends on the presence of heparan sulfate receptors. To
evaluate the role of heparan sulfates on the uptake of non-conjugated TAT peptide,
wild type CHO cells (CHO-K1 cell line) or mutant cells lacking surface heparan sulfate
(pgs-A745 and pgs-D677 cell lines) were incubated with 10µM fluorescein labeled TAT
peptide for 30 min. Peptide uptake was significantly inhibited in mutant cells lacking
surface heparan sulfates compared to wild type CHO cells (Fig 2A,B). However a
significant amount of peptide was internalized even in the absence of heparan sulfate
receptors (Fig. 2A). In control experiments no decrease in uptake of the lypophilic dye
FM 4-64, a heparan sulfate independent marker of endocytosis, was observed in both
mutant cell lines in comparison with wild type cells (Fig. 2B). The involvement of
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

heparan sulfate receptors in TAT peptide was confirmed with another experimental
approach. The pre-treatment of cells with heparinase III inhibited TAT uptake in wild
type cells, but not in heparan sulfate receptor-deficient cells (Fig. 2C,D). While these
data indicate the importance of heparan sulfate receptors for the uptake of TAT peptide
in wild type CHO cells, Fig. 2 also shows TAT internalization in the absence of heparan
sulfate proteoglycans, indicating the existence of heparan sulfate independent
mechanisms of entry.
TAT peptide uptake involves caveolin-independent pathways of endocytosis.
The uptake of TAT peptide fused to proteins has been recently attributed specifically to
caveolin-dependent pathway (21,22). In contrast, in our earlier work (19) efficient
uptake of unconjugated TAT peptide was observed in a number of cell lines, deficient in
caveolin-1 expression, including Jurkat T cell line and HeLa cells (27). Likewise, we
observed an ATP- and temperature- sensitive uptake of TAT in HepG2 hepatoma cells
(data not shown), which have been reported to lack caveolin-1 (28). To specifically test
whether internalization of free TAT peptide involves caveolin-dependent endocytosis
we explored the effects of nystatin and filipin III – both known inhibitors of caveolin
dependent uptake (29). Both inhibitors had little effect on the uptake of
AlexaFluor488-tagged TAT peptide in HeLa cells (data not shown). That is not
surprising considering the low level of caveolin-1 expression in these cells and the
inefficient internalization of a well-established marker of raft/caveolin-dependent
endocytic pathway, BODIPY-lactosylceramide (BODIPY-LacCer) (27). Importantly, in
CHO cells, which efficiently express caveolin-1 and actively internalize BODIPY-LacCer
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

(27), uptake of AlexaFluor488-tagged TAT peptide incubation also was not significantly
affected in the presence of 50 µg/ml nystatin (Fig. 3A, D), while in the same conditions
uptake of BODIPY-LacCer was inhibited by ~30% (Fig. 3B, D). As expected, nystatin
had no effect on uptake of Fluorescein-tagged transferrin (Fig. 3C, D). Taken together
these results indicate that cellular uptake of unconjugated TAT peptide does not require
raft/caveolin-dependent endocytic pathway.
Cellular uptake of TAT peptide involves clathrin-dependent endocytosis. To test
whether TAT uptake involves clathrin coated pit mediated endocytosis, TAT uptake
was measured in the presence of known inhibitors of this endocytic pathway,
potassium depletion and chlorpromazine. Treatment of HeLa cells with chlorpromazine
(30-min preincubation in the presence of 30µM chlorpromazine followed by 30-min
incubation with both 30µM chlorpromazine and 1µM AlexaFluor488-tagged TAT
peptide) resulted in 50% inhibition of peptide uptake (Fig. 4A, C). Similarly, 30-min
preincubation of HeLa cells in potassium-free buffer followed by 5-min hypotonic
shock and 30-min incubation with 1µM AlexaFluor488-tagged TAT peptide in
potassium free buffer resulted in 40% decrease in peptide uptake (Fig. 4B, C). In control
experiments significant inhibition was observed for the uptake of fluorescently labeled
transferrin, a well-established marker of clathrin-dependent endocytosis (Fig. 4C).
Inhibition of the uptake of AlexaFluor488-tagged TAT peptide and fluorescein-tagged
transferrin in potassium-free buffer was completely reversible upon replenishment of
potassium in the media (Fig. 4B, C). These data indicate that in the absence of active
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

caveolin-dependent endocytosis TAT peptide is efficiently internalized by clathrin-
dependent endocytic pathway.
To test involvement of clathrin-dependent pathway in cells efficiently expressing
caveolin-1 we explored the effect of potassium depletion on the TAT peptide uptake in
CHO cells, which efficiently express caveolin-1 (27). Cellular internalization of
AlexaFluor488-tagged TAT peptide (Fig. 5A, C) was inhibited by ~55% in potassium-
free buffer as compared to ~85% inhibition of Fluorescein-tagged transferrin uptake
(Fig. 5B, C). Again, as with HeLa cells inhibition for both AlexaFluor488-tagged TAT
peptide and fluorescein-tagged transferrin was completely reversible upon
replenishment of potassium in the media (Fig. 5). These data indicate that even in cells
efficiently expressing caveolin-1 uptake of unconjugated TAT peptide involves clathrin-
dependent pathway of endocytosis.
Internalized TAT peptide is targeted to acidic compartments. As endocytosed
proteins pass from early to late endosomes and to lysosomes, the vesicular
compartments undergo progressive acidification. Even early endosomes show a
reduction in pH (5.9-6.0) to promote the release of ligands bound to receptors.
Acidification of these luminal compartments is blocked by agents such as monensin
(30). To test whether endocytosed TAT peptide is targeted into acidified compartments,
we took advantage of the fact that the fluorescence of fluorescein significantly decreases
at pH=5-6 typical for endosomes and lysosomes as compared to fluorescence at pH=7.4
of cell culture media. We incubated HeLa cells with 2µM fluorescein-tagged TAT for 60
min followed by 30 min incubation in the presence or in the absence of 50µM monensin
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

at 4°C. As shown on Fig 6B,C, incubation in the presence of monensin resulted in a
significant increase in the cell-associated fluorescence. To verify that the higher
fluorescent signal was due to a pH-dependent increase in the fluorescence of TAT-
fluorescein within acidified compartments; the fluorescein tag was replaced with an
Alexa Fluor488 probe, which does not alter with pH (in the 5-8 range). In contrast to
cells with TAT-fluorescein, fluorescence of cells incubated with TAT-Alexa Fluor488
was not affected by monensin (Fig. 6A,C). These experiments indicate that a significant
fraction of the TAT peptide is delivered into acidic cellular compartments.
DISCUSSION
While the number of publications confirming the great potential of PTD’s for
cellular delivery of many important compounds is increasing each year (1), the
mechanism of PTD-mediated delivery, remains elusive. For about a decade it was
commonly accepted that despite their highly polar nature PTD’s are able to rapidly and
efficiently translocate directly through the lipid bilayer, thus delivering their cargo into
the cytoplasm and nucleus in a receptor- and energy- independent way (14,15). This
model was supported by a large number of indirect evidences, including temperature-,
ATP- and receptor independence of cellular uptake (11-14), but attempts to directly
validate this model on protein-free lipid bilayer systems gave contradictory results
(31,32). Moreover, it was recently shown, that apparent ATP- and temperature-
independence as well as fast kinetics of PTD’s uptake result from artifacts of cell
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

fixation and incomplete removal of surface-bound peptide (18,19). These recent findings
have led to a major reevaluation of key characteristics of PTD’s internalization and the
role of endocytosis in PTD mediated uptake.
Our present results indicate that fluorescently labeled TAT peptide, as full-length
TAT protein (16) or TAT-streptavidin conjugate (20), uses heparan sulfate receptors for
internalization. However, the uptake of small basic peptides such as the TAT PTD
might be also mediated by other pathways as suggested by the fact that the
internalization of TAT peptide is not completely inhibited in cells lacking surface
heparan sulfates due to mutations or enzymatic treatment. The observed promiscuity of
TAT peptide in terms of receptor use is not very surprising given the highly cationic
nature of the peptide and the abundance of negatively charged moieties on the cell
surface. Therefore one might expect that in addition to specific interactions with
heparan sulfate proteoglycans there may well be a high level of electrostatic interactions
with other negatively charged cell surface determinants. The importance of cell surface
receptors for TAT uptake, mentioned above, as well as ATP- and temperature-
dependence, as shown in this and earlier works (18,19), demonstrate that TAT peptide
uptake involves endocytosis. In some of the recent reports arguing for the crucial role of
endocytosis in the uptake of constructs in which the TAT peptide was fused to proteins,
the uptake has been attributed specifically to caveolae-dependent pathway (21,22). On
the other hand, we found cells deficient in caveolin-1 expression or lacking caveolin-1
cells to be able to efficiently internalize unconjugated TAT peptide. This indicates that
caveolin-dependent pathway is not required for TAT peptide uptake and alternative
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

pathways might be utilized for uptake of TAT in those cells. We therefore tested the
effects of various inhibitors of clathrin- and caveolae- dependent endocytosis on the
uptake of unconjugated TAT peptide. For both caveolin-1 expressing and caveolin-1
deficient cells inhibitors of clathrin-dependent pathway (potassium depletion and
chlorpromazine) significantly decrease TAT peptide uptake, while inhibitors of
raft/caveolin-dependent pathway (nystatin and filippin III) have no significant effect on
the uptake. These findings support an involvement of endocytosis in the TAT
internalization and show that clathrin-dependent pathway is involved in cellular
uptake of unconjugated TAT, while raft/caveolin-dependent pathway does not play a
significant role. Along the same lines, the full-size TAT protein is internalized by a
clathrin-dependent pathway (33). In contrast to our results, recent reports on the
cellular uptake of TAT peptide conjugated to proteins in HeLa and CHO cells have
attributed uptake of these conjugates to caveolin-dependent pathway (21,22). This
apparent discrepancy suggests that free TAT peptide and TAT peptide conjugated to
macromolecules utilize different pathways for internalization, possibly due to
difference in receptor usage. Interestingly, recent report on the uptake of TAT
conjugated to Cre recombinase in T cells, which are deficient in caveolin-1, implicates
caveolin-independent lipid raft macropinocytosis in transduction of TAT-Cre
recombinase into cells (34). Very recently lipid raft macropinocytosis also has been
implicated in the uptake of unconjugated TAT peptide (35). However methyl-β-
cyclodextrin, which was used in this work to inhibit raft/caveolin dependent pathway,
has been shown to affect clathrin-dependent pathway (36), thus complicating
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

interpretation of the data. Taken together these data indicate that alternative pathways
could be used for TAT uptake depending on cell type and nature (and presence) of
conjugated cargo. It is possible, that while free fluorescently labeled TAT peptide is
mainly directed into constitutive clathrin-dependent pathway, and TAT peptide
conjugated to macromolecular cargo upon binding to cellular receptors induces
signaling cascades activating caveolin-dependent endocytosis or macropinocytosis.
Whatever the case, our results confirm that endocytosis is a major route of TAT
peptide internalization. It is generally accepted that endocytosed material is targeted to
late endosomes or lysosomes - compartments that contain active proteases and are
involved in the degradation of internalized proteins and other macromolecules. In our
experiments with monensin – an inhibitor of endosome acidification, we found that
within 1h at least a part of the TAT peptide is delivered into acidic compartments.
Trapping and possible degradation of TAT peptides within these acidic vesicles is at
odds with effective delivery of bioactive transported cargo within cytoplasm and/or
nucleus. Nevertheless the delivery of functionally active proteins into the cytoplasm
and the nucleus (7,9,10) has been well documented. This apparent discrepancy could be
explained by different sensitivities of the experimental approaches. While in our
experiments we measure the total amount of internalized peptide, the delivery of only a
negligible fraction of total TAT conjugate to the cytoplasm or to the nucleus might be
sufficient for functional detection of the transduction with very sensitive biological
assays as an end-point. It is possible therefore that only a small fraction of all
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

internalized conjugated cargo molecules is actually delivered into the cytoplasm or
nucleus.
There are several hypothetical mechanisms by which PTDs can facilitate
transduction of macromolecules into the cell. First, it is possible that once inside
endosomes, TAT and other cationic PTD’s destabilize the endosomal membrane to leak
a portion of the enclosed macromolecules into the cytoplasm as reported for
polyethyleneimine (37). TAT-induced destabilization of endosomes might involve TAT
interactions with proteins or lipids specific to endosomal compartments. These
interactions can be promoted by acidic pH inside endosomes. It is also possible that a
functionally relevant fraction of the TAT peptide utilizes the caveolin-dependent
endocytic pathway that bypasses acidic cellular compartments. This pathway is used by
some viruses as recently described for SV 40 (38). Finally, one cannot exclude the
possibility that a small, but nonetheless all-important, fraction of the TAT peptide
directly crosses the lipid bilayer to deliver conjugated cargo into the cytoplasm. This
latter mechanism was additionally supported by the recent study, demonstrating direct
translocation of penetratin (another well studied PTD) through protein-free synthetic
lipid bilayers (39). Translocation of penetratin was shown to be lipid composition
dependent, requiring the presence of anionic lipids and high (>100mV) transmembrane
potentials and was observed in a much slower time frame than previously expected
(several hours rather than minutes) (39). This slow direct translocation of PTD through
membranes might be of importance for the release of PTD from endosomes and
lysosomes into the cytosol.
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

To conclude, our experiments further substantiate the key role of endocytosis in
the PTD entry into living cells and provide new insights into utilized endocytic
pathways and surface receptors. We believe that these results along with future work
involving sensitive functional assays will bring a better understanding of the
mechanisms by which relatively short PTD deliver large cargos to the cytosol and
nucleus.
Acknowledgments
We thank Dr. E. Vives for providing the fluorochrome-tagged TAT peptides used
in this study and for valuable discussions. We also want to thank Dr. S. Kinet for
providing lymphocytes. We would like to thank Dr. D. Marks for his valuable advice on
caveolin-depndent endocytosis and Dr. J.C. Grivel for his kind and expert help in
performing some of the FACS experiments. This work has been supported by grants
from the Association pour la Recherche contre le Cancer (5919) and from the European
Commission (QLK3-CT-2002-01989) to B. Lebleu.
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

REFERENCES
1. Lindsay, M. A. (2002) Curr Opin Pharmacol 2, 587‐594
2. Matsui, H., Tomizawa, K., Lu, Y. F., and Matsushita, M. (2003) Curr Protein Pept
Sci 4, 151‐157
3. Wadia, J. S., and Dowdy, S. F. (2003) Curr Protein Pept Sci 4, 97‐104
4. Rothbard, J. B., Garlington, S., Lin, Q., Kirschberg, T., Kreider, E., McGrane, P. L.,
Wender, P. A., and Khavari, P. A. (2000) Nat Med 6, 1253‐1257
5. Astriab‐Fisher, A., Sergueev, D., Fisher, M., Shaw, B. R., and Juliano, R. L. (2002)
Pharm Res 19, 744‐754
6. Shibagaki, N., and Udey, M. C. (2002) J Immunol 168, 2393‐2401
7. Nagahara, H., Vocero‐Akbani, A. M., Snyder, E. L., Ho, A., Latham, D. G., Lissy,
N. A., Becker‐Hapak, M., Ezhevsky, S. A., and Dowdy, S. F. (1998) Nat Med 4,
1449‐1452
8. Schwarze, S. R., Ho, A., Vocero‐Akbani, A., and Dowdy, S. F. (1999) Science 285,
1569‐1572
9. Joshi, S. K., Hashimoto, K., and Koni, P. A. (2002) Genesis 33, 48‐54
10. Peitz, M., Pfannkuche, K., Rajewsky, K., and Edenhofer, F. (2002) Proc Natl Acad
Sci U S A 99, 4489‐4494
11. Futaki, S., Suzuki, T., Ohashi, W., Yagami, T., Tanaka, S., Ueda, K., and Sugiura,
Y. (2001) J Biol Chem 276, 5836‐5840
12. Derossi, D., Calvet, S., Trembleau, A., Brunissen, A., Chassaing, G., and
Prochiantz, A. (1996) J Biol Chem 271, 18188‐18193
13. Suzuki, T., Futaki, S., Niwa, M., Tanaka, S., Ueda, K., and Sugiura, Y. (2002) J Biol
Chem 277, 2437‐2443
14. Vives, E., Brodin, P., and Lebleu, B. (1997) J Biol Chem 272, 16010‐16017
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

15. Derossi, D., Joliot, A. H., Chassaing, G., and Prochiantz, A. (1994) J Biol Chem 269,
10444‐10450
16. Tyagi, M., Rusnati, M., Presta, M., and Giacca, M. (2001) J Biol Chem 276, 3254‐
3261
17. Lundberg, M., and Johansson, M. (2002) Biochem Biophys Res Commun 291, 367‐
371
18. Lundberg, M., Wikstrom, S., and Johansson, M. (2003) Mol Ther 8, 143‐150
19. Richard, J. P., Melikov, K., Vives, E., Ramos, C., Verbeure, B., Gait, M. J.,
Chernomordik, L. V., and Lebleu, B. (2003) J Biol Chem 278, 585‐590
20. Console, S., Marty, C., Garcia‐Echeverria, C., Schwendener, R., and Ballmer‐
Hofer, K. (2003) J Biol Chem 278, 35109‐35114
21. Ferrari, A., Pellegrini, V., Arcangeli, C., Fittipaldi, A., Giacca, M., and Beltram, F.
(2003) Mol Ther 8, 284‐294
22. Fittipaldi, A., Ferrari, A., Zoppe, M., Arcangeli, C., Pellegrini, V., Beltram, F., and
Giacca, M. (2003) J Biol Chem 278, 34141‐34149
23. Drin, G., Cottin, S., Blanc, E., Rees, A. R., and Temsamani, J. (2003) J Biol Chem
278, 31192‐31201
24. Sakai, N., and Matile, S. (2003) J Am Chem Soc 125, 14348‐14356
25. Hariton‐Gazal, E., Rosenbluh, J., Graessmann, A., Gilon, C., and Loyter, A. (2003)
J Cell Sci 116, 4577‐4586
26. Dardalhon, V., Jaleco, S., Kinet, S., Herpers, B., Steinberg, M., Ferrand, C., Froger,
D., Leveau, C., Tiberghien, P., Charneau, P., Noraz, N., and Taylor, N. (2001) Proc
Natl Acad Sci U S A 98, 9277‐9282
27. Singh, R. D., Puri, V., Valiyaveettil, J. T., Marks, D. L., Bittman, R., and Pagano, R.
E. (2003) Mol Biol Cell 14, 3254‐3265
28. Fujimoto, T., Kogo, H., Nomura, R., and Une, T. (2000) J Cell Sci 113 Pt 19, 3509‐
3517
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

29. Schnitzer, J. E., Oh, P., Pinney, E., and Allard, J. (1994) J Cell Biol 127, 1217‐1232
30. Monsigny, M., Roche, A. C., and Midoux, P. (1984) Biol Cell 51, 187‐196
31. Thoren, P. E., Persson, D., Karlsson, M., and Norden, B. (2000) FEBS Lett 482, 265‐
268
32. Kramer, S. D., and Wunderli‐Allenspach, H. (2003) Biochim Biophys Acta 1609,
161‐169
33. Vendeville, A., Rayne, F., Bonhoure, A., Bettache, N., Montcourrier, P., and
Beaumelle, B. (2004) Mol Biol Cell 15, 2347‐2360
34. Wadia, J. S., Stan, R. V., and Dowdy, S. F. (2004) Nat Med 10, 310‐315
35. Kaplan, I. M., Wadia, J. S., and Dowdy, S. F. (2004) J Control Release in press
36. Rodal, S. K., Skretting, G., Garred, O., Vilhardt, F., van Deurs, B., and Sandvig, K.
(1999) Mol Biol Cell 10, 961‐974
37. Boussif, O., Lezoualcʹh, F., Zanta, M. A., Mergny, M. D., Scherman, D.,
Demeneix, B., and Behr, J. P. (1995) Proc Natl Acad Sci U S A 92, 7297‐7301
38. Pelkmans, L., Kartenbeck, J., and Helenius, A. (2001) Nat Cell Biol 3, 473‐483
39. Terrone, D., Sang, S. L., Roudaia, L., and Silvius, J. R. (2003) Biochemistry 42,
13787‐13799
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

FIGURE LEGENDS
Fig. 1. Cellular uptake of TAT is inhibited at 4 °C and by depletion of cellular
ATP in primary cells. Panel A, HUVEC cells were incubated for 30 min in the presence
of 500nM AlexaFluor488-tagged TAT at 37 °C (red curve) or at 4 °C (green curve) or after
depletion of the cellular ATP pool (blue curve). The black curve corresponds to cells
incubated in the absence of TAT. Panel B, Effect of 4 °C (red) or depletion of the cellular
ATP pool (green) on the AlexaFluor488-tagged TAT internalization in HUVEC and
human macrophages. Uptake is expressed as median of cell fluorescence distribution in
experiment normalized to the cell fluorescence distribution median in untreated control
(internalization at 37 °C). Mean and S.D. are indicated. Values significantly (p<0.05)
different from control (internalization at 37 °C) are marked with asterisk.
For depletion of cellular ATP pool cells were preincubated 1h with 10 mM Na azide and
6 mM 2-deoxy-D-glucose.
Fig. 2. Involvement of heparan sulfate receptors in the cellular uptake of TAT
peptide. Panel A, CHO-K1 (red curve), pgsA-745 (green curve) or pgsD-677 (blue curve)
cells were incubated for 30 min in the presence of 10µM FITC-tagged TAT at 37 °C.
Panel B, CHO-K1 (red), pgsA-745 (green) or pgsD-677 (blue) cells were incubated with
10µM FITC-tagged TAT or 10µM FM4-64 for 30 min. Uptake is expressed as median of
cell fluorescence distribution in experiment normalized to the cell fluorescence
distribution median in CHO-K1 cells. Mean and S.D. are indicated. Values significantly
(p<0.05) different from control (uptake in CHO-K1 cells) are marked with asterisk.
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Panel C, CHO-K1 were preincubated for 40 min at 37 °C in Na/Pi containing 0.1%
bovine serum albumine, 0.2% gelatin, 0.1% glucose with (in blue) or without (in red) 10
mlU/ml of heparinase III. Subsequently cells were incubated for 30 min in the presence
of 10µM FITC-tagged TAT at 37 °C. Panel D, Effect of heparinase treatment on the
uptake of FITC-tagged TAT (in blue) in CHO-K1 and pgsA-745 cells. Untreated control
is shown in red. Uptake is expressed as median of cell fluorescence distribution in
experiment normalized to the cell fluorescence distribution median in untreated
control. Mean and S.D. are indicated. Values significantly (p<0.05) different from
untreated control are marked with asterisk.
Fig. 3. Effects of inhibitors of caveolin dependent endocytosis. Panel A, CHO
cells were incubated for 30 min in the presence or in the absence of 50µg/ml nystatin at
10°C in a water bath. Then, they were incubated for 10 min at 37°C in the presence
(redcurves) or in the absence (bluecurves) of 50µg/ml nystatin and 500nM AlexaFluor488-
tagged TAT. The black curve corresponds to cells incubated in the absence of TAT.
Membrane-bound peptide was removed by a pronase treatment before FACS analysis
Panel B, CHO cells were incubated with 500nM BODIPY-LacCer, in presence or absence
of nystatin using the same protocol as in Panel A. Membrane-bound material was
removed according to the protocol described in (27). Panel C, CHO cells were incubated
with 25µg/ml fluorescein-labelled Transferrin and washed as described in panel A.
Panel D, Effect of nystatin treatment on the internalization of AlexaFluor488-tagged
TAT (in blue), BODIPY-LacCer (in green) and fluorescein-tagged Transferrin (in red).
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Uptake is expressed as median of cell fluorescence distribution in experiment
normalized to the cell fluorescence distribution median in untreated control. Mean and
S.D. are indicated. Values significantly (p<0.05) different from untreated control are
marked with asterisk.
Fig. 4. Effect of inhibitors of clathrin dependent endocytosis on HeLa cells.
Panel A, HeLa cells were preincubated for 30 min in the presence or in the absence of
30µM chlorpromazine. Then, they were incubated for 30 min in the presence (blue
curves) or in the absence (red curves) of 30µM chlorpromazine and 1 µM AlexaFluor488-
tagged TAT. The black curves correspond to cells incubated in the absence of TAT. Panel
B, HeLa cells were preincubated for 60 min in regular buffer or in K+ free-buffer (140
mM NaCl, 20 mM HEPES, 1mM CaCl2, 1mM MgCl2, 1mg/ml D-glucose, pH 7.4). Then,
they were incubated 5 min in a hypotonic buffer (K+ free-buffer diluted 1:1 with
distilled water) in order to induce a hypotonic shock, and finally incubated in K+ free-
buffer (red curves) or in regular buffer (blue curves) in the presence of 500nM
AlexaFluor488 tagged TAT for 60 min. The final incubation step was done in regular
buffer (green curves) when testing for reversibility. The black curve corresponds to cells
incubated in the absence of TAT. Panel C, Effect of chlorpromazine (in blue), K+-
depletion (in green) and K+-replenishment (in red) on the internalization of
AlexaFluor488-tagged TAT and fluorescein-tagged Transferrin. Uptake is expressed as
median of cell fluorescence distribution after treatment normalized to the cell
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

fluorescence distribution median in untreated control. Mean and S.D. are indicated.
Values significantly (p<0.05) different from untreated control are marked with asterisk.
Fig. 5. Effect of K+ depletion on clathrin dependent endocytosis in CHO cells.
Panel A, CHO cells were preincubated for 60 min in regular buffer or in K+ free-buffer
(140 mM NaCl, 20 mM HEPES, 1mM CaCl2, 1mM MgCl2, 1mg/ml D-glucose, pH 7.4).
Then, they were incubated 5 min in a hypotonic buffer (K+ free-buffer diluted 1:1 with
distilled water) in order to induce a hypotonic shock, and finally incubated in K+ free-
buffer (red curves) or in regular buffer (blue curves) in the presence of 500nM
AlexaFluor488 tagged TAT for 60 min. When testing for reversibility the final
incubation step was done in regular buffer (green curves).The black curve corresponds to
cells incubated in the absence of TAT. Panel B, Same experiments in CHO cells with
25µg/ml fluorescein-tagged Transferrin. Panel C, Effect of K+-depletion (in blue) and K+-
replenishment (in red) on the internalization of AlexaFluor488-tagged TAT and
fluorescein-tagged Transferrin. Uptake is expressed as median of cell fluorescence
distribution after treatment normalized to the cell fluorescence distribution median in
untreated control. Mean and S.D. are indicated. Values significantly (p<0.05) different
from untreated control are marked with asterisk.
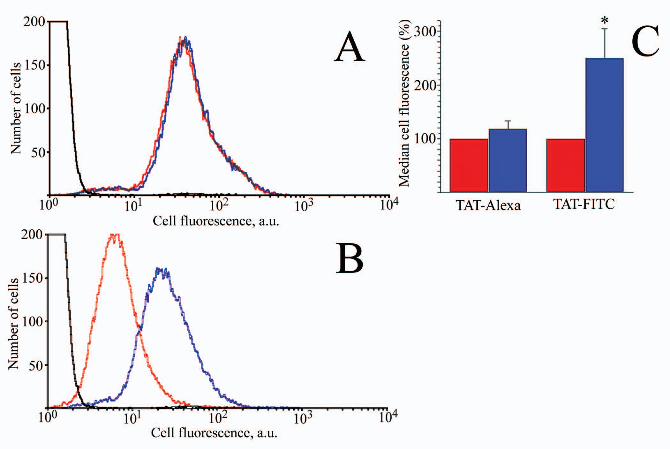
Fig. 6. Monensin treatment. HeLa cells were incubated for 60 min in the
presence of 2µM AlexaFluor488-(Panel A) or Fluorescein (Panel B) tagged TAT peptide.
After incubation cells were washed with NaCl/Pi, and treated with trypsin.
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Subsequently, they were incubated for 30 min in the presence (blue curves) or in the
absence (red curves) of 50µM monensin at 4°C. Following this, they were washed again
with NaCl/Pi, and resuspended in 500µl NaCl/Pi for FACS analysis. The black curve
corresponds to cells incubated in the absence of TAT. Panel C, Effect of monensin
treatment (in blue) on the internalization of AlexaFluor488-tagged TAT and FITC-
tagged TAT. Untreated control is shown in red. Uptake is expressed as median of cell
fluorescence distribution in experiment normalized to the cell fluorescence distribution
median in untreated control. Mean and S.D. are indicated. Values significantly (p<0.05)
different from untreated control are marked with asterisk.
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Leonid V. ChernomordikJean Philippe Richard, Kamran Melikov, Hilary Brooks, Paul Prevot, Bernard Lebleu and
and heparan sulfate receptorsCellular uptake of unconjugated TAT peptide involves clathrin-dependent endocytosis
published online February 1, 2005J. Biol. Chem.
10.1074/jbc.M401604200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on February 2, 2019http://w
ww
.jbc.org/D
ownloaded from


















