A New Species of Agama (Sauria: Agamidae) from Mauritania
9
HERPETOLOGICAL JOURNAL, Vol. 15, pp. 27-35 (2005) A NEW SPECIES OF AGAMA (SAURIA: AGAMIDAE) FROM MAURITANIA JOSÉ M. PADIAL 1,2 1 Museo de Historia Natural Noel Kempff Mercado (Area Zoología), Santa Cruz de la Sierra, Bolivia 2 Department of Biodiversity and Evolutionary Biology, Museo Nacional de Ciencias Naturales – CSIC, C/José Gutierrez Abascal 2, 2800 Madrid, Spain A new agama species of the Agama agama species group, is described from the Adrar Mountains of Mauritania, in the Meridional Sahara. This species is morphologically similar and genetically related to Agama impalearis. It is characterized by small size (snout-vent length of adults: 67.6-74.88 mm); long hind legs; gular region with brown irregular longitudinal lines; 10 preanal pores; fourth finger longer than first; small nuchal crest (composed of six spines) and absence of caudal crest; reddish eyelid in males; smooth head scales; regular keeled and mucronate dorsal scales; ventral scales smaller than dorsals; 55-63 scales around mid body; 10- 12 surpralabials; 9-12 infralabials; 8-9 group of spines between the anterior margin of the ear opening and the shoulder; 12 lamellae under fourth finger; 19-21 lamellae under fourth toe. It is a solitary rock dweller inhabiting extremely dry habitats with scarce vegetation. Key words: Africa, Agama castroviejoi, lizard, mtDNA, Sahara INTRODUCTION Information on north-west African reptiles, from the Mediterranean to the Saharan region, has increased no- tably in the last 20 years (Schleich et al., 1996). Countries like Morocco, Western Sahara and Mali have received special attention (Joger, 1981; Böhme et al., 1996; Joger & Lambert, 1996; Bons & Geniez, 1996; Geniez et al., 2000), and recently also Mauritania (Ineich, 1997; Böhme, 2000; Böhme et al., 2001). As a result, taxonomic work has been carried out on several complex groups and some new species have been de- scribed (Salvador, 1982; Joger, 1984; Mateo et al., 1998; Wilms & Böhme, 2001). Nevertheless, new spe- cies are discovered only occasionally (Joger, 1980; Joger & Lambert, 1996; Geniez & Foucart, 1995; Brown et al., 2002), indicating that we have still not reached a complete knowledge of the reptile diversity in north-western Africa, above all in the Saharan region, where vast areas still remain unexplored. Four Agama species have been cited for Mauritania: Agama agama (Linnaeus, 1758), Agama boueti Chabanaud, 1917, Agama boulengeri Lataste, 1886 and Agama impalearis (Duméril & Duméril, 1851) (Chabanaud, 1917; Dekeyser & Villiers, 1956; Ineich, 1997; Lambert & Mullié, 1998; Böhme et al., 2001). During field-work carried out in Mauritania, only the first three species were found. A. agama is a typical spe- cies from Sahel savannah, but it also inhabits human settlements (Joger, 1979) and is common in Southern Mauritania (pers. obs.). A. boueti is also a typical inhab- itant of the sandy savannahs of southern Mauritania. Although Chabanaud (1917) reported A. impalearis for Mauritania (“Mauritanie Saharienne”), Joger (1979) stated that this species did not occur there and that the nearest record would be that of Seguia el Hamra (West- ern Sahara). Despite A. agama being cited for the Village Chinguetti in the Adrar region (Dekeyser & Villiers, 1956) it was not found by the author. Only A. boulengeri and five specimens of an unidentified Agama species could be collected in this area. These five specimen were discovered on a tableland (“Dahr”) from the Adrar Mountains, in the Sahara Region. They are morphologically similar to A. impalearis, but some morphological differences in scalation count characters where found. Nevertheless, some works have demon- strated great morphological and genetic variation among populations of A. impalearis (Brown & Znari, 1998; Brown et al., 1999; Brown et al., 2002) and have recognized a vicariant linage as a putative new species (Brown et al., 2002). In fact, the diversity of the group is higher than previously supposed. Therefore, a genetic comparison among the aforementioned species was also necessary to clarify the identity of Adrar population. The results indicate the specimens collected in the Adrar Mountains do not correspond to any of the known North African agamid species. The aim of this paper is to de- scribe these specimens and to clarify their possible relationship with A. impalearis. MATERIAL AND METHODS Specimens of the new species are deposited in Museo Nacional de Ciencias Naturales (MNCN), Madrid (Spain). Other specimens examined are listed in Appen- dix 1. Geographical localities were obtained with a Garmin E-Trek GPS receiver. Colour slides of the speci- mens were taken in the field. All measurements of the specimens are in millimetres and were taken with a dig- ital calliper (to the nearest 0.01mm). Specimens were collected the 20 May 2002 during a field-trip to the Adrar plateau of Mauritania. They were sacrificed by injection of a dilution of nicotine, fixed with 10 % for- malin and preserved in 70% ethanol. Tissue samples for Correspondence: J. M. Padial, Department of Biodiversity and Evolutionary Biology, Museo Nacional de Ciencias Naturales – CSIC, C/José Gutierrez Abascal 2, 2800 Madrid, Spain. E-mail: [email protected]
Transcript of A New Species of Agama (Sauria: Agamidae) from Mauritania

HERPETOLOGICAL JOURNAL Vol 15 pp 27-35 (2005)
A NEW SPECIES OF AGAMA (SAURIA AGAMIDAE) FROM MAURITANIA
JOSEacute M PADIAL12
1Museo de Historia Natural Noel Kempff Mercado (Area Zoologiacutea) Santa Cruz de la Sierra Bolivia
2Department of Biodiversity and Evolutionary Biology Museo Nacional de Ciencias Naturales ndash CSIC CJoseacute GutierrezAbascal 2 2800 Madrid Spain
A new agama species of the Agama agama species group is described from the AdrarMountains of Mauritania in the Meridional Sahara This species is morphologically similar andgenetically related to Agama impalearis It is characterized by small size (snout-vent length ofadults 676-7488 mm) long hind legs gular region with brown irregular longitudinal lines 10preanal pores fourth finger longer than first small nuchal crest (composed of six spines) andabsence of caudal crest reddish eyelid in males smooth head scales regular keeled andmucronate dorsal scales ventral scales smaller than dorsals 55-63 scales around mid body 10-12 surpralabials 9-12 infralabials 8-9 group of spines between the anterior margin of the earopening and the shoulder 12 lamellae under fourth finger 19-21 lamellae under fourth toe It isa solitary rock dweller inhabiting extremely dry habitats with scarce vegetation
Key words Africa Agama castroviejoi lizard mtDNA Sahara
INTRODUCTION
Information on north-west African reptiles from theMediterranean to the Saharan region has increased no-tably in the last 20 years (Schleich et al 1996)Countries like Morocco Western Sahara and Mali havereceived special attention (Joger 1981 Boumlhme et al1996 Joger amp Lambert 1996 Bons amp Geniez 1996Geniez et al 2000) and recently also Mauritania(Ineich 1997 Boumlhme 2000 Boumlhme et al 2001) As aresult taxonomic work has been carried out on severalcomplex groups and some new species have been de-scribed (Salvador 1982 Joger 1984 Mateo et al1998 Wilms amp Boumlhme 2001) Nevertheless new spe-cies are discovered only occasionally (Joger 1980Joger amp Lambert 1996 Geniez amp Foucart 1995Brown et al 2002) indicating that we have still notreached a complete knowledge of the reptile diversity innorth-western Africa above all in the Saharan regionwhere vast areas still remain unexplored
Four Agama species have been cited for MauritaniaAgama agama (Linnaeus 1758) Agama bouetiChabanaud 1917 Agama boulengeri Lataste 1886 andAgama impalearis (Dumeacuteril amp Dumeacuteril 1851)(Chabanaud 1917 Dekeyser amp Villiers 1956 Ineich1997 Lambert amp Mullieacute 1998 Boumlhme et al 2001)During field-work carried out in Mauritania only thefirst three species were found A agama is a typical spe-cies from Sahel savannah but it also inhabits humansettlements (Joger 1979) and is common in SouthernMauritania (pers obs) A boueti is also a typical inhab-itant of the sandy savannahs of southern MauritaniaAlthough Chabanaud (1917) reported A impalearis forMauritania (ldquoMauritanie Sahariennerdquo) Joger (1979)stated that this species did not occur there and that the
nearest record would be that of Seguia el Hamra (West-ern Sahara) Despite A agama being cited for theVillage Chinguetti in the Adrar region (Dekeyser ampVilliers 1956) it was not found by the author Only Aboulengeri and five specimens of an unidentifiedAgama species could be collected in this area Thesefive specimen were discovered on a tableland (ldquoDahrrdquo)from the Adrar Mountains in the Sahara Region Theyare morphologically similar to A impalearis but somemorphological differences in scalation count characterswhere found Nevertheless some works have demon-strated great morphological and genetic variationamong populations of A impalearis (Brown amp Znari1998 Brown et al 1999 Brown et al 2002) and haverecognized a vicariant linage as a putative new species(Brown et al 2002) In fact the diversity of the group ishigher than previously supposed Therefore a geneticcomparison among the aforementioned species was alsonecessary to clarify the identity of Adrar populationThe results indicate the specimens collected in the AdrarMountains do not correspond to any of the known NorthAfrican agamid species The aim of this paper is to de-scribe these specimens and to clarify their possiblerelationship with A impalearis
MATERIAL AND METHODS
Specimens of the new species are deposited in MuseoNacional de Ciencias Naturales (MNCN) Madrid(Spain) Other specimens examined are listed in Appen-dix 1 Geographical localities were obtained with aGarmin E-Trek GPS receiver Colour slides of the speci-mens were taken in the field All measurements of thespecimens are in millimetres and were taken with a dig-ital calliper (to the nearest 001mm) Specimens werecollected the 20 May 2002 during a field-trip to theAdrar plateau of Mauritania They were sacrificed byinjection of a dilution of nicotine fixed with 10 for-malin and preserved in 70 ethanol Tissue samples for
Correspondence J M Padial Department of Biodiversityand Evolutionary Biology Museo Nacional de CienciasNaturales ndash CSIC CJoseacute Gutierrez Abascal 2 2800 MadridSpain E-mail jmpadialyahoocom
DNA studies were obtained from four specimens of thenew species (MNCN 41776-79) and preserved in abso-lute ethanol
The following 27 morphological characters were ob-tained from each specimen (1) 16 linear bodydimensions SVL snout-vent length TAL tail lengthTAH tail height (at the base of the tail) TAW tailwidth (at the base of the tail) RTAL relative tail length( of SVL) HL head length HW maximum headwidth HH maximum head height FOOT foot length(from heel to the tip of the longest toe including theclaw) HAND hand length (from wrist to the tip of thelongest finger including the claw) FINGER length ofthe fourth finger (from the joint with the hand to the tipincluding the claw) TOE fourth toe length (from thejoint with the foot to the tip including the claw) TLtibia length FL femur length (shank length) EN eye-nostril distance (from the posterior margin of the nostrilto the anterior corner of the eye) EE distance ear open-ing-eye (from the posterior corner of the eye to theanterior margin of the ear opening) (2) 11 scalationcounts characters VE number of ventral scales (fromthe inguinal region to the level of the axilla) EMBnumber of scales around midbody SL number ofsupralabials scales IL number of infralabial scales SOnumber of supraocular scales RSLE rows of scales be-tween supralabials and eye GNS groups of neck spinesIP number of inguinal pores LFF number of lamellaeunder fourth finger LFT number of lamellae underfourth toe SC number of spines of the nuchal crest
Scalation count characters of males of the two formsof A impalearis were provided by Brown (in litt)Comparisons with A boueti and A boulengeri werebased on data from Joger (1979) and personal observa-tions and on A agama from personal observations
Partial DNA sequences of the 16S rRNA mitochon-drial gene were obtained for four specimens of the newspecies and compared with previously published 16SrRNA sequences of A impalearis from Morocco(Brown et al 2002) Laudakia atricollis from Tanzaniawas used as outgroup (Brown et al 2002)
Accession numbers for the MNCN specimens se-quenced are as follows MNCN 41776=AY522926MNCN 41777= AY522927 MNCN 41778=AY522928 MNCN 41779= AY522929 Total genomicDNA extraction followed standard protocols describedelsewhere (Carranza et al 1999 2000) Primers usedin both amplification and sequencing were L2510 (5rsquo-CGCCTGTTTATCAAAAACAT-3rsquo) and H3062(5rsquo-CCGGTTTGAACTCAGATCA-3rsquo) from Lenk etal (2001)
Mitochondrial sequences were aligned in ClustalX(Thompson et al 1997) with default parameters Onlytwo gaps had to be postulated to align the new specieswith A impalearis In total 464 base pairs of 16S rRNAmitochondrial DNA were included in the phylogeneticanalysis of which 84 were variable and 27 parsimony-informative The results of the alignment are available
from the author upon request The aligned data set wasanalyzed using maximum-parsimony (MP) in PAUP40B10 (Swofford 2002) and include heuristicsearches involving tree bisection and reconnection(TBR) branch swapping with 100 random stepwise ad-ditions of taxa Gaps were included as fifth state Nodalsupport for the MP tree was assessed using bootstrapanalysis (Felsenstein 1985) involving 1000 pseudo-rep-lications
RESULTS
AGAMA CASTROVIEJOI SP NOV FIGS 1-8
Types Holotype MNCN-41779 an adult male fromDahr Chinguettti on the road between Atar and Tidjikja(20deg 26547rsquoN12deg 49407rsquoW) Wilaya of Adrar Col-lected by Joseacute M Padial 20 May 2002
Paratypes MNCN-41778 an adult female MNCN-41780 a subadult female and MNCN-41776 41777 twoyoung females (same data as the holotype)
Etymology This species is a patronym for Dr JavierCastroviejo (Galicia Spain) for his help and encour-agement and in recognition of his great effort in fundingthe study and conservation of biodiversity in Spain Af-rica and Latin America
Diagnosis A species morphologically similar toAgama impalearis with the following combination of
J M PADIAL
FIG 1 Male (holotype MNCN 41779) of Agamacastroviejoi sp nov Picture taken in the Adrar MountainsMauritania
FIG 2 Dorsal view of the male MNCN 41779 (holotype) ofAgama castroviejoi
28
NEW AGAMA SPECIES FROM MAURITANIA
FIG 3 Lateral view of the head of the male MNCN41779(holotype) of Agama castroviejoi
FIG 4 Frontal view of the snout of the male MNCN 41779(holotype) of Agama castroviejoi showing the orientation ofthe nostrils
FIG 5 Ventral view of the gular region of the male MNCN41779 (holotype) of Agama castroviejoi
FIG 6 Dorsal view of an adult female MNCN 41778(paratype) of Agama castroviejoi
FIG 7 Lateral view of the head of an adult female MNCN41778 (paratype) of Agama castroviejoi
FIG 8 Frontal view of the snout of an adult female MNCN41778 (paratype) of Agama castroviejoi showing theorientation of the nostrils
29
J M PADIAL
FIG 9 Strict consensus of 19 equally parsimonious trees showing the phylogenetic relationships between A impalearis and Acastroviejoi Tree-length= 97 steps CI= 0750 and RI= 0922 (both under exclusion of uninformative sites) Bootstrap supportvalues are shown above the nodes Haplotype numbers refer to Brown et al (2002)
characters (see Figs 2-8) long narrow and low headscales on head smooth small nuchal crest nine groupsof spines on each side of the head nostrils orienteddorsolaterally plicate throat with two gular folds andone mite pocket on each side of the neck regular keeledand mucronate scales in dorsum keels converging to-ward midbody less keeled and mucronate dorsal scalesin females relative length of fingers 3gt4gt5=2gt1 rela-tively long hind legs (always reaching the anteriormargin of the ear opening some reaching the eye) baseof the tail weakly compressed tail scales keeled andmucronate absence of caudal crest tail one and 34 totwice as long as SVL gular region with brown irregularlongitudinal lines ten preanal pores live male with red-dish eyelid ventral regions of the body tail andextremities immaculate white dorsal surfaces of femalelight (sandy) brown with dark brown subocular and la-bial bars and some transverse dark brown bands onback males with dorsal surfaces grey-blue with lightbrown spots Distinguished from the north-west Africanspecies (characters of the other species in parentheses)(see also Table 2) from Agama impalearis by relativelength of finger and toes smaller nuchal crest and ab-sence of caudal crest reddish eyelid in males anddifferent colour pattern some scalation count charac-ters (Table 3) and 43 difference in the 16S rRNAmitochondrial region sequenced for this study Thephylogenetic analysis presented in Fig 9 shows that Acastroviejoi and A impalearis form two reciprocally
monophyletic groups supported by relatively highbootstrap values Distinguished from Agama boueti bygular region with brown irregular longitudinal lines (im-maculate) red eyelid in males (brown) 8-9 groups ofneck spines (4-7) 10 preanal pores (12) 12 scales onfourth finger (9-10) 19-21 scales on fourth toe (15-17)different relative length of fingers and toes longer hindlegs Distinguished from Agama agama by very smallersize and very different colour pattern and sexual dimor-phism (metallic blue body and bright orange on heads ofadult males) Distinguished from Agama boulengeri byits smaller size very different colour pattern (speciallysexual dimorphism) fewer number of scales aroundmidbody smooth head scales higher number of groupof spines on the sides of the head and shoulder and by ab-sence of caudal crest
DESCRIPTION OF THE HOLOTYPE
An adult male with head scales smooth occipitalscale big dorsal scales homogeneous large rhomboidalkeeled and mucronate all keels converging towards thevertebral line keels on arms and limbs mucronate throatplicate with two gular folds and one mite pocket on eachside of the neck ventral scales smooth relative fingerlength 3gt4gt5=2gt1 tail twice as long as SVL base of thetail weakly compressed tail scales keeled and mucro-nate nostrils oriented upwards and backwards
Measurements (mm) Snout-vent length 7488 taillength 11670 head length 1984 head width 1599
30
TABLE 1 Linear body measurements (mm) and scalation count characters of the type series of Agama castroviejoi sp nov Seemethodology for abbreviations
Linear body MNCN MNCN MNCN MNCN MNCN Range measurement 41779 41777 41778 41776 41780and dimension (male) (juv female) (female) (juv female) (female)
SVL 7421 5991 7488 5931 6760 5931-7488TAL 11640 1051 - - 9017 9017-1164TAH 881 692 725 675 687 692-881TAW 1146 820 1055 813 984 813-1146RTAL 6375 570 - - 7497 570-7497HL 1984 1716 1965 1591 1840 1591-1984HW 1599 1358 1567 1321 1511 1321-1599HH 969 888 967 867 986 867-969EN 476 30 398 311 435 30-476EE 601 437 480 461 533 437-601HAND 1455 1204 1280 1153 1414 1153-1455FOOT 2364 1969 2104 1882 2261 1882-2364FINGER 677 623 575 519 667 519-677TOE 1202 964 975 920 1174 920-1202TL 2121 2031 2175 1977 2134 1977-2175FL 1936 1758 1986 1544 2029 1544-2029HLSVL 027 029 026 027 027 026-029HBSVL 022 023 021 022 022 021-023HLHB 124 126 125 120 122 120-126TLSVL 029 034 029 033 032 029-034FLSVL 026 029 027 026 030 026-030ENSVL 024 017 020 020 024 017-024EESVL 030 025 024 029 029 024-030TOESVL 016 016 013 016 017 013-017FOOTSVL 032 033 028 032 033 028-033
Scalation countcharacters
VE 63 59 61 58 59 58-63EMB 61 58 55 60 63 55-63SL 10 11 11 11 12 10-12IL 9 11 11 11 12 9-12SO 6 7 6 7 7 6-7RSLE 4 3 4 4 4 3-4GNS 9 8 8 8 9 8-9IP 10 - - - - -LFF 12 12 12 12 12 -LFT 19 19 21 19 20 19-21SC 6 6 7 7 7 6-7
NEW AGAMA SPECIES FROM MAURITANIA
head height 969 hand length 1455 (from posterior endof wrist to top of longest finger) foot length 2364 (fromposterior end of heel to top of longest toe) limb length1936 tibia length 2121 length of fourth finger (to joinwith third) 677 length of fourth toe 1202 eye-nostrillength (from the anterior border of the eye orbit to thenostril) 476 ear-eye length (from the anterior border ofthe tympanum to the posterior border of the eye orbit)601 tail width 1146 tail height (at the base) 881
Pholidosis 63 ventral scales between inguinal foldand the beginning of the arm 61 scales around midbody11 supralabials 9 infralabials 6 rows of supraoculars 4
rows of scales between supralabials and suboculars (in-cluding the subocular row) 9 groups of spines betweenthe anterior margin of the ear opening and the shoulderIn each group the central spines are largest 10 analpores 12 lamellae under fourth finger 19 lamellae un-der fourth toe nuchal crest composed of 6 spines
Colour in life Ventral regions of the body tail andextremities immaculate white dorsal surfaces grey-blue with dark brown subocular and labial bars gularregion with brown irregular longitudinal lines sometransverse dark brown bands on the back a perpendicu-lar thin beige band in the anterior margin of the back
31
J M PADIAL
TABLE 2 Diagnostic characters for Agama castroviejoi sp nov Agama impalearis and A boueti
A castroviejoi A impalearis A boueti
SVL 676-7488 to 120 to 99TL 9017-1164 150 to 160EMB 55-63 50-62Dorsal scales keeled keeled keeledHead scales smooth smooth smoothNuchal crest small strong smallGNS 8-9 3-9 4-8Caudal crest absent present absent
Relative length 3gt4gt5gt2gt1 3gt4gt5gt2gt1 3gt4gt2gt5gt1of fingersRelative length 4gt3gt5gt2gt1 3=4gt5gt2gt1 3gt4gt5gt2gt1of toes
Gular region brown lines brown lines white to yellowSexual dicromatism low low lowSociability solitary solitary solitaryHabitat desert rocky areas semidesert rocky areas sandy savannah
TABLE 3 Some scalation counts characters of male (holotype) of Agama castroviejoi and two geographical forms of Aimpalearis (a) southern form (Tan Tan Morocco) (b) northern form (Chefchauen Morocco) See methodology for abbreviations
SL IL GNS LFF LFT IP
Holotype 10 9 9 12 19 10a (n=6) 11-13 10-15 10-11 14-16 16-18 12-14b (n=4) 11-13 11-13 10-11 15 16-19 12-13
and one in the posterior margin two beige irregular cir-cumferences in the middle of the back
Colour in alcohol Ventral regions of the body tailand extremities immaculate white gular region withbrown irregular longitudinal lines all dorsal and dorso-lateral regions grey-blue (slate grey) with some lightbrown spot on the head back and forearms hands yel-low a perpendicular thin beige band in the anteriormargin of the back and one in the posterior margin twobeige irregular circumferences in the middle of the back
VARIATION
For variation in morphometrics see Table 1 The onlymales has more ventral scales and fewer labial scalesthan females The dorsal scales of the male are moremucronate than in females There is colour differencebetween sexes females being paler than males In lifefemales have almost no colour differences they havelight brown head with dark brown subocular stripes thedorsal region is beige to light brown with transversedark brown bands from the scapular region to the middleof the tail each dark band is composed of 3-7 scalesthey have a thin longitudinal light stripe The ventral re-gion is immaculate white with subgular longitudinalbrown stripes In alcohol the colour becomes faded butthe pattern is retained the feet and hands become lightyellow
From the genetic point of view there is a single basepair difference (and A-G transition) between specimenMNCN 41778 and specimens MNCN 41776 MNCN41777 MNCN 41779 in the 464 of the 16S rRNA mito-chondrial region sequenced for this study (Fig 9)
DISTRIBUTION AND ECOLOGY
Specimens are only known from the type localityThey were found on a tableland (dahr) 679 m above sealevel in the Adrar region in Mauritania This region ispart of the ldquoSahara Meridional Occidentalrdquo and is char-acterized by annual rainfall of about 100 mm andaverage annual temperatures of 30degC with minimumtemperatures in January rarely descending below 10degC(Le Houeacuterou 1990) The habitat is a rocky plain withlittle vegetation composed of sparse bushes and somesmall trees (Acacia spp) All specimens were foundalone and separate by few kilometres from each otherThe first specimen encountered was active at 0920 hr(air temperature of 286degC) The other specimens werefound perching around the middle of the day and wereeasily caught when they moved under solitary stones
DISCUSSION
Among the four Agama species (A agama Aboulengeri A boueti A impalearis) inhabiting the aridand semi-arid regions of NW Africa the last two show
32
superficial resemblance with A castroviejoi Neverthe-less morphological quantitative and meristic charactersdistinguish A castroviejoi well from the other speciesrecorded for Mauritania (A boulengeri A agama andA boueti) and also from the two Moroccan forms of Aimpalearis MtDNA analyses demonstrate the identityof A castroviejoi and its close relationship to Aimpalearis and the vicariant form from south and east ofthe Atlas mountains
Furthermore distributional social and ecologicalfactors also allow a clear separation among species Acastroviejoi is solitary and inhabits extremely dry rockyareas of the Adrar plateau Agama boueti is solitary andinhabits the semi-desert sandy plains and savannahs ofthe Sahel region of southern Mauritania and the AtlanticCoast Agama impalearis is a typical solitary rockdweller (Schleich et al 1996) Its distribution rangesfrom the Mediterranean coast (Bons amp Geniez 1996) toSeguia el Hamra in Western Sahara (Geniez et al2000) Although it was cited for ldquoMauritanieSahariennerdquo (Chabanaud 1917) and despite appearingwidely distributed across the country in Le Berrersquos(1989) distribution map for this species there are actu-ally no confirmed records for Mauritania MoreoverJoger (1979) noted that this species did not occur in theAdrar and that the nearest record corresponded toSeguia el Hamra The presence of this species in thecountry is still plausible in rocky areas similar to thoseof the Zemmour Mountains (Geniez et al 2000) butthis requires confirmation A boulengeri is a speciesfrom the Sahel Savannah that inhabits rocky areas andforms social groups We found this species as far northas Oued Choucircm (21deg 226N12deg 586W) near the bor-der with Western Sahara At this latitude it is alwaysassociated to wet rocky gorges (Gueltas) in the moun-tains A agama is another social species reported to beassociated with human settlements in the Adrar region(Dekeyser amp Villiers 1956) In the south of the countrythis species prefers trees from the Sahel savannahs but itshows high habitat plasticity (pers obs)
Miocene vicariance events have been found to be re-sponsible of the variation and differentiation in Aimpalearis in NW Africa (Brown amp Znari 1998 Brownet al 2002) The uplift of the Atlas mountains consti-tuted a great geographical barrier causing allopatricfragmentation of the populations A castroviejoi is iso-lated in a rocky tableland surrounded by lowland sandyplains of the Sahara that could serve as a barrier for arock dweller Also in situ selection-mediated responsesto the ecological current conditions of the isolatedpopulations are responsible of the morphological varia-tion of A impalearis (Brown amp Znari 1998) Presentecological conditions of the Adrar are very extreme andis probably one of the hottest areas of the Sahara (LeHoueacuterou 1990) As demonstrated by Brown amp Znari(1998) variation in scalation is significantly associatedwith different temperature conditions Therefore bothisolation and ecological differentiation could be impli-cated in the differentiation of A castroviejoi
Although the Adrar belongs to the Meridional Sahara(Le Houeacuterou 1990) this area shows a complex historywith its faunas and floras comprising a mixture of Medi-terranean Saharan and Sahelian species (Dekeyser ampVilliers 1956) Probably A castroviejoi constitutes anexample of a Saharan endemic of the Adrar Mountainsas some invertebrate species and also vertebrate subspe-cies (Dekeyser amp Villiers 1956) Nevertheless thephylogeographic affinities of A castroviejoi remain un-clear since it could have a Sahelian or Saharan origin Amore thorough phylogenetic comparison among NorthAfrican agamas will help to resolve this question
ACKNOWLEDGEMENTS
This work could not have been carried out withoutthe effort of J Castroviejo and the people of AsociacioacutenAmigos de Dontildeana I am very gratefully to S Carranza(BMNH) who did the genetic work and helped me toimprove the manuscript To I de la Riva (MNCN) forhis help in many aspects of this work To Dr NAnderson (BMNH) who revised the draft version andthe English of the manuscript To R Garciacutea M UrceraC Carballo M O Deida and E Mohamed O Saleh fortheir help and companion in Mauritania and J Peacuterez EAvila S Castroviejo-Fisher and A Quintana for theirfriendship and companion in the first trip to MauritaniaP Geniez J A Mateo H Nickel and W Boumlhme allhelped us in some aspect of this work W Wuster andtwo anonymous reviewers critically commented an ear-lier draft of the manuscript R Brown kindly providedraw scalation data for A impalearis from Morocco andcontributed to the improvement of the manuscript
REFERENCES
Boumlhme W (2000) Die wechselvolle Geschichte derSahara Untersuchung von Reptilienzoumlnosen entlangeines westsaharischen Transektes mit einemuumlberraschenden Fund im Suumldosten Mauritaniens Tierund Museum 7 11-21
Boumlhme W Meinig H amp Roumldel M A (1996) Newrecords of amphibians and reptiles from Burkina Fasoand Mali British Herpetological Society Bulletin 56 7-26
Boumlhme W Wilms T Nickel H amp Merz M (2001)Bericht uumlber eine herpetologische Forschungsreise indie westliche Sahara und ihre Randgebiete Zeitschriftdes Koumllner Zoo 44 117-131
Bons J amp Geniez P (1996) Anfibios y reptiles deMarruecos (Incluido Saacutehara Occidental) Atlasbiogeograacutefico Barcelona Asociacioacuten HerpetoloacutegicaEspantildeola
Brown R P amp Znari M (1998) Morphological variationin Agama impalearis from Morocco evidence forhistorical population vicariance and current climaticeffects Ecography 21 605-612
Brown R P Znari M El Mouden E H amp Harris P(1999) Estimating asymptotic size and geographicvariation in Agama impalearis from MoroccoEcography 22 277-284
NEW AGAMA SPECIES FROM MAURITANIA 33
J M PADIAL
Accepted 22304
Brown R P Pestano J amp Suaacuterez N M (2002) TheAtlas mountains as a biogeographical divide in North-West Africa evidence from mtDNA evolution in theAgamid lizard Agama impalearis MolecularPhylogenetics and Evolution 24 324-332
Carranza S Arnold E N Thomas R H Mateo J ampLopez-Jurado L F (1999) Status of the extinct giantlacertid lizard Gallotia simonyi simonyi (ReptiliaLacertidae) assessed using mtDNA sequences frommuseum specimens Herpetological Journal l9 83-86
Carranza S Arnold E N Mateo J amp Lopez-Jurado LF (2000) Long-distance colonization and radiation ingekkonid lizards Tarentola (Reptilia Gekkonidae)revealed by mitochondrial DNA sequencesProceedings of the Royal Society Series B 267 637-649
Chabanaud P (1917) Enumeacuteration des reptiles nonencore eacutetudieacutes de lrsquoAfrique occidentale appartenantaux collections du Museacuteum avec la description desespeacuteces nouvelles Bulletin du Museacuteum NationaldrsquoHistoire Naturelle de Paris Serie Zoologique 2383-105
Dekeyser P L amp Villiers A (1956) Contribution alrsquoeacutetude du peuplement de la Mauritanie Notationseacutecologiques et biogeacuteographiques sur la faune delrsquoAdrar Memoires de lrsquoInstitut Franccedilais de lrsquoAfriqueNoire 44 222 p 25 pl
Felsenstein J (1985) Confidence-limits on phylogenies ndashan approach using the bootstrap Evolution 39 783-791
Geniez Ph amp Foucart A (1995) Un nouvelAcanthodactyle en Algeacuterie Acanthodactylustaghitensis n sp (Reptilia Sauria Lacertidae)Bulletin du Museacuteum National drsquoHistoire NaturelleParis 4deg seacuterie 17 3-9
Geniez P Mateo J A amp Bons J (2000) A checklist ofthe amphibians and reptiles of Western Sahara(Amphibia Reptilia) Herpetozoa 13 149-163
Ineich I (1997) Les amphibiens et reptiles du littoralmauritanien In Environnement et littoralmauritanien 93-99 Colas P (Ed) MontpellierCIRAD
Joger U (1979) Zur Oumlkologie und Verbreitung wenigbekannter Agamen Westafrikas (Reptilia SauriaAgamidae) Salamandra 15 31-52
Joger U (1980) Eine neue Art der Gattung Tarentola(Reptilia Sauria Gekkonidae) aus WestafrikaAmphibia-Reptilia 1 137-147
Joger U (1981) Zur Herpetofaunistik WestafrikasBonner zoologische Beitraumlge 32 297-340
Joger U (1984) Taxonomische Revision der GattungTarentola (Reptilia Gekkonidae) Bonner zoologischeBeitraumlge 35 129-174
Joger U amp Lambert M R K (1996) Analysis of theherpetofauna of the Republic of Mali I Annotatedinventory with description of a new Uromastyx(Sauria Agamidae) Journal of African Zoology 11021-51
Lambert M R K amp Mullieacute W C (1998) Sexualdichromatism of Agama boulengeri observed insouthern Mauritania British Herpetological SocietyBulletin 65 42-44
Le Berre M (1989) Faune du Sahara I Poissons-Amphibiens-Reptiles Paris Lechevalier-R Chabaud
Le Houeacuterou H N (1990) Deacutefinition et limitesbioclimatiques du Sahara Segravecheresse 1 246-259
Lenk P Kalyabina S Wink M amp Joger U (2001)Evolutionary relationships among the true vipers(Reptilia Viperidae) inferred from mitochondrial DNAsequences Molecular Biology and Evolution 19 94-104
Mateo J A Geniez Ph Loacutepez-Jurado J L amp Bons J(1998) Chorological analysis and morphologicalvariation of saurians of the genus Uromastyx (ReptiliaAgamidae) in western Sahara Description of two newtaxa Revista Espantildeola de Herpetologiacutea 12 97-109
Salvador A (1982) A revision of the lizards of the genusAcanthodactylus (Sauria Lacertidae) Bonnerzoologische Beitraumlge Monographien 16167 p
Schleich H H Kaumlstle W amp Kabisch K (1996)Amphibians and reptiles of North Africa KoenigsteinKoeltz Scientific Publications
Swofford D L (2002) PAUP phylogenetic analysisusing parsimony (and others methods) v40b10Sinauer Associates Sunderland MA
Thompson J D Gibson T J Plewniak F JeanmouginF amp Higgins D G (1997) The ClustalX windowsinterface flexible strategies for multiple sequencealignment aided by quality analysis tools NucleicAcids Research 24 4876-4882
Wilms T amp Boumlhme W (2001) Revision der Uromastyxacanthinura ndash Artengruppe mit Beschreibung einerneuen Art aus der Zentralsahara (Reptilia SauriaAgamidae) Zoologische Abhandlungen StaatlichesMuseum fuumlr Tierkunde Dresden 51 73-104
34
APPENDIX 1
Additional specimens of Agama species examinedfor this work EBD Estacioacuten Bioloacutegica de Dontildeana Se-ville Spain CET Centro de Estudios TropicalesSevilla Spain Locality and coordinates in parentheses
Agama agama CET(RIM)-028 (near Bouli 15deg25803 N11deg 55562 W Mauritania) CET(RIM)-328-329 (El Wad-Zoueina15deg 42689 N 9deg 39906 WMauritania) CET(RIM)-331-335 (El Wad-Foulania15deg 31682 N9deg 48896 W Mauritania)
Agama boueti CET(RIM)-287-291 336 (Ayoucircn elAtroucircs 16deg 37412 N9deg 37441 W Mauritania)
Agama boulengeri CET(RIM)-016-018 (Bou-gari16deg 32034 N10deg 47892 W Mauritania)CET(RIM)-021-022 (between Timbedgha and AyoucircnEl Atroucircs 16deg 26890 N9deg 14690 W Mauritania)CET(RIM)-066 (Terjicirct 20deg 15578 N13 deg 05854 WMauritania) CET(RIM)-094-96 (Guelta Molomhar20deg 34873 N13deg 07630 W Mauritania) CET(RIM)-116-117 (Oued Choucircm 21deg 22654 N12deg 58581 WMauritania) CET(RIM)-174-175 (Zig between Lekh-cheb and Ticircchicirct 18deg 34487 N9deg 48379 WMauritania) CET(RIM)-256-261 (Guelta Oumm Leb-are 16deg 29472 N10deg 49822 W Mauritania)CET(RIM)-338 (Ayoucircn el Atroucircs 16deg 37412 N9deg37441 W Mauritania) CET(RIM)-344 (Guelta Mat-mata17deg 53571 N12deg 07467 W Mauritania)
Agama impalearis EBD-11352 (Mount Marcan-Kbir Tetuaacuten Morocco) EBD-20222 (TaklimMorocco) EBD-22084 (Yassinen Morocco) EBD-24515 (Bab Bou Ichir-Fritissa Morocco) EBD-6552(El Aioum Western Sahara)
NEW AGAMA SPECIES FROM MAURITANIA 35

DNA studies were obtained from four specimens of thenew species (MNCN 41776-79) and preserved in abso-lute ethanol
The following 27 morphological characters were ob-tained from each specimen (1) 16 linear bodydimensions SVL snout-vent length TAL tail lengthTAH tail height (at the base of the tail) TAW tailwidth (at the base of the tail) RTAL relative tail length( of SVL) HL head length HW maximum headwidth HH maximum head height FOOT foot length(from heel to the tip of the longest toe including theclaw) HAND hand length (from wrist to the tip of thelongest finger including the claw) FINGER length ofthe fourth finger (from the joint with the hand to the tipincluding the claw) TOE fourth toe length (from thejoint with the foot to the tip including the claw) TLtibia length FL femur length (shank length) EN eye-nostril distance (from the posterior margin of the nostrilto the anterior corner of the eye) EE distance ear open-ing-eye (from the posterior corner of the eye to theanterior margin of the ear opening) (2) 11 scalationcounts characters VE number of ventral scales (fromthe inguinal region to the level of the axilla) EMBnumber of scales around midbody SL number ofsupralabials scales IL number of infralabial scales SOnumber of supraocular scales RSLE rows of scales be-tween supralabials and eye GNS groups of neck spinesIP number of inguinal pores LFF number of lamellaeunder fourth finger LFT number of lamellae underfourth toe SC number of spines of the nuchal crest
Scalation count characters of males of the two formsof A impalearis were provided by Brown (in litt)Comparisons with A boueti and A boulengeri werebased on data from Joger (1979) and personal observa-tions and on A agama from personal observations
Partial DNA sequences of the 16S rRNA mitochon-drial gene were obtained for four specimens of the newspecies and compared with previously published 16SrRNA sequences of A impalearis from Morocco(Brown et al 2002) Laudakia atricollis from Tanzaniawas used as outgroup (Brown et al 2002)
Accession numbers for the MNCN specimens se-quenced are as follows MNCN 41776=AY522926MNCN 41777= AY522927 MNCN 41778=AY522928 MNCN 41779= AY522929 Total genomicDNA extraction followed standard protocols describedelsewhere (Carranza et al 1999 2000) Primers usedin both amplification and sequencing were L2510 (5rsquo-CGCCTGTTTATCAAAAACAT-3rsquo) and H3062(5rsquo-CCGGTTTGAACTCAGATCA-3rsquo) from Lenk etal (2001)
Mitochondrial sequences were aligned in ClustalX(Thompson et al 1997) with default parameters Onlytwo gaps had to be postulated to align the new specieswith A impalearis In total 464 base pairs of 16S rRNAmitochondrial DNA were included in the phylogeneticanalysis of which 84 were variable and 27 parsimony-informative The results of the alignment are available
from the author upon request The aligned data set wasanalyzed using maximum-parsimony (MP) in PAUP40B10 (Swofford 2002) and include heuristicsearches involving tree bisection and reconnection(TBR) branch swapping with 100 random stepwise ad-ditions of taxa Gaps were included as fifth state Nodalsupport for the MP tree was assessed using bootstrapanalysis (Felsenstein 1985) involving 1000 pseudo-rep-lications
RESULTS
AGAMA CASTROVIEJOI SP NOV FIGS 1-8
Types Holotype MNCN-41779 an adult male fromDahr Chinguettti on the road between Atar and Tidjikja(20deg 26547rsquoN12deg 49407rsquoW) Wilaya of Adrar Col-lected by Joseacute M Padial 20 May 2002
Paratypes MNCN-41778 an adult female MNCN-41780 a subadult female and MNCN-41776 41777 twoyoung females (same data as the holotype)
Etymology This species is a patronym for Dr JavierCastroviejo (Galicia Spain) for his help and encour-agement and in recognition of his great effort in fundingthe study and conservation of biodiversity in Spain Af-rica and Latin America
Diagnosis A species morphologically similar toAgama impalearis with the following combination of
J M PADIAL
FIG 1 Male (holotype MNCN 41779) of Agamacastroviejoi sp nov Picture taken in the Adrar MountainsMauritania
FIG 2 Dorsal view of the male MNCN 41779 (holotype) ofAgama castroviejoi
28
NEW AGAMA SPECIES FROM MAURITANIA
FIG 3 Lateral view of the head of the male MNCN41779(holotype) of Agama castroviejoi
FIG 4 Frontal view of the snout of the male MNCN 41779(holotype) of Agama castroviejoi showing the orientation ofthe nostrils
FIG 5 Ventral view of the gular region of the male MNCN41779 (holotype) of Agama castroviejoi
FIG 6 Dorsal view of an adult female MNCN 41778(paratype) of Agama castroviejoi
FIG 7 Lateral view of the head of an adult female MNCN41778 (paratype) of Agama castroviejoi
FIG 8 Frontal view of the snout of an adult female MNCN41778 (paratype) of Agama castroviejoi showing theorientation of the nostrils
29
J M PADIAL
FIG 9 Strict consensus of 19 equally parsimonious trees showing the phylogenetic relationships between A impalearis and Acastroviejoi Tree-length= 97 steps CI= 0750 and RI= 0922 (both under exclusion of uninformative sites) Bootstrap supportvalues are shown above the nodes Haplotype numbers refer to Brown et al (2002)
characters (see Figs 2-8) long narrow and low headscales on head smooth small nuchal crest nine groupsof spines on each side of the head nostrils orienteddorsolaterally plicate throat with two gular folds andone mite pocket on each side of the neck regular keeledand mucronate scales in dorsum keels converging to-ward midbody less keeled and mucronate dorsal scalesin females relative length of fingers 3gt4gt5=2gt1 rela-tively long hind legs (always reaching the anteriormargin of the ear opening some reaching the eye) baseof the tail weakly compressed tail scales keeled andmucronate absence of caudal crest tail one and 34 totwice as long as SVL gular region with brown irregularlongitudinal lines ten preanal pores live male with red-dish eyelid ventral regions of the body tail andextremities immaculate white dorsal surfaces of femalelight (sandy) brown with dark brown subocular and la-bial bars and some transverse dark brown bands onback males with dorsal surfaces grey-blue with lightbrown spots Distinguished from the north-west Africanspecies (characters of the other species in parentheses)(see also Table 2) from Agama impalearis by relativelength of finger and toes smaller nuchal crest and ab-sence of caudal crest reddish eyelid in males anddifferent colour pattern some scalation count charac-ters (Table 3) and 43 difference in the 16S rRNAmitochondrial region sequenced for this study Thephylogenetic analysis presented in Fig 9 shows that Acastroviejoi and A impalearis form two reciprocally
monophyletic groups supported by relatively highbootstrap values Distinguished from Agama boueti bygular region with brown irregular longitudinal lines (im-maculate) red eyelid in males (brown) 8-9 groups ofneck spines (4-7) 10 preanal pores (12) 12 scales onfourth finger (9-10) 19-21 scales on fourth toe (15-17)different relative length of fingers and toes longer hindlegs Distinguished from Agama agama by very smallersize and very different colour pattern and sexual dimor-phism (metallic blue body and bright orange on heads ofadult males) Distinguished from Agama boulengeri byits smaller size very different colour pattern (speciallysexual dimorphism) fewer number of scales aroundmidbody smooth head scales higher number of groupof spines on the sides of the head and shoulder and by ab-sence of caudal crest
DESCRIPTION OF THE HOLOTYPE
An adult male with head scales smooth occipitalscale big dorsal scales homogeneous large rhomboidalkeeled and mucronate all keels converging towards thevertebral line keels on arms and limbs mucronate throatplicate with two gular folds and one mite pocket on eachside of the neck ventral scales smooth relative fingerlength 3gt4gt5=2gt1 tail twice as long as SVL base of thetail weakly compressed tail scales keeled and mucro-nate nostrils oriented upwards and backwards
Measurements (mm) Snout-vent length 7488 taillength 11670 head length 1984 head width 1599
30
TABLE 1 Linear body measurements (mm) and scalation count characters of the type series of Agama castroviejoi sp nov Seemethodology for abbreviations
Linear body MNCN MNCN MNCN MNCN MNCN Range measurement 41779 41777 41778 41776 41780and dimension (male) (juv female) (female) (juv female) (female)
SVL 7421 5991 7488 5931 6760 5931-7488TAL 11640 1051 - - 9017 9017-1164TAH 881 692 725 675 687 692-881TAW 1146 820 1055 813 984 813-1146RTAL 6375 570 - - 7497 570-7497HL 1984 1716 1965 1591 1840 1591-1984HW 1599 1358 1567 1321 1511 1321-1599HH 969 888 967 867 986 867-969EN 476 30 398 311 435 30-476EE 601 437 480 461 533 437-601HAND 1455 1204 1280 1153 1414 1153-1455FOOT 2364 1969 2104 1882 2261 1882-2364FINGER 677 623 575 519 667 519-677TOE 1202 964 975 920 1174 920-1202TL 2121 2031 2175 1977 2134 1977-2175FL 1936 1758 1986 1544 2029 1544-2029HLSVL 027 029 026 027 027 026-029HBSVL 022 023 021 022 022 021-023HLHB 124 126 125 120 122 120-126TLSVL 029 034 029 033 032 029-034FLSVL 026 029 027 026 030 026-030ENSVL 024 017 020 020 024 017-024EESVL 030 025 024 029 029 024-030TOESVL 016 016 013 016 017 013-017FOOTSVL 032 033 028 032 033 028-033
Scalation countcharacters
VE 63 59 61 58 59 58-63EMB 61 58 55 60 63 55-63SL 10 11 11 11 12 10-12IL 9 11 11 11 12 9-12SO 6 7 6 7 7 6-7RSLE 4 3 4 4 4 3-4GNS 9 8 8 8 9 8-9IP 10 - - - - -LFF 12 12 12 12 12 -LFT 19 19 21 19 20 19-21SC 6 6 7 7 7 6-7
NEW AGAMA SPECIES FROM MAURITANIA
head height 969 hand length 1455 (from posterior endof wrist to top of longest finger) foot length 2364 (fromposterior end of heel to top of longest toe) limb length1936 tibia length 2121 length of fourth finger (to joinwith third) 677 length of fourth toe 1202 eye-nostrillength (from the anterior border of the eye orbit to thenostril) 476 ear-eye length (from the anterior border ofthe tympanum to the posterior border of the eye orbit)601 tail width 1146 tail height (at the base) 881
Pholidosis 63 ventral scales between inguinal foldand the beginning of the arm 61 scales around midbody11 supralabials 9 infralabials 6 rows of supraoculars 4
rows of scales between supralabials and suboculars (in-cluding the subocular row) 9 groups of spines betweenthe anterior margin of the ear opening and the shoulderIn each group the central spines are largest 10 analpores 12 lamellae under fourth finger 19 lamellae un-der fourth toe nuchal crest composed of 6 spines
Colour in life Ventral regions of the body tail andextremities immaculate white dorsal surfaces grey-blue with dark brown subocular and labial bars gularregion with brown irregular longitudinal lines sometransverse dark brown bands on the back a perpendicu-lar thin beige band in the anterior margin of the back
31
J M PADIAL
TABLE 2 Diagnostic characters for Agama castroviejoi sp nov Agama impalearis and A boueti
A castroviejoi A impalearis A boueti
SVL 676-7488 to 120 to 99TL 9017-1164 150 to 160EMB 55-63 50-62Dorsal scales keeled keeled keeledHead scales smooth smooth smoothNuchal crest small strong smallGNS 8-9 3-9 4-8Caudal crest absent present absent
Relative length 3gt4gt5gt2gt1 3gt4gt5gt2gt1 3gt4gt2gt5gt1of fingersRelative length 4gt3gt5gt2gt1 3=4gt5gt2gt1 3gt4gt5gt2gt1of toes
Gular region brown lines brown lines white to yellowSexual dicromatism low low lowSociability solitary solitary solitaryHabitat desert rocky areas semidesert rocky areas sandy savannah
TABLE 3 Some scalation counts characters of male (holotype) of Agama castroviejoi and two geographical forms of Aimpalearis (a) southern form (Tan Tan Morocco) (b) northern form (Chefchauen Morocco) See methodology for abbreviations
SL IL GNS LFF LFT IP
Holotype 10 9 9 12 19 10a (n=6) 11-13 10-15 10-11 14-16 16-18 12-14b (n=4) 11-13 11-13 10-11 15 16-19 12-13
and one in the posterior margin two beige irregular cir-cumferences in the middle of the back
Colour in alcohol Ventral regions of the body tailand extremities immaculate white gular region withbrown irregular longitudinal lines all dorsal and dorso-lateral regions grey-blue (slate grey) with some lightbrown spot on the head back and forearms hands yel-low a perpendicular thin beige band in the anteriormargin of the back and one in the posterior margin twobeige irregular circumferences in the middle of the back
VARIATION
For variation in morphometrics see Table 1 The onlymales has more ventral scales and fewer labial scalesthan females The dorsal scales of the male are moremucronate than in females There is colour differencebetween sexes females being paler than males In lifefemales have almost no colour differences they havelight brown head with dark brown subocular stripes thedorsal region is beige to light brown with transversedark brown bands from the scapular region to the middleof the tail each dark band is composed of 3-7 scalesthey have a thin longitudinal light stripe The ventral re-gion is immaculate white with subgular longitudinalbrown stripes In alcohol the colour becomes faded butthe pattern is retained the feet and hands become lightyellow
From the genetic point of view there is a single basepair difference (and A-G transition) between specimenMNCN 41778 and specimens MNCN 41776 MNCN41777 MNCN 41779 in the 464 of the 16S rRNA mito-chondrial region sequenced for this study (Fig 9)
DISTRIBUTION AND ECOLOGY
Specimens are only known from the type localityThey were found on a tableland (dahr) 679 m above sealevel in the Adrar region in Mauritania This region ispart of the ldquoSahara Meridional Occidentalrdquo and is char-acterized by annual rainfall of about 100 mm andaverage annual temperatures of 30degC with minimumtemperatures in January rarely descending below 10degC(Le Houeacuterou 1990) The habitat is a rocky plain withlittle vegetation composed of sparse bushes and somesmall trees (Acacia spp) All specimens were foundalone and separate by few kilometres from each otherThe first specimen encountered was active at 0920 hr(air temperature of 286degC) The other specimens werefound perching around the middle of the day and wereeasily caught when they moved under solitary stones
DISCUSSION
Among the four Agama species (A agama Aboulengeri A boueti A impalearis) inhabiting the aridand semi-arid regions of NW Africa the last two show
32
superficial resemblance with A castroviejoi Neverthe-less morphological quantitative and meristic charactersdistinguish A castroviejoi well from the other speciesrecorded for Mauritania (A boulengeri A agama andA boueti) and also from the two Moroccan forms of Aimpalearis MtDNA analyses demonstrate the identityof A castroviejoi and its close relationship to Aimpalearis and the vicariant form from south and east ofthe Atlas mountains
Furthermore distributional social and ecologicalfactors also allow a clear separation among species Acastroviejoi is solitary and inhabits extremely dry rockyareas of the Adrar plateau Agama boueti is solitary andinhabits the semi-desert sandy plains and savannahs ofthe Sahel region of southern Mauritania and the AtlanticCoast Agama impalearis is a typical solitary rockdweller (Schleich et al 1996) Its distribution rangesfrom the Mediterranean coast (Bons amp Geniez 1996) toSeguia el Hamra in Western Sahara (Geniez et al2000) Although it was cited for ldquoMauritanieSahariennerdquo (Chabanaud 1917) and despite appearingwidely distributed across the country in Le Berrersquos(1989) distribution map for this species there are actu-ally no confirmed records for Mauritania MoreoverJoger (1979) noted that this species did not occur in theAdrar and that the nearest record corresponded toSeguia el Hamra The presence of this species in thecountry is still plausible in rocky areas similar to thoseof the Zemmour Mountains (Geniez et al 2000) butthis requires confirmation A boulengeri is a speciesfrom the Sahel Savannah that inhabits rocky areas andforms social groups We found this species as far northas Oued Choucircm (21deg 226N12deg 586W) near the bor-der with Western Sahara At this latitude it is alwaysassociated to wet rocky gorges (Gueltas) in the moun-tains A agama is another social species reported to beassociated with human settlements in the Adrar region(Dekeyser amp Villiers 1956) In the south of the countrythis species prefers trees from the Sahel savannahs but itshows high habitat plasticity (pers obs)
Miocene vicariance events have been found to be re-sponsible of the variation and differentiation in Aimpalearis in NW Africa (Brown amp Znari 1998 Brownet al 2002) The uplift of the Atlas mountains consti-tuted a great geographical barrier causing allopatricfragmentation of the populations A castroviejoi is iso-lated in a rocky tableland surrounded by lowland sandyplains of the Sahara that could serve as a barrier for arock dweller Also in situ selection-mediated responsesto the ecological current conditions of the isolatedpopulations are responsible of the morphological varia-tion of A impalearis (Brown amp Znari 1998) Presentecological conditions of the Adrar are very extreme andis probably one of the hottest areas of the Sahara (LeHoueacuterou 1990) As demonstrated by Brown amp Znari(1998) variation in scalation is significantly associatedwith different temperature conditions Therefore bothisolation and ecological differentiation could be impli-cated in the differentiation of A castroviejoi
Although the Adrar belongs to the Meridional Sahara(Le Houeacuterou 1990) this area shows a complex historywith its faunas and floras comprising a mixture of Medi-terranean Saharan and Sahelian species (Dekeyser ampVilliers 1956) Probably A castroviejoi constitutes anexample of a Saharan endemic of the Adrar Mountainsas some invertebrate species and also vertebrate subspe-cies (Dekeyser amp Villiers 1956) Nevertheless thephylogeographic affinities of A castroviejoi remain un-clear since it could have a Sahelian or Saharan origin Amore thorough phylogenetic comparison among NorthAfrican agamas will help to resolve this question
ACKNOWLEDGEMENTS
This work could not have been carried out withoutthe effort of J Castroviejo and the people of AsociacioacutenAmigos de Dontildeana I am very gratefully to S Carranza(BMNH) who did the genetic work and helped me toimprove the manuscript To I de la Riva (MNCN) forhis help in many aspects of this work To Dr NAnderson (BMNH) who revised the draft version andthe English of the manuscript To R Garciacutea M UrceraC Carballo M O Deida and E Mohamed O Saleh fortheir help and companion in Mauritania and J Peacuterez EAvila S Castroviejo-Fisher and A Quintana for theirfriendship and companion in the first trip to MauritaniaP Geniez J A Mateo H Nickel and W Boumlhme allhelped us in some aspect of this work W Wuster andtwo anonymous reviewers critically commented an ear-lier draft of the manuscript R Brown kindly providedraw scalation data for A impalearis from Morocco andcontributed to the improvement of the manuscript
REFERENCES
Boumlhme W (2000) Die wechselvolle Geschichte derSahara Untersuchung von Reptilienzoumlnosen entlangeines westsaharischen Transektes mit einemuumlberraschenden Fund im Suumldosten Mauritaniens Tierund Museum 7 11-21
Boumlhme W Meinig H amp Roumldel M A (1996) Newrecords of amphibians and reptiles from Burkina Fasoand Mali British Herpetological Society Bulletin 56 7-26
Boumlhme W Wilms T Nickel H amp Merz M (2001)Bericht uumlber eine herpetologische Forschungsreise indie westliche Sahara und ihre Randgebiete Zeitschriftdes Koumllner Zoo 44 117-131
Bons J amp Geniez P (1996) Anfibios y reptiles deMarruecos (Incluido Saacutehara Occidental) Atlasbiogeograacutefico Barcelona Asociacioacuten HerpetoloacutegicaEspantildeola
Brown R P amp Znari M (1998) Morphological variationin Agama impalearis from Morocco evidence forhistorical population vicariance and current climaticeffects Ecography 21 605-612
Brown R P Znari M El Mouden E H amp Harris P(1999) Estimating asymptotic size and geographicvariation in Agama impalearis from MoroccoEcography 22 277-284
NEW AGAMA SPECIES FROM MAURITANIA 33
J M PADIAL
Accepted 22304
Brown R P Pestano J amp Suaacuterez N M (2002) TheAtlas mountains as a biogeographical divide in North-West Africa evidence from mtDNA evolution in theAgamid lizard Agama impalearis MolecularPhylogenetics and Evolution 24 324-332
Carranza S Arnold E N Thomas R H Mateo J ampLopez-Jurado L F (1999) Status of the extinct giantlacertid lizard Gallotia simonyi simonyi (ReptiliaLacertidae) assessed using mtDNA sequences frommuseum specimens Herpetological Journal l9 83-86
Carranza S Arnold E N Mateo J amp Lopez-Jurado LF (2000) Long-distance colonization and radiation ingekkonid lizards Tarentola (Reptilia Gekkonidae)revealed by mitochondrial DNA sequencesProceedings of the Royal Society Series B 267 637-649
Chabanaud P (1917) Enumeacuteration des reptiles nonencore eacutetudieacutes de lrsquoAfrique occidentale appartenantaux collections du Museacuteum avec la description desespeacuteces nouvelles Bulletin du Museacuteum NationaldrsquoHistoire Naturelle de Paris Serie Zoologique 2383-105
Dekeyser P L amp Villiers A (1956) Contribution alrsquoeacutetude du peuplement de la Mauritanie Notationseacutecologiques et biogeacuteographiques sur la faune delrsquoAdrar Memoires de lrsquoInstitut Franccedilais de lrsquoAfriqueNoire 44 222 p 25 pl
Felsenstein J (1985) Confidence-limits on phylogenies ndashan approach using the bootstrap Evolution 39 783-791
Geniez Ph amp Foucart A (1995) Un nouvelAcanthodactyle en Algeacuterie Acanthodactylustaghitensis n sp (Reptilia Sauria Lacertidae)Bulletin du Museacuteum National drsquoHistoire NaturelleParis 4deg seacuterie 17 3-9
Geniez P Mateo J A amp Bons J (2000) A checklist ofthe amphibians and reptiles of Western Sahara(Amphibia Reptilia) Herpetozoa 13 149-163
Ineich I (1997) Les amphibiens et reptiles du littoralmauritanien In Environnement et littoralmauritanien 93-99 Colas P (Ed) MontpellierCIRAD
Joger U (1979) Zur Oumlkologie und Verbreitung wenigbekannter Agamen Westafrikas (Reptilia SauriaAgamidae) Salamandra 15 31-52
Joger U (1980) Eine neue Art der Gattung Tarentola(Reptilia Sauria Gekkonidae) aus WestafrikaAmphibia-Reptilia 1 137-147
Joger U (1981) Zur Herpetofaunistik WestafrikasBonner zoologische Beitraumlge 32 297-340
Joger U (1984) Taxonomische Revision der GattungTarentola (Reptilia Gekkonidae) Bonner zoologischeBeitraumlge 35 129-174
Joger U amp Lambert M R K (1996) Analysis of theherpetofauna of the Republic of Mali I Annotatedinventory with description of a new Uromastyx(Sauria Agamidae) Journal of African Zoology 11021-51
Lambert M R K amp Mullieacute W C (1998) Sexualdichromatism of Agama boulengeri observed insouthern Mauritania British Herpetological SocietyBulletin 65 42-44
Le Berre M (1989) Faune du Sahara I Poissons-Amphibiens-Reptiles Paris Lechevalier-R Chabaud
Le Houeacuterou H N (1990) Deacutefinition et limitesbioclimatiques du Sahara Segravecheresse 1 246-259
Lenk P Kalyabina S Wink M amp Joger U (2001)Evolutionary relationships among the true vipers(Reptilia Viperidae) inferred from mitochondrial DNAsequences Molecular Biology and Evolution 19 94-104
Mateo J A Geniez Ph Loacutepez-Jurado J L amp Bons J(1998) Chorological analysis and morphologicalvariation of saurians of the genus Uromastyx (ReptiliaAgamidae) in western Sahara Description of two newtaxa Revista Espantildeola de Herpetologiacutea 12 97-109
Salvador A (1982) A revision of the lizards of the genusAcanthodactylus (Sauria Lacertidae) Bonnerzoologische Beitraumlge Monographien 16167 p
Schleich H H Kaumlstle W amp Kabisch K (1996)Amphibians and reptiles of North Africa KoenigsteinKoeltz Scientific Publications
Swofford D L (2002) PAUP phylogenetic analysisusing parsimony (and others methods) v40b10Sinauer Associates Sunderland MA
Thompson J D Gibson T J Plewniak F JeanmouginF amp Higgins D G (1997) The ClustalX windowsinterface flexible strategies for multiple sequencealignment aided by quality analysis tools NucleicAcids Research 24 4876-4882
Wilms T amp Boumlhme W (2001) Revision der Uromastyxacanthinura ndash Artengruppe mit Beschreibung einerneuen Art aus der Zentralsahara (Reptilia SauriaAgamidae) Zoologische Abhandlungen StaatlichesMuseum fuumlr Tierkunde Dresden 51 73-104
34
APPENDIX 1
Additional specimens of Agama species examinedfor this work EBD Estacioacuten Bioloacutegica de Dontildeana Se-ville Spain CET Centro de Estudios TropicalesSevilla Spain Locality and coordinates in parentheses
Agama agama CET(RIM)-028 (near Bouli 15deg25803 N11deg 55562 W Mauritania) CET(RIM)-328-329 (El Wad-Zoueina15deg 42689 N 9deg 39906 WMauritania) CET(RIM)-331-335 (El Wad-Foulania15deg 31682 N9deg 48896 W Mauritania)
Agama boueti CET(RIM)-287-291 336 (Ayoucircn elAtroucircs 16deg 37412 N9deg 37441 W Mauritania)
Agama boulengeri CET(RIM)-016-018 (Bou-gari16deg 32034 N10deg 47892 W Mauritania)CET(RIM)-021-022 (between Timbedgha and AyoucircnEl Atroucircs 16deg 26890 N9deg 14690 W Mauritania)CET(RIM)-066 (Terjicirct 20deg 15578 N13 deg 05854 WMauritania) CET(RIM)-094-96 (Guelta Molomhar20deg 34873 N13deg 07630 W Mauritania) CET(RIM)-116-117 (Oued Choucircm 21deg 22654 N12deg 58581 WMauritania) CET(RIM)-174-175 (Zig between Lekh-cheb and Ticircchicirct 18deg 34487 N9deg 48379 WMauritania) CET(RIM)-256-261 (Guelta Oumm Leb-are 16deg 29472 N10deg 49822 W Mauritania)CET(RIM)-338 (Ayoucircn el Atroucircs 16deg 37412 N9deg37441 W Mauritania) CET(RIM)-344 (Guelta Mat-mata17deg 53571 N12deg 07467 W Mauritania)
Agama impalearis EBD-11352 (Mount Marcan-Kbir Tetuaacuten Morocco) EBD-20222 (TaklimMorocco) EBD-22084 (Yassinen Morocco) EBD-24515 (Bab Bou Ichir-Fritissa Morocco) EBD-6552(El Aioum Western Sahara)
NEW AGAMA SPECIES FROM MAURITANIA 35

NEW AGAMA SPECIES FROM MAURITANIA
FIG 3 Lateral view of the head of the male MNCN41779(holotype) of Agama castroviejoi
FIG 4 Frontal view of the snout of the male MNCN 41779(holotype) of Agama castroviejoi showing the orientation ofthe nostrils
FIG 5 Ventral view of the gular region of the male MNCN41779 (holotype) of Agama castroviejoi
FIG 6 Dorsal view of an adult female MNCN 41778(paratype) of Agama castroviejoi
FIG 7 Lateral view of the head of an adult female MNCN41778 (paratype) of Agama castroviejoi
FIG 8 Frontal view of the snout of an adult female MNCN41778 (paratype) of Agama castroviejoi showing theorientation of the nostrils
29
J M PADIAL
FIG 9 Strict consensus of 19 equally parsimonious trees showing the phylogenetic relationships between A impalearis and Acastroviejoi Tree-length= 97 steps CI= 0750 and RI= 0922 (both under exclusion of uninformative sites) Bootstrap supportvalues are shown above the nodes Haplotype numbers refer to Brown et al (2002)
characters (see Figs 2-8) long narrow and low headscales on head smooth small nuchal crest nine groupsof spines on each side of the head nostrils orienteddorsolaterally plicate throat with two gular folds andone mite pocket on each side of the neck regular keeledand mucronate scales in dorsum keels converging to-ward midbody less keeled and mucronate dorsal scalesin females relative length of fingers 3gt4gt5=2gt1 rela-tively long hind legs (always reaching the anteriormargin of the ear opening some reaching the eye) baseof the tail weakly compressed tail scales keeled andmucronate absence of caudal crest tail one and 34 totwice as long as SVL gular region with brown irregularlongitudinal lines ten preanal pores live male with red-dish eyelid ventral regions of the body tail andextremities immaculate white dorsal surfaces of femalelight (sandy) brown with dark brown subocular and la-bial bars and some transverse dark brown bands onback males with dorsal surfaces grey-blue with lightbrown spots Distinguished from the north-west Africanspecies (characters of the other species in parentheses)(see also Table 2) from Agama impalearis by relativelength of finger and toes smaller nuchal crest and ab-sence of caudal crest reddish eyelid in males anddifferent colour pattern some scalation count charac-ters (Table 3) and 43 difference in the 16S rRNAmitochondrial region sequenced for this study Thephylogenetic analysis presented in Fig 9 shows that Acastroviejoi and A impalearis form two reciprocally
monophyletic groups supported by relatively highbootstrap values Distinguished from Agama boueti bygular region with brown irregular longitudinal lines (im-maculate) red eyelid in males (brown) 8-9 groups ofneck spines (4-7) 10 preanal pores (12) 12 scales onfourth finger (9-10) 19-21 scales on fourth toe (15-17)different relative length of fingers and toes longer hindlegs Distinguished from Agama agama by very smallersize and very different colour pattern and sexual dimor-phism (metallic blue body and bright orange on heads ofadult males) Distinguished from Agama boulengeri byits smaller size very different colour pattern (speciallysexual dimorphism) fewer number of scales aroundmidbody smooth head scales higher number of groupof spines on the sides of the head and shoulder and by ab-sence of caudal crest
DESCRIPTION OF THE HOLOTYPE
An adult male with head scales smooth occipitalscale big dorsal scales homogeneous large rhomboidalkeeled and mucronate all keels converging towards thevertebral line keels on arms and limbs mucronate throatplicate with two gular folds and one mite pocket on eachside of the neck ventral scales smooth relative fingerlength 3gt4gt5=2gt1 tail twice as long as SVL base of thetail weakly compressed tail scales keeled and mucro-nate nostrils oriented upwards and backwards
Measurements (mm) Snout-vent length 7488 taillength 11670 head length 1984 head width 1599
30
TABLE 1 Linear body measurements (mm) and scalation count characters of the type series of Agama castroviejoi sp nov Seemethodology for abbreviations
Linear body MNCN MNCN MNCN MNCN MNCN Range measurement 41779 41777 41778 41776 41780and dimension (male) (juv female) (female) (juv female) (female)
SVL 7421 5991 7488 5931 6760 5931-7488TAL 11640 1051 - - 9017 9017-1164TAH 881 692 725 675 687 692-881TAW 1146 820 1055 813 984 813-1146RTAL 6375 570 - - 7497 570-7497HL 1984 1716 1965 1591 1840 1591-1984HW 1599 1358 1567 1321 1511 1321-1599HH 969 888 967 867 986 867-969EN 476 30 398 311 435 30-476EE 601 437 480 461 533 437-601HAND 1455 1204 1280 1153 1414 1153-1455FOOT 2364 1969 2104 1882 2261 1882-2364FINGER 677 623 575 519 667 519-677TOE 1202 964 975 920 1174 920-1202TL 2121 2031 2175 1977 2134 1977-2175FL 1936 1758 1986 1544 2029 1544-2029HLSVL 027 029 026 027 027 026-029HBSVL 022 023 021 022 022 021-023HLHB 124 126 125 120 122 120-126TLSVL 029 034 029 033 032 029-034FLSVL 026 029 027 026 030 026-030ENSVL 024 017 020 020 024 017-024EESVL 030 025 024 029 029 024-030TOESVL 016 016 013 016 017 013-017FOOTSVL 032 033 028 032 033 028-033
Scalation countcharacters
VE 63 59 61 58 59 58-63EMB 61 58 55 60 63 55-63SL 10 11 11 11 12 10-12IL 9 11 11 11 12 9-12SO 6 7 6 7 7 6-7RSLE 4 3 4 4 4 3-4GNS 9 8 8 8 9 8-9IP 10 - - - - -LFF 12 12 12 12 12 -LFT 19 19 21 19 20 19-21SC 6 6 7 7 7 6-7
NEW AGAMA SPECIES FROM MAURITANIA
head height 969 hand length 1455 (from posterior endof wrist to top of longest finger) foot length 2364 (fromposterior end of heel to top of longest toe) limb length1936 tibia length 2121 length of fourth finger (to joinwith third) 677 length of fourth toe 1202 eye-nostrillength (from the anterior border of the eye orbit to thenostril) 476 ear-eye length (from the anterior border ofthe tympanum to the posterior border of the eye orbit)601 tail width 1146 tail height (at the base) 881
Pholidosis 63 ventral scales between inguinal foldand the beginning of the arm 61 scales around midbody11 supralabials 9 infralabials 6 rows of supraoculars 4
rows of scales between supralabials and suboculars (in-cluding the subocular row) 9 groups of spines betweenthe anterior margin of the ear opening and the shoulderIn each group the central spines are largest 10 analpores 12 lamellae under fourth finger 19 lamellae un-der fourth toe nuchal crest composed of 6 spines
Colour in life Ventral regions of the body tail andextremities immaculate white dorsal surfaces grey-blue with dark brown subocular and labial bars gularregion with brown irregular longitudinal lines sometransverse dark brown bands on the back a perpendicu-lar thin beige band in the anterior margin of the back
31
J M PADIAL
TABLE 2 Diagnostic characters for Agama castroviejoi sp nov Agama impalearis and A boueti
A castroviejoi A impalearis A boueti
SVL 676-7488 to 120 to 99TL 9017-1164 150 to 160EMB 55-63 50-62Dorsal scales keeled keeled keeledHead scales smooth smooth smoothNuchal crest small strong smallGNS 8-9 3-9 4-8Caudal crest absent present absent
Relative length 3gt4gt5gt2gt1 3gt4gt5gt2gt1 3gt4gt2gt5gt1of fingersRelative length 4gt3gt5gt2gt1 3=4gt5gt2gt1 3gt4gt5gt2gt1of toes
Gular region brown lines brown lines white to yellowSexual dicromatism low low lowSociability solitary solitary solitaryHabitat desert rocky areas semidesert rocky areas sandy savannah
TABLE 3 Some scalation counts characters of male (holotype) of Agama castroviejoi and two geographical forms of Aimpalearis (a) southern form (Tan Tan Morocco) (b) northern form (Chefchauen Morocco) See methodology for abbreviations
SL IL GNS LFF LFT IP
Holotype 10 9 9 12 19 10a (n=6) 11-13 10-15 10-11 14-16 16-18 12-14b (n=4) 11-13 11-13 10-11 15 16-19 12-13
and one in the posterior margin two beige irregular cir-cumferences in the middle of the back
Colour in alcohol Ventral regions of the body tailand extremities immaculate white gular region withbrown irregular longitudinal lines all dorsal and dorso-lateral regions grey-blue (slate grey) with some lightbrown spot on the head back and forearms hands yel-low a perpendicular thin beige band in the anteriormargin of the back and one in the posterior margin twobeige irregular circumferences in the middle of the back
VARIATION
For variation in morphometrics see Table 1 The onlymales has more ventral scales and fewer labial scalesthan females The dorsal scales of the male are moremucronate than in females There is colour differencebetween sexes females being paler than males In lifefemales have almost no colour differences they havelight brown head with dark brown subocular stripes thedorsal region is beige to light brown with transversedark brown bands from the scapular region to the middleof the tail each dark band is composed of 3-7 scalesthey have a thin longitudinal light stripe The ventral re-gion is immaculate white with subgular longitudinalbrown stripes In alcohol the colour becomes faded butthe pattern is retained the feet and hands become lightyellow
From the genetic point of view there is a single basepair difference (and A-G transition) between specimenMNCN 41778 and specimens MNCN 41776 MNCN41777 MNCN 41779 in the 464 of the 16S rRNA mito-chondrial region sequenced for this study (Fig 9)
DISTRIBUTION AND ECOLOGY
Specimens are only known from the type localityThey were found on a tableland (dahr) 679 m above sealevel in the Adrar region in Mauritania This region ispart of the ldquoSahara Meridional Occidentalrdquo and is char-acterized by annual rainfall of about 100 mm andaverage annual temperatures of 30degC with minimumtemperatures in January rarely descending below 10degC(Le Houeacuterou 1990) The habitat is a rocky plain withlittle vegetation composed of sparse bushes and somesmall trees (Acacia spp) All specimens were foundalone and separate by few kilometres from each otherThe first specimen encountered was active at 0920 hr(air temperature of 286degC) The other specimens werefound perching around the middle of the day and wereeasily caught when they moved under solitary stones
DISCUSSION
Among the four Agama species (A agama Aboulengeri A boueti A impalearis) inhabiting the aridand semi-arid regions of NW Africa the last two show
32
superficial resemblance with A castroviejoi Neverthe-less morphological quantitative and meristic charactersdistinguish A castroviejoi well from the other speciesrecorded for Mauritania (A boulengeri A agama andA boueti) and also from the two Moroccan forms of Aimpalearis MtDNA analyses demonstrate the identityof A castroviejoi and its close relationship to Aimpalearis and the vicariant form from south and east ofthe Atlas mountains
Furthermore distributional social and ecologicalfactors also allow a clear separation among species Acastroviejoi is solitary and inhabits extremely dry rockyareas of the Adrar plateau Agama boueti is solitary andinhabits the semi-desert sandy plains and savannahs ofthe Sahel region of southern Mauritania and the AtlanticCoast Agama impalearis is a typical solitary rockdweller (Schleich et al 1996) Its distribution rangesfrom the Mediterranean coast (Bons amp Geniez 1996) toSeguia el Hamra in Western Sahara (Geniez et al2000) Although it was cited for ldquoMauritanieSahariennerdquo (Chabanaud 1917) and despite appearingwidely distributed across the country in Le Berrersquos(1989) distribution map for this species there are actu-ally no confirmed records for Mauritania MoreoverJoger (1979) noted that this species did not occur in theAdrar and that the nearest record corresponded toSeguia el Hamra The presence of this species in thecountry is still plausible in rocky areas similar to thoseof the Zemmour Mountains (Geniez et al 2000) butthis requires confirmation A boulengeri is a speciesfrom the Sahel Savannah that inhabits rocky areas andforms social groups We found this species as far northas Oued Choucircm (21deg 226N12deg 586W) near the bor-der with Western Sahara At this latitude it is alwaysassociated to wet rocky gorges (Gueltas) in the moun-tains A agama is another social species reported to beassociated with human settlements in the Adrar region(Dekeyser amp Villiers 1956) In the south of the countrythis species prefers trees from the Sahel savannahs but itshows high habitat plasticity (pers obs)
Miocene vicariance events have been found to be re-sponsible of the variation and differentiation in Aimpalearis in NW Africa (Brown amp Znari 1998 Brownet al 2002) The uplift of the Atlas mountains consti-tuted a great geographical barrier causing allopatricfragmentation of the populations A castroviejoi is iso-lated in a rocky tableland surrounded by lowland sandyplains of the Sahara that could serve as a barrier for arock dweller Also in situ selection-mediated responsesto the ecological current conditions of the isolatedpopulations are responsible of the morphological varia-tion of A impalearis (Brown amp Znari 1998) Presentecological conditions of the Adrar are very extreme andis probably one of the hottest areas of the Sahara (LeHoueacuterou 1990) As demonstrated by Brown amp Znari(1998) variation in scalation is significantly associatedwith different temperature conditions Therefore bothisolation and ecological differentiation could be impli-cated in the differentiation of A castroviejoi
Although the Adrar belongs to the Meridional Sahara(Le Houeacuterou 1990) this area shows a complex historywith its faunas and floras comprising a mixture of Medi-terranean Saharan and Sahelian species (Dekeyser ampVilliers 1956) Probably A castroviejoi constitutes anexample of a Saharan endemic of the Adrar Mountainsas some invertebrate species and also vertebrate subspe-cies (Dekeyser amp Villiers 1956) Nevertheless thephylogeographic affinities of A castroviejoi remain un-clear since it could have a Sahelian or Saharan origin Amore thorough phylogenetic comparison among NorthAfrican agamas will help to resolve this question
ACKNOWLEDGEMENTS
This work could not have been carried out withoutthe effort of J Castroviejo and the people of AsociacioacutenAmigos de Dontildeana I am very gratefully to S Carranza(BMNH) who did the genetic work and helped me toimprove the manuscript To I de la Riva (MNCN) forhis help in many aspects of this work To Dr NAnderson (BMNH) who revised the draft version andthe English of the manuscript To R Garciacutea M UrceraC Carballo M O Deida and E Mohamed O Saleh fortheir help and companion in Mauritania and J Peacuterez EAvila S Castroviejo-Fisher and A Quintana for theirfriendship and companion in the first trip to MauritaniaP Geniez J A Mateo H Nickel and W Boumlhme allhelped us in some aspect of this work W Wuster andtwo anonymous reviewers critically commented an ear-lier draft of the manuscript R Brown kindly providedraw scalation data for A impalearis from Morocco andcontributed to the improvement of the manuscript
REFERENCES
Boumlhme W (2000) Die wechselvolle Geschichte derSahara Untersuchung von Reptilienzoumlnosen entlangeines westsaharischen Transektes mit einemuumlberraschenden Fund im Suumldosten Mauritaniens Tierund Museum 7 11-21
Boumlhme W Meinig H amp Roumldel M A (1996) Newrecords of amphibians and reptiles from Burkina Fasoand Mali British Herpetological Society Bulletin 56 7-26
Boumlhme W Wilms T Nickel H amp Merz M (2001)Bericht uumlber eine herpetologische Forschungsreise indie westliche Sahara und ihre Randgebiete Zeitschriftdes Koumllner Zoo 44 117-131
Bons J amp Geniez P (1996) Anfibios y reptiles deMarruecos (Incluido Saacutehara Occidental) Atlasbiogeograacutefico Barcelona Asociacioacuten HerpetoloacutegicaEspantildeola
Brown R P amp Znari M (1998) Morphological variationin Agama impalearis from Morocco evidence forhistorical population vicariance and current climaticeffects Ecography 21 605-612
Brown R P Znari M El Mouden E H amp Harris P(1999) Estimating asymptotic size and geographicvariation in Agama impalearis from MoroccoEcography 22 277-284
NEW AGAMA SPECIES FROM MAURITANIA 33
J M PADIAL
Accepted 22304
Brown R P Pestano J amp Suaacuterez N M (2002) TheAtlas mountains as a biogeographical divide in North-West Africa evidence from mtDNA evolution in theAgamid lizard Agama impalearis MolecularPhylogenetics and Evolution 24 324-332
Carranza S Arnold E N Thomas R H Mateo J ampLopez-Jurado L F (1999) Status of the extinct giantlacertid lizard Gallotia simonyi simonyi (ReptiliaLacertidae) assessed using mtDNA sequences frommuseum specimens Herpetological Journal l9 83-86
Carranza S Arnold E N Mateo J amp Lopez-Jurado LF (2000) Long-distance colonization and radiation ingekkonid lizards Tarentola (Reptilia Gekkonidae)revealed by mitochondrial DNA sequencesProceedings of the Royal Society Series B 267 637-649
Chabanaud P (1917) Enumeacuteration des reptiles nonencore eacutetudieacutes de lrsquoAfrique occidentale appartenantaux collections du Museacuteum avec la description desespeacuteces nouvelles Bulletin du Museacuteum NationaldrsquoHistoire Naturelle de Paris Serie Zoologique 2383-105
Dekeyser P L amp Villiers A (1956) Contribution alrsquoeacutetude du peuplement de la Mauritanie Notationseacutecologiques et biogeacuteographiques sur la faune delrsquoAdrar Memoires de lrsquoInstitut Franccedilais de lrsquoAfriqueNoire 44 222 p 25 pl
Felsenstein J (1985) Confidence-limits on phylogenies ndashan approach using the bootstrap Evolution 39 783-791
Geniez Ph amp Foucart A (1995) Un nouvelAcanthodactyle en Algeacuterie Acanthodactylustaghitensis n sp (Reptilia Sauria Lacertidae)Bulletin du Museacuteum National drsquoHistoire NaturelleParis 4deg seacuterie 17 3-9
Geniez P Mateo J A amp Bons J (2000) A checklist ofthe amphibians and reptiles of Western Sahara(Amphibia Reptilia) Herpetozoa 13 149-163
Ineich I (1997) Les amphibiens et reptiles du littoralmauritanien In Environnement et littoralmauritanien 93-99 Colas P (Ed) MontpellierCIRAD
Joger U (1979) Zur Oumlkologie und Verbreitung wenigbekannter Agamen Westafrikas (Reptilia SauriaAgamidae) Salamandra 15 31-52
Joger U (1980) Eine neue Art der Gattung Tarentola(Reptilia Sauria Gekkonidae) aus WestafrikaAmphibia-Reptilia 1 137-147
Joger U (1981) Zur Herpetofaunistik WestafrikasBonner zoologische Beitraumlge 32 297-340
Joger U (1984) Taxonomische Revision der GattungTarentola (Reptilia Gekkonidae) Bonner zoologischeBeitraumlge 35 129-174
Joger U amp Lambert M R K (1996) Analysis of theherpetofauna of the Republic of Mali I Annotatedinventory with description of a new Uromastyx(Sauria Agamidae) Journal of African Zoology 11021-51
Lambert M R K amp Mullieacute W C (1998) Sexualdichromatism of Agama boulengeri observed insouthern Mauritania British Herpetological SocietyBulletin 65 42-44
Le Berre M (1989) Faune du Sahara I Poissons-Amphibiens-Reptiles Paris Lechevalier-R Chabaud
Le Houeacuterou H N (1990) Deacutefinition et limitesbioclimatiques du Sahara Segravecheresse 1 246-259
Lenk P Kalyabina S Wink M amp Joger U (2001)Evolutionary relationships among the true vipers(Reptilia Viperidae) inferred from mitochondrial DNAsequences Molecular Biology and Evolution 19 94-104
Mateo J A Geniez Ph Loacutepez-Jurado J L amp Bons J(1998) Chorological analysis and morphologicalvariation of saurians of the genus Uromastyx (ReptiliaAgamidae) in western Sahara Description of two newtaxa Revista Espantildeola de Herpetologiacutea 12 97-109
Salvador A (1982) A revision of the lizards of the genusAcanthodactylus (Sauria Lacertidae) Bonnerzoologische Beitraumlge Monographien 16167 p
Schleich H H Kaumlstle W amp Kabisch K (1996)Amphibians and reptiles of North Africa KoenigsteinKoeltz Scientific Publications
Swofford D L (2002) PAUP phylogenetic analysisusing parsimony (and others methods) v40b10Sinauer Associates Sunderland MA
Thompson J D Gibson T J Plewniak F JeanmouginF amp Higgins D G (1997) The ClustalX windowsinterface flexible strategies for multiple sequencealignment aided by quality analysis tools NucleicAcids Research 24 4876-4882
Wilms T amp Boumlhme W (2001) Revision der Uromastyxacanthinura ndash Artengruppe mit Beschreibung einerneuen Art aus der Zentralsahara (Reptilia SauriaAgamidae) Zoologische Abhandlungen StaatlichesMuseum fuumlr Tierkunde Dresden 51 73-104
34
APPENDIX 1
Additional specimens of Agama species examinedfor this work EBD Estacioacuten Bioloacutegica de Dontildeana Se-ville Spain CET Centro de Estudios TropicalesSevilla Spain Locality and coordinates in parentheses
Agama agama CET(RIM)-028 (near Bouli 15deg25803 N11deg 55562 W Mauritania) CET(RIM)-328-329 (El Wad-Zoueina15deg 42689 N 9deg 39906 WMauritania) CET(RIM)-331-335 (El Wad-Foulania15deg 31682 N9deg 48896 W Mauritania)
Agama boueti CET(RIM)-287-291 336 (Ayoucircn elAtroucircs 16deg 37412 N9deg 37441 W Mauritania)
Agama boulengeri CET(RIM)-016-018 (Bou-gari16deg 32034 N10deg 47892 W Mauritania)CET(RIM)-021-022 (between Timbedgha and AyoucircnEl Atroucircs 16deg 26890 N9deg 14690 W Mauritania)CET(RIM)-066 (Terjicirct 20deg 15578 N13 deg 05854 WMauritania) CET(RIM)-094-96 (Guelta Molomhar20deg 34873 N13deg 07630 W Mauritania) CET(RIM)-116-117 (Oued Choucircm 21deg 22654 N12deg 58581 WMauritania) CET(RIM)-174-175 (Zig between Lekh-cheb and Ticircchicirct 18deg 34487 N9deg 48379 WMauritania) CET(RIM)-256-261 (Guelta Oumm Leb-are 16deg 29472 N10deg 49822 W Mauritania)CET(RIM)-338 (Ayoucircn el Atroucircs 16deg 37412 N9deg37441 W Mauritania) CET(RIM)-344 (Guelta Mat-mata17deg 53571 N12deg 07467 W Mauritania)
Agama impalearis EBD-11352 (Mount Marcan-Kbir Tetuaacuten Morocco) EBD-20222 (TaklimMorocco) EBD-22084 (Yassinen Morocco) EBD-24515 (Bab Bou Ichir-Fritissa Morocco) EBD-6552(El Aioum Western Sahara)
NEW AGAMA SPECIES FROM MAURITANIA 35

J M PADIAL
FIG 9 Strict consensus of 19 equally parsimonious trees showing the phylogenetic relationships between A impalearis and Acastroviejoi Tree-length= 97 steps CI= 0750 and RI= 0922 (both under exclusion of uninformative sites) Bootstrap supportvalues are shown above the nodes Haplotype numbers refer to Brown et al (2002)
characters (see Figs 2-8) long narrow and low headscales on head smooth small nuchal crest nine groupsof spines on each side of the head nostrils orienteddorsolaterally plicate throat with two gular folds andone mite pocket on each side of the neck regular keeledand mucronate scales in dorsum keels converging to-ward midbody less keeled and mucronate dorsal scalesin females relative length of fingers 3gt4gt5=2gt1 rela-tively long hind legs (always reaching the anteriormargin of the ear opening some reaching the eye) baseof the tail weakly compressed tail scales keeled andmucronate absence of caudal crest tail one and 34 totwice as long as SVL gular region with brown irregularlongitudinal lines ten preanal pores live male with red-dish eyelid ventral regions of the body tail andextremities immaculate white dorsal surfaces of femalelight (sandy) brown with dark brown subocular and la-bial bars and some transverse dark brown bands onback males with dorsal surfaces grey-blue with lightbrown spots Distinguished from the north-west Africanspecies (characters of the other species in parentheses)(see also Table 2) from Agama impalearis by relativelength of finger and toes smaller nuchal crest and ab-sence of caudal crest reddish eyelid in males anddifferent colour pattern some scalation count charac-ters (Table 3) and 43 difference in the 16S rRNAmitochondrial region sequenced for this study Thephylogenetic analysis presented in Fig 9 shows that Acastroviejoi and A impalearis form two reciprocally
monophyletic groups supported by relatively highbootstrap values Distinguished from Agama boueti bygular region with brown irregular longitudinal lines (im-maculate) red eyelid in males (brown) 8-9 groups ofneck spines (4-7) 10 preanal pores (12) 12 scales onfourth finger (9-10) 19-21 scales on fourth toe (15-17)different relative length of fingers and toes longer hindlegs Distinguished from Agama agama by very smallersize and very different colour pattern and sexual dimor-phism (metallic blue body and bright orange on heads ofadult males) Distinguished from Agama boulengeri byits smaller size very different colour pattern (speciallysexual dimorphism) fewer number of scales aroundmidbody smooth head scales higher number of groupof spines on the sides of the head and shoulder and by ab-sence of caudal crest
DESCRIPTION OF THE HOLOTYPE
An adult male with head scales smooth occipitalscale big dorsal scales homogeneous large rhomboidalkeeled and mucronate all keels converging towards thevertebral line keels on arms and limbs mucronate throatplicate with two gular folds and one mite pocket on eachside of the neck ventral scales smooth relative fingerlength 3gt4gt5=2gt1 tail twice as long as SVL base of thetail weakly compressed tail scales keeled and mucro-nate nostrils oriented upwards and backwards
Measurements (mm) Snout-vent length 7488 taillength 11670 head length 1984 head width 1599
30
TABLE 1 Linear body measurements (mm) and scalation count characters of the type series of Agama castroviejoi sp nov Seemethodology for abbreviations
Linear body MNCN MNCN MNCN MNCN MNCN Range measurement 41779 41777 41778 41776 41780and dimension (male) (juv female) (female) (juv female) (female)
SVL 7421 5991 7488 5931 6760 5931-7488TAL 11640 1051 - - 9017 9017-1164TAH 881 692 725 675 687 692-881TAW 1146 820 1055 813 984 813-1146RTAL 6375 570 - - 7497 570-7497HL 1984 1716 1965 1591 1840 1591-1984HW 1599 1358 1567 1321 1511 1321-1599HH 969 888 967 867 986 867-969EN 476 30 398 311 435 30-476EE 601 437 480 461 533 437-601HAND 1455 1204 1280 1153 1414 1153-1455FOOT 2364 1969 2104 1882 2261 1882-2364FINGER 677 623 575 519 667 519-677TOE 1202 964 975 920 1174 920-1202TL 2121 2031 2175 1977 2134 1977-2175FL 1936 1758 1986 1544 2029 1544-2029HLSVL 027 029 026 027 027 026-029HBSVL 022 023 021 022 022 021-023HLHB 124 126 125 120 122 120-126TLSVL 029 034 029 033 032 029-034FLSVL 026 029 027 026 030 026-030ENSVL 024 017 020 020 024 017-024EESVL 030 025 024 029 029 024-030TOESVL 016 016 013 016 017 013-017FOOTSVL 032 033 028 032 033 028-033
Scalation countcharacters
VE 63 59 61 58 59 58-63EMB 61 58 55 60 63 55-63SL 10 11 11 11 12 10-12IL 9 11 11 11 12 9-12SO 6 7 6 7 7 6-7RSLE 4 3 4 4 4 3-4GNS 9 8 8 8 9 8-9IP 10 - - - - -LFF 12 12 12 12 12 -LFT 19 19 21 19 20 19-21SC 6 6 7 7 7 6-7
NEW AGAMA SPECIES FROM MAURITANIA
head height 969 hand length 1455 (from posterior endof wrist to top of longest finger) foot length 2364 (fromposterior end of heel to top of longest toe) limb length1936 tibia length 2121 length of fourth finger (to joinwith third) 677 length of fourth toe 1202 eye-nostrillength (from the anterior border of the eye orbit to thenostril) 476 ear-eye length (from the anterior border ofthe tympanum to the posterior border of the eye orbit)601 tail width 1146 tail height (at the base) 881
Pholidosis 63 ventral scales between inguinal foldand the beginning of the arm 61 scales around midbody11 supralabials 9 infralabials 6 rows of supraoculars 4
rows of scales between supralabials and suboculars (in-cluding the subocular row) 9 groups of spines betweenthe anterior margin of the ear opening and the shoulderIn each group the central spines are largest 10 analpores 12 lamellae under fourth finger 19 lamellae un-der fourth toe nuchal crest composed of 6 spines
Colour in life Ventral regions of the body tail andextremities immaculate white dorsal surfaces grey-blue with dark brown subocular and labial bars gularregion with brown irregular longitudinal lines sometransverse dark brown bands on the back a perpendicu-lar thin beige band in the anterior margin of the back
31
J M PADIAL
TABLE 2 Diagnostic characters for Agama castroviejoi sp nov Agama impalearis and A boueti
A castroviejoi A impalearis A boueti
SVL 676-7488 to 120 to 99TL 9017-1164 150 to 160EMB 55-63 50-62Dorsal scales keeled keeled keeledHead scales smooth smooth smoothNuchal crest small strong smallGNS 8-9 3-9 4-8Caudal crest absent present absent
Relative length 3gt4gt5gt2gt1 3gt4gt5gt2gt1 3gt4gt2gt5gt1of fingersRelative length 4gt3gt5gt2gt1 3=4gt5gt2gt1 3gt4gt5gt2gt1of toes
Gular region brown lines brown lines white to yellowSexual dicromatism low low lowSociability solitary solitary solitaryHabitat desert rocky areas semidesert rocky areas sandy savannah
TABLE 3 Some scalation counts characters of male (holotype) of Agama castroviejoi and two geographical forms of Aimpalearis (a) southern form (Tan Tan Morocco) (b) northern form (Chefchauen Morocco) See methodology for abbreviations
SL IL GNS LFF LFT IP
Holotype 10 9 9 12 19 10a (n=6) 11-13 10-15 10-11 14-16 16-18 12-14b (n=4) 11-13 11-13 10-11 15 16-19 12-13
and one in the posterior margin two beige irregular cir-cumferences in the middle of the back
Colour in alcohol Ventral regions of the body tailand extremities immaculate white gular region withbrown irregular longitudinal lines all dorsal and dorso-lateral regions grey-blue (slate grey) with some lightbrown spot on the head back and forearms hands yel-low a perpendicular thin beige band in the anteriormargin of the back and one in the posterior margin twobeige irregular circumferences in the middle of the back
VARIATION
For variation in morphometrics see Table 1 The onlymales has more ventral scales and fewer labial scalesthan females The dorsal scales of the male are moremucronate than in females There is colour differencebetween sexes females being paler than males In lifefemales have almost no colour differences they havelight brown head with dark brown subocular stripes thedorsal region is beige to light brown with transversedark brown bands from the scapular region to the middleof the tail each dark band is composed of 3-7 scalesthey have a thin longitudinal light stripe The ventral re-gion is immaculate white with subgular longitudinalbrown stripes In alcohol the colour becomes faded butthe pattern is retained the feet and hands become lightyellow
From the genetic point of view there is a single basepair difference (and A-G transition) between specimenMNCN 41778 and specimens MNCN 41776 MNCN41777 MNCN 41779 in the 464 of the 16S rRNA mito-chondrial region sequenced for this study (Fig 9)
DISTRIBUTION AND ECOLOGY
Specimens are only known from the type localityThey were found on a tableland (dahr) 679 m above sealevel in the Adrar region in Mauritania This region ispart of the ldquoSahara Meridional Occidentalrdquo and is char-acterized by annual rainfall of about 100 mm andaverage annual temperatures of 30degC with minimumtemperatures in January rarely descending below 10degC(Le Houeacuterou 1990) The habitat is a rocky plain withlittle vegetation composed of sparse bushes and somesmall trees (Acacia spp) All specimens were foundalone and separate by few kilometres from each otherThe first specimen encountered was active at 0920 hr(air temperature of 286degC) The other specimens werefound perching around the middle of the day and wereeasily caught when they moved under solitary stones
DISCUSSION
Among the four Agama species (A agama Aboulengeri A boueti A impalearis) inhabiting the aridand semi-arid regions of NW Africa the last two show
32
superficial resemblance with A castroviejoi Neverthe-less morphological quantitative and meristic charactersdistinguish A castroviejoi well from the other speciesrecorded for Mauritania (A boulengeri A agama andA boueti) and also from the two Moroccan forms of Aimpalearis MtDNA analyses demonstrate the identityof A castroviejoi and its close relationship to Aimpalearis and the vicariant form from south and east ofthe Atlas mountains
Furthermore distributional social and ecologicalfactors also allow a clear separation among species Acastroviejoi is solitary and inhabits extremely dry rockyareas of the Adrar plateau Agama boueti is solitary andinhabits the semi-desert sandy plains and savannahs ofthe Sahel region of southern Mauritania and the AtlanticCoast Agama impalearis is a typical solitary rockdweller (Schleich et al 1996) Its distribution rangesfrom the Mediterranean coast (Bons amp Geniez 1996) toSeguia el Hamra in Western Sahara (Geniez et al2000) Although it was cited for ldquoMauritanieSahariennerdquo (Chabanaud 1917) and despite appearingwidely distributed across the country in Le Berrersquos(1989) distribution map for this species there are actu-ally no confirmed records for Mauritania MoreoverJoger (1979) noted that this species did not occur in theAdrar and that the nearest record corresponded toSeguia el Hamra The presence of this species in thecountry is still plausible in rocky areas similar to thoseof the Zemmour Mountains (Geniez et al 2000) butthis requires confirmation A boulengeri is a speciesfrom the Sahel Savannah that inhabits rocky areas andforms social groups We found this species as far northas Oued Choucircm (21deg 226N12deg 586W) near the bor-der with Western Sahara At this latitude it is alwaysassociated to wet rocky gorges (Gueltas) in the moun-tains A agama is another social species reported to beassociated with human settlements in the Adrar region(Dekeyser amp Villiers 1956) In the south of the countrythis species prefers trees from the Sahel savannahs but itshows high habitat plasticity (pers obs)
Miocene vicariance events have been found to be re-sponsible of the variation and differentiation in Aimpalearis in NW Africa (Brown amp Znari 1998 Brownet al 2002) The uplift of the Atlas mountains consti-tuted a great geographical barrier causing allopatricfragmentation of the populations A castroviejoi is iso-lated in a rocky tableland surrounded by lowland sandyplains of the Sahara that could serve as a barrier for arock dweller Also in situ selection-mediated responsesto the ecological current conditions of the isolatedpopulations are responsible of the morphological varia-tion of A impalearis (Brown amp Znari 1998) Presentecological conditions of the Adrar are very extreme andis probably one of the hottest areas of the Sahara (LeHoueacuterou 1990) As demonstrated by Brown amp Znari(1998) variation in scalation is significantly associatedwith different temperature conditions Therefore bothisolation and ecological differentiation could be impli-cated in the differentiation of A castroviejoi
Although the Adrar belongs to the Meridional Sahara(Le Houeacuterou 1990) this area shows a complex historywith its faunas and floras comprising a mixture of Medi-terranean Saharan and Sahelian species (Dekeyser ampVilliers 1956) Probably A castroviejoi constitutes anexample of a Saharan endemic of the Adrar Mountainsas some invertebrate species and also vertebrate subspe-cies (Dekeyser amp Villiers 1956) Nevertheless thephylogeographic affinities of A castroviejoi remain un-clear since it could have a Sahelian or Saharan origin Amore thorough phylogenetic comparison among NorthAfrican agamas will help to resolve this question
ACKNOWLEDGEMENTS
This work could not have been carried out withoutthe effort of J Castroviejo and the people of AsociacioacutenAmigos de Dontildeana I am very gratefully to S Carranza(BMNH) who did the genetic work and helped me toimprove the manuscript To I de la Riva (MNCN) forhis help in many aspects of this work To Dr NAnderson (BMNH) who revised the draft version andthe English of the manuscript To R Garciacutea M UrceraC Carballo M O Deida and E Mohamed O Saleh fortheir help and companion in Mauritania and J Peacuterez EAvila S Castroviejo-Fisher and A Quintana for theirfriendship and companion in the first trip to MauritaniaP Geniez J A Mateo H Nickel and W Boumlhme allhelped us in some aspect of this work W Wuster andtwo anonymous reviewers critically commented an ear-lier draft of the manuscript R Brown kindly providedraw scalation data for A impalearis from Morocco andcontributed to the improvement of the manuscript
REFERENCES
Boumlhme W (2000) Die wechselvolle Geschichte derSahara Untersuchung von Reptilienzoumlnosen entlangeines westsaharischen Transektes mit einemuumlberraschenden Fund im Suumldosten Mauritaniens Tierund Museum 7 11-21
Boumlhme W Meinig H amp Roumldel M A (1996) Newrecords of amphibians and reptiles from Burkina Fasoand Mali British Herpetological Society Bulletin 56 7-26
Boumlhme W Wilms T Nickel H amp Merz M (2001)Bericht uumlber eine herpetologische Forschungsreise indie westliche Sahara und ihre Randgebiete Zeitschriftdes Koumllner Zoo 44 117-131
Bons J amp Geniez P (1996) Anfibios y reptiles deMarruecos (Incluido Saacutehara Occidental) Atlasbiogeograacutefico Barcelona Asociacioacuten HerpetoloacutegicaEspantildeola
Brown R P amp Znari M (1998) Morphological variationin Agama impalearis from Morocco evidence forhistorical population vicariance and current climaticeffects Ecography 21 605-612
Brown R P Znari M El Mouden E H amp Harris P(1999) Estimating asymptotic size and geographicvariation in Agama impalearis from MoroccoEcography 22 277-284
NEW AGAMA SPECIES FROM MAURITANIA 33
J M PADIAL
Accepted 22304
Brown R P Pestano J amp Suaacuterez N M (2002) TheAtlas mountains as a biogeographical divide in North-West Africa evidence from mtDNA evolution in theAgamid lizard Agama impalearis MolecularPhylogenetics and Evolution 24 324-332
Carranza S Arnold E N Thomas R H Mateo J ampLopez-Jurado L F (1999) Status of the extinct giantlacertid lizard Gallotia simonyi simonyi (ReptiliaLacertidae) assessed using mtDNA sequences frommuseum specimens Herpetological Journal l9 83-86
Carranza S Arnold E N Mateo J amp Lopez-Jurado LF (2000) Long-distance colonization and radiation ingekkonid lizards Tarentola (Reptilia Gekkonidae)revealed by mitochondrial DNA sequencesProceedings of the Royal Society Series B 267 637-649
Chabanaud P (1917) Enumeacuteration des reptiles nonencore eacutetudieacutes de lrsquoAfrique occidentale appartenantaux collections du Museacuteum avec la description desespeacuteces nouvelles Bulletin du Museacuteum NationaldrsquoHistoire Naturelle de Paris Serie Zoologique 2383-105
Dekeyser P L amp Villiers A (1956) Contribution alrsquoeacutetude du peuplement de la Mauritanie Notationseacutecologiques et biogeacuteographiques sur la faune delrsquoAdrar Memoires de lrsquoInstitut Franccedilais de lrsquoAfriqueNoire 44 222 p 25 pl
Felsenstein J (1985) Confidence-limits on phylogenies ndashan approach using the bootstrap Evolution 39 783-791
Geniez Ph amp Foucart A (1995) Un nouvelAcanthodactyle en Algeacuterie Acanthodactylustaghitensis n sp (Reptilia Sauria Lacertidae)Bulletin du Museacuteum National drsquoHistoire NaturelleParis 4deg seacuterie 17 3-9
Geniez P Mateo J A amp Bons J (2000) A checklist ofthe amphibians and reptiles of Western Sahara(Amphibia Reptilia) Herpetozoa 13 149-163
Ineich I (1997) Les amphibiens et reptiles du littoralmauritanien In Environnement et littoralmauritanien 93-99 Colas P (Ed) MontpellierCIRAD
Joger U (1979) Zur Oumlkologie und Verbreitung wenigbekannter Agamen Westafrikas (Reptilia SauriaAgamidae) Salamandra 15 31-52
Joger U (1980) Eine neue Art der Gattung Tarentola(Reptilia Sauria Gekkonidae) aus WestafrikaAmphibia-Reptilia 1 137-147
Joger U (1981) Zur Herpetofaunistik WestafrikasBonner zoologische Beitraumlge 32 297-340
Joger U (1984) Taxonomische Revision der GattungTarentola (Reptilia Gekkonidae) Bonner zoologischeBeitraumlge 35 129-174
Joger U amp Lambert M R K (1996) Analysis of theherpetofauna of the Republic of Mali I Annotatedinventory with description of a new Uromastyx(Sauria Agamidae) Journal of African Zoology 11021-51
Lambert M R K amp Mullieacute W C (1998) Sexualdichromatism of Agama boulengeri observed insouthern Mauritania British Herpetological SocietyBulletin 65 42-44
Le Berre M (1989) Faune du Sahara I Poissons-Amphibiens-Reptiles Paris Lechevalier-R Chabaud
Le Houeacuterou H N (1990) Deacutefinition et limitesbioclimatiques du Sahara Segravecheresse 1 246-259
Lenk P Kalyabina S Wink M amp Joger U (2001)Evolutionary relationships among the true vipers(Reptilia Viperidae) inferred from mitochondrial DNAsequences Molecular Biology and Evolution 19 94-104
Mateo J A Geniez Ph Loacutepez-Jurado J L amp Bons J(1998) Chorological analysis and morphologicalvariation of saurians of the genus Uromastyx (ReptiliaAgamidae) in western Sahara Description of two newtaxa Revista Espantildeola de Herpetologiacutea 12 97-109
Salvador A (1982) A revision of the lizards of the genusAcanthodactylus (Sauria Lacertidae) Bonnerzoologische Beitraumlge Monographien 16167 p
Schleich H H Kaumlstle W amp Kabisch K (1996)Amphibians and reptiles of North Africa KoenigsteinKoeltz Scientific Publications
Swofford D L (2002) PAUP phylogenetic analysisusing parsimony (and others methods) v40b10Sinauer Associates Sunderland MA
Thompson J D Gibson T J Plewniak F JeanmouginF amp Higgins D G (1997) The ClustalX windowsinterface flexible strategies for multiple sequencealignment aided by quality analysis tools NucleicAcids Research 24 4876-4882
Wilms T amp Boumlhme W (2001) Revision der Uromastyxacanthinura ndash Artengruppe mit Beschreibung einerneuen Art aus der Zentralsahara (Reptilia SauriaAgamidae) Zoologische Abhandlungen StaatlichesMuseum fuumlr Tierkunde Dresden 51 73-104
34
APPENDIX 1
Additional specimens of Agama species examinedfor this work EBD Estacioacuten Bioloacutegica de Dontildeana Se-ville Spain CET Centro de Estudios TropicalesSevilla Spain Locality and coordinates in parentheses
Agama agama CET(RIM)-028 (near Bouli 15deg25803 N11deg 55562 W Mauritania) CET(RIM)-328-329 (El Wad-Zoueina15deg 42689 N 9deg 39906 WMauritania) CET(RIM)-331-335 (El Wad-Foulania15deg 31682 N9deg 48896 W Mauritania)
Agama boueti CET(RIM)-287-291 336 (Ayoucircn elAtroucircs 16deg 37412 N9deg 37441 W Mauritania)
Agama boulengeri CET(RIM)-016-018 (Bou-gari16deg 32034 N10deg 47892 W Mauritania)CET(RIM)-021-022 (between Timbedgha and AyoucircnEl Atroucircs 16deg 26890 N9deg 14690 W Mauritania)CET(RIM)-066 (Terjicirct 20deg 15578 N13 deg 05854 WMauritania) CET(RIM)-094-96 (Guelta Molomhar20deg 34873 N13deg 07630 W Mauritania) CET(RIM)-116-117 (Oued Choucircm 21deg 22654 N12deg 58581 WMauritania) CET(RIM)-174-175 (Zig between Lekh-cheb and Ticircchicirct 18deg 34487 N9deg 48379 WMauritania) CET(RIM)-256-261 (Guelta Oumm Leb-are 16deg 29472 N10deg 49822 W Mauritania)CET(RIM)-338 (Ayoucircn el Atroucircs 16deg 37412 N9deg37441 W Mauritania) CET(RIM)-344 (Guelta Mat-mata17deg 53571 N12deg 07467 W Mauritania)
Agama impalearis EBD-11352 (Mount Marcan-Kbir Tetuaacuten Morocco) EBD-20222 (TaklimMorocco) EBD-22084 (Yassinen Morocco) EBD-24515 (Bab Bou Ichir-Fritissa Morocco) EBD-6552(El Aioum Western Sahara)
NEW AGAMA SPECIES FROM MAURITANIA 35
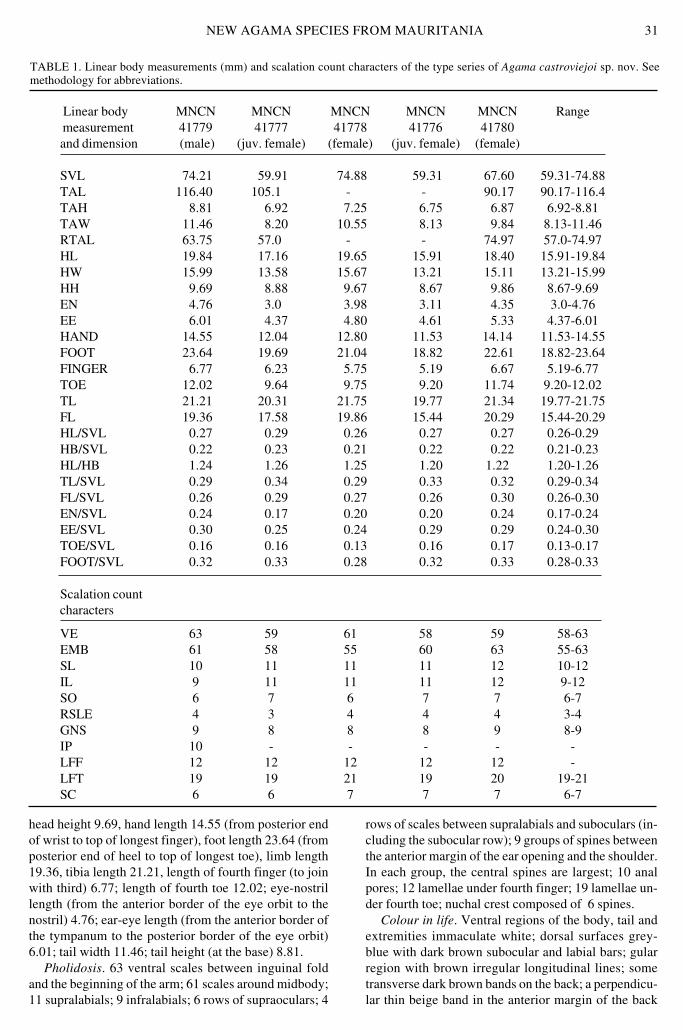
TABLE 1 Linear body measurements (mm) and scalation count characters of the type series of Agama castroviejoi sp nov Seemethodology for abbreviations
Linear body MNCN MNCN MNCN MNCN MNCN Range measurement 41779 41777 41778 41776 41780and dimension (male) (juv female) (female) (juv female) (female)
SVL 7421 5991 7488 5931 6760 5931-7488TAL 11640 1051 - - 9017 9017-1164TAH 881 692 725 675 687 692-881TAW 1146 820 1055 813 984 813-1146RTAL 6375 570 - - 7497 570-7497HL 1984 1716 1965 1591 1840 1591-1984HW 1599 1358 1567 1321 1511 1321-1599HH 969 888 967 867 986 867-969EN 476 30 398 311 435 30-476EE 601 437 480 461 533 437-601HAND 1455 1204 1280 1153 1414 1153-1455FOOT 2364 1969 2104 1882 2261 1882-2364FINGER 677 623 575 519 667 519-677TOE 1202 964 975 920 1174 920-1202TL 2121 2031 2175 1977 2134 1977-2175FL 1936 1758 1986 1544 2029 1544-2029HLSVL 027 029 026 027 027 026-029HBSVL 022 023 021 022 022 021-023HLHB 124 126 125 120 122 120-126TLSVL 029 034 029 033 032 029-034FLSVL 026 029 027 026 030 026-030ENSVL 024 017 020 020 024 017-024EESVL 030 025 024 029 029 024-030TOESVL 016 016 013 016 017 013-017FOOTSVL 032 033 028 032 033 028-033
Scalation countcharacters
VE 63 59 61 58 59 58-63EMB 61 58 55 60 63 55-63SL 10 11 11 11 12 10-12IL 9 11 11 11 12 9-12SO 6 7 6 7 7 6-7RSLE 4 3 4 4 4 3-4GNS 9 8 8 8 9 8-9IP 10 - - - - -LFF 12 12 12 12 12 -LFT 19 19 21 19 20 19-21SC 6 6 7 7 7 6-7
NEW AGAMA SPECIES FROM MAURITANIA
head height 969 hand length 1455 (from posterior endof wrist to top of longest finger) foot length 2364 (fromposterior end of heel to top of longest toe) limb length1936 tibia length 2121 length of fourth finger (to joinwith third) 677 length of fourth toe 1202 eye-nostrillength (from the anterior border of the eye orbit to thenostril) 476 ear-eye length (from the anterior border ofthe tympanum to the posterior border of the eye orbit)601 tail width 1146 tail height (at the base) 881
Pholidosis 63 ventral scales between inguinal foldand the beginning of the arm 61 scales around midbody11 supralabials 9 infralabials 6 rows of supraoculars 4
rows of scales between supralabials and suboculars (in-cluding the subocular row) 9 groups of spines betweenthe anterior margin of the ear opening and the shoulderIn each group the central spines are largest 10 analpores 12 lamellae under fourth finger 19 lamellae un-der fourth toe nuchal crest composed of 6 spines
Colour in life Ventral regions of the body tail andextremities immaculate white dorsal surfaces grey-blue with dark brown subocular and labial bars gularregion with brown irregular longitudinal lines sometransverse dark brown bands on the back a perpendicu-lar thin beige band in the anterior margin of the back
31
J M PADIAL
TABLE 2 Diagnostic characters for Agama castroviejoi sp nov Agama impalearis and A boueti
A castroviejoi A impalearis A boueti
SVL 676-7488 to 120 to 99TL 9017-1164 150 to 160EMB 55-63 50-62Dorsal scales keeled keeled keeledHead scales smooth smooth smoothNuchal crest small strong smallGNS 8-9 3-9 4-8Caudal crest absent present absent
Relative length 3gt4gt5gt2gt1 3gt4gt5gt2gt1 3gt4gt2gt5gt1of fingersRelative length 4gt3gt5gt2gt1 3=4gt5gt2gt1 3gt4gt5gt2gt1of toes
Gular region brown lines brown lines white to yellowSexual dicromatism low low lowSociability solitary solitary solitaryHabitat desert rocky areas semidesert rocky areas sandy savannah
TABLE 3 Some scalation counts characters of male (holotype) of Agama castroviejoi and two geographical forms of Aimpalearis (a) southern form (Tan Tan Morocco) (b) northern form (Chefchauen Morocco) See methodology for abbreviations
SL IL GNS LFF LFT IP
Holotype 10 9 9 12 19 10a (n=6) 11-13 10-15 10-11 14-16 16-18 12-14b (n=4) 11-13 11-13 10-11 15 16-19 12-13
and one in the posterior margin two beige irregular cir-cumferences in the middle of the back
Colour in alcohol Ventral regions of the body tailand extremities immaculate white gular region withbrown irregular longitudinal lines all dorsal and dorso-lateral regions grey-blue (slate grey) with some lightbrown spot on the head back and forearms hands yel-low a perpendicular thin beige band in the anteriormargin of the back and one in the posterior margin twobeige irregular circumferences in the middle of the back
VARIATION
For variation in morphometrics see Table 1 The onlymales has more ventral scales and fewer labial scalesthan females The dorsal scales of the male are moremucronate than in females There is colour differencebetween sexes females being paler than males In lifefemales have almost no colour differences they havelight brown head with dark brown subocular stripes thedorsal region is beige to light brown with transversedark brown bands from the scapular region to the middleof the tail each dark band is composed of 3-7 scalesthey have a thin longitudinal light stripe The ventral re-gion is immaculate white with subgular longitudinalbrown stripes In alcohol the colour becomes faded butthe pattern is retained the feet and hands become lightyellow
From the genetic point of view there is a single basepair difference (and A-G transition) between specimenMNCN 41778 and specimens MNCN 41776 MNCN41777 MNCN 41779 in the 464 of the 16S rRNA mito-chondrial region sequenced for this study (Fig 9)
DISTRIBUTION AND ECOLOGY
Specimens are only known from the type localityThey were found on a tableland (dahr) 679 m above sealevel in the Adrar region in Mauritania This region ispart of the ldquoSahara Meridional Occidentalrdquo and is char-acterized by annual rainfall of about 100 mm andaverage annual temperatures of 30degC with minimumtemperatures in January rarely descending below 10degC(Le Houeacuterou 1990) The habitat is a rocky plain withlittle vegetation composed of sparse bushes and somesmall trees (Acacia spp) All specimens were foundalone and separate by few kilometres from each otherThe first specimen encountered was active at 0920 hr(air temperature of 286degC) The other specimens werefound perching around the middle of the day and wereeasily caught when they moved under solitary stones
DISCUSSION
Among the four Agama species (A agama Aboulengeri A boueti A impalearis) inhabiting the aridand semi-arid regions of NW Africa the last two show
32
superficial resemblance with A castroviejoi Neverthe-less morphological quantitative and meristic charactersdistinguish A castroviejoi well from the other speciesrecorded for Mauritania (A boulengeri A agama andA boueti) and also from the two Moroccan forms of Aimpalearis MtDNA analyses demonstrate the identityof A castroviejoi and its close relationship to Aimpalearis and the vicariant form from south and east ofthe Atlas mountains
Furthermore distributional social and ecologicalfactors also allow a clear separation among species Acastroviejoi is solitary and inhabits extremely dry rockyareas of the Adrar plateau Agama boueti is solitary andinhabits the semi-desert sandy plains and savannahs ofthe Sahel region of southern Mauritania and the AtlanticCoast Agama impalearis is a typical solitary rockdweller (Schleich et al 1996) Its distribution rangesfrom the Mediterranean coast (Bons amp Geniez 1996) toSeguia el Hamra in Western Sahara (Geniez et al2000) Although it was cited for ldquoMauritanieSahariennerdquo (Chabanaud 1917) and despite appearingwidely distributed across the country in Le Berrersquos(1989) distribution map for this species there are actu-ally no confirmed records for Mauritania MoreoverJoger (1979) noted that this species did not occur in theAdrar and that the nearest record corresponded toSeguia el Hamra The presence of this species in thecountry is still plausible in rocky areas similar to thoseof the Zemmour Mountains (Geniez et al 2000) butthis requires confirmation A boulengeri is a speciesfrom the Sahel Savannah that inhabits rocky areas andforms social groups We found this species as far northas Oued Choucircm (21deg 226N12deg 586W) near the bor-der with Western Sahara At this latitude it is alwaysassociated to wet rocky gorges (Gueltas) in the moun-tains A agama is another social species reported to beassociated with human settlements in the Adrar region(Dekeyser amp Villiers 1956) In the south of the countrythis species prefers trees from the Sahel savannahs but itshows high habitat plasticity (pers obs)
Miocene vicariance events have been found to be re-sponsible of the variation and differentiation in Aimpalearis in NW Africa (Brown amp Znari 1998 Brownet al 2002) The uplift of the Atlas mountains consti-tuted a great geographical barrier causing allopatricfragmentation of the populations A castroviejoi is iso-lated in a rocky tableland surrounded by lowland sandyplains of the Sahara that could serve as a barrier for arock dweller Also in situ selection-mediated responsesto the ecological current conditions of the isolatedpopulations are responsible of the morphological varia-tion of A impalearis (Brown amp Znari 1998) Presentecological conditions of the Adrar are very extreme andis probably one of the hottest areas of the Sahara (LeHoueacuterou 1990) As demonstrated by Brown amp Znari(1998) variation in scalation is significantly associatedwith different temperature conditions Therefore bothisolation and ecological differentiation could be impli-cated in the differentiation of A castroviejoi
Although the Adrar belongs to the Meridional Sahara(Le Houeacuterou 1990) this area shows a complex historywith its faunas and floras comprising a mixture of Medi-terranean Saharan and Sahelian species (Dekeyser ampVilliers 1956) Probably A castroviejoi constitutes anexample of a Saharan endemic of the Adrar Mountainsas some invertebrate species and also vertebrate subspe-cies (Dekeyser amp Villiers 1956) Nevertheless thephylogeographic affinities of A castroviejoi remain un-clear since it could have a Sahelian or Saharan origin Amore thorough phylogenetic comparison among NorthAfrican agamas will help to resolve this question
ACKNOWLEDGEMENTS
This work could not have been carried out withoutthe effort of J Castroviejo and the people of AsociacioacutenAmigos de Dontildeana I am very gratefully to S Carranza(BMNH) who did the genetic work and helped me toimprove the manuscript To I de la Riva (MNCN) forhis help in many aspects of this work To Dr NAnderson (BMNH) who revised the draft version andthe English of the manuscript To R Garciacutea M UrceraC Carballo M O Deida and E Mohamed O Saleh fortheir help and companion in Mauritania and J Peacuterez EAvila S Castroviejo-Fisher and A Quintana for theirfriendship and companion in the first trip to MauritaniaP Geniez J A Mateo H Nickel and W Boumlhme allhelped us in some aspect of this work W Wuster andtwo anonymous reviewers critically commented an ear-lier draft of the manuscript R Brown kindly providedraw scalation data for A impalearis from Morocco andcontributed to the improvement of the manuscript
REFERENCES
Boumlhme W (2000) Die wechselvolle Geschichte derSahara Untersuchung von Reptilienzoumlnosen entlangeines westsaharischen Transektes mit einemuumlberraschenden Fund im Suumldosten Mauritaniens Tierund Museum 7 11-21
Boumlhme W Meinig H amp Roumldel M A (1996) Newrecords of amphibians and reptiles from Burkina Fasoand Mali British Herpetological Society Bulletin 56 7-26
Boumlhme W Wilms T Nickel H amp Merz M (2001)Bericht uumlber eine herpetologische Forschungsreise indie westliche Sahara und ihre Randgebiete Zeitschriftdes Koumllner Zoo 44 117-131
Bons J amp Geniez P (1996) Anfibios y reptiles deMarruecos (Incluido Saacutehara Occidental) Atlasbiogeograacutefico Barcelona Asociacioacuten HerpetoloacutegicaEspantildeola
Brown R P amp Znari M (1998) Morphological variationin Agama impalearis from Morocco evidence forhistorical population vicariance and current climaticeffects Ecography 21 605-612
Brown R P Znari M El Mouden E H amp Harris P(1999) Estimating asymptotic size and geographicvariation in Agama impalearis from MoroccoEcography 22 277-284
NEW AGAMA SPECIES FROM MAURITANIA 33
J M PADIAL
Accepted 22304
Brown R P Pestano J amp Suaacuterez N M (2002) TheAtlas mountains as a biogeographical divide in North-West Africa evidence from mtDNA evolution in theAgamid lizard Agama impalearis MolecularPhylogenetics and Evolution 24 324-332
Carranza S Arnold E N Thomas R H Mateo J ampLopez-Jurado L F (1999) Status of the extinct giantlacertid lizard Gallotia simonyi simonyi (ReptiliaLacertidae) assessed using mtDNA sequences frommuseum specimens Herpetological Journal l9 83-86
Carranza S Arnold E N Mateo J amp Lopez-Jurado LF (2000) Long-distance colonization and radiation ingekkonid lizards Tarentola (Reptilia Gekkonidae)revealed by mitochondrial DNA sequencesProceedings of the Royal Society Series B 267 637-649
Chabanaud P (1917) Enumeacuteration des reptiles nonencore eacutetudieacutes de lrsquoAfrique occidentale appartenantaux collections du Museacuteum avec la description desespeacuteces nouvelles Bulletin du Museacuteum NationaldrsquoHistoire Naturelle de Paris Serie Zoologique 2383-105
Dekeyser P L amp Villiers A (1956) Contribution alrsquoeacutetude du peuplement de la Mauritanie Notationseacutecologiques et biogeacuteographiques sur la faune delrsquoAdrar Memoires de lrsquoInstitut Franccedilais de lrsquoAfriqueNoire 44 222 p 25 pl
Felsenstein J (1985) Confidence-limits on phylogenies ndashan approach using the bootstrap Evolution 39 783-791
Geniez Ph amp Foucart A (1995) Un nouvelAcanthodactyle en Algeacuterie Acanthodactylustaghitensis n sp (Reptilia Sauria Lacertidae)Bulletin du Museacuteum National drsquoHistoire NaturelleParis 4deg seacuterie 17 3-9
Geniez P Mateo J A amp Bons J (2000) A checklist ofthe amphibians and reptiles of Western Sahara(Amphibia Reptilia) Herpetozoa 13 149-163
Ineich I (1997) Les amphibiens et reptiles du littoralmauritanien In Environnement et littoralmauritanien 93-99 Colas P (Ed) MontpellierCIRAD
Joger U (1979) Zur Oumlkologie und Verbreitung wenigbekannter Agamen Westafrikas (Reptilia SauriaAgamidae) Salamandra 15 31-52
Joger U (1980) Eine neue Art der Gattung Tarentola(Reptilia Sauria Gekkonidae) aus WestafrikaAmphibia-Reptilia 1 137-147
Joger U (1981) Zur Herpetofaunistik WestafrikasBonner zoologische Beitraumlge 32 297-340
Joger U (1984) Taxonomische Revision der GattungTarentola (Reptilia Gekkonidae) Bonner zoologischeBeitraumlge 35 129-174
Joger U amp Lambert M R K (1996) Analysis of theherpetofauna of the Republic of Mali I Annotatedinventory with description of a new Uromastyx(Sauria Agamidae) Journal of African Zoology 11021-51
Lambert M R K amp Mullieacute W C (1998) Sexualdichromatism of Agama boulengeri observed insouthern Mauritania British Herpetological SocietyBulletin 65 42-44
Le Berre M (1989) Faune du Sahara I Poissons-Amphibiens-Reptiles Paris Lechevalier-R Chabaud
Le Houeacuterou H N (1990) Deacutefinition et limitesbioclimatiques du Sahara Segravecheresse 1 246-259
Lenk P Kalyabina S Wink M amp Joger U (2001)Evolutionary relationships among the true vipers(Reptilia Viperidae) inferred from mitochondrial DNAsequences Molecular Biology and Evolution 19 94-104
Mateo J A Geniez Ph Loacutepez-Jurado J L amp Bons J(1998) Chorological analysis and morphologicalvariation of saurians of the genus Uromastyx (ReptiliaAgamidae) in western Sahara Description of two newtaxa Revista Espantildeola de Herpetologiacutea 12 97-109
Salvador A (1982) A revision of the lizards of the genusAcanthodactylus (Sauria Lacertidae) Bonnerzoologische Beitraumlge Monographien 16167 p
Schleich H H Kaumlstle W amp Kabisch K (1996)Amphibians and reptiles of North Africa KoenigsteinKoeltz Scientific Publications
Swofford D L (2002) PAUP phylogenetic analysisusing parsimony (and others methods) v40b10Sinauer Associates Sunderland MA
Thompson J D Gibson T J Plewniak F JeanmouginF amp Higgins D G (1997) The ClustalX windowsinterface flexible strategies for multiple sequencealignment aided by quality analysis tools NucleicAcids Research 24 4876-4882
Wilms T amp Boumlhme W (2001) Revision der Uromastyxacanthinura ndash Artengruppe mit Beschreibung einerneuen Art aus der Zentralsahara (Reptilia SauriaAgamidae) Zoologische Abhandlungen StaatlichesMuseum fuumlr Tierkunde Dresden 51 73-104
34
APPENDIX 1
Additional specimens of Agama species examinedfor this work EBD Estacioacuten Bioloacutegica de Dontildeana Se-ville Spain CET Centro de Estudios TropicalesSevilla Spain Locality and coordinates in parentheses
Agama agama CET(RIM)-028 (near Bouli 15deg25803 N11deg 55562 W Mauritania) CET(RIM)-328-329 (El Wad-Zoueina15deg 42689 N 9deg 39906 WMauritania) CET(RIM)-331-335 (El Wad-Foulania15deg 31682 N9deg 48896 W Mauritania)
Agama boueti CET(RIM)-287-291 336 (Ayoucircn elAtroucircs 16deg 37412 N9deg 37441 W Mauritania)
Agama boulengeri CET(RIM)-016-018 (Bou-gari16deg 32034 N10deg 47892 W Mauritania)CET(RIM)-021-022 (between Timbedgha and AyoucircnEl Atroucircs 16deg 26890 N9deg 14690 W Mauritania)CET(RIM)-066 (Terjicirct 20deg 15578 N13 deg 05854 WMauritania) CET(RIM)-094-96 (Guelta Molomhar20deg 34873 N13deg 07630 W Mauritania) CET(RIM)-116-117 (Oued Choucircm 21deg 22654 N12deg 58581 WMauritania) CET(RIM)-174-175 (Zig between Lekh-cheb and Ticircchicirct 18deg 34487 N9deg 48379 WMauritania) CET(RIM)-256-261 (Guelta Oumm Leb-are 16deg 29472 N10deg 49822 W Mauritania)CET(RIM)-338 (Ayoucircn el Atroucircs 16deg 37412 N9deg37441 W Mauritania) CET(RIM)-344 (Guelta Mat-mata17deg 53571 N12deg 07467 W Mauritania)
Agama impalearis EBD-11352 (Mount Marcan-Kbir Tetuaacuten Morocco) EBD-20222 (TaklimMorocco) EBD-22084 (Yassinen Morocco) EBD-24515 (Bab Bou Ichir-Fritissa Morocco) EBD-6552(El Aioum Western Sahara)
NEW AGAMA SPECIES FROM MAURITANIA 35

J M PADIAL
TABLE 2 Diagnostic characters for Agama castroviejoi sp nov Agama impalearis and A boueti
A castroviejoi A impalearis A boueti
SVL 676-7488 to 120 to 99TL 9017-1164 150 to 160EMB 55-63 50-62Dorsal scales keeled keeled keeledHead scales smooth smooth smoothNuchal crest small strong smallGNS 8-9 3-9 4-8Caudal crest absent present absent
Relative length 3gt4gt5gt2gt1 3gt4gt5gt2gt1 3gt4gt2gt5gt1of fingersRelative length 4gt3gt5gt2gt1 3=4gt5gt2gt1 3gt4gt5gt2gt1of toes
Gular region brown lines brown lines white to yellowSexual dicromatism low low lowSociability solitary solitary solitaryHabitat desert rocky areas semidesert rocky areas sandy savannah
TABLE 3 Some scalation counts characters of male (holotype) of Agama castroviejoi and two geographical forms of Aimpalearis (a) southern form (Tan Tan Morocco) (b) northern form (Chefchauen Morocco) See methodology for abbreviations
SL IL GNS LFF LFT IP
Holotype 10 9 9 12 19 10a (n=6) 11-13 10-15 10-11 14-16 16-18 12-14b (n=4) 11-13 11-13 10-11 15 16-19 12-13
and one in the posterior margin two beige irregular cir-cumferences in the middle of the back
Colour in alcohol Ventral regions of the body tailand extremities immaculate white gular region withbrown irregular longitudinal lines all dorsal and dorso-lateral regions grey-blue (slate grey) with some lightbrown spot on the head back and forearms hands yel-low a perpendicular thin beige band in the anteriormargin of the back and one in the posterior margin twobeige irregular circumferences in the middle of the back
VARIATION
For variation in morphometrics see Table 1 The onlymales has more ventral scales and fewer labial scalesthan females The dorsal scales of the male are moremucronate than in females There is colour differencebetween sexes females being paler than males In lifefemales have almost no colour differences they havelight brown head with dark brown subocular stripes thedorsal region is beige to light brown with transversedark brown bands from the scapular region to the middleof the tail each dark band is composed of 3-7 scalesthey have a thin longitudinal light stripe The ventral re-gion is immaculate white with subgular longitudinalbrown stripes In alcohol the colour becomes faded butthe pattern is retained the feet and hands become lightyellow
From the genetic point of view there is a single basepair difference (and A-G transition) between specimenMNCN 41778 and specimens MNCN 41776 MNCN41777 MNCN 41779 in the 464 of the 16S rRNA mito-chondrial region sequenced for this study (Fig 9)
DISTRIBUTION AND ECOLOGY
Specimens are only known from the type localityThey were found on a tableland (dahr) 679 m above sealevel in the Adrar region in Mauritania This region ispart of the ldquoSahara Meridional Occidentalrdquo and is char-acterized by annual rainfall of about 100 mm andaverage annual temperatures of 30degC with minimumtemperatures in January rarely descending below 10degC(Le Houeacuterou 1990) The habitat is a rocky plain withlittle vegetation composed of sparse bushes and somesmall trees (Acacia spp) All specimens were foundalone and separate by few kilometres from each otherThe first specimen encountered was active at 0920 hr(air temperature of 286degC) The other specimens werefound perching around the middle of the day and wereeasily caught when they moved under solitary stones
DISCUSSION
Among the four Agama species (A agama Aboulengeri A boueti A impalearis) inhabiting the aridand semi-arid regions of NW Africa the last two show
32
superficial resemblance with A castroviejoi Neverthe-less morphological quantitative and meristic charactersdistinguish A castroviejoi well from the other speciesrecorded for Mauritania (A boulengeri A agama andA boueti) and also from the two Moroccan forms of Aimpalearis MtDNA analyses demonstrate the identityof A castroviejoi and its close relationship to Aimpalearis and the vicariant form from south and east ofthe Atlas mountains
Furthermore distributional social and ecologicalfactors also allow a clear separation among species Acastroviejoi is solitary and inhabits extremely dry rockyareas of the Adrar plateau Agama boueti is solitary andinhabits the semi-desert sandy plains and savannahs ofthe Sahel region of southern Mauritania and the AtlanticCoast Agama impalearis is a typical solitary rockdweller (Schleich et al 1996) Its distribution rangesfrom the Mediterranean coast (Bons amp Geniez 1996) toSeguia el Hamra in Western Sahara (Geniez et al2000) Although it was cited for ldquoMauritanieSahariennerdquo (Chabanaud 1917) and despite appearingwidely distributed across the country in Le Berrersquos(1989) distribution map for this species there are actu-ally no confirmed records for Mauritania MoreoverJoger (1979) noted that this species did not occur in theAdrar and that the nearest record corresponded toSeguia el Hamra The presence of this species in thecountry is still plausible in rocky areas similar to thoseof the Zemmour Mountains (Geniez et al 2000) butthis requires confirmation A boulengeri is a speciesfrom the Sahel Savannah that inhabits rocky areas andforms social groups We found this species as far northas Oued Choucircm (21deg 226N12deg 586W) near the bor-der with Western Sahara At this latitude it is alwaysassociated to wet rocky gorges (Gueltas) in the moun-tains A agama is another social species reported to beassociated with human settlements in the Adrar region(Dekeyser amp Villiers 1956) In the south of the countrythis species prefers trees from the Sahel savannahs but itshows high habitat plasticity (pers obs)
Miocene vicariance events have been found to be re-sponsible of the variation and differentiation in Aimpalearis in NW Africa (Brown amp Znari 1998 Brownet al 2002) The uplift of the Atlas mountains consti-tuted a great geographical barrier causing allopatricfragmentation of the populations A castroviejoi is iso-lated in a rocky tableland surrounded by lowland sandyplains of the Sahara that could serve as a barrier for arock dweller Also in situ selection-mediated responsesto the ecological current conditions of the isolatedpopulations are responsible of the morphological varia-tion of A impalearis (Brown amp Znari 1998) Presentecological conditions of the Adrar are very extreme andis probably one of the hottest areas of the Sahara (LeHoueacuterou 1990) As demonstrated by Brown amp Znari(1998) variation in scalation is significantly associatedwith different temperature conditions Therefore bothisolation and ecological differentiation could be impli-cated in the differentiation of A castroviejoi
Although the Adrar belongs to the Meridional Sahara(Le Houeacuterou 1990) this area shows a complex historywith its faunas and floras comprising a mixture of Medi-terranean Saharan and Sahelian species (Dekeyser ampVilliers 1956) Probably A castroviejoi constitutes anexample of a Saharan endemic of the Adrar Mountainsas some invertebrate species and also vertebrate subspe-cies (Dekeyser amp Villiers 1956) Nevertheless thephylogeographic affinities of A castroviejoi remain un-clear since it could have a Sahelian or Saharan origin Amore thorough phylogenetic comparison among NorthAfrican agamas will help to resolve this question
ACKNOWLEDGEMENTS
This work could not have been carried out withoutthe effort of J Castroviejo and the people of AsociacioacutenAmigos de Dontildeana I am very gratefully to S Carranza(BMNH) who did the genetic work and helped me toimprove the manuscript To I de la Riva (MNCN) forhis help in many aspects of this work To Dr NAnderson (BMNH) who revised the draft version andthe English of the manuscript To R Garciacutea M UrceraC Carballo M O Deida and E Mohamed O Saleh fortheir help and companion in Mauritania and J Peacuterez EAvila S Castroviejo-Fisher and A Quintana for theirfriendship and companion in the first trip to MauritaniaP Geniez J A Mateo H Nickel and W Boumlhme allhelped us in some aspect of this work W Wuster andtwo anonymous reviewers critically commented an ear-lier draft of the manuscript R Brown kindly providedraw scalation data for A impalearis from Morocco andcontributed to the improvement of the manuscript
REFERENCES
Boumlhme W (2000) Die wechselvolle Geschichte derSahara Untersuchung von Reptilienzoumlnosen entlangeines westsaharischen Transektes mit einemuumlberraschenden Fund im Suumldosten Mauritaniens Tierund Museum 7 11-21
Boumlhme W Meinig H amp Roumldel M A (1996) Newrecords of amphibians and reptiles from Burkina Fasoand Mali British Herpetological Society Bulletin 56 7-26
Boumlhme W Wilms T Nickel H amp Merz M (2001)Bericht uumlber eine herpetologische Forschungsreise indie westliche Sahara und ihre Randgebiete Zeitschriftdes Koumllner Zoo 44 117-131
Bons J amp Geniez P (1996) Anfibios y reptiles deMarruecos (Incluido Saacutehara Occidental) Atlasbiogeograacutefico Barcelona Asociacioacuten HerpetoloacutegicaEspantildeola
Brown R P amp Znari M (1998) Morphological variationin Agama impalearis from Morocco evidence forhistorical population vicariance and current climaticeffects Ecography 21 605-612
Brown R P Znari M El Mouden E H amp Harris P(1999) Estimating asymptotic size and geographicvariation in Agama impalearis from MoroccoEcography 22 277-284
NEW AGAMA SPECIES FROM MAURITANIA 33
J M PADIAL
Accepted 22304
Brown R P Pestano J amp Suaacuterez N M (2002) TheAtlas mountains as a biogeographical divide in North-West Africa evidence from mtDNA evolution in theAgamid lizard Agama impalearis MolecularPhylogenetics and Evolution 24 324-332
Carranza S Arnold E N Thomas R H Mateo J ampLopez-Jurado L F (1999) Status of the extinct giantlacertid lizard Gallotia simonyi simonyi (ReptiliaLacertidae) assessed using mtDNA sequences frommuseum specimens Herpetological Journal l9 83-86
Carranza S Arnold E N Mateo J amp Lopez-Jurado LF (2000) Long-distance colonization and radiation ingekkonid lizards Tarentola (Reptilia Gekkonidae)revealed by mitochondrial DNA sequencesProceedings of the Royal Society Series B 267 637-649
Chabanaud P (1917) Enumeacuteration des reptiles nonencore eacutetudieacutes de lrsquoAfrique occidentale appartenantaux collections du Museacuteum avec la description desespeacuteces nouvelles Bulletin du Museacuteum NationaldrsquoHistoire Naturelle de Paris Serie Zoologique 2383-105
Dekeyser P L amp Villiers A (1956) Contribution alrsquoeacutetude du peuplement de la Mauritanie Notationseacutecologiques et biogeacuteographiques sur la faune delrsquoAdrar Memoires de lrsquoInstitut Franccedilais de lrsquoAfriqueNoire 44 222 p 25 pl
Felsenstein J (1985) Confidence-limits on phylogenies ndashan approach using the bootstrap Evolution 39 783-791
Geniez Ph amp Foucart A (1995) Un nouvelAcanthodactyle en Algeacuterie Acanthodactylustaghitensis n sp (Reptilia Sauria Lacertidae)Bulletin du Museacuteum National drsquoHistoire NaturelleParis 4deg seacuterie 17 3-9
Geniez P Mateo J A amp Bons J (2000) A checklist ofthe amphibians and reptiles of Western Sahara(Amphibia Reptilia) Herpetozoa 13 149-163
Ineich I (1997) Les amphibiens et reptiles du littoralmauritanien In Environnement et littoralmauritanien 93-99 Colas P (Ed) MontpellierCIRAD
Joger U (1979) Zur Oumlkologie und Verbreitung wenigbekannter Agamen Westafrikas (Reptilia SauriaAgamidae) Salamandra 15 31-52
Joger U (1980) Eine neue Art der Gattung Tarentola(Reptilia Sauria Gekkonidae) aus WestafrikaAmphibia-Reptilia 1 137-147
Joger U (1981) Zur Herpetofaunistik WestafrikasBonner zoologische Beitraumlge 32 297-340
Joger U (1984) Taxonomische Revision der GattungTarentola (Reptilia Gekkonidae) Bonner zoologischeBeitraumlge 35 129-174
Joger U amp Lambert M R K (1996) Analysis of theherpetofauna of the Republic of Mali I Annotatedinventory with description of a new Uromastyx(Sauria Agamidae) Journal of African Zoology 11021-51
Lambert M R K amp Mullieacute W C (1998) Sexualdichromatism of Agama boulengeri observed insouthern Mauritania British Herpetological SocietyBulletin 65 42-44
Le Berre M (1989) Faune du Sahara I Poissons-Amphibiens-Reptiles Paris Lechevalier-R Chabaud
Le Houeacuterou H N (1990) Deacutefinition et limitesbioclimatiques du Sahara Segravecheresse 1 246-259
Lenk P Kalyabina S Wink M amp Joger U (2001)Evolutionary relationships among the true vipers(Reptilia Viperidae) inferred from mitochondrial DNAsequences Molecular Biology and Evolution 19 94-104
Mateo J A Geniez Ph Loacutepez-Jurado J L amp Bons J(1998) Chorological analysis and morphologicalvariation of saurians of the genus Uromastyx (ReptiliaAgamidae) in western Sahara Description of two newtaxa Revista Espantildeola de Herpetologiacutea 12 97-109
Salvador A (1982) A revision of the lizards of the genusAcanthodactylus (Sauria Lacertidae) Bonnerzoologische Beitraumlge Monographien 16167 p
Schleich H H Kaumlstle W amp Kabisch K (1996)Amphibians and reptiles of North Africa KoenigsteinKoeltz Scientific Publications
Swofford D L (2002) PAUP phylogenetic analysisusing parsimony (and others methods) v40b10Sinauer Associates Sunderland MA
Thompson J D Gibson T J Plewniak F JeanmouginF amp Higgins D G (1997) The ClustalX windowsinterface flexible strategies for multiple sequencealignment aided by quality analysis tools NucleicAcids Research 24 4876-4882
Wilms T amp Boumlhme W (2001) Revision der Uromastyxacanthinura ndash Artengruppe mit Beschreibung einerneuen Art aus der Zentralsahara (Reptilia SauriaAgamidae) Zoologische Abhandlungen StaatlichesMuseum fuumlr Tierkunde Dresden 51 73-104
34
APPENDIX 1
Additional specimens of Agama species examinedfor this work EBD Estacioacuten Bioloacutegica de Dontildeana Se-ville Spain CET Centro de Estudios TropicalesSevilla Spain Locality and coordinates in parentheses
Agama agama CET(RIM)-028 (near Bouli 15deg25803 N11deg 55562 W Mauritania) CET(RIM)-328-329 (El Wad-Zoueina15deg 42689 N 9deg 39906 WMauritania) CET(RIM)-331-335 (El Wad-Foulania15deg 31682 N9deg 48896 W Mauritania)
Agama boueti CET(RIM)-287-291 336 (Ayoucircn elAtroucircs 16deg 37412 N9deg 37441 W Mauritania)
Agama boulengeri CET(RIM)-016-018 (Bou-gari16deg 32034 N10deg 47892 W Mauritania)CET(RIM)-021-022 (between Timbedgha and AyoucircnEl Atroucircs 16deg 26890 N9deg 14690 W Mauritania)CET(RIM)-066 (Terjicirct 20deg 15578 N13 deg 05854 WMauritania) CET(RIM)-094-96 (Guelta Molomhar20deg 34873 N13deg 07630 W Mauritania) CET(RIM)-116-117 (Oued Choucircm 21deg 22654 N12deg 58581 WMauritania) CET(RIM)-174-175 (Zig between Lekh-cheb and Ticircchicirct 18deg 34487 N9deg 48379 WMauritania) CET(RIM)-256-261 (Guelta Oumm Leb-are 16deg 29472 N10deg 49822 W Mauritania)CET(RIM)-338 (Ayoucircn el Atroucircs 16deg 37412 N9deg37441 W Mauritania) CET(RIM)-344 (Guelta Mat-mata17deg 53571 N12deg 07467 W Mauritania)
Agama impalearis EBD-11352 (Mount Marcan-Kbir Tetuaacuten Morocco) EBD-20222 (TaklimMorocco) EBD-22084 (Yassinen Morocco) EBD-24515 (Bab Bou Ichir-Fritissa Morocco) EBD-6552(El Aioum Western Sahara)
NEW AGAMA SPECIES FROM MAURITANIA 35

superficial resemblance with A castroviejoi Neverthe-less morphological quantitative and meristic charactersdistinguish A castroviejoi well from the other speciesrecorded for Mauritania (A boulengeri A agama andA boueti) and also from the two Moroccan forms of Aimpalearis MtDNA analyses demonstrate the identityof A castroviejoi and its close relationship to Aimpalearis and the vicariant form from south and east ofthe Atlas mountains
Furthermore distributional social and ecologicalfactors also allow a clear separation among species Acastroviejoi is solitary and inhabits extremely dry rockyareas of the Adrar plateau Agama boueti is solitary andinhabits the semi-desert sandy plains and savannahs ofthe Sahel region of southern Mauritania and the AtlanticCoast Agama impalearis is a typical solitary rockdweller (Schleich et al 1996) Its distribution rangesfrom the Mediterranean coast (Bons amp Geniez 1996) toSeguia el Hamra in Western Sahara (Geniez et al2000) Although it was cited for ldquoMauritanieSahariennerdquo (Chabanaud 1917) and despite appearingwidely distributed across the country in Le Berrersquos(1989) distribution map for this species there are actu-ally no confirmed records for Mauritania MoreoverJoger (1979) noted that this species did not occur in theAdrar and that the nearest record corresponded toSeguia el Hamra The presence of this species in thecountry is still plausible in rocky areas similar to thoseof the Zemmour Mountains (Geniez et al 2000) butthis requires confirmation A boulengeri is a speciesfrom the Sahel Savannah that inhabits rocky areas andforms social groups We found this species as far northas Oued Choucircm (21deg 226N12deg 586W) near the bor-der with Western Sahara At this latitude it is alwaysassociated to wet rocky gorges (Gueltas) in the moun-tains A agama is another social species reported to beassociated with human settlements in the Adrar region(Dekeyser amp Villiers 1956) In the south of the countrythis species prefers trees from the Sahel savannahs but itshows high habitat plasticity (pers obs)
Miocene vicariance events have been found to be re-sponsible of the variation and differentiation in Aimpalearis in NW Africa (Brown amp Znari 1998 Brownet al 2002) The uplift of the Atlas mountains consti-tuted a great geographical barrier causing allopatricfragmentation of the populations A castroviejoi is iso-lated in a rocky tableland surrounded by lowland sandyplains of the Sahara that could serve as a barrier for arock dweller Also in situ selection-mediated responsesto the ecological current conditions of the isolatedpopulations are responsible of the morphological varia-tion of A impalearis (Brown amp Znari 1998) Presentecological conditions of the Adrar are very extreme andis probably one of the hottest areas of the Sahara (LeHoueacuterou 1990) As demonstrated by Brown amp Znari(1998) variation in scalation is significantly associatedwith different temperature conditions Therefore bothisolation and ecological differentiation could be impli-cated in the differentiation of A castroviejoi
Although the Adrar belongs to the Meridional Sahara(Le Houeacuterou 1990) this area shows a complex historywith its faunas and floras comprising a mixture of Medi-terranean Saharan and Sahelian species (Dekeyser ampVilliers 1956) Probably A castroviejoi constitutes anexample of a Saharan endemic of the Adrar Mountainsas some invertebrate species and also vertebrate subspe-cies (Dekeyser amp Villiers 1956) Nevertheless thephylogeographic affinities of A castroviejoi remain un-clear since it could have a Sahelian or Saharan origin Amore thorough phylogenetic comparison among NorthAfrican agamas will help to resolve this question
ACKNOWLEDGEMENTS
This work could not have been carried out withoutthe effort of J Castroviejo and the people of AsociacioacutenAmigos de Dontildeana I am very gratefully to S Carranza(BMNH) who did the genetic work and helped me toimprove the manuscript To I de la Riva (MNCN) forhis help in many aspects of this work To Dr NAnderson (BMNH) who revised the draft version andthe English of the manuscript To R Garciacutea M UrceraC Carballo M O Deida and E Mohamed O Saleh fortheir help and companion in Mauritania and J Peacuterez EAvila S Castroviejo-Fisher and A Quintana for theirfriendship and companion in the first trip to MauritaniaP Geniez J A Mateo H Nickel and W Boumlhme allhelped us in some aspect of this work W Wuster andtwo anonymous reviewers critically commented an ear-lier draft of the manuscript R Brown kindly providedraw scalation data for A impalearis from Morocco andcontributed to the improvement of the manuscript
REFERENCES
Boumlhme W (2000) Die wechselvolle Geschichte derSahara Untersuchung von Reptilienzoumlnosen entlangeines westsaharischen Transektes mit einemuumlberraschenden Fund im Suumldosten Mauritaniens Tierund Museum 7 11-21
Boumlhme W Meinig H amp Roumldel M A (1996) Newrecords of amphibians and reptiles from Burkina Fasoand Mali British Herpetological Society Bulletin 56 7-26
Boumlhme W Wilms T Nickel H amp Merz M (2001)Bericht uumlber eine herpetologische Forschungsreise indie westliche Sahara und ihre Randgebiete Zeitschriftdes Koumllner Zoo 44 117-131
Bons J amp Geniez P (1996) Anfibios y reptiles deMarruecos (Incluido Saacutehara Occidental) Atlasbiogeograacutefico Barcelona Asociacioacuten HerpetoloacutegicaEspantildeola
Brown R P amp Znari M (1998) Morphological variationin Agama impalearis from Morocco evidence forhistorical population vicariance and current climaticeffects Ecography 21 605-612
Brown R P Znari M El Mouden E H amp Harris P(1999) Estimating asymptotic size and geographicvariation in Agama impalearis from MoroccoEcography 22 277-284
NEW AGAMA SPECIES FROM MAURITANIA 33
J M PADIAL
Accepted 22304
Brown R P Pestano J amp Suaacuterez N M (2002) TheAtlas mountains as a biogeographical divide in North-West Africa evidence from mtDNA evolution in theAgamid lizard Agama impalearis MolecularPhylogenetics and Evolution 24 324-332
Carranza S Arnold E N Thomas R H Mateo J ampLopez-Jurado L F (1999) Status of the extinct giantlacertid lizard Gallotia simonyi simonyi (ReptiliaLacertidae) assessed using mtDNA sequences frommuseum specimens Herpetological Journal l9 83-86
Carranza S Arnold E N Mateo J amp Lopez-Jurado LF (2000) Long-distance colonization and radiation ingekkonid lizards Tarentola (Reptilia Gekkonidae)revealed by mitochondrial DNA sequencesProceedings of the Royal Society Series B 267 637-649
Chabanaud P (1917) Enumeacuteration des reptiles nonencore eacutetudieacutes de lrsquoAfrique occidentale appartenantaux collections du Museacuteum avec la description desespeacuteces nouvelles Bulletin du Museacuteum NationaldrsquoHistoire Naturelle de Paris Serie Zoologique 2383-105
Dekeyser P L amp Villiers A (1956) Contribution alrsquoeacutetude du peuplement de la Mauritanie Notationseacutecologiques et biogeacuteographiques sur la faune delrsquoAdrar Memoires de lrsquoInstitut Franccedilais de lrsquoAfriqueNoire 44 222 p 25 pl
Felsenstein J (1985) Confidence-limits on phylogenies ndashan approach using the bootstrap Evolution 39 783-791
Geniez Ph amp Foucart A (1995) Un nouvelAcanthodactyle en Algeacuterie Acanthodactylustaghitensis n sp (Reptilia Sauria Lacertidae)Bulletin du Museacuteum National drsquoHistoire NaturelleParis 4deg seacuterie 17 3-9
Geniez P Mateo J A amp Bons J (2000) A checklist ofthe amphibians and reptiles of Western Sahara(Amphibia Reptilia) Herpetozoa 13 149-163
Ineich I (1997) Les amphibiens et reptiles du littoralmauritanien In Environnement et littoralmauritanien 93-99 Colas P (Ed) MontpellierCIRAD
Joger U (1979) Zur Oumlkologie und Verbreitung wenigbekannter Agamen Westafrikas (Reptilia SauriaAgamidae) Salamandra 15 31-52
Joger U (1980) Eine neue Art der Gattung Tarentola(Reptilia Sauria Gekkonidae) aus WestafrikaAmphibia-Reptilia 1 137-147
Joger U (1981) Zur Herpetofaunistik WestafrikasBonner zoologische Beitraumlge 32 297-340
Joger U (1984) Taxonomische Revision der GattungTarentola (Reptilia Gekkonidae) Bonner zoologischeBeitraumlge 35 129-174
Joger U amp Lambert M R K (1996) Analysis of theherpetofauna of the Republic of Mali I Annotatedinventory with description of a new Uromastyx(Sauria Agamidae) Journal of African Zoology 11021-51
Lambert M R K amp Mullieacute W C (1998) Sexualdichromatism of Agama boulengeri observed insouthern Mauritania British Herpetological SocietyBulletin 65 42-44
Le Berre M (1989) Faune du Sahara I Poissons-Amphibiens-Reptiles Paris Lechevalier-R Chabaud
Le Houeacuterou H N (1990) Deacutefinition et limitesbioclimatiques du Sahara Segravecheresse 1 246-259
Lenk P Kalyabina S Wink M amp Joger U (2001)Evolutionary relationships among the true vipers(Reptilia Viperidae) inferred from mitochondrial DNAsequences Molecular Biology and Evolution 19 94-104
Mateo J A Geniez Ph Loacutepez-Jurado J L amp Bons J(1998) Chorological analysis and morphologicalvariation of saurians of the genus Uromastyx (ReptiliaAgamidae) in western Sahara Description of two newtaxa Revista Espantildeola de Herpetologiacutea 12 97-109
Salvador A (1982) A revision of the lizards of the genusAcanthodactylus (Sauria Lacertidae) Bonnerzoologische Beitraumlge Monographien 16167 p
Schleich H H Kaumlstle W amp Kabisch K (1996)Amphibians and reptiles of North Africa KoenigsteinKoeltz Scientific Publications
Swofford D L (2002) PAUP phylogenetic analysisusing parsimony (and others methods) v40b10Sinauer Associates Sunderland MA
Thompson J D Gibson T J Plewniak F JeanmouginF amp Higgins D G (1997) The ClustalX windowsinterface flexible strategies for multiple sequencealignment aided by quality analysis tools NucleicAcids Research 24 4876-4882
Wilms T amp Boumlhme W (2001) Revision der Uromastyxacanthinura ndash Artengruppe mit Beschreibung einerneuen Art aus der Zentralsahara (Reptilia SauriaAgamidae) Zoologische Abhandlungen StaatlichesMuseum fuumlr Tierkunde Dresden 51 73-104
34
APPENDIX 1
Additional specimens of Agama species examinedfor this work EBD Estacioacuten Bioloacutegica de Dontildeana Se-ville Spain CET Centro de Estudios TropicalesSevilla Spain Locality and coordinates in parentheses
Agama agama CET(RIM)-028 (near Bouli 15deg25803 N11deg 55562 W Mauritania) CET(RIM)-328-329 (El Wad-Zoueina15deg 42689 N 9deg 39906 WMauritania) CET(RIM)-331-335 (El Wad-Foulania15deg 31682 N9deg 48896 W Mauritania)
Agama boueti CET(RIM)-287-291 336 (Ayoucircn elAtroucircs 16deg 37412 N9deg 37441 W Mauritania)
Agama boulengeri CET(RIM)-016-018 (Bou-gari16deg 32034 N10deg 47892 W Mauritania)CET(RIM)-021-022 (between Timbedgha and AyoucircnEl Atroucircs 16deg 26890 N9deg 14690 W Mauritania)CET(RIM)-066 (Terjicirct 20deg 15578 N13 deg 05854 WMauritania) CET(RIM)-094-96 (Guelta Molomhar20deg 34873 N13deg 07630 W Mauritania) CET(RIM)-116-117 (Oued Choucircm 21deg 22654 N12deg 58581 WMauritania) CET(RIM)-174-175 (Zig between Lekh-cheb and Ticircchicirct 18deg 34487 N9deg 48379 WMauritania) CET(RIM)-256-261 (Guelta Oumm Leb-are 16deg 29472 N10deg 49822 W Mauritania)CET(RIM)-338 (Ayoucircn el Atroucircs 16deg 37412 N9deg37441 W Mauritania) CET(RIM)-344 (Guelta Mat-mata17deg 53571 N12deg 07467 W Mauritania)
Agama impalearis EBD-11352 (Mount Marcan-Kbir Tetuaacuten Morocco) EBD-20222 (TaklimMorocco) EBD-22084 (Yassinen Morocco) EBD-24515 (Bab Bou Ichir-Fritissa Morocco) EBD-6552(El Aioum Western Sahara)
NEW AGAMA SPECIES FROM MAURITANIA 35

J M PADIAL
Accepted 22304
Brown R P Pestano J amp Suaacuterez N M (2002) TheAtlas mountains as a biogeographical divide in North-West Africa evidence from mtDNA evolution in theAgamid lizard Agama impalearis MolecularPhylogenetics and Evolution 24 324-332
Carranza S Arnold E N Thomas R H Mateo J ampLopez-Jurado L F (1999) Status of the extinct giantlacertid lizard Gallotia simonyi simonyi (ReptiliaLacertidae) assessed using mtDNA sequences frommuseum specimens Herpetological Journal l9 83-86
Carranza S Arnold E N Mateo J amp Lopez-Jurado LF (2000) Long-distance colonization and radiation ingekkonid lizards Tarentola (Reptilia Gekkonidae)revealed by mitochondrial DNA sequencesProceedings of the Royal Society Series B 267 637-649
Chabanaud P (1917) Enumeacuteration des reptiles nonencore eacutetudieacutes de lrsquoAfrique occidentale appartenantaux collections du Museacuteum avec la description desespeacuteces nouvelles Bulletin du Museacuteum NationaldrsquoHistoire Naturelle de Paris Serie Zoologique 2383-105
Dekeyser P L amp Villiers A (1956) Contribution alrsquoeacutetude du peuplement de la Mauritanie Notationseacutecologiques et biogeacuteographiques sur la faune delrsquoAdrar Memoires de lrsquoInstitut Franccedilais de lrsquoAfriqueNoire 44 222 p 25 pl
Felsenstein J (1985) Confidence-limits on phylogenies ndashan approach using the bootstrap Evolution 39 783-791
Geniez Ph amp Foucart A (1995) Un nouvelAcanthodactyle en Algeacuterie Acanthodactylustaghitensis n sp (Reptilia Sauria Lacertidae)Bulletin du Museacuteum National drsquoHistoire NaturelleParis 4deg seacuterie 17 3-9
Geniez P Mateo J A amp Bons J (2000) A checklist ofthe amphibians and reptiles of Western Sahara(Amphibia Reptilia) Herpetozoa 13 149-163
Ineich I (1997) Les amphibiens et reptiles du littoralmauritanien In Environnement et littoralmauritanien 93-99 Colas P (Ed) MontpellierCIRAD
Joger U (1979) Zur Oumlkologie und Verbreitung wenigbekannter Agamen Westafrikas (Reptilia SauriaAgamidae) Salamandra 15 31-52
Joger U (1980) Eine neue Art der Gattung Tarentola(Reptilia Sauria Gekkonidae) aus WestafrikaAmphibia-Reptilia 1 137-147
Joger U (1981) Zur Herpetofaunistik WestafrikasBonner zoologische Beitraumlge 32 297-340
Joger U (1984) Taxonomische Revision der GattungTarentola (Reptilia Gekkonidae) Bonner zoologischeBeitraumlge 35 129-174
Joger U amp Lambert M R K (1996) Analysis of theherpetofauna of the Republic of Mali I Annotatedinventory with description of a new Uromastyx(Sauria Agamidae) Journal of African Zoology 11021-51
Lambert M R K amp Mullieacute W C (1998) Sexualdichromatism of Agama boulengeri observed insouthern Mauritania British Herpetological SocietyBulletin 65 42-44
Le Berre M (1989) Faune du Sahara I Poissons-Amphibiens-Reptiles Paris Lechevalier-R Chabaud
Le Houeacuterou H N (1990) Deacutefinition et limitesbioclimatiques du Sahara Segravecheresse 1 246-259
Lenk P Kalyabina S Wink M amp Joger U (2001)Evolutionary relationships among the true vipers(Reptilia Viperidae) inferred from mitochondrial DNAsequences Molecular Biology and Evolution 19 94-104
Mateo J A Geniez Ph Loacutepez-Jurado J L amp Bons J(1998) Chorological analysis and morphologicalvariation of saurians of the genus Uromastyx (ReptiliaAgamidae) in western Sahara Description of two newtaxa Revista Espantildeola de Herpetologiacutea 12 97-109
Salvador A (1982) A revision of the lizards of the genusAcanthodactylus (Sauria Lacertidae) Bonnerzoologische Beitraumlge Monographien 16167 p
Schleich H H Kaumlstle W amp Kabisch K (1996)Amphibians and reptiles of North Africa KoenigsteinKoeltz Scientific Publications
Swofford D L (2002) PAUP phylogenetic analysisusing parsimony (and others methods) v40b10Sinauer Associates Sunderland MA
Thompson J D Gibson T J Plewniak F JeanmouginF amp Higgins D G (1997) The ClustalX windowsinterface flexible strategies for multiple sequencealignment aided by quality analysis tools NucleicAcids Research 24 4876-4882
Wilms T amp Boumlhme W (2001) Revision der Uromastyxacanthinura ndash Artengruppe mit Beschreibung einerneuen Art aus der Zentralsahara (Reptilia SauriaAgamidae) Zoologische Abhandlungen StaatlichesMuseum fuumlr Tierkunde Dresden 51 73-104
34
APPENDIX 1
Additional specimens of Agama species examinedfor this work EBD Estacioacuten Bioloacutegica de Dontildeana Se-ville Spain CET Centro de Estudios TropicalesSevilla Spain Locality and coordinates in parentheses
Agama agama CET(RIM)-028 (near Bouli 15deg25803 N11deg 55562 W Mauritania) CET(RIM)-328-329 (El Wad-Zoueina15deg 42689 N 9deg 39906 WMauritania) CET(RIM)-331-335 (El Wad-Foulania15deg 31682 N9deg 48896 W Mauritania)
Agama boueti CET(RIM)-287-291 336 (Ayoucircn elAtroucircs 16deg 37412 N9deg 37441 W Mauritania)
Agama boulengeri CET(RIM)-016-018 (Bou-gari16deg 32034 N10deg 47892 W Mauritania)CET(RIM)-021-022 (between Timbedgha and AyoucircnEl Atroucircs 16deg 26890 N9deg 14690 W Mauritania)CET(RIM)-066 (Terjicirct 20deg 15578 N13 deg 05854 WMauritania) CET(RIM)-094-96 (Guelta Molomhar20deg 34873 N13deg 07630 W Mauritania) CET(RIM)-116-117 (Oued Choucircm 21deg 22654 N12deg 58581 WMauritania) CET(RIM)-174-175 (Zig between Lekh-cheb and Ticircchicirct 18deg 34487 N9deg 48379 WMauritania) CET(RIM)-256-261 (Guelta Oumm Leb-are 16deg 29472 N10deg 49822 W Mauritania)CET(RIM)-338 (Ayoucircn el Atroucircs 16deg 37412 N9deg37441 W Mauritania) CET(RIM)-344 (Guelta Mat-mata17deg 53571 N12deg 07467 W Mauritania)
Agama impalearis EBD-11352 (Mount Marcan-Kbir Tetuaacuten Morocco) EBD-20222 (TaklimMorocco) EBD-22084 (Yassinen Morocco) EBD-24515 (Bab Bou Ichir-Fritissa Morocco) EBD-6552(El Aioum Western Sahara)
NEW AGAMA SPECIES FROM MAURITANIA 35

APPENDIX 1
Additional specimens of Agama species examinedfor this work EBD Estacioacuten Bioloacutegica de Dontildeana Se-ville Spain CET Centro de Estudios TropicalesSevilla Spain Locality and coordinates in parentheses
Agama agama CET(RIM)-028 (near Bouli 15deg25803 N11deg 55562 W Mauritania) CET(RIM)-328-329 (El Wad-Zoueina15deg 42689 N 9deg 39906 WMauritania) CET(RIM)-331-335 (El Wad-Foulania15deg 31682 N9deg 48896 W Mauritania)
Agama boueti CET(RIM)-287-291 336 (Ayoucircn elAtroucircs 16deg 37412 N9deg 37441 W Mauritania)
Agama boulengeri CET(RIM)-016-018 (Bou-gari16deg 32034 N10deg 47892 W Mauritania)CET(RIM)-021-022 (between Timbedgha and AyoucircnEl Atroucircs 16deg 26890 N9deg 14690 W Mauritania)CET(RIM)-066 (Terjicirct 20deg 15578 N13 deg 05854 WMauritania) CET(RIM)-094-96 (Guelta Molomhar20deg 34873 N13deg 07630 W Mauritania) CET(RIM)-116-117 (Oued Choucircm 21deg 22654 N12deg 58581 WMauritania) CET(RIM)-174-175 (Zig between Lekh-cheb and Ticircchicirct 18deg 34487 N9deg 48379 WMauritania) CET(RIM)-256-261 (Guelta Oumm Leb-are 16deg 29472 N10deg 49822 W Mauritania)CET(RIM)-338 (Ayoucircn el Atroucircs 16deg 37412 N9deg37441 W Mauritania) CET(RIM)-344 (Guelta Mat-mata17deg 53571 N12deg 07467 W Mauritania)
Agama impalearis EBD-11352 (Mount Marcan-Kbir Tetuaacuten Morocco) EBD-20222 (TaklimMorocco) EBD-22084 (Yassinen Morocco) EBD-24515 (Bab Bou Ichir-Fritissa Morocco) EBD-6552(El Aioum Western Sahara)
NEW AGAMA SPECIES FROM MAURITANIA 35