







Languages
Pages
Legal


UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR
DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA
POSGRADO EN CIENCIAS MARINAS Y COSTERAS
TESIS
PROPUESTA DE ZONACIÓN DEL GOLFO DE CALIFORNIA CON
BASE EN VARIABLES OCEANOGRÁFICAS Y DISTRIBUCIÓN DE
MACROINVERTEBRADOS
QUE COMO REQUISITO PARA OBTENER EL GRADO DE
MAESTRO EN CIENCIAS MARINAS Y COSTERAS
CON ORIENTACIÓN EN BIOLOGÍA MARINA
PRESENTA:
DAVID PETATÁN RAMÍREZ
DIRECTOR:
DR. HÉCTOR REYES BONILLA
LA PAZ, B.C.S. ENERO DE 2015


A Denisse, Luis Ángel, Eduardo y Alondra.

Agradecimientos
Gracias a CONACyT por la beca de manutención (267516) y la beca mixta otorgada
para movilidad nacional y en el extranjero (290748 y 290749), a los proyectos de
CONABIO (JM065) y CONACYT-CB (183534) a cargo del Dr. Héctor Reyes Bonilla, al
COSCyT y al XIV Ayuntamiento de La Paz por el apoyo económico para la asistencia
a congresos.
Gracias a mi comité de tesis, en especial a mi director, el Dr. Héctor Reyes, gracias
por todo el apoyo, tiempo y paciencia que me tuviste, no pude elegir a un mejor guía
que tú para entrar a este mundo nuevo para mí. A mis asesores, la Dra. Laura Sánchez
y el Dr. Genaro Martínez, lo mismo, gracias por el apoyo y las valiosas observaciones
que enriquecieron el desarrollo de este trabajo. De verdad gracias!
Gracias a Rick Brusca, por recibirme en la Universidad de Arizona para la estancia, sin
duda por algo eres el papá de los pollitos, gracias por todo tu apoyo y tus comentarios
que lograron darme una idea más clara de lo que había que hacer. Por su puesto un
enorme gracias a la familia Frisby (Matty, Tommy, Iván, Grecia y Jorge), que se
convirtió en mi familia adoptiva durante mi estancia, de verdad muchísimas gracias por
todos los buenos recuerdos.
Al Dr. Saúl Álvarez Borrego, gracias Saúl por creer en mí, por aceptarme para la
estancia en CICESE, por tu tiempo, tus observaciones y tus consejos, sin duda eres
un gran investigador y una gran persona. Gracias a Orión y Cecilia, por recibirme en
mis idas a Ensenada, por su apoyo, por su amistad, ¡Gracias!
A mis maestros y compañeros de clases, que me ayudaron a conocer un poco de todo
esto, y desde luego, a todos mis compañeros de laboratorio, con los cuales he
aprendido mucho más que solo ciencia, gracias por apoyarme, por ayudarme, por
aguantarme, aunque hubo momentos en los que quería abandonar todo, al final, lo
hemos logrado. Agradecimiento especial a Gaby, Triana, Sara y Daniel, por su ayuda
y su amistad.
A los amigos, Isma, Jorge, Andrés, Adrián, Denise y Juan por su amistad, por su apoyo
incondicional, por sus consejos, por todos los buenos momentos que hemos vivido y
los que están por venir. Gracias…
A todas aquellas personas que he ido conociendo en esta nueva etapa, de las cuales
solo he recibido apoyo, crítica constructiva, que me han brindado su amistad, su ayuda
y su conocimiento para enriquecer este trabajo. Esta tesis no es más que el resultado
de ese apoyo colectivo, muchas gracias.

i
ÍNDICE GENERAL
Página
ÍNDICE GENERAL ....................................................................................................................................... i
LISTA DE FIGURAS .................................................................................................................................... iii
LISTA DE TABLAS ....................................................................................................................................... v
RESUMEN ................................................................................................................................................. vi
INTRODUCCIÓN ....................................................................................................................................... 1
ANTECEDENTES ........................................................................................................................................ 5
JUSTIFICACIÓN ....................................................................................................................................... 15
OBJETIVOS.............................................................................................................................................. 17
Objetivo General ................................................................................................................................ 17
Objetivos Específicos .......................................................................................................................... 17
ÁREA DE ESTUDIO .................................................................................................................................. 18
METODOLOGÍA ...................................................................................................................................... 21
Información oceanográfica ................................................................................................................ 22
Información oceanográfica vía sensores remotos ......................................................................... 23
Información oceanográfica in situ .................................................................................................. 24
Información Biológica......................................................................................................................... 25
Regionalización del Golfo de California.............................................................................................. 27
Regionalización oceanográfica ....................................................................................................... 27
Regionalización biológica ............................................................................................................... 28
Biomas del Golfo de California ....................................................................................................... 29
RESULTADOS .......................................................................................................................................... 30
Regionalización oceanográfica ........................................................................................................... 30
Características de las regiones oceanográficas .............................................................................. 33
Variabilidad mensual de las regiones oceanográficas ................................................................... 38
Variables medidas in situ ............................................................................................................... 46
Regionalización biológica ................................................................................................................... 47
Regionalización a nivel phylum ...................................................................................................... 47
Regionalización general de macroinvertebrados ........................................................................... 53

ii
Biomas del Golfo de California ........................................................................................................... 55
DISCUSIÓN ............................................................................................................................................. 57
Regionalización oceanográfica ........................................................................................................... 57
Regionalización biológica ................................................................................................................... 63
Los biomas del Golfo de California. .................................................................................................... 67
CONCLUSIONES ...................................................................................................................................... 70
RECOMENDACIONES ............................................................................................................................. 72
BIBLIOGRAFÍA ........................................................................................................................................ 73
ANEXO I .................................................................................................................................................. 86
ANEXO II ................................................................................................................................................. 87
ANEXO III ................................................................................................................................................ 88
ANEXO IV ............................................................................................................................................... 89
ANEXO V ................................................................................................................................................. 90
ANEXO VI ............................................................................................................................................... 91
ANEXO VII .............................................................................................................................................. 92
ANEXO VIII ............................................................................................................................................. 93
ANEXO IX ................................................................................................................................................ 94
ANEXO X ................................................................................................................................................. 95
ANEXO XI ................................................................................................................................................ 96

iii
LISTA DE FIGURAS
Página
Figura 1 - Regiones biogeoquímicas del océano propuestas por Longhurts (1995).
Modificado de IOCCG (2009). ..................................................................................... 2
Figura 2 - Ecorregiones del mundo. Modificado de Spalding y colaboradores (2007).3
Figura 3 - División geográfica de acuerdo con: (A) Sverdrup, 1941 con líneas rojas.
Roden y Emilsson, 1979 con líneas verdes. (B) Walker, 1960. (C) Thomson y
colaboradores, 2000. ................................................................................................... 6
Figura 4 - División geográfica de acuerdo con: (A) Gilbert y Allen, 1943. (B) Round,
1967. ......................................................................................................................... 10
Figura 5 - División geográfica de acuerdo con: (A) Santamaría del Ángel y
colaboradores, 1994. (B) Lluch-Cota y Arias-Aréchiga, 2000. (C) Lavín y Marinone,
2003. ......................................................................................................................... 11
Figura 6 - División geográfica de acuerdo con: (A) Kahru y colaboradores, 2004. (B)
Hidalgo-González y Álvarez-Borrego, 2004, con líneas roja regionalización de invierno,
con línea azul regionalización de verano. (C) Ulloa y colaboradores, 2006. ............. 12
Figura 7 - División geográfica de acuerdo con: (A) SEMARNAT, 2006. (B) Wilkinson y
colaboradores, 2009. ................................................................................................. 13
Figura 8 - El Golfo de California, limitado al sur por una línea entre Cabo San Lucas,
BCS, y Bahía de Banderas, Nayarit. ......................................................................... 18
Figura 9 - Rejilla de trabajo para la regionalización del Golfo de California a 9km2 de
resolución espacial. ................................................................................................... 22
Figura 10 - Distribución espacial de las mediciones in situ del periodo 1939 – 2007
obtenida de la NODC. ............................................................................................... 25
Figura 11 - Índice de k – promedios para calcular el número de regiones
oceanográficas del Golfo de California. ..................................................................... 30
Figura 12 - Regionalización oceanográfica del Golfo de California. ......................... 31
Figura 13 - Cobertura de las regiones oceanográficas. ............................................ 32
Figura 14 - Dendrograma de similitud oceanográfica a nivel región. ........................ 33

iv
Figura 15 - Variabilidad mensual de temperatura del Golfo de California a nivel región.
.................................................................................................................................. 39
Figura 16 - Variabilidad mensual de clorofila a del Golfo de California a nivel región.
.................................................................................................................................. 40
Figura 17 - Variabilidad mensual de atenuación de la luz del Golfo de California a nivel
región. ....................................................................................................................... 41
Figura 18 - Variabilidad mensual de radiación fotosintéticamente activa en el Golfo de
California a nivel región. ............................................................................................ 42
Figura 19 - Variabilidad anual del carbono inorgánico particulado en el Golfo de
California a nivel región. ............................................................................................ 43
Figura 20 - Variabilidad mensual de carbono orgánico particulado en el Golfo de
California a nivel región. ............................................................................................ 44
Figura 21 - Variabilidad mensual de la materia coloreada disuelta en el Golfo de
California a nivel región. ............................................................................................ 45
Figura 22 - Índice de k – promedios para los cinco fila seleccionados de
macroinvetebrados del Golfo de California. .............................................................. 47
Figura 23 - Riqueza específica de macroinvertebrados en el Golfo de California. ... 48
Figura 24 - A) Regionalización del phylum Mollusca. B) Dendrograma de similitud
biológica entre regiones. C) Riqueza de específica de especies a nivel región. ....... 49
Figura 25 - (A) Regionalización del phylum Echinodermata. (B) Dendrograma de
similitud biológica entre regiones. (C) Riqueza de específica de especies a nivel región.
.................................................................................................................................. 50
Figura 26 - (A) Regionalización del phylum Cnidaria. (B) Dendrograma de similitud
biológica entre regiones. (C) Riqueza de específica de especies a nivel región. ...... 51
Figura 27 - (A) Regionalización del phylum Artrhopoda. (B) Dendrograma de similitud
biológica entre regiones. (C) Riqueza de específica de especies. ............................ 52
Figura 28 - (A) Regionalización del phylum Annelida. (B) Dendrograma de similitud
biológica entre regiones. (C) Riqueza de específica de especies a nivel región. ...... 53
Figura 29 - (A) Regionalización biológica del Golfo de California. (B) Dendrograma de
similitud biológica entre regiones. (C) Riqueza de específica de especies a nivel región.
.................................................................................................................................. 54

v
Figura 30 - (A) Biomas del Golfo de California. (B) Dendrograma de similitud
oceanográfica entre biomas. (C) Dendrograma de similitud biológica entre biomas. (D)
Riqueza de específica de especies a nivel bioma. .................................................... 56
Figura 31 - Componentes principales de las variables oceanográficas para la
regionalización del Golfo de California. ..................................................................... 61
Figura 32 - Sedimentología del Golfo de California. (modificado de INAPESCA, 1994).
.................................................................................................................................. 65
LISTA DE TABLAS
Página
TABLA I - Variables oceanográficas utilizadas para la regionalización. ................... 23
TABLA II - Número de Invertebrados del Golfo de California por nivel taxonómico. 26
TABLA III - Lista de regiones oceanográficas del Golfo de California. ..................... 32
TABLA IV - valores promedio de las variables medidas in situ................................. 46

vi
RESUMEN
El Golfo de California (GC) es uno de los mares más diversos y productivos del mundo,
de gran importancia económica para el país por las pesquerías y el turismo, por lo que
se han realizado esfuerzos enfocados a su regionalización desde hace más de 50
años, con base en aspectos como la oceanografía o la distribución de algunos grupos
faunísticos, sin que se lograra hasta el momento una regionalización única generada
a partir de la combinación de los dos factores (oceanográfico/biológico) los cuales
resultarían en regiones con mayor sustento al englobar ambas características. Es por
esto que el objetivo de este trabajo es crear una zonación que fusione las dos
características para la generación de los biomas marinos del Golfo de California, los
cuales son regiones naturales que comparten las mismas características ambientales
y biológicas, por lo que con base en información oceanográfica obtenida por sensores
remotos y datos históricos obtenidos in situ, se generó una nueva zonación
oceanográfica con 11 variables físico-químicas del cual se obtuvieron 14 regiones
oceanográficas. Por otra parte, para la zonación biológica se generaron mapas de
distribución geográfica de cinco grupos de invertebrados (Mollusca, Arthropoda,
Cnidaria, Echinodermata y Annelida) con más de 3,400 especies y se generó la
zonación que tiene cada grupo en el GC para finalmente generar la zonación general
de macroinvertebrados, generada a partir de las distintas zonas obtenidas de cada
grupo ponderado a partir de la contribución (número de especies) de cada uno de los
fila. Se encontró que las regiones siguen un patrón estacional para la mayoría de las
variables ambientales, mientras que la distribución de especies mantiene un gradiente
de sur a norte altamente relacionado con el tipo de sedimentos, la profundidad y el tipo
de costa. Finalmente, para la generación de los biomas marinos se consideró a la
zonación oceanográfica con un 50% de contribución y a los fila con el otro 50%
obteniendo un total de 10 biomas, de los cuales cuatro corresponden a la zona costera
y seis a la zona oceánica, estos últimos presentaron menor variabilidad ambiental,
además de una diversidad más baja, relacionados a la presencia de cuencas.
Respecto a los biomas costeros, el cambio es principalmente latitudinal, todos con
presencia en la costa peninsular y la continental.

1
INTRODUCCIÓN
Históricamente los mares del mundo han sido subdivididos en zonas o regiones
para poder estudiarlos, entenderlos y administrarlos. Las zonificaciones obedecen a
diversos objetivos y por ello han incluido por separado parámetros físicos o biológicos,
buscando la organización de un área o zonas con características similares u
homogéneas que funcionen a escalas definidas (Córdova et al., 2006; Callejas-
Jiménez et al., 2012).
Estas divisiones han sido efectuadas incluso a escala planetaria, y se siguieron
diferentes enfoques. Por ejemplo, Dietrich (1957) propuso una regionalización de los
océanos del mundo por las condiciones ambientales de la superficie, ya que las
consideró de mayor importancia para el ser humano; el autor, delimitó siete regiones
hidrográficas, descartando mares marginales al considerarlos de transición del margen
costero, y dejando solamente a los océanos. Longhurst y colaboradores en 1995
realizaron estimaciones de la producción primaria global con datos de concentración
pigmentaria obtenidos del sensor CZCS (Coastal Zone Color Scanner) y mediciones
en campo, determinaron a partir de un análisis numérico 57 provincias repartidas en
cuatro dominios (fig. 1). Finalmente, Spalding y colaboradores (2007) con base en la
distribución de la biota bentónica y pelágica crearon una regionalización global de la
zona costera y plataforma continental ya que las consideran de mayor diversidad de
especies y de alta productividad, del cual obtuvieron 12 reinos, 62 provincias y 232
ecorregiones (fig. 2).
Para realizar una aproximación de regionalización marina, distintos trabajos
alrededor del mundo han hecho uso de variables ambientales que pueden venir de dos
fuentes: datos directos, los cuales son medidos in situ, o indirectos, derivados de
sensores que cuantifican el color del océano (Millán-Núñez et al., 2006). Estos últimos
tienen la desventaja de que los sensores son capaces de tomar datos únicamente de
la primera profundidad óptica del océano (aproximadamente el 22% de la zona
eufótica; Álvarez-Borrego, 1995; Hidalgo-González y Álvarez-Borrego, 2004), pero por
otra parte dan la opción de tener información con un corto retraso de tiempo y a
grandes escalas espaciales (Santamaría del Ángel et al., 2010). En contraste, los datos

2
medidos in situ son precisos pero presentan el problema de no contar con un monitoreo
espacio temporal adecuado, debido al alto costo operativo que tiene el llevar a cabo la
toma de muestras (Callejas-Jiménez et al., 2012). Por lo anterior, la mayoría de los
estudios de regionalización marina basados en aspectos ambientales se han llevado
a cabo con datos medidos por satélite. (Callejas-Jiménez et al., 2012).
Figura 1 - Regiones biogeoquímicas del océano propuestas por Longhurts (1995). Modificado de IOCCG (2009).
La regionalización basada en el medio físico es generalmente un buen predictor
de las regiones bióticas (Pielou, 1979; Hayden, et al., 1984). Por ejemplo, el gradiente
latitudinal de riqueza de especies (más alto en los trópicos) es en principio una
respuesta a las condiciones climáticas, aunque las condiciones locales pueden
provocar alteraciones en dicho patrón (Campero et al., 2003; Okolodkov, 2010). Los
trabajos de regionalización biológica se presentan en un sistema jerárquico de áreas
geográficas en términos de similitud de su biota o bien de patrones de especies
endémicas (Escalante, 2009).

3
Figura 2 - Ecorregiones del mundo. Modificado de Spalding y colaboradores (2007).
En el caso de México, existen trabajos de regionalización biogeográfica de la
cobertura terrestre, los cuales dividen al país en cinco componentes bióticos (neártico
californiano, neártico continental, mexicano de montaña, antillano y mesoamericano),
los cuales fueron divididos en 14 provincias (Morrone y Márquez, 2003). En el medio
marino, la zona económica exclusiva mexicana abarca ocho de las ecorregiones
propuestas por la Comisión para la Cooperación Ambiental de América del Norte (Lara-
Lara et al., 2008, Wilkinson et al., 2009); También se han hecho estudios sobre las
regiones prioritarias marinas (CONABIO-CONANP-TNC-PRONATURA, 2007), el cual
es un tipo de regionalización donde se identifican las zonas costeras y oceánicas con
alta biodiversidad para su conservación con base en criterios geomorfológicos,
fisiográficos y ecológicos, siendo un marco de referencia para el desarrollo de los
planes de manejo, uso, investigación y conservación (Arriaga et al., 1998; Contreras,
2002).
Por último, se han realizado esfuerzos para la creación de arreglos espaciales
utilizados como parte esencial de los ordenamientos ecológicos en México, los cuales
han seguido cuatro visiones del océano: oceánica, costera, de especies de importancia
ecológica y comercial, y socio económica (Córdova et al., 2006; CONABIO-CONANP-
TNC-PRONATURA, 2007), los cuales deben partir de un conocimiento adecuado de
la biodiversidad tomando en cuenta tres factores importantes: distribución geográfica,

4
batimetría y hábitat o sustrato utilizado por las especies (Hendrickx, 2008). Para que
todos estos tipos de esfuerzos sean exitosos, es necesario contar con bases de datos
de información biológica, pero también información sobre impactos antropogénicos,
mapas georreferenciados de las áreas, de la batimetría, y una extensa serie de
registros ambientales (Zacharias y Roff, 2000; Córdova et al., 2006; Hendrickx, 2008).
En México, el Golfo de California es una de las áreas más estudiadas
oceanográfica y biológicamente, del cual se puede concluir que es un ecosistema en
buen estado de conservación (Hendrickx y Brusca, 2007), de alta biodiversidad
(Brusca y Hendrickx, 2010), en el que existen comunidades marinas de afinidades
templadas y tropicales (López-Martínez et al., 2012); estas comunidades se componen
a su vez de miles de especies, las cuales desempeñan un papel estratégico en los
múltiples procesos ecológicos y biológicos (Hendrickx y Brusca, 2007), debido a estas
características se han elaborado varios trabajos de regionalización, sin embargo, estos
estudios se han realizado de manera aislada, sin una combinación de factores que
regionalicen de una manera integral al Golfo de California, por lo que existe una
necesidad de agrupar la información biología y oceanográfica para definir patrones que
en conjunto ayuden a delimitar espacios geográficos los cuales podrían ser útiles para
el manejo de recursos (Córdova et al., 2006).

5
ANTECEDENTES
Debido a la importancia biológica y económica del Golfo de California se ha
realizado una gran cantidad de estudios sobre sus condiciones bióticas y
oceanográficas, haciéndolo uno de los mares más estudiados del mundo (Molina-Cruz,
1984). A manera de resumen puede decirse que en un principio, los trabajos se
efectuaron con el fin de determinar si el GC es una región biológica y oceanográfica
diferente al Pacifico Oriental Tropical, y a partir de estos estudios se encontraron los
patrones espacio-temporales que siguen las diferentes variables oceanográficas.
Posteriormente hubo más interés en revisar aspectos biogeográficos, y se prestó
atención a los intervalos de distribución de las especies mejor caracterizadas
taxonómicamente. Finalmente, en las últimas décadas el enfoque cambió con el uso
de sensores remotos, lo que dirigió la investigación hacia la revisión de indicadores
ligados al color del océano.
Uno de los primeros estudios de regionalización del Golfo basada en aspectos
físicos o químicos corresponde a Sverdrup (1941), quien encontró que la temperatura
disminuye de sur a norte y decrece con la profundidad, sin embargo, en las islas
Tiburón y Ángel de la Guarda (Grandes Islas) se tiene un comportamiento diferente
(fig. 3A). Posteriormente, Roden y Emilsson (1979), en un estudio sobre la estructura
vertical termohalina del GC, determinan cuatro regiones: la primera desde la
desembocadura del Río Colorado hasta la Isla Tiburón (caracterizada por fuertes
corrientes de marea); el Canal de las Ballenas con condiciones oceanográficas
inusuales producto de continuas surgencias; el Bajo Golfo, el cual se localiza entre el
sur de Isla Tiburón hasta Cabo San Lucas y Mazatlán; y finalmente la entrada del Golfo,
la cual es un área triangular señalada entre Cabo Corrientes, Mazatlán y Cabo San
Lucas, la cual cuenta con una estructura termohalina complicada, causada por la
presencia de frentes y giros, además de la confluencia de tres diferentes corrientes
(fig. 3A).

6
Figura 3 - División geográfica de acuerdo con: (A) Sverdrup, 1941 con líneas rojas. Roden y Emilsson, 1979 con líneas verdes. (B) Walker, 1960. (C) Thomson y colaboradores, 2000.
El estudio de la fauna del GC también ha sugerido la existencia de patrones de
distribución espacial. A partir de la distribución geográfica de la ictiofauna, Walker
(1960) encontró que las especies de peces del GC presentan un aislamiento de la
fauna tropical al sur por dos barreras: la gran extensión de arena entre Guaymas y
Mazatlán que tiende a limitar la distribución de especies que habitan en orillas rocosas,
y la boca del Golfo que por la presencia de frentes oceanográficos limita el flujo de
larvas y organismos hacia el interior. El autor determinó que en el Golfo hay cuatro
agrupaciones de peces (fig. 3B), el Alto Golfo, desde Bahía San Francisquito al oeste
y la isla Tiburón en el lado continental, hasta el delta del Río Colorado, zona
caracterizada por poca variedad de especies pero la presencia de muchas endémicas
residentes de profundidades moderadas. Luego está el centro del Golfo, región que
cubre la zona aledaña a Guaymas y un gran tramo de la península de Baja California,
hasta La Paz, y aproximadamente a la misma latitud está el Golfo Sureste que abarca
desde Guaymas hasta Cabo Corrientes. Esta es la región menos distintiva ya que
cuenta con una costa rocosa muy pobre. Finalmente, en el sur hay otra zona que cubre
el sur de la península y de Mazatlán a Cabo Corrientes en el este, y que se caracteriza

7
por contar con especies afines al Pacifico Oriental Tropical y del Pacífico Occidental,
las cuales normalmente se encuentran en islas oceánicas adyacentes a las Américas.
Thomson y colaboradores (2000) sugieren una división en la distribución de
peces en solo tres zonas (fig. 3C), al norte resaltan las especies de afinidad templada,
mientras que en el sur sobresalen las especies típicas del Pacifico Oriental Tropical,
sobre todo en aguas profundas dado que parte de las masas de agua tienen las
mismas propiedades físico-químicas que las del Océano Pacífico (Castro-Aguirre et
al., 1996). El traslape biogeográfico de la fauna se debe a que el Golfo de California
es una zona transicional en donde se manifiestan especies con afinidad templada,
tropical y de transición templado-cálido (López-Martínez et al., 2012).
En el norte del GC, González-Cuellar (2012) realizó un estudio sobre las
comunidades de peces de arrecifes rocosos empleando datos de abundancias y
composición de especies con el fin de analizar la estructura comunitaria, encontrando
una regionalización que coincide con algunos trabajos previos como el de Santamaría
del Ángel y colaboradores (1994, que será descrita en detalle más adelante) y la de
Wilkinson y colaboradores (2009). Además diversos trabajos realizados con larvas de
peces y conectividad (Sánchez-Velasco et al., 2012; Sánchez-Velasco et al., 2013; y
Sánchez-Velasco et al., 2014), definen hábitats de larvas de peces que son similares
con las divisiones geográficas propuestas por Gilbert y Allen (1943) y Round (1967),
ya que la composición larvaria se diferencia en el Alto Golfo, la región Norte del Golfo,
Grandes Islas y Golfo sur, finalmente la entrada del Golfo es un hábitat diferente siendo
la zona de confluencia entre el Pacifico tropical transicional y el GC.
Otro estudio de la fauna que concuerda con regionalizaciones previas es el de
Szteren y Aurioles-Gamboa (2011), que se realizó con 13 colonias del lobo marino de
California (Zalophus californianus), con base en nueve variables de índole
demográfico, biológico y ambiental, encontraron una diferenciación de las colonias en
cuatro grupos principales: Golfo Norte, Ángel de la Guarda, Golfo Central y Golfo Sur,
estos grupos fueron coincidentes con la regionalización oceanográfica propuesta por
Lluch-Cota y Arias-Aréchiga (2000; detalles en páginas posteriores).

8
Por otra parte, estudios basados en la distribución de crustáceos eufáusidos por
Brinton y Townsend (1980) y anfípodos por Siegel-Causey (1982) concuerdan en que
existe una división del Golfo de California en dos regiones limitadas por las islas
Tiburón y Ángel de la Guarda, coincidiendo con la propuesta hidrográfica de Sverdrup
(1941). Además, las obras citadas destacan la sucesión estacional de especies
templadas y tropicales en la zona sur del Golfo.
Otro estudio que enfocado en la distribución geográfica de crustáceos
(Hendrickx et al., 2002) encontró que la riqueza disminuye tanto batimétrica y
latitudinalmente, siendo más diversas las zonas costeras, con una disminución desde
la boca del Golfo con una riqueza de 777 especies, hacia el norte del Golfo, con 498
especies, lo que la convierte en la zona con menor número de especies. Este patrón
de disminución de la diversidad de la fauna de sur a norte es consistente también en
el grupo de los moluscos (Hendrickx et al., 2007). Por otra parte, Simison en 2006 con
base en un estudio sobre la distribución de gasterópodos del Pacífico Oriental Tropical,
determina que el endemismo y la diversidad indican que el GC no es un ecosistema
aislado como sugieren otros autores, en el cual se esperaría encontrar baja diversidad
y un alto endemismo, sin embargo, presenta una alta diversidad, y aunque presenta
un endemismo del 17%, en zonas aledañas al Golfo es posible encontrar una
proporción de endemismo similar.
El Golfo de California y la costa occidental de la península de Baja California
han sido las que mayor esfuerzo de muestreo (oceanográfico y biológico) han recibido
en el Pacífico mexicano (Zamorano y Hendrickx, 2007), uno de los primeros estudios
fue en el crucero Albatross en 1891, en el cual se colectaron algunas especies de
profundidad. En años más recientes se han realizado estudios con el proyecto TALUD
(2000-2001) en el cual se colectaron especies bentónicas y pelágicas (Zamorano et
al., 2006; Massin y Hendrickx, 2011; Hendrickx, 2012). Además encontraron que el
oxígeno disuelto puede ser una barrera para la dispersión de especies que no toleren
condiciones de hipoxia. En el año 2008, se llevó a cabo una investigación de
caracterización geológica del fondo marino en las cuencas abisales y escarpes de
fallas transformes del Golfo, a bordo del crucero oceanográfico Atlantis, en el cual, se

9
sumergió un vehículo no tripulado en 26 localidades. Como producto alternativo de
este proyecto, se obtuvo la caracterización de la fauna abisal, encontrando algunos
equinodermos, crustáceos, moluscos y peces (Ayón-Parente et al., 2014; Hendrickx et
al., 2014).
La distribución y abundancia de organismos planctónicos también han sugerido
una división en el Golfo de California. Allen (1937) y Cupp y Allen (1938) dividieron el
Golfo en tres regiones geográficas al comparar la abundancia de diatomeas. La parte
sur está caracterizada por tener las poblaciones más pequeñas de fitoplancton en todo
el Golfo, mientras que la central cuenta con densidades más altas de productores
primarios. Sin embargo las más grandes abundancias de diatomeas se registraron en
la región norte; Posteriormente Gilbert y Allen (1943) agregan una cuarta región en el
Golfo Norte, esta división surgió porque corresponde a los antinodos encontrados por
Sverdrup (1941) quien determinó que las aguas superficiales de estas zonas se
encuentran segregadas por una onda estacionaria interna a lo largo del eje del Golfo
(fig. 4A). Otro trabajo (Round, 1967) estudió la depositación de diatomeas en los
sedimentos superficiales (50 m de profundidad), y también distingue cuatro zonas (fig.
4B), con pequeñas diferencias a la zonación de Gilbert y Allen (1943), por lo que se
concluyó que el desplazamiento lateral del fitoplancton durante la sedimentación no es
muy grande. Estudiando el mismo grupo, Gaxiola-Castro y colaboradores (1995)
encuentran tres zonas de distribución del fitoplancton durante el invierno de acuerdo a
la estratificación de la columna de agua: una bien mezclada cerca de las Grandes Islas
(asociada a baja temperatura, alta clorofila a superficial, y baja productividad primaria),
una zona parcialmente mezclada al centro del Golfo, y una porción estratificada al sur.
Estas últimas dos presentaron alta productividad primaria.

10
Figura 4 - División geográfica de acuerdo con: (A) Gilbert y Allen, 1943. (B) Round, 1967.
En los últimos años, la implementación de tecnología de percepción remota ha
permito realizar estudios de regionalización a gran escala, incluido el Golfo de
California, el cual es con frecuencia una zona libre de nubes, lo que lo hace un sitio
ideal para el uso de esta tecnología (Kahru et al., 2004). El primer ejemplo es el de
Santamaría del Ángel y colaboradores (1994) quienes con imágenes de alta resolución
del sensor del satélite CZCS estudiaron la variación espacio-temporal de la
concentración pigmentaria del GC, con el fin de encontrar una división geográfica que
fuera ecológicamente significativa. Para ello utilizaron 33 pixeles de las imágenes
satelitales en promedios semanales de 1978 hasta 1986. De ahí derivaron 14 regiones
biogeográficas dentro del Golfo (fig. 5A), cada una relacionada con aspectos como la
pesquería de escama y camarón, la distribución de algunos mamíferos marinos, y los
sitios de crianza importantes para peces. Siguiendo la misma línea de investigación,
en un estudio a meso escala Arias-Aréchiga (1998) utiliza la misma información del
sensor CZCS y propone tres grandes regiones con distintos niveles de productividad
primaria: el Golfo Norte, el cual comprende desde el delta del Río Colorado hasta el
sur de las Grandes Islas y es una zona influenciada por mareas. La región centro
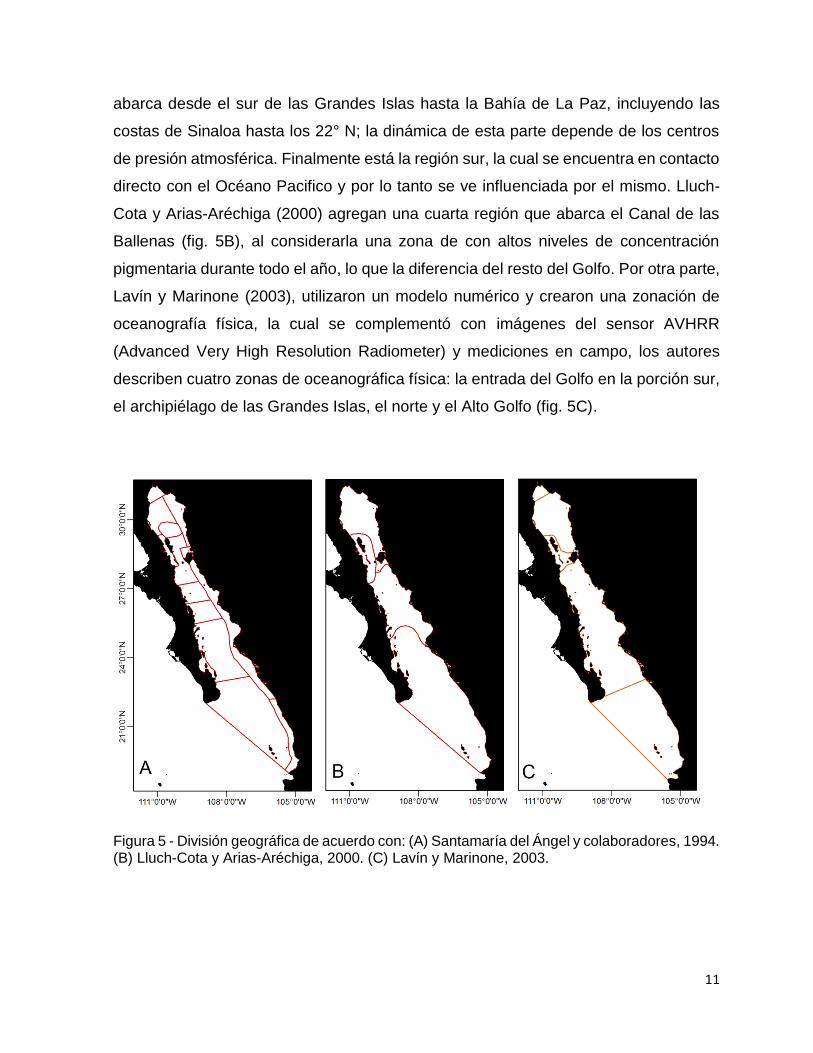
11
abarca desde el sur de las Grandes Islas hasta la Bahía de La Paz, incluyendo las
costas de Sinaloa hasta los 22° N; la dinámica de esta parte depende de los centros
de presión atmosférica. Finalmente está la región sur, la cual se encuentra en contacto
directo con el Océano Pacifico y por lo tanto se ve influenciada por el mismo. Lluch-
Cota y Arias-Aréchiga (2000) agregan una cuarta región que abarca el Canal de las
Ballenas (fig. 5B), al considerarla una zona de con altos niveles de concentración
pigmentaria durante todo el año, lo que la diferencia del resto del Golfo. Por otra parte,
Lavín y Marinone (2003), utilizaron un modelo numérico y crearon una zonación de
oceanografía física, la cual se complementó con imágenes del sensor AVHRR
(Advanced Very High Resolution Radiometer) y mediciones en campo, los autores
describen cuatro zonas de oceanográfica física: la entrada del Golfo en la porción sur,
el archipiélago de las Grandes Islas, el norte y el Alto Golfo (fig. 5C).
Figura 5 - División geográfica de acuerdo con: (A) Santamaría del Ángel y colaboradores, 1994. (B) Lluch-Cota y Arias-Aréchiga, 2000. (C) Lavín y Marinone, 2003.
12
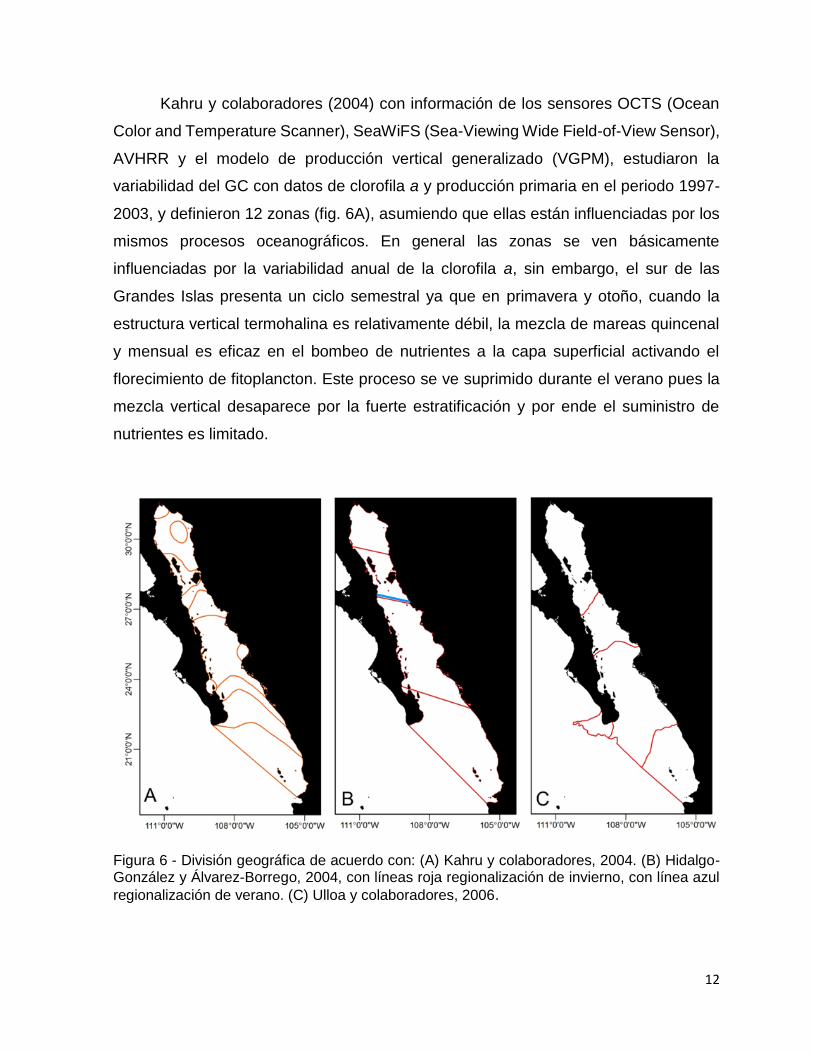
Kahru y colaboradores (2004) con información de los sensores OCTS (Ocean
Color and Temperature Scanner), SeaWiFS (Sea-Viewing Wide Field-of-View Sensor),
AVHRR y el modelo de producción vertical generalizado (VGPM), estudiaron la
variabilidad del GC con datos de clorofila a y producción primaria en el periodo 1997-
2003, y definieron 12 zonas (fig. 6A), asumiendo que ellas están influenciadas por los
mismos procesos oceanográficos. En general las zonas se ven básicamente
influenciadas por la variabilidad anual de la clorofila a, sin embargo, el sur de las
Grandes Islas presenta un ciclo semestral ya que en primavera y otoño, cuando la
estructura vertical termohalina es relativamente débil, la mezcla de mareas quincenal
y mensual es eficaz en el bombeo de nutrientes a la capa superficial activando el
florecimiento de fitoplancton. Este proceso se ve suprimido durante el verano pues la
mezcla vertical desaparece por la fuerte estratificación y por ende el suministro de
nutrientes es limitado.
Figura 6 - División geográfica de acuerdo con: (A) Kahru y colaboradores, 2004. (B) Hidalgo-González y Álvarez-Borrego, 2004, con líneas roja regionalización de invierno, con línea azul
regionalización de verano. (C) Ulloa y colaboradores, 2006.
13
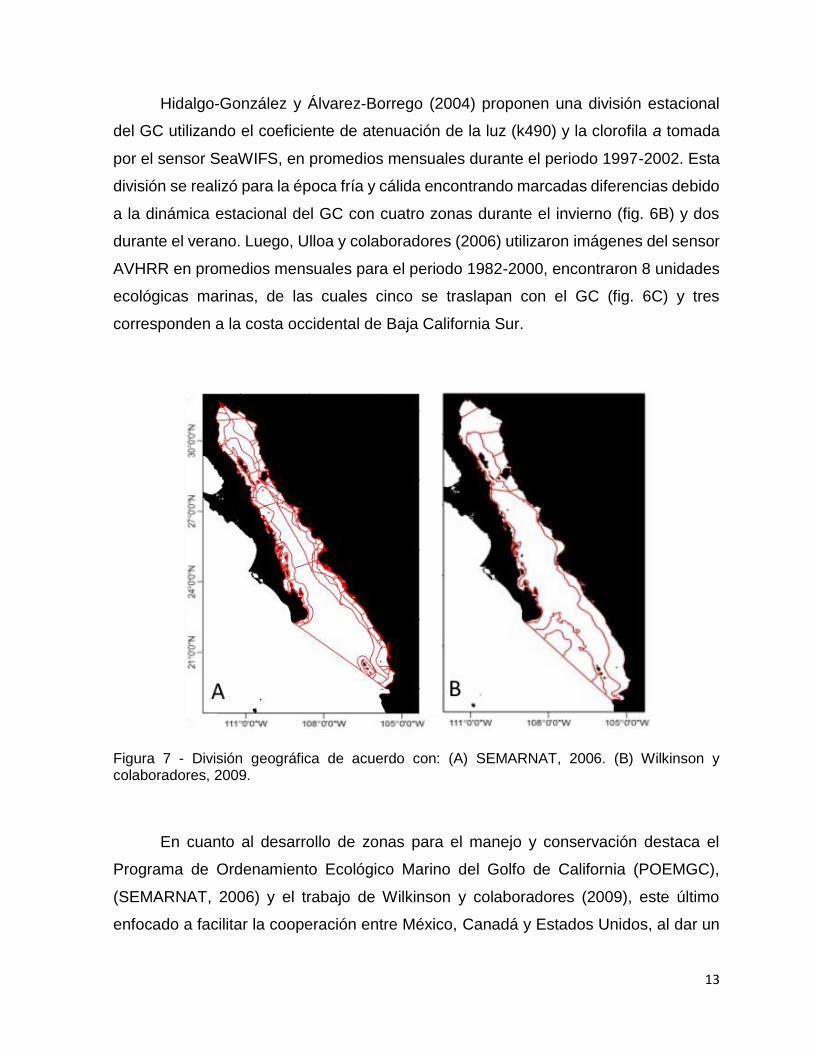
Hidalgo-González y Álvarez-Borrego (2004) proponen una división estacional
del GC utilizando el coeficiente de atenuación de la luz (k490) y la clorofila a tomada
por el sensor SeaWIFS, en promedios mensuales durante el periodo 1997-2002. Esta
división se realizó para la época fría y cálida encontrando marcadas diferencias debido
a la dinámica estacional del GC con cuatro zonas durante el invierno (fig. 6B) y dos
durante el verano. Luego, Ulloa y colaboradores (2006) utilizaron imágenes del sensor
AVHRR en promedios mensuales para el periodo 1982-2000, encontraron 8 unidades
ecológicas marinas, de las cuales cinco se traslapan con el GC (fig. 6C) y tres
corresponden a la costa occidental de Baja California Sur.
Figura 7 - División geográfica de acuerdo con: (A) SEMARNAT, 2006. (B) Wilkinson y colaboradores, 2009.
En cuanto al desarrollo de zonas para el manejo y conservación destaca el
Programa de Ordenamiento Ecológico Marino del Golfo de California (POEMGC),
(SEMARNAT, 2006) y el trabajo de Wilkinson y colaboradores (2009), este último
enfocado a facilitar la cooperación entre México, Canadá y Estados Unidos, al dar un

14
marco de referencia con el objetivo de servir de base común para ayudar en las
decisiones de manejo y conservación del medio marino. Con base en variables
oceanográficas, fisiográficas y biológicas generales determinaron al GC como una
ecorregión (Spalding et al., 2007), la cual segmentan en regiones más pequeñas (fig.
7B) con ayuda del conocimiento de expertos, distinguieron las zonas que podrían
beneficiarse de formas de manejo y medidas de conservación similares. Por otra parte
el POEMGC surge como un instrumento de política ambiental dirigido a lograr un mejor
balance entre las actividades productivas y la protección del GC, la caracterización del
GC se consiguió mediante el uso de indicadores en diferentes sectores de interés,
como lo es la pesca industrial, pesca ribereña, turismo y la conservación, siguiendo
una ponderación de expertos, se analizaron niveles de presión y fragilidad regional,
delimitando un total de 22 unidades de gestión ambiental en todo el GC (fig. 7A) con
características homogéneas en términos de los patrones regionales de presión,
fragilidad y vulnerabilidad. (SEMARNAT, 2006).

15
JUSTIFICACIÓN
Los estudios previos sobre regionalización del Golfo de California han seguido
enfoques con líneas específicas, es decir, se basaron por separado en condiciones
oceanográficas, biológicas o de manejo de recursos. Estos trabajos resultaron útiles
para los fines para los cuales fueron elaborados, permitiendo un mejor entendimiento
sobre el GC y sus características. Sin embargo, los avances tecnológicos y la cada
vez mayor disponibilidad de información permiten actualmente lograr una
regionalización más precisa e integral basada en información sobre una mayor
cantidad de variables bióticas y abióticas. Esto representa una mejora sensible sobre
trabajos que usaron, por ejemplo, únicamente la clorofila a, la temperatura, o la
distribución de algunos grupos taxonómicos del plancton.
En la parte biológica también hay un gran avance, al existir nutridas bases de
datos y listas taxonómicas actualizadas para diversos grupos de especies de peces e
invertebrados que habitan en el GC; un buen ejemplo es la base de datos de
macroinvertebrados marinos del Golfo de California (Brusca y Hendrickx, 2008),
producto de años de esfuerzos de investigadores y organizaciones, que incluye datos
sobre clasificación taxonómica, distribución geográfica y preferencias de hábitat
(Brusca, 2010).
Diferentes autores hacen referencia que para una regionalización es necesario
tomar en cuenta elementos perdurables en el tiempo, como sustrato, sedimentos,
incluso la fauna bentónica (Hayden et al., 1984; Zacharias y Roff, 2000; Arias-
Aréchiga, 1998; Campero, 2003) dado que a diferencia de otros grupos, la dispersión
de las especies bentónicas depende esencialmente de la presencia/ausencia de
barrera físicas o ecológicas, y de la capacidad de dispersión de sus estadios larvarios,
si es que los presentan (Hendrickx, 2008), por lo que regionalizar la distribución de los
macroinvertebrados, los cuales son en su mayoría bentónicos, nos ayudaría a
representar mejor los patrones generales de la distribución de la biota, ya que es uno
de los grupos más diversos en el GC (Brusca et al., 2005)
Además de la disponibilidad de información, en la actualidad existen distintas
herramientas para el procesamiento de la información como los sistemas de

16
información geográfica (SIG) que permiten la realización operaciones geoespaciales y
el manejo de gran cantidad de datos georreferenciados para su representación
cartográfica. (Córdova et al., 2006).
Contar con regionalizaciones oceanográficas y biológicas nos permitirá
determinar la correspondencia existente entre ambas, y al integrar ambos factores,
encontrar los biomas del Golfo de California, los cuales son regiones naturales que
comparten características de clima, flora y fauna, por lo que se obtendría una zonación
con un mayor sustento, útil para la ciencia básica, la conservación y el manejo.

17
OBJETIVOS
Objetivo General
Proponer una regionalización del Golfo de California, con base en el conjunto de la
distribución geográfica de macroinvertebrados y las condiciones ambientales del
océano.
Objetivos Específicos
A) Generar una regionalización oceanográfica, a partir de variables ambientales.
B) Generar una regionalización biológica a partir de la distribución geográfica de
grupos selectos de macroinvertebrados.
C) Generar biomas marinos a partir de la fusión de las regionalizaciones anteriores.

18
ÁREA DE ESTUDIO
El Golfo de California es un cuerpo de agua que separa a la península de Baja
California del macizo continental en el occidente de México, tiene una edad de entre
6.5 y 13 millones de años (Simison, 2006). Se extiende de noroeste a sureste con una
extensión de 1,070 km, variando en ancho de 100 a 200 km (Lindsay y Engstrand,
2002). Es uno de los cuerpos oceánicos más jóvenes geológicamente hablando, y
llega casi a los 4,000 m de profundidad en las fosas abisales (Álvarez-Borrego y Lara-
Lara, 1991). El Golfo abarca la costa peninsular de los estados de Baja California y
Baja California Sur (la cual es principalmente rocosa), mientras en la costa continental,
en los estados de Sonora, Sinaloa y Nayarit, presenta mayor diversidad de hábitats
como playas arenosas, estuarios y zonas lagunares (Castro-Aguirre et al., 1996; Lluch
Cota et al,. 2005; Simison, 2006).
Figura 8 - El Golfo de California, limitado al sur por una línea entre Cabo San Lucas, BCS, y Bahía de Banderas, Nayarit.

19
El patrón de circulación de las aguas superficiales está relacionado al sistema
de vientos locales, las mareas, distribución de la densidad, además del intercambio de
calor y agua con la atmosfera y el forzamiento con el Océano Pacífico (Lavín et al.,
1997; Ripa, 1997). El patrón de vientos, los cuales vienen del noroeste en invierno, y
del sureste en verano, introduce aire húmedo del sur hacia el GC, creando condiciones
climáticas más tropicales (Merrifield y Winant, 1989) y originando precipitaciones en
ciertas épocas del año (Torres-Orozco, 1993). Se ha registrado la presencia de la
Corriente de California, la cual ingresa al GC durante los meses de enero y diciembre,
y sale durante el mes de marzo, en Mayo se detecta la presencia de la Corriente
Costera de Costa Rica, la Corriente Norecuatorial entra de agosto a septiembre, en
Octubre la Corriente del Golfo la cual fluye hacia el Océano Pacífico.
En la superficie del GC se encuentran dos masas de agua, las cuales llegan
hasta los 150 m de profundidad, la primera corresponde al Agua del Golfo de California,
la cual se localiza en la región norte y se distingue por su alta salinidad, la segunda es
el Agua Superficial Ecuatorial, la cual abarca desde la entrada del Golfo hasta la
cuenca de Guaymas en verano y en invierno solo se encuentra en la boca del GC
(Molina-Cruz, 1986; Lavín et al., 1997). Entre los 150 y 500 m se encuentra el Agua
Subsuperfial Subtropical (ASS), hasta la isoterma de los 9 °C con una salinidad menor
a los 35 unidades prácticas de salinidad (UPS; Lavín et al., 1997), durante el otoño
presenta la máxima intrusión en la parte norte del Golfo (Arias-Aréchiga, 1998).
Después del ASS se encuentra entre los 500 y 1200 m de profundidad el Agua
Intermedia del Pacífico, la cual constituye el 33% del volumen del Golfo de California
con una temperatura entre 4 y 9 °C, el límite norte de esta masa de agua se localiza
en la cuenca Tiburón (Torres-Orozco, 1993; Lavín et al., 1997). Finalmente desde los
1200 m y hasta el fondo delimitado por la isoterma de 4 °C, se encuentra el Agua
Profunda del Pacífico, la cual ocupa el 41% del volumen del GC, y presenta mayor
estabilidad debido a que está aislada de la influencia meteorológica que afecta a las
capas superficiales (Torres-Orozco, 1993; Lavín et al., 1997).
El Golfo de California presenta tres mecanismos de enriquecimiento: La mezcla
por mareas, la circulación termohalina y las surgencias inducidas por el viento. Al norte

20
del Golfo destaca la mezcla por mareas, principalmente en la región de las Grandes
Islas, las surgencias costeras inducidas por el viento se presentan en ambas costas
(en la occidental durante el verano, y en la oriental durante el invierno), y la circulación
termohalina se produce por la mezcla de temperatura y salinidad de las distintas masas
de agua del Golfo (Álvarez-Borrego, 2002). Presenta una estacionalidad oceanográfica
muy marcada (Álvarez-Borrego, 2010), siendo afectado interanualmente por el
fenómeno niño y la niña (Soto-Mardones et al., 1999) principalmente en la boca del
Golfo, por lo que disminuye significativamente la clorofila a y producción primaria
(Escalante et al., 2013). El GC es reconocido como uno de los cinco mares más
productivos y diversos del mundo (Brusca, 2010) y dadas sus características ha sido
nombrado como una ecorregión por Comisión para la Cooperación Ambiental de
América del Norte (Wilkinson et al., 2009), ya que en el existe gran diversidad de
ecosistemas marinos y costeros, que incluyen alrededor de 350,000 hectáreas de
manglares, cinco géneros de tortugas marinas, 32 especies de mamíferos marinos
(que representan el 38% de las especies de cetáceos que se conocen en el mundo),
181 aves acuáticas, y 875 especies de peces de los cuales 77 son considerados
endémicos (SEMARNAT, 2006). A la gran biodiversidad de la ecorregión se le suma
su complejidad fisiográfica, ya que hay más de 920 islas e islotes desde la
desembocadura del Río Colorado hasta Cabo Corrientes, en Jalisco, muchas de ellas
habitadas por especies terrestres endémicas o especies amenazadas (Brusca et al.,
2005; Ulloa et al., 2006). Los macroinvertebrados son el grupo más numeroso al contar
con más de 4,900 especies, aunque se estima que este dato representa solo el 70%
del número de especies que realmente habitan la zona, del cual un 16% (782 especies)
de invertebrados endémicos se consideran endémicos (Brusca et al., 2005; Brusca,
2010).
El Golfo de California es una de las regiones más importantes para la pesquería
en México, ya que el valor de la producción pesquera equivale al 70% del rubro
nacional (unos 300 millones de dólares), con un volumen de medio millón de toneladas
anuales, lo representa el 50% del peso total de la captura nacional, esta actividad
genera más de 50 mil empleos (Ulloa et al., 2006).

21
METODOLOGÍA
Para regionalizar un área, primero es necesario definir los límites geográficos
de la misma. En el caso del GC los limites varían dependiendo del autor, por ejemplo,
anteriormente se estableció que el límite sur es a los 23° 38’N (International
Hydrographic Organization, 1953), ubicado al norte de Mazatlán, mientras otros
autores, lo extendieron hasta Cabo Corrientes (Walker, 1960; Roden, 1964; Roden y
Emilsson, 1979), este límite sur ha sido ampliamente debatido, sobre todo debido a
que el GC no necesariamente coincide con los límites de la provincia zoo geográfica
del Cortés que han documentado algunos autores (Hendrickx et al., 2002; Robertson
y Cramer, 2009; Brigss y Bowen, 2012). Por lo que se tomó como referencia los límites
geográficos establecidos en los polígonos del Programa de Ordenamiento Ecológico
del Golfo de California (SEMARNAT, 2006), límite establecido por una línea entre Cabo
San Lucas y el límite norte de Bahía de Banderas.
Una vez determinada el área de estudio, es necesario segmentarla en unidades
más pequeñas para caracterizar cada unidad, en este caso desde la perspectiva
oceanográfica y biológica, y de ese modo poder establecer patrones de similitud entre
ellas. Con el uso de la herramienta “Fishnet” del software ArcMap 10.2 se creó una
rejilla con un tamaño de celda de 9 km2, para desarrollar todos los análisis. Se usó
dicha resolución espacial debido a que la información oceanográfica medida por
sensores a partir del color del océano en nivel tres se encuentra a 4 y 9km2; sin
embargo, al tratar de trabajar con gradículas de 4 km2, el tiempo de procesamiento,
alto requerimiento de memoria y recursos del sistema, hizo que el análisis se tornara
inviable. Como resultado de este proceso se obtuvieron de 3,502 celdas a 9 km2 las
cuales sirvieron de base para la creación de la base de datos ambiental (variables
oceanográficas) y biológica (distribución de macroinvertebrados).

22
Figura 9 - Rejilla de trabajo para la regionalización del Golfo de California a 9km2 de resolución espacial.
Información oceanográfica
Considerando que el mar es un ambiente muy dinámico, es necesario tener
series de tiempo extensas para tener una buena caracterización de sus condiciones a
través de promedios climatológicos y de las posibles regionalizaciones que el océano
presente (Millán-Núñez et al., 2006), ya que de este modo es posible conocer los
efectos de la variabilidad temporal e interanual, y de algunos fenómenos de gran
escala como la Oscilación del Sur de El Niño (Escalante et al., 2013). En esta tesis,
para la caracterización oceanográfica del GC se utilizó dos fuentes de información
(tabla I), los datos medidos por sensores remotos, e información histórica in situ,
almacenada en la base de datos del Centro Nacional de Datos Oceanográficos (NODC
por sus siglas en inglés) de la NOAA. (National Ocean and Atmospheric Administration;
http://www.nodc.noaa.gov/).

23
TABLA I - Variables oceanográficas utilizadas para la regionalización.
Variable Unidad Fuente Promedio
Materia coloreada disuelta m-1 MODIS – Aqua Mensual (2003 - 2012)
Clorofila a mg m−3 MODIS – Aqua Mensual (2003 - 2012)
Atenuación de la luz m-1 MODIS – Aqua Mensual (2003 - 2012)
Radiación fotosintéticamente
activa
Einstein m2
D
MODIS – Aqua Mensual (2003 - 2012)
Carbono inorgánico mol m-3 MODIS – Aqua Mensual (2003 - 2012)
Carbono orgánico mg m-3 MODIS – Aqua Mensual (2003 - 2012)
Temperatura superficial °C MODIS – Aqua Mensual (2003 - 2012)
Fosfato µmol/l NODC Anual (1939 - 2007)
Salinidad UPS NODC Anual (1939 - 2007)
Silicatos µmol/l NODC Anual (1939 - 2007)
Oxígeno disuelto ml/l NODC Anual (1939 - 2007)
Información oceanográfica vía sensores remotos
Los datos del color del océano se obtuvieron de la Agencia Aeroespacial
Nacional de los Estados Unidos (NASA, por sus siglas en inglés), en la página de
internet del Servicio de Visualización en línea de datos radiométricos del color del
océano (Ocean Color Radiometry Online Visualization and Analysis, GIOVANNI;
http://disc.sci.gsfc.nasa.gov/giovanni) adquiridos del sensor MODIS – Aqua a 9 km2, a
nivel superficial del mar y en promedios mensuales de un periodo de 10 años (2003 -
2012). Los datos se descargaron en formato ASCII, los cuales fueron introducidos al
software ArcMap 10.2 para su transformación a formato raster, el cual, es un formato
de archivo usado por los sistemas de información geográfica para almacenar
información en una matriz de datos, en este caso, se utilizó como base la rejilla de
trabajo de 3,502 pixeles validos definida previamente.
Las siete variables descargadas del portal GIOVANNI (tabla I), generaron 840
archivos rasters (120 por variable) y para el trabajo se calculó el año tipo de cada
variable, el cual es el promedio climatológico de cada mes del año a lo largo del periodo

24
(por ejemplo, el promedio de todos los meses de enero, generan el enero del año tipo).
Esto permitió representar la tendencia mensual que tuvo el GC en los últimos 10 años
para cada aspecto estudiado del océano. Una vez establecido el año tipo se obtuvieron
los valores máximos y mínimos por pixel para cada variable, para representar las
condiciones extremas que se dan estacionalmente en el GC y utilizar estos datos para
la regionalización oceanográfica.
Información oceanográfica in situ
Las variables que no pueden ser medidas por el color del océano, como lo son
fosfatos, silicatos, oxígeno disuelto y salinidad fueron descargadas del NODC. Se
obtuvieron 2,024 sitios de muestreo distribuidos en toda el área de estudio, los cuales
fueron tomados in situ entre 1939 y 2007 por diferentes cruceros oceanográficos en
distintos meses de año. Dado que para la toma de muestras no se lleva a cabo de
manera sistemática ni periódica para todo el Golfo de California (fig. 10), no se pudo
determinar el año tipo para estas variables, por lo que se obtuvo únicamente el
promedio anual interpolando los valores de las variables usando la rutina de
ponderación inversa a la distancia (“inverse distance weighting”), el cual ha
demostrado ser un método de interpolación eficiente para conservar la variabilidad de
las condiciones oceanográficas además de minimizar los errores (Sheng, 1999; Torres
et al., 2008). Estas nuevas capas de información fueron añadidas a la base de datos
océano gráfica, por lo que fueron creadas una escala homogénea a la rejilla de trabajo
establecida previamente.

25
Figura 10 - Distribución espacial de las mediciones in situ del periodo 1939 – 2007 obtenida de la NODC.
Información Biológica
Para generar los registros de distribución de las especies se utilizó la base de
datos de invertebrados del Golfo de California (Brusca y Hendrickx, 2008), la cual
incluye información geográfica y batimétrica para 4,944 especies (tabla II). Se
eliminaron las especies que no tuvieran información sobre los límites geográficos o
batimétricos completos, dado que no es posible representar una distribución
geográfica confiable sin estos datos. Aunque la base de datos citada revisa todos los
fila de invertebrados presentes en el Golfo, (para la tesis) se optó por manejar solo los
cinco grupos con mayor cantidad de especies registradas. Los fila seleccionados
fueron: Arthropoda, Mollusca, Annelida, Echinodermata y Cnidaria, los cuales
acumulan más del 90% de los macroinvertebrados y son los mejor representados en
la base de datos (Brusca, comunicación personal, julio 2013).

26
TABLA II - Número de Invertebrados del Golfo de California por nivel taxonómico.
Con la lista de especies depurada se obtuvieron 3,496 especies (tabla II) las
cuales representan el 70% de las especies totales de macroinvertebrados registrados
en el GC. Con este listado, se creó un script en R con el cual, a partir de los límites
geográficos y batimétricos, generó un raster con la distribución geográfica de cada
especie. La matriz final presenta valores de 0 y 1, para señalar la presencia o ausencia
de cada especie en cada uno de los 3,502 sitios.
Phylum Clases Ordenes Familias Géneros Especies Procesados
Annelida 2 15 62 302 736 556
Arthropoda 3 12 168 540 1,065 903
Brachipoda 2 2 2 2 5 -
Bryozoa 2 3 58 97 170 -
Chaetognatha 1 2 4 12 20 -
Chordata 2 4 10 20 42 -
Cnidaria 3 16 73 129 262 189
Ctenophora 2 3 3 4 4 -
Echinodermata 3 26 66 157 264 244
Echiura 1 1 2 4 4 -
Hemichordata 1 - 1 3 3 -
Mollusca 6 27 221 693 2,199 1,605
Nemertea 2 4 7 12 17 -
Platyhelminthes 1 1 8 19 22 -
Porifera 2 14 38 69 120 -
Spincula 2 4 5 8 11 -
Total 35 134 728 2,071 4,944 3,496

27
Regionalización del Golfo de California
Regionalización oceanográfica
Una vez generada la base de datos ambiental, compuesta por las variables
medidas por sensores y las variables medidas in situ, los datos oceanográficos fueron
estandarizados para reducir el intervalo de los datos de 0 a 1 en todas las variables y
tener una escala homogénea para evitar distorsión de resultados (Prieto-Guerra,
2006). Para generar la regionalización se utilizó el análisis de similitud, el cual es uno
de los métodos que se utiliza para establecer grupos de individuos, unificando dentro
de un mismo grupo a aquellos elementos con características similares (Linares, 1990).
El coeficiente de similitud o disimilitud de los elementos se selecciona de acuerdo al
tipo de variables a analizar, ya que la confiabilidad de los resultados dependerán del
método utilizado (Miranda, 1997). Este análisis, se llevó a cabo con las variables
oceanográfica estandarizadas y acomodadas en una matriz de 3,502 renglones (rejilla
del GC) y 18 columnas (siete variables en dos temporadas del año para representar la
variabilidad estacionaria del GC, y los cuatro promedios anuales de los nutrientes, los
cuales fueron medidos in situ). El método utilizado para calcular la similitud
oceanográfica del Golfo de California fue el de distancia euclidiana, una vez obtenida
la distancia entre los sitios, el agrupamiento de estos se realizó por el método de Ward
(1963) el cual minimiza la varianza entre los sitios y los agrupa, si se tiene más de una
variable, en lugar de la varianza se unen los grupos cuya inercia (suma la diagonal
principal de la matriz de varianzas y covarianzas) sea la mínima (Álvarez-Cáceres,
1994; Zar, 1999). En resumen, este método es útil porque maximiza la homogeneidad
dentro de los grupos. Finalmente para determinar el número de grupos a formar, se
analizó la tabla mediante el índice k-promedios, el cual es un proceso iterativo en el
cual las agrupaciones se definen con base en la distancia euclidiana entre los
elementos. Este coeficiente permite reducir la variabilidad de los elementos dentro de
cada grupo, al tiempo que maximiza la variabilidad entre ellos (Kintigh y Ammerman,
1982; Chollett et al., 2012).

28
Regionalización biológica
Una vez obtenido el raster de distribución geográfica de cada especie, se
crearon cinco tablas, una para cada phylum indiando con 0 y 1 la presencia o ausencia
de las especies en cada sitio. Para tener un criterio de la cantidad de regiones a formar
en cada phylum, se obtuvo la suma de cuadrados de k - promedios (similar al realizado
en la zonación oceanográfica) y una vez establecido el número de grupos, se calculó
el índice de similitud de Bray-Curtis (Krebs, 1989) y se formaron grupos por el método
de Ward (1963). El procedimiento fue el mismo aplicado a las variables oceanográficas
y se utilizó porque los métodos no son sensibles a valores extremos, tendiendo a
formar grupos compactos y de igual tamaño. Al final se obtuvieron cinco
regionalizaciones biológicas (una por cada phylum). Luego, para obtener una
regionalización única de invertebrados, se creó una matriz de 3,502 renglones y cinco
columnas. Cada una de las columnas representan de manera cualitativa las regiones
que fueron asignadas por el análisis previo de cada zonación biológica, esta matriz se
analizó mediante el coeficiente de similitud de Gower (1971), el cual permite la
utilización de variables cualitativas, cuantitativas y binarias. Mediante este coeficiente
de similitud se puede determinar el grado de semejanza o similitud entre elementos
(en este caso, pixeles); Entre las características de este coeficiente se encuentra que
cuando todos los caracteres son cualitativos, es equivalente al coeficiente de
coincidencias simple, definido como la relación del número total de coincidencias y el
número total de caracteres, además de que es posible ponderar las variables, dando
mayor o menor peso a las distintas variables, dependiendo el papel que juegue cada
una en la ordenación de grupos (Prieto-Guerra, 2006). En este caso, la ponderación
se realizó por el porcentaje de especies que aportó cada phylum, siendo el phylum
Mollusca el de mayor contribución con el 46%, seguido por Arthropoda (25%) y
Annelida (16%), y finalmente el phylum Echinodermata y Cnidaria fueron las de menor
peso con 7 y 5% respectivamente, una vez ponderado el aporte de cada phylum se
obtuvo la regionalización biológica única. Posteriormente se crearon matrices de
presencia/ausencia de todas las especies a nivel región, para conocer la similitud de
especies entre regiones, además de la cantidad de especies dentro de cada una de

29
ellas (riqueza específica) y mostrar la diferenciación en la cantidad y composición de
especies de cada región.
Biomas del Golfo de California
Para generar los biomas se incluyeron ambos factores (biológico y
oceanográfico) y se siguió un proceso similar al de la obtención de la regionalización
de invertebrados, ponderando la contribución de cada elemento para la regionalización
final; Al tratarse de datos de naturaleza distinta, se dejó al grupo biológico con el 50%
(con su respectiva ponderación a nivel phylum) y el otro 50% a la regionalización
oceanográfica previamente obtenida, de esta forma, se realizó la regionalización de
forma integral, considerando la caracterización oceanográfica con la distribución de las
especies.

30
RESULTADOS
Regionalización oceanográfica
El análisis de k - promedios indicó que el número de grupos para formar las
regiones dentro del GC por condiciones oceanográficas fue de 12 ya que con esta
cantidad de grupos se alcanza la asíntota de la curva de la suma de cuadrados (fig.
11), sin embargo, el análisis de agrupamiento de los datos encontró dos regiones con
dos secciones cada una pero geográficamente separados; es decir, la diferencia de
las características entre los sitios era mínima, pero dado que espacialmente no eran
contiguos, aquí se tomaron en cuenta como grupos diferentes. Por este motivo se
decidió subir a 14 el número de regiones (tabla III). De las 14 regiones, siete están
paralelas a la línea de costa, por lo que se consideran costeras y el resto en mar
abierto, es decir, oceánicas.
Figura 11 - Índice de k – promedios para calcular el número de regiones oceanográficas del Golfo de California.
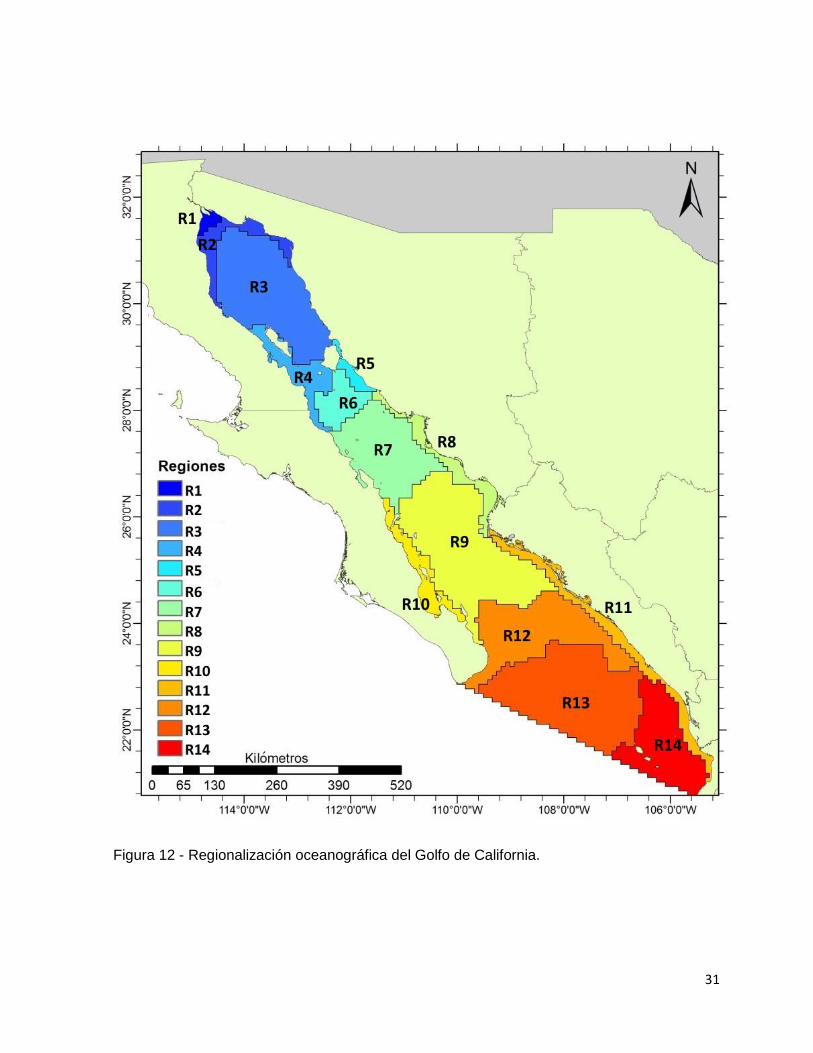
31
Figura 12 - Regionalización oceanográfica del Golfo de California.

32
TABLA III - Lista de regiones oceanográficas del Golfo de California.
Figura 13 - Cobertura de las regiones oceanográficas.
R11%
R23%
R313%
R44% R5
1%
R63%
R710%
R84%
R917%R10
3%
R116%
R1211%
R1316%
R148%
Regiones Oceanográficas del Golfo de California
Región Oceánica Costera
Norte del GC R1) Delta del Río Colorado
R2) Alto Golfo
R3) Golfo Norte
R4) Grandes Islas
R5) Isla Tiburón
R6) Grandes Islas sur
Centro del GC R7) Cuenca de Guaymas
R8) Sur de Sonora
R9) Cuenca Carmen - Farallón
R10) La Paz – Loreto
Sur del GC R11) Sinaloa – Nayarit
R12) Cuenca Pescadero
R13) Boca del Golfo
R14) Islas Marías

33
Figura 14 - Dendrograma de similitud oceanográfica a nivel región.
Características de las regiones oceanográficas
Los resultados indican que las regiones oceánicas ocupan la mayor parte del
GC siendo aproximadamente el 78% de la superficie total (fig. 13); el resto (22%)
corresponde a las siete regiones asociadas a la costa. La similitud entre regiones,
como se aprecia en la figura 14, se asocian en 4 grandes grupos. En el primero se
encuentran las regiones oceánicas del sur del Golfo, desde la boca hasta la región de
la cuenca Carmen - Farallón. El grupo dos comprende el margen costero desde Nayarit
hasta Sonora, incluyendo la región de las Grandes Islas. El grupo tres contiene
solamente dos regiones: el Alto Golfo y el delta del Río Colorado, y finalmente el grupo

34
cuatro, incluye a las regiones oceánicas del centro y norte del Golfo, e incluye la región
costera de La Paz – Loreto.
A manera de información se denotarán las características de las 14 regiones
oceanográficas designadas, en orden de norte a sur (fig. 12).
La región 1, el Delta del Río Colorado abarca la zona norte de la reserva Alto
Golfo de California y Delta del Río Colorado, y corresponde al 0.77% del GC en
superficie (fig. 13). Esta sección presenta la mayor variabilidad de temperatura con un
intervalo superior a 15°C (de 15.31 a 31.85 °C; fig. 15), con concentraciones altas de
clorofila a la mayor parte del año (de 3.22 a 3.66 mg m−3; fig. 16), y de carbono
inorgánico (de 0.027 a 0.048 mol m-3; fig. 19), y de carbono orgánico particulado (de
331 a 358 mg m-3; fig. 20). Los niveles de atenuación de la luz son elevados la mayor
parte del año, y con poca variabilidad (de 0.23 a 0.28 m-1, fig. 17). La región presenta
la mayor salinidad de todo el GC con valores superiores a los 36 UPS (tabla IV).
La región 2, correspondiente al Alto Golfo abarca desde la bahía de San Luis
Gonzaga en B.C. hasta El Desemboque, Son. Esta región ocupa aproximadamente el
3% de la superficie del GC (fig. 13), y al igual que la región Detal del Río Colorado
presenta un amplio intervalo de temperatura (de 16.12 a 31.6 °C; fig. 15). Esta región
presenta el intervalo más amplio de radiación fotosintéticamente activa (de 24.45 a
59.57 Einstein m2 D; fig. 18) además de la mayor concentración de materia coloreada
disuelta de todo el GC (de 4.93 a 6.71 m-1; fig. 21), y es la segunda más salina de
todas con 35.71 UPS en promedio (tabla IV).
La región 3, el Golfo Norte es la de mayor área en latitudes superiores a los
28°N y ocupa el 13% de la superficie del GC (fig. 13), colinda con la región Alto Golfo
y su límite al sur alcanza el Canal de Ballenas y las Grandes Islas, incluyendo parte de
la costa central del estado de Sonora. La temperatura oscila entre 16.79° y 30.79 °C
(fig. 15). Esta región presenta la mayor radiación fotosintéticamente activa en promedio
con amplio intervalo (de 25.84 a 60.34 Einstein m2 D; fig. 18), así como la menor
variación en la concentración de materia coloreada disuelta (de 3.57 a 4.9 m-1; fig. 21).
Es la segunda región con mayor oxígeno disuelto con 5.64 ml/l.

35
La región 4, las Grandes Islas comprende desde el Canal de Ballenas hasta
San Francisquito, BCS, limitan al este con la Isla Tiburón, y ocupa el 4% de la superficie
del Golfo de California. Es la tercera zona con las temperaturas más frías (16.37°C;
fig. 15) solo superada por la región Delta del Río Colorado y el Alto Golfo, es la segunda
con la mayor radiación fotosintéticamente activa, solo por debajo de región Golfo
Norte; además tiene el máximo intervalo de irradiancia (de 27.29 a 60 Einstein m2 D;
fig. 18). También tiene la menor variabilidad en la concentración de materia coloreada
disuelta (de 3.96 a 5.43 m-1; fig. 21). Esta es la región de mayor concentración de
silicatos 19.71 µmol/l (tabla IV).
La región 5, Isla Tiburón, está situada entre la isla del mismo nombre y el macizo
continental, abarca el Canal del Infiernillo hasta el norte de la Isla San Pedro Nolasco,
y es una de las secciones con menor área (1% del GC). Esta región presenta la
temperatura promedio más alta con amplio intervalo de variabilidad (de 16.53 a 32 °C;
fig. 15). La concentración de clorofila a varía de 1.68 a 7.56 mg m−3 (fig. 16). También
es la segunda región con mayor concentración de carbono orgánico particulado con
concentraciones que van desde 296.89 hasta 700 mg m-3 (fig. 20). Presenta la mayor
atenuación de la luz (de 0.18 a 0.54 m-1; fig. 17). La salinidad media de la región es de
35.24 UPS y es la de mayor concentración de fosfatos con 1.35 µmol/l, la segunda con
los valores más altos de silicatos 16.80 µmol/l y la de mayor concentración de oxígeno
disuelto con 5.70 ml/l (tabla IV).
La región 6, las Grandes Islas sur se sitúa al sur de las Islas de la Cintura, entre
la región de las Grandes Islas y la región Isla Tiburón, con límites al sur hasta San
Francisquito, BCS, e incluye una zona oceánica donde se encuentra la Isla San Pedro
Mártir. Abarca aproximadamente el 3% del Golfo de California (fig. 13) y presenta altos
valores de radiación fotosintéticamente activa (59.74 Einstein m2 D), solo superada por
la región Golfo Norte y la región Grandes Islas (fig. 18). Presenta además valores más
bajos de concentración de materia coloreada disuelta y es la segunda con mayor
concentración de fosfatos (1.32 µmol/l), solo por debajo de la región Isla Tiburón (tabla
IV).

36
La región 7, la Cuenca de Guaymas corresponde a la zona oceánica que se
ubica en el centro del Golfo de California y además abarcan la zona costera peninsular
desde el sur de Bahía San Francisquito hasta el norte de Isla del Carmen, limitando al
este con la región Sur de Sonora e incluye el área de la cuenca de Guaymas. La región
ocupa el 10% del GC (fig. 13), y siendo una zona oceánica presenta características
oceanográficas intermedias (temperatura de promedio anual de 24.39°C) respecto a
las condiciones del Golfo (temperatura promedio anual de 24.7 °C). Por lo que puede
decirse que es una zona de transición entre las regiones con características tropicales
(regiones 11 a la 14) y templadas (regiones 1 a la 6).
La región 8, Sonora Sur limita al norte con la región Isla Tiburón y al sur con el
norte del estado de Sinaloa es una zona costera que limita al oeste con la región
Cuenca de Guaymas. Esta región comprende el 4% de la superficie del Golfo de
California (fig. 13) y es una de las regiones con la mayor temperatura superficial del
mar, alcanzando su máximo durante el mes de agosto con 31.94 °C (fig. 15). Esta zona
presenta uno los mayores valores de clorofila a de todo el Golfo con 7.25 mg m−3, esto
ocurre en el mes de marzo (fig. 16), tiene además altos valores de carbono orgánico
particulado (de 160.73 a 700.83 mg m-3; fig. 20) y altos valores de atenuación de la luz
(de 0.10 a 0.53 m-1; fig. 17), con el máximo alcanzado en el mes de marzo.
La región 9, la Cuenca Carmen – Farallón es una región oceánica que limita al
norte con la región Cuenca de Guaymas, va desde la Isla del Carmen hasta Punta
Arenas en Baja California Sur, esta región abarca las cuencas de Carmen y Farallón.
La región ocupa la mayor área del Golfo de California con el 17% de la superficie total
(fig. 13); presenta bajos niveles de clorofila a (de 0.23 a 1.54 mg m−3; fig. 16) con un
promedio anual de 0.78 mg m−3. La región tiene además bajos valores de atenuación
de la luz (de 0.04 a 0.13 m-1; fig. 17) con un promedio anual de 0.088 m-1, y es de las
zonas con mayor intervalo en sus valores de concentración de materia coloreada
disuelta (de 1.8 a 5.02 m-1; fig. 21), el valor más alto lo alcanza durante el mes de abril
y el mínimo ocurre durante el mes de agosto. También es la segunda región con los
valores más bajos de silicatos (4.6 µmol/l) solo por encima de la región La Paz-Loreto
(tabla IV).

37
La región 10 (La Paz – Loreto) ocupa el 3% de la superficie del GC y abarca la
zona costera desde Isla Coronados hasta la Isla Cerralvo, incluyendo a todas las islas
en ese tramo y la Bahía de La Paz. Ahí, la temperatura fluctúa de 20.23° a 30.5 °C (fig.
15) con un promedio anual de 25.11 °C, alcanza su valor máximo de temperatura en
el mes de septiembre y el mínimo durante el mes de febrero. Presenta además valores
medios de clorofila a respecto a otras regiones, con valores de 0.37 a 2.10 mg m−3 (fig.
16). Esta región presenta un amplio intervalo en la concentración carbono orgánico
particulado (87.89 - 479.09 mg m-3; fig. 20) y es una de las tres regiones que alcanza
la mayor concentración de materia coloreada disuelta en el Golfo con 5.9 m-1, esto
ocurre durante el mes de abril (fig. 21). Esta región presenta la menor concentración
de silicatos de todos las regiones (3.07 µmol/l; tabla IV), muy por abajo del promedio
para el Golfo de California el cual se sitúa arriba de 10 µmol/l.
Región 11 (Sinaloa – Nayarit) abarca la zona costera de los estados de Sinaloa
y Nayarit. Es una de las áreas con menor variación de temperatura con valores desde
21.68° hasta 31.77 °C (fig. 15) con una media anual de 26.68 °C. Similarmente, es una
de las tres regiones con menor intervalo de radiación fotosintéticamente activa (de
30.83 a 57.23 Einstein m2 D; fig. 18) y con pequeñas variaciones en la cantidad de
carbono inorgánico particulado con valores de 0.003 a 0.005 mol m-3 (fig. 19), presenta
además variaciones en la clorofila a con valores desde 1.30 hasta 6.76 mg m−3 (fig.
16) y un intervalo en el coeficiente de atenuación de la luz al registrar valores de 0.145
a 0.490 m-1(fig. 17). La región presenta una las mayores concentraciones de materia
coloreada disuelta en todo el Golfo con 6.15 m-1, esto ocurre en el mes de marzo y
este valor solo es superado por la región del Alto Golfo (fig. 21) en el mes de abril.
La región 12 (Cuenca Pescadero) abarca la zona costera sur de la península de
Baja California, desde Punta Arenas hasta Cabo San Lucas, y abarcan la cuenca El
Pescadero. Ocupa el 11% de la superficie del GC y es una de las tres regiones con
menor variabilidad de temperatura (con valores de 21.33° hasta 30.81°C; fig. 15), con
un promedio anual de 26.11 °C. Se caracteriza también por ser una de las dos zonas
con menor concentración de clorofila a (registra de 0.19 a 1.25 mg m−3; fig. 16) con un
promedio anual de 0.63 mg m−3, y por presentar bajos valores de carbono inorgánico

38
particulado (de 0.0001 a 0.0005 mol m-3; fig. 19), también presenta valores bajos de
carbono orgánico particulado (de 62.83 a 234.68 mg m-3; fig. 20), y además baja
atenuación de la luz (de 0.046 a 0.119 m-1; fig. 17). Esta es la tercera región con menor
salinidad con 34.94 UPS y de concentración de fosfatos con 0.509 µmol/l (tabla IV).
La región 13 (Boca del Golfo) es la zona con mayor influencia del Océano
Pacífico y tiene limitada variabilidad en la temperatura (de 21.55°a 29.06 °C). Esta
porción del GC presenta los valores más bajos de clorofila a (de 0.164 a 0.597 mg m−3;
fig. 16), valores bajos de carbono orgánico (de 50.74 a126.03 mg m-3; fig. 20) e
inorgánico particulado (de 0.0001 a 0.0003 mol m-3; fig. 19), por otra parte tiene
además la más baja variabilidad de radiación fotosintéticamente activa (de 30.16 a
54.44 Einstein m2 D; fig. 17), de atenuación de la luz (de 0.037 a 0.073 m-1 ; fig. 18), y
de materia colorada disuelta (de 1.32 a 4.28 m-1; fig. 21). Es una de las tres regiones
con menor concentración de silicatos (5.01 µmol/l) y la de menor salinidad 33.22 UPS
y oxígeno disuelto con 4.63 ml/l (tabla IV).
Finalmente la región 14 (Islas Marías) situada entre las Islas Marías y el macizo
continental, es una región oceánica que presenta la menor variabilidad de temperatura
con valores de 24.4° a 31.6 °C (fig. 15) y un promedio anual de 28.10 °C con lo que se
convierte en la región del Golfo con el promedio más alto de temperatura anual. Es
una de las tres zonas que alcanza los valores más bajos de concentración de carbono
inorgánico al presentar 0.00017 mol m-3 (fig. 19), alcanza este valor durante el mes de
noviembre, y tiene además baja variabilidad de radiación fotosintéticamente activa (de
33.36 a 58.39 Einstein m2 D; fig. 18). Esta es la zona del Golfo de California con la
menor concentración de fosfatos con 0.322 µmol/l, y tiene baja concentración de
oxígeno disuelto con 4.83 ml/l (tabla IV).
Variabilidad mensual de las regiones oceanográficas
A continuación se describe la variabilidad mensual a nivel región de las variables
medidas por sensores remotos, las variables medidas in situ se describen en otro
apartado, porque para esas variables no se pudo obtener el año tipo. Para facilitar la

39
interpretación, se siguen los mismos colores asignados en la regionalización
oceanográfica.
La temperatura superficial del Golfo de California (fig. 15, anexo I) muestra un
patrón estacional, siendo la temporada cálida durante los meses de junio a octubre y
las temperaturas más frías de diciembre a Febrero. En todas las regiones se
encontraron los valores más altos durante el mes de agosto, apreciándose mayor
variabilidad en la región del Alto Golfo con intervalos de hasta 15°C. Las zonas del
centro del Golfo (desde región 7 hasta la 10) presentan un comportamiento estacional
muy similar, mientras que las de la entrada (región 11 a la 14) son las zonas que
presentan la menor variabilidad.
Figura 15 - Variabilidad mensual de temperatura del Golfo de California a nivel región.
La temperatura anual promedio de las regiones del norte del Golfo (R1-R6) es
de 23.38 °C, mientras el promedio de las del centro del Golfo (R7-R10) es de 24.95 °C
y las regiones del sur (R11-R14) presentan un promedio de 26.56 °C.
14
16
18
20
22
24
26
28
30
32
Te
mp
era
tura
(°C
)
R1 R2 R3 R4 R5 R6 R7
R8 R9 R10 R11 R12 R13 R14

40
El año tipo de la clorofila a (fig.16, anexo ll) por su parte, presenta los valores
más altos en las regiones 5, 8, y 11, (es decir la zona costera desde Isla Tiburón hasta
Nayarit), esto se presenta especialmente durante el invierno con valores entre 6 y 7.5
mg m−3. Las cifras descienden a sus valores mínimos durante los meses cálidos (junio
– octubre). Las regiones del norte del Golfo (región 1 y 2) presentan poca variabilidad
con valores de 2.5 a 3.5 mg m−3 la mayor parte del año. Las regiones de la boca del
Golfo (regiones 13 y 14) presentan los valores más bajos de todo el Golfo de California
durante todo el año, normalmente inferiores a 1.5 mg m−3.
Figura 16 - Variabilidad mensual de clorofila a del Golfo de California a nivel región.
Las regiones del norte del Golfo (R1-R6) presentan un promedio anual de 2.63
mg m-3, las del centro del Golfo (R7-R10) presentan 1.58 mg m-3 y finalmente el
promedio en las regiones del sur (R11-R14) es de 1.49 mg m-3.
0
1
2
3
4
5
6
7
8
Clo
rofi
la a
(m
g m
−3)
R1 R2 R3 R4 R5 R6 R7
R8 R9 R10 R11 R12 R13 R14

41
El año tipo de la atenuación de la luz (fig. 17, anexo III), observando un patrón
similar al presentado por la clorofila a. Presenta en las regiones 5, 8 y 11 (desde Isla
Tiburón hasta Nayarit) valores más altos durante la temporada fría (0.5 – 0.1 m-1) y
alcanza los valores mínimos en los meses de junio a agosto (fig. 17). La región Delta
del Río Colorado muestra los valores menos variables (entre 0.25 y 0.29 m-1) a lo largo
del año. Por otra parte las regiones del centro y hacia el sur del Golfo (principalmente
las oceánicas) presentan números bajos y poca variabilidad durante el año. En general
los valores bajos se aprecian al sur del Golfo, y las regiones de mayor variabilidad
(región 3, 5, 8 y 11) se encuentran del lado del macizo continental.
Figura 17 - Variabilidad mensual de atenuación de la luz del Golfo de California a nivel región.
El promedio anual de la atenuación de la luz para las regiones del norte del
Golfo (R1-R6) es de 0.20 m-1, mientras que en el centro del Golfo (R7-R10) es de 0.14
m-1 y para las regiones del sur (R11-R14) es de 0.13 m-1.
0
0.1
0.2
0.3
0.4
0.5
0.6
Ate
nu
ac
ión
de
la
lu
z (
m-1
)
R1 R2 R3 R4 R5 R6 R7
R8 R9 R10 R11 R12 R13 R14

42
El año tipo de la radiación fotosintéticamente activa (fig. 18, anexo IV) muestra
un patrón estacional, con los valores más altos en la primavera/verano (durante mayo-
junio), y estos descienden al mínimo durante los meses de diciembre y enero. Las
regiones del norte del Golfo (regiones 1, 2 y 3) presentan los valores más bajos la
mayor parte del año (octubre a marzo), por el contrario, en las regiones del sur se
tienen los valores más altos durante ese mismo periodo. Las regiones del centro del
Golfo presentan valores intermedios respecto al norte y al sur.
Figura 18 - Variabilidad mensual de radiación fotosintéticamente activa en el Golfo de California a nivel región.
En las regiones del norte del Golfo (R1-R6) el promedio anual es de 43.86
Einstein m2 D, en las regiones del centro (R7-R10) es de 45.49 Einstein m2 D, y en las
regiones del sur del Golfo (R11-R14) se alcanza un promedio anual de 45.28 Einstein
m2 D.
20
25
30
35
40
45
50
55
60
65
R.F
.A.
(Ein
ste
in m
2D
)
R1 R2 R3 R4 R5 R6 R7
R8 R9 R10 R11 R12 R13 R14

43
El año tipo del carbono inorgánico particulado (fig. 19, anexo V) presenta los
valores más altos y más variables en una zona del norte del Golfo, específicamente la
región del Delta del Río Colorado y en el Alto Golfo (regiones 1 y 2) esto probablemente
sea un dato mal calculado debido a que es una zona muy somera y la reflectancia del
fondo incorpore ruido a la medida tomada por el sensor. Las regiones del centro
presentaron poca variabilidad y las del sur concentraron los valores más bajos de todos
(menores a 0.01 mol m-3).
Figura 19 - Variabilidad anual del carbono inorgánico particulado en el Golfo de California a nivel región.
El promedio anual más alto se presenta en las regiones del norte del Golfo (R1-
R6) con 0.0085 mol m-3, el más bajo en las regiones del centro (R7-R10) con 0.0010
mol m-3, y un valor de 0.0014 mol m-3 en las regiones del sur (R11-R14).
0
0.01
0.02
0.03
0.04
0.05
C.I.P
. (m
ol
m-3
)
R1 R2 R3 R4 R5 R6 R7
R8 R9 R10 R11 R12 R13 R14
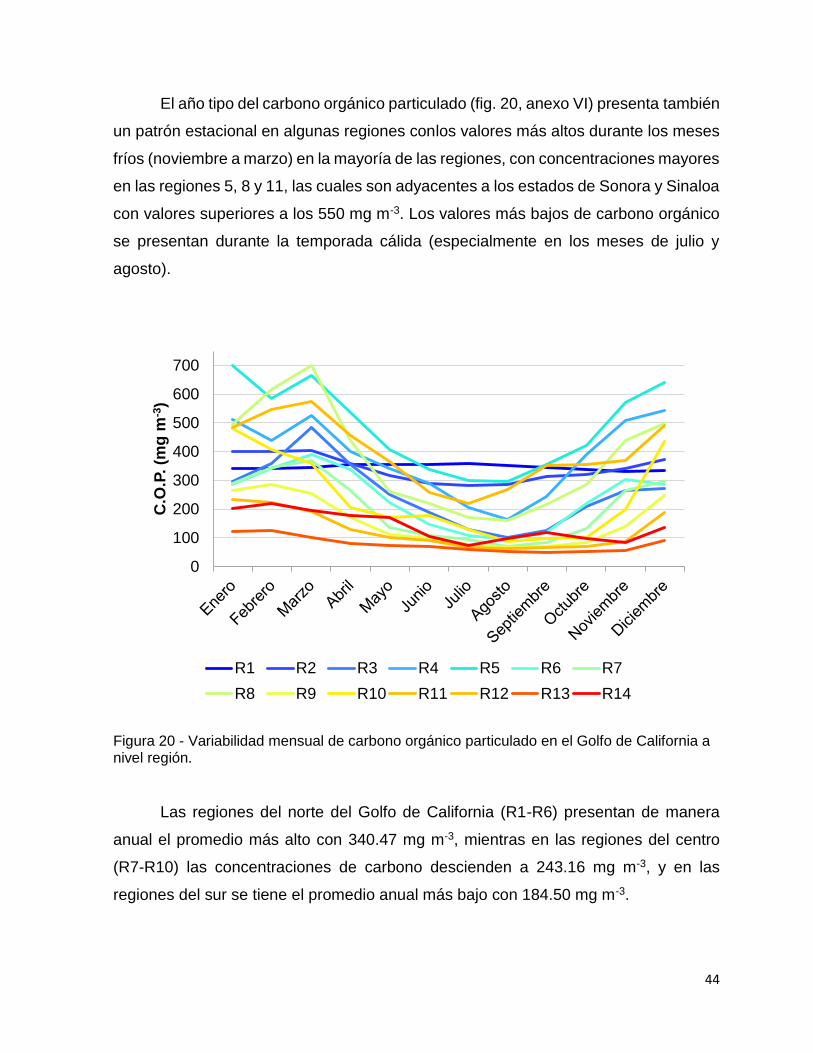
44
El año tipo del carbono orgánico particulado (fig. 20, anexo VI) presenta también
un patrón estacional en algunas regiones conlos valores más altos durante los meses
fríos (noviembre a marzo) en la mayoría de las regiones, con concentraciones mayores
en las regiones 5, 8 y 11, las cuales son adyacentes a los estados de Sonora y Sinaloa
con valores superiores a los 550 mg m-3. Los valores más bajos de carbono orgánico
se presentan durante la temporada cálida (especialmente en los meses de julio y
agosto).
Figura 20 - Variabilidad mensual de carbono orgánico particulado en el Golfo de California a nivel región.
Las regiones del norte del Golfo de California (R1-R6) presentan de manera
anual el promedio más alto con 340.47 mg m-3, mientras en las regiones del centro
(R7-R10) las concentraciones de carbono descienden a 243.16 mg m-3, y en las
regiones del sur se tiene el promedio anual más bajo con 184.50 mg m-3.
0
100
200
300
400
500
600
700
C.O
.P. (m
g m
-3)
R1 R2 R3 R4 R5 R6 R7
R8 R9 R10 R11 R12 R13 R14
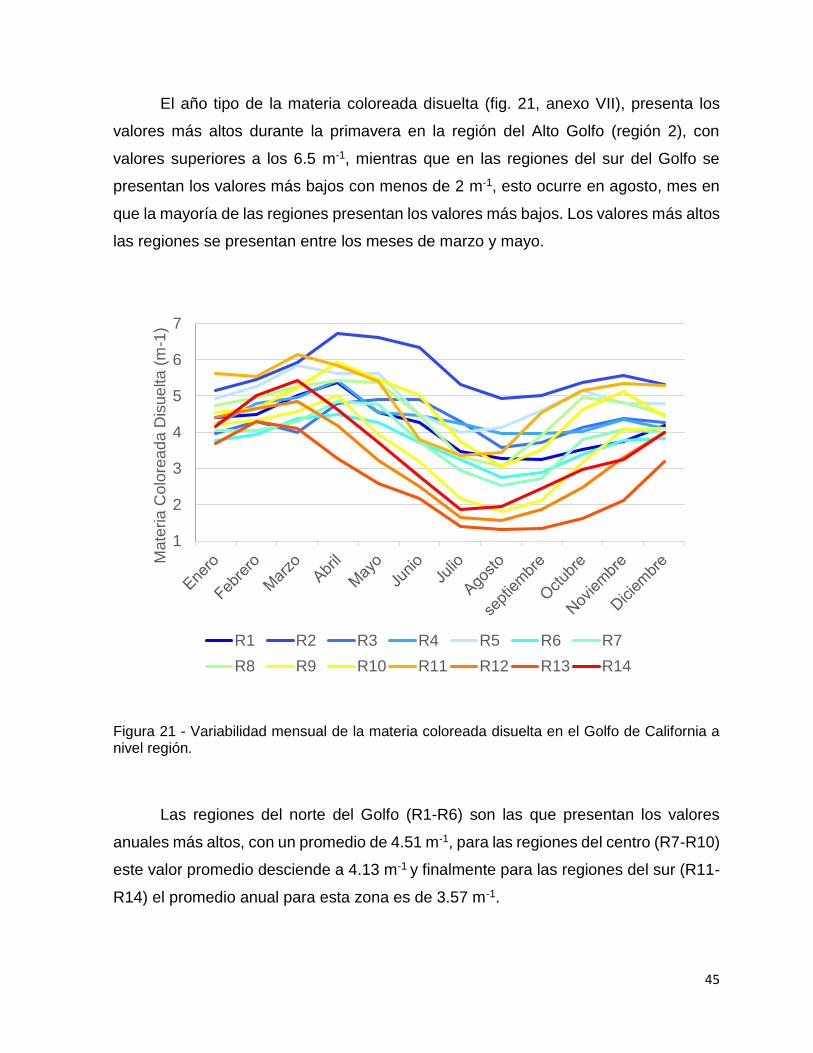
45
El año tipo de la materia coloreada disuelta (fig. 21, anexo VII), presenta los
valores más altos durante la primavera en la región del Alto Golfo (región 2), con
valores superiores a los 6.5 m-1, mientras que en las regiones del sur del Golfo se
presentan los valores más bajos con menos de 2 m-1, esto ocurre en agosto, mes en
que la mayoría de las regiones presentan los valores más bajos. Los valores más altos
las regiones se presentan entre los meses de marzo y mayo.
Figura 21 - Variabilidad mensual de la materia coloreada disuelta en el Golfo de California a nivel región.
Las regiones del norte del Golfo (R1-R6) son las que presentan los valores
anuales más altos, con un promedio de 4.51 m-1, para las regiones del centro (R7-R10)
este valor promedio desciende a 4.13 m-1 y finalmente para las regiones del sur (R11-
R14) el promedio anual para esta zona es de 3.57 m-1.
1
2
3
4
5
6
7
Ma
teria
Colo
rea
da
Dis
ue
lta
(m
-1)
R1 R2 R3 R4 R5 R6 R7
R8 R9 R10 R11 R12 R13 R14
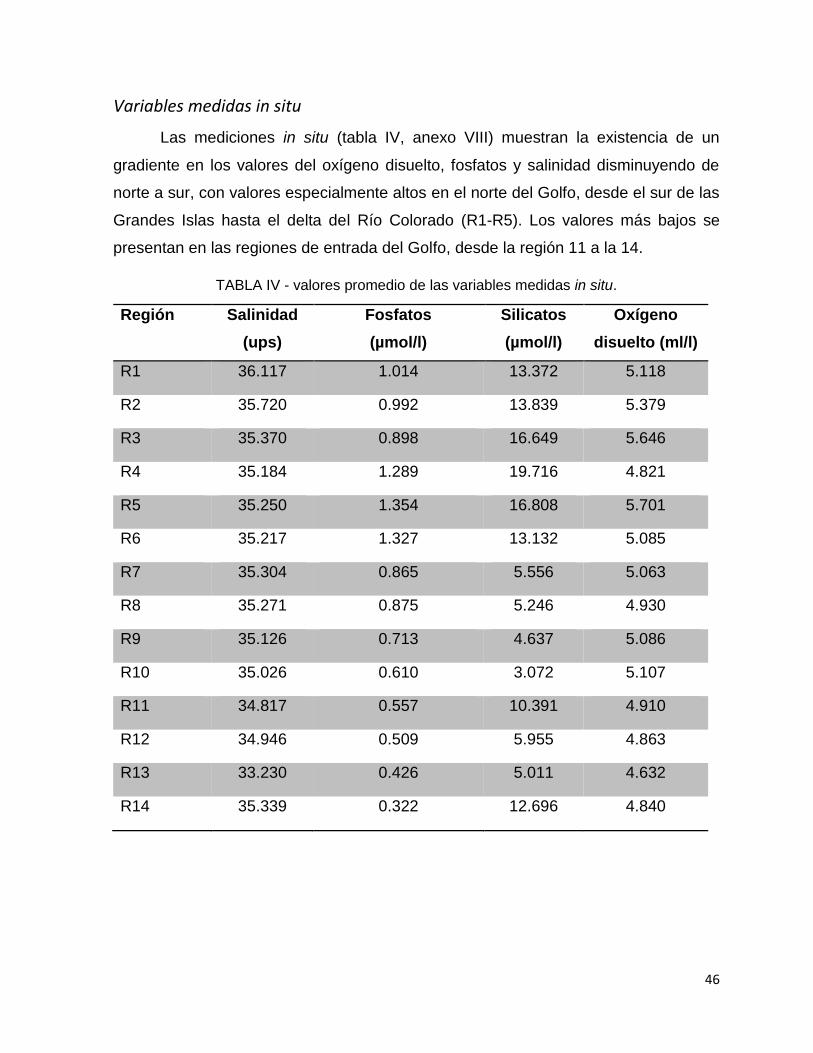
46
Variables medidas in situ
Las mediciones in situ (tabla IV, anexo VIII) muestran la existencia de un
gradiente en los valores del oxígeno disuelto, fosfatos y salinidad disminuyendo de
norte a sur, con valores especialmente altos en el norte del Golfo, desde el sur de las
Grandes Islas hasta el delta del Río Colorado (R1-R5). Los valores más bajos se
presentan en las regiones de entrada del Golfo, desde la región 11 a la 14.
TABLA IV - valores promedio de las variables medidas in situ.
Región Salinidad
(ups)
Fosfatos
(µmol/l)
Silicatos
(µmol/l)
Oxígeno
disuelto (ml/l)
R1 36.117 1.014 13.372 5.118
R2 35.720 0.992 13.839 5.379
R3 35.370 0.898 16.649 5.646
R4 35.184 1.289 19.716 4.821
R5 35.250 1.354 16.808 5.701
R6 35.217 1.327 13.132 5.085
R7 35.304 0.865 5.556 5.063
R8 35.271 0.875 5.246 4.930
R9 35.126 0.713 4.637 5.086
R10 35.026 0.610 3.072 5.107
R11 34.817 0.557 10.391 4.910
R12 34.946 0.509 5.955 4.863
R13 33.230 0.426 5.011 4.632
R14 35.339 0.322 12.696 4.840

47
Regionalización biológica
Regionalización a nivel phylum
El análisis de k – promedios para cada phylum, y la exploración visual de las
regionalizaciones obtenidas, indicó un total de siete grupos o regiones para los
moluscos y equinodermos, ocho para artrópodos y anélidos y seis para los cnidarios
(fig. 22).
Figura 22 - Índice de k – promedios para los cinco fila seleccionados de macroinvetebrados del Golfo de California.
Para ejemplificar la distribución de los macroinvertebrados en el Golfo de
California, se presenta un mapa de riqueza específica (fig. 23), el cual es la suma de
todos los rasters de distribución geográfica para las 3,496 especies y expresa cuantas
de ellas habitan en cada una de las 3,502 celdas de 9 km2. Es claro el gradiente de
riqueza de sur a norte, y de este a oeste, siendo más el número de especies en zonas
tropicales y en la plataforma continental desde Guaymas hasta el estado de Nayarit.

48
También es claro que la mayor riqueza se presenta en la zona costera, disminuyendo
hacia las zonas más profundas, y especialmente en las cuencas y en la entrada del
Golfo, la cual presenta la mayor profundidad del área y conecta con el Océano Pacífico.
Figura 23 - Riqueza específica de macroinvertebrados en el Golfo de California.

49
La regionalización del phylum Mollusca presenta siete divisiones, tres en zona
costera y cuatro localizadas en zonas oceánicas (fig. 24A). La composición de las
especies señala una diferenciación por grupos (fig. 24B) estando dentro de una mismo
rama las regiones costeras (R1, R3 y R5), y las oceánicas (R2, R4, R6 y R7) en otra
rama del dendrograma. La mayor riqueza de especies (fig. 24C) se encuentra en R5
que abarca desde Guaymas, hasta Bahía de Banderas. En general la zona costera
presenta la mayor riqueza con regiones con más de 600 especies cada una, mientras
las zonas oceánicas presentan menos de 250 especies. Incluso hay zonas donde no
se han llevado estudios sobre este phylum y hay aparente ausencia de especies en
algunas de las cuencas y zonas más profundad del Golfo de California.
Figura 24 - A) Regionalización del phylum Mollusca. B) Dendrograma de similitud biológica entre regiones. C) Riqueza de específica de especies a nivel región.

50
La regionalización de los equinodermos (fig. 25A) incluye siete secciones que
se diferencian en riqueza y composición. La mayor similitud está entre las regiones 2,
4 y 6 (fig. 25B) las cuales presentan la mayor riqueza con más de 100 especies (fig.
25C). Esos sitios abarcan la zona costera del Golfo, excepto la región del Alto Golfo,
la cual se agrupa con las regiones 3, 5 y 7 que corresponden a zonas oceánicas con
una riqueza de especies inferior a las 100 especies.
Figura 25 - (A) Regionalización del phylum Echinodermata. (B) Dendrograma de similitud biológica entre regiones. (C) Riqueza de específica de especies a nivel región.
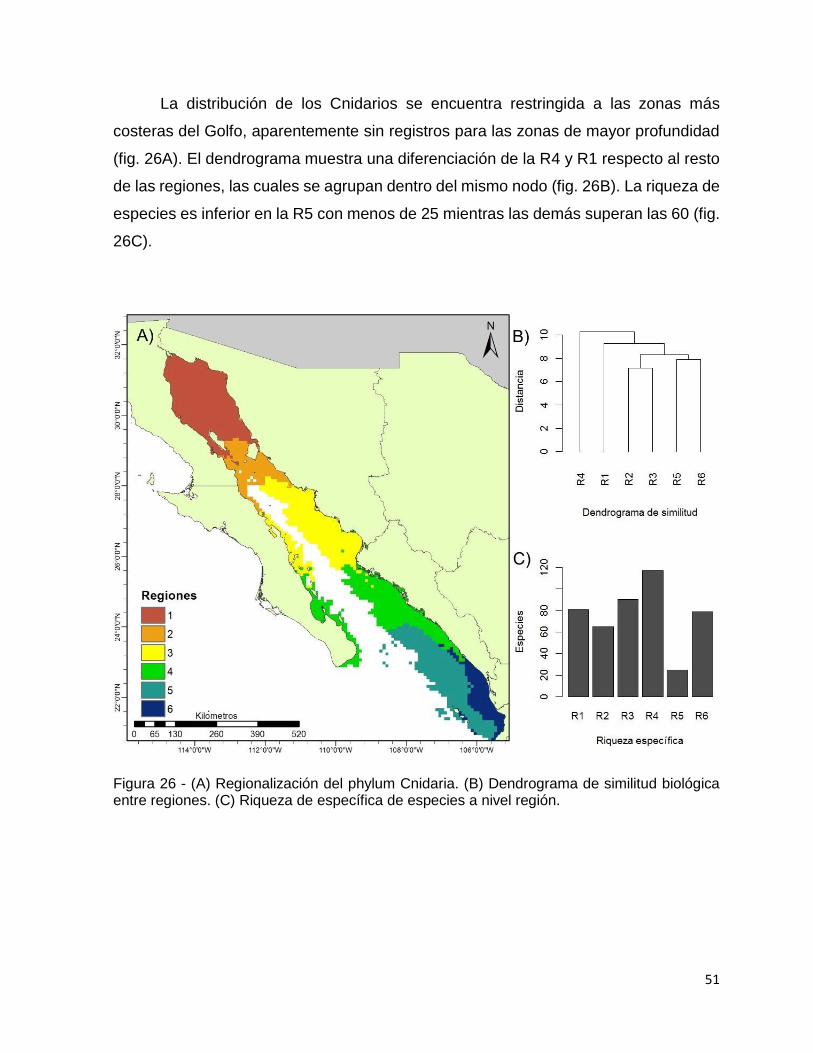
51
La distribución de los Cnidarios se encuentra restringida a las zonas más
costeras del Golfo, aparentemente sin registros para las zonas de mayor profundidad
(fig. 26A). El dendrograma muestra una diferenciación de la R4 y R1 respecto al resto
de las regiones, las cuales se agrupan dentro del mismo nodo (fig. 26B). La riqueza de
especies es inferior en la R5 con menos de 25 mientras las demás superan las 60 (fig.
26C).
Figura 26 - (A) Regionalización del phylum Cnidaria. (B) Dendrograma de similitud biológica entre regiones. (C) Riqueza de específica de especies a nivel región.
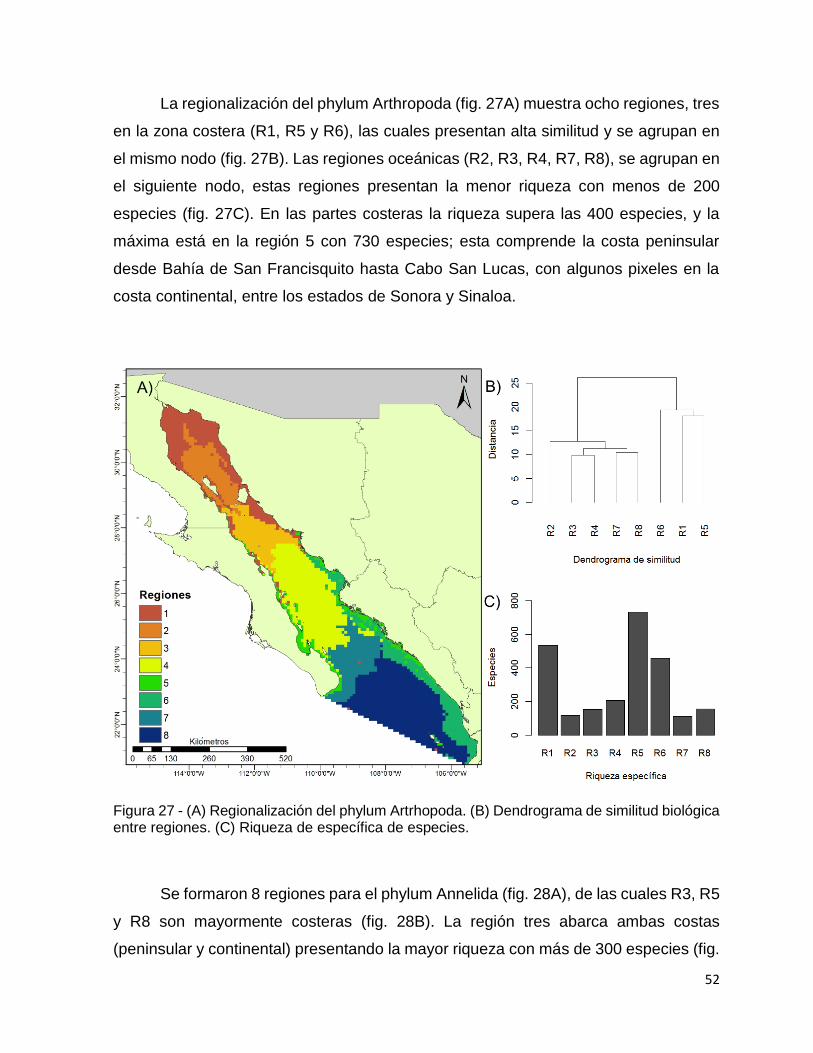
52
La regionalización del phylum Arthropoda (fig. 27A) muestra ocho regiones, tres
en la zona costera (R1, R5 y R6), las cuales presentan alta similitud y se agrupan en
el mismo nodo (fig. 27B). Las regiones oceánicas (R2, R3, R4, R7, R8), se agrupan en
el siguiente nodo, estas regiones presentan la menor riqueza con menos de 200
especies (fig. 27C). En las partes costeras la riqueza supera las 400 especies, y la
máxima está en la región 5 con 730 especies; esta comprende la costa peninsular
desde Bahía de San Francisquito hasta Cabo San Lucas, con algunos pixeles en la
costa continental, entre los estados de Sonora y Sinaloa.
Figura 27 - (A) Regionalización del phylum Artrhopoda. (B) Dendrograma de similitud biológica entre regiones. (C) Riqueza de específica de especies.
Se formaron 8 regiones para el phylum Annelida (fig. 28A), de las cuales R3, R5
y R8 son mayormente costeras (fig. 28B). La región tres abarca ambas costas
(peninsular y continental) presentando la mayor riqueza con más de 300 especies (fig.

53
28C). Finalmente, las regiones que abarcan aguas oceánicas (R1, R2, R4, R6, R7)
presentan menor riqueza respecto a las costeras, siempre con menos de 200 especies
en cada región.
Figura 28 - (A) Regionalización del phylum Annelida. (B) Dendrograma de similitud biológica entre regiones. (C) Riqueza de específica de especies a nivel región.
Regionalización general de macroinvertebrados
La propuesta de regionalización general de los macroinvertebrados del Golfo de
California (fig. 29A) obtenida mediante el coeficiente de Gower (1971) a partir de la
ponderación de las cinco regionalizaciones biológicas presentadas anteriormente,
presenta 9 grupos; 6 en las zonas de gran profundidad y 3 en la zona costera. Las
regiones costeras fueron la R1, R5 y R7, la primera rodea el norte del Golfo, desde
San Francisquito hasta Guaymas, la R5 se encuentra en dos segmentos, uno incluye
la zona costera continental, desde Guaymas hasta Topolobampo, y el otro la costa
península que llega hasta la Isla Cerralvo. La R7 incluye el resto de la costa peninsular

54
desde la isla Cerralvo hasta Cabo San Lucas y en el lado continental desde
Topolobampo hasta Bahía de Banderas. Estas tres regiones costeras presentaron alta
similitud biológica (fig. 29B), y riqueza de especies superior a las 2,000 a nivel región
(fig. 29C).
Figura 29 - (A) Regionalización biológica del Golfo de California. (B) Dendrograma de similitud
biológica entre regiones. (C) Riqueza de específica de especies a nivel región.
Para las regiones oceánicas, hubo dos grupos importantes, las R2 y R3
ubicadas en el norte y centro del Golfo, y el grupo de las regiones del sur (R4, R6, R8,
R9). En general, las regiones oceánicas presentaron la menor riqueza de especies (no
más de 700 especies en total) siendo la R6 (cuencas oceánicas) la de menor
diversidad al tener menos de 250 especies.

55
Biomas del Golfo de California
Finalmente, al unir las características oceanográficas y biológicas mediante la
ponderación de las distintas regionalizaciones (con ponderación del 50%
oceanográficas y 50% para las regiones biológicas) presentadas previamente para
formar biomas marinos, se encontraron un total de 10 para el Golfo de California (fig.
30A), seis biomas oceánicos y cuatro costeros. Todos los primeros presentaron
segmentos en la península y en el macizo continental.
De norte a sur, los biomas costeros son el B1, el cual abarca el área del Alto
Golfo desde la Bahía de San Luis Gonzaga hasta el norte del Canal del Infiernillo. El
bioma B2 se presenta desde el Canal de Ballenas hasta San Francisquito en el lado
peninsular, y desde el Canal del Infiernillo hasta Guaymas en su parte continental. El
bioma B6 ocupa desde San Francisquito hasta Punta Arenas en la península, y desde
Guaymas hasta los 23°N en el lado continental. Finalmente el bioma B8, que va desde
Punta Arenas hasta Cabo San Lucas y desde los 23°N hasta Bahía de Banderas en el
lado continental. Estos cuatro biomas costeros (B1, B3, B6 y B8) en el análisis de
similitud biológica (fig. 30C) están dentro del mismo grupo, con B6 y B8 en un subgrupo
y B3 con B1 en el otro, los cuales geográficamente están contiguos. Mientras que por
similitud oceanográfica solo agruparon en el mismo nodo el bioma B6 con B3. Los
biomas B6 y B8 presentaron la mayor riqueza, siendo superior a 2,000 especies cada
uno, seguidas por el bioma B1 y B3, las cuales tienen una riqueza entre 1,000 y 1,700
especies cada uno (fig. 30D). Por otra parte dentro de los biomas oceánicos (B2, B4,
B5, B7, B9 y B10), el bioma B2 está ubicado en el norte del Golfo y se agrupó por
similitud oceanográfica con los biomas costeros B1 y el B8 (fig. 30B), mientras que
biológicamente lo hizo con los biomas oceánicos B4 y B5 (fig. 30C). Por otra parte
oceanográficamente los biomas B4 y B5 presentan mayor similitud con el bioma B7,
estos tres biomas se encuentran en el centro del Golfo de California. Finalmente los
biomas B9 y B10 ubicados al sur del Golfo, oceanográficamente se agrupan en el
mismo nodo, mientras que por similitud biológica se agrupan también con el bioma B7
(fig. 30C). Los biomas oceánicos presentan una riqueza de menos de 500 especies
cada una, es decir un 20% del número de especies que habitan los biomas costeros
(fig. 30D).
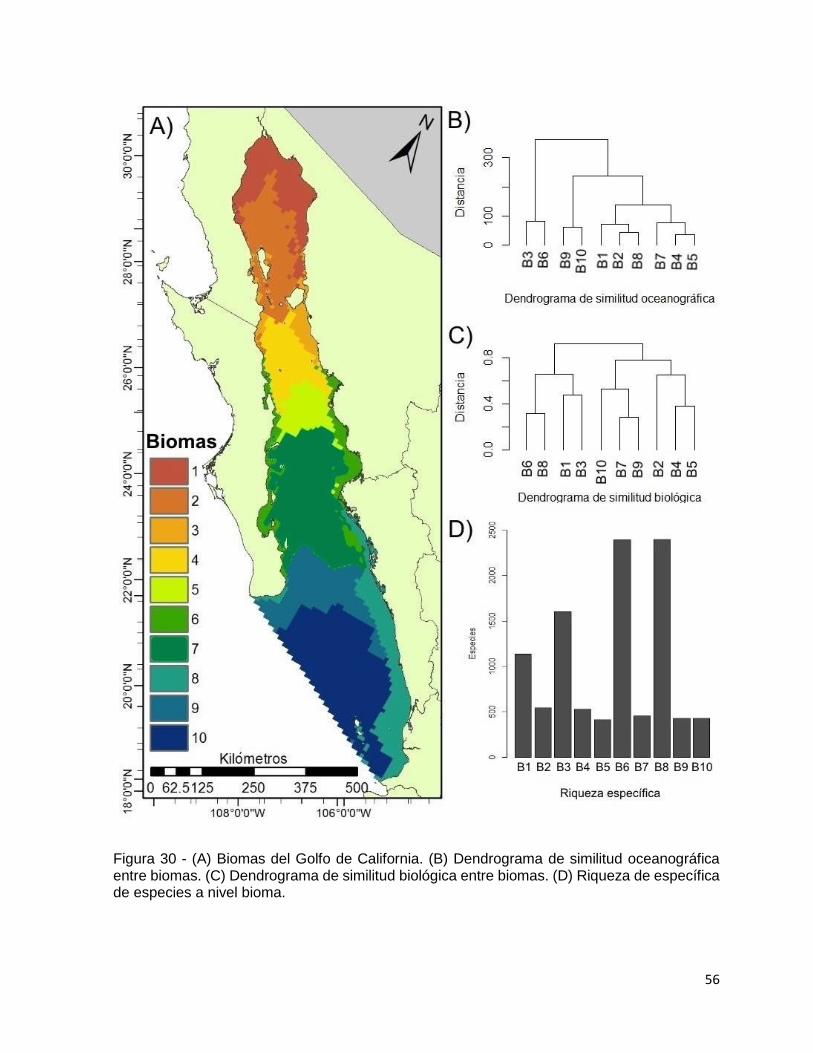
56
Figura 30 - (A) Biomas del Golfo de California. (B) Dendrograma de similitud oceanográfica entre biomas. (C) Dendrograma de similitud biológica entre biomas. (D) Riqueza de específica de especies a nivel bioma.

57
DISCUSIÓN
Regionalización oceanográfica
El Golfo de California presenta características bastante diferenciables en la
distribución de la fauna y en las características oceanográficas, y es por ello que
diversos autores desde hace más de 70 años lo han subdivido en regiones para
simplificar el análisis de la complejidad biológica y ambiental del mismo. En el aspecto
oceanográfico se han descrito de 2 a 14 regiones, a partir de mediciones in situ (Allen,
1937; Cupp y Allen, 1938; Sverdrup, 1941; Gilbert y Allen 1943; Round 1967; Roden y
Emilson, 1979; Lavin y Marinone, 2003), datos de sensores remotos (Santamaría del
Ángel et al., 1994; Gaxiola-Castro et al., 1995; Arias-Arechiga, 1998; Lluch-Cota y
Arias-Arechiga, 2000; Kahru et al., 2004, Hidalgo-Gonzalez y Alvarez-Borrego, 2004;
Ulloa et al., 2006) e incluso simulaciones de corrientes superficiales y conectividad
hidrográfica (Marinone, 2012; Santiago-García et al., 2014). Las variables utilizadas
para realizar esas zonaciones fueron la temperatura, el fitoplancton, pigmentos
fotosintéticos, atenuación de la luz y modelos de corrientes marinas. En ese sentido,
la propuesta de zonación oceanográfica presentada en este trabajo (fig. 12), contiene
una serie de variables (materia coloreada disuelta, carbono orgánico e inorgánico
particulado y radiación fotosintéticamente activa) que no fueron consideradas
anteriormente. Además, incorpora por primera vez variables químicas del océano para
su regionalización en el Golfo de California, estas variables son los silicatos, fosfatos,
oxígeno disuelto y finalmente la salinidad. El uso de estas características físico-
químicas da un mayor sustento a la regionalización, ya que no se enfoca en un solo
tipo de variables y aunque no se pudo determinar el año tipo de las variables químicas,
debido que no se lleva un monitoreo adecuado espacial y temporal para todo el Golfo,
al acumular los registros históricos, promediarlos e interpolarlos, los resultados
muestran la amplia variabilidad en la concentración de nutrientes en el Golfo, y
demuestran un gradiente de sur a norte de estos valores (tabla IV, anexo VIII), para
reforzar así la existencia de una diferenciación latitudinal en el Golfo de California.
Algunas de las regiones oceanográficas encontradas en este trabajo que ya han
sido señaladas por otros autores son, la región Delta del Río Colorado (R1) la cual

58
aparece en el trabajo de Santamaría del Ángel y colaboradores en 1994, Lavín y
Marinone en 2003, y Kahru y colaboradores en 2004, los autores encontraron esta
región al trabajar con valores de temperatura y clorofila a, pese a que la regionalización
de esta tesis se realizó con un conjunto de 11 variables oceanográficas, características
como la variabilidad de la temperatura, de carbón inorgánico particulado y alta
salinidad hacen de esta zona una región con características únicas dentro del Golfo
de California, esta región también aparece en el trabajo de Marinone (2012) y
Santiago-García y colaboradores (2014), al presentar pequeños giros y corrientes
residuales. Otra región aquí propuesta, y que ha sido delimitada como región por otros
autores es la zona de las Grandes Islas, la cual aparece en los trabajos de Lluch-Cota
y Arias Arechiga (2000), Lavin y Marinone (2003) y Kahru y colaboradores (2004).
Lavin y Marinone (2003) establecen esta zona por sus características físicas, la cual
presenta una fuerte mezcla por mareas, por lo que se puede deducir que esta región
es de las más distintivas del GC ya que aparece al analizar no solo la oceanografía
física, sino también por las variables medidas por el color del océano y además por la
alta concentración de nutrientes. Respecto a la oceanografía física, las regiones
oceánicas definidas en la tesis son consistentes con los giros oceanográficos del Golfo
de California reportados por Lavín y colaboradores, 1997, Pegau y colaboradores en
2002 y Zamudio y colaboradores en 2008. Los primeros autores describen un gran
remolino estacional reversible y que abarca la región Golfo Norte. Al centro y sur del
Golfo, describen la presencia de varios remolinos que cambian con la circulación
estacional del Golfo, ya que los patrones de corrientes son generalmente ciclónicos en
verano y anticiclónicos en inverno (Sánchez-Velasco et al., 2009). La regiones
oceánicas delimitadas en esta tesis pertenecen a zonas de presencia de continuos
remolinos a mesoescala, y probablemente estas regiones sean el reflejo del efecto del
afloramiento de nutrientes, la mezcla de las distintas masas de agua (de origen y
composición distinta), especialmente en la entrada y centro del Golfo (Trasviña et al.,
1999) que ayudaron a delimitar las regiones en mar abierto de este trabajo.
Si bien, aunque esta regionalización diferencia las zonas costeras de las
oceánicas, pero concuerda en términos generales con el patrón de tres grandes
regiones: norte, centro y sur, es decir, regiones de templado a tropical (López-Martínez

59
et al., 2012) con las que es segmentado el Golfo de California tradicionalmente. Es
entendible que regularmente se use un número limitado de regiones con cortes a lo
ancho del Golfo, debido a que son más fáciles de entender y que en los primeros
estudios no se contaba con información detallada y a gran escala como en la
actualidad, prueba de ello es que con el primer trabajo con imágenes de satélite para
el GC (Santamaría del Ángel et al., 1994), el número de regiones se incrementó a 14
cuando con anterioridad se manejaban de 2 a 4 zonas. Como se puede apreciar en
estos resultados (fig. 15-21, anexo I-X) las zonas costeras y oceánicas poseen
características distintas, no solo en las condiciones ambientales, sino también en la
distribución de algunas especies (fig. 24-28) por lo que una regionalización adecuada,
debe diferenciar las zonas costeras y las oceánicas.
Los resultados indicaron que la formación de las regiones se debe a un intenso
cambio en la mayoría de las variables ambientales que se da estacionalmente, donde,
cada región responde con poca o alta variabilidad, dependiendo de su latitud y si se
trata de una zona oceánica o costera, por ejemplo, en las condiciones de temperatura
y clorofila a, usadas en las regionalizaciones marinas donde la temperatura se toma
como el componente físico, y la clorofila a como el componente biológico (González-
Silvera et al., 2004) los intervalos son más altos en el alto Golfo, para las temperaturas,
mientras que la concentración de clorofila también se presenta un patrón estacional,
especialmente en la las regiones costeras de los estados de Sonora y Sinaloa,
mientras al norte del Golfo se mantienen valores altos la mayor parte del año y las
concentraciones bajas de clorofila se localizan al sur. Se encontró además una alta
correlación entre variables (anexo XI), por ejemplo la Clorofila a con la atenuación, en
invierno cuando se tiene los valores más altos de esta variable, se correlacionan en un
99% con la atenuación de la luz, y superior al 90% con el carbono orgánico disuelto,
debido a la contribución que hacen las proliferaciones de fitoplancton en el carbono, el
cual se ha estimado que es entre el 70 y 80% (Hernández-Alfonso, et al., 2007)
mientras que en temporada cálida y con menor cantidad de clorofila en el Golfo, el
fitoplancton se correlacionó en un 90% con el carbono inorgánico particulado, debido
a la incorporación de carbono inorgánico por los cocolitofóridos (Fernández et al.,
1993). Por otra parte, la temperatura superficial en temporada fría se correlacionó con

60
la radiación fotosintéticamente activa, probablemente debido a que en invierno el
florecimiento del fitoplancton se incrementa en el Golfo, relacionado con las bajas
temperaturas. Este incremento en los pigmentos fotosintéticos hacen que la actividad
fotosintética de estos organismos sea mejor detectada por los sensores. Además de
la radiación fotosintética, las temperaturas bajas se correlacionaron negativamente
con las concentraciones de fosfato (-89%) y es que estos últimos son especialmente
altos al norte del Golfo, mientras que las temperaturas son bajas en esa zona y altas
al sur, donde se tienen características templado-tropicales y las concentraciones de
fosfatos disminuyen, sin embargo, este efecto de disminución del fosfato
aparentemente no es causalidad de las altas temperaturas, contrario a lo que ocurre
con la salinidad con la que tiene presenta una correlación del 72% y es que las altas
temperaturas en verano hacen del Golfo una cuenca de evaporación (Roden, 1958),
haciendo el agua hasta 2.8 UPS (Tabla IV) más salina en las regiones del norte que
en la boca, la cual está en contacto con el pacífico. Para facilitar la interpretación de
las correlaciones y la variabilidad de las condiciones ambientales del Golfo, se
presenta un análisis de componentes principales (fig. 31) en la que se muestran los
dos primeros componentes, los cuales explican en conjunto la variabilidad del 66% de
los datos (49.8 y 16.6 % respectivamente), se presentan los eigenvectores de las
distintas variables oceanográficas en sus condiciones máximas y mínimas, así como
la localización en el eigenespacio generado para las 14 regiones oceanográficas. En
la gráfica se muestra la amplia variabilidad estacional de las condiciones, así como las
correlaciones descritas anteriormente, además de la distancia entre regiones, por lo
que se fundamenta la necesaria utilización de distintas variables físico-químicas en
mares de alta variabilidad como lo es el Golfo de California (Soto-Mardones et al.,
1999; Álvarez-Borrego, 2010; Escalante et al., 2013).

61
Figura 31 - Componentes principales de las variables oceanográficas para la regionalización del Golfo de California(regiones indicadas en la Tabla I, en rojo las regiones costeras, en azul las regiones oceánicas) Donde: sst_M=Temperatura máxima, sst_m=Temperatura mínima, Chla_M=Clorofila a máxima, Chla_m=Clorofila a mínima, pic_M=Carbono inórganico máximo, pic_m=Carbono inorgánico mínimo, poc_M=Carbono orgánico máximo, poc_m=carbono orgánico mínimo, par_M=Radiación fotosintética máxima, par_m= Radiación fotosintética mínima, k490_M=atenuación de luz máxima, k490_m=atenuación de luz mínima, cdom_M=Materia coloreada máxima, cdom_m=Materia coloreada mínima, Sal=Salinidad, Po4=Fosfatos, SiO2=Silicatos, O2=Oxígeno disuelto.
La separación de las regiones oceánicas y las costeras se presenta en zonas
poco variables estacionalmente y con los valores más altos de temperatura durante el
invierno, es decir las regiones localizadas en las latitudes más tropicales, y es que las
temperaturas mínimas (invierno) tuvo una correlación negativa con el componente

62
principal uno, también la radiación fotosintética respondió en el mismo sentido ya que
fue baja debido a una menor concentración de fitoplancton en aguas oceánicas del sur
del GC. Por otra parte, en el componente uno se asociaron las variables de Clorofila
a, carbono orgánico particulado, atenuación de la luz y materia coloreada, todas de
manera positiva, y en ella se asociaron las regiones que presentaron altos valores y
amplio intervalo estacional para estas variables, es decir, las regiones costeras a
excepción de la región R3 (Golfo Norte) la cual se consideró oceánica en este trabajo,
sin embargo presenta valores altos de clorofila y por eso se asocia con las regiones
costeras en este análisis de componentes principales.
Para el segundo componente, se aprecia una correlación positiva con el
carbono inorgánico particulado y la clorofila a, en el que se asocia la región (R1) Delta
del Río Colorado, ya que presenta altos valores durante todo el año (fig. 19) mientras
que de manera negativa está relacionado con las variables de carbono orgánico
particulado y de radiación fotosintéticamente activa, también es por ello que la región
R3 (Golfo Norte) se asocia en ese espacio, debido a que presenta altos valores de
radiación fotosintética la mayor parte del año (anexo IV).
En resumen, las regiones del GC se caracterizan por tener una estacionalidad
muy marcada en la zona costera y con intervalos de variabilidad bajos en las zonas
oceánicas, además del gradiente latitudinal existente entre las Grandes Islas y el norte
del Golfo con alta concentración de nutrientes, lo que genera gran cantidad de
alimento, para sostener un ecosistema en el que se realiza la mayor parte de las
pesquerías (Ulloa et al., 2006), mientras que en el sur la concentración de nutrientes y
la productividad primaria es más baja, sumado a la baja variabilidad de las condiciones
oceanográficas.

63
Regionalización biológica
La regionalización biológica se realizó por separado (una por cada phylum) para
evitar que al acumular a todas las especies indistintamente, la tendencia representara
al phylum con mayor número de especies, en este caso los moluscos, sin conocer los
patrones espaciales de los demás fila, cuando lo que se busca es generar los patrones
de distribución de las especies más representativas, y es que como se aprecia en
regionalizaciones previas de peces (Walker, 1960; Robertson y Cramer, 2009), y
crustáceos (Brinton y Townsend, 1980; Siegel-Causey 1982; Boschi, 2000), cada
grupo de especies se distribuye de manera distinta.
La regionalización biológica de este trabajo (fig. 29A), utiliza por primera vez la
distribución de miles de especies para el Golfo de California, y además con una
resolución espacial con el más alto nivel de detalle hasta ahora (cinco minutos
geográficos), por ejemplo, otro trabajo que utiliza una gran cantidad de especies es el
de Robertson y Cramer (2009), el cual analiza la distribución de 1,139 especies de
peces costeros del Pacífico Oriental Tropical, de los cuales, solo 698 especies
pertenecen a la provincia zoo geográfica del cortés, la cual incluye al Golfo de
California. Ese trabajo se realizó con una resolución espacial de 1° geográfico, por lo
que los resultados de esta tesis representan una mejora en el estudio de la
biogeografía del Golfo de California, al dar patrones espaciales para más de 3,400
especies de cinco distintos fila, a una mejor resolución espacial y que confirman los
cambios en la diversidad tanto latitudinal como batimétrico (fig. 24-28), y establece
además nuevos límites geográficos, especialmente en la zona oceánica. Por ejemplo,
los principales puntos de convergencia entre regiones de los distintos fila se presentan
en San Luis Gonzaga para moluscos y equinodermos, Guaymas para los moluscos,
equinodermos, cnidarios y artrópodos, Bahía de los Ángeles para moluscos, cnidarios
y anélidos, la Bahía de San Francisquito para equinodermos, anélidos, y artrópodos.
También la Bahía de La Paz es una zona de convergencia de regiones para los
moluscos, equinodermos y anélidos (al igual que para los peces de acuerdo con
Walker, 1960 y Thompson, 2000). La Bahía San Esteban lo es para los equinodermos,
cnidarios y anélidos. Por otra parte resulta interesante que las regiones biológicas de
mar profundo coincidan latitudinalmente con la presencia de cuencas, por ejemplo para

64
las cuencas Carmen y Farallón, los equinodermos, artrópodos y anélidos presentaron
una región con la misma extensión geográfica, lo mismo ocurre con la cuenca de
Guaymas, donde una región de los equinodermos y de artrópodos coinciden de nuevo
con esa cuenca, además en la cuenca Pescadero ocurre algo similar con los
artrópodos y anélidos. Probablemente las fallas y cuencas sirven de barreras en la
distribución de algunos grupos de especies (Hendrickx, 2008) y es por eso que los
límites geográficos de las regiones biológicas se relacionan con la presencia de ellas
en las zonas más profundas del Golfo.
Al unificarse las regionalizaciones de cada phylum en una regionalización de
macroinvertebrados (fig. 29A), se puede observar que solo tres regiones son las
abarcan la zona costera, la primera coincide con las regiones oceanográficas de alta
variabilidad (R1-R5), las cuales se encuentran al norte del Golfo, presentan
características templadas y de mayor endemismo (Hendrickx y Brusca, 2007;
Robertson y Cramer, 2009; Petatán-Ramírez et al., 2014) debido al aislamiento y al
cambio en las condiciones ambientales. La segunda región costera corresponde a la
costa rocosa (la cual incluye algunas playas arenosas) de la península de Baja
California, desde Bahía de San Francisquito hasta Cabo San Lucas, y de Guaymas a
Bahía San Esteban, donde se encuentran zonas principalmente arenosas. Esta región
presenta una mayor similitud biológica con la región que corresponde a la zona costera
al norte del Golfo, debido a la proximidad geográfica y a que comparten un tipo de
sedimento similar, terrígeno mezclado con arena y limo (INAPESCA, 1994), lo que
limita al tipo de especies que pueden habitar ambas zonas. Finalmente, el margen
costero desde Sinaloa hasta Nayarit, el cual presenta la mayor riqueza de especies,
probablemente debido a la gran diversidad de hábitats, pues cuenta con playas
arenosas, ecosistemas de manglar, lagunas costeras, estuarios, entre otros (Castro-
Aguirre et al., 1996, Ulloa et al., 2006). Tiene además una menor variabilidad en las
condiciones oceanográficas, con características tropicales, y junto con el Alto Golfo,
presenta una de las mayores plataformas continentales del Golfo de California
(SEMARNAT, 2006), lo que representa más espacio para la distribución de
organismos.

65
Figura 32 - Sedimentología del Golfo de California. (modificado de INAPESCA, 1994).
Es de destacar que las tres regiones biológicas costeras previamente descritas,
aparentemente coinciden geográficamente en áreas ocupadas por determinados
sedimentos, por ejemplo, los límites geográficos de la región 1, ubicada al norte del
Golfo, es coincidente los límites del limo del Alto Golfo (fig. 32), luego, la región 5, que
abarca la costa peninsular desde San Francisquito hasta Cabo San Lucas y desde
Guaymas hasta Bahía San Esteban, tiene el mismo tipo de sedimento en ambas
costas (limo) y finalmente, la región 7, que abarca desde Bahía San Esteban hasta
Nayarit, presenta un único tipo de sedimento (arena). Aunque la información de
sedimentos no se utilizó para la zonación biológica ya que estos datos no se
encontraron georreferenciados de manera precisa, esta aparente coincidencia de las
regiones biológicas con los sedimentos y el tipo de costa, es consistente con lo que

66
han mencionado otros autores, que además de los factores ambientales del océano,
la materia orgánica en sedimentos, el tamaño de grano, la latitud y los pigmentos del
sedimento son indicadores importantes en la distribución de la macrofauna (Hendrickx,
2008; Méndez, 2012).
Por otra parte, en las regiones de aguas profundas podemos observar una
mayor variación latitudinal en las regiones, en tres grandes grupos. El primer grupo
está dado por la región 2 y 3, las cuales abarcan desde la cuenca Wagner hasta la
cuenca de Guaymas y presentan alta similitud (fig. 29B), seguidas por la región 4 y 8
que están geográficamente contiguas, y finamente el grupo con las regiones de mayor
profundidad, una de las cuales incluyen las cuencas del centro del Golfo (región 6) y
la región 9 que está en contacto directo con el Océano Pacífico. Estas últimas zonas
presentan la diversidad más baja de todas las regiones, con menos de 250 especies,
e incluso, en algunas zonas de estas regiones destaca la ausencia de registros para
algunos grupos de especies como los cnidarios y moluscos (fig. 24, 26). Esta
información es importante ya que es la primera en su tipo para el Golfo de California,
además es información base que podría servir de referencia para la determinación
zonas para la investigación y planeación territorial (Ray y Hayden 1992) como la
Reserva de la Biosfera Zona Marina Profunda Golfo de California, la cual propone
nueve polígonos en las cuencas y fallas del Golfo, ya que se infiere que estos
ecosistemas presentan un excelente estado de conservación con una enorme
diversidad biológica, (CONANP, 2011). Por otra parte, a diferencia de las regiones
costeras, en las regiones oceánicas no hubo coincidencia exacta con los distintos tipos
de sedimentos, el cual, en la zona oceánica es aparentemente más diversa en cuanto
al tipo de sedimentos que la zona costera (fig. 32), aunque para una mejor
comparación, es necesaria una actualización en la información del tipo de sedimentos
del Golfo de California.

67
Los biomas del Golfo de California.
Este trabajo utiliza únicamente datos de la superficie del mar para denotar a las
condiciones oceánicas como el componente ambiental y la distribución de
macroinvertebrados como el componente biológico para una primera aproximación en
la generación de biomas marinos del Golfo de California, el cual no había sido
abordado previamente en ningún trabajo de regionalización de esta zona. Aunque
varios de los limites propuestos en la regionalización oceanográfica y las
regionalizaciones biológicas presentadas anteriormente en esta tesis ya habían sido
propuestos en otros trabajos, estos límites variaron entre los distintos grupos de
especies y la regionalización oceanográfica también presentó diferencias respecto a
otras regionalizaciones que se basaron únicamente en variables físicas o químicas del
Golfo. Esta es la razón de por qué los 10 biomas encontrados al integrar toda la
información no coinciden de manera exacta con trabajos previos, los cuales se basaron
únicamente en algunas variables oceanográficas o en un número reducido de
especies, también puede deberse al hecho de que muchos trabajos de regionalización
no utilizaron herramientas de análisis espacial, ni información al nivel de detalle de
este trabajo.
El único trabajo nombrado como una bioregionalización del Golfo de California
es el de Santamaría del Ángel y colaboradores (1994), el cual analiza la variabilidad
espacio-temporal de los pigmentos fotosintéticos a nivel superficial como un indicador
biológico, ese trabajo presentó algunas similitudes con la propuesta de regionalización
oceanográfica de esta tesis, sin embargo, al compararlo con la regionalización
biológica los cinco grupos de invertebrados aquí seleccionados, los patrones no
coincidieron, por lo que tampoco coincide con esta propuesta de biomas del Golfo de
California, probablemente debido a que aquí se consideran una mayor cantidad de
variables físico-químicas y además de fauna que habita desde superficie hasta
profundidades superiores a los 4,000 m, donde se encuentra la masa de Agua
Profunda del Pacífico, la cual posee características muy distintas a la de Agua del
Golfo de California y el Agua Superficial Ecuatorial (Torres-Orozco, 1993; Lavín et al.,
1997; CONANP, 2011) encontradas en la superficie.

68
Los biomas del Golfo de California (fig. 30A), como se pudo apreciar, muestra
cuatro regiones costeras con cierta similitud a la propuesta de Walker (1960) para
peces, sin embargo lo importante de este trabajo es que es el primero que se realiza
a partir del análisis espacial y numérico de datos oceanográficos y biológicos, como
han sugerido algunos autores (Córdova et al., 2006; Hendrickx, 2008). Además de
localizar por primera vez los biomas de las zonas profundas del Golfo de California,
del cual aún se tiene poco conocimiento. Esta información puede resultar útil para
mejorar la planeación sistemática del medio marino, ya que se pueden usar de
contexto para la preservación de especies y ecosistemas (Lourie y Vincent, 2004).
Aunque en los biomas, ya que son el conjunto de características ambientales y
biológicas, se podría esperar que en un análisis de similitud, el acomodo de los biomas
en la parte ambiental dentro de un dendrograma correspondiera al acomodo que
tendrían esos biomas en la similitud biológica, esto no ocurre en el Golfo, como se
aprecia en la fig. 30 B y C, los biomas presentaron una similitud oceanográfica que
difiere de la similitud biológica, mientras que las especies se concentran en dos
grandes grupos, en un nodo los de alta diversidad (costeros) los cuales se acomodan
con su bioma contiguo, lo mismo ocurre con la similitud biológica de los biomas de
aguas profundas, dos grandes grupos (norte y sur) con alta similitud entre biomas
contiguos a diferencia del acomodo de los biomas por similitud oceanográfica, donde
la similitud no ocurre siempre entre los biomas contiguos, por ejemplo, aparecen dentro
del mismo nodo los biomas 1 y 2 (norte del Golfo) con la costa de Sinaloa, el cual
presenta altos valores de clorofila en ciertos meses del año (fig. 16), es decir, presenta
una estacionalidad marcada como ocurre con las regiones del norte y por ello esa
similitud. Sin embargo, en lo general se denota como los biomas van cambiando
latitudinalmente, así como los cambios que se dan en la similitud y diversidad biológica
(riqueza específica), en cada uno de ellos, además de en las condiciones ambientales
y es curioso que los lugares que ocupan estas zonas, sobre todo los biomas costeros
tengan aparente correspondencia con los sedimentos y tipo de costa a pesar de no
haberse usado esta información como insumo para la regionalización.

69
En este punto se puede decir que, a partir de la similitud, biológica y
oceanográfica, la zona templada del GC empieza Guaymas en el lado continental y en
la Bahía de San Francisquito, BCS, en el lado peninsular e incluye la zona profunda
desde la cuenca de Guaymas, ya que es notorio el cambio en la biodiversidad (fig. 30
D), la cual disminuye a pesar de tratarse zonas costeras (B1 y B3), este cambio de
diversidad biológica y el aislamiento de las características subtropicales del sur hacen
que la zona presente un alto endemismo (Hendrickx y Brusca, 2007). Una baja
diversidad y un alto endemismo son características de ecosistemas aislados, si bien la
parte sur del Golfo comparte mucha de la biodiversidad con zonas aledañas en
algunos grupos de especies (Walker, 1960; Simison, 2006), la parte norte del Golfo sí
puede considerarse como un ecosistema aislado.
Por otra parte, la Zona Mínima de Oxígeno (ZMO) el cual es una característica
ambiental importante ya que representa una barrera fisiológica para la migración
vertical en la columna de agua de las especies, por ejemplo, se sabe que en algunas
regiones, las bajas concentraciones de oxigeno genera zonas prácticamente azoicas
(sin fauna) (Serrano y Hendrickx, 2011). Esta variable se aborda en esta tesis solo con
los valores superficiales de oxígeno disuelto, sin embargo la ZMO se localiza en las
masas de agua profunda, y no se aborda en esta tesis debido a que en la actualidad
se cuenta con poca información al respecto, debido al alto costo operativo que se tiene
al llevar a cabo la toma de muestras, pues requieren un gasto de $5,000 a $50,000
USD o más por día de operación (Hendrickx, 2012), sin embargo sería importante
incorporar información de la columna de agua para generar resultados más exactos.
El Golfo de California es el segundo ecosistema marino y costero prioritario en
materia de conservación a nivel mundial (CONANP, 2011). En ese sentido, estos
resultados representan un avance substancial ya que este trabajo brinda información
base sobre la distribución y agrupamiento de especies y condiciones ambientales.

70
CONCLUSIONES
Con la información y las herramientas disponibles en la actualidad es posible
generar regionalizaciones con mayor sustento, para simplificar la complejidad del
océano. En el caso del Golfo de California, aunque se han realizado varios intentos de
regionalización oceanográfico y biológico, aún existe la necesidad de información
oceanográfica y de la distribución de las especies más detallada que los complemente,
sin embargo con este trabajo puede decirse que:
Resultó evidente que la zona costera y la zona oceánica presentan
características muy distintas (anexo I-X), por lo que un trabajo de regionalización debe
de separar ambas zonas, especialmente en mares de alta variabilidad oceanográfica
espacial y temporal como lo es el GC y no en cortes a lo ancho como con frecuencia
se hace.
Existen 14 regiones oceanográficas en el Golfo de California, siete
correspondientes a la zona costera y el resto a las zonas oceánicas, con un marcado
patrón estacional.
Cada phylum presenta distinto número de regiones, así como límites
geográficos variables.
En la mayoría de las regionalizaciones de los cinco fila, es notorio que las zonas
de mayor profundidad presentaron una mayor cantidad de regiones respecto a la
cantidad de zonas para la zona costera, a pesar de la baja biodiversidad de estos
sitios.
Los macroinvertebrados en su totalidad presentan una zonación de nueve
regiones dentro del Golfo de California, de los cuales, tres corresponden a la zona
costera, con una disminución en la biodiversidad de sur a norte, sin embargo, se
presenta un incremento en el endemismo.
Las regiones oceanográficas de mar abierto y las regionalizaciones de las
especies están altamente relacionado a las cuencas y fallas del Golfo.

71
Se determinó la existencia de 10 biomas marinos en el Golfo de California, el
cual es el resultado de fusionar las regiones oceanográficas con las biológicas en una
regionalización única.
Se encontró que los biomas costeros están limitados no solo por las condiciones
ambientales sino por el tipo de costa y de sedimentos.
Por otra parte, los biomas oceánicos presentaron relación con la complejidad
batimétrica de las zonas profundas del Golfo.

72
RECOMENDACIONES
Para mejorar este trabajo se recomienda incorporar información oceanográfica
de la columna de agua, especialmente del nivel de oxígeno disuelto, para determinar
la variación espacio temporal de la Zona Mínima de Oxígeno, el cual se considera
menor a 0.5 ml/l, esto es importante ya que una baja concentración limita la distribución
de organismos aeróbicos, los cuales pueden ser potencialmente explotables (Serrano,
2012). Existen pocas publicaciones acerca de la ZMO en el Pacífico Mexicano y el
Golfo de California (Parker, 1963; Gaxiola-Castro et al., 2002; Sánchez-Velasco et al.,
2004; Hendrickx y Serrano, 2010; Serrano y Hendrickx, 2011; y Serrano, 2012).
Aunque es poca la información, se sabe que en México la ZMO es muy amplia en el
sur del Golfo de California y a lo largo de la costa suroeste de México (Hendrickx, 2001;
Hendrickx y Serrano, 2010).
Específicamente en el Golfo de California, se sabe que en el sur, la ZMO
empieza a los 74 m (en las costas del estado de Sinaloa) y hacia el norte en la zona
central del Golfo de California empieza una profundidad de 255 m con un espesor casi
constante de 1,167 m (Serrano, 2012). Otro motivo por el cual es necesario la
integración de información oceanográfica de la columna de agua para generar un
mejor escenario de regionalización oceanográfica es que el Golfo está compuesto por
cinco masas de agua, las cuales tiene características fisicoquímicas muy distintivas
(Lavín y Marinone, 2003). Por lo que una mejora a este trabajo sería incluir esa
información ya que en esta tesis solo trabaja con datos oceanográficos de superficie,
sin embargo estos datos tienen que ser medidos in situ, que como se mencionó en un
principio, son escasos y solo se lleva monitoreo espacio temporal adecuado en pocas
partes del mundo (Callejas-Jiménez et al., 2012). Por otra parte, también es necesario
mejorar el conocimiento de la macrofauna y su distribución, especialmente en las
zonas más profundas del Golfo de California, pues los estudios son escasos, con lo
que se podría realizar una actualización en la distribución geográfica y batimétrica de
las especies para generar una mejor representación cartográfica y en consecuencia
una mejor regionalización biológica.

73
BIBLIOGRAFÍA
Allen, W. E. 1937. Plankton diatoms of the Gulf of California obtained by the G. Allan
Hancock Expedition of 1936. University of Southern California Press. Hancock
Pacific Exp. 3: 47-59.
Alvarez-Borrego, S. y J. R. Lara-Lara. 1991. The physical environment and primary
productivity of the Gulf of California. En: The Gulf and Peninsular Province of
the Californias.(J. P. Dauphin y B. R. T Simoneit, eds.). American Association
of Petroleum Geologist. Memoir, 47: 603-616.
Alvarez-Borrego, S. 1995. Satellite derived photosynthetic pigment surveys: A review
of marine phytoplankton biomass and productivity. Geofísica Internacional,
35(1).
Alvarez-Borrego, S. 2002. Physical Oceanography. En: T.J. Case, M.L. Cody y E.
Ezcurra (eds.). A new island biogeography of the sea of Cortes. Oxford
University Press. New York. 669 pp.
Alvarez Borrego, S. 2010. Physical, chemical, and biological oceanography of the Gulf
of California. En: Richard C. Brusca, Editor, The Gulf of California: biodiversity
and conservation. University of Arizona Press. ISBN: 2009046445. 24-48 p.
Álvarez Cáceres, R. 1994. Estadística multivariante y no paramétrica con SPSS.
Aplicación a las ciencias de la salud. Ed. Díaz de Santos. Madrid.
Arias-Arechíga, J.P. 1998. Regionalización del Golfo de California: Una propuesta a
partir de concentración de pigmentos fotosintéticos. Tesis de Licenciatura.
Universidad Autónoma de Baja California Sur. 57p.
Arriaga, L., E. Vázquez, J. González, R. Jiménez, E. Muñoz y V. Aguilar. 1998.
Regiones prioritarias marinas de México. Comisión Nacional para el
Conocimiento y Uso de la Biodiversidad, México.
Ayón-Parente, M., M.E. Hendrickx y A. Hinojosa-Corona. 2014. Análisis de las
comunidades observadas en transectos. Capítulo 7. Pp 224-243. En:
Explorando el mar profundo del Golfo de California. CICESE, México

74
Boschi, E. 2000. Species of decapod crustaceans and their distribution in the American
marine zoogeographic provinces. Rev. Invest. Des. Pesq. 13:7–136
Briggs, J. C., y Bowen, B. W. 2012. A realignment of marine biogeographic provinces
with particular reference to fish distributions. Journal of Biogeography, 39(1),
12-30.
Brinton, E. y A. W. Townsend. 1980. Euphasiids in the Gulf of California, the 1957
Cruises. Calif. Coop. Oceanic Fish. Invest. Rep. 21: 211-236
Brusca, R. C., Findley, L. T., Hastings, P. A., Hendrickx, M. E., Cosio, J. T., y van der
Heiden, A. M. 2005. Macrofaunal diversity in the Gulf of California. Biodiversity,
ecosystems, and conservation in Northern Mexico, 179.
Brusca, R. C. y M. E. Hendrickx. 2008. The Gulf of California Invertebrate Database:
The Invertebrate Portion of the Macrofauna Golfo Database.
http://www.desertmuseum.org/center/seaofcortez/database.php.
Brusca, R. C. y M. E. Hendrickx. 2010. Invertebrates biodiversity and conservation. En:
The Gulf of California: biodiversity and conservation. Brusca, R. C. (Ed.).
(2010). University of Arizona Press.
Brusca, R. C. (Ed.). (2010). The Gulf of California: biodiversity and conservation.
University of Arizona Press.
Callejas-Jimenez, M., Santamaría-del-Angel, E., Gonzalez-Silvera, A., Millán-Nuñez,
R., y Cajal-Medrano, R. 2012. Dynamic Regionalization of the Gulf of Mexico
based on normalized radiances (nLw) derived from MODIS-Aqua. Continental
Shelf Research, 37, 8-14.
Campero, M., Van Damme, P., Arévalo, B., y Claros, D. 2003. Análisis de
correspondencia canónica vs. regresión múltiple: ¿pueden mostrar diferencias
entre hidroecoregiones?. Revista Boliviana de Ecología, 13, 55-64.
Castro-Aguirre JL, Balart EF y Arvizu-Martínez J. 1995. Contribución al conocimiento
del origen y distribución de la ictiofauna del Golfo de California, México.
Hidrobiología. 5: 1-2

75
Chollett, I., Mumby, P. J., Müller-Karger, F. E., y Hu, C. 2012. Physical environments
of the Caribbean Sea. Limnology and Oceanography, 57(4), 1233.
CONABIO-CONANP-TNC-PRONATURA. 2007. Análisis de vacíos y omisiones en
conservación de la biodiversidad marina de México: océanos, costas e islas.
Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Conabio),
Conanp, The Nature Conservancy, Pro Natura, Semarnat, CBD, México, DF
Comisión Nacional de Áreas Naturales Protegidas (CONANP). 2011. Estudio Previo
Justificativo para el establecimiento del área natural protegida con la categoría
de Reserva de la Biosfera Zona Marina Profunda Golfo de California, localizada
frente a las costas de los Estados de Baja California, Baja California Sur,
Sonora, Sinaloa, Nayarit y Jalisco, México, 117 páginas + 4 anexos. Marzo
2012.
Contreras, E. F. 2002, Importancia de la pesca ribereña en México. Departamento de
Hidrobiología, División de Ciencias Biológicas y de la Salud, Universidad
Autónoma Metropolitana-Iztapalapa, ContactoS 46: 5-14.
Córdova, A., Vázquez, F. R. V., Hernández, G. E., y De la Torre, B. H. 2006.
Ordenamiento ecológico marino. Visión integrada de la regionalización.
Instituto Nacional de Ecología. México 226p
Cupp, E. E., y W. E. Allen. 1938. Plankton diatoms of the Gulf of California obtained by
the Allan Hancock Pacific Expedition of 1937, in Hancock Pacific Expedition,
Publ. 3, pp. 61-74, Univ. of South. Calif., Los Angeles.
De la Lanza-Espino, G. 2001. Características Físico-Químicas de los Mares de México.
Temas Selectos de Geografía en México. Instituto de Geografía, UNAM,
México.
Dietrich, D. 1957. General Oceanography: An Introduction. Interscience Publishers,
New York.
Escalante, F., Valdez-Holguín, J. E., Álvarez-Borrego, S., y Lara-Lara, J. R. 2013.
Variación temporal y espacial de temperatura superficial del mar, clorofila a y

76
productividad primaria en el golfo de California. Ciencias marinas, 39 (2), 203-
215.
Escalante, T. 2009. Un ensayo sobre regionalización biogeográfica. Revista mexicana
de biodiversidad, 80(2), 551-560.
Fernández, E., P. Boyd, P.M. Holligan y D.S. Harbour. 1993. Production of organic and
inorganic carbon within a large scale coccolithophore bloom in the North Atlantic
ocean. Marine Ecology Progress Seris. 97: 271-285
Gaxiola-Castro G., S. Álvarez-Borrego, S. Nájera-Martínez y A. R. Zirino. 2002. Efecto
de las ondas internas en el fitoplancton del golfo de California. Ciencias Marinas
28: 297-309
Gaxiola-Castro G., Gracía-Córdova J., Valdez-Holguín J.E. y. Botello-Ruvalcaba M.
1995. Spatial distribution of chlorophyll a and primary productivity in relation to
winter physical structure in the Gulf of California. Continental Shelf Research.
15: 1043-1059.
Lindsey, George E. y Iris H. W. Engstrand. 2002. History of scientific exploration in the
Sea of Cortés. In: Ted J. Case, Martin L. Cody y Exequiel Ezcurra (eds.), A New
Island Biogeography of the Sea of Cortés. New York: Oxford University Press.
pp. 3-13
Gilbert, J. Y. y W. E. Allen. 1943. The phytoplankton of the Gulf of California obtained
by the "E. W. Scripps" in 1939 and 1940, J. Mar. Res., 5(2), 89-110.
González-Cuellar, O. 2012.Estructura comunitaria y zonación geográfica de peces de
arrecifes rocosos del norte del Golfo de California. Tesis de licenciatura.
Universidad Autónoma de Baja California Sur.
González-Silvera, A., E. Santamaría-del-Ángel, V. M. T. García, C. A. E García, R.
Millán-Núñez, F. Müller-Karger. 2004. Biogeographically regions of the
tropical and subtropical Atlantic Ocean off South America: classification
based on pigment (CZCS and chlorophyll-a (SeaWiFS) variability.
Continental Shelf Research. 24, 983-1000

77
Gower, J. C. 1971. A general coefficient of similarity and some of its
properties. Biometrics, 857-871.
Hayden, B. P., Ray, G. C., y Dolan, R. 1984. Classification of coastal and marine
environments. Environmental Conservation, 11(03), 199-207.
Hendrickx, M.E. 2001. Occurrence of a continental slope decapod crustacean
community along the edge of the minimum oxygen zone in the southeastern
Gulf of California, Mexico. Belgian Journal of Zoology131 (Suppl. 2): 95-109.
Hendrickx, M. E. 2012. Operaciones oceanográficas en aguas profundas: los retos del
pasado, del presente y del proyecto TALUD en el Pacífico mexicano (1989-
2009). Biodiversidad y comunidades del talud continental del Pacífico
mexicano. P. Zamorano, ME Hendrickx y M. Caso (eds.). Secretaría del Medio
Ambiente y Recursos Naturales (SEMARNAT), Instituto Nacional de Ecología
(INE), 23-104.
Hendrickx, M. E., Brusca, R. C. y Ramírez-Reséndiz, G. 2002. Biodiversity of
macrocrustaceans in the Gulf of California, Mexico. Contributions to the Study
of East Pacific Crustaceans. Instituto de Ciencias del Mar y Limnologıa, UNAM,
Mexico.
Hendrickx, M. E., Brusca, R. C., Cordero, M. y Ramírez, G. 2007. Marine and brackish-
water molluscan biodiversity in the Gulf of California, Mexico. Scientia
Marina, 71(4), 637-647.
Hendrickx, M.E., D.L. Pawson, B.E. Mejía-Mercado, M. Ayón-Parente y A. Hinojosa-
Corona. 2014. Registros de especies de Holothuroidea (Echinodermata).
Capítulo 5. Pp. 193-203. En: Explorando el mar profundo del golfo de California
2008-2014. CICESE, Baja California, México.
Hendrickx, M. E. y R. C. Brusca. 2007. Distribución de invertebrados marinos
endémicos en el golfo de California, México. Memorias del XII Congreso Latino-
Américano de Ciências do Mar XII COLACMAR, 15-19 abril, Florianópolis. p. 1-
4

78
Hendrickx, M.E. y D. Serrano. 2010. Impacto de la zona de mínimo de oxígeno sobre
los corredores pesqueros en el Pacifico mexicano. Interciencia 35:12-18
Hendrickx, M.E. 2008. Biodiversidad y ecosistemas: El caso del Pacífico Mexicano. En:
Córdova, Rosete, Enríquez, Fernández (Compiladores), Marine Ecologic
Ordering. Thematic Vision of Regionalization. México.
Hernández-Alfonso, J. R., Martínez-López, A., y Maeda-Martínez, A. N. 2007.
Variabilidad del carbono orgánico particulado en una laguna somera de Baja
California. Hernández de la Torre B., G. Gaxiola (eds). Carbono en
Ecosistemas Acuáticos de México. Instituto Nacional de Ecología y CICESE.
México, 159-170.
Hidalgo-González, R. M., y Alvarez-Borrego, S. 2004. Total and new production in the
Gulf of California estimated from ocean color data from the satellite sensor
SeaWIFS. Deep Sea Research Part II: Topical Studies in Oceanography, 51(6),
739-752.
Instituto Nacional de Pesca. 1994. Atlas Pesquero de México. ISBN 968-817-319-3
International Hydrographic Organization (IHO).1953. Limits of Oceans and Seas,
International Hydrographic Organization. Bremerhaven, PANGAEA .
IOCCG, 2009. Partition of the Ocean into Ecological Provinces: Role of Ocean-Colour
Radiometry. Dowell, M. y Platt, T. (eds.), Reports of the International Ocean
Colour Coordinating Group, No. 9, IOCCG, Dartmouth, Canada.
Kahru, M., Marinone, S. G., Lluch-Cota, S. E., Parés-Sierra, A., y Greg Mitchell, B.
2004. Ocean-color variability in the Gulf of California: scales from days to
ENSO. Deep Sea Research Part II: Topical Studies in Oceanography,51(1),
139-146.
Kintigh, K. W., y Ammerman, A. J. 1982. Heuristic approaches to spatial analysis in
archaeology. American Antiquity, 31-63.
Krebs CJ. 1989. Ecological methodology. Harper y Row, NY, USA.

79
Lara Lara, J. R., V. Arenas Fuentes, M. D. C. Bazan Guzman, M. De La Cruz García
Abad, V. M. Diaz Castañeda, E. Escobar Briones, G. Gaxiola Castro, E. G.
Robles Jarero, R. Sosa Avalos, L. A. Soto González, M. Tapia García y J. E.
Valdez Holguín. 2008. Los ecosistemas marinos. En: Jorge Soberón, Gonzalo
Halffter y Jorge Llorente, Editores, "Capital Natural de Mexico. Vol. I:
Conocimiento actual de la biodiversidad". Comisión Nacional para el
Conocimiento y Uso de la Biodiversidad.
Lavín, M.F., E. Beier, y A. Badan. 1997. Estructura hidrográfica y circulación del Golfo
de California: Escalas estacional e interanual. En M.F. Lavín. Ed.
Contribuciones a la Oceanografía Física en México. Monografía No. 3. Unión
Geofísica Mexicana.
Lavín, M. F., y Marinone, S. G. 2003. An overview of the physical oceanography of the
Gulf of California. In Nonlinear processes in geophysical fluid dynamics.
Springer Netherlands.
Linares Fleites, G. 1990. Análisis de datos. Universidad de La Habana. Facultad de
Matemática-Cibernética, ENPES, MES, 321-47.
Lourie, S. A., y Vincent, A. C. 2004. Using biogeography to help set priorities in marine
conservation. Conservation Biology, 18(4), 1004-1020.
Lluch-Cota SE, Arias-Aréchiga JP. 2000. Sobre la importancia de considerar la
existencia de centros de actividad biológica para la regionalización del océano:
El caso del Golfo de California. En: Lluch-Belda D, Elorduy-Garay J,Lluch-Cota
SE, Ponce-Díaz G (eds.), BAC Centros de Actividad Biológica del Pacífico
Mexicano. CIB, CICIMAR, CONACYT. La Paz, México.
Longhurst, A., Sathyendranath, S., Platt, T., y Caverhill, C. 1995. An estimate of global
primary production in the ocean from satellite radiometer data. Journal of
Plankton Research, 17(6), 1245-1271.
López-Martínez J, Acevedo-Cervantes A., Herrera-Valdivia E., Rodríguez-Romero J. y
Palacios-Salgado D.S. 2012. Composición taxonómica y aspectos

80
zoogeográficos de peces de profundidad (90-540m) del Golfo de California,
México. Rev. Biol. Trop. (Int. J. Trop. Biol. ISSN-0034-7744). 60, (1). 347-360.
Marinone, S. G. 2012. Seasonal surface connectivity in the Gulf of California.Estuarine,
Coastal and Shelf Science, 100, 133-141.
Massin, C., y Hendrickx, M. E. 2011. Deep-water Holothuroidea (Echinodermata)
collected during the TALUD cruises off the Pacific coast of Mexico, with the
description of two new species. Revista Mexicana de Biodiversidad, 82(2), 413-
443.
Méndez, N. 2012. Poliquetos (Annelida, Polychaeta) del talud continental suroriental
del golfo de California y su relación con algunas variables ambientales.
Biodiversidad y comunidades del talud continental del Pacífico mexicano.
Instituto Nacional de Ecología, Secretaría del Medio Ambiente y Recursos
Naturales (Semarnat), 161-222.
Merrifield, M. A. y C. D. Winant. 1989. Shelf circulation in the Gulf of California: A
description of the Variability. Journal of Geophysical Research, 94: 18133-
18160
Millán-Núñez, R., Santamaría-del-Angel, E., González-Silvera, A., 2006. Methods for
the regionalization of the ocean zone through biologic aspects. In: Córdova,
Rosete, Enríquez, Fernández (Compiladores), Marine Ecologic Ordering.
Thematic Vision of Regionalization. México.
Miranda, I. 1997. Análisis de cluster como estrategia multivariada de clasificación.
Solución a un problema taxonómico. Tesis para optar por el título de Master en
Matemática Aplicada a las Ciencias Agropecuarias. CENSA- ISAAC
Molina-Cruz A. 1984. Evolución oceanográfica de la Boca del Golfo de California.
Anales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional.
Morrone, J. J. y J. Márquez. 2003. Aproximación a un atlas biogeográfico mexicano:
Componentes bióticos principales y provincias biogeográficas. En Una

81
perspectiva latinoamericana de la biogeografía, J. J. Morrone y J. Llorente
(eds.). Las Prensas de Ciencias, UNAM, México, D.F. p. 1-4.
Okolodkov, Y. 2010. Biogeografía Marina. Centro EPOMEX-UAC.
Parker, R.H. 1963. Zoogeography and ecology of some macro-invertebrates,
particularly mollusks, in the Gulf of California and the continental slope off
Mexico. Vidensk avelige fra Dansk Medd Naturhistorisk Forening 126: 1-178
Pegau, W. S., Boss, E., y Martínez, A. 2002. Ocean color observations of eddies during
the summer in the Gulf of California. Geophysical Research Letters, 29(9), 6-1.
Petatán-Ramirez D., R. Brusca, H. Reyes-Bonilla y J.L. García-Corona. “Spatial
patterns of mollusc species richness in the Gulf of California and a proposal of
biogeographic zonation”. Congreso Mollusca. Ciudad de México, Junio 2014.
Pielou, E.C. 1979. Biogeography. Wiley-Interscience, New York, EE.UU. 351 pp.
Prieto Guerra, R.E., 2006.Técnicas estadísticas de clasificación, un ejemplo de análisis
cluster. Tesis de licenciatura. Universidad Autónoma del estado de Hidalgo.
Ray, G.C. y B.P. Hayden. 1992. Coastal zones ecotones. En: A. J. Hansen y F. di
Castri (eds). Landscape Boundaries, Consequences for Biotic Diversity and
Ecological Flows. Springer-Verlag, New York, EE.UU. 452 pp.
Ripa P. 1997. Towards a physical explanation of the seasonal dynamics and
thermodynamics of the Gulf of California Journal of Physical Oceanography 27
597-614
Robertson, D. R., y Cramer, K. L. 2009. Shore fishes and biogeographic subdivisions
of the Tropical Eastern Pacific. Marine Ecology Progress Series,380, 1-17.
Roden, G.I.1958. Oceanographic and meteorological aspects of the Gulf of California.
Pac. Sci., (12)21–45
Roden, G. I., y I. Emilsson. 1979. Physical oceanography of the Gulf of California, paper
presented at the Gulf of California Symposium, Univ. Nac. Autón. de Méx.,
Mazatlán, Sinaloa, México.

82
Round, F. E. 1967. The phytoplankton of the Gulf of California, I, Its composition,
distribution and contribution to the sediments, J. Exp. Mar. Biol. Ecol., 1, 76--
97.
Sánchez-Velasco, L., V.M. Godínez-Sandoval, B. Shirasago, y M. Peguero-Icaza.
2004. Informe Técnico 16560. Serie Oceanografía Física. CICESE. México. A1-
28. 12 pp.
Sánchez Velasco Laura, Lavín M.L., Peguero-Icaza M., León-Chávez C.A., Contreras-
Catala F., Marinoe S.G., Gutiérrez-Palacios I.V., y Godínez V.M. 2009.
Seasonal changes of larval fish assemblages in a semi-enclosed sea (Gulf of
California). Continental Shelf Research. 29(14): 1697-1710.
Sánchez-Velasco, L., Lavin, M. F., Jiménez-Rosenberg, S. P. A., Montes, J. M., y Turk-
Boyer, P. J. 2012. Larval fish habitats and hydrography in the Biosphere
Reserve of the Upper Gulf of California (June 2008). Continental Shelf
Research, 33, 89-99.
Sánchez-Velasco, L., Lavín, M. F., Jiménez-Rosenberg, S. P. A., Godínez, V. M.,
Santamaría-del-Angel, E., y Hernández-Becerril, D. U. 2013. Three-
dimensional distribution of fish larvae in a cyclonic eddy in the Gulf of California
during the summer. Deep Sea Research Part I: Oceanographic Research
Papers, 75, 39-51.
Sánchez-Velasco, L., Lavín, M. F., Jiménez-Rosenberg, S. P. A., y Godínez, V. M.
2014. Preferred larval fish habitat in a frontal zone of the northern Gulf of
California during the early cyclonic phase of the seasonal circulation (June
2008). Journal of Marine Systems, 129, 368-380.
Santamaría del Angel, E., Alvarez-Borrego, S., y Müller‐Karger, F. E. 1994. Gulf of
California biogeographic regions based on coastal zone color scanner imagery.
Journal of Geophysical Research: Oceans (1978–2012), 99(C4), 7411-7421.
Santamaría del Angel, E., Millán-Núñez, R., González-Silvera, A., y Cajal-Medrano, R.
2010. Comparison of In Situ and Remotely-Sensed Chl-a concentrations: a
Statistical Examination of the Match-up Approach. Chap. 15, 221-238.

83
Santiago García, M. W., S. G. L. Marinone Moschetto y O. U. Velasco Fuentes. 2014.
Three-dimensional connectivity in the Gulf of California based on a numerical
model. Progress in Oceanography.
Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT). 2006. Programa
de Ordenamiento Ecológico Marino del Golfo de California. Secretaría de Medio
Ambiente y Recursos Naturales, México, 129 pp.
Serrano, D. y M.E. Hendrickx. 2011. Debajo de la zona del mínimo de oxígeno.
Investigación y Ciencia 417: 13-15.
Serrano, D. 2012. La zona del mínimo de oxígeno en el Pacífico
mexicano.Biodiversidad y comunidades del talud continental del Pacífico
mexicano. Instituto Nacional de Ecología, Secretaría del Medio Ambiente y
Recursos Naturales (Semarnat), 101-115.
Siegel-Causey, D. 1982. Factors determining the distribution of hyperiid Amphipoda
inthe Gulf of California. Ph. D. Dissertation, Univ. Arizona.
Simison, W. B. 2006. Patterns of endemism for nearshore gastropod molluscs: a new
view of the Gulf of California. Journal of molluscan studies,72(1), 53-63.
Soto-Mardones, L., Marinone, S. G., y Parés-Sierra, A. 1999. Variabilidad
espaciotemporal de la temperatura superficial del mar en el golfo de california
time and spatial variability of sea surface temperature in the gulf of
california. Ciencias Marinas, 25(1), 1-30.
Sheng, Y. S. Y. 1999. Visualizing upwelling at Monterey Bay in an integrated
environment of GIS and scientific visualization. Marine Geodesy,22(2), 93-103.
Spalding, M. D., Fox, H. E., Allen, G. R., Davidson, N., Ferdaña, Z. A., Finlayson, M.
A. X., y Robertson, J. 2007. Marine ecoregions of the world: a bioregionalization
of coastal and shelf areas. BioScience, 57(7), 573-583.
Sverdrup, H. U. 1941. The Gulf of California: Preliminary discussion of the cruise of the
EW Scripps in February and March 1939. In Proc. 6th Pac. Sci. Congr (Vol. 3,
pp. 161-166).

84
Szteren, D., y Aurioles-Gamboa, D. 2011. Ecological regionalization of Zalophus
californianus rookeries, as a tool for conservation in the Gulf of California
Regionalización ecológica de las colonias reproductivas de Zalophus
californianus, como herramienta para su conservación en el Golfo de
California.Ciencias Marinas, 37(3), 349-368.
Thomson DA, Findley LT y Kersthich A. 2000. Reef fishes of the sea of Cortez. Wiley.
New York. 374 p
Torres, L. G., Read, A. J., y Halpin, P. 2008. Fine-scale habitat modeling of a top marine
predator: do prey data improve predictive capacity. Ecological Applications,
18(7), 1702-1717.
Torres-Orozco E. 1993. Análisis volumétrico de las masas de agua del golfo de
California. MSc thesis, Centro de Investigación científica y de Educación
Superior de Ensenada, México, 80 pp
Trasviña A., D. Lluch-Cota, A.E. Filonov y A. Gallegos. 1999. “Oceanografía y El Niño”.
En : V. Magaña, V. (ed.) Los Impactos del Niño en México. UNAM.
Ulloa, R., Torre, J., Bourillón, L., Gondor, A., y Alcanzar, N. 2006. Planeación
ecorregional para la conservación marina: Golfo de California y costa occidental
de Baja California Sur. Final report to the Nature Conservancy. Guaymas
(Mexico): Comunidad y Biodiversidad, AC.
Walker BW. 1960. The distribution and affinities of the marine fish fauna of the Gulf of
California. Systematic Zoology. 9: 123-133
Ward, J.H.J., 1963. Hierarchical grouping to optimize an objective function. J. Am.Stat.
Assoc. 58, 236–244.
Wilkinson T, Wiken E, Bezaury Creel J, Hourigan T, Agardy H, Herrmann H,
Janishevski L, Madden C, Morgan L y Padilla M. 2009. Ecoregiones Marinas de
América del Norte. Comisión para la cooperación ambiental. Montreal. 200 p.
Zacharias, M y J. Roff. 2000. A hierarchical approach to conserving marine biodiversity.
Conservation Biology, 14(5):1327-1334.

85
Zamorano, P., Hendrickx, M. E., Ríos-Jara, E., Esqueda-González, M. C., y Galván-
Villa, C. M. 2007. Biocenosis y distribución de los moluscos de aguas profundas
en el Pacífico mexicano: una evaluación de los avances. Estudios Sobre la
Malacología y conquiliología en México, 48-49.
Zamorano, P., Hendrickx, M. E., y Toledano-Granados, A. 2007. New geographic and
depth records for deep-water mollusks in the Gulf of California, Mexico. Revista
Mexicana de Biodiversidad, 78(2), 311-318.
Zamudio, L., P. Hogan, y E. J. Metzger. 2008. Summer generation of the Southern Gulf
of California eddy train, J. Geophys. Res., 113
Zar, H. J. 1999. Biostatistical Analisis, Prentice Hall, USA, New Jersey, 123 pp.

86
ANEXO I
Año tipo de la temperatura superficial en promedios mensuales (2003 – 2012).

87
ANEXO II
Año tipo de la Clorofila a superficial en promedios mensuales (2003 – 2012).

88
ANEXO III
Año tipo de la atenuación de la luz en promedios mensuales (2003 – 2012).
.

89
ANEXO IV
Año tipo de la radiación fotosintética en promedios mensuales (2003 – 2012)

90
ANEXO V
Año tipo del carbono inorgánico superficial en promedios mensuales (2003 – 2012)
91
ANEXO VI
Año tipo del carbono orgánico superficial en promedios mensuales (2003 – 2012).

92
ANEXO VII
Año tipo de la materia coloreada en promedios mensuales (2003 – 2012).

93
ANEXO VIII
Valor promedio histórico (1939-2007) de fosfato, silicato, oxígeno disuelto y salinidad.

94
ANEXO IX
Valores máximos de las regiones en las variables medidas por el color del océano.
Región Temperatura Clorofila
a
Carbono
Inorgánico
Carbono
Orgánico
Radiación
Fotosintética
Atenuación Materia
Coloreada
Delta del Río Colorado 31.859 3.666 0.04816 358.349 57.487 0.29 5.381
Alto Golfo 31.600 3.373 0.01249 405.101 59.575 0.25 6.720
Golfo Norte 30.794 3.473 0.00134 483.693 60.341 0.26 4.904
Grandes islas 29.852 3.194 0.00083 542.942 60.001 0.24 5.435
Isla Tiburón 32.007 7.569 0.00571 700.649 59.203 0.55 5.839
Grandes islas sur 31.026 2.781 0.00091 390.358 59.744 0.22 4.489
Cuenca de Guaymas 31.067 2.068 0.00077 367.588 59.506 0.17 4.850
Sur de Sonora 31.946 7.256 0.00385 700.838 59.076 0.54 5.434
Cuencas Carmen -
Farallón
31.094 1.545 0.00067 286.241 59.119 0.14 5.024
La Paz-Loreto 30.504 2.107 0.00069 479.095 59.058 0.18 5.931
Sinaloa – Nayarit 31.772 6.761 0.00569 575.430 57.323 0.49 6.150
Cuenca Pescadero 30.818 1.259 0.00052 234.684 58.029 0.12 4.849
Boca del Golfo 29.061 0.598 0.00036 126.037 54.446 0.07 4.289
Islas Marías 31.608 1.430 0.00066 221.263 58.395 0.13 5.434
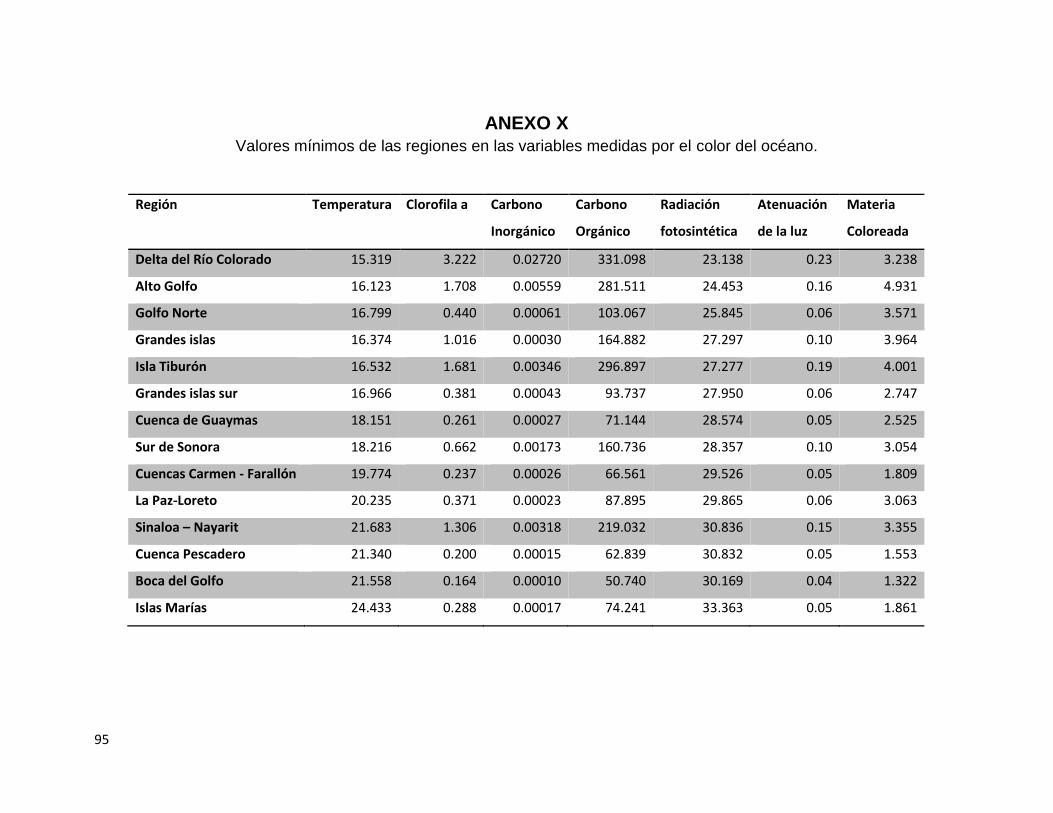
95
ANEXO X Valores mínimos de las regiones en las variables medidas por el color del océano.
Región Temperatura Clorofila a Carbono
Inorgánico
Carbono
Orgánico
Radiación
fotosintética
Atenuación
de la luz
Materia
Coloreada
Delta del Río Colorado 15.319 3.222 0.02720 331.098 23.138 0.23 3.238
Alto Golfo 16.123 1.708 0.00559 281.511 24.453 0.16 4.931
Golfo Norte 16.799 0.440 0.00061 103.067 25.845 0.06 3.571
Grandes islas 16.374 1.016 0.00030 164.882 27.297 0.10 3.964
Isla Tiburón 16.532 1.681 0.00346 296.897 27.277 0.19 4.001
Grandes islas sur 16.966 0.381 0.00043 93.737 27.950 0.06 2.747
Cuenca de Guaymas 18.151 0.261 0.00027 71.144 28.574 0.05 2.525
Sur de Sonora 18.216 0.662 0.00173 160.736 28.357 0.10 3.054
Cuencas Carmen - Farallón 19.774 0.237 0.00026 66.561 29.526 0.05 1.809
La Paz-Loreto 20.235 0.371 0.00023 87.895 29.865 0.06 3.063
Sinaloa – Nayarit 21.683 1.306 0.00318 219.032 30.836 0.15 3.355
Cuenca Pescadero 21.340 0.200 0.00015 62.839 30.832 0.05 1.553
Boca del Golfo 21.558 0.164 0.00010 50.740 30.169 0.04 1.322
Islas Marías 24.433 0.288 0.00017 74.241 33.363 0.05 1.861

96
ANEXO XI
Correlación de las variables oceanográficas. Donde: sst_M=Temperatura superficial máxima, sst_m=Temperatura Superfcial mínima,
Chla_M=Clorofila a máxima, Chla_m=Clorofila a mínima, pic_M=Carbono inórganico máximo, pic_m=Carbono inorgánico mínimo, poc_M=Carbono
orgánico máximo, poc_m=carbono orgánico mínimo, par_M=Radiación fotosintética máxima, par_m= Radiación fotosintética mínima,
k490_M=atenuación de luz máxima, k490_m=atenuación de luz mínima, cdom_M=Materia coloreada máxima, cdom_m=Materia coloreada mínima,
Sal=Salinidad, Po4=Fosfatos, SiO2=Silicatos, O2=Oxígeno.
Variables sst_M Chla_M pic_M poc_M par_M k490_M cdom_M sst_m Chla_m pic_m poc_m par_m k490_m cdom_m sal Po4 Sio2 O2
sst_M 1.00
Chla_M 0.64 1.00
pic_M 0.39 0.18 1.00
poc_M 0.47 0.90 0.01 1.00
par_M 0.34 0.21 -0.18 0.50 1.00
k490_M 0.64 1.00 0.19 0.90 0.20 1.00
cdom_M 0.53 0.50 0.23 0.51 0.25 0.49 1.00
sst_m -0.17 -0.35 -0.45 -0.47 -0.50 -0.34 -0.14 1.00
Chla_m 0.49 0.47 0.90 0.32 -0.04 0.47 0.51 -0.56 1.00
pic_m 0.39 0.18 1.00 0.01 -0.19 0.19 0.20 -0.44 0.89 1.00
poc_m 0.58 0.67 0.72 0.52 0.08 0.66 0.66 -0.59 0.94 0.71 1.00
par_m -0.20 -0.27 -0.66 -0.31 -0.31 -0.26 -0.21 0.93 -0.71 -0.64 -0.67 1.00
k490_m 0.58 0.65 0.77 0.48 -0.01 0.65 0.61 -0.54 0.97 0.77 0.99 -0.64 1.00
cdom_m 0.37 0.60 0.28 0.70 0.54 0.57 0.73 -0.70 0.59 0.26 0.75 -0.68 0.66 1.00
sal 0.72 0.29 0.51 0.36 0.70 0.29 0.45 -0.53 0.54 0.50 0.52 -0.54 0.50 0.55 1.00
Po4 0.19 0.44 0.21 0.57 0.58 0.43 0.10 -0.89 0.43 0.21 0.51 -0.70 0.46 0.66 0.46 1.00
Sio2 0.16 0.32 0.23 0.34 0.40 0.29 0.24 -0.51 0.47 0.23 0.53 -0.47 0.47 0.64 0.42 0.63 1.00
O2 0.44 0.41 0.15 0.47 0.57 0.40 0.33 -0.59 0.32 0.14 0.45 -0.57 0.39 0.62 0.50 0.53 0.45 1.00
Top Related