






Languages
Pages
Legal

8/12/2019 Cell Membrane Final-Dr.urvashi
1/75
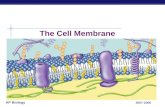
Cell Membrane
Dr.Urvashi Kumar Kohli
8/12/2019 Cell Membrane Final-Dr.urvashi
2/75
crucial to the life of the cell
encloses the cell,
defines its boundaries, and
maintains the essential differences between the cytosol and the
extracellular environment.
Cell Membrane
8/12/2019 Cell Membrane Final-Dr.urvashi
3/75
1.1 Functions and properties of membrane
Membrane is a selectively permeablebarrierbetween the cell and the external environment.
Its function is to maintainHomeostasis. Its selectivepermeability allows the cell to maintain aconstant internal environment.
Plasma membranes formcompartments(compartmentation) within cells.
8/12/2019 Cell Membrane Final-Dr.urvashi
4/75
Functions of Cell Membrane are to:
1. regulate cell volume
(control water in/ out)
2. maintain intracellular pH(H+regulation)
3. selectively regulate ionic
composition(e.g. Na+
, K+
)4. concentrate metabolic fuel
(nutrients, ATP, etc)
,
8/12/2019 Cell Membrane Final-Dr.urvashi
5/75
Membrane transport system ought to (continued):
5. concentrate and move building blocks (amino
acids, etc)
6. remove toxic compounds (detoxification, trap
and/or pump out)7. generate ionic gradientsto maintain excitability
of nerve and muscle cells
8. control theflow of informationwithin the cell,
between cells and their environment.
8/12/2019 Cell Membrane Final-Dr.urvashi
6/75
A BRIEF HISTORY OF STUDIES ON
PLASMA MEMBRANE STRUCTURE
Ernst Overton(1890s) - Chemical nature of the outer boundary layer
To test the permeability of the outer boundary layer, Overton placed plant root hairs
into hundreds of different solutions containing a diverse array of solutes. He discovered
that the more lipid soluble the solute, the more rapidly it would enter the root hair
cells
Remarks------dissolving power of the outer boundary layer of the cell
matched that of a fatty oil
8/12/2019 Cell Membrane Final-Dr.urvashi
7/75
-- the polar groups of each molecular layer (or leaflet) were directed outward
toward the aqueous environment
E. Gorter and F. Grendel(1925)
Ratio of the surface area of water covered by the extracted lipid to the surface area
calculated for the red blood cells from which the lipid was extracted varied between1.8 to 1 and 2.2 to 1 and concluded that the plasma membrane contained a
bimolecular layer of lipids, that is, a lipid bilayer
8/12/2019 Cell Membrane Final-Dr.urvashi
8/75
In 1935, Hugh Davson and James Danielli proposed that the plasmamembrane was composed of a lipid bilayer that was lined on both its inner and
outer surface by a layer of globular proteins
Davson and Danielli suggested that, in addition
to the outer and inner protein layers, the lipid
bilayer was also penetrated by protein-lined
pores, which could provide conduits for polar
solutes and ions to enter and exit the cell.
8/12/2019 Cell Membrane Final-Dr.urvashi
9/75
Fluid-mosaic model proposed in 1972 by S. Jonathan Singer and Garth
Nicolson
A current representation of the plasma membrane showing the same basic organization as that proposed bySinger and Nicolson
The fluid-mosaic model of membrane structure as initially
proposed by Singer and Nicolson in 1972.
8/12/2019 Cell Membrane Final-Dr.urvashi
10/75
All biological membranes have a common general
structure:Lipid and protein molecules, held together mainly by noncovalent
interactions.Characteristics:dynamic, fluid structures, and most of their molecules are able to move about in
the plane of the membrane
lipid molecules - double layer about 6 nm thick
- bilayer provides the basic structure of the membrane
- serves as a relatively impermeable barrier to the
passage of most water-soluble molecules.
Protein molecules - transport specific molecules
- catalyzing membrane associated reactions, such as ATP
synthesis- structural links - connect the membrane to the cytoskeleton
and/or to either the extracellular matrix or
an adjacent cell,
- receptors
Carbohydrates -cell membranes are asymmetrical structures
8/12/2019 Cell Membrane Final-Dr.urvashi
11/75
Figure - 1. Three views of a cell membrane. (A) An electron micrograph of a plasma
membrane (of a human red blood cell) seen in cross-section. (B and C) Schematic
drawings showing twodimensional and three-dimensional views of a cell
membrane.
8/12/2019 Cell Membrane Final-Dr.urvashi
12/75
8/12/2019 Cell Membrane Final-Dr.urvashi
13/75
Phospholipids(Amphipathic)
Sphingolipids(amphipathic)
8/12/2019 Cell Membrane Final-Dr.urvashi
14/75
Figure -2. The parts of a phospholipid molecule. Phosphatidylcholine, represented
schematically (A), in formula (B), as a space-filling model (C), and as a symbol (D). The kink
due to the cis-double bond is exaggerated in these drawings for emphasis
8/12/2019 Cell Membrane Final-Dr.urvashi
15/75
Figure 10-3. A lipid micelle and a lipid bilayer seen in cross-section. Lipid molecules form
such structures spontaneously in water. The shape of the lipid molecule determines
which of these structures is formed. Wedge-shaped lipid molecules (above) form micelles,
whereas cylindershaped phospholipid molecules (below) form bilayers.
8/12/2019 Cell Membrane Final-Dr.urvashi
16/75
Figure - 4 . Phospholipid mobility. The types of movement possible for phospholipid
molecules in a lipid bilayer.
8/12/2019 Cell Membrane Final-Dr.urvashi
17/75
The Fluidity of a Lipid Bilayer Depends on Its Composition
Phase Transition
- Hydrocarbon chains(short and Double bond)
- cholestrolInhibit phase transition
Composition A. phospholipids ( 4 major)
- phosphatidylcholine, sphingomyelin,
phosphatidylserine, and
phosphatidylethanolamine.
- inositol phospholipidsB. Cholestrol
C. Glycolipids
8/12/2019 Cell Membrane Final-Dr.urvashi
18/75
Figure - 5. Influence of cis-double bonds in hydrocarbon chains. The double bonds
make it more difficult to pack the chains together and therefore make the lipid bilayer
more difficult to freeze.
8/12/2019 Cell Membrane Final-Dr.urvashi
19/75
Figure - 6. The structure of cholesterol. Cholesterol is represented by a formula in
(A), by a schematic drawing in (B), and as a space-filling model in (C).
8/12/2019 Cell Membrane Final-Dr.urvashi
20/75
Figure - 7. Cholesterol in a lipid bilayer. Schematic drawing of a cholesterol molecule
interacting with two phospholipid molecules in one leaflet of a lipid bilayer.
The Lipid Bilayer Is Asymmetrical
8/12/2019 Cell Membrane Final-Dr.urvashi
21/75
Figure - 8. Four major phospholipids in mammalian plasma membranes. Note that different
head groups are represented by different symbols in this figure and the next. All of the lipid
molecules shown are derived from glycerol except for sphingomyelin, which is derived fromserine
The Lipid Bilayer Is Asymmetrical
8/12/2019 Cell Membrane Final-Dr.urvashi
22/75
Figure - 9. The asymmetrical distribution of phospholipids and glycolipids in the lipidbilayer of human red blood cells. The symbols used for the phospholipids are those
introduced in Figure- 8. In addition, glycolipids are drawn with hexagonal polar head
groups (blue). Cholesterol (not shown) is thought to be distributed about equally in both
monolayers.
The Lipid Bilayer Is Asymmetrical
8/12/2019 Cell Membrane Final-Dr.urvashi
23/75
Plasma membrane, are synthesized in the endoplasmic reticulum (ER), andit is here that the phospholipid asymmetry is generated by the phospholipid
translocators in the ER that move specific phospholipid molecules from one
monolayer to the other.
8/12/2019 Cell Membrane Final-Dr.urvashi
24/75
Sphingolipids (amphipathic)
8/12/2019 Cell Membrane Final-Dr.urvashi
25/75
Glycolipids Are Found on the Surface of All Plasma Membranes
5% of the lipid molecules in the outer monolayer
most complex of the glycolipids, the gangliosides - sialic acid
- most abundant in the plasma membrane of nerve cells, where they
constitute 5-10% of the total lipid mass,
8/12/2019 Cell Membrane Final-Dr.urvashi
26/75
Figure 10-12. Glycolipid molecules. Galactocerebroside (A) is called a neutral glycolipid because
the sugar that forms its head group is uncharged. A ganglioside (B) always contains one or more
negatively charged sialic acid residues (also called N-acetylneuraminic acid, or NANA), whose structure is shown in (C). Whereas in bacteria
and plants almost all glycolipids are derived from glycerol, as are most phospholipids, in animal cells they are almost always produced from
sphingosine, an amino alcohol derived from serine, as is the case for the phospholipid sphingomyelin (see Figure- . Gal = g
8/12/2019 Cell Membrane Final-Dr.urvashi
27/75
functions of glycolipids
In the plasma membrane of epithelial cells, -glycolipids protect the membrane from
the harsh conditions (such as low pH and degradative enzymes).
Charged glycolipids, such as gangliosides, (electrical effects): their presence will alter
the electrical field across the membrane and the concentrations of ions- especially
Ca2+ - at its external surface.
Glycolipids may also play a role in electrical insulation, since in the myelin
membrane, which electrically insulates nerve cell axons, the noncytoplasmic half of
the bilayer is filled with them.
They are also thought to function in cell-recognition processes.
Nature and Importance of the Lipid Bilayer
8/12/2019 Cell Membrane Final-Dr.urvashi
28/75
Table -1 .Approximate Lipid Compositions of Different Cell Membranes
Nature and Importance of the Lipid Bilayer
8/12/2019 Cell Membrane Final-Dr.urvashi
29/75
The Nature and Importance of the Lipid Bilayer
1. diversity in composition in different bacteria---physical state depends upon lipid
composition
2.Thickness - 60 Angstron
3.Continuous unbroken structure
4.self resealing property
5.flexible-shape changes (during locomotion n cell division)
6.lipid bilayer- facilitate regulated fusion and budding of membranes
7.maintaining the proper internal composition of a cell,
8. in separating electric charges across the plasma membrane
9. Self Assemble( PC Liposomes )
2 Proteins
8/12/2019 Cell Membrane Final-Dr.urvashi
30/75
2. Proteins
THE STRUCTURE AND FUNCTIONS OF MEMBRANE PROTEINS
- Asymmetry of Proteins orientation in Membrane(membrane "sidedness")
- membrane protein- has a defined orientation relative to the cytoplasm.
-Therefore properties of one surface of a membrane are very different from those
of other surface the other surface
Three Classes of Proteins
a. Integral membrane
Proteins
b. Peripheral Proteins
c. GPI anchored Proteins
2 Proteins
8/12/2019 Cell Membrane Final-Dr.urvashi
31/75
Figure - Six ways in which membrane proteins associate with the lipid bilayer. Most transmembrane proteins are
thought to extend across the bilayer as a single helix (1) or as multiple helices (2); some of these "single-pass"
and "multipass" proteins have a covalently attachedfatty acid chain inserted in the cytoplasmic monolayer (1).
Other membrane proteins are attached to the bilayer solely by a covalently attached lipid - either a fatty acid
chain or prenyl group - in the cytoplasmic monolayer (3) or, less often, via an oligosaccharide, to a minor
phospholipid, phosphatidylinositol, in the noncytoplasmic monolayer (4). Finally, many proteins are attached to
the membrane only by noncovalent interactions with other membrane proteins (5) and (6). How
the structure in (3) is formed is illustrated in next Figure
Membrane Proteins Can Be Associated with the Lipid Bilayer in Various Ways
2. Proteins
Integral Membrane Protein
8/12/2019 Cell Membrane Final-Dr.urvashi
32/75
Figure 10-14. The covalent attachment of either of two types of lipid groups can help localize a
water-soluble protein to a membrane after its synthesis in the cytosol. (A) A fatty acid chain
(either myristic or palmitic acid) is attached via an amide linkage to an amino-terminal glycine. (B)
A prenyl group (either farnesyl or a longer geranylgeranyl group - both related to cholesterol) is
attached via a thioether linkage to a cysteine residue that is four residues from the carboxyl
terminus. Following this prenylation, the terminal three amino acids are cleaved off and the new
carboxyl terminus is methylated before insertion into the membrane. The structures of two lipid
anchors are shown underneath: (C) a myristyl anchor (a 14-carbon saturated fatty acid chain),
and (D) a farnesyl anchor (a 15-carbon unsaturated hydrocarbon chain).
In Most Transmembrane Proteins the Polypeptide Chain Is Thought to Cross the Lipid
8/12/2019 Cell Membrane Final-Dr.urvashi
33/75
Figure 10-15. A segment of a transmembrane polypeptide chain crossing the lipid
bilayer as an alpha helix. Only the - carbon backbone of the polypeptide chain is
shown, with the hydrophobic amino acids in green and yellow.
yp p g p
Bilayer in an alpha-helical Conformation
8/12/2019 Cell Membrane Final-Dr.urvashi
34/75
Figure . The three-dimensional structure of a porin trimer of Rhodobacter capsulatus
determined by x-ray crystallography. (A) Each monomer consists of a 16-stranded antiparallel b-
barrel that forms a transmembrane water-filled channel. (B) The monomers tightly associate to
form trimers, which have three separate channels for the diffusion of small solutes through the
bacterial outer membrane. A long loop of polypeptide chain (shown in red), which connects two b-strands, protrudes into the lumen of each channel, narrowing it to a cross-section of 0.6 x 1 nm.
Porins Are Pore-forming Transmembrane Proteins That Cross the Lipid Bilayer as a
beta- Barrel
Synthesis of TMM (Asymmetry of proteins)
8/12/2019 Cell Membrane Final-Dr.urvashi
35/75
Figure 10-17. A typical single-pass transmembrane protein. Note that the polypeptide chain
traverses the lipid bilayer as a right-handed alpha- helix and that the oligosaccharide chains and
disulfide bonds are all on the noncytosolic surface of the membrane. Disulfide bonds do not form
between the sulfhydryl groups in the cytoplasmic domain of the protein because the reducingenvironment in the cytosol maintains these groups in their reduced (-SH) form.
8/12/2019 Cell Membrane Final-Dr.urvashi
36/75
location of proteins- reflects the functionIMM(TM)- Functions on both sides- transport molecules across it.e.g. cell-surface receptors-TM proteins.Peripheral membrane proteins-e.g. protiens involved in intracellular signaling (cytosolic half).
TMM(IMM)Unique Orientation(Text)
---non-Cytoplasmic, membrane- spanning domain and cytoplasmic domain
8/12/2019 Cell Membrane Final-Dr.urvashi
37/75
Figure 10-18. A detergent micelle in water, shown in cross-section. Because
they have both polar and nonpolar ends, detergent molecules are
amphipathic.
Membrane Proteins Can Be Solubilized and Purified in Detergents
8/12/2019 Cell Membrane Final-Dr.urvashi
38/75
Figure 10-19. Solubilizing membrane proteins with a mild detergent. The detergent disrupts the
lipid bilayer and brings the proteins into solution as protein-lipid-detergent complexes. The
phospholipids in the membrane are also solubilized by the detergent.
8/12/2019 Cell Membrane Final-Dr.urvashi
39/75
Figure 10-20. The structures of two commonly used detergents. Sodium dodecyl sulfate (SDS) is
an anionic detergent, and Triton X-100 is a nonionic detergent. The hydrophobic portion of each
detergent is shown in green, and the hydrophilic portion is shown in blue. Note that the bracketedportion of Triton X-100 is repeated about eight times
8/12/2019 Cell Membrane Final-Dr.urvashi
40/75
Figure . The use of mild detergents for solubilizing, purifying, and reconstituting functional
membrane protein systems. In this example functional Na+-K+ ATPase molecules are purified and
incorporated into phospholipid vesicles. The Na+-K+ ATPase is an ion pump that is present in the
plasma membrane of most animal cells; it uses the energy of ATP hydrolysis to pump Na+ out ofthe cell and K+ in,
Cells Can Confine Proteins and Lipids to Specific Domains
8/12/2019 Cell Membrane Final-Dr.urvashi
41/75
Figure . Diagram of an epithelial cell showing how a plasma membrane protein is restricted
to a particular domain of the membrane. Protein A (in the apical membrane) and protein B (in the
basal and lateral membranes) can diffuse laterally in their own domains but are prevented from
entering the other domain, at least partly by the specialized cell junction called a tight junction.
Lipid molecules in the outer (noncytoplasmic) monolayer of the plasma membrane are likewise
unable to diffuse between the two domains; lipids in the inner (cytoplasmic) monolayer, however,
are able to do so (not shown).
Within a Membrane
8/12/2019 Cell Membrane Final-Dr.urvashi
42/75
Six major functions of membrane proteins:
Transport
Enzymatic activity
Signal transduction
Cell-cell recognition
Intercellular joining
Attachment to the cytoskeleton and extracellular
matrix (ECM)
LE 7-9a
8/12/2019 Cell Membrane Final-Dr.urvashi
43/75
EnzymesSignal
ReceptorATP
Transport Enzymatic activity Signal transduction
LE 7-9b
8/12/2019 Cell Membrane Final-Dr.urvashi
44/75
Glyco-protein
Cell-cell recognition Intercellular joining Attachment to the
cytoskeleton and extra-cellular matrix (ECM)
8/12/2019 Cell Membrane Final-Dr.urvashi
45/75
The Role of Membrane Carbohydrates
8/12/2019 Cell Membrane Final-Dr.urvashi
46/75
The Role of Membrane Carbohydratesin Cell-Cell Recognition
Cells recognize each other by binding to surface
molecules, often carbohydrates, on the plasma
membrane
Membrane carbohydrates may be covalently bonded to
lipids (forming glycolipids) or more commonly toproteins (forming glycoproteins)
Carbohydrates on the external side of the plasma
membrane vary among species, individuals, and even
cell types in an individual
Plasma membrane is a semi-permeable selective)
8/12/2019 Cell Membrane Final-Dr.urvashi
47/75
Lipid bilayer
Small hydrophobic molecules: O2,CO2, N2, benzene
Small uncharged polar molecules: H2O,ethanol, glycerol
Larger uncharged polar molecules:glucose, amino acid, nucleotides
membrane/ Membrane structure results inselective permeability
Ions: H+, Na+, HCO3-, K+, Ca+,Mg2+, CL-, etc. Transporters or
channels
The process of diffusionof water is called osmosis.
But why some can diffuse while some cannot? What criteria are controlling the diffusion of
molecules across the membrane ?
Plasma membrane is a semi-permeable (selective)
http://en.wikipedia.org/wiki/Diffusionhttp://en.wikipedia.org/wiki/Osmosishttp://en.wikipedia.org/wiki/Osmosishttp://en.wikipedia.org/wiki/Diffusion8/12/2019 Cell Membrane Final-Dr.urvashi
48/75
Plasma membrane is a semi-permeable (selective) membrane
The process of diffusionof
water is called osmosis.
But why some can diffuse
while some cannot? What
criteria are controlling the
diffusion of moleculesacross the membrane ?
Figure 11-1. The relative permeability of a synthetic lipid bilayer to different classes of molecules.
The smaller the molecule and, more important, the fewer hydrogen bonds it makes with water, the
more rapidly the molecule diffuses across the bilayer.
Protein - free Lipid Bilayers Are Highly
Impermeable to Ions
http://en.wikipedia.org/wiki/Diffusionhttp://en.wikipedia.org/wiki/Osmosishttp://en.wikipedia.org/wiki/Osmosishttp://en.wikipedia.org/wiki/Diffusion8/12/2019 Cell Membrane Final-Dr.urvashi
49/75
Simple diffusion : cell membranes allow water and non-polar molecules to
permeate
Channel protein- Passive diffusion
Carrier Protein- Active or Passive
Two classes of membrane transport proteins
8/12/2019 Cell Membrane Final-Dr.urvashi
50/75
Figure 11-3. A schematic view of the two classes of membrane transport proteins. A carrier
protein is thought to alternate between two conformations, so that the solute binding site is
sequentially accessible on one side of the bilayer and then on the other. In contrast, a channel
protein is thought to form a water-filled pore across the bilayer through which specific ions can
Diffuse.
Two classes of membrane transport proteins
8/12/2019 Cell Membrane Final-Dr.urvashi
51/75
Figure 11-4. Comparison of passive transport down an electrochemical gradient with active
transport against an electrochemical gradient. Whereas simple diffusion and passive transport by
membrane transport proteins (facilitated diffusion) occur spontaneously, active transport requires
an input of metabolic energy. Only carrier proteins can carry out active transport, but both carrier
proteins and channel proteins can mediate facilitated diffusion.
Three types of carrier-mediated transport26
8/12/2019 Cell Membrane Final-Dr.urvashi
52/75
Figure - Three types of carrier-mediated transport. The schematic diagram shows carrier
proteins functioning as uniports, symports, and antiports.
Uniport-
Glucose carriers in
animal cells take upglucose from
extracellular fluid.
(passive transport)
Symport -glucose with Na+
intestinal cells and Kidneycellstake up glucose from
the lumen of the intestine
and kidney tubules(low)
Antiport
Band 3 protein
(anion carrier)
In human RBC- exchange Cl-
for HCO3-)
Conformational change in a carrier protein could mediate the facilitated diffusion of a
8/12/2019 Cell Membrane Final-Dr.urvashi
53/75
Figure . A hypothetical model showing how a conformational change in a carrier protein could
mediate the facilitated diffusion of a solute. The carrier protein shown can exist in two
conformational states: in state "pong" the binding sites for solute A are exposed on the outside of
the bilayer; in state "ping" the same sites are exposed on the other side of the bilayer. The
transition between the two states is proposed to occur randomly and to be completely reversible.
Therefore, if the concentration of A is higher on the outside of the bilayer, more A will bind to the
carrier protein in the pong conformation than in the ping conformation, and there will be a net
transport of A down its electrochemical gradient.
solute
8/12/2019 Cell Membrane Final-Dr.urvashi
54/75
Table 11-1. Comparison of Ion Concentrations
Inside and Outside a Typical Mammalian Cell
Pumping cycle of the Na+-K+ ATPasePrimary Acitve Transport
8/12/2019 Cell Membrane Final-Dr.urvashi
55/75
Figure . A schematic model of the pumping cycle of the Na+-K+ ATPase. The binding of Na+ (1) and the subsequent phosphorylation by ATP of
the cytoplasmic face of the ATPase (2) induce the protein to undergo a conformational change that transfers the Na+ across the membrane and
releases it on the outside (3). Then the binding of K+on the extracellular surface (4) and the subsequent dephosphorylation (5) return the protein
to its original conformation, which transfers the K+ across the membrane and releases it into the cytosol (6). These changes in conformation are
analogous to the ping pong transitions shown in Figure - except that here the Na+- dependent phosphorylation and the K+-dependent
dephosphorylation of the protein cause the conforma-tional transitions to occur in an orderly manner, enabling the protein to do useful work.
Although for simplicity only one Na+- and one K+-binding site are shown, in the real pump there are thought to be three Na+- and two K+-
binding sites. Moreover, although the ATPase is shown as alternating between two conformational states, there is evidence that it goes through
a more complex series of conformational changes during the actual pumping cycle.
8/12/2019 Cell Membrane Final-Dr.urvashi
56/75
Figure 11-10. The Na+-K+ ATPase. This carrier protein actively pumps Na+ out of and K+ into a
cell against their electrochemical gradients. For every molecule of ATP hydrolyzed inside the cell,
three Na+ are pumped out and two K+ are pumped in. The specific pump inhibitor ouabain and K+
compete for the same site on the external side of the ATPase.
8/12/2019 Cell Membrane Final-Dr.urvashi
57/75
Secondary Active Transport: Ion Gradients
Na+Glucose
Cl-
HClO3-
8/12/2019 Cell Membrane Final-Dr.urvashi
58/75
Cells maintain voltage across
8/12/2019 Cell Membrane Final-Dr.urvashi
59/75
Cells maintain voltage acrossplasma membranes
Cells maintain voltage across plasmamembranes.
Cytoplasm negative compared toopposite side of membrane(membrane potential - ranges from
-50 to -200 millivolts)
8/12/2019 Cell Membrane Final-Dr.urvashi
60/75
8/12/2019 Cell Membrane Final-Dr.urvashi
61/75
Membrane potential favors passivetransport of cations (positive ions)into cell and anions (negative ions)out of cell.
Creates an electrochemical
gradient across membrane.
8/12/2019 Cell Membrane Final-Dr.urvashi
62/75
Some organisms have proton pumpsthat actively pump H+out of cell(i.e. plants, bacteria, and fungi)
8/12/2019 Cell Membrane Final-Dr.urvashi
63/75
Maintenance of Membrane Potential
8/12/2019 Cell Membrane Final-Dr.urvashi
64/75
Maintenance of Membrane Potential
by Ion Pumps
Membrane potential is the voltage difference across amembrane
Two combined forces, collectively called the
electrochemical gradient, drive the diffusion of ions
across a membrane: A chemical force (the ions concentration gradient)
An electrical force (the effect of the membrane
potential on the ions movement)
8/12/2019 Cell Membrane Final-Dr.urvashi
65/75
An electrogenic pump is a transport protein thatgenerates the voltage across a membrane
The main electrogenic pump of plants, fungi, and
bacteria is a proton pump
Ionophores
8/12/2019 Cell Membrane Final-Dr.urvashi
66/75
Figure . A mobile ion carrier and a channel-forming ionophore. In both cases net ion flow
occurs only down an electrochemical gradient.
Valinomycin
(K+)Gramicidin A
Ionophores
8/12/2019 Cell Membrane Final-Dr.urvashi
67/75
Figure 11-6. The structure of a gramicidin channel. The channel is formed by the association of
two identical peptides at their amino-terminal ends. Each chain is folded into a b helix, which
resembles a rolled-up b pleated sheet. (A) is a side view and (B) a top view. The peptide
backbones that line the channel are shown in blue and dark green, while the light green
represents the protruding hydrophobic side chains. The lipid bilayer is shown in gray. (C) shows
the size of unhydrated K+ions, while (D) shows a membrane-spanning a helix in top view for
comparison with the b helix.
Bulk transport across the plasma
8/12/2019 Cell Membrane Final-Dr.urvashi
68/75
p p
membrane occurs by exocytosis and
endocytosis
Small molecules and water enter or leave the cell
through the lipid bilayer or by transport proteins Large molecules, such as polysaccharides and proteins,
cross the membrane via vesicles
8/12/2019 Cell Membrane Final-Dr.urvashi
69/75
Exocytosis In exocytosis, transport vesicles migrate to the
membrane, fuse with it, and release their contents
Many secretory cells use exocytosis to export their
products
8/12/2019 Cell Membrane Final-Dr.urvashi
70/75
Endocytosis In endocytosis, the cell takes in macromolecules by
forming vesicles from the plasma membrane
Endocytosis is a reversal of exocytosis, involving
different proteins
8/12/2019 Cell Membrane Final-Dr.urvashi
71/75
Three types of endocytosis:
Phagocytosis (cellular eating): Cell engulfs particlein a vacuole
Pinocytosis (cellular drinking): Cell creates vesiclearound fluid
Receptor-mediated endocytosis: Binding of ligandsto receptors triggers vesicle formation
LE 7-20c
8/12/2019 Cell Membrane Final-Dr.urvashi
72/75
Receptor
RECEPTOR-MEDIATED ENDOCYTOSIS
Ligand
Coated
pit
Coated
vesicle
Coat protein
Coat
protein
Plasma
membrane
0.25 m
A coated pit
and a coated
vesicle formed
during
receptor-
mediated
endocytosis(TEMs).
8/12/2019 Cell Membrane Final-Dr.urvashi
73/75
Synthesis and Sidedness of Membranes
Asymmetry of plasma membranes
8/12/2019 Cell Membrane Final-Dr.urvashi
74/75
Asymmetry of plasma membranes
1. Membrane separates interior and exterior side of the cell
and the 2 faces of lipid bilayer are different in compositionand structure, with different proteins and phospholipids.2. The plasma membrane core contain 1/3 cholesterol and 2/3
phospholipids and sphingolipids, the outer leaflet contains5% glycolipids.
3. Oligosaccharide chains are attached at outer face of lipidsor proteins.
4. Shingomyelin and phosphatidylcholine are mainly at theouter face of the bilayer.
5. Phosphotidylethanolamine and phosphatidylserine aremainly in the inner face.
6. Asymmetry of proteins- Oligosaccharides and Disulphidebonds in the protein on non-cytoplasmis side
Ch i l d h i l ti f l
8/12/2019 Cell Membrane Final-Dr.urvashi
75/75
1. Sheet-likestructure consists of proteins,lipids, with carbohydrates attached to them.
2. Non-covalent assemblies of lipids and proteins.3. The proteins serve the functions as
transporters, channels, enzymes, signaltransducers, etc.
4. The lipid molecules areamphipathicmolecules, they have both hydrophilic (polar)
and hydrophobic (non-polar) moieties.5. Barriersto the flow of charged molecules.
Chemical and physical properties of plasma
membranes
Top Related