Unidad 2. Las Células Del Sistema Nervioso
31
Unidad 2 Las células del sistema Nervioso Historia del estudio de la neurona El nombre de neurona fue dado a la célula nerviosa por Waldeyer. Su descubrimiento se acredita a Ehrenberg, quien la describió en la corteza cerebral y en los ganglios espinales, en 1883. Fontana en 1871, hizo la primera descripción detallada de la fibra nerviosa. Remark, entre 1836 y 1844, identifico las fibras mielínicas y amielínicas (estas últimas se llamaron más tarde fibras de Remark) y sugirió la posible continuidad entre la célula y la fibra nerviosa, como hacen notar Clarear y O'Malley (1968, citado en López Antúnez, 1986). El advenimiento del microscopio acromático compuesto hacia la tercera década del siglo pasado y el perfeccionamiento de las técnicas de fijación y coloración del tejido nervioso, hicieron posible estudiar detalladamente la célula nerviosa. La neuroanatomía de la segunda mitad del siglo XIX se caracterizó por la famosa controversia entre los que postulaban la continuidad citoplasmica entre las células nerviosas a través de una red de fibras y los partidarios de la independencia anatómica de las neuronas, cuya relación sería solamente por contigüidad. Los reticulistas más notables fueron Golgi, Gerlach, Held y Lenhossek, entre otros. Por la otra parte, los principales representantes de la teoría neuronista fueron His, Forel y principalmente Cajal. Los estudios de Esclerosis sobre la degeneración nerviosa confirmaron el concepto de la individualidad anatómica de las neuronas, al demostrar que la lesión de una célula nerviosa no afecta a las que están en relación con ella. His, al investigar el desarrollo de las neuronas y Forel mediante el método de degeneración retrógrada y la técnica de impregnación de plata de Golgi, contribuyeron considerablemente a fundamentar la teoría neuronista cuyo
-
Upload
rafaelcalvillogarcia -
Category
Documents
-
view
38 -
download
1
description
Las Células Del Sistema Nervioso
Transcript of Unidad 2. Las Células Del Sistema Nervioso

Unidad 2Las células del sistema Nervioso
Historia del estudio de la neurona
El nombre de neurona fue dado a la célula nerviosa por Waldeyer. Su descubrimiento se acredita a Ehrenberg, quien la describió en la corteza cerebral y en los ganglios espinales, en 1883. Fontana en 1871, hizo la primera descripción detallada de la fibra nerviosa. Remark, entre 1836 y 1844, identifico las fibras mielínicas y amielínicas (estas últimas se llamaron más tarde fibras de Remark) y sugirió la posible continuidad entre la célula y la fibra nerviosa, como hacen notar Clarear y O'Malley (1968, citado en López Antúnez, 1986).
El advenimiento del microscopio acromático compuesto hacia la tercera década del siglo pasado y el perfeccionamiento de las técnicas de fijación y coloración del tejido nervioso, hicieron posible estudiar detalladamente la célula nerviosa.
La neuroanatomía de la segunda mitad del siglo XIX se caracterizó por la famosa controversia entre los que postulaban la continuidad citoplasmica entre las células nerviosas a través de una red de fibras y los partidarios de la independencia anatómica de las neuronas, cuya relación sería solamente por contigüidad. Los reticulistas más notables fueron Golgi, Gerlach, Held y Lenhossek, entre otros. Por la otra parte, los principales representantes de la teoría neuronista fueron His, Forel y principalmente Cajal.
Los estudios de Esclerosis sobre la degeneración nerviosa confirmaron el concepto de la individualidad anatómica de las neuronas, al demostrar que la lesión de una célula nerviosa no afecta a las que están en relación con ella. His, al investigar el desarrollo de las neuronas y Forel mediante el método de degeneración retrógrada y la técnica de impregnación de plata de Golgi, contribuyeron considerablemente a fundamentar la teoría neuronista cuyo máximo exponente fue Cajal, quien aportó pruebas concluyentes de la misma.
Hacia 1891, Waldeyer después de hacer una evaluación de las evidencias derivadas de las investigaciones mencionadas, enuncio la doctrina neuronal aplicando al sistema nervioso la teoría celular que habían propuesto independientemente Schwann y Schleiden, en 1838 y 1839, respectivamente. Aunque Waldeyer como señalan Clarear y O'Malley, no hizo contribuciones originales al respecto, coopero enormemente en divulgar el concepto de la neurona como unidad embrionaria, morfológica y funcional. La introducción del concepto sinapsis por Sherrington, para denotar el sitio en que dos neuronas se ponen en contacto y la subsecuente comprobación, mediante la microscopia electrónica, del espacio que las separa a ese nivel, confirmaron las ideas neuronistas.
En 1906 fue otorgado conjuntamente el premio nobel de medicina y fisiología a Cajal y a Golgi, los investigadores que defendieron conceptos antagónicos,

dirigiendo la controversia científica que tan fecundos resultados aportó al conocimiento de la estructura del sistema nervioso.
La Doctrina Neuronal. Tal como se enuncia actualmente, comprende cuatro postulados:
a) La neurona es la unidad genética del sistema nervioso.
b) La neurona es la unidad anatómica del sistema nervioso.
c) La neurona es la unidad funcional del sistema nervioso.
d) La neurona es la unidad trófica del sistema nervioso.
a)La neurona se desarrolla de acuerdo con un proceso que a partir de un neuroblasto la lleva, pasando por diferentes etapas, hasta el estado adulto. Esto significa que cada neurona proviene de una célula primitiva, el neuroblasto, y como pierde su capacidad para reproducirse, si por alguna razón es destruida, no puede ser sustituida. Por su origen y su proceso de diferenciación ulterior, se considera a la neurona como la unidad genética o embrionaria del sistema nervioso.
b)Cada neurona constituye una entidad estructural morfológicamente independiente de las demás células nerviosas. La relación entre ellas es por contigüidad, sin continuidad citoplasmica. El sitio en que dos neuronas se ponen en contacto se llama sinapsis y en está siempre hay un espacio que separa las membranas de las células vecinas, lo que significa que la relación entre las neuronas es únicamente funcional. Por esto se considera a la neurona como la unidad anatómica del sistema nervioso.
c)La neurona es una célula especializada capaz de conducir impulsos nerviosos. Aunque, fisiológicamente, una célula nerviosa aislada no tiene significación pues se requieren cuando menos dos neuronas para constituir un sistema que integre la relación neuronal más elemental, todas las vías nerviosas están formadas por neuronas, ya que estas representan el elemento más simple a través del cual pueden cursar los impulsos. Por esto se le considera la unidad funcional del sistema nervioso.
d)Todas las partes de la neurona dependen para su nutrición de su relación con el cuerpo celular: por consiguiente si una parte de la célula nerviosa queda separada del mismo, se degenera. Por ello se considera a la neurona como la unidad trófica del sistema nervioso (López Antúnez, 1986).
La célula como unidad de organización del tejido nervioso
El cerebro es el órgano que mueve los músculos. Esta afirmación puede parecer un poco simple, pero en última instancia el movimiento --o más precisamente la conducta-- constituye la función principal del sistema nervioso. Para que los movimientos sean eficaces, el cerebro debe saber lo que ocurre en su entorno. De

este modo, el organismo contiene células especializadas en la detección de los acontecimientos del entorno y células especializadas en la producción de movimientos. Naturalmente los animales complejos, como nosotros mismos, no reaccionamos automáticamente ante los acontecimientos ambientales; nuestros cerebros son lo suficientemente flexibles como para poder comportarnos de formas diferentes de acuerdo no sólo con las circunstancias presentes sino también con las experimentadas en nuestro pasado. Además de percibir y actuar, somos capaces de recordar y decidir. Todas estas capacidades son posibles gracias a los miles de millones de células que se encuentran en nuestro sistema nervioso.
La, información ya sea en forma de luz, ondas sonoras, olores, gusto o contacto con objetos, es captada del entorno mediante células especializadas denominadas neuronas sensoriales. Los movimientos tienen lugar mediante la contracción de los músculos, controlados a su vez por neuronas motoras (el término motor alude a movimiento y o a un motor mecánico.). Entre las neuronas sensoriales y las motoras se halan todas las interneuronas-- neuronas que se encuentran totalmente en el sistema nervioso central--. Los circuitos de interneuronas (principalmente del encéfalo) son responsables de las funciones de percepción aprendizaje, recuerdo, decisión y control de las conductas complejas. ¿Cuántas neuronas hay en el sistema nervioso humano? Podemos hacer una estimación de entre 100,000 millones y 1 billón, pero todavía nadie las ha contado (Carlson, 2006).
Técnicas neurocitológicas
La estructura intrínseca de la neurona, aunque es básicamente como la de las otras células, tienen algunas características especiales. Los conocimientos de neurocitología se han acumulado a lo largo de muchas décadas debido a la aplicación de diversos métodos de tinción para la microscopia de luz, y más recientemente, por el uso del microscopio electrónico.
Las tinciones de hematoxilina y eosina son muy útiles en la mayoría de los tejidos, tienen poca aplicación en el estudio del sistema nervioso normal por medio de la microscopia de luz. E cambio se usan técnicas especiales, de las cuales las más importantes son las siguientes.
Tinciones catiónicas, como el violeta de cresilo, el azul de toluidina, y tionina, llamadas con frecuencia "tinciones de Nissl”, que se fijan a los ácidos nucleicos. Por tanto destacan el núcleo y el nucléolo, y también la sustancia cromatofílica (de Nissl) que se encuentra en el citoplasma de las neuronas.
En la Figura 1 se observa un corte delgado de cerebro de gato teñido según dos procedimientos: la tinción de Nissl, que tiñe todos los cuerpos celulares de violeta y la tinción de Golgi que muestra los perfiles de algunas neuronas teñidos de negro observándose las siluetas de los cuerpos celulares y sus prolongaciones. El método de Golgi tiñe el 5 por ciento o menos de las neuronas, si se tiñeran todas las neuronas del tejido el corte parecería casi totalmente negro (Fig. 1).

Figura 1 Tinción de Nissl y Golgi en neuronas. Fuente: ht.org.ar.
El método de Golgi tiene muchas variantes, es muy valioso en el estudio de la morfología neuronal, especialmente de las dendritas. En el método original, las porciones de tejido se tratan en secuencia con soluciones que contienen bicromato de potasio y nitrato de plata: los cortes se preparan de 100μm a 200μm de grueso. Algunas neuronas, incluyendo las finas ramas de sus dendritas destacan en café o negro en contraste con un fondo claro. Ocasionalmente se manifiestan células de la neuroglia, pero en general sus axones no se tiñen. La principal ventaja de estos métodos es que se tiñe una pequeña cantidad de células, ya que si todas se oscurecieran sería imposible determinar los detalles estructurales (Fig. 1).
Los métodos de reducción argéntica o de plata, que producen depósitos oscuros de plata coloidal en varias estructuras. Las técnicas más usadas son las de Ramón y Cajal, de Bielschowsky, de Bodian y de Holmes (para axones). Otros métodos especialmente los de Ramón y Cajal, del Rio Hortega, y Penfield, son útiles para la demostración selectiva de tiempos diferentes de células de neuroglia.
Las tinciones para la mielina destacan las fibras de los tractos más importantes a diferencia de los pocos que consisten exclusivamente de axones amielínicos. Con menor aumento se ven los tractos mielinizados en azul oscuro mientras que las áreas celulares, como los núcleos, permanecen incoloras. La combinación de tinciones para la mielina y de Nissl se usan mucho en neuropatología.
Métodos histioquímicos e inmunocitoquímicos son valiosos para localizar sustancias que se encuentran en poblaciones específicas de neuronas. Por supuesto que estas sustancias incluyen a los neurotransmisores (por ejemplo: norepinefrina, dopamina, serotonina y varios péptidos) y enzimas involucradas en la síntesis o degradación de neurotransmisores (ejemplos: dopamina, colina

acetiltransferasa y acetilcolinesterasa). Algunos sistemas de neuronas que no habían sido reconocidas fueron identificadas en animales de laboratorio, y resulta razonable presumir que también existen sistemas equivalentes en los humanos.
Microscopia electrónica. Muestra detalles más finos de la estructura interna de las neuronas y la especificación que existe en las uniones sinápticas. Sin embargo la necesidad de usar cortes ultra finos hace difícil reconstruir la neurona en tres dimensiones. La combinación de microscopia de luz y electrónica proporciona un panorama fácil de comprender la estructura neuronal y la neuroglia (Barr & Kiernan, 2001).
Organización funcional de la neurona
La neurona (célula nerviosa) es la unidad elemental de procesamiento y transmisión de la información en el sistema nervioso (Fig. 2).
Hay neuronas de diferentes formas y variedades, dependiendo del tipo de tarea especializada que llevan a cabo. De una forma u otra, generalmente tienen las cuatro estructuras o regiones siguientes: (1) cuerpo celular o soma; (2) dendritas; (3) axón; (4) botones terminales (Carlson, 2006).
Figura 2 Estructura del cuerpo neuronal y sus prolongaciones. Fuente: es.wikipedia.org

El cuerpo celular o soma varía extraordinariamente tanto en forma como en sus dimensiones. Puede ser redondeado, triangular, fusiforme, estrellado, piramidal, piriforme, etc. Por otra parte, hay neuronas que poseen un cuerpo muy pequeño, de unas cuantas micras; en cambio, otras tienen somas de 70 u 80 micras; entre ambos extremos pueden encontrarse numerosas dimensiones intermedias. En general el cuerpo de la neurona es muy semejante en su estructura al resto de las células del organismo. En la figura 2 se representa un esquema de una célula nerviosa con sus principales formaciones consideradas a nivel de la microscopia electrónica. Es conveniente consultar este esquema a medida que se vayan describiendo sus características morfológicas (López Antúnez, 1986).
Sustancia cromatofílica (sustancia de Nissl). Cuando el tejido nervioso es teñido con un colorante catiónico, como el violeta de cresilio o tionina, se observan grumos de material basófilo en la mayoría de las neuronas. (Fig. 3) Tales grumos fueron conocidos como los cuerpos de Nissl, en honor de Franz Nissl (1860-1919), un neurólogo de Heidelberg. La cantidad de sustancia cromatofílica (de Nissl) aumenta con el tamaño de la neurona, y su distribución varia de un tipo de neurona a otra. Por ejemplo los grumos de sustancia cromatofílica son toscos en la neurona motora, en tanto que el material basófilico o cromatofílico está distribuido más finamente en las neuronas sensoriales. La sustancia cromatofílica se extiende dentro de las prolongaciones dendrílicas de la neurona, pero no se encuentra en el colículo axónico, o sea, en la zona periférica del pericarión en donde el axón emerge pero tampoco se encuentra en al propio axón. La sustancia cromatofílica es la sustancia basófila, algunas veces llamada ergastoplasma o sustancia cromoidal, en el citoplasma de las células secretorias.

Figura 3 La sustancia de Nissl, son regiones basófilas (flechas rojas). Fuente: ht.org.ar
Por microscopia electrónica la sustancia cromatofílica se observa en ordenadas formaciones de retículo endoplásmatico granular. Esto constituye un sistema de cisternas aplanadas o vesículas a las que se adhieren partículas ribosómicas en la superficie exterior, y con polirribosomas en la matriz citoplasmática adyacente. Los ribosomas contienen ácido ribonucleico (RNA), lo que explica los basofilia; participan en la síntesis de las proteínas estructurales y enzimáticas, de ahí que se explica la abundancia de sustancia cromatofílica en las neuronas de mayores dimensiones que tienen más cantidad de citoplasma que mantiene a las prolongaciones largas. Se ha demostrado por diversos métodos, entre ellos la autorradiografía que se realiza después de marcar a los aminoácidos con sustancias radioactivas, ya que las proteínas de las neuronas se sintetizan predominantemente en el pericarión (Barr & Kiernan, 2001).
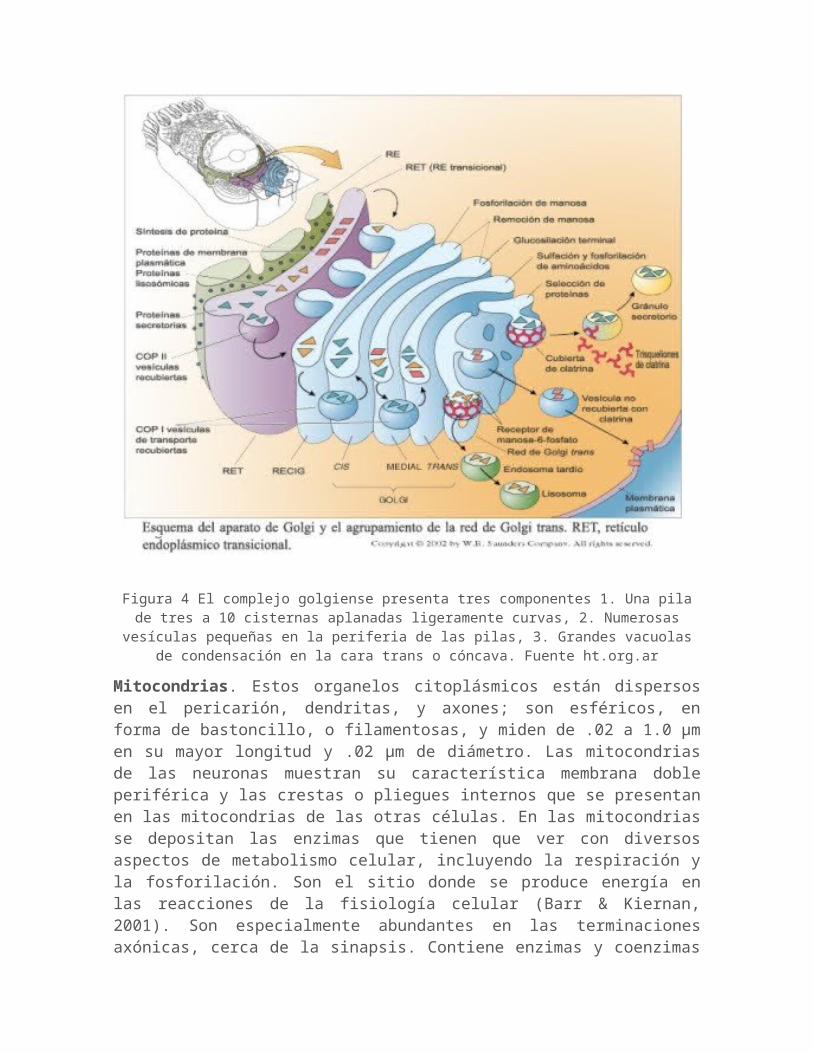
Complejo golgiense (aparato reticular interno o aparato de Golgi), retículo endoplásmatico liso, y lisomas. El complejo golgiense es un organelo universal del citoplasma de interés histórico especial en relación a las neuronas, ya que fue demostrado por primera vez en ellas por el histólogo italiano Camilo Golgi (1843-1926) Con tinciones para microscopía óptica, el complejo golgiense aparece como una red obscura e irregular, que está dispuesta frecuentemente alrededor del núcleo. El complejo golgiense aparece en las micrografías electrónicas como cúmulos de cisternas aplanadas, estrechamente, yuxtapuestas, las cuales se encuentran apiladas y rodeadas por muchas vesículas pequeñas. Este sistema de membranas es continuo con retículo endoplásmatico agranular o de superficie lisa, y este último es continuo con retículo endoplásmatico granular. La superficie del complejo golgiense es el área donde se adhieren los carbohidratos de algunas proteínas, que posteriormente se transforman en gluco-proteínas. Estas sustancias están rodeadas por diversos tipos de vesículas que se fijan a la membrana para ser transportadas en dirección distal a lo largo de las prolongaciones citoplasmáticas. Las vesículas sirven para renovar las vesículas sinápticas en los bulbos terminales de las terminaciones axónicas y también contribuyen la renovación de la membrana neuronal. Los lisomas, que derivan del retículo endoplásmatico liso y del aparato golgiense, son grandes vesículas fijas a las membranas y contienen enzimas que catalizan las descomposición de moléculas grandes no necesarias. En las neuronas normales, los lisosomas son más numerosos y conspicuos lo que no acontece en las células lesionadas o enfermas (Barr & Kiernan, 2001).

Figura 4 El complejo golgiense presenta tres componentes 1. Una pila de tres a 10 cisternas aplanadas ligeramente curvas, 2. Numerosas vesículas pequeñas en la
periferia de las pilas, 3. Grandes vacuolas de condensación en la cara trans o cóncava. Fuente ht.org.ar
Mitocondrias. Estos organelos citoplásmicos están dispersos en el pericarión, dendritas, y axones; son esféricos, en forma de bastoncillo, o filamentosas, y miden de .02 a 1.0 μm en su mayor longitud y .02 μm de diámetro. Las mitocondrias de las neuronas muestran su característica membrana doble periférica y las crestas o pliegues internos que se presentan en las mitocondrias de las otras células. En las mitocondrias se depositan las enzimas que tienen que ver con diversos aspectos de metabolismo celular, incluyendo la respiración y la fosforilación. Son el sitio donde se produce energía en las reacciones de la fisiología celular (Barr & Kiernan, 2001). Son especialmente abundantes en las terminaciones axónicas, cerca de la sinapsis. Contiene enzimas y coenzimas respiratorias, lo que indica su importante función en la producción y transporte energético de una célula que, como la neurona, tiene grandes exigencias metabólicas (López Antúnez, 1986).
Lisosomas. Son pequeños sacos que contienen enzimas hidrolíticas entre las que tiene particular importancia la fosfatasa ácida. Intervienen en los procesos de

digestión intracelular. Son muy abundantes en las neuronas. Actualmente se supone que del complejo de Golgi derivan vesículas que constituyen lisosomas primarios, los cuales transportan enzimas hidrolíticas a los lisosomas secundarios derivados de los cuerpos multivesiculares que están en relación con el retículo endoplásmatico liso (Novikoff, 1967 citado en López Antúnez, 1986). Los cuerpos multivesiculares son pequeñas estructuras rodeadas de membrana, que parecen ser portadores de vesículas. (López Antúnez, 1986).
Neurofibrillas, neurofilamentos, microtúbulos y microfilamentos. Cuando se emplean ciertas tinciones de plata, el citoplasma visto con microscopio fotónico contiene Neurofibrillas, algunas veces agrupadas en fascículos, que van a través del pericarión y penetran a las prolongaciones celulares (Barr & Kiernan, 2001). Su función no ha sido determinada aún, pero se ha supuesto que están en relación con el transporte de iones, con el mantenimiento de la forma de la neurona, y con las vesículas sinápticas, pero hace falta evidencia experimental que aclare su función (López Antúnez, 1986). En las micrografías electrónicas de las células nerviosas, el citoplasma contiene neurofilamentos, que miden de 7.5 a 10 nm de espesor. Los Neurofilamentos están constituidos por proteínas estructurales similares a las que forman los filamentos intermedios de otros tipos de células; probablemente son la causa de que las Neurofibrillas se observen así en la microscopía fotónica. El microscopio electrónico también muestra microtúbulos de 25 nm de diámetro externo, semejantes a aquellos de otros tipos de células, estos participan en la transportación rápida de las moléculas proteínicas y pequeñas particulas en ambas direcciones a lo largo de los axones y dendritas. Los microfilamentos (4 nm) están formados de moléculas de proteína actina que es contráctil. Se presentan en el interior del plasmalema y son particularmente numerosos en los extremos de los axones en crecimiento (Barr & Kiernan, 2001).
Pigmentos. El pericarión puede contener inclusiones citoplásmicas (depósitos de material inerte, a diferencia de los organelos), de las cuales las más conspicuas son los gránulos de pigmentos. El pigmento lipofucsina (lipocromo) se encuentra como grumos de gránulos de color pardo amarillento. Muestras de este pigmento aparecen en las neuronas de la médula espinal y el bulbo alrededor del octavo año de vida, y en las células de los ganglios espinales y simpáticos aproximadamente a la misma edad. La cantidad de pigmento aumenta con los años; el papel de la lipofucsina es desconocido, excepto que se relaciona con el proceso de envejecimiento y que se encuentra en otros sitios incluyendo los miocitos cardiacos. Algunos tipos de neuronas, por ejemplo, los neurocitos piriformes del cerebelo (células de Purkinje) localizados en la corteza, no acumulan lipofucsina, ni siquiera en la vejez (Barr & Kiernan, 2001).
Actualmente se considera que los gránulos de lipofucsina son un tipo de lisomas en que se ha almacenado el pigmento, ya que manifiestan actividad de fosfatasa ácida; se consideran cuerpos residuales. Issidorides y Shanklin (1961, citado en López Antúnez, 1986) han estudiado la distribución de la lipofucsina en diversos núcleos del sistema nervioso: oliva inferior, núcleo del XII, núcleo arcuato del bulbo, corteza cerebelosa, y núcleo dentado del cerebro, etc. Encontraron que en la oliva inferior los gránulos de lipofucsina están constituidos por capas

concéntricas de lípidos y proteínas y demostraron una alta concentración de cistina, sustancia que se encuentra también en las células neurosecretoras del hipotálamo, pero no en otros núcleos. Estos autores suponen una relación entre el material de Nissl y la lipofucsina y postulan que este pigmento podría ser un material portador de alguna sustancia producida por las neuronas, como el caso de la neuro-secreción del hipotálamo. En el cerebelo y en la propia oliva inferior, la lipofucsina se halla en los intrersticios entre las células, en áreas muy ricas en sinapsis. La significación funcional de esto se desconoce, pero la lipofucsina, según estos autores, no puede ser considerada como material de desecho de la neurona, sino posiblemente, como un producto de secreción (López Antúnez, 1986).
La presencia de pigmento oscuro de melanina en el citoplasma se restringe a unos pocos grupos celulares, las principales se encuentran en la sustancia negra del mesencéfalo y locus ceruleus (mancha azulosa o cerúlea del puente). El precursor metabólico de este pigmento es la dihidroxifenililalanina (DOPA), que se convierte a melanina por una serie de oxidaciones seguidas de polimerización. La DOPA es también precursora de la dopamina, un neurotransmisor usado por las neuronas pigmentosas de la sustancia negra y de la norepinefrina, que es el neurotransmisor de las células de locus ceruleus. La melanina puede acumularse como productos de desintegración de la síntesis de dopamina y norepinefrina. El pigmento de la sustancia negra aparece al final del primer año, aumenta en la pubertad y permanece después relativamente constante (Barr & Kiernan, 2001).
Foley y Baxter (1958, citado en López Antúnez, 1986), postulan que la melanina no se encuentra en los cerebros de animales jóvenes y que su acumulación aumenta con la edad. Marsden (1961, citado en López Antúnez, 1986), estudió la sustancia negra en cuarenta y nueve especies de mamíferos correspondientes a once órdenes y encontró que el pigmento se halla en una gran concentración de esos órdenes. Observó que la mayor pigmentación se presenta en los primates y que en ellos la riqueza de melanina aumenta a medida que si posición en la escala se aproxima a la del hombre, en la que la sustancia negra es desarrollada. Se ignora la significación funcional que esto pueda tener (López Antúnez, 1986).
Cilios y centriolos. En diferentes sitios del sistema nervioso se han descrito cilios y centriolos. Sotelo y Play (1968, citado en López Antúnez, 1986), observaron cilios en algunas células del núcleo vestibular de la rata y Del Cerro y Snider (1967, 1969, citado en López Antúnez, 1986) encontraron cilios y cuerpos basales en las células de Purkinje del cerebelo de ratas inmaduras y adultas. Los cilios presentan habitualmente la fórmula 9 X 10, es decir, nueve pares periféricos de microtúbulos sin que exista el par central, aunque en ocasiones la fórmula puede ser 8 X 1. Ambas disposiciones de los microtúbulos se consideran propias de los cilios que carecen de movimiento. A veces están relacionados con un centriolo que contiene 9 tripletes de microtúbulos. Se desconoce la significación funcional de los cilios en las células nerviosas (López Antúnez, 1986).
Gránulos neurosecretorios. En algunas células, por ejemplo en las de los núcleos supraóptico y paraventricular del hipotálamo, se encuentran en el
citoplasma gránulos de una sustancia d aspecto coloide, que parece ser un producto de secreción de la neurona, que posteriormente para a la neurohipófisis.
Núcleo. El núcleo de la neurona se halla comúnmente en posición central, aunque en algunas puede estar situado cerca de la membrana plasmática. En general, es grande, redondo u oval y de aspecto vesiculoso, pero su forma puede variar bastante. A veces ocupa gran parte del cuerpo celular. La neurona no tiene la capacidad para reproducirse; la cromatina se encuentra dispersa en el núcleo como ocurre en las células que están en interfase. En algunos casos se forman grumos en que el material cromatínico está acumulado, pero no se observan cromosomas. La membrana nuclear se continúa con el retículo endoplásmatico. Por lo general se ve un nucléolo, a veces dos, que se tiñen fuertemente con colorantes básicos. En el nucléolo se distinguen dos partes: el nucleolonema que tiene una estructura fibrilar o filamentosa, y la pars amorfa. Cerca del nucléolo frecuentemente unido a él, se halla un pequeño corpúsculo denso, el satélite nuclear o cuerpo de Barr que se supone contiene la cromatina sexual (cromosoma X). En el núcleo se halla el DNA de la neurona y el nucléolo es particularmente rico en RNA.
Dendritas. También llamadas prolongaciones citoplasmáticas (Véase Fig. 5). Parten del cuerpo celular por lo que su estructura es semejante a la del citoplasma: presentan retículo endoplásmatico rugoso y liso, mitocondrias, material de Nissl. Aparato de Golgi, neurofilamentos y microtúbulos, organelos que tienden a disponerse paralelamente al eje mayos de la dendrita. Un componente importante de las dendritas son las espinas o espículas, de forma y dimensiones variables, muy notables en algunos tipos de células, por ejemplo, en las de Purkinje del cerebelo y en las pirámides de la corteza cerebral donde puede haber varios miles de ellas por célula; en las espinas se establecen contactos sinápticos.
Las neuronas que poseen varias dendritas se llaman multipolares; ejemplo de estas son las del asta ventral de la medula espinal. Las que tienen solamente una dendrita se denominan bipolares, como las células que los ganglios del VIII nervio craneal o las bipolares de la retina.

Figura 5 Esquema 3D de un soma con sus dendritas. Observe como numerosos botones sinápticos de otras neuronas se ponen en contacto con el soma y las
dendritas. Fuente: ht.org.ar
Las dendritas son sitios en los que la célula nerviosa recibe impulsos; asimismo, el cuerpo celular constituye otra zona a la que llega información. Por esto el conjunto de dendritas y cuerpo celular representan el polo receptor de la neurona, al cual se le llama también área dendrítica.
Debido a que la célula nerviosa descarga los impulsos por el axón y sus colaterales, que constituyen so polo efector, se le considera polarizada dinámica. Esto significa que el impulso nervioso cursa en la neurona del área dendrítica hacia el axón y puede ser transmitido a otras neuronas a través de este y de sus colaterales. Es fácil comprender que la orientación de las dendritas posiblemente esté en relación con la fuente de los impulsos que recibe de la neurona. Dos ejemplos de esto son: las neuronas del asta ventral de la medula y las células de Purkinje del cerebelo. En la primera, las dendritas están orientadas en todas direcciones, ya que el área sináptica de estas células esta potencialmente abierta a impulsos que pueden llegar a ella desde diferentes sitios del espacio; en cambio las células de Purkinje poseen un verdadero penacho dendrítico que se orienta hacia la superficie de la corteza del cerebelo (capa molecular) que es por donde estas células reciben gran parte de su información.
El Axón. El axón es una prolongación generalmente única, que parte del cuerpo de la neurona, recorre un trayecto variable, emite colaterales y termina en arborizaciones finas llamadas telodentricas.
El axón y sus colaterales constituyen el polo efector de la neurona, a través del cual descarga impulsos nerviosos. Se originan en una elevación del cuerpo celular llamada cono axónico en el cual no existe material de Nissl. En la proximidad del cono axónico frecuentemente se establecen sinapsis inhibidoras.
Las fibras nerviosas se clasifican en 3 tipos, según Erlanger y Gasser. Sin embargo la velocidad e conducción no depende únicamente del espesor del axón, sino de su grado de mielinización y de otros factores.
Las terminaciones del axón y de sus colaterales pueden establecer las siguientes relaciones: a) con el área dendrítica de otras neuronas, para intentar integrar sinapsis axo-dendríticas o axo-somáticas, b) con otros axones con los que pueden construir sinapsis axo-axónicas., c) con fibras musculares estiradas, con las que forman las uniones neuro-musculares, d) con fibras musculares lisas o glándulas, como ocurre en las neuronas viscerales postganglionares.
Desde el punto de vista ultraesctructural, el axón posee axoplasma que es continuación del citoplasma del cuerpo neuronal, desde el cual parece haber una corriente constante hacia aquél. Se encuentran numerosas mitocondrias, particularmente cerca de las terminaciones axónicas que forman parte de la sinapsis. En el segmento inicial del axón existe retículo endoplásmatico rugoso, pero no en otras partes de la fibra donde no hay tampoco aparato de Golgi; se observan neurofilamentos y microtúbulos, siendo aquellos más abundantes que

los microtúbulos en las neuronas adultas; sin embargo a medida que los axones se ramifican disminuye progresivamente el número de neurofilamentos y en las más finas ramificaciones se ven solamente microtúbulos. No se conoce aún la significación funcional de estas estructuras, pero parecen corresponder a las neurofibrillas o axofibrillas que se observan cuando se usan tinciones de plata.
Las terminaciones axónicas frecuentemente se presentan como dilataciones bulbosas, los botones terminales que hacen contacto, por ejemplo, con el soma de otras neuronas en las sinapsis. Asimismo, desde el punto de vista funcional se puede considerar tres partes en el axón: el segmento inicial, comprendido entre el cono axónico y el primer nodo de Ranquiver, sitio en el que se origina el potencial de acción; la porción conductora, formada por la mayor parte del axón y sus colaterales, y la porción transmisora constituida por las terminaciones axónicas a través de las cuales el impulso nervioso se transmite a otras neuronas (a nivel de sinapsis), o a los efectores.

Figura 6 Principales estructuras del Axón. Fuente: neurocirugiadf.blogspot.mx
Vainas del axón. En el sistema nervioso central, los axones poseen vaina de mielina. En los nervios periféricos además de dicha vaina, las fibras están cubiertas por la vaina de Schwann o neurilema y por la vaina endoneural de Key Retzius (Fig. 6). Las fibras postganglionares del simpático, son llamadas amielínicas para indicar que carecen de vaina mielínica. Por otra parte, los cuerpos celulares de las neuronas aferentes periféricas de los ganglios espinales y craneales, así como los de las neuronas eferentes de los ganglios viscerales, están cubiertos por las llamadas células satélites, que los aíslan del medio extracelular.
Vaina de la mielina. La mielina es una sustancia lipoproteica que contiene cerebrósidos, fosfatidos, colesterol y esfingo-mielina. Está en contacto con el axón, pero presenta interrupciones situadas a intervalos regulares a lo largo del mismo, llamados nodos de Ranvier; en estos sitios se originan las colaterales del axón. Los nodos de Ranvier proporcionan a la vaina de mielina una disposición segmentaria a lo largo de la fibra nerviosa. El espacio comprendido entre nodo y nodo se llama segmento intermodal.
Fibras amielínicas. Esta denominación indica que carecen de mielina. Un ejemplo lo constituyen los axones de las neuronas de los ganglios viscerales que van a inervar músculo liso, musculo cardiaco o flándulas.
Secuencia de la mielinización. La formación de la mielina se inicia poco antes de la vida intrauterina, pero continua hasta después del nacimiento y hay datos que indican que la vaina mielínica aumenta en grosor desde la infancia hasta la edad madura. Los haces espinotalánicos y espinocerebelosos, por ejemplo presentan ya la mielinización hacia el principio del cuarto mes de vida intrauterina, mientras que los haces córticoespinales comienzan a adquirir su mielina hacia el final de la gestación, pero el proceso tarda varios años en completarse. En un sistema de fibras determinado, la mielinización parece estar en relación con la secuencia en que aparecen los axones en dicho sistema. Existen datos que hacen posible afirmar que la capacidad funcional óptima de un sistema neuronal no se alcanza sino hasta que se ha completado su mielinización. En este sentido hay en la patología neurológica un grupo de padecimientos que se ha caracterizado por la alteración, degradación y pérdida de la mielina: son las llamadas enfermedades desmielinizantes, cuya etología se desconoce.
Mielinización. Aunque el origen de la mielina central no está perfectamente aclarado, hay suficiente evidencia experimental para suponer que en este caso intervienen los oligodendrocitos interfasciculares, que se comportarían respecto a las fibras centrales de manera semejante a como lo hacen las células de Schwann en las periféricas; es decir, arrollándose su membrana en torno a los axones, con la diferencia de que, mientras la célula de Schwann forma mielina en un solo axón, los oligodendrocitos lo hacen en un número variable de fibras, quedando en ocasiones el cuerpo de la célula glíal unido a los axones por puentes de citoplásmicos (López Antúnez, 1986).

Los componentes de la glía son la neuroglia y microglía.
La neuroglia puede dividirse en central, constituida por células que se hallan en los centros nerviosos y periféricos, que tienen relación con las neuronas situadas fuera del sistema nervioso central. La neuroglia central está formada por 3 tipos de células: células ependimarias, astrocitos (astroglía) y oligodendrocitos (oligodendroglía). La neuroglia periférica está constituida por: células satélites o capsulares y células de Schwann. Toda neuroglia es de origen ectodérmico: la célula proviene del neuroepitelio del tubo neural y la periférica del ectodermo de la cresta neutral.
La microglía (llamada también mesoglía), es de origen mesodérmico, originándose de la pared de los vasos sanguíneos.
Origen de la glía. La microglía proviene del mesodermo. Río Hortega señalo zonas a las que llamó fuentes de microglía en las cuales se originan estas células: el endotelio de los vasos sanguíneos, las leptomeninges y los plexos coroideos. A partir de estas estructuras se produce migración de células que invaden el sistema nervioso en desarrollo. A favor de este concepto esta el hecho de que la microglía aparece cuando existe la trama vascular de los centros nerviosos en los cuales representaría al sistema retículo endotelial.
Algunos autores piensan que tiene un origen común con el resto de las células de la neuroglia, a partir de la capa subependimaria. Estudios de radio autográficos usando timidina triada inyectada intraventricularmente en ratas (Lewis, 1968 citado en López Antúnez, 1986), han demostrado que en la capa subependimaria a nivel de los ventrículos laterales y en menor proporción, del tercero, hay actividad de síntesis de DNA después del nacimiento e incluso en animales adultos y que a partir de ella pueden derivarse elementos gliales incluyendo la microglía, cuyo origen en las fuentes de Rio Hortega, se pone en duda. Esto plantearía el origen de la glía.
Características estructurales y relaciones de la glía.
Astrocitos. Son células de aspecto trillado. Se consideran dos tipos principales: fibrosos y citoplasmáticos. Los primero se hallan principalmente en la sustancia blanca; los segundos en la gris. Los fibrosos tienen prolongaciones finas; los citoplasmáticos, gruesas y nudosas muy ramificadas a partir del cuerpo celular.
La microscopia electrónica ha demostrado que los astrocitos también tienen relación con la pía, con la cual forman una barrera liquido encefálica y además, con las neuronas, en las que constituyen la barrera sináptica.
Desde el punto de vista estructural, el citoplasma de los astrocitos, de aspecto pálido, contiene abundante retículo endoplásmatico rugoso, aparato de Golgi, mitocondrias, gliosomas y gliofibrillas. Mori y Lablond (1969, citado en López Antúnez, 1986), estudiaron los astrocitos del cuerpo calloso de las rata mediante la técnica de cloruro de oro sublimado de Cajal, y consideran que las principales características de los astrocitos son: la existencia de haces de microfilamentos que corresponden a las gliofibrillas que se observan con el microscopio de luz; los

gliosomas, cuerpos densos, redondeados, que se supone son de origen lisosomal y la presencia de gránulos de glucógeno en el citoplasma. Según estos investigadores, los astrocitos son capaces de reproducirse en animales y no solamente en cultivo de tejidos. Finalmente suponen que pueden sufrir degeneraciones en condiciones normales, siendo contrarrestada esta situación por su capacidad de dividirse.
Los astrocitos son las únicas células neuróglicas que presentan esta estructura fibrilar (Fig. 7). Estudiando los procesos de maduración de los astrocitos fibrosos en el medio óptico de la rata, Vaughn y Peters (1967, citado en López Antúnez, 1986), han observado que en las células inmaduras hay gran abundancia de microtúbulos y son escasos los filamentos; pero a medida que los astrocitos se aproximan a su estado adulto, esta relación se invierte, de modo de que en las células maduras hay gran numero de filamentos y solo ocasionalmente se ven microtúbulos. Por otra parte, Rodríguez, Echandia, Piezzi y Rodríguez (1968, citado en López Antúnez, 1986), han hecho notar que los microtúbulos son un componente constantemente de las neuronas y de algunas células gliales y además han observado en su interior gránulos cuya significación es oscura, suponiendo estos investigadores que pueden corresponder a material que fluye en ellos. Parece que la afinidad que los astrocitos tienen por el cloruro de oro sublimado de Cajal, que constituye un método de tinción clásico para estas células, se debe a los abundantes filamentos que tienen sus prolongaciones.
Figura 7 Estructura del astrocito, vaso sanguíneo y la neurona. Fuente: voxpopulidelaciencia.blogspot.mx
Oligodendrocitos. Son células de cuerpo pequeño, con escaso citoplasma y prolongaciones finas y poco numerosas que emergen del cuerpo celular orientándose en diferentes direcciones y acodándose en ángulos pronunciados, abundan en la sustancia gris (Fig. 8). Según su situación, se clasifican en perineuronales o satélites, e interfasciculares. Los primeros se encuentran en

relación con el cuerpo de la neurona, al que tienden a rodear formándole in revestimiento que sugiere la disposición de las células satélites en las neuronas ganglionares. Los interfasciculares establecen relación con las fibras nerviosas de la sustancia blanca.
Figura 8 Oligodendrocito y las vainas de mielina. Fuente: guiasdeneuro.com.ar
Según Mori u Lblond (1970, citado en López Antúnez, 1986), las características estructurales de los oligodendrotocitos son: citoplasma rico en ribosomas y microtúbulos, pero gliofibrillas ni gránulos de glucógeno. Su núcleo es regular y presenta numerosos poros en su membrana con la cromatina condensada cerca de ellos. Estos autores proponen 3 tipos de oligodendrocitos de acuerdo con la densidad del núcleo y citoplasma; claros, opacos y oscuros. Los claros presentan figuras de mitosis más frecuentemente que las demás variedades. En la medula espinal de los gatos pequeños, Bunge, Bunge y Pappas (1962, citado en López Antúnez, 1986), demostraron la existencia de conexiones entre la glía (probablemente oligodendrocitos) y la vaina de mielina en el sistema nervioso en

desarrollo. Puede haber cierta distancia entre el soma de la célula glial y la vaina mielínica, estando unidas dichas estructuras por los puntes citoplasmáticos formados por las prolongaciones gliales.
Células satélites. Rodean a los cuerpos de las neuronas de los ganglios espinales, craneales y viscerales, formado una verdadera capsula, por lo que también se les llama capsulares. Suelen formar en los cuerpos en los cuerpos neuronales un revestimiento un revestimiento constituido por la superposición de varias capas de membrana plegada (Pineda, Maxwell y Kruger, 1967 citado en López Antúnez, 1986), disposición que recuerda a la vaina de la mielina de las fibras periféricas. La superficie externa de las células satélites está cubierta por membrana basal que se continúa de una célula a otra y reviste a la célula de Schwann del primer internodo. A veces las células satélites se prolongan sobre el segmento inicial del axón.
Células de Schwann. Están situadas en relación con las fibras nerviosas periféricas y terminan formando digitaciones a nivel de los nodos de Ranvier. Están revestidas exteriormente de membrana basal. Existe una sola célula de Schwann en cada segmento internodal.
Microglía. Son células de cuerpo pequeño, con prolongaciones finas que emiten colaterales que se desprenden en ángulo recto de las ramas principales. Mori y Leblond (1969, citado en López Antúnez, 1986), mediante la técnica del carbonato de plata débil de Río-Hortega, identifican 2 tipos de microglía: pericital, que se halla cubierta por una membrana basal de los capilares, e intersticial, que se encuentra diseminada en el tejido nervioso. Tienen núcleo pequeño con masas densas de cromatina y núcleoplasma claro; el nucléolo es muy visible. Su escaso citoplasma forma un anillo en torno al núcleo. Una característica ultraesctructural de la microglía es la existencia de cuerpos densos en el citoplasma rodeados de un halo de lisosomas, lo que explicaría su capacidad fagocitaria; el retículo endoplásmico es escaso. No presenta fibrillas ni gránulos de glucógeno.
Reproducción de la glía. Hommes y Leblond (1967, citado en López Antúnez, 1986), mediante técnicas de marcaje con timidina en ratas adultas normales, notaron que el isotopo se hallaba en el núcleo de células gliales y además identificaron figuras mitóticas en células que suponen pueden ser oligodendrocitos o espongioblastos precursores de estos; también se observo este fenómeno en células posiblemente micróglicas, aunque Mori y Leblond (1969, citado en López Antúnez, 1986) marcando células de este último tipo, concluyen que no se reproducen, salvo en condiciones de emergencia.
Funciones de la glía. La mayor parte de sus posibles actividades son aún motivo de investigación y en consecuencia, esperan comprobación experimental, existen datos suficientes que sugieren la participación de la glía en diversas funciones, todas de gran importancia. Por otra parte, la correlación que comienza a establecerse entre las modificaciones morfo funcionales de la glía y las alteraciones que se observan en ciertos estados patológicos, hacen posible comprender algunos aspectos de la patogenia de padecimientos neurológicos a los que el médico se enfrenta frecuentemente. En esta forma ha surgido el

concepto de que las células nerviosas y el tejido glial constituyen una verdadera unidad que interviene tanto en la regulación de actividades metabólicas como, probablemente, en los fenómenos mismos de la electrogénesis neural.
Metabolismo. Con el microscopio se afirmó la idea de que probablemente representan una vía por la cual los iones y metabolitos procedentes de la sangre, llegan hasta la neurona, cuyos productos de desecho pueden, a su vez, ser llevados a los capilares o al líquido cefalorraquídeo. Se planteó así mismo la probable participación de la glía en los fenómenos que ocurren a nivel de la sinapsis. Roberts y Gerschenfeld señalan la participación de la glía en la formación de las barredas hemato-encefálica, liquido-encefálica, y sináptica. Por otra parte, estudios sobre el metabolismo de la neurona y la glía, llevados a cabo por numerosos investigadores, entre los que hay que destacar a Hydén y colaboradores (1967, citado en López Antúnez, 1986), han demostrado una relación estrecha entre las síntesis de RNA y las actividades enzimáticas de la cadena respiratoria en ambos tipos de células. En condiciones de estimulación, hay aumento tanto en la síntesis de RNA como de enzimas respiratorias en la neurona, pero no en la glía, en donde se ha observado un incremento de la glicólisis anaeróbica; Hydén interpreta esto en el sentido de que la neurona tiene prioridad para obtener la energía disponible de la cadena respiratoria cuando las demandas funcionales aumentan, mientras a través de la glicólisis anaeróbica. Piensa que, en consecuencia, la neurona y la glía están acopladas en un sistema que satisface las exigencias metabólicas del tejido nervioso. Hydén y Lange (1962, citado en López Antúnez, 1986), estudiando la actividad de una enzima respiratoria, la succinocidasa, en células del núcleo de Deiters de gatos sometidos a estimulación vestibular, encontraron que mientras la neurona reacciona aumentando la capacidad del sistema de transporte de electrones, lo que indica un aumento en el consumo de energía bajo efecto de la estimulación, la glía por el contrario, no manifiesta cambios al respecto, lo cual está de acuerdo con la diferencia de velocidad de las actividades eléctricas en los dos tipos de células: los potenciales de las células gliales son muy lentos comparados con los de las neuronas.
Las células satélites parecen tener una relación estrecha con la actividad metabólica de las neuronas ganglionares. Schwyn (1967, citado en López Antúnez, 1986), mediante marcaje con timidina tritiada y aumentando la actividad sináptica por estimulación de las fibras preganglionares que llegan al ganglio cervical superior del gato, observo que el marcador se incorpora al núcleo de las células satélites, lo cual podría indicar actividad de síntesis de DNA condicionada al aumento de la actividad neuronal debido a la estimulación. Dixon (1969, citado en López Antúnez, 1986), estudiando las células satelitales del ganglio cervical superior del conejo después de producir cromatólisis de las neuronas ganglionares seccionando sus axones, notó en ellas importantes modificaciones: numerosas vesículas de pinocitosis, aumentando del retículo endoplásmatico y de la longitud de las mitocondrias, y abundantes cisternas bajo la membrana celular, así como la aparición de numerosos microfilamentos. Interpreta estos cambios como resultado del aumento en el transporte de materiales del medio extracelular hacia la neurona a través de la célula satélite, así como de una mayor actividad y de síntesis de

proteínas; también se observó que cuando el cuerpo neuronal aumenta de tamaño correlativamente a los fenómenos de cromatólisis, las células satelitales se ajustan al mayor volumen de la neurona y forman una sola capa en torno a ella. El aumento de los microfilamentos puede interpretarse como una adaptación del citoesqueleto de la célula satélite a las nuevas condiciones derivadas de la modificación en el tamaño de la neurona, ya que desaparece cuando esta recobra su volumen normal. Lasek y Embree (1969, citado en López Antúnez, 1986), después de seccionar las fibras preganglionares del ganglio cervical superior de la rata, observaron aumento del DNA y del RNA, persistiendo por mucho mayor tiempo el primero, lo que interpreta como actividad de las células gliales del ganglio, mientras que los cambios en el RNA los relacionan con fenómenos metabólicos de las neuronas.
Aspectos funcionales de la glía. Se especula acerca del papel que las células gliales podrían desempeñar en la adquisición de la posición definitiva que las neuronas ocuparán en el sistema nervioso. En este sentido son interesantes los trabajos de Rakic (1971, citado en López Antúnez, 1986) que al estudiar la diferenciación y migración de las células granulosas de la corteza cerebelosa del macaco Rhesus, supone que dichos fenómenos son favorecidos por las fibras de Bergmann, pues se observa una relación constante entre la célula granulosa y la fibra glial, dando la impresión de que la segunda orienta la migración de la neurona, ya que esta desciende siempre adosada de la fibra glial, dando la impresión de que la segunda orienta la migración de que de la neurona, ya que esta desciende siempre adosada a la fibra de Bergmann y simultáneamente va diferenciándose; su axón se alarga y posteriormente se divide en T en la capa molecular. Es probable, que la función de la fibra de Bergmann no sea exclusivamente mecánica, sino también metabólica.
Para terminar con la glía téngase presente que los tumores malignos del sistema nervioso son de origen glial. Los gliomas se clasifican incluso según los tipos celulares que los forman, por ejemplo: astrocitomas, oligodendrogliomas, ependimomas, cuando predominan la variedad celular correspondiente; si existen diferentes tipos de células se describen varios gliomas, entre ellos, el glioblastoma multiforme, que probablemente sea el tumor cerebral más frecuente.
Glía y sinapsis. La posible participación de la glía en la función de la sinapsis se discute plenamente en un terreno puramente especulativo. Desde el punto de vista morfológico, se ha demostrado la existencia de prolongaciones gliales que llegan a los lados del espacio sináptico y en ocasiones se han visto complejos de unión entre dichas prolongaciones y los botones sinápticos (Gonzales de Aguilar y De Robertis, 1963 citado en López Antúnez, 1986), lo que constituye la base morfológica de la barrera sináptica, como ha sido propuesta por De Robertis.
La situación de las células gliales cerrando los lados del espacio sináptico, ha sido interpretada como una disposición que evita que el mediador se difunda fuera de dicho espacio. Algunos autores han emitido la hipótesis de que la glía contigua a la sinapsis probablemente contiene colinesterasa (Koelle, 1962; Fredricsson y Sjoquist, 1952 citado en López Antúnez, 1986).

Glía y electrogénesis. Vallecalle y Svaetichin como resultado de estudios hechos en la retina de ciertas variedades de peces, cuyos elementos gliales son suficientemente grandes de modo que es posible introducir electrodos intracelulares, suponen que además de los fenómenos de excitabilidad de la membrana, de conducción rápida de impulsos y de transmisión sináptica característicos de la actividad neuronal. La glía es capaz de mantener potenciales estables y actuar como elemento controlador de la función de los receptores y de las neuronas. Postulan que la organización de la visión cromática en los peces, depende de dicha interacción entre los elementos no neurales (gliales) y los elementos receptores y conductores. El control de la excitabilidad retiniana se haría a través de un mecanismo de retroalimentación estabilizador entre el elemento de entrada: el receptor y el elemento conductor: la neurona. Una característica de los elementos controladores es que su membrana es eléctricamente inexcitable, dependiendo de su acción de la difusión de la actividad a nivel molecular, entre las zonas de contacto de la membrana.
Neurona- glía y aprendizaje. Una de las hipótesis de trabajo más interesantes ha sido propuesta por Hydén y Clos. Este investigador hace notar que una de las características funcionales de la unidad metabólica neurona-glía, es la intensa actividad de síntesis de macromoléculas, especialmente de RNA, en cuya formación ninguna otra célula somática puede competir con la neurona. Hydén ha observado que en condiciones de estimulación, el contenido en enzimas respiratorias y en RNA aumenta en la neurona y disminuye en la glía, en la que, como se indicó previamente, se incrementa la glicolisis anaeróbica. En cambio, en ratas sometidas a determinados procesos de aprendizaje, el contenido de RNA de tipo particular, rico en adenina y pobre en citosina. Concluye que este tipo de RNA, que se sintetiza durante el proceso de aprendizaje, es cromosomal y que, en consecuencia, es posible que los factores ambientales, en este caso la situación de aprendizaje a que es sometido el animal, induzcan una estimulación del genoma de la glía y de las neuronas implicadas. Plantea la posibilidad de que pueda transferencia de RNA entre la neurona y la glía, lo cual sugeriría la participación de esta última conjuntamente con la neurona en los procesos de almacenamiento de memoria. Esto dependería, según este autor, de cambios a nivel macromolecular, expresados en diferentes tipos de RNA mensajero, que condicionaría la síntesis de proteínas diferentes (López Antúnez, 1986).
Bibliografía
1.Barr, M. L., & Kiernan, J. A. (2001). El sistema Nervioso Humano: un punto de vista anatómico. México: Mc Graw Hill Interamericana.
2.Carlson, N. R. (2006). Fisiología de la conducta. España: Pearson.
3.López Antúnez, L. (1986). Anatomía Funcional del Sistema Nervioso. México: Limusa.