Uncovering BRD4 hyperphosphorylation associated with ...Significance BRD4 plays a vital role in...
10
Uncovering BRD4 hyperphosphorylation associated with cellular transformation in NUT midline carcinoma Ranran Wang a , Xing-Jun Cao b , Katarzyna Kulej b , Wei Liu a , Tongcui Ma a , Margo MacDonald a , Cheng-Ming Chiang c,d,e , Benjamin A. Garcia b , and Jianxin You a,1 a Department of Microbiology, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA 19104; b Epigenetics Program, Department of Biochemistry and Biophysics, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA 19104; c Simmons Comprehensive Cancer Center, University of Texas Southwestern Medical Center, Dallas, TX 75390; d Department of Biochemistry, University of Texas Southwestern Medical Center, Dallas, TX 75390; and e Department of Pharmacology, University of Texas Southwestern Medical Center, Dallas, TX 75390 Edited by Peter M. Howley, Harvard Medical School, Boston, MA, and approved May 24, 2017 (received for review February 24, 2017) The epigenetic reader BRD4 plays a vital role in transcriptional regulation, cellular growth control, and cell-cycle progression. Dysregulation of BRD4 function has been implicated in the pathogenesis of a wide range of cancers. However, how BRD4 is regulated to maintain its normal function in healthy cells and how alteration of this process leads to cancer remain poorly under- stood. In this study, we discovered that BRD4 is hyperphosphory- lated in NUT midline carcinoma and identified CDK9 as a potential kinase mediating BRD4 hyperphosphorylation. Disruption of BRD4 hyperphosphorylation using both chemical and molecular inhibitors led to the repression of BRD4 downstream oncogenes and abrogation of cellular transformation. BRD4 hyperphosphor- ylation is also observed in other cancers displaying enhanced BRD4 oncogenic activity. Our study revealed a mechanism that may regulate BRD4 biological function through phosphorylation, which, when dysregulated, could lead to oncogenesis. Our finding points to strategies to target the aberrant BRD4 signaling specifi- cally for cancer intervention. bromodomain-containing protein 4 | NUT midline carcinoma | CDK9 | cellular transformation | cancer B RD4 (bromodomain-containing protein 4) is a member of the bromodomain and extraterminal (BET) family. The tandem bromodomains (BD1 and BD2) of BRD4 specifically recognize acetylated histones H3 and H4 on chromatin (1). Through this interaction, BRD4 actively recruits P-TEFb (posi- tive transcription elongation factor b), mediators and other transcriptional activators to facilitate gene activation (2–4). P-TEFb is a heterodimer of CDK9 and cyclin T1, which together phosphorylate serine 5 in the C-terminal domain of RNA poly- merase II (RNAPII) to support transcription elongation (5–7). Therefore, BRD4 functions as an epigenetic reader that plays a central role in transcriptional regulation, cellular growth control, and cell-cycle progression (2, 8, 9). Dysregulation of BRD4 function has been implicated in the path- ogenesis of a wide range of cancers including acute myeloid leukemia (AML), multiple myeloma, Burkitt’ s lymphoma, diffuse large B-cell lymphoma, breast cancer, colon cancer, and ovarian carcinoma (4, 10– 14). It is also the target of a genetic translocation between chromo- somes 15 and 19, denoted as t (15;19), which results in the formation of a novel fusion oncogene BRD4-NUT that accounts for the highly lethal NUT (nuclear protein in testis) midline carcinoma (NMC) (15). Although BRD4 has emerged as a critical therapeutic target for a wide variety of cancers (4, 10–14), the mechanisms that regulate BRD4 function have not been clearly elucidated. How alteration of BRD4 function leads to cancer development re- mains largely unknown. BRD4 has been shown to enrich dis- proportionately at a subset of key oncogenic and lineage-specific genes such as c-MYC and selectively stimulates their expression to drive cellular proliferation in cancers (4, 14, 16–18). Blocking bromodomain binding to acetylated histones with BET inhibitors, including (+)-JQ1 (19) and I-BET (20), specifically down-regulate these oncogenes (4, 16, 17). Addiction of the tumor cells to high- level expression of these oncogenes provides the basis for using BET inhibitors to abrogate BRD4 function for treating these cancers (4, 14, 16, 17). Multiple BET inhibitors have thus entered clinical trials. Early clinical trials have shown promising results, especially for hematological malignancies (21), highlighting the potential of targeting BRD4 in anticancer treatment. However, resistance to BET inhibitors has also emerged (22, 23), revealing the therapeutic limitations of BET inhibitors and the complexity of BRD4 regulation mechanisms. More importantly, we and others have shown that BRD4 also plays an important role in noncan- cerous systems such as mouse embryonic stem cells, preimplanta- tion embryos, and keratinocyte differentiation (9, 24–26). There are growing concerns regarding the consequences of disrupting BRD4 function in the normal cells by using BET inhibitors (27). Therefore, it is critical to elucidate the molecular mechanisms that regulate BRD4’s biological function in both normal and dis- ease settings so that therapeutic interventions can be developed to switch off the oncogenic activity of BRD4 specifically in cancer cells while sparing the normal BRD4 function in healthy cells. NMC is a highly lethal tumor typically caused by translocation t(15;19). The translocation breakpoint splits BRD4 in half, resulting in the in-frame fusion of BRD4 bromodomains and extraterminal domain with nearly the entire sequence of the NUT gene (15, 28). NMCs represent the most lethal subset of squamous cell carcinomas (15). They metastasize rapidly and are extremely aggressive; patients have a median survival of <7 mo (15). Translocation t(15;19), which causes the formation of the Significance BRD4 plays a vital role in cellular growth control. Because BRD4 is dysregulated in a wide range of aggressive malig- nancies, it is being increasingly implicated as a major driver of oncogenic growth and a novel target for cancer therapy. However, how BRD4 is regulated to maintain its normal func- tion in healthy cells and how alteration of this process leads to cancer remain poorly understood. We discovered that BRD4 is hyperphosphorylated in cancers and that this hyperphos- phorylation may be a general mechanism to support its onco- genic activities. Our study shows how dysregulation of BRD4 function could lead to tumorigenesis. Our discovery also pro- vides the rationale for investigating how cellular signaling pathways modulate BRD4 phosphorylation to control its func- tion during cancer development. Author contributions: R.W., X.-J.C., B.A.G., and J.Y. designed research; R.W., X.-J.C., W.L., T.M., and M.M. performed research; C.-M.C. and B.A.G. contributed new reagents/analytic tools; R.W., X.-J.C., K.K., and J.Y. analyzed data; and R.W. and J.Y. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. 1 To whom correspondence should be addressed. Email: [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1703071114/-/DCSupplemental. E5352–E5361 | PNAS | Published online June 19, 2017 www.pnas.org/cgi/doi/10.1073/pnas.1703071114 Downloaded by guest on August 27, 2020
Transcript of Uncovering BRD4 hyperphosphorylation associated with ...Significance BRD4 plays a vital role in...

Uncovering BRD4 hyperphosphorylation associatedwith cellular transformation in NUT midline carcinomaRanran Wanga, Xing-Jun Caob, Katarzyna Kulejb, Wei Liua, Tongcui Maa, Margo MacDonalda, Cheng-Ming Chiangc,d,e,Benjamin A. Garciab, and Jianxin Youa,1
aDepartment of Microbiology, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA 19104; bEpigenetics Program, Department ofBiochemistry and Biophysics, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA 19104; cSimmons Comprehensive Cancer Center,University of Texas Southwestern Medical Center, Dallas, TX 75390; dDepartment of Biochemistry, University of Texas Southwestern Medical Center, Dallas,TX 75390; and eDepartment of Pharmacology, University of Texas Southwestern Medical Center, Dallas, TX 75390
Edited by Peter M. Howley, Harvard Medical School, Boston, MA, and approved May 24, 2017 (received for review February 24, 2017)
The epigenetic reader BRD4 plays a vital role in transcriptionalregulation, cellular growth control, and cell-cycle progression.Dysregulation of BRD4 function has been implicated in thepathogenesis of a wide range of cancers. However, how BRD4 isregulated to maintain its normal function in healthy cells and howalteration of this process leads to cancer remain poorly under-stood. In this study, we discovered that BRD4 is hyperphosphory-lated in NUT midline carcinoma and identified CDK9 as a potentialkinase mediating BRD4 hyperphosphorylation. Disruption ofBRD4 hyperphosphorylation using both chemical and molecularinhibitors led to the repression of BRD4 downstream oncogenesand abrogation of cellular transformation. BRD4 hyperphosphor-ylation is also observed in other cancers displaying enhancedBRD4 oncogenic activity. Our study revealed a mechanism thatmay regulate BRD4 biological function through phosphorylation,which, when dysregulated, could lead to oncogenesis. Our findingpoints to strategies to target the aberrant BRD4 signaling specifi-cally for cancer intervention.
bromodomain-containing protein 4 | NUT midline carcinoma | CDK9 |cellular transformation | cancer
BRD4 (bromodomain-containing protein 4) is a member ofthe bromodomain and extraterminal (BET) family. The
tandem bromodomains (BD1 and BD2) of BRD4 specificallyrecognize acetylated histones H3 and H4 on chromatin (1).Through this interaction, BRD4 actively recruits P-TEFb (posi-tive transcription elongation factor b), mediators and othertranscriptional activators to facilitate gene activation (2–4).P-TEFb is a heterodimer of CDK9 and cyclin T1, which togetherphosphorylate serine 5 in the C-terminal domain of RNA poly-merase II (RNAPII) to support transcription elongation (5–7).Therefore, BRD4 functions as an epigenetic reader that plays acentral role in transcriptional regulation, cellular growth control,and cell-cycle progression (2, 8, 9).Dysregulation of BRD4 function has been implicated in the path-
ogenesis of a wide range of cancers including acute myeloid leukemia(AML), multiple myeloma, Burkitt’s lymphoma, diffuse large B-celllymphoma, breast cancer, colon cancer, and ovarian carcinoma (4, 10–14). It is also the target of a genetic translocation between chromo-somes 15 and 19, denoted as t(15;19), which results in the formationof a novel fusion oncogene BRD4-NUT that accounts for the highlylethal NUT (nuclear protein in testis) midline carcinoma (NMC) (15).Although BRD4 has emerged as a critical therapeutic target
for a wide variety of cancers (4, 10–14), the mechanisms thatregulate BRD4 function have not been clearly elucidated. Howalteration of BRD4 function leads to cancer development re-mains largely unknown. BRD4 has been shown to enrich dis-proportionately at a subset of key oncogenic and lineage-specificgenes such as c-MYC and selectively stimulates their expressionto drive cellular proliferation in cancers (4, 14, 16–18). Blockingbromodomain binding to acetylated histones with BET inhibitors,including (+)-JQ1 (19) and I-BET (20), specifically down-regulate
these oncogenes (4, 16, 17). Addiction of the tumor cells to high-level expression of these oncogenes provides the basis for usingBET inhibitors to abrogate BRD4 function for treating thesecancers (4, 14, 16, 17). Multiple BET inhibitors have thus enteredclinical trials. Early clinical trials have shown promising results,especially for hematological malignancies (21), highlighting thepotential of targeting BRD4 in anticancer treatment. However,resistance to BET inhibitors has also emerged (22, 23), revealing thetherapeutic limitations of BET inhibitors and the complexity ofBRD4 regulation mechanisms. More importantly, we and othershave shown that BRD4 also plays an important role in noncan-cerous systems such as mouse embryonic stem cells, preimplanta-tion embryos, and keratinocyte differentiation (9, 24–26). There aregrowing concerns regarding the consequences of disruptingBRD4 function in the normal cells by using BET inhibitors (27).Therefore, it is critical to elucidate the molecular mechanisms thatregulate BRD4’s biological function in both normal and dis-ease settings so that therapeutic interventions can be developed toswitch off the oncogenic activity of BRD4 specifically in cancer cellswhile sparing the normal BRD4 function in healthy cells.NMC is a highly lethal tumor typically caused by translocation
t(15;19). The translocation breakpoint splits BRD4 in half,resulting in the in-frame fusion of BRD4 bromodomains andextraterminal domain with nearly the entire sequence of theNUT gene (15, 28). NMCs represent the most lethal subset ofsquamous cell carcinomas (15). They metastasize rapidly and areextremely aggressive; patients have a median survival of <7 mo(15). Translocation t(15;19), which causes the formation of the
Significance
BRD4 plays a vital role in cellular growth control. BecauseBRD4 is dysregulated in a wide range of aggressive malig-nancies, it is being increasingly implicated as a major driver ofoncogenic growth and a novel target for cancer therapy.However, how BRD4 is regulated to maintain its normal func-tion in healthy cells and how alteration of this process leads tocancer remain poorly understood. We discovered that BRD4 ishyperphosphorylated in cancers and that this hyperphos-phorylation may be a general mechanism to support its onco-genic activities. Our study shows how dysregulation of BRD4function could lead to tumorigenesis. Our discovery also pro-vides the rationale for investigating how cellular signalingpathways modulate BRD4 phosphorylation to control its func-tion during cancer development.
Author contributions: R.W., X.-J.C., B.A.G., and J.Y. designed research; R.W., X.-J.C., W.L.,T.M., and M.M. performed research; C.-M.C. and B.A.G. contributed new reagents/analytictools; R.W., X.-J.C., K.K., and J.Y. analyzed data; and R.W. and J.Y. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1703071114/-/DCSupplemental.
E5352–E5361 | PNAS | Published online June 19, 2017 www.pnas.org/cgi/doi/10.1073/pnas.1703071114
Dow
nloa
ded
by g
uest
on
Aug
ust 2
7, 2
020

BRD4-NUT fusion oncogene in NMC (15), has been described inpediatric head and neck tumors as well as in lung cancers (29).All NMCs carry an intact BRD4 locus and simultaneously ex-press BRD4 and the BRD4-NUT fusion oncogene (30, 31), pro-viding a unique tumor model to investigate how alteration ofBRD4 function by oncogenic mutation leads to cancer. TheBRD4-NUT fusion oncoprotein is also tethered to acetylatedchromatin by the bromodomains (31, 32). It causes malignancyby blocking NMC differentiation while driving tumor growth (15,19). We and others demonstrated that BRD4-NUT stimulatesBRD4 transcription function to activate specifically the expres-sion of oncogenes such as SOX2 (28, 30) and c-MYC (18), whichcollectively drive the potent NMC transforming activity. How-ever, the molecular mechanisms by which BRD4-NUT modu-lates BRD4 function to induce such highly aggressive carcinomasremain to be elucidated.In this study, we found that BRD4 is hyperphosphorylated in
NMC tumors and that this hyperphosphorylation is linked to itsability to drive oncogene expression and cellular transformation.We found that BRD4 is hyperphosphorylated in other BRD4-associated cancers as well. Our study revealed a cellular mech-anism that could regulate BRD4’s biological function through phos-phorylation, which, when dysregulated could lead to oncogenesis.
ResultsBRD4 Is Hyperphosphorylated in NMC Tumors. From our previousNMC studies (28, 30, 31), we observed that BRD4 isolated fromNMC cells, including HCC2429, 10-15, 14169, and Ty-82 cells,migrates more slowly in SDS/PAGE than do a number of non–NMC cells, such as HEK293, C33A, HeLa, U2OS, andA549 cells (Fig. 1A). Because both types of cells carry the wild-type BRD4 gene, our observation indicates that BRD4 has dif-ferent posttranslational modification(s) in NMC and non–NMCcells. Therefore, we investigated the BRD4 phosphorylationstatus in these cells. Whole-cell lysates isolated from bothHCC2429 and HEK293 cells were reactive to a phospho-BRD4 antibody, α-pS484/488 (Fig. S1A), which was raisedagainst diphosphorylated S484 and S488 residues of BRD4 (33).This result suggested that BRD4 is phosphorylated in both celltypes. To understand the different mobility shift of BRD4 in thesecell types, we analyzed BRD4 proteins from HCC2429 andHEK293 cells and from noncancerous cells such as human dermalfibroblasts (HDFs) on an acrylamide gel containing the chemicalPhos-tag that binds specifically to phosphorylated proteins andselectively retards their migration during electrophoresis (34).Remarkably, BRD4 isolated from HCC2429 cells migrates to amuch higher position than BRD4 isolated from HEK293 cells orfrom HDFs (Fig. 1B). More importantly, after dephosphorylationtreatment with lambda protein phosphatase (λPP) or calf in-testinal alkaline phosphatase, BRD4 proteins isolated fromHCC2429 and HEK293 cells and from HDFs migrated to thesame position in Phos-tag gel, which was much lower the positionof untreated BRD4 from any of the cell types (Fig. 1C and Fig.S1B). This mobility shift was reversed when phosphatase inhib-itors (PIs) were added to the phosphatase reaction. These studiesdemonstrated that BRD4 is phosphorylated in all three celltypes, but the phosphorylation in HCC2429 NMC cells is clearlydifferent, most likely from hyperphosphorylation.That the dephosphorylated BRD4 proteins in HCC2429 and
HEK293 cells and HDFs migrated to the same position in Phos-tag gel also confirmed that the unphosphorylated BRD4 pro-teins in these three cell types have the same molecular weight.The result suggested that the unique cellular environment inHCC2429 NMC cells might induce BRD4 hyperphosphorylation.To test this possibility, we transfected a construct expressing HAand FLAG dual-tagged BRD4 into HCC2429 and HEK293 cells.At 2 d posttransfection, the whole-cell lysates were analyzed on aPhos-tag gel (Fig. 1D). As observed with the endogenous BRD4,
HA-FLAG-BRD4 expressed in HCC2429 cells migrated to amuch higher position than the same protein expressed in HEK293cells (Fig. 1D). Together, our study demonstrated that BRD4 ishyperphosphorylated in NMC cells.
CDK9 Is a Potential Kinase That Mediates BRD4 Hyperphosphorylation.We then performed an in vitro kinase assay to confirm that BRD4is hyperphosphorylated in NMC cells compared with HEK293cells. Recombinant BRD4-TII protein, in which BRD4 is fused totwo IgG-binding domains of protein A through a tobacco etchvirus (TEV) protease cleavage site, was expressed in Escherichiacoli and affinity purified on IgG beads. The BRD4-TII beads wereincubated with an equal amount of nuclear proteins isolated fromHCC2429 or HEK293 cells to immunoprecipitate the kinases forBRD4. The immunocomplexes captured on the beads were thensubjected to an in vitro kinase assay (Fig. 2A). Compared with thebuffer control, BRD4-TII was phosphorylated in both reactionsusing either HCC2429 or HEK293 nuclear extract, but muchmore [γ-32P]ATP was incorporated into BRD4-TII incubated withthe HCC2429 nuclear extract (Fig. 2A). Thus, this in vitro systemwas able to mimic the situation in the cells and confirmed that ahigher level of BRD4 phosphorylation was induced by the cellularfactors from NMC cells.Kinases often bind directly to their substrates (35). The data
shown in Fig. 2A also suggest that BRD4 kinases can be isolatedusing the BRD4-TII beads. We then used SILAC (stable isotopelabeling with amino acids in cell culture)-based proteomic
Fig. 1. BRD4 is hyperphosphorylated in NMCs. (A) Whole-cell lysates ofnon–NMCs and NMCs were resolved on SDS/PAGE and immunoblotted withBRD4 C-terminal domain (BRD4 C) and NUT antibodies. Brackets mark thedifference in the position of NMC and non–NMC BRD4. (B) Whole-cell lysatesof HCC2429 and HEK293 cells and HDFs were resolved on a Phos-tag gel andimmunoblotted with BRD4 C antibody. Molecular weight markers are shownon the right. However, as indicated by the manufacturer, the molecularweight markers are frequently distorted during Phos-tag gel electrophoresis,and therefore they can be used only as rough estimates of the molecularweights. (C) BRD4 immunoprecipitated from HCC2429 and HEK293 cells andHDFs was mixed with buffer, λPP, or PIs. Samples were incubated at 4 °C or30 °C as indicated. Proteins then were analyzed on Phos-tag gel andimmunoblotted with BRD4 C antibody. (D) HCC2429 and HEK293 cells weretransfected with a construct expressing HA-FLAG–tagged BRD4. At 2 dposttransfection, whole-cell lysates were analyzed on Phos-tag gel andimmunoblotted with HA antibody. Because of the different transfectionefficiencies of the two cell lines, the lysates of HCC2429 and HEK293 wereloaded at the ratios of 3:1, 5:1, and 8:1 as indicated. Lysates from untrans-fected HCC2429 and HEK293 cells were also loaded on the same Phos-tag gelto show the endogenous BRD4 controls. The samples were immunoblottedwith BRD4 C antibody. See also Fig. S1.
Wang et al. PNAS | Published online June 19, 2017 | E5353
CELL
BIOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
Aug
ust 2
7, 2
020

analysis to identify the BRD4-interacting kinases that contributeto its hyperphosphorylation in NMC cells (Fig. 2B). BRD4-TIIwas used as the bait protein, and TII was used as the tag control(Fig. 2B and Fig. S2A). An in vitro kinase assay confirmed thatTII did not pull down any detectable kinase activity, but BRD4-TII coimmunoprecipitated more kinase activity from HCC2429cells than from HEK293 cells (Fig. S2A). HCC2429 cells thenwere cultured either in normal (light) medium or medium con-taining heavy isotope (13C15N)-labeled arginine and lysine. Nu-clear extracts from light-labeled cells were coimmunoprecipitatedwith TII tag protein immobilized on IgG beads, and the nuclearextracts from heavy-labeled cells were coimmunoprecipitated withBRD4-TII beads (Fig. 2B). After extensive washing to removeunbound proteins, the two samples were combined at a 1:1 ratioand subjected to mass spectrometry. Following the principles ofSILAC technology (36), the proteins with a heavy-to-light (H/L)ratio of more than 1 as determined by mass spectrometry wereidentified as BRD4-associated proteins (Fig. 2C). From this study,several protein kinases, including CDK9, the kinase component ofP-TEFb, PRKDC, CKIIα, and TRIM28 (37), were identified asBRD4-interacting proteins in NMCs (Fig. 2C and Fig. S2B).Among these, CDK9 was found to be the most abundant BRD4-associated kinase with the highest H/L ratio (Fig. 2C and Fig. S2B);the levels of the other three kinases were much lower in the BRD4-binding proteins (Fig. 2C).In parallel with the proteomic study, we also performed a
minicompound screening to determine the effect of numerousdrugs on BRD4 hyperphosphorylation. Among all the kinaseinhibitors tested, only the known CDK9 inhibitors could blockBRD4 hyperphosphorylation in NMC cells (Table S1). In aPhos-tag gel phosphorylation assay, all three CDK9 inhibitorstested—LDC000067, flavopiridol, and PHA767491—showeda significant inhibition of BRD4 hyperphosphorylation inHCC2429 NMC cells (Fig. 2D and Fig. S2C). In addition, twoother less specific CDK9 inhibitors, 5,6-dichloro-1-β-D-ribofur-anosylbenzimidazole (DRB) (38) and apigenin (39), also re-duced BRD4 hyperphosphorylation in HCC2429 cells (Fig.S2C). All the inhibitors tested also moderately inhibited BRD4phosphorylation in HEK293 cells (Fig. 2D and Fig. S2C). Incontrast, treatment with a casein kinase II-specific inhibitor,4,5,6,7-tetrabromobenzotriazole (TBB), which has been shownto inhibit BRD4 phosphorylation (33), did not affect BRD4phosphorylation in either HCC2429 NMC cells or HEK293 cellsin the Phos-tag gel assay (Fig. S2D).Notably, BRD4 from the NMC cells treated with these
CDK9 inhibitors migrated to the same position as the lessphosphorylated BRD4 in the untreated HEK293 cells (Fig. 2Dand Fig. S2C) but not to the lower position of completelydephosphorylated BRD4 as shown in Fig. 1C. This result sug-gested that other kinase(s) besides CDK9 could be involved inBRD4 phosphorylation. In addition, the CDK9 inhibitors wereable to block BRD4 hyperphosphorylation efficiently in NMCcells but inhibited BRD4 phosphorylation in HEK293 cells to alesser degree (Fig. 2D and Fig. S2C). These results indicated thatan NMC-specific mechanism might allow CDK9 to hyper-phosphorylate BRD4 in these cells.In the in vitro kinase assay using nuclear extract of NMC cells to
phosphorylate recombinant BRD4, the CDK9 kinase inhibitors alsodramatically inhibited BRD4 phosphorylation, whereas TBB hadno appreciable effect (Fig. 2E). However, TBB was able to inhibit invitro phosphorylation of the short isoform of BRD4, BRD4S (2),which contains the N-terminal bromodomains and the extra-terminal domain (Fig. S2E). Together, these results suggest that
Fig. 2. CDK9 contributes to BRD4 hyperphosphorylation in NMC. (A) BRD4-TII expressed in E. coli was affinity purified on IgG beads. The beads wereeither kept in buffer or coimmunoprecipitated with equal amounts of nu-clear proteins from HCC2429 or HEK293 cells. After washing, beads weresubjected to kinase assay. BRD4-TII samples then were analyzed on SDS/PAGE and visualized by autoradiography (Autorad). BRD4-TII extracted fromthe beads before kinase assay was analyzed on SDS/PAGE and stained withCoomassie Brilliant Blue (CBB). Asterisks indicate the full-length BRD4-TII.Arrows mark a BRD4-TII fragment purified from E. coli. (B) Flow diagramof the SILAC experiment for identifying BRD4-associated proteins.HCC2429 cells were cultured in light medium (with normal arginine and ly-sine) or in heavy medium (with 13C- and 15N-labeled arginine and lysine).Nuclear extracts from light-labeled cells were coimmunoprecipitated withbeads containing cross-linked TII tag protein, which served as the back-ground control. Nuclear extracts from heavy-labeled cells were coimmuno-precipitated with beads with cross-linked BRD4-TII. After extensive washing,the two samples were mixed at a 1:1 ratio and subjected to mass spec-trometry. (C) Plot showing the normalized SILAC ratio intensity. Shown is theSILAC H/L ratio versus the corresponding protein intensity distribution. Reddots indicate BRD4-associated kinases identified with log2(ratio H/L)>0. Bluedots highlight cyclin T1, cyclin T2, and components of the NELF complex.PRKDC encodes the catalytic subunit of the DNA-dependent protein kinase(DNA-PK). The total SILAC data are shown in Dataset S1. (D) HEK293 andHCC2429 cells were treated with DMSO or 5 μM LDC000067 for 1 h. Whole-cell lysates were analyzed on Phos-tag gel and immunoblotted with BRD4 Cantibody. (E) BRD4-TII expressed in E. coli was affinity purified on IgG beadsand used to coimmunoprecipitate nuclear proteins from HCC2429 cells.Equal amounts of immunocomplexes were subjected to in vitro kinase assayin the presence of DMSO, 50 μMDRB, 50 μM apigenin, 50 μM TBB, or 2 μM or10 μM LDC000067. The samples then were analyzed on SDS/PAGE and vi-sualized by autoradiography. The asterisk indicates the full-length BRD4-TII.The arrow marks a BRD4-TII fragment purified from E. coli. (F) BRD4-TIIexpressed in E. coli was used to coimmunoprecipitate GST, GST-CDK9, GST-cyclin T1, MBP (Maltose-binding protein), or MBP-CDK9 from E. coli lysates asindicated. The immunocomplexes were subjected to in vitro kinase assay.The samples were resolved on SDS/PAGE and visualized by autoradiography.
Phosphorylated BRD4-TII, cyclin T1, and CDK9 are marked with red, green,and blue asterisks, respectively. The arrow marks a nonspecific band presentin all the samples. See also Fig. S2, Table S1, and Dataset S1.
E5354 | www.pnas.org/cgi/doi/10.1073/pnas.1703071114 Wang et al.
Dow
nloa
ded
by g
uest
on
Aug
ust 2
7, 2
020

CDK9 plays a critical role in BRD4 hyperphosphorylation in NMCcells. Therefore, we focused on CDK9 in the rest of this study.In a previous study, P-TEFb purified from mammalian cells
was shown to phosphorylate BRD4 protein isolated fromSf9 insect cells in vitro (40). To test whether CDK9 could directlyphosphorylate BRD4, recombinant BRD4-TII expressed inE. coli and immunoprecipitated on IgG beads was used to coim-munoprecipitate CDK9 and cyclin T1, which also were expressedin E. coli (Fig. S2F). Western blotting showed that BRD4-TIIcould pull down CDK9 and cyclin T1 efficiently, individually orin complex (Fig. S2G). Interestingly, only the immunocomplexescontaining both CDK9 and cyclin T1 demonstrated strong kinaseactivity for BRD4 in the in vitro phosphorylation assay (Fig. 2F).The results provided direct evidence that CDK9 in the P-TEFbcomplex functions as a BRD4 kinase. Various CDK9 inhibitorsalso dramatically inhibited BRD4 phosphorylation in this in vitrophosphorylation assay, whereas TBB and palbociclib, a selectiveinhibitor of CDK4 and CDK6, had moderate effects on BRD4phosphorylation (Fig. S2H). This in vitro experiment using allrecombinant proteins further confirmed the specific role of CDK9in BRD4 hyperphosphorylation.
The Molecular Mechanism Underlying BRD4 Hyperphosphorylation inNMC. We then set out to understand the mechanism by whichCDK9 leads to BRD4 hyperphosphorylation in NMCs. We first
examined the expression level of relevant genes in HCC2429NMC cells and compared them with HEK293 cells by RT-qPCR(Fig. 3A). Because NMC cells carry only one copy of the intactBRD4 gene, it was not surprising that the BRD4 transcript levelwas about half that observed in HEK293 cells (Fig. 3A). Incontrast, the CDK9 transcript level was nearly doubled inHCC2429 cells compared with HEK293 cells (Fig. 3A). Theexpression levels of cyclin T1 and cyclin T2 were comparable inHCC2429 and HEK293 cells. Hexamethylene bisacetamide(HMBA)-inducible protein 1 (HEXIM1) is a P-TEFb inhibitorthat binds to cyclin T1 and sequesters the P-TEFb complex intoan inhibitory complex containing the 7SK small nuclear ribo-nucleoprotein (41). Interestingly, the level of HEXIM1 was sig-nificantly lower in HCC2429 cells (Fig. 3A). Western blottinganalysis of these proteins in the two cell types confirmed thehigher level of CDK9 and lower level of HEXIM1 inHCC2429 cells (Fig. S3). In addition, we consistently detected ahigher level of cyclin T1 protein in HCC2429 cells than inHEK293 cells (Fig. 3 B and C and Fig. S3). Therefore, the higherlevel of P-TEFb and the reduced level of the P-TEFb inhibitorHEXIM1 may contribute to BRD4 hyperphosphorylation inNMC cells.We also examined the BRD4–P-TEFb interaction in HCC2429
and HEK293 cells. An antibody specific for the N terminus ofBRD4 pulled down a similar amount of BRD4 from HCC2429
Fig. 3. Enrichment of CDK9 in BRD4–NUT–inducedhyperacetylated chromatin foci causes BRD4 hy-perphosphorylation. (A) Relative mRNA levels inHEK293 and HCC2429 cells were measured by RT-qPCR and normalized to GAPDH mRNA levels. Thevalues for HEK293 mRNA levels were set as 1. Valuesrepresent the average of three independent experi-ments; error bars indicate SD. ***P < 0.001. (B) Nuclearextracts of HEK293 and HCC2429 cells were immu-noprecipitated with normal rabbit (NR) or Brd4N-terminal domain (Brd4 N) antibodies. The sampleswere immunoblotted with the indicated antibodies.(C) Nuclear extracts of HEK293 and HCC2429 cellswere immunoprecipitated with normal rabbit (NR) orCDK9 antibodies. The samples were immunoblottedwith the indicated antibodies. (D) The mechanismsunderlying BRD4 hyperphosphorylation in NMC.BRD4-NUT recruits and activates p300 to induce his-tone hyperacetylation in discrete chromatin foci inNMC. This hyperacetylation leads to the sequestra-tion and enrichment of BRD4 and associated P-TEFbin the hyperacetylated chromatin domains, lead-ing to BRD4 hyperphosphorylation. (E) HCC2429 cellswere transfected with control (CO) or NUT siRNA. At30 h posttransfection, whole-cell lysates were ana-lyzed on Phos-tag or SDS/PAGE gels and wereimmunoblotted with the indicated antibodies. Thebracket marks the BRD4 bands with reduced levelsof phosphorylation. (F) HEK293 cells were trans-fected with the empty vector control or a constructexpressing Xpress-tagged BRD4-NUT (XBRD4-NUT).At 2 d posttransfection, whole-cell lysates were an-alyzed on Phos-tag or SDS/PAGE gels and wereimmunoblotted with the indicated antibodies. Thebracket marks the hyperphosphorylated BRD4bands. In both E and F, lysates from untransfectedHCC2429 and HEK293 cells were also loaded on thesame Phos-tag gel to show the endogenous BRD4.See also Fig. S3.
Wang et al. PNAS | Published online June 19, 2017 | E5355
CELL
BIOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
Aug
ust 2
7, 2
020

and HEK293 cells, but the amount of CDK9 and cyclin T1coimmunoprecipitated with BRD4 was dramatically greater inHCC2429 than in HEK293 cells (Fig. 3B). In the reciprocalcoimmunoprecipitation experiment, a CDK9 antibody also pulleddown a significantly increased level of BRD4 from HCC2429 cellscompared with HEK293 cells (Fig. 3C). Although these coim-munoprecipitation results suggested an increased interaction be-tween P-TEFb and BRD4 in HCC2429 NMC cells compared withHEK293 cells, this increase might also be caused in part by thehigher levels of CDK9 and cyclin T1 detected in HCC2429 cells(Fig. 3 B and C).We also examined whether other cellular factors such as local
kinase and substrate concentrations might contribute to BRD4hyperphosphorylation in NMC cells. In previous studies (28, 30,31), we showed that in NMC cells BRD4-NUT localizes to dis-crete chromatin foci containing hyperacetylated histones andrecruits histone acetyltransferases (HATs), such as p300, tostimulate more histone hyperacetylation in these chromatin foci.The hyperacetylated histones provide binding sites for the ac-cumulation of the BRD4 and P-TEFb complexes (Fig. 3D). Wealso have shown that P-TEFb is enriched in these hyper-acetylated chromatin foci (31). We therefore speculated thatBRD4 enrichment and associated P-TEFb in these hyper-acetylated BRD4-NUT foci might increase the local concentra-tions of both the kinase and substrate, thus leading to BRD4
hyperphosphorylation in NMC cells (Fig. 3D). BRD4-NUTknockdown in NMC cells disperses the hyperacetylated chro-matin foci, causing BRD4 to be released from the punctatechromatin foci and to diffuse throughout chromatin (31). Im-portantly, BRD4-NUT knockdown in HCC2429 NMC cells alsocaused a partial reduction of BRD4 hyperphosphorylation (Fig.3E). Conversely, BRD4-NUT expressed ectopically in non–NMCcells induces the formation of histone-hyperacetylated nuclearfoci in non–NMC cells and recruits BRD4 to punctate chromatinfoci (31). We found that in HEK293 cells transfected with aBRD4-NUT expression construct a portion of the endogenousBRD4 also became hyperphosphorylated and migrated to thesame position as BRD4 isolated from HCC2429 NMC cells (Fig.3F). These studies supported the role of BRD4-NUT inpromoting BRD4 hyperphosphorylation.We also demonstrated that BRD4-NUT enriched in the hyper-
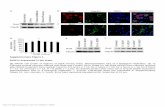
acetylated chromatin domain strongly stimulates the abnormalactivation of the stem cell marker SOX2 to support the aberrantstem cell-like proliferation and highly aggressive transforming ac-tivity of NMC (30). Inhibition of BRD4 hyperphosphorylation inNMC cells by BRD4-NUT knockdown also correlates with SOX2repression (30), suggesting that BRD4 hyperphosphorylation in-duced by BRD4-NUT might contribute to the oncogene trans-activation that supports NMC transformation.
Fig. 4. Dispersion of BRD4-NUT foci reduces BRD4hyperphosphorylation, inhibits downstream onco-gene expression, and abolishes NMC transformation.(A) HCC2429 cells were treated with 20 μM C646 for1 h, 1 μM (+)-JQ1 for 1 h, or 5 mM HMBA for 12 h.Untreated control cells and treated cells were fixedand immunostained with BRD4 C and p300 anti-bodies and were counterstained with DAPI. (Scalebars: 10 μm.) (B) HEK293 and HCC2429 cells weretreated with DMSO or 20 μM C646 for 1 h. Whole-cell lysates were analyzed on Phos-tag or SDS/PAGEgel and were immunoblotted with the indicatedantibodies. (C) HEK293 and HCC2429 cells weretreated with 1 μM (−)-JQ1 or (+)-JQ1 for 1.5 h.Whole-cell lysates were analyzed on Phos-tag or SDS/PAGE gel and were immunoblotted with the in-dicated antibodies. (D) HCC2429 cells were seeded inthe presence of 100 nM (−)-JQ1 or (+)-JQ1 for thesoft agar assay. The percentage of cells formingcolonies was calculated from 8,000 seeded cells.Values represent the average of three independentexperiments; error bars indicate SD. (E) HEK293 andHCC2429 cells were treated with or without 5 mMHMBA for 12 h. Whole-cell lysates were analyzed onPhos-tag or SDS/PAGE gel and were immunoblottedwith the indicated antibodies. (F) HCC2429 cellswere treated with 3 mM or 5 mM HMBA. Untreatedcontrol (CO) or treated cells were harvested at theindicated times. Whole-cell lysates were immuno-blotted with the indicated antibodies. (G) HCC2429cells were seeded in the presence or absence ofHMBA at the indicated concentration for the sphere-formation assay. The percentage of cells formingspheres was calculated from 80 seeded cells. Valuesrepresent the average of three independent exper-iments; error bars indicate SD. (H) HCC2429 cellswere seeded in the absence or presence of 3 mMHMBA for the soft agar assay. The percentage ofcells forming colonies was calculated from 8,000seeded cells. Values represent the average of threeindependent experiments; error bars indicate SD.***P < 0.001. See also Fig. S4.
E5356 | www.pnas.org/cgi/doi/10.1073/pnas.1703071114 Wang et al.
Dow
nloa
ded
by g
uest
on
Aug
ust 2
7, 2
020

Inhibition of BRD4 Hyperphosphorylation by Chemical CompoundsRepresses Oncogene Expression and Cellular Transformation. Be-cause we discovered that BRD4 recruitment to hyperacetylatedchromatin foci stimulates its hyperphosphorylation, we testedwhether blocking the formation of these foci in NMC cells couldblock BRD4 hyperphosphorylation and downstream oncogeneexpression. Both BRD4-NUT knockdown and overexpressionprovided evidence that BRD4–NUT–induced hyperacetylatedchromatin foci are critical for triggering BRD4 hyperphos-phorylation in NMC cells. However, the low transfection effi-ciency for BRD4-NUT siRNA and expression construct resultedin very moderate changes in BRD4 hyperphosphorylation in thetransfected cells (Fig. 3 E and F). We therefore used the chemicalapproach using three different compounds to examine furtherwhether blocking BRD4–NUT–induced HAT recruitment andhistone hyperacetylation prevents BRD4 hyperphosphorylation inNMC cells (Fig. 4).We first started by disrupting the BRD4–NUT/p300–induced
hyperacetylated chromatin foci (Figs. 3D and 4A). In the un-treated HCC2429 cells, both BRD4 and p300 were sequesteredin the hyperacetylated, distinct chromatin foci (Fig. 4A). Treat-ment of HCC2429 cells with the p300 inhibitor C646 (30) foronly 1 h efficiently dispersed the hyperacetylated BRD4-NUTchromatin foci in all the treated cells (Fig. 4A). Remarkably,C646 treatment also resulted in a significant reduction of
BRD4 hyperphosphorylation in HCC2429 cells without dra-matically affecting the BRD4 phosphorylation level inHEK293 cells (Fig. 4B). We, and others, have demonstrated thatBRD4-NUT and BRD4 are recruited to induce abnormal acti-vation of oncogenes such as SOX2 (30) and c-MYC (18) withinthe hyperacetylated chromatin domains in NMC. By inhibitingp300 activity with C646, we observed a significant repression ofSOX2 (30). Both the mRNA and protein levels of c-MYC werealso reduced in C646-treated NMC cells (Fig. S4). These resultsindicate that BRD4–NUT–recruited p300 plays an importantrole in BRD4 hyperphosphorylation, which correlates withdownstream oncogene expression.We also reasoned that if BRD4 binding to the hyperacetylated
chromatin foci is critical for enriching P-TEFb to stimulateBRD4 hyperphosphorylation (Fig. 3D), dissociation of BRD4from acetylated histones by the BET inhibitor (+)-JQ1 (19) mightalso inhibit its hyperphosphorylation by dispersing the foci con-taining both highly concentrated CDK9 kinase and BRD4 sub-strate. Indeed, HCC2429 cells treated with (+)-JQ1 showedsignificantly reduced BRD4 hyperphosphorylation compared withthe cells treated with the inactive stereoisomer (−)-JQ1 (Fig. 4C).In contrast, the BRD4 phosphorylation level in HEK293cells was not reduced but instead was slightly increased by(+)-JQ1 treatment (Fig. 4C), indicating that it might be regulatedthrough a different mechanism, such as BET inhibitor-induced
Fig. 5. Blocking the BRD4–CDK9 interaction reducesBRD4 hyperphosphorylation, inhibits downstreamoncogene expression, and abolishes cellular trans-formation. (A) BRD4–C229 functions as a DNI toblock the BRD4–CDK9 interaction. (B) Nuclear ex-tracts of HCC2429 cells were coimmunoprecipitatedwith normal rabbit (NR), CDK9, or cyclin T1 anti-bodies in the presence of 1.3 μg purified GST control(CO) or GST–C229. The immunocomplexes wereimmunoblotted with the indicated antibodies.(C) Nuclear extracts of HCC2429 cells were coimmu-noprecipitated with normal rabbit (NR) or BRD4 Nantibodies in the presence of 2, 4, 8, or 16 μg purifiedGST or GST–C229. The immunocomplexes wereimmunoblotted with the indicated antibodies.(D) HCC2429 cells carrying a vector control (CO) or aninducible Xpress-C229 construct were treated withDox and harvested as indicated. Cell lysates wereanalyzed on Phos-tag or SDS/PAGE gels and blot-ted with the indicated antibodies. (E) Growthcurves of HCC2429 control (CO) or C229 induciblecells after treating with or without 1 μg/mL Dox.(F). HCC2429 control (CO) or C229 inducible cellswere seeded in the presence or absence of 1 μg/mLDox for the sphere-forming assay. The percent-age of cells forming spheres was calculated from240 seeded cells. Values represent the average offour independent experiments; error bars indicateSD. (G) HCC2429 control (CO) or C229 inducible cellswere seeded in the presence or absence of 1 μg/mLDox for the soft agar assay. The percentage of cellsforming colonies was calculated from 12,000 seededcells. Values represent the average of three in-dependent experiments; error bars indicate SD.(H) Representative images of soft agar plates for theexperiments shown in G. (Scale bar: 1 mm.) **P <0.01; ***P < 0.001. See also Fig. S5.
Wang et al. PNAS | Published online June 19, 2017 | E5357
CELL
BIOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
Aug
ust 2
7, 2
020

adaptive kinome reprogramming (42). The (+)-JQ1 treatment ofNMC cells also led to SOX2 repression, abrogation of the aber-rant stem cell-like proliferation, and complete disruption of col-ony formation in soft agar plates by cancer cells (Fig. 4D) (30).Together, these results provided additional evidence that BRD4hyperphosphorylation is linked to its downstream oncogene ex-pression during NMC cellular transformation.Another chemical compound, HMBA, has been used to in-
duce cancer cell differentiation for treating a number of cancers(43). The mechanism of HMBA action involves inducing theexpression of the P-TEFb inhibitor HEXIM1 (41). In addition,HMBA also acts as a selective bromodomain inhibitor and dis-places BET proteins from chromatin (43). We therefore testedthis early anticancer agent in NMC cells. HCC2429 cells treatedwith HMBA indeed showed diffusion of BRD4 away from thepunctate foci observed in untreated cells, indicating that HMBAcould dissociate BRD4 from hyperacetylated chromatin domain(Fig. 4A). HMBA also efficiently decreased BRD4 hyper-phosphorylation in HCC2429 cells without dramatically affectingthe BRD4 phosphorylation level in HEK293 cells (Fig. 4E). Thiseffect of HMBA may be attributable to its ability to induceHEXIM1, which then can negatively regulate the CDK9/cyclinT1 complex (41). More importantly, HMBA treatment also led toa dramatic repression of the BRD4 downstream oncogenes SOX2and c-MYC while inducing the cellular differentiation markerscytokeratin and involucrin (Fig. 4F). As we reported previously(30), NMC cells can grow into stem cell-like spheres in culturedishes and form colonies in soft agar. We therefore performedsphere-formation and soft agar transformation assays to de-termine if HMBA inhibits NMC proliferation and cellular trans-formation. Unlike the untreated NMC cells, HMBA-treated NMCcells could no longer grow into cancer stem cell-like spheres or astransformed colonies in soft agar plates (Fig. 4 G and H).In summary, three different chemical compounds used in this
study, with different mechanisms of action for blocking BRD4recruitment to the hyperacetylated chromatin foci, were all ableto inhibit BRD4 hyperphosphorylation. We also demonstrated acorrelation between BRD4 hyperphosphorylation and its abilityto drive downstream oncogenes that directly contribute to NMCtumorigenesis. This study further indicates that BRD4 hyper-phosphorylation may be an important factor for sustaining targetoncogene expression and cellular transformation.
A Dominant Negative Inhibitor of BRD4–CDK9 Interaction AbrogatesBRD4 Hyperphosphorylation, Oncogene Expression, and CellularTransformation in NMC. Because CDK9 binds BRD4 through itsC-terminal domain (44), we tested if the BRD4 C-terminal do-main could be used as a dominant negative inhibitor (DNI) toblock the BRD4 and CDK9 interaction competitively and toinhibit BRD4 hyperphosphorylation (Fig. 5A). A set of con-structs encoding varied lengths of the BRD4 C-terminal domainsfused to the Xpress tag were transfected into HCC2429 cells.The fragment spanning amino acids 1134–1362 showed thehighest expression level in HCC2429 cells when expressed fromtwo different vectors (Fig. S5A). Therefore, it was chosen for therest of this study and was named “C229.” We first tested theeffect of C229 on the P-TEFb–BRD4 interaction by performingcoimmunoprecipitation. Because C229 was degraded veryquickly in the HCC2429 cell lysates used for coimmunoprecipi-tation, we performed the coimmunoprecipitation using HCC2429cell lysates supplemented with or without recombinant GST-C229 expressed in E. coli (Fig. S5B). Both CDK9 and cyclinT1 antibodies coimmunoprecipitated BRD4 in the absence of theC229, but the antibodies pulled down only C229 when it was addedinto the reaction (Fig. 5B). C229 also caused a dose-dependentreduction of P-TEFb coimmunoprecipitated with BRD4 antibody(Fig. 5C). NMC cells then were transduced with lentivirusesencoding the doxycycline (Dox)-inducible C229 to test how
blocking the P-TEFb–BRD4 interaction affects BRD4 hyper-phosphorylation and downstream gene expression. Remarkably,expression of C229 in NMC cells effectively inhibited BRD4hyperphosphorylation and repressed its oncogenic target genesSOX2 and MYC (Fig. 5D and Fig. S5C). On the other hand,C229 expression led to significant induction of NMC cell differ-entiation, as indicated by the robustly increased differentiationmarkers cytokeratin, involucrin, and c-FOS (Fig. 5D and Fig.S5C). The C229-expressing HCC2429 cells also showed a clearlydifferentiated morphology, with nearly all the cells becoming flatand enlarged (Fig. S5 D and E). C229 expression also significantlyinhibited the growth of NMC tumor cells (Fig. 5E). The inducedC229 expression diminished the ability of NMC cells to grow asstem cell-like spheres and soft agar colonies, abolishing NMCtransformation (Fig. 5 F–H). In contrast, C229 expression inHEK293 and C33A cells, which do not show BRD4 hyper-phosphorylation, did not significantly repress cellular proliferation,differentiation, or transformation (Fig. S5 F–K). Although C229expression in HEK293 cells did not dramatically change the numberof colonies grown in soft agar, the size of these colonies was reduced(Fig. S5K). The underlying mechanism remains to be investigated.Thus, the effect of C229 on BRD4 hyperphosphorylation and cel-lular transformation appeared to be specific to NMC. Taken to-gether, our results showed that C229 functions as a DNI to block theP-TEFb–BRD4 interaction, inhibiting BRD4 hyperphosphorylation,oncogene expression, and NMC transformation.
BRD4 Is Also Hyperphosphorylated in Non–NMC Cancers Associatedwith High BRD4 Oncogenic Activity. Our study suggested thatBRD4 hyperphosphorylation is linked to oncogene expressionand cellular transformation in NMC tumors. Because BRD4 is akey therapeutic target that drives oncogene expression in mul-tiple cancers (4, 14), we decided to examine if BRD4 is alsohyperphosphorylated in other cancer cells that have demon-strated enhanced BRD4 oncogenic activity. Activation of theBRD4-dependent core transcriptional program is the key onco-genic driver for various leukemia diseases (10, 45, 46). BRD4 isalso implicated in the pathogenesis of Burkitt’s lymphoma (12). We
Fig. 6. BRD4 is also hyperphosphorylated in non–NMC cancer cells associ-ated with BRD4 dysfunction. (A) Whole-cell lysates from various leukemiaand lymphoma cells were analyzed on a Phos-tag gel and compared with thesamples from HCC2429 and HEK293 cells and HDFs. The samples wereimmunoblotted with BRD4 C antibody. Brackets mark the hyper-phosphorylated BRD4 bands. The samples were also analyzed on SDS/PAGEand immunoblotted with GAPDH. Because the BRD4 protein level in HDFs isextremely low, seven times more HDF protein sample was loaded in thePhos-tag gel than in the SDS/PAGE for the GAPDH blot. The images shownare representative of three independent experiments. (B) Leukemia andlymphoma cells were treated with DMSO or 10 μM LDC000067 for 2 h.Whole-cell lysates were analyzed on Phos-tag or SDS/PAGE gel and wereimmunoblotted with the indicated antibodies. Lysates from untreatedHCC2429 and HEK293 cells were also loaded on the same Phos-tag gel toshow the endogenous BRD4. See also Fig. S6.
E5358 | www.pnas.org/cgi/doi/10.1073/pnas.1703071114 Wang et al.
Dow
nloa
ded
by g
uest
on
Aug
ust 2
7, 2
020

therefore examined the BRD4 phosphorylation status in a numberof leukemia and lymphoma cell lines, including K562, MV4;11,Jurkat, THP-1, HL-60, Namalwa, and Raji cells (Fig. 6). Using thePhos-tag gel assay, we compared the BRD4 phosphorylation statusin these cancer cells and in normal cells such as HDFs, as well as inHCC2429 and HEK293 cells. BRD4 in the normal HDFs migratedto the lowest position in Phos-tag gel, showing very little signalfor phosphorylated BRD4 bands (Fig. 6A). BRD4 is phosphory-lated to some degree in HEK293 cells but shows robust hyper-phosphorylation in HCC2429 cells (Fig. 6A). We detected variousdegrees of BRD4 hyperphosphorylation in K562, Jurkat, Namalwa,Raji, and HL-60 cells, because some of the BRD4 isolated fromthese cancer cells migrated to a higher position in the Phos-tag gelthan BRD4 isolated from HDFs and HEK293 cells (Fig. 6A). Onthe other hand, MV4;11 and THP-1 cells showed much lesshyperphosphorylated BRD4 (Fig. 6A). Notably, treatment with theCDK9 inhibitor LDC000067 dramatically inhibited BRD4 hyper-phosphorylation in these non–NMC cancer cells (Fig. 6B and Fig.S6). These results showed that, in addition to NMC cells, BRD4 isalso hyperphosphorylated in some other cancer cells displaying highlevels of BRD4 oncogenic function and that CDK9 may be a keykinase that contributes to BRD4 hyperphosphorylation.
DiscussionBRD4 is dysregulated in a range of aggressive malignancies (4,10–12, 14, 16, 17) and is being increasingly implicated as a major
driver of oncogenic growth. Hence, it is under intense investigationas a target for cancer therapy. BRD4 also plays a fundamental rolein the regulation of transcription, cell-cycle progression, and cellularproliferation. However, how BRD4 activity is regulated to maintainits normal function in healthy cells and how dysregulation of thisprocess leads to cancer remain poorly understood. Therefore, it hasbeen challenging to develop anticancer strategies that specificallytarget the aberrant BRD4 oncogenic activity in cancer cells withoutaffecting the normal BRD4 function in healthy cells.In this study, we tackled the important question regarding the
mechanistic regulation of BRD4 function during cancer devel-opment. We discovered that BRD4 is hyperphosphorylated in NMCtumors and identified CDK9 as a potential kinase that contributes toBRD4 hyperphosphorylation in NMC (Figs. 1 and 2). HighCDK9 expression levels and robust P-TEFb–BRD4 interaction inNMC may contribute to BRD4 hyperphosphorylation (Fig. 3). Also,in NMC cells, the BRD4 and P-TEFb complexes are enriched at theBRD4–NUT–induced histone-hyperacetylated chromatin domain(31), thus stimulating BRD4 hyperphosphorylation (Fig. 3D). BRD4-NUT knockdown, disrupting the histone-hyperacetylated chromatinfoci, inhibited BRD4 hyperphosphorylation, whereas induction ofhistone-hyperacetylated chromatin domain by expressing BRD4-NUT in non–NMC cells stimulated BRD4 hyperphosphorylation(Fig. 3). Using both chemical and molecular inhibitors to disperseBRD4–NUT–induced hyperphosphorylated chromatin foci or todisrupt the P-TEFb–BRD4 interaction in NMC cells abrogated
Table 1. Summary of in vivo studies using compounds that inhibit BRD4 hyperphosphorylation
Compound Target In vivo studies and references
Flavopiridol CDK1/2/4/6/7/9 Flavopiridol significantly inhibits tumor growth in one of the twoNMC xenograft lines (47).
Flavopiridol prolongs survival in murine transplanted models ofMLL-ENL–transformed cells (48).
Flavopiridol has potent antitumor activity in human leukemia andlymphoma xenografts (49).
Flavopiridol has modest to significant activities in relapsed CLL inclinical trials (50, 51).
PHA-767491 CDC7/CDK9 PHA-767491 has antitumor activity in rodent models of humanleukemia, human colon carcinoma, and DMBA-induced mammarycarcinomas (52).
C646 p300 C646 suppresses in vivo growth of transplanted AE9a leukemia blastsin mice (53).
C646 suppresses tumor growth in a lung cancer xenograft and aleukemia xenograft (54).
(+)-JQ1 BRD4/BRD4-NUT JQ1 promotes tumor regression and prolongs survival in murinemodels of NMC (19). Therapeutic effects for JQ1 were observedin mouse models of hematological malignancies, such as acutemyeloid leukemia, multiple myeloma, and lymphoma, and insome solid tumors, such as neuroblastoma, breast cancer,and prostate cancer (10–14, 55, 56).
OTX015/MK-8628(A JQ1 analog)
BRD4/BRD4-NUT Among four patients with advanced-stage NMC treated withOTX015/MK-8628, two patients responded rapidly with tumorregression and symptomatic relief, and a third had meaningfuldisease stabilization with a minor metabolic response (57).
HMBA Releases CDK9 from its inactive complex;induces HEXIM1; targets p300 and BRD4 (43)
HMBA induces a sharp decrease in tumorigenicity in a murinemodel of human colon cancer (58).
HMBA slightly promotes tumor regression and prolongs survival inmurine models of human oral squamous cell carcinoma (59).
A phase II clinical trial was conducted using HMBA to treat patientswith MDS or AML. Of the 41 patients in the trial, HMBA induced acomplete remission in three patients and a partial remission in sixpatients (60).
AE9a, AML1-ETO 9a fusion gene; AML, acute myelogenous leukemia; CLL, chronic lymphocytic leukemia; DMBA, 7,12-dimethylbenz(a)anthracene; MDS,myelodysplastic syndrome; MLL-ENL, mixed-lineage leukemia (MLL) oncogenic fusion caused by the translocation between chromosomes 11 and 19 (alsocalled eleven nighteen leukemia, ENL); NMC, NUT midline carcinoma.
Wang et al. PNAS | Published online June 19, 2017 | E5359
CELL
BIOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
Aug
ust 2
7, 2
020

BRD4 hyperphosphorylation, downstream oncogene expression,and NMC cellular transformation (Figs. 3–5). Therefore, our studysuggested that BRD4 hyperphosphorylation is linked to its ability todrive downstream oncogene expression and cellular transformationin NMC tumors.Up-regulation of a specific set of oncogenic target genes
driven by BRD4 has been observed in a variety of cancers (4, 14).Therefore, we compared the BRD4 phosphorylation status inthese BRD4-associated cancers with that in normal cells. Wediscovered that BRD4 is also hyperphosphorylated in a numberof non–NMC cancers associated with enhanced BRD4 onco-genic transcription activities (Fig. 6), suggesting that this hyper-phosphorylation may be a general mechanism that supportsBRD4 oncogenic function. In line with this notion, the chemicalcompounds that we found to cause reduction of BRD4 hyper-phosphorylation could significantly repress the growth of bothNMC and non–NMC tumor growth in vivo, with several ofthem demonstrating potent antitumor activities in clinical trials(Table 1). Therefore, it is tempting to speculate that thesecompounds may function, at least in part, by blocking BRD4hyperphosphorylation.Our study suggests that CDK9 is one of the major kinases that
potentially could phosphorylate BRD4 in NMCs. In NMCs,BRD4–NUT–induced enrichment of CDK9 at the hyperace-tylated chromatin foci may lead to BRD4 hyperphosphorylation. Inaddition, p300 recruited by BRD4-NUT to the hyperacetylatedchromatin foci also has the ability to activate CDK9 (61), and thisactivation could contribute to BRD4 hyperphosphorylation inNMC cells. Treating additional BRD4-associated cancers with aCDK9 inhibitor also blocked BRD4 hyperphosphorylation in thesecancer cells (Fig. 6B), suggesting that CDK9 could also mediateBRD4 hyperphosphorylation in these cancers. However, othercellular kinases and protein phosphatases may also regulate BRD4phosphorylation under different physiological conditions. For ex-ample, from the SILAC BRD4 proteomic study described above,we identified a number of kinases and protein phosphatases asBRD4-binding partners (Fig. 2 and Dataset S1). Protein phospha-tases such as PP2A frequently have been found to be geneticallymutated or functionally inactivated in many cancers and thereforehave become attractive targets for anticancer therapy (62). In fact,PP2A has been identified as a BRD4 serine phosphatase in triple-negative breast cancer (23). Building on our finding and these re-cent studies, we reason that coordinated actions of multiple cellularkinases and phosphatases provide a tightly controlled molecularmechanism to regulate BRD4 phosphorylation and function innormal cells and that abnormal activation of these kinases or in-hibition of phosphatases may lead to aberrant BRD4 phosphory-lation, thus inducing BRD4 activity in oncogenes to activatetumorigenesis in NMC and other cancers. Further studies are crit-ical to identify the additional kinases and phosphatases that regulateBRD4 phosphorylation and/or function. Identification of theseBRD4 regulatory factors may allow us to examine how dysregula-tion of the relevant signaling pathways contributes to BRD4 hyper-phosphorylation in BRD4-associated cancers.So far, our study showed only that BRD4 hyperphosphorylation
correlates with downstream oncogene expression and cellulartransformation. Whether phosphorylation of BRD4 itself is thekey event driving cellular transformation or whether the hyper-phosphorylated BRD4 recruits P-TEFb to stimulate downstreamoncogene expression is unclear and remains an important questionfor future studies. To prove definitively that BRD4 hyper-phosphorylation supports its ability to activate the oncogenes that
drive cellular transformation, we will need to identify BRD4 res-idues that are hyperphosphorylated in cancers and test if muta-genesis of BRD4 hyperphosphorylation sites abolishes cellulartransformation. Compared with normal cells, BRD4 is hyper-phosphorylated in a number of BRD4-associated cancers (Fig. 6).Also, the wide range of BRD4 mobility shifts observed in thesecancers suggested that BRD4 is phosphorylated at multiple sites.Indeed, using the KinasePhos 2.0 web server (63), we analyzed theBRD4 protein sequence and identified more than 40 consensusphosphorylation sites for kinases that potentially could phos-phorylate BRD4. As described above, BRD4 also could bephosphorylated by different kinases at different sites in variouscancers. The complexity of BRD4 phosphorylation pattern pre-vents us from mutating a specific residue of BRD4 to test itsfunction in cellular transformation. Our current effort focuses onusing quantitative mass spectrometry to identify systematicallythe BRD4 residues differentially phosphorylated in various can-cers. Identifying the BRD4 hyperphosphorylation sites that areuniquely present in the relevant cancers will allow us to developphosphospecific antibodies that recognize hyperphosphorylatedBRD4 as new diagnostic markers to detect abnormal BRD4function in related cancers. The BRD4 phosphorylation patternswill also be critical for future mechanistic studies to elucidate howBRD4 phosphorylation may dynamically regulate its function inboth normal and cancer cells.In summary, our study uncovered a mechanism by which
BRD4 function could be regulated by cellular kinases and sig-naling pathways, providing insights into how dysregulation of thisimportant epigenetic reader could lead to tumorigenesis. AML,NMC, and other BRD4-associated cancers are often resistant toconventional chemotherapy (15, 64). Therefore, to develop noveltherapeutic approaches, it is imperative to determine the onco-genic mechanisms of these cancers. BRD4 and other BET pro-teins provide ideal “druggable” targets for epigenetic therapy.Elucidation of the mechanism underlying BRD4 hyperphos-phorylation will reveal novel targets, allowing the design of moreeffective anticancer therapeutic strategies to modulate BRD4phosphorylation specifically, thereby turning off its oncogenicactivity in the relevant cancers while sparing the normal BRD4functions described in previous studies from our group andothers (9, 24–26). Our studies also open opportunities for de-veloping BET inhibitors and kinase inhibitors as combinatorialtreatment approaches for BRD4-associated cancers.
Materials and MethodsIn vitro kinase and phosphatase assays, Western blot analyses, immunopre-cipitation, immunofluorescent staining, RT-qPCR, soft agar growth andsphere-formation assays, and SILAC studies were performed using standardmethods. Phos-tag gel analysis was performed using 5% PAGE gel containing10 μM Phos-tag acrylamide AAL-107 (Wako Chemicals) and 40 μM MnCl2following the manufacturer’s instructions. A full description of reagents andmethods used in this study can be found in SI Materials and Methods.
ACKNOWLEDGMENTS. We thank Dr. Junpeng Yan (Case Western ReserveUniversity) for providing plasmids; Drs. Martin P. Carroll (University ofPennsylvania), Erle S. Robertson (University of Pennsylvania), Thao P. Dang(Vanderbilt University), and Christopher A. French (Harvard Medical School)for providing cell lines; Dr. Chao-Xing Yuan and Anne Lehman for techni-cal support; and the members of our laboratories for helpful discussion.This work was supported by NIH Grants R01CA148768, R01CA142723,R01CA187718, R01GM110174, P01CA196539, and R01CA103867; CancerPrevention Research Institute of Texas Grants RP110471 and RP140367;and Welch Foundation Grant I-1805.
1. Dey A, Chitsaz F, Abbasi A, Misteli T, Ozato K (2003) The double bromodomain
protein Brd4 binds to acetylated chromatin during interphase and mitosis. Proc Natl
Acad Sci USA 100:8758–8763.2. Wu SY, Chiang CM (2007) The double bromodomain-containing chromatin adaptor
Brd4 and transcriptional regulation. J Biol Chem 282:13141–13145.
3. Rahman S, et al. (2011) The Brd4 extraterminal domain confers transcription activa-
tion independent of pTEFb by recruiting multiple proteins, including NSD3. Mol Cell
Biol 31:2641–2652.4. Lovén J, et al. (2013) Selective inhibition of tumor oncogenes by disruption of super-
enhancers. Cell 153:320–334.
E5360 | www.pnas.org/cgi/doi/10.1073/pnas.1703071114 Wang et al.
Dow
nloa
ded
by g
uest
on
Aug
ust 2
7, 2
020

5. Marshall NF, Peng J, Xie Z, Price DH (1996) Control of RNA polymerase II elongationpotential by a novel carboxyl-terminal domain kinase. J Biol Chem 271:27176–27183.
6. Czudnochowski N, Bösken CA, Geyer M (2012) Serine-7 but not serine-5 phosphory-lation primes RNA polymerase II CTD for P-TEFb recognition. Nat Commun 3:842.
7. Yang Z, et al. (2005) Recruitment of P-TEFb for stimulation of transcriptional elon-gation by the bromodomain protein Brd4. Mol Cell 19:535–545.
8. Yang Z, He N, Zhou Q (2008) Brd4 recruits P-TEFb to chromosomes at late mitosis topromote G1 gene expression and cell cycle progression. Mol Cell Biol 28:967–976.
9. Houzelstein D, et al. (2002) Growth and early postimplantation defects in mice de-ficient for the bromodomain-containing protein Brd4. Mol Cell Biol 22:3794–3802.
10. Zuber J, et al. (2011) RNAi screen identifies Brd4 as a therapeutic target in acutemyeloid leukaemia. Nature 478:524–528.
11. Delmore JE, et al. (2011) BET bromodomain inhibition as a therapeutic strategy totarget c-Myc. Cell 146:904–917.
12. Mertz JA, et al. (2011) Targeting MYC dependence in cancer by inhibiting BET bro-modomains. Proc Natl Acad Sci USA 108:16669–16674.
13. Puissant A, et al. (2013) Targeting MYCN in neuroblastoma by BET bromodomaininhibition. Cancer Discov 3:308–323.
14. Shi J, Vakoc CR (2014) The mechanisms behind the therapeutic activity of BET bro-modomain inhibition. Mol Cell 54:728–736.
15. French CA (2012) Pathogenesis of NUT midline carcinoma. Annu Rev Pathol 7:247–265.
16. Chapuy B, et al. (2013) Discovery and characterization of super-enhancer-associateddependencies in diffuse large B cell lymphoma. Cancer Cell 24:777–790.
17. Dawson MA, et al. (2011) Inhibition of BET recruitment to chromatin as an effectivetreatment for MLL-fusion leukaemia. Nature 478:529–533.
18. Grayson AR, et al. (2014) MYC, a downstream target of BRD-NUT, is necessary andsufficient for the blockade of differentiation in NUT midline carcinoma.Oncogene 33:1736–1742.
19. Filippakopoulos P, et al. (2010) Selective inhibition of BET bromodomains. Nature 468:1067–1073.
20. Nicodeme E, et al. (2010) Suppression of inflammation by a synthetic histone mimic.Nature 468:1119–1123.
21. Berthon C, et al. (2016) Bromodomain inhibitor OTX015 in patients with acute leu-kaemia: A dose-escalation, phase 1 study. Lancet Haematol 3:e186–e195.
22. Fong CY, et al. (2015) BET inhibitor resistance emerges from leukaemia stem cells.Nature 525:538–542.
23. Shu S, et al. (2016) Response and resistance to BET bromodomain inhibitors in triple-negative breast cancer. Nature 529:413–417.
24. Liu W, et al. (2014) BRD4 regulates Nanog expression in mouse embryonic stem cellsand preimplantation embryos. Cell Death Differ 21:1950–1960.
25. Wu S-Y, et al. (2016) BRD4 phosphorylation regulates HPV E2-mediated viral tran-scription, origin replication, and cellular MMP-9 expression. Cell Reports 16:1733–1748.
26. Bolden JE, et al. (2014) Inducible in vivo silencing of Brd4 identifies potential toxicitiesof sustained BET protein inhibition. Cell Reports 8:1919–1929.
27. Andrieu G, Belkina AC, Denis GV (2016) Clinical trials for BET inhibitors run ahead ofthe science. Drug Discov Today Technol 19:45–50.
28. Wang R, You J (2015) Mechanistic analysis of the role of bromodomain-containingprotein 4 (BRD4) in BRD4-NUT oncoprotein-induced transcriptional activation. J BiolChem 290:2744–2758.
29. Haruki N, et al. (2005) Cloned fusion product from a rare t(15;19)(q13.2;p13.1) inhibitS phase in vitro. J Med Genet 42:558–564.
30. Wang R, et al. (2014) Activation of SOX2 expression by BRD4-NUT oncogenic fusiondrives neoplastic transformation in NUT midline carcinoma. Cancer Res 74:3332–3343.
31. Yan J, Diaz J, Jiao J, Wang R, You J (2011) Perturbation of BRD4 protein function byBRD4-NUT protein abrogates cellular differentiation in NUT midline carcinoma. J BiolChem 286:27663–27675.
32. Reynoird N, et al. (2010) Oncogenesis by sequestration of CBP/p300 in transcription-ally inactive hyperacetylated chromatin domains. EMBO J 29:2943–2952.
33. Wu SY, Lee AY, Lai HT, Zhang H, Chiang CM (2013) Phospho switch triggersBrd4 chromatin binding and activator recruitment for gene-specific targeting. MolCell 49:843–857.
34. Kinoshita E, Kinoshita-Kikuta E, Koike T (2009) Separation and detection of largephosphoproteins using Phos-tag SDS-PAGE. Nat Protoc 4:1513–1521.
35. Fernandes N, et al. (2005) DNA damage-induced association of ATM with its targetproteins requires a protein interaction domain in the N terminus of ATM. J Biol Chem280:15158–15164.
36. Chen X, Wei S, Ji Y, Guo X, Yang F (2015) Quantitative proteomics using SILAC:Principles, applications, and developments. Proteomics 15:3175–3192.
37. Manning G, Whyte DB, Martinez R, Hunter T, Sudarsanam S (2002) The protein kinasecomplement of the human genome. Science 298:1912–1934.
38. Wang S, Fischer PM (2008) Cyclin-dependent kinase 9: A key transcriptional regulatorand potential drug target in oncology, virology and cardiology. Trends Pharmacol Sci29:302–313.
39. Polier G, et al. (2011) Wogonin and related natural flavones are inhibitors ofCDK9 that induce apoptosis in cancer cells by transcriptional suppression of Mcl-1. CellDeath Dis 2:e182.
40. Devaiah BN, Singer DS (2012) Cross-talk among RNA polymerase II kinases modulatesC-terminal domain phosphorylation. J Biol Chem 287:38755–38766.
41. Yik JH, et al. (2003) Inhibition of P-TEFb (CDK9/Cyclin T) kinase and RNA polymerase IItranscription by the coordinated actions of HEXIM1 and 7SK snRNA. Mol Cell 12:971–982.
42. Kurimchak AM, et al. (2016) Resistance to BET bromodomain inhibitors is mediated bykinome reprogramming in ovarian cancer. Cell Reports 16:1273–1286.
43. Nilsson LM, et al. (2016) Cancer differentiating agent hexamethylene bisacetamideinhibits BET bromodomain proteins. Cancer Res 76:2376–2383.
44. Yan J, Li Q, Lievens S, Tavernier J, You J (2010) Abrogation of the Brd4-positivetranscription elongation factor B complex by papillomavirus E2 protein contributesto viral oncogene repression. J Virol 84:76–87.
45. Valent P, Zuber J (2014) BRD4: A BET(ter) target for the treatment of AML? Cell Cycle13:689–690.
46. Knoechel B, et al. (2014) An epigenetic mechanism of resistance to targeted therapyin T cell acute lymphoblastic leukemia. Nat Genet 46:364–370.
47. Beesley AH, et al. (2014) Comparative drug screening in NUT midline carcinoma. Br JCancer 110:1189–1198.
48. Garcia-Cuellar MP, et al. (2014) Efficacy of cyclin-dependent-kinase 9 inhibitors in amurine model of mixed-lineage leukemia. Leukemia 28:1427–1435.
49. Arguello F, et al. (1998) Flavopiridol induces apoptosis of normal lymphoid cells,causes immunosuppression, and has potent antitumor activity In vivo against humanleukemia and lymphoma xenografts. Blood 91:2482–2490.
50. Byrd JC, et al.; Cancer and Leukemia Group B (2005) Treatment of relapsed chroniclymphocytic leukemia by 72-hour continuous infusion or 1-hour bolus infusion offlavopiridol: Results from Cancer and Leukemia Group B study 19805. Clin Cancer Res11:4176–4181.
51. Lin TS, et al. (2009) Phase II study of flavopiridol in relapsed chronic lymphocyticleukemia demonstrating high response rates in genetically high-risk disease. J ClinOncol 27:6012–6018.
52. Montagnoli A, et al. (2008) A Cdc7 kinase inhibitor restricts initiation of DNA repli-cation and has antitumor activity. Nat Chem Biol 4:357–365.
53. Gao XN, et al. (2013) A histone acetyltransferase p300 inhibitor C646 induces cell cyclearrest and apoptosis selectively in AML1-ETO-positive AML cells. PLoS One 8:e55481.
54. Ogiwara H, et al. (2016) Targeting p300 addiction in CBP-deficient cancers causessynthetic lethality by apoptotic cell death due to abrogation of MYC expression.Cancer Discov 6:430–445.
55. Shi J, et al. (2014) Disrupting the interaction of BRD4 with diacetylated Twist sup-presses tumorigenesis in basal-like breast cancer. Cancer Cell 25:210–225.
56. Asangani IA, et al. (2014) Therapeutic targeting of BET bromodomain proteins incastration-resistant prostate cancer. Nature 510:278–282.
57. Stathis A, et al. (2016) Clinical response of carcinomas harboring the BRD4–NUT on-coprotein to the targeted bromodomain inhibitor OTX015/MK-8628. Cancer Discov 6:492–500.
58. Schroy P, Winawer S, Friedman E (1989) Effect on in vivo tumorigenicity of lengthyexposure of human colon cancer cells to the differentiation agent hexamethylenebisacetamide. Cancer Lett 48:53–58.
59. Naito S, et al. (2006) Enhancement of antitumor activity of herpes simplex virusgamma(1)34.5-deficient mutant for oral squamous cell carcinoma cells by hexamethylenebisacetamide. Cancer Gene Ther 13:780–791.
60. Andreeff M, et al. (1992) Hexamethylene bisacetamide in myelodysplastic syndromeand acute myelogenous leukemia: A phase II clinical trial with a differentiation-inducing agent. Blood 80:2604–2609.
61. Fu J, Yoon H-G, Qin J, Wong J (2007) Regulation of P-TEFb elongation complex activityby CDK9 acetylation. Mol Cell Biol 27:4641–4651.
62. Sangodkar J, et al. (2016) All roads lead to PP2A: Exploiting the therapeutic potentialof this phosphatase. FEBS J 283:1004–1024.
63. Wong YH, et al. (2007) KinasePhos 2.0: A web server for identifying protein kinase-specific phosphorylation sites based on sequences and coupling patterns. NucleicAcids Res 35:W588–594.
64. O’Brien JA, Rizzieri DA (2013) Leukemic stem cells: A review. Cancer Invest 31:215–220.
65. Wang R, Li Q, Helfer CM, Jiao J, You J (2012) Bromodomain protein Brd4 associatedwith acetylated chromatin is important for maintenance of higher-order chromatinstructure. J Biol Chem 287:10738–10752.
66. You J, Croyle JL, Nishimura A, Ozato K, Howley PM (2004) Interaction of the bovinepapillomavirus E2 protein with Brd4 tethers the viral DNA to host mitotic chromo-somes. Cell 117:349–360.
67. Li Q, et al. (2004) A modified mammalian tandem affinity purification procedure toprepare functional polycystin-2 channel. FEBS Lett 576:231–236.
68. Nyormoi O, Sinclair JH, Klein G (1973) Isolation and characterization of an adherent,8-azaguanine resistant variant of the Burkitt lymphoma cell line, Raji. Exp Cell Res 82:241–251.
69. Liu W, et al. (2016) Identifying the target cells and mechanisms of Merkel cell poly-omavirus infection. Cell Host Microbe 19:775–787.
70. Dignam JD, Lebovitz RM, Roeder RG (1983) Accurate transcription initiation by RNApolymerase II in a soluble extract from isolated mammalian nuclei. Nucleic Acids Res11:1475–1489.
71. Villén J, Gygi SP (2008) The SCX/IMAC enrichment approach for global phosphoryla-tion analysis by mass spectrometry. Nat Protoc 3:1630–1638.
72. Cox J, Mann M (2008) MaxQuant enables high peptide identification rates, in-dividualized p.p.b.-range mass accuracies and proteome-wide protein quantification.Nat Biotechnol 26:1367–1372.
73. Cox J, et al. (2011) Andromeda: A peptide search engine integrated into theMaxQuant environment. J Proteome Res 10:1794–1805.
Wang et al. PNAS | Published online June 19, 2017 | E5361
CELL
BIOLO
GY
PNASPL
US
Dow
nloa
ded
by g
uest
on
Aug
ust 2
7, 2
020