The origin of pituitary cysts in the rostral pars distalis of the nine-spined stickleback, Pungitius...
14
Cell Tissue Res (1981) 214:417-430 Cell and Tissue Research Springer-Verlag 1981 The Origin of Pituitary Cysts in the Rostral Pars Distalis of the Nine-Spined Stickleback, Pungitius pungitius L. Michael Benjamin Department of Anatomy, UniversityCollege, Cardiff, Wales, U.K. Swmnary. Pituitary cysts in the nine-spined stickleback, Pungitius pungitius L., develop next to blood vessels passing through the prolactin zone of the rostral pars distalis to the connective tissue capsule at its periphery. Cysts were most frequent when pituitaries were large compared with body lengths. However, the incidence of cysts could not be closely related to body length alone. As the rostral pars distalis is more highly vascularised in fish with large pituitaries, and as there was no evidence of accumulating secretion around these blood vessels, it is suggested that cysts develop when vascular demands (or supplies) become excessive. Associated with the greater vascularity of the rostral pars distalis in large pituitaries was a remarkable development of non-granulated cells. Indeed cysts may originate as enlarged intercellular spaces between such cells, as the latter often surround blood vessels. The non-granulated cells are also important in enlarging cyst cavities (by phagocytosing prolactin-cell debris) and perhaps for maintaining their structural integrity. It is suggested that the macrophages within cyst cavities are derived from the non-granulated cells. Key words: Pituitary - Cyst - Prolactin cell - Non-granulated cell - Blood vessels. There are large, fluid-filled cysts in the prolactin zone of the rostral pars distalis (RPD) of the nine-spined stickleback, Pungitius pungitius L. (Benjamin 1978, 1979; Benjamin and Williams 1979a, b). Their diameter varies from 6-200 gm and they are virtually restricted to animals > 20 mm in length. It is thus unlikely that they are of any embryological significance, as are the pituitary cysts in Xiphophorus maculatus (Schreibman 1966). Although present in freshwater animals at all times of the year, they are particularly common in June-August, i.e., towards the end, and just after, the breeding season (Benjamin and Williams 1979 a). As cysts are usually associated with small, inactive, prolactin cells, it seems unlikely that they are stores for prolactin released by holocrine secretion (Benjamin 1979). Send offprint requeststo: Dr. MichaelBenjamin, Department of Anatomy, UniversityCollege, Cardiff CF1 1XL, Wales, U.K. 0302-766 X/81/0214/0417/$02.80
-
Upload
michael-benjamin -
Category
Documents
-
view
212 -
download
0
Transcript of The origin of pituitary cysts in the rostral pars distalis of the nine-spined stickleback, Pungitius...

Cell Tissue Res (1981) 214:417-430 Cell and Tissue Research �9 Springer-Verlag 1981
The Origin of Pituitary Cysts in the Rostral Pars Distalis of the Nine-Spined Stickleback, Pungitius pungitius L.
Michael Benjamin
Department of Anatomy, University College, Cardiff, Wales, U.K.
Swmnary. Pituitary cysts in the nine-spined stickleback, Pungitius pungitius L., develop next to blood vessels passing through the prolactin zone of the rostral pars distalis to the connective tissue capsule at its periphery. Cysts were most frequent when pituitaries were large compared with body lengths. However, the incidence of cysts could not be closely related to body length alone. As the rostral pars distalis is more highly vascularised in fish with large pituitaries, and as there was no evidence of accumulating secretion around these blood vessels, it is suggested that cysts develop when vascular demands (or supplies) become excessive. Associated with the greater vascularity of the rostral pars distalis in large pituitaries was a remarkable development of non-granulated cells. Indeed cysts may originate as enlarged intercellular spaces between such cells, as the latter often surround blood vessels. The non-granulated cells are also important in enlarging cyst cavities (by phagocytosing prolactin-cell debris) and perhaps for maintaining their structural integrity. It is suggested that the macrophages within cyst cavities are derived from the non-granulated cells.
Key words: Pituitary - Cyst - Prolactin cell - Non-granulated cell - Blood vessels.
There are large, fluid-filled cysts in the prolactin zone of the rostral pars distalis (RPD) of the nine-spined stickleback, Pungitius pungitius L. (Benjamin 1978, 1979; Benjamin and Williams 1979a, b). Their diameter varies from 6-200 gm and they are virtually restricted to animals > 20 mm in length. It is thus unlikely that they are of any embryological significance, as are the pituitary cysts in Xiphophorus maculatus (Schreibman 1966). Although present in freshwater animals at all times of the year, they are particularly common in June-August, i.e., towards the end, and just after, the breeding season (Benjamin and Williams 1979 a). As cysts are usually associated with small, inactive, prolactin cells, it seems unlikely that they are stores for prolactin released by holocrine secretion (Benjamin 1979).
Send offprint requests to: Dr. Michael Benjamin, Department of Anatomy, University College, Cardiff CF1 1XL, Wales, U.K.
0302-766 X/81/0214/0417/$02.80

418 M. Benjamin
A l t h o u g h the u l t ras t ruc ture o f p i tu i ta ry cysts has been descr ibed (Benjamin and Wi l l i ams 1979 b), the s tudy was rest r ic ted to large cysts visible at au topsy , with a low-power b inocu la r microscope . Thus we know little abou t the stages o f cyst fo rmat ion . The present l ight and e lect ron microscope s tudy repor ts an assoc ia t ion between b l o o d vessels o f the RPD, non -g ranu la t ed (stellate) cells, and the origin o f p i tu i t a ry cysts. I t a lso examines in detai l the re la t ionship between body length, p i tu i t a ry size and cyst incidence.
Materials and Methods
Nine-spined sticklebacks (Pungitius pungitius L.) of both sexes were collected from freshwater reens at Marshfield, Gwent, in September - February. The fish were measured from the tip of the snout to the base of the tail, and their lengths were recorded to the nearest millimetre. All fish were killed soon after collection.
Light and electron microscopy procedures for stickleback pituitaries have been described previously (Benjamin 1978; Benjamin and Williams 1979b). For light microscopy 6 btm serial sections of 340 pituitaries were stained with alcian blue - PAS - orange G. The size of the pituitary was estimated using a projection microscope at x 350 magnification. Every tenth section was traced on to good quality paper. The relative weights of the paper outlines reflected the relative volumes of the pituitary glands. In correlating cyst incidence with pituitary size, only small cysts not occupying more than one third of the RPD, were considered. However, in relating cyst incidence to body length, no pituitaries were excluded. The percentage of the RPD occupied by blood vessels was estimated by the point-counting method of Weibel (1969). The chosen test system consisted of a quadratic lattice of lines, whose cross-points served as markers for point-counting. On sections seen with a projection microscope, the distance between the lines was equivalent to 15.6 tam. The numerical data was assessed with Student's t-test.
The ultrastructural findings were based on a study of 20 pituitaries.
Results
The basic m o r p h o l o g y o f the p i tu i t a ry g land in Pungitius has been descr ibed prev ious ly (Benjamin 1978). The pro lac t in cells form a p rominen t mass domina t ing the R P D and lying ventra l to a smal ler g roup o f A C T H cells (Fig. 1). I t is within this p ro lac t in zone o f the p i tu i t a ry tha t cysts m a y form. They are fluid-fil led cavities tha t vary cons iderab ly in size (6-200 ktm diameter) and do no t usual ly conta in much solid, s ta inable mater ia l . Large cysts m a y expand into the A C T H zone and the p rox ima l pars distalis. A smal l cyst is i l lus t ra ted in Fig. 1 and a por t ion o f a large one in Fig. 11. The reader should consul t Benjamin (1979) for a l ight mic rog raph o f a large cyst.
The Relation Between Pituitary Cysts, Blood Vessels, and Non-granulated Cells. Thin-wal led b l o o d vessels pass f rom the neurohypophys i s , th rough or pos te r ior to the A C T H region, and into the pro lac t in zone (Fig. 2). Serial sections showed that they were con t inuous with b l o o d vessels a t the per iphery o f the R P D in the thin, connect ive tissue capsule. Nine ty five per cent o f p i tu i ta ry cysts were closely assoc ia ted with b lood vessels in the pro lac t in zone o f the RPD. The smaller cysts (and thus p r o b a b l y the newly- fo rmed ones) were usual ly next to those b l o o d vessels lying within the pro lac t in zone, ra ther than those in the connect ive tissue at its per iphery. B lood vessels were invar iab ly at the per imeter o f smal ler cysts (Figs. 2

The Origin of Pituitary Cysts in Pungit~us 419
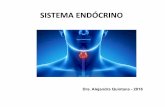
and 3), but also ran through the middle of larger ones. It seems likely that the central position of some blood vessels within larger cysts, either results from the fusion of smaller cysts associated with different blood vessels, or from the origin of cysts from adjacent regions of the same blood vessel (Fig. 7). Occasional cavities were packed with red blood cells (Fig. 8).
The blood vessels of the RPD typically consisted of a single layer of endothelial cells resting upon a basal lamina that was continuous with a similar structure extending away from the blood vessels and among the surrounding non-granulated cells (Fig. 9). The basal lamina may split and enclose surrounding pericytes (Fig. 9). Blood vessels associated with pituitary cysts were particularly thin-walled, and either lacked a basal lamina or shared one with the cyst lining (Fig. 10). There were enlarged intercellular spaces between neighbouring cells near the peripheral blood vessels of cysts (Fig. 10). Fenestrations in the endothelial cells were especially obvious in vessels associated with cysts, Such fenestrations were small pores, closed by a thin, membranous diaphragm (Fig. 10 inset). In the peripheral blood vessels of cysts, fenestrations were not restricted to the side facing the cyst cavity. There was no evidence of secretory-granule release from prolactin cells that were near the blood vessels of cysts. Neither were micro-pinocytotic vesicles conspicuous features of any endothelial cell. Both tight junctions and desmosomes were seen between adjacent endothelial cells in the vessel walls (Fig. 10).
Blood vessels at the periphery of the RPD, and those running through the region, were often surrounded by a sheath of chromophobic cells, From electron micrographs, it seems that such cells were probably non-granulated (stellate) cells, rather than partially de-granulated protactin cells (Fig. 9). They were identical to, and continuous with, the network o f non-granulated cells that ramified between the protactin cells. Their cytoplasm was pale in both light and electron micrographs. Although a few secretory granules were seen (Fig. 17), most cells were truly non- granular and characterised by an abundance of mitochondria and free ribosomes, occasional fragments of rough endoplasmic reticulum, groups of dense bodies (presumptive lysosomes), and a small Golgi apparatus of flattened cisternae and small vesicles (Figs. 15 and 17). Their nuclei were similar to those of the prolactin cells and were predominantly of eu-chromatin (Fig. 15). There could be elaborate in foldings of the plasma membranes of these non-granulated cells, parlicularly near blood vessels (Fig. 9), but also elsewhere. On occasions when such infoldings were less obvious, ~here were enlarged intercellular spaces betweer~ the ~on-granulated ceils, and between these and the prolactin cells (Fig. 15).
Thus, small cysts often lay next to blood vessels, and these frequently had sheaths of non-granulated cells. [t is not surprising therefore that a thin zone of non-granulated cells could separate the cyst cavity from the adjacent prolactin cells (Figs. 1, 4 and 11). In small, but not large cysts, many of the neighbouring prolactin cells were elongated and arranged radially with respect to the cyst cavity (Figs. 1, 2, 4 and 11). Particularly in large pituitaries, enormous collections of non-granulated cells could dominate the RPD and greatly reduce the numbers of prolactin cells (Fig. 5). Other pituitaries also had unusual numbers of non-granulated cells, but these were interspersed with prolactin cells and did not restrict the latter to the periphery (Fig. 6). It is perhaps significant that in many pituitaries with cysts, the remainder of~he prolactin zone also had highly consp~cueus non-granalated ce/ls.

420 M. Benjamin
Figs. 1-3, 5-8, 13. All these light micrographs are sagittal sections of the pituitary gland, stained with alcian blue-PAS-orange G, and photographed with a turquoise filter - unless otherwise stated
Fig. 1. The topographical relationship of the neurohypophysis (N) to the ACTH (A) and prolactin (P) cells. A small cyst (C) in the prolactin zone has a layer of non-granulated cells (NG) separating the cyst cavity from the adjacent prolactin cells. The immediately neighbouring prolactin cells are elongated, and arranged radially with respect to the cyst cavity (arrowheads). Calibration bar: 30 lim

The Origin of Pituitary Cysts in Pungitius 421
Few heal thy cells were ever seen within the cyst cavity when it was lined by a distinct membrane . Those present were phagocyt ic cells tha t rested on the basal lamina and were strikingly similar to non-granula ted cells, especially when the lat ter lay next to enlarged intercellular spaces (Figs. 14 and 15). The similarity is greatly s t rengthened by the finding o f non-granula ted cells in which numerous vacuoles conta ined dense masses resembling prolact in granules denuded o f their l imiting membrane . There were identical cells in the cyst cavity and in the lumina o f b lood vessels.
In some pituitaries with R P D cysts, occasional prolact in cells near the cyst cavity had very dense cytoplasm, poor ly defined nuclei, and swollen or burs t mi tochondr ia . Some o f these cells were phagocy tosed b y non-granu la ted cells (Fig. 16). Changes in mi tochondr i a m a y be a m o n g the earliest signs o f degenera t ion in prolact in cells, for enormous ly vacuola ted mi tochondr ia are very c o m m o n in otherwise heal thy- looking cells (Fig. 12). In the vast number o f pituitaries examined by light microscopy, only rarely were there large collections o f degenerat ing cells in the remaining R P D o f glands with cysts. These degenerat ing cells were packed with PAS-posi t ive droplets (Fig. 13). I t was impossible to decide whether they were prolact in cells, non-granu la ted cells or both . N o n e o f the cysts examined by light or electron microscopy were ever collapsed, i.e., they did not burs t and release their contained fluid.
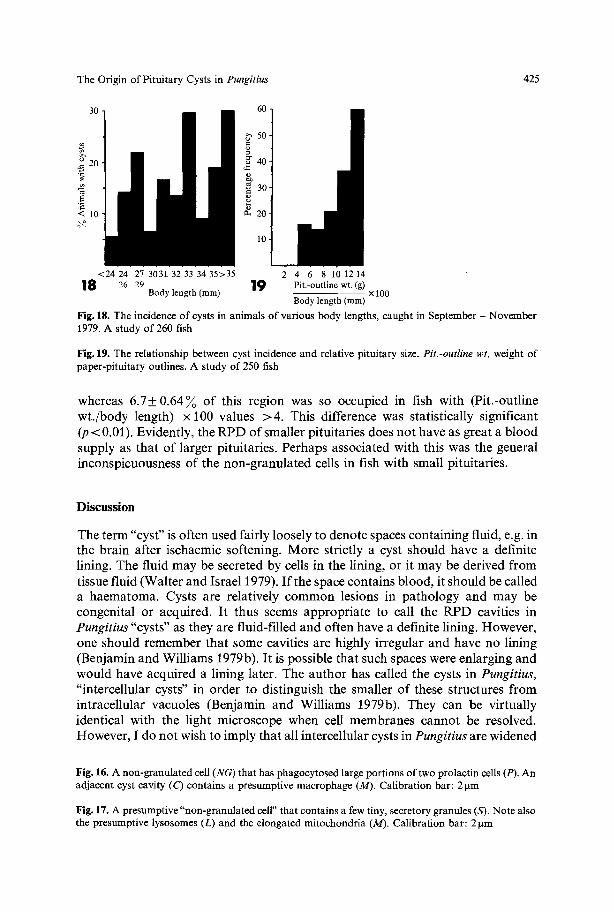
Cyst Incidence, Fish Length and Pituitary Size. In fish collected during Sep tember - November , there was no obvious t rend towards an increased incidence o f cysts in larger fish (Fig. 18), a l though they were virtually restricted to animals > 20 m m in length, as originally repor ted by Benjamin and Will iams (1979a). Cyst incidence was governed more clearly by pi tui tary size, as they were m o r e c o m m o n in animals whose pituitaries were large c o m p a r e d with their b o d y length (Fig. 19). I t should be noted f rom Fig. 19 tha t when the value for (Pit.-outline wt . /body length) • 100 fell below 4, no pituitaries had cysts, even though the fish were 17-32 m m in length. In such fish only 2 .7+ 0.67 ~ ( m e a n + S.E.) o f the R P D was occupied by b lood vessels,
Fig. 2. A blood vessel (BV) passing from the neurohypophysis (N), posterior to the ACTH (A) cell zone, has entered the region of prolactin cells (P) and is there associated with a small cyst (C). The blood vessel lies at the periphery of the cyst and the neighbouring prolactin cells are radially arranged. Calibration bar: 30tim
Fig.3. Blood vessels (BI/) lying at the periphery of two small cysts (C) among the prolactin cells (P). Calibration bar: 30 ~tm. Green filter
Pig. 4. The immediate neighbourhood of a small cyst cavity (C), such as that illustrated in Fig. 1. Noia- granulated cells (NG) separate the cyst cavity from radially arranged prolactin cells (P). Calibration bar: 1 ~tm
Fig. 5. An enormous collection of non-granulated cells (NG) dominating the prolactin zone (P) of the RPD. A ACTH cells. Calibration bar: 30~tm
Fig. 6. As in Fig. 5, large numbers of non-granulated cells (NG) dominate the RPD. In this very large pituitary, however, they are intermingled among the prolactin cells (P) and thus do not form one large, central mass. Calibration bar: 30 ~tm
422 M, Benjamin
Fig. 7. Two small cysts (arrows) arising from adjacent regions of a longitudinally sectioned blood vessel (BV). Calibration bar: 301am
Fig. 8. A cavity in the pralactin zane packed with red blood ceils (RBC). CaIibratlon bar: 30 ~xm

The Origin of Pituitary Cysts in Pungitius 423
Fig. 11. A survey electron micrograph to show a large cyst within the prolactin zone (P) of the RPD. The cyst cavity is lined throughout by a basement membrane and non-granulated cells (arrowheads), and contains an isolated macrophage (M). There is no evidence of radially arranged prolactin cells. B Blood vessel; C capsule marking the outside of the pituitary. Calibration bar: 5 lxm
Fig. 12. A vacuolated mitochondrion (M) in an otherwise healthy prolactin cell- note the well preserved rough endoplasmic reticulum (RER). Calibration bar: 1 ~tm
Fig. 13. Degenerating cells next to a cyst cavity (C). Some of these cells are packed with PAS-positive droplets (PAS). BV blood vessel; P healthy prolactin cells. Calibration bar: 30 ~tm. Green filter
Fig. 9. A blood vessel (BV) surrounded by a sheath of non-granulated cells (NG) within the prolactin zone (PRL) of the RPD. Cysts form in areas such as these. The endothelial cells (E) rest on a basal lamina (BL) that splits to enclose surrounding pericytes (P) and is continuous with a similar structure (arrow) extending among the non-granulated cells (NG). Note ~he elaborately infolded cell membranes (IM) of the non-granulated cells. Calibration bar: 2 ~tm
Fig. 10. A thin-walled blood vessel at the periphery of a cyst cavity (C). Only parts of its endothelial cells (E) are associated with a basal lamina (BL). This basal lamina is continuous with that lining the cyst cavity. Non-granulated cells (NG) near the blood vessel are separated by wide intercellular spaces (S). Note also the desmosome (D) between adjacent endothelial cells. Calibration bar: 2~tm. Inset: An enlargement of the area enclosed within the rectangle, to show fenestrations within the endothelial cells (arrows). Calibration bar: 2~tm

424 M. Benjamin
Fig, 14. Two presumptive macrophages (M) lying within a cyst cavity (C) that is lined by a basal lamina (B) Calibration bar: 2~tm
Fig, 15. Note the similarity between the non-gramflatcd cell (NG) separated from its neighbours by locally enlarged intercellular spaces (S), and lhe presumptive macrophage~ in Fig 14. G Golgi apparatus; M mitochan/ria; P prolactin ceil;/~E/~ rough endopiasmic reticulum. Calibration bar: 2 •m
The Origin of Pituitary Cysts in Pungitius 425
30
.= 20
7 < 10,
o=
<24 24 27 3031. 32 33 34 35>35 2 4 6 8 10 12 14 1 8 26 29 1 9 Pit.-outline wt. (g)
Body length (ram) x 100 Body length (ram)
Fig. 18. The incidence of cysts in animals of various body lengths, caught in September - November 1979. A study of 260 fish
Fig. 19. The relationship between cyst incidence and relative pituitary size. Pit.-outline wt. weight of paper-pituitary outlines. A study of 250 fish
whereas 6 .7_+0.64~ o f this region was so occupied in fish with (Pit.-outline wt . /body length) • 100 values >4 . This difference was statistically significant (p < 0.01). Evidently, the R P D o f smaller pituitaries does no t have as great a b lood supply as that o f larger pituitaries. Perhaps associated with this was the general inconspicuousness o f the non-granula ted cells in fish with small pituitaries.
Di scuss ion
The term "cyst" is often used fairly loosely to denote spaces containing fluid, e.g. in the brain after ischaemic softening. More strictly a cyst should have a definite lining. The fluid m a y be secreted by cells in the lining, or it may be derived f rom tissue fluid (Walter and Israel 1979). I f the space contains blood, it should be called a haematoma. Cysts are relatively c o m m o n lesions in pa thology and may be congenital or acquired. I t thus seems appropr ia te to call the R P D cavities in Pungitius "cysts" as they are fluid-filled and often have a definite lining. However , one should remember that some cavities are highly irregular and have no lining (Benjamin and Williams 1979b). It is possible that such spaces were enlarging and would have acquired a lining later. The author has called the cysts in Pungitius, "intercellular cysts" in order to distinguish the smaller o f these structures f rom intracellular vacuoles (Benjamin and Williams 1979b). They can be virtually identical with the light microscope when cell membranes cannot be resolved. However , I do no t wish to imply that all intercellular cysts in Pungitius are widened
Fig. 16. A non-granulated cell (NG) that has phagocytosed large portions of two prolactin cells (P). An adjacent cyst cavity (C) contains a presumptive macrophage (M). Calibration bar: 2 ~tm
Fig. 17. A presumptive "non-granulated cell" that contains a few tiny, secretory granules (S). Note also the presumptive lysosomes (L) and the elongated mitochondria (M). Calibration bar: 2 lam

426 M. Benjamin
intercellular spaces - merely that they often start that way. On purely light- microscopy evidence, it is difficult to say whether the large, blood-filled spaces seen on rare occasions in the RPD of Pungitius, are dilated, but intact blood vessels, true haematomas, or cysts that are the site of a secondary haemorrhage.
As the cysts in Pungitius have no mucous-secreting cells in their lining, their contents may be largely tissue fluid that leaks from the thin-walled and fenestrated, blood vessels, that are so typically associated with them. Thus they differ fundamentally from the pituitary cysts in Xiphophorus maculatus (Schreibman 1966), not only in being of no embryological significance (see Introduction), but also in the nature and origin of their fluid contents. In Xiphophorus, the cysts are lined by goblet cells that secrete mucus into the cavity. This is also the case with Lampetrafluviatilis (Bage and Fernholm 1975) and Myxine glutinosa (Fernhotm 1972).
One should also note that cysts in Pungitius have a more constant position than those in Xiphophorus (Schreibman 1966). They are generally restricted to the protactin zone of the RPD and usually form within the gland rather than at its periphery. In Xiphophorus they are found throughout the adenohypophysis and form within the surrounding connective tissue capsule. Pituitary cysts affect gonad development in Xiphophorus, but not in Pungitius (Benjamin 1980). This is undoubtedly associated with the high frequency of cysts in the PPD of Xiphophorus (Schreibman 1966). Whereas there is usually only one cyst per gland in Xiphophorus, there are often more in Pungitius, and large cysts may form by the confluence of smaller ones.
It has previously been shown (Benjamin and Williams 1979a) that cysts are virtually restricted to animals > 20 mm in length. To what extent therefore is the body length of larger fish a reliable indicator of cyst incidence ? The present results suggest it is a poor guide and that the size of the pituitary gland is far better. Cysts seem more common in animals with large pituitaries. One could justifiably ask whether pituitaries might be enlarged by the mere presence of cysts? In the case of the largest cavities, such as those illustrated by Benjamin and Williams (1979 b), this is undoubtedly so, for the fluid contents may cause the surrounding wall to bulge beyond the general confines of the pituitary. For this reason, pituitaries with large cysts were ignored in this part of the present study. Only pituitaries with small cysts, where at least two thirds of the RPD remained intact, were measured. As it is possible to induce cysts in laboratory animals without increasing the size of their pituitaries (Benjamin 1980), the present conclusions are probably valid.
Pituitary cysts in Pungitius commonly form around blood vessels. This is clear from the present work. It is therefore likely that cysts are more common in large pituitaries because the RPDs of such glands are more highly vascularised. The more blood vessels there are, the greater the number of potential sites for cysts to develop. Similarly, cysts may be absent from fish < 20 mm long, because these animals have small pituitaries that do not require a rich blood supply. It is unlikely that the development of cysts results in an increased blood supply to the RPD, as the cysts do not contain viable cells, other than macrophages. One can suggest that the RPD in large Pungitius pituitaries requires a rich blood supply, and that cysts develop later, either when vascular demands or vascular supplies finally become too great. It is difficult to decide which is more likely. Perhaps it is significant that similar cysts

The Origin of Pituitary Cysts in Pungitius 427
have never been reported in the closely related stickleback, Gasterosteus aculeatus, and that the RPD of this fish has few blood vessels (Leatherland 1970; Benjamin 1974). Thomas et al. (1978) have reported cysts in the endocrine pancreas of the hagfish, Myxine glutinosa, and regard them as products of an inadequate blood supply. Zelander and Kirkeby (1977) noticed a close association between blood vessels and ultimobranchial cysts in the guinea-pig thyroid, but did not comment on the significance of their finding.
Perhaps associated with the greater vascularity of the RPD in Pungitius than in Gasterosteus, is the remarkable development of what are assumed to be non- granulated (stellate) cells within the prolactin zone of some of the larger Pungitius pituitaries. Although Leatherland and Percy (1976) rightly regard the non- granulated cells of Gasterosteus as very characteristic components of the RPD, they can be more more conspicuous in Pungitius, Whereas most of these cells in Pungitius lack secretory granules other than those sequestered within lysosomes, they can have a few. It is not known whether such granules contain prolactin. This seems an important point, for perhaps non-granulated cells can be derived by the degranulation and transformation of endocrine cells, as suggested by Horvath et al. (1974).
The non-granulated cells are otherwise very similar to those in other fish (Leatherland 1970; Abraham 1971; Bage and Fernholm 1975; Leatherland and Percy 1976; Rawdon 1978; Schultz et al, 1979). Their function is unclear and several possibilities have been suggested. They may play a role in transporting substances to and from blood vessels (Leatherland and Percy 1976), or phagocytose degenerating cells (Farquhar et al. 1975; Bage and Fernholm 1975; Rawdon 1978), or aid in mechanical support within the pituitary (Cardell 1969; Schechter 1969). The present study and that of Benjamin and Williams (1979b) offer support for all three suggestions. The great increase in the blood supply to the RPD of large pituitaries in Pungitius is accompanied by an increase in the number of non- granulated cells, particularly in the immediate vicinity of the blood vessels. A similar enveloping of blood vessels by these cells has been noticed in the RPD of Gasterosteus (Leatherland and Percy 1976). Complex interdigitations of non- granulated cells, which would greatly increase the available surface for the transport of material, are particularly prominent near blood vessels in Pungitius pituitaries lacking cysts. When cysts are present, such infoldings are less obvious - perhaps because the RPD is breaking down. Indeed it is when cysts are present that non-granulated cells are obviously phagocytic, for they may enclose even large portions of prolactin cells within phagosomes. Thus, non-granulated cells play a role in cavity enlargement as well as cavity formation. The similarity between macrophages roaming the cyst cavity and the non-granulated cells in intact portions of the RPD, suggests that macrophages are derived from non-granulated cells, rather than from cells of the blood (Benjamin and Williams 1979b). Furthermore, the presence of phagocytic cells containing partially digested secretory granules, within the lumen of RPD blood vessels, must mean that such cells were being transported away from the RPD rather than towards it. Evidently, cell debris is not allowed to accumulate in large quantities within cyst cavities, for they are often devoid of solid material. This contrasts markedly with many reports of cysts in other animals, e.g., the parathyroid (Hatakeyema et al, 1970) and thyroid

428 M. Benjamin
(Christov et al. 1973) cysts of rats, the thyroid cyst of a dog (Booth and Ghoshal 1978), the parathyroid cysts of the house shrew, Suncus murinus (Swarup et al. 1978) and cysts in the endocrine pancreas of the hagfish (Thomas et al. 1978).
Some phagocytosis by non-granulated cells in Pungitius occurs outside cyst cavities, within the remaining RPD. It is proposed that cysts initially develop because fluid accumulates between non-granulated cells around blood vessels. They further enlarge because prolactin cells degenerate and are phagocytosed by non- granulated cells. These losen their attachments to neighbours and migrate towards blood vessels to be transported away from the pituitary, Occasional non-granulated cells move into the cyst cavity and act as macrophages to remove any debris discharged therein. As cysts are usually associated with small prolactin cel~s (Benjamin 1978, 1979; Benjamin and Williams 1979a), perhaps their shrinkage contributes to an enlargement of the perivascular spaces, and thus to the initiation of cysts, Cerlainly prolactin cells neighbouring small cyst cavities are frequently spindle-shaped, as if moving (or being moved) in relation to the cyst cavity. This contrasts directly with earlier findings (Benjamin and Williams 1979 b) - b u t these related mainly to larger cysts and perhaps this difference is significant. However the most likely explanation for the spindle-shape of the prolactin cells is that it is a consequence of cavity formation and not the cause. It seems a general principle that cavities impose a polarity on surrounding cells in glands (e,g,, pancreatic acini; RPD follicles m Anguilla), Benjamin and Williams (1979b) have previously discussed the similarities between RPD cysts in Pungitius and the secondary follicles or persistent oro-hypophysiaI ducts in other teleosts. The analogies are more striking when the shape of the neighbouring prolactin cells in small RPD cysts is also considered. The absence ofexoeytotic figures near the blood vessels of cysts, and the inactivity of the remaining prolactin cells (Benjamin 1979), argues against the idea that the cavities originate as"retention cysts" because of some selective blockage of prolactin release into the blood_ This contrasts with the ideas of Selye et al. (1964) on the origin of parathyroid cysts in the rat.
Only in non-granulated cells lining very large cysts are there conspicuous bundles of microfilaments (Benjamin and Williams 1979b). One may cautiously suggest that in large cysts there is little left to phagocytose, that transport of material between the remaining cells and blood vessels poses fewer problems, and that the non-granulated cells now serve mainly to lend structural stability to a large, unyieidy bag of fluid.
It is intriguing to note that cysts in Pungitius are primarily lesions of prolactin cells, yet they often originate within areas of non-granulated cells. Such cells seem eminently suited to lining a variety of fluid or colloid-filled cavities, e.g., the lumen of the ultimobranchial body in Salmo gairdneri (Hooker et al. 1979), the orohypophysial duct of Alosa pseudoharengus (Cook et al. 1973), follicles in the pars intermedia of Meriones unguiculatus (Bhattacharjee et al. 1980), the hypophysial cleft of the rat (Vanha-Perttula and Arstila 1970) or mouse (Dingemans and Feltkamp 1972) and cyst-like cavities in the endocrine pancreas of Myxine (Thomas et al. 1978). Falkmer etal , (1973) also remark on an inherent lumen-forming tendency of non-granulated cells.
Acknowledgements. I wish to thank Mrs. D. Hayman ~nd Mrs, G. O'ghea for their technical assislance.

The Origin of Pituitary Cysts in Pungitius 429
References
Abraham M (1971) The ultrastructure of the cell types and of the neurosecretory innervation in the pituitary of Mugil cephalus L. from fresh water, the sea, and a hypersaline lagoon. Gen Comp Endocrinol 17:334-350
Bage G, Fernholm B (1975) Ultrastructure of the pro-adenohypophysis of the river lamprey, Lampetra fluviatilis, during gonad maturation. Acta Zool (Stockh) 56:95-118
Benjamin M (1974) Seasonal changes in the prolactin cell of the pituitary gland of the freshwater stickleback, Gasterosteus aculeatus, form leiurus. CeU Tissue Res 152:93-102
Benjamin M (1978) Cytological changes in prolactin, ACTH, and growth hormone cells of the pituitary gland of Pungitius pungitius L. in response to increased environmental salinities. Gen Comp Endocrinol 36:48-58
Benjamin M (1979) Comparative ultrastructural studies on the prolactin ceils in the pituitary of nine- spined sticklebacks, Pungitius pungitius L., with and without adenohypophyseal cysts. Zoomorpho- logie 93:125-135
Benjamin M (1980) Factors associated with the seasonal variation in the incidence of pituitary cysts in nine-spirted sticklebacks, Pungitius pungitius L. Acta Zool (Stockh) (In Press)
Benjamin M, Williams JG (1979 a) Pituitary cysts in the nine-spined stickleback, Pungitiuspungitius L. I. Seasonal changes and salinity transfer experiments. Acta Zool (Stockh) 60:233-240
Benjamin M, Williams JG (1979b) Pituitary cysts in the nine-spined stickleback, Pungitiuspungitius L. I I. Light and electron microscopy. Acta Zool (Stockh) 60:241-250
Bhattacharjee DK, Chatterjee P, Holmes RL (1980) Follicles and related structures in the pars intermedia of the adenohypophysis of the jird (Meriones unguiculatus). J Anat (Lond.) 130:63-67
Booth KK, Ghoshal NG (1978)Atypical thyroid follicles arising from an ultimobranchial-like cyst in a postnatal canine thyroid gland. Acta Anat 102: 405-410
Cardell RR (1969) The ultrastructure of stellate cells in the pars distalis of the salamander pituitary gland. Am J Anat 126:429-456
Christov K, Bollmann R, Thomas C (1973) Ultimobranchial follicles and cysts in the rat thyroid during postnatal development. Beitr Path 149:47-59
Cook H, Rusthoven JJ, Vogelzang NJ (1973) The rostral pars distalis of the pituitary gland of the freshwater and marine alewife (Alosa pseudoharengus). A light and electron microscope study. Z Zellforsch 141 : 145-159
Dingemans KP, Feltkamp CA (1972) Non-granulated cells in the mouse adenohypophysis. Z Zellforsch 124:387-405
Falkmer S, Emdin S, Havu N, Lundgren G, Marques M, Ostberg Y, Steiner DF, Thomas NW (1973) Insulin in invertebrates and cyclostomes. Am Zool 13:625-638
Farquhar MG, Skutelsky EH, Hopkins CR (1975) Structure and function of the anterior pituitary and dispersed cells. In vitro studies. In: Tixier-Vidal A, Farquhar MG (eds) The anterior pituitary. Academic Press New York, p 83-137
Fernholm B (1972) The ultrastructure of the adenohypophysis of Myxine glutinosa. Z Zellforsch 132:451-472
Hatakeyama S, Tuchweber B, Blascheck JA, Garg BD, Kovacs K (1970) Parathyroid cyst formation induced by dihydrotachysterol and calcium acetate. An electron microscopic study. Endocrinol Jpn 17:355-363
Hooker WM, McMillan PJ, Thaete LG (1979) Ultimobranchial gland of the trout (Salmogairdneri) II. Fine structure. Gen Comp Endocrinol 38:275-284
Horvath E, Kovacs K, Penz G, Ezrin C (1974) Origin, possible function and fate of"follicular ceils" in the anterior lobe of the human pituitary. Am J Path 77:199-212
Leatherland JF (1970) Seasonal variation in the structure and ultrastructure of the pituitary of the marine form (Trachurus) of the threespine stickleback, Gasterosteus aculeatus L. 1. Rostral pars distalis. Z Zellforsch 104:301-317
Leatherland JF, Percy R (1976) Structure of the nongranulated cells in the hypophyseal rostral pars distalis of cyclostomes and actinopterygians. Cell Tissue Res 166:185 200
Rawdon BB (1978) Ultrastructure of the non-granulated hypophysial cells in the teleost Pseudocrenil- abrus philander (Hemihaplochromis philander) with particular reference to cytological changes in culture. Acta Zool (Stockh) 59:25-33
Schechter J (1969) The ultrastructure of the stellate cell in the rabbit pars distalis. Am J Anat 126: 477- 487

430 M. Benjamin
Schreibman MP (1966) Hypophysial cysts in a teleost fish. J Exp Zool 162:57-68 Schultz H J, Patzner RA, Adam H (1979) Fine structure of the agranular adenohypophysial cells in the
hagfish, Myxine glutinosa (Cyclostomata). Cell Tissue Res 204:67-75 Selye H, Ortega MR, Tuchweber B (1964) Experimental production of parathyroid cysts. Am J Path
45:251 256 Swarup K, Srivastav AK, Tewari NP (1978) Occurrence ofcalcitonin cells and cysts in the parathyroid of
the house shrew, Suncus murinus. Acta Anat 101:340-345 Thomas NW, Ostberg Y, Falkmer S (1978) Cell degeneration and cavity formation in the endocrine
pancreas of the hagfish Myxine glutinosa. Acta Zool (Stockh) 59:119 123 Vanha-Perttula T, Arstila AU (1970) On the epithelium of the rat pituitary residual lumen. Z Zellforsch
108:487-500 Walter JB, Israel MS (1979) General Pathology. 5th edition. Churchill, Livingstone London New York Weibel ER (1969) Stereological principles for morphometry in electron microscopic cytology. Int Rev
Cytol 26:235-302 Zelander T, Kirkeby S (1977) Fine structure of the ultimobranchial cysts in the thyroid of the adult
guinea pig. Cell Tissue Res 183:343-351
Accepted September 24, 1980