
THE OF CHEMISTRY Vol. No. Issue June 15, pp. … JOURNAL OF BIOLOGICAL CHEMISTRY Q 1986 by The...
5
THE JOURNAL OF BIOLOGICAL CHEMISTRY Q 1986 by The American Society of Biological Chemists, Inc. Vol. 261, No. 17, Issue of June 15, pp. 7621-7625,1986 Printed in U.S.A. Transcriptional and Post-transcriptional Regulation of L-type Pyruvate Kinase Gene Expression in Rat Liver* (Received for publication, December 26, 1985) Sophie Vaulont, Arnold Munnich, Jean-Franqois Decaux, and Axel Kahn From the Laboratoire de Recherche en Ginitigue etPathologie Moliculaires, Institut National de la Sante et de la Recherche Medicale U 129, Chu Cochin, 24 Rue du Faubourg St. Jacques, 75674 Paris, Cedex 14, France The effects of starvation, refeeding a diet high in carbohydrate, administration of glucagon and cyclic AMP, thyroidectomy, and adrenalectomy on transcrip- tion of the gene for liver L-type pyruvate kinase and on the accumulation of cytoplasmic mRNA for L-type pyruvate kinase were investigated in rat. Transcription of the gene was undetectable in either fasted or protein-fed rats. Refeeding fasted rats a car- bohydrate-rich diet stimulated an increase in L-type pyruvate kinase mRNA, preceded by an increase in the gene transcription. Transcription was maximal at 12 h of refeeding, decreasing to 10% of maximum at 72 h. The level of L-type pyruvate kinase mRNA remained constant at 50% of maximum for at least 120 h. Neither thyroidectomy nor adrenalectomy affectedgene tran- scription in fasted rats refed the carbohydrate-rich diet, despite a decrease in mRNA abundance to 40 and 20%, respectively, of controls fed a normal diet. Glu- cagon or cyclic AMP totally blocked the increase in transcription of the L-type pyruvate kinase gene caused by feeding a carbohydrate-rich diet to previ- ously fasted rats. Nevertheless, the level of L-type pyruvate kinase mRNA remained high for 3 h after glucagon administration.After 3 h, the mRNA de- creased rapidly with a half-life less than l h. Thus, expression of the gene for L-type pyruvate kinase is regulated at both transcriptional and post- transcriptional levels. The transcription is regulated by two major effectors, one positive, namely carbohy- drates, and one negative, namely glucagon (via cyclic AMP). Both agents probably act at the level of the mRNA stability as well. Glucocorticoids and thyroid hormones do not regulate transcription of the gene for L-type pyruvate kinase but do appear to be required for a normal accumulation of the transcripts in the cytoplasm. L-type pyruvate kinase (ATPpyruvate 02-phosphotrans- ferase, EC 2. 7. 1. 40), the predominant pyruvate kinase isozyme in liver, is regarded as a key enzyme in the glycolytic pathway. The effects of both dietary and hormonal agents on enzyme activity inthe liver have been studied extensively (1- 10). During fasting, the activity of the enzyme rapidly de- creases; it is restored by refeeding a carbohydrate-rich diet (5). The increase in activity is due to an increase in the rate of enzyme synthesis which, in turn, is proportional to an * This work was supported by Grants 955 and 1971 from Action Thkmatique Programme6 Centre National de la Recherche Scienti- fique. The costs of publication of this articlewere defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. increase in the amount of functional L-type pyruvate kinase mRNA (11). We have used a cDNA probe complementary to rat L-type pyruvate kinase mRNA (12) to demonstrate that feeding a carbohydrate-rich diet to fasted animals causes a 40-100-fold increase in the L-type pyruvate kinase mRNAs in the liver (11). All three species of L-type pyruvate kinase mRNA, 3.2, 2.2, and 2 kb,’ accumulate to about the same extent. These different mRNA species have the same 5‘ end and coding sequence and differ by the length of their 3‘-untranslated extension.* Our in uiuo studies suggest that the maximum stimulation of accumulation of these mRNAs by glucose re- quires at least two hormonal features: decreased glucagon levels and normal levels of the permissive hormones, insulin, glucocorticoids, and thyroid hormones (13, 14). However, the level and mechanism of the regulation of the gene expression remain unknown. In order to determine the respective roles of transcriptional and post-transcriptional events in the dietary and hormonal regulation of L-PK mRNA accumulation, we compared changes in the rate of gene tran- scription in isolated nuclei to changes in the level of the specific mRNA in the liver of fasted and carbohydrate-fed rats. The results show that in vivo control of expression of the L-PK gene is both transcriptional and post-transcrip- tional. The transcriptional control is either positive, mediated by carbohydrates, or negative, mediated by glucagon via cyclic AMP. In addition, in rats refed carbohydrates, L-PK mRNA appears to be very stable, whereas in glucagon-treated rats, it is unstable. Glucocorticoids andthyroid hormones arein- volved at a post-transcriptional level: ablation of these glands has no effect on gene transcription but decreases mRNA abundance in liver. EXPERIMENTAL PROCEDURES Materials Chemicals of the highest purity available were from Merck, Serva, Sigma, and Boehringer Mannheim. Cortisone acetate was from Rous- sel; 3,3’,5-triiodo-~-thyronine and iV,@’-dibutyryl cyclic AMP were from Sigma. Long-acting glucagon (zinc-glucagon and zinc-prota- mine-glucagon)was a gift from Novo Ind. Nitrocellulosepaper (BA85) was obtained from Schleicher & SchuelI. Nylon GeneScreen Plus Filters were from New England Nuclear. Formaldehyde was from Fluka, and methylmercury hydroxide from Ventron Alpha Laboratory [~I-~~PI~CTP (3,000 Ci/mmol) and [CY-~~P]UTP (>400 Ci/mmol) were supplied by Amersham Corp., and Biofluor by New England Nuclear. Restriction enzymes and DNA polymerase large fragment according to Klenow were obtained from Amersham Corp. Royal X-Omat AR film was from Eastman-Kodak. ~~ ‘The abbreviations used are: kb, kilobase pair; L-PK, L-type pyruvate kinase. Marie, J., et al. (1986) Eur. J. Biochern., in press. 7621
-
Upload
dinhkhuong -
Category
Documents
-
view
216 -
download
0
Transcript of THE OF CHEMISTRY Vol. No. Issue June 15, pp. … JOURNAL OF BIOLOGICAL CHEMISTRY Q 1986 by The...

THE JOURNAL OF BIOLOGICAL CHEMISTRY Q 1986 by The American Society of Biological Chemists, Inc.
Vol. 261, No. 17, Issue of June 15, pp. 7621-7625,1986 Printed in U.S.A.
Transcriptional and Post-transcriptional Regulation of L-type Pyruvate Kinase Gene Expression in Rat Liver*
(Received for publication, December 26, 1985)
Sophie Vaulont, Arnold Munnich, Jean-Franqois Decaux, and Axel Kahn From the Laboratoire de Recherche en Ginitigue et Pathologie Moliculaires, Institut National de la Sante et de la Recherche Medicale U 129, Chu Cochin, 24 Rue du Faubourg St. Jacques, 75674 Paris, Cedex 14, France
The effects of starvation, refeeding a diet high in carbohydrate, administration of glucagon and cyclic AMP, thyroidectomy, and adrenalectomy on transcrip- tion of the gene for liver L-type pyruvate kinase and on the accumulation of cytoplasmic mRNA for L-type pyruvate kinase were investigated in rat.
Transcription of the gene was undetectable in either fasted or protein-fed rats. Refeeding fasted rats a car- bohydrate-rich diet stimulated an increase in L-type pyruvate kinase mRNA, preceded by an increase in the gene transcription. Transcription was maximal at 12 h of refeeding, decreasing to 10% of maximum at 72 h. The level of L-type pyruvate kinase mRNA remained constant at 50% of maximum for at least 120 h. Neither thyroidectomy nor adrenalectomy affected gene tran- scription in fasted rats refed the carbohydrate-rich diet, despite a decrease in mRNA abundance to 40 and 20%, respectively, of controls fed a normal diet. Glu- cagon or cyclic AMP totally blocked the increase in transcription of the L-type pyruvate kinase gene caused by feeding a carbohydrate-rich diet to previ- ously fasted rats. Nevertheless, the level of L-type pyruvate kinase mRNA remained high for 3 h after glucagon administration. After 3 h, the mRNA de- creased rapidly with a half-life less than l h.
Thus, expression of the gene for L-type pyruvate kinase is regulated at both transcriptional and post- transcriptional levels. The transcription is regulated by two major effectors, one positive, namely carbohy- drates, and one negative, namely glucagon (via cyclic AMP). Both agents probably act at the level of the mRNA stability as well. Glucocorticoids and thyroid hormones do not regulate transcription of the gene for L-type pyruvate kinase but do appear to be required for a normal accumulation of the transcripts in the cytoplasm.
L-type pyruvate kinase (ATPpyruvate 02-phosphotrans- ferase, EC 2. 7. 1. 40), the predominant pyruvate kinase isozyme in liver, is regarded as a key enzyme in the glycolytic pathway. The effects of both dietary and hormonal agents on enzyme activity in the liver have been studied extensively (1- 10). During fasting, the activity of the enzyme rapidly de- creases; it is restored by refeeding a carbohydrate-rich diet ( 5 ) . The increase in activity is due to an increase in the rate of enzyme synthesis which, in turn, is proportional to an
* This work was supported by Grants 955 and 1971 from Action Thkmatique Programme6 Centre National de la Recherche Scienti- fique. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
increase in the amount of functional L-type pyruvate kinase mRNA (11).
We have used a cDNA probe complementary to rat L-type pyruvate kinase mRNA (12) to demonstrate that feeding a carbohydrate-rich diet to fasted animals causes a 40-100-fold increase in the L-type pyruvate kinase mRNAs in the liver (11). All three species of L-type pyruvate kinase mRNA, 3.2, 2.2, and 2 kb,’ accumulate to about the same extent. These different mRNA species have the same 5‘ end and coding sequence and differ by the length of their 3‘-untranslated extension.* Our in uiuo studies suggest that the maximum stimulation of accumulation of these mRNAs by glucose re- quires at least two hormonal features: decreased glucagon levels and normal levels of the permissive hormones, insulin, glucocorticoids, and thyroid hormones (13, 14).
However, the level and mechanism of the regulation of the gene expression remain unknown. In order to determine the respective roles of transcriptional and post-transcriptional events in the dietary and hormonal regulation of L-PK mRNA accumulation, we compared changes in the rate of gene tran- scription in isolated nuclei to changes in the level of the specific mRNA in the liver of fasted and carbohydrate-fed rats. The results show that in vivo control of expression of the L-PK gene is both transcriptional and post-transcrip- tional. The transcriptional control is either positive, mediated by carbohydrates, or negative, mediated by glucagon via cyclic AMP. In addition, in rats refed carbohydrates, L-PK mRNA appears to be very stable, whereas in glucagon-treated rats, it is unstable. Glucocorticoids and thyroid hormones are in- volved at a post-transcriptional level: ablation of these glands has no effect on gene transcription but decreases mRNA abundance in liver.
EXPERIMENTAL PROCEDURES
Materials Chemicals of the highest purity available were from Merck, Serva,
Sigma, and Boehringer Mannheim. Cortisone acetate was from Rous- sel; 3,3’,5-triiodo-~-thyronine and iV,@’-dibutyryl cyclic AMP were from Sigma. Long-acting glucagon (zinc-glucagon and zinc-prota- mine-glucagon) was a gift from Novo Ind. Nitrocellulose paper (BA85) was obtained from Schleicher & SchuelI. Nylon GeneScreen Plus Filters were from New England Nuclear. Formaldehyde was from Fluka, and methylmercury hydroxide from Ventron Alpha Laboratory [ ~ I - ~ ~ P I ~ C T P (3,000 Ci/mmol) and [CY-~~P]UTP (>400 Ci/mmol) were supplied by Amersham Corp., and Biofluor by New England Nuclear. Restriction enzymes and DNA polymerase large fragment according to Klenow were obtained from Amersham Corp. Royal X-Omat AR film was from Eastman-Kodak.
~~
‘The abbreviations used are: kb, kilobase pair; L-PK, L-type pyruvate kinase.
Marie, J., et al. (1986) Eur. J. Biochern., in press.
7621

7622 Regulation of L-type Pyruvate Kinase Gene Expression
Animak For the dietary studies, 3-month-old male Wistar rats weighing
approximately 250 g were starved for 72 h. Then, the animals were given a solid diet containing 65% sucrose and water containing 200 g/liter fructose for various periods of time ranging from 7 to 120 h.
For the starvation studies, rats refed the high-carbohydrate diet for 12-18 h were deprived of food for 48 h.
Some animals underwent bilateral adrenalectomy or thyroparathy- roidectomy under pentobarbital anesthesia, as previously described (13, 15). Plasma corticosterone in adrenalectomized animals was 1.4 f 0.6 ng/ml (normal: 70 zk 0.6 ng/ml). Thyroidectomized animals were retained for experiments when their plasma thyrostimulating hormone exceeded 4 microunits/ml (normal: <2 microunits/ml). Some thyroidectomized rats received one single dose of triiodo-L- thyronine (50 pg) intraperitoneally.
The time course of the effect of glucagon was studied in rats submitted to a nycterohemeral inversion for 6 weeks. Since rats usually do not eat during the day, inversion was planned to ensure they would eat during glucagon administration. Rats were injected at 8 a.m. intraperitoneally with both rapid glucagon (500 pg/kg) + zinc- glucagon (7.5 mg/kg) and subcutaneously with long-acting glucagon (zinc-protamine-glucagon, 7.5 mg/kg) in order to obtain a rapid and sustained hormonal impregnation over the refeeding period (16).
The dose-response effect of glucagon was studied in rats refed the carbohydrate-rich diet for 18 h. The animals were given various doses of glucagon * insulin or dibutyryl cyclic AMP intravenously via the cannulated jugular vein as previously described (13).
Rats were killed by decapitation, and blood was taken for plasma glucose and hormone determination. Food ingestion to the stomach and gut was checked by post-mortem examination, and the whole liver was removed, weighed, and homogenized for subsequent purifi- cation of nuclei and RNA.
General Methods RNA Isolation and Quantitation-Total cellular RNAs were iso-
lated from rat liver by the modified guanidium chloride procedure (12, 13, 17). Integrity of the RNA preparations was analyzed system- atically by electrophoresis and visualization of 18 S and 28 S ribo- somal RNAs by ethidium bromide staining; the ratio of 28 S to 18 S was always close to 2 without any detectable low molecular weight forms.
Nuclear RNAs were obtained from isolated nuclei using the same procedure. Nuclei were prepared by a modification of the citric acid procedure (18). One gram of liver was homogenized in 10 ml of ice- cold 5% citric acid in a motor-driven Potter-Elvehjem homogenizer. The homogenate was filtered through two layers of cheese-cloth and centrifuged at 600 X g for 20 min. The nuclear pellet was suspended in 2 ml of 1.5% citric acid, 0.25 M sucrose; layered onto a 2-ml cushion of 1.5% citric acid, 1 M sucrose; and centrifuged at 1,000 X g for 20 min at 4 "C. This step was repeated twice. Then, the nuclear pellet was washed in 4 ml of 1.5% citric acid, 0.25 M sucrose, 0.5% Triton X-100 and centrifuged at 600 X g for 10 min at 4 "C. This procedure yielded approximately 10' nuclei/g of liver, wet weight.
Subsequent analysis and quantitation of L-PK RNA was per- formed using either Northern blot or dot-blot procedure. Nuclear and cytoplasmic RNAs were electrophoresed in vertical methylmercury hydroxide-agarose gels (13, 19) and then transferred to Genescreen Plus nylon filters as previously described (12, 13). In some experi- ments, the intensity of the mRNA bands detected on the Northern blot was measured by scanning the autoradiograms using a Shimadzu densitometer. Cytoplasmic RNA quantitation was performed by dot- blot hybridization and subsequent assay of the individual dots for radioactivity by liquid scintillation spectrometry (11, 13).
cDNA Probe and Hybridization-cDNA probes for rat liver L-type pyruvate kinase have been cloned in our laboratory (12). In the liver, one of the L-type pyruvate kinase cDNA clones (clone llC6) recog- nizes three distinct mRNAs of 3.2, 2.2, and 2 kb, whereas another recombinant clone (clone 2B8) hybridizes with the 3.2-kb transcript only (12). A 388-base pair TqI-PstI restriction fragment of llC6, corresponding to the 144 last coding nucleotides and the first 244 3'- noncoding nucleotides of llC6, was subcloned into the single-strand phage M13 (20). The recombinant phage was used as template for the synthesis of the complementary strand in the presence of deox- ynucleotide including a limiting amount of [ L U - ~ ~ P I ~ C T P . After strand separation, a highly labeled probe was obtained (21): Specific activity
DeKeyzer et al., personal communication.
was about lo9 cpm/pg DNA. The 2B8-L-PK cDNA probe consisted of purified cDNA inserts labeled by nick translation as described (13) a t a specific activity of 2 X 10' cpmlpg DNA. The filters were hybridized as previously described (13) at 65 "C in 3 X SSC (1 X SSC = 0.15 M NaC1, 0.015 M Tris sodium citrate) with 3 X lo6 cpm/ml probe and 0.5 ml/l0-cmz hybridization mixture. Final wash was at 65 "C in 0.2 X SSC, 1% sodium dodecyl sulfate.
In Vitro Transcription and Hybridization-The elongation of nas- cent RNA transcripts on the L-PK gene was measured by a "run-on" transcription assay in isolated nuclei. This experiment is a means to investigate the in vivo rate of transcription initiation of a specific gene at the time the animal is killed. The in vitro initiation of new transcripts in isolated nuclei is, in fact, inefficient (22, 23) so that labeling of RNAs only reflects in vitro elongation of RNA molecules whose synthesis has been initiated in vivo under the metabolic con- ditions studied.
Isolation of nuclei, in vitro nuclear transcription, purification of 32P-labeled RNAs, hybridization to Gene Screen Plus filter-bound cDNA, and elution of specific radioactivity from the filters were performed as described (14, 24). Transcription rates, expressed as ppm, were calculated from the following equation: specific cpm eluted from the filters/total cpm of radiolabeled RNA X (lOO/hybridization efficiency) X the length of the gene/the length of the cDNA insert in base pairs.
This corrects for the size of the cDNA probe relative to gene length and for the efficiency of hybridization which was on average 40%. The L-PK gene was 10 kb long4, and we have used three overlapping cDNA clones covering 2 kb of the mRNA sequence.
RESULTS AND DISCUSSION
Analysis of L-PK Nuclear Transcripts in Fasted and Car- bohydrate-fed Rats-Fig. l shows a Northern blot analysis of hepatic nuclear RNAs. In addition to the 3.2-kb mature form of L-PK mRNA, two 4.2- and 4.5-kb species, absent in liver of fasted rats, were detected by 7-12 h of refeeding and disappeared by 24 h of refeeding while the mature mRNA accumulated. The two high molecular weight RNA species are likely precursors which are being processed to form the mature messenger RNA. It is unlikely that the 4.5-kb nuclear species represents the unspliced primary transcript since the size of the L-PK gene is about 10 kb.4 These results are consistent with transcriptional activation of the L-PK gene or with
Kb
.4.5 -42 '3.2
1 2 3 4 5 FIG. 1. Northern blot analysis of cytoplasmic and nuclear
L-PK RNA in the liver of fasted and carbohydrate-fed rats. Liver nuclear RNAs (20 pg) from 48-h fasted rats (lane I); fasted rats refed the high-carbohydrate diet for 9 h (lane 21, 12 h ( l a n e 3 ) , or 24 h ( l a n e 4); and cytoplasmic RNA (4 pg) from fasted rats refed the carbohydrate-rich diet ( l a n e 5) were electrophoresed in 10 mM meth- ylmercury hydroxide-agarose gels, transferred to nylon filters, and hybridized with a nick-translated [32P]-labeled 2B8-L-PK cDNA probe (3 X lo6 cpm/ml). This probe is complementary to the extreme 3' end of the 3.2-kb L-PK mRNA and does not recognize the lighter 2- and 2.2-kb species (11,12). Autoradiography was performed for 12 h with intensifying screens at -80 "C.
M. Cognet, et al., manuscript in preparation.

Regukation of L-type Pyruvate Kinase Gene Expression 7623
increased stability of the RNA precursors in rats fed the carbohydrate-rich diet. Therefore, a direct assessment of gene transcription was undertaken.
Determination of L-PK Gene Transcription in Fasted, Pro- tein-fed, and Carbohydrate-fed Rats-Our measurements of transcription indicate an undetectable rate of transcription of the L-PK gene in the liver of fasted or protein-fed rats, accounting for the absence of precursor RNAs in their nuclei. Refeeding fasted animals with a carbohydrate-rich diet stim- ulates a rapid increase in the rate of transcription (Fig. 2). The maximum rate is not achieved until 12 h after refeeding is initiated. The lag period can signify that transcriptional stimulation is not a direct effect of carbohydrates or of insulin whose secretion is quickly stimulated by carbohydrates (25), but rather needs complex metabolic modifications which are secondary consequences of the carbohydrate feeding. Tran- scription decreases after 18-24 h and is only one-fifth to one- tenth of maximum at 72 h of refeeding. This low rate of gene transcription is sufficient to maintain a level of L-PK mRNA which is about 50% of maximum for at least 120 h. Thus, in addition to their transcriptional effect, carbohydrates may decrease the rate of L-PK mRNA degradation, as recently suggested by Goldman et al. (26) for avian malic enzyme mRNA. The carbohydrate-rich diet increases the levels of translatable mRNAs for glucokinase (27), glucose-6-P dehy- drogenase (28), 6-phosphogluconate dehydrogenase (28), malic enzyme (29), and fatty acid synthetase (30). When cloned cDNA probes have been used, these increases in mRNA translational activities paralleled increases in the levels of cytoplasmic mRNAs (31-34). Finally, in a limited number of instances, carbohydrate diets have been shown to increase transcription of some genes (26,35). This is the case for the pyruvate kinase gene. It is, however, difficult to determine from these experiments whether decreased gluca- gon secretion, increased insulin secretion, or changes in the levels of some glucidic metabolite(s) is the mechanism of this transcriptional activation.
A mixed carbohydrate diet was used in these experiments because full induction of L-PK mRNA is obtained with rats refed either a maltose-rich or a fructose-rich diet (13). How- ever, in fasted normal rats, glucose is a more powerful stim- ulator of transcription than fructose, suggesting that post- transcriptional events may allow fructose to stimulate mRNA accumulation to the same extent as glucose? The level of L-
- 0 12 24 48 72 120
7ime of catt~ohydrate mfeeding (hours) FIG. 2. Time course effect of carbohydrate refeeding on the
level of L-PK gene transcription and mRNA content in fasted rat liver. The L-PK gene transcription (o"--o) and mRNA abun- dance (0- - -0) were measured in the liver of animals refed the carbohydrate-rich diet for various periods of time ranging from 0 to 120 h after a 48-h starvation. Data are the mean of two distinct experiments. They are expressed as a percentage of the maximum values. The results obtained in protein-fed rats where the same as those shown here for fasted rats.
A. Munnich et al., manuscript in preparation.
PK mRNA was increased about 5-fold in the liver of diabetic rats fed fructose, whereas glucose feeding had no effect (36). Interestingly, in diabetic animals, induction of L-PK mRNA by fructose was not associated with an increase in transcrip- tion (37). In these diabetic rats fed a dextrin-containing diet, insulin strongly increased gene transcription by a probably indirect mechanism (37). These results emphasize the post- transcriptional effect of fructose on L-PK gene expression in diabetic rats and parallel those reported above for fasted normal rats. The mechanisms of stimulation of L-PK gene expression by fructose alone and glucose + insulin are clearly different. The relative roles of glucose and insulin in the control of L-PK gene expression are difficult to determine using in vivo models because insulin must be administered with carbohydrates and because insulin secretion is regulated by glucagon and glucagon secretion by insulin in normal rats. In vitro models are needed to determine whether insulin is an activator of L-PK gene expression by itself or is permissive for stimulation by glucose.
Glucagon and cAMP Effects on L-PK Gene Transcription and mRNA Leuel-Glucagon, in addition to its role in inhib- iting L-PK enzyme activity, also decreases the amount of L- PK mRNA (13) by inhibiting gene transcription (14). This transcriptional inhibition by glucagon occurs at both high doses (250 pg/rat) and low doses (1 pg/rat) (Table I), sug- gesting that in fasting, feeding protein-rich diets, or diabetes, hyperglucagonemia plays a major role in regulating gene tran- scription. Moreover, glucagon inhibits gene transcription even if administered together with insulin (Table I). So far, all of our experiments suggest that insulin cannot counteract the inhibitory effect of glucagon. As expected, CAMP (5 mg of dibutyryl cAMP/rat) has the same effect on L-PK gene tran- scription as glucagon (Table I). The hypothesis that glucagon (via synthesis of its second messenger, cyclic AMP) is a dominant regulator of L-PK gene expression whose effect is not easily reversed by carbohydrates and insulin was first proposed by Munnich et al. (13). Noguchi et al. (37) have confirmed this hypothesis, indicating in addition that the insulin effect is a relatively delayed phenomenon requiring active protein synthesis and may be indirect.
The inhibitory effect of glucagon on L-PK gene transcrip- tion is rapid (it is complete 15 min after glucagon administra- tion), suggesting a "direct effect" of cAMP or of a CAMP- dependent protein kinase on a transcriptional factor. Fig. 3 shows the time course effect of glucagon on the level of the 3.2-kb mRNA species as compared to the level of gene tran- scription. No significant decrease of L-PK mRNA is observed 1 h after glucagon administration, whereas gene transcription is already completely inhibited. The mRNA level remains
TABLE I Dose-response effect of glucagon administration on L-PK gene
transcription in the liver of 18-h carbohydrate-fed rats, eithm in the presence or in the absence of insulin
Regular glucagon, dibutyryl cyclic AMP, or insulin was adminis- tered intravenously to 18-h carbohydrate-fed rats. The animals were killed after 20 min, and their livers were immediately removed for nuclei extraction.
n" L-PK gene % of transcription control
PPm Carbohydrate-fed controls Fed + glucagon (1 pg)
4 121 f 21 3 60.5 f 3.5 50.0
Fed + glucagon (1 pg) 3 55.3 + 10 45.7 + insulin (2 units) Fed + glucagon (250 pg) 3 5 + 3 4.1 Fed + dibutyryl cAMP (5 mg) 1 ND ND n, number of animals; ND, not detectable.
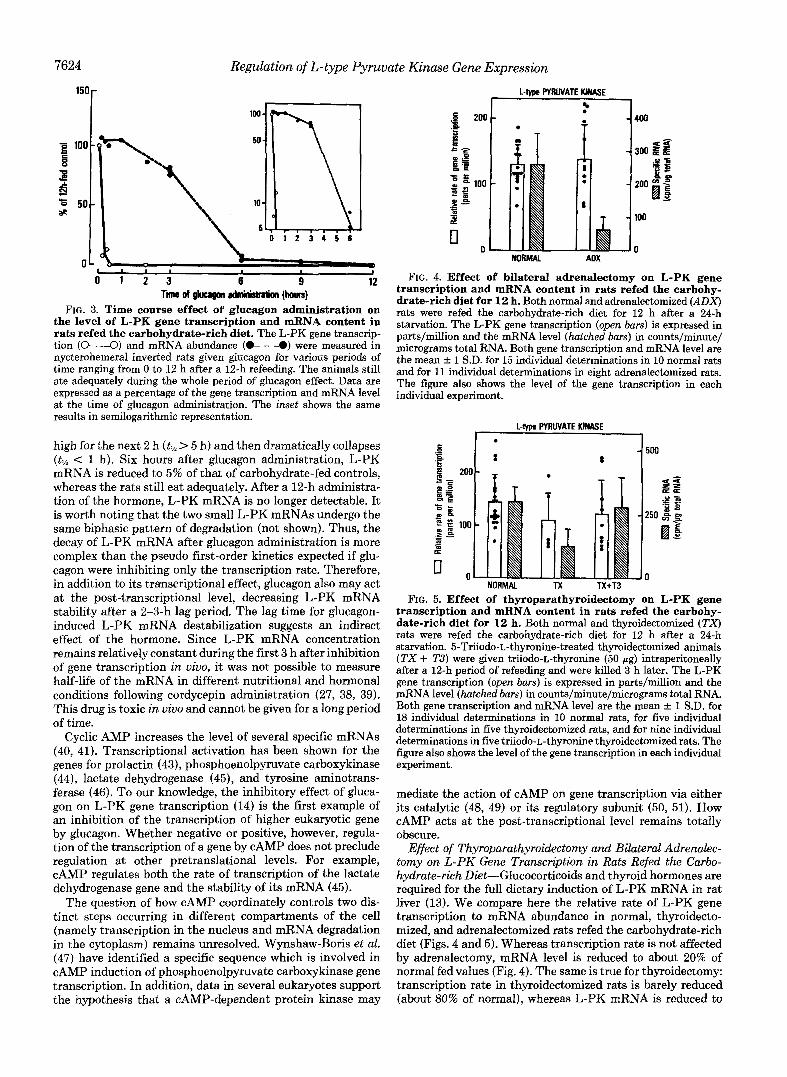
7624 Regulation of L-type Pyruvate Kinase Gene Expression
150r
0 0 1 2 3 6 9 12
Time of glucagon ahnhimh {huts)
FIG. 3. Time course effect of glucagon administration on the level of L-PK gene transcription and mRNA content in rats refed the carbohydrate-rich diet. The L-PK gene transcrip- tion ( O " - - O ) and mRNA abundance (0- - -@) were measured in nycterohemeral inverted rats given glucagon for various periods of time ranging from 0 to 12 h after a 12-h refeeding. The animals still ate adequately during the whole period of glucagon effect. Data are expressed as a percentage of the gene transcription and mRNA level at the time of glucagon administration. The inset shows the same results in semilogarithmic representation.
high for the next 2 h (tIh > 5 h) and then dramatically collapses (tlh < 1 h). Six hours after glucagon administration, L-PK mRNA is reduced to 5% of that of carbohydrate-fed controls, whereas the rats still eat adequately. After a 12-h administra- tion of the hormone, L-PK mRNA is no longer detectable. It is worth noting that the two small L-PK mRNAs undergo the same biphasic pattern of degradation (not shown). Thus, the decay of L-PK mRNA after glucagon administration is more complex than the pseudo first-order kinetics expected if glu- cagon were inhibiting only the transcription rate. Therefore, in addition to its transcriptional effect, glucagon also may act at the post-transcriptional level, decreasing L-PK mRNA stability after a 2-3-h lag period. The lag time for glucagon- induced L-PK mRNA destabilization suggests an indirect effect of the hormone. Since L-PK mRNA concentration remains relatively constant during the first 3 h after inhibition of gene transcription i n vivo, it was not possible to measure half-life of the mRNA in different nutritional and hormonal conditions following cordycepin administration (27, 38, 39). This drug is toxic in vivo and cannot be given for a long period of time.
Cyclic AMP increases the level of several specific mRNAs (40, 41). Transcriptional activation has been shown for the genes for prolactin (43), phosphoenolpyruvate carboxykinase (44), lactate dehydrogenase (45), and tyrosine aminotrans- ferase (46). To our knowledge, the inhibitory effect of gluca- gon on L-PK gene transcription (14) is the first example of an inhibition of the transcription of higher eukaryotic gene by glucagon. Whether negative or positive, however, regula- tion of the transcription of a gene by cAMP does not preclude regulation at other pretranslational levels. For example, cAMP regulates both the rate of transcription of the lactate dehydrogenase gene and the stability of its mRNA (45).
The question of how cAMP coordinately controls two dis- tinct steps occurring in different compartments of the cell (namely transcription in the nucleus and mRNA degradation in the cytoplasm) remains unresolved. Wynshaw-Boris et al. (47) have identified a specific sequence which is involved in cAMP induction of phosphoenolpyruvate carboxykinase gene transcription. In addition, data in several eukaryotes support the hypothesis that a CAMP-dependent protein kinase may
L-tvpc PYRUVATE W E I I
a
UZ 300 B E
g g
I 200
100
NORMAL ADX 0
FIG, 4. Effect of bilateral adrenalectomy on L-PK gene transcription and mRNA content in rats refed the carbohy- drate-rich diet for 12 h. Both normal and adrenalectomized (ADX) rats were refed the carbohydrate-rich diet for 12 h after a 24-h starvation. The L-PK gene transcription (open bars) is expressed in parts/million and the mRNA level (hutched bars) in counts/minute/ micrograms total RNA. Both gene transcription and mRNA level are the mean f 1 S.D. for 15 individual determinations in 10 normal rats and for 11 individual determinations in eight adrenalectomized rats. The figure also shows the level of the gene transcription in each individual experiment.
1-m PYRUVATE KINASE
: -I 5w
FIG. 5. Effect of thyroparathyroidectomy on L-PK gene transcription and mRNA content in rats refed the carbohy- date-rich diet for 12 h. Both normal and thyroidectomized (TX) rats were refed the carbohydrate-rich diet for 12 h after a 24-h starvation. 5-Triiodo-~-thyronine-treated thyroidectomized animals
after a 12-h period of refeeding and were killed 3 h later. The L-PK (TX + T3) were given triiodo-L-thyronine (50 pg) intraperitoneally
gene transcription (open bars) is expressed in parts/million and the mRNA level (hatched bars) in counts/minute/micrograms total RNA. Both gene transcription and mRNA level are the mean f 1 S.D. for 18 individual determinations in 10 normal rats, for five individual determinations in five thyroidectomized rats, and for nine individual determinations in five triiodo-L-thyronine thyroidectomized rats. The figure also shows the level of the gene transcription in each individual experiment.
mediate the action of cAMP on gene transcription via either its catalytic (48, 49) or its regulatory subunit (50, 51). How cAMP acts at the post-transcriptional level remains totally obscure.
Effect of Thyroparathyroidectomy and Bilateral Adrenalec- tomy on L-PK Gene Transcription in Rats Refed the Carbo- hydrate-rich Diet-Glucocorticoids and thyroid hormones are required for the full dietary induction of L-PK mRNA in rat liver (13). We compare here the relative rate of L-PK gene transcription to mRNA abundance in normal, thyroidecto- mized, and adrenalectomized rats refed the carbohydrate-rich diet (Figs. 4 and 5). Whereas transcription rate is not affected by adrenalectomy, mRNA level is reduced to about 20% of normal fed values (Fig. 4). The same is true for thyroidectomy: transcription rate in thyroidectomized rats is barely reduced (about 80% of normal), whereas L-PK mRNA is reduced to

Regulation of L-type Pyrul
about 40% of the normal fed value (Fig. 5). Therefore, gluco- corticoids and thyroid hormones probably regulate mRNA concentration at a post-transcriptional level. We cannot en- tirely exclude, however, the possibility that these hormones modify elongation of the nascent RNA chains, although this mechanism is not generally thought to play a significant role in the regulation of eukaryotic gene expression.
The effects of glucocorticoids and thyroid hormones on L- PK mRNA concentration first reported by Munnich et al. (13) and confirmed in this paper relate to the impairment of normal induction in fasted rats refed a mixed glucose-fructose diet for 12-18 h. By contrast, Noguchi et al. (37) did not observe a decrease in L-PK mRNA in adrenalectomized rats fed a fructose diet without prior fasting. The reason for this discrepancy could be that glucocorticoids are mainly involved in the early phenomena occurring when the L-PK gene, totally switched off in fasted rats, is activated by carbohydrate re- feeding and are not necessary to maintain mRNA concentra- tion during steady state conditions.
Whereas transcriptional effects of glucocorticoids are now well documented (52-58), there are several recent reports of the action of these hormones at the level of pre-RNA proc- essing (59), transport from nucleus to cytoplasm (60), or mRNA stability (61). The same is true for thyroid hormones which, in addition to their action on gene transcription (57, 62), may modify the post-transcriptional processing of some transcripts (42).
In conclusion, our results indicate that diet and hormones regulate L-PK gene expression at both transcriptional and post-transcriptional levels. Essentially the same is true for two other carbohydrate-induced hepatic genes which are cur- rently under investigation, namely aldolase B (15) and a 5.4- kb mRNA species (35). Transcription is regulated by two major effectors, one positive, namely carbohydrates in the presence of insulin, and one negative, namely glucagon (via CAMP); both agents probably act at the level of mRNA stability as well. Glucocorticoids and thyroid hormones alter pyruvate kinase gene expression at the post-transcriptional level.
Acknowledgments-We thank Claudine Brunner for typing the manuscript and Allan Strickland for his help in preparing it.
REFERENCES
late Kinase Gene Expression 7625 13. Munnich, A,, Marie, J., Reach, G., Vaulont, S., Simon, M.-P., and Kahn,
A. (1984) J. Biol. Chem. 259,10228-10231 14. Vaulont, S., Munnich, A., Marie, J., Reach, G., Pichard, A. L., Simon, M.
P.. Besmond. C.. Barbrv. P.. and Kahn. A. (1984) Blochem. Bio~hvs. Res. Commun. 125,135-147 '
C., and Kahn, A. (1985) J. Chn. Invest. 7 5 , 1045-1052
, . . ~"
15. Munnich, A., Besmond, C., Darquy, S., Reach, G., Vaulont, S., Dreyfus, J.
16. Mahmood, H. A., Wood, P. J., and Marks, V. (1978) Diabetologia 14,405-
17. Kahn, A,, Cottreau, D., Daegelen, D., and Dreyfus, J. C. (1981) Eur. J.
18. Busch, H. (1967) Methods Enzymol. 12,421-448 19. Bailey, J. M., and Davidson, N. (1976) Anal. Biochern. 70,75-85 20. Messing, J. (1983) Methods EnzymoL 101,ZO-79 21. Church, G. M., and Gilbert, W. (1984) Proc. Natl. Acad. Sei. U. S. A. 8 1 ,
22. Reeder, R. H., and Roeder, R. G. (1972) J. Mol. Bid. 67,433-441 23. Zylber, E. A., and Penman, S. (1971) Proc. Natl. Acad. Sci. U. S. A. 6 8 ,
24. Tuil, D., Vaulont, S., Levin, M. J., Munnich, A,, Moguilewsky, M., Bouton, 2861-2865
M. M., Brissot, P., Dreyfus, J. C., and Kahn, A. (1985) FEBS Lett. 189, 21 n-21 A
412
Biochem. 116,7-I2
1991-1995
25.
26.
27. 28. 29.
30.
Williams, R. H., and Porte, D. (1974), Textbook of Endocrinobgy, p. 515,
Goldman, M. J., Back, D. W., and Goodridge, A. G. (1985) J. Biol. Chem.
Sibrowski, W., and Seitz, H. J. (1984) J. Biol. Chem. 2 5 9 , 343-346 Miksicek R. J. and Towle H. C. (1982) J . Bid. Chern. 257,11829-11835 Siddiqui,'U. A.: Golaam, k., and Goodridge, A. G. (1981) J. Biol. Chem.
Pry, T. A., and Porter, J. W. (1981) Biochem. Biophys. Res. Cornmun. 100 ,
v.Ly
W. B. Saunders Co., Philadelphia
260,4404-4408
256,4544-4550
1nn2-1009 31. Kletzien, R. F., Prostko, C. R., Stumpo D. J. McClung, J. K., and Dreher,
32. Whberry L. K., Morris S. M. Jr., Fisch, J. E., Glynias, M. J., Jenik, R. K. L. (1985) J. Biol. Chem. 260,562'1-5624
33. Nepokroeff, C. M., Adachi, K., Yan, C., and Porter, J. W. (1984) Eur. J. A,, and'Goodridge, A. k. (19873) J. Biol. Chem. 258,1337-1342
34. SUI, H. S., Wise, L. S., Brown, M. L., and Rubin, C. S. (1984) J. Bid. Chern. Biochem. 140 ,441445
35. Pichard A. L., Munnich A., Meienhofer, M. C., Vaulont, S., Simon, M. P., 259,1201-1205
Marie', J., Dreyfus, J. h., and Kahn, A. (1985) Biochem. J. (Tokyo) 226 , 637-644
36. Inoue H., Noguchi T., and Tanaka, T. (1984) J. Biochem. 96, 145-1462 37. Nogulhi, T., Inoue: H., and Tanaka, T. (1985) J. Biol. Chem. 260,14393-
38. Cimbala M. A., Lamers, W. H., Nelson K., Monahan, J. E., Yoo-Warren,
39. Ernest, M. J. (1982) Biochemistry 21,6761-6767 40. Firestone, G. L., and Heath, E. C. (1981) J. Bid. Chem. 256,1396-1403 41. Lewis, E. J., Tank, A. W., Weiner, N., and Chikaraishi, D. M. (1983) J.
42. Narayan P., and Towle H. C. (1985) Mol. Cell. Bzol. 5 , 2642-2646 Biol. Chem. 258,14632-14637
43. Maurer, R. A. (1981) NAture 294,94-97 44. Lamers W. H., Hanson, R. W., and Meisner, H. M. (1982) Proc. Natl.
45. Jungmann, R. A,, Kelley, D. C., Miles, M. F., and Milkowski, D. M. (1983) Acad.'Sci. U. S. A. 7 9 , 5137-5141
46. Hashimoto, S., Schmid, W., and Schutz, G. (1984) Proc. Natl. Acad. Sci. U. J. Biol. Chem. 258,5312-5318
47. Wynshaw-Boris, A,, Lugo, T. G., Short, J. M., Fournier, R. E. K., and 5'. A. 81,6637-6641
48. Boney, C., Fink, D., Schlichter, D., Carr, K., and Wicks, W. D. (1983) J. Hanson, R. W. (1984) J. Bid. Chem. 259,12161-12169
Bid. Chem. 258,4911-4918
50. Nagamine, Y., and Reich. E. (1985) Proc. Natl. Acad. Sci. U. S. A. 82. 49. Waterman, M., and Murdoch, G. H. (1985) Science 229,267-269
- . - - . .
14397
H., and Hanson, R. W. (1982) J. Biol.' Chem. 257 , 7629-7636
2. 1.
3.
4.
5.
6. 7. 8.
10. 9.
11.
12.
Weber, G., Stamm, N. B., and Fisher, E. (1965) Science 149,65-67 Freedland, R. A,, Cunliffe, T. L., and Zinkl, J. G. (1966) J. Biol. Chem.
Sillero, A., Sillero, M. A. G., and Sols, A. (1969) Eur. J. Biochern. 10, 351-
Kohl, E. A., and Cottam, G. L. (1976) Arch. Biochem. Biophys. 176 , 671-
Cladaras, C., and Cottam, G. L. (1980) Arch. Biochem. Biophys. 200,426-
Johnson, M. L., and Veneziale, C. M. (1980) Biochemistry 19, 2191-2195 Cladaras, C., and Cottam, G. L. (1980) J. Biol. Chem. 255,11499-11503
Miyanaga, O., Nagono, M., and Cottam, G. L. (1982) J. Biol. Chem. 2 5 7 ,
Poole, G. P., and Bloxham D. P. (1982) Biochern. J. 204,89-95 Poole, G. P., Postle, A. D.: and Bloxham, D. P. (1982) Biochem. J. 2 0 4 ,
Weber, A., Marie, J., Cottreau, D., Simon, M.-P., Besmond, C., Dreyfus, J.
Simon, M.-P., Besmond, C., Cottreau, D., Weber, A., Chaumet-Riffaud P.
241,5448-5451
354
682
433
10617-10623
81-87
C., and Kahn, A. (1984) J. Bud. Chem. 259,1798-1802
Dreyfus, J. C., Sala-Trepat, J., Marie, J., and Kahn, A. (1983) J. Chem. 2 5 8 , 14576-14584
4606-4610 51. Constantinou, A. I., Squinto, S. P., and Jungmann, R. A, (1985) Cell 4 2 ,
52. Hager L. J., and Palmiter R. D. (1981) Nature 291,340-342 53. Ebedine, J. H., and Roberts, J. L. (1984) J. Biol. Chem. 2 5 9 , 2166-2170 54. Danesch, U., Hashimoto, S., Renkawitz, R., and Schiitz, G. (1983) J. Biol.
55. Turcotte, B., Guertin, M., Chevrette, M., and Belanger, L. (1985) Nucleic Chem. 258,4750-4753
56. Hynes, N. E., Rahmsdorf, U., Kennedy, N., Fabiani, L., Michalides, R., Acids Res. 13,2387-2398
57. Spindler, S. R., Mellon, S. H., and Baxter, J. D. (1982) J. Bid. Chem. 2 5 7 , Nusse, R., and Groner, B. (1981) Gene (Amst.) 15,307-317
58. Sasaki, K., Cripe, T. P., Koch S. R. Andreone, T. L. Petersen D. D. 11627-11632
59. Vannice, J. L., Taylor, J. M., and Ringold, G. M. (1984) Proc. Natl. Acad. Beale, E. G., and Granner, D.'K. ( l 9 b ) J. Bid. Chem. 259,15248-1525i
60. Fulton, R., Birnie, G. D., and Knowler, J. T. (1985) Nucleic Acids Res. 13 , Sei. U. S. A. 81,4241-4245
6467-6482
62. Loose, D. S., Cameron, D. K., Short, H. P., and Hanson, R. W. (1985) 61. Diamond, D. J., and Goodman, H. M. (1985) J. Mol. Biol. 181 , 41-62
429-437
Biochemistry 24,4509-4512


















