
The main goal of .me resolved x-ray crystallography is to...
74
1
Transcript of The main goal of .me resolved x-ray crystallography is to...

1

Themaingoalof.meresolvedx-raycrystallographyistobeableto“film”aproteininac.on.Theideaistofollowthedifferentstructuralchangesoccurringduringanenzyma.creac.on,anallostericchange,theaccessofaligandorsubtratetoabindingsiteorac.vesite,thereleaseofaproductorligandfromtheac.vesiteorbindingsite,etc…Forenzyma.creac.on,theatomicresolu.onisrequired,asweneedtobeabletoassesspreciselyalltheinteratomicinterac.on(hydrogenbonds,saltbridges,vanderWaalscontacts,etc…).Lowerresolu.onmaybeenoughtomonitorglobalstructuralchanges(allostericchanges).Wealsoneedtoobtaininforma.onattherelevant.mescale,dependingonthebiologicalsystemstudied.
2

3

The.mescaleatwhichwecanobservestructuralchangesisextremelylargeandbasicallygoesfrom100fs(atomicmovementinducedbyatomicorelectronicexcita.on)uptothesforunfoldingprocesses,thusarangeofatleast13orderofmagnitude.Mostofmacromoleculefunc.onarepossiblethankstolargeramplitudecollec.vemovements.Thetemperaturehasastrongimpactonthedynamicandamplitudeofmovementinmacromolecules.Thepresenceofcollec.vemovementisillustratedbythechangeinslopeatagiventemperature,whenthemeansquaredatomicdisplacement<∆u2>isploRedasafunc.onoftemperature.Onthediagram,weclearlyobservetwodifferentbehaviors,eitherbeloworabove180-200K.Thismeansthatwemayblockcollec.vemo.onbydecreasingthetemperaturebelow180Kandthustraptheproteininagivenstate.Liquidnitrogentemperature(100K)isappropriateforsuchatrapping.
4

Thetwotechniquesthatallowtogetstructuralinforma.onatthemostappropriate.mescalesareX-raydiffrac.onandSmall(orWide)AngleX-rayscaRering(SAXSorWAXS).Themaincharacteris.csofthetwotechniquesthatmakethemwellsuitedfor.meresolvedstructuralstudiesaremen.onedonthisslideandthenextone.
5

6

Howtotriggerareac.on?Oneofthemainissuewillbetotriggerthereac.onofallmoleculeswithinthesampleattheverysame.me(i.e.ina.methatismuchshorterthanthereac.on.me)inordertohavethereac.ongoingoninaperfectlysynchronouswayonallmolecules.1) Thediffusionofsubstrate,co-factor,etc…canbedonewithinthecrystal.Itis
possibleinmanycases,sinceproteincrystalsdocontainasignificantamountofsolvent:thevolumeofaproteincrystaloccupiedbysolventmayvaryfrom30to80%,withsome.melargechannelinwhichligandscaneasilydiffuse.Thedrawbackisthatitisarela.velyslowprocess(about100msfor30x40x50µm3crystal),thatcanbeusedonlyforquiteslowreac.on.Theuseofverysmallcrystals,thankstoalwaysmorepowerfulX-raysourcemayspeedupthetriggering.
2) Endogenousphotosen.vity.Lightistheperfecttoolstotriggerareac.onasitcantrulyac.vateallthemoleculesatthesame.me.Insomecases,theproteinisendogenouslyac.vatedbylight(presenceofachromophore,protein-ligandinterac.onthatcanbebrokenbyphotons,…
3) Onecanuseacagedcompound,thatdobindclosetotheac.vesite,andthatcanbeuncagedbylight.Inthatcasealso,wecanthustriggerthereac.onatthesame.meinallmoleculesofthesample.
4) TheX-raysthemselvescaninduceareac.on(radiolysis,foranexample).Inprinciple,itmayworkasgoodaswithlighttriggering,butoneshouldpayaRen.onthatX-raysusedforthestructuralinves.ga.onsarenotalsosignificantlyac.va.ngthereac.on.
7

8

9
Some reminder about classical crystallography. With first need to produce crystals, step which remains one of the major bottleneck today. The main idea is to find conditions in which macromolecules interact with each other in order to form an ordered solid: the crystal. This conditions cannot be predicted from the amino-acid sequence and lots of trials should often me made, with no guaranty of success. Ultimately, when crystals of suitable size are obtained, they are plunged in a cryoprotecting solution and frozen in liquid or gazeous nitrogen.

10
Either laboratory X-ray generators or synchrotron sources (much more intense beam) can be used. The typical wavelength is around 1 Å, that correspond to a photon energy of about 10 keV. The full 3D diffraction spectra (series of fine angular slices, 0.05° to 1° step, depending on the equipment used) is collected on CCD or direct photon detection detector. Different techniques can be used to retrieve the phase information which is lost during the data collection process (only intensities of the diffracted waves are recorded). These phases allow the calculation of the electron density map at atomic resolution, in which the atomic model can be built. The main message is that the structure obtained represent an average over the data collection time (typically 1 mn, with today’s synchrotron, but can be up to a few hours for less intense X-ray sources) and over all the molecules present in the crystal (1014-1018). The data collection is clearly not fast enough to image an enzymatic reaction.

ThesetupforSAXSorWAXSexperimentsisabitdifferent.LikeforX-raydiffrac.on,either laboratory X-ray generators or synchrotron sources (much more intense beam) can be used.However,thesampleisinsolu.onandisflowninacapillarythroughtheX-raybeam.Dataarecollectedatlowdiffrac.ngangle(typically:λ≈1Å;0<q<0.5Å-1;0<2θ<4.5°;dmax≈13Å).Sincemoleculesformingthesampleareinrandomorienta.onandmoverapidly,thediffusionpaRernhasacircularsymmetry,andthediffusionintensitycanthusbeaveragetoa1Ddiffusioncurve.
11
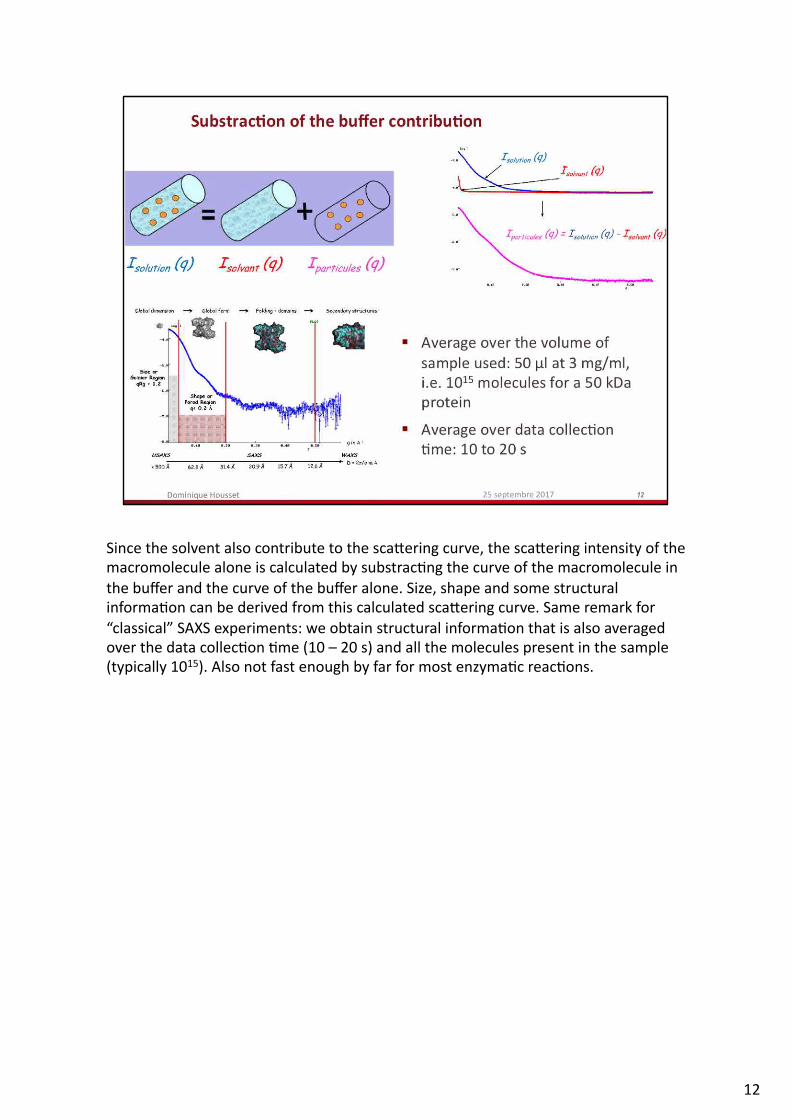
SincethesolventalsocontributetothescaReringcurve,thescaReringintensityofthemacromoleculealoneiscalculatedbysubstrac.ngthecurveofthemacromoleculeinthebufferandthecurveofthebufferalone.Size,shapeandsomestructuralinforma.oncanbederivedfromthiscalculatedscaReringcurve.Sameremarkfor“classical”SAXSexperiments:weobtainstructuralinforma.onthatisalsoaveragedoverthedatacollec.on.me(10–20s)andallthemoleculespresentinthesample(typically1015).Alsonotfastenoughbyfarformostenzyma.creac.ons.
12

13

CeRetechniqueconsisteàbaisserlatempératurepourpiégerlamacromoléculedansunétatpar.culier,enbloquantunetrèsgrandepar.edesmouvements.Eneffet,lafonc.ondenombreusesprotéineestdirectementliéàl’existencedemouvementscollec.fsdelargeamplitudequisontobservésaudelàd’unecertainetempérature,typiquementdel’ordrede180K.Onpeutdonc“figer”uneprotéinedansunétatpar.culierenlacongelantàlatempératuredel’azoteliquide(100K).Audessousd’unecertainetempérature,lesenzymesnefonc.onnentplus,carlamoléculesnepossèdeplusl’énergiethermiquesuffisanteetnepeutplusfranchirlesbarrièresd’énergied’ac.va.on.Celarevientàdiminuerfortementlaconstantecataly.quekcat,quisuituneloid’Arrheniusenfonc.ondelatempérature.
14

Plusieursstratégiessontenvisageablespourpiégerunétatintermédiaire:1) Trigger-Freezze:ondéclenchelaréac.on,puisonbaisselatempératureàun
momentprécis(possiblepourlesréac.onssuffisammentlentes)2) Freeze-trigger:onbaisselatempérature,puisondéclenchelaréac.on,enfixantla
températuredesortequelapremièreétapedelaréac.onsoitpossible,maispaslasuivante.
3) SteadyState:idemque2)maisonmain.entuneac.va.oncon.nue(permetd’empêcherquel’étatintermédiairesedépeupleàcausedelaréac.onensensinverse)
4) Lauce-trapping:Leréseaucristallinpeutempêchercertainmouvement,augmentercertainesénergiesd’ac.va.onetdoncbloquercertainesréac.onsdansunétatintermédiairedonné.Icionjouedoncsurlamodifica.ondel’énergied’ac.va.on,etéventuellementsurlatempérature,sibesoin.
15

Onpêcheuncristalquel’onmetdansunesolu.onquidéclenchelaréac.on.Aprèsuntempsdéterminé,onre-pêchelecristaletonlecongèledansl’azoteliquideOnu.lisececristalcongelépourlesétudesstructuralesetspectroscopiquesIlfautquelaréac.onsoitassezlente(minute).Onasouventdesmélangesd’états.Lacombinaisond’étudesstructuralesetspectroscopiquespermetdemieuxiden.fierlesétatsetlesespèceschimiquesprésentesdansl’échan.llon.
16

LaSORestunemetalloenzyme(Fe)quel’ontrouvechezuncertainnombredebactéries(e.g.Treponemapallidum)etarchae(e.g.Pyrococcusfuriosus)sensiblesàl’oxygène.Ellepar.cipeàunevoiealterna.ved’élimina.ondel’ionsuperoxyde,différentedecelletrouvéechezlesbactériesaérobies,quifaitintervenirlasuperoxydedismutase(SOD),lacatalaseetdesperoxydases.Danslesiteac.fdelaSOR,leFerestcoordonnépar4his.dineséquatorialesetunecystéineaxiale.Ici,nousallonsregarderplusendétailuneétudequicombinecristallographieciné.queetspectroscopieRaman(Katonaetal.,2007)pourcomprendrelemécanismeréac.onneldelaSOR.
17

L’idéedebaseestdepiégerlecomplexeFe-peroxo.Pourcefaire,ilsuffitdemeRreenprésencelaformeréduitedelaSOR(Fe2+)etH2O2.Cependant,pouréviterlaforma.onderadicauxhydroxyltrèsréac.fs,laSORestd’abordoxydéeaveclesodiumhexachloroiridate(IV),puismisenprésenced’H2O2(Pourcetravail,c’estlaréac.oninversequiestdéclenchée,maisl’étatintermédiairerecherchéerestelemême).Pourbloquerlaréac.onaustadeoùlecomplexeFe-peroxoestformé,unmutantspécifiqueGlu114Alaestu.lisépourstabiliserunétatintermédiaire.
18

LecomplexeFe-peroxydeestvudans3unitésasymétriquessur4etsaprésenceestconfirméeparspectroscopieRamanensolu.onetincristallo.LaLys48quiformeunesortedecouvercle(lid)ausiteac.festunrésidutrèsconservéchezlesSORetl’onvoiticisonrôledanslastabilisa.ondesmoléculesd’eauetduperoxo.Lescalculsdechimiequan.queparlathéoriedefonc.onnelledensité(DFT)ontpermisdemontrerquelesmonomèresBetDétaientprotonéssurl’oxygènedistalduperoxo.(A) Lidlocked-open:Lys48estdésordonnée(B) Lidlocked-closed:Lysinteragitavecleperoxo.Wat10etWat11par.cipentà
l’interac.on.Lachargeposi.veduNH3+aureleperoxoetfragiliselaliaisonFe-
peroxo(C) Lidlocked-closed:Lys48sedéplaceunpeu.Wat10etWat11aussi.Wat11est
maintenantdansuneposi.onoùellepeutdonnerunprotonàl’oxygèneproximalduperoxo,pourformerH2O2quiseralibéré.C’estcequidis.nguelesSORdesenzymesàhèmes,oùc’estl’oxygènedistalquiestprotonéundeuxièmefois,avecrelargaged’H2Oetcréa.ond’uncomplexeoxo-feryl,toxique.
19

D’aprèslesdifférentsétatsdéterminés,un“film”dumécanismeréac.onnelpeutêtreproposé.Toutefois,lesconstantesdetempsnesontpasaccessibles.
20

Unvueplusdétailléeetconven.onnelledumécanismeréac.onnelproposépourl’élimina.onO2
-parlaSOR.
21

Un.l2013,thevastmajorityof.meresolvedcrystallographicstudieswereperformedusingtheLauediffrac.ontechnique.Insteadofusingamonochroma.cX-raybeamusedforclassicalstructuralinves.ga.on,apolychroma.cbeamisused.Theadvantageistocollectmuchmoredataperimageforeachorienta.onofthecrystal.Therefore,thedatacollec.onrequireslessimagesinordertoobtainacompletedataset.Thedrawbackisthatsomespotsdooverlapwhichmakesthedataprocessingabitmorecomplex.TheconceptofLaue.meresolvedX-raycrystallographystudiesistoac.vatethemoleculesofthecrystalbyalaserpulse,andthentocollectthestructuraldatawithanX-raypulse.ThedelaybetweenthelaserpulseandtheX-raypulsecanbechosenbytheuser,inordertoinves.gatedifferentstatesalongthereac.onpathway.Theminimum.mescalethatcanbeinves.gateddependonthedura.onofeitherthelaserpulseortheX-raypulse:thelongestofbothdefiningtheminimum.mescale.Agoodsynchroniza.onbetweenlaserandX-raypulseisrequired.Onlyreversiblereac.onsareinves.gatedusingthisexperimentalsetup,sinceonecrystalisusedforcollec.ngseveralimages(infactanen.redataset).Thedatacollec.onismadeatroomtemperature,inordertoallowstructuralchangestohappenandthereturntothe“dark”stateforthecollec.onofthenextimage.
22

Illustra.onbytheEwalddiagramofreflec.onscollectedforone“s.ll”image.Foramonochroma.cexperiment,onlythenodesofthereciprocalspace(greyorbluepoints)thatcrossestheEwaldsphere(letsaytheredone,ifdatacollec.onismadeatλmax)willgeneratespotsonthediffrac.onimage.Ifapolychroma.cradia.onisused,e.g.allwavelengthbetweenλminandλmax,allthenodesofthereciprocalspacethatarelocatedinbetweentheredandthegreencircle(areashadedingrey)willgeneratespotsonthediffrac.onimage:thus,muchmorethanwithamonochroma.cbeam.
23

SchemeoftheexperimentalsetupusedforLaue.meresolvedexperimentsandpictureofsucharealsetuponasynchrotronbeam.YoucanseethattheenvironmentofthecrystalispreRycrowded.
24

25

Examplesof.meresolvedstructuralstudiesfocusonmyoglobinorhemoglobin.Here,thegoalistounderstandwhathappenwhenthebondbetweentheiron(inthecenterofthehemegroup)andtheligand(CO,bindingmore.ghtlythannaturalligandsO2,CO2)isbrokenbyphotolysis.Ifallostericchangesareknowntobetriggeredbyligandbindingorunbinding,thekine.cofsuchevents,andthedetailedstructuralmechanismsunderlyingallostericchangesremainedtobeinves.gated.
26

In2003,ans.meresolvedstudyofSpermwhale(Cachalotinfrench)myoglobinhasbeenperformed.Twosnapshots,one3nsaÇerphotolysisoftheFe-CObond,one316nsaÇerphotolysisoftheFe-CObondhavebeenanalyzed.
27

Theanalysisofthedifferenceelectrondensitymaps{Fphotolysed–Fdark}showsthatsignificantchangesoccurredaÇer3ns.TheCOisnolongerboundtotheFe(largenega.vedensity,inred).Tworesidues,Tyr29andGln64havemovedsignificantlyfromtheirini.alposi.oninyellowtotheirnewposi.oninblue.TheCOseemstohavemovedtotheXe4posi.on.AÇer316ns,nomajorfurtherchangesintheimmediatesurroundingoftheheme.TheCOhasnowmovedontheothersideofthehem,inposi.onXe1
28

ExcepttheCOposi.on,themaindifferencedifferencebetweenstructuresat3nsand316nsisobservedquitefarfortheac.vesite.Collec.vedisplacementsareslowerandoccursattheµs.mescale.
29

Onthebasesofthistwosnapshots,ananima.onoftheCOphotolysiscanbeproposedbytheauthors.
30

Scapharcainaequivalvis,akindofclam(palourdeorclaminfrench,ashellfishfromthebivalvesfamily)possessadimerichemoglobin.Asforhumanhemoglobin(whichistetrameric),thishemoglobinundergoanallosterictransi.on,withtwowelliden.fiedstates:theR(relaxed)stateandtheT(tense)state,whichhavedifferentaffini.esforligands.Here,themaingoalistounveilthemainstructuralintermediatesalongthestructuraltransi.on.
31

In2006,a5ns.meresolvedstructuralstudyunveiloveraquitelarge.mescale(5ns–80µs).Asforthepreviousstudy,theelectrondensitymapshowsthat5nsaÇerlaserpulse,40%oftheCOmoleculesarenolongerboundtotheFeandhavemovedtoadistalsitelabeledCO*inthefigure.TheFeatomhasalsobeendisplacedbyabout0.4Åtowardstheproximalhis.dine.
32

CeRefigureillustrelemouvementdel’atomedeFer,aucentredel’hème.Dansunepremièrephase(5ns-1µs)l’atomedefersedéplacede0.4Åperpendiculairementàl’hèmeversl’his.dineproximale(posi.on≠deRetT).Peudedéplacementdansplandel’hème:onrestedansl’étatR.Puisaprès1ms,l’atomedeFerevientdansleplan(mouvementperpendiculairementauplan)maissondéplacementdansleplandel’hèmemontrequ’iladoptel’étatTets’éloignesignifica.vementdel’étatR(≈0.9Å).
33

Lacourbeduhautmontrequ’entre5nset200ns,lesitedistalduCO(CO*)estoccupéalorsquelesiteproximalest(engrandepar.e)inoccupé.Apar.rde1µs,lesiteproximalretrouve(quasitotalement)sonCOetlesitedistalleperd.Lechangementdeconforma.ondelaphénylalanineF4estplustardif:peudechangementavant1µs,puistransi.onversle«Tstate».LedépartduCOdéclenchedoncunecascadedechangementsstructurauxquiperdurentbienaprèsleretourduCOàsaposi.onini.ale.
34

Thisanimatedpictureshowsthedifferentstepsfrom5nsto60µs.At60µs,mostofthedifferenceelectrondensityisseenattheinterfacebetweensubunits,possiblyindica.ngaemergingallostericchange,whilethedifferenceelectrondensityintheFeimmediatesurroundingshasbasicallyvanished,indica.ngthattheCOisnowbacktoitsini.alposi.on.ThemaintakehomemessageisthatthestructuralcycleofCOphotolysisonhemoglobincanbedescribedbyseriesofeventsoccurringatdifferent.mescales.TheFe-COphotolysis,whichoccurveryrapidlyaÇerthelaserpulse(<5ns),triggerstructuralchanges(movementofsidechains,movementofsecondarystructures,rota.onofmonomer)thatcon.nueatthe10µs–ms.mescale,whiletheCOhascomebacktoitsini.alposi.oninlessthat1µs.At60µs,theallostericchangeisfarfrombeingcompleted(0.6°rota.onreachedinsteadof3°forthewholerela.vechangeoforienta.onofthetwomonomersintheTstate,comparedtotheRstate)Fasttriggeringeventcantriggerlonglas.ngstructuralevents.
35

Thetranstocisisomerisa.onreac.onconcernstheC1-C2-C3-C1’dihedral,whichistransinthedarkstate(anglecloseto180°)andcisintheac.vatedstate(anglecloserto0°).Thetrans-to-cisisomerisa.onisthoughttohappenveryrapidly,inthefstops.mescale,accordingtospectroscopicstudies.However,globalstructuralchangesarerequiredforsignaltransduc.onshouldoccuratamuchlonger.mescale.Manyworkshavebeendoneonthisprotein(wewillgothroughsomeoftheminthenextslides),asPYPisgoodcandidateforacasestudy.Butthemainbiologicalgoalwastounveilthestructuralmechanismthatistriggeredbythephotonabsorp.on,thateventuallytellsthebacteriatochangeofdirec.ontoescapeblueradia.ons.
36

Thefirst.meresolvedstructuralinves.ga.onofPYPwasmade20yearsago.The.mescaleis10ms,sinceathat.me,theX-pulsewasgeneratedbyamechanicalshuRer:onecannotgotomuchlower.mescalewithsuchshuRers.Thesamecrystalisusedforabout300images:itresistpreRywelltoradia.ondamageatroomtemperature,whichisnotalwaysthecaseforproteincrystals.Twostructuresarecompared:darkstate(nolaserillumina.on)andbleachedstate(datacollected2msaÇertheendofa200mslaserpulse).From10to15X-raypulse(10ms)areusedeachorienta.on,inordertohaveenoughsignal.A30srecovery.meisrequiredbetweeneachexposure,forthePYPtoreturnintothedarkstate.
37

Theelectrondensityomitmap(thechromophorehasnotbeentakenintoaccountfortheFcalcandphasecalcula.ontoavoidanypossiblebias)ofpanelCandD(bleachedstate)showsthattwostatesarepresentinthecrystal.Thismeansthatthereac.onhasnotbeentriggeredinallthemoleculesofthecrystal,butonabout50%ofthem.Sincetheresolu.onishighenough(1.9Å),thetwostatescanbeseenandmodeledanyway(yellowforthedarkstate,whiteforthebleachedstate).PanelAandBshowthedifferenceelectrondensitymapFbleached–Fdark.Posi.veregions(blue)indicatewheretomovethechromophoreaÇerac.va.onandcorrespondtothebleachedstate(whitemodelbuildinit)andnega.veregions(red)indicatethatfulloccupancyofthedarkstateconforma.onofthechromophoreisnotmaintainedaÇerac.va.on.
38
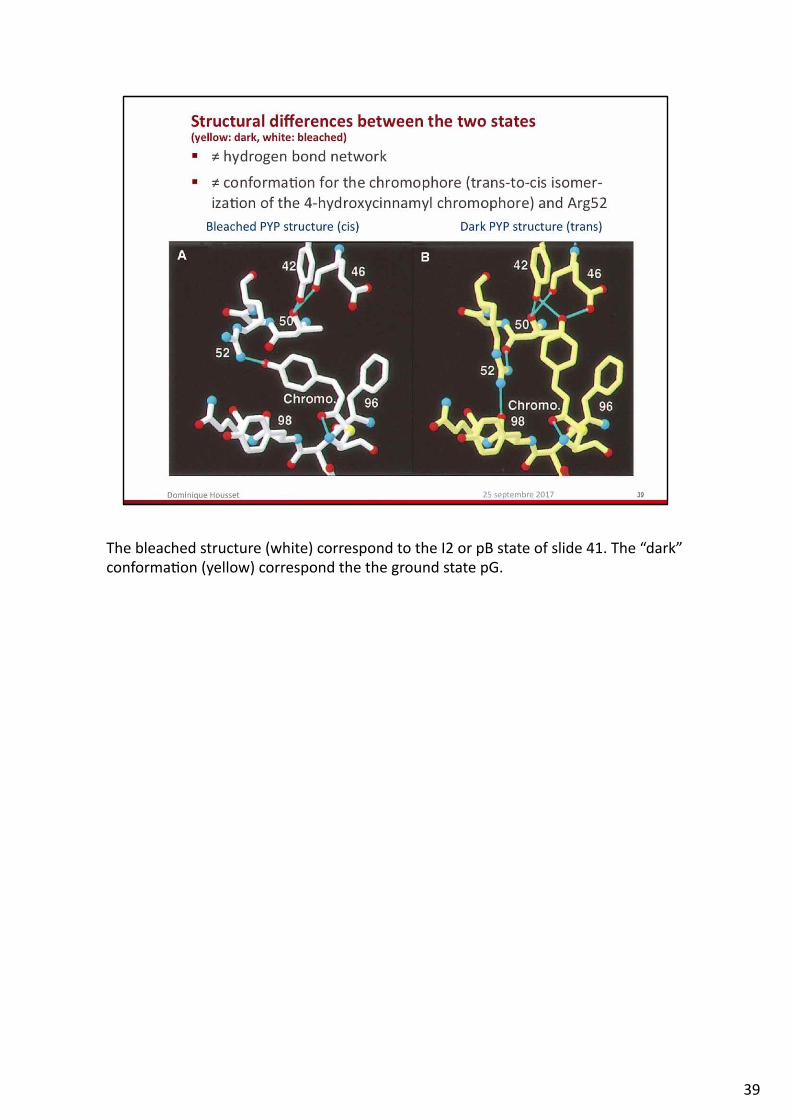
Thebleachedstructure(white)correspondtotheI2orpBstateofslide41.The“dark”conforma.on(yellow)correspondthethegroundstatepG.
39

Astoldbefore,PYPhasbeenwidelystudiedun.lrecently,sinceitisaverygoodproteintotesttheprogressofthedifferenttechniques.4studies(from2005un.l2016)willbeanalyzedinthefollowingslides.Forthefirstthree,performedwithLauediffrac.on,youcanseetheimprovementsinboththe.meresolu.onandthenumberofsnapshots,comparetothepioneeringstudy.Theyrepresentthebestwecandowithsynchrotronsourcesofar.
40

In2005,sevenyearslater,ansstudy(insteadof10msforthepreviousone)ofthePYPmechanismismadepossiblebytheuseofanslaserandofsynchrotronX-raypulses.Amuchmoredetailedviewofthecyclecanbeproposed(comparetotheoneproposedin1998fromspectroscopicdata),whith4differentstates:Icp(cis-planarintermediate,α,possiblycorrespondingtoI0#),onepRstatewithtwopossibleconforma.ons(β)andtwopBstates(γandδ)andthe.meconstantsassociatedtoeachstate.
41

Accordingtothechangesintheresidualelectrondensitymap({Fobs–Fcalc},bluerepresentposi.vedifferences,i.e.posi.onwheremoreelectron(atoms)shouldplaced;redrepresentnega.vedifferences,i.e.posi.onswhereatomsshouldnolongerbe,atleastpar.ally,Fcalciscalculatedwiththemodelcorrespondingtothedarkstate),at1ns,thetrans-to-cistransi.onhasalreadyoccurredonthechromophore,butchangesatlimitedtothecloseneighborhoodofthec1-c2-c3-C1’dihedral.Significantchangesofthechromophoreringposi.onoccursattheµsscale,withthetwopRconforma.onobserved.Thechromophoreconforma.onalchangeiscompletedatthemsscale.Butlargerscalemovementhavebeentriggered.Thedisplacementofanhelix,quitedistantfromtheac.vesiteisobserveaÇer50ms(δstate,panelJ).
42

Here,thedifferentstateshavebeenmodeledintotheelectrondensity(panelsA,B,C,D)andthesuperposi.onwiththeground(dark)stateisshowninthepanesE,F,G,H.Thetwopossibleconforma.onsofthepRstateareshown:oneinwhichtheringofthechromophorehasnotmovedmuch,andoneinwhichithasmoveclosertoposi.onadoptedinpBstate.
43

Asimplerviewofthedifferentstructuraltransi.onsobserved.Theblackarrowsindicatethemovementthatwilloccurtoreachthenextstate.
44

Thankstothese47snapshots,thepopula.onofthedifferentstatescanbeploRedonadiagram,asafunc.onof.me.ThepresentdiagramshowsthatthemechanismofPYPspanatleastfromthenstos.mescale,thepB2state,withchangeintheter.arystructure(movementofanhelix),beingthoughttosignalthephotonabsorp.ontothebacteria.However,itisclearthatsomechangeshavealreadyoccurredatthens.mescale,atthatshorter.mescalesarerequiredtoknowthefullstory.
45

In2012,a.meresolvedstudyat100ps,thesmaller.mescalepossiblewithpresentsynchrotrons,hasbeencarriedoutonPYP.Themainmessageprovidedbytheelectrondensitymapsshownhereisthatthetrans-to-cistransi.onhasalreadyoccurredaÇer100ps,confirmingthatthistransi.onoccursveryrapidlyaÇerphotonabsorp.on.
46

AbeReriden.fica.onoftheearlystepscanbemade,andaslightlydifferentschemeofthereac.oncycleisproposed.TwopRstatesareproposed,pR1andpR2,thatcorrespondtoabeRer.meresolveddescrip.onofthepRcwtransi.ondescribedinthepreviousstudy.Here,theotherconforma.onofthepRstateisnotobserved,illustra.ngpossiblevaria.onbetweensamples.
47

Another100psstudy,in2013,confirmsthattwopathwaysexistsforpRstates,withtwodifferentconforma.ons,similartotheonesproposedinthe2005study.
48

Theexamplespresentedbeforeshowedthebest.meresolu.onthatcanbeobtainedwithsynchrotronsX-raysources.However,anewsourceofX-ray,theX-rayfreeelectronlaser(X-FEL),isemerging.Afewarealreadyopera.ngintheworld(Japan,USA)andonehasjuststartedinEurope(Hamburg)inseptember2017.Themainadvantagesare(i)theextremepeakbrilliancethatcanbeobtained(1035photons/s/mrad2/mm2/0.1%bw;today’ssynchrotronsareoffering1026,andshouldbeabletoreach1033intwoyears),and(ii)theshortX-raypulse:10fsinsteadof100ps:againof4ordersofmagnitudein.mescale.
49

ThankstoaverywellfocusedX-raybeam(≈1µm–100nm)andthehighbrilliance,structuraldatacanbecollectedonverysmallcrystals(1µmto100nmineachdimension).Serialcrystallographyhasthusbeendeveloped.Usingasinglecrystaltoobtainacompletedatasetisnolongerpossible,sincetheX-raydosereceivedbyacrystalistoohighandbasicallydestroyit.Eachcrystalisusedtocollectonlyonediffrac.onimage.Sincethepulseisveryshort(10-100fs),datacanbecollectedbeforedamagesoccur.Actually,nodamageinducedbytheintensepulseisobservedonthediffrac.onpaRern.Thedamageswilloccuratalarger.mescale,whenthedataarealreadycollected.Tocollectacompletedataset,wethusneedalotasimilarsmallcrystals,thatareinjectedasacon.nuousflowtroughthelaserandX-raybeams.Thissetupenablestoinves.gateirreversiblereac.ons,sinceeachcrystalisusedonlyonce.
50

WithX-FELandserialcrystallography,thechallengeistocollectdatabeforedestruc.on.Aseminalpaperin2000showedthatamolecule,becauseoftheiner.aassociatedtoatomicmovements,cannotexplodebeforeafewtensoffs.Thus,isthepulseisshortenough,itsintensitycanbeashighasneeded.Theyalsoes.matedthatstudyonasinglemoleculeinsteadofacrystalcanbeenvisagedifthebrilliancereachesatleast1040.Wearenotthereyet,butnotthatfarIfprogressinX-raysourcearecon.nuingatthesamerate,wemayhopetogetsuchsourcesin5to10years..
51

OneofthefirststructuralstudydonewithanX-FELsource,onphotosystemI,in2011.Theyexperimentallyconfirmedthataproteincrystalcouldresista700MGydoseifthepulsewasshortenough(<100fs),whileinprinciple,adoseof30MGyisconsideredasthemaximumpossibleformostproteincrystalsstudiedonasynchrotronsource.Ofcourse,serialcrystallographyischallenging,ifoneconsiderthequan.tyofcrystalsneeded,thenumberofimagestobecollectedanddataprocessing.Butthisstudydemonstratedthatitwasactuallyfeasible.Andmuchprogresseshavebeenperformedsince.
52

Anotherstep(theul.mate?)in.meresolvedstructuralinves.ga.onofPYP,at100fsin2016usingX-FEL.Here,theLauetechniqueusedwithsynchrotronisnopossible,astheX-raysaremonochroma.c.However,sincecrystalsareexposedonlyonce,themonochroma.capproachcanbeusedwithoutanyproblems.Thetechnicaldataoftheexperimentsareshownontheslide.Inthisstudy,theauthorshavefirstmadeaposi.vecontrolwithadelayof200nsbetweenlaserpulseandX-raypulsetoconfirmtheresultsobtainedinpreviousstudiesmadewithsynchrotronsources.Thus,thenewsetupisnotintroducingunexpectedar.facts.ThepresenceoftwoRstates,R1(β1andβ2features)andR2(γ1andγ2features),withtwodifferentconforma.onshasbeenconfirmed.
53

Before500fs,thechromophoreremainsintransconforma.on,butslightmovementsareobserved.Thecisconforma.onisclearlyseenaÇer500fs.Thelife.meoftheelectronicexcitatedstateisafewhundredfemtosecond.Thereturntoelectronicgroundisassociatedtotran-cistransi.on.Ifthereac.oniselectronicallytriggeredinabout100fs,itsthermallydrivenaÇerwardsonamuchwider.mescale.Theenergyofonebluephoton(2.76eV)isten.melargerthantheonerequiredtomovethechromophore(0.2eV)
54

X-FELX-raysourcescanreallyenabletoexploretheveryearlyeventsofthechromophoreac.va.on,witha.mescalecloseto100fs.Andthe.meatwhichthetrans-cistransi.oncanbedeterminedprecisely.Thesuccessionofelectronicandstructuraleventscanthusbusobservedoveralarge.merange.Asforhemoglobin,understandingthephotoac.va.oncycleofPYPrequirestoinves.gateawiderangeof.mescale.Here,itspansover13orderofmagnitudes,fromthetrans-to-cistransi.onoftheC1-C2-C3-C1’dihedralofthechromophore,in≈500fs,tochangesinter.arystructure(movementsofhelices)thattakeplaceinthe10ms–1s.mescale,thatcaninturntriggerbacteriasignalingpathways.Thankstothesenewtools,thefullcycleamanyreac.onsmaybestudiedandalltheuseful.mescalescannowbeinves.gated.
55

SerialfemtosecondcrystallographyhasbeenusedtostudythestructuralchangesoccurringinaphotoswitchablefluorescentproteinfromtheGreenFluorescentProtein(GFP)familly,rsEGFP2.Theseproteinsarewidlyusedincellularimagingandunderstandingtheirmechanismmayhelpinimprovingormodifyingtheircharacteris.csinordertopushthelimitofcellularimaging.AsforPYP,thechangestakingplaceonthechromophorearefast,andthefemto-topicosecondscaleisrequiredtoinves.gatethisphotochemicalreac.on.
56
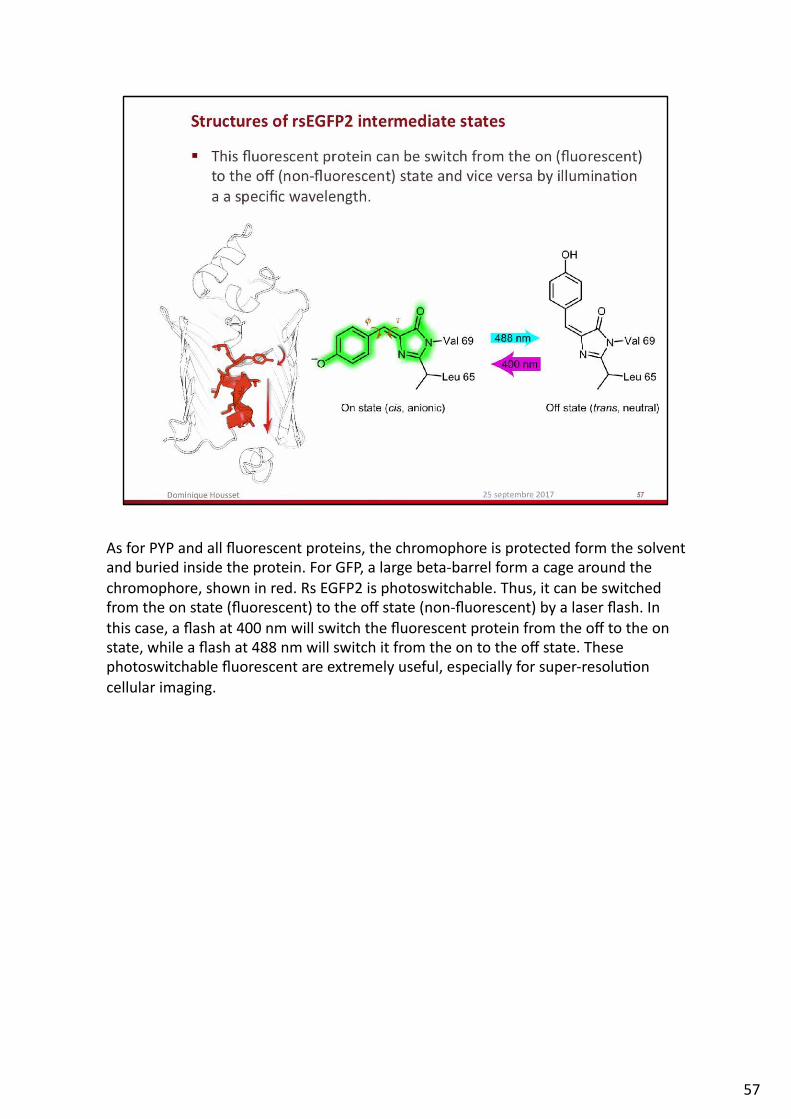
AsforPYPandallfluorescentproteins,thechromophoreisprotectedformthesolventandburiedinsidetheprotein.ForGFP,alargebeta-barrelformacagearoundthechromophore,showninred.RsEGFP2isphotoswitchable.Thus,itcanbeswitchedfromtheonstate(fluorescent)totheoffstate(non-fluorescent)byalaserflash.Inthiscase,aflashat400nmwillswitchthefluorescentproteinfromtheofftotheonstate,whileaflashat488nmwillswitchitfromtheontotheoffstate.Thesephotoswitchablefluorescentareextremelyuseful,especiallyforsuper-resolu.oncellularimaging.
57

Asforpreviousstudies,a.meresolvedstructuralstudycombinesabsorp.onspectroscopy,structuralinves.ga.onandpossiblysimula.ons.Here,ultrafasttranscientabsorp.onspectroscopymadeatdifferent.melapseaÇeralaserpulseof150fs(thattriggerthephotoac.vatedswitchreac.on).Here,thedifferenceabsorp.onspectraisshownatdifferent.mesaÇerthelaserpulse.Thereferencespectraistheonewithlaseroff,i.e.theonecorrespondingtotheoffstate.Thedifferencespectrareachhismaximumatabout0.5ps.Itshowseitherdeple.onorpopula.onofdifferentspecies(iden.fiedbyagivenwavelength)asafunc.onof.me.
58

Eachstatehasaspecificabsorp.onsignatureanditspopula.onordeple.oninfunc.onof.mecanbefollowed,witharesolu.onin.meof≈150ps.Aglobalfitwith3exponen.alsandaconstant,convolutedbyaGaussianshapedpulseof150fshasbeenmadeandhasprovided3.meconstantsof90fs,0.9psand3.7ps.Thankstothisanalysis,a“reac.on”schemecanbeproposed.The90fs.mescalecorrespondtothebuildupoftwointermediatestates,I*1andI*2.ThenthedecayofI*1andI*2towardstrans-offstateandcis-onstate,respec.velyisassociatedwith.meconstant0.92and3.65ps.Indeed,thedecayofthenega.vebroadpeak(500-700nm)hadtobefitwithtwoexponen.als(orange(530nm)andred(650nm)curves),implyingtheexistenceoftwoexitedstates,I*1andI*2.At20ps,thecis-protonatedstateispopulatedanditstrasi.ontothecis-anioniconstatetakeplaceatmuchlonger.mescales(>ns).
59

Thestructuresofthenn,offandtwoexcitedstates(at1psand3ps)weredeterminedbeseraialfemtosecondX-raycrystallography.
60

∆1ps:P(Planar)andT(Twisted)statecorrespondtoabout6-7%.Thisdemonstratethatcrystallographyisabletoprovidepreciseinforma.ononstatesthatrepresentonlyaminorfrac.onofthecrystalcontent.Rela.veoccupancyisabout0.6forPand0.4forT.PisclosetotheoffstatewhileTishalfwaybetweenoffandonstate.Tosummarize,at1ps,twoexcitedstatescanbeobserved,PandT,alongwiththetrans-offstate;At3ps,thePexcitedhasalreadydisappeared,theexcitedTstateiss.llthere,butpar.allydepleted.Thecis-onstatehasbeenpopulatedandthetrans-offstatehasbeenrepopulated,infullagreementwiththereac.onschemeofslide59.
61

Simula.onsbyQM/MMandmoleculardynamicsapproachesareconsistentwithexperimentalresults.Thetranstocistransi.onoccursviathehula-twistmo.on(seeslide48onPYP).This.meresolvedstructuralstudyhasallowedtoiden.fyamutantofrsEGFP2,V151A,whichhasabeRerquantumyield.
62

ThemainideaistouseX-FELor4thgenera.onsynchrotronX-raysourcescombinedwithserialcrystallographytoinves.gatereac.onstriggeredbydiffusionofthesubstrate.Theproblemisthatthediffusionshouldbemuchfasterthanthereac.on.me,inordertobeabletohaveallthemoleculesofthesamplesynchronized.Typically,enzyma.creac.onwillbeatthems.mescaleorslower.In2013Schmidtcalculatedthatthediffusion.meofaligandinacrystaldrama.callydependonthesizeofthelaRer,withtheseordersofmagnitude:10sfor300x400x500µm3;15msfor10x20x30µm3,1msfor3x4x5µm3,150µsfor1x2x3µm3,17µsfor0.5x0.5x0.5µm3,1.5µsfor0.1x0.2x0.3µm3Thus,microornanocrystalsseemssuitableforsuchstudiesatthems.mescale.Moreover,smallcrystalscanacceptlargerstructuralchangesofthemoleculeswithinthecrystal.
63

Theassocia.onofX-FELandserialcrystallographyallowtocollectdataonsuchmicroornanocrystals,atroomtemperature,withnoneedtoreusethesamecrystaltwice(non-reversiblereac.oncannowbeinves.gated).Deviceshavebeendevelopedtodeliverthecrystalinwhichthesubstratehasdiffusedindue.me(alsofasterthanthereac.on.mescale):seenextslide.ComparisonoftheMixandinjectsetup(leÇ)andthelasertriggeredsetup(right)areshownontheslide.Oneshouldnotethatinthecase“Mixandinject”approach,synchrotronsourcesthatproduce≈150psX-raypulsescanbeusedsincethepulseismuchshorterthanthe.mescaleofthereac.oninves.gated.
64

Hereisthefirststudypublishedusingthe“Mixandinject”technique,appliedonariboswitch.
65

First,thestructurewasdeterminedintheapostate.Sincetheasymmetricunitcontainedtwoindependentmolecules,twodifferentconforma.onsoftheriboswitchcouldberefined.Intheapostate,theriboswitchcanadoptanensembleofconforma.onsandthecrystalhasstabilizedtwoofthem.Inthepresentcaseonlyoneseemsabletobindtheligand,theadeninmolecule.
66

ThestructureofthisriboscwitchhasthenbeendeterminedusingX-FELserialcrystallography10sand10minaÇermixingtheribiswitchcrystalsandadenin.AÇer10s,onlyoneconforma.on,apo2,hasboundtheadeninligand(a&b),withlimitedstructuralchanges.Thiscorrespondstotheintermediateadenin-boundstateIB.Theconforma.onapo1,A23preventthebindingofadenin(c).
67

AÇer10min,bothriboswitchmoleculesoftheasymmetricunithavetheanadeninboundtothem.Largestructuralchangeshaveinducedbythebindingofadenin.Thestructuralchangesaresoimportantthanthecrystallaucehaschanged:thespacegroupintheapoformisP21(2moleculesperasymmetricunit)whilethespacegroupwhenadeninisboundtoallriboswitchmolecules(aÇer10minofadeninpresence)isP21212,(1moleculeintheasymmetricunit).Surprisingly,thischangetakesplacewithoutdestroyingthecrystallineorder.Aplotofthepopula.onofthedifferentspecies,asafunc.onof.mecanthenbemade.Thistechniquenotonlyprovidesthestructureofintermediatestates(aswiththecryo-trappingtechnique),italsoprovidesdetailedkine.cinforma.onaboutthereac.oninves.gated.
68

Asmen.onedintheintroduc.on,.meresolvedstudiescanalsobydonebySAXSorWAXStechniques.Thesetupisquitesimilartotheoneusedforserialcrystallography,exceptthatproteinisinsolu.oninsteadofbeingpackedinsmallcrystals.SAXSorWAXSexperimentscanonlyprovideinforma.ononglobalchangesoftheprotein(changesinradiusofgyra.on,shapeforSAXS,changesindomainandsecondarystructuresforWAXS).Sincesolu.onstudiesonlyprovide1Dinforma.on(ascaReringcurve),insteadof3Dforcrystallography(a3Delectrondensitymap),modelingisrequiredtointerpretchangesinscaReringcurves.
69

Here,thedifferencescaReringcurvesSlight(q,Δt)-Sdark(q)atdifferent.messhowthatthemajorvaria.onsoccurfor1.5<q<2.5Å-1,indica.ngchangeswithindomains,i.e.movementofsecondarystructures,inthe3–300ps.mescale.Usingmoleculardynamics,thesechangescanbeexplainedbymovementthetransmembranehelices,asshownonthefigure.
70

Inthis.meresolvedSAXSstudyofbacteriorhodopsin,moderatechangesintheshape(q<0.6Å-1)areobservedbetween360nsand100ms.StructuralmodelingalsosuggestsshiÇsinsomeregionsoftransmembranehelices.
71

72

73

74


















