
The Magnitude of Akt/Phosphatidylinositol 3 -Kinase ... · The Magnitude of...
8
of September 6, 2018. This information is current as CD45 Expression in Human Myeloma Cells -Kinase Proliferating Signaling Is Related to ′ The Magnitude of Akt/Phosphatidylinositol 3 Szpak, Régis Bataille, Nelly Robillard and Martine Amiot Géraldine Descamps, Catherine Pellat-Deceunynck, Yann http://www.jimmunol.org/content/173/8/4953 doi: 10.4049/jimmunol.173.8.4953 2004; 173:4953-4959; ; J Immunol References http://www.jimmunol.org/content/173/8/4953.full#ref-list-1 , 15 of which you can access for free at: cites 31 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2004 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on September 6, 2018 http://www.jimmunol.org/ Downloaded from by guest on September 6, 2018 http://www.jimmunol.org/ Downloaded from
Transcript of The Magnitude of Akt/Phosphatidylinositol 3 -Kinase ... · The Magnitude of...

of September 6, 2018.This information is current as
CD45 Expression in Human Myeloma Cells-Kinase Proliferating Signaling Is Related to
′The Magnitude of Akt/Phosphatidylinositol 3
Szpak, Régis Bataille, Nelly Robillard and Martine AmiotGéraldine Descamps, Catherine Pellat-Deceunynck, Yann
http://www.jimmunol.org/content/173/8/4953doi: 10.4049/jimmunol.173.8.4953
2004; 173:4953-4959; ;J Immunol
Referenceshttp://www.jimmunol.org/content/173/8/4953.full#ref-list-1
, 15 of which you can access for free at: cites 31 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2004 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on September 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on September 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Magnitude of Akt/Phosphatidylinositol 3�-KinaseProliferating Signaling Is Related to CD45 Expression inHuman Myeloma Cells1
Geraldine Descamps,* Catherine Pellat-Deceunynck,* Yann Szpak,* Regis Bataille,*Nelly Robillard, † and Martine Amiot 2*
In multiple myeloma, the Akt/PI3K pathway is involved in the proliferation of myeloma cells. In the current study, we haveinvestigated the impact of the CD45 phosphatase in the control of Akt/PI3K activation. We show that Akt activation in responseto insulin-like growth factor-1 (IGF-1) is highly variable from one human myeloma cell line to another one. Actually, Aktactivation is highly related to whether CD45 is expressed or not. Indeed, both the magnitude and the duration of Akt phosphor-ylation in response to IGF-1 are more important in CD45� than in CD45� myeloma cell lines. We next demonstrate a physicalassociation between CD45 and IGF-1 receptor (IGF-1R) suggesting that CD45 could be involved in the dephosphorylation of theIGF-1R. Furthermore, the growth of CD45� myeloma cell lines is mainly or even totally controlled by the PI3K pathway whereasthat of CD45� myeloma cell lines is modestly controlled by it. Indeed, wortmannin, a specific PI3K inhibitor, induced a dramaticgrowth inhibition in the CD45 � myeloma cell lines characterized by a G1 growth arrest, whereas it has almost no effect on CD45�
myeloma cell lines. Altogether, these results suggest that CD45 negatively regulates IGF-1-dependent activation of PI3K. Thus,strategies that block IGF-1R signaling and consequently the Akt/PI3K pathway could be a priority in the treatment of patientswith multiple myeloma, especially those lacking CD45 expression that have a very poor clinical outcome.The Journal of Im-munology, 2004, 173: 4953–4959.
M ultiple myeloma (MM)3 is a rapidly fatal plasma cellmalignancy that evolves mainly in the bone marrow.MM is a very heterogeneous disease in that the clinical
features and subsequent outcome are quite variable because thesurvival of patients may range from a few days to�15 years (1).IL-6 is known to be an essential growth and survival factor in thismalignancy (2, 3). Besides this well-characterized growth factor, itis now clear that insulin-like growth factor-1 (IGF-1) also plays animportant role in the growth and survival of human myeloma celllines (HMCLs) (4–6). In MM, activation of the IGF-1 receptor(IGF-1R) induces both insulin receptor substrate-1 (IRS-1) andShc activation that ultimately results in activation of both thePI3K/Akt and the MAPK signaling cascades (5). Indeed, activa-tion of IRS-1 triggers its association with and activation of PI3K.Activation of PI3K leads to generation of phosphatidylinositol andsubsequent to activation of Akt by phosphorylation on Thr308 and
Ser473. Although the Thr308 phosphorylation strictly governs theactivation of Akt, the Ser473phosphorylation is necessary to obtaina fully active multiphosphorylated Akt enzyme (7). A significantsupport for a role of PI3K/Akt pathway as a potential mediator oftumor expansion in MM was recently demonstrated. Indeed, se-lective inhibition of the Akt pathway resulted in inhibition of MMcell proliferation (8) or in sensitization to apoptosis (9). It wasshown that Ser473Akt phosphorylation was frequently activated inMM cells and the frequency of activation correlated with diseaseactivity (10). Phosphatase and tensin homolog deleted on chromo-some 10 (PTEN) is a recently identified tumor suppressor gene thatencodes a phosphatase that mainly dephosphorylates phosphati-dylinositol 3,4,5-trisphosphate and is an important negative regu-lator controlling the Akt activation (11). It was recently shown thatloss of PTEN expression is responsible for uncontrolled Ser473Aktphosphorylation in two HMCLs (12). The critical role of PTEN intumor growth was clearly demonstrated for OPM-2 HMCL inSCID mice. The lack of PTEN expression in OPM-2 cells facili-tated in vivo tumoral growth in 100% of SCID mice. Furthermore,tumor growth could be reversed in a dose-dependent fashion byincreasing amount of PTEN (13).
IGF-1R has a high degree of homology with the insulin receptor,and their cytoplasmic signaling after receptor autophosphorylationappears to be similar. In the U266 HMCL, it has been demon-strated that CD45, a transmembrane protein tyrosine phosphatase,can negatively regulate insulin receptor signal transduction (14)and that the expression of CD45 is induced by IL-6 stimulation(15). In other cell types, such as HepG2 or kidney fibroblast cells,CD45 functions as a negative modulator of IGF-1 signaling (16,17). Based on these data, it is conceivable that CD45 could actdirectly on IGF-1 signaling in MM cells. Thus, it appears thatPI3K/Akt activation could be very heterogeneous in MM and un-der the influence of several phosphatases like PTEN and CD45.
*Institut National de la Sante et de la Recherche Medicale Unite 601, Ligue Nationalecontre le Cancer Label, and†Laboratoire d’Hematologie, Institut de Biologie, Nantes,France
Received for publication November 21, 2003. Accepted for publication August2, 2004.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by the Ligue Nationale Contre le Cancer (equipe labelise´e2001).2 Address correspondence and reprint requests to Dr. Martine Amiot, U601, Institutde Biologie, 9, Quai Moncousu, 44093 Nantes cedex 01, France. E-mail address:[email protected] Abbreviations used in this paper: MM, multiple myeloma; HMCL, human myelomacell lines; IGF-1, insulin-like growth factor-1; IGF-1R, IGF-1 receptor; IRS-1, insulinreceptor substrate-1; MFR, mean fluorescence ratio; PTEN, phosphatase and tensinhomolog deleted on chromosome 10; Akt-P, Akt phosphorylation; Erk-P, Erkphosphorylation.
The Journal of Immunology
Copyright © 2004 by The American Association of Immunologists, Inc. 0022-1767/04/$02.00
by guest on September 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Because the role of PTEN in Akt activation was clearly demon-strated, the purpose of this study was to elucidate the impact ofCD45 expression in the control of the PI3K/Akt pathway. Theexpression of CD45 in myeloma cells is directly related to theirproliferation rate and differentiation status (18, 19). The heteroge-neity of CD45 expression is also found on HMCLs, thus CD45�
and CD45� HMCLs have been identified, leading to an analysis ofPI3K/Akt pathway for each group of cell lines.
Materials and MethodsmAbs and reagents
Anti-actin was purchased from Chemicon International (Temecula, CA).Anti-phospho-Akt (Thr308), anti-PTEN (A2B1), and anti-IGF-1R� arefrom Santa Cruz Biotechnology (Tebu-Bio, Le Perray en Yvelines,France.) Anti-phospho Akt (Ser473) and anti-phospho-p44/42 MAPK fromCell Signaling Technology (Ozyme, Saint Quentin Yvelines, France). Anti-
STAT3 and anti-CD45 are from BD Biosciences (Le Pont de Claix,France). Wortmannin and LY294002 were from Alexis (Tebu-Bio). Hu-man recombinant IGF-1 and vanadate were purchased from Sigma-Aldrich(St. Louis, MI).
HMCLs and culture conditions
The XG-1, XG-6, BCN, MDN HMCLs have been previously established inour laboratory and are cultured in the presence of 3 ng/ml r-IL-6 (NovartisPharmaceuticals, Basel, Switzerland). LP-1, L363, and NCI-H929 HMCLswere purchased from Deutsche Sammlung von Mikroorganismen andZellkulturen (Braunschweig, Germany) and U266 from the American TypeCulture Collection (Manassas, VA). Cell lines were maintained in RPMI1640 medium supplemented with 10% FCS, 2 mM glutamine, antibiotics,and 5 � 10�5 M 2-ME.
Immunofluorescence analysis
Cells (0.5 � 106) were sampled in each condition and washed with PBSbefore incubation with anti-CD45-FITC (Immunotech, Marseilles, France)
FIGURE 1. A, PTEN expression in CD45� and CD45� HMCLs. Equivalent amounts of cell lysates were separated by SDS-PAGE, then immunoblottedwith anti-PTEN Ab. B, Akt phosphorylation induced by IGF-1 is stronger in CD45� than in CD45� HMCLs. Serum-starved cells for 18 h were untreatedor treated with IGF-1 for 30 min. Equivalent amounts of cell lysates were separated by SDS-PAGE, then immunoblotted with anti-phospho-Akt Abs. Proteinloading was controlled with an anti-actin. Relative Akt-P in each cell line compared with the highest value of BCN was calculated using densitometry.
4954 Akt SIGNALING IS DEPENDENT ON CD45 EXPRESSION IN MYELOMA CELLS
by guest on September 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

or IGF-1R-PE (BD Biosciences) for 20 min. After two washes, cells werefixed in 1% formaldehyde. Flow cytometry analysis was performed on aFACSCalibur using the CellQuest program (BD Biosciences, San Jose,CA). The ratio of fluorescence was determined by dividing the mean flu-orescence intensity by the mean fluorescence intensity of the respectivecontrol.
Cell viability
Cell viability was determined by vital dye (0.4% eosin) exclusion andassessed by visual inspection in an hemocytometer.
Preparation of pervanadate solution
The vanadate solution was prepared in incubation buffer (30 mM HEPES(pH 7.5), 150 mM NaCl, 4 mM KCl, 0.8 mM MgSO4, 1.8 mM CaCl2, 10mM glucose). Pervanadate was obtained by mixing vanadate with 1 mMH2O2 for 15 min at 22°C. This was followed by the addition of catalase(200 �M) to remove residual H2O2.
Immunoblot analysis
A total of 4 � 106 cells were resuspended in lysis buffer (10 mM Tris-HCl(pH 7.6), 150 mM NaCl, 5 mM EDTA, 1 mM PMSF, 2 mM Na3VO4, 1mM NaF, 2 �g/ml aprotinin, leupeptin 1 �g/ml, and 0.5% Nonidet P-40).After 40 min on ice, lysates were cleared by centrifugation at 12,000 � gfor 30 min at 4°C. Protein concentration was measured using bicinchoninicacid (BCA protein assay; Pierce, Rockford, IL). A total of 100 �g ofproteins was loaded for each lane. The proteins were separated by 10%SDS-PAGE and then electrotransferred to polyvinylidene difluoride mem-branes. Western blot analysis was performed by standard techniques withECL detection (Pierce).
Immunoprecipitation
Cells (20 � 106) were lysed in 1% CHAPS-containing lysis buffer. Wholecell lysates were obtained, precleared with protein A-Sepharose, and then
incubated overnight with 2 �g of the specific Ab. The immunocomplexeswere captured with protein A-agarose. Beads were pelleted, washed threetimes, and boiled in SDS sample buffer. The presence of immune com-plexes was determined by Western blotting.
Cell cycle distribution
For cell cycle analysis, cells were fixed in 70% cold ethanol for 30 min at4°C, washed twice in PBS, and stained with propidium iodide (20 �g/ml)for 10 min at room temperature. Flow cytometry analysis was performedon a FACSCalibur using CellQuest Pro software (BD Biosciences). Datawere gated on the FL2-Area vs FL2-Width cytogram to excluded doubletsand aggregates, and a minimum of 2 � 104 gated cells were collected persample. Analysis of the cell cycle was performed using the Modfit LT (BDBiosciences).
Statistical analysis
For statistical analysis, we used the nonparametric Wilcoxon rank sum testor the Student t test.
ResultsAkt activation is higher in CD45� than in CD45� humanmyeloma cells regardless of their expression of both IGF-1Rand PTEN
Given the potential role of CD45 as a negative regulator of IGF-1signaling in HepG2 (14), we searched for a correlation between theexpression of CD45 and the level of Akt-P in MM cells. We se-lected eight HMCLs to analyze Akt activation in response toIGF-1, three expressing CD45 on 100% of cells (XG-1, XG-6,MDN), one expressing CD45� on a majority of cells (�90%)(U266), and four lacking CD45 expression (LP-1, NCI-H929,BCN, L363). Of note, all these cell lines express PTEN proteins asshown by Western blotting analysis using an Ab recognizing the Cterminus of PTEN (Fig. 1A). Following overnight serum starva-tion, we investigated Thr308 and Ser473 Akt phosphorylation(Akt-P) in response to IGF-1. The level of Akt-P was variable, thehighest levels for both Ser and Thr phosphorylation were observedfor the four CD45� cell lines. In contrast, CD45� HMCLs had aweak to a moderate level of Akt-P, especially for the Thr308 phos-phorylation in which the difference is significant ( p � 0.05). In
FIGURE 2. IGF-1R expression on HMCL. Cells were stained with iso-type-matched control-PE mAb (black histogram) or with anti-IGF-1R PE(gray histogram).
FIGURE 3. Comparison of Akt-P induced by IGF-1 after pervanadatetreatment in XG-1 CD45� and LP-1 CD45�. Serum-starved cells for 18 hwere treated or not by 100 �M pervanadate before being stimulated 30 minby IGF-1. Equivalent amounts of cell lysates were separated by SDS-PAGE, then immunoblotted with anti-phospho-Akt. Protein loading wascontrolled with an anti-actin. Akt-P was quantified by densitometric scan-ning and presented as the percentage of Akt-P in response to IGF-1 withoutpervanadate treatment.
4955The Journal of Immunology
by guest on September 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

conclusion, the level of Thr308 Akt-P appears to discriminateCD45� from CD45� cell lines.
Then, the functional impact of the expression of the IGF-1R onAkt-P was evaluated by determining the level of IGF-1R by flowcytometry. The results in Fig. 2 indicate that all cell lines expressthe IGF-1R. Whereas LP-1 has the highest level of IGF-1R (meanfluorescence ratio (MFR) � 11), and BCN and MDN have theweaker level (MFR � 1.8), the ratio of IGF-1R expression is al-most identical for other cell lines (3.2 � MFR � 4). Thus, there isno discernable association between the level of IGF-1R expressionand the level of Akt-P.
Pervanadate treatment dramatically increases Akt-P in CD45�
HMCL whereas it has no effect in CD45� HMCL
Pervanadate, a well-known powerful inhibitor of tyrosine phos-phatases, inhibits CD45 phosphatase activity at the concentration
of 100 �M (20). XG-1 (CD45�) and LP-1 (CD45�) were treatedby pervanadate before IGF-1 stimulation. In XG-1, both Ser473 andThr308 Akt-P were dramatically increased by 3- and 2-fold, re-spectively, whereas in LP-1 only a very weak increase was ob-served (Fig. 3). Thus, these results indicate that Akt-P is controlledby a phosphatase in CD45� cell lines only.
Induction of CD45 expression in LP-1 HMCL reduced theIGF-1 signaling
Because IL-6 has been shown to induce CD45 expression on U266(15), LP-1 cells were cultured with 10 ng/ml IL-6. After twomonths of culture with IL-6, LP-1 expressed CD45 on 50% ofcells, as shown in Fig. 4A. In LP-1 IL-6-treated cells, both Ser473
and Thr308 Akt-P was reduced by 50 and 25%, respectively, andconversely, Erk phosphorylation (Erk-P) was also reduced by 32%(Fig. 4B). These data clearly indicate that PI3K/Akt and ERK/
FIGURE 4. Comparison of both Akt-P and Erk-Pinduced by IGF-1 in LP-1 cultured or not with IL-6.A, Flow cytometry analysis of CD45 expression inLP-1 cultured with or without IL-6. B, Serum-starvedLP-1 or IL-6-treated LP-1 cells for 18 h were treatedwith IGF-1 for 30 min. Equivalent amounts of celllysates were separated by SDS-PAGE, then immuno-blotted with anti-phospho-Akt and anti-phospho-p44/42MAPK. Protein loading was controlled with an anti-actin. Akt-P and Erk-P were quantified by densitometricscanning and presented as the percentage of Akt-P orErk-P in response to IGF-1 in LP-1.
FIGURE 5. Kinetic of Akt-P after IGF-1 stimulation. A, Serum-starved cells were stimulated by IGF-1 for 30 min then washed and incubated for varioustimes in RPMI 1640 0.5% BSA. Equivalent amounts of cell lysates were separated by SDS-PAGE, then immunoblotted with anti-phospho-Akt. B, Akt-Pwas quantified by densitometric scanning and presented as the percentage of Akt phosphorylated at t � 0 for each HMCL.
4956 Akt SIGNALING IS DEPENDENT ON CD45 EXPRESSION IN MYELOMA CELLS
by guest on September 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

MAPK signaling in response to IGF-1 are reduced in CD45� celllines.
In response to IGF-1, the kinetic of Ser473 Akt activation differsin CD45� from CD45� HMCLs
To compare the kinetic of Akt-P in response to IGF-1 in bothCD45� and CD45� HMCLs, we selected two CD45� (XG-6,XG-1) and two CD45� (LP-1, L363) cell lines. After 30 min ofIGF-1 stimulation, cells were washed and incubated in RPMI 16400.5% BSA for different times. In response to IGF-1, the Thr308
Akt-P was short lasting in both CD45� and CD45� HMCL but thepersistency of the Ser473 Akt-P was very different between CD45�
and CD45� cell lines (Fig. 5). Indeed, after 60 min the level ofSer473 Akt-P was almost undetectable in the CD45� XG-6 andXG-1 cells (�3% of the initial signal), whereas 30% and 36% ofthe initial signal was still observed in CD45� LP-1 and L363 cells,respectively. Moreover, in the CD45� HMCLs, Ser473 Akt-P stillremained detectable 3 h after IGF-1 stimulation. These resultsdemonstrate that Ser473 Akt-P remained activated for a long time(�3 h) after IGF-1 stimulation in CD45� HMCLs in contrast toCD45� HMCLs, where Ser473 Akt-P returned to baseline level inaround 1 h.
IGF-1R is physically associated to CD45 in myeloma cells
Because CD45 molecules modulate IGF-1 signaling, we wonderedwhether CD45 and IGF-1R could be physically associated. Immu-noprecipitation assays were conducted using Abs directed toIGF-1R or STAT3 as a control in XG-6 cells. The results shown inFig. 6 demonstrate that CD45 is coimmunoprecipitated with theIGF-1R, whereas CD45 is not found associated to STAT3. Thephysical interaction between IGF-1R and CD45 let us think thatCD45 could directly dephosphorylate IGF-1R or linked molecules.
Inhibition of proliferation by wortmannin is stronger in CD45�
than in CD45� HMCLs
To evaluate the consequences of Akt activation, we next comparedthe efficiency of two PI3K inhibitors, i.e., wortmannin andLY294002 in NCI-H929 cells. Wortmannin induced a completeinhibition of Akt-P at the concentration of 1 �M, whereas theinhibition induced by LY294002 at 50 �M was not complete (Fig.7). Consequently, wortmannin appeared more efficient thanLY294002 to inhibit Akt-P confirming the results obtained by Da-vies et al. (21). Moreover, it was shown that LY294002 inhibits theCasein kinase 2 (21), a kinase involved in multiple pathways.
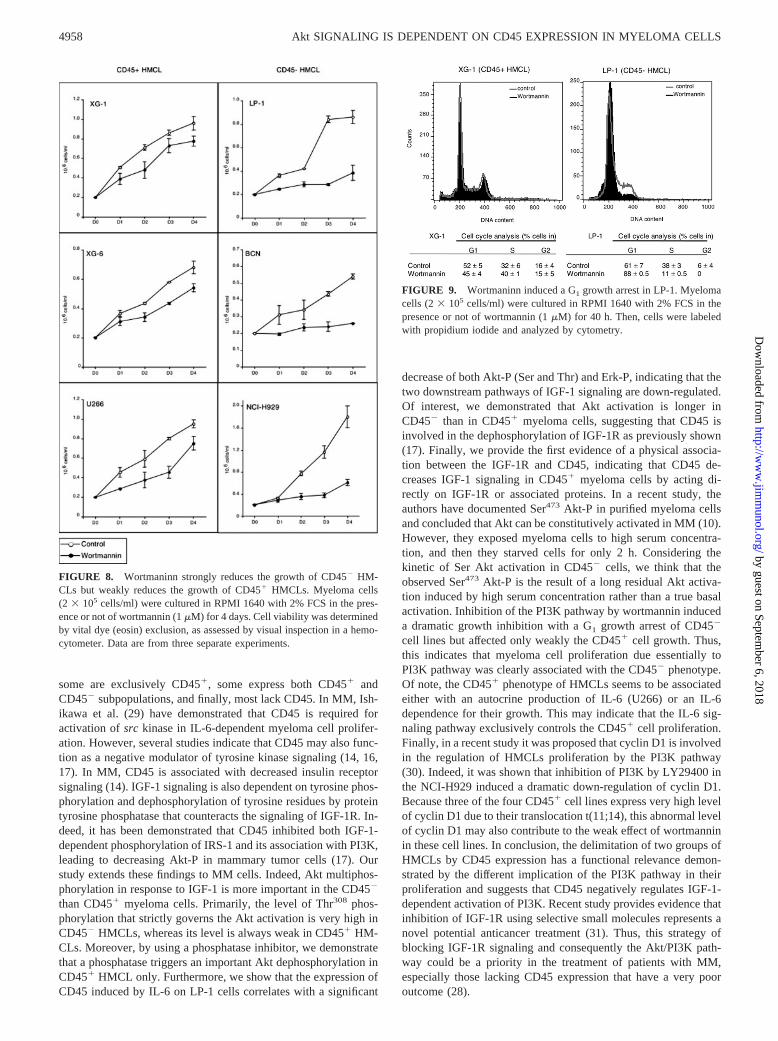
Thus, we selected wortmannin to evaluate the importance of thePI3K pathway in myeloma cell proliferation and survival of threeCD45� cell lines (LP-1, NCI-H929, and BCN) and three CD45�
cell lines (U266, XG-1, and XG-6). Four-day wortmannin treat-ment does not induce significant apoptosis in any of the cell lines(data not shown). In contrast, wortmannin induces an importantgrowth inhibition of CD45� HMCLs of 70, 85, and 75% for LP-1,BCN, and NCI-H929, respectively, whereas this inhibition wasweaker in CD45� cell lines being 21, 25, and 25% for XG-6,XG-1, and U266, respectively (Fig. 8). The growth inhibition issignificantly different between CD45� and CD45� cell lines ( p �0.001). This result indicates that the growth of CD45� cell lineswas mainly controlled by the PI3K pathway, whereas the growthof CD45� cell lines was modestly controlled by the PI3K pathwayand thus correlated well with the magnitude of Akt-P in responseto IGF-1.
Wortmaninn induces G1 growth arrest in CD45� but not inCD45� HMCLs
The effect of wortmannin on the cell cycle was investigated inXG-1 and LP-1. After 40 h of wortmannin treatment, the cell cyclewas unchanged in XG-1 but impaired in LP-1 (Fig. 9). Indeed, inLP-1, S phase was reduced (38 vs 11%) ( p � 0.001) whereas G1
phase was significantly increased (61 vs 88%) ( p � 0.001). Ofnote, no significant hypodiploid DNA peak was observed, indicat-ing that wortmannin did not induce any apoptosis. Altogether,these results indicated that wortmannin induces a significantgrowth arrest in the CD45� HMCLs only.
DiscussionCD45, initially described as leukocyte common Ag, is a transmem-brane protein-tyrosine phosphatase that plays a crucial role forboth T and B cell activation through the corresponding Ag recep-tors (22). CD45 appears to be a positive mediator of cellular ty-rosine phosphorylation by activating members of the src familyprotein-tyrosine kinases (23). The expression of CD45 has beenshown to decline during normal plasma cell differentiation (24). InMM cells, the expression of CD45 is variable and correlates withthe proliferation rate and differentiation status (18, 19, 25). In the5T2MM mouse model, CD45� cell populations have the mostimportant invasive, proliferative, and angiogenic capacities (26,27). As in this model, in humans, the loss of expression of CD45is associated with advanced disease and reflects the phenotype ofmore malignant cells (19). Finally, we recently demonstrated thatthis CD45� phenotype is a strong predictor of poor clinical out-come in humans (28), as in the 5T2MM myeloma model (26). Inour study, we show that HMCLs reflect this heterogeneity because
FIGURE 6. Coimmunoprecipitation of IGF-1R and CD45 on CD45�
HMCL (XG-6). Immunoprecipitations were conducted with agarose-con-jugated IGF-1R� or STAT-3 polyclonal Abs. Western blotting analysiswith an anti-CD45, anti-STAT-3, or anti-IGF-1R Abs was conducted oneach precipitate.
FIGURE 7. Inhibition of Akt-P by PI3K inhibitors in NCI-H929 cellsafter IGF-1 stimulation. Serum-starved cells were incubated or not witheither wortmannin or LY294002 for 150 min, then IGF-1 was added for thelast 30 min. Equivalent amounts of cell lysates were separated by SDS-PAGE, then immunoblotted with anti-phospho-Akt. Protein loading wascontrolled with an anti-actin.
4957The Journal of Immunology
by guest on September 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
some are exclusively CD45�, some express both CD45� andCD45� subpopulations, and finally, most lack CD45. In MM, Ish-ikawa et al. (29) have demonstrated that CD45 is required foractivation of src kinase in IL-6-dependent myeloma cell prolifer-ation. However, several studies indicate that CD45 may also func-tion as a negative modulator of tyrosine kinase signaling (14, 16,17). In MM, CD45 is associated with decreased insulin receptorsignaling (14). IGF-1 signaling is also dependent on tyrosine phos-phorylation and dephosphorylation of tyrosine residues by proteintyrosine phosphatase that counteracts the signaling of IGF-1R. In-deed, it has been demonstrated that CD45 inhibited both IGF-1-dependent phosphorylation of IRS-1 and its association with PI3K,leading to decreasing Akt-P in mammary tumor cells (17). Ourstudy extends these findings to MM cells. Indeed, Akt multiphos-phorylation in response to IGF-1 is more important in the CD45�
than CD45� myeloma cells. Primarily, the level of Thr308 phos-phorylation that strictly governs the Akt activation is very high inCD45� HMCLs, whereas its level is always weak in CD45� HM-CLs. Moreover, by using a phosphatase inhibitor, we demonstratethat a phosphatase triggers an important Akt dephosphorylation inCD45� HMCL only. Furthermore, we show that the expression ofCD45 induced by IL-6 on LP-1 cells correlates with a significant
decrease of both Akt-P (Ser and Thr) and Erk-P, indicating that thetwo downstream pathways of IGF-1 signaling are down-regulated.Of interest, we demonstrated that Akt activation is longer inCD45� than in CD45� myeloma cells, suggesting that CD45 isinvolved in the dephosphorylation of IGF-1R as previously shown(17). Finally, we provide the first evidence of a physical associa-tion between the IGF-1R and CD45, indicating that CD45 de-creases IGF-1 signaling in CD45� myeloma cells by acting di-rectly on IGF-1R or associated proteins. In a recent study, theauthors have documented Ser473 Akt-P in purified myeloma cellsand concluded that Akt can be constitutively activated in MM (10).However, they exposed myeloma cells to high serum concentra-tion, and then they starved cells for only 2 h. Considering thekinetic of Ser Akt activation in CD45� cells, we think that theobserved Ser473 Akt-P is the result of a long residual Akt activa-tion induced by high serum concentration rather than a true basalactivation. Inhibition of the PI3K pathway by wortmannin induceda dramatic growth inhibition with a G1 growth arrest of CD45�
cell lines but affected only weakly the CD45� cell growth. Thus,this indicates that myeloma cell proliferation due essentially toPI3K pathway was clearly associated with the CD45� phenotype.Of note, the CD45� phenotype of HMCLs seems to be associatedeither with an autocrine production of IL-6 (U266) or an IL-6dependence for their growth. This may indicate that the IL-6 sig-naling pathway exclusively controls the CD45� cell proliferation.Finally, in a recent study it was proposed that cyclin D1 is involvedin the regulation of HMCLs proliferation by the PI3K pathway(30). Indeed, it was shown that inhibition of PI3K by LY29400 inthe NCI-H929 induced a dramatic down-regulation of cyclin D1.Because three of the four CD45� cell lines express very high levelof cyclin D1 due to their translocation t(11;14), this abnormal levelof cyclin D1 may also contribute to the weak effect of wortmanninin these cell lines. In conclusion, the delimitation of two groups ofHMCLs by CD45 expression has a functional relevance demon-strated by the different implication of the PI3K pathway in theirproliferation and suggests that CD45 negatively regulates IGF-1-dependent activation of PI3K. Recent study provides evidence thatinhibition of IGF-1R using selective small molecules represents anovel potential anticancer treatment (31). Thus, this strategy ofblocking IGF-1R signaling and consequently the Akt/PI3K path-way could be a priority in the treatment of patients with MM,especially those lacking CD45 expression that have a very pooroutcome (28).
FIGURE 8. Wortmaninn strongly reduces the growth of CD45� HM-CLs but weakly reduces the growth of CD45� HMCLs. Myeloma cells(2 � 105 cells/ml) were cultured in RPMI 1640 with 2% FCS in the pres-ence or not of wortmannin (1 �M) for 4 days. Cell viability was determinedby vital dye (eosin) exclusion, as assessed by visual inspection in a hemo-cytometer. Data are from three separate experiments.
FIGURE 9. Wortmaninn induced a G1 growth arrest in LP-1. Myelomacells (2 � 105 cells/ml) were cultured in RPMI 1640 with 2% FCS in thepresence or not of wortmannin (1 �M) for 40 h. Then, cells were labeledwith propidium iodide and analyzed by cytometry.
4958 Akt SIGNALING IS DEPENDENT ON CD45 EXPRESSION IN MYELOMA CELLS
by guest on September 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

References1. Bataille, R., and J. L. Harousseau. 1997. Multiple myeloma: recent advances in
the biology and management. N. Engl. J. Med. 336:1657.2. Kawano, M., T. Hirano, T. Matsuda, T. Taga, Y. Horii, K. Iwato, H. Asaoku,
B. Tang, O. Tanabe, H. Tanaka, et al. 1988. Autocrine generation and require-ment of BSF-2/IL-6 for human multiple myeloma. Nature 332:83.
3. Klein, B., X. G. Zhang, M. Jourdan, J. Content, F. Houssiau, L. Aarden,M. Piechaczyk, and R. Bataille. 1989. Paracrine rather than autocrine regulationof myeloma-cell growth and differentiation by interleukin-6. Blood 73:517.
4. Georgii-Hemming, P., H. J. Wiklund, O. Ljunggren, and K. Nilsson. 1996. In-sulin-like growth factor I is a growth and survival factor in human multiplemyeloma cell lines. Blood 88:2250.
5. Ge, N. L., and S. Rudikoff. 2000. Insulin-like growth factor I is a dual effector ofmultiple myeloma cell growth. Blood 96:2856.
6. Ferlin, M., N. Noraz, C. Hertogh, J. Brochier, N. Taylor, and B. Klein. 2000.Insulin-like growth factor induces the survival and proliferation of myeloma cellsthrough an interleukin-6-independent transduction pathway. Br. J. Haematol.111:626.
7. Scheid, M. P., and J. R. Woodgett. 2003. Unravelling the activation mechanismsof protein kinase B/Akt. FEBS Lett. 546:108.
8. Tu, Y., A. Gardner, and A. Lichtenstein. 2000. The phosphatidylinositol 3-kinase/AKT kinase pathway in multiple myeloma plasma cells: roles in cytokine-de-pendent survival and proliferative responses. Cancer Res. 60:67.
9. Hideshima, T., N. Nakamura, D. Chauhan, and K. C. Anderson. 2001. Biologicsequelae of interleukin-6 induced PI3-K/Akt signaling in multiple myeloma. On-cogene 20:5991.
10. Hsu, J., Y. Shi, S. Krajewski, S. Renner, M. Fisher, J. C. Reed, T. F. Franke, andA. Lichtenstein. 2001. The AKT kinase is activated in multiple myeloma tumorcell. Blood 98:2853.
11. Cantley, L. C., and B. G. Neel. 1999. New insights into tumor suppression: PTENsuppresses tumor formation by restraining the phosphoinositide 3-kinase/AKTpathway. Proc. Natl. Acad. Sci. USA 96:4240.
12. Hyun, T., A. Yam, S. Pece, X. Xie, J. Zhang, T. Miki, J. S. Gutkind, and W. Li.2000. Loss of PTEN expression leading to high Akt activation in human multiplemyelomas. Blood 96:3560.
13. Ge, N. L., and S. Rudikoff. 2000. Expression of PTEN in PTEN-deficient mul-tiple myeloma cells abolishes tumor growth in vivo. Oncogene 19:4091.
14. Kulas, D. T., G. G. Freund, and R. A. Mooney. 1996. The transmembrane pro-tein-tyrosine phosphatase CD45 is associated with decreased insulin receptorsignaling. J. Biol. Chem. 271:755.
15. Mahmoud, M. S., H. Ishikawa, R. Fujii, and M. M. Kawano. 1998. Induction ofCD45 expression and proliferation in U-266 myeloma cell line by interleukin-6.Blood 92:3887.
16. Way, B. A., and R. A. Mooney. 1993. Activation of phosphatidylinositol-3-ki-nase by platelet-derived growth factor and insulin-like growth factor-1 is inhib-ited by a transmembrane phosphotyrosine phosphatase. J. Biol. Chem.268:26409.
17. Mooney, R. A., G. G. Freund, B. A. Way, and K. L. Bordwell. 1992. Expressionof a transmembrane phosphotyrosine phosphatase inhibits cellular response toplatelet-derived growth factor and insulin-like growth factor-1. J. Biol. Chem.267:23443.
18. Joshua, D., A. Petersen, R. Brown, B. Pope, L. Snowdon, and J. Gibson. 1996.The labelling index of primitive plasma cells determines the clinical behaviour ofpatients with myelomatosis. Br. J. Haematol. 94:76.
19. Bataille, R., N. Robillard, C. Pellat-Deceunynck, and M. Amiot. 2003. A cellularmodel for myeloma cell growth and maturation based on an intra-clonal CD45hierarchy. Immunol. Rev. 193:1.
20. Imbert, V., J. F. Peyron, D. Farahi Far, B. Mari, P. Auberger, and B. Rossi. 1994.Induction of tyrosine phosphorylation and T-cell activation by vanadate peroxide,an inhibitor of protein tyrosine phosphatases. Biochem. J. 297:163.
21. Davies, S. P., H. Reddy, M. Caivano, and P. Cohen. 2000. Specificity and mech-anism of action of some commonly used protein kinase inhibitors. Biochem. J.351:95.
22. Justement, L., V. K. Brown, and J. Lin. 1994. Regulation of B-cell activation byCD45: a question of mechanism. Immunol. Today 15:309.
23. Hermiston, M. L., Z. Xu, and A. Weiss. 2002. CD45: a critical regulator ofsignaling thresholds in immune cells. Annu. Rev. Immunol. 21:107.
24. Medina, F., C. Segundo, A. Campos-Caro, I. Gonzalez-Garcia, and J. A. Brieva.2002. The heterogeneity shown by human plasma cells from tonsil, blood, andbone marrow reveals graded stages of increasing maturity, but local profiles ofadhesion molecule expression. Blood 99:2154.
25. Fujii, R., H. Ishikawa, M. S. Mahmoud, H. Asaoku, and M. M. Kawano. 1999.MPC-1�CD49e� immature myeloma cells include CD45� subpopulations thatcan proliferate in response to IL-6 in human myelomas. Br. J. Haematol.105:131.
26. Vanderkerken. K., K. Asosingh, P. Croucher, and B. Van Camp. 2003. Multiplemyeloma biology: lesson from the 5TMM models. Immunol. Rev. 194:196.
27. Asosingh, K., H. De Raeve, E. Menu, I. Van Riet, E. Van Marck, B. Van Camp,and K. Vanderkerken. 2004. Angiogenic switch during 5T2MM murine myelomatumorigenesis: role of CD45 heterogeneity. Blood 103:3131.
28. Moreau, P., N. Robillard, H. Avet-Loiseau, D. Pineau, N. Morineau, N. Milpied,J. L. Harousseau, and R. Bataille. 2004. Patients with CD45 negative multiplemyeloma receiving high-dose therapy have a shorter survival than those withCD45 positive multiple myeloma. Haematologica 89:547.
29. Ishikawa, H., N. Tsuyama, S. Abroun, S. Liu, F. J. Li, O. Taniguchi, andM. M. Kawano. 2002. Requirements of src family kinase activity associated withCD45 for myeloma cell proliferation by interleukin-6. Blood 99:2172.
30. Pene, F., Y. E. Claessens, O. Muller, F. Viguie, P. Mayeux, F. Dreyfus,C. Lacombe, and D. Bouscary. 2002. Role of the phosphatidylinositol 3-kinase/Akt and mTOR/P70S6-kinase pathways in the proliferation and apoptosis in mul-tiple myeloma. Oncogene 21:6587.
31. Mitsiades, C. S., N. S. Mitsiades, C. J. McMullan, V. Poulaki, R. Shringarpure,M. Akiyama, T. Hideshima, D. Chauhan, M. Joseph, T. A. Libermann, et al.2004. Inhibition of the insulin-like growth factor receptor-1 tyrosine kinase ac-tivity as a therapeutic strategy for multiple myeloma, other hematologic malig-nancies, and solid tumors. Cancer Cell 5:221.
4959The Journal of Immunology
by guest on September 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from


















