
THE JOURNAL OF BIOUXICAL Cmsrnu 269, No. 6, 11, 4626-4633
8
THE JOURNAL OF BIOUXICAL Cmsrnu 0 1994 by The American Society for Bioebemistry and Molecular Biology, Inc. Vol. 269, No. 6, Issue of February 11, pp. 4626-4633, 1994 Printed in USA. Lipoprotein Lipase MOLECULAR MODEL BASED ON THE PANCREATIC LIPASE X-RAY STRUCTURE: CONSEQUENCES FOR HEPARIN BINDING AND CATALYSIS* (Received forpublication, September 16, 1993) Herman van TilbeurghS, Main Roussel, Jean-Marc Lalouelln, and Christian Cambillaull From the Laboratory of Cristallisation et Macromolecules Biologiques-Centre National de la Recherche Scientifique, Facultk de Mbdecine Nord, 13916 Marseille Cedex 20, France and the $Howard Hughes Medical Institute, University of Utah Health Sciences Center, Salt Lake City, Utah 84132 Lipoprotein lipase and pancreatic lipase have about 30% sequence identity, suggesting a similar tertiary fold. Three-dimensionalmodelsoflipoproteinlipasewere constructed, based upon two recently determined x-ray crystal structures of pancreatic lipase, in which the ac- tive site was in an open and closed conformation, respec- tively. These models allow us to propose a few hypoth- eses on the structural determinants of lipoprotein lipase which are responsible for heparin binding, dimer forma- tion, and phospholipase activity. The folding of the pro- tein assembles a number of positive charge clusters at the back of the molecule, opposite the active site. These clusters probably form the heparin binding site, as con- firmed by recent site-directed mutagenesis experi- ments. The active sites of lipoprotein lipase and pancre- atic lipase look very similar, except for the lid (a surface loop covering the catalytic serine in the inactive state). A different open (active) conformation of the lid in both enzymes may be responsible for their differing substrate specificities. Predictions of the nature of the lipoprotein lipasedimerremainelusive,althoughourmodelen- abled us to propose a few possibilities. Lipoprotein lipase is responsible for the hydrolysis of plasma lipids in very low density lipoproteins and chilomicrons and causes transfer of fatty acids to tissues (1, 2). It has a high affinity for heparin and sulfate proteoglycans, explaining its physiological location on the endothelial cell wall of capillaries and on the surfaceof adipose tissue cells. Lipoprotein lipase is a noncovalent dimer and is activated by apolipoprotein C-11, a small protein of 79 residues (3). Several distinctregions of the enzyme must be involved in these interactions. Pancreatic lipase is secreted in the duodenum and is active on dietary triglycerides. The pancreatic lipase structure con- sists of two distinct domains (4). The N-terminal domain (resi- dues 1-335) containing the catalytic triad is made of a central p-sheet surrounded by a-helices (the so-called dp-hydrolase fold) (5). The active site is shielded by a surface loop that moves over 30 A upon substrate binding (6). The C-terminal domain (residues ‘336449) has a p-sandwich fold and forms the main * This work was supported by the EEC BRIDGE-Lipase project, the Centre National de la Recherche Scientifique-IMABIO program and the PACA region. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. Pharmacie, Blt. D, 15, avenue Charles Flahault, F-34060 Montpellier j: Permanent address: Centre de Biochimie Structurale, Facult6 de Cedex, France. 1 Investigator of the Howard Hughes Medical Institute. 11 To whomcorrespondenceshould be addressed. Tel.: 91-69-89-09; Fax: 91-69-89-13; E-mail: [email protected]. binding site for colipase (7), a 11-kDa protein that counteracts the inhibition of pancreatic lipase by bile salts. Pancreatic li- pase and lipoprotein lipase share about 30% sequence identity. Together with hepatic lipase they form the so-called mamma- lian triglyceride lipase family (8). This has led to the prediction of a common overall fold for these three enzymes and to the structural interpretation of some gene mutations (9). Despite considerable sequence homology and so probably a similar overall fold, lipoprotein lipase and pancreatic lipase display some important functional differences. Both enzymes are active on triglycerides, whereas phospholipids are sub- strates only for lipoprotein lipase (phospholipase A-1 activity) (2). The phospholipase A-1 activity of lipoprotein lipase is lower than its triglyceride activity and depends on the lipoprotein particle that harbors the substrate(10). Colipase anchors the pancreatic lipase to the lipidwater interface but does not in- fluence the catalytic rate (7). In contrast, apoC-II,l the lipopro- tein lipase cofactor, does increase the turnover number. The mechanism of this activation is poorly understood (11). There is nosequence homology between apoC-I1 and colipase. Other important differences from lipoprotein lipase are that pancre- atic lipase has not been reported to have a high affinity for heparin-like molecules and that it is active as a monomer. The presumed similar tertiary fold but very different bio- chemical properties of lipoprotein lipase and pancreatic lipase make of lipoprotein lipase a challenging target for molecular modeling studies. In this paper we describe molecular models of lipoprotein lipase, basedupon two different crystal structures: one with pancreatic lipase alone and one with a ternary pan- creatic lipase-procolipase-phospholipid complex (4, 6). In our analysis of the molecular lipoprotein lipase model we focus on the following questions. (i) Is there an extended cluster of posi- tive charges on the surface of lipoprotein lipase which could form the interaction site for heparin? (ii) Are there any differ- ences in and around the active site which could explain the phospholipase activity of lipoprotein lipase compared with pan- creatic lipase? We pay special attention here to a surface loop that covers the active site (the lid). (iii) What regions in the molecule could be involved in dimer formation and cofactor binding? MATERIALS AND METHODS The sequences were aligned according to Ref. 8, except for the C- terminal domain, in which we made slight structure-based modifica- tions. The actual sequence alignment used for the construction of our model is represented in Fig. 1. All structure manipulations used the molecular graphics program TURBO (12). The inevitable bad side chain contacts were relieved by torsion angle manipulations. Two lipoprotein lipase models were con- The abbreviation used is: apoC, apolipoprotein C. 4626
Transcript of THE JOURNAL OF BIOUXICAL Cmsrnu 269, No. 6, 11, 4626-4633

THE JOURNAL OF BIOUXICAL C m s r n u 0 1994 by The American Society for Bioebemistry and Molecular Biology, Inc.
Vol. 269, No. 6 , Issue of February 11, pp. 4626-4633, 1994 Printed in U S A .
Lipoprotein Lipase MOLECULAR MODEL BASED ON THE PANCREATIC LIPASE X-RAY STRUCTURE: CONSEQUENCES FOR HEPARIN BINDING AND CATALYSIS*
(Received for publication, September 16, 1993)
Herman van TilbeurghS, Main Roussel, Jean-Marc Lalouelln, and Christian Cambillaull From the Laboratory of Cristallisation et Macromolecules Biologiques-Centre National de la Recherche Scientifique, Facultk de Mbdecine Nord, 13916 Marseille Cedex 20, France and the $Howard Hughes Medical Institute, University of Utah Health Sciences Center, Salt Lake City, Utah 84132
Lipoprotein lipase and pancreatic lipase have about 30% sequence identity, suggesting a similar tertiary fold. Three-dimensional models of lipoprotein lipase were constructed, based upon two recently determined x-ray crystal structures of pancreatic lipase, in which the ac- tive site was in an open and closed conformation, respec- tively. These models allow us to propose a few hypoth- eses on the structural determinants of lipoprotein lipase which are responsible for heparin binding, dimer forma- tion, and phospholipase activity. The folding of the pro- tein assembles a number of positive charge clusters at the back of the molecule, opposite the active site. These clusters probably form the heparin binding site, as con- firmed by recent site-directed mutagenesis experi- ments. The active sites of lipoprotein lipase and pancre- atic lipase look very similar, except for the lid (a surface loop covering the catalytic serine in the inactive state). A different open (active) conformation of the lid in both enzymes may be responsible for their differing substrate specificities. Predictions of the nature of the lipoprotein lipase dimer remain elusive, although our model en- abled us to propose a few possibilities.
Lipoprotein lipase is responsible for the hydrolysis of plasma lipids in very low density lipoproteins and chilomicrons and causes transfer of fatty acids to tissues (1, 2). It has a high affinity for heparin and sulfate proteoglycans, explaining its physiological location on the endothelial cell wall of capillaries and on the surface of adipose tissue cells. Lipoprotein lipase is a noncovalent dimer and is activated by apolipoprotein C-11, a small protein of 79 residues (3). Several distinct regions of the enzyme must be involved in these interactions.
Pancreatic lipase is secreted in the duodenum and is active on dietary triglycerides. The pancreatic lipase structure con- sists of two distinct domains (4). The N-terminal domain (resi- dues 1-335) containing the catalytic triad is made of a central p-sheet surrounded by a-helices (the so-called dp-hydrolase fold) (5) . The active site is shielded by a surface loop that moves over 30 A upon substrate binding (6). The C-terminal domain (residues ‘336449) has a p-sandwich fold and forms the main
* This work was supported by the EEC BRIDGE-Lipase project, the Centre National de la Recherche Scientifique-IMABIO program and the PACA region. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Pharmacie, Blt. D, 15, avenue Charles Flahault, F-34060 Montpellier j: Permanent address: Centre de Biochimie Structurale, Facult6 de
Cedex, France. 1 Investigator of the Howard Hughes Medical Institute. 11 To whom correspondence should be addressed. Tel.: 91-69-89-09;
Fax: 91-69-89-13; E-mail: [email protected].
binding site for colipase (7), a 11-kDa protein that counteracts the inhibition of pancreatic lipase by bile salts. Pancreatic li- pase and lipoprotein lipase share about 30% sequence identity. Together with hepatic lipase they form the so-called mamma- lian triglyceride lipase family (8). This has led to the prediction of a common overall fold for these three enzymes and to the structural interpretation of some gene mutations (9).
Despite considerable sequence homology and so probably a similar overall fold, lipoprotein lipase and pancreatic lipase display some important functional differences. Both enzymes are active on triglycerides, whereas phospholipids are sub- strates only for lipoprotein lipase (phospholipase A-1 activity) (2). The phospholipase A-1 activity of lipoprotein lipase is lower than its triglyceride activity and depends on the lipoprotein particle that harbors the substrate (10). Colipase anchors the pancreatic lipase to the lipidwater interface but does not in- fluence the catalytic rate (7). In contrast, apoC-II,l the lipopro- tein lipase cofactor, does increase the turnover number. The mechanism of this activation is poorly understood (11). There is no sequence homology between apoC-I1 and colipase. Other important differences from lipoprotein lipase are that pancre- atic lipase has not been reported to have a high affinity for heparin-like molecules and that it is active as a monomer.
The presumed similar tertiary fold but very different bio- chemical properties of lipoprotein lipase and pancreatic lipase make of lipoprotein lipase a challenging target for molecular modeling studies. In this paper we describe molecular models of lipoprotein lipase, based upon two different crystal structures: one with pancreatic lipase alone and one with a ternary pan- creatic lipase-procolipase-phospholipid complex (4, 6). In our analysis of the molecular lipoprotein lipase model we focus on the following questions. (i) Is there an extended cluster of posi- tive charges on the surface of lipoprotein lipase which could form the interaction site for heparin? (ii) Are there any differ- ences in and around the active site which could explain the phospholipase activity of lipoprotein lipase compared with pan- creatic lipase? We pay special attention here to a surface loop that covers the active site (the lid). (iii) What regions in the molecule could be involved in dimer formation and cofactor binding?
MATERIALS AND METHODS
The sequences were aligned according to Ref. 8, except for the C- terminal domain, in which we made slight structure-based modifica- tions. The actual sequence alignment used for the construction of our model is represented in Fig. 1.
All structure manipulations used the molecular graphics program TURBO (12). The inevitable bad side chain contacts were relieved by torsion angle manipulations. Two lipoprotein lipase models were con-
The abbreviation used is: apoC, apolipoprotein C.
4626

Molecular Model of the Lipoprotein Lipase Structure 4627
K E V C Y E R L G C F S D D S P W S G I T E R P L H I L P W WHPL A D Q R R D 6LPL
S P K D V N T R F L L Y T N E N P . . . N N F Q E V A A D S 56HPL F I . - I E S K - - A - R - P - D T A E D T C H L I P G V A E 37LPL
S S I S G S N F K T N R K T R F I I H G F I D K G E . E N W 85HPL - - V A T C H . - N H S S - - - - F M V - - - - W T V T - M Y " - 64LPL
L A N V C K N L F K V E . . S V N C I C V D W K G G S R T G 113HPL V P K L V A A - Y - R - - P D - - . - V - V - - - - L S R A Q E H 93LPL
Y T Q A S Q N I R I V G A E V A Y F V E F L Q S A F G Y S P 143HPL - - P V S A G Y T K L - - - - Q D - - - R - - I N W M E E E - N - P L 123LPL
S N V H V I G H S L G A H A A G E A G R . R T N G T I G R I 172HPL D.- _ _ _ _ L ~ - y _ _ _ _ - _ _ _ _ _ _ _ _ _ _ I __ __ s . . _ _ _ _ K K V N - - 1 . 5 2 ~ ~ ~
T G L D P A E P C F Q G T P E L V R L D P S D A K F V D V I ZOZHPL -. " . . " -. . - G - . N - . E Y A E A P S _ _ _ _ S - . D _ _ _ _ D L 182LPL
H T D G A P I V P N L G F G M S Q V V G H L D F F P N G G V 232HPL " " . F T R G S - - G R S I - I Q K P - - - - " - 1 Y - - - - - - - T 211LPL
E M P G C K K N I L S Q I V D I D G I W E G T R D F A A C N 262HPL F Q - - - - - - N I G E A I R V I A E R - L . G D V D Q L V K - S 24OLPL
H L R S Y K Y Y T D S I V N P D G . F A G F P C A S Y N V F 291HPL - E - - 1 H L F I _. - - L L - - E E N P S K A Y R - S - K E A - 270LPL
T A N K C F P C P S G G C P Q M G H Y A D R Y P G K T N D V 321HPL E K G L - L S - R K N R - N N L - Y E 1 N K V R A - R . . . 297LPL
G Q K F Y L D T G D A S N F A R W R Y K V S V T L S G K K . 3H)HPL S S - M - - - K - - R S Q M P Y K V F H - Q - K I H F - - - - T E S 327LPL
V T G . H I L V S L F G N K G N S K Q Y E I F K G T L K 378HPL E T H - N Q A F E I - - - - Y - T v A E - - E N I P F T L P E V S 3 5 7 ~ ~ ~
P D S T H S N E F D S D V D V G D L Q M V K F I W . Y N N . 405HPL T N K - - Y - - F L I Y T E - - - I L - - L K - - K S D S Y 387LPL
V I N P . T . L . P R V G A S K I I V E T . N V G K Q F N F 432HPL F S W S D W W S S P G F A I Q - - - - R - - K A G E T Q - K V I - 416LPL
C S P E T V R E E . V L L T L T P C - - - - R - - K - - S H L Q K G K A P A V F V K - - H D K S L N K K S G 4 4 8 L P L
449 HPL
(HPL) and human lipoprotein lipase (LPL) (4, 8). Identical resi- FIG. 1. Alignment of the sequences of human pancreatic lipase
dues are indicated by a double hyphen sign, and dots are introduced to match the alignment where necessary.
structed. The first was based on the structure of human pancreatic lipase alone (4). This structure is essentially the same as that of the pancreatic lipase as observed in the complex with procolipase (7). In these two structures pancreatic lipase has its active site hidden from the solvent by a surface loop, and the corresponding lipoprotein lipase model will hereafter be called the closed form. A second model of lipo- protein lipase (the open form) was based on the structure of pancreatic lipase co-crystallized with colipase and phospholipidmile salt micelles (6). The resulting models, without water or ligands, were subjected to 500 cycles of energy minimization using the conjugate gradient algo- rithm implied in XPLOR (13). The structures were superimposed with the rigid body superimposition option in the molecular graphics pro- gram TURBO.
The electrostatic potential was calculated using the procedure of Getzoff and co-workers (14, 15). The charges used were those from the XPLOR force field. The calculation procedure was implemented in a series of programs (CHARGES and POTENTIAL (16)). The electrostatic field and potential are calculated on surface points (program MS) lo- cated 14 A from the Ca atoms using either a uniform dielectric constant (e = 80, for bulk water) or a distance-dependent one (4ri). Different electrostatic field surfaces were generated according to (15) and visu- alized using the program TURBO-FRODO using a procedure outlined by Tegoni et al. (16). Ten sets of electric field arrows are generated according to a sorting procedure based on increasing values of the electrostatic potential. The program allows visualization of each set of arrows independently.
RESULTS
Overall Structure of Lipoprotein Lipase Analysis of the cDNA sequences showed that lipoprotein li-
pase, hepatic lipase, and pancreatic lipase belong to the same family and suggested that all three enzymes must have the same overall fold (8). Lipoprotein lipase and pancreatic lipase are of about the same length (450 amino acids), but compared with pancreatic lipase, lipoprotein lipase has an N-terminal deletion of 25 amino acids and a C-terminal extension of 10 residues. The sequences of pancreatic lipase and lipoprotein
lipase have 31% of their residues identical in the N-terminal domain (40% similar) and 28% in the C-terminal domain (38% similar). The alp-fold of the catalytic domain belongs to the best conserved regions of the mammalian lipase family (17). Inser- tions and deletions are mainly found in loops connecting the secondary structure elements. The homology in the C-terminal domain is slightly lower than in the N-terminal domain, but careful analysis of the hydrophobicity patterns by means of hydrophobic cluster analysis (18,19) suggests that this domain in lipoprotein lipase has a P-sandwich structure similar to the one observed in pancreatic lipase (results not shown). Since we did not model the C-terminal extension of lipoprotein lipase, our lipoprotein lipase models are missing 10 residues.
For the closed lipoprotein lipase model most residues are situated in the allowed regions of the Ramchandran plot. Al- most the same result was obtained for lipoprotein lipase in the open conformation. We superimposed the lipoprotein lipase models with the corresponding structures of pancreatic lipase. As can be seen in Fig. 2, pancreatic lipase and the lipoprotein lipase model have the same overall fold. The root mean square deviation between lipoprotein lipase and pancreatic lipase of the Ca! atom positions of residues belonging to the alp-fold of the catalytic domain is 0.72 A for the closed form and 0.86 A for the open form.
Electrostatic Potential Calculation The electrostatic properties of lipoprotein and pancreatic li-
pase were investigated qualitatively using the method outlined above. The electrostatic potential surface of lipoprotein lipase, calculated in uacuo, is represented in Fig. 3. The positive elec- trostatic potential mapped onto the surface of lipoprotein lipase has a pronounced maximum, situated in front of a loop at the junction between the two domains and opposite the active site. This region is at the center of a series of LydArg clusters which are indicated in Fig. 3. Cluster 1 is Arg294-Lys296-Arg297-Lys300; cluster 2 is Arg263-Arg279-Lys280-Arg282; cluster 3 is Lys147- L y ~ ' ~ ~ - a n d and finally the C-terminal domain cluster 4 is Lys319-Arg405-Lys407-Lys414. The positive electrostatic poten- tial maximum is not only caused by the relative abundance of positive charges at the backside of lipoprotein lipase but also by the scarcity of compensating negative charges in this region of the molecule. Pancreatic lipase, which has a more symmetric charge distribution, has a less pronounced electrostatic poten- tial surface (results not shown). Its positive maximum is situ- ated in a region coinciding with the colipase binding region.
The Active Site Region The recently determined structure of pancreatic lipase-pro-
colipase complex in the presence of lipidhile salt micelles showed considerable rearrangements of two surface loops in the active site of pancreatic lipase (6). These conformational changes, which move some residues over a distance of 29 A, render the catalytic serine accessible to substrate. These mo- bile surface loops (residues 76-84 (p5-loop) and residues 237- 261 (the lid) in pancreatic lipase) are also present in lipoprotein lipase (residues 54-63 and residues 216-239). Any discussion on the mechanism and active site of lipoprotein lipase will have to take into consideration the flexibility of this region. We used the open pancreatic lipase structure to model an open form of the active site of lipoprotein lipase and compared the different constituents of the active site. The energy-minimized open and closed lipoprotein lipase structures are virtually the same when the mobile surface loops are not taken into account.
The Catalytic Diad-The regions surrounding the catalytic triad residues are among the best conserved between the mem- bers of the lipase family (8). It is thus not surprising to observe that the arrangements of the catalytic triads of lipoprotein
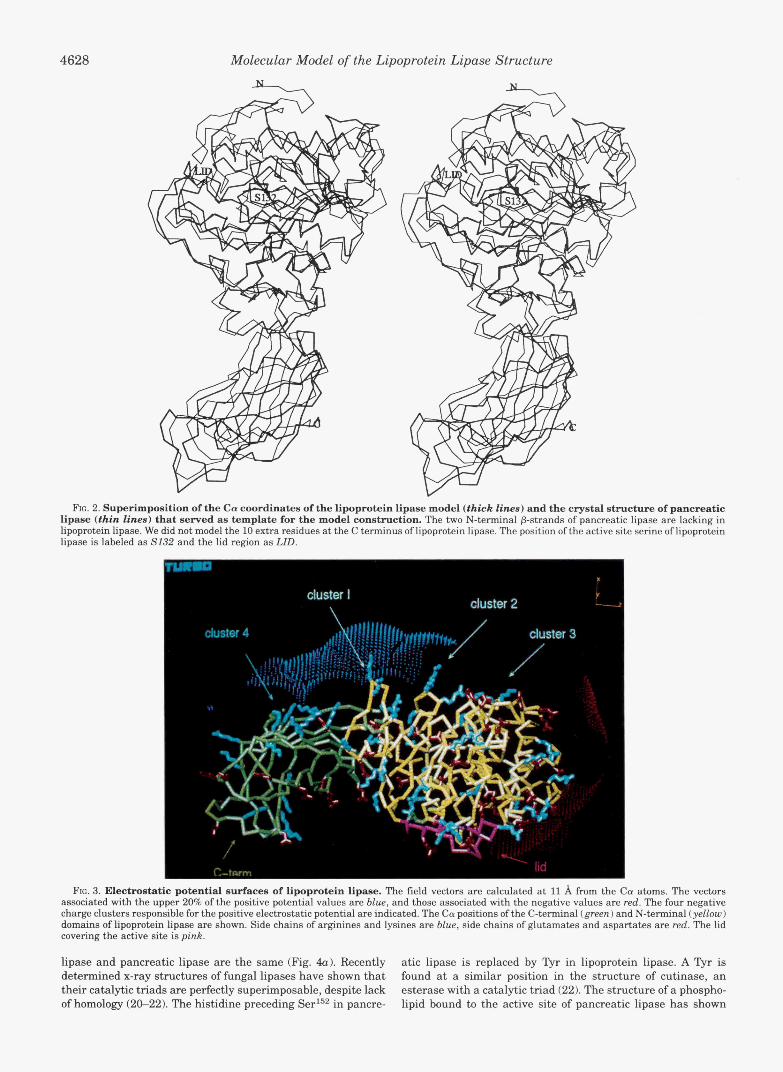
4628 Molecular Model of the Lipoprotein Lipase Structure
RG. 2. Superimposition of the Ca coordinates of the lipoprotein lipase model (thick lines) and the crystal structure of pancreatic lipase (thin lines) that served as template for the model construction. The two N-terminal p-strands of pancreatic lipase are lacking in lipoprotein lipase. We did not model the 10 extra residues at the C terminus of lipoprotein lipase. The position of the active site serine of lipoprotein lipase is labeled as S132 and the lid region as LID.
FIG. 3. Electrostatic potential surfaces of lipoprotein lipase. The field vectors are calculated at 11 A from the Ca atoms. The vectors associated with the upper 20% of the positive potential values are blue, and those associated with the negative values are red. The four negative charge clusters responsible for the positive electrostatic potential are indicated. The Ca positions of the C-terminal (green) and N-terminal (yellow) domains of lipoprotein lipase are shown. Side chains of arginines and lysines are blue, side chains of glutamates and aspartates are red. The lid covering the active site is pink.
lipase and pancreatic lipase are the same (Fig. 4u). Recently atic lipase is replaced by "yr in lipoprotein lipase. A Tyr is determined x-ray structures of fungal lipases have shown that found at a similar position in the structure of cutinase, an their catalytic triads are perfectly superimposable, despite lack esterase with a catalytic triad (22). The structure of a phospho- of homology (20-22). The histidine preceding Ser152 in pancre- lipid bound to the active site of pancreatic lipase has shown

Molecular Model of the Lipoprotein Lipase Structure 4629
a
FIG. 4. Panel a, superimposition of the catalytic triads of the lipoprotein lipase model (thick lines; 0156, H241, S132) and that of pancreatic lipase (thin lines). Aspzo5 of pancreatic lipase is also represented (D205#). Panel b, superimposition of active site loops and lid of the lipoprotein lipase model (thick lines) and those of pancreatic lipase (thin lines). The marked residues belong to lipoprotein lipase. We also superimposed a model of a phosphatidylcholine molecule (thickest, labeled P ) as observed in the crystal structure of the ternary pancreatic lipase-procolipase-phospholipid complex (6). This model could be fitted into the active site of lipoprotein lipase with only minor changes of torsion angles of protein side chains and of the lipid acyl chains.
that the residue at position 151 could interact with the polar part of a bound lipid substrate (6). The H i m substitution in lipoprotein lipase could therefore influence the catalytic activ- ity. Asp156, the third triad residue of lipoprotein lipase, may be stabilized by main chain nitrogens of T h P 4 and PhelE5.
The Lid-The surface loops, covering the active site in pan- creatic lipase (residues 237-261) and lipoprotein lipase (resi- dues 216-239) are contained between 2 conserved cysteines but do not display sequence homology between the two enzymes. The lid in lipoprotein lipase (and hepatic lipase) has one dele- tion compared with pancreatic lipase. We have modeled the open lipoprotein lipase lid using the structure of the open lid of pancreatic lipase (Fig. 4b) . The open conformation of the lid in pancreatic lipase is stabilized by hydrogen bonds between the lid and the core of the catalytic domain and between the lid and colipase (6). The absence of these interactions in lipoprotein lipase may induce a different conformation and orientation of the lid.
The p5-Loop-We showed that upon opening of the lid of pancreatic lipase, the p5-100~ (residues 76-85) folds back upon the core of the protein. This rearrangement increases even further the access to the active site and brings one of the elec- trophiles of the oxyanion hole (the main chain nitrogen of P h e T into a catalytically competent position. The sequence of this loop (residues 54-64) is not well conserved in lipoprotein lipase and contains one insertion (Fig. 1). However, the nature of the substitutions is more conservative than for the lid. Therefore this loop may adopt a similar structure in the open
conformation for pancreatic lipase and lipoprotein lipase. The oxyanion hole in our lipoprotein lipase model is formed by the main chain nitrogens of Leu133 and Trp55, which correspond to Leu153 and Phe77 in pancreatic lipase. Phe77 is hanging over the glycerol moiety of a bound phospholipid at the active site of pancreatic lipase (6). The substitution by “q in lipoprotein lipase may influence catalytic activity. In the open form of pancreatic lipase, Phe77 is part of a sharp turn and is followed by Ile78. Rhizomucor lipase and cutinase, which have a surface loop very similar to the open p5-loop in pancreatic lipase, have Thr at that position, exactly like lipoprotein lipase (22,23). The movement of the p5-loop upon binding to the interface brings GlyE1 very close to the body of the catalytic domain. The fact that this glycine is conserved in lipoprotein lipase means that the p5-100~ may adopt the same conformation as in pancreatic lipase. Any other residue at this position would prevent this for steric reasons.
The Lipid Binding Site--To explore possible important de- terminants of lipoprotein lipase activity and specificity, apart from the catalytic triad residues, we superimposed the phos- phatidylcholine molecule, as observed in the tertiary pancre- atic lipase-procolipase-phospholipid complex structure (6), onto the model of the lipoprotein lipase active site (Fig. 4b). The sn-1 aliphatic chain of the phospholipid in the complex with pancre- atic lipase is lying in the hydrophobic groove formed by side chains of surface loops, whereas the sn-2 is interacting with residues from the open lid. The phosphatidyl moiety is pointing toward the solvent and was hardly visible into the electron

4630 Molecular Model of the Lipoprotein Lipase Structure
density map of the complex. Although the detailed sequences of the surface loops differ between lipoprotein lipase and pancre- atic lipase, the hydrophobic nature of the active site groove is well conserved. By analogy with pancreatic lipase, we propose that the lipoprotein lipase binding site for the sn-1 (or sn-3) alkyl chain of a triglyceride substrate is formed by the active site loops contained between Glngl and Prog5, between Pro157 and Pro160, and between ArgIa7 and IleIg6. The following resi- dues may display hydrophobic interactions with the substrate Tyrg4-Pro157-Ala158-Ile193. As is the case for pancreatic lipase, the active site of lipoprotein lipase is not particularly adapted for the harboring of the phosphoryl moiety of a phospholipid. Except for the catalytic triad, most residues in the active site are hydrophobic, and their main chain atoms are not accessible for hydrogen bonding.
The C-terminal Domain The sequence similarity between lipoprotein lipase and pan-
creatic lipase of the noncatalytic C-terminal domain is some- what lower than for the N-terminal domain. Hydrophobic clus- ter analysis using the method of Gaboriaud et al. (181, however, indicates that this domain must also have a similar overall structure in the two enzymes (results not shown). Modeling of this domain was trickier because of a few insertions in the lipoprotein lipase sequence. Most of these insertions are situ- ated in the loops that connect the p-strands. Although the structure of the C-terminal domain of lipoprotein lipase and pancreatic lipase may be more divergent than for the N-termi- nal domain, we are confident that the overall p-sandwich fold is conserved in lipoprotein lipase. Our lipoprotein lipase model is in that sense confirming that the interior of the sandwich is composed of hydrophobic residues and that there are no buried charges. It was demonstrated recently that this domain can be cleaved in bovine lipoprotein lipase by mild chymotrypsin treatment (24). The cleavage site (Phe388, human lipoprotein lipase numbering) is situated in our model in a very exposed hydrophobic loop, explaining its vulnerability to chymotrypsin treatment.
DISCUSSION
The three-dimensional model of lipoprotein lipase, based on the x-ray structures of pancreatic lipase, is a good starting point to discuss structure-function relationships for this impor- tant enzyme. A series of site-directed mutagenesis studies on lipoprotein lipase have appeared recently in the literature, and our model allows us to give a more rational explanation for these observations (25-32). Now we will use our model to dis- cuss a few functional properties of lipoprotein lipase.
Heparin Binding-Glycosaminoglycan interaction with pro- teins regulates a number of metabolic processes (for review, see Ref. 33). The binding of lipoprotein lipase onto the extracellular matrix is relatively well documented physiologically (34). This interaction keeps lipoprotein lipase fixed onto the luminal side of the endothelial cell wall, localizing the hydrolysis of plasma lipids. Recent reports indicate that lipoprotein lipase could also be involved in the receptor-mediated cellular uptake of lipopro- teins, a process that is linked to the heparan sulfate binding properties of lipoprotein lipase (35-37). Heparin-like polysac- charides are heavily negatively charged molecules expected to interact with positively charged regions on the lipoprotein li- pase surface. Lipoprotein lipase is eluted from a heparin- Sepharose column using high NaCl concentrations, indicating the electrostatic nature of the interaction (38). To examine the structural determinants for heparin binding, starting from a molecular model, we calculated the electrostatic potential sur- face of lipoprotein lipase. Fig. 3 shows the mapping of these potentials onto the surface of the molecular lipoprotein lipase
model. The positive electrostatic potential maximum of lipopro- tein lipase is concentrated around a few positive charge clus- ters at the back of the molecule. Pancreatic lipase, which was not reported to interact significantly with heparin, has a more symmetrical charge distribution than lipoprotein lipase.
The majority of lysine and arginine residues at the back of lipoprotein lipase are grouped into four clusters (clusters 1-4 in Fig. 3). The maximum positive electrostatic potential in lipo- protein lipase is situated in front of positive charge clusters containing Arg263-Arg279-Lys280-Arg282 (cluster I), Lys296-Arg297-Lys300 (cluster 2). This region contains two hy- pothetical heparin binding consensus sequences (B-B-X-B and B-X-B-B-X-X-B, where B is a positively charged residue and X a small neutral one), present in the heparin binding peptides of a number of proteins such as fibroblast growth factors, platelet factor 4, and apolipoprotein E and B (39, 40). Both consensus sequences are part of an irregular surface loop at the junction of the N- and C-terminal domains. The presence of two disul- fide bridges (Cys27s-Cys2s3 and Cys264-Cys275) may confer ri- gidity to this loop and probably prevents a drastic rearrange- ment of this region upon heparin binding, as was observed for apolipoprotein E (41). Interestingly, the folding of the protein brings the two consensus sequences close together on the sur- face of our lipoprotein Ppase model (Ca distance between resi- dues 281 and 296: 10 A). The amino acids belonging to cluster 1 (residues 279-282) in avian lipoprotein lipase (human lipo- protein lipase sequence numbering) were subjected to exten- sive site-directed mutagenesis experiments (25,421. A decrease of the total positive charge in this region caused a decrease in affinity for heparin. The triple mutant Arg279 + Gln/LysZso +
Gl1dArg2~~ + Gln saw its heparin affinity diminished but not abolished and interacted poorly with heparan sulfate proteo- glycans. Our model structure shows that this hypothetical hep- arin binding site has a supplementary positive charge cluster (2) in the neighborhood (residues 294-300), which may explain why the triple mutant retains considerable heparin affinity. These mutations should not seriously perturb the tertiary structure because only well exposed hydrophilic residues are involved. A third consensus sequence for heparin binding is found between residues 148 and 152 (cluster 3). This region is contained in a surface loop, situated close to the previous charge clusters, and could therefore also contribute to heparin binding. Binding studies with size-fractionated heparin have shown that lipoprotein lipase is released by saccharides that contained at least 8 units, indicating that the lipoprotein lipase heparin binding site may be extended (43). An octosaccharide would be more than sufficiently long to span the distance be- tween the various consensus peptides, the aver?ge length of a tetrasaccharide in the heparin chain being 16.7 A (44). Hata et al. (25) have mutagenized the charges contained in cluster 1,2, and 3 of human lipoprotein lipase. These experiments demon- strated that the charges in clusters 1 and 2 are important for heparin binding, which confirms the results obtained with avian lipoprotein lipase for cluster 1. Mutations in cluster 3, however, did not affect heparin binding. Combined mutations in cluster 1 and 2 did not completely abolish heparin binding, SO the authors suspected the presence of further determinants for this interaction.
A fourth positive charge cluster (cluster 4, Fig. 3) is situated in the C-terminal domain and accommodates Lys319-Ly~403- Ar$05-Lys407-Lys414-Lys415. This cluster is different from the previous ones in the sense that it is not originating from a contiguous peptide stretch. The charged residues of cluster 4 come from three neighboring p-strands and are at the hydro- philic exposed side of the P-sheet pointing roughly in the same direction as residues of the three other clusters. Cluster 4 is also relatively close to the ArgLys cluster 1 of the N-terminal

Molecular Model of the Lipoprotein Lipase Structure 4631
a A
FIG. 5. Panel a, some schematic dimerization models of lipoprotein lipase. The catalytic domain is represented as a shaded sphere, the C-terminal domain as a white lobe, the lid covering the active site as an L. Heparin molecules are shown as rectang&ar bores. Models A and B represent the back and front views, respectively, of the head to head dimer and show how this dimer could form a single heparin binding site. Model C shows a head to tail model in which the active sites are face to face. This model is more difficult to reconcile with structural data. Model D, head to tail dimer, in with the individual monomers are rotated along their longitudinal axis compared with model C. This alternative model for the dimer would leave enough freedom for the lid to open upon interfacial activation. This type of association is observed in a crystal contact of the ternary pancreatic lipase-procolipase-phospholipid complex. Panel b, stereo view of the hypothetical dimer that is schematically represented in panel a (model D). A heparin dodecamer model is represented as thick lines. A model for the dimer was constructed by taking the crystallographic dimer from the crystal structure of the open form of the pancreatic lipase and superimposing the lipoprotein lipase coordinates onto it. Since this result is only illustrative no attempt was made to optimize the interactions between the monomers or between the dimer and heparin.
domain (Co distance between residues 414 and 280: 16 A). Chymotrypsin cleavage of bovine lipoprotein lipase yields a truncated protein (residues 1-392) that has kept its heparin affinity intact (24). Cluster 4 therefore does not seem to be important for heparin binding. However, Davis et al. (31) have studied the properties of a chimaeric construct consisting of the hepatic lipase C-terminal domain and the lipoprotein lipase N-terminal domain. This chimaeric lipase had the same affin- ity for heparin as the intact hepatic lipase, which is consider- ably lower than for the intact lipoprotein lipase. The authors therefore assigned heparin binding to the C-terminal domain. None of the sequences of the positive charge clusters of lipo- protein lipase is absolutely conserved in hepatic lipase. We propose that the ensemble of positive clusters may be involved in the in vivo interaction of lipoprotein lipase with heparan proteoglycans. Which cluster dominates in binding may depend
upon the detailed distribution of charges and the nature of the bound polysaccharide ligand. A last cluster of positive charges is contained in the C-terminal extension of lipoprotein lipase. Because this region has no equivalent in pancreatic lipase, we did not model its structure. The presence of 3 lysines out of 10 residues and its localization at the back of the molecule make it also a candidate for heparin interaction.
Dimer Formation-It has been shown that the active form of lipoprotein lipase is a dimer (45). Inactive monomers elute from a heparin-Sepharose column at much lower NaCl concentra- tions than the native dimer (25). High affinity binding of lipo- protein lipase to heparin seems to necessitate the native dimer conformation. This suggests that the various positively charged clusters of the individual monomers form a single dimeric hep- arin binding site. We have depicted some hypothetical lipopro- tein lipase dimers schematically in Fig. 5a. In model A the

4632 Molecular Model of the Lipoprotein Lipase Structure
dimer contacts are exclusively situated in the catalytic domain, and the heparin binding site is formed by the sum of the posi- tive charge clusters of the individual monomers. This model would be in accordance with recent biochemical results claim- ing that chymotryptic cleavage of the C-terminal domain does not influence dimer formation (24). Proteolytic cleavage at resi- due 392 would leave one sheet of the p-sandwich domain un- covered to the solvent. Therefore presumably the partial pro- teolysis will heavily perturb the structure of the remaining part of this domain, in agreement with the conclusion that the C- terminal domain is not involved in dimer formation. Head to tail lipoprotein lipase dimers, which have been proposed (32) in analogy with native pancreatic lipase noncrystallographic dimers (4), also seem unlikely. This type of dimer, represented in Fig. 5a, model C , would block the opening of the lid and is more difficult to conciliate with heparin binding at the back of the active site (arrows in Fig. 5a, model C ) . If the active form of lipoprotein lipase is a head to tail dimer, it would need to give enough space to the lid for a conformational change upon sub- strate binding. Head to tail association was observed in a crys- tallographic dimer of the open pancreatic lipase-colipase-phos- pholipid complex (Fig. 5a, model D ) . In this type of dimer, monomers are associating side to side and are leaving the lid free to move. An analogous hypothetical model for lipoprotein lipase, bound to heparin is shown in Fig. 5b.
Active Site and Lid-Lipoprotein lipase is capable of hydro- lyzing the primary acyl bond of phospholipids in triglyceride- rich lipoproteins (10, 46). Pancreatic lipase on the other hand has a negligible activity toward these substrates (less than 1%, compared with triglycerides) (471, whereas hepatic lipase is the most potent phospholipase of the mammalian lipase family (48). Modeling of the active site of lipoprotein lipase using the structure of pancreatic lipase with a phospholipid bound into the active site (6) is meaningful because of the high sequence similarity of the two enzymes in this region. The active site of pancreatic lipase can be schematically divided into a catalytic triad, an oxyanion hole, a very hydrophobic groove that is prob- ably binding the aliphatic chain of the acyl-enzyme intermedi- ate,2 and the lid. The catalytic triad residues of lipoprotein lipase are situated in highly conserved peptides and occupy very similar spatial positions in the lipoprotein lipase model and the pancreatic lipase structure. The identity and impor- tance of the catalytic triad residues of lipoprotein lipase and pancreatic lipase were recently confirmed by site-directed mu- tagenesis experiments (28, 49). Also, the environment of the catalytic triad in lipoprotein lipase and pancreatic lipase seems almost identical, and the different catalytic properties can probably not be explained at this level. The oxyanion hole in pancreatic lipase is formed by the main chain nitrogen atoms of Leu153 and Phe77, and we predict a similar role for the main chain nitrogens of Leu132 and Trp55 in lipoprotein lipase.
The reorganization of the lid in pancreatic lipase gives access to a very hydrophobic groove that binds one of the alkyl chains of a lipid substrate. This groove is made by the hydrophobic side chains of a few active site loops (Fig. 4b). The first loop in pancreatic lipase (residues 91-95 in lipoprotein lipase) con- tains a tyrosine (Tyrg4) that makes important hydrophobic con- tacts with a bound lipid. This tyrosine is conserved in lipopro- tein lipase and hepatic lipase and so may have the same function in these enzymes. The second loop (residues 157-160 in lipoprotein lipase), contained between 2 conserved prolines, follows Asp156 of the catalytic triad. Pro160 and Ala158 make up part of the hydrophobic binding groove. A missense mutation Pro157 + Arg in lipoprotein lipase (lipoprotein lipase-Nijme- gen) leads to a totally inactive enzyme (48). Arginine at this
M. P. Egloff et al., unpublished results.
position must indeed seriously perturb the conformation of this loop and even the position of the catalytic Asp156. A third loop (residues 187-196 in lipoprotein lipase) has a twisted confor- mation. Residues at the base of this loop interact with the lipid in pancreatic lipase (especially Phe215, whose side chain rotates more than 100" upon opening of the lid, is replaced by Ilelg4 in lipoprotein lipase). The preceding Gly is conserved between the two enzymes; any other residue would seriously interfere with the catalytic His-Ser interaction. The rest of the loop protrudes from the active site and may be interacting with the lipidwater interface. It is therefore important to note that the sequence of lipoprotein lipase in that region is much more hydrophilic than for pancreatic lipase, a factor that may influence interfacial behavior.
The sequence and architecture of the active site loops of pancreatic lipase and our lipoprotein lipase model are rather similar, in contrast to the very different sequences of their respective lids (4, 6). In the closed form of pancreatic lipase, which is probably the structure in solution, the lid has a short a-helical region that blocks the active site and is connected to the core of the protein by two elongated strands (4). The open form of the pancreatic lipase lid, determined with a lipid at the active site, reveals the presence of two amphipathic helices connected by a p-turn. One of the helices is preceded by a short stretch of strand structure. The open lid forms an elbow that sticks out from the active site and therefore is certainly in- volved in binding to the lipidwater interface. One side of the main helix also interacts with the lipid bound at the active site, suggesting a double role for the lid: interface interaction and substrate binding. Secondary structure prediction proposes for the lipoprotein lipase lid two highly amphiphilic helices (resi- dues 217-228 and 232-238) connected by a p-turn (26). Since our model of the open lipoprotein lipase lid is based on the open pancreatic lipase lid (Fig. 5b) , it is not in accordance with secondary structure predictions as regards the first 4 residues of the lid (in elongated conformation). The helical wheel pro- jections of the two crystallographically observed helices (i.e. without the first 4 residues) in the open pancreatic lipase lid look very similar to those of the presumed helices in the lipo- protein lipase lid (result not shown). There is a reasonable chance that the entire lipoprotein lipase lid has a helix-turn- helix motif. This would cause a considerable deviation of the orientation of the lid between pancreatic and lipoprotein lipase relative to the entrance of the active site. Another problem with our pancreatic lipase-based model of the lipoprotein lipase lid concerns Lys238. This residue corresponds to Ala260 in pancre- atic lipase. Ala260 has p-conformational angles and causes a 90" kink between an a-helix of the core and the second amphipathic helix of the open lid. This conformation of Lys238 in lipoprotein lipase projects the side chain into the active site and causes some steric hindrance around the catalytic triad. A different conformation of this residue would change the angle of the helical kink between lipoprotein lipase and pancreatic lipase and relieve some of the steric stress caused by its side chain. To summarize, we think that the open conformation of the lipo- protein lipase lid could consist entirely of a helix-turn-helix motif with a different orientation compared with the helix- turn-helix motif as observed in the open pancreatic lipase lid. Recent mutagenesis experiments have shown the importance of the lid for lipoprotein lipase activity (30). A lipoprotein lipase mutant in which the lid was exchanged for the pancreatic li- pase lid yielded a totally inactive enzyme toward insoluble substrates (16, 30). Site-directed permutation mutants that changed the amphiphilicity of the two presumed helices were still active on monodisperse tributyrin but lost their capability of hydrolyzing insoluble triolein (26). This demonstrates that the conformation and the hydrophobic profile of the lid play a

Molecular Model of the Lipoprotein Lipase Structure 4633 crucial role for activity against insoluble substrates. The open lid in pancreatic lipase is stabilized by many hydrogen bonds with the core of the enzyme and with colipase. These interac- tions would not be present in the case of a hybrid lipoprotein lipase molecule onto which one has grafted a pancreatic lipase lid. Hepatic lipase, whose natural substrates resemble those of lipoprotein lipase, has a lid sequence that is homologous to that of lipoprotein lipase. Lid exchange between those two enzymes was less deleterious for activity than was the case for the lid exchange between lipoprotein lipase and pancreatic lipase (26).
These facts suggest that the specificity of mammalian lipases toward insoluble substrates is determined by the lid. This hy- pothesis is further supported by the recent determination of the guinea pig pancreatic lipase sequence (50), which has 70% strict identity with human pancreatic lipase. This lipase is almost identical to the human pancreatic lipase in the region around the active site, except for the lid which is reduced to only 5 amino acids. Guinea pig lipase has high phospholipase A-1 activity and is not interfacially activated, which make it distinct from all other known pancreatic lipases. It is tempting to explain its different behavior by the absence of an extended lid structure.
C-terminal Domain-Recent crystallographic studies have shown that the C-terminal domain of pancreatic lipase is bind- ing its cofactor colipase (7). Lipoprotein lipase is also activated by a proteinic cofactor, apoC-11. ApoC-I1 shares no sequence homology to colipase. Our model study proposes that the C- terminal domains of lipoprotein lipase and pancreatic lipase, despite a number of insertions and deletions, have the same overall fold. Since this domain seems structurally conserved in the mammalian lipase family, it would be logical to assign apoC-I1 binding to this domain. However, several lines of bio- chemical evidence go against the presence of the apoC-I1 bind- ing site of lipoprotein lipase on this domain. Hepatic lipase, which also has this C-terminal domain, has no cofactor. Chi- maeric constructs consisting of the catalytic domain of hepatic lipase and the C-terminal domain of lipoprotein lipase are not activated by apoC-I1 (31). Chymotrypsin cleavage of the C- terminal domain does not hamper apoC-I1 activation (24). Little is known, however, about lipoprotein lipase-apoC-I1 in- teraction, let alone the structural explanation of the activation phenomenon. X-ray crystallography on the lipase-colipase sys- tem has shown that the activation of pancreatic lipase by coli- pase involves interactions of the cofactor with both the C-ter- minal and N-terminal domains (6). An unexpected outcome of this work was that the interactions between colipase and the N-terminal domain are only realized upon binding of the com- plex to the lipidlwater interface. Whether a similar complicated phenomenon is happening in the lipoprotein lipase activation by apoC-I1 remains to be seen.
Acknowledgments-We thank Dr. R. Verger for critical reading of the manuscript and for valuable discussions and Drs. A. Imberty and S. Cros for providing a heparin model.
REFERENCES 1. Garfinkel, A. S., and Schotz, M. (1987) in Plasma Lipoproteins (Gotto, A. M.,
2. Olivecrona, T., and Bengtsson-Olivecrona, G. (1987) in Lipoprotein Lipase
3. Jackson, R. L., Baker, H. N., Gilliam, E. B., and Gotto, A. M., Jr. (1977) Proc.
4. Winkler, F. K., DArcy, A,, and Hunziker, W. (1990) Nature 343, 771-774 5. Ollis, D. L., Cheah, E., Cygler, M., Dijkstra, B., Fmlow, F., Franken, S . M.,
Harel, M., Remington, S. J., Silman, I., Schrag, J., Sussman, J., Versch- ueren, K., and Goldman, A. (1992) Protein Eng. 5, 197-211
6. van Tilbeurgh, H., Egloff, M.-P., Martinez, C., Rugani, N., Verger, R., and Cambillau, C. (1993) Nature 362,814420
Jr., ed) Elsevier Scientific Publishing Co., New York
(Borenszjatin, J., ed) Evener Publishers, Chicago
Natl. Acad. Sci. U. S. A. 74, 1942-1945
7.
8.
10. 9.
11. 12.
13.
14.
15.
16.
17.
18.
19. 20.
21. 22.
23.
24.
25.
26.
27.
28.
29.
30. 31.
32.
33. 34.
35.
36.
37.
38.
39. 40.
41.
42. 43.
44.
45.
46.
47.
48.
49. 50.
van Tilbeurgh, H., Sarda, L., Verger, R., and Cambillau, C. (1992) Nature 369, 159-162
Kirchgessner, T. D., Chuat, J.-C., Heinzmann, C., Etienne, J., Guilhot, S. , Svenson, K., Ameis, D., Pilon, C., DAuriol, L., Andalibi, A.. Schotz, M., Galibert, F., and Lusis, A. J. (1989) P m . Natl. Acad. Sci. U. S. A 86, 9647-9651
Derewenda, Z. S., and Cambillau, C. (1991) J. Biol. Chem. 266,23112-23119 Deckelbaum, R. J., Ramakrishnan, R., Eisenberg, S. , Olivemna, T., and
Clarke, A. R., and Holbmok, J. J. (1985) Biochim. Biophys. Acta 827,35W68 Roussel, A,, and Cambillau, C. (1991) Silicon Graphics Directory, Silicon
Briinger, A. T. (199O)XF'LOR Version 2.1 Manual, Yale University Press, New
Getzoff, E. D., Tainer, J. A,, Weiner, P. K., Kollman, P. A,, Richardson, J. S. , and
Roberts, V. A,, Freeman, H. C., Olson, A. J., Tainer, J. A,, and Getzoff, E. D.
Tegoni, M., White, S. A,, Roussel, A., Mathews, F. S., and Camhillau, C. (1993)
Persson, B., Jomvall, H., Olivecrona, T., and Bengtsson-Olivecmna, G. (1991)
Gaboriaud, C., Bissery, V., Benchetrit T., and Mornon, J. P. (1987) FEBS Lett.
Woodcock, S., Mornon, J. P., and Henrissat, B. (1992) Protein Eng. 5,629-635 Brady, L., Bnozowski, A. M., Derewenda, Z. S. , Dodson, E., Dodson, G., Tolley,
S., Turkenburg, J. P., Christiansen, L., Huge-Jensen, B., Norskov, L., Thim, L., and Menge, U. (1990) Nature 343, 767-770
Schrag, J., Winkler, F., and Cygler, M. (1992) J. Biol. Chem. 267,4300-4303 Martinez, C., De Geus, P., Lauwereys, M., Matthysens, G., and Cambillau, C.
Brzozowski, A. M., Derewenda, U., Derewenda, Z. S., Dodson, G. G., Lawson, (1992) Nature 366,615418
D. M., Turkenburg, J. P., Bjorkling, F., Hugedensen, B., Patkar, S . A,, and Thim, L. (1991) Nature 351,491494
h k e n e , A,, and Bengtsson-Olivecrona, G. (1993) Eur J. Biochem. 213,185- 194
Hata, A,, Ridiger, D. N., Sutherland, S., Emi, M., Shuhua, Z., Myers, R. L., Ren, K., Cheng, T., Inoue, I., Wilson, D. E., Iverius, P.-H., and Lalouel, J:M. (1993) J. Biol. Chem. 268,8447-8457
Dugi, K. A., Dichek, H. L., Talley, G. D., Brewer, H. B., and Santamarina-Fojo,
Semenkovich, C. F., Luo, C.-C., Nakanishi, M. K., Chen, S.-H., Smith, L. C., S. (1992) J. Biol. Chem. 267,2508&25091
Faustinella, F., Smith, L. C., Semenkovich, C. F., and Chan, L. (1991) J. Biol. and Chan, L. (1990) J. Biol. Chem. 265,5429-5433
Emmerich, J., Beg, 0. U., Peterson, J., Prevatio, L., Brunzell, J. D., Brewer, H. Chem. 266,9481-9485
Faustinella, F., Smith, L. C., and Chan, L. (1992) Biochemistry 31,7219-7223 B., and Santamarina-Fojo, S . (1992) J. Biol. Chem. 267,41614165
Davis, R. C., Wong, H., Nikazy, J., Wang, K., Han, Q., and Schotz, M. C. (1992)
Wong, H., Davis, R. C., Nikazy, J., Seehart, K. E., and Schotz, M. C. (1991)
Jackson, R. L., Buscb, S . J., and Cardin A. D. (1991)Physiol. Reu. 71,481-539 Shimada, K., Gill, P. J., Silbert, J. E., Douglas, W. H. J., and Fanburg B. L.
Beisiegel, U., Weber, W., and Bengtsson-Olivemna, G. (1991) Proc. Natl. (1981) J. Clin Invest. 68,995-1002
Mulder, M., Lombardi, P., Jansen, H., van Berkel, T. J. C., Franta, R. R., and Acad. Sei. U. S. A. 88,8342-8346
Eisenberg, S., Sehayek, E., Olivecrona, T., and Vlodavsky, I. (1992) J. Clin. Havekes, L. M. (1992) Biochem. Biophys. Res. Commun. 185, 582687
Iverius, P.-H., and Ostlund-Lindqvist, A,". (1976) J. Biol. Chem. 251,7791- Inuest. SO, 2013-2021
7795 Cardin, A. D., and Weintraub, H. J. R. (1989) Arteriosclerosis 9, 21-32 Stuckey, J. A,, St. Charles, R., and Edwards, B. F. P. (1992) Proteins 14,
277-287 Cardin, A. D., and Jackson, R. L. (1988) in Eicosanoids, Apolipoproteins, Li-
poproteins, Lipoprotein Particles, and Atherosclerosis (Malmendier C. L., Alaupovic, P., eds) pp. 157-163, Plenum Publishing Co., New York
Bengtsson-Olivecrona, G. (1992) Biochemistry 31,8544-8551
Graphics, Mountain View, CA
Haven, CT
Richardson, D. C. (1983) Nature 306, 287-290
(1991) J. Biol. Chem. 266, 13341-13444
Proteins 16, 408-422
FEBS Lett. 288,33-36
224, 149-155
J. Biol. Chem. 267,21499-21504
Proc. Natl. Acad. Sci. U. S. A. 88, 11290-11294
Berryman, D. E., and Bensadoun, A. (1993) J. Biol. Chem. 268,327223276 Liu, G., Hultin, M., Ostergaard, P., and Olivecmna, T. (1992) Biochem. J. 285,
731-736 Atkins, E. D. T., and Nieduszynski, I. A. (1976) in Heparin Chemistry and
York Clinical Usage (Kakkar, V. V., and Thomas, D. P., eds) Academic Press, New
Osborne, J. C., Jr., Bentgsson-Olivecrona, G., Lee, N. S. , and Olivecrona, T.
Shirai, K., Fitzhams, J. T., Shinomiya, M., Muntz, G. H., Harmony, J. A. K., (1985) Biochemistry 24,5606-5611
Verger, R. (1984) in Lipases (Borgstriim, B., and Bmkman, H. L., eds) Elsevier Jackson, R. L., and Quinn, D. M. (1983) J. Lipid Res. 24, 721-730
Bruin, T., Kastelein, J. J. P., van Dierman, D. E., Ma, Y., Henderson, E. H., Science Publishing Co., Amsterdam
Stuyt, P. M. J., Stalenhoef, A. F. H., Sturk, A,, Brunzell, J. D., and Hayden, M. R. (1992) Eur J. Biochem. 208, 267-272
Lowe, M. E. (1992) J. Biol. Chem. 267, 17069-17073 Hjorth, A., CarriBre, F., Cudrey, C., Woldike, H., Boel, E., Lawson, D. M.,
Biochemistry 32, 47024707 Ferrato, F., Cambillau, C., Dodson, G. G. , Thim, L., and Verger, R. (1993)


















