
THE JOURNAL of 269, No. 33, Issue OF The and in U.S.A ... · Mapping the ABPl Auxin-binding Site...
5
THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc. Vol . 269, No. 33, Issue of August pp. 2113621140, 1994 Printed in U.S.A. Mapping the Auxin-binding Site of Auxin-binding Protein l* (Received for publication, May 6, 1994, and in revised form, June 13, 1994) Justin C. Brown and Alan M. Jones$ From the Department of Biology, University of North Carolina, Chapel Hill, North Carolina 27599-3280 Auxin-binding protein 1 (ABP1) is a putative receptor for the class of plant growth hormones designated aux- ins of which indole-3-acetic acid (IAA) is the predomi- nant endogenous member. ABPl is a homodimeric gly- coprotein consisting of subunits of 163 amino acid residues. We have performed a structural study of ABPl that has localized a region along its primary sequence that is involved in hormone binding.We have used the photoaffinity labeling agent, 5-[7-SH]azidoindole-3-ace- tic acid (5-[SHlNsIAA), an active auxin analog, to co- valently label residues that are within, or near, the aux- in-binding site. Photolabeled ABPl was digested to completion with trypsin, and the resulting peptides were purified by reverse phase high performance liquid chromatography. When 5-ISHINsIAA was used at a con- centration of 0.5 p~ (one order of magnitude below the Kd for 5-NsIAA) only one peptide was labeled at a high specific activity. Labeling was blocked by the presence of 50 p~ lAA, indicating that the interaction is specific. Sequenceanalysisdeterminedthatthistrypticfrag- ment was derived fromIlelS0 to Leu’4s of ABP1. We sug- gest that residue AsplS4 is the specjfic target of the pho- tolabeling and is within 1.48 A of the postulated hydrophobic platform of the auxin-binding site. We pro- pose that Trp1s6 may serve as this hydrophobic platform in the binding site for the aromatic rings of auxins. Auxins are a class of plant hormones defined by their ability to promote cellular elongation, but they are also involved in other plant responses such as cell division, differentiation, po- larity, and wound responsiveness (Davies, 1987). Indole-3-ace- tic acid (MI1 is the most extensively studied member of the auxin class. While auxin responses have been well character- ized, the exact biochemical mechanism of auxin action remains unclear, including how auxins are perceived by the cell. There are several auxin-binding proteins (ABP) that have been iden- tified and characterized (Jones, 1994). On the basis of a com- parison between growth induction and binding affinities for a large set of compounds (Ray et al., 1977), one of these, now ~ ~~~ * This work was supported by grants from the University of North J. C. B.), the United States Department of Agriculture National Com- Carolina at Chapel Hill Undergraduate Research Award Program (to petitive Research Initiative Grants Program, and the National Science Foundation (Research Experience for Undergraduates) (to A. M. J.). The costs of publication of this article were defrayed in part by the “aduertisement” in accordance with 18 U.S.C.Section 1734 solely to payment of page charges. Thisarticle must therefore be hereby marked indicate-thisfact. Deut. of Bioloev. Universitv of North Carolina. Chapel Hill. NC 27599- $To whom correspondence should be addressed: 318 Coker Hall, 3280. Tel.: 919-962-6932; Fax: 919-962-1625; E-mail: alan-jones@ unc.edu. The abbreviationsused are: IAA, indole-3-acetic acid; ABPI, auxin- TPCK, L-1-tosylamido-2-phenylethyl chloromethyl ketone; PAGE, poly- bindingprotein 1; 5-C3HIN,IAA, tritiated 5-azidoindole-3-aceticacid; acrylamide gel electrophoresis; HPLC, high performance liquid chro- matography; NAA, naphthalene-I-acetic acid. designatedABP1, is probably involved in mediating some of the rapid auxin effects on cell elongation. ABPl is a low abundance glycoprotein ubiquitously foundin plants. It is a 163-amino acid homodimeric protein with sub- units having a molecular mass of 22 kDa. ABPl binds the most active auxins with Kd values in the range of 0.1-10 m~ (Ray, et al., 1977) and ,with a narrow pH optimum of 5.5 (Lobler and Klambt, 1985). The mature protein contains a single high man- nose glycosylation site at AsnS8. ABPI also exists in several isoforms that differ slightly in their amino acid sequences at the amino terminus (e.g. Schwob et al. (1993)). Greater than 90% of the cellular ABPl is located within the endoplasmic reticulum lumen (Jones et al., 1989; Shimomura et al., 1988; Napier et al., 19921, consistent with thepresence of the endo- plasmic reticulum retention signal, KDEL, at its carboxyl ter- minus. While most of the ABPl pool is found in the endoplasmic reticulum, monospecific antibodies against ABPl have been used to detect low levels of ABPl on the Golgi, plasma mem- brane, and within the cell wall space (Jones and Herman, 1993). This is consistent with indirect evidence suggesting that some ABPl is located extracellularly and binds extracellular auxin and that this ABPl/auxin complex interacts with a plasma membrane partner to modulate channel or pump ac- tivities (Barbier-Brygoo, et al., 1989; Ruck, et al., 1993; Thiel, et al., 1993). Presumably, these changes in ion movement lead to changes in cell wall structure, one of the rapid responses that auxins evoke to cause cell elongation (Senn and Goldsmith, 1988; Cleland, 1987). It is generally agreed that ABP1 is one of the candidate receptors for auxin-induced growth (discussedin Jones (1994)). We have initiated a study on ABPl structure to more fully understand the function of this putativehormone receptor and to provide information for a structure-based design of new growth regulators. We report here the first direct evidence for the position of residue(s) in the auxin-binding site within ABPl using thephotoaffinity labeling reagent 5-[7-3Hlazidoindole-3- acetic acid(5-[3H]N31AA) and microsequence analysis of photo- labeled ABPl tryptic fragments. We use this mapping data to extend a recent model of the binding site of ABPl (Edgerton et al., 1994). 5-N,IAA, has been shown to be an active auxin in several plants (Melhado et al., 1981) and can be a specific photoaffinity labeling reagent (Jones and Venis, 1989). 5-N31AA, like IAA, is transported in maize sections at the same rate as IAAand with the same unique polar movement (Jones et al., 1991). Finally, 5-N31AAbindsto ABPl with the same affinity (Kd= 5 pd as IAA (Jones et al., 1984). EXPERIMENTAL PROCEDURES Plant Material and Chemicals-Maize seeds (hybrid57710, Jacques Seed Co., Prescott, WI) were planted in moist vermiculite and grown at 25 “C in the dark for 5 days. EDTA, magnesium chloride, hydrochloric acid, calcium chloride, I-butanol, sodium chloride, citric acid, magnesium sulfate, dtrapure urea, and Scintiverse BD were purchased from Fisher. The urea was purified further using mixed bed resin. Phenylmethylsulfonyl fluoride, 21136
Transcript of THE JOURNAL of 269, No. 33, Issue OF The and in U.S.A ... · Mapping the ABPl Auxin-binding Site...

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol . 269, No. 33, Issue of August pp. 2113621140, 1994 Printed in U.S.A.
Mapping the Auxin-binding Site of Auxin-binding Protein l*
(Received for publication, May 6, 1994, and in revised form, June 13, 1994)
Justin C. Brown and Alan M. Jones$ From the Department of Biology, University of North Carolina, Chapel Hill, North Carolina 27599-3280
Auxin-binding protein 1 (ABP1) is a putative receptor for the class of plant growth hormones designated aux- ins of which indole-3-acetic acid (IAA) is the predomi- nant endogenous member. ABPl is a homodimeric gly- coprotein consisting of subunits of 163 amino acid residues. We have performed a structural study of ABPl that has localized a region along its primary sequence that is involved in hormone binding. We have used the photoaffinity labeling agent, 5-[7-SH]azidoindole-3-ace- tic acid (5-[SHlNsIAA), an active auxin analog, to co- valently label residues that are within, or near, the aux- in-binding site. Photolabeled ABPl was digested to completion with trypsin, and the resulting peptides were purified by reverse phase high performance liquid chromatography. When 5-ISHINsIAA was used at a con- centration of 0.5 p~ (one order of magnitude below the Kd for 5-NsIAA) only one peptide was labeled at a high specific activity. Labeling was blocked by the presence of 50 p~ lAA, indicating that the interaction is specific. Sequence analysis determined that this tryptic frag- ment was derived from IlelS0 to Leu’4s of ABP1. We sug- gest that residue AsplS4 is the specjfic target of the pho- tolabeling and is within 1.48 A of the postulated hydrophobic platform of the auxin-binding site. We pro- pose that Trp1s6 may serve as this hydrophobic platform in the binding site for the aromatic rings of auxins.
Auxins are a class of plant hormones defined by their ability to promote cellular elongation, but they are also involved in other plant responses such as cell division, differentiation, po- larity, and wound responsiveness (Davies, 1987). Indole-3-ace- tic acid ( M I 1 is the most extensively studied member of the auxin class. While auxin responses have been well character- ized, the exact biochemical mechanism of auxin action remains unclear, including how auxins are perceived by the cell. There are several auxin-binding proteins (ABP) that have been iden- tified and characterized (Jones, 1994). On the basis of a com- parison between growth induction and binding affinities for a large set of compounds (Ray et al., 1977), one of these, now
~ ~~~
* This work was supported by grants from the University of North
J. C. B.), the United States Department of Agriculture National Com- Carolina at Chapel Hill Undergraduate Research Award Program (to
petitive Research Initiative Grants Program, and the National Science Foundation (Research Experience for Undergraduates) (to A. M. J.). The costs of publication of this article were defrayed in part by the
“aduertisement” in accordance with 18 U.S.C. Section 1734 solely to payment of page charges. This article must therefore be hereby marked
indicate-this fact.
Deut. of Bioloev. Universitv of North Carolina. Chapel Hill. NC 27599- $To whom correspondence should be addressed: 318 Coker Hall,
3280. Tel.: 919-962-6932; Fax: 919-962-1625; E-mail: alan-jones@ unc.edu.
The abbreviations used are: IAA, indole-3-acetic acid; ABPI, auxin-
TPCK, L-1-tosylamido-2-phenylethyl chloromethyl ketone; PAGE, poly- binding protein 1; 5-C3HIN,IAA, tritiated 5-azidoindole-3-acetic acid;
acrylamide gel electrophoresis; HPLC, high performance liquid chro- matography; NAA, naphthalene-I-acetic acid.
designatedABP1, is probably involved in mediating some of the rapid auxin effects on cell elongation.
ABPl is a low abundance glycoprotein ubiquitously found in plants. I t is a 163-amino acid homodimeric protein with sub- units having a molecular mass of 22 kDa. ABPl binds the most active auxins with Kd values in the range of 0.1-10 m~ (Ray, et al., 1977) and ,with a narrow pH optimum of 5.5 (Lobler and Klambt, 1985). The mature protein contains a single high man- nose glycosylation site at AsnS8. ABPI also exists in several isoforms that differ slightly in their amino acid sequences a t the amino terminus (e.g. Schwob et al. (1993)). Greater than 90% of the cellular ABPl is located within the endoplasmic reticulum lumen (Jones et al., 1989; Shimomura et al., 1988; Napier et al., 19921, consistent with the presence of the endo- plasmic reticulum retention signal, KDEL, at its carboxyl ter- minus. While most of the ABPl pool is found in the endoplasmic reticulum, monospecific antibodies against ABPl have been used to detect low levels of ABPl on the Golgi, plasma mem- brane, and within the cell wall space (Jones and Herman, 1993). This is consistent with indirect evidence suggesting that some ABPl is located extracellularly and binds extracellular auxin and that this ABPl/auxin complex interacts with a plasma membrane partner to modulate channel or pump ac- tivities (Barbier-Brygoo, et al., 1989; Ruck, et al., 1993; Thiel, et al., 1993). Presumably, these changes in ion movement lead to changes in cell wall structure, one of the rapid responses that auxins evoke to cause cell elongation (Senn and Goldsmith, 1988; Cleland, 1987). I t is generally agreed that ABP1 is one of the candidate receptors for auxin-induced growth (discussed in Jones (1994)).
We have initiated a study on ABPl structure to more fully understand the function of this putative hormone receptor and to provide information for a structure-based design of new growth regulators. We report here the first direct evidence for the position of residue(s) in the auxin-binding site within ABPl using the photoaffinity labeling reagent 5-[7-3Hlazidoindole-3- acetic acid (5-[3H]N31AA) and microsequence analysis of photo- labeled ABPl tryptic fragments. We use this mapping data to extend a recent model of the binding site of ABPl (Edgerton et al., 1994).
5-N,IAA, has been shown to be an active auxin in several plants (Melhado et al., 1981) and can be a specific photoaffinity labeling reagent (Jones and Venis, 1989). 5-N31AA, like IAA, is transported in maize sections at the same rate as IAAand with the same unique polar movement (Jones et al., 1991). Finally, 5-N31AAbinds to ABPl with the same affinity (Kd= 5 pd as IAA (Jones et al., 1984).
EXPERIMENTAL PROCEDURES Plant Material and Chemicals-Maize seeds (hybrid 57710, Jacques
Seed Co., Prescott, WI) were planted in moist vermiculite and grown at 25 “C in the dark for 5 days.
EDTA, magnesium chloride, hydrochloric acid, calcium chloride, I-butanol, sodium chloride, citric acid, magnesium sulfate, dtrapure urea, and Scintiverse BD were purchased from Fisher. The urea was purified further using mixed bed resin. Phenylmethylsulfonyl fluoride,
21136

Mapping the ABPl Auxin-binding Site 21137
2-mercaptoethanol, sucrose, TMD-8 mixed bed resin, dithioerythreitol, iodoacetamide, goat IgG directed against rabbit IgG and conjugated with alkaline phosphatase, and indole-3-acetic acid were purchased from Sigma. IAA was further purified by recrystallization in 95% etha- nol. Tris was purchased from Life Technologies, Inc. Sodium citrate was purchased from the Baker Co. TPCK-treated trypsin was purchased from Worthington. Trifluoroacetic acid was purchased from Pierce. HPLC-grade acetonitrile was purchased from EM Science Industries Inc.
[7-3Hl, 5-azidoindole-3-acetic acid was synthesized as described by Melhado et al. (1982). The polyclonal antibodies were prepared against the 22-kDa ABPl protein as described by Jones et al. (1991).
22-kDa ABPl Preparation-The purification ofABPl was camed out using a modified version of the protocol described by Viola (1991). Briefly, maize shoots (typically 1 kg) were homogenized in a large War- ing blender with 1 ml of homogenizing buffer 1 (HBl: 50 mM Tris-C1, pH 8.0, 1 mM EDTA, 0.1 mM MgCI,, 1.5 mM phenylmethylsulfonyl fluoride, and 1.5 mM 2-mercaptoethanol) per 2 g of tissue. The homogenate was filtered through one layer of nylon cloth and clarified by centrifugation a t 3,000 x g for 10 min. Calcium chloride was added to the supernatant to make a final concentration of 10 mM. The microsomal fraction, col- lected by centrifugation a t 20,000 x g for 30 min, was resuspended in HB1 using one-tenth of the original HB1 volume. One volume of l-bu- tanol (at -20 "C) was added to the resuspension with vigorous stirring on ice. The bottom aqueous phase was dialyzed overnight against 100 volumes of HB1. The dialyzed resuspension was loaded onto a Pharma- cia fast flow Q-Sepharose column (2.0 x 23 cm) equilibrated with ho- mogenizing buffer 2 (HB2: 50 mM Tris, pH 8.0, 1 mM EDTA, and 1 mM MgCI,). Proteins were eluted a t a rate of 1.0 mumin, using a 0-1.0 M NaCl continuous gradient in HB2, into 5-ml fractions (0-0.5 M over 60 min and then 0.5-1.0 M over 20 rnin). Fractions enriched in ABPl were combined. The pH was adjusted to 5.5 using 0.5 N citric acid, and then the fractions were added to a phenylacetic acid-Sepharose column (3 x 3 cm, prepared using epoxy-activated Sepharose 6B from Pharmacia Biotech Inc. by manufacturer's protocol) equilibrated with binding buffer (BB: 10 mM sodium citrate, pH 5.50,5 mM MgCl,). The slurry was rotated for 20 min followed by two washes each of 5 volumes of 1 M NaCl (in BB) followed by BB. ABPl was eluted using one volume of elution buffer (50 mM Tris, pH 9.0,5 mM MgSO,). The pH of the elution fractions was immediately adjusted to 7.5, and the fractions were concentrated to less than 1.0 ml using a Centriprep 10 concentrator a t 2500 x g, 90 min.
SDS-PAGE and Immunoblot Analysis-Confirmation of the identity of the purified protein and estimation of the yield and purity were carried out by Western blot analysis using anti-ABP1 polyclonal rabbit antibodies (NC04, 1:2000 dilution) and silver staining, respectively. SDS-PAGE was performed using 15% polyacrylamide gels. Samples for immunoblot analysis were transferred to nitrocellulose and processed with anti-ABP1 polyclonal antibodies. Goat anti-rabbit immunoglobulin conjugated with alkaline phosphatase was the secondary reporter. The protein yield was estimated by comparing the silver stain intensity of the 22-kDa ABPl band with the staining intensity of known quantities of carbonic anhydrase.
Photoaffinity Labeling-All manipulations involving 5-[3HlN,IAA were performed under dim red light in open 3.0-ml glass reaction vials. Separate labeling reactions using approximately 30 pg of ABPl were each performed using 5-[3H]N31AA both with and without the presence of 50 p~ IAA. The ABPl solution was prepared for labeling by adding one-half volume of 3 x modified RM Buffer (750 mM sucrose, 150 mM sodium citrate, pH 5.4), which reduced the reaction pH to 5.5. In a parallel reaction, IAA was added as an ethanolic stock of IAA for a final IAA concentration of 50 PM. 5-[3HlN,IAA was also added from an eth- anolic stock for a final concentration of 0.5 or 5 p ~ . The ethanolic stocks were added first to the reaction vials, and the ethanol was evaporated to near dryness under a stream of N, gas. The modified protein solu- tions were added to the prepared reaction vials, mixed by swirling, and incubated on ice for 30 min. Photolysis (60 s) was carried out by placing the reaction vials between two UV sources mounted 15 cm apart in opposition (Jones and Venis, 1989). The UV sources were a 254-nm Spectronics XX15-F (mounted above) and a 300-nm Spectronics XX15-B (mounted below) that generated an irradiance of approximately 10 milliwatts/cm2 a t the sample level. In the experiment testing various auxin analogs for inhibition of photolabeling (see Fig. 2), the reaction vials were rapidly frozen in liquid nitrogen prior to photolysis. One percent by volume of each reaction was subjected to SDS-PAGE (15%) and was exposed to preflashed Kodak XAR-5 film for 3-7 days a t -80 "C. Gels prepared for fluorography were Coomassie-stained for 2 h and soaked in 1 M salicylic acid for 1 h.
1 2 3 4 5 6 7
FIG. 1. Pure AFSPl is specifically photolabeled. The left panel (lanes 1 4 ) is the silver-stained gel of the purification of ABPl from maize coleoptile; the central panel (lane 5 ) is an immunoblot; and the right panel (lanes 6 and 7) is a fluorogram. Lane 1 , solubilized micro- somal proteins prior to anion exchange separation; lane 2, proteins eluted from the anion exchange column in fractions that are enriched in auxin binding and ABP1; lane 3, affinity column flow-through; lane 4, concentrated affinity column elution containing ABPl at the purity used in this study; lane 5, immunoblot analysis of ABPl (sample as in lane 4 ) probed with polyclonal anti-ABP1 (NC04) used a t a concentra- tion of 1:2000; lane 6, ABPl (as in lane 4 ) photolabeled with 0.5 PM 5-[3H]N31AA in the presence of 50 PM competing IAA, lane 7, proteins photolabeled with 0.5 p~ 5-[3HlN31AA without IAA. In all cases, 1 5 1 acrylamide gels were used in SDS-PAGE.
'Ilypsin Digestion-Photolyzed samples were lyophilized to complete dryness, redissolved in 4 M urea, 250 mM Tris, pH 8.6, and then reduced and alkylated by incubation for 30 min a t 37 "C in 50 mM dithioeryth- reitol followed by 1 h a t 23 "C in the dark in 130 mM iodoacetamide. The solution was diluted 10 times in water and TPCK-treated trypsin was added for a trypsidABP1 mass ratio of approximately 1:30. The diges- tion reactions were incubated overnight a t 37 "C.
Peptide Separation and Identification-Tryptic fragments were separated using a reverse phase Microsorb-MV C,, HPLC column a t a flow rate of 1 mumin. Peptides were eluted along a continuous gradi- ent of 0-100% CH,CN in 0.1% trifluoroacetic acid. Amino acid se- quencing analysis of the appropriate fraction was performed by auto- mated Edman degradation on a gas-phase sequencer (model 4704 Applied Biosystems).
RESULTS AND DISCUSSION ABPl was purified to near homogeneity using anion ex-
change and affinity chromatographies by the method of Viola (1991) with slight modifications described under "Experimen- tal Procedures." The narrow pH optimum for auxin binding enabled us to elute ABPl from the phenylacetic acid Sepharose column using high pH buffer instead of excessive amounts of free ligand. While 5 m~ naphthalene-1-acetic acid (NAA, a synthetic auxin) will elute ABPl from phenylacetic acid-Sepha- rose, the difficulty in removing NAA from the samples compli- cated subsequent photoaffinity labeling experiments. Silver staining and immunoblot analysis indicates that ABPl is pure and undegraded (Fig. 1). ABPl is photoaffinity labeled using 5-[3H]N31AA at 0.5 VM. Labeling is specific because 50 PM IAA, a concentration equivalent to 10 times the Kd (90% occupancy), substantially blocks the incorporation of 5-[3H]N31AA. This suggests that 5-[3H]N31AAinserts at a single site within or near enough to the binding site such that occupation by IAA in this site is sufficient to displace 5-[3HlN31AA.
Photoaffinity labeling by 5-[3H]N31AA is pH-dependent. As shown in Table I, photoaffinity labeling is reduce 10-fold by a 16-fold change in H+ concentration. This pH dependence is the same as that previously shown for NAA binding to purified ABPl using an equilibrium technique (Lobler and Klambt, 1985), further supporting the idea that photoaffinity labeling ABPl by 5-[3H]N31AA, at least at concentrations below the K,, is dependent on occupancy of the binding site. A small set of auxins and one compound with similar structure that is not an
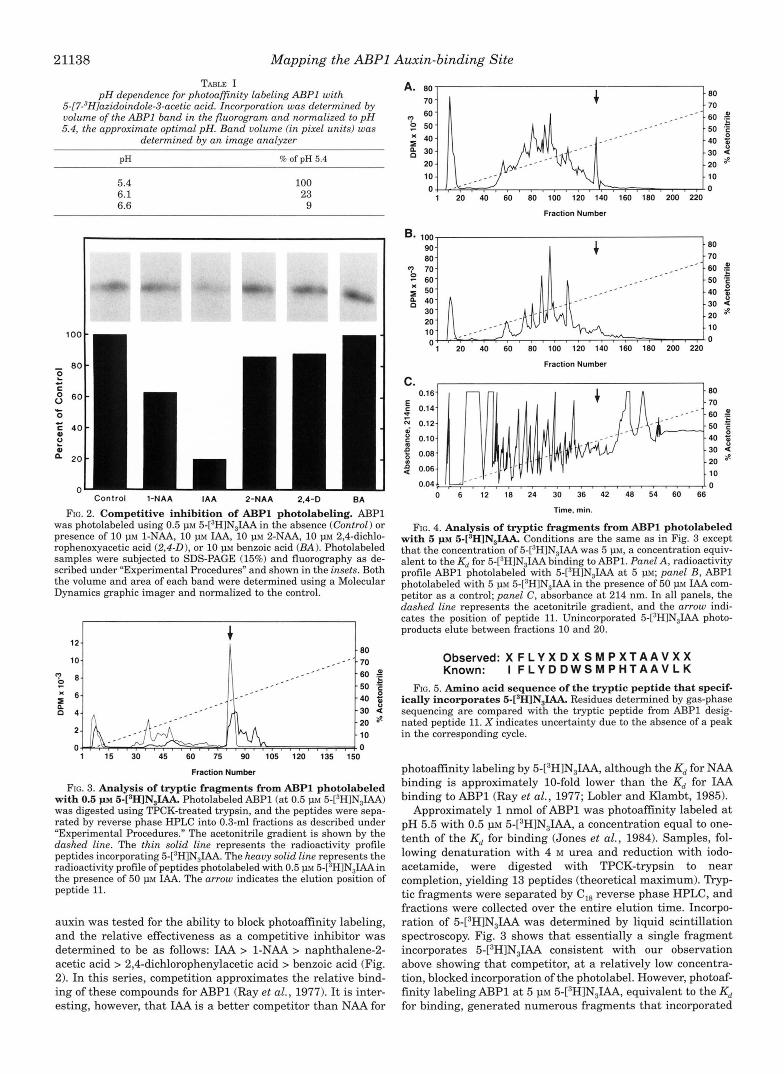
21138 Mapping the ABPl Auxin-binding Site TABLE I
pH dependence for photoafinity labeling ABPl with 5-[7-3H]azidoindole-3-acetic acid. Incorporation was determined by volume of the U P 1 band in the fluorogram and normalized to pH 5.4, the approximate optimal pH. Band volume f in pixel units) was
determined by an image analyzer
PH % of pH 5.4
5.4 6.1 6.6
100 23 9
Control 1-NAA IAA 2-NAA 2.4-D BA
FIG. 2. Competitive inhibition of ABPl photolabeling. AJ3Pl was photolabeled using 0.5 p~ 5-[’HlN,IAA in the absence (Control) or presence of 10 p~ 1-NAA, 10 PM IAA, 10 PM 2-NAA, 10 p~ 2,4-dichlo- rophenoxyacetic acid (2,4-0), or 10 p~ benzoic acid (BA) . Photolabeled samples were subjected to SDS-PAGE (15%) and fluorography as de- scribed under “Experimental Procedures” and shown in the insets. Both the volume and area of each band were determined using a Molecular Dynamics graphic imager and normalized to the control.
I 8 0
E n ”
1 15 30 45 60 75 90 105 120 135 150
I IW t 10
Fraction Number
FIG. 3. Analysis of tryptic fragments from ABPl photolabeled with 0.5 p~ 5 - [ s H ] N , ~ Photolabeled ABPl (at 0.5 PM 5-[3HlN31AA) was digested using TPCK-treated trypsin, and the peptides were sepa- rated by reverse phase HPLC into 0.3-ml fractions as described under ”Experimental Procedures.” The acetonitrile gradient is shown by the dashed line. The thin solid line represents the radioactivity profile peptides incorporating 5-[’H]N,IAA. The heavy solid line represents the radioactivity profile of peptides photolabeled with 0.5 p~ 5-[’H]N,IAAin the presence of 50 p~ IAA. The arrow indicates the elution position of peptide 11.
auxin was tested for the ability to block photoaffinity labeling, and the relative effectiveness as a competitive inhibitor was determined to be as follows: IAA > 1-NAA > naphthalene-2- acetic acid > 2,4-dichlorophenylacetic acid > benzoic acid (Fig. 2). In this series, competition approximates the relative bind- ing of these compounds for ABPl (Ray et al., 1977). I t is inter- esting, however, that IAA is a better competitor than NAA for
? 60- 50-
40- ”
’
” _ - -””.60 _.-
-50 f a
_ _ ” - 4 0 -30 4 -20 8 - 10 0 k . . . . . , .~
1 20 40 60 80 100 120 140 160 180 200 220
Fraction Numbet
B.
8 x
E O
100 ~
90. .c .80 - 70 80 - 70. 60 - 50’
”” ” 6 0 $ - 5 0 ‘E -40 5 -30 -l
1 ”-”
-20 8. . l o 0 o ” ” ’ ” ” ” ’ ’ ” ” ” ” ”
1 20 40 60 80 100 120 140 160 180 200 220
Fraction Number
c. I 1
0.16
0.14
N 0.12 i
-t F
e 0.10
g 0.08 v) 2 0.06
0.04 0 6 12 18 24 30 36 42 48 54 60 66
Time. min.
FIG. 4. Analysis of tryptic fragments from ABPl photolabeled with 5 p~ 5-[sH]N,IAA. Conditions are the same as in Fig. 3 except that the concentration of 5-[”H]N,IAA was 5 p ~ , a concentration equiv- alent to the Kd for 5-?H]N,IAA binding to ABP1. Panel A, radioactivity profile ABPl photolabeled with 5-[’H1N3IAA a t 5 PM; panel B , ABPl photolabeled with 5 p~ 5-[3H]N,IAA in the presence of 50 p~ IAA com- petitor as a control; panel C , absorbance a t 214 nm. In all panels, the dashed line represents the acetonitrile gradient, and the arrow indi- cates the position of peptide 11. Unincorporated 5-[3H]N31AA photo- products elute between fractions 10 and 20.
Observed: X F L Y X D X S M P X T A A V X X Known: I F L Y D D W S M P H T A A V L K
FIG. 5. Amino acid sequence of the tryptic peptide that specif- ically incorporates 5-[’H]N,IAA. Residues determined by gas-phase sequencing are compared with the tryptic peptide from ABPl desig- nated peptide 11. X indicates uncertainty due to the absence of a peak in the corresponding cycle.
photoaffinity labeling by 5-[3H]N,IAA, although the Kd for NAA binding is approximately 10-fold lower than the Kd for IAA binding to AJ3Pl (Ray et al., 1977; Lobler and Klambt, 1985).
Approximately 1 nmol of ABPl was photoaffinity labeled a t pH 5.5 with 0.5 1.1~ 5-[3H]N31AA, a concentration equal to one- tenth of the Kd for binding (Jones et al., 1984). Samples, fol- lowing denaturation with 4 M urea and reduction with iodo- acetamide, were digested with TPCK-trypsin to near completion, yielding 13 peptides (theoretical maximum). nyP- tic fragments were separated by C,, reverse phase HPLC, and fractions were collected over the entire elution time. Incorpo- ration of 5-[3H]N31AA was determined by liquid scintillation spectroscopy. Fig. 3 shows that essentially a single fragment incorporates 5-[3H]N31AA consistent with our observation above showing that competitor, a t a relatively low concentra- tion, blocked incorporation of the photolabel. However, photoaf- finity labeling ABPl at 5 p~ 5-[3H]N31AA, equivalent to the Kd for binding, generated numerous fragments that incorporated

Mapping the ABPl Auxin-binding Site 21139
FIG. 6. Comparison of three ABPl sequences. ABPl sequences were de- duced from cDNAs. The accession num- bers are as follows: Maize, X16309; Ara- bidopsis, S40550; Tobacco, X70903. Identical residues are shaded. Box A and peptide 11 are indicated by boxes. The di- amond indicates the three conserved tryptophans. Trp136, located in peptide 11, is the proposed hydrophobic platform of the auxin-binding site. k g 8 , located in Box A, is the proposed carboxylic acid binding group of the auxin-binding site.
1 10 2 0 3 0 40 s o . o Maize: -SCVRDWSL--V~ DISQMPQSSY QIEGLSWITV AGALNWGMKE VEVWLQTISPG Arabidopsis: -PC-PINGLPIWR N'ISDLPQDNY GRPGESHMTV AGSVLHGMKE VEIWLQTFAPG Tobacco : -HC-SINGLPLVR NfsELPQENV GRSGLSHTTI AGSVLHGMKE IEVWLQTFAPG
6 0 7 0 80 90 100 110
Box A
5-[3H]N31AA as shown in Fig. 4. Only one of these is not labeled in the presence of 50 VM MA, again demonstrating under much higher stringency that this fragment specifically incorporates 5-[3H]N31AA. We interpret this observation of only a single peptide specifically incorporating 5-L3H1N3IAA, even at a high concentration of 5-L3H1N3IAA, to mean that ABPl contains only a single type of 5-[3H]N,IAA-binding site.
Approximately 1 pmol of this specifically labeled fragment was sequenced by automated Edman degradation, and it was determined, as shown in Fig. 5, that the sequence matched a 17-mer tryptic fragment fromAl3Pl designated peptide 11. We conclude that at least one determinant of the auxin-binding site resides within this contiguous stretch of residues. Further- more, since the average lenph of the C-N bond of the photo- generated nitrene is 1.48 A, we conclude that one or more residues on peptide 11 lie within this distance from the binding pocket and are, therefore, likely to form at least part of the binding site.
We do not know specifically which residue within peptide 11 is covalently modified by 5-[3HlN3LU. However, we suggest that Asp'34 is the site of attack based on two indirect argu- ments. First, all known aryl nitrenes are electrophillic and, therefore, are likely to attack nucleophillic amino acids such as those containing carboxylic acids. Within peptide 11 there are two candidates, Asp'34 and Asp135. Second, at this level of sen- sitivity, Asp135 was clearly detected, yet Asp'34 was not, suggest- ing that this residue was modified by covalent attachment of the photogenerated nitrene. The C-H bond on the a carbon is the probable site for insertion due to its relatively lower bond energy. Note that Trp136 and Hid4' were also not identified in the corresponding Edman cycles; therefore, we do not exclude either of these residues from being photolytically tagged. There is precedent that His can be the target of an electrophillic attack by a nitrene (Brinegar, et al., 1988).
Fig. 6 shows the amino acid sequences of ABPl from three plants. Peptide 11, derived by trypsin cleavage near the car- boxyl terminus, is 65% identical among the three sequences, and, for the most part, differences are due to conservative re- placements. The exception is Val'" (maize sequence and num- bering). Edgerton et al. (1994) have proposed that the binding site contains a hydrophobic platform upon which the pheny-, indole-, or naphthalene rings of auxins rests. We suggest that one of the three conserved aromatic residues found in peptide 11, could serve as this critical element of the binding site model. Trp'36 is the most hydrophobic of these and the most suitable to serve as an indole-binding platform.
Another highly conserved region of ABPl is located between Thr54 and Gly7'. This region, designated Box A (Jones, 1994),
Peptide 11 has been proposed to contain the auxin-binding site by Venis et al. (1992). Based on our evidence here, we conclude that the structural requisites for the auxin-binding site do not lie en- tirely within Box A as previously proposed.
However, we do not preclude a structural role for Box Ain the auxin-binding site. Rather, we propose a working hypothesis that incorporates both the model of the auxin-binding site pro- posed by Edgerton et al. (1994) and the model by Venis et al. (1992). These two models are briefly described here.
Edgerton et al. (1994) used an active analog approach to form a molecular model of the auxin-binding site. They introduced the term phytophore, analogous to the term pharmacophore, to describe the idealized auxin ligand conformation within the binding site. They proposed that the phytophoric conformation is one in which the plane of the carboxylic acid group is per- pendicular to the plane of the aromatic ring system. The car- boxylic acid oxygens are positioned directly astride the ring and interact with a positive point charge or charge cluster that has a somewhat flexible position. Their model of the auxin-binding site describes three regions of the protein: a planer aromatic ring-binding platform, a carboxylic acid binding site, and a hydrophobic transition region that separates the two binding sites. The two critical elements of this model that pertain to the current study are the aromatic ring-binding platform and the carboxylic acid binding site. Edgerton et al. (1994) proposed that the ring system of auxins interacted in parallel fashion through T electron sharing with another aromatic ring, prob- ably the side chain of a Trp residue. The carboxylic acid-binding group, probably an amide of an Arg residue, is located above or below this plane and would serve to orient binding (providing specificity) and increase the free energy change (increasing affinity). Other models of the auxin-binding site, all formulated using auxin-induced growth data instead of auxin-binding data (Rakhaminova et al., 1978; Katekar, 1979; Chiriac et al., 1983), incorporate the hydrophobic binding platform and/or the car- boxylic acid binding site features as described by the Edgerton et al. (1994) model.
The model of Venis et al. (1992) is based on previous studies indicating a possible role in auxin binding for His, Cys, Tyr, or Lys (Venis, 1977) and Arg (Nave and Benveniste, 1984). They rationalized that Box A is the binding site because it contains most of the residues implicated by the chemical modification experiments (see Fig. 6). To test this, they prepared antibodies to a synthetic Box A and found that these antibodies, in the absence of auxin, stimulate hyperpolarization of tobacco proto- plast plasma membranes in a manner similar to auxin stimu- lation. This result lead these authors to conclude that the an- tibodies are auxin agonists, that Box A is the auxin-binding

21140 Mapping the ABPl Auxin-binding Site
site, and that ABPl acts on the external face of the plasma membrane.
We propose that the two key binding groups in the Edgerton et al. model of the auxin-binding site are formed by residues located in both Box A and peptide 11. Specifically, the hydro- phobic platform could be formed by ! I '~P '~~ (from peptide 111, and the carboxylic acid binding site could be formed by the positive charge cluster of His-Ar8'-His (from Box A) or byAr8' alone.
The identification of the region of ABPl spanning from Ile"' to as a region of functional significance is an important step toward understanding the precise structural and chemical nature of the ABPl binding site for auxin and is prerequisite to a structure-based design of agrichemicals that regulate growth.
Acknowledgments-We thank Kara Hiller and Dr. Dave mapper, both of University of North Carolina, for advice concerning trypsin digestion and separation of the tryptic fragments; Dr. Jane Walent, University of Wisconsin Biotechnology Center for sequence analysis; and Susan Whitfield, University of North Carolina, for preparing the illustrations.
REFERENCES
Edgerton, M. D., Tropsha, A,, and Jones, A. M. (1994) Phytochemistry 35, 1111-
Jones, A. M. (1994) Annu. Reu. Plant Physiol. Plant Mol. Biol. 45, 393420 Jones, A. M., and Herman, E. (1993) Plant Physiol. (Bethesda) 101,595-606 Jones, A. M., and Venis, M. A. (1989) Proc. Natl. Acad. Sci. U. S. A. 86,6153-6156 Jones, A. M., Melhado, L. L., Ho, T.-H. D., and Leonard, N. J. (1984) Plant Physiol.
Jones, A. M., Lamerson, P., and Venis, M. A. (1989) Plunta 179, 409-414 Jones, A. M, Cochran, D. S., Lamerson, P. L., Cohen J., and Evans, M. (1991) Plant
Katekar, G. F. (1979) Phytochemistry 18, 223-233 Lobler, M., and Klambt, D. (1985) J. Biol. Chem. 260,9848-9853 Melhado, L. L., Jones, A. M., Leonard, N. J., and Vanderhoef L. N. (1981) Plant
Melhado, L. L., Pearce, C. J., DAlarco, M., and Leonard, N. J. (1982) Phytochem-
Napier, R. M., Fowke, L. C., Hawes, C., Lewis, M., Pelham, H. R. B. (1992) J. Cell
Nave, J.-F., and Benveniste, P. (1984) Plant Physiol. (Bethesda) 74, 1035-1040 Rakhaminova, A. B., Khavkin, E. E., and Yaguzhinskii, L. S. (1978) Biochemistry
Ray, P. M., Dohrmann, U., and Hertel, R. (1977) Piant Physiol. (Bethesda) 60,
Ruck, A,, Palme, K., Venis, M. A,, Napier, R. M., and Felle, H. H. (1993) Plant J. 4,
Schwob, E. Choi,, S-Y, Simmons, C., Migliaccio, F., Ilag, L., Hesse, T., Palme, K.,
Senn. A. P.. and Goldsmith, M. H. M. (1988) Plant Physiol. (Bethesda) 88,131-38
opment, Martinus Nijhoff, Boston, MA
1123
(Bethesda) 74, 294302
Physiol. (Bethesdaj 97,352-358
PhysioZ. (Bethesda) 68, 469475
istry 21,2879-2885
Sei. 102,261-271
(Engl. l h z n s . Bwkhimiya) 43,639-653
585-591
41-46
and SOU, D. (1993) Plant J. 4,423-432
Barbier-Brygoo, H., Epfitikhine, G., K h h t , D., Ghis la , M., and G ~ ~ ~ , J. Shimomura, s., Inohma, N., Fukui, T., and Futai, M.-(1988) Plants 175,558-566 (1989) Proc. Natl. Acad. Sci. U. S. A. 86, 891-895 Thiel, G., Blatt, M. R., Fricker, M. D., White, I. R., andMillner, P. (1993)Proc. Natl.
Brinegar,A. C., Cooper, G., Stevens,A., Hauer, C. R., Shabanowitz, J., Hunt, D. F., h a d . sei . U. s. A. So, 11493-11497 Fox, J. E. (1988jProc. Natl. Acad. Sci., U. S. A. 86, 5927-5931 Venis, M. A. (1977) Planta 134, 145-49
Chiriac, A,, Chiriac, V., Ciubotariu, D., Holban, S., and Simon Z. (1983)Eur: J. Med. Venis, M. A,, Napier, R. M., Barhier-Brygoo, H., Maurel, C., Perrot-Rechenmann,
. .
r h o - C W W - K l . ? C.. and Guern, J. (1992) Proc. Natl. Acad. Sci. U. S. A. 89, 7208-7212 Cleland, R. E. (1987) Physiology of Cell Expansion during Plant Growth, pp. 18-27, Viola, G. (1991) Proteine aus maize (Zea mays L . ) und Lowenmaulchen (Antirrhi-
Davies, F? J. (1987) Plant Hormones and Their Role in Plant Growth and Deuel- tein und Nucleinsaureebene. Ph.D. Thesis, University of Bonn num m j u s L,): Untersuchungen zur Isolierung und Charakterisierung auf Pro-
.,," ,,._ ") ". ""
American Society of Plant Physiologists, hckville, MD














![ABPL 90228 Post-Renaissance Architecture - Miles Lewis · Colosseum, RAD2Rome, AD 72-80 wall elevation and cutaway view Trewin Copplestone [ed], World Architecture (London 1963) 591963),](https://static.fdocuments.net/doc/165x107/5ad80ab37f8b9a6b668dc03f/abpl-90228-post-renaissance-architecture-miles-rad2rome-ad-72-80-wall-elevation.jpg)



