
The Immunosuppressant FK-506 Specifically Inhibits Mitogen ...
14
Vol. 11, No. 8 MOLECULAR AND CELLULAR BIOLOGY, Aug. 1991, p. 4074-4087 0270-7306/91/084074-14$02.00/0 Copyright © 1991, American Society for Microbiology The Immunosuppressant FK-506 Specifically Inhibits Mitogen- Induced Activation of the Interleukin-2 Promoter and the Isolated Enhancer Elements NFIL-2A and NF-AT1 SUNANDITA S. BANERJI, JANEY N. PARSONS, AND MICHAEL J. TOCCI* Department of Molecular Immunology, Merck, Sharp and Dohme Research Laboratories, P.O. Box 2000, Rahway, New Jersey 07065 Received 7 November 1990/Accepted 7 May 1991 The macrolide FK-506, like the cyclic undecapeptide cyclosporin A (CsA), is a potent immunosuppressant that interferes with the transcriptional activation of several early-phase genes in T lymphocytes, including that for interleukin-2 (IL-2). We compared the effects of FK-506 and CsA on transcription from the 5' upstream activating sequences (UAS) of the human IL-2 gene and several cellular and viral UAS to define cis-acting sites which may be responsive to FK-506. The UAS surveyed included the human IL-2 receptor a-chain, human metallothionein II, simian virus 40 early, human cytomegalovirus immediate-early, adenovirus major late, and Rous sarcoma virus long terminal repeat UAS. In addition, we studied multimers of several defined promoter elements (NFIL-2A, NF-KcB, or NF-AT1) which are found in the UAS of the human IL-2 gene and which have been reported to be responsive to CsA when linked to a minimal promoter element (TATA box and transcription start site). Each promoter-regulatory region was fused to the bacterial chloramphenicol acetyltransferase gene and used to transiently transfect Jurkat cells. Quantitative chloramphenicol acetyltrans- ferase assay determinations indicated that the transcriptional activity of each UAS induced upon T-cell activation was (i) completely sensitive, (ii) partially sensitive, or (iii) resistant to inhibition by CsA and FK-506. The induced transcription driven by the IL-2 promoter elements NF-AT1 and NFIL-2A could be blocked completely by FK-506 or CsA. Gel mobility shift assays indicated that the binding activities of the factors specifically interacting with these sequences were detected in activated cells regardless of whether the cells were treated with FK-506 or CsA. The results suggest that FK-506 or CsA inhibits a transacting mechanism(s) without disrupting the binding activities of these transcription factors. The degree to which each UAS was resistant to FK-506 was consistent with the level of transcription induced by phorbol myristate acetate, while UAS which were sensitive to inhibition by FK-506 were dependent on the presence of both phorbol myristate acetate and ionomycin. The naturally occurring macrolide FK-506 possesses ex- tremely potent immunosuppressive activity in a variety of in vitro and in vivo assays (27, 34, 35, 49). The immunosup- pressive properties of FK-506 are similar to those of cyclo- sporin A (CsA), a cyclic undecapeptide which has been used extensively to prevent graft rejection in human organ trans- plants (20, 24, 54, 59, 66). CsA and FK-506, while structur- ally unrelated, each bind to and inhibit the enzymatic activ- ities of distinct ubiquitous cytosolic proteins which catalyze cis-trans prolyl isomerization (peptidyl-prolyl isomerases [PPlases]) (13, 23, 57, 61). While similar in their enzymatic activities, the CsA and FK-506 binding proteins share little or no amino acid homology (67). Despite this intriguing coincidence, the role that these proteins play in T-cell activation and immunosuppression remains unclear. In addition to their ability to bind to specific PPlases, both FK-506 and CsA block the induction of a select group of early (E)-phase activation genes in T lymphocytes (6, 19, 20, 38, 62). These genes include c-myc and those for a variety of lymphokines, such as interleukin-2 (IL-2), IL-3, IL-4, gran- ulocyte-macrophage colony-stimulating factor, tumor necro- sis factor a, and gamma interferon. Expression of the mRNAs for immediate-early (IE)-, delayed-early-, or late- phase genes as well as constitutive class genes appears to be unaffected by these agents (6, 19, 20, 62). The ability of * Corresponding author. FK-506 or CsA to block T-cell activation can be overcome by the addition of IL-2 to the T cells, demonstrating that the inhibition of IL-2 production is crucial to the mode of action of these compounds. Nuclear run-on transcription experi- ments with nuclei from T cells activated in the presence or absence of CsA or FK-506 have demonstrated that these compounds inhibit transcription of the IL-2 gene (37, 62). Thus, the ability of these compounds to modulate the transcription of a specific class of eukaryotic genes provides a model system with which to explore the process of T-cell activation and the mechanism(s) of action of these agents. Expression of the IL-2 gene requires a signal from the T-cell receptor (TcR) and the activation of protein kinase C (PKC) (for a review, see references 6 and 29). The TcR may be activated by an antigen or molecules which cross-link it, such as anti-TcR antibodies or lectins. The requirement for TcR activation may be circumvented by Ca2+ ionophores. Activation signals are transmitted through the TcR to the nucleus of the T cell, where they induce IL-2 gene transcrip- tion. A 275-bp region located in the 5'-flanking region of the human IL-2 gene has been shown to faithfully regulate the expression of this gene in Jurkat T-lymphoblastoid cells (10). Within the upstream activating sequences (UAS) of the IL-2 gene (UASIL-2) are specific sites, antigen receptor response elements (ARREs), that respond to the recognition of anti- gens (11). The AAREs are major elements controlling tran- scriptional activation of the IL-2 gene after triggering of the 4074 on April 12, 2018 by guest http://mcb.asm.org/ Downloaded from
Transcript of The Immunosuppressant FK-506 Specifically Inhibits Mitogen ...

Vol. 11, No. 8MOLECULAR AND CELLULAR BIOLOGY, Aug. 1991, p. 4074-40870270-7306/91/084074-14$02.00/0Copyright © 1991, American Society for Microbiology
The Immunosuppressant FK-506 Specifically Inhibits Mitogen-Induced Activation of the Interleukin-2 Promoter and the
Isolated Enhancer Elements NFIL-2A and NF-AT1SUNANDITA S. BANERJI, JANEY N. PARSONS, AND MICHAEL J. TOCCI*
Department of Molecular Immunology, Merck, Sharp and Dohme Research Laboratories,P.O. Box 2000, Rahway, New Jersey 07065
Received 7 November 1990/Accepted 7 May 1991
The macrolide FK-506, like the cyclic undecapeptide cyclosporin A (CsA), is a potent immunosuppressantthat interferes with the transcriptional activation of several early-phase genes in T lymphocytes, including thatfor interleukin-2 (IL-2). We compared the effects of FK-506 and CsA on transcription from the 5' upstreamactivating sequences (UAS) of the human IL-2 gene and several cellular and viral UAS to define cis-acting siteswhich may be responsive to FK-506. The UAS surveyed included the human IL-2 receptor a-chain, humanmetallothionein II, simian virus 40 early, human cytomegalovirus immediate-early, adenovirus major late, andRous sarcoma virus long terminal repeat UAS. In addition, we studied multimers of several defined promoterelements (NFIL-2A, NF-KcB, or NF-AT1) which are found in the UAS of the human IL-2 gene and which havebeen reported to be responsive to CsA when linked to a minimal promoter element (TATA box andtranscription start site). Each promoter-regulatory region was fused to the bacterial chloramphenicolacetyltransferase gene and used to transiently transfect Jurkat cells. Quantitative chloramphenicol acetyltrans-ferase assay determinations indicated that the transcriptional activity of each UAS induced upon T-cellactivation was (i) completely sensitive, (ii) partially sensitive, or (iii) resistant to inhibition by CsA and FK-506.The induced transcription driven by the IL-2 promoter elements NF-AT1 and NFIL-2A could be blockedcompletely by FK-506 or CsA. Gel mobility shift assays indicated that the binding activities of the factorsspecifically interacting with these sequences were detected in activated cells regardless of whether the cells weretreated with FK-506 or CsA. The results suggest that FK-506 or CsA inhibits a transacting mechanism(s)without disrupting the binding activities of these transcription factors. The degree to which each UAS wasresistant to FK-506 was consistent with the level of transcription induced by phorbol myristate acetate, whileUAS which were sensitive to inhibition by FK-506 were dependent on the presence of both phorbol myristateacetate and ionomycin.
The naturally occurring macrolide FK-506 possesses ex-tremely potent immunosuppressive activity in a variety of invitro and in vivo assays (27, 34, 35, 49). The immunosup-pressive properties of FK-506 are similar to those of cyclo-sporin A (CsA), a cyclic undecapeptide which has been usedextensively to prevent graft rejection in human organ trans-plants (20, 24, 54, 59, 66). CsA and FK-506, while structur-ally unrelated, each bind to and inhibit the enzymatic activ-ities of distinct ubiquitous cytosolic proteins which catalyzecis-trans prolyl isomerization (peptidyl-prolyl isomerases[PPlases]) (13, 23, 57, 61). While similar in their enzymaticactivities, the CsA and FK-506 binding proteins share littleor no amino acid homology (67). Despite this intriguingcoincidence, the role that these proteins play in T-cellactivation and immunosuppression remains unclear.
In addition to their ability to bind to specific PPlases, bothFK-506 and CsA block the induction of a select group ofearly (E)-phase activation genes in T lymphocytes (6, 19, 20,38, 62). These genes include c-myc and those for a variety oflymphokines, such as interleukin-2 (IL-2), IL-3, IL-4, gran-ulocyte-macrophage colony-stimulating factor, tumor necro-sis factor a, and gamma interferon. Expression of themRNAs for immediate-early (IE)-, delayed-early-, or late-phase genes as well as constitutive class genes appears to beunaffected by these agents (6, 19, 20, 62). The ability of
* Corresponding author.
FK-506 or CsA to block T-cell activation can be overcomeby the addition of IL-2 to the T cells, demonstrating that theinhibition of IL-2 production is crucial to the mode of actionof these compounds. Nuclear run-on transcription experi-ments with nuclei from T cells activated in the presence orabsence of CsA or FK-506 have demonstrated that thesecompounds inhibit transcription of the IL-2 gene (37, 62).Thus, the ability of these compounds to modulate thetranscription of a specific class of eukaryotic genes providesa model system with which to explore the process of T-cellactivation and the mechanism(s) of action of these agents.
Expression of the IL-2 gene requires a signal from theT-cell receptor (TcR) and the activation of protein kinase C(PKC) (for a review, see references 6 and 29). The TcR maybe activated by an antigen or molecules which cross-link it,such as anti-TcR antibodies or lectins. The requirement forTcR activation may be circumvented by Ca2+ ionophores.Activation signals are transmitted through the TcR to thenucleus of the T cell, where they induce IL-2 gene transcrip-tion.A 275-bp region located in the 5'-flanking region of the
human IL-2 gene has been shown to faithfully regulate theexpression of this gene in Jurkat T-lymphoblastoid cells (10).Within the upstream activating sequences (UAS) of the IL-2gene (UASIL-2) are specific sites, antigen receptor responseelements (ARREs), that respond to the recognition of anti-gens (11). The AAREs are major elements controlling tran-scriptional activation of the IL-2 gene after triggering of the
4074
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

INHIBITION OF IL-2 TRANSCRIPTION BY FK-506 4075
TcR. Two ARREs have been identified and defined as thenuclear factor binding sites IL-2A (-69 to -93 bp) andIL-2E (-255 to -285 bp). The inhibition of IL-2 transcrip-tion by CsA has been shown to be mediated through theARREs located in the UASIL 2 (11). The IL-2A and IL-2Eelements contain binding sites for a constitutively expressedoctamer binding transcription factor (13a, 31, 33, 43, 54a)and an inducible transcription factor termed nuclear factorfrom activated T cells (NF-AT1), respectively (11, 12, 53). Inaddition, several other sequence elements in the UASIL 2have been implicated in the activation of IL-2 transcription.These include an AP-1, ATF/CREB-like element and NFIL-2C, an NF-KB-like element (4, 21, 25, 42, 52, 55).To further define the mechanism of action of FK-506, we
examined its effects on the transcriptional activities of anumber of natural and artificial UAS, including the UASIL 2and multimers of the NFIL-2A, NF-KB, and NF-AT1 bind-ing sites. The activity of each of the UAS examined could becategorized as (i) completely sensitive, (ii) partially sensi-tive, or (iii) resistant to inhibition by FK-506. These studiesdemonstrate that FK-506, like CsA, completely inhibits theactivation of the UASIL-2, as well as that of multimers of theNFIL-2A or NF-AT1 elements in Jurkat cells. DNA bindingstudies did not detect significant differences in NF-AT1,NF-KB, and NFIL-2A DNA binding activities in nuclearextracts from induced cells treated with FK-506 or CsA.Lastly, we demonstrate that FK-506 interferes with a Ca2+_associated signal that leads to increased transcription of theIL-2 gene.The selective effects of FK-506 and CsA on T-cell gene
expression suggest that the phorbol myristate acetate(PMA)-induced signal which synergizes with Ca2+ iono-phores is different from that which mediates the induction ofgenes activated by PMA alone and that this signal transduc-tion pathway is restricted to the induction of E-phase acti-vation genes. Understanding the biochemical mechanism ofaction of FK-506 in T-cell suppression could lead to thedesign of uniquely targeted immunosuppressive compoundsand further define the components of the signal transductionpathway involved in T-cell activation.
(Portions of this study were presented at the UCLASymposia on Molecular and Cell Biology, SteamboatSprings, Colo., 1 to 7 April 1990.)
MATERIALS AND METHODS
Cell culture. Jurkat cells, a human leukemic T-cell line (65,69), were grown in RPMI 1640 medium containing 10% fetalcalf serum, 50 U of penicillin per ml, and 50 ,ug of strepto-mycin per ml. For the standard induction of IL-2, cells wereincubated with 25 ng of PMA per ml and 3.5 ,ug of ionomycinper ml or 10 pLg of the lectin concanavalin A (ConA) per ml.The amount of IL-2 secreted into the medium followinginduction by the T-cell activators was determined by anenzyme-linked immunosorbent assay (ELISA) (Collabora-tive Research). Monolayer cultures of MRC-5 and HeLacells were grown as described above in Dulbecco's modifiedEagle's medium containing 10% (vol/vol) fetal calf serum.All cells were grown in a humidified 37°C incubator at 5%CO2.
Plasmid DNAs. The UASIL 2 consisted of a 483-bp frag-ment of the human IL-2 gene 5'-flanking sequence encom-passing nucleotides -440 to +3 linked to the bacterialchloramphenicol acetyltransferase (CAT) gene. Comple-mentary oligonucleotides containing the basal promoter ele-ment from the UASIL 2 were synthesized on an Applied
Biosystems 380A DNA synthesizer and purified by prepar-ative gel electrophoresis. The basal promoter element con-sists of a region of the UASIL 2 containing the IL-2 TATAbox sequence and transcription start site (-39 to +3 bp) withHindlIl and EcoRI ends (lowercasing indicates linker se-quences): 5'aattcACAGTATAAATTGCATCTCTTGTTCAAGAGTTCa3'. The oligonucleotides were phosphorylatedwith [_-32P]ATP and T4 polynucleotide kinase, annealed,and ligated to the 232-bp EcoRI-PvuII fragment from pUC19to insert the basal promoter element upstream of the multiplecloning region. The ligated fragment was cut with HindlIl toremove vector sequences and multiple linker additions. Theresulting 94-bp fragment was cloned into pSVO-SEAP. Dou-ble-stranded (ds) oligodeoxyribonucleotides containing theNF-AT1 (5'gatccGGAGGAAAACTGTTTCATACAGAAGGCGa3') and NFIL-2A (5'gatccGAAAATATGTGTAATATGTAAAACATTTTGa3') sites (11) with BamHI and BgIIHends were prepared for cloning as described above. Inaddition, oligodeoxyribonucleotides containing the sequencefrom -310 to -286 (5'gatccGTATCCCCACCCCCTTAAAGAAAGa3') were prepared. BamHI-restricted fragmentswere cloned upstream of the basal promoter element at theBamHI site of the pUC19 polylinker. Isolated clones werescreened by sequencing with the Sequenase system (U.S.Biologicals) to determine the number and orientation of theinserted elements. For use with the bacterial CAT markergene, the HindIll fragment. from these clones was purifiedand cloned into pNHCAT, a pSVO (18)-derived vectorcontaining a 23-bp polylinker (gift from N. Hutchinson,Merck, Sharp and Dohme Research Laboratories [MSDRL],Rahway, N.J.). The human cytomegalovirus (CMV) IE UAS(UASCMV IE) CAT (pCAT760) contained the regulatory se-quences from -760 to +7 of the CMV-IE promoter-regula-tory region. The pCAT760 plasmid and the NF-KB trimerCAT plasmids were gifts from Mark Stinski of the Universityof Iowa, Iowa City (26). The IL-2Ra-CAT plasmid was a giftfrom W. C. Greene, Duke University, Durham, N.C. (4).The UAScmv-,E-p-galactosidase construct was a gift fromEric Nordby (MSDRL, Rahway, N.J.).DNA transfections. Transient DNA transfections were
performed on Jurkat cells by the DEAE-dextran method asdescribed by Fujita et al. (16) with slight modifications. Inbrief, cells were collected by centrifugation and washed inSTBS (25 mM Tris-HCl [pH 7.4], 137 mM NaCl, 5 mM KCl,0.6 mM Na2HPO4, 0.7 mM CaCl2, 0.5 mM MgCl2). Cells(107) were resuspended in 1 ml of a DNA-DEAE-dextranmixture containing 100 ,ug of DEAE-dextran and 5 to 10 ,ugof DNA in STBS. For cotransfection experiments, 2 p.g ofthe cotransfected reporter plasmid was added to the mixture.Cells were incubated at 37°C for 1 h. Cells were washed withserum-free medium and cultivated in complete medium for24 h at a density of 3.5 x 105 cells per ml before furthermanipulation. For adherent HeLa and MRC-5 cells, DEAE-dextran transfections were conducted on monolayers whichhad been plated at 65% confluence the previous day. Cellswere incubated with the DNA-DEAE-dextran mixture for 1h, after which serum-free medium was added and cells werecultivated for an additional 6 h. Cells were washed withserum-free medium and cultivated for 24 h in completemedium before further manipulation. DNAs were preparedfor use in transfection experiments by two centrifugationsthrough CsCl2 equilibrium gradients.Enzyme assays. CAT enzyme analyses were carried out
essentially as described by Gorman et al. (18). Cell lysatesfor CAT enzyme analyses was prepared by three cycles offreezing-thawing in 250 mM Tris-HCl (pH 7.5). The protein
VOL . 1 l, 1991
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

4076 BANERJI ET AL.
contents of each lysate were determined by the Bradfordmethod (BioRad). Equivalent amounts of protein were usedin the 150-pul assay mixtures, which contained 0.1 ,uCi of14C-chloramphenicol, 250 mM Tris-HCl (pH 7.5), and 2.75mM acetyl coenzyme A. Acetylation occurred at 37°C formore than 4 to 16 h. The reaction was linear throughout thistime. The labeled material was extracted into ethyl acetateand analyzed by thin-layer chromatography. Thin-layerchromatography plates were analyzed on a computer-con-trolled radioanalytic imaging system (AMBIS, San Diego,Calif.). ,-Galactosidase assays were performed as describedby Hollon and Yoshimura (24a) with 5 mM chlorophenolred-p3-D-galactopyranoside as the substrate. Reactions wereperformed in a total volume of 100 ,ul containing 50 ,ug ofcellular lysate at 37°C for 4 h. The resulting P-galactosidaseactivity was read at 574 nm and compared with that in areaction run at the same time with a standard amount of theenzyme.
Preparation of nuclear extracts. Cells were harvested bycentrifugation 2 h after treatment with PMA and ConA. Cellswere swelled by exposure to a buffer containing 10 mM Tris(pH 7.9), 15 mM KCl, 2 mM MgCl2, 0.2 mM EDTA, 1 mMdithiothreitol, and 1 mM phenylmethylsulfonyl fluoride andDounce homogenized with a loose-fitting B pestle. Nucleiwere collected by centrifugation at 25,000 x g for 10 min.Nuclear proteins were extracted from chromatin in a buffercontaining either 0.42 M NaCl (7) or 0.3 M (NH4)2SO4 andprecipitated with 0.2 g of (NH4)2SO4 per ml (46). Extractscontaining 5 to 10 mg of protein per ml were dialyzed againsta buffer containing 20 mM Tris (pH 7.9), 50 mM KCl, 20%glycerol, 0.2 mM EDTA, 1 mM dithiothreitol, and 1 mMphenylmethylsulfonyl fluoride.
Gel electrophoretic mobility shift assays. DNA bindingactivities were detected essentially as described previously(14, 58, 60). Approximately 10 ,ug of nuclear extract wasused in each assay. For competition experiments, oligode-oxyribonucleotides homologous to the sequences of interestwere prepared as described above and the complementarystrands were annealed. The amount of ds DNA was deter-mined by Hoechst dye binding in 2 M NaCl with a Hoeferdedicated fluorimeter. Oligonucleotides used in the gel shiftassays included NF-AT1 and NFIL-2A described above andoligonucleotides corresponding to sites in the IL-2 promoterfor NF-KB-like and AP-1-like sites. The sequence for theNF-KB site was -206 to -180 bp from the IL-2 promoter. Ads oligodeoxyribonucleotide containing the NF-KB site fromthe K light-chain enhancer (58) was also prepared for use incompetitive binding assays. Gels from these experimentswere analyzed on a phosphoimager from Molecular Dynam-ics, Sunnyvale, Calif.), to quantitate the amount of boundradiolabeled oligonucleotide present in each reaction lane.
Reagents. FK-506 and rapamycin were obtained fromRobert Boris, and CsA was synthesized and purified byPhilip Durrette (both at MSDRL, Rahway, N.J.) (62). Stocksolutions of each immunosuppressant were prepared in 95%ethanol from crystalline compounds that were determined tobe greater than 99% pure by high-pressure liquid chromatog-raphy and mass spectrometry.
RESULTS
Effect of FK-506 on the activity of cellular and viralpromoter-regulatory sequences. The striking similarities in theimmunosuppressive activities of CsA and FK-506 promptedus to ask whether FK-506 also inhibits IL-2 transcriptionthrough the UASIL2. Previous studies showed that a reporter
gene linked to the 275-bp UASIL2 (10, 11, 16, 56, 68) istranscriptionally regulated like the endogenous IL-2 gene bysignals emanating from the TcR. To determine whether FK-506 specifically blocked IL-2 transcription from these se-quences or whether inhibition was universal, we examinedthe ability of a variety of cellular (human metallothionein IIUAS [UASMTI11 and human IL-2 receptor a-chain UAS[UASIL2RaJ) and viral (UASCMV-IE, simian virus 40 earlyUAS [UASSV40], Rous sarcoma virus long terminal repeatUAS [UASRSV], and adenovirus major late UAS [UASAdMLI)promoter-regulatory regions to drive induced expression ofthe bacterial CAT gene in the presence or absence of thecompound. These UAS were chosen either on the basis ofbeing regulatory regions for genes known to be induced uponT-cell activation (UASCMV IE [26] and UASIL2Rao [4]) orbecause they contain regulatory regions which respond to theagents used for in vitro activation of T cells (UASSV40 [15],UASRSV [64], and UASMT II [1]). For our experiments, thehuman T-lymphoblastoid cell line Jurkat was used because itfaithfully mimics the early stages of T-cell activation, whichare sensitive to CsA and FK-506 (65, 69).
In these experiments, Jurkat cells were transiently trans-fected with plasmids containing the CAT gene driven bydifferent UAS and activated with the phorbol ester PMA andeither the Ca2+ ionophore ionomycin or the lectin ConA (38,45, 47, 52, 66). Following a 24-h recovery, cells from a singletransfection were stimulated with PMA plus ionomycin anddivided into equal aliquots. The stimulated cells were treatedwith FK-506, CsA, or an equivalent volume of the solventvehicle (95% [vol/vol] ethanol). The results of these initialexperiments (Table 1) showed that all the UAS surveyedgenerated inducible CAT activity upon T-cell activation. Theinhibition of CAT activity by the immunoregulants, how-ever, was selective and did not correlate with the strength ofa particular UAS. The UASIL2 directed a very low level ofCAT activity in uninduced Jurkat cells. Following activationof the cells with PMA and ionomycin, the level of CATactivity directed from the UASIL-2 increases approximately20-fold. In the presence of FK-506 or CsA, this increase wascompletely inhibited. CAT activity expressed by theUASMT-II increased approximately fivefold over the basallevel following activation. In the presence of either immu-noregulant, this induction was only twofold (50% of the levelin the non-drug-treated, induced control). The behavior ofthe UASSV40 and the UASRSV was similar to that of theUASMT-II (data not shown). The UASIL2R, promoted a lowlevel of CAT expression in uninduced Jurkat cells whichincreased 10-fold following T-cell activation. Induction ofthe UASILo2Ra was unaffected by either FK-506 or CsA.Similarly, the activity of the UASCMV IE, while highly induc-ible in Jurkat cells (18-fold), was unaffected by these immu-noregulants. The UASAdML, which exhibited a high level ofconstitutive CAT activity, was resistant to inhibition byFK-506 or CsA in both uninduced and induced Jurkat cells(data not shown).To determine the dose dependence of this effect on each of
the viral and cellular UAS, we treated aliquots of cells froma single transfection with increasing concentrations of eitherFK-506 (0.01 to 10 ng/ml) or CsA (0.1 to 100 ng/ml). Theimmunoregulants were added at the time of induction, andthe cells were harvested 24 h later. Cell lysates were assayedfor CAT activity, and the resulting thin-layer chromatogramswere analyzed on an AMBIS radioanalytic imaging system.To correlate the effect of each compound on the activity ofthe transfected reporter gene with the behavior of theendogenous IL-2 gene, we determined the levels of IL-2
MOL. CELL. BIOL.
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

INHIBITION OF IL-2 TRANSCRIPTION BY FK-506 4077
TABLE 1. Bacterial CAT activity expressed by UAS and multimerized enhancer elements in uninduced and PMA-ionomycin-inducedJurkat cells treated with FK-506 or CsA
% Acetylation' in:
UAS' (no. of experiments) Uninduced cells Induced cells
Not FK-506 CsA Not FK-506 CsAtreatedc treated treated treatedc treated treated
IL-2-CAT (7) 0.6 ± 0.3 0.4 ± 0.3 0.4 ± 0.2 10.8 ± 2.9 0.6 ± 0.4 0.4 ± 0.3IL-2Rt-CAT (5) 0.5 + 0.2 0.4 ± 0.3 0.5 ± 0.3 5.4 ± 0.8 4.6 ± 0.5 5.1 ± 0.7CMV-IE-CAT (7) 2.8 ± 1.7 2.3 ± 1.3 2.4 ± 1.3 38.3 ± 7.0 35.1 ± 7.2 36.6 ± 4.7MT-II-CAT (3) 4.2 ± 1.7 3.6 ± 1.4 3.9 ± 1.4 20.6 ± 4.1 12.8 ± 3.1 13.8 ± 2.9NFIL-2A (2X)-CAT (7) 0.3 ± 0.3 0.7 ± 0.5 0.9 ± 0.5 7.1 ± 0.8 0.7 ± 0.6 0.6 ± 0.6NF-AT1 (2X)-CAT (7) 0.4 ± 0.2 0.6 ± 0.4 0.6 ± 0.3 10.1 ± 3.1 0.6 ± 0.4 0.7 ± 0.6NF-KB (3X)-CAT (5) 0.4 ± 0.3 0.2 ± 0.2 0.3 ± 0.3 8.0 ± 2.1 3.9 ± 0.5 5.1 ± 1.0-285 to -310 bp (3X)-CAT (3) 1.2 ± 0.3 1.3 ± 0.3 1.3 ± 0.4 1.7 ± 0.4 1.2 + 0.3 1.4 ± 0.4
a 2X, dimer; 3X, trimer.b Jurkat cells (10') were transfected with 10 p.g of plasmid DNA and allowed to recover for 24 h. Aliquots from each transfection were induced for 16 h with
PMA (25 ng/ml) and ionomycin (3.5 j.g/ml) or with these inducers plus 10 ng of FK-506 per ml or 100 ng of CsA per ml. Uninduced cells were treated with anequal volume of the solvent vehicle (95% ethanol). Cells were harvested and prepared for CAT assays as described in the text. Equivalent amounts of proteinwere assayed for CAT activity, and the resulting acetylated products were resolved by thin-layer chromatography and quantitated with the AMBIS system.Conversion from mock-transfected cells was considered background and was subtracted from the percent acetylation of each sample. Each value represents theaverage + the standard deviation for three to seven different transfection experiments.
c Percent acetylation was calculated (counts in acetylated material/total counts in the loaded lane), and the amount of acetylation from each UAS in uninducedcells without an immunoregulant was set as 1 U. Remaining values for each set of transfections were normalized to this value.
secreted into the culture medium by cells from each set oftransfections by an ELISA.
In Jurkat cells, the CAT activities expressed by thevarious transfected UAS were (i) completely sensitive, (ii)partially sensitive, or (iii) resistant to inhibition by thesecompounds (Fig. 1). FK-506 and CsA completely inhibitedthe expression of CAT activity by the UASIL-2 in a dose-dependent manner (Fig. 1A), with 50% inhibition at approx-imately 0.03 and 0.4 ng/ml, respectively. Similarly, the levelof IL-2 secreted by the endogenous gene was inhibited byeither FK-506 or CsA (50% inhibitory concentrations[IC50s], and 0.6 to 1.0 ng/ml, respectively). These IC50s aresimilar to those obtained previously for both the inhibition ofIL-2 mRNA accumulation and the inhibition of T-cell prolif-eration (8, 9, 62). FK-506 was approximately 20 times morepotent than CsA in these experiments. We also observedthat in Jurkat cells, CAT activity expressed from the tumornecrosis factor a UAS (UASTNFa), like that expressed fromthe UASIL 2, required both PMA and ionomycin for induc-tion and that induction was blocked by either FK-506 orCsA. However, in the HL60 and THP-1 monocytic cell lines,the UASTNF. can be activated by PMA alone and itsinduction is not sensitive to inhibition by either immunoreg-ulant (2b).As expected, neither FK-506 nor CsA affected the CAT
activity expressed by the UASIL2Ro, (Fig. 1B) (62). Theconstitutive and induced CAT activities expressed by theviral UASCMVIE were also resistant to the effects of thesecompounds (Table 1 and Fig. 1C). FK-506 partially blockedthe activities expressed by the UASMT.II (Fig. 1D), theUASRSV (Fig. 1E), and the UASSv4O (Fig. 1F). Partialinhibition was most evident at high concentrations of eitherFK-506 or CsA and averaged from 45 to 60% inhibition.These results confirmed that FK-506, like CsA, selectivelyblocked the expression of a specific set of genes and did notcause an overall cessation of inducible CAT expression fromthese UAS.
Previous studies demonstrated that rapamycin, a macro-lide closely related in structure to FK-506, while possessingsome immunosuppressive activity of its own, was alsocapable of reversing the immunosuppressive effects of FK-
506 (8, 9). We showed previously that rapamycin alone doesnot block IL-2 mRNA expression (62). Furthermore, ourearlier results indicated that rapamycin slightly enhancedIL-2 mRNA expression (by approximately twofold) in acti-vated T cells. We tested the effects of rapamycin on theUASIL 2 to determine whether this phenomenon was medi-ated by signals transmitted through the IL-2 enhancer. Inthis experiment, Jurkat cells were cotransfected with theUASIL2-CAT and UAScmv-,E-p-galactosidase reportergene plasmids, induced with PMA and ionomycin, dividedinto aliquots, and treated with different amounts of rapamy-cin. The cells and culture media from each sample wereharvested 24 h after treatment and assayed for CAT andP-galactosidase activities and for IL-2 secretion, respec-tively. The results of this experiment (Fig. 2B) demonstratedthat rapamycin slightly enhanced CAT activity driven by theUASIL2 at the same concentrations at which FK-506 exhib-ited its inhibitory effect (0.10 to 10.0 ng/ml). Despite thenearly twofold enhancement in IL-2 mRNA accumulation(62) and IL-2 promoter activity in rapamycin-treated Jurkatcells, the secretion of IL-2 (Fig. 2C) was significantly de-creased by high concentrations (100 to 1,000 ng/ml) ofrapamycin as compared with the vehicle (95% ethanol).P-Galactosidase activity (Fig. 2A) was slightly decreased atthe highest concentrations of rapamycin (100 to 1,000 ng/ml),even though equal amounts of protein were used to assayeach sample, suggesting probable toxicity associated withhigh levels of this compound.
Effect of FK-506 on defined promoter-enhancer elements intransfected Jurkat cells. Activation of the IL-2 gene requiresthe complex interaction of various transcription factorswhich bind to both proximal promoter sequences and distalenhancer elements. The most well-characterized regulatorysites within the UASIL2 are the two ARREs, the IL-2A (-65to -93 bp) and IL-2E (-255 to -285 bp) sites, which havebeen shown to be involved in antigen-inducible activation ofIL-2 transcription (5, 11, 53). Recombinant promoters con-taining multimers of either IL-2A (NFIL-2A binding site) orIL-2E (NF-AT1 binding site) linked to a basal promoterelement activate transcription in transfected Jurkat cells in amanner identical to that of the entire UASIL2 and are
VOL. 11, 1991
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from
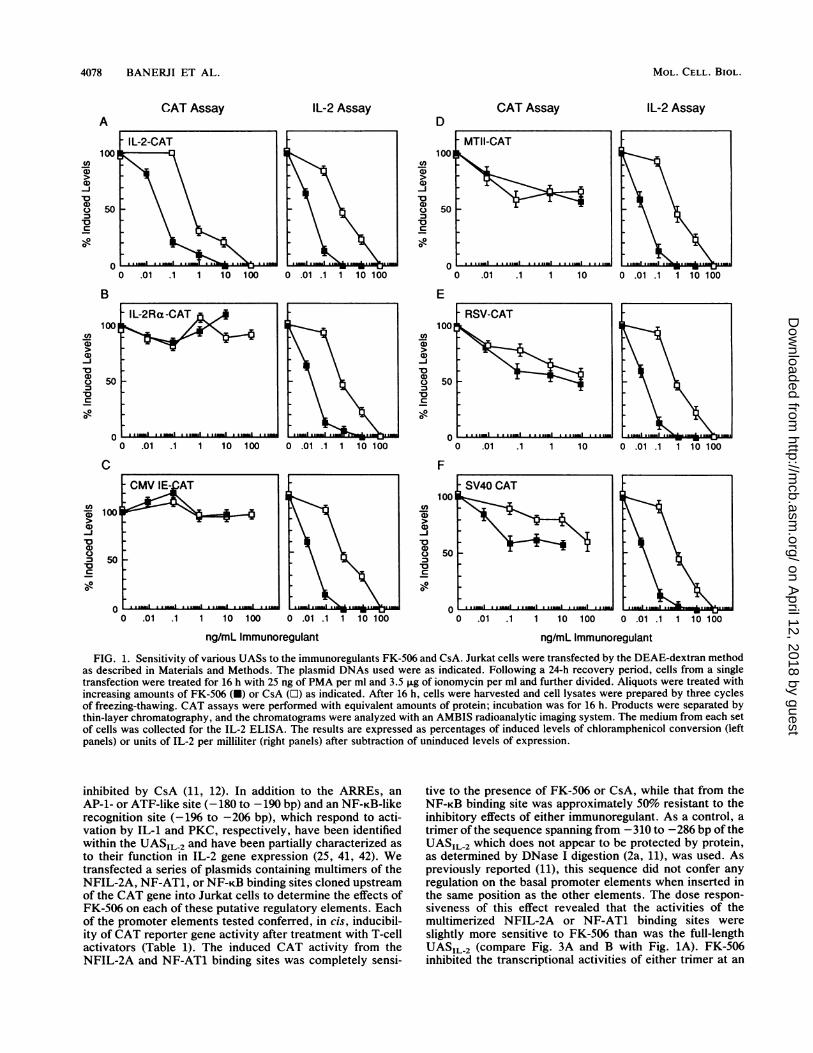
4078 BANERJI ET AL.
CAT Assay
50s-
0 .01 .1 1 10 100
IL-2 AssayD
1 c1Coa)a)-J'0o(D0 C
0--
E
C')7a)'a)-J'0a)C.)
C
-0
C
CMV IE- AT
(0
75100 oCD
a)
0
:3 50"O
0 .
0A I I '--' - MI_I
0 .01 .1 1 10 100 0 .01 .1
F
(I)
a)
a)
-J
c0a)
0
0-
100
50
0
ng/mL Immunoregulant
SV40 CAT
0 .01 .1 1 10 100 0 .01 .1 1 10 100
ng/mL Immunoregulant
FIG. 1. Sensitivity of various UASs to the immunoregulants FK-506 and CsA. Jurkat cells were transfected by the DEAE-dextran methodas described in Materials and Methods. The plasmid DNAs used were as indicated. Following a 24-h recovery period, cells from a singletransfection were treated for 16 h with 25 ng of PMA per ml and 3.5 ,ug of ionomycin per ml and further divided. Aliquots were treated withincreasing amounts of FK-506 (-) or CsA (LI) as indicated. After 16 h, cells were harvested and cell lysates were prepared by three cyclesof freezing-thawing. CAT assays were performed with equivalent amounts of protein; incubation was for 16 h. Products were separated bythin-layer chromatography, and the chromatograms were analyzed with an AMBIS radioanalytic imaging system. The medium from each setof cells was collected for the IL-2 ELISA. The results are expressed as percentages of induced levels of chloramphenicol conversion (leftpanels) or units of IL-2 per milliliter (right panels) after subtraction of uninduced levels of expression.
inhibited by CsA (11, 12). In addition to the ARREs, anAP-1- or ATF-like site (-180 to -190 bp) and an NF-KB-likerecognition site (-196 to -206 bp), which respond to acti-vation by IL-1 and PKC, respectively, have been identifiedwithin the UASIL-2 and have been partially characterized as
to their function in IL-2 gene expression (25, 41, 42). Wetransfected a series of plasmids containing multimers of theNFIL-2A, NF-AT1, or NF-KB binding sites cloned upstreamof the CAT gene into Jurkat cells to determine the effects ofFK-506 on each of these putative regulatory elements. Eachof the promoter elements tested conferred, in cis, inducibil-ity of CAT reporter gene activity after treatment with T-cellactivators (Table 1). The induced CAT activity from theNFIL-2A and NF-AT1 binding sites was completely sensi-
tive to the presence of FK-506 or CsA, while that from theNF-KB binding site was approximately 50% resistant to theinhibitory effects of either immunoregulant. As a control, a
trimer of the sequence spanning from -310 to -286 bp of theUASIL 2 which does not appear to be protected by protein,as determined by DNase I digestion (2a, 11), was used. Aspreviously reported (11), this sequence did not confer anyregulation on the basal promoter elements when inserted inthe same position as the other elements. The dose respon-siveness of this effect revealed that the activities of themultimerized NFIL-2A or NF-AT1 binding sites were
slightly more sensitive to FK-506 than was the full-lengthUASIL-2 (compare Fig. 3A and B with Fig. 1A). FK-506inhibited the transcriptional activities of either trimer at an
ACAT Assay
100
50
0
B
100
IL-2 Assay
(Aa)
a1)-J-oa)
C
0-
a)a)-jl0a1)
C.
0-
0
MOL. CELL. BIOL.
IL-2Ra-CAT
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

INHIBITION OF IL-2 TRANSCRIPTION BY FK-506 4079
CAT AssayA
U)7a)a)-J0Da)
70V
0-.001 .01 .1 1 10 100 1000 10000
100
50
0
B
u)a)
-J40-j'aC.
10-
1000 10000
0 .01 .11 10 100
.° 2-
o C.CO..N0
C
V
.001 .01 .1 1 10 100 1000 10000ng/mI
FIG. 2. Effect of rapamycin on UASIL2-CAT expression andIL-2 secretion. Jurkat cells were cotransfected with 15 jig ofUASIL2-CAT and 5 pLg of UAScmv,E--3-galactosidase plasmids,induced with PMA and ionomycin as described in the text, dividedinto aliquots, and incubated with different concentrations of FK-506, CsA, or rapamycin (RAP) for 24 h. The cells were harvested,lysed, and assayed for CAT and P-galactosidase activities. Theculture medium was also harvested and assayed for secreted IL-2 bythe ELISA. Results are expressed as percentages of activity presentin PMA-ionomycin-induced cells incubated in the absence of drug(percent of induced control). (A) 3-Galactosidase (b-gal) activity;100o equals 625 U/ml. (B) CAT activity; 100%o equals 27% conver-sion (18,500 + 143 cpm) of ['4C-chloramphenicol to acetylatedforms. (C) IL-2 secreted; 100%o equals 400 U/ml.
approximate IC50 of 0.05 ng/ml. In comparison, the IC50 ofCsA was approximately 0.5 ng/ml under the same condi-tions. Thus, the NFIL-2A and NF-AT1 dimers mimic thegeneral behavior of the entire UASIL 2 but differ slightly intheir magnitude of induction (Table 1) and sensitivity toFK-506 or CsA (compare Fig. 3 with Fig. 1). The transcrip-tional activity of the NF-KB trimer (Fig. 3C) in the presenceof FK-506 (IC50, >5 ng/ml) or CsA (IC50, >100 ng/ml) wascomparable to that of the complete UASSV4O or UASRSV,which was partially inhibited by these compounds. TheUASIL2 and its element multimers were completely inactivein HeLa cells or MRC-5 fibroblasts treated with PMA andionomycin, indicating that the transacting factors necessaryfor transcription or the pathway necessary for induction was
(A 1001aDa)
-J
a)
0° 50c
o0
0
0 .01 .1 1 10 100 0 .01 .1 1 10 100
ng/mL ImmunoregulantFIG. 3. Sensitivity of isolated promoter elements to FK-506 and
CsA. Jurkat cells were transfected by the DEAE-dextran method asdescribed in Materials and Methods. Cells from a single transfectionwere subsequently treated for 16 h with 25 ng ofPMA per ml and 3.5F±g of ionomycin per ml and further divided. Aliquots were treatedwith increasing amounts of FK-506 (-) or CsA (O) as indicated.After 16 h, cells were harvested, prepared for CAT analysis, andanalyzed as described in the text. The medium from each set of cellswas collected for the IL-2 ELISA. The results are expressed as
percentages of induced levels of chloramphenicol conversion (leftpanels) or units of IL-2 per milliliter (right panels) after subtractionof uninduced levels of expression.
not operative in these cell types (data not shown). In HeLacells, the NF-KB-driven CAT activity was induced under thesame conditions as those used in Jurkat cells; however, thisinduction was not blocked by FK-506 and required onlyPMA (data not shown).FK-506 specifically inhibits transcription activated by an
ionomycin-induced signal which acts in synergy with a PMA-induced signal in Jurkat cells. FK-506 selectively blocks theactivation of IL-2 and other E-phase activation genes. Oneexplanation for this selectivity is that the induction of eachclass of genes is dependent upon different signal transduc-tion pathways operating in activated T cells (22, 28). For
A. 1oo
80'
:-*"60-00
Xi O- 40D6 20'
-0O-- CsA-*-- FK506
* RAP
NFIL-2A-CAT
.-ads-
lm
IL-2 Assay
B. 200-175 -
150 -
.e'7 125 -
0 00coo275.
0*
250 .01 .1
.001 .01 .1
C.
-.1.--CsA\ \ * ~~~FK506L=~~~~A
_ . 1 -1
1 10 100
NF-KB-CAT
VOL. 11, 1991
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

4080 BANERJI ET AL.
IL-2-CAT CMV IE-CAT SV40-CAT-- u-n FK-506T
O[ + FK-506
60 T
None PMA lono PMA +lono
NF-AT1 -CAT8- -FK-506
O + FK-506
6I
4m
2-
None PMA lono PMA+lono
El - FK-506Q_+ FK-506
20
10
None PMA lono PMA +lono
NFIL-2A-CAT
None PMA lono PMA +
lono
None PMA lono PMA +lono
NF-KB-CAT
None PMA lono PMA +
lono
FIG. 4. Different activation pathways in Jurkat cells. Jurkat cells were transfected as described in Materials and Methods. After recovery
for 24 h, cells from a single transfection were divided into aliquots and treated with the solvent vehicle (None), 25 ng of PMA per ml (PMA),3.5 ,ug of ionomycin per ml (lono), both PMA and ionomycin (PMA + lono), or each of the conditions plus 1.0 ng of FK-506 per ml. Cellswere collected after an additional 16 h and prepared for CAT analysis. Results from the chromatography were quantified with the AMBISsystem. Results are expressed as percent acetylated material after subtraction of percent acetylated material from a cell lysate derived frommock-transfected cells.
example, in Jurkat cells the combination of PMA and iono-mycin or ConA is necessary for the activation of T cells andIL-2 gene expression (29, 65), while PMA alone activates theUASIL2Rao (4, 55). Studies in HeLa cells have shown that theUASMT-II requires PMA for induction (1). PMA can alsoactivate the NF-KB elements in several cell types, includingthose of lymphoid origin (44). To determine whether thedegree of sensitivity to FK-506 was associated with activa-tion by particular stimuli, we examined the effects of FK-506on transcription from several UAS stimulated with PMA orionomycin alone or together in transfected Jurkat cells. Ithas been shown that the addition of PMA to Jurkat cellsenhances the activity of PKC without changing the intracel-lular Ca2+ concentration. Similarly, treatment with Ca2+ionophores modulates intracellular Ca2+ stores without sig-nificantly affecting PKC activity (29). Upon activation, thesesecondary messenger systems, and possibly others, bringabout the cellular changes that typify a T-cell response toantigen.We chose to study (i) the UASIL 2 and multimers of the
NF-AT1 or NFIL-2A binding sites because each mimics thepattern of expression of the endogenous IL-2 gene, (ii) theUASSV40 and multimers of the NF-KB binding site becauseof their intermediate sensitivity to CsA and FK-506, and (iii)the UASCMV-IE because of its resistance to CsA and FK-506.Jurkat cells were transfected with the individual CAT con-structs, and the resulting transfectants were incubated withPMA alone, ionomycin alone, or both PMA and ionomycinin the presence or absence of FK-506 (Fig. 1, 3, and 4). TheUASIL2 and NF-AT1 and NFIL-2A multimers mimicked the
behavior of the endogenous IL-2 gene in that they requiredboth PMA and ionomycin for maximal induction and werecompletely sensitive to FK-506 (Fig. 4). IL-2 was not de-tected in the medium from uninduced cells or in cells treatedwith PMA or ionomycin, regardless of the UAS-CAT con-struct used to transfect the cells. Jurkat cells treated withboth PMA and ionomycin secreted over 300 U of IL-2 per mlinto the culture medium. As expected, activated cells treatedwith FK-506 did not secrete IL-2.PMA alone activated the UASCMV-IE, and the addition of
ionomycin did not increase its transcriptional activity (Fig.4). The UASSV40 and NF-KB trimer were also activated byPMA. However, the addition of ionomycin further enhancedthe transcriptional activities of these sequences. This en-hancement can not be explained simply by Ca2'-mediatedeffects on PKC because PMA-induced transcription of theUASCMVIE was not enhanced in the presence of ionomycin.Therefore, as with the UASIL-2, the combination of PMAand ionomycin provided a signal which synergistically en-
hanced transcription from the UASSV40 and the NF-KBelement. FK-506 had no effect on the transcriptional activityof the UASSV40 or the NF-KB element in PMA-treated cellsor on the induced expression from the UASCMV-IE- How-ever, in PMA- and ionomycin-treated cells, FK-506 reducedthe amount of transcription from the UASSV40 or the NF-KBtrimer by approximately 50%. These results suggest that thesensitivity of a particular UAS or transcriptional regulatoryelement to FK-506 is dependent on a specific signal trans-duction pathway which mediates the activation of thatregulatory sequence. The signaling pathway activated by
c0
01)
MOL. CELL. BIOL.
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

INHIBITION OF IL-2 TRANSCRIPTION BY FK-506 4081
PMA alone is resistant to FK-506, while that requiring bothPMA and ionomycin is completely sensitive.The binding activities of NFIL-2A, NF-AT1, and NF-KB-
like factors are present in induced Jurkat cells treated withFK-506 or CsA. To determine whether FK-506 modified thebehavior of the specific trans-acting factors which interactwith sites important in IL-2 gene regulation, we comparedNFIL-2A, NF-AT1, and NF-KB DNA binding activities innuclear extracts from Jurkat cells treated with or not treatedwith FK-506 (0.5 ng/ml) or CsA (25 ng/ml). For this experi-ment, each immunosuppressant was used at a concentrationthat inhibited IL-2 gene expression by greater than 95% (Fig.3A). To detect the presence of NFIL-2A, NF-AT1, andNF-KB DNA binding activities, we assayed each of theseextracts by a gel electrophoretic mobility shift assay (Fig. 5).NFIL-2A DNA binding activity was present in extracts
from either induced or uninduced Jurkat cells. The levels ofthis activity did not change when the induced cells weretreated with FK-506 or CsA (Fig. 5A). The specificity of theNFIL-2A DNA binding activity in each extract was demon-strated by competition experiments (Fig. 5B). NFIL-2ADNA binding was specifically competed for by an unlabeledoligonucleotide representing the NFIL-2A sequence but notby an oligonucleotide corresponding to the NF-AT1 site.NF-AT1 DNA binding activity (Fig. SC) was not present
in extracts prepared from uninduced Jurkat cells (5, 11, 12,53). Extracts prepared from activated Jurkat cells containeda binding activity which interacted with the NF-AT1 oligo-nucleotide (Fig. SC). Competition assays (Fig. SD) showedthat excess unlabeled NF-AT1 oligonucleotide specificallycompeted for this activity (band A). An additional activity(band A') also appeared upon activation and was competedfor specifically. The appearance of this band, however, wasvariable between preparations, and this band may representa degradation product of the higher-molecular-weight spe-cies. As with NFIL-2A, these activities were not affected bylevels of CsA (25 ng/ml) or FK-506 (0.5 ng/ml) that inhibitedCAT activity from the multimers of the NF-AT1 element.The DNA binding activity designated band B in Fig. SC andD was present in uninduced cells and was induced to higherlevels following the activation of Jurkat cells. The identity ofband B has not been determined.
Binding studies with the NF-KB sequence yielded a com-plex pattern of band-shift products resembling those re-ported by others (4, 25, 50) and suggesting multiple DNAbinding species. A specific NF-KB binding complex wasdetected in induced Jurkat cells (Fig. SE). This bindingcomplex consisted of multiple species and differed slightly inmobility from that observed in extracts prepared from rest-ing cells (Fig. SE). None of the inducible NF-KB DNAbinding activities were affected by levels of FK-506 (0.5ng/ml) or CsA (25 ng/ml) that inhibited 50% of the CATactivity from NF-KB multimers. Binding to the NF-KB-likesequence was also competed for by a 400-fold excess ofunlabeled oligonucleotide corresponding to the NF-KB con-sensus sequence from the immunoglobulin K light chain butnot by an oligonucleotide corresponding to the ATF- orCREB-like sequence (-185 to -170) from the UASIL 2 (datanot shown). Similar studies have been reported by otherswith CsA (11) and, more recently, with FK-506 (see Discus-sion and references 3, 21, 39, and 50). In summary, thesestudies suggest that specific trans-acting factors or com-plexes, such as NFIL-2A, NF-KB, and NF-AT1, are able tobind their appropriate recognition sequences in FK-506-treated Jurkat cells but are unable to activate transcription.
DISCUSSION
Like CsA, FK-506 has been shown to selectively block thetranscription of the IL-2 gene and other E-phase T-cellactivation genes while having little effect on the activation ofother gene classes expressed during T-cell activation (62).We have compared the effects of FK-506 and CsA on thetranscriptional activities of UASIL2, the UAS of severalviral and cellular genes, and three promoter-enhancer ele-ments contained within the UASIL-2. Previously, deletionand functional analyses of multimerized elements driving abasal promoter showed these three elements were critical inregulating the induction of IL-2 transcription following T-cellactivation (5, 10-12, 16, 25, 42, 53, 54a-56). The studiespresented here demonstrate that induced transcriptiondriven by the UASIL2 or by dimers of the NF-AT1 orNFIL-2A binding site is completely sensitive to FK-506.In contrast, induced CAT expression from multimerizedNF-KB sites, the UASRSV, the UASSV40, and the UASMT IIis approximately 50% resistant to FK-506. Activation of theCAT gene driven by the UASCMV IE, the UASAdML, or theUASIL-2Ra was not significantly affected by this compound.While there has been a report that the UASCMv-IE 'ssensitive to CsA (45), the levels of CsA used in that studywere 1 to 10 jLg/ml, 100 to 1,000 times higher than the levelnecessary to completely block T-cell activation (62). Theseobservations clearly demonstrate that diverse UAS exhibitdifferent sensitivities to these compounds on the basis of thesignals required for induction. The ability of FK-506 toinhibit the transcription of a specific class of genes withoutaffecting others suggests that there are multiple pathways forthe activation of each class of genes following T-cell stimu-lation. Each pathway may affect a different transcriptionaltarget characterized by different levels of sensitivity toFK-506.The UAS examined in our studies, like endogenous T-cell
genes, were not equally affected by FK-506. The differencesbetween the responses of the UAS to FK-506 and CsA mayreflect differences in the dependence of individual sequenceson secondary messenger systems utilized during T-cell acti-vation (22, 28). Each of these pathways most likely has aspecific molecular target that responds differently to theseimmunoregulants. The activation of the TcR results in thestimulation of PKC activity and an increase in intracellularCa2 . In vitro, the addition of PMA stimulates PKC activity,while the requirement for increased intracellular Ca2+ can besatisfied by the addition of a Ca2+ ionophore. The IL-2 generequires both PMA and ionomycin for induction (29, 65, 69),while the CMV IE gene (26, 48) and genes dependent onNF-KB (41, 44) require only PMA. Our studies suggest thatthe PMA-associated pathway in Jurkat cells is resistant tothese immunoregulants, while the PMA-ionomycin-ionomy-cin-dependent pathway is completely sensitive to theseimmunoregulants. It is noteworthy that the activation of Tcells through the CD28 complex-PMA-dependent pathwayor the IL-1-PMA-dependent pathway is not sensitive toinhibition by CsA or FK-506 and does not elicit increases inintracellular Ca2+ (30, 61a). In this study, we could notdistinguish whether FK-506 inhibits the Ca2+ signal, therebyeliminating the synergy with PMA, or inhibits a commonpoint along the pathway.
While the complete cascade of signal transduction is notunderstood, it is generally known that the binding of appro-priate extracellular ligands to the surface of T cells ulti-mately triggers the expression of specific genes which to-gether constitute the program for growth and differentiation.
VOL . 1 l, 1991
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

4082 BANERJI ET AL.
B.NF-IL2A
' 5 9*09Jr
+ R-
R S
+ -S
S+C
+ F
S+F
Competitors:NF-AT1 400xNF-IL2A 200xNF-IL2A 400xNE
w_
Free i-
IL-2 units:
Phosphor units:
106
Free b
~~~~~~~~.~ :u *i
n~~~~~~~~~~~~~~~~~~~~~j
* :i .'s~~~~~~~~~~~~~~~~~~~~~~~~~~~~A
as_6 106 8 10
1.3 1.1 1.4 1.2
C.
Probe: NF-AT1 G
NE 1? , 5x 5V
D.
R_ +
SS +
S+C
+ F-
S+F
Competitors:NF-IL2A 400xNF-AT1 200xNF-AT1 400xNE
A V- M.1X
A' w..
.3...
B W- __
Free 0
IL-2 units:
Phosphor units:
105
8 >1000 80 20A 7.1 7.86.8A' 1.3 1.5 1.2B 7.5 11.9 8.1 9.2
A.Probe:
NE
s.
A' o.
B 0
Free >.
i.
MOL. CELL. BIOL.
::
.z,l:I
i.
it .1.1.&' A.
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

INHIBITION OF IL-2 TRANSCRIPTION BY FK-506 4083
F.E.Probe: NF-kBNE V? s 5Jr%Or"
A -
B 10
Free >
IL-2 units:Phosphor units:
105
4 D.. B A
&L.
6 106 8 10A. 08 - - -B. 2.3 - - -B' - 6.3 5.9 5.7C. 7.0 7.3 6.7 6.0D. - 2.3 2.0 2.5
Competitors:- + -_ + -_ + -_ + - NF-kB 400x- - + - - + - - + - - + NF-IL2A 400x
R S S+C S+F NE
*w " t"..
Free >-
FIG. 5. Electrophoretic mobility shift assays. (A, C, and E) Sensitivity ofDNA binding activities to FK-506 or CsA. Nuclear extracts (NE)were prepared as described in Materials and Methods. ds oligonucleotide probes were end labeled with [.y-32P]ATP, and 10 pmol was usedin binding assays containing 10 ,ug of protein, 20 mM Tris (pH 7.5), 12% glycerol, 25 mM KCI, 50 mM NaCl, and 5 j±g of poly(dI dC).Following 1 h of incubation, bound products were resolved from the free probe on a 4% polyacrylamide (30:0.8) Tris-glycine gel system.Extracts were prepared from Jurkat cells treated with the solvent vehicle alone (R, resting), 25 ng of PMA and 10 ng of ConA per ml (S,stimulated), the mitogens plus 0.5 ng of FK-506 per ml (S+F), or the mitogens plus 25 ng of CsA per ml (S+C). (A) NFIL-2A. (C) NF-AT1.(E) NF-KB. Free, unbound oligonucleotide probe. Arrowheads indicate the positions of specific shift-up species. The units of IL-2 producedby an aliquot of cells that were incubated for 24 h and that were used to prepare the 2-h extracts are indicated below each lane. For eachsample, the bands corresponding to specific shift-up products were analyzed on a Molecular Dynamics phosphoimager to quantitate theamount of radioactivity present in each band. The amount of radioactivity present in each band is expressed as linear phophor units beloweach lane. (B, D, and F) Competition experiments for DNA binding activities in extracts from activated Jurkat cells. Nuclear extracts (NE)were prepared as described above. ds oligonucleotide probes were end labeled with [.y-32P]ATP, and 10 pmol was used in binding assayscontaining 10 ,ug of protein from induced cells in the presence or absence of a 200- or 400-fold molar excess of the unlabeled ds oligonucleotideindicated. Following 20 min of incubation, bound products were resolved from the free probe on a 4% polyacrylamide (30:0.8) Tris-glycinegel system. Competitor oligonucleotides used in each assay are indicated. (B) NFIL2-A, (D) NF-AT1, (F) NF-KB.
Among the genes regulated upon T-cell activation are thoserequired for cell cycle progression, such as c-fos and c-myc,and those related to tissue-specific function, such as genesfor various lymphokines. Signal transduction from the cellmembrane to the nucleus involves the activation of nuclearfactors which interact with the UAS of the appropriategenes, thereby modulating their expression. The UASIL 2contains several such sequence elements, which have beenshown genetically to be important for regulation or whichinteract with DNA binding proteins.The region containing the NF-KB-like site in the IL-2
promoter is protected from DNase I digestion in nuclearextracts from stimulated T cells (5, 11). It has been shownthat NF-KB is present but repressed in the cytoplasm ofmany cell types by IKB until treatment of the cells withphorbol esters (2). In the current study, we demonstratedthat a related binding activity in Jurkat cells is inducedfollowing T-cell activation but is not inhibited by eitherFK-506 or CsA. Such results are consistent with earlier
observations that the immediate events in T-cell activationare not blocked by either of these compounds (62). Atconcentrations of FK-506 (0.5 ng/ml) or CsA (25 ng/ml) thatcompletely inhibit IL-2 mRNA expression, the functionalactivity of the isolated NF-KB elements is partially inhibited,while the DNA binding activity does not appear to besignificantly affected. These results suggest that a later eventmay be necessary for this binding activity to become capableof transactivation. Interpretation of these phenomena isfurther complicated by recent results which suggest that theNF-KB-like activity in T cells may be composed of at leastthree or four factors which can form different DNA bindingcomplexes in response to various cellular activation signals(41). Thus, it is not yet clear how the NF-KB-like site in theIL-2 gene participates in the induction of IL-2 gene expres-sion following antigen receptor recognition (11, 25, 55). Inaddition to the elements studied here, the full-length IL-2promoter contains other sequence elements which havesignificant homologies to other PMA-inducible elements,
VOL. 11, 1991
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

4084 BANERJI ET AL.
such as AP-1 (1, 5, 11, 52) and AP-3 (52), which may act inconcert with other factors to regulate IL-2 transcriptionfollowing T-cell activation.
In contrast to the situation with the NF-KB element, CATactivity driven by multimers of the NFIL-2A element wascompletely inhibited by FK-506 and CsA. DNA bindingassays, however, revealed that the binding activity associ-ated with the NFIL-2A site was present in uninduced cellsand was not altered by induction or by the presence ofFK-506 or CsA (12) (Fig. 5). Thus, the DNA binding activityof this factor may not be a direct target for change uponT-cell activation. Instead, the transacting domain of thisfactor may be affected. Alternatively, NFIL-2A may requirean ancillary factor to activate transcription but not binding.Levels or modifications of such an accessory factor may bethe target of regulation. Support for this notion is theobservation that an octamer binding transcription factor siteis contained within the NFIL-2A site (13a, 31, 33, 54a). In aheterologous system, the herpes simplex virus transactivat-ing protein VP16 interacts with the OCT-1 protein in vitro,suggesting that VP16 converts OCT-1 into a transactivatorby providing it with a transactivating domain (17, 36). ADNase I-hypersensitive site is present in the chromatin ofmitogen-induced T cells immediately upstream of theNFIL-2A site. This alteration in chromatin structure couldresult from a change in DNA-factor interactions.The NF-AT1 element, like the NFIL-2A site, has also
been shown by deletion analysis to be important for theactivation of transcription of the IL-2 gene through theantigen receptor. Like the NFIL-2A region, the NF-AT1region is near a (second) DNase I-hypersensitive region inmitogen-induced T cells (56). NF-AT1 binding activity hasnot been detected in uninduced cells or in extracts ofinduced cells treated with inhibitors of protein synthesis(53), indicating that NF-AT1 is expressed de novo uponactivation. The immunoregulants FK-506 and CsA at con-centrations two or three times higher than that required tocompletely inhibit IL-2 mRNA expression and promoteractivation do not significantly affect the appearance ordecrease the level of NF-AT1 binding activity in Jurkat cells2 h after activation. Thus, like the expression of other IEgenes, NF-AT1 expression in Jurkat cells does not appear tobe affected by physiologically relevant concentrations ofFK-506 or CsA. However, the ability of NF-AT1 to functionas a transactivator appears to be dependent on a later eventin T-cell activation which is sensitive to FK-506 and CsA.Alternatively, NF-AT1 may need to interact with an ancil-lary factor(s) which is regulated through the immunoregu-lant-sensitive signal transduction pathway.During the review of this manuscript, several reports
indicating that FK-506 and CsA interfere with the synthesisof NF-AT1, NF-KB, AP-3, and AP-1 appeared (3, 12, 21, 39).In these studies, decreased DNA binding activities wereobserved in Jurkat cells or peripheral blood T lymphocytestreated with high levels of FK-506 or CsA. Thus, at concen-trations 10 to 1,000 times higher than that required tocompletely block IL-2 gene activation, these compoundsmay affect the expression of AP-1, NF-AT1, and NF-KB. Inthese studies (3, 12, 21, 39, 40), binding activities wereassayed 4 to 6 h after T-cell activation and distal to the keyinhibitory event(s) (62). In our study, the DNA bindingactivities for NFIL-2A, NF-AT1, and NF-KB were assayedduring the time period when inhibition by FK-506 or CsA hasbeen shown to occur (62), between 0 and 2 h postinduction.It is during this time period that maximum IL-2 genetranscription is achieved in activated Jurkat cells, 2 to 4 h
after activation. Thus, transcriptional complexes are pre-sumably assembled during the 2-h period after induction.Furthermore, the concentrations of each immunoregulantused in our experiments were only two or three times higherthan that required to completely block IL-2 expression. Onepossible explanation for these apparent differences may bethat specific factors, while inducible, may have shorterhalf-lives in the absence of E-phase gene expression. It isalso important to determine whether decreased binding ofkey transcription factors results from toxicity. Thus, at thephysiological levels at which these compounds mediate theirimmunosuppressive effects, neither CsA nor FK-506 ap-pears to significantly affect the DNA binding of NFIL-2A,NF-AT1, or NF-KB.
In light of these results, it is important to define theprimary mechanistic events versus secondary or tertiaryeffects associated with the use of these compounds. BothFK-506 and CsA are extremely potent entities and produceimmunosuppressive effects at remarkably low concentra-tions in vitro and in vivo. In several species, the levels ofFK-506 in serum are approximately 1 nglml (59), and inhumans and some lower primates, circulating levels areoften below detection limits (<100 pglml) (59, 63). Tissueculture studies have shown that in normal T cells or T-lym-phoblastoid cell lines of human or murine origin, IL-2expression is completely blocked by as little as 0.01 ng ofFK-506 per ml or 10 ng of CsA per ml (3, 12, 39, 62).Furthermore, FK-506 and CsA do not accumulate to highenough levels in cells or tissues to completely saturate andinhibit the activities of the major cytosolic receptors identi-fied to date (51).
In addition to the NFIL-2A, NF-AT1, and NF-KB sites,the IL-2 promoter contains other sequences which sharehomology with the binding sites of previously describedtranscription factors. How factor availability and the spatialarrangement of these binding sites affect IL-2 transcriptionalregulation has not yet been determined (5, 11, 16, 21). Manypromoter studies have revealed that a single factor is usuallynot sufficient to regulate all the activities exhibited by asingle promoter. The tightly controlled regulation of the IL-2gene and the complexity of the IL-2 promoter have shownthat IL-2 expression can be controlled by a variety ofstimuli, each of which affects one or more signal transduc-tion pathways. In turn, these different signals appear tomediate effects on the IL-2 promoter through one or moretranscription factors, some of which may be common tomore than one pathway. The activation of the IL-2 gene byphytohemagglutinin and IL-1 is dependent on an AP-1-likesequence which is not necessary for induction by PMA andphytohemagglutinin (42). Similarly, induction by PMA andionomycin is dependent on the presence of NF-AT1 andNFIL-2A (11). The inhibition of IL-2 expression by FK-506has not been correlated with the presence of a singlesequence element. However, when taken out of context ofthe complete UASIL-2, the NF-AT1 and NFIL-2A sites arecompletely sensitive to FK-506. The UASCMV-IE, UASIL29and UASIL-2RO, sequences all contain NF-KB elements (4, 5,12, 25, 26), yet each displays a different sensitivity toinhibition by FK-506 and CsA. In addition, the UASsv40contains an octamer transcription factor binding site (15) butis only partially sensitive to FK-506 and CsA. These obser-vations are consistent with the concepts that multiple inter-actions may occur in a complex 5'-regulatory sequence andthat only specific interactions are able to drive the properexpression of individual promoters and contribute to theinhibition of promoter activation by FK-506 and CsA.
MOL. CELL. BIOL.
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

INHIBITION OF IL-2 TRANSCRIPTION BY FK-506 4085
The immunoregulants FK-506 and CsA both inhibit theearly stage of T-cell activation by blocking the transcrip-tional induction of a set of genes which includes those forIL-2 and other lymphokines. While these immunoregulantsare structurally unrelated, the proteins which bind them inthe cytoplasm appear to be related in function. Cyclophilin,the binding protein of CsA, has been shown to be a 17-kDaprotein which exhibits PPIase activity (13, 61). The FK-506binding protein is 11 to 13 kDa and has been shown to exhibita similar enzymatic activity. The binding of the respectiveimmunosuppressant to its corresponding binding proteininhibits this enzymatic activity (23, 57). These findingssuggest that the immunoregulants FK-506 and CsA may playa role in altering the secondary structure of certain proteins,perhaps DNA binding proteins or ancillary proteins, to bringabout changes in their ability to function properly. Thispotential explanation for the mechanism of action of thesecompounds has been brought into question by recent obser-vations that the macrolide rapamycin, which is related instructure to FK-506, does not inhibit IL-2 gene activationand can reverse the effects of FK-506 in activated T cells (8,9). In addition, rapamycin can compete with FK-506 for thebinding of the FK-506 binding protein and is also able toblock its PPIase activity at the same concentration asFK-506 (3, 51). In addition, the antagonistic effect of rapa-mycin is highly dependent on the concentration of ionomy-cin used to activate the T cells in vitro. Thus, under someconditions, rapamycin appears to be related mechanisticallyto CsA and FK-506 in its ability to specifically modulate theevents mediated by a rise in intracellular calcium. Onepossible explanation for these observations is that thesecompounds mimic a natural ligand that acts as a regulator ofE-phase activation genes in T cells. These results providefurther evidence that rapamycin affects two different meta-bolic pathways, one involved in E-phase activation eventsand the other involved in late-phase events, such as cellularproliferation and perhaps protein transport and secretion.The selective effects of FK-506 and CsA on gene expres-
sion suggest the existence of a signaling pathway that isrestricted to the induction of a group of E-phase activationgenes in T cells, including that for IL-2, which is necessaryfor autocrine growth of the activated T-cell population.Detailed understanding of this pathway should provide in-sights into the mechanism(s) of T-cell activation. The studyof the inhibitory mechanism(s) of compounds such as FK-506, rapamycin, and CsA can be used to further dissect thebiochemical events involved in this differentiation process.
ACKNOWLEDGMENTS
We thank E. A. O'Neill, N. I. Hutchinson, S. Molineaux, M.Stinski, G. Wiederrecht, B. Frantz, and E. Nordby for criticalreading of the manuscript and helpful discussions. We are grateful toK. Collier, N. I. Hutchinson, and P. Kwok for early participation inthis project, to M. Stinski for the gift of the CMV IE-derivedplasmids, and to A. R. Williamson and N. Sigal for support.
S.S.B. is supported by a Merck, Sharp and Dohme postdoctoralfellowship.
REFERENCES1. Angel, P., M. Imagawa, R. Chiu, B. Stein, R. J. Imbra, H. J.
Rhamsdorf, C. Jonat, P. Herrlich, and M. Karin. 1987. Phorbolester-inducible genes contain a common cis element recognizedby a TPA-modulated trans-acting factor. Cell 49:729-739.
2. Baeuerle, P. A., and D. Baltimore. 1988. IKB: a specific inhibitorof NF-KB transcription factor. Science 242:540-546.
2a.Banerji, S. Unpublished observation.
2b.Banerji, S., A. Siddiqui, D. Mark, and M. Tocci. Unpublisheddata.
3. Bierer, B. E., P. S. Mattila, R. F. Staandaert, L. A. Herzenberg,S. J. Burakoff, G. Crabtree, and S. L. Schreiber. 1990. Twodistinct single transmission pathways in T lymphocytes areinhibited by complexes formed between an immunophilin andeither FK-506 or rapamycin. Proc. Natl. Acad. Sci. USA87:9231-9235.
4. Bohnlein, E., D. W. Ballard, H. Bogerd, N. J. Peffer, J. W.Lowenthal, and W. C. Greene. 1989. Induction of interleukin-2receptor a gene expression is regulated by post-translationalactivation of KB specific DNA binding proteins. J. Biol. Chem.264:8475-8478.
5. Brunvand, M. W., A. Schmidt, and U. Siebenlist. 1988. Nuclearfactors interacting with the mitogen-responsive regulatory re-gion of the interleukin-2 gene. J. Biol. Chem. 263:18904-18910.
6. Crabtree, G. R. 1989. Contingent genetic regulatory events in Tlymphocyte activation. Science 243:355-361.
7. Dignam, J. D., R. M. Lebovitz, and R. G. Roeder. 1983.Accurate transcription initiation by RNA polymerase II in asoluble extract from isolated mammalian nuclei. Nucleic AcidsRes. 11:1475-1489.
8. Dumont, F. J., M. R. Melino, M. J. Staruch, S. L. Koprak, P. A.Fischer, and N. H. Sigal. 1990. The immunosuppressive macro-lides FK-506 and rapamycin act as reciprocal antagonists inmurine T cells. J. Immunol. 144:1418-1424.
9. Dumont, F. J., M. J. Strauch, S. L. Koprak, M. R. Melino, andN. H. Sigal. 1990. Distinct mechanisms of suppression of murineT cell activation by the related macrolides FK-506 and rapamy-cin. J. Immunol. 144:251-258.
10. Durand, D. B., M. R. Bush, J. G. Morgan, A. Weiss, and G. R.Crabtree. 1987. A 275 basepair fragment at the 5' end of theinterleukin-2 gene enhances expression from a heterologouspromoter in response to signals from the T cell antigen receptor.J. Exp. Med. 165:395-407.
11. Durand, D. B., J. P. Shaw, M. R. Bush, R. E. Replogle, R.Belagaje, and G. R. Crabtree. 1988. Characterization of antigenreceptor response elements within the interleukin-2 enhancer.Mol. Cell. Biol. 8:1715-1724.
12. Emmel, E. A., C. L. VerweU, D. B. Durand, K. M. Higgins, E.Lacy, and G. R. Crabtree. 1989. Cyclosporin A specificallyinhibits function of nuclear proteins involved in T-cell activa-tion. Science 246:1617-1620.
13. Fischer, G., L. B. Wittman, K. Lang, T. Kiefhaber, and F. X.Schmid. 1989. Cyclophilin and peptidyl-prolyl cis-trans isomer-ase are probably identical proteins. Nature (London) 337:476-478.
13a.Frantz, B., E. Nordby, M. Tocci, and E. A. O'Neill. Submittedfor publication.
14. Fried, M. G., and D. M. Crothers. 1981. Equilibria and kineticsfor LAC-repressor-operator interactions by polyacrylamide gelelectrophoresis. Nucleic Acids Res. 9:6505-6520.
15. Fromental, C., M. Kanno, H. Nomiyama, and P. Chambon.1988. Cooperativity and hierarchical levels of functional orga-nization in the SV40 enhancer. Cell 54:943-953.
16. Fujita, T., H. Shibuya, T. Ohashi, K. Yamanishi, and T.Taniguchi. 1986. Regulation of the human IL-2 gene: functionalDNA sequences in the 5'-flanking region for gene expression inactivated T-lymphocytes. Cell 46:401-405.
17. Gerster, T., and R. G. Roeder. 1988. A herpesvirus transacti-vating protein interacts with transcription factor OTF-1 andother cellular proteins. Proc. Natl. Acad. Sci. USA 85:6347-6351.
18. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982.Recombinant genomes which express chloramphenicol acetyl-transferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051.
19. Granelli-Piperino, A., L. Andrus, and R. M. Steinman. 1986.Lymphokine and non-lymphokine mRNA levels in stimulatedhuman T cells: kinetics, mitogen requirements and effects ofcyclosporin A. J. Exp. Med. 163:922-937.
20. Granelli-Piperino, A., and M. Keane. 1988. Effects of cyclospo-rine A on T lymphocytes and accessory cells from human blood.Transplant. Proc. 20:136-142.
VOL. 11, 1991
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

4086 BANERJI ET AL.
21. Granelli-Piperino, A., P. Nolan, K. Inaba, and R. M. Steinman.1990. The effects of immunosuppressive agents on the inductionof nuclear factors that bind to sites on the interleukin-2 pro-moter. J. Exp. Med. 172:1869-1872.
22. Gunter, K. C., S. G. Irving, P. F. Zipfel, U. Siebenlist, and K.Kelly. 1989. Cyclosporin A-mediated inhibition of mitogen-induced gene transcription is specific for the mitogen stimulusand cell type. J. Immunol. 142:3286-3291.
23. Harding, M. W., A. Galat, D. E. Wehling, and S. L. Schreiber.1989. A receptor for the immunosuppressant FK-506 is acis-trans peptidyl-prolyl isomerase. Nature (London) 341:758-760.
24. Herold, K. C., D. W. Lancki, R. L. Moldwin, and F. W. Fitch.1986. Immunosuppressive effects of cyclosporine A on cloned Tcells. J. Immunol. 136:1315-1321.
24a.Hollon, T., and F. K. Yoshimura. 1989. Variation in enzymatictransient gene expression assays. Anal. Biochem. 182:411-418.
25. Hoyos, B., D. W. Ballard, E. Bohnlein, M. Siekevitz, and W. C.Greene. 1989. Kappa-B-specific DNA binding proteins: role inthe regulation of human IL-2 gene expression. Science 244:457-460.
26. Hunninghake, G. W., M. M. Monick, B. Liu, and M. F. Stinski.1989. The promoter-regulatory region of the major immediate-early gene of human cytomegalovirus responds to T-lymphocytestimulation and contains functional cyclic AMP-response ele-ments. J. Virol. 63:3026-3033.
27. Inamura, N., M. Hashimoto, K. Nakahara, H. Aoki, I. Yamagu-chi, and M. Kohsaka. 1988. Immunosuppressive effect of FK-506 on collagen-induced arthritis in rats. Clin. Immunol. Immu-nopathol. 46:82-90.
28. Irving, S. G., C. H. June, P. F. Zipfel, U. Siebenlist, and K.Kelly. 1989. Mitogen-induced genes are subject to multiplepathways of regulation in the initial stages of T-cell activation.Mol. Cell. Biol. 9:1034-1040.
29. Isakov, N., M. I. Mally, W. Scholz, and A. Altman. 1987.T-lymphocyte activation: the role of protein kinase C and thebifurcating inositol phospholipid signal transduction pathway.Immunol. Rev. 95:89-111.
30. June, C. H., J. A. Ledbetter, M. M. Gillespie, T. Lindsten, andC. B. Thompson. 1987. T-cell proliferation involving the CD28pathway is associated with cyclosporine-resistant interleukin 2gene expression. Mol. Cell. Biol. 7:4472-4481.
31. Kamps, M. P., L. Corcoran, J. H. LeBowitz, and D. Baltimore.1990. The promoter of the human interleukin-2 gene containstwo octamer-binding sites and is partially activated by theexpression of Oct-2. Mol. Cell. Biol. 10:5464-5472.
32. Kay, J. E., and C. R. Benzie. 1990. Lymphocyte-T activationthrough the C28 pathway is insensitive to inhibition by theimmunosuppressive drug FK-506. Immunol. Lett. 23:155-159.
33. Kemler, I., and W. Schaffner. 1990. Octamer transcriptionfactors and the cell type-specificity of immunoglobulin geneexpression. FASEB J. 4:1444 1449.
34. Kino, T., H. Hatanaka, M. Hashimoto, M. Nishiyama, T. Goto,M. Okuhara, M. Kohsaka, H. Aoki, and H. Imanaka. 1987.FK-506, a novel immunosuppressant isolated from a Streptomy-ces. I. Fermentation, isolation, and physico-chemical and bio-logical characteristics. J. Antibiot. 40:1249-1255.
35. Kino, T., H. Hatanaka, S. Miyata, N. Inamura, M. Nishiyama,T. Yajima, T. Goto, M. Okuhara, M. Kohsaka, H. Aoki, and T.Ochia. 1987. FK-506, a novel immunosuppressant isolated froma Streptomyces. II. Immunosuppressive effect of FK-506 invitro. J. Antibiot. 40:1256-1265.
36. Kristie, T. M., J. H. LeBowitz, and P. A. Sharp. 1989. Theoctamer binding proteins form multi-protein-DNA complexeswith the HSV aTIF regulatory protein. EMBO J. 8:4229-4238.
37. Kronke, M., W. J. Leonard, J. Depper, S. Arya, F. Wong-Stahl,R. C. Gallo, T. Waldman, and W. C. Greene. 1984. CyclosporinA inhibits T-cell growth factor expression at the level of mRNAtranscription. Proc. Natl. Acad. Sci. USA 81:5214-5218.
38. Kronke, M., W. J. Leonard, J. M. Depper, and W. C. Greene.1985. Sequential expression of genes involved in human T-lym-phocyte growth and differentiation. J. Exp. Med. 161:1593-1598.
39. Mattila, P. R., K. S. Ullman, S. Fiering, E. A. Emmel, M.McCutcheon, G. R. Crabtree, and L. A. Herzenberg. 1990. Theactions of cyclosporine A and FK-506 suggest a novel step in theactivation of T-lymphocytes. EMBO J. 9:4425-4433.
40. Mizushima, Y., H. Kosaka, S. Sakuma, K. Kanda, K. Itoh, T.Osugi, A. Mizushima, T. Hamaoka, H. Yoshida, K. Sobue, andK. Fujiwara. 1987. Cyclosporin A inhibits late steps of Tlymphocyte activation after transmembrane signaling. J. Bio-chem. 102:1193-1201.
41. Molitor, J., W. Walker, S. Doerre, D. W. Ballard, and W. C.Greene. 1990. NF-KB: a family of inducible and differentiallyexpressed enhancer binding proteins in human T cells. Proc.Natl. Acad. Sci. USA 87:10028-10032.
42. Muegge, K., T. M. Wifliams, J. Kant, M. Karin, R. Chiu, A.Schmidt, U. Siebenlist, H. A. Young, and S. K. Durum. 1989.Interleukin-1 costimulatory activity on the interleukin-2 pro-moter via AP-1. Science 246:249-251.
43. Nabel, G. J., C. Gorka, and D. Baltimore. 1988. T-cell specificexpression of interleukin-2: evidence for a negative regulatorysite. Proc. Natl. Acad. Sci. USA 85:2934-2938.
44. Nelsen, B., L. Hellman, and R. Sen. 1988. The NF-KB-bindingsite mediates phorbol ester-inducible transcription in nonlym-phoid cells. Mol. Cell. Biol. 8:3526-3531.
45. Niller, H. H., and L. Hennighausen. 1990. Phytohemagglutinin-induced activity of cyclic AMP (cAMP) response elements fromcytomegalovirus is reduced by cyclosporine and synergisticallyenhanced by cAMP. J. Virol. 64:2388-2391.
46. Ohlsson, H., and T. Edlund. 1986. Sequence-specific interac-tions of nuclear factors with the insulin gene enhancer. Cell45:35-44.
47. Rice, G. P., R. D. Schrier, and M. B. Oldstone. 1984. Cytomeg-alovirus infects human lymphocytes and monocytes: virusexpression is restricted to immediate early gene products. Proc.Natl. Acad. Sci. USA 81:6134-6138.
48. Sambucetti, L. C., J. M. Cherrington, G. W. Wilkinson, andE. S. Mocarski. 1989. NF-KB activation of the cytomegalovirusenhancer is mediated by a viral transactivator and by T cellstimulation. EMBO J. 8:4251-4258.
49. Sawada, S., G. Suzuki, Y. Kawase, and F. Takaku. 1987. Novelimmunosuppressive agent, FK-506, in vitro effects on clonedT-cell activation. J. Immunol. 139:1797-1803.
50. Schmidt, A., L. Hennighausen, and U. Siebenlist. 1990. Induciblenuclear factor binding to the KB elements of the human immu-nodeficiency virus enhancer in T cells can be blocked bycyclosporin A in a signal-dependent manner. J. Virol. 64:4037-4041.
51. Schreiber, S. L. 1991. Chemistry and biology of the immuno-philins and their immunosuppressive ligands. Science 251:283-287.
52. Serfling, E., R. Barthelmas, I. Pfeuffer, B. Schenk, S. Zarius, R.Swoboda, F. Mercurio, and M. Karin. 1989. Ubiquitous andlymphocyte-specific factors are involved in the induction of themouse interleukin 2 gene in T lymphocytes. EMBO J. 8:465-473.
53. Shaw, J. P., P. J. Utz, D. B. Durand, J. J. Toole, E. A. Emmel,and G. R. Crabtree. 1988. Identification of a putative regulatorof early T-cell activation genes. Science 241:202-215.
54. Shevach, E. M. 1985. The effects of cyclosporin A on theimmune system. Annu. Rev. Immunol. 3:397-423.
54a.Shibuya, H., and T. Taniguchi. 1989. Identification of multiplecis-elements and trans-acting factors involved in the inducedexpression of the human IL-2 gene. Nucleic Acids Res. 17:9173-9184.
55. Shibuya, H., M. Yonegama, and T. Taniguchi. 1984. Involve-ment of a common transcription factor in the regulated expres-sion of IL-2 and IL-2 receptor genes. Int. J. Immunol. 1:43-49.
56. Siebenlist, U., D. B. Durand, P. Bressler, N. J. Holbrook. C. A.Norris, M. Kamoun, J. A. Kant, and G. R. Crabtree. 1986.Promoter region of interleukin-2 gene undergoes chromatinstructure changes and confers inducibility on chloramphenicolacetyltransferase gene during activation of T cells. Mol. Cell.Biol. 6:3042-3049.
57. Siekierka, J. J., S. H. Hung, M. Poe, C. S. Lin, and N. H. Sigal.
MOL. CELL. BIOL.
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

INHIBITION OF IL-2 TRANSCRIPTION BY FK-506 4087
1989. A cytosolic binding protein for the immunosuppressantFK-506 has peptidyl-prolyl isomerase activity but is distinctfrom cyclophilin. Nature (London) 341:755-757.
58. Singh, H., R. Sen, D. Baltimore, and P. A. Sharp. 1986. Anuclear factor that binds to a conserved sequence motif intranscriptional control elements of immunoglobulin genes. Na-ture (London) 319:154-158.
59. Starzl, T. E., S. Todo, J. Fung, A. J. Demetris, R. Venkataram-man, and A. Jain. 1989. FK-506 for liver, kidney, and pancreastransplantation. Lancet 1i:1000-1004.
60. Strauss, F., and A. Varshavsky. 1984. A protein binds to asatellite DNA repeat at three specific sites that would bebrought into mutual proximity by DNA folding in the nucleo-some. Cell 37:889-901.
61. Takahashi, N., T. Hayano, and M. Suzuki. 1989. Peptidyl-prolylcis-trans isomerase is the cyclosporin A-binding protein cyclo-philin. Nature (London) 337:473-475.
61a.Tocci, M., and F. Dumont. Unpublished observations.62. Tocci, M. J., D. A. Matkovich, K. A. Colier, P. Kwok, F.
Dumont, C. S. Lin, S. DeGudicibus, J. J. Siekierka, J. Chin, andN. I. Hutchinson. 1989. The immunosuppressant FK-506 selec-tively inhibits expression of early T cell activation genes. J.Immunol. 143:718-726.
63. Todo, S., Y. Ueda, J. A. Demeris, 0. Inventarza, M. Nalesnik, R.Venkataranan, L. Makowka, and T. E. Starzl. 1988. Immuno-suppression of canine, monkey and baboon allografts by FK-506: with special reference to synergism with other drugs andtolerance induction. Surgery 104:239-249.
64. Vacca, A., S. Martinotti, I. Screpanti, M. Maroder, M. P. Felr,A. R. Farina, A. Gismondi, A. Santoni, L. Frati, and A. Gulino.1990. Transcriptional regulation of the interleukin-2 gene byglucocorticoid hormones. Role of steroid receptor and antigen-responsive 5'-flanking sequences. J. Biol. Chem. 265:8075-8080.
65. Weiss, A., R. L. Wiskocil, and J. D. Stobo. 1984. The role of T3surface molecules in the activation of human T cells: a twostimulus requirement for IL-2 production reflects events occur-ring at a pre-translational level. J. Immunol. 133:123-128.
66. White, D. J. G., and R. Y. Calne. 1982. The use of cyclosporinA: immunosuppression in organ grafting. Immunol. Rev. 65:115-131.
67. Wiederrecht, G., L. Brizuela, K. Elliston, N. H. Sigal, and J. J.Siekierka. 1991. FKBI encodes a nonessential FK 506-bindingprotein in Saccharomyces cerevisiae and contains regions sug-gesting homology to the cyclophillins. Proc. Natl. Acad. Sci.USA 88:1029-1033.
68. Williams, T. M., L. Eisenberg, J. E. Burlein, C. A. Norris, S.Pancer, D. Yao, S. Burger, M. Kamoun, and J. A. Kant. 1988.Two regions within the human IL-2 gene promoter are impor-tant for inducible IL-2 expression. J. Immunol. 141:662-666.
69. Wiskocil, R., A. Weiss, J. Imboden, R. Kamin-Lewis, and J.Stobo. 1985. Activation of a human T-cell line: a two stimulusrequirement in the pre-translational events involved in thecoordinate expression of interleukin-2 and y-interferon genes. J.Immunol. 134:1599-1603.
VOL . 1 l, 1991
on April 12, 2018 by guest
http://mcb.asm
.org/D
ownloaded from


















