
Sympathetic Control Esophageal Sphincter Function in the Cat
9
Sympathetic Control of Lower Esophageal Sphincter Function in the Cat ACTION OF DIRECT CERVICAL AND SPLANCHNIC NERVE STIMULATION JACQUES FOURNET, WILLIAM J. SNAPE, JR., and SIDNEY COHEN, Gastrointestinal Section of the Department of Medicine, University of Pennsylvania at the Hospital of the University of Pennsylvania, Philadelphia, Pennsylvania 19104 A B S T R A C T The purpose of this study was to de- termine the effect of direct stimulation of the sympa- thetic nerves on the lower esophageal sphincter (LES) in the anesthetized cat. Neither unilateral nor bilateral cervical sympathectomy, or splanchnicectomy signif- icantly modified basal LES pressure in animals with intact vagi, or animals having undergone bilateral cer- vical vagotomy. Electrical stimulation of the cut, pe- ripheral, cervical sympathetic trunk increased mean arterial blood pressure, but had no effect on LES pres- sure or LES relaxation as induced by vagal stimulation. Stimulation of the central end of the cervical sympa- thetic trunk had no effect on LES pressure. Stimulation of the central end of the cut splanchnic nerve produced a decrease in LES pressure with a maximal response of 69.1 ± 16.0% (mean± SEM). This inhibitory response was not modified by either propranolol or bilateral cer- vical vagotomy. Stimulation of the peripheral end of the cut, greater splanchnic nerve gave an increase in LES pressure with a maximal response of 38.2 + 7.19 mm Hg. Guanethidine, in the presence or absence of the adrenal glands, significantly augmented this excita- tory response. This response was also slightly increased by phentolamine alone at 10 V, 1 Hz, but was not altered by propranolol. The excitatory response was completely antagonized by atropine or by trimethaphan camsylate. Stimulation of the peripheral end of the splanchnic nerve inhibited LES relaxation as induced by vagal stimulation. The results of this study suggest that: (a) the LES in the cat is not affected by either central or Dr. Foumet is supported by funds from the University of Grenoble, Grenoble, France. Received for publication 24 July 1978 and in revised form 8 December 1978. 562 peripheral stimulation of the cervical sympathetic trunk; (b) the central portion of the splanchnic nerve carries an afferent inhibitory response to the LES through yet unknown pathways; (c) the peripheral splanchnic nerve carries an atropine-sensitive excita- tory response to the LES; and (d) the splanchnic nerves may modulate LES relaxation as induced by vagal stimulation. INTRODUCTION In recent studies, the role of the vagal nerves in the control of lower esophageal sphincter (LES)' function has been emphasized. The vagi serve a major role in the initiation of LES relaxation during swallowing (1, 2). Excitatory cholinergic pathways in the vagi have been difficult to demonstrate in the opossum, but may be present in the cat (3, 4). Our understanding of the influence of vagal function on the LES had developedl through studies designed to inhibit or stimulate the vagi directly (1, 2). These studies allowed the identi- fication of neural pathways of the nonadrenergic in- hibitory or purinergic type that were not previously recognized during earlier studies, in which only agonists or antagonists were parenterally administered (5). At the present time, our understanding of the sympathetic control of LES function is based upon studies in which alpha or beta adrenergic agonists or antagonists were given (6, 7). Recent studies using accurate measures of LES function have not been per- formed with direct stimulation of the sympathetic in- nervation to the LES (8, 9). 'Abbreviation used in this paper: LES, lower esophageal sphincter. J. Clin. Invest. © The American Society for Clinical Investigation, Inic. (0021-9738/79/0410562/09 $1.00 Volume 63 April 1979 562-570
Transcript of Sympathetic Control Esophageal Sphincter Function in the Cat

Sympathetic Control of Lower Esophageal SphincterFunction in the Cat
ACTION OF DIRECT CERVICAL ANDSPLANCHNIC
NERVESTIMULATION
JACQUES FOURNET, WILLIAM J. SNAPE, JR., and SIDNEY COHEN, GastrointestinalSection of the Department of Medicine, University of Pennsylvania at the Hospitalof the University of Pennsylvania, Philadelphia, Pennsylvania 19104
A B S T RA C T The purpose of this study was to de-termine the effect of direct stimulation of the sympa-thetic nerves on the lower esophageal sphincter (LES)in the anesthetized cat. Neither unilateral nor bilateralcervical sympathectomy, or splanchnicectomy signif-icantly modified basal LES pressure in animals withintact vagi, or animals having undergone bilateral cer-vical vagotomy. Electrical stimulation of the cut, pe-ripheral, cervical sympathetic trunk increased meanarterial blood pressure, but had no effect on LES pres-sure or LES relaxation as induced by vagal stimulation.Stimulation of the central end of the cervical sympa-thetic trunk had no effect on LES pressure. Stimulationof the central end of the cut splanchnic nerve produceda decrease in LES pressure with a maximal response of69.1 ± 16.0% (mean± SEM). This inhibitory responsewas not modified by either propranolol or bilateral cer-vical vagotomy. Stimulation of the peripheral end ofthe cut, greater splanchnic nerve gave an increase inLES pressure with a maximal response of 38.2 + 7.19mmHg. Guanethidine, in the presence or absence ofthe adrenal glands, significantly augmented this excita-tory response. This response was also slightly increasedby phentolamine alone at 10 V, 1 Hz, but was not alteredby propranolol. The excitatory response was completelyantagonized by atropine or by trimethaphan camsylate.Stimulation of the peripheral end of the splanchnicnerve inhibited LES relaxation as induced by vagalstimulation. The results of this study suggest that: (a)the LES in the cat is not affected by either central or
Dr. Foumet is supported by funds from the University ofGrenoble, Grenoble, France.
Received for publication 24 July 1978 and in revised form8 December 1978.
562
peripheral stimulation of the cervical sympathetictrunk; (b) the central portion of the splanchnic nervecarries an afferent inhibitory response to the LESthrough yet unknown pathways; (c) the peripheralsplanchnic nerve carries an atropine-sensitive excita-tory response to the LES; and (d) the splanchnic nervesmay modulate LES relaxation as induced by vagalstimulation.
INTRODUCTION
In recent studies, the role of the vagal nerves in thecontrol of lower esophageal sphincter (LES)' functionhas been emphasized. The vagi serve a major role inthe initiation of LES relaxation during swallowing (1,2). Excitatory cholinergic pathways in the vagi havebeen difficult to demonstrate in the opossum, but maybe present in the cat (3, 4). Our understanding of theinfluence of vagal function on the LES had developedlthrough studies designed to inhibit or stimulate thevagi directly (1, 2). These studies allowed the identi-fication of neural pathways of the nonadrenergic in-hibitory or purinergic type that were not previouslyrecognized during earlier studies, in which onlyagonists or antagonists were parenterally administered(5). At the present time, our understanding of thesympathetic control of LES function is based uponstudies in which alpha or beta adrenergic agonists orantagonists were given (6, 7). Recent studies usingaccurate measures of LES function have not been per-formed with direct stimulation of the sympathetic in-nervation to the LES (8, 9).
'Abbreviation used in this paper: LES, lower esophagealsphincter.
J. Clin. Invest. © The American Society for Clinical Investigation, Inic. (0021-9738/79/0410562/09 $1.00Volume 63 April 1979 562-570

The purpose of this study is threefold: first, to de-termine the effect of direct stimulation or section ofthe cervical sympathetic or greater splanchnic nerves;second, to identifv the nieural transmitter involved inmediating these changes in LES pressure; and third,to study the interaction between the sympathetic andvagal pathways involved in the regulation of LESfunction.
NIETHODS
Studies were performedl in 34 adult cats of either sex, weigh-ing between 2.0 anid 4.5 kg. Each animal was fasted for 12-18 hbefore the manometric study and was initially anesthetizedwith sodium pentobarbital (50 mg/kg, i.p.) or ketamine (10mg/kg). Intravenous anesthesia was given as needed into acatheter in the femoral vein (pentobarbital, 10 mg; or keta-mine, 2.0 mg/kg). After induction of the anesthesia, a tracheot-omy was performed for assisted ventilation with a respirator.A cannula inserted into the femoral artery was filled withheparin and connected to an external transducer (StathamP231A, Statham Instruments, Inc., Oxnard, Calif.) for constantblood pressure monitoring. The rectal temperature of theanimal was maintained at a constant level with a heat lamp.
The neck was explored through a midline incision. Theright and left vago-sympathetic trunks were identified withinthe carotid sheath between the internal jugular vein andthe carotid artery. The sympathetic and vagal trunks werethan isolated during microscopic examination and securedwith two loops of umbilical tape. The sympathetic trunk wascharacterized as being smaller than the cervical vagus. Thesympathetic trunk had prominent ganglia and rami coimmlluIni-cantes going to the spinal cord. Nerve identity was confirmedat postmortem examiniation by determining continuity withthe throacic sympathetic trunk.
The abdomen was opened through a midline inlcisioIn. Theright or left greater splanchnic nerve was iclentified andisolated as it transversed the diaphragm before its entry intothe celiac ganglion. The trunk was then secured with twoloops of umbilical tape. Nerve identity was again confirmedat postmortem examination by identifying its proximal con-nection to the thoracic svmpathetic trunk and its distalconnection to the celiac ganglion.
Esophageal manometric studies were performed using atube assembly consisting of three polyvinyl catheters (1.4 mminside diameter) vith side orifices of similar size spaced at5-cm intervals. Each catheter was continuously infused withdistilled water at a rate of 1.2 ml/min. This rate of infusiongave a >200 mmHg/s rise in pressure upon occlusion of therecording orifice. The intraluminal pressures were transmittedto external transducers with outputs graphed on a Beckmnanirectilinear ink writing recorder (Beckman Instruments, Inc.,Fullertoni, Calif.).
In each animal, the recording catheter was passed throughthe mouth into the stomach. The catheter was withdrawnwhile recording pressure at 0.5-cm intervals for a 1.0-minperiod. Upon completion of the pull through, the recordingtube was anchored with the middle orifice in the LES at thepoint of maximum recorded pressure. After the pull through,the tube was secured at the jaw with tape, and within theabdomen with small arterial clips fixed at the distal esoph-agus and most proximal stomach. This arrangement preventedaxial movement of the LES in relation to the tube orifice.This was verified using a separate tube with axial openingsat 1.0-cm intervals. The LES high pressure remained at thedesired catheter opening. Also, manual orad movement of
the esophagus did not displace the LES from the recordingorifice.
LES pressure was recorded as millimeters of mercury, withgastric pressure used as a zero reference; the values wereobtained as the midrespiratory value. The results were ex-pressed as a percentage of change in the sphincter pressurefor the sphincter relaxation, and as absolute values for theexcitatory motor responses. The percentage by which LESpressure decreased during swallowing was calculated as rest-ing LES pressure minus the nadir of LES pressure duringswallowing, divided by resting LES pressure times 100. Thestomach was intermittently aspirated to prevent gastricdistension.
After recording the basal LES pressure, the right or leftcervical sympathetic trunk was sectioned at a site 1.0 cm proxi-mal to the superior cervical ganglion. LES pressure was con-tinuously recorded during all procedures. After 30 min, theother cervical sympathetic trunk was sectioned. A similar pro-cedure was used for the splanchnic nerve studies. The greatersplanchnic nerves were sectioned at a site 1.0 cm proximalto the celiac ganglion.
Electrical stimulation of the cervical sympathetic trunk, thecervical vagi, or the greater splanchnic nerves, was performedwith a platinum wire bipolar electrode (0.04 mmdiameter)which was mounted in polyvinyl tubing (11.0 mmlength,5.0 mmdiameter) and was sealed with bonded adhesive.The distance between stimulating points was 5.0 mm. Thedistal or proximal ends of the nerves were placed into thetrough-shaped electrode, covered with cotton, and saturatedwith mineral oil. Care was taken to prevent current leakageby elevating the nerve from adjacent tissues with cottonpledgets. An electrical stimulus was delivered with a Grassstimulator (Model 88, with stimulus isolation unit SIU 5, GrassInstrument Co., Quincy, Mass.). Square wave pulses of 1.0-msstimulus duration, were delivered at a frequency range of0.50-50 Hz and a voltage range of 5 or 10 V. The lengthof the stimulus train was 5 s. The trains were separated byintervals of at least 5 min. A special procedure was carriedout for the simultaneous stimulation of the cervical vagal trunkand the sympathetic nerves. The length of the stimulus trainwas 15 s, either for the cervical sympathetic trunk or the greatersplanchnic nerves; simultaneous vagal stimulation was per-formed 10 s after the onset of sympathetic stimulation. Re-sponses to vagal or splanchnic nerve stimulation began withthe onset of nerve stimulation and persisted for the durationof stimulation, returning to basal values upon the terminationof the stimulus.
Drugs were administered through an intravenous catheterinto the femoral vein. The agents were given as single intra-venous boluses over 60 s or by continuous infusion. Normalsaline was used to flush the catheter after each drug adminis-tration. The LES response to splanchnic nerve stimulationwas measured in the same animals before and after administra-tion of the different antagonists. A period of 60 min for stabili-zation to base-line levels was observed after the administrationof each pharmacological agent. 1.5 mg/kg phentolamine(Regitine, CIBA-Geigy Corporation, Summit, N. J.) waschosen as the dose that antagonized the maximal LES responseto phenylephrine at 24.0 jig/kg. 1.0 mg/kg propranolol (In-deral, Ayerst Laboratories, New York) was given as the dosethat antagonized the maximal LES response to 40 ,ug/kg iso-proterenol. 30.0 ,ug/kg atropine sulfate (The Vitarine Co., Inc.,New York) was selected as the dosage that antagonized com-pletely the maximal LES response to 5.0 mg bethanechol.Additional studies with 10.0 and 100.0 ,Ag/kg atropine wereperformed. Trimethaphan camsylate (Arfonad, Roche Labora-tories, Div. Hoffmann-LaRoche Inc., Nutley, N. J.), 4.0 mg/min,was given at a dosage that inhibited the maximal blood pres-
Sympathetic Control of Lotwer Esophageal Sphincter Function 563

sure response during splanchnic nerve stimulation or the LESresponse to nicotine sulfate (50.0 ,ug/kg). 5.0-10.0 mg/kgguanethidine (Ismelinf, CIBA-Geigy Corporation) was givenat dosages previously reported to be effective in loweringblood pressure and blocking adrenergic neural function.Guanethidine effect was evaluated at 10-60 min after itsadminstration. Alone, guanethidine gave significant reductionin blood pressure in all animals.
To eliminate the effect of preganglionic splanchnic nervestimulation upon the adrenal glands, a group of animals werealso studied after bilateral adrenalectomy.
Statistical analysis was performed with the paired t testfor observations made in a minimum of seven animals. Allrecords were read in a blinded fashion without knowledgeof the nerve stimulation or of the drug being employed.
RESULTS
Basal LES pressure in response to cervical or greatersplanchnic nerve section. Cervical sympathetic sec-tion (unilateral or bilateral) at a site proximal to thecervical ganglion failed to significantly alter LES pres-sure. Greater splanchnic nerve section (unilateral orbilateral) at a site proximal to the celiac ganglion alsodid not significantly change the basal LES pressure.The control LES pressure was 15.66± 1.41 mmHg(mean±SEM) as compared with 16.02±1.28 mmHgafter bilateral cervical sympathectomy (P > 0.05) and18.00±2.10 mmHg after bilateral splanchnic nervesection (P > 0.05). LES pressures were similar witheither pentobarbital or ketamine.
Cervical vagotomy (unilateral or bilateral) also failedto alter the basal LES tone. The LES pressure in controlanimals was 15.54+2.06 mmHg as compared with15.37±2.15 mmHg after unilateral cervical vagotomy(P > 0.05), and 15.64±2.31 mmHg after bilateral cer-vical vagotomy (P±0.05). The basal LES pressure wasalso obtained in animals undergoing bilateral cervicalvagotomy with either unilateral or bilateral cervicalsympathetic or greater splanchnic nerve sectioning.Vagotomy plus sympathectomy also did not lead tosignificant alterations in basal LES tone (P > 0.05).
Effect of peripheral cervical sympathetic nerve stim-ulation on LES pressure, vagal-induced LES relaxationand blood pressure. Fig. 1 shows the percent de-crease in LES pressure (above) and the change in meanblood pressure in mmHg (below) for a full frequencyrange at 5 V (left) and 10 V (right). Cervical sympatheticnerve stimulation did not alter LES pressure at either5 or 10 V over the entire frequency range. Cervicalsympathetic nerve stimulation did give a small increasein mean arterial blood pressure. Cervical vagal stimu-lation gave a decrease in LES pressure and mean bloodpressure. Simultaneous stimulation of the cervical vagusand cervical sympathetic trunks did not alter eitherthe percentage decrease in LES pressure or the meandecrease in arterial blood pressure at either voltageover the full frequency range.
Effect of stimulation of the central portion of the
100
80(I)
-J
n)cx
0
50
20
0
0w +20
4Al(L + 10
E - 10
E -20
*-3 0
o,
I.- I (>0-
?,1. ...4... ........ ...... //......
-4 *v--o V's
4. s
, 1 0 30' 50 1510 20 30 50
Frequency Of Stimulation (Hz)(5 Volts) (10 Volts)
FIGURE 1 Effect of electrical stimulation of the distal end ofthe sectioned cervical sympathetic trunks on LES pressure(above), and on mean blood pressure (below). Sympatheticstimulation(s) produced no effect on sphincter pressure overthe entire frequency range at 5 and 10 V; it induced aslight increase in mean blood pressure. Peripheral vagal stim-ulation (V) induced a frequency-related sphincter relaxationand a decrease in blood pressure. Simultaneous stimulationof the sympathetic nerve (V + S) did not modify sphincterrelaxation and blood pressure responses as induced by vagalstimulation.
cervical sympathetic or splanchnic nerves on LES pres-sure; action of antagonist agents. Fig. 2 shows theeffect of electrical stimulation of the central end ofthe cut cervical sympathttic or greater splanchnicnerves. Data is expressed as a percent decrease in LES
6L
a-(nw-J
c
0)
0
°OOr80
60
40
20
*CONTROL(0-4*)BILATERAL CERVICAL VAGOTOMY(0 ...0 SPLANCHNIC
d;rr 7 - PROPRANOLOL(Img/kg) (--*4
Y 0 -0 CERVICAL SYMPATHETIC (0-O)
1 5 10 20 30 50Frequency Of Stimulation (H!)
FIGURE 2 Effect of electrical stimulation of the central end ofthe sectioned cervical sympathetic or splanchnic nerve onLES pressure. Electrical stimulation at 10 V was applied overan entire frequency range. Stimulation of the central end ofthe cut cervical sympathetic nerve gave no change in sphincterpressure. Stimulation of the central end of the cut splanchnicnerve produced a decrease in sphincter pressure. Bilateralcervical vagotomy or propranolol did not modify this response.
564 J. Fournet, W. J. S nape, Jr., and S. Cohen
p........&.......*... // ...0
* 0
pressure at 10 V over the full frequency range. Centralcervical sympathetic nerve stimulation did not alterLES pressure at any point. Stimulation of the centralportion of the greater splanchnic nerve gave a decreasein LES pressure; maximum inhibition of 69.1±16.0%was noted at a frequency of 10 Hz.
The inhibition in LES pressure noted during centralstimulation of the cut splanchnic nerve was next stud-ied during bilateral cervical vagotomy, or after theadministration of propranolol (1.0 mg/kg, i.v.). Thisdose of propranolol was selected as the lowest dosewhich produced complete antagonism of the maximuminhibitory effect of isoproterenol (4.0 /Lg/kg, i.v.) onLES pressure. Change in LES pressure was similar atall frequencies (P > 0.05) after either vagotomy orpropranolol.
In additional studies, not shown here, it was demon-strated that bilateral cervical sympathectomy or cuttingof the contralateral splanchnic nerve also failed to alterthe response to central splanchnic nerve stimulation.
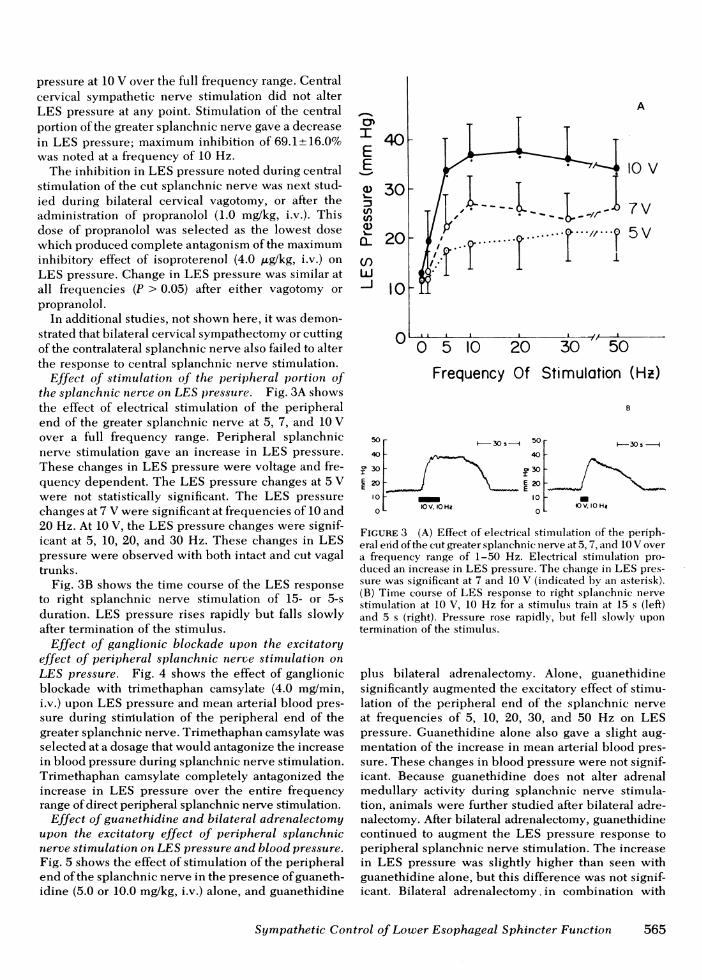
Effect of stimulation of the peripheral portion ofthe splanchnic nerve on LES pressure. Fig. 3A showsthe effect of electrical stimulation of the peripheralend of the greater splanchnic nerve at 5, 7, and 10 Vover a full frequency range. Peripheral splanchnicnerve stimulation gave an increase in LES pressure.These changes in LES pressure were voltage and fre-quency dependent. The LES pressure changes at 5 Vwere not statistically significant. The LES pressurechanges at 7 V were significant at frequencies of 10 and20 Hz. At 10 V, the LES pressure changes were signif-icant at 5, 10, 20, and 30 Hz. These changes in LESpressure were observed with both intact and cut vagaltrunks.
Fig. 3B shows the time course of the LES responseto right splanchnic nerve stimulation of 15- or 5-sduration. LES pressure rises rapidly but falls slowlyafter termination of the stimulus.
Effect of ganglionic blockade upon the excitatoryeffect of peripheral splanchnic nerve stimulation onLES pressure. Fig. 4 shows the effect of ganglionicblockade with trimethaphan camsylate (4.0 mg/min,i.v.) upon LES pressure and mean arterial blood pres-sure during stiniulation of the peripheral end of thegreater splanchnic nerve. Trimethaphan camsylate wasselected at a dosage that would antagonize the increasein blood pressure during splanchnic nerve stimulation.Trimethaphan camsylate completely antagonized theincrease in LES pressure over the entire frequencyrange of direct peripheral splanchnic nerve stimulation.
Effect of guanethidine and bilateral adrenalectomyupon the excitatory effect of peripheral splanchnicnerve stimulation on LES pressure and blood pressure.Fig. 5 shows the effect of stimulation of the peripheralend of the splanchnic nerve in the presence of guaneth-idine (5.0 or 10.0 mg/kg, i.v.) alone, and guanethidine
I
EE
0)
-j
tn
a)CL
Ji
40F
A
A410v
.--- ,,,- 7V
..11-1-,,---. 5 V
30 -
20F
1o0
0 5 10 20 30 50
Frequency Of Stimulation (Hz)
B
50- i30s--- 50 30
401- 40-
E 20-J E 2o -E E10 100 10V. 0 Hi
0IOV. 10 Hz
FIGURE 3 (A) Effect of electrical stimulation of the periph-eral eyid of the cut greater splanchnic inerve at 5, 7, aind 10 V overa frequency range of 1-50 Hz. Electrical stimulation pro-duced an increase in LES pressure. The change in LES pres-sure was significant at 7 and 10 V (indicated by an asterisk).(B) Time course of LES response to right splanchnic nervestimulation at 10 V, 10 Hz for a stimulus train at 15 s (left)and 5 s (right). Pressure rose rapidly, but fell slowly upontermination of the stimulus.
plus bilateral adrenalectomy. Alone, guanethidinesignificantly augmented the excitatory effect of stimu-lation of the peripheral end of the splanchnic nerveat frequencies of 5, 10, 20, 30, and 50 Hz on LESpressure. Guanethidine alone also gave a slight aug-mentation of the increase in mean arterial blood pres-sure. These changes in blood pressure were not signif-icant. Because guanethidine does not alter adrenalmedullary activity during splanchnic nerve stimula-tion, animals were further studied after bilateral adre-nalectomy. After bilateral adrenalectomy, guanethidinecontinued to augment the LES pressure response toperipheral splanchnic nerve stimulation. The increasein LES pressure was slightly higher than seen withguanethidine alone, but this difference was not signif-icant. Bilateral adrenalectomy in combination with
Sympathetic Control of Lower Esophageal Sphincter Function
OL
565

E 40E~~30-
CONTROL (@_)TRIMETHAPHANCAMSYLATE(4mg/min) (O--O)
L20 -
w
I-J 0
~40[8EEc~20 S-
a -O-0_ 0_ -0-4 4O--V/--OX 0 5 10 20 30 50
Frequency Of Stimulation (Hz)FIGURE 4 Effect of trimethaphan camsylate on the frequencyresponse curve of peripheral splanchnic nerve stimulationupon LES pressure (above), and mean blood pressure (below),at 10 V. Trimethaphan camsylate completely suppressed theexcitatory effect on both the sphincteric and blood pressureresponses.
-
80E
60-
40-
cn
20-l
0
E 40°I 0' 20
*01&10
E
* GUANETHIDINE+BILATERAL ADRENALECTOMY(0 0)
P ti GUANETHIDINE (5 nd 0mOg/kg) ( 0)
P CONTROL (0-10
*P' 0.05
.. I - -GUANETHIDINE (5 and 10mg/kg)
J~A-----i-- -4 CONTROL(O_)
GUANETHIDINE (5,ond Omg/kg)+- qF- -- - BILATERAL ADNIENALECTOMY(0--O)
5 10 20 30 50Frequency Of Stimulation (Hz)
FIGURE 5 Effect of guanethidine on the frequency responsecurve of peripheral splanchnic nerve stimulation upon LESpressure (above), and mean blood pressure (below), at 10 V.Guanethidine (5 and 10 mg/kg) increased the excitatory effectof splanchnic nerve stimulation on LES and blood pressure.Guanethidine (5 and 10 mg/kg) plus bilateral adrenalectomyabolished the change in blood pressure, but continued to aug-ment the LES pressure response to peripheral splanchnicnerve stimulation.
guanethidine completely abolished the changes inmean blood pressure during peripheral splanchnicnerve stimulation. Adrenalectomy alone reduced theblood pressure but not the LES response to splanchnicstimulation.
These observations suggested that the inhibition ofcatecholamine release by guanethidine augmented theincrease in LES pressure during peripheral splanchnicnerve stimulation. These changes in LES pressurecould be dissociated from the alterations in blood pres-sure noted during nerve stimulation.
Effect of alpha adrenergic antagonism with phentol-amine upon the excitatory effect of peripheral splanch-nic nerve stimulation on LES pressure and mean bloodpressure. Fig. 6 shows the effect of phentolamine (1.5mg/kg, i.v.) upon the excitatory effect of peripheralsplanchnic nerve stimulation on LES pressure andmean blood pressure. Phentolamine was selected atthe lowest dosage that would completely antagonizethe maximal excitatory action of phenylephrine (24.0p,g/kg, i.v.) on the LES. Phentolamine augmented theeffect on the LES response to peripheral splanchnicnerve stimulation at 10 V, 1 Hz. There was no effectat other stimulus parameters. Phentolamine did com-pletely antagonize the increase in mean blood pressurenoted during splanchnic nerve stimulation.
Thus, alpha adrenergic antagonism with phentol-amine, at a dosage sufficient to block the maximal ex-citatory action of phenylephrine on the LES and the
PHENTOLAMINE(1.5Img/kg)
CONTROL
CONTROL
l ----_ - _ -s ..--- -4----4 PHENTOLAMINE (1.5mg/kg)0 0.5 1 3 5 10 20 50
Frequency Of Stimulation (Hz)
FIGURE 6 Effect of phentolamine on the frequency responsecurve of peripheral splanchnic nerve stimulation upon LESpressure (above), and mean blood pressure (below), at 10 V.Phentolamine increased the LES pressure response to periph-eral splanchnic nerve stimulation at 1 Hz, but had no signif-icant effect at the other frequencies. Phentolamine didcompletely antagonize the increase in mean blood pressurenoted during splanchnic nerve stimulation.
566 J. Fournet, W. J. Snape, Jr., and S. Cohen
fX0 E 408oE
m ,c, V.
effect of splanchnic nerve stimulation on blood pres-sure, failed to alter the excitatory action of nervestimulation on LES pressure.
Effect of combined alpha and beta adrenergic an-tagonism upon the excitatory effect of peripheralsplanchnic nerve stimulation on LES pressure. Fig. 7shows the effect of propranolol (1.0 mg/kg, i.v.) alone,and propranolol plus phentolamine upon the excitatoryeffect of peripheral splanchnic nerve stimulation.Neither propranolol alone, nor propranolol plus phentol-amine significantly altered the LES response tosplanchnic nerve stimulation. Blood pressure re-sponses were inhibited during propranolol and phentol-amine administration, but not with propranolol alone.
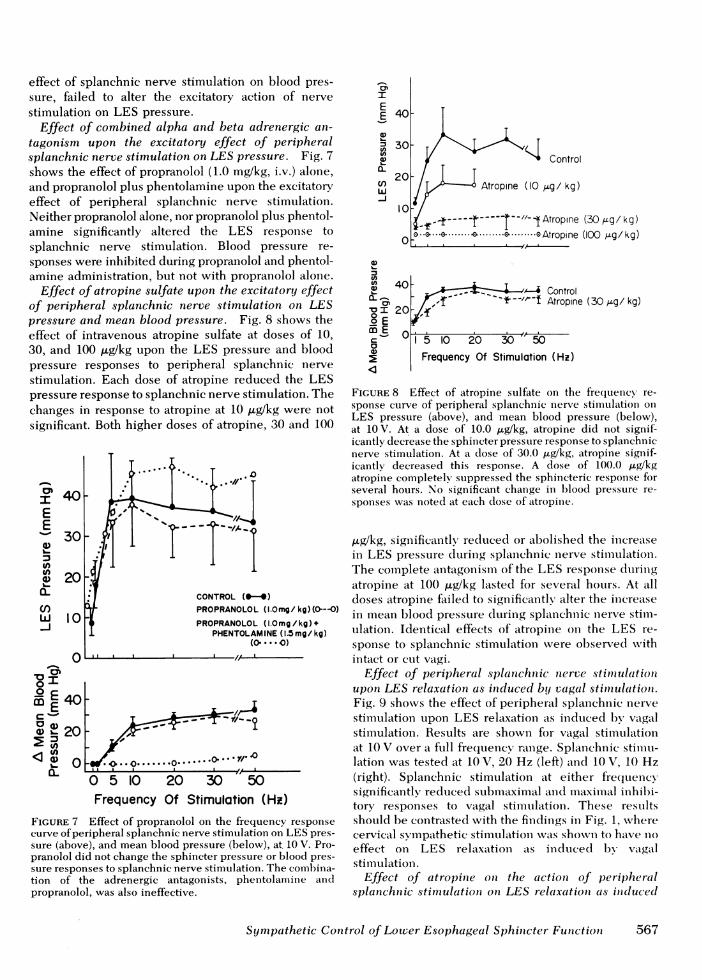
Effect of atropine sulfate upon the excitatory effectof peripheral splanchnic nerve stimulation on LESpressure and mean blood pressure. Fig. 8 shows theeffect of intravenous atropine sulfate at doses of 10,30, and 100 ,ug/kg upon the LES pressure and bloodpressure responses to peripheral splanchnic nervestimulation. Each dose of atropine reduced the LESpressure response to splanchnic nerve stimulation. Thechanges in response to atropine at 10 ,ug/kg were notsignificant. Both higher doses of atropine, 30 and 100
IEE1-
0~CL(nw
IJ
O.-
40rm E 40E
X0 0 20C 20
a- O. .o..... 0..o
I.. I , A-- .
0 5 10 20 30 50Frequency Of Stimulation (Hi)
FIGURE 7 Effect of propranolol on the frequency responsecurve of peripheral splanchnic nerve stimulation on LES pres-sure (above), and mean blood pressure (below), at. 10 V. Pro-pranolol did not change the sphincter pressure or blood pres-sure responses to splanchnic nerve stimulation. The comhibina-tion of the adrenergic antagonists, phentolamine andpropranolol, was also ineffective.
IEE 401-a)U) 30
1020
o Ea.
2 00-..
20
m00
a)
-J > Control
Atropine (10 9Lg/ kg)
f - ----- "-1Atropine (30 ,g/ kg).o..-.............-.o.--.oAtropine (100 4g/kg)
> Control-t--"- ~i Atropine (30 ,4g/ kg)
5 10 20 30 t0Frequency Of Stimulation (Hz)
FIGURE 8 Effect of atropine sulfate on the frequency re-sponse curve of peripheral splanchnic nerve stimnulation onLES pressure (above), and mean blood pressure (below),at 10 V. At a dose of 10.0 ,ug/kg, atropine did not signif-icantly decrease the sphincter pressure response to splanchnicnerve stimulation. At a dose of 30.0 ,gWkg, atropine signif-icantly decreased this response. A dose of 100.0 ,Lg/kgatropine completely! suppressed the sphincteric response forseveral hours. No significant change in hloocl pressure re-sponses was noted at each dose of atropine.
,ug/kg, significantly reclticed or abolished the inereatsein LES pressure dIurinig splaciehiuic nerve stimltulationi.The complete antagonismii of the LES response duringatropine at 100 Ag/kg lasted for several hours. At alldoses atropine failed to significantly alter the inereasein meean blood pressure during splanchnici nerve stimll-ulation. Identical effects of atropine on the LES re-sponse to splanchiuic stimulationv were observed withintact or cut vagi.
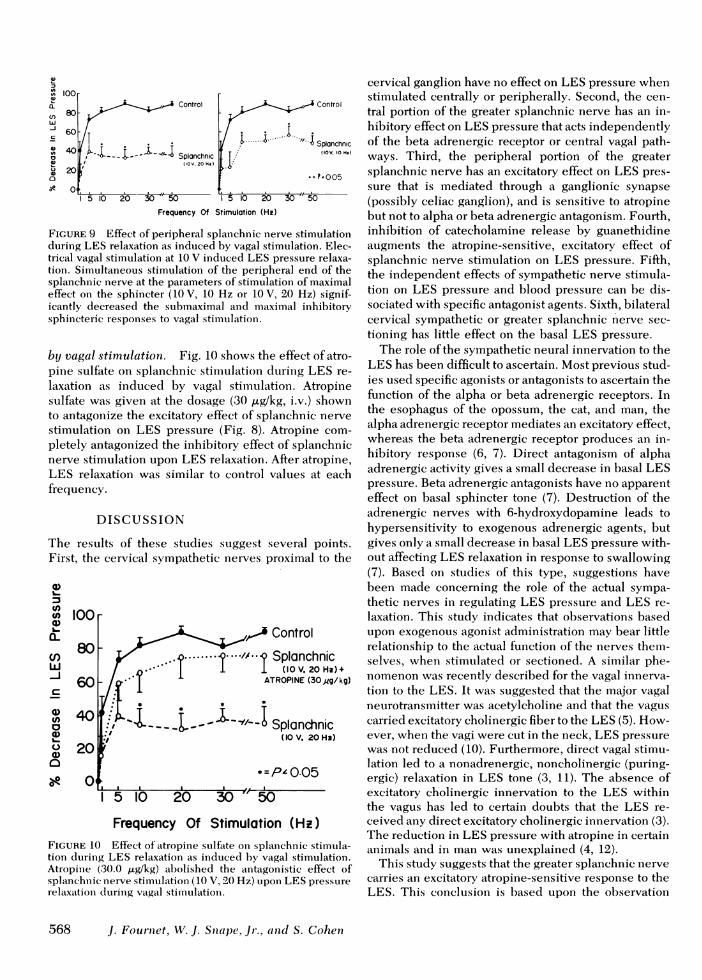
Effect of peripheral splanchn ic nerve stim ulationupon LES relaxation as induced by vagal stitmiulationi.Fig. 9 shows the effect of peripheral splanehiic nervestimulation upon LES relaxation as induced by vagalstimulation. Results are shown for vagal stimiiulationiat 10 V over a full frecquency range. Splanchiuic stiniu-lation was tested at 10 V, 20 Hz (left) aind 10 V, 10 Hz(right). Splanchnic stimulation at either frecquencysignificantly reduced subimcaximiail and milaximiail inhibi-tory responses to vagal stimulaltioni. These resuiltsshould be contrasted with the findinigs in Fig. 1, wherecervical symipathetic stimulaltion was showin to have n1oeffect on LES relaxation as iniduicedl by vatgalstimulation.
Effect of atropitne oni the actiotn of peripheralsplanchnic stimulation oni LES relaxation as iniducedc
Sympathetic Control of Lower Esophageal Sphincter Function 6567
a .-- i Control ,. Control80
-j 60-
.e. .i Splanchnic
40o 20 (IOV, 20 Hi
Q .= P'~~~~~~~~~~005__.___. _. _..
5 10 20 30 50 I5 10 20 3 ' 50Frequency Of Stimulation (Hz)
FIGURE 9 Effect of peripheral splanchnic nerve stimulationduring LES relaxation as induced by vagal stimulation. Elec-trical vagal stimulation at 10 V induced LES pressure relaxa-tion. Simultaneous stimulation of the peripheral end of thesplanchnic nerve at the parameters of stimulation of maximaleffect on the sphincter (10 V, 10 Hz or 10 V, 20 Hz) signif-icantly decreased the submaximal and maximal inhibitorysphincteric responses to vagal stimnulation.
by vagal stimulation. Fig. 10 shows the effect of atro-pine sulfate on splanchnic stinmulation during LES re-laxation as induced by vagal stimulation. Atropinesulfate was given at the dosage (30 Ag/kg, i.v.) shownto antagonize the excitatory effect of splanchnic nervestimulation on LES pressure (Fig. 8). Atropine com-pletely antagonized the inhibitory effect of splanchnicnerve stimulation upon LES relaxation. After atropine,LES relaxation was similar to control values at eachfrequency.
DISCUSSION
The results of these studies suggest several points.First, the cervical sympathetic nerves proximal to the
01)L.
U)U)lU)L._o
o)w
a)(I)
0)
a)0N
100
80
60
40
20
Frequency Of Stimulation (Hz)FIGURE 10 Effect of atropine sulfate on splanchnic stimula-tion during LES relaxationi as induced by vagal stimulation.Atropine (30.0 jig/kg) abolished the antagonistic effect ofsplanichniic nerve stimulation (10 V, 20 Hz) upon LES presssure
relaxationl durinlg vagal stimulationi.
cervical ganglion have no effect on LES pressure whenstimulated centrally or peripherally. Second, the cen-tral portion of the greater splanchnic nerve has an in-hibitory effect on LES pressure that acts independentlyof the beta adrenergic receptor or central vagal path-ways. Third, the peripheral portion of the greatersplanchnic nerve has an excitatory effect on LES pres-sure that is mediated through a ganglionic synapse(possibly celiac ganglion), and is sensitive to atropinebut not to alpha or beta adrenergic antagonism. Fourth,inhibition of catecholamine release by guanethidineaugments the atropine-sensitive, excitatory effect ofsplanchnic nerve stimulation on LES pressure. Fifth,the independent effects of sympathetic nerve stimula-tion on LES pressure and blood pressure can be dis-sociated with specific antagonist agents. Sixth, bilateralcervical sympathetic or greater splanchnic nerve sec-tioning has little effect on the basal LES pressure.
The role of the sympathetic neural innervation to theLES has been difficult to ascertain. Most previous stud-ies used specific agonists or antagonists to ascertain thefunction of the alpha or beta adrenergic receptors. Inthe esophagus of the opossum, the cat, and man, thealpha adrenergic receptor mediates an excitatory effect,whereas the beta adrenergic receptor produces an in-hibitory response (6, 7). Direct antagonism of alphaadrenergic activity gives a small decrease in basal LESpressure. Beta adrenergic antagonists have no apparenteffect on basal sphincter tone (7). Destruction of theadrenergic nerves with 6-hydroxydopamine leads tohypersensitivity to exogenous adrenergic agents, butgives only a small decrease in basal LES pressure with-out affecting LES relaxation in response to swallowing(7). Based on studies of this type, suggestions havebeen made concerning the role of the actual sympa-thetic nerves in regulating LES pressure and LES re-laxation. This study indicates that observations basedupon exogenous agonist administration may bear littlerelationship to the actual function of the nerves them-selves, when stimulated or sectioned. A similar phe-nomenon was recently described for the vagal innerva-tion to the LES. It was suggested that the major vagalneurotransmitter was acetylcholine and that the vaguscarried excitatory cholinergic fiber to the LES (5). How-ever, when the vagi were cut in the neck, LES pressurewas not reduced (10). Furthermore, direct vagal stimu-lation led to a nonadrenergic, noncholinergic (puring-ergic) relaxation in LES tone (3, 11). The absence ofexcitatory cholinergic innervation to the LES withinthe vagus has led to certain doubts that the LES re-ceived any direct excitatory cholinergic innervation (3).The reduction in LES pressure with atropine in certainanimals and in man was unexplained (4, 12).
This study suggests that the greater splanchnic nervecarries an excitatory atropine-sensitive response to theLES. This conclusion is based upon the observation
568 J. Fourtnet, W. J. Sniape, Jr., anid S. Cohen
(L
vl

that atropine sulfate; but not phentolamine, proprano-lol, or guanethidine; antagonizes the increase in LESpressure during the peripheral splanchnic nerve stimu-lation. It is uncertain whether the action of atropineis upon the muscarinic receptor in the muscle mem-brane, or whether atropine acts at the ganglion. It ismost likely that the action of atropine is at the musclemembrane.
Even at small doses, atropine suppressed the effectof splanchnic nerve stimulation for several hours. Atthose doses, atropine is a highly selective antagonistof muscarinic agents on smooth muscle (13). The actionof small doses of atropine makes the other possibleaction of this drug less tenable. Under appropriate ex-perimental conditions and in high dose, atropine isknown to have an alpha sympatholytic effect (14). Like-wise, atropine in high doses may antagonize the nico-tinic action of acetylcholine at the autonomic ganglion(13). On the other hand, muscarinic antagonism at aganglionic synapse does not usually abolish a responseas observed in these studies. It should be emphasizedthat ganglionic blockade with trimethaphan camsylateabolished the LES response to splanchnic stimulation,but this drug also antagonized the increase in bloodpressure as well. Atropine reduced the LES responsewithout altering blood pressure.
Cholinergic pathways in the sympathetic nerveshave been described in other studies with differentend organ responses (15-20). However, this is the ini-tial demonstration of an excitatory cholinergic responseobtained during sympathetic nerve stimulation. Usu-ally, mixed, cholinergic, and adrenergic responses aredescribed (18, 20). The neural pathway of this choliner-gic response is not clear; the simplest explanationswould be the existence of cholinergic fibers within thesplanchnic nerve, or some connection with the para-sympathetic nerves (15, 18). The excitatory action ofsplanchnic nerve stimulation on the LES was specificfor this nerve and was not observed during cervicalsymnpathetic nerve stimulation.
Guanethidine in the presence or absence of the ad-renal glands augmented the LES pressure response tosplanchnic nerve stimulation. This response was inde-pendent of the drug's effect on blood pressure. Theaction of guanethidine in augmnenting the excitatoryaction of peripheral splanchnic nerve stimulation wasunexpected. Guanethidine is known to cause adrener-gic neuroblockade, noradrenaline potentiation, andsympathomiimetic effects by release of noradrenaline(21, 22). Noradrenaline potentiation seems to be un-related to its action on adrenergic nerves (23). Theincrease in the sensitivity of tissues to circulating cate-cholamines was suggested by the increased response inblood pressure during splanchnic nerve stimulation.This response was suppressed after bilateral adrenalec-tomv. Guanethidine has no effect on the release of
catecholamines from the adrenal medulla (24). Adre-nalectomy did not modify the LES response to guaneth-idine during splanchnic nerve stimulation, thus sug-gesting that the effect on the LES was not a result ofpotentiation of the action of released catecholamines.A possible sympathomimetic effect of guanethidineseems unlikely as no change in resting LES pressurewas noted after drug injection. The action of guanethi-dine does not seem to be related to noradrenaline po-tentiation or a sympathomimetic effect, as its effectswere not modified by an alpha blocking agent, phentol-amine. It is possible that guanethidine augmented theLES response to splanchnic stimulation by the blockageof a simultaneous inhibitory adrenergic response elic-ited during splanchnic stimulation (25-27). However,the demonstration that effective alpha, but not beta,blockage only minimally augments the excitatory re-sponse to splanchnic nerve stimulation makes this pos-sible explanation less tenable. Therefore, the mecha-nism by which guanethidine augmented the LESresponse to splanchnic nerve stimulation remainsundefined.
The splanchnic nerves, when stimulated at the pe-ripheral end, also seemed capable of inhibiting LESrelaxation in response to direct vagal stimulation. Thisinhibitory response was not seen during cervical sym-pathetic nerve stimulation. The physiological role ofthe splanchnic nerves in altering the LES response tovagal stimulation is unclear. This study does not definethe significance of this response in health or in disease,but simply indicates the presence of a neural pathwaywithin the splanchnic nerves that can limit the abilityof the vagi to induce LES relaxation. This splanchnicresponse was sensitive to atropine but not to the otherneural antagonists. A similar reduction in LES relaxa-tion as induced by vagal stimulation has been shownduring intravenous dopamine infusion and during sub-cutaneous addition of bethanechol (28, 29). The presentstudy suggests that the magnitude of LES relaxationmay be reduced by splanchnic nerve stimulation aswell as by certain drugs.
Stimulation of the central end of the splanchnic nervegave a decrease in LES pressure. This response wasunchanged after bilateral cervical sympathetic or bi-lateral vagal sectioning in the neck. Additionally, directstimulation of the cervical sympathetic nerve also failedto elicit this response. The central splanchnic responsewas not antagonized by a seemingly effective dose of abeta adrenergic antagonist, propranolol. Also, sectionof the contralateral splanchnic nerve failed to abolishthis inhibitory response. These results would suggestthat the inhibitory response was transmitted by a non-adrenergic neurotransmitter that is carried to the LESthrough unknown pathways. It is possible that the cen-tral portion of the splanchnic nerve connects with in-hibitory pathways in the vagus nerves within the thorax.
Symnpathetic Cotntrol of Low;er Esophageal Sphincter Function 569

This mlay occur without connection to the brain stem.It has been shown that many small branches of thevagus within the chest can initiate LES relaxation whenstimulated directly (30).
In summary, this study emphasizes several imlportanitpoints conieerning the sympathetic iinnervation of theLES. First, studies dturinig direct nieuiral stimulationmlay give results (uite differenit thani would be pre-dicted from studies usinlg the exogenous administra-tion of agonist or antagonist comnpound(Is. Second, theLES does have an atropine-sensitive excitatory in-nervation that is carried in the greater splanchnic nerve.Third, the sympathetic innervatioin miay modulate theaction of direct vagal inihibitioIn on1 the LES. Fourth,although adrenergic inhibition with guanethidine aug-ments the excitatory, atropine-sensitive increase inLES pressure during splanchnic inerve stimulation, thetrue intrinsic role of the alpha anid beta adrenergic re-ceptors in the regulation of LES function remainsunclear.
ACKNOWVLEDGMENTS
The authors wish to thank Ms. Marv Carroll for secretarialassistance.
This work was supported by Research grant ROI AM19379from the National Institutes of Health.
REFERENCES
1. Rattan, S., and R. K. Goyal. 1974. Neural control of thelower esophageal sphincter. Influence of the vagusnerves.J. Clin. Invest. 54: 889-906.
2. Matarazzo, S. A., W. J. Snape, Jr., J. P. Ryan, and S. Cohen.1976. Relationship of cervical and abdominal vagal ac-tivity to lower esophageal sphincter function. Gastroen-terology. 71: 999-1003.
3. Goyal, R. K., and S. Rattan. 1975. Nature of the vagalinhibitory innervation to the lower esophageal sphincter.
J. Clin. Invest. 55: 1119-1126.4. Gonella, J., J. P. Niel, and C. Roman. 1977. Vagal control
of lower esophageal sphincter motility in the cat.J. Phys-iol. (Lond.). 273: 647-664.
5. Christensen, J. 1968. Pharmacology of the esophagus.Handb. Physiol. 4(Sect. 6): 2325-2330.
6. Christensen, J., and E. E. Daniel. 1968. Effects of someautonomic drugs on circular esophageal smooth muscle.
J. Pharmacol. Exp. Ther. 159: 243-247.7. DiMarino, A. J., and S. Cohen. 1973. The adrenergic con-
trol of lower esophageal sphincter function.J. Clin. Invest.52: 2264-2271.
8. Carlson, A. J., T. E. Boyd, and J. F. Pearcy. 1922. Theinnervation of the cardia and the lower end of the esoph-agus in mammals. Am. J. Physiol. 61: 14-4 1.
9. Ingelfinger, F. J. 1958. Esophageal motility. Physiol. Rev.38: 533-584.
10. Ryan, J. P., W. J. Snape, Jr., and S. Cohen. 1977. Effectof vagal cooling on esophageal peristalsis and sphincterrelaxation. AmJ. Physiol. 1: E159- E 165.
11. Tuch, A., and S. Cohen. 1973. Lower esophageal sphincterrelaxation: studies on the neurogenic inhibitory mecha-nism. J. Clin. Invest. 52: 14-20.
12. Lind, J. F., J. S. Crispin, and D. K. McIver. 1968. Theeffect of atropinie on the gastroesophageal sphincter. Catn.
J. Physiol. Pharmacol. 46: 233-238.13. Goodman, L. S., and A. Gilmani, editors. 1970. Drugs in-
hibiting the actioni of acetylcholinie on structure in-nervated by postganiglionic parasynmpathetic nerves. InThe Pharmacological Basis of Therapeutics 1970. Mac-millan & Co. Ltd., Londoni. 521.
14. Ludvena, F. B., and M. J. Braninl. 1966. Adrenolytic ac-tivity of atropinie (+) hyoscvaminie, atroscine, homatro-pine and related compounds. J. Pharm. Sci. 55: 280-284.
15. Kure, K., and NI. Fujii. 1933. Influenice of spinal para-sympathetic on 1lood vessels aindl external secretion ofpancreas. Q. J. Exp). Phiysiol. Cogii. Med. Sci. 22:323-328.
16. Kure, K., K. Kanida, A. WVakabavaski, anid S. Okinaka. 1933.Parasympathetic nerve supply of the kidney. Q. J. Exp.Physiol. Cogn. Med. Sci. 22: 315-321.
17. Dale, H. H., and XV. Feldberg. 1934. The chemical tranis-mission of secretory impulses to the sweat glandls of thecat. J. Physiol. (Lond.). 82: 121- 128.
18. Gillespie, J. S., and B. R. NMacKenna. 1961. The inhibitoryactioni of the sympathetic nerves on the smooth muscleof the rabbit gut, its reversal by reserpine andl restora-tion by catecholaminies ailcl by dopa. J. Physiol. (Lond.).156: 17-34.
19. Barlow, T. E., J. R. Greeniwell, A. A. Harper, andcl T.Scratchers. 1974. The influence of the splanchnic nerveson the external secretion, blood flow and electrical coni-ductance of the cat pancreas. J. Physiol. (Lond.). 236:421-433.
20. Persson, C. G. A. 1973. Dual effects on the sphincter ofOddi and gallbladder induced by stimiiulation of the rightgreat splanchnic nerve. Acta Physiol. Scand. 87: 334-343.
21. Goodman, L. S., and A. Gilman, editors. 1970. In ThePharmacologic Basis of Therapeutics 1970. Macmillan &Co. Ltd., London. 570.
22. Forster, R. W., D. S. Shah, and R. C. Small. 1978. A studyof the sympathomimetic action of guanethidine on theisolated anococcygeus muscle of the rat. Br.J. Pharmacol.62: 307-313.
23. Maxwell, R. A. 1965. Conicerninig the mode of action ofguanethidine and some derivatives in augmenting thevasomotor action of adrenergic amines in vascular tissuesof the rabbit. J. Pharmacol. Exp. Ther. 148: 320-328.
24. Abercrombie, G. F., and B. N. Davies. 1963. The action ofguanethidine with particular reference to the symiipatheticnervous system. Br. J. Pharmacol. 20: 171-177.
25. Paton, W. D. M., and E. S. Vizi. 1969. The inhibitoryaction of noradrenaline and adrenaline on acetylcholineoutput by guinea pig ileum longitudinal muscle strip.Br. J. Pharmacol. 35: 10-28.
26. De Groat, W. C., and W. R. Saum. 1972. Sympatheticinhibition of the urinary bladder and of pelvic ganglionictransmission in the cat.J. Physiol. (Lond.). 220: 297-314.
27. Gillespie, J. S., and M. A. Khoyi. 1975. A pharmacologicalanalysis of the inhibitory effects of the sympathetic nerveson the rabbit colon.J. Physiol. (Lond.). 244: 42p-43p.
28. Doody, P., S. Rattan, and R. K. Goyal. 1976. Adrenergicmodulation of the vagal inhibitory action: evidence fordopamine receptors. Gastroenterology. 70: 139. (Abstr.)
29. Fournet, J., W. J. Snape, Jr., and S. Cohen. 1978. Modula-tion of lower esophageal sphincter relaxation. Clin. Res.26: 318A. (Abstr.)
30. Kravitz, J. J., W. J. Snape, Jr., and S. Cohen. 1978. Effectof thoracic vagotomy and vagal stimulation on esophagealfunction. Am. J. Physiol. 234: E359-E364.
570 J. Fournet, W. J. Snape, Jr., and S. Cohen


















