
Structure of Ustilago maydis Killer Toxin KP6 α-Subunit A ...

THE JOURNAL OF U~OLOCICAL CHEMISTRY Vol. 248, No. 13,Issue of July 10, pp. 4584-4590, 1973
Printed in U.S.A.
Subunit Structure of Fragment D from
Fibrinogen and Cross-Linked Fibrin*
(Received for publication, September 6, 1972)
SALVATORE V. PIZZO,$ LLOYD M. TAYLOR, JR., MARTIN L. SCHWARTZ,$. ROBERT L. HILL, AND PATRICK A. MCKEE$
From the Department of Biochemistry, D&e University Medical Center, and the Department of Medicine, Vet- erans Administration Hospital, Durham, North Carolina %TiO
SUMMARY
The different species of Fragment D were purified from plasmin digests of either fibrinogen or highly cross-linked fibrin and examined electrophoretically in sodium dodecyl sulfate on polyacrylamide gels before and after reduction by ,&mercaptoethanol. It was found that Fragment D from cross-linked fibrin is approximately twice the size of that from fibrinogen, which indicates that the COOH-terminal cross-links in the y chain remnants are not cleaved during fibrin digestion by plasmin. The first Fragment D to appear in plasmin digests of fibrinogen is designated Fragment D1 which is composed of a /3” chain (44,500 molecular weight), a y’ chain (42,000 molecular weight), and an (Y” chain (-15,000 molecular weight) linked together by disulfide bonds. Fragment D from fibrin differs in that it has a y’ dimer (81,000 molecular weight) in place of y’ chains and that it contains two each of the o(’ and 0” chains. Further digestion of Fragment D, from fibrinogen results in cleavage of the y’ chain to give first the y” chain (37,000 molecular weight) and finally the 7”’ chain (27,000 molecular weight). Neither the y” chain nor the y”’ chain contained the COOH- terminal cross-links found in the intact y chain or the y’ chain.
It was possible to show by sodium dodecyl sulfate poly- acrylamide gel electrophoresis that fibrinogen Fragment D1 could be cross-linked in the presence or absence of native fibrinogen through the formation of y-y’ or y’- y’ dimers.
The digestion of human fibrinogen by plasmin results in five terminal digestion products identified as Fragments A, 13, C, D, and E (l-5). Recently tEe appearance of these fragments have been correlated with changes in the subunit structure of fibrin-
* This work was supported by research grants from the Na- tional Heart Institute (HE-06400) and National Institute of Neu- rological Diseases and Stroke (NS-06233), National Institutes of Health, and a Clinical Investigatorship from the Veterans Ad- ministration.
$ Predoctoral Fellow, Medical Scientist Training Program, National Institute of General Medical Sciences (GM-01678).
0 Clinical Investigator, Durham, Veterans ~ Administration Hospital.
ogen and fibrin during plasmin digestion (1, 2). Fragments A, B, and C are derived from the Q(A) chains of fibrinogen while both Fragments D and E contain polypeptide chains derived from each of the three subunit chains of fibrinogen. Fragment E is derived from the NH&erminal region of the molecule and contains three pairs of low molecular weight pept,ides held to- gether by disulfide bonds (1, 2). The three chains composing Fragment D are also held together by disulfide bonds, although fewer are probably present than in Fragment E since the NH2- terminal region is particularly rich in disulfide bonds (6). In Fragment D the subunit derived from the a(A) chains is des- ignated as the CX” chain and has a molecular weight of about 15,000; that derived from the P(B) chains is called the B” chain and has a molecular weight of 44,500. Finally, depending on the extent of proteolysis by plasmin, any one of three types of degraded y chain may be present in different species of Frag- ment D and designated as the y’, y”, and 7”’ chains with mo- lecular weights of 42,000, 37,000, and 27,000, respectively. The corresponding Fragment D species are called Fragments D1, 02, and Da, respectively.
Recent studies of fibrin digestion by plasmin (2) suggest that Fragment E from fibrinogen and Fragment E from cross-linked fibrin are identical in structure. However, Fragment D from fibrinogen and cross-linked fibrin differ since the latter con- tains intermolecular COOH-terminal cross-links between its y chains. The formation of these cross-links prevents cleavage in this region of the y dimers. We now wish to report additional data on the structures of Fragment D derived from digests of fibrinogen or highly cross-linked fibrin. Highly purified Frag- ment D from both sources was examined by sodium dodecyl sulfate polyacrylamide gel electrophoresis before and after reduc- tion. Moreover, the ability of various types of fibrinogen Frag- ment D to cross-link with itself or native fibrinogen was also examined. The results of these studies support models pre- viously presented for the digestion of fibrinogen and highly cross- linked fibrin (1, 2).
EXPERIMENTAL PROCEDURE
Reagents-Purified human fibrinogen, fibrin-stabilizing fac- tor, and plasminogen were prepared as previously described (1, 2, 7) or, as in the case of urokinase (Calbiochem), obtained from commercial sources. Purified human thrombin was kindly supplied by Dr. D. L. Aronson, Division of Niological Standards, National Institutes of Health.
4584
by guest on June 22, 2018http://w
ww
.jbc.org/D
ownloaded from

4585
Yur$icatian of Pibrinogerb and Fibrin Degradation Products- Fibrinogen as well as fibrin degradation products were purified by a modification of the method reported by Nussenzweig et al. (4) for the purification of fibrinogen degradation products. A lOO-mg sample of fibrinogen was dissolved in 20 ml of 0.2 M so-
dium chloride following which 0.1 ml of a 6 mg per ml (20 CTA units per A.& plasmiuogen solut’ion (2) and 0.4 ml of a 460 Ploug unit per ml urokinase solution (2) were added. The sample was digested at 37” for 18 hours and then soybean trypsin in- hibitor was added to give a final concent,ration of 2 mg per ml. Cross-linked fibrin was prepared by the addition of 25 units of human thrombin (2) and 0.75 ml of 0.8 M calcium chloride to 20 ml of a fibrinogen solution containing 5 mg per ml. The mixture was allowed to clot for 3 hours which was sufficient time for the formation of both y dimers and o( polymers (8). The plasminogen and urokinase solutions were then added to the cross-linked fibrin clot which was broken up with a glass stirring rod. At the end of 18 hours of digestion, soybean trypsin in- hibitor was added to the digest.
The fibrinogen or fibrin digests were then dialyzed for 24 hours at 4” against three changes of 100 volumes of 0.01 M so-
dium carbonate, pH 8.9. At the end of this time the pH and conductivity of the solution were identical with the sodium car- bonate buffer. Purification of the fibrinogen and fibrin digest products was achieved by elution from a DEAE-cellulose col- umn, 50 x 1.5 cm, equilibrated with 0.01 M sodium carbonate, pH 8.9. Following the application of the sample, a linear gra- dient of 0 to 0.2 M sodium chloride in 0.01 M sodium carbonate buffer, pH 8.9, was used to develop the column. After collecting about 800 ml, the gradient was discontinued since no more pro- tein appeared in the effluent, and a final wash of 0.5 M sodium chloride in 0.01 M sodium carbonate, pH 8.9, was used to remove Fragment E from the column. Protein peaks were pooled as indicated, dialyzed for 24 hours at 4” against three changes of 40 volumes of distilled water, and then lyophilized. SDS’- polyacrylamide gel eleclrophoresis was used to identify the digestion products (l-5).
Since it was found that the Fragment D obtained from the l&hour fibrinogen digest already showed extensive degradation of the y chains, the purification of fibrinogen Fragment D was subsequently further modified. The digestion of fibrinogen with plasmin was allowed to proceed for only 2 hours and then soybean trypsin inhibitor was added to a final concentration of 0.3 mg per ml. This digest mixture was also purified by gra- dient elution from a DEAE-cellulose column, 50 X 1.5 cm, except that the linear gradient was from 0.04 t,o 0.4 M sodium chloride in 0.01 RI sodium carbonate, pH 8.9. No attempt was made to recover Fragment E in this experiment. Samples were pooled as indicated, dialyzed, and lyophilized as described above, fol- lowing which SDS-gel electrophoresis was again used to identify the products. Moreover, standard electrophoresis at pH 3.2 in the presence of urea (9) was used t)o examine these peaks.
Fibrinogen Fragment D which was found mainly in Peak 4 was further purified by gel filtration as follows. A 25-mg sam- ple was dissolved in 10 ml of 0.2 M sodium phosphate, pH 7.2, and applied to a Sephadex G-100 column, 100 X 1.5 cm, equil- ibrated in the same buffer. The first peak was pooled, dialyzed against distilled water, and lyophilized as described above; a final yield of 15 mg was obtained. SDS-gel electrophoresis was used to identify this fraction as Fragment D.
Immunocli$usion Studies-The above preparation of fibrinogen
The abbreviation used is: SDS, sodium dodecyl sulfate.
Fragment D was also compared with the purified fibrin Frap- ment D by microimmunodiffusion in 1% agarose (lo), using antisera against human fibrinogen or fibrinogen Fragment D. Both antisera were prepared and kindly supplied to us by Dr. J. Gormsen, Sundby Hospital, Copenhagen, Denmark.
Cross-Linking of Fibrinogen Fragment D with Fibrin-Purified fibrinogen (1, 2) and Fragment D1 (Fig. 3) were each made up to a concentration of 10 mg per ml in 0.05 M Tris-HCl buffer, pH 7.4, containing 0.15 M sodium chloride. Purified fibrin- stabilizing factor (9) was dissolved in 0.5 M sodium citrate-O.16 M
potassium chloride, pH 7.1, to give an absorbance at 280 nm of 0.5 per ml. A,series of mixtures of native fibrinogen and Frag- ment D1 were prepared as shown in Table I. In each case 5 ,ul of human thrombin solution, 25 NIH units per ml (2), 5 ~1 of 0.8 M calcium chloride, and 5 ~1 of fibrin-stabilizing factor solu- tion were added and the samples were then incubated at room temperature for 4 hours. Visible clot formation occurred in all tubes except that containing Fragment D1 only; however, at a Fragment D1 to fibrinogen molar ratio of 40:1, the clot was extremely small and fragmented. Thrombin and fibrin-sta- bilizing factor activities were stopped in each reaction mixture by the addition of 5 ~1 of 0.5 M EDTA, pH 7.1, and 0.9 ml of 2% SDS-5 M urea-:!% /3-mercaptoethanol. Following overnight incubation at room temperature, these samples were then ex- amined by SCS-gel electrophoresis.
SDS-Gel Electrophoresis-Polyacrylamide gel electrophoresis in SDS was performed as previously described (1, 2). In all cases 7.5% polyacrylamide gels were used.
EXPERIMENTAL RESULTS
Purification of Fragment D from Fibrinogen and Fibrin--Fig. 1A shows the purification of the fragments from fibrinogen digests by DEAE-cellulose chromatography. As previously reported by Nussenzweig et al. (4), Peak 1 contains Fragment A; Peak d, Fragments B and C; Peak S, Fragment D ; and Peak 5, Fragment E. In the present study an additional peak, labe!ed 4, is soybean trypsin inhibitor which is present in a n,n~ch higher concentration than criginally used (4). Fig. 1B shows the puri- fication of the fragments from a digest of fibrin by the same method. Peaks 1 and 2 which contained Fragments A, B, and C in the case of fibrinogen digests are not observed. Otherwise, the patterns are very similar except that fibrin Fragment D (Peak S) eluted at a higher salt concentration. Fig. 2 shows the gel electrophoretic patterns of fibrinogen Fragment D and fibrin Fragment D before and after reduction. As can be seen the mobility cf unreduced fibrinogen Fragment D on Gel a is sig- nificantly greater than that of unreduced fibrin Fragment D on Gel b. Moreover, as shown on Gel c, the reduced subunit pat- terns differ in that the y’ chains of fibrinogen Fragment D1 have undergone extensive degradation to give y” chain of 37,000 molecular weight and 7”’ chain of 27,000 molecular weight, respectively. On the other hand, Gel d shows that reduced fibrin Fragment D contains an 81,000 molecular weight degrada- tion product of the y dimer (2). Even in prolonged digests of fibrin, only the one species of fibrin Fragment D (Fig. 4) was produced.
The identity of Peak 4 as soybean trypsin inhibitor was es- tablished by comparison to pure soybean trypsin inhibitor sub- jected to electrophoresis under the same conditions. The identification of Peak 5 as Fragment E was determined by com- parison to the Fragment E seen on electrophoresis of terminal fibrinogen and fibrin digests (1, 2) and by comparison to purified Fragment E prepared by a different method (2).
by guest on June 22, 2018http://w
ww
.jbc.org/D
ownloaded from

4586
-0
-0
-C
-C
-c
-c
1.5
1.3 G s
I.1 g !I
5
P
j.5
I.3
).I
ELUTION VOLUME (ml)
FIG. 1. A, DEAE-cellulose gradient elution of fibrinogen di- gestion fragments. The digestion proceeded for 18 hours and was stopped by the addition of soybean trypsin inhibitor. Frac- tions were pooled as indicated by the horizontal bars. B, DEAE- cellulose gradient elution profile for the purification of highly cross-linked fibrin digestion products. The fibrin formed for 3 hours before urokinaie was added to the clot. The digestion conditions were then identical with those described for fibrinogen.
Fibrinogen Fragment D appeared to consist of several species which had slightly different mobilities on SDS-gel electrophoresis. The lower molecular weight species were formed as a function of digestion time and were arbitrarily designated as D1, Dz, DI, and Dq according to their order of appearance. Because the fibrinogen Fragment D species isolated in the above experi- ment consisted mainly of DP and DD, the method was modif?ed by allowing plasmin to digest fibrinogen for only 2 hours before the addition of soybean trypsin inhibitor and subsequent purifi- cation by DEAE-cellulose chromatography. It should also be noted that the concentration of soybean trypsin inhibitor was reduced 7-fold since the higher concentration was found un- necessary. Fig. 3 shows the chromatographic pattern for this preparation. Once again Peaks 1 and 2 contain Fragments A, and B and C, respectively. The soybean trypsin inhibitor was present in Peak 6. As can be seen by examining the electro- phoretic patterns shown in Fig. 4, Fragment D species were found in Peaks S, 4, and 5. The species present in Peak 3 was
the parent Fragment D1; Peak 4 contained primarily Fragment D1 as well as a small amount of Fragments Dt and Dt; Peak 5 contained mainly Fragments Dz and DP. In addition to SDS- gel electrophoretic analysis, acrylamide gel electrophoresis at pH 3.2 in urea (9) was used to examine the fibrinogen Fragment D species contained in Peaks S, 4, and 5. As can be seen on the middle three gels in Fig. 4, at least five fibrinogen Fragment D species are resolved on the basis of charge by this method. The gel pattern for Peak S, which essentially consists of Fragment D1, reveals two major and three minor bands. The gel for Peak 4, which contains primarily Fragment D1 and much smaller
FIG. 2. SDS-gel electrophoretic patterns for fibrinogen and fibrin Fragment D species prepared as shown in Fig. 1. Gel a shows the electrophoretic pattern of fibrinogen Fragment D spe- cies. This preparation shows little Fragment DI but a large amount of Fragments Dz and Da. Gel b shows the pattern of fibrin Fragment D. The mobility of this sample is considerably less than the mobilities of any of the fibrinogen Fragment D spe- cies seen on Gel a. Gel c shows the reduced electrophoretic pat- tern of the same fibrinogen Fragment D species shown on Gel a. Gel d shows the reduced electrophoretic pattern of fibrin Frag- ment D.
ELUTKW) VOLUME
FIG. 3. DEAE-cellulose gradient elution of fibrinogen diges- tion fragments. The digestion proceeded for only 2 hours be- fore adding soybean trypsin inhibitor. The column was eluted by a linear gradient of 0.04 to 0.4 M sodium carbonate buffer, pH 8.9. No attempt was made to recover Fragment E in this study. Fractions were pooled as indicated in the figure.
amounts of Fragments DQ and D1, shows the same two major bands and an increased amount of three minor bands. Thus, the two major bands observed by this electrophoretic method must correspond to Fragment D1 while the three minor bands apparently represent Fragments Dz and Da.
As shown in Fig. 5, Peak 4, which contained the largest amount of the parent fibrinogen Fragment D1 species, was subsequently rechromatographed on Sephadex G-100 to remove small amounts of low molecular weight contaminants. The subunit structure of purified fibrinogen Fragment D1, which is known to be com- posed of an CX” chain, a /3” chain, and the VI-chain, all held to- gether by disulfide bonds (l), is shown on Gel a in Fig. 4. The subunit structures of Fragments Dz and Da are shown on Gels c and d, in Fig. 2, and in place of the y’ chain of 42,000 molecular weight as found in Fragment D1, the y” chain of 37,000 mo- lecular weight is found in Fragment Dz, and the 7”’ chain of 27,000 molecular weight is present in Fragment Da. Finally, it should be noted that Gel b of Fig. 4 shows that the /3” chain of 44,500 molecular weight is the only subunit of any of the Frag-
by guest on June 22, 2018http://w
ww
.jbc.org/D
ownloaded from
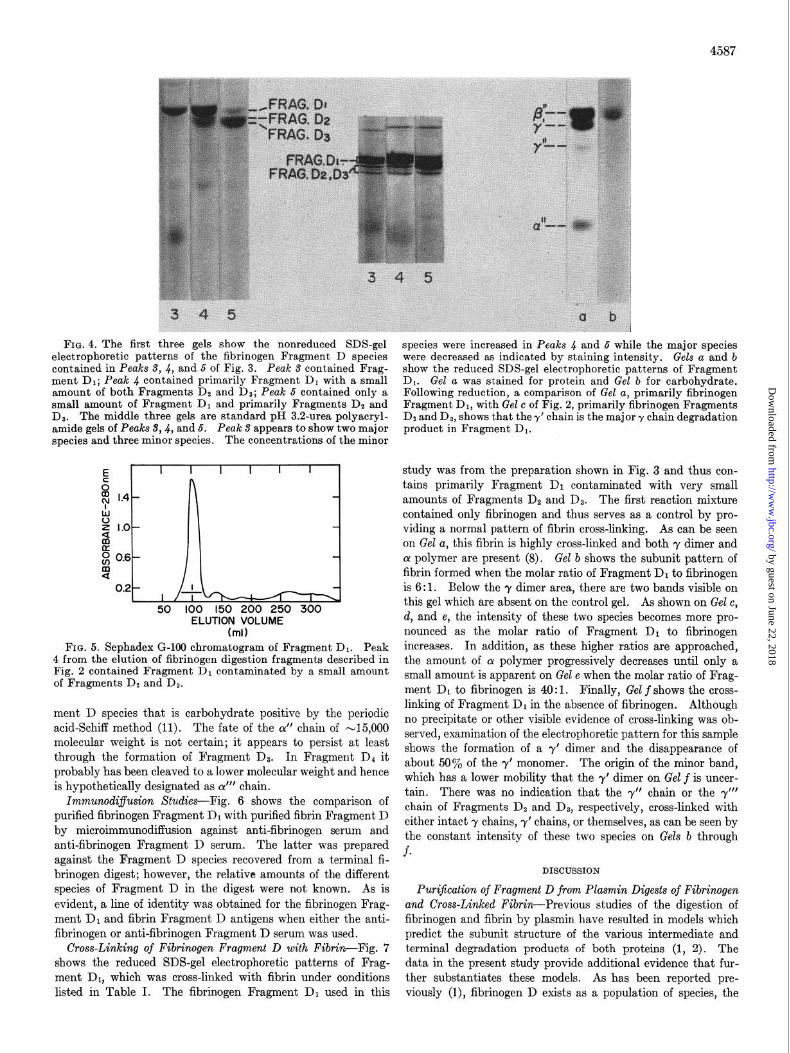
4587
FIG. 4. The first three gels show the nonreduced SDS-gel electrophoretic patterns of the fibrinogen Fragment D species contained in Peaks S, 4, and 5 of Fig. 3. Peak S contained Frag- ment Dr; Peak .J contained primarily Fragment DI with a small amount of both Fragments De and I).?: Peak 5 contained onlv a small amount of Fr&gment Dr and primarily Fragments DI and Da. The middle three gels are standard pl!f 3.2~urea polyacryl- amide gels of Peaks $4, and 5. Peak 3 appears to show two major species and three minor species. The concentrations of the minor
w I I I I
ig[ lJIj 50 100 150 200 250 300
ELUTION VOLUME (ml)
FIG. 5. Sephadex G-100 chromatogram of Fragment D1. Peak 4 from the elution of fibrinogen digestion fragments described in Fig. 2 contained Fragment D, contaminated by a small amount of Fragments DZ and Da.
ment D species that is carbohydrate positive by the periodic acid-Schiff method (11). The fate of the CI” chain of -15,000 molecular weight is not certain; it appears to persist at least through the formation of Fragment DI. In Fragment II4 it probably has been cleaved to a lower molecular weight and hence is hypothetically designated as CY”’ chain.
Immunocli$usion Studies-Fig. 6 shows the comparison of purified fibrinogen Fragment Dr with purified fibrin Fragment D by microimmunodiffusion against anti-fibrinogen serum and anti-fibrinogen Fragment D serum. The latter was prepared against the Fragment D species recovered from a terminal fi- brinogen digest; however, the relative amounts of the different species of Fragment D in the digest were not known. As is evident, a line of identity was obtained for the fibrinogen Frag- ment Di and fibrin Fragment D antigens when either the anti- fibrinogen or anti-fibrinogen Fragment D serum was used.
Cross-Linking of Fibrinogen Fragment D with Fibrin-Fig. 7 shows the reduced SDS-gel electrophoretic patterns of Frag- ment Dr, which was cross-linked with fibrin under conditions listed in Table I. The fibrinogen Fragment Dr used in this
species were increased in Peaks 4 and 5 while the major species were decreased as indicated by staining intensity. Gels a and b show the reduced SDS-gel electrophoretic patterns of Fragment Dr. Gel a was stained for protein and Gel b for carbohydrate. Following reduction, a comparison of Gel a, primarily fibrinogen Fragment Dr. with Gel c of Fig. 2. nrimarilv fibrinogen Fraaments De and D8, shows that they’ chain is the major y chayn degr;dation product in Fragment Dr.
study was from the preparation shown in Fig. 3 and thus con- tains primarily Fragment Di contaminated with very small amounts of Fragments DP and Da. The first reaction mixture contained only fibrinogen and thus serves as a control by pro- viding a normal pattern of fibrin cross-linking. As can be seen on Gel a, this fibrin is highly cross-linked and both y dimer and Q! polymer are present (8). Gel b shows the subunit pattern of fibrin formed when the molar ratio of Fragment Dr to fibrinogen is 6: 1. Below the y dimer area, there are two bands visible on this gel which are absent on the control gel. As shown on Gel c, d, and e, the intensity of these two species becomes more pro- nounced as the molar ratio of Fragment Dr to fibrinogen increases. In addition, as these higher ratios are approached, the amount of CY polymer progressively decreases until only a small amount is apparent on Gel e when the molar ratio of Frag- ment Dr to fibrinogen is 40: 1. Finally, Gel f shows the cross- linking of Fragment Di in the absence of fibrinogen. Although no precipitate or other visible evidence of cross-linking was ob- served, examination of the electrophoretic pattern for this sample shows the formation of a y’ dimer and the disappearance of about 50% of the y’ monomer. The origin of the minor band, which has a lower mobility that the y’ dimer on Gel, f is uncer- tain. There was no indication that the y” chain or the y”’ chain of Fragments Dz and D3, respectively, cross-linked with either intact y chains, y’ chains, or themselves, as can be seen by the constant intensity of these two species on Gels b through f.
DISCUSSION
Purijication of Fragment D from Plasmin Digests of Fibrinogen and Cross-Linked Fibrin-Previous studies of the digestion of fibrinogen and fibrin by plasmin have resulted in models which predict the subunit structure of the various intermediate and terminal degradation products of both proteins (1, 2). The data in the present study provide additional evidence that fur- ther substantiates these models. As has been reported pre- viously (l), fibrinogen D exists as a population of species, the
by guest on June 22, 2018http://w
ww
.jbc.org/D
ownloaded from

4588
FIG. 6. (left). Microimmmodiffusion comparing fibrinogen Fragment Dr to fibrin Fragment D. Well a contained antiserum to human fibrinogen and Well d, antiserum to fibrinogen Frag- ment D. In both cases, Well b contained fibrinogen Fragment D1 and Well c, fibrin Fragment D.
FIG. 7. (right). The reduced SDS-gel electrophoretic patterns for the cross-linking of fibrinogen Fragment Di with fibrinogen. Gel a shows the electrophoretic pattern of highly cross-linked fibrin formed in the absence of Fragment Di. This gel shows a normal cross-linking pattern with virtually complete cross-link- ing of the y chains and cross-linking of most of the o( chains. Gel b shows the fibrin formed from a molar ratio of Fragment Dr to fibrinogen of 6:l; Gel c, a molar ratio of 10: 1; Gel d, a molar ratio
of 2O:l; and Gel e shows a molar ratio of 4O:l. Gel f shows the cross-linking of Fragment Dr in the absence of fibrinogen. Gel b through e shows two bands below the y dimer which represent an intact y chain cross-linked to a y’ chain, and two y’ chains cross- linked together. These bands increase in intensity in going from left to right. Gel f shows only the y’ dimer since no intact y chains were present in this sample. The origin of the minor high molec- ular weight band is uncertain. Gels b through f show no evidence that y” or 7” chains can participate in cross-linking, either with themselves or with intact y chains. Furthermore no evidence was found to suggest that the 01” chains can cross-link with them- selves or with intact a: chains.
TABLE I TABLE II
Cross-linking of puri.fied fragment D1 with and wilhout Jibrinogen Polypeptide chain composition of Fragment D species
Volume
Fibrinogen ‘ragment III
nd ml
0.10 0.00
0.09 0.01 0.08 0.02
0.07 0.03
0.06 0.04 0.05 0.05
0.04 0.06
0.03 0.07 0.02 0.08
0.01 0.09
0.00 0.10 -
Molar ratio of Fragment D1° to
fibrinogen
0
0.5 1 2
3 4 6
10 20
- I Formation of visible clot
40 co
+ (poor clot) -
- a Based on an assumed molecular weight of 80,000 for Fragment
D, and 330,000 for fibrinogen.
size of which largely depends on the extent of y chain digestion. As shown in Fig. 3, the first species of Fragment D to appear in plasmin digests is Fragment Dr u hich has the following subunit structure: an (Y” chain of about 15,000 molecular weight, a 0” chain of 44,500 molecular weight, and a y’ chain of 42,000 mo- lecular weight. However, as seen in Fig. 4, by lengthening the period of digestion, further cleavage of Fragment Di results in the formation of Fragments Dz and Da which increase in inten- sity as Fragment Di gradually disappears. In the chromatogram shown in Fig. lA, the majority of the Fragment D species existed as Fragment Da which contained the y” chain of 37,000 mo-
Fragment D species
Polypeptik chain subunits
Dl
D:!
D3
D3
-
TYPO Mol wt
01”
P ,I
Y’ a”
P r,
Y I,
d
P ,I
Y I,,
alll
P I,
Y 11,
-15,000 44,500
42,000 -15,000
44,500
37,000 -15,000
44,500
27,000 < 15,000
44,500
27,000
lecular weight instead of the y’ chain; and Fragment Da which contained the 7”’ chain of 27,000 molecular weight instead of the y’ or y” chains. At still longer digestion times than shown in this study Fragment Da can be completely digested with a concomitant increase in Fragment D1. In addition, with fur- ther degradation, another lower molecular weight species of Fragment D, D+ can be isolated whose subunit structure is thought to vary as a result of (Y” chain digestion, since there appears to be no change in molecular weight or decrease in in- tensity of the /3” or 7”’ chains (2). Following even the most prolonged digestion, no species of fibrinogen Fragment D ever showed evidence of /3” chain digestion. For convenience of the reader, Table II summarizes the subunit structures of the various
by guest on June 22, 2018http://w
ww
.jbc.org/D
ownloaded from

4589
fibrinogen Fragment D species. As indicated in the accompany- ing paper, the species of Fragment D derived from non-cross- linked fibrin had identical subunits to those of Fragment D from fibrinogen (2).
In contrast to the multiple forms of fibrinogen Fragment D, cross-linked fibrin Fragment D (Fig. 1B) was isolated as only a single species, even after prolonged digestions. As shown in Figs. 4 and 5, fibrin Fragment D showed a much lower mobility than fibrinogen Fragment D when the two proteins were com- pared by SDS-gel electrolhoresis. The difference in mobilities is easily explained, since fibrin Fragment D is essentially com- posed of two fibrinogen Fragment D1 molecules held together by cross-links in the COOH-terminal portion of the y’ chains. Hence, the 81,000 molecular weight subunit found in fibrin Frag- ment D must result from the partial digestion of y dimer. As a result of this cross-linked subunit species, the molecular weight of fibrin Fragment D would therefore be twice that reported for fibrinogen Fragment D (5). Additional support for this con- clusion is provided by our observation that fibrin Fragment D contained one pair of e-(y-glutamyl)lysyl cross-links as opposed to fibrinogen Fragment D1 which, as expected, contained none (12).2 These findings are in disagreement with the report by Dudek et al. that fibrinogen Fragment D and fibrin Fragment D have virtually identical molecular weights (13). Conceivably their fibrin Fragment D could have been recovered from a digest of non-cross-linked fibrin, thus explaining bhe discrepancy be- tween their results and ours.
When the species of fibrinogen Fragment D were examined for charge heterogeneity as shown in Fig. 5, at least five dif- ferent forms were identified. It appeared that Fragment D1 itself contained at least two major species by charge and that Fragment Dz or Dt also contained more than one species
of charge. Catanzaro et al. (14) have recently reported the identification of eight bands on standard polyacrylamide gel electrophoresis at pH 8.6 for a Fragment D species which con- tained three subunits of molecular weights of 44,500, 25,000, and 10,000. This Fragment D species presumably corresponds to our Fragment Da in light of the subunit molecular weights and the fact that an 1%hour digestion time was used. These results would seem to indicate that all or, at least several, of the Fragment D species may be very similar in size but different in charge.
Immunodi.ffusion Studies-The immunodiffusion studies in Fig. 6 show that Fragment D1 from fibrinogen and Fragment D from fibrin gave a line of identity when diffused against anti- serum to fibrinogen Fragment D. Thus the presence of inter- molecular cross-links in 1 he COOH-terminal portions of the y’ chains does not alter the reactivity of fibrin Fragment D with an antibody to fibrinogen Fragment D,. Therefore attempts to use such antiserum to distinguish between fibrinogen Frag- ment D1 and fibrin Fragment D could lead to the erroneous conclusion that the 2 molecules are identical.
Cross-Linking of Fibrinogen Fragment D with Fibrincgen-The studies presented in Fig. 7 show that fibrincgen Fragment Dr can be cross-linked bcth to fibrin mcncmers and itself if fibrin- stabilizing factor is present. However, only Fragment D 1 which contains y’ chains of 42,000 molecular weight is capable of form- ing cross-links since neither the 7” chain ncr the y”’ chain dis- appeared as cross-linked forms appeared. Moreover, as further confirmation of this observation, a mixture of Fragments C2 and Da, or a preparation containing primarily Fragment D3, were
substituted for Fragment D1, and still no evidence of cross-link formation was noted when these species were incubated with thrombin, calcium, fibrin-stabilizing factor, and fibrinogen. Thus neither the y” chain of Fragment DB nor the y”’ chain of Fragment DO can cross-link with intact y chains to form a higher molecular weight subunit as was the case with the y’ chain of Fragment D1. However, when fibrin was formed from a mix- ture of Fragment D1 and fibrinogen, it was possible to identify the normal y dimer fibrin subunit, and two new subunits con- sisting of a y chain cross-linked to a y’ chain or two y’ chains cross-linked to give a y’ dimer (Fig. 6). When Fragment D1 was cross-linked in the absence of fibrin monomer, about 50% of the -y’ chain disappeared with the concomitant formation of a y’ dimer. It should te emphasized that no visible precipitation or gelation cccurred in this sample despite the formation of cross- linked species. The above data indicate that y’ chains must be formed by removal of an lS Hz-terminal peptide from the y chain as previously suggested by our studies of fibrinogen and fibrin digestion by plasmin (1, 2). On the other hand, Fragments DP and Da n hich contain t.he y” chain of 37,000 molecular weight and the 7”’ chain of 27,000 molecular weight, respectively, do not cross-link and hence probably do not contain the COOH terminal cross-link region (15-17) as previously suggested (1, 2).
These results also explain the reports that Fragment D is an inhibitor of fibrin formation (18-24). Previously it was spec- ulated that cross-linking of Fragment D to fibrin monomers could account for the inhibition of clot formation (2529). The present study confirms that this in fact is the mechanism of inhibition. More recently, data have been presented showing that a molar ratio of Fragment D1 to fibrinogen of 40: 1 markedly inhibited clotting (24, 30). In the present study it was con- firmed that clotting was delayed at this ratio and in fact, only a granular precipitate was observed. However, SDS-gel elec- trophoretic analysis of this sample (Fig. 7, Gel f) showed that while LY polymerizaticn was only partial with some (Y chains remaining non-cross-linked, all of the intact y chains of fibrino- gen became cross-linked to anot.her intact y chain or to a 7’ chain of Fragment D1. All of the y’ chains did not become cross- linked, possibly as a result of the conformation of Fragment DI being altered in such a way that the rate of cross-linking was decreased. It must be kept in mind that at high ratios of fibrino- gen Fragment D to fibrinogen, tl-e y chain of a fibrin monomer will more likely encounter a -y’ chain of a Fragment I~1 species than a y chain of another fibrin monomer, thus markedly slow- ing the rate of fibrin polymerization.
REFERENCES
1. PIZZO, S. V., SCHWARTZ, M. L., HILL, R. L. & MCKEE, P. A. (1972) J. Biol. Chem. 247, 636-645
2. PIZZO, S., SCHWARTZ, M., HILL, R. & MCKEE, P. (1973) J. Biol. Chem. 246, 4574-4583
3. NUSSENZWEIG, V. & SELIGMANN, M. (1960) Rev. Hematol.16, 451-466
4. NUSSENZWEIG, V., SELIGMANN, M., PELMONT, J. & GRABAR, P. (1961) Ann. Inst. Pasteur Paris 100, 377-389
5. MARDER, V.J., SHULMAN,N. R. &CARROLL, W. R. (1969)J. Biol. Chem. 244, 2111-2119
6. BLOMB;~CK, B., BLOMB~CK, M., HESSEL, B. & IWANAGA, S. (1967) Nature 216, 1445-1448
7. SCHWARTZ, M. L., PIZZO, S. V., HILL, R. L. & MCKEE, P. A. (1971) J. Biol. Chem. 246, 5851-5854
8. MCKEE, P. A., MATTOCK, P. & HILL, R. L. (1970) Proc. Nat. Acad. Sci. U. S. A. 66. 738-744
2 A. P. Ball, S. V. Pizza, and P. A McKee, unpublished observa- 9. PANYIM, S. & CHALKLEG, R. (1969) Arch. Biochem. Biophys. t,ions. 130, 337-346
by guest on June 22, 2018http://w
ww
.jbc.org/D
ownloaded from

4590
10. OUCHTERLQNY, 0. (1968) Handbook of Immunodifusion and Immunoelectrophoresis. 215 p., Ann Arbor Science Pub- lishers, Inc., Ann Arbor, Michigan
11. ZACHARIUS. R. M.. ZELL. T. E.. MORRISON. J. H. & WOOD- LOCK, J. j. (1969’) Anal: Biochkm. 30, 14%i52
12. PISANO, J. J., FINLAYSON, J. S. & PEYTON, M. P. (1969) Bio- chemistry 8, 871-876
13. DUDEK, G. A., KLOCZEWIAK, M., BUDZYASKI, A. Z., LATAL~O, Z. S. & KOPE~, M. (1970) Biochem. Biophys. Acta 214, 44- 51
14. CATANZARO, A., HATHAWAY, G., STRATHERN, J. & EDGINGTON, T. (1972) Proc. Sot. Exp. Biol. Med. 139, 1401-1406
15. CHEN, R. & DOOLITTLE, R. F. (1970) Proc. Nat. Acad. Sci. U. S. A. 66, 472-479
16. CHEN, R. & DOOLITTLE, R. F. (1971) Biochemistry 10, 4486- 4491
17. DOOLITTLF,, R. F., CHEN, R. & LAU, F. (1971) Biochem. Bio- phys. Res. Commun. 44, 94-100
18. TRIANTAPHYLLOPOULOS, D. C. (1958) Can. J. Biochem. Phys- iol. 36, 249-259
19. NIEWIAROWSKI, S. & KO~.U~SKI, E. (1958) Rev. HematoZ. 13, 320-328
20. TRIANTAPHYLLOPOULOS, E. & TRIANTAPHYLLOPOULOS, D. C. (1965) Amer. J. Phusiol. 208, 521-727
21. TR~T.~PHYLLOPCIUL&, E. & ‘TRIANTAPHYLLOPOULOS, D. C. (1965) Brit. J. Haematol. 11, 331-341
22. TRIANTAPHYLLOPOULOS. E. & TRISNTAPHYLLOPOULOS. D. C. (1967) Brit. J. Haematol. 13, 28-41
23. MARDER, V. J., SHULMAN, N. R. & CARROLL, W. R. (1967) Trans. Ass. Amer. Phusicians. Philadelphia 80. 156-167
24. MARDER, V. J. & SHULM~N, N. &. (1969) j. Biol. .Chem. 244, 2120-2124
25. FLETCHER, A. P., ALKJAERSIG, N. &. SHERRY, 8. (1962) J. Clin. Invest. 41, 896-916
26. ALKJAERSIG, N., FLETCHER, A. P. & SHERRY, S. (1962) J. Clin. Invest. 41, 917-934
27. BANG, N. U., FLETCHER, A. P., ALICJAERSIG, N. &. SHERRY, S. (1962) 3’. Clin. Znvek. 41, kI35-948
28. LATA~EO. Z. S.. FLETCHER. A. P.. ALIGJAERSIG. N. & SHERRY. S. (196i) dmkr. J. Physiol. 2Oi, 681-686 ’
29. FLETCHER, A. P., ALKJAERSIG, N., FISHER, S. & SHERRY, S. (1966) J. Lab. Clin. Med. 68, 780-802
30. MARDER, V. J. (1970) Thromb. Diath. Haemorrh. 39, (suppl), 187-195
by guest on June 22, 2018http://w
ww
.jbc.org/D
ownloaded from

A. McKeeSalvatore V. Pizzo, Lloyd M. Taylor, Jr., Martin L. Schwartz, Robert L. Hill and Patrick
Subunit Structure of Fragment D from Fibrinogen and Cross-Linked Fibrin
1973, 248:4584-4590.J. Biol. Chem.
http://www.jbc.org/content/248/13/4584Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/248/13/4584.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on June 22, 2018http://w
ww
.jbc.org/D
ownloaded from


















