
Subcellular localization of the von Hippel-Lindau disease gene product is cell cycle-dependent
8
SUBCELLULAR LOCALIZATION OF THE VON HIPPEL-LINDAU DISEASE GENE PRODUCT IS CELL CYCLE-DEPENDENT Ying Y E 1 , Sandip V ASAVADA 1 , Igor KUZMIN 2 , Thomas STACKHOUSE 2 , Berton ZBAR 2 and Bryan R.G. WILLIAMS 1 * 1 Department of Cancer Biology, Lerner Research Institute, Cleveland Clinic Foundation, Cleveland, OH, USA 2 Laboratory of Immunobiology, Biological Carcinogenesis and Development Program, National Cancer Institute, Frederick, MD, USA The von Hippel-Lindau gene product (pVHL) interacts with and inhibits the cellular transcription factor elongin. How- ever, the subcellular localization of pVHL has remained uncertain. Naturally occurring pVHL mutants which fail to interact with elongin have been described in patients with VHL disease or sporadic renal cell carcinoma (RCC). Here, we have examined the cellular expression pattern of endog- enous pVHL in different RCC cell lines by immunocytochem- istry and confocal microscopy. Both anti-N-terminal and anti-C-terminal pVHL antibodies were able to recognize endogenous wild-type pVHL expressed by the RCC cells studied. A C-terminal truncated VHL mutant expressed by RCC cell line A498 was detected only by the N-terminal antibody but not by the C-terminal antibody as expected. The overall staining patterns of these cell lines are similar, with a predominant nuclear speckled pattern and a moderate cyto- plasmic staining in subconfluent cell cultures. Interestingly, when cells reached confluency, more prominent nuclear staining with little or no cytoplasmic expression was ob- served. By using double labeling with anti-VHL and anti- bromodeoxyuridine (BrdU) antibodies and cell cycle analy- ses, we found that in the G 1 /G 0 -phase, pVHL was localized exclusively in the nucleus associated with distinctive large subnuclear structures, whereas the majority of the cells in S-phase of the cell cycle also showed a diffuse cytoplasmic staining. Our results indicate that subcellular localization of pVHL is regulated in a cell cycle-dependent manner. Int. J. Cancer 78:62–69, 1998. r 1998 Wiley-Liss, Inc. The von Hippel-Lindau (VHL) syndrome is an autosomal dominant inherited cancer syndrome characterized by hemangio- blastomas of the retina and central nervous system as well as malignant neoplasms of the kidney and adrenal gland. Patients with VHL disease have a median life expectancy of 49 years and the most common cause of death in these patients is renal cell carcinoma (RCC) (Maher et al., 1991). The VHL tumor suppressor gene is located on human chromosome 3p25-26 (Latif et al., 1993). A partial cDNA was cloned which encodes 3 exons and predicts a 24-kDa protein product (pVHL) (Latif et al., 1993). Subsequent mutational analyses have indicated that approximately 57% of patients with sporadic clear cell RCC have somatic VHL mutations (Gnarra et al., 1994; Osaka et al., 1994). The gene is silenced by methylation in a further 10% of patients (Herman et al., 1994). Reintroduction of wild-type VHL into 786-0 cells, a RCC cell line harboring a VHL mutant, inhibited their ability to form tumors in nude mice (Iliopoulos et al., 1995b). These findings indicate that the VHL gene has a critical role in the development of RCC. At present, the precise biological function of the VHL protein product has not been elucidated, although there is biochemical evidence that pVHL functions as a negative regulator of the transcriptional elongation factor elongin (SIII) (Aso et al., 1995; Kibel et al., 1995; Duan et al., 1995b). Elongin is a heterotrimer composed of subunits A, B and C (Aso et al., 1995). Elongin A, a catalytic subunit, is capable of activating RNA polymerase II (pol II) elongation and its specific activity is greatly increased when it is complexed with regulatory subunit C. Elongin B serves as a chaperone to facilitate assembly and to enhance the stability of the elongin complex. pVHL has been shown to compete with elongin A by binding tightly and specifically to elongin B/C (Aso et al., 1995; Kibel et al., 1995) and to inhibit transcriptional activity of elongin in vitro (Duan et al., 1995b). The binding site of pVHL to elongin B/C was mapped to a 32-amino acid (aa) C-terminal region of pVHL (Kishida et al., 1995). This binding domain appears to be the region where mutations frequently occur in the germline of VHL patients (the region contains the aa 167 hot spot mutation) as well as in sporadic RCC. These data suggest that the binding of pVHL to these 2 proteins is essential for cellular growth regulation, and failure of this binding may contribute to the tumorigenesis charac- teristic of VHL disease. However, it is obvious that VHL is not a global regulatory inhibitor of transcriptional elongation since constitutional cells expressing mutated VHL are viable. Moreover, several VHL-associated cellular proteins have yet to be identified. It is possible that pVHL targets other biochemical pathways in addition to transcription elongation or that its function is under the regulation of other cellular components (Kishida et al., 1995; Tsuchiya et al., 1996). The vascular endothelial growth factor (VEGF) has emerged as a potential target of pVHL (Siemeister et al., 1996). VEGF is a key mediator in the induction of angiogen- esis, and its receptors, VEGFR-1 and VEGFR-2, are preferentially expressed in the vessels lining and penetrating the tumors (Martiny- Baron and Marme, 1995). The expression of VEGF was deregu- lated by inactive mutant VHL and this was reverted by the introduction of wild-type VHL (Siemeister et al., 1996). pVHL also binds the cdc53-related protein CUL2 and may play a role in cell cycle exit (Pause et al., 1997). The repression of VEGF by pVHL requires an interaction with the transcription factor Sp1 and may also involve a direct interaction with and inhibition of isoforms of protein kinase C (Mukhopadhyay et al., 1997; Pal et al., 1997). Since pVHL is apparently directly involved in regulation of transcriptional process, its subcellular localization was presumed to be in the nucleus. It is now known that pVHL is indeed a nuclear protein, but it can also be found in cytoplasm. This has led to speculation that pVHL may be able to translocate between nuclear and cytosolic compartments. Lee et al. (1996) have identified a putative nuclear localization signal (NLS), PR[R/K] . . . RPRPV, in the first 60 and 28 aa of the human and rat VHL protein, respectively, and have suggested that the nuclear transport of pVHL is determined by cell density. Here, we examined the cellular expression pattern of endog- enous pVHL in several RCC cell lines. In agreement with other reports (Tsuchiya et al., 1996; Siemeister et al., 1996; Martiny- Baron and Marme, 1995; Lee et al., 1996), pVHL can be detected in both the nucleus and cytoplasm of RCC cells. We also observed that the subcellular localization of pVHL is influenced by cell density and cell cycle stage, and identified the cell population that expressed exclusive nuclear VHL as in the G 1 /G 0 -phase of the cell cycle. Cells with both nuclear and cytosolic staining were in the S-phase of the cell cycle. Thus, the subcellular localization of pVHL is cell cycle regulated. Grant sponsor: USPHS; Grant number: DK47101. *Correspondence to: Department of Cancer Biology, NN10, Lerner Research Institute, Cleveland Clinic Foundation, 9500 Euclid Avenue, Cleveland, OH 44195, USA. Fax: (216) 445-6269. E-mail: [email protected] Received 5 January 1998; Revised 6 April 1998 Int. J. Cancer: 78, 62–69 (1998) r 1998 Wiley-Liss, Inc. Publication of the International Union Against Cancer Publication de l’Union Internationale Contre le Cancer
Transcript of Subcellular localization of the von Hippel-Lindau disease gene product is cell cycle-dependent

SUBCELLULAR LOCALIZATION OF THE VON HIPPEL-LINDAU DISEASE GENEPRODUCT IS CELL CYCLE-DEPENDENTYing YE1, Sandip VASAVADA 1, Igor KUZMIN2, Thomas STACKHOUSE2, Berton ZBAR2 and Bryan R.G. WILLIAMS 1*1Department of Cancer Biology, Lerner Research Institute, Cleveland Clinic Foundation, Cleveland, OH, USA2Laboratory of Immunobiology, Biological Carcinogenesis and Development Program, National Cancer Institute, Frederick, MD, USA
The von Hippel-Lindau gene product (pVHL) interacts withand inhibits the cellular transcription factor elongin. How-ever, the subcellular localization of pVHL has remaineduncertain. Naturally occurring pVHL mutants which fail tointeract with elongin have been described in patients withVHL disease or sporadic renal cell carcinoma (RCC). Here,we have examined the cellular expression pattern of endog-enous pVHL in different RCC cell lines by immunocytochem-istry and confocal microscopy. Both anti-N-terminal andanti-C-terminal pVHL antibodies were able to recognizeendogenous wild-type pVHL expressed by the RCC cellsstudied. A C-terminal truncated VHL mutant expressed byRCC cell line A498 was detected only by the N-terminalantibody but not by the C-terminal antibody as expected. Theoverall staining patterns of these cell lines are similar, with apredominant nuclear speckled pattern and a moderate cyto-plasmic staining in subconfluent cell cultures. Interestingly,when cells reached confluency, more prominent nuclearstaining with little or no cytoplasmic expression was ob-served. By using double labeling with anti-VHL and anti-bromodeoxyuridine (BrdU) antibodies and cell cycle analy-ses, we found that in the G1/G0-phase, pVHL was localizedexclusively in the nucleus associated with distinctive largesubnuclear structures, whereas the majority of the cells inS-phase of the cell cycle also showed a diffuse cytoplasmicstaining. Our results indicate that subcellular localization ofpVHL is regulated in a cell cycle-dependent manner. Int. J.Cancer 78:62–69, 1998.r 1998 Wiley-Liss, Inc.
The von Hippel-Lindau (VHL) syndrome is an autosomaldominant inherited cancer syndrome characterized by hemangio-blastomas of the retina and central nervous system as well asmalignant neoplasms of the kidney and adrenal gland. Patients withVHL disease have a median life expectancy of 49 years and themost common cause of death in these patients is renal cellcarcinoma (RCC) (Maheret al.,1991). TheVHL tumor suppressorgene is located on human chromosome 3p25-26 (Latifet al.,1993).A partial cDNA was cloned which encodes 3 exons and predicts a24-kDa protein product (pVHL) (Latifet al., 1993). Subsequentmutational analyses have indicated that approximately 57% ofpatients with sporadic clear cell RCC have somaticVHL mutations(Gnarraet al., 1994; Osakaet al., 1994). The gene is silenced bymethylation in a further 10% of patients (Hermanet al., 1994).Reintroduction of wild-typeVHL into 786-0 cells, a RCC cell lineharboring aVHL mutant, inhibited their ability to form tumors innude mice (Iliopouloset al., 1995b). These findings indicate thattheVHL gene has a critical role in the development of RCC.
At present, the precise biological function of the VHL proteinproduct has not been elucidated, although there is biochemicalevidence that pVHL functions as a negative regulator of thetranscriptional elongation factor elongin (SIII) (Asoet al., 1995;Kibel et al., 1995; Duanet al., 1995b). Elongin is a heterotrimercomposed of subunits A, B and C (Asoet al.,1995). Elongin A, acatalytic subunit, is capable of activating RNA polymerase II (polII) elongation and its specific activity is greatly increased when it iscomplexed with regulatory subunit C. Elongin B serves as achaperone to facilitate assembly and to enhance the stability of theelongin complex. pVHL has been shown to compete with elongin Aby binding tightly and specifically to elongin B/C (Asoet al.,1995;Kibel et al.,1995) and to inhibit transcriptional activity of elonginin vitro (Duanet al.,1995b). The binding site of pVHL to elongin
B/C was mapped to a 32-amino acid (aa) C-terminal region ofpVHL (Kishidaet al.,1995). This binding domain appears to be theregion where mutations frequently occur in the germline of VHLpatients (the region contains the aa 167 hot spot mutation) as wellas in sporadic RCC. These data suggest that the binding of pVHL tothese 2 proteins is essential for cellular growth regulation, andfailure of this binding may contribute to the tumorigenesis charac-teristic of VHL disease. However, it is obvious that VHL is not aglobal regulatory inhibitor of transcriptional elongation sinceconstitutional cells expressing mutated VHL are viable. Moreover,several VHL-associated cellular proteins have yet to be identified.It is possible that pVHL targets other biochemical pathways inaddition to transcription elongation or that its function is under theregulation of other cellular components (Kishidaet al., 1995;Tsuchiya et al., 1996). The vascular endothelial growth factor(VEGF) has emerged as a potential target of pVHL (Siemeisteretal., 1996). VEGF is a key mediator in the induction of angiogen-esis, and its receptors, VEGFR-1 and VEGFR-2, are preferentiallyexpressed in the vessels lining and penetrating the tumors (Martiny-Baron and Marme, 1995). The expression of VEGF was deregu-lated by inactive mutant VHL and this was reverted by theintroduction of wild-type VHL (Siemeisteret al.,1996). pVHL alsobinds the cdc53-related protein CUL2 and may play a role in cellcycle exit (Pauseet al.,1997). The repression of VEGF by pVHLrequires an interaction with the transcription factor Sp1 and mayalso involve a direct interaction with and inhibition of isoforms ofprotein kinase C (Mukhopadhyayet al.,1997; Palet al.,1997).
Since pVHL is apparently directly involved in regulation oftranscriptional process, its subcellular localization was presumed tobe in the nucleus. It is now known that pVHL is indeed a nuclearprotein, but it can also be found in cytoplasm. This has led tospeculation that pVHL may be able to translocate between nuclearand cytosolic compartments. Leeet al. (1996) have identified aputative nuclear localization signal (NLS), PR[R/K] . . . RPRPV, inthe first 60 and 28 aa of the human and rat VHL protein,respectively, and have suggested that the nuclear transport of pVHLis determined by cell density.
Here, we examined the cellular expression pattern of endog-enous pVHL in several RCC cell lines. In agreement with otherreports (Tsuchiyaet al., 1996; Siemeisteret al., 1996; Martiny-Baron and Marme, 1995; Leeet al.,1996), pVHL can be detectedin both the nucleus and cytoplasm of RCC cells. We also observedthat the subcellular localization of pVHL is influenced by celldensity and cell cycle stage, and identified the cell population thatexpressed exclusive nuclear VHL as in the G1/G0-phase of the cellcycle. Cells with both nuclear and cytosolic staining were in theS-phase of the cell cycle. Thus, the subcellular localization ofpVHL is cell cycle regulated.
Grant sponsor: USPHS; Grant number: DK47101.
*Correspondence to: Department of Cancer Biology, NN10, LernerResearch Institute, Cleveland Clinic Foundation, 9500 Euclid Avenue,Cleveland, OH 44195, USA. Fax: (216) 445-6269.E-mail: [email protected]
Received 5 January 1998; Revised 6 April 1998
Int. J. Cancer:78,62–69 (1998)
r 1998 Wiley-Liss, Inc.
Publication of the International Union Against CancerPublication de l’Union Internationale Contre le Cancer

MATERIAL AND METHODS
Cell lines and antibodies
Cultures of human RCC lines CCF-RC1, A498 and UMRC6were maintained in RPMI 1640 medium with 10% fetal calf serum(FCS) as described previously (Hashimuraet al.,1989; Chenet al.,1995). A498 (ATCC) has a 4-base deletion at nucleotides 639–642of theVHL gene (Latifet al.,1993). UMRC6 containing a 10-basedeletion (717–726) (Latifet al.,1993) was transfected with pCEP4plasmid (Invitrogen, San Diego, CA) coding for full-length wild-type pVHL under control of the CMV promoter using lipofectin(Life Technologies, Gaithersburg, MD) as described (Grossmanetal., 1985). Hygromycin-resistant clones were analyzed for thepresence of the 700 nucleotide exogenous VHL transcript (byNorthern blot) and wild-type pVHL (by Western blot). No VHLmutations were identified in RC1 cells by single-strand conforma-tion polymorhism (SSCP) analyses using 6 sets of polymerasechain reaction (PCR) primers covering the known open readingframe (Blencoweet al.,1994).
Polyclonal antibodies to VHL N-terminal peptide (aa 1–16) andto C-terminal peptide (aa 198–213, which specifically recognizeN-terminal and C-terminal VHL peptides in Western blots) wereraised in New Zealand white rabbits and affinity purified (Gnarraetal., 1994). Anti-WT1 (C19), anti-tubulin (a-tubulin) and anti-bromodeoxyuridine (anti-BrdU) antibodies were obtained commer-cially from Santa Cruz Biotechnology (Santa Cruz, CA), Sigma(St. Louis, MO) and Amersham (Arlington Heights, IL), respec-
tively. Mouse monoclonal anti-spliceosome antibody was obtainedfrom Oncogene Research Products (Cambridge, MA).
Immunocytochemistry and confocal microscopyCells grown on coverslips were washed with phosphate-buffered
saline (PBS) and fixed in cold acetone for 10 min and air dried.Coverslips were rehydrated in PBS prior to staining or kept at220°C for later use. Indirect immunofluorescent staining wasperformed by preincubating cells with blocking solution [3%bovine serum albumin (BSA), 0.1% Tween 20/PBS] for 45 min,followed by incubation with primary antibodies for 1 hr at roomtemperature and rinsed with PBS. Secondary antibodies [fluores-cein isothiocyanate (FITC)-conjugated anti-rabbit IgG and/or rho-damine-conjugated anti-mouse IgG] were applied for 45 min andcells were rinsed and mounted on glass slides with Vectashied(Vector, Burlingame, CA).
For double staining of VHL and BrdU, cells labeled with BrdU(Amersham) were washed with PBS 3 times, fixed with 1%formaldehyde/PBS for 20 min, washed with PBS and permeabi-lized with 0.1% Triton-X-100/PBS for 5 min prior to blocking.Anti-VHL antibody (1:100 dilution) was first incubated with cellsfor 1 hr, rinsed in PBS and treated with freshly prepared 2N-HClfor 5 min followed by a PBS rinse. Anti-BrdU antibody was appliedfor 1 hr at 37°C, then processed as described above. Double-labeledcells were analyzed by fluorescent microscopy using standardfilters for simultaneous detection of FITC and rhodamine (Texas
FIGURE 1 – Indirect immunostaining of CCF-RC1(a), UMRC6 (b) and A498(c) cells with anti-N-terminal VHL antibody, and CCF-RC1(d) and A498(e)cells with anti-C-terminal VHL antibody. Bars: 25 µm(a,b,d); 10 µm(c).
63LOCALIZATION OF pVHL

red). The detailed cellular localization of proteins was furtheranalyzed by confocal laser scanning microscopy.
Cell cycle analysisRC1 cells were synchronized by growing cells to confluency,
then serum starving in RPMI 1640 medium containing 0.3% FCSfor 48 hr. Cells were harvested and kept in cryoprotective media(RPMI containing L-glutamine and 25 mM Hepes, 20% new borncalf serum (NBCS), 10% dimethylsulfoxide and gentamicin) at270°C for subsequent cell cycle analyses by flow cytometry. Cellswere stained and analyzed according to the manufacturer’s proto-col (Becton Dickinson, Mountain View, CA). Synchronized G1/G0-phase cells were induced to progress by replacing fresh mediumwith 10% FCS. After 7–28 hr incubation, cell populations in dif-ferent stages of the cell cycle were determined by flow cytometry.
RESULTS
pVHL expression in RCC cell cultures
One sporadic RCC strain (RC1) and 2 cell lines (A498 andUMRC6) were used for this study. As reported previously (Latifetal., 1993), A498 has a 5-nucleotide (nt) VHL deletion (638–642)that created a frameshift mutation resulting in a C-terminaltruncated protein. UMRC6, harboring a 10-nt VHL deletion(717–726), was transfected with pCEP4 plasmids carrying afull-length wild-type VHL minigene. Overexpression of theVHLtransgene in UMRC6 was confirmed by Northern and Westernanalyses (data not shown). Through SSCP analyses of genomic
DNA, we determined that RC1 contains a wild-typeVHL gene.Two polyclonal antibodies raised against the N- and C-terminalregions of pVHL were used for immunofluorescence staining (seeMaterial and Methods). Most staining was performed on subconflu-ent cell cultures fixed with acetone. The N-terminal antibody thatrecognizes all forms of pVHL, including the endogenous, exog-enous wild-type VHL and C-end truncated mutant, stained pVHLexpressed by RC1, UMRC6 and A498, respectively (Fig. 1a–c). Asexpected, the C-terminal antibody reacts only with the full-lengthwild-type pVHL expressed by RC1 (UMRC6 was not tested) butnot with mutant pVHL in A498 (Fig. 1d,e). Despite the differentforms of pVHL, the overall staining patterns are similar, withpVHL preferentially expressed in the nucleus in large punctatebodies and to a lesser extent in the cytoplasm where staining ismore diffuse and less intense.
pVHL does not colocalize with splicing factorsThe nuclear staining pattern of pVHL appears similar to the
‘‘speckled’’ pattern described for splicing factors (Nymanet al.,1986; Spectoret al., 1991). Splicing factors along with the RNAcomponent are located in the nuclear matrix to form the ‘‘splicingapparatus’’, a highly organized structure that also provides theframework for nuclear shape and conformity (Berezney andCoffey, 1974; Blencoweet al., 1994). Confocal microscopy hasmade possible the high-resolution analysis of protein colocalizationin whole cells. For example, the Wilms tumor suppressor geneproduct WT1 has been shown to be colocalized with severalsplicing factors as well as p53in vivo by confocal analyses
FIGURE 2 – Localization of WT1, VHL and spliceosomes. RC1 cells were labeled with anti-WT1 antibody(a) and double-stained withanti-VHL (b,d) or anti-spliceosome(c,e)antibodies. Images(d–f) were captured by confocal microscopy. Image( f ) was created by overlayingimages(d) and(e).Bars: 25 µm(a–c); 10 µM (d–f).
64 YE ET AL.
(Larssonet al., 1995; Ye et al., 1996). To investigate whetherpVHL is also associated with splicing factors, we first comparedstaining patterns of pVHL with WT1 in RC1 cells. WT1 is alsoexpressed predominantly in the nucleus (Fig. 2a), but the nuclearstains are relatively diffuse or fine granules compared to the brightnuclear body clusters of pVHL viewed (Fig. 1a) under fluorescentmicroscopy. To determine whether pVHL and splicing factorscolocalized, RC1 cells were double-labeled with anti-VHL andanti-spliceosome antibodies and the labeled cells analyzed underimmunofluorescent (Fig. 2b,c) and confocal microscopy. Confocalanalyses revealed that while both pVHL and spliceosomes arelocalized in the nucleus and exist as a large number of bright foci(Fig. 2d,e), the patterns of positive foci are not identical althoughthere are few small overlapping areas shown as yellow from thesuperimposed image (Fig. 2f). This staining demonstrates thatalthough pVHL and spliceosome are colocalized in the nucleus,major molecular interactions do not likely occur because of thenon-overlapping staining patterns.
Subcellular localization of pVHL is cell cycle-dependentDuring the course of the study, we observed that in confluent cell
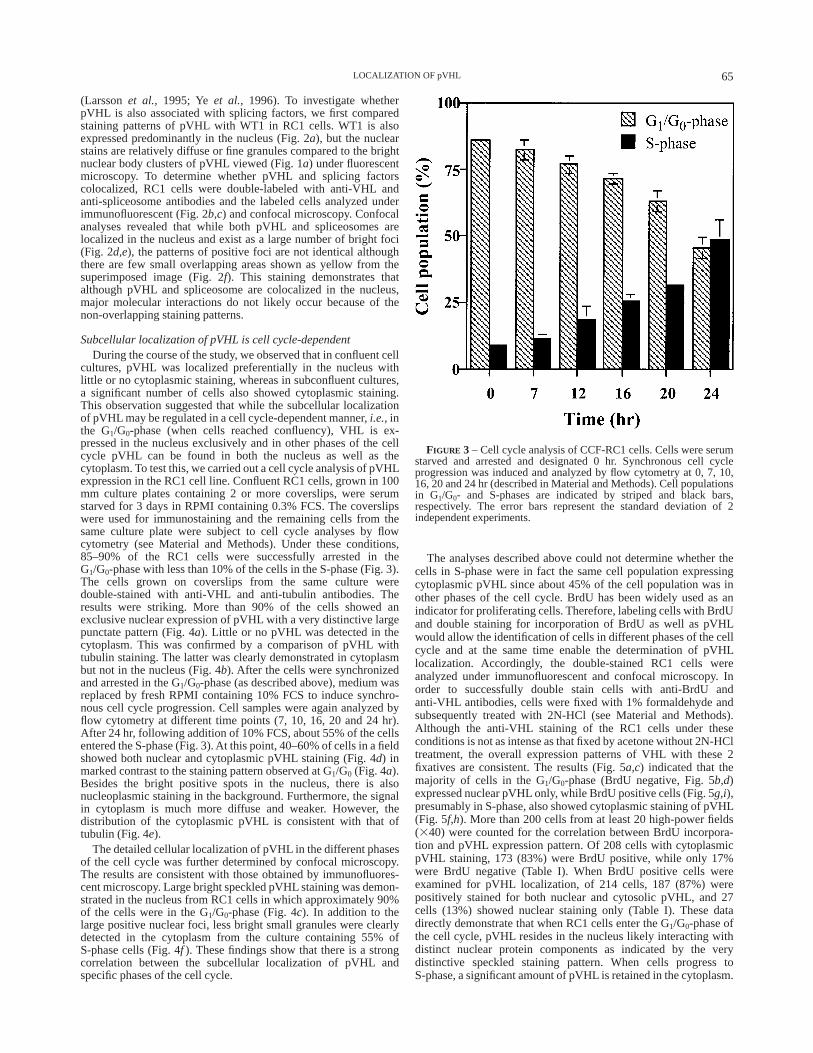
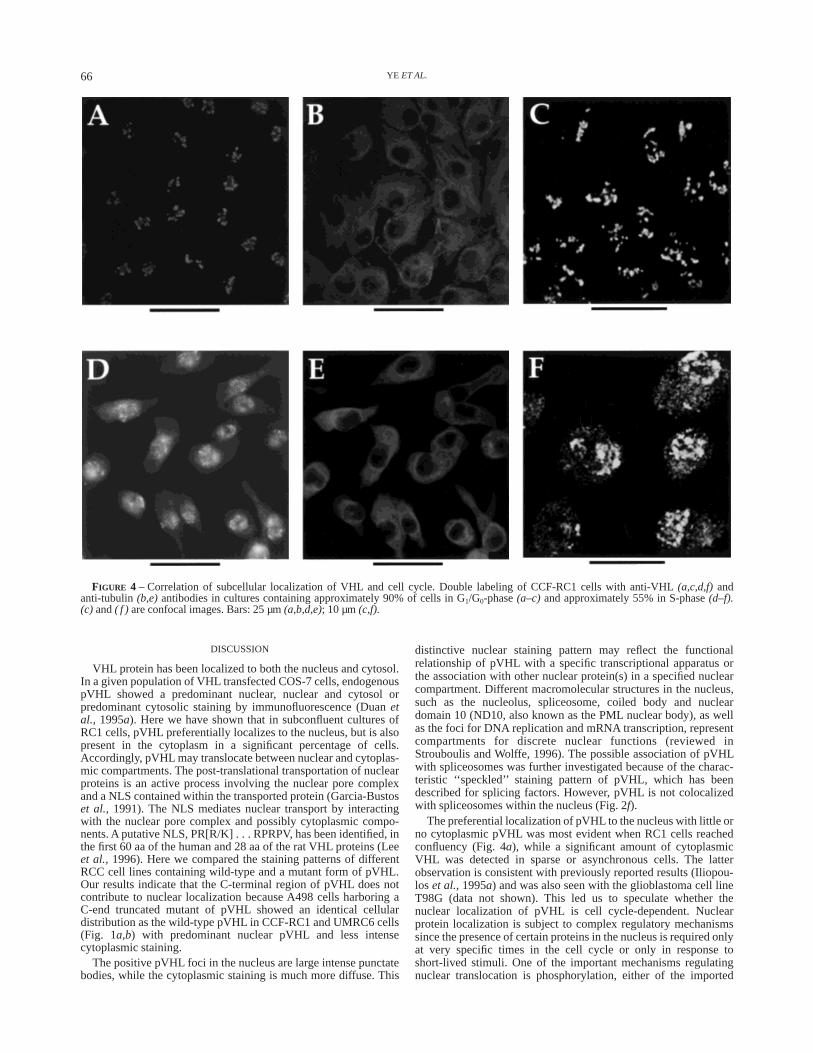
cultures, pVHL was localized preferentially in the nucleus withlittle or no cytoplasmic staining, whereas in subconfluent cultures,a significant number of cells also showed cytoplasmic staining.This observation suggested that while the subcellular localizationof pVHL may be regulated in a cell cycle-dependent manner,i.e.,inthe G1/G0-phase (when cells reached confluency), VHL is ex-pressed in the nucleus exclusively and in other phases of the cellcycle pVHL can be found in both the nucleus as well as thecytoplasm. To test this, we carried out a cell cycle analysis of pVHLexpression in the RC1 cell line. Confluent RC1 cells, grown in 100mm culture plates containing 2 or more coverslips, were serumstarved for 3 days in RPMI containing 0.3% FCS. The coverslipswere used for immunostaining and the remaining cells from thesame culture plate were subject to cell cycle analyses by flowcytometry (see Material and Methods). Under these conditions,85–90% of the RC1 cells were successfully arrested in theG1/G0-phase with less than 10% of the cells in the S-phase (Fig. 3).The cells grown on coverslips from the same culture weredouble-stained with anti-VHL and anti-tubulin antibodies. Theresults were striking. More than 90% of the cells showed anexclusive nuclear expression of pVHL with a very distinctive largepunctate pattern (Fig. 4a). Little or no pVHL was detected in thecytoplasm. This was confirmed by a comparison of pVHL withtubulin staining. The latter was clearly demonstrated in cytoplasmbut not in the nucleus (Fig. 4b). After the cells were synchronizedand arrested in the G1/G0-phase (as described above), medium wasreplaced by fresh RPMI containing 10% FCS to induce synchro-nous cell cycle progression. Cell samples were again analyzed byflow cytometry at different time points (7, 10, 16, 20 and 24 hr).After 24 hr, following addition of 10% FCS, about 55% of the cellsentered the S-phase (Fig. 3). At this point, 40–60% of cells in a fieldshowed both nuclear and cytoplasmic pVHL staining (Fig. 4d) inmarked contrast to the staining pattern observed at G1/G0 (Fig. 4a).Besides the bright positive spots in the nucleus, there is alsonucleoplasmic staining in the background. Furthermore, the signalin cytoplasm is much more diffuse and weaker. However, thedistribution of the cytoplasmic pVHL is consistent with that oftubulin (Fig. 4e).
The detailed cellular localization of pVHL in the different phasesof the cell cycle was further determined by confocal microscopy.The results are consistent with those obtained by immunofluores-cent microscopy. Large bright speckled pVHL staining was demon-strated in the nucleus from RC1 cells in which approximately 90%of the cells were in the G1/G0-phase (Fig. 4c). In addition to thelarge positive nuclear foci, less bright small granules were clearlydetected in the cytoplasm from the culture containing 55% ofS-phase cells (Fig. 4f ). These findings show that there is a strongcorrelation between the subcellular localization of pVHL andspecific phases of the cell cycle.
The analyses described above could not determine whether thecells in S-phase were in fact the same cell population expressingcytoplasmic pVHL since about 45% of the cell population was inother phases of the cell cycle. BrdU has been widely used as anindicator for proliferating cells. Therefore, labeling cells with BrdUand double staining for incorporation of BrdU as well as pVHLwould allow the identification of cells in different phases of the cellcycle and at the same time enable the determination of pVHLlocalization. Accordingly, the double-stained RC1 cells wereanalyzed under immunofluorescent and confocal microscopy. Inorder to successfully double stain cells with anti-BrdU andanti-VHL antibodies, cells were fixed with 1% formaldehyde andsubsequently treated with 2N-HCl (see Material and Methods).Although the anti-VHL staining of the RC1 cells under theseconditions is not as intense as that fixed by acetone without 2N-HCltreatment, the overall expression patterns of VHL with these 2fixatives are consistent. The results (Fig. 5a,c) indicated that themajority of cells in the G1/G0-phase (BrdU negative, Fig. 5b,d)expressed nuclear pVHL only, while BrdU positive cells (Fig. 5g,i),presumably in S-phase, also showed cytoplasmic staining of pVHL(Fig. 5f,h). More than 200 cells from at least 20 high-power fields(340) were counted for the correlation between BrdU incorpora-tion and pVHL expression pattern. Of 208 cells with cytoplasmicpVHL staining, 173 (83%) were BrdU positive, while only 17%were BrdU negative (Table I). When BrdU positive cells wereexamined for pVHL localization, of 214 cells, 187 (87%) werepositively stained for both nuclear and cytosolic pVHL, and 27cells (13%) showed nuclear staining only (Table I). These datadirectly demonstrate that when RC1 cells enter the G1/G0-phase ofthe cell cycle, pVHL resides in the nucleus likely interacting withdistinct nuclear protein components as indicated by the verydistinctive speckled staining pattern. When cells progress toS-phase, a significant amount of pVHL is retained in the cytoplasm.
FIGURE 3 – Cell cycle analysis of CCF-RC1 cells. Cells were serumstarved and arrested and designated 0 hr. Synchronous cell cycleprogression was induced and analyzed by flow cytometry at 0, 7, 10,16, 20 and 24 hr (described in Material and Methods). Cell populationsin G1/G0- and S-phases are indicated by striped and black bars,respectively. The error bars represent the standard deviation of 2independent experiments.
65LOCALIZATION OF pVHL
DISCUSSION
VHL protein has been localized to both the nucleus and cytosol.In a given population of VHL transfected COS-7 cells, endogenouspVHL showed a predominant nuclear, nuclear and cytosol orpredominant cytosolic staining by immunofluorescence (Duanetal., 1995a). Here we have shown that in subconfluent cultures ofRC1 cells, pVHL preferentially localizes to the nucleus, but is alsopresent in the cytoplasm in a significant percentage of cells.Accordingly, pVHL may translocate between nuclear and cytoplas-mic compartments. The post-translational transportation of nuclearproteins is an active process involving the nuclear pore complexand a NLS contained within the transported protein (Garcia-Bustoset al., 1991). The NLS mediates nuclear transport by interactingwith the nuclear pore complex and possibly cytoplasmic compo-nents. A putative NLS, PR[R/K] . . . RPRPV, has been identified, inthe first 60 aa of the human and 28 aa of the rat VHL proteins (Leeet al., 1996). Here we compared the staining patterns of differentRCC cell lines containing wild-type and a mutant form of pVHL.Our results indicate that the C-terminal region of pVHL does notcontribute to nuclear localization because A498 cells harboring aC-end truncated mutant of pVHL showed an identical cellulardistribution as the wild-type pVHL in CCF-RC1 and UMRC6 cells(Fig. 1a,b) with predominant nuclear pVHL and less intensecytoplasmic staining.
The positive pVHL foci in the nucleus are large intense punctatebodies, while the cytoplasmic staining is much more diffuse. This
distinctive nuclear staining pattern may reflect the functionalrelationship of pVHL with a specific transcriptional apparatus orthe association with other nuclear protein(s) in a specified nuclearcompartment. Different macromolecular structures in the nucleus,such as the nucleolus, spliceosome, coiled body and nucleardomain 10 (ND10, also known as the PML nuclear body), as wellas the foci for DNA replication and mRNA transcription, representcompartments for discrete nuclear functions (reviewed inStrouboulis and Wolffe, 1996). The possible association of pVHLwith spliceosomes was further investigated because of the charac-teristic ‘‘speckled’’ staining pattern of pVHL, which has beendescribed for splicing factors. However, pVHL is not colocalizedwith spliceosomes within the nucleus (Fig. 2f).
The preferential localization of pVHL to the nucleus with little orno cytoplasmic pVHL was most evident when RC1 cells reachedconfluency (Fig. 4a), while a significant amount of cytoplasmicVHL was detected in sparse or asynchronous cells. The latterobservation is consistent with previously reported results (Iliopou-los et al.,1995a) and was also seen with the glioblastoma cell lineT98G (data not shown). This led us to speculate whether thenuclear localization of pVHL is cell cycle-dependent. Nuclearprotein localization is subject to complex regulatory mechanismssince the presence of certain proteins in the nucleus is required onlyat very specific times in the cell cycle or only in response toshort-lived stimuli. One of the important mechanisms regulatingnuclear translocation is phosphorylation, either of the imported
FIGURE 4 – Correlation of subcellular localization of VHL and cell cycle. Double labeling of CCF-RC1 cells with anti-VHL(a,c,d,f)andanti-tubulin(b,e)antibodies in cultures containing approximately 90% of cells in G1/G0-phase(a–c) and approximately 55% in S-phase(d–f).(c) and( f ) are confocal images. Bars: 25 µm(a,b,d,e); 10 µm(c,f).
66 YE ET AL.
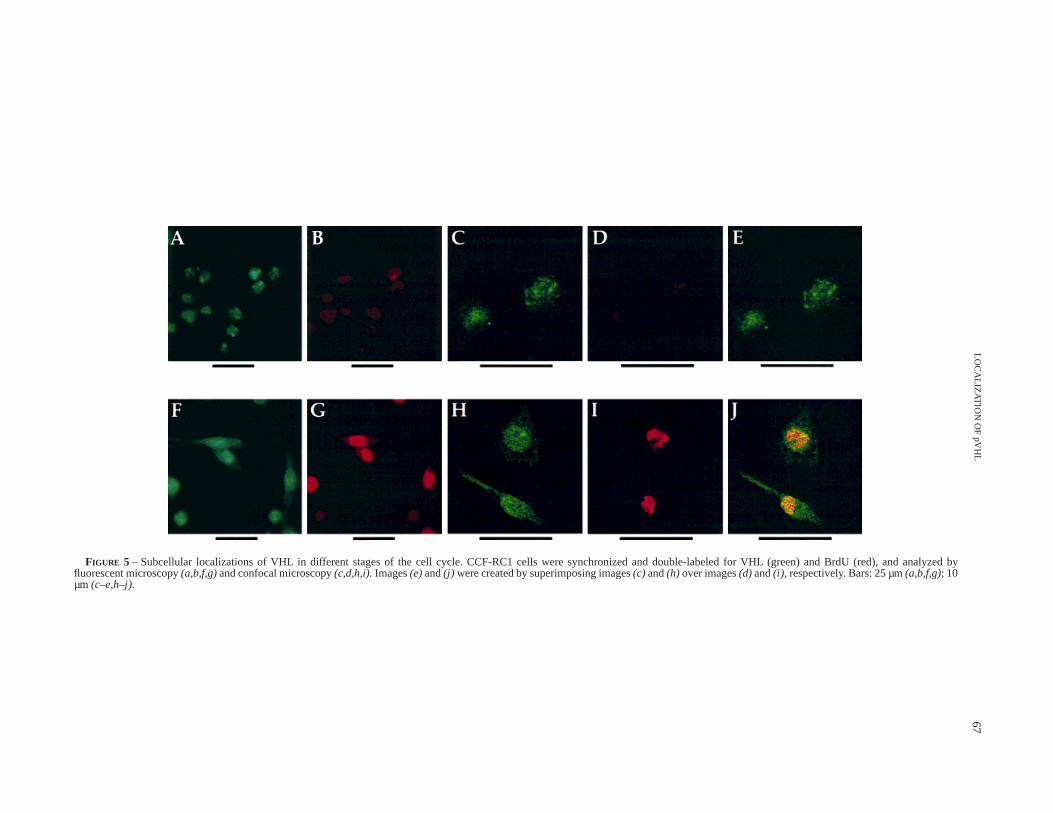
FIGURE 5 – Subcellular localizations of VHL in different stages of the cell cycle. CCF-RC1 cells were synchronized and double-labeled for VHL (green) and BrdU(red), and analyzed byfluorescent microscopy(a,b,f,g)and confocal microscopy(c,d,h,i).Images(e)and(j) were created by superimposing images(c) and(h) over images(d) and(i), respectively. Bars: 25 µm(a,b,f,g); 10µm (c–e,h–j).
67LO
CA
LIZAT
ION
OF
pVH
L

protein itself or of a protein with which it interacts. For example,phosphorylation of SW15, a yeast transcription factor, at sites nearor within the NLS by Cdc2 prevents nuclear localization (Molletal., 1991), while phosphorylation of SV40 T-antigen by caseinkinase II (CKII) can accelerate nuclear import (Rihset al.,1991).We have previously shown that phosphorylation also regulates thenuclear translocation of the WT1 protein (Yeet al.,1996). Cdc2 is ahighly conserved cell cycle-regulated protein kinase, a member ofthe cyclin-dependent kinase (CDKs) family. In a complex withcyclins, CDKs play important roles in the regulation of theeukaryotic cell cycle. CKII is also required for cell cycle progres-sion (Hannaet al., 1995) and its activity fluctuates in a cellcycle-dependent manner in mammalian cells (Carroll and Marshak,1989; DeBenedette and Snow, 1991). Thus nuclear translocation isoften closely linked to the cell cycle. For pVHL, we havedemonstrated a strong correlation between subcellular localizationand specific phase of the cell cycle. In RC1 cultures whenapproximately 90% of the cells are in the G1/G0-phase, pVHL isalmost exclusively localized to the nucleus. When synchronizedcells were allowed to progress and were double-labeled withanti-VHL and anti-BrdU antibodies, 83% of cells with cytoplasmicpVHL were in S-phase (BrdU positive) and 87% of the S-phasecells showed cytoplasmic VHL staining. Thus the nuclear localiza-tion of pVHL is cell cycle-regulated. Cell cycle-regulated nuclearlocalization has also been shown for CKII (Filhol-Cochetet al.,1994). However, CKII becomes preferentially associated with thenuclear compartment in actively growing cells, but is evenlydistributed in growth-arrested cells, making it unlikely that CKII isinvolved in pVHL translocation.
Given the proposed role of pVHL as a transcription elongationregulator, the nuclear localization is not surprising. Although thespecific gene(s) negatively regulated by pVHL remain(s) to bedescribed (with the possible exception of VEGF), it is tempting to
speculate that these are likely involved in proliferation as pVHL issequestrated in the cytoplasm potentially in an inactive form duringrapid cell proliferation or S-phase. In response to intracellular orextracellular negative signals during the G0/G1-phase, pVHL thenundergoes nuclear translocation where it functions to inhibitelongation of cell cycle progression of growth-promoting genes.This process could be activated or regulated by phosphorylation, aswith CKII and CDKs. Although the putative NLS in VHL flanks 3CKII consensus phosphorylation sites in human sequence (Leeetal., 1996) for the reasons discussed above, pVHL is unlikely to beregulated by CKII. However, a potential Cdc2 phosphorylation siteis also found in residue at 80, suggesting this kinase may be acandidate for regulating the rate of nuclear import of pVHL. Atightly regulated, cell density-dependent transport of VHL haspreviously been observed (Leeet al., 1996). However, in thisreport, the VHL protein was found predominantly in the cytoplasmin densely grown cells, whereas in sparse cultures, most of theprotein could be detected in the nucleus. The differences betweenour results and those of Leeet al. (1996) may be attributed to the 2different cell culture systems used. In the present study, we usedRCC cells which express endogenous wild-type VHL, whereasthey examined transiently transfected COS-7 cells and stablytransfected NRK and NIH 3T3 cells expressing FLAG-tagged VHLprotein. Although RC1 cells expressed wild-type pVHL, it remainspossible that there may be mutations upstream or downstream ofVHL in these cells.
Both nuclear transport and cytoplasmic sequestration are subjectto specific regulatory mechanisms. Interaction with other cytoplas-mic elements, as an alternative or in addition to phosphorylation,could be a general mechanism to retain otherwise nuclear protein inthe cytoplasm until their function is required (Hunt, 1989). pVHLhas been shown to associate with a number of cellular proteins inaddition to elongins B and C (Kishidaet al.,1995; Tsuchiyaet al.,1996). Further identification and characterization of these cellularproteins should elucidate the mechanisms involved in VHL cellulartranslocation.
ACKNOWLEDGEMENTS
We thank Dr. J. Drazba for her assistance with the confocalmicroscopy. This work was supported in part by USPHS grantDK47101 to BRGW.
REFERENCES
ASO, T., LANE, W.S., CONAWAY, J.W. and CONAWAY, R.C., Elongin (SIII): amultisubunit regulator of elongation by RNA polymerase II.Science,269,1439–1443 (1995).
BEREZNEY, R. and COFFEY, D.S., Identification of a nuclear protein matrix.Biochem. Biophys. Res. Comm.,60,1410–1417 (1974).
BLENCOWE, B.J., NICKERSON, J.A., ISSNER, R., PENMAN, S. and SHARP, P.A.,Association of nuclear matrix antigens with exon-containing splicingcomplexes.J. Cell Biol.,127,593–607 (1994).
CARROLL, D. and MARSHAK, D.R., Serum-stimulated cell growth causesoscillations in casein kinase II activity.J. biol. Chem.,264, 7345–7348(1989).
CHEN, F., KISHIDA, T., DUH, F.-M., RENBAUM, P., ORCUTT, M.L., SCHMIDT, L.and ZBAR, B., Suppression of growth of renal carcinoma cells by the vonHippel-Lindau tumor suppressor gene.Cancer Res.,55,4804–4807 (1995).
DEBENEDETTE, M. and SNOW, E.C., Induction and regulation of caseinkinase II during B lymphocyte activation.J. Immunol.,147, 2839–2845(1991).
DUAN, D.R., HUMPHREY, J.S., CHEN, D.Y.T., WENG, Y., SUKEGAWA, J., LEE,S., GNARRA, J.R., LINEHAN, W.M. and KLAUSNER, R.D., Characterization ofthe VHL tumor suppressor gene product: localization, complex formation,and the effect of natural inactivating mutations.Proc. nat. Acad. Sci.(Wash.),92,6459–6463 (1995a).
DUAN, D.R., PAUSE, A., BURGESS, W.H., ASO, T., CHEN, D.Y.T., GARRETT,K.P., CONAWAY, R.C., CONAWAY, J.W., LINEHAN, W.M. and KLAUSNER,
R.D., Inhibition of transcription elongation by the VHL tumor suppressorprotein.Science,269,1402–1406 (1995b).
FILHOL-COCHET, O., LOUE-MACKENBACH, P., COCHET, C. and CHAMBAZ ,E.M., Casein kinase 2 and the cell response to growth factors.Cell mol.Biol. Res.,40,529–537 (1994).
GARCIA-BUSTOS, J., HEITMAN, J. and HALL , M.N., Nuclear protein localiza-tion. Biochim. biophys. Acta,1071,83–101 (1991).
GNARRA, J.R., TORY, K., WENG, Y., SCHMIDT, L., WEI, M.H., LATIF, F., LI, H.,LATIF, F., CHEN, F. and DUH, F.M., Mutations of the VHL tumour suppressorgene in renal carcinoma.Nature (Genet.),7, 85–90 (1994).
GROSSMAN, H.B., WEIDEMEYER, G. and REN, L., Human renal carcinoma:characterization of five new cell lines.J. surg. Oncol.,28,237–244 (1985).
HANNA, D.E., RETHINASWAMY, A. and GLOVER, C.V.C., Casein kinase II isrequired for cell cycle progression during G1 and G2/M in Saccharomycescerevisiae. J. biol. Chem.,43,25905–25914 (1995).
HASHIMURA, T., TUBBS, R.R., CONNELLY, R., CAULFIELD, M.J., TRINDADE,C.S., MCMAHON, J.T., GALETTI, T.P., EDINGER, M., SANDBERG, A.A., DALCIN, P., SAIT, S.J. and PONTES, J.E., Characterization of two cell lines withdistinct phenotypes and genotypes established from a patient with renal cellcarcinoma.Cancer Res.,49,7064–7071 (1989).
HERMAN, J.G., LATIF, F., WENG, Y., LERMAN, M.I., ZBAR, B., LIU, S., SAMID ,D., DUAN, D.-S.R., GNARRA, J.R., LINEHAN, W.M. and BAYLIN , S.B.,Silencing of the VHL tumor-suppressor gene by DNA methylation in renalcarcinoma.Proc. nat. Acad. Sci. (Wash.),91,9700–9704 (1994).
TABLE I – DOUBLE STAINING OF CCF-RC1 CELLS WITH ANTI-VHLAND ANTI-BrdU ANTIBODIES
Cytosolic VHL positive(208 cells)
BrdU positive(214 cells)
BrdU (1) BrdU (2) Cytosolic VHL (1) Cytosolic VHL (2)83% (173) 17% (35) 87% (187) 13% (27)
68 YE ET AL.

HUNT, T., Cytoplasmic anchoring proteins and the control of nuclearlocalization.Cell, 59,949–951 (1989).ILIOPOULOS, O., GRAY, S., KIBEL, A. and KAELIN, W.G., Characterization ofpVHL, the product of the von Hippel-Lindau (VHL) disease gene.Proc.Amer. Ass. Cancer Res.,36,194 (1995a).ILIOPOULOS, O., KIBEL, A., GRAY, S. and KAELIN, W.G., JR., Tumoursuppression by the human von Hippel-Lindau gene product.Nature (Med.),8, 822–826 (1995b).KIBEL, A., ILIOPOULOS, O., DECAPRIO, J.A. and KAELIN, W.G., Binding ofthe von Hippel-Lindau tumor suppressor protein to elongins B and C.Science,269,1444–1446 (1995).KISHIDA, T., STACKHOUSE, T.M., CHEN, F., LERMAN, M.I. and ZBAR, B.,Cellular proteins that bind the von Hippel-Lindau disease gene product:mapping of binding domains and the effect of missense mutations.CancerRes.,55,4544–4548 (1995).LARSSON, S., CHARLIEU, J.-P., MIYAGAWA , K., ENGELKAMP, D., RASSOULZA-DEGAN, M., ROSS, A., CUZIN, F., VAN HEYNINGEN, V. and HASTIE, N.D.,Subnuclear localization of WT1 in splicing or transcription factor domainsis regulated by alternative splicing.Cell, 81,391–401 (1995).LATIF, F. and 30OTHERS, Identification of the von Hippel-Lindau diseasetumor suppressor gene.Science,260,1317–1320 (1993).LEE, S., CHEN, D.Y.T., HUMPHREY, J., GNARRA, J.R., LINEHAN, W.M. andKLAUSNER, R.D., Nuclear/cytoplasmic localization of the von Hippel-Lindau tumor suppressor gene product is determined by cell density.Proc.nat. Acad. Sci. (Wash.),93,1770–1775 (1996).MAHER, E.R., ISELIUS, L., YATES, J.R., LITTLER, M., BENJAMIN, C., HARRIS,R., SAMPSON, J., WILLIAMS , A., FERGUSON-SMITH, M.A. and MORTON, N.,Von Hippel-Lindau disease: a genetic study.J. med. Genet.,28, 443–447(1991).MARTINY-BARON, G. and MARME, D., VEGF-mediated tumour angiogen-esis: a new target for cancer therapy.Curr. Opin. Biotechnol.,6, 675–680(1995).MOLL, T., TEBB, G., SURANA, U., ROBITSCH, H. and NASMYTH, K., The roleof phosphorylation and the CDC28 protein kinase in cell cycle-regulatednuclear import of theS. cerevisiaetranscription factor SW15.Cell, 66,743–758 (1991).MUKHOPADHYAY, D., KNEBELMANN, B., COHEN, H.T., ANANTH, S. andSUKHATME, V.P., The von Hippel-Lindau tumor suppressor gene product
interacts with Sp1 to repress vascular endothelial growth factor promoteractivity. Mol. cell. Biol.,17,5629–5639 (1997).
NYMAN , U., HALLMAN , H., HADLACZKY, G., PETTERSSON, I., SHARP, G. andRINGERTZ, N.R., Intranuclear localization of snRNP antigens.J. Cell Biol.,102,137–144 (1986).
OSAKA, M., NAGASHIMA, Y., KITAMURA , H., LATIF, F., ZBAR, B., LERMAN,M.I. and YAO, M., Frequent somatic mutations and loss of heterozygosity ofthe von Hippel-Lindau tumor suppressor gene in primary human renal cellcarcinomas.Cancer Res.,54,2852–2855 (1994).
PAL, S., CLAFFEY, K.P., DVORAK, H.F. and MUKHOPADHYAY, D., The vonHippel-Lindau gene product inhibits vascular permeability factor/vascularendothelial growth factor expression in renal cell carcinoma by blockingprotein kinase C pathways.J. biol. Chem.,272,27509–27512 (1997).
PAUSE, A., LEE, S., WORRELL, R.A., CHEN, D.Y., BURGESS, W.H., LINEHAN,W.M. and KLAUSNER, R.D., The von Hippel-Lindau tumor-suppressor geneproduct forms a stable complex with human CUL-2, a member of the Cdc53family of proteins.Proc. nat. Acad. Sci. (Wash.),94,2156–2161 (1997).
RIHS, H.-P., JANS, D.A., FAN, H. and PETERS, R., The rate of nuclearcytoplasmic protein transport is determined by the casein kinase II siteflanking the nuclear localization sequence of the SV40 T-antigen.EMBO J.,10,633–639 (1991).
SIEMEISTER, G., WEINDEL, K., MOHRS, K., BARLEON, B., MARTINY-BARON,G. and MARME, D., Reversion of deregulated expression of vascularendothelial growth factor in human renal carcinoma cells by von Hippel-Lindau tumor suppressor protein.Cancer Res.,56,2299–2301 (1996).
SPECTOR, D.L., FU, X. and MANIATIS, T., Associations between distinctpre-mRNA splicing components and the cell nucleus.EMBO J., 10,3467–3481 (1991).
STROUBOULIS, J. and WOLFFE, A.P., Functional compartmentalization of thenucleus.J. Cell. Sci.,109,1991–2000 (1996).
TSUCHIYA, H., ISEDA, T. and HINO, O., Identification of a novel protein(VBP-1) binding to the von Hippel-Lindau (VHL) tumor suppressor geneproduct.Cancer Res.,56,2881–2885 (1996).
YE, Y., RAYCHAUDHURI, B., GURNEY, A., CAMPBELL, C.E. and WILLIAMS ,B.R.G., Regulation of WT1 by phosphorylation: inhibition of DNAbinding, alteration of transcriptional activity and cellular translocation.EMBO J.,15,5606–5615 (1996).
69LOCALIZATION OF pVHL





![Von Hippel Lindau Disease [VHL]: Magnetic Resonance Imaging](https://static.fdocuments.net/doc/165x107/620636d38c2f7b17300580b0/von-hippel-lindau-disease-vhl-magnetic-resonance-imaging.jpg)












