
Studies on nebulin in amphioxus striated muscle
54
Studies on nebulin in amphioxus striated muscle July 2009 Akira Hanashima Graduate School of Science CHIBA UNIVERSITY
Transcript of Studies on nebulin in amphioxus striated muscle

Studies on nebulin in amphioxus striated muscle
July 2009
Akira Hanashima
Graduate School of Science
CHIBA UNIVERSITY

(千葉大学学位申請論文)
Studies on nebulin in amphioxus striated muscle
2009 年 7 月
千葉大学大学院理学研究科
地球生命圏科学専攻生物学コース
花島 章

1
Contents
Chapter 1. Primary structure, localization and binding proteins of
the C-terminus of amphioxus nebulin
1. Abstract---------------------------------------------------------------3
2. Introduction----------------------------------------------------------4
3. Materials and Methods-------------------------------------------5
4. Results-----------------------------------------------------------------11
5. Discussion------------------------------------------------------------15
6. Conclusion------------------------------------------------------------17
7. References------------------------------------------------------------18
8. Figures-----------------------------------------------------------------22
Chapter 2. Structure of the amphioxus nebulin gene and
evolution of the nebulin family genes
1. Abstract----------------------------------------------------------------31
2. Introduction-----------------------------------------------------------32
3. Materials and Methods--------------------------------------------34
4. Results------------------------------------------------------------------35
5. Discussion--------------------------------------------------------------39
6. Conclusion-------------------------------------------------------------43
7. References-------------------------------------------------------------44
8. Figures------------------------------------------------------------------47

2
Chapter 1
Primary structure, localization and binding proteins of
the C-terminus of amphioxus nebulin

3
1. Abstruct
Nebulin is a large protein located along the thin filaments of vertebrate
skeletal muscle. I isolated nebulin cDNA from the striated muscle of
cepharochordate (invertebrate) amphioxus. Although the sequence homology
is 26% with the human, the C-terminal of amphioxus nebulin has similar
structural motifs of 35 amino acid nebulin repeats and an SH3 domain. With
indirect immunofluorescence analysis by using specific antibody raised to
the bacteria produced recombinant peptide, I confirm that this nebulin
fragment is located in the Z-line of the sarcomere similar to human nebulin.
Pull-down and co-sedimentation assays in vitro showed that the C-terminal
region binds to actin, α-actinin and connectin (titin). These results suggest
that the C-terminal region of amphioxus nebulin plays a role in maintaining
striate muscle structure similar to that of human. This is the first report that
nebulin is not vertebrate-only protein but chordate protein.

4
2. Introduction
Nebulin is a large, 773-kDa protein found in vertebrate skeletal muscles
(Wang, 1982; McElhinny et al., 2003). The primary structure of human
nebulin consists of a N-terminal glutamic acid-rich domain, followed by 185
(M1–185) contiguous nebulin repeats of approximately 35 amino acids with a
central SDXXYK consensus sequence, a serine-rich region and a C-terminal
Src homology 3 (SH3) domain (Labeit and Kolmerer, 1995). Nebulin repeats
(M9–162) in the I-band region of the sarcomere consist of sets of
seven-repeats. These seven-repeats comprise approximately 245 amino-acid
residues (i.e., 35 amino-acid residues per repeat x 7 repeats) to form a
super-repeat, there are 22 super-repeats (SR1–22).
Within the sarcomere, the N-terminal region is localized in the pointed end
of the thin filaments, the central region along the thin filaments and the
C-terminal region in the Z-line (Wang and Wright, 1988). Binding assays
have revealed that each nebulin repeat (SDXXYK) binds to actin (Chen et al.,
1993, Lukoyanova et al., 2002). Furthermore, the nebulin repeat 1–3, repeat
163–170, repeat 185–SH3 domain and the SH3 domain bind to tropomodulin,
desmin, connectin (also called titin), and myopalladin and b-actinin (also
called CapZ), respectively (Bang et al., 2001; Bang et al., 2002; Jin and Wang,
1991; McElhinny et al., 2001; Witt et al., 2006).
Existence of nebulin in non-vertebrate has not been reported; however,
sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE)
revealed that bands corresponding to the size of nebulin are present in
amphioxus striated muscle (Hu et al., 1986; Locker and Wild, 1986). In 1997,

5
Kimura et al. reported that one of several nebulin antibodies reacted with a
750-kDa protein found in amphioxus striated muscle, and in 1999, Fock and
Hinssen prepared an antibody against this protein and showed that it was
located in the I-Z-I region of the sarcomere. However, because these findings
could have resulted from a nebulin-mimicking epitope or simply from
cross-immunoreactivity with an SH3 domain protein, this 750-kDa protein
was not proven to be nebulin.
In this study, for the first time, I cloned the C-terminal region of the
750-kDa protein identified in amphioxus striated muscle, examined its
functional properties and compared it to vertebrate nebulin.
3. Materials and Methods
3.1. cDNA cloning, sequencing and analysis
Approximately 500 specimens of adult amphioxus (Branchiostoma
belcheri) were collected in the Enshu-Nada Sea, Japan (Kubokawa et al.,
1998). Nerve cord and muscle tissues were used to produce a cDNA library
that was constructed in lambda ZAP II (Stratagene, La Jolla, CA, USA)
using oligo (dT) primers. The fragments encoding the homologue of nebulin
were obtained from the cDNA library using an EST-analysis system
established by the sequencing laboratory in RIKEN, Kobe, Japan (Mineta et
al., 2003). Three clones containing the nebulin homologues were isolated
from approximately 7000 clones of the cDNA library. The domains were
aligned with reference to Labeit and Kolmerer (1995). The sequence analysis
and homology search were performed using GENETYX-WIN Ver. 4.0.4

6
(Genetyx, Tokyo, Japan).
3.2. SDS-PAGE and immunoblotting
A piece of freshly excised amphioxus striated muscle was dissolved in 5
volumes of a solution containing 10% SDS, 40 mM dithiothreitol, 10 mM
EDTA and 100 mM Tris-HCl (pH 8.0). The solution with the dissolved muscle
was boiled for 3 min and clarified by centrifugation at 13,500rpm for 10 min.
The supernatant proteins were separated by SDS-PAGE using 2–6%
gradient polyacrylamide gels (acrylamide:methylenebisacrylamide, 30:1.5;
w/w) or 2–15% gradient polyacrylamide gels
(acrylamide:methylenebisacrylamide, 30:0.8; w/w) (Laemmli, 1970). The
protein bands were electrically transferred onto a nitrocellulose membrane
(Towbin et al., 1979) and treated with primary antibody and horseradish
peroxidase-conjugate anti-rabbit IgG secondary antibody (Code number:
P-0399, Dako, Carpinteria, CA, USA).
3.3. Antibody production
The cDNA fragment of amphioxus nebulin (1007–1848 bp of clone 5361)
was generated by restriction enzyme digestion (SmaI/SalI) and inserted into
the PvuII/SalI site of the pGEX4T-3 vector (GE Healthcare, Piscataway, NJ,
USA). Recombinant GST-tagged protein was expressed in Escherichia coli (E.
coli) XL1 Blue-MRF’ with 3 l of LB + ampicillin culture medium under the
conditions of OD600 = 0.5, IPTG 0.1mM, 37°C and 3 h. The bacteria were
harvested by centrifugation (4000rpm, 4°C, 10min), dialysed with 120ml

7
PBS, treated with 80ml sample buffer, boiled for 10min and subjected to 227
sheets of 10% SDS-PAGE. The bands of recombinant protein were excised
and the recombinant protein was electrically extracted from the gels into a
running buffer solution (0.1% SDS, 25mM Tris, 192mM glycine) at 400mA for
16h. The protein was dialysed with PBS, and GST was deleted using
thrombin (Sigma-Aldrich Japan, Tokyo, Japan) (1/1000 volume of
recombinant protein solution) at 22°C for 16h. The solution was added to the
sample buffer and subjected to 88 sheets of 12.5% SDS-PAGE. The nebulin
fragment without GST was electrically extracted from the gel, dialyzed with
PBS, conjugated with an equal volume of Freund’s incomplete adjuvant
(Difco Laboratories, Detroit, MI, USA) and injected into three times (0.25,
0.9 and 0.1mg protein) into a rabbit. The antiserum was separated from
blood by centrifugation.
3.4. Immunofluorescence microscopy
After skinning the amphioxus specimens and collecting the muscle tissue
under a stereoscopic microscope, the myofibers were stretched with relaxing
buffer (50 mM KCl, 10 mM EGTA, 10 mM NaPO4, 3 mM ATP and 0.5%
Triton X-100; pH 7.5) and fixed with buffer (3.5% formaldehyde, 45 mM KCl,
9 mM EGTA, 9 mM NaPO4, 2.7 mM ATP and 0.45% Triton X-100; pH 7.5) for
1 min. The tissue was homogenized five times for 2 s each in phosphate
buffer saline (PBS) containing 0.5 mM leupeptin using Ultra-turrax T-25
(IKA-Labortechnik, Staufen, Germany) and then fixed on a glass slide. The
samples were fixed with 3.7% formaldehyde in PBS for 15min, washed two

8
times with PBS for 5min and blocked with 1% bovine serum albumin(BSA)
in PBS for 15min. The fixed samples were stained with amphioxus nebulin
(1:50) and a-actinin A7811 (1:800; Sigma-Aldrich Japan, Tokyo, Japan)
antibodies for 12h, washed three times with PBS for 10 min and reacted with
Alexa 488 (1:2500) and Alexa 546 (1:4000) conjugated secondary antibodies
(Invitrogen, Carlsbad, CA, USA). The samples were then washed three times
with PBS for 15min, fixed with 3.7% formaldehyde in PBS for 15 min and
washed with PBS for 15 min. Anti-fador was added to the samples and they
were covered with a cover glass. Fluorescence was observed with
fluorescence microscope (Zeiss Axioskop 2 plus; Carl Zeiss, Oberkochen,
Germany).
3.5. Protein preparation
The cDNA fragments of amphioxus nebulin repeats 3–9 (AN3–9) and the
unique region of the SH3 domain (U–SH3) generated by restriction enzyme
digestion were cloned into the pGEX6P series (GE Healthcare, Piscataway,
NJ, USA). The recombinant proteins tagged with GST were expressed in E.
coli BL21 (DE3) pLysS in 250ml LB containing ampicillin. The conditions
were OD600 = 0.5, IPTG 0.5 mM, 37°C and 3 h. The bacteria were harvested
by centrifugation (3000rpm, 4°C, 10min). The bacteria with the GST-fusion
proteins were dissolved in PBS (140 mM NaCl, 10 mM Na2HPO4, 2.7 mM
KCl and 1.8 mM KH2PO4; pH7.3), frozen at –80°C for 20 min, sonicated at
output 2 for 1min (TOMY UD-200; Tomy Seiko, Tokyo, Japan) and
centrifuged at 13,000rpm for 30min at 4°C. The supernatant was loaded onto

9
a 2ml glutathione Sepharose 4B column (GE Healthcare, Piscataway, NJ,
USA), washed with PBS and eluted with 10mM glutathione (reduced) in 50
mM Tris-HCl (pH 8.0). For the co-sedimentation assay, GST was deleted
from GST–AN3–9 on a column using PreScission Protease (GE Healthcare,
Piscataway, NJ, USA) with buffer (50 mM Tris, 150 mM NaCl, 1 mM EDTA
and 1 mM DTT; pH7.0) at 4°C for 12 h. Actin was prepared from acetone
powder of rabbit skeletal muscle using the protocol described by Spudich and
Watt (1971). G-actin was polymerized in 0.1 M KCl. a-Actinin was prepared
from rabbit skeletal muscle by the method of Goll et al. (1972). Connectin
was prepared according to Kimura et al. (1992).
3.6. Far-western blot assay
GST-fusion proteins mixed with GST, and the bacteria with the His-tag
proteins were dissolved in SDS sample buffer, subjected to SDS-PAGE and
transferred onto a nitrocellulose membrane. The membranes were blocked
with 20 mg/ml BSA in buffer (80 mM KCl, 2mM MgCl2, 10 mM Hepes, 1 mM
DTT and 2% Triton X-100, pH 7.3, for α-actinin or connectin) or 0.5% casein
in buffer (2 mM Tris, 0.01% NaN3, 0.1 mM CaCl2 and 100 mM KCl, pH8.0,
for F-actin) at 25°C for 1 h. The membranes were incubated with F-actin (0.3
mg/ml), α-actinin (10 µg/ml) or connectin (80 µg/ml) in each buffer at 25°C for
1.5 h. After washing two times with TBS for 10 min, binding was detected
with anti-actin (Sigma A4700; Sigma-Aldrich Japan), anti α-actinin (Sigma
A7811; Sigma-Aldrich Japan) or anti-connectin (Pc1200) (Kimura et al.,
1992) antibody.

10
3.7. Yeast two hybrid assay
The cDNA fragments coding the repeat sequence 1–9 of amphioxus nebulin
or each of the regions of the chicken a-actinin 2 were generated by restriction
enzyme digestion and inserted into pGBT9 or pGAD424 vector (Clontech
Labratories, Palo Alto, CA, USA). The nebulin clone was co-transformed with
the a-actinin clones into Saccharomyces cerevisiae Y170. The transformants
grown on –Leu, –Trp plates were transferred on to a nitrocellulose
membrane and their b-galactosidase activity was examined.
3.8. Co-sedimentation assay
F-actin (2.5 mM) or BSA(5 mM) was mixed with AN3–9 (12.5 mM) in 500 ml
buffer (2 mM Tris, 0.1 mM CaCl2, 0.1M KCl and 0.01% NaN3; pH 8.0) at 25°C
for 1 h. The mixture was centrifuged (13,000 rpm, 4°C, 30 min), and the
supernatant and pellet were diluted with SDS sample buffer and subjected
to SDS-PAGE.
3.9. GST pull down assay
GST–AN3–9 (2mg for a-actinin pull down) or GST–U–SH3 (2mg for
connectin pull-down) were bound to 20 ml glutathione Sepharose 4B beads
according to the manufacture’s method. The beads were washed two times
with 150ml buffer (80 mM KCl, 2 mM MgCl2, 10 mM HEPES, 1 mM DTT and
2% TritonX-100; pH7.3), and bound with a-actinin (4 mg, 20 ml) or connectin
(5 mg, 150 ml) in buffer at 4°C for 1h. They were washed four times with 150
ml buffer, dissolved in 20 ml SDS sample buffer, and subjected to SDS-PAGE.

11
The gels were stained with Coomassie Brilliant Blue.
4. Results
4.1. Primary structure of amphioxus nebulin C-terminus
Three clones were obtained from a mixed cDNA library of amphioxus
muscle tissues and nerve cord. Sequencing revealed a 2527-bp sequence
containing a poly-A tail and a stop codon, suggesting that it contains the
C-terminal region of the protein (DDBJ accession no. AB244086). The
amphioxus sequence contains 13 nebulin repeats and an SH3 domain with
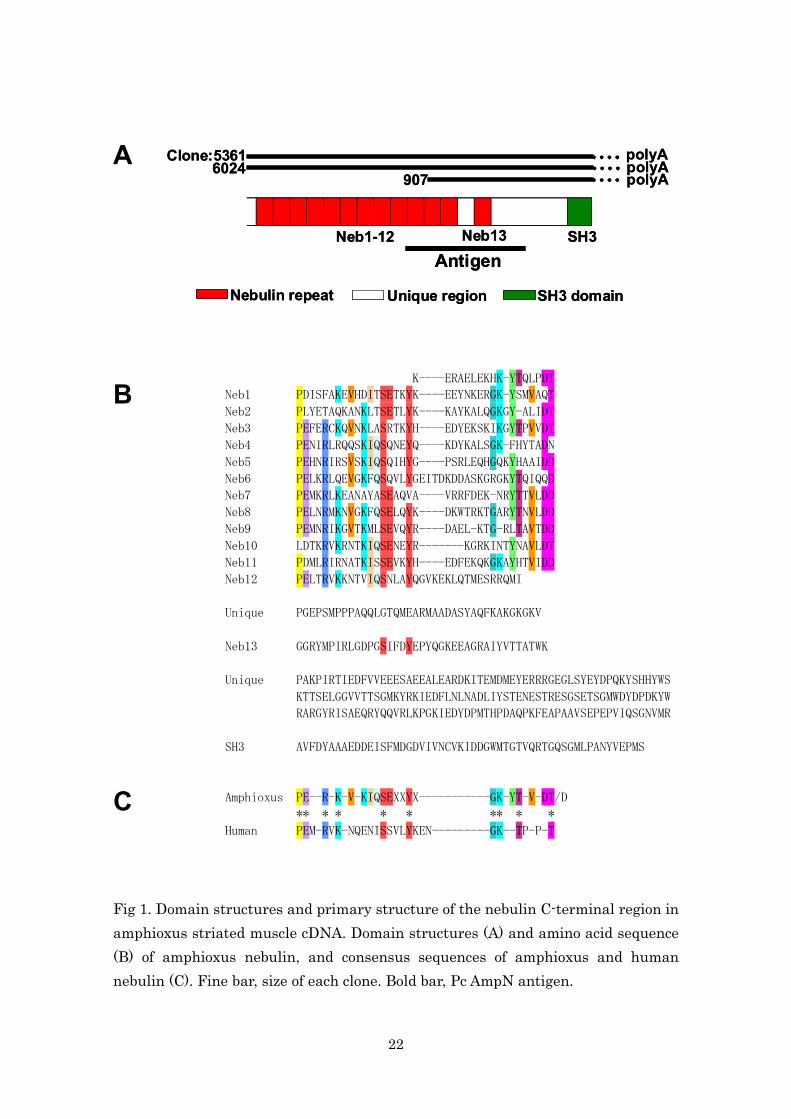
unique regions between these modules (Fig. 1A).
Amphioxus nebulin repeats were assigned the numbers Neb-1–13 for
descriptive purposes. The consensus sequence observed in each nebulin
repeat was a PEXXRXKXVXKIQ motif in the N-terminal region and a
GKXYTXVXDT/D motif in the C-terminal region, as well as a SE and a Y in
the SEXXYX-motif (Fig. 1B,C). P1, E2, R5 and K7 in the N-terminal and S14
and Y18 in the SEXXYX motif of the amphioxus nebulin repeats correspond
to the human C-terminal nebulin repeats (M172–185) (Fig. 1C). The
SSVLYKEN-box, which is present in human M174–181, was not found in the
amphioxus nebulin repeats.
4.2. Comparison between amphioxus and vertebrate nebulins
The amphioxus nebulin repeats shared an approximately 26% homology
with human C-terminal nebulin repeats. I predicted the secondary structure
of the amphioxus nebulin repeat region using the Multivariate Linear

12
Regression Combiner secondary structure prediction program
(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_mlrc.ht
ml). The secondary structure was composed of 48% a-helix, 0% b-structure
and 47% random coil. The composition was similar to that of the secondary
structure of the C-terminal region of the human nebulin repeat (41% a-helix,
0% b-structure and 52% random coil) predicted using the same program.
Calculation of the isoelectric points (pIs) based on the amphioxus C-terminal
amino acid sequence revealed pIs of 5.6 and 7.1–10.8 (total pI=9.9) for Neb-1
and Neb-2–13, respectively indicating that the pIs of the amphioxus nebulin
repeats are in accordance with those of the human nebulin repeats (pI = 5.6
for M172 and pI = 10.0 for M173–185) (Labeit and Kolmerer, 1995) (Fig. 2).
The PGSIFDYEP of the last nebulin repeat (Neb-13) in amphioxus (Fig.
1B) is conserved across species, and corresponds to the last human nebulin
repeat M185 (X83957) (Labeit and Kolmerer, 1995), chicken nebulin repeat
M65 (AB024330) (Suzuki et al., 2000), the zebrafish nebulin-like protein
(AL974314) and the nebulin-like protein (nebulette) of vertebrate cardiac
muscle (Y16350) (Millevoi et al., 1998; Moncman and Wang, 1995).
Furthermore, when I compared the C-terminal SH3 domain of amphioxus
nebulin to human (X83957), chicken (AB024330) and zebrafish (AL974314), I
observed a high homology (approximately 65%) with human nebulin (Fig. 2),
revealing that the amino acid sequence of the SH3 domain was well
conserved across species. However, two unique regions, which are between
Neb-12 and Neb-13 and between Neb-13 and the SH3 domain, showed no
homology with any region of the human nebulin (Fig. 2).

13
The domain structure, secondary structure and the pIs of the C-terminal
region of amphioxus nebulin are similar to those of vertebrate nebulin.
4.3. Localization of the amphioxus nebulin C-terminal region in the
sarcomere
To confirm that the cDNA was from amphioxus nebulin and to localize the
C-terminal region within the sarcomere, I prepared an antibody (PcAmpN)
using the recombinant protein from Neb 10 to the former half of the unique
region as an antigen (Figs. 1A, 3A). Immunoblot analysis showed that the
antibody reacted only with the 750-kDa band and not with other proteins
such as nebulette (107 kDa) (Fig. 3B,C). This confirmed the sequence to be
that of amphioxus nebulin cDNA.
Double immunostaining of amphioxus myofibrils with PcAmpN and an
a-actinin monoclonal antibody revealed that the C-terminal region of
amphioxus nebulin is localized to the Z-line of the sarcomere and co-localizes
with a-actinin (Fig. 4).
4.4. Binding of amphioxus nebulin to F-actin, a-actinin and connectin
It is known that vertebrate nebulin is localized along thin filaments and
the nebulin repeats binds to actin. To clarify whether amphioxus nebulin
repeats bind to actin, I examined the binding of nebulin repeats 3–9 to
F-actin by far-western blot analysis and a co-sedimentation assay. The
far-western blot analysis showed that F-actin binds to the amphioxus
nebulin repeats, similar to human nebulin repeats (Fig. 5A). Furthermore,

14
the results of the co-sedimentation assay showed that F-actin alone did not
precipitate after centrifugation at 13,000 rpm for 30 min, however, F-actin
co-precipitated with the nebulin repeats, indicating that amphioxus nebulin
repeats bind to F-actin (Fig. 5B).
Next, I examined binding of the amphioxus nebulin repeats with purified
a-actinin by far-western blot analysis, a yeast two-hybrid assay and a GST
pull-down assay. The results of the far-western blot analysis showed that
a-actinin bind to nebulin repeats 3-9 (Fig. 6A). I also created cDNA
fragments encoding each region of a-actinin 2 in the striated muscles, and
examined their binding ability to repeats 1–9 of amphioxus nebulin using the
yeast two-hybrid system. The 343–804 amino-acid fragments of a-actinin
showed b-galactosidase activity, while the fragments without this region did
not (Fig. 6B), demonstrating the direct binding of 343–804 amino-acid
fragments of a-actinin to amphioxus nebulin. Furthermore, the results of the
GST pull-down assay showed that a-actinin precipitated when added to the
GST-fusion protein with nebulin repeats 3–9 (lane 5 in Fig. 6C), but not
when it was added only to GST (lane 6 in Fig. 6C). The results of a control
experiment showed that amphioxus nebulin repeats 3–9 do not bind to BSA
(Fig. 6D).
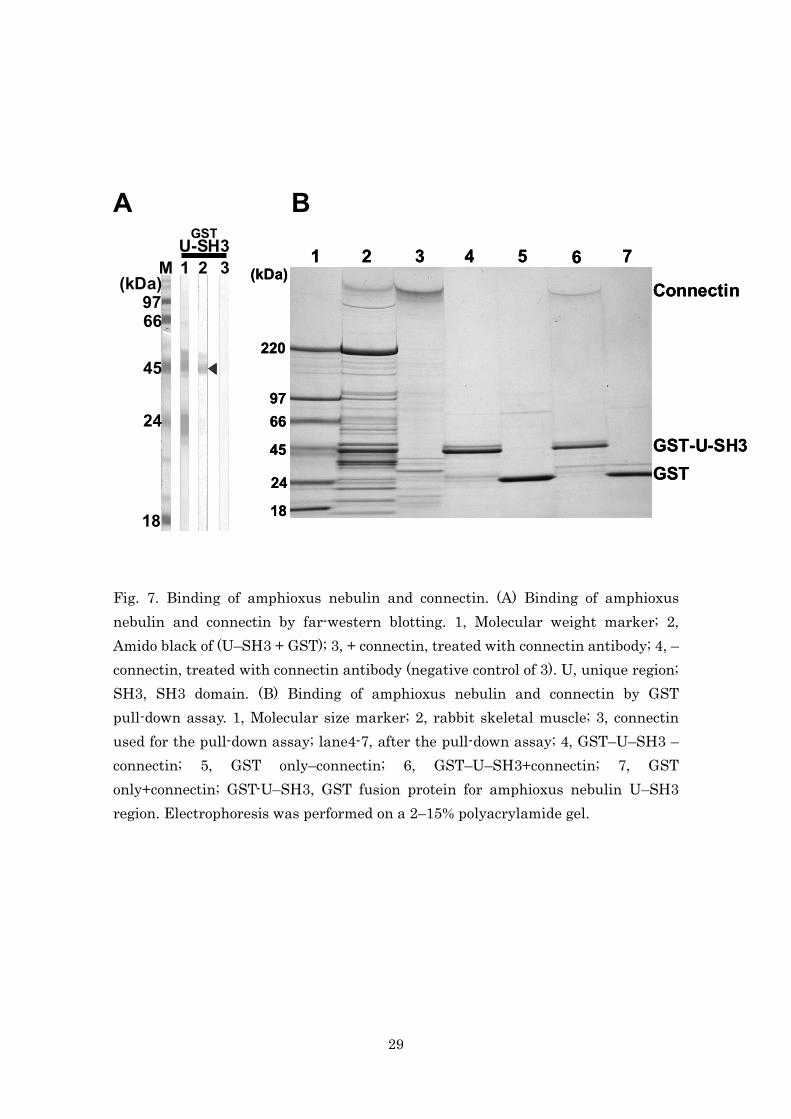
I examined binding of the SH3 domain from the unique region (U–SH3) in
amphioxus nebulin with purified rabbit connectin by far-western blot
analysis and a pull-down assay. The results of the far-western blot analysis
showed that connectin binds to the U–SH3 domain (Fig. 7A). Furthermore,
the result of the GST pull-down assay revealed that connectin binds to the

15
U–SH3 domain of amphioxus nebulin, but not to GST alone (Fig. 7B).
5. Discussion
The present study elucidated the primary structure, localization and
binding proteins of the C-terminal region of amphioxus nebulin. These
results confirm earlier findings that nebulin is expressed in amphioxus and
suggest that it plays a role similar to that of vertebrate nebulin.
In human nebulin, repeats M167–172 are negatively charged and repeats
M173–185 are positively charged under physiological conditions. Similarly,
in amphioxus nebulin, Neb-1 is negatively charged and Neb-2–13 is
positively charged. Thus, the charge transition is conserved between human
and amphioxus nebulins. Although the function of the charge transition
region is unknown, its localization within or around the Z-line suggests that
it is involved in the formation and structural maintenance of the Z-line.
a-Actinin is the main component of the Z-line of the sarcomere, and is also
the location of the N-terminus of connectin (titin) (Tskhovrebova and Trinick,
2003). I showed that amphioxus nebulin bind to a-actinin and connectin
(titin) and that the C-terminal region of amphioxus nebulin is localized in
the Z-line of the sarcomere (Fig. 4). These results suggest that amphioxus
nebulin maintains the structure of the Z line by binding to a-actinin and
connectin, similar to the C-terminal region of human nebulin.
I predicted that the secondary structure of the amphioxus nebulin repeat
region was 48% a-helix and 0% b-structure. The secondary structure was
similar to that of the C-terminal region of the human nebulin repeat. This is

16
because both amphioxus and human nebulins have the consensus sequences
PEXXRXK at the N terminal, SXXXYX in the middle and GKXXTXXXXT at
the C-terminal of each repeat. As a result, amphioxus nebulin functions
similar to human nebulin, even though there is only a 26% homology at the
sequence level. Because the helical part of the protein can present the
conserved surface and interact to other proteins (McLachlan and Karn,
1982).
The thin filaments in the striated muscle of arthropods and molluscs do
not have a uniform length of 1 mm as in vertebrate skeletal muscle, whereas
the thin filaments of amphioxus striated muscle have a uniform length of 1
mm (Hagiwara et al., 1971). Although amphioxus is not a vertebrate and is
the phylogenetically lowest chordate, its nebulin has a similar function to
vertebrate nebulin, which might explain the uniform 1 μm size of the thin
filaments in amphioxus striated muscle. Taken together with my result of
the interaction of amphioxus nebulin with actin, a-actinin and connectin, the
thin filaments in amphioxus striated muscle might be maintained in a
manner identical or similar to that in the thin filaments of vertebrate
skeletal muscle.
The existence of nebulin protein have not be seen in invertebrate without
amohioxus. In fact, nebulin gene cannot be seen in Drosophila melanogaster,
Caenorhabditis elegans and Strongylocentrotus purpuratus genome.
However urochordate ciona, which is evolved from cepharochordate, have
nebulin gene in its genome. These insights indicate chordate (vertebrate,
urochordate and cepharochordate) have nebulin but the other creatures don’t

17
have nebulin.
6. Conclusion
Invertebrate amphioxus have nebulin and its structure and functions are
similar to human nebulin. These insights show that nebulin is not
vertebrate-only protein but chordate protein. Detailed elucidation of the
differences of thin filaments between chordate and non-chordate will lead to
a better understanding of the nebulin function such as length regulator of
the thin filaments.

18
7. References
Bang, M. L., Mudry, R. E., McElhinny, A. S., Trombitas, K., Geach, A. J.,
Yamasaki, R., Sorimachi, H., Granzier, H., Gregorio, C. C. and Labeit, S.
(2001). Myopalladin, a novel 145-kilodalton sarcomeric protein with multiple
roles in Z-disc and I-band protein assemblies. J. Cell Biol. 153, 413-427.
Bang, M. L., Gregorio, C. and Labeit, S. (2002) Molecular dissection of the
interaction of desmin with the C-terminal region of nebulin. J. Struct. Biol.
137, 119-127.
Chen, M. J., Shih, C. L. and Wang, K. (1993) Nebulin as an actin zipper. A
two-module nebulin fragment promotes actin nucleation and stabilizes actin
filaments. J. Biol. Chem. 268, 20327-20334.
Fock, U. and Hinssen, H. (1999) Identification and localisation of nebulin as
a thin filament component of invertebrate chordate muscles. J Comp Physiol
169, 555-560.
Goll, D. E., Suzuki, A., Temple, J. and Holmes, G. R. (1972) Studies on
purified a-actinin. I. Effect of temperature and tropomyosin on the
a-actinin-F-actin interaction. J. Mol. Biol. 67, 469-488.
Hagiwara, S., Henkart, P. M. and Kidokoro, Y. (1971) Excitation-contraction
coupling in amphioxus muscle cells. J. Physiol. 219, 233-251.
Hu, D. H., Kimura, S. and Maruyama, K. (1986) Sodium dodecyl sulfate gel
electrophoresis studies of connectin-like high molecular weight proteins of
various types of vertebrate and invertebrate muscles. J. Biochem. 99,
1485-1492.
Jin, J. P. and Wang, K. (1991) Cloning, expression, and protein interaction of

19
human nebulin fragments composed of varying numbers of sequence
modules. J. Biol. Chem. 266, 21215-21223.
Kimura, S., Matsuura, T., Ohtsuka, S., Nakauchi, Y., Matsuno, A. and
Maruyama, K. (1992) Characterization and localization of alpha-connectin
(titin 1): an elastic protein isolated from rabbit skeletal muscle. J. Muscle
Res. Cell Motil. 13, 39-47.
Kimura, S., Kawamura, Y., Watanabe, A., Kubokawa, K. and Maruyama K.
(1997) A connectin-like protein in amphioxus striated muscle. Zool. Sci. 14,
54.
Kubokawa, K., Azuma, N. and Tomiyama, M. (1998) A new population of the
amphioxus (Branchiostoma belcheri) in the Enshu-Nada sea in Japan. Zool.
Sci. 15, 799-803.
Labeit, S. and Kolmerer, B. (1995) The complete primary structure of human
nebulin and its correlation to muscle structure. J. Mol. Biol. 248, 308-315.
Laemmli, U. K. (1970) Cleavage of structural proteins during the assembly of
the head of bacteriophage T4. Nature 227, 680-685.
Locker, H. R. and Wild, D. J. C. (1986) A comparative study of high molecular
weight proteins in various types of muscle across the animal kingdom. J.
Biochem. 99, 1473-1484.
Lukoyanova, N., VanLoock, M. S., Orlova, A., Galkin, V. E., Wang, K. and
Egelman E. H. (2002) Each actin subunit has three nebulin binding sites:
implications for steric blocking. Curr. Biol. 12, 383-388.
McElhinny, A. S., Kolmerer, B., Fowler, V. M., Labeit, S and Gregorio, C. C.
(2001) The N-terminal end of nebulin interacts with tropomodulin at the

20
pointed ends of the thin filaments. J. Biol. Chem. 276, 583-592.
McElhinny, A. S., Kazmierski, S. T., Labeit, S. and Gregorio, C. C. (2003)
Nebulin: the nebulous, multifunctional giant of striated muscle. Trends
Cardiovasc. Med. 13, 195-201.
McLachlan, A. D. and Karn, J. (1982) Periodic charge distributions in the
myosin rod amino acid sequence match cross-bridge spacing in muscle.
Nature 299,226-231.
Millevoi, S., Trombitas, K., Kolmerer, B., Kostin, S., Schaper, J., Pelin, K.,
Granzier, H. and Labeit, S. (1998) Characterization of nebulette and nebulin
and emerging concepts of their roles for vertebrate Z-discs. J. Mol. Biol. 28,
111-123.
Mineta, K., Nakazawa, M., Cebria, F., Ikeo, K., Agata, K. and Gojobori, T.
(2003) Origin and evolutionary process of the CNS elucidated by comparative
genomics analysis of planarian ESTs. Proc. Natl. Acad. Sic. USA 13,
7666-7671.
Moncman, C. L. and Wang, K. (1995) Nebulette: a 107 kD nebulin-like
protein in cardiac muscle. Cell Motil. Cytoskeleton 32, 205-225.
Spudich, J. A. and Watt, S. (1971) The regulation of rabbit skeletal muscle
contraction. I. Biochemical studies of the interaction of the
tropomyosin-troponin complex with actin and the proteolytic fragments of
myosin. J. Biol. Chem. 246, 4866-4871.
Suzuki, T., Yajima, H., Maruyama, K. and Kimura, S. (2000) A 7.5-kb 3'
terminal cDNA sequence of chicken skeletal muscle nebulin reveals its actin
binding regions. Zool. Sci. 17, 1095-1099.

21
Towbin, H., Staehelin, T. and Gordon, J. (1979) Electrophoretic transfer of
proteins from polyacrylamide gels to nitrocellulose sheets: procedure and
some applications. Proc. Natl. Acad. Sci. USA 76, 4350-4354.
Tskhovrebova, L. and Trinick, J. (2003) Titin: properties and family
relationship. Nat. Rev. Mol. Cell Biol. 4, 679-689.
Wang, K. (1982) Purification of titin and nebulin. Methods Enzymol. 85,
264-274.
Wang, K. and Wright, J. (1988) Architecture of the sarcomere matrix of
skeletal muscle: immunoelectron microscopic evidence that suggests a set of
parallel inextensible nebulin filaments anchored at the Z line. J. Cell Biol.
107, 2199-2212.
Witt, C. C., Burkart, C., Labeit, D., McNabb, M., Wu, Y., Granzier, H. and
Labeit, S. (2006) Nebulin regulates thin filament length, contractility, and
Z-disk structure in vivo. EMBO J. 25, 3843-3855, 2006.
22
A
B
C Fig 1. Domain structures and primary structure of the nebulin C-terminal region in amphioxus striated muscle cDNA. Domain structures (A) and amino acid sequence (B) of amphioxus nebulin, and consensus sequences of amphioxus and human nebulin (C). Fine bar, size of each clone. Bold bar, Pc AmpN antigen.
Neb1-12 Neb13 SH3
Unique region SH3 domain
Antigen
Clone:53616024
907polyApolyApolyA
Nebulin repeat
Neb1-12 Neb13 SH3
Unique region SH3 domain
Antigen
Clone:53616024
907polyApolyApolyA
Nebulin repeat
K----ERAELEKHK-YTQLPDT
Neb1 PDISFAKEVHDITSETKYK----EEYNKERGK-YSMVAQT
Neb2 PLYETAQKANKLTSETLYK----KAYKALQGKGY-ALIDT
Neb3 PEFERCKQVNKLASRTKYH----EDYEKSKIKGYTPVVDT
Neb4 PENIRLRQQSKIQSQNEYQ----KDYKALSGK-FHYTADN
Neb5 PEHNRIRSVSKIQSQIHYG----PSRLEQHGQKYHAAIDD
Neb6 PELKRLQEVGKFQSQVLYGEITDKDDASKGRGKYTQIQQD
Neb7 PEMKRLKEANAYASEAQVA----VRRFDEK-NRYTTVLDD
Neb8 PELNRMKNVGKFQSELQYK----DKWTRKTGARYTNVLDD
Neb9 PEMNRIKGVTKMLSEVQYR----DAEL-KTG-RLTAVTDD
Neb10 LDTKRVKRNTKIQSENEYR-------KGRKINTYNAVLDT
Neb11 PDMLRIRNATKISSEVKYH----EDFEKQKGKAYHTVIDD
Neb12 PELTRVKKNTVIQSNLAYQGVKEKLQTMESRRQMI
Unique PGEPSMPPPAQQLGTQMEARMAADASYAQFKAKGKGKV
Neb13 GGRYMPIRLGDPGSIFDYEPYQGKEEAGRAIYVTTATWK
Unique PAKPIRTIEDFVVEEESAEEALEARDKITEMDMEYERRRGEGLSYEYDPQKYSHHYWS
KTTSELGGVVTTSGMKYRKIEDFLNLNADLIYSTENESTRESGSETSGMWDYDPDKYW
RARGYRISAEQRYQQVRLKPGKIEDYDPMTHPDAQPKFEAPAAVSEPEPVIQSGNVMR
SH3 AVFDYAAAEDDEISFMDGDVIVNCVKIDDGWMTGTVQRTGQSGMLPANYVEPMS
Amphioxus PE--R-K-V-KIQSEXXYX-----------GK-YT-V-DT/D
** * * * * ** * *
Human PEM-RVK-NQENISSVLYKEN---------GK--TP-P-T

23
Fig 2. Comparison of the C-terminal region between amphioxus and human nebulins. The domain structure and isoelectric point (pI) are presented according to Labeit and Kolmerer (1995), and the I-Z border is presented according to Millevoi et al. (1998). Amphioxus striated muscle nebulin, Amphioxus; Human skeletal muscle nebulin, Human.
N2-12N1N13
M169-172
SH3
SH3
pI=10.0pI=5.6
pI=9.9pI=5.6
26% 11% 65%
Z-line
Amphioxus
HumanM173-185
Unique region SH3 domainNebulin repeat
N2-12N1N13
M169-172
SH3
SH3
pI=10.0pI=5.6
pI=9.9pI=5.6
26% 11% 65%
Z-line
Amphioxus
HumanM173-185
Unique region SH3 domainNebulin repeat

24
Fig 3. SDS-PAGE and immunoblot detection of amphioxus striated muscle. Antigen for amphioxus nebulin (A). Molecular size marker, lane M; Pc AmpN antigen (bold bar of Fig. 1A), lane 1. Immunoblot detection of amphioxus striated muscle using Pc AmpN (B, C). Electrophoresis was performed on 2–6% (B) or 2–15% (C) polyacrylamide gradient gels. Molecular size marker, lane M; amido black stains, lane 1; immunoblot detection by Pc AmpN, lane 2. Connectin, C; Nebulin, N; Myosin heavy chain, MHC.
2209766
45
24
18
(kDa) M 1A B
N
MHC
C
21C
220
9766
45
2418
(kDa) M 1 2
N220
9766
45
24
18
(kDa) M 1A
2209766
45
24
18
2209766
45
24
18
(kDa) M 1A B
N
MHC
C
21B
N
MHC
C
21
N
MHC
C
21C
220
9766
45
2418
(kDa) M 1 2
N
C
220
9766
45
2418
(kDa) M 1 2
N

25
Fig. 4. Immunofluorescence microscopic observations of amphioxus striated muscle sarcomeres using Pc AmpN (B) and Mc a-actinin (C). Phase-contrast image (A) and merged image of Pc AmpN and Mc a-actinin (D). Arrow head, Z line. Bar, 10 mm.
A
C
D
B
A
C
D
B

26
A B Fig. 5. Binding of amphioxus nebulin and F-actin. (A) Binding of amphioxus nebulin and F-actin by far-western blotting. 1, Molecular weight marker; 2, Amido black of (AN3-9 + GST); 3, + F-actin, treated with actin antibody; 4, – F-actin, treated with actin antibody (negative control of 3). (B) Binding of amphioxus nebulin and F-actin by co-sedimentation. Actin only, Actin; amphioxus nebulin repeats 3–9 dissolved in GST, AN3–9; amphioxus nebulin repeats 3–9 + actin, AN3–9 + Actin; amphioxus nebulin repeats 3–9 + BSA, AN3–9 + BSA. Before centrifugation, W; supernatant after centrifugation, S; pellet after centrifugation, P. Electrophoresis was performed on a 12.5% polyacrylamide gel.
W S P W S P W S P W S PActin AN3-9 AN3-9+ActinAN3-9+BSA
BSAActin
AN3-9
220
9766
45
24
(kDa)
GST3-9
1 2 3M

27
A B
C D
1 2 3 4 5 6220
97
66
45
24
(kDa)
a-Actinin
GST-AN3-9
GST
1 2 3 4 5 6220
97
66
45
24
(kDa)
a-Actinin
GST-AN3-9
GST
18
9766
45
24
(kDa)
His1-21 2 1 2 1 2 1 2
His3-9
His10-U
HisU-SH3
M
220
97
66
45
24
(kDa)1 2 3 4
CH SPEC EFEFSPECSPECSPECCHa-act
343-897343-804426-804343-729
Amphioxus Nebulin binding-++-
343-897 343-804 426-804343-729
CH SPEC EFEFSPECSPECSPECCHa-act
343-897343-804426-804343-729
Amphioxus Nebulin binding-++-
343-897 343-804 426-804343-729

28
Fig. 6. Binding of amphioxus nebulin and a-actinin. (A) Binding of amphioxus nebulin C-terminal region and a-actinin by far-western blotting. 1, treated with polyhistidine antibody; 2, treated with a-actinin antibody. U, unique region; SH3, SH3 domain. (B) Binding of amphioxus nebulin and a-actinin by the yeast two-hybrid system. a-act, Domain structure of a-actinin; CH, calponin homology domain; SPEC, spectrin repeat; EF, EF hand. +, b-Gal (b-Galactosidase) positive; –, b-Gal negative. (C) Binding of amphioxus nebulin and a-actinin by GST pull-down assay. 1, Molecular size marker; 2, a-actinin used for the pull-down assay; lane3-6, after the pull-down assay.; 3, GST fusion protein for amphioxus nebulin repeats 3–9–a-actinin; 4, GST only–a-actinin; 5, GST fusion protein for amphioxus nebulin repeats 3–9+a-actinin; 6, GST only+a-actinin; GST–AN3–9, GST fusion protein for amphioxus nebulin repeats 3–9. Electrophoresis was performed on a 10% polyacrylamide gel. (D) Binding of amphioxus nebulin repeats 3−9 and BSA as a control for binding of nebulin repeats 3−9 and α-actinin, by a GST pull-down assay. 1, Molecular size marker; 2, BSA used for pull-down assay; lane3 and 4, after pull-down assay; 3, GST fusion protein for amphioxus nebulin repeats 3−9 + BSA; 4, GST fusion protein for amphioxus nebulin repeats 3−9 − BSA; GST−AN3−9, the GST fusion protein for amphioxus nebulin repeats 3−9. Electrophoresis was performed on a 10% polyacrylamide gel.
29
A B Fig. 7. Binding of amphioxus nebulin and connectin. (A) Binding of amphioxus nebulin and connectin by far-western blotting. 1, Molecular weight marker; 2, Amido black of (U–SH3 + GST); 3, + connectin, treated with connectin antibody; 4, – connectin, treated with connectin antibody (negative control of 3). U, unique region; SH3, SH3 domain. (B) Binding of amphioxus nebulin and connectin by GST pull-down assay. 1, Molecular size marker; 2, rabbit skeletal muscle; 3, connectin used for the pull-down assay; lane4-7, after the pull-down assay; 4, GST–U–SH3 – connectin; 5, GST only–connectin; 6, GST–U–SH3+connectin; 7, GST only+connectin; GST-U–SH3, GST fusion protein for amphioxus nebulin U–SH3 region. Electrophoresis was performed on a 2–15% polyacrylamide gel.
1 2 3 4 5 6 7
220
9766
45
24
(kDa)
18
Connectin
GST-U-SH3GST
1 2 3 4 5 6 7
220
9766
45
24
(kDa)
18
Connectin
GST-U-SH3GST
18
9766
45
24
(kDa)2 3
GSTU-SH3
M 1

30
Chapter 2
Structure of the amphioxus nebulin gene and evolution of
the nebulin family genes

31
1. Abstruct
Nebulin family genes are believed to have diverged from a single gene
during the evolution of vertebrates. I determined the structure of the
amphioxus nebulin gene and showed that in addition to the features of the
human nebulin gene, this gene had a LIM domain, secondary super repeats
and a giant exon with 98 nebulin repeats containing unique sequences. A
transcript of this gene amplified by reverse transcriptase-polymerase chain
reaction had an LIM domain, three nebulin repeats and an SH3 domain.
This transcript was similar to an isoform of human nebulette (Lasp-2).
Phylogenetic analysis using the LIM and SH3 domains of the nebulin family
proteins showed that amphioxus nebulin is located outside the vertebrate
nebulin family group in the phylogenetic tree. These results indicated that
the amphioxus nebulin gene had a unified structure among nebulin,
nebulette, lasp-1 and N-RAP of vertebrates, and that these nebulin family
genes diverged from the amphioxus nebulin gene during the course of
vertebrate evolution.

32
2. Introduction
The nebulin family of proteins includes Nebulin, Nebulette, N-RAP and
Lasp-1 actin binding proteins, which are characterized by nebulin repeats.
Nebulin is a large protein (molecular mass: 773 kDa) found on the thin
filaments of vertebrate skeletal muscles (Wang, 1982; McElhinny et al.,
2003). The primary structure of human nebulin is an N-terminal glutamic
acid-rich domain, followed by 185 (M1–185) contiguous nebulin repeats of
approximately 35 amino acids with a central SDXXYK consensus sequence, a
C-terminal serine-rich region and an Src homology 3 (SH3) domain. Nebulin
repeats (M9–162) in the I-band region of the sarcomere comprise 22 sets of
seven-repeats, called super repeats (SR1-22) (Labeit and Kolmerer, 1995).
The gene for the production of human nebulin is located in chromosome
2q21-22 and is composed of 183 exons spanning 249 kb of the genome
sequence, an area that contains 29 equivalent super repeats (Donner et al.
2004).
Nebulette is a 107 kDa protein located in the Z-line of cardiac muscle
(Moncman and Wang. 1995). The primary structure of human nebulette is a
N-terminus glutamate acid-rich domain, followed by 23 nebulin repeats, a
C-terminal ser-rich region and an SH3 domain (Millevoi et al. 1998). A
34-kDa splicing isoform of nebulette is Lasp-2 (LIM-nebulette), observed in
various tissues and has an N-terminus LIM domain, followed by 3 nebulin
repeats, a C-terminus ser-rich region and an SH3 domain (Katoh and Katoh.
2003; Terasaki et al. 2004; Li et al. 2004). The gene producing human
nebulette is located in chromosome 10p12 and spans 393 kb—an area

33
containing 28 exons (exon 1–28) for nebulette protein (Arimura et al. 2000)
and four additional 5’ exons (1a-4a) only expressed in Lasp-2 protein
(Katoh and Katoh. 2003). Lasp-2 is coded by exons 1a–4a, 24, 27 and 28.
N-RAP is a 196 kDa protein that has been reported in striated muscles
(Luo et al. 1997). The N-terminus of human N-RAP is a LIM domain, and is
followed by 11 nebulin repeats and 5 super repeats (35 nebulin repeats). The
gene responsible for the production of human N-RAP is located in
chromosome 10q25.3 and spans 75 kb, an area that contains 42 exons
(Mohiddin et al. 2003).
Lasp-1 is a 38 kDa protein (Tomasetto et al. 1995) that is highly expressed
within epithelial and brain tissues. It consists of an N-terminus LIM domain,
followed by two nebulin repeats, a C-terminal unique region and an SH3
domain. The gene encoding human Lasp-1 production is located in
chromosome 17q12-21 and spans 52 kb.
The genes of the above four proteins are believed to have diverged from a
single gene by gene duplication during vertebrate evolution. In order to
explore this hypothesis thoroughly, it was deemed necessary to find a gene
that was common to the nebulin family in amphioxus, a cephalochordate (i.e.,
an invertebrate chordate), and the nearest ancestor to the vertebrates. In
chapter 1, I demonstrated the existence of amphioxus nebulin by revealing
its C-terminal primary structure, localization and binding protein. In
chapter 2, I reveal the structure of the amphioxus nebulin gene and analyze
the hypothesis that the nebulin family arose from a single gene.

34
3. Materials and Methods
3.1. Genome analysis
The scaffold coding amphioxus nebulin gene was obtained from the
Branchiostoma floridae genome assembly release v2.0 (Putnam et al. 2008)
on the JGI portal site (http://genome.jgi-psf.org/Brafl1/Brafl1.home.html) by
BLAST search using the sequence of the SH3 domain of the amphioxus
(Branchiostoma belcheri) nebulin (Chapter 1). The complementary sequence
of No. 205 in 398 connected sets of allelic scaffolds was identified (amino acid
homology 98%), and translated to 3 frame amino acids. The exons were
visually determined referred to the C-terminus sequence of amphioxus
nebulin (Chapter 1), an EST sequence of B. floridae (BW805004) and the
sequence of human nebulin (Labeit and Kolmerer. 1995). The connecting
error of the No. 205 set was proofed by comparison with scaffold 169 of the B.
floridae v1.0 assembly.
3.2. Reverse transcriptase-polymerase chain reaction (RT-PCR) and
sequence
Collection of amphioxus, isolation of RNA and construction of the first
strand of cDNA by reverse transcription (RT) were performed by Mizuta and
Kubokawa (2007). PCR reactions were performed using a forward primer
(cgcggatccATGAATCCGAAGTGTGCACG; BamHI site plus N-terminus of
LIM domain) and a reverse primer
(cgcgaattcCGACATGGGCTCCACGTAGT; EcoRI site plus C-terminus of SH3
domain) with a condition schedule of 1 min at 95°C, followed by 30 s at 95°C,

35
30 s at 50°C and 1 min at 72°C for 25 cycles, followed by 3 min at 72°C; the
product obtained was stored at 4°C. Sequences were analyzed using an ABI
PRISM 310 sequencer (Applied Biosystems, Foster City, CA, USA).
3.3. Phylogenetic analysis
The amino acid sequences of the LIM and SH3 domains were aligned in
various species and proteins and 1000 bootstrap values were calculated
using the CLUSTALW algorithm (Thompson et al. 1994). Phylogenetic trees
were visualized using TreeView (Page, 1996).
4. Results
4.1. The structure of the amphioxus nebulin gene
Using the amino acid sequence of the SH3 domain of amphioxus nebulin
(Chapter 1), I searched for an amphioxus nebulin gene in the B. floridae v2.0
genome assembly (Putnam et al. 2008) and identified a complementary
sequence of No. 205 in 398 connected sets of allelic scaffolds encoding
nebulin genes. The exons encoding nebulin repeats were visually determined
with reference to human nebulin sequences (Labeit and Kolmerer, 1995),
whereas the exons encoding the unique sequence of the amphioxus nebulin
C-terminus and the SH3 domain were searched using homology with the
C-terminal sequence of amphioxus nebulin (Chapter 1). Furthermore, exons
encoding the N-terminal LIM domain and unique sequences were searched
based on homology with the EST sequence(BW805004).
The predicted amphioxus nebulin gene comprised 208 kb with 283 exons

36
(Fig. 1), and was smaller and had more exons than the human nebulin gene
(249 kb with 183 exons). The size range of the exons were very various from
18 b (exon 277) to 15 kb (exon 260).
All exons were connected and aligned (Fig.2), and exons 1 and 2, encoding
the LIM domain, were not observed in the human nebulin gene. These exons
were separated by 24.7 kb from the other exons.
Exon 3–269 encoded nebulin repeats, while in exon 9–245, nebulin super
repeats were observed, including those super repeats observed in human
nebulin. However, the amphioxus nebulin gene has 2 sets of similar super
repeats, one containing 21 super repeats (SR01–21) and the other containing
20 super repeats (SR22–41). These sets were different from those in the
human nebulin gene, which has one set of 22 super repeats. Each set of super
repeats had amino acids that were highly conserved. In particular, repeat 2
among the super repeats was more similar than other repeats within the
super repeats. Furthermore, the WLKGIGW motif observed in the super
repeat sequences of human nebulin was not observed in that of amphioxus
nebulin.
Exon 260 is a large exon with 98 nebulin repeats containing unique
sequences at the N- and C-terminus, which show no similarities to other
proteins in the database (Fig. 3). This exon is located between the exons that
encode Neb04/05 and Neb05/06, and is not observed at the C-terminus
sequence of either amphioxus (Chapter 1) or human nebulin. Expression of
the C-terminus of this exon was observed in the neurula stage EST clones of
B. floridae (DDBJ accession BW855343; underlined in Fig. 3).

37
Exons 266–267 were two additional nebulin repeats that could not be
observed in the sequence of the amphioxus nebulin C-terminus
(Neb00/01–Neb11/12) (Chapter 1). The C-terminus of human nebulin has a
splicing isoform region called a ‘hot spot’, but the C-terminus of amphioxus
nebulin has few alternatively spliced exons compared to that of human
nebulin. Exon 270–282 encoded unique sequences and exon 283 encoded the
SH3 domain. While exons 271–283 are expressed in amphioxus nebulin
(Chapter 1), exon 270 is not.
These results indicate that the amphioxus nebulin gene has exons
encoding a LIM domain, a set of secondary super repeats and 98 nebulin
repeats containing unique sequences, in addition to having exons that are
present in the human nebulin gene.
4.2. A small transcript from the amphioxus nebulin gene
In order to search for a transcript that had an LIM domain in the
amphioxus nebulin gene that was not observed in human nebulin, I
performed RT-PCR using amphioxus (B. belcheri) total RNA and primers
that encoded the N-terminus of the LIM domain as well as the C-terminus of
the SH3 domain. A transcript was identified (Fig. 4A) that had an LIM
domain and a unique sequence at the N-terminus, three nebulin repeats and
a unique sequence and an SH3 domain at the C-terminus (Fig. 4B). This
sequence was encoded by exons 1, 2, 3, 258, 269, 270, 282 and 283 (Fig. 1). In
particular, exons 258, 269, 282 and 283 were visible on the C-terminus of
amphioxus nebulin (Chapter 1), but exon 270 was not. Domain structures

38
and splicing pathways observed in this transcript were not observed in
human nebulin. Instead, they were similar to human Lasp-2, which is a
splicing isoform of nebulette (Li et al. 2004; Terasaki et al. 2004) (Fig. 4C).
These results indicate that the nebulette gene is merged with the nebulin
gene in amphioxus.
4.3. The evolution of vertebrate nebulin family genes from the amphioxus
nebulin gene
The results show that the amphioxus nebulin gene has many sequences in
addition to those observed in human nebulin, and that a small transcript
from the amphioxus nebulin gene is similar to the Lasp-2 isoform of human
nebulette. Therefore, I investigated the possibility that vertebrate nebulin
family genes diverged from an amphioxus nebulin gene.
Molecular phylogenetic analysis using amino acid sequences of the SH3
domains in nebulin, nebulette and Lasp-1 showed that each vertebrate
protein, from fish to human, formed its own specific group (i.e., a vertebrate
nebulin group, a vertebrate nebulette group and a vertebrate Lasp-1 group),
and that each group formed a vertebrate super-group (Fig. 5A). Furthermore,
the analysis showed that Ciona nebulin (urochordate) and amphioxus
nebulin (cephalochordate) were located outside of the vertebrate
super-group.
Similar analysis using amino acid sequences of LIM domains in nebulette,
Lasp-1 and N-RAP showed that each protein of vertebrate formed one group,
and that amphioxus nebulin was located outside the vertebrate group (Fig.

39
5B).
These results indicate that the genes of vertebrate nebulin, nebulette,
Lasp-1 and N-RAP diverged from an amphioxus nebulin gene.
5. Discussion
In this study, I have determined the structure of the amphioxus nebulin
gene and have shown that this gene has exons encoding the LIM domain,
secondary super repeats and 98 nebulin repeats containing unique
sequences, in addition to having exons similar to those of the human nebulin
gene. I have also identified a small isoform from this amphioxus nebulin
gene and have shown that the isoform is similar to the Lasp-2 isoform of
human nebulette. Furthermore, I have revealed the evolution of nebulin
family proteins from the amphioxus nebulin gene.
5.1. LIM domain
The presence of the exons encoding an LIM domain in the amphioxus
nebulin gene has been demonstrated and I have also demonstrated the
expression of these exons in a small isoform. These results indicate that in
amphioxus, the nebulette gene is the same as the nebulin gene. However,
this insight did not lead to the conclusion that the N-terminus of amphioxus
nebulin is an LIM domain. In fact, the exon encoding the LIM domain is 24.7
kb away from other exons. Among exons, this is a relatively long distance
and a similar great amount of separation is only observed in a small isoform
of human nebulette, (Lasp-2; 152.2 kb away from other exons). Instead of an

40
LIM domain, nebulette has a unique sequence at the N-terminus. This
indicates that nebulette does not require an LIM domain, even if the exons at
the N-terminus of the gene encode an LIM domain. I performed an analysis
of the N-terminal amino acids within the amphioxus nebulin protein, using a
750-kDa band transferred onto a polyvinylidene fluoride membrane; however,
I could not determine the sequence of amphioxus nebulin due to the high
molecular weight of nebulin (data not shown). Thus, the determination of the
N-terminal sequence of amphioxus nebulin is a future task.
5.2. Super repeats
My data show that the amphioxus nebulin gene has two super repeat
groups: one contains 21 super repeats and the other contains 20 super
repeats. In human nebulin, there are 29 super repeats (Donner et al. 2004).
Twenty-two of those are expressed in skeletal muscles (Labeit and Kolmerer.
1995) and the other seven are expressed only in foetal muscles (Donner et al.
2004). Since the molecular weight of nebulin in amphioxus striated muscles
(750 kDa, Chapter 1) is slightly lower than that in human adult skeletal
muscles (773 kDa, Labeit and Kolmerer 1995), the expression of one group of
21 or 20 super repeats is possible in amphioxus nebulin. On the other hand,
the expression of both groups of 21 and 20 super repeats is unlikely in
amphioxus nebulin, because the length of thin filament (about 1 mm) in
amphioxus striated muscle has similar length to that in human skeletal
muscle (Hagiwara et al. 1971). My phylogenetic analysis showed that the
N-RAP gene diverged from the amphioxus nebulin gene. This may mean that

41
one group of the super repeats expresses nebulin, whereas the other
expresses N-RAP. Two unresolved issues for future studies are: the reasons
for the presence of two groups of super repeats in the amphioxus nebulin
gene, and identification of the group from these two groups that is expressed
in amphioxus nebulin.
All amino acid sequences in the super repeats from the amphioxus nebulin
gene lack a WLKGIGW-motif, which is assumed to be a
tropomyosin/troponin binding motif. However, because the second repeat in
every repeat is more conservative than those between the super repeats of
each group, this second repeat may be important for binding to the
tropomyosin/troponin complex.
5.3. Giant exon
Here, I report the detection of a giant exon in amphioxus that encodes 98
nebulin repeats and has unique sequences at the N- and C- terminals. The
expression of this exon was observed in the EST sequence (BW855343). This
giant exon does not exist in nebulin, nebulette or N-RAP genes of vertebrates.
There are three possible reasons for the expression of this giant exon. The
first possibility is that partial exons within the giant exon are expressed and
play a role similar to that of the hot spot of alternatively spliced exons in the
human nebulin C-terminus. The reason for considering this possibility is
that this giant exon is inserted between the exons encoding Neb3/4 and
Neb4/5 of the C-terminus of nebulin amphioxus (Chapter 1) and human
nebulin, and that this region is a hot spot of alternatively spliced exons. The

42
second possibility is that this giant exon is expressed with other exons,
especially with the SH3 domain, and it performs the functions of amphioxus
nebulette, because like human nebulette, this giant exon has no super repeat.
The third possibility is that only this giant exon is expressed, it performs
functions specific to the striated muscles of amphioxus and these functions
differ from those of nebulin, nebulette, N-RAP and Lasp-1. Future
investigations of the expression and functional analysis of this exon should
lead to new insights in amphioxus striated muscle.
5.4. Gene separation
It is believed that vertebrates have more genes than their closest
invertebrate relatives (amphioxus) because of gene duplication (Holland et al.
1994, 2003). This belief leads to the hypothesis that nebulette, N-RAP and
lasp-1 genes emerged through nebulin gene duplication at an early stage of
vertebrate evolution. This hypothesis is partially supported by the
observation that the cardiac muscle of primitive vertebrates, such as jawless
lamprey and hagfish, have nebulin, but that this nebulin has changed to
nebulette in further evolved vertebrates, such as sharks (Fock and Hinssen
2002). However, there is no direct evidence for this hypothesis at the gene
level, and my results show that there are nebulette exons and possibly
N-RAP exons in the amphioxus nebulin gene. Furthermore, other than the
nebulin gene, I could not find genes corresponding to the production of
nebulette, N-RAP or lasp-1 in the amphioxus genome. My results are the
first direct data to support the fact that nebulin, nebulette, N-RAP and

43
lasp-1 are present in a single gene, and that these results will assist in the
analysis of nebulin family evolution in vertebrates.
6. Conclusion
The amphioxus nebulin gene has many exons that are observed not only in
vertebrate nebulin, but also in other nebulin family genes in vertebrates.
Elucidation of this gene and its transcripts will lead to a better
understanding of the evolution of nebulin family proteins and their
functions.

44
7. References
Arimura, T., Nakamura, T., Hiroi, S., Satoh, M., Takahashi, M., Ohbuchi, N.,
Ueda, K., Nouchi, T., Yamaguchi, N., Akai, J., Matsumori, A., Sasayama, S.,
Kimura, A. (2000) Characterization of the human nebulette gene: a
polymorphism in an actin-binding motif is associated with nonfamilial
idiopathic dilated cardiomyopathy. Hum. Genet. 107, 440-451.
Donner, K., Sandbacka, M., Lehtokari, V.L., Wallgren-Pettersson, C., Pelin,
K. (2004) Complete genomic structure of the human nebulin gene and
identification of alternatively spliced transcripts. Eur. J. Hum. Genet. 12,
744-751.
Fock, U., Hinssen, H. (2002) Nebulin is a thin filament protein of the cardiac
muscle of the agnathans. J. Muscle Res. Cell Motil. 23, 205-213.
Hagiwara, S., Henkart, M.P., Kidokoro, Y. (1971) Excitation-contraction
coupling in amphioxus muscle cells. J. Physiol. 219, 233-251.
Holland, P.W., Garcia-Fernàndez, J., Williams, N.A., Sidow, A. (1994) Gene
duplications and the origins of vertebrate development. Dev. Suppl. :125-133
Holland, P.W. (2003) More genes in vertebrates? J. Struct. Funct. Genomics 3,
75-84.
Katoh, M., Katoh, M. (2003) Identification and characterization of LASP2
gene in silico. Int. J. Mol. Med. 12, 405-410.
Labeit, S., Kolmerer, B. (1995) The complete primary structure of human
nebulin and its correlation to muscle structure. J. Mol. Biol. 248, 308-315.
Li, B., Zhuang, L., Trueb, B. (2004) Zyxin interacts with the SH3 domains of
the cytoskeletal proteins LIM-nebulette and Lasp-1. J. Biol. Chem. 279,

45
20401-20410.
Luo, G., Zhang, J.Q., Nguyen, T.P., Herrera, A.H., Paterson, B., Horowits, R.
(1997) Complete cDNA sequence and tissue localization of N-RAP, a novel
nebulin-related protein of striated muscle. Cell Motil. Cytoskeleton 38,
75-90.
McElhinny, A.S. Kazmierski, S.T., Labeit, S., Gregorio, C.C. (2003) Nebulin:
the nebulous, multifunctional giant of striated muscle. Trends Cardiovasc.
Med. 13, 195-201.
Millevoi, S., Trombitas, K., Kolmerer, B., Kostin, S., Schaper, J., Pelin, K.,
Granzier, H., Labeit, S. (1998) Characterization of nebulette and nebulin and
emerging concepts of their roles for vertebrate Z-discs. J. Mol. Biol. 28,
111-123.
Mizuta, T., Kubokawa, K. (2007) Presence of sex steroids and cytochrome
P450 genes in amphioxus. Endocrinology 148, 3554-3565.
Mohiddin, S. A., Lu, S., Cardoso, J.P., Carroll, S., Jha, S., Horowits, R.,
Fananapazir, L. (2003) Genomic organization, alternative splicing, and
expression of human and mouse N-RAP, a nebulin-related LIM protein of
striated muscle. Cell Motil. Cytoskeleton 55, 200-212.
Moncman, C.L., Wang, K. (1995) Nebulette: a 107 kD nebulin-like protein in
cardiac muscle. Cell Motil. Cytoskeleton 32, 205-225.
Page, R.D. (1996) TreeView: an application to display phylogenetic trees on
personal computers. Comput. Appl. Biosci. 12, 357-358.
Putnam, N.H., Butts, T., Ferrier, D.E., Furlong RF, Hellsten U, Kawashima
T, Robinson-Rechavi M, Shoguchi E, Terry A, Yu JK, Benito-Gutiérrez EL,

46
Dubchak I, Garcia-Fernàndez J, Gibson-Brown JJ, Grigoriev IV, Horton AC,
de Jong PJ, Jurka J, Kapitonov VV, Kohara Y, Kuroki Y, Lindquist E, Lucas
S, Osoegawa K, Pennacchio LA, Salamov AA, Satou Y, Sauka-Spengler T,
Schmutz J, Shin-I T, Toyoda A, Bronner-Fraser M, Fujiyama A, Holland LZ,
Holland PW, Satoh N, Rokhsar DS (2008) The amphioxus genome and the
evolution of the chordate karyotype. Nature 453, 1064-1071.
Terasaki, A.G., Suzuki, H., Nishioka, T., Matsuzawa, E., Katsuki, M.,
Nakagawa, H., Miyamoto, S., Ohashi, K. (2004) A novel LIM and SH3
protein (Lasp-2) highly expressing in chicken brain. Biochem. Biophys. Res.
Commun. 313, 48-54.
Tomasetto, C., Moog-Lutz, C., Régnier, C.H., Schreiber, V., Basset, P., Rio,
M.C. (1995) Lasp-1 (MLN 50) defines a new LIM protein subfamily
characterized by the association of LIM and SH3 domains. FEBS Lett. 373,
245-249.
Thompson, J.D., Higgins, D.G., Gibson, T.J. (1994) CLUSTAL W: improving
the sensitivity of progressive multiple sequence alignment through sequence
weighting, position-specific gap penalties and weight matrix choice. Nucleic
Acids Res. 22, 4673-4680.
Wang, K. (1982) Purification of titin and nebulin. Methods Enzymol. 85,
264-274.

47
Fig. 1 The structure of the amphioxus nebulin gene.The exons are shown as boxes and the introns as lines. Yellow boxes, untranslated regions; red, nebulin repeats; purple, LIM domains; dark orange, nebulin repeats (i.e. super repeat set 1); light orange, nebulin repeats (i.e. super repeat set 2); pink, a large exon with 98 nebulin repeats containing unique sequences at the N- and C-terminus; grey, unique regions; green, SH3 domain. Double solidus marks, connecting error sites of allelic scaffolds. Arrowheads, exons coding an isoform of nebulin (shown in Fig. 4).
5‘UTR
10 20 30
40 50 60
70 80 90
100 110 120
130 140 150
160 170 180
190 200210
SR1 SR2 SR3 SR4 SR5 SR6 SR7 SR8 SR9 SR10
SR11 SR12 SR13 SR14 SR15 SR16 SR17 SR18 SR19
SR20 SR21 SR22 SR23 SR24 SR25 SR26 SR27 SR28 SR29 SR30
SR31 SR32 SR33 SR34 SR35 SR36 SR37 SR38 SR39
kb
SR40 SR41
LIM
SH33‘UTR
5‘UTR
10 20 30
40 50 60
70 80 90
100 110 120
130 140 150
160 170 180
190 200210
SR1 SR2 SR3 SR4 SR5 SR6 SR7 SR8 SR9 SR10
SR11 SR12 SR13 SR14 SR15 SR16 SR17 SR18 SR19
SR20 SR21 SR22 SR23 SR24 SR25 SR26 SR27 SR28 SR29 SR30
SR31 SR32 SR33 SR34 SR35 SR36 SR37 SR38 SR39
kb
SR40 SR41
LIM
SH33‘UTR

48
Fig. 2 The connected sequence of amphioxus nebulin exons. The tyrosines conserved among the nebulin repeats are marked with black. The common amino acids conserved among super repeats of each group are marked with grey. LIM, LIM domain; Neb, nebulin repeat; SR01–42, super repeat 01–42; Uni, unique region; SH3, SH3 domain. The exons coded by Chapter 1 are shown as <Neb00/01> – <SH3>. Exon 270, which codes a unique sequence, but was not expressed in Chapter 1 was not added.
(LIM) MNPKCARCEKTVYPMEKLNCLDKYWHKGCFNCEECRMTLNMKNYKGFNKKPYCNA
(Neb) HYPQSKPTVVSETPEIARVMQQSQQQSL
(Neb) *TNKQTNMLTVVLMFQADQPWDTPETKRVREMGTVQSE
(Neb) *AEMMAEMTAGSGKRG DQSWETPEMVRCREMKDLQSE
(Neb) RVYKEKAKERMKAYTHHPDAPEMARLKEIGKVCSE
(Neb) VKYHEAYEKSKAKGYTSVVDTPENIRVKESGKIASQ
(Neb) *GTRQAKCIEDFEKLPKPGMLPADTSEIVRMKKVQQIQSE
(SR01) IPYKKEVKESTGKFFDVQDTPGYQHNLQLTKYQSD-----NVYMDEAKYIMEKGHSQYPFTPEMTRVQGAEKVKNE--RLYKESYEKYKSTFTLPPDTPGIVHAKNMGALHSEMKYHELY-QKELGQANKLPPDAILLRGAKQAVKIASEIEYSKKALQETRKMPYTAV-DSLAYMTAKAVSGLISQVNYTKQLGQALSHYNITADTPAIKQAKKAQQIQSQNTYSKDRPTKYTTVYDSPAMKQASNLSKIQSD
(SR02) IPYKQEVKESTGKFFPVQDTPTYQHHLQLTKYESD-----NVYKDEAKHIMQQGHSQYPFTPEMFLVGDAERIKSE--RLYKANYEKWKSTYTMHADEPSITRAKDMGTLSSENTYRKMY-EDSKAQGNTLPADAINIKRAKEAMEISSEINYVKKARK--DNKPHIAV-DSLSYMTAKAVNGLVSQVKYKENLDKALASYSLPSDTPEILLARKAQQLQSQHRYTKDKPSKYTTVYETPLMMQASNVSKMQSD
(SR03) IPYKREVKENTGSFFQVQDTPTYQHHLQLTKYESD-----HRYKDEAKFIMEKGHSQYPFTPEMSRVEGAEKIKSE--RLYKENYEKWKSRYNMGVDDPQIVHAKEVASLQSENKYKEQY-EKDKEQGKSLTVDSVNIQRAKDAMEMASEIKYMREAQKVAQKTPFNAV-ESLTYMTAKAVSNLNSQLNYTAKLREALSQYQLTADIPAIKEARRAQQIQSQHNYTKDRPTKYTTVPDTPLMLQASNVAKLQSD
(SR04) IPYKKEVKESTGKFFPVQDTPTYQHMLELTKYESD-----HKYKDEAKFIMQKGHSQYPFTPEMTLVEDAEKIKSQ--RLYKENYEKWKSRYTMQVDDPQIVHAKEVATLQSENKYKEQY-EKDKEQGKSLTVDSVNIQRAKDAMEMASEINYVKDAKKVAQKNTHGAV-DSLSYLTAKAVSNLTSQVKYTEKLHEALTQYQVTPDMPAILEARRVQQIQSQHNYTKDKPTKYTTVYDTPLMLQASNVAKLQSD
(SR05) IPYKKEVKESTGKFFPVQVTPTYQHQLELTKYESD-----HKYKDEAKFIMEKGHSQYPFTPEMTLVEDAEKIKSQ--RLYKENYEKWKSRYTMQVDDPQIVHAKEVATLQSENKYKEQY-EKDKEQGKSLTVDSVNVQRAKNAMELASEINYMKDAKKVAQQNTHGAV-DSLSYLTAKAVSNLTSQVKYTEKLREALTQYQVTPDMPAILEARRVQQIQSQHNYTKDRPTNYTTVLDTPLLKQAANAAKWQSD
(SR06) VPYKKDVKESTGKFFPVQVTPTYQHMLELTKYESD-----HKYKDEAKFIMEKGHSQYPFTPEMFRVDDAEKIKSE--RLYKANYEKWKTGYTMLVDDPNIIHAREVAALQSENKYKEQY-EKDKGQGKSLTVDAVIIRGAKDAMELASEIKYVEESRKEAQKMPFNAV-DSLAYLTVKAVSNLNSQLKYTEKLPEALSQYHISADIPAIKEARRAQEIQSQYNYTKDRPTKYTTVYETPLMKQAELAAKHQSD
(SR07) SLYKKEVKESTGKFFPVQVTPTYQHMLELTKYESD-----RNYKEEAKYIMEQGHSQYPFTPEFTRVEDAEKVKSE--RLYKENYEKWKSRYNMQVDDPQIVHAKEMASLQSENKYKEQY-EKDKEQGKSLTVDSVNIQRAKDAMEMASEINYVKEAKKGAQKNTHGAV-DSLSYLTAKAVSNLTSQVKYTEKLREALTHYHITADMPAIHDARRAQEIQSQHNYTRDRPTSYTTVYETPLMKQAELVAKLQSD
(SR08) IPYKKDVREAEVKQFPVHDTPTYEHMLQLTKYQSD-----RVYKDDAKYIMEKGHSQYPFTPEISRVEDAEKVKSE--RLYKENYEKWKSRYTMLVDDPQIVHARDVALLQSENKYKEQY-EKDKEHGKSLTVDSMNVQRAKDAMEMASEINYVKEAKKVAQKNVHGAV-DSLSYLTAKAVSSLNSQVKYTEKLREALTHYQITHDIPAIKDARRAQEIQSQHYYTKDRPTSYTTVYDTPLMKQAELVARLQSD
(SR09) IPYKKEMREAEVKQFPVHDTPTYEHMLQLTKYQSD-----HVYKDEAKEIMEKGHSQYPFTPEMFRVDDAEKIKSE--RLYKENYEKWKSRYTMLVDDPQIVHARDVALLQSENKYKEQY-EKDKEQGKSLTVDSVNIQRAKDAMEMASEINYVKEARKEAQRKPFSAV-DSLTYLTAKAVSELNSQVKYKEKLREALTQYHITADMPAIHDARKAQEIQSQHNYTKDRPTSYTTVYDTPLMKQAELVARLQSD
(SR10) IPYKKEMREAEVKQFPVHDTPTYEHMLQLTKYQSD-----RVYKDEARYIMEKGHSQYPFTPEMFLVEDAEKIKSQ--RLYKENYEKWKSRYTMLVDDPNIIHAKEVALLQSENKYKEQY-EKDKEQGKSLTVDSVNIQRAKDAMEMASEINYAKDARSEAHSKAYNAV-DSLPYMTAKAAAEISSEVKYTEKLREALSQYLITHDVPAIQEARRAQQIQSQHNYTKDRPTSYTTVYDTPLMKQAELVARLQSD
(SR11) IPYKKEMKEAEVKQFPVHDTPTYEHMLQLTKYQSD-----HVYKDEAKYIMEKGHSQYPFTPEMSRVEDAEKIKSE--RLYKANYEKWRADYSKSVDDPDIIHAREVGEQQSENKYRELY-KKERGKTKALPPDTMDNVRAKAASEISSDVRYTQDAQKEARSKPFNAM-MSLATLTAKVVAGLTSQKKYTDRAARIRSRFNMPADTPTIQEAKRVQELQSEYLYTKDRPTTYTTVYDTPYIKQAEQVAKLQSD
(SR12) IPYKKEMKEAEVKQFPVHETPVYLNQLQVSGFQSD-----HVYKDEAREIMEKGHSQYPFTPEMSRVEEAEKIKSE--KLYKEAFEKQRGHFTVRVDDPQITHAKETTDLQSENKYKEQY-RQDKLKGHMLPPDAMDNMRAKEAMDIASEVKYRKKTRDEMQNQMFTSV-DTLGNLTAKACMDMISMVKYKEEAEKMRPQLKMPPDLPEVKLAKEVQQLQSYQNYTKDRPTSYTTVYDTPLMKQAELVARLQSD
(SR13) IPYKKEMKEAEVKQFPVHETPVYLNQLQVSGYQSD-----HVYRDEAKDIMEKGHSQYPFTPEMTRVEGAEKLKSE--KLYREAFEKQRGHFTVRVDDPQITHAKETTDLQSENKYKEQY-RQDKLKGHMLPLDAMENMRAKEAMDIASEVKYVKKTRDEMQNQMFTSV-DTLGNLTAKACMDMISMVKYKEEAEKMRPQLKMPPDLPEVKLAKEVQQLQSQQNYTKDRPTSYTTVYDTPFIKQAELVAQLQSD
(SR14) IPYRREVKEAEVKQFPVHETPVYLNQLQVSGYQSD-----RNYRDEAKYIMEKGHSQYPFTPEMSRVEETEKLKSE--KLYKEGFEKQRGHFTVRVDDPQITHAKETTDLQSENKYKEQY-LQDRGKGQMLPLDAMDNMRAKEAMDIASEVKYVKKTRDEMQNQMFTSV-DSLSNLTAKACMDMVSQVKYKEEAEKMRPQLKMPPDLPEVKLAKEVQQLQSYKNYTKDRPTSYTTVYDTPFIKQAELVAQLQSD
(SR15) IPYKKEMKEAEVKQFPVHETEQYKYMLQVSGYQSE-----NNYKDEAKYIMHQGHSQYPFTPEMSRVEETEKLKSE--KLYKENFEKQRGQYSVRVDDPQILHAKEADSLQSENKYKEQY-LQDKLKSQFLPLDAMDNMRAKEAMDIASEVKYTRPTRDEMQNQMFTSV-DTLGNLTAKACMDMISQVKYKEAAEKMRPQLKMPADLPEVKLAKEVQQLQSHQNYTKDRPTSYTTVYDTPLIKQAELVAKLQSD
(SR16) IPYKKEMKEAEVKQFPVHETPVYLNQLQVSGYQSD-----HVYRDEAKTIMEEGHSQYPFTPEMTRVEEAEKLKSD--KLYKENYEKMKGQFSVRLDHPALIHAKEVADLQSERKYKEQY-LQDKLKSQFYPVDAMENIRAKEATDIASEVKYTDKARKEMQNQMYLAV-DSLTNMSAKLVHDLVSQVKYKEAAEKMRPQLKMPPDLPEVKWAKEVQELQSYKNYTKDRPTNYMTVYDTPAIKQAEQATMIQSD
(SR17) IPYKKDVKEAEVKQFPVHETSQYEFMLQVSGYQSD-----HNYRDEAKYIMEKGHSQYPFTPEMTRVEEAEKLKSE--KLYKENSEKQRGHYSVRLDDPQIQHAKEADSLQSENKYREQY-LKEKGQGLMLPLDSMDNQRAKEVSELVSEVKYAQDARREAQEKMYNTV-DSLTSMTAQVVSQLTSQVKYKEKADKERSQHKVPADTPEVAWAKEVQQLQSTKSYTKDRPTNYIPVYDSPHIKQAEEATKLQSS
(SR18) LQYSDRDAGTLHSEFRGQRYTPLETLEKKHTDDIRNIVSDVVYKDEAKEIMQKGHSQYPFTPEMTMVEEAEKLKSN--HLYQENYKKYLTNFTIRADYPEFVFAKETSDLQSENKYREQY-NKEKGQGLILPLDTMDNLHAKEAVALASEVKYTQDARKEAQEKMYSPV-NSLTLMTAKMVTDLTSQAKYREQVQSTLAQHKMPPDTPEILWAKEVGKLQSQNEYIKGKSTQYTPVFDTPAIRQADEASRLQSD
(SR19) IPYKYEVKNLVGKQFPVHETEQYKYMLQVSGYQSD-----HVYRDEAKTIMEEGHSQYPFTPEMTRVEEAEKLKSD--HLYKENYEKWKQKYTSHPNQLGFVTAKEIAEVQSESNYREKY-LKERGTGYFVPADNLEAVRVKEATELASEVKYSQDALNEAHNKKFDAVGDSVVVKTAQEVSELTSEVKYKEQAEKIKSQHKVPADAPDFVWAKEVGDLQNQRLYAKDKSDKYVPVLDTPVNLHAKEAGALQSH
(SR20) LPYSDRNAGTLFSEYRGQRYTPLVTLEKKHTDEVQSMISDVIYKDEAKEIMQKGHSQYPFTPEMSRVEEAEKLKSE--NLYKANYEKHKQVYTQTPNTAHITACKEAAELQSEMKYREQY-EKGKGHGYFLPADSMENVRAKEASQLASQILYTHEAQKEAQTKPYNPV-SSLTLLTAQAISELTSEVKYSEEAEKTKSQHKMPPDAPEIQRAKEVGQLQSQVEYAKNKTNQYTAVLDTPSNRHAVEATRLTSE
(SR21) LPYKEEAEDLIRNKAFAAQDTPVMDRVKKVSDLQSD----VVYKADAKHIMSLGHSQYPWTPDMARVQEAETIKSE--RLYTRDSNDQKKKFTILPETPEVALAKEVAKLQSEKQYKEQY-QKERGKGLHLPPDAVENLRAKEANQLASQIKYTEEARKSAEGKKFDAVAETPVFKTAVEAAENASETNYMKDYEKGKTKYNLSSDTPLIKTARQSANLASEFKYQQPVKGAKFEQICDGMREVHCATVSKLQSD
(SR22) IEYKQPLE-NYTVITDTPEFRKLASLQEIVSD--------IFYKEEAKEIIERCCSQAPIYLEMDRMARLQDLLNM—-TQYREKWEMDKKKYTVLTDSPEIIRVVEAGKQQSDLEYKKGM—QKQQKKGYALVPDSTSFVAAKEAAKIASDVEYKTKDLIDMRKASFKNVHESPWYLQAKLSAENASGIKYIKDYEKEKVHYLTPVDRVDFVAAKAVSGMTD-RKYRQKLGKPISIPDTPEIVRSKKLEKLKD
(SR23) QTYKQEPG-PFISTPDSMEMDRVRTLQQITSE--------QMYKEDAEWLVQRCLSQYPFSVEIDRVTKVKFQNDP--RVYQWEADRDKQKYTVLADSPEITRVVNAAKMQSDRVYHQGY-EKQKQGGYRLAADAPTFTTAKEAAKNASDYNYQSGLREA-RGKGFPNFQESPLFLQNKLSAQLASGAEYTKDYEKQKVKYTLPADRVDMVTAKNILELSD-KKYRQEPGKFISIPDTMEQVRCTKAAKLASD
(SR24) KHYKQEPG-AFISTPDSMEMERVKTLQEITSD--------QYYKEDAQWLVQRCLSQYPFSVELDRVNKVKFQNDP--RIYQWEADKEKQKYTVLADSPEITRVVNAAKMQSDRLYHQDY-EKQKEGGYRLAADAPVFLTAKEAAINASDHVYQSGLREA-RGKGFPNFQESPLFLQNKLSAQLASGAEYTKDYEKQKVKYTLPADRVDMVTAKNILEQSD-RKYRQEPGKFISIPDTMEQVRCTKAAKLASD
(SR25) KHYKQEPG-AFISTPDSMEMERVRTLQDITSD--------QYYKEDAQWLVQRCLSQYPFSVELDRVNKVKFQNDP--RLYQEEADREKQKYTVLADSPEITRVVNAAKMQSDRVYHEGY-EKQKQGGYRLAADAPVFLTAKEAAINASDYNYQSGLREA-RGKGFPNFQESPLFLQNKLSAQLASGAVYTKDYEQQKLKYTLPADRVDMVAAKAVLEQSD-KKYRQKPGKFISIPDNMEQVRCNAVAKLASD
(SR26) KHYKQEPG-AFISIPDSMEMERVKTLQEITSD--------QYYKEDAQWLVQRCLSQYPYSVELDRVNKVKFQNDP--QLYREEWDRDRQKYTVLADSPEVTRVCEVAKLQSDLVYHQGY-EKQKQAGYKLAADAPAFLTAKESAKNASDYNYQSGLREA-RGKGFPNFQESPLFLQNKLSAQLASGALYTKDYEQQKLKYTLPADRVDMVAAKNILELSD-RKYRKEPGKFISIPDNVEQVRCNAVAKLASD
(SR27) KNYKQEPG-KFISIPDSMEMERVKSLQEITSD--------QYYKEDAQWLVQRCLSQYPYSVELDRVNKVKFQNDP--QLYREEWDRDRQKYTVLADSPEVTRVCEVAKLQSDLVYHQGY-EKQKQAGYKLAADAPAFLTAKESAKNASDYNYQSGLREA-RGRGFPNFQESPLFLQNKLSAQLASGALYTKDYEQQKLKYTLPADRVDMVAAKNILDLSE-RKYRQRPGKFISIPDNVEQIRCNAVAKLASD
(SR28) KHYKQEPG-KFISIPDSMEMERVKTLQEITSD--------LYYKEDAKELVERCLSQYPFSVELDRVNKVSMMQDP--RLYREEWDRDRQNYTVLADSPEVTRVCEVAKLQSDLVYHQGY-EKQKQAGYKLAADAPAFLTAKESAKNASDYNYQTGLREA-RGRGFPNFQESPLFLQNKLSAQLASGALYTKDYQQQKLKYTLPADRVDMVAAKAVLDLSD-KKYRQEPGKFISIPDNVEQVRCNAVAKLASD
(SR29) KYYKQEPG-KFISIPDSMEIERVKTLQDITSD--------YYYKEEAKELVERCLSQYPFSVEIDRVNKVSLMQDP--RLYREEWDRDRQKYTVLADSPEVTRVCEVAKLQSDLVYHQGY-EKQKQAGYKLAADAPLFLTAKESAKNASDYNYQTGLREA-RGRGFPNFQESPLFLQNKLSAQLASGALYVKDYQQQKLKYTLPADRVDMKAAKAVLDLSD-RKYKQQPGKPISIPDTVEQVRCNEVAKLASD
(SR30) KHYKQEPG-RFISTNDTPEILRLKSLQEIVSD--------HYYKEEAKEIVERCLSQYPFSVEIDRVHKVSEMQDP--RLYREEWDRDRQKYTVIPDSPEVARVQEVAKMQSDLVYRQGY-EKQKQGGYKLAADAPLFLTAKESAKNASDYNYQTGLRAA-RGSGFPNFQESPLFLQNKLSAQLASGALYIKDYQQQKLKYTLPADRVDMVAAKAVMDLSD-RKYKQKPAVPKLPPDSVEQVHFKSLAPLKD
(SR31) KVYKHEPG-GFTSEIDTPETRRLKSLQEIVSD--------LYYKEEAKEIVNKCDSQYPFSVEIDRVHKVSEMQDP--RLYREEWDRDRQKYTVIPDSPEVARVCEVAKLQSDRIYQLGY-EKQKQEGYKMIPDTPLFLQCRESARNASDYVYQTALKEA-RGRGFPNFQESPLFLQNKLSAQLASGALYTKDYEQQKLKYTLPADRVDMVAAKNILDLSD-RKYKQKPAVPKVFPDSVEQVHFKSLAPLKD
(SR32) KVYKHEPG-GFTSEIDTPETRRMRTLLETVSD--------YYYKEEAKEIVEKCCSQFPFTVEIERVEKVTEMQDD--RRYRADYERARHNYTVIPDSPEVARVIEAARMQSDRFYHEGY-EKQKQEGYKMVSDTPLFLQCRESAWNASDYNYKTALEEA-RKSQYKNVHESPLFLQNKLSAQLASGPLYVKDYEQQKLKYTLPADRVDMVAAKNILDLSD-RKYKQKPGVPKVFPDSVEQLHVKSLAPLKD
(SR33) KVYKHEPG-GFTSEIDTPETRRMKSLWEMVSD--------YYYKEEAKEIVEKCCSQFPFTVEIERVEKVTEMQDD--RRYRADYERDRHNYKVIPDSPEVARVIEANRMQSDLYYKEGF-EQQKKEGYKMVSDTPLFNQCRDSARNASDYNYKTAIKQA-QEAQYKNVHESPLYLQNKLSAQLASGPLYVKDYEQQKLKYTLPADRVDMVAAKNILDLSE-RKYKQKPGVPKVFPDSVEQLHVKSLDPLKD
(SR34) KVYKSEPG-RFTSEIDTPETRRMKSLWEMVSD--------HYYKEEAREIVEKCCSQFPFTVEIERVEKVSEMQDA--KKYREKYDEEKQKYTVIPDSPEVLRVIEAAKMQSDLHYKDGF-EQQKKEGYKMVSDTPLYNQCRDSARNASDYIYKTAIKQA-QEAQYKNVHESPLYLQNKLSAQLASGPLYVKDYEQQKLKYTLPADRVDMVAAKNILDLAD-RKYKQKPGVPKVFPDSVEQLHVKSLDPLKD
(SR35) KVYKSEPG-RFTSEIDTPETRRMKSLWEMVSD--------HYYKEEAREIVEKCCSQFPFTVEIERVEKVSEMQDA--KKYREKYDEEKQKYTVIPDSPEVLRVIEAARMQSDLHYKEGL-EKQKKEGYKMVSDTPLFLQARESAWNASDYNYKTALKEA-QEKQYKNVHESPLYLQNKFSAQLASGALYIKDYETGKVKYALPPDRVDIVAAKNYQNFSD-RKYKQKPGVPKVFPDTIEGVRVKSLEPLKD
(SR36) KIYKQEPG-RFTSEIDTPETRRMKSLLEIVSD--------HYYKEEAKEIVEKCCSQFPFTVEIERVEKVSEMQDA--KKYREAWDEAKQNYTVIPDSPEVLRVLEANRMQSDRIYHEGL-LKQRDEGYKMVSDTPEYHRARFAAALASDYLYKSALKKA-QETQYKNVHESPLYLQNKLSAELSSGARYIKDYGENKAKYQLPADVWDMVAAKEAIARSD-RHYKQKLGSPKVFPDSIEGVRVKSLEPMKD
(SR37) KVYKQEPG-RFTSEIDTPETRRLKSLLEIVSD--------HYYKEEAKEIVEKCCSQYPFTVEIERLEKVAEMQDA--KKYREKWDEDKQLYKVIPDSPEVLRVLEAAKMQSDRLYQKGL-DKQRDEGYKLVADTPEYRRILQTTAQASDYLYKAALREA-REKQYKNVHESPLYLQNKLSAEMASGARYVKDYETMKVKYSCPADRVDLKTAKLVAGFSD-LKYKHIEARYTPVADTVQNEHAKTVAGFSD
(SR38) LKYVQKVSAKSLSETDTPVMRRIKSLQEITSD--------NLYKEEAKEIVEKCCSQYPFTVEMERFEKLLELNDKASKLYKKDYEEERQKYHVYPDTPEVKRVIDAAILQSDAKYREKYEKEMKGKGNFPQHITPVY-ASQKEAAGLTDAVYSRDAREKMQEKSYKNFHESPLYRTLKDDTDMASMSNYVSDWEKRKVDYKVVSDTPDIKRVKKAGTLASPRTYVQKQEKYETVADTPANILANESGKLVGD
(SR39) RHYKQVVKGKGIMDTDTPEMRKIKSLREITSD--------NIYKEEAKEIVEKCCTQYPFTVDMERFEKLMELMDQ--KRYKKNFEEERNNYHVYPDAPDIKRTLAMAQLYSDLKYKEIYESQIKGKGNFPQHITPVY-KSQIEAAKITDAVYKKDAHDEMQKKSYKNFHESPLYRTLKDDTDMASMTRYRDQYEKEKTNYKITTDTPDIKRVKKGGILASERLYKQKCDSYETVADTPANIHAKETGKLHSD
(SR40) LPYSNRDAGTLRSVYRGQRFTPFDTPESNRLKSLRELASDNVYKEEAKELYEKCCTQFPYTVEMERLDKLKELYDK--GLYMKLVDEARQ-YHLFPDCPEVKRVIDADKIQSDLVYKKAY-EEARGKGYFKVTETPLYKTLVKAALDASPDHYSRQALTEMREKNYTAVHESPLYKMQKENTDNQSPHKYNKFPETRDNF-KTPVDTPDLKRVKKAGDIASEIKYQKDKQTEKYIPIADTVAVRHAQHMGEIQGY
(SR41) LPYEKHDAGTVRARYRGQRFTPVDTPESNRLKSLKEIVSDTIYKEEAKEINEKCCSQFPLTPEMERFWRLQDLYSD--ADYRQKALDTRGDFHLYPDIPEFERLNKMNDVQSDLKYQERYMKELRGKGYKLTPDMPLYQRMKRVTAQNSDNVYKGEAKKLLGKSIDVTQTPEYARQQEAQKISSQ--SEYVAKDKERTHQYETTPDTPEYRRQRKVAKFTPDKAYQTAKPDLDKYHPVKDTPSNERADQAQKIQSD
(Neb) LWYQEQGRDLKGKAYTEVDTPVNRRVKSLQEITSD
(Neb) TFYKEEAQHLWHVACTQYPYTPDMERHEDNLRKFSD
(Neb) VLYKNKYEMNKHEYHPVPDSPELNLAKENAVRHSG
(Neb) FKYREKYEELKKKPKFNIAETQLYRRMRQVAADTCD
(Neb) DNYMGDYNKNIKGKGVANYAETPLYQRNKEAGDASSE
(Neb) LKYGKDLKKIREGKGYTPTADTPQHRHVKKMEEIQSS
(Neb) VKYPKALKDIAKYATVLDTPDNQRAISTYKNQST
(Neb) LYYGADAKNEMIGKAFKEVNETPASAHHKKVLDMSD
(Neb) VVYKDEAKRLHTVACTQYPYTPENLRMAELSRTQSD
(Neb) QMYKERGELEKHKYTTLANTPDLLHAKEVHDITSE (Neb00/01)
(Neb) NKYKEEYNKERGKYSMVAQTPLYETAQKANKLTSE (Neb01/02)
(Neb) TLYKKAYKALQGKGYALIDTPEFERCKQVNKLASR (Neb02/03)
(Neb) TKYHEDYEKSKIKGYTPVVDTPENIRLRQQSKIQSQ (Neb03/04)
(Neb) NEYQKGYKALAGKFSYTADNPEHNRIRSVSKIQSQ (Neb04/05)
(98 nebulin repeats with Unique sequence)
(Neb) IHYGHTRLEQHGQKYHAAIDDPELKRLQEVGKFQSQ (Neb05/06)
(Neb) VLYGEIKDKDDASKGRGKYTTIQQDPEMKRLKEANAYASE (Neb06/07)
(Neb) AQVAVRKFDEKNRYTTVLDDPELNRMKNVGKFQSE (Neb07/08)
(Neb) LQYRDKKTRKTGARYTNVLDDPEMNRIKGVTKMLSE (Neb08/09)
(Neb) VQYRDRELKAGRLTAVTDDLDTARVKRNTKIQSE (Neb09/10)
(Neb) LQYRDEELKVGKLHVVTDDIDAERIRRNTKIQSE
(Neb) TLYRKDFSKGRGTFTGTNGIMDSLDYERIKRASKLCSE (Neb) NEYRKGRKINMYNAVLDTPDMARIRNATKISSE (Neb10/11)
(Neb) VKYHEDFEKQKGKAYHTVIDDPELTRVKKNTVIQSN (Neb11/12)
(Uni) LAYQGVKEKLQTMESRRQMIPGEHPMPQPAGQQLGQMEARMAADASYAQFKAKGKGKVGG (Neb12/Uni)
(Neb) RYMPIRLGDPGSIFDYEPYQGKEEAGRAIY (Neb13)
(Uni) VTTATWKPAKPIRTIEDFVVEESTDVSAAKVEEAMEARDKLTEMDMEYERRQGEGLSYEYDPEKYSHHYWSKTTGELGGVVTTSGMKYRKIEDFLNLNADLIYTTENESSTRESASESSGMWDYDPDKYWKARGYRISAEQRYQQVRLKPGKIEDYDPMTHPDAQQPKFEAPAAVSEPEPTIQSG (Uni)
(SH3) QNVMRAVFDYTAAEDDEISFMDGDVIVNCVKIDDGWMTGTVQRTGQSGMLPANYVEPMS (SH3)

49
Fig. 3 The giant exon of 98 nebulin repeats with a unique sequence. The tyrosines conserved among the nebulin repeats are marked with black. Underlined area, coding site in a B. floridae neurula stage EST clone (DDBJ accession BW855343).
MSHPKFDSQESAPYQQGAMWVLTGCSRLALVNCNNNNNNG
SSDDENDGNGNAQPGEHETPAAYNSEQNPGQEDTESDNAD
QAEGNGAFGTEGVQDQGQGDDSPPPGEAQEEEPPQESPSI
EVTPPEDSPPQQDVFEEETQPKEPEGPIIEMAPSPTVEEA
GIEQAMEAEEGPSEQFTELPLSTDAPLEPIRTPVVEEPEE EQPAEYEEAEDALIDFDLPSPPPEKVSPDEYRAVSPREQP
TPEERPVSPRATSPAEEAVPPERPPSPMEEDVFVEPAFPS
PAEATGRPDVPTPPYMEQAPSPVVPDALMEAVEPEVPETP
MEPDFPSPPSYNTLTEDVVRDVPVHVEKPSQRTASVFTNP
DELDEEEVGNPHPLAGNLGHKTGNPHPHNPGNPHPHNPGT PPPIEEKSSSPQPVQASGGRLVKTGSYSSYQYEESVREVR
SSTKQTIREVVSSDGKVISKQESEETSHAEASQARAKSET
SRSDGKQTVTTKREAEESMQALEMQKKKKTTSGGVTTVEP
TQGMRAMEMERKAKASTTVQQAVGAAMEVEPPPPIPPLPV EIGQVTIKQEPGTLRGISLKGVSKSRGGVKHTIKTTNALS
EERSSQVTTSAMSEQRSSHVTTSSQSEQRASHVSSSSMSQ
ERASHVATSAMTQERASHVSTSAMSQERASHVTSVRAAKA
VHTGVTKKVESQPKIVVARRTETVKIQLPRGSRERVLRIK
IRFGKKDKPNRLTAYAWSQLRWPAITEVKTQEDVSEDTRY VYILLKEISPDRVAKESCLRVQVPPVIEQSIMKISAETSR
MLVQQQKVIYKAHERKYGSGQKYKFTVQQAVPPELDESPT
LSARDAKSRFEMARAEELRQRRHVTRHEVGQMKEMFERGA
SDAEDYEREPAQITRQEVGEKKEAFERGVSEVRMAEREEI QVARQDIGQKKQLFEKVAEGHWASRYMDSDEGGEAPERPP
PPRFDVGQKKEIFENVGQSWRRTDSETEEALQMITQQDVV
RKRELFERGRSGSEEETPERPPPPRQDVGQKAELFEKVAV
GQFVRQEGEERERVIPIRPPTPSHDMIQKREMFQQFSQGR
WSSSETEEREEIQIAAGVSTKKELFEKAAEAKLLPPPSPR RMRRRIRSDSFRRSMSSDDEGTPERPPPPRPPPPAPAPPT
PSPPASPVTTAPPSRPPPPPEGEEGEDWMASVSKCVEMYE
REVQIRGERMKLKTIRRDPTPVRRQPRSWPPTPPSSPAEP
ISPVRRTRRSPSPGPPVADRPRGREPPPVARRFRDRSPAV HREHSSERRLTNLAQNILSQEGFEEYRAVPQDTYQVVVDP
FLQKAQEVTKLLSR
AEYEKEAKDRMWRYQGGLPDEAEARRRAELARMQSD
VRYRDYPRYLEAKPGMVAVVDRWMIGQKKQTHEQST
VEYPRDHPKDMTWRLKYQSTPDDTVTKRAQEVTHISSE LEYQKRAKQTKDRYTAITDELQLETQKDAARMQSS
VSYVKDHPLAMEKRHAYTSVEEDPSSKHAKDAYKMSSQ
LEYEKRYKQEVLGHVGYIELDTPEQERIRRMKTLQSQ
LEYQKDYPMDFERRKDYQSVHDDPNTIRAKEATHLASK VEYESRYHKEKDIYKTVADTPEGVRVKDVGKMQSE
IEYQKPDKERRQYYQTITDAKDSQRLKEVGHLQSQ
IEYQKDHPMGFEGRRDYQAVSDDPETLRVKDVTKLSSG
VEYVRKDKPRREVYQTVTDDPEGKRMKEVGKLHSQ
LEYQKDHPMTFEGRLDYTSIADDPETRRVKDVTMMASG IEYERKDKPRREVYQTVTDDMESKRMKEVGHLQSQ
IEYQRDHPMTLEGRLDYTAVADDPETRRVKDMTIMASG
IEYERKDKPRREIYQTVTDDMESNRMREVGKLHSQ
IEYQKDHPMTLEGRLDYTAVADDPETLRVKDMTLMASG
VEYQRKDKPRREIYQTVTDDMESNRMKEVGKLHSQ
IEYQKDHPMTLEGRLDYTAIADDPETRRVKDVTMMASG
IEYERKDKPRREVYQTVTDDMESKRMKEVGHMQSQ
IEYQRDHPMTLEGRLDYTAVADDPETRRVKDMTIMASG
VEYERKDKPRREMYQTVTDDMESNRMREVGKLHSQ
VEYQKDHPMTLEGRLDYTAIADDPETRRVKDVTMMASE
IEYERKDKPRREIYQTVTDDMESKRMKEVGKLHSQ
MEYQKDHPMTLEGRLDYTAVADDPETLRVKDVTKLSSG IEYQKKDKPRREIYQSVTDDVATVQAKDSQRLQSQ
LEYERLYREKKGTGIFTMIADDKNLQHVKDVSKLGSQ
QGYTKDFPMELEKRGGYTSVEADPTTERAKEVYQFQSD
VEYQRNFRQRRGSFTAITDDAATKHAQESGKVSNE LAYTKNFRTRRGSFTAVVDTPEAQRVKEVNRYQSE
LEYKKRALQRRGSFTAVVDDPVTERVKDSTRLASD
TIYKAAAHERKQAYTIALDDPNTERVKNVSKMVSD
FEYQRMYREKRGTGIFTSIADDPFIKHAKETAEYLSS
NVYKKDFPENMERRSGYTTIVDASGLKHAKEASEIQSL IKYYEKGKEGKSKYTLVTDDPESMRMKDVNKMQSR
IEYEKDYEASKDKYTMVGDDPQTRRVQEMTKLQSQ
YEYQRTFLEERGLGWYTAWLDYDKEMERQLGMKHMYSQ
VKYKKDFPEQMTGREGYQMVADDVQTVHASDMSKMQSR VEYEKDYHEKKGLYTTTTDEMRATHAKEVGKLASS
VEYPVDHPMEWERRRKYTAVADDPNTQRVKETTKYQSP
YEYEREHRRKKGQYTMVADDREQVRVVEMNKLHSL
VQYHETFEKEKGQFTTVLFDPETKRVVELSKLQSM
NYYQQEFEKNKGNYITVLFDPESQRYKEVGKLASD
VEYQKEFKDLRGKYTATADTVATKHATEAGKLQSQ
LEYQKDFPQQMTERAGYTPVTDKETIRVVEMSQLHNL
IKYREQFEKDKDRFTTVLFDPETKRVVELSSIQSV
IQYEKKYQANKDKYTTVMFDPETSRTMEMSKLQSL
NEYHLQFERDKGQYQLVTDDPVSERIREVSRHQSQ
YEYVRKWLEERGIYHSAPDDKEHMRIVEVNKLQSL
LHYTDEYNKEKGKYTTILFDPETKRVVELSKLQSA
VQYHEEFEKNKGNYITVLFDPESKRYKEVGKWQSD
VEYQKEFRGLRGKYTSVEEDVQTKRAKDVGKYQSQ LEYQKDFPQEFEGRTGYTPVADKETVRVVEMSQLHSL
LKYQEEFEKDKGKYTTILFDPETKRVVELSSIQSA
IAYQKEYQKDKDQFTTILYDPETQRFTELSKIQSL
VSYHDEYEKEKGKYHSVADDIDSKRLKDVGKFASE TEYQKEYQKLKAFYTMVPDKEMLRVIEMSQLHSV
VKYHEEYEKDKGDFTTVLYDPETRRVVELSSIQSI
NKYQEEFEKNKGKYTTILFDPETERVQEVGKIQSN
VEYQKAYHEMKGSYRTVTDDPESERLKSVSKWQSM
SEYQKDFPSKFEGRPEYQSLAADKETLRIVDVTKLQSL IKYQEQHKDDIGKFTTVLFDPETKRVLELSKVQSL
LKYKEEFESEKGGYTTVLYDPETQRFIDLSSKQSD
VHYHEQYEKEKGQYTSVVDDVETRRQTGITKVQSS
VVYPKDYPLRYEGRSGYTTIPDDKELLRVVNLQKIHSL
IRYNKEYEDEKGAYTTVMFDPETKRLIELSKIQSL
IKYQEQFQKEKGSFTTVLFDPETKRAIEVGKIQSQ
VEYQKDFKEGRAAQSGAEADKEILRLVDMSQLHSI LKYQEEFEQGKGKYTTILYDPETQRVVELSSIQSA
VKYEDEYKKNKGQYTTVMFDPETKRLLELSEIQSA
FKYQEDFERGKSIYSQIPDDVQTKRAEEVSHFQST
VEYGKEARAQLPQYTPVADEKEVVRMVNMTKLHSL
INYHERYEKERGGYTTILFDPETRRTMELSNVQSL VKYQEQFQQQKGNYTTVLYDPETKRVVELSKTQSQ
LEYQKDFPVAMEGRDAGVIDDQDKEVLRLVDISKLQSL
SKYQEEFQRGKGNYTTVLYDPETKRVVEVSSIQSL
IKYEEDYRRSKGQYTTIMFDPETQRLMEMSEIQSA
FKYHEDFEKGKGNIMQITDDVETRRATDVTHLQST
FEYRRDYPLEYQQRPGYTSSTTDKEMVRLVEMSKLHSL
AEYRSEYEKGRGGYTTILFDPETKRLLEISRVQSL
INYQEQFYREKGNYTTVLWDPRTKTAVESTKLQSQ
IQYQKDFPVGMEGRGGKGADEADKEILRLVDVSKLQSL
AQYHEEFKKDKGKYTTVLFDPETQRVIELSAVQSL
VKYEDEYKRNVGQYQTVLYDPETQRVIEMQKMQSV
FKYHEQFEQEKGQYTLATDSIDSRRAQEASRVQST
LLYRRDFPQEFEGRAAYIPIPDDKETLRIIELTKLQSL IQYQKEYDLSKGDYTTCMYDPETQRVLELTRIQSY
VNYKKNFEKEKGHYATIIYDPETRRCIELSKTESQ
IGYKKDFPITFEGRPHISTYGDKELLRLIDISNVQSV
IKYQEEFLRDKDKYTTVMYDPETKRVVELSRILSL
SKYQEEHQKGRGKYSTILFDPETQRALDLTQVQSS
VRYRKDYPQTMEGRAAYSASIDMSILRVIEMSQAIGV
TRYHEDYDIAKANYNAILYDPETRHVMEISKIQHL
DKYKEAFTHQVGPHVAVLYDPATKKAMNLSSISSQ
KDFRNEYVETFETIVKRAKNKVVLRLIDLTRVDNL
VAFEEELIIETSNYRTLLFNSENSRVIEISRLDSF
VNYEKRFKETKGSYTTVLYDPETQRALTLSKVTAQ
AEYQRQVFESSFGDRVSESTQFEEPQWSLDIESMA
KFYEDVASGKHREPEIPSYTEGTSSSKGAPKYRGKKKVGFSL
SGEELRGVDSAGKAELEVIYKGGDVVETKEQAEEVKQTIEQV GLKAAGAVKDEKPGVSVTSRRSDTPPEALGDGGQEEDDDTPM
TQDMVRRLIELHEQSMRGIVRTLDEDLAALNGNNNNNKVVEA
TRYRQPTWTVCTMADPEALTYCQVHGPRNYPLTVLNS

50
Fig. 4 A small transcript from the amphioxus nebulin gene. (A) Reverse transcriptase-polymerase chain reaction (RT-PCR) product using LIM and SH3 domain primers. (B) Amino acid sequence of the RT-PCR product. (C) Domain structure of the RT-PCR product and the product’s homology with human Lasp-2.
LIM METKCARCQKTVYPMEKLNCLDKYWHKGCFNCEVCHMTLNMKNYKGYNKIPYCNAUni HYPQSKPTVVSETNeb PEIARVMQQSQQQSLTKYHEDYEKSKIKGYTPVVDTNeb PENIRLRQQAKIQSQVKYHEDFEKQKGKAYHTVIDDNeb PELTRVKKNTVIQSNLAYQGVKEKLQTMESRRQMIUni PGEPSMPPPAQQLGTLMDPTHGMESMSIQDGHERMMQQQQQEEDRHLASHRRQE
NPPISIQSYQQVRLKPGKIEDYDPMTHPDAQPKFEAPAAVSEPEPVIQSGNVMRSH3 AVFDYAAAEDDEISFMDGDVIVNCVKIDDGWMTGTVQRTGQSGMLPANYVEPMS
337 aa, 1011 bp, Molecular weight 38,677
(B)
(C)
(A)
1000
500
(bp)M AM
1000
500
(bp)M AM
80%
46%
50%
53%
40%
65%
7%
AmphioxusLasp-like protein
Human Lasp-2
80%
46%
50%
53%
40%
65%
7%
AmphioxusLasp-like protein
Human Lasp-2
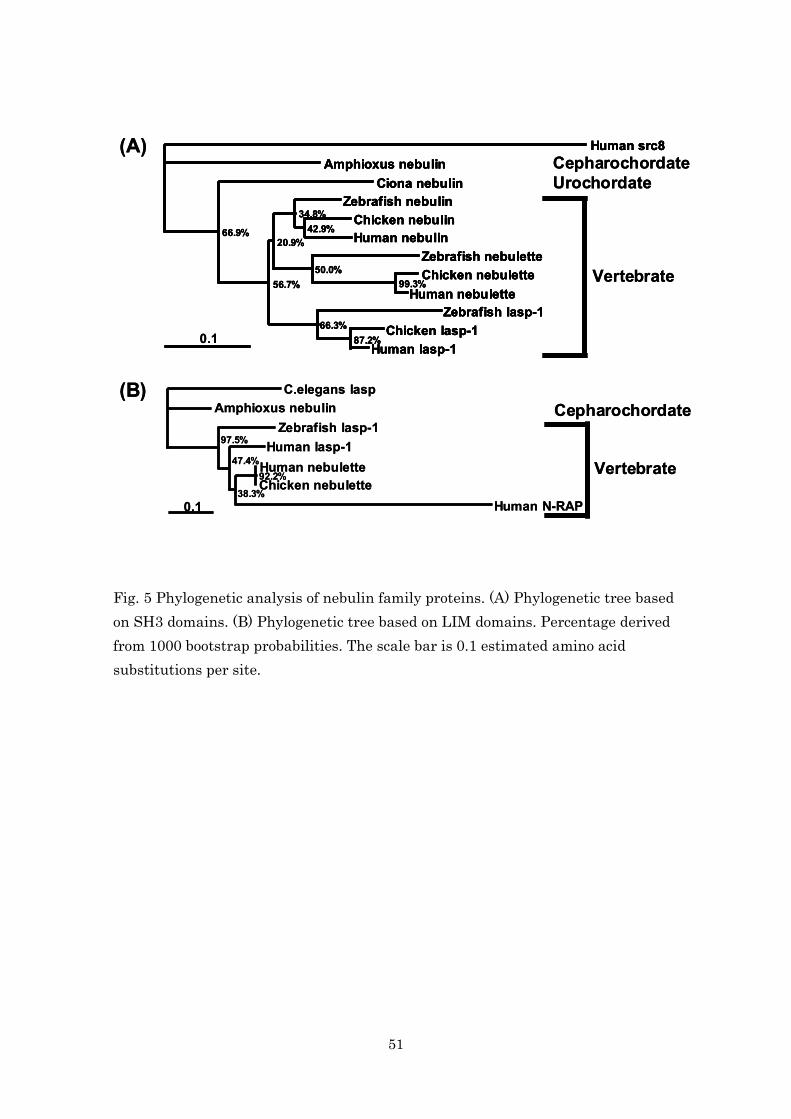
51
Fig. 5 Phylogenetic analysis of nebulin family proteins. (A) Phylogenetic tree based on SH3 domains. (B) Phylogenetic tree based on LIM domains. Percentage derived from 1000 bootstrap probabilities. The scale bar is 0.1 estimated amino acid substitutions per site.
Human src8Amphioxus nebulin
Ciona nebulinZebrafish nebulin
Chicken nebulinHuman nebulin
Zebrafish nebuletteChicken nebulette
Human nebulette
Human lasp-1Chicken lasp-1
Zebrafish lasp-1
34.8%42.9%
20.9%
50.0%56.7% 99.3%
66.3%87.2%
66.9%
(A)
Vertebrate
CepharochordateUrochordate
0.1
Human src8Amphioxus nebulin
Ciona nebulinZebrafish nebulin
Chicken nebulinHuman nebulin
Zebrafish nebuletteChicken nebulette
Human nebulette
Human lasp-1Chicken lasp-1
Zebrafish lasp-1
34.8%42.9%
20.9%
50.0%56.7% 99.3%
66.3%87.2%
66.9%
Human src8Amphioxus nebulin
Ciona nebulinZebrafish nebulin
Chicken nebulinHuman nebulin
Zebrafish nebuletteChicken nebulette
Human nebulette
Human lasp-1Chicken lasp-1
Zebrafish lasp-1
Human src8Amphioxus nebulin
Ciona nebulinZebrafish nebulin
Chicken nebulinHuman nebulin
Zebrafish nebuletteChicken nebulette
Human nebulette
Human lasp-1Chicken lasp-1
Zebrafish lasp-1
34.8%42.9%
20.9%
50.0%56.7% 99.3%
66.3%87.2%
66.9%
(A)
Vertebrate
CepharochordateUrochordate
0.1
C.elegans laspAmphioxus nebulin
Zebrafish lasp-1Human lasp-1
Human nebuletteChicken nebulette
Human N-RAP
(B)
97.5%
47.4%92.2%
38.3%
Vertebrate
Cepharochordate
0.1
C.elegans laspAmphioxus nebulin
Zebrafish lasp-1Human lasp-1
Human nebuletteChicken nebulette
Human N-RAP
(B)
97.5%
47.4%92.2%
38.3%
Vertebrate
Cepharochordate
0.1

52
Acknowledgements
I am indebted to Professor S. Kimura of Chiba University for her
continuing guidance and stimulating advice during my course of study. I
wish to express my gratitude to Professor K. Ohashi of Chiba University for
his thoughtful and helpful discussions. I am grateful to Professor K.
Kubokawa of University of Tokyo for collaborations on this study. Finally, let
me thank the members of Professor Kimura’s laboratory for their technical
assistances and useful discussions.


















