
Steady-State Visually Evoked Potential Correlates of ... · Chapter 1 Introduction ... related ERP...
220
Steady-State Visually Evoked Potential Correlates of Object Recognition Memory Thesis for Doctorate of Philosophy Andrew Pipingas, BAppSc(Dist) February, 2003 Brain Sciences Institute, Swinburne University of Technology
Transcript of Steady-State Visually Evoked Potential Correlates of ... · Chapter 1 Introduction ... related ERP...

Steady-State Visually Evoked Potential
Correlates of Object Recognition
Memory
Thesis for Doctorate of Philosophy
Andrew Pipingas, BAppSc(Dist)
February, 2003
Brain Sciences Institute,
Swinburne University of Technology

Declaration
This thesis contains no material which has been accepted for the award of any other
degree at any University and to the best of my knowledge and belief contains no
material previously published or written by another person or persons except where due
reference is made.
Andrew Pipingas
February, 2003

Acknowledgements
The author would like to acknowledge the following people whose contribution made
this work possible.
My supervisor and mentor, Prof. Richard Silberstein, for his invaluable assistance,
guidance and patience throughout the project and for providing me with the intellectual
inspiration to undertake a PhD project in the area of brain electrophysiology.
Mr David Simpson for the design and development of the instrumentation used to
conduct this work and for his assistance with many other computer-related and technical
matters.
Mr Geoff Nield for his assistance with the development of a suite of software programs
that were used to analyse the data.
Ms Barbara Livett for generously giving up so much time to assist with various drafts
and the English expression for this thesis.
Assoc. Prof. Aina Puce and Assoc. Prof. David Crewther for their helpful comments on
earlier drafts of the manuscript.
My parents for providing me with the motivation and support to pursue a higher degree
and, together with my brother, for providing continual encouragement and support.
Finally, I would like to thank my wife and daughter for their encouragement, support
and patience during the highs and lows associated with a PhD project.

Contents
List of figures.................................................................................................................... v
List of equations .............................................................................................................vii
List of abbreviations and units .....................................................................................viii
Abstract ............................................................................................................................ix
Chapter 1 Introduction..............................................................................................1
Chapter 2 Neural correlates of object recognition memory.....................................8
2.1 Object recognition memory: a brief overview..............................................9
2.1.1 Inferotemporal cortex and primate studies of object recognition .............9
2.1.2 The temporal lobes and object recognition memory in human and other
primates .................................................................................................................13
2.1.3 A brief summary of haemodynamic (PET and fMRI) neuroimaging
correlates of episodic memory retrieval..................................................................18
2.2 Human electrophysiological correlates of episodic memory retrieval .....21
2.2.1 EEG and ERP functional electrophysiological techniques: a brief
background..............................................................................................................22
2.2.2 EEG changes associated with episodic memory retrieval.......................25
2.2.3 Event-related potential changes associated with episodic memory
retrieval .................................................................................................................32
2.2.3.1 ERP correlates of episodic memory retrieval: evidence from
continuous recognition paradigms ......................................................................32
2.2.3.2 ERP correlates of episodic memory retrieval: evidence from study-test
paradigms ............................................................................................................41
2.2.3.3 Differentiation between transient item-related and sustained task-
related ERP correlates of episodic memory retrieval..........................................56
2.3 Summary of neural correlates of object recognition memory ..................58
Chapter 3 Steady-State Probe Topography ............................................................62
i

3.1 Steady-state evoked potentials compared with transient evoked potentials
.........................................................................................................................62
3.2 Steady-state evoked potentials in the study of cognitive processes...........64
3.3 Steady-State Probe Topography (SSPT).....................................................65
3.4 SSPT and recording artifacts.......................................................................70
3.5 Overview of investigations utilizing the SSPT technique ..........................70
3.5.1 Visual vigilance.......................................................................................72
3.5.2 Planning - Wisconsin Card Sorting Task................................................73
3.5.3 Attention - Continuous Performance Task..............................................74
3.5.4 Clinical application of SSPT - ADHD....................................................74
3.5.5 Spatial working memory .........................................................................75
3.6 Conclusions ....................................................................................................76
3.7 Formulation of hypotheses for the present investigation ..........................76
Chapter 4 Methods ..................................................................................................79
4.1 Introduction...................................................................................................79
4.2 Cognitive task design ....................................................................................80
4.3 Task presentation and stimulus parameters ..............................................83
4.4 Subjects ..........................................................................................................84
4.5 Probe stimulus ...............................................................................................85
4.6 Recording.......................................................................................................86
4.6.1 Electrodes and recording setup ...............................................................87
4.6.2 Recording instrumentation and phase locked data acquisition ...............88
4.7 Analysis of behavioural data........................................................................90
4.8 Offline signal processing...............................................................................90
4.8.1 Extraction of the 13Hz SSVEP from the EEG signal .............................90
4.8.2 Automatic detection of artifact in the EEG signal ..................................92
4.8.3 Calculations for modal and transient effects...........................................93
4.8.3.1 Long averaging period (sustained effects) ..........................................94
4.8.3.2 Short averaging period (transient changes).........................................94
ii

4.8.4 Group averaging......................................................................................96
4.8.4.1 Group averaging: long averaging period (sustained effects) ..............97
4.8.4.2 Group averaging: short averaging period (transient changes) ............98
4.8.5 Topographic mapping of SSVEP data ..................................................100
4.8.6 Statistical analysis and Significance Probability Mapping ...................101
Chapter 5 Results...................................................................................................103
5.1 Behavioural data .........................................................................................104
5.1.1 Individual subject ..................................................................................104
5.1.2 Group data.............................................................................................105
5.2 Electrophysiological data ...........................................................................107
5.2.1 Sustained effects: long averaging period ..............................................107
5.2.1.1 Individual subjects ............................................................................107
5.2.1.2 Group data.........................................................................................109
5.2.2 Transient changes: short averaging period............................................113
5.2.2.1 Changes with memory load...............................................................114
5.2.2.2 Targets versus non-targets.................................................................124
Chapter 6 Discussion.............................................................................................128
6.1 Behavioural results: the effect of increasing memory load on retrieval
accuracy and response time ...................................................................................128
6.2 Sustained SSVEP amplitude and latency changes and retrieval mode..130
6.2.1 The topography of sustained SSVEP changes ......................................131
6.2.2 Interpretation of sustained SSVEP amplitude and latency reductions..134
6.3 Transient SSVEP amplitude and latency changes: the effect of memory
load on successful memory retrieval .....................................................................137
6.3.1 Transient parietal effects .......................................................................139
6.3.2 Transient right frontal effects................................................................143
6.3.3 Transient effects and retrieval effort .....................................................147
6.3.4 Interpretation of transient SSVEP amplitude and latency reductions...148
6.4 Transient SSVEP amplitude and latency changes: target versus non-
target objects............................................................................................................151
6.5 Conclusions and future directions .............................................................154
iii

Appendix A. Task instructions ..............................................................................157
Appendix B. Determination of optimum luminance of light-emitting diode (LED)
arrays ...........................................................................................................160
Appendix C. Behavioural results for practice tasks .............................................162
Appendix D. The amplitude of the SSVEP produced by turning on the probe
stimulus ...........................................................................................................165
Appendix E. Sustained SSVEP effects for each memory load condition relative to
the baseline task: practice tasks...................................................................................170
Appendix F. Retrieval of everyday objects............................................................173
Publications by the author ...........................................................................................175
Bibliography .................................................................................................................189
iv

List of figures
Figure 3.1 The effect of various forms of artifact on an SSVEP amplitude time series
________________________________________________________ 71
Figure 4.1 Study-test experimental design _______________________________ 80
Figure 4.2 Experimental task design ___________________________________ 81
Figure 4.3 Calculation of modulation depth______________________________ 86
Figure 4.4 Sixty-four scalp recording locations ___________________________ 87
Figure 4.5 Experimental recording arrangement__________________________ 88
Figure 4.6 Single cycle and averaged Fourier coefficients __________________ 95
Figure 5.1 Mean response times for target objects for each memory load for an
individual subject ____________________________________________________ 105
Figure 5.2 Mean response time and mean number of errors for target objects versus
memory load for 40 subjects ____________________________________________ 106
Figure 5.3 Sustained effects: Mean SSVEP amplitude and phase topography for the
baseline task and each of the memory load conditions calculated with a long (40s)
averaging period for an individual subject_________________________________ 108
Figure 5.4 Sustained effects: Mean SSVEP amplitude and phase topography for the
baseline condition and each of the memory load conditions averaged across 40 subjects
_______________________________________________________ 110
Figure 5.5 Sustained effects: Topographic differences in SSVEP amplitude and
latency between the average of the 3 memory load conditions and the baseline task
averaged across 40 subjects ____________________________________________ 111
Figure 5.6 Sustained effects: Topographic differences in SSVEP amplitude and
latency between memory load conditions averaged across 40 subjects ___________ 112
Figure 5.7 Normalized SSVEP amplitude time series for each of the memory load
conditions calculated across 40 subjects __________________________________ 116
Figure 5.8 SSVEP phase time series for each of the memory load conditions
calculated across 40 subjects ___________________________________________ 117
Figure 5.9 Normalized SSVEP amplitude time series for each of the memory load
conditions calculated across 40 subjects __________________________________ 118
v

Figure 5.10 SSVEP phase time series for each of the memory load conditions
calculated across 40 subjects ___________________________________________ 119
Figure 5.11 Topographic differences in SSVEP amplitude and latency between
memory load conditions for target objects and Hotelling's T statistic (averaged across
40 subjects) _______________________________________________________ 122
Figure 5.12 Topographic differences in SSVEP amplitude and latency between
memory load conditions for non-target objects and Hotelling's T statistic (averaged
across 40 subjects) ___________________________________________________ 123
Figure 5.13 Topographic differences in SSVEP amplitude and latency between
correctly identified target and non-target objects, averaged separately for each memory
load condition: pooled across 40 subjects_________________________________ 125
Figure B.1 Determination of optimum luminance of LED arrays_____________ 161
Figure C.1 Practice tasks: Mean response times for target objects versus memory
load for an individual subject ___________________________________________ 163
Figure C.2 Practice tasks: Mean response time and errors for target objects versus
memory load for 40 subjects ____________________________________________ 164
Figure D.1 SSVEP amplitude and phase during the stimulus-off and stimulus-on
intervals for an individual subject________________________________________ 167
Figure D.2 Mean SSVEP amplitude during the stimulus-off and stimulus-on intervals
for an individual subject _______________________________________________ 167
Figure D.3 SSVEP amplitude and phase in response to switching-on probe stimulus
averaged across 40 subjects ____________________________________________ 168
Figure D.4 Mean SSVEP amplitude topography during the stimulus-off and
stimulus-on intervals averaged across 40 subjects___________________________ 169
Figure D.5 SSVEP amplitude difference topography and Hotelling’s T values for
stimulus-on relative to stimulus-off conditions averaged across 40 subjects_______ 169
Figure E.1 Mean SSVEP amplitude and phase topography for the baseline task and
each of the practice memory load tasks for an individual subject _______________ 171
Figure E.2 Mean SSVEP amplitude and phase topography for the baseline task and
each of the practice memory load tasks averaged across 40 subjects ____________ 172
Figure F.1 Retrieval of everyday objects _______________________________ 174
vi

List of equations
Equation 4.1 Calculation of unamplified EEG amplitude ___________________ 89
Equation 4.2 Calculation of single cycle Fourier coefficients ________________ 91
Equation 4.3 Calculation of SSVEP amplitude and phase ___________________ 91
vii

List of abbreviations and units
BA ……………………………………………………………….….… Brodmann’s area
Cd/m2 …………..………………………………………..……. candela per square metre
dB ………………………………………………………………………..………. decibel
DC ………………………………………………………………………... direct-coupled
ECG ……………………………………………………..…………… electrocardiogram
EEG ………….………………………………………...…..…….. electroencephalogram
EOG ………………………………………………………………… electro-occulogram
EMG …………………………………………………………………… electromyogram
ERP ……………………………………………………...………. event-related potential
fMRI ………………...…………………...……. functional magnetic resonance imaging
Hz ………….…………...……………………………………………………..……. hertz
IAF ………………………………………………………... individual’s alpha frequency
LPC …………………………………………………………….. late positive component
MEG ………………………………………………………….... magnetoencephalogram
MΩ ……...……………………………………….…………………………… mega-ohm
ms ………….………………...……………………………………………… millisecond
µV …………………………………………………………...………………… microvolt
ORM …………………………...…………………………… object recognition memory
kΩ ……...……………………………………….……………………………… kilo-ohm
PET ……………………………...…………………….... positron emission tomography
s …………………………………...………………………...……………………. second
SSPT ……………………………………………….……. steady-state probe topography
SSVEP …...…………………………...…………. steady-state visually evoked potential
VEP ……………………………………...……………………… visual evoked potential
viii

Abstract
Object recognition memory (ORM) refers to both recognition of an object and the
memory of having seen it before. In humans, ORM has been investigated using
functional neuroimaging and electrophysiological techniques with tests of episodic
memory retrieval involving recollection of previously studied items. Processes involved
in the maintenance of a mental state adopted for the performance of a retrieval task
(retrieval mode) appear to involve right frontal neural regions. More transient processes
occurring at the time of item recollection (retrieval success) have shown scalp activity
over parietal and right frontal regions. This activity is thought to originate in the medial
temporal lobes and the underlying right frontal cortex respectively. The aforementioned
findings have been derived mainly from studies using verbal stimuli. It is uncertain
whether the same neural regions are involved in object recollection. It is also uncertain
whether sustained modal and transient item-related activity involve the same or
different right frontal regions. In this study, steady-state probe topography (SSPT) was
used to investigate both sustained and transient processes involved in the retrieval of
abstract pictorial objects from memory. The ability to vary the evaluation period of the
steady-state visually evoked potential (SSVEP) allows investigation of cognitive
processes occurring over different time scales. Neural regions involved in sustained
modal processes were identified by examining the SSVEP values averaged over the
duration of a memory retrieval task. Sustained SSVEP effects were observed over right
fronto-temporal regions. Neural regions involved in transient retrieval success processes
were identified by comparing the transient SSVEP responses to tasks with different
memory loads. Comparison of a higher with a lower memory load condition showed
SSVEP effects over parieto-temporal and right inferior frontal regions. Larger
differences between memory loads gave effects that were larger and more right
lateralized. Retrieval mode and retrieval success processes showed SSVEP effects over
different right frontal regions. It was also found that, in contrast to the left lateralized
parietal ERP response to recollected verbal stimuli, the SSVEP effects produced with
abstract pictorial shapes showed a more bilateral pattern. This was considered to reflect
the relatively non-verbalizable pictorial nature of the stimuli.
ix

Chapter 1 Introduction
This study uses the technique of steady-state probe topography (SSPT) to investigate the
neural correlates of object recognition memory (ORM). Object recognition memory is
defined as the ability to recognize or remember previously encountered objects. Object
recognition memory is only one facet of the multifaceted function we term ‘memory,’
and is usually considered to involve three main stages: encoding, storage and retrieval.
For an event to be remembered it must first be encoded. This process can be thought of
as the formation of new memory traces in the brain. Storage involves maintenance of
these memory traces over time, and retrieval refers to the accessing of these memory
traces.
It appears that memories can be encoded into different stores, characterized by the
length of time that the memory traces are maintained. Some memories may be
maintained only fleetingly. For example, while watching a movie, each individual still
frame is remembered long enough for a succession of these to make sense (Baddeley
1999). This type of memory is sometimes referred to as immediate, or sensory, memory.
Memories which are maintained for slightly longer, for example, a telephone number
that is remembered only long enough to dial it, are stored in short term, or working,
memory. In contrast, other memories may last several minutes, days, or even a lifetime.
The store for these is referred to as long term memory.
Long term memory is generally considered to consist of two broad and largely
independent types of memory referred to as explicit and implicit memory (eg. Squire
and Zola-Morgan 1991; Tulving 1983). Explicit memory, also known as declarative
memory, encompasses those memories that involve conscious recollection, such as, the
experience of eating steak for dinner the previous night, or the fact that the Eiffel Tower
is in Paris. These two examples represent a further subdivision of explicit memory into
what is termed episodic and semantic memory (Tulving 1983). Episodic memory
consists of context-specific memories of experiences within one’s personal past,
whereas semantic memory consists of knowledge of facts that can be stated in words.
Conversely, implicit memory, also known as non-declarative memory, encompasses

Introduction
those memories that do not necessarily require conscious recollection. These include
skills, such as riding a bicycle, and habits. Proficiency is usually measured in terms of
accuracy or speed of response.
Object recognition memory (ORM) refers to both recognition of an object and the
memory of having seen it before. Object recognition memory can be part of sensory
memory, working memory, and long term memory processes. However, in human
studies, the longer term memory aspects of ORM have usually been studied, and tests of
episodic retrieval have frequently been used. In such tests, objects are studied and then
identified some time later. In a more general sense, the experiencing of an object can be
thought of as an event in one’s personal past. However, items may be recognized
because of their familiarity, rather than because the actual encounter is remembered (eg.
Jacoby 1991; Mandler 1981). It has been suggested that when recognition is based on
familiarity, implicit memory processes may be involved. It has also been suggested,
however, that memory retrieval based on familiarity may also form a part of a larger
explicit, or declarative, memory system (eg. Moscovitch 1992; Moscovitch 1994;
Squire 1994). While the distinction between familiarity and the recollection of an
experience could complicate the study of ORM, tests have been designed to influence
the extent to which familiarity or recollection processes are used. With such tests it has
been possible to distinguish between processes involved in familiarity-based and
recollection-based recognition.
The neural regions involved in ORM have been extensively investigated in non-human
primates, mainly by examining how ablating various parts of the brain affects ORM
function. Such studies have shown that a series of cortical regions beginning at the
primary visual cortex and ending within the inferior temporal lobe, the so-called ventral
pathway, are important in object perception and recognition processes. Awareness that
something has been seen previously has been shown to involve interaction of these
ventral pathway regions with adjacent medial temporal lobe regions, including the
hippocampus and perirhinal cortex.
Lesion studies in humans have indicated that neural regions equivalent to those in non-
human primates are involved in human ORM. However, because lesions in human
subjects generally result from accident, disease or surgery, they are generally larger and
2

Introduction
less precisely localized than those that can be produced experimentally in animals.
Thus, confidence in conclusions drawn about functions associated with specific brain
regions must be limited. Furthermore, it is generally not clear whether memory
impairment stems from deficits in encoding or retrieval. Thus, differentiation of the
neural correlates of encoding and retrieval on the basis of lesion data is difficult.
Despite the problems inherent in such studies, findings have indicated that the same
structures that are important in non-human primate recognition memory, namely, the
medial temporal lobe, the hippocampus and perirhinal cortex, are also important in
human recognition memory.
During the last two decades, functional neuroimaging and non-invasive
electrophysiological techniques have enabled memory processes in the undamaged
brains of normally functioning humans to be studied. These techniques have permitted
the monitoring of brain activity at a large number of neural sites simultaneously, whilst
subjects perform tasks designed to activate regions involved in ORM.
Functional neuroimaging detects changes in cerebral blood flow and can localize brain
activations to within a few millimetres, but it has relatively poor temporal resolution.
These techniques, used in the study of ORM, have had temporal resolutions ranging
from a few seconds to a few minutes. They have been used mainly to investigate
sustained, or task-related, cognitive states that are initiated by task instructions and that
persist throughout the task.
On the other hand, electrophysiological techniques can monitor neural activity with a
temporal resolution in the order of milliseconds. However, spatial resolution is limited
to gross brain regions. Electrophysiological techniques have mainly been used to
investigate more transient, or item-related, processes. These processes are initiated by
the presentation of each stimulus item and may continue throughout the duration of
presentation.
In both functional neuroimaging and electrophysiological studies, recognition memory
processes have been investigated using tests of episodic memory retrieval. Although,
these studies have mainly used word rather than object stimuli, findings have
nevertheless aided the understanding of memory retrieval processes.
3

Introduction
Functional neuroimaging studies have identified seven main regions considered to be
involved in episodic retrieval, namely, prefrontal, medial temporal, medial parieto-
occipital, lateral parietal, anterior cingulate, occipital, and cerebellar regions (Cabeza
and Nyberg 2000). Findings of medial temporal lobe activity support findings from
human lesions studies. It has been shown that medial temporal lobe activity increases
with increasing recognition accuracy. However, activity in the right prefrontal region
appears to be the most frequently reported finding in episodic retrieval studies. While
activity in this region appears to be present under a number of experimental conditions,
the most consistently reported finding is that activity in this region is sustained for the
duration of the retrieval task, that is, the activity is task-related. This sustained right
prefrontal activity may be necessary for the maintenance of the mental set, or state that
accompanies conscious recollection. This has been termed retrieval mode by Tulving
(1983).
A major focus of electrophysiological research into episodic retrieval has been the
difference between the brain’s electrical response to correctly recognized, previously
presented, or old, words compared with the response to the presentation of words not
previously presented, or new, words. This is known as the ERP old/new effect. This
effect has been used to investigate processes associated with retrieval success. Most
studies that used verbal materials as stimuli found the largest ERP old/new effect over
left parietal regions, beginning approximately 400ms after the appearance of the
stimulus and lasting throughout its duration. Transient item-related right frontal effects
are found in situations when the context of item presentation has to be recalled, that is,
when recognition is based on recollection rather than purely on familiarity, and are
thought to be associated with both recollection and monitoring processes.
Despite the wealth of information that has been attained through use of these functional
neuroimaging and electrophysiological techniques, a number of important unresolved
issues remain. Firstly, it is not clear whether the right frontal activity found in functional
neuroimaging and electrophysiological studies is sustained task-related activity, or more
transient item-related activity. What appears to be task-related activity may, in fact, be
the result of the summed intermittent activity occurring in response to individual items.
Alternatively, both sustained and transient activity may be present. Furthermore,
4

Introduction
because of the limitations of the respective techniques, it is not be possible to determine
whether the same or different adjacent regions are involved if both types of activity are
present. Secondly, it is not clear whether non-verbal stimuli produce patterns of activity
essentially the same as those observed with verbal stimuli. This is an important issue in
the present study as the focus is on the retrieval of objects, rather than words, from
memory. Thirdly, activity associated with retrieval success is generally investigated by
comparing the activity associated with correctly retrieved old items with the activity
associated with correctly identified new items. Because a comparison is made with a
secondary task that entails the identification of new items, which may itself produce its
own characteristic neural activity patterns, this may confound interpretation. Duzel et al.
(1999) have explored the matter of whether activity associated with a word-based
episodic retrieval is task and/or item-related by using a combination of functional
neuroimaging and electrophysiological techniques. They found that the right prefrontal
region, BA10, is active throughout the course of a retrieval task, whereas the left medial
temporal lobe becomes active intermittently whenever individual words are presented
and is more active for familiar than for novel words. However, this study does not
address the fact that in many electrophysiological studies, transient item-related, right
prefrontal effects have also been reported. Furthermore, it is not clear whether non-
verbal stimuli produce the same task and item-related effects that verbal stimuli
produce. The present study attempts to address the three aforementioned issues using
the technique of steady-state probe topography (SSPT).
The SSPT technique was developed by Silberstein and colleagues (Silberstein et al.
1995a; Silberstein et al. 1990b). With this technique, the effects of mental activity on
the steady-state evoked potential (SSVEP) generated by a rapidly repeating irrelevant,
or probe, stimulus are examined at multiple scalp recording sites. A major advantage of
SSPT over traditional electrophysiological techniques is that it permits temporal
continuity as well as a range of time scales over which processes can be studied. This
makes it a valuable tool for the investigation of both sustained task and transient item-
related effects. In this study, the probe stimulus was a 13Hz visual flicker. The SSVEP
produced in response to the visual flicker was recorded via 64 scalp electrodes. The
spatial resolution of SSPT using 64 electrodes, although not as good as that obtained
using functional neuroimaging methods, appears satisfactory for investigating neural
activity within gross brain regions. Variations in the SSVEP amplitude and phase have
5

Introduction
been shown to reflect a range of cognitive processes, including working memory
processes (Silberstein et al. 2001).
The main aim of the present study was to use SSPT to investigate both sustained
retrieval mode activity and transient activity occurring during the actual recollection
process. In this study, an episodic retrieval task was used in which previously studied,
abstract, two-dimensional objects had to be identified when presented within a sequence
containing a larger number of unstudied distractor objects. The memory load was varied
by changing the number of objects that had to be remembered. To investigate sustained
task-related processes, the average of the SSVEP values across all memory load
conditions was compared with the average of the SSVEP values obtained in a separate
non-episodic control task. To investigate more transient item-related processes, SSVEP
amplitude and phase values obtained for different memory load conditions were
compared at a point in time when subjects were considered to be engaged in the
memory retrieval process. It was anticipated that increases in memory load would result
in increased utilization of those neural regions necessary for retrieval success. This
approach has the advantage over the traditional approach of comparing old and new
items because it avoids any spurious effects that might result from the ‘new item’
comparison task. The use of abstract objects enabled exploration of the question of
whether patterns of activity associated with the retrieval of object stimuli are the same
as those associated with verbal stimuli.
Sustained, task-related, SSVEP effects were most prominent over right frontal regions.
This is consistent with functional neuroimaging and electrophysiological studies that
have associated activations in this region with the maintenance of a retrieval mode. The
right frontal effects in the present study were characterized mainly by reductions in
SSVEP latency. Memory-load-dependent transient SSVEP amplitude and latency
reductions were found over parieto-temporal regions bilaterally and over right inferior
frontal regions. In addition, increases in memory load led to larger and more right-
lateralized SSVEP effects. These transient item-related effects and the ERP old/new
effects are apparent during essentially the same time interval post stimulus onset.
However, the parietal ERP old/new effect is usually reported as being left-lateralized,
whereas a right-lateralized effect was observed in the present study. This right
lateralization, however, is consistent with the abstract pictorial nature of the stimuli
6

Introduction
used. The spatio-temporal patterns of the item-related right frontal effects found in the
present study are similar to those of the right frontal ERP old/new effects associated
with remembering past events. While both sustained task and transient item-related
effects occurred over the right frontal region, they appeared to be localized to different
parts of the region. Furthermore, sustained task-related effects were characterized
mainly by SSVEP latency reductions, whereas transient item-related effects were
characterized by both amplitude and latency reductions that increased with increasing
memory load.
This thesis is in six chapters. Chapter 1 comprises the introduction. Chapter 2 reviews
the literature on the neural correlates of object recognition memory. The chapter begins
with a brief overview of the way in which primate lesion studies have indicated the
neural structures and pathways involved in ORM. This is followed by an overview of
PET and fMRI functional neuroimaging studies of episodic memory retrieval. These
overview sections are then followed by a review of studies investigating
electrophysiological correlates of episodic memory retrieval. Chapter 3 comprises a
description of the SSPT methodology and a formulation of the hypotheses of the present
study. Chapter 4 describes in detail the experimental methods that were employed in
this study. Task design, recording and analysis are explained in this chapter. Chapter 5
deals with results of the present study. Task and item-related findings are then discussed
in Chapter 6.
7

Chapter 2 Neural correlates of object recognition
memory
This chapter outlines findings from studies concerned with the representation of object
recognition memory (ORM) in the human brain. The chapter consists of two sections.
The first is an introduction to the neural regions involved in ORM (section 2.1), and the
second is a comprehensive review of human electrophysiological studies that have
attempted to define and localize neural substrates of ORM (section 2.2).
The introduction to ORM (section 2.1) is not intended as an extensive review of the
literature. Rather, it is included as background for the reader not familiar with this area.
This section begins with a brief description of the neural areas involved in visual
perception and recognition, with a particular focus on the so-called ventral pathway that
connects visual areas with the inferotemporal cortex (section 2.1.1). The functions of
regions involved in perception and recognition have been determined largely from
electrophysiological recordings and lesion studies in non-human primates. A brief
discussion then follows outlining the more specific role of the medial temporal lobes in
ORM (section 2.1.2). This is discussed within the context of the declarative model of
memory. This model has been derived predominantly from findings of lesion studies in
non-human primates, and of neuropsychological lesion studies with humans. Finally,
there is a brief overview of functional neuroimaging studies, focusing on those studies
that have utilized PET and fMRI techniques with tests of episodic memory retrieval to
investigate neural regions involved with ORM (section 2.1.3). Studies using these
imaging techniques have largely examined modal task-related activity, that is, activity
that is maintained for the duration of the imaging period.
Section 2.2 contains the primary focus of this review: human electrophysiological
correlates of object recognition memory. In human electrophysiological studies, visual
recognition memory has been examined using tests of memory retrieval in which
previously presented words or pictures are later identified. In addition to structures of
the ventral pathway and the medial temporal lobe, recognition of items during a

Neural correlates of object recognition memory
retrieval task also involves other regions, such as the temporal, parietal, and frontal
regions. Electrophysiological techniques that achieve a high temporal resolution have
been used to examine the fast neural processes that occur during memory retrieval. A
major review of electrophysiological correlates of episodic memory retrieval is
presented in sections 2.2.2 and 2.2.3. These sections are concerned with EEG and ERP
correlates respectively.
2.1 Object recognition memory: a brief overview
The following overview of ORM and neural regions involved in ORM processes
focuses on the visual pathways, and in particular, on the role of the inferior temporal
cortex in visual perception (section 2.2.1). This section is followed by a discussion of
ORM within the context of a model of declarative (explicit) memory. The focus here is
on the role of the medial temporal lobes in ORM (section 2.1.2). More precise
localization of neural regions involved in the retrieval of items from memory has been
investigated using PET and fMRI neuroimaging. A short summary of the main findings
is provided in section 2.1.3.
2.1.1 Inferotemporal cortex and primate studies of object recognition
A number of stages of visual processing precede object recognition. As visual
recognition memory depends on visual perception, it is important to consider the brain
regions involved in the early stages of visual processing. Visual information from the
retina reaches the cortex via two main pathways: one includes the lateral geniculate
nucleus, which projects almost exclusively to the primary visual cortex (also known as
V1, area 17 or striate cortex), and the other includes the superior colliculus and
pulvinar, which projects much more extensively (eg. Kandel et al. 2000; Reid 1999).
Cortical regions that receive projections from the latter pathway are not exclusive to
vision and are also associated with functions such as somatosensory, auditory and motor
processing. The areas receiving the densest input from the lateral geniculate nucleus and
pulvinar appear to be area 17, and the extrastriate cortex (also known as visual
association or areas 18 and 19). Area 17 is mainly confined to the calcarine fissure and
9

Neural correlates of object recognition memory
includes parts of the cuneus and lingual gyrus. Areas 18 and 19 are organized
concentrically around area 17 and also receive direct inputs from area 17 (Kolb and
Whishaw 1996 among others).
Parietal and temporal regions also receive a rich supply of connections from area 17 and
appear to perform a number of important visual functions. In fact, the visual cortex
projects to a large proportion of the total cortical area. Felleman and van Essen (1991)
report that in the primate brain, 55% of the whole cortical surface is involved in vision,
whereas only 11% is involved in somatosensory processing, and 3% in auditory
processing.
Findings from a series of electrophysiological studies on non-human primates indicate
that sensory information from the primary visual cortex reaches the parietal and
temporal lobes via a number of cortico-cortical stages (Ungerleider and Mishkin 1982).
Two relatively distinct pathways, or streams, were noted. One pathway passes dorsally
into the extrastriate cortex and terminates in the posterior parietal lobule, while the other
passes ventrally through the extrastriate cortex and terminates within the inferotemporal
cortex. The authors proposed that the dorsal pathway is concerned with ‘where’ visual
information is located, and the ventral pathway is concerned with ‘what’ the visual
information is. The inferior temporal cortex represents the final cortical stage in the
‘what’ pathway. Evidence for the existence of distinct ventral and dorsal visual
pathways, the so-called ‘what ‘ and where’ pathways in humans has been found using
functional neuroimaging techniques (Courtney et al. 1996; Haxby et al. 1991; Kohler et
al. 1995; Ungerleider and Haxby 1994).
Interconnections between primary visual areas and regions associated with these two
visual pathways, or streams, have been extensively studied in the non-human primate
brain. Felleman and Van Essen (1991) reported that 32 visual and visual association
areas can be differentiated in the non-human primate brain. Almost half of these areas
have now been mapped in the human brain (Sereno et al. 1995; Tootell et al. 1996;
Tootell et al. 1997). In many respects, the organisation of visual areas in the human and
non-human primate cortex appears to be similar (Tootell et al. 1996). Furthermore, as
indicated earlier, PET imaging studies have revealed that the dorsal and ventral
processing streams are also similar. Although there are differences between the visual
10

Neural correlates of object recognition memory
systems of humans and non-human primates, the non-human primate brain provides a
good model for the investigation of the human visual system (Tootell et al. 1997).
It is the ventral pathway, the so-called ‘what’ pathway, that is of particular relevance to
this study. A brief description of its interconnections will follow, with a focus on the
flow of visual information through this pathway to the inferior temporal cortex.
In non-human primates, the ventral pathway includes a number of cortical regions that
appear to be hierarchically organized, beginning with the primary visual cortex (V1).
This area projects to all other visual areas. The second level in the hierarchy is V2
(secondary visual area in the cerebral cortex), and this also projects to all other visual
areas. There are three main projections from V2: to the parietal cortex in the dorsal
pathway, and to the superior temporal sulcus and inferior temporal cortex in the ventral
pathway (Kandel et al. 2000).
A number of studies suggest that the last exclusively visual stage of the ventral pathway
is located in the inferior temporal cortex (see Logothetis and Sheinberg 1996 for
review). The inferior temporal cortex extends ventrally from just anterior to the inferior
occipital sulcus to within a few millimetres posterior to the temporal pole, and from the
fundus of the occipito-temporal sulcus to the fundus of the superior temporal sulcus.
This region includes Brodmann’s areas 20 and 21, or area TE, named by Von Bonin and
Bailey (1947). Area TE was later subdivided into two cytoarchitectonically distinct
cortical regions, TEO posteriorly and TE anteriorly (Iwai and Mishkin 1969; Von Bonin
and Bailey 1950). Cortical regions, roughly corresponding to TEO and TE have also
been shown to be functionally specialized. Lesions within TEO result in recognition
deficits for simple patterns, while lesions within TE lead to associative and visual
memory deficits (Iwai 1978; Iwai 1981; Iwai 1985).
Areas TEO and TE of the visual ventral stream project to many cortical and sub-cortical
regions. Of particular interest are the interconnections between the inferotemporal
cortex and parts of the medial temporal cortex, in particular, the hippocampus, the
amygdala, and the entorhinal and perirhinal cortices. These structures, as will be
discussed later, are all implicated in various aspects of memory. Area TEO receives
feedforward cortical inputs from the secondary visual areas V2, V3, and V4, including
11

Neural correlates of object recognition memory
contralateral connections from these areas via the corpus callosum. These secondary
areas also receive feedback projections from area TEO. Area TEO projects in a
feedforward fashion to several other cortical areas, designated TEm, TEa, and IPa, all of
which also send feedback projections back to TEO (eg. Rolls 2000; Van Essen 2002).
Areas designated TH and TG, and Brodmann’s area 36 also provide feedback
projections to TEO. Area TE projects to regions designated TH, TF, STP, FEF and area
46 (dorsolateral prefrontal cortex). TE also projects directly to the amygdala and to the
hippocampus. In addition, the hippocampus receives an indirect projection from TE via
the parahippocampal gyrus (eg. Rolls 2000; Van Essen 2002). There are also indirect
projections from TE to the entorhinal cortex via the perirhinal and parahippocampal
cortices. Area TE is also interconnected, both directly and indirectly, with limbic
structures (Desimone and Duncan 1995; Kolb and Whishaw 1996; Logothetis and
Sheinberg 1996 among others). Visual association areas of TE project to two prefrontal
regions, one on the dorsolateral surface and one in the orbital region (Kolb and
Whishaw 1996 p. 289). Areas TEO and TE also make connections with a large number
of subcortical areas (see Webster et al. 1993).
A large proportion of the inferotemporal cortex responds selectively to shapes. In fact,
more than 85% of neurons in the inferotemporal cortex appear to respond to simple or
complex visual patterns (Desimone et al. 1984). Tanaka (1993) reported that the
inferotemporal cortex consists of ‘elaborate’ cells that respond only to composite
shapes. Unlike the primary visual cortex, the inferotemporal cortex does not appear to
be organised retinotopically. Instead, inferotemporal neurons are systematically
organised such that neurons with similar response properties are assembled into
modules extending through the thickness of the cortex (Tanaka 1993). These modules
appear to be tuned to respond to similar combinations of shapes and other stimulus
characteristics. These findings, among others, suggest that the general class of an object
is represented by the combined activity of different modules in the inferotemporal
cortex, whereas fine discriminations are represented by differences in the activity of
neurons within a single module. (For a more detailed description see Fujita et al. 1992;
Gawne and Richmond 1993; Tanaka 1993; Young 1993)
The inferotemporal cortex, where the final stage of the ‘what’ visual pathway is located,
lies adjacent to the medial temporal cortex, which has been shown to be important in
12

Neural correlates of object recognition memory
memory. In other words, the inferior temporal cortex plays a crucial role in identifying
‘what’ an object is, whereas the medial temporal cortex plays a crucial role in
‘remembering’ whether it has been seen before. The brain regions believed to be
involved in visual recognition memory, particularly the medial temporal lobe, have been
determined by examining the consequences of lesions in humans and of focally
produced lesions in non-human primates. This will be discussed in the next section.
2.1.2 The temporal lobes and object recognition memory in human and other
primates
Although the study of the neural basis of memory began in the mid-nineteenth century
with the first descriptions of memory disorders (Ebbinghaus 1964), it wasn’t until the
1980s that memory was considered to consist of distinct components that depend on
different brain systems (Schacter 1987; Schacter 1992; Schacter and Crovitz 1977). A
distinction has been made between a capacity for the conscious recollection of facts and
events (declarative memory) and non-conscious performance of previously learned
behaviours (non-declarative memory) (Squire and Zola 1996 among others). This came
about because amnesic patients with bilateral medial temporal lobe damage presented
with deficits in recall or recognition, but performed normally on tasks requiring a
capacity for skill and habit learning and for priming (Squire and Zola 1996). These
neuropsychological findings indicated that the memory processes that were intact in this
form of amnesia utilized brain regions other than those that were damaged. Visual
recognition memory, which is considered to be one aspect of declarative memory, has
been shown to be compromised with damage to the medial temporal lobes.
Perhaps the most famous account of declarative memory loss is that of Scoville and
Milner’s patient HM (Scoville and Milner 1957). This patient had both medial temporal
lobes surgically removed to alleviate his chronic epilepsy. Following this procedure,
and to this day, this patient has exhibited anterograde amnesia. That is, memories prior
to the temporal lobe resection remain intact, whereas no subsequent long-term
memories have been established. In contrast, patient HM still retains non-declarative
memory function.
13

Neural correlates of object recognition memory
Petri and Mishkin (1994) described neural systems for explicit and implicit memory
based on human and animal studies. For explicit memory, the regions involved include,
not only the temporal lobes, but also the prefrontal cortex, thalamus, basal forebrain and
neocortex. In general, experiments in monkeys and rats have indicated that the rhinal
cortex is involved in object memory, the hippocampus in spatial memory, and the
amygdala in emotional memory. The differing effects of brain damage on HM and other
amnesic patients are considered to result from damage to different combinations of
connections in the Petri and Mishkin (1994) model.
In non-human primates, bilateral lesions to specific parts of the hippocampus (CA1 and
CA2) resulted in impairments in a delayed non-matching to sample task with a delay of
approximately 10 minutes (Zola-Morgan et al. 1992). In this task, a presented object
must be selected if it fails to match an object that had been presented previously.
Furthermore, the more extensive the lesion to the hippocampus and adjacent regions, the
greater the impairment.
Analysis of a number of other monkey studies also suggested that the larger the lesion
within the medial temporal lobe, the greater the memory impairment (Squire and Zola
1996). While damage confined to the hippocampus resulted in significant memory
impairment, lesions that included the adjacent parahippocampal and entorhinal cortices
in addition to the hippocampus resulted in greater impairment, and when the damaged
region extended still further into the perirhinal cortex, the severity of memory
impairment was even greater.
The extent of damage is not the only factor that determines the degree of memory
impairment. Various studies have indicated that certain parts of the medial temporal
lobe are more important in memory than others. Firstly, it appears that the entorhinal
cortex, considered to be part of the hippocampal formation, is not essential for memory
(Squire and Zola 1996 among others). Most of the sensory input to the hippocampus
passes through the entorhinal cortex, while the entorhinal cortex receives a large
proportion of its input from the perirhinal and the parahippocampal cortices (see Squire
and Zola 1996). When only the entorhinal cortex is damaged, only mild memory
impairment results (Meunier et al. 1993). Furthermore, when monkeys with entorhinal
lesions were re-administered delayed non-matching to sample memory tests 6-13
14

Neural correlates of object recognition memory
months later, they were found to perform normally at all delay intervals (Leonard et al.
1995). Also, localized lesions to the amygdala did not appear to affect performance in
the delayed non-matching to sample task (Zola-Morgan et al. 1989).
On the other hand, it appears that the perirhinal and parahippocampal cortices,
structures adjacent to the hippocampus and entorhinal cortex, are particularly important
in memory. Damage to these regions results in a lasting behavioural impairment in the
delayed non-matching to sample task. The degree of impairment came close to that
occurring with a larger medial temporal lobe lesion that includes the perirhinal and
parahippocampal cortices as well as the hippocampus and the amygdala (Squire and
Zola 1996).
In fact, there are many pathways to the medial temporal lobe from other parts of the
neocortex in which small specific lesions can affect performance on specific memory
tasks (Squire and Zola 1996). Of particular relevance to this study is the fact that the
perirhinal cortex receives greater visual input than does the parahippocampus, a finding
determined using retrograde tracer techniques (Suzuki and Amaral 1994; Webster et al.
1991). It was found that lesions within the perirhinal cortex affected visual memory
more so than lesions at any other single site within the medial temporal lobe (Horel et
al. 1987 among others; Meunier et al. 1993). More recently, Parker and Gaffan (1998)
demonstrated with monkeys that the frontal lobe must interact with the perirhinal cortex
in the same hemisphere for ORM to occur. There are many pathways, both direct and
indirect from the perirhinal cortex to the frontal lobes. When specific points in a number
of these pathways were ablated, ORM was seriously impaired.
Area TE, in the inferior temporal lobe, is a unimodal visual area situated adjacent and
lateral to the perirhinal cortex (Von Bonin and Bailey 1947), and was mentioned earlier
as being important in ORM as well as in object perception. In monkeys, lesions within
this area resulted in impaired visual perception as measured, for example, by pattern
discrimination, and in impaired performance on the delayed non-matching to sample
task. With this latter task, neurons in the inferotemporal cortex continued to discharge
during the ‘memory’ period for objects that had to be remembered (Fuster and Jervey
1982 p.298). It was subsequently suggested, however, that in virtually all previous
studies that described impairment resulting from lesions within TE there was also
15

Neural correlates of object recognition memory
damage to the perirhinal cortex (see Squire and Zola 1996). Squire (1996) showed that
the function of TE and the perirhinal cortex can be dissociated. When monkeys were
tested on various tasks, prior to and following lesioning, it was found that the perirhinal
region is particularly important in declarative memory, whereas TE in the inferior
temporal lobe is more important in visual perception.
The perirhinal cortex is located at the interface of TE and the medial temporal lobe.
While precisely located bilateral lesions can be made in monkeys, damage to medial
temporal lobes in humans is usually confined to one hemisphere and varies in size and
location. Nevertheless, despite a lack of consistent human neuropsychological data,
there have been some important observations confirming that regions already described
as being important in ORM in non-human subjects are also important for humans. As
mentioned earlier, patient HM had undergone bilateral medial temporal lobe resection,
which included removal of the entire temporal lobes. Scoville and Milner’s (1957)
initial interpretation was that damage to the hippocampus was responsible for his
anterograde amnesia. However, subsequent experiments on monkeys and rats have
indicated that the hippocampal damage would account for only the spatial impairments
displayed by HM. Other medial temporal lobe regions such as the amygdala, which is
responsible for emotional memory, and the perirhinal cortex, responsible for object
recognition memory, also appear to be responsible for his impairment (Kolb and
Whishaw 1996).
An analysis of two studies that described neuropsychological changes in patients with
damage to the medial temporal lobe has indicated that the more extensive the damage,
the greater the memory impairment (Squire and Zola 1996). All the patients were less
severely impaired in tests of declarative memory than HM, and the damage to
hippocampal and adjacent cortical regions was also less extensive than in HM. All
presented with moderately severe anterograde amnesia and two with extensive
retrograde amnesia also. All had damage to the CA1 area of the hippocampus. Those
whose damaged areas included the entorhinal and perirhinal cortices showed the most
severe memory impairment.
Buffalo et al. (1998) examined recognition memory for complex visual stimuli in two
patients with extensive damage to the perirhinal cortex, and in six other amnesic
16

Neural correlates of object recognition memory
patients with damage confined to the hippocampus and other diencephalic structures.
Immediate and long-term memory were tested separately, using delays from 0 to 40s in
a delayed recognition memory task. They found that both patient groups exhibited intact
recognition memory at delays of 0 to 2s, and both groups also displayed delay-
dependent memory impairment for delays greater than 6s. Furthermore, with delays
greater than 25s, the performance of the two patients with damage to the perirhinal
cortex was worse than the performance of the other amnesic patients. It was thus
concluded that the perirhinal cortex is not important for visual perception or immediate
memory. However, like the other medial temporal lobe structures, the perirhinal cortex
appears to be involved in longer-term memory processes.
It is now thought that the two temporal lobes have their own specialized functions.
Milner (Milner 1958; Milner 1968; Milner 1970) found that in patients who had
undergone temporal lobe removal, the specific types of memory deficit depended on the
side of the lesion. Patients with damage to the left temporal lobe showed deficits in
verbal memory in, for example, tests of recall of previously presented stories and pairs
of words and on recognition of words or numbers (see Kolb and Whishaw 1996).
Patients with right temporal lobe lesions showed deficits in non-verbal memory. These
patients found it difficult to recall complex geometric figures, recognize nonsense
figures, recognize tunes, and to recognize previously viewed photographs of faces (see
Kolb and Whishaw 1996 p. 369). Kolb and Whishaw (1996 p.291) cautioned that
although it appears that the two temporal lobes appear to have specialized functions,
there is also a high degree of functional overlap. While an association of the left
temporal lobe with verbal memory appears to be reasonably well accepted, the precise
role of the right temporal lobe in memory is less clear.
Neuropsychological investigations with humans and other primates have thus indicated
that the inferotemporal cortex and the medial temporal lobes, and in particular, the
perirhinal cortex, are important in visual recognition memory. Furthermore, these areas
project to other cortical and sub-cortical regions such as the frontal lobes, which need to
be intact for preserved memory functioning.
17

Neural correlates of object recognition memory
In recent years, a number of functional neuroimaging techniques have been used with
humans in attempts to localize more precisely the neural substrates involved in ORM.
Studies using these techniques will be summarized in the following section.
2.1.3 A brief summary of haemodynamic (PET and fMRI) neuroimaging
correlates of episodic memory retrieval
Neural regions involved in memory retrieval processes have been identified in humans
using functional neuroimaging techniques such as positron emission tomography (PET)
and functional magnetic resonance imaging (fMRI). These techniques have identified
neural regions involved in memory retrieval processes through the monitoring of
cerebral blood flow changes while subjects are engaged in tests of memory retrieval.
While these techniques possess excellent spatial resolution, temporal resolution is
relatively poor. Notwithstanding this limitation, important findings have included the
identification of neural regions that are considered to be involved in the maintenance of
retrieval mode, that is, those regions that remain active for the duration of a retrieval
task, and that are considered to facilitate successful memory retrieval.
The premise underlying PET and fMRI neuroimaging techniques is that regional
increases in blood flow or oxygenation reflect local neural activity. In the case of PET,
blood-flow changes are assessed by monitoring the uptake of a short-half-life
radiotracer injected into the bloodstream. Functional magnetic resonance imaging
techniques assess blood flow changes through changes in blood oxygenation. Shortly
after a neural region is activated, the relative concentration of oxy-haemoglobin
increases and deoxyhaemoglobin decreases, producing an overall increase in the fMRI
signal. For both of these techniques, image reconstruction methods are employed to
localize these changes to within a few millimetres (eg. Schmitt et al. 1998).
While the spatial resolution of these techniques is excellent, the temporal resolution is
relatively poor. Typically, in the case of PET, an image can only be formed reliably
every 20 to 30s. For this reason, PET studies have only been used to assess task-related
changes. Episodic memory retrieval studies have used blocked task designs in which
two separate task blocks are presented. One task block constitutes the target task and the
18

Neural correlates of object recognition memory
other the reference task. Neural regions showing increased activity during the target task
relative to the reference task are normally termed activations. These activations are
considered to reflect cognitive processes that are utilized to a greater extent by the target
task than by the reference task. Functional MRI techniques can, in theory, achieve a
temporal resolution better than 1s. However, a haemodynamic lag (time for local uptake
of blood) of around 6s cannot be measured accurately and varies depending on the
neural region being examined. Consequently most fMRI-based episodic retrieval studies
to date have used block designs similar to those used in PET studies. While the recently
developed event-related fMRI technique can achieve a temporal resolution better than
1s (eg. see Menon and Kim 1999), the present author is not aware of any episodic
memory retrieval studies in which they have been used. It must however be noted that
while activations observed using a blocked design may well be indicative of modal
activity, we have to be aware that summed responses to individual items may also
contribute to what appears to be sustained activity.
Functional neuroimaging studies have mainly used tests of episodic memory retrieval to
identify neural regions involved in ORM. Episodic memory retrieval refers to the recall
of events in one’s personal past (Tulving 1993) and is generally considered to be one
aspect of declarative memory. Regions involved in episodic memory retrieval include
the medial temporal lobe and frontal regions.
Functional neuroimaging studies have, in the majority of cases, examined episodic
memory retrieval using verbal stimuli, and in only a few instances have they used
objects. A very brief summary of PET and fMRI studies into episodic memory retrieval,
with both verbal and non-verbal stimuli, will follow. A more extensive discussion of
this body of literature can be found in a selection of review articles (Buckner and
Tulving 1995; Cabeza and Nyberg 2000; Nyberg et al. 1998b; Rugg and Wilding 2000).
In a recent publication by Cabeza and Nyberg (2000) the authors reviewed 275 PET and
fMRI studies, of which, a large number were concerned with episodic memory retrieval.
This paper summarizes findings for all aspects of episodic memory retrieval that have
been investigated using PET and fMRI techniques to the year 2000 and clarifies the
consensus of opinion on a number of issues. The section in this paper on episodic
memory retrieval is the main source of information for the following discussion of PET
19

Neural correlates of object recognition memory
and fMRI findings (remainder of section 2.1.3). The discussion focuses mainly on
neural regions involved in retrieval success, retrieval effort, and retrieval mode.
Activations have been noted consistently in seven main regions in PET and fMRI
studies dealing with episodic retrieval: prefrontal, medial-temporal, medial parieto-
occipital, lateral parietal, anterior cingulate, occipital, and cerebellar regions.
Although bilateral frontal activations are sometimes observed, right prefrontal
activations have been a consistent finding in PET and fMRI studies. Moreover, the
majority of these studies have indicated a role for these regions in establishing and
maintaining the mental set for episodic retrieval, or retrieval mode. Right prefrontal
(BA10) activation reflecting retrieval mode is usually observed by contrasting an
episodic retrieval task with a non-episodic retrieval task. Other studies have associated
bilateral activation of prefrontal regions (BA 10, 9, 46) with processes reflecting
retrieval success, and left prefrontal regions (BA 47, 10) with retrieval effort.
Although medial temporal activations are lateralized during episodic encoding, they
occur bilaterally during episodic retrieval. Importantly, these medial temporal lobe
activations occur bilaterally regardless of whether stimulus items are verbal or non-
verbal. Medial temporal lobe activations have been associated with retrieval success,
but not with retrieval mode or retrieval effort. It was therefore suggested that activation
of the medial temporal lobes reflects successful memory retrieval, given also the
findings of one study in which a strong relationship between medial temporal lobe
activation and recognition accuracy was reported. Activation of the medial temporal
lobes has also been associated with reactivation of stored memory representations and
with conscious recollection.
Episodic memory retrieval tasks have also produced activation of medial parieto-
occipital regions. Activation of the precuneus, located within this region, has been
associated with imagery operations and retrieval success. Lateral parietal activations
have been associated with the processing of spatial information during episodic
retrieval, and with perceptual aspects of recognition. Activation of the occipital cortex
has been associated with non-verbal retrieval involving memory-related imagery
20

Neural correlates of object recognition memory
operations, and activation of the cerebellum has been associated with self-initiated
retrieval operations.
A comparison of the retrieval of object location relative to object identity yielded
activation of inferior parietal regions. The opposite comparison (object identity minus
object location) yielded activation of fusiform regions. This result suggests that the
distinction between the dorsal and ventral pathways proposed by Ungerleider and
Mishkin (1982), via which, respectively, an object’s location and identity are
recognized, also applies to episodic retrieval.
2.2 Human electrophysiological correlates of episodic memory retrieval
Findings from neuropsychological and functional neuroimaging studies in humans have
provided important information about brain regions involved in recognition memory
(section 2.1). Furthermore, functional neuroimaging studies using PET and fMRI
techniques have shown that seven main regions (as discussed in section 2.1.3) are active
while subjects perform memory retrieval tasks. While these techniques can locate
activated neural regions relatively accurately, information regarding the timing of these
activations is limited. The majority of studies have, in fact, used PET methods, which
are suitable for the investigation of sustained modal activity, that is, activations
maintained for the duration of the imaging period. Such studies, while contributing to
the understanding of memory processes such as retrieval mode, do not necessarily
indicate which neural regions are transiently activated during successful retrieval.
However, electrophysiological techniques would appear to possess the required
temporal resolution required for monitoring these fast memory processes, albeit with a
relatively poor spatial resolution. These techniques have been used to investigate the
spatio-temporal patterns of brain electrical activity associated with the retrieval of items
from memory.
While the main focus of this study is ORM, few electrophysiological studies, so far,
have focused on ORM processes as such. Human electrophysiological studies have dealt
more extensively with the retrieval from memory of verbal stimuli and therefore involve
processes specific to humans. Retrieval of objects from memory has also been studied
21

Neural correlates of object recognition memory
using electrophysiological techniques, although the number of studies is far fewer. The
retrieval of verbal and object items has mainly been investigated using tests for episodic
memory retrieval. Episodic memory retrieval is defined as the explicit recollection of
events in one’s personal past (eg. Tulving 1993). Studies that discuss findings in terms
of episodic retrieval have not always been designed so that processes involving
recollection are specifically investigated. In fact, many experimental designs may
promote the tapping into an unconscious form of memory (implicit memory), in which
item retrieval is based on familiarity, rather than on recollection. This does not
necessarily mean, however, that familiarity processes involve different neural regions
(although see Klimesch et al. 2001b, for example). The term episodic memory retrieval
will therefore be used in a broad sense in the following review, indicating retrieval from
some form of longer-term memory. Recognition memory may be a more useful term
than episodic memory retrieval because it does not differentiate between whether items
were recognized on the basis of familiarity or recollection. Recognition memory and
episodic retrieval are terms that will be used interchangeably. The following review will
therefore examine human electrophysiological correlates of episodic memory retrieval
for verbal and non-verbal material.
Electrophysiological methodologies will be outlined in section 2.2.1. EEG correlates of
episodic memory retrieval are reviewed in section 2.2.2. In section 2.2.3, ERP correlates
of episodic memory retrieval are discussed. This section is sub-divided into three parts.
Sections 2.2.3.1 and 2.2.3.2 contain the bulk of the ERP review and focus on transient
item-related effects observed using two different experimental task designs, the
continuous recognition design and the study-test design. Section 2.2.3.3 is a small
section that deals with one study that used the DC-ERP technique to investigate
sustained task-related effects.
2.2.1 EEG and ERP functional electrophysiological techniques: a brief
background
In 1929, when Berger recorded electrical activity from the scalp of a human subject, he
noted that a component of the EEG in the 8-12Hz band was present when the subject's
eyes were closed but disappeared with they were open (Berger 1929). This frequency
22

Neural correlates of object recognition memory
band is now known as the alpha band. He subsequently showed various reliable
correlations between the alpha power and the level of attention (Gloor 1994). Berger’s
major finding was that increased task demand attenuates the alpha power. In so doing,
Berger had shown that the EEG was potentially a powerful tool for studying brain
function; however, it took many decades before the true value of this was realized. More
recently, computers have permitted the analysis of vast quantities of data, which hitherto
had not been practicable. In particular, the use of multi-channel recordings with large
arrays of electrodes has resulted in greatly improved spatial resolution. These
advancements in technology have therefore made the exploration of cognitive activity
using EEG methods more feasible.
The alpha rhythm first described by Berger (1929) is the most prominent feature of the
normal EEG. This rhythm, however, represents only a small proportion of the total EEG
frequency spectrum. The EEG is usually discussed over the 1Hz to approximately 80Hz
range (Regan 1989). The main frequency bands of interest have traditionally been
defined as slow wave delta and theta, from 1 to 3Hz and 4 to 7Hz respectively, alpha
from 8 to 12Hz, and fast beta from 12 to 30Hz. These classifications have been made to
simplify comparisons of power spectra between subjects and subject groups. The
boundaries of these frequency bands vary somewhat between laboratories, and bands
may be further broken down into sub-bands. For example, beta can be separated into
beta1 and beta2, representing the lower and upper regions of the beta band. More
recently, however, because of significant inter-subject variation, frequency bands have
been defined for individual subjects based on the individual’s alpha frequency (IAF).
Other EEG frequency components are then ‘anchored’ to the IAF (see Klimesch et al.
1993; Klimesch et al. 1994 for more details).
Dawson (1947) first reported that distinct electrical responses produced by external
stimulation could be extracted from the EEG. It was subsequently found that similar
responses could also occur without an external stimulus, instead occurring with
internally generated events. These event-related potentials (ERPs) are very difficult to
observe in the EEG as amplitudes are typically between 1/100th to 1/10th of
background (Duffy and McAnulty 1988). To achieve a satisfactory signal to noise ratio,
signal averaging is usually applied to the recorded EEG. Multiple individual responses
(20-2000) may be summed to obtain a waveform representative of the averaging period
23

Neural correlates of object recognition memory
(see Regan 1989, for review). This type of analysis is ideal for observing millisecond
changes that occur during the processing of a stimulus, assuming that the processes
being monitored do not change over the averaging period (Silberstein et al. 1990a).
ERP components that occur less than 200ms after stimulus onset are usually regarded as
exogenous components generated by stimulation of the senses and related to brain
activity associated with perceptual processes. For example, clicks to the auditory
system, flashes to the visual system and electric shocks to peripheral nerves will
produce exogenous potentials (see Regan 1989, for review). Exogenous ERPs are used
clinically to test the integrity of these sensory systems. For example, visually evoked
potentials to a checkerboard stimulus can be used to test optic nerve lesions associated
with multiple sclerosis (eg. see Halliday et al. 1972). ERPs with longer latencies are
usually generated internally by events, such as, for example, surprise, detection of an
infrequent stimulus, or a remembered item. Unlike the earlier potentials, these later
components are independent of the evoking stimulus and represent information
processing after the initial sensory volley. These late potentials have therefore been
associated with cognitive processes (Sutton et al. 1965).
ERPs, therefore, consist of a series of electrical potentials that vary in amplitude and
latency, and are thought to represent the activity of brain regions involved in processing
exogenous or endogenous stimuli (eg. Picton et al. 1995; Regan 1989). For example, the
endogenous P3 or P300 potential (Sutton et al. 1965), which has been extensively
investigated, is a target detection response, and has been linked to attentional and
memory processes (Iragui et al. 1993; Pritchard 1981). This potential is termed P3 or
P300 because it is positive with respect to the reference electrode (eg. Picton et al. 1995;
Regan 1989) and occurs around 300ms after the evoking stimulus has been presented.
Manipulation of attentional and memory parameters have been shown to alter both the
amplitude and latency of the P3 potential (eg. Dujardin et al. 1993).
Electrical signals recorded at the scalp reflect the average activity of synchronously
active neural populations underlying the scalp (eg. Kutas and Dale 1997). However,
only EEG/ERP components generated by neural regions with the dipole orientated
perpendicular to the skull can be detected at the scalp (Picton et al. 1995). Moreover, the
CSF, skull and scalp act to smear the activity generated in underlying neural areas (eg.
24

Neural correlates of object recognition memory
Nunez 1981). Therefore, these techniques permit only gross localization of neural
activity, which must be discussed at the level of, for example, right pre-frontal,
temporo-parietal, occipital regions, etc. These gross patterns of activity have
nevertheless proven useful in differentiating between spatio-temporal patterns evoked
by different classes of experimental stimuli. Compared with other imaging techniques,
EEG/ERP methodologies are popular because of the relatively low cost and the
convenience associated with a non-invasive procedure. Thus recordings in normal and
clinical subjects can be conducted comparatively easily and frequently (Johnson 1995).
A major limitation of these electrophysiological methodologies, however, is that they do
not permit the kind of three-dimensional, high spatial resolution imaging that can be
obtained using fMRI or PET. However, techniques exist (eg. Scherg 1990) that utilize a
large number of electrodes (at least 64) to help locate underlying neural generators from
scalp data. This is known as the inverse problem (eg. Nunez 1981).
2.2.2 EEG changes associated with episodic memory retrieval
In addition to a decrease in alpha amplitude with mental activity, Berger (1929) also
observed a frequency increase. Martinson (1939) made similar observations. Moreover,
alpha activity was found to be selectively reduced in the hemisphere that is dominant in
performing a specific type of task (Adrian and Matthews 1934). More recently, these
early findings have been corroborated (Donchin et al. 1977; Kinsbourne and Hiscock
1983) and extended (Osaka 1984). Osaka (1984) showed that alpha frequency increases
more for difficult tasks than for easy tasks, and only in the hemisphere engaged in the
task.
Pfurtscheller and his colleagues confirmed these aforementioned characteristics of alpha
band power and frequency changes. In addition, they observed transient alpha
attenuation (desynchronization) that appeared to reflect regional increases in cortical
activity associated with the performance of cognitive or motor tasks (eg. Pfurtscheller
and Aranibar 1977b; Pfurtscheller and Klimesch 1990; Pfurtscheller and Klimesch
1991).
25

Neural correlates of object recognition memory
Early studies following Berger’s observations had failed to find any correlations
between alpha characteristics and memory processes, despite the many accounts in the
literature of the association between alpha activity and mental activity (see Klimesch et
al. 1993). In 1985, however, Saletu and Grünberger (1985) found a positive correlation
between alpha frequency and memory performance, although the authors could not rule
out the possible confounding influence of vigilance; that is, good performers may have
been more attentive.
Klimesch et al. (1990b), obtained a similar positive relationship between mean alpha
frequency and memory performance. They later addressed the possible confounding
influence of attention on memory performance in a study where both memory and
attentional demands were varied (Klimesch et al. 1993). A clear relationship between
the individual alpha frequency (IAF) and memory performance was observed. Good
performers exhibited an IAF 1.25Hz higher than poor performers during the retrieval
period of a short-term memory task. Increased attentional and task demands also tended
to reduce IAF although to a much lesser degree. Furthermore, poor performers showed a
greater attenuation of power at this IAF than did good performers.
In a more recent study, Klimesch and colleagues (1994), found that the EEG produced
during the retrieval part of a semantic memory task differed from the EEG produced
during the retrieval part of an episodic task. Semantic memory was defined as ‘pure
long-term memory’ by the authors, and has been described as a store of impersonal
knowledge in the broadest sense (eg. knowledge about language or geography)
(Doppelmayr et al. 1998). Episodic memory was defined as the intentional or incidental
storage of ‘new information’ that can subsequently be retrieved. Klimesch et al. (1994)
used two retrieval tasks (Kroll and Klimesch 1992) to examine firstly, semantic and
secondly, episodic memory. Subjects were firstly presented with a semantic congruency
task in which they had to decide whether sequentially presented ‘concept-feature’ pairs
such as ‘eagle-claws’ or ‘canary-blue’ were semantically congruent or incongruent. The
second task tested episodic memory. In this, subjects were presented with new concept-
feature pairs interspersed with word pairs identical to those presented in the preceding
semantic task, and were required to identify those pairs that had been presented in the
semantic congruency task. Subjects were not informed beforehand that they would be
performing this subsequent recognition task so that they would not be inclined to use
26

Neural correlates of object recognition memory
semantic encoding strategies. In this way episodic memory demands were maximized
(Klimesch et al. 1994). Because both semantic and episodic retrieval processes would
have occurred only after the feature word (second item in the pair) was presented, only
the EEG recorded after this time point would have reflected memory retrieval processes.
Klimesch et al. (1994) predicted that the cognitive demands of the semantic memory
task would be reflected in a decrease in alpha power. They also noted that a number of
converging lines of research suggested that episodic retrieval would be characterized by
theta band changes. Firstly, the late positive component (LPC) of event-related brain
potentials has been associated with memory processes (eg. Fabiani et al. 1990 among
others), and the frequency decomposition of these potentials reveals predominantly
theta frequencies (Klimesch et al. 1994). Therefore, these theta band changes might
reflect episodic memory processes. Secondly, the hippocampus has been shown to be
involved in the generation of the P300 (eg. Halgren et al. 1980), and episodic memory is
compromised with lesions to the hippocampal formation (eg. Scoville and Milner 1957).
Therefore, this area may be focal in episodic memory processing. Finally, theta band
activity has been shown to be the dominant frequency of the hippocampal formation
(eg. Arnolds et al. 1980). This hippocampal theta activity could be transmitted to the
scalp via hippocampo-cortical longitudinal pathways (Klimesch et al. 1994).
Klimesch et al. (1994) found that semantic memory processes were associated with an
attenuation of upper alpha band power. This was consistent with a previous study
(Klimesch et al. 1992) which showed that lower alpha band power was related to non-
task related activity such as attention, whereas upper alpha band power was related to
cognitive processes. In contrast, episodic memory retrieval was reflected by power
changes in the theta band and not in the upper alpha band. Increased theta band power,
or synchronization, was significant only in the episodic task, and only after the feature
word was presented. This dissociation in EEG frequency bands could not have been due
to task difficulty because alpha attenuation would then have been greater for the more
difficult episodic memory task, and this was not the case. The greater alpha attenuation
was exhibited during the semantic congruency task. The authors therefore speculated
that different EEG frequencies reflect different cognitive processes (Klimesch et al.
1994). Klimesch (1996) suggested that short-term episodic memories are reflected by
27

Neural correlates of object recognition memory
oscillations in the anterior limbic system, and long-term semantic processes are
reflected by oscillations in a posterior-thalamic system.
Klimesch et al. (1997) investigated the relationship between theta band power and
episodic encoding and retrieval. Their task involved the intentional memorization of 96
words presented for 1s each, followed by the subsequent recognition of these words
presented amongst 96 semantically similar distractor words (Kroll and Klimesch 1992).
During the encoding phase, only words that were later correctly recognized produced a
significant increase in synchronization of theta activity. Similarly, in the recognition
task, only correctly identified target words produced a significant increase in theta band
power. These increases in theta band power were most pronounced at frontal sites and
smallest at occipital sites. Target words that were not remembered and distractor words
that were correctly identified did not produce any increase in theta power. During the
encoding phase, theta band synchronization appeared to be mirrored by lower alpha
band desynchronization. That is, words that were later remembered produced a marked
desynchronization at lower alpha band frequencies. Words that were not subsequently
remembered produced a significant desynchronization in the upper alpha band.
Therefore, a reduction in lower alpha band power for later remembered words was
consistent with previous findings by this group, thus supporting their proposal that
lower alpha band power reflects increased attention. It would be reasonable to assume
that subjects had paid more attention to those words that were later remembered. The
authors also argued that the upper alpha desynchronization observed with words that
could not subsequently be remembered was due to a triggering of semantic associations
that had produced a detrimental effect on recognition performance. Alpha band
desynchronization in general was most pronounced at occipital sites and smallest at
frontal sites. As already mentioned, theta band synchronization was largest frontally and
smallest occipitally.
The same task design was used in a more recent study by Doppelmayr et al. (1998). The
aim of this investigation, however, was to examine the relationship between EEG band
power and performance in an episodic retrieval task. Band power was observed over the
interval between 500 and 1000ms after the presentation of correctly identified words.
Both high and low performance subject groups showed significant theta synchronization
at all recording sites. Furthermore, high performance subjects showed a higher level of
28

Neural correlates of object recognition memory
theta synchronization in the right hemisphere than did poorly performing subjects.
Greater right hemisphere synchronization of theta band power for high performance
subjects was described by the authors as being consistent with the selective activation
hypothesis (Ibatoullina et al. 1994), wherein high performance subjects show a more
localized and restricted pattern of activation. The authors suggested that poorly
performing subjects utilized additional resources in the left hemisphere to increase their
retrieval accuracy, resulting in a more diffuse pattern of theta band synchronization. It
was suggested that results for high performance subjects are consistent with Tulving’s
HERA (hemispheric encoding/retrieval asymmetry) model (Tulving et al. 1994a). In the
HERA model, the left hemisphere is postulated to be involved in episodic encoding,
whereas the right hemisphere is involved in episodic retrieval. Most PET studies have
indicated that right hemisphere activation during episodic retrieval is restricted to the
pre-frontal cortex (for recent review see Nyberg et al. 1996a). However, Doppelmayr et
al. (1998) observed significant theta synchronization at all right hemisphere sites. They
suggested that the majority of PET studies may have failed to show activation at other
right hemisphere sites because the control task used may have produced a pattern of
activation similar to that of the main task at these sites. This would have reduced any
differences between episodic retrieval and control task activations. In fact, a small
number of PET studies have shown additional involvement of right hemisphere central
and parietal sites in episodic retrieval (Nyberg et al. 1996a; Tulving et al. 1994b).
Doppelmayr and colleagues (2000) also used a similar recognition paradigm to
investigate the relative contributions to theta band power of time-locked evoked and
non-time locked induced theta components, and the relationship of these components to
memory performance. The authors found that the early component of the post stimulus
interval (<400ms) is dominated by an evoked component, whereas the late part
(>400ms) is dominated by the induced theta component. Induced theta for hits was
significantly larger than for correct rejections for the high performance subjects only.
Theta ERPs were analysed separately in an attempt to investigate absolute band power
and possible group differences in the time locked theta response. The authors found that
the theta ERP was much larger for the high performance than the low performance
subject groups. For the high performance group, there was also a correspondence in
time between peaks of the theta ERP and the standard ERP (coincident with the P3
ERP). They suggested that for high performance subjects, theta peaks occur in a
29

Neural correlates of object recognition memory
‘preferred time window’ after the target is presented, and given that they occur
maximally when the P3 can be observed, act as an orienting response that is necessary
for successful episodic recognition. The authors also suggested that the induced theta
component that occurs much later, and is coincident with the late positive component, is
related more to the actual processes of episodic retrieval.
The relationship between induced theta band power and the P3 component was further
highlighted in a study (Klimesch et al. 2000) wherein a memory–related increase in
ERP positivity appeared to be functionally related to a significant increase in both theta
and delta induced band power. Both timing (375-750ms) and topography (largest at left
occipital sites) of these phenomena were similar. The authors suggested that
hippocampal theta activity, transferred to the cortex via limbic-hippocampal-cortical re-
entrant loops, generates the induced theta power and occurs simultaneously with a
transient de-activation of cortical regions which generates the P3 component (see
Klimesch et al. 2000, for further detail).
Studies reviewed thus far where EEG band power has been used to investigate episodic
retrieval have employed verbal stimuli. Klimesch et al. (2001a) used pictures instead of
words in a task design similar to that used by Klimesch et al. (1997). Once again, theta
band changes associated with episodic encoding and retrieval were examined. In this
study, however, changes occurring within the 250 to 750ms interval post stimulus onset
were examined, rather than within the entire 1s interval of Klimesch et al. (1997). The
reduced time frame was employed because this interval has been shown to be important
for memory sensitive ERP components (eg. see Rugg et al. 2000, for review). Klimesch
et al. (2001a) attempted to determine whether theta synchronization during retrieval is
related to the access of a stored code or to more general processes. Furthermore, by
using pictures and comparing findings with those of earlier studies of Klimesch and
collegues (Klimesch et al. 1997; Klimesch et al. 1994), it should have been possible to
determine whether theta band power during encoding and retrieval is different for
pictures and words and therefore stimulus dependent. Theta synchronization was greater
for retrieval than for encoding; however, there was no significant difference between
hits and correct rejections. Because the degree of theta synchronization for hits and
correct rejections was not significantly different, the authors suggested that theta
synchronization during retrieval is related to the attempt to retrieve, rather than to the
30

Neural correlates of object recognition memory
actual access of, a stored memory trace. The authors also concluded that theta changes
during encoding and retrieval do not depend on the nature of the stimulus. However, the
magnitude of theta synchronization during retrieval was several times greater than that
reported in previous studies utilizing verbal material. As greater theta synchronization
has been linked to better performance (Doppelmayr et al. 1998), the authors argued that
the greater theta synchronization for pictures was related to the fact that picture memory
is superior to verbal memory.
Theta changes were also assessed in a word-based retrieval task in which the level of
conscious awareness was manipulated (Klimesch et al. 2001b). Subjects made
remember/know judgements by indicating whether they recollected, or remembered, the
specific event of word presentation, or whether they recognized, or knew, a word based
on familiarity alone without recollection of the event. Theta synchronization was
observed for both ‘remember’ and ‘know’ judgements, but in different temporal
windows. Theta synchronization and a more positive ERP (see section 2.2.3) was
associated with ‘knowing’ in the 300 to 450ms interval post onset, and with
‘remembering’ in the 450 to 625ms interval. The authors suggested that knowing based
on familiarity is associated with activity within the perirhinal cortex and medial dorsal
nucleus of the thalamus, whereas remembering, which occurs later, is associated with
hippocampal activity.
In summary, episodic retrieval of verbal stimuli is associated with increased
synchronization in the EEG theta band, particularly at frontal sites. Moreover, theta
synchronization appears to be related to task performance; high performance subjects
show greater theta band synchronization in the right hemisphere than do low
performance subjects. These theta band changes appear to be specific to episodic
memory retrieval as they do not occur with semantic retrieval. Semantic retrieval, in
contrast, is associated with upper-alpha band desynchronization. However, this theta
band finding has more recently been questioned in a study indicating that theta
synchronization reflects the attempt to retrieve, rather than the actual access of stored
information. Notwithstanding a number of important studies that have indicated a
relationship between EEG theta band power and episodic memory retrieval, the neural
regions involved with generating and modulating theta changes, and the precise
31

Neural correlates of object recognition memory
relationship between theta band power and episodic memory retrieval processes remains
unresolved.
2.2.3 Event-related potential changes associated with episodic memory retrieval
Compared to PET and fMRI imaging techniques, the superior temporal resolution of
ERP techniques has allowed investigation of faster neural changes associated with
successful memory retrieval. These transient neural changes have been investigated
using a number of experimental designs and different stimulus types. Most studies that
will be reviewed used scalp electrodes, although a smaller number of clinical studies
have used depth electrodes located within the brain. In most cases ERPs elicited by
previously studied and unstudied items (ERP old/new effect) are compared in attempts
to determine the neural regions associated with successful retrieval processes. This type
of approach has shown consistent parietal and right frontal ERP differences within
certain time intervals following item presentation.
Sections 2.2.3.1 and 2.2.3.2 contain the bulk of the review of ERP studies and focus on
transient item-related effects observed using two different experimental task designs,
the continuous recognition design and the study-test design. Section 2.2.3.3 is a small
section that deals with one study that used the ERP technique to investigate sustained
task-related effects.
2.2.3.1 ERP correlates of episodic memory retrieval: evidence from continuous
recognition paradigms
This section deals with studies utilizing the continuous recognition experimental task
design to investigate ERP correlates of episodic memory retrieval. With this design,
single stimulus items, such as words or pictures, are presented in a continuous sequence,
and subjects are required to identify those items that are presented for the first time
(new) and those that are repeated (old). Correct identification of repeated items
presumably taps into some form of recognition memory. This type of experimental
design has traditionally been used to explore the transfer from immediate memory, also
32

Neural correlates of object recognition memory
known as primary memory, to short or long term memory, also known as secondary
memory. This is done by varying the time lag between repeated items and observing the
change in various behavioural parameters, for example, the decay of recognition
accuracy (Friedman 1990a). Findings from continuous recognition studies are reviewed
separately from findings from study-test studies (section 2.2.3.2) as there are distinct
differences in the two experimental paradigms (eg. see Friedman 1990a; Johnson 1995,
for review). The main difference lies in the separation of encoding and retrieval phases
in the study-test paradigm. These phases occur concurrently in the continuous
recognition paradigm. This is a particularly important distinction when sustained, task-
related processes (see sections 2.2.3.3 and 2.2.4) are being investigated. In the
continuous recognition paradigm each new stimulus must be encoded for possible later
recognition. This intermixing of retrieval and encoding processes is therefore a
confounding factor when investigating sustained task effects associated solely with
retrieval. Furthermore, while repetition lags of less than 1 minute are common in
continuous recognition tasks, intervals between encoding and retrieval phases of
approximately 5 minutes are usual in study-test tasks. It has been suggested that ERPs
elicited using short repetition lags may not reflect longer-term episodic memory. Puce et
al. (1991) suggest that a lag greater than 45s is necessary for transfer into long-term
memory. However, definitions and models of memory are varied, and there are many
other variables such as rate of presentation, familiarity, distraction, age, etc. (eg.
Baddeley 1999) that may be important in considering the exact types of memory
processes being accessed.
A review of studies that have used continuous recognition tasks to investigate neural
correlates of episodic memory retrieval follows. A review of studies where ERPs have
been elicited by verbal stimuli will firstly be presented, followed by a discussion of their
possible relevance to episodic memory retrieval. Attempts have been made to
distinguish between the ERPs generated in the continuous recognition paradigm and
other ERPs, such as the P300, which have been associated with other cognitive
processes. A small number of studies that employed both scalp and depth electrode
recordings in epileptic patients, and utilized pictorial stimuli of both a verbalizable and
abstract non-verbalizable nature, will then be reviewed.
33

Neural correlates of object recognition memory
Early studies utilizing the continuous recognition paradigms found differences between
ERPs elicited by new and repeated verbal stimuli (eg. Johnson et al. 1985; Sanquist et
al. 1980). The main finding reported in these studies is that ERPs elicited by repeated
items were more positive than those elicited by new items. It was suggested that this
was due to an enhancement of the P3 and related late positive components (eg. Neville
et al. 1986).
Smith et al. (1986) described a negative-positive ERP complex, N400-P600, which was
originally evoked during a verbal study-test task, and was also reported in a number of
continuous recognition studies (eg. Nagy and Rugg 1989; Rugg and Nagy 1989; Smith
and Halgren 1988). In all of these studies, compared with the presentation of new items,
repeated stimuli produced an attenuated N400 component and an enhanced P600
component. These differences in ERP components generated by new and repeated items
is known as the ERP old/new effect, or sometimes the ERP repetition effect. Rugg
(1990) referred to the modulation of the N400 potential, occurring between 300 and
500ms, as the ‘early’ ERP old/new effect, and changes to the P600 (P3), occurring
between 500 and 800ms, as the ‘late’ ERP old/new effect. Since both early and late
components appeared to vary in the same direction, it was suggested that an overlapping
increase in positivity or decrease in negativity mediated the old relative to the new
effect (Halgren and Smith 1987).
The ERP old/new effect for words has been described as being most prominent over
central and posterior electrode sites, and larger over the left than the right hemisphere
(Johnson 1995). The late ERP old/new effect for verbal stimuli is more uniformly
distributed from frontal to occipital regions (Johnson 1995), although in some studies
larger differences have been reported over left parietal and left temporal regions
(Neville et al. 1986; Rugg and Doyle 1992).
ERPs with the approximate timing and polarity of the N400 and P600 have been
reported in a number of other studies not dealing with episodic retrieval. The N400, a
negative centroparietal potential, occurring at approximately 400ms in response to
semantic incongruities in sentences, was first reported by Kutas and Hillyard (1980). An
N400 has also been recorded with other task paradigms involving, for example, picture
naming and lexical decisions (eg. see for review Johnson 1995; Puce et al. 1991), while
34

Neural correlates of object recognition memory
attenuation of the N400 amplitude has been reported in unstudied, or incidental, word
repetition tasks that do not involve explicit memory processes (see for example Paller
and Kutas 1992; Rugg 1987). The P600 potential is reported as having a similar
topography to the P300 (Sutton et al. 1965), a potential elicited, for example, in the
detection of rare tones in an auditory discrimination task (eg. see Picton 1992, for
review).
Friedman (1990a) argued that the scalp distributions of early and late ERPs recorded in
continuous recognition paradigms are different from those of the aforementioned N400
and P300 components. Friedman (1990a) reported a frontal N400 ERP old/new effect,
and argued that this pattern was quite different from the many other scalp N400
distributions that had been previously reported. These previously reported distributions
were characterized mainly by parietal, frontal, or both parietal and frontal negativities.
Besson et al. (1992), however, noted that the spatio-temporal pattern of the early
negativity evoked in their study closely matched the N400. Halgren and Smith (1987)
suggested that the N400 represents ‘associative activation’ resulting in the formation or
the retrieval of a memory traces. More recently though, it was suggested that the early
old/new effect is not related to longer-term episodic memory processes because this
effect disappears with lags between repeated items greater than 2mins (see for example,
Nagy and Rugg 1989). Johnson (1995) in fact argued that the early ERP old/new effect,
as indexed by the N400, has little to do with explicit memory processes.
Friedman (1990a) also indicated that the P600 recorded using continuous recognition
designs is distinct from the P300 recorded using target detection designs and suggested
that these are not generated by the same processes. Traditionally the P300 has been
characterized by a parietal maximum. Friedman (1990a), however, reported a frontal
positive maximum which occurred significantly later than the P300. A distinction
between these two positive waveforms has been further supported by many other studies
(eg. Friedman 1990b; Paller and Kutas 1992; Puce et al. ; Rugg 1987; Smith 1993;
Smith and Guster 1993). Smith and Guster (1993) demonstrated a distinction between
the late P600 potential and a separate, temporally overlapping, P3b potential. The
authors concluded that, unlike the P3b component, the P600 component was not related
to the ‘targetness’ of the verbal stimuli but rather to retrieval from secondary memory.
Friedman (1990b), among others, also ruled-out the possibility that modulation of the
35

Neural correlates of object recognition memory
late positive potential is due to stimulus probability effects; a well documented P300
effect (eg. see Pritchard 1981, for review). Puce et al (1991), using depth electrodes
located inside the medial temporal lobes, demonstrated a double dissociation between
the late positivity evoked in continuous recognition paradigms, and the P300 evoked in
an oddball task. These reported functional differences between the P300 component and
the late positive component have added weight to the argument that the late ERP
old/new effect reflects memory retrieval processes (Johnson 1995).
Despite the majority of studies equating the late ERP old/new effect with recognition
memory processes, there has been considerable debate regarding the more specific type
of memory that is being accessed. Dual process models of memory (Jacoby 1991;
Mandler 1981) attempt to address, for example, the frequently recounted experience that
one can recognize someone’s face but not necessarily recollect that person’s name, or
when or where that person was previously encountered. Dual-process models explain
this phenomenon by incorporating two distinct components of memory: recognition
based on familiarity and recollection. Familiarity has been described as being automatic
and unconscious, whereas recollection is characterized by conscious processing
requiring retrieval of context specific information. These main characteristics have led
researchers to equate recognition based on familiarity and recollection with implicit and
explicit memory respectively.
Rugg and his colleagues (Rugg 1990; Rugg and Doyle 1992; Young and Rugg 1992)
argued that the ERP old/new effect is a familiarity effect and hence based on implicit
memory. They reported the absence of a late ERP old/new effect for high frequency
words but not low frequency words, and therefore concluded that repetition enhances
the ease with which an item is perceived and thus increases its ‘local’ familiarity.
Further support for the familiarity argument was provided by Potter et al. (1992). They
administered the anticholinergic agent scopolamine because of its known detrimental
effect on explicit memory processes. If the ERP old/new effect is based on explicit
memory processes, then the magnitude of this effect should have be been reduced with
scopolamine. However, the ERP old/new effect was unaltered despite a decrement in
recognition performance. Potter et al. therefore concluded that the ERP old/new effect is
not mediated by cholinergic neural activity. Instead, they concluded that the
36

Neural correlates of object recognition memory
scopolamine had an adverse effect on recollection, resulting in an increased reliance on
relative familiarity as a cue for recognition judgements.
Other investigators have equated the ERP old/new effect with explicit memory
processes (eg. Smith 1993; Smith and Guster 1993). Smith and Halgren (1989) first
suggested this in the light of the results of a clinical study which used the study-test
paradigm. Here, patients, who had undergone left temporal lobectomy, failed to show a
late ERP old/new difference. This indicated that the hippocampus, a component of the
left medial temporal lobe, is involved in the generation of the old/new difference.
Because previous findings had implicated the hippocampal formation in explicit
memory function (Squire and Zola-Morgan 1991) it was concluded that the ERP
old/new effect reflects recollective processes rather than familiarity-based implicit
memory processes. Johnson (1995) also argued that if the ERP old/new effect was based
on aspects of implicit memory such as familiarity, the amplitude of the late component
of the ERP old/new effect should steadily increase with continued repetition of items.
Instead, Johnson (1995) noted that the magnitude of the ERP old/new effect does not
change with repetition, a finding consistent with a basis in explicit memory processes.
Smith (1993) tackled this issue more directly using a study-test experimental design in
which subjects were required to indicate whether they consciously recollected the
presentation of an item, or whether they simply knew that it had been presented. The
P600 component was more positive going for correctly recollected items than for those
simply known to have been presented. Many subsequent study-test experiments have
attempted to clarify the issue of whether the ERP old/new effect is based on familiarity
or recollection. A more detailed review of these studies is presented in the next section
(section 2.2.3.2).
As already indicated, there are only a few studies on episodic memory retrieval that
have used non-verbal stimuli. Friedman (1990a) used pictures of common objects
presented in a task designed to determine whether the scalp ERP old/new effect would
be the same as that described in verbal studies. They also hoped to determine whether
representation of the picture would be transferred from primary to secondary memory.
The authors used 3 different repetition lags of 2, 8 and 32 items. They argued that,
according to the model of Waugh and Norman (1965), ERP changes should indicate a
transfer from primary memory, when the lag is 2 items, to secondary memory, when the
37

Neural correlates of object recognition memory
lag is 8 or 32 items. As was the case with ERPs with verbal stimuli, an early negativity
(N300) followed by a late positivity (P600) was observed, and again, repeated stimuli
produced a smaller N300 and larger P600 component. Unlike verbal studies, which
showed spatially diffuse ERP old/new differences between novel and repeated items,
this pictorial study showed more localized differences that were largest at frontal sites
for the N300, and at frontal and central sites for the P600. Friedman suggested that the
N300 reflected memory retrieval processes, as this component was significantly larger
when evoked by new items than by old items. He also argued that the P600 was more
likely to reflect processes relating to the organization and execution of the response
rather than to those involved in the recognition and/or classification of a picture.
However, Friedman failed to find a difference between ERPs obtained with different lag
intervals. This may have been partly because pictures require less elaboration than
words during encoding, and partly because they are more differentiable and are less
likely to interfere with other pictures. Friedman suggested that pictures might be
transferred from primary to secondary memory more readily than words.
Findings of clinical studies using the continuous recognition task design have indicated
possible neural structures involved in the generation and modulation of ERP old/new
components associated with memory retrieval processes. Puce et al. (1991) recorded
ERPs from epilepsy patients prior to surgery using depth electrodes located within the
temporal lobes. They used a continuous recognition task with lags to repeated items
greater than 45s. A relatively long lag time was used in order to engage longer-term
recognition memory processes. Unlike Friedman (1990a), however, who used common
objects as stimuli, Puce et al. used both abstract non-verbalizable stimuli and verbal
stimuli, arguing that the ERPs evoked using abstract non-verbalizable stimuli were more
likely to reflect ‘pure visuospatial recognition memory.’ The N400 was larger for novel
stimuli, and the P600 larger for repeated stimuli, for both verbal and non-verbal items,
findings similar to those of Smith et al. (1986). Puce et al. (1991) also found no
significant difference in the ERPs evoked by verbal and non-verbal stimuli, with both
stimulus types evoking larger right hemisphere ERPs. Also, polarity reversals within the
mid and posterior hippocampus and hippocampal gyrus were observed, suggesting a
local generator in medial temporal regions; a finding also reported by Smith et al.
(1986). For 3 patients with unilateral white matter lesions, the N400 and P600
components were absent. Puce et al. (1991) discussed this finding in terms of a possible
38

Neural correlates of object recognition memory
interruption in the visual recognition pathways (Mishkin 1982) caused by the white
matter lesions in these patients (see section 2.1.1).
Guillem et al. (1995b) extended previous findings by examining depth ERPs recorded at
a larger number of sites, including a number in parietal, occipital and frontal regions.
They used verbalizable pictorial stimuli, and examined ERPs to novel and repeated
stimuli with a lag of 6 intervening items. They found that anterior temporal, prefrontal,
and possibly parietal regions, were involved in generating and modulating N400 and
P600 components. In addition, the authors suggested that the regions responsible for
generating the resultant N400/P600 scalp ERPs were likely to interact with each other to
modulate these ERPs.
Halgren et al. (1992) proposed that activity originating in anterior temporal, prefrontal
and parietal regions reflects three different aspects of cognitive processes involving
three different sub-systems. The first of these is involved in the generation of the scalp
P3a and includes neural regions contributing to the parieto-frontal attentional system.
The second sub-system is involved in the generation and modulation of the scalp N400
and includes medial temporal structures and parietal structures. The third sub-system is
involved in the production of the P600 (P300 or P3b) component. This component is
mediated by temporal and superior parietal structures, and reflects cognitive closure
processes (see Guillem et al. 1995b, for more specific details of brain regions involved;
Halgren et al. 1992).
ERP data obtained using a longer lag between repeated items was presented in a follow-
up paper by Guillem et al. (1996). Once again, the ERP old/new effect was observed at
various sites within the brain using depth electrodes. This study specifically examined
the involvement of temporal regions in the generation and/or modulation of the
N400/P600 scalp potentials. In addition, the relative contribution of temporal regions to
short-term and longer-term memory processes was examined by using lags of 6 and 19
items respectively. Once again, verbalizable pictorial stimuli were used. A strong N400
old/new effect was observed in all temporal lobe structures examined, although the
magnitude of the effect varied with the recording site and lag. Because the magnitude of
the N400 old/new effect differed with the different lags, the authors speculated that
temporal lobe structures contribute differently to short-term and long-term memory
39

Neural correlates of object recognition memory
processes. In contrast, the P600 old/new effect was not consistently observed. The
authors suggested that the nature of the stimuli might have contributed to this outcome.
Friedman (1990a), who also used highly verbalizable stimuli, also reported that the
P600 old/new effect was less pronounced than the N400 old/new effect. Because there
was a large N400 old/new difference at posterior temporal regions for a lag of 6, but not
for a lag of 19, Guillem et al. suggested that the posterior temporal regions are
specifically involved in short-term memory processes. The authors also found that the
amygdala, hippocampus and anterior temporal regions contribute to both short-term and
long-term memory processes. Moreover, the authors suggested that these regions also
play an independent role in long-term memory processes in general.
In summary, the majority of verbal and pictorial studies utilizing the continuous
recognition paradigm reported an increased ERP positivity for repeated items compared
to new items in the 300-800ms post onset interval. The early ERP old/new effect,
although consistently observed in continuous recognition tasks, decays rapidly, and
therefore appears to be unrelated to memory retrieval. The late ERP old/new effect
appears to be related to recognition memory; however, the precise form of memory has
not been conclusively determined in continuous recognition studies. This issue has been
more thoroughly investigated using the study-test experimental design and is discussed
in the following section. The scalp ERP old/new effect occurs, in the majority of cases,
over parietal and central regions with a left hemisphere bias for verbal stimuli, and over
frontal and central regions for verbalizable pictorial stimuli. Depth electrode recordings
have identified a number of brain regions involved in generating and modulating the
late ERP old/new effect. These regions appear to respond differently to verbal stimuli
and to both verbalizable and abstract non-verbalizable pictorial stimuli. While a number
of brain regions have been implicated in the ERP old/new effect, involvement of medial
temporal lobes has been reported consistently for both verbal and non-verbal stimuli.
The additional involvement of parietal and prefrontal sites has also been observed with
verbalizable stimuli. Due to the paucity of continuous recognition studies utilizing
pictorial stimuli, it is not yet clear if the spatial distribution of the ERP old/new effect at
the scalp is essentially the same for verbal and non-verbal stimuli.
40

Neural correlates of object recognition memory
2.2.3.2 ERP correlates of episodic memory retrieval: evidence from study-test
paradigms
Although the continuous recognition task design has been used to study episodic
memory retrieval, this design suffers from certain disadvantages. Firstly, in the
continuous recognition design (section 2.2.3.1), encoding and retrieval processes are
intermixed in the one task making separation of encoding and retrieval processes
difficult. Furthermore, lags between repeated items are generally short, thereby reducing
dependence on long-term memory. The study-test experimental design does not suffer
from the aforementioned disadvantages. With this design, words or pictures are studied
in an encoding task and are later retrieved in a task that is usually presented
approximately five minutes later (although see Duzel et al. 1999, who used a lag of
30s). Thus, encoding and retrieval occur in separate phases, and the relatively long time
lag between encoding and retrieval also increases dependence on longer-term memory
processes.
A number of variants of the study-test format have been used to determine more
precisely the type of memory processes associated with the ERP old/new effect. Many
studies have investigated whether retrieval processes are based on recollection or on
familiarity. Most studies focusing on recollection have used task designs that maximize
the contribution of explicit memory processes, thereby facilitating the investigation of
ERP changes associated with this aspect of memory. Under these conditions, the ERP
old/new effect at parietal sites is largest and is accompanied by a right frontal effect that
occurs over a similar time frame, that is, it begins approximately 500ms after stimulus
appearance and lasts longer than 1s. While the parietal ERP old/new effect appears to be
obligatory, and largest when retrieval is based on recollection, the presence of a right
frontal effect depends on the task used and reflects monitoring processes. While most
experimental designs and subsequent analyses have focused on the investigation of
processes associated with retrieval success, as indexed by the ERP old/new effect, other
aspects of retrieval processes such as retrieval effort and retrieval strategy have also
been investigated and will be reviewed in this section. In these studies, the contribution
of frontal regions is highlighted. Finally, clinical studies will be reviewed. These studies
provide further insights into the origin of ERP old/new effects and the memory
processes that they might reflect.
41

Neural correlates of object recognition memory
Paller and Kutas (1992) were perhaps the first to test directly the nature of the cognitive
activity associated with the ERP old/new effect. They used ‘a levels-of-processing
manipulation’ design, wherein subjects performed two tasks designed to give identical
priming, which involves implicit memory, in the study phase but different recall
associations during the subsequent test phase. Subjects had to study words in relation to
their meaning in one task and letter identity in the other. In the test phase, subjects
performed a word-identification task in which words were presented very briefly (33-
50ms). While behaviourally the frequency of identification of old words was the same
for both study categories, there was dissociation in ERP activity. The authors argued
that they had demonstrated an electrophysiological correlate of recollection because the
physical characteristics of the words from the two study groups were identical, priming
was the same, and the behavioural responses were not significantly different. The
authors therefore hypothesized that the ERP old/new effect represents an
electrophysiological correlate of conscious recollection, distinct from priming processes
or other confounding influences.
Smith (1993) conducted the first study that attempted to determine whether the ERP
old/new difference reflects explicit or implicit memory processes. He used an
experimental paradigm developed by Gardiner (1988) designed to investigate the
relationship between recognition and conscious awareness. In the test phase, in addition
to making an old/new judgement for words that had been presented in the study phase,
subjects also had to indicate whether they consciously remembered (R) each word, or
simply knew (K) that the word was old. Gardiner (1988) had equated R responses with
explicit memory and K responses with implicit memory. In Smith’s study, these
corresponded to recall based on recollection and familiarity respectively. Smith found
that R words elicited larger and more positive ERPs than K words. Both R and K words
were, however, more positive than correctly identified new unrepeated words.
Furthermore, old words that were incorrectly identified did not show the ERP changes
characteristic of correctly identified old words. The authors argued on this basis that
both R and K ERPs were related to explicit memory retrieval processes rather than
implicit memory processes. However, Johnson (1995) later suggested that differing
spatio-temporal patterns of R-K and K-new differences in Smith’s study represented
distinct memory processes emanating from different brain regions. Johnson noted that
42

Neural correlates of object recognition memory
the spatio-temporal patterns of the R-K difference and the ERP old/new effects were
similar, that is, the R-K difference was small and equipotential over frontal, central and
parietal scalp regions, with an onset at around 550ms. The K-new difference was,
however, characterized by a centro-parietal distribution, similar to that of the P300, and
with an onset at around 400ms. Johnson (1995) thus concluded that the R-K difference
depends on conscious explicit memory processes, whereas the K-new difference may
depend on implicit memory processes.
Duzel et al. (1997) used the R/K paradigm to explore conscious awareness of memory
for past events. They argued that R responses reflect ‘autonoetic’ awareness, that is, a
re-living of one’s personal past (episodic memory), and K responses reflect ‘noetic’
awareness and reflect one’s interaction with the environment in the present. They
argued that true and false R judgements should produce the same ERP changes, given
that they both reflect the subject’s subjective awareness, independent of whether the
targets were presented or not. To test this hypothesis, they used an experimental
paradigm in which subjects responded to old words that were previously presented in
the study phase (true targets), new words that were not presented in the study phase but
were semantically related to words presented in the study phase (false targets), or words
that were new and not semantically related to words presented in the study phase.
Subjects had to decide which of the three categories each word belonged to. Firstly, they
responded to each word in the test phase by indicating whether the word was old or
new, and then, if old, whether they remembered (R) the event of the word’s presentation
or simply knew (K) that the word had been in the study list. Duzel et al. argued that
ERPs derived using this three-way classification of subjects’ subjective assessment
should reflect three distinct states of consciousness: (i) memory of the event of the
word’s presentation in the study list (remember or autonoetic awareness); (ii) memory
that that word was presented, but no memory of the event (know or noetic awareness);
(iii) no awareness of any kind that the item was presented (unawareness of the past).
ERPs associated with autonoetic and noetic awareness were compared with ERPs
associated with correct rejection of new words. Autonoetic awareness was associated
with a late positivity occurring between 600 and 1000ms post onset over left temporo-
parietal and bifrontal regions. ERPs associated with noetic awareness displayed an
earlier bilateral temporo-parietal positivity between 300 and 600ms, and a later bifrontal
43

Neural correlates of object recognition memory
negativity between 600 and 1000ms. Moreover, consistent with their hypothesis, the
ERPs generated by true and false recognition were identical. Duzel et al. noted that the
spatio-temporal ERP patterns associated with autonoetic awareness were similar to the
previously reported patterns associated with the late positive component (P600). A
different pattern of activity for K judgments led the authors to suggest that the late
positive component is related to autonoetic conscious recollection. Furthermore, given
the similarity between the ERPs elicited by true and false targets, it was concluded that
these ERPs were not sensitive to whether the words were studied or not, rather, they
reflected neural changes associated with the subjects’ subjective awareness.
Wilding and his colleagues (1995) used a study-test design to explore the extent to
which the ERP old/new effect stems from familiarity-based retrieval and the extent to
which it stems from recollection-based retrieval. During the study phase, subjects
performed a lexical decision task in which they were presented with words, visually and
auditorily, and had to make word/non-word judgements. Subjects were unaware there
would be a subsequent test phase. During the test phase, subjects had to make a forced
old/new decision, and for items judged as old, a further forced decision regarding the
context in which the word was presented, namely visual or auditory. To ensure that test
modality did not confound the results, test phase words were presented visually in one
experiment and auditorily in a second. The authors argued that ERPs elicited by old
items that had been correctly assigned contextually reflected recollection, whereas
correctly judged old items with incorrect assignment of study context must have been
judged old on the basis of familiarity rather than recollection.
In both experiments, that is, when the test phase words were presented both visually and
auditorily, the ERPs elicited by words correctly assigned to study modality were more
positive than the ERPs elicited by new unstudied words. When the test phase words
were visually presented, the ERP old/new effect was absent for words incorrectly
assigned to study modality, whereas, when they were presented auditorily, it was
present, although in a restricted time interval (400-800ms). The authors argued that the
absence of an ERP old/new effect for incorrectly assigned words with the visually
presented test phase words was due to poor performance, and therefore was not used in
further interpretation of the data. The ERPs elicited by studied words, whether correctly
or incorrectly assigned to study context, were more positive than the ERPs elicited by
44

Neural correlates of object recognition memory
new unstudied words. Between 400 and 800ms post stimulus onset, the ERPs for the
studied words, regardless of whether they were correctly or incorrectly assigned, were
essentially the same. However, the correctly assigned words generated an extended
positivity 800 to 1100ms post stimulus onset that was not seen with the incorrectly
assigned words. This difference, 800 to 1100ms post-onset, led Wilding et al. to
conclude that the late ERP old/new effect for correct contextual judgements was based
on recollection rather than familiarity. However, they noted that this finding was open
to interpretation because of the similarity of the ERPs evoked by correct and incorrect
contextual judgements in the 400 to 800ms interval. Wilding and Rugg (1996) later
proposed that the similarity of ERPs in this range was consistent with the proposal of
Johnson et al. (1993), namely, that words for which study modality was incorrectly
specified had engendered weak or partial recollection, insufficient for recollection of
modality but sufficient for recollection of its earlier presentation.
Wilding et al. (1995) discussed their findings in relation to a model that suggests that
both familiarity-based recognition and recollection may rely on declarative memory
systems involving both hippocampal and frontal regions (Moscovitch 1992; Moscovitch
1994; Squire 1994). In this model, familiarity-based recognition can be achieved solely
with hippocampal output. Recollection, involving retrieval of contextual information, is
achieved through further integration with the prefrontal cortex. In line with this
proposal, Wilding et al. (1995) suggested that because ERPs elicited by correct and
incorrect contextual judgements had similar spatial distributions in the 400 to 800ms
post-onset interval, they might both be generated by the hippocampal formation.
Moreover, the prolongation of the ERP old/new effect for words for which study
modality had been correctly specified might reflect the involvement of the frontal lobes
in recollection. However, the authors cautioned that further investigation was necessary
because, for the correctly assigned words, the difference between the spatial
distributions of the ERP old/new effect within the two time intervals was not
statistically significant.
In case fluency effects occurring when the test and study phase words were in the same
modality had contributed to correct word assignation, rather than recollection per se,
Wilding and Rugg (1996) performed a further study where this wouldn’t occur. In the
study phase, all the words were auditorily presented, and in the test phase, all the words
45

Neural correlates of object recognition memory
were visually presented. In the study phase, subjects listened to words spoken by either
a male or female voice. In the recognition phase, in addition to deciding whether the
visually presented words were old or new, subjects also had to decide whether words
judged old had been spoken by a male or female voice. The ERPs elicited by old words
for which the study context was correctly specified should have reflected processes
involving recollection rather than familiarity-based recognition.
The ERPs elicited by words correctly judged old were more positive than those elicited
by correctly identified new items. This ERP old/new effect could be dissociated into
two spatially and temporally distinct components, a large left parietal component
between approximately 400 and 1000ms post stimulus onset, and a right frontal
component beginning at about the same time and sustained for the length of the
recording epoch (>1400ms). The parietal difference was likened to the widely reported
old/new modulation of the P600 potential (Neville et al. 1986; Rugg 1995), while the
right frontal component did not appear to have been reported previously. ERPs elicited
by correctly assigned old words were more positive than ERPs for old words incorrectly
assigned, although this difference was not statistically significant. The authors
concluded, as did Wilding et al. (1995), that ERPs elicited by words for which the
context was correctly and incorrectly specified were qualitatively the same, and were
likely have been generated by the same combination of neural structures. The authors
also concluded that the increased positivity over left parietal areas for old items relative
to new reflects simple memory of prior occurrence, whereas the right frontal increased
positivity for old items relative to new, an effect not previously reported, may reflect
recovery of contextual information. Moreover, the right frontal changes could have
indicated the involvement of the prefrontal cortex. This would be consistent with
neuroimaging evidence of activation within the right dorsolateral prefrontal cortex
during tasks requiring episodic retrieval, and the clinical observations that patients with
prefrontal lesions have poor source memory (Schacter et al. 1991), that is, they have
poor recollection of the details of an event while still knowing of the occurrence of the
event.
In both studies by Wilding and colleagues (Wilding et al. 1995; Wilding and Rugg
1996), the authors concluded that the ERP old/new effect is more likely to reflect
recollection of old items rather than familiarity with them. Moreover, given the
46

Neural correlates of object recognition memory
similarity of ERP amplitudes for correct and incorrect contextual judgments, it was
suggested in both papers that recollection is a graded rather than an all-or-none process.
The authors argued that their results were consistent with Rugg’s (1995) proposal that
the ERP old/new effect reflects the amount or quality of information retrieved in
response to the old item. To tackle this issue more directly, Wilding (2000) also used
the presentation context of male versus female voice, but subjects were also required to
retrieve further information about the words encountered during the study phase. During
the study phase, in addition to listening to words spoken in male or female voice,
subjects had to make either an ‘action’ or ‘liking’ judgement about each word. That is,
they had to decide whether the word was either active or passive, or pleasant or
unpleasant. During the test phase, for words judged old, subjects also had to decide
whether they had been spoken in a male or female voice, and whether an action or
liking judgement had been made. The magnitude of the parietal ERP old/new effect
increased with the number of accurate source judgments. Wilding therefore concluded
that the parietal ERP old/new effect indexes recollection in a graded fashion.
Wilding (1999) used a similar approach to explore possible variations in the spatio-
temporal patterns of ERPs associated with different retrieval strategies. Here, the ERPs
associated with the two retrieval tasks, namely, deciding whether the voice was male or
female, and whether an action or liking judgement had been made, were compared. The
task was designed to explore possible variations in ERP spatio-temporal patterns
associated with retrieval strategy. Similar ERP old/new effects were seen at parietal
electrodes for the two retrieval tasks. Moreover, given the similarity between the ERPs
associated with correct and incorrect source judgements, the authors concluded, as
previously, that the parietal ERP old/new effect indexes the amount or quality of
information retrieved. The right frontal ERP old/new effect was, however, larger for the
word association condition than for the voice condition. The authors argued that the
parietal ERP old/new effect not only reflected successful retrieval, but also the type of
source information that was retrieved from memory.
Donaldson and Rugg (1998) used an associative recognition task format to vary the
likelihood that items would be recollected. This was modelled on the context-based
recognition design used by Wilding and Rugg (1996), but was designed in such a way
that word associations probed recollective processes. In the study phase, word pairs
47

Neural correlates of object recognition memory
were shown. In the test phase, three categories of word pairs were shown: the same
word pairs shown in the study phase, re-arranged word pairs made from new
combinations of studied words, and new word pairs using previously unstudied words.
For each word pair presented in the test phase, subjects initially made an old/new
judgment on whether the word pair contained old studied words or new unstudied
words. For word pairs containing studied words, subjects made a further judgment,
indicating whether the word pair was the same pairing as that presented in the study
phase or whether the studied words had been re-arranged. The authors argued that same
pairs should engage recollection processes to a greater extent than rearranged pairs
because correct identification of same pairs would be indicative of recollection of the
actual pair occurring in the study phase.
Compared with the ERPs for unstudied new pairs, the ERPs for same pairs exhibited an
early left parietal and bilateral frontal positivity between 600 and 1200ms. By the end of
the recording epoch (>1200ms), the left parietal positivity remained and was
accompanied by a frontal positivity, now lateralized to the right. The spatio-temporal
patterns of these positivities were similar to the context-based (male/female voice) ERP
old/new effect reported by Wilding and Rugg (1996). Rearranged pairs showed patterns
of activity that were qualitatively similar, but markedly smaller. Donaldson and Rugg
argued that larger ERPs in response to same pairs compared with rearranged pairs was
consistent with their proposal that correct identification of same pairs involves
recollection to a greater extent than rearranged pairs. Moreover, the similarity of these
ERP old/new effects to those of Wilding and Rugg (1996) suggested that associative
recognition memory might involve the same recollective processes as those utilized in
context-based recognition memory.
In a second experiment performed by Donaldson and Rugg (1998), designed to
investigate recognition in the absence of association, subjects were only required to
make old/new judgments. They were not required to judge whether the word pairs were
the same as those presented in the study phase or were re-arranged. A similar pattern of
results was again evident, namely, a sustained left parietal positivity followed by a
prominent right frontal positivity, although the ERP old/new differences were smaller
than those observed in the first experiment. The occurrence of a prominent right frontal
ERP old/new effect in both experiments indicated that the additional associative
48

Neural correlates of object recognition memory
judgement made in the first experiment did not cause the right frontal effect. Instead,
Donaldson and Rugg (1998) suggested that the right frontal effect may be obligatory in
nature and could be related to the ‘richness’ or amount of information that is retrieved.
They noted that this view is supported by similar right-frontal findings using picture
stimuli that attracted high levels of recognition accuracy (Schloerscheidt and Rugg
1997).
Observations by Rugg et al. (1996) provide further evidence that the parietal ERP
old/new effect in an associative recognition task reflects recollection-based retrieval.
They used an associative verbal task in which word pairs were presented in the study
phase, and subjects were required to incorporate each pair of words into a sentence. In
the test phase, the first word of each studied pair was intermixed with an equal number
of unstudied words. Subjects were required to make old/new recognition judgments, and
for each word judged old, they had to recall the specific word that it had been associated
with in the study phase. A strongly left-lateralized ERP old/new pattern was observed
only for words judged old for which the associated word was also correctly recalled.
The authors concluded that the findings support the view that the parietal ERP old/new
effect reflects neural activity associated with the recollection of specific past episodes,
and may therefore reflect medial temporal lobe involvement in episodic retrieval.
In the studies discussed above, context-based paradigms were primarily used to
investigate whether familiarity-based and recollection-based retrieval processes involve
the same or different neural systems. There are, however, other aspects of retrieval, such
as strategies used and effort involved, which Rugg et al. (2000) have termed retrieval set
effects. Rugg et al. studied retrieval strategy and effort using different encoding
conditions to examine the effect of depth of encoding on retrieval processes. In the
study phase of their task designed to promote only ‘shallow’ encoding, subjects had to
decide whether the first and last letters of the study words were in alphabetical order. In
the task designed to promote ‘deep’ encoding, subjects were required to incorporate
each word into a meaningful sentence.
In the interval between 300 and 800ms post stimulus onset, the ERP old/new effects
associated with the deep encoding task were most prominent over left parietal and right
frontal regions, and were therefore described as being qualitatively the same as those
49

Neural correlates of object recognition memory
observed in context-based studies. The only ERP old/new effects generated by the
shallow encoding task occurred during the 800 to 1400ms post stimulus interval and
were restricted to right frontal regions. Recognition accuracy for shallowly studied
words was significantly lower than for words studied in the deep encoding task.
Because other studies had reported a prominent early left parietal old/new effect
associated with recollective processes, its absence with the shallowly encoded words
was considered consistent with the reduced recognition accuracy for these. It was
suggested, however, that the right frontal ERP old/new effect for the shallow task was
inconsistent with this interpretation, given also its occurrence with context-based
retrieval and recollection. Rugg et al. argued that the right frontal effect could be
explained, however, by a necessity for more extensive monitoring for shallowly studied
items prior to decision; a theory also supported by the longer response times associated
with these items.
Retrieval set effects were investigated by comparing ERPs elicited by the new words in
both the shallow encoding and deep encoding test blocks. ERPs elicited by new words
presented in the shallow test block were found to be more positive than those in the
deep test block. This result suggests that different retrieval sets are adopted when
attempting to retrieve words that have been encoded using different linguistic attributes.
It was also suggested that the prominent differences observed at left frontal sites
reflected a greater retrieval effort. Rugg et al. likened this result to Ranganath and
Paller’s (1999) observation of a greater positivity elicited over left frontal regions by the
more difficult of two retrieval tasks.
Only few studies have used pictorial stimuli to investigate scalp-derived ERP correlates
of object recognition memory. For example, Schloerscheidt and Rugg (1997) used a
study-test design to compare ERPs elicited by the recognition of words and pictures of
objects. In the encoding phase, subjects were asked to imagine if each represented word
or picture item would, in real life, be larger than the computer monitor that they were
viewing. Clinical data (Smith 1989) had suggested that damage to the left medial
temporal lobe results in relatively greater impairment to verbal than to pictorial
memory, and conversely, damage to the right medial temporal lobe results in relatively
greater deficits in pictorial memory. Given this neuropsychological evidence, and the
50

Neural correlates of object recognition memory
possibility that the parietal ERP old/new effect reflects medial temporal lobe
involvement in recognition memory (Rugg et al. 1996), Schloerscheidt and Rugg (1997)
hypothesized that, in contrast to left lateralized parietal ERP old/new effect observed
with word stimuli, the retrieval of pictorial stimuli would be characterized by a more
bilateral distribution of the parietal old/new effect. However, both pictures and words
elicited similar early ERP old/new effects that were lateralized to left parietal sites. As
this result was contrary to their hypothesis, it was suggested that the parietal old/new
effect may not be reflecting medial temporal lobe function. In addition, a right frontal
ERP old/new effect occurred with the picture recognition but not with the word
recognition task. This effect was initially left-lateralized, with an onset 100ms prior to
that of the left parietal ERP old/new effect, and shifted to right frontal regions after 1s.
This right frontal finding is consistent with the proposal that pictures are more richly
encoded than words, and are therefore more likely to engage post-retrieval processes.
An early left frontal ERP old/new effect, which preceded the left parietal effect, was
previously reported by Tendolkar et al. (1997). Thus, Schloerscheidt and Rugg
concluded that the left parietal ERP old/new effect is not necessarily the earliest ERP
correlate of successful retrieval.
Paller and colleagues (Paller et al. 1999; Paller et al. 2000) also used a study-test format
to investigate ERP correlates of successful retrieval using photographs of faces. During
the encoding phase, subjects were shown photographs of faces that they were asked to
memorize. Accompanying some of the photographs was a short simulated voice of the
individuals shown stating who they were and what they did (‘remember faces’). Other
faces were presented without voices and others with instructions to forget the faces
(‘forget faces’). During the test phase, subjects were required to identify faces that were
shown previously (‘remember’ and ‘forget faces’), presented in amongst photographs of
new faces. The authors reported that ERPs elicited by ‘remember faces’ were
significantly different from those elicited by ‘forget faces.’ In a separate behavioural
study, the degree of priming for the two types of studied faces was found to have been
equal, although ‘remember faces’ were much more accurately recognized. Paller and
colleagues therefore concluded that differences between the ERPs elicited by the two
types of studied faces represents a neural correlate of recollection.
51

Neural correlates of object recognition memory
Ranganath and Paller (2000) hypothesized that frontal effects previously associated with
episodic retrieval might also reflect the recall from memory of perceptual detail of
studied items. They tested this hypothesis using a study-test design with two test phase
retrieval tasks: a general task in which subjects were required to make old/new
recognition judgements of previously studied line drawings, and a more specific task in
which an additional judgement was required, namely, whether the test phase drawings
were larger or smaller than those previously studied. Frontal ERPs were more positive
for the specific test than for the general test, and were largest within the 600 to 800ms
interval post stimulus onset. Because this result was significant for both new and old
items, the authors concluded that frontal activity not only reflects successful retrieval,
but also reflects evaluative processing of specific stimulus attributes retrieved from
memory. In addition, given the different requirements for the general and the more
difficult specific retrieval tests, the authors proposed that both left and right prefrontal
regions are engaged when retrieval demands and evaluation of perceptual detail are
relatively high. A left parietal positivity was also observed for the specific relative to the
general task, but only for old items. Ranganath and Paller likened this observation to the
left parietal ERP old/new effect, and therefore concluded that this pattern of activity
reflected reactivation of stored information.
A number of clinical studies utilizing the study-test design have provided further insight
into the brain structures which could be responsible for the generation and modulation
of late positive ERPs and the specific memory processes that they reflect. For example,
when the response to verbal stimuli was recorded using depth electrodes sited within the
temporal lobes of pre-surgery patients, N400- and P300-like components appeared to be
generated and modulated by old items within medial temporal lobe structures in the left
hemisphere only (Smith et al. 1986). It was suggested that the left-lateralized generation
and modulation of these potentials was consistent with the involvement of the left
medial temporal lobe in verbal memory. Johnson (1995) noted that the ERP components
recorded in Smith et al.’s study resembled those recorded at the scalp surface, but a
causal relationship was not demonstrated. A subsequent study added further weight to
the view that there is a relationship between the parietal ERP old/new effect and explicit
memory (Smith and Halgren 1989). Patients with intractable epilepsy who had
undergone left temporal lobectomy failed to show a late ERP old/new effect. This result
contrasted with findings from patients who had undergone right temporal lobectomy
52

Neural correlates of object recognition memory
and a group of control subjects. Both of these groups showed preserved ERPs.
Moreover, left temporal lobectomy patients showed impaired recognition accuracy
compared to the other groups. In the light of these results that indicate the importance of
the hippocampal formation in the generation of the ERP old/new effect, and in
conjunction with previous findings implicating the hippocampus in the formation of
explicit memories, Smith and Halgren concluded that the ERP old/new effect reflects
explicit recollective processes rather than implicit familiarity processes.
Guillem et al. (1995a) investigated the effects of temporal and extra-temporal (parietal
and frontal) epileptogenic lesions on recognition memory-related hippocampal activity
related to recognition memory. ERPs were elicited using verbalizable pictures, and were
recorded using depth electrodes sited within the hippocampal formation. The authors
observed ERP components with waveforms similar to those of the scalp-derived N400
and P600 potentials reported in previous studies. In addition, they recorded polarity
reversals within the hippocampus for both N400 and P600 components, a finding
consistent with other studies (eg. Heit et al. 1990; Puce et al. 1991; Smith et al. 1986).
Polarity reversals indicate local generation rather than volume-conducted activity
generated by distant sources. It was noted that the more extensive the damage, the
greater the effect on the N400 and the P600 components. These components were not
dramatically affected in patients with unilateral temporal lobe epilepsy. Moreover, the
N400 component was preserved both ipsilaterally and contralaterally to the seizure
focus. However, patients with multi-focal lesions showed greatly diminished N400 and
P600 components in both left and right hippocampi. These results were compared to
those reported by Puce et al. (1991), who reported absent N400/P600 components in
patients with large white matter lesions. The fact that the greatly diminished N400 and
P600 components recorded by Guillem et al. (1995a) were nevertheless identifiable,
was attributed to less extensive lesions in their multi-focal patients. Guillem et al.
concluded that N400 and P600 components are modulated by distributed, yet highly
interconnected, brain regions, including hippocampal, parietal and frontal regions. This
conclusion was also drawn by this group from the findings obtained using a continuous
recognition memory design (Guillem et al. 1995b) (see section 2.2.3.1).
Two other clinical studies, using continuous recognition rather than study-test formats,
are worthy of note as they contradict the study-test findings described above. In the first
53

Neural correlates of object recognition memory
of these studies, Rugg et al. (1991) used verbal stimuli in a continuous recognition task,
and reported reduced ERP old/new effects in patients who had undergone either left or
right temporal lobectomy. Furthermore, they found no relationship between the
magnitude of the ERP old/new effect and scores of verbal memory performance across
all subjects investigated. In contrast to Smith et al. (1986), Rugg et al. concluded that
the cognitive processes reflected by their ERP data were not lateralized to one
hemisphere, and moreover, because of the lack of correlation with verbal memory
scores, they are not necessary for normal verbal memory functioning. Furthermore, it
was suggested that while the temporal lobes appear to be involved in generating the
ERPs that show the old/new differences, the primary locus of these ERPs does not
appear to lie within medial temporal lobe structures (Rugg et al. 1991). This finding
contradicts those of a number of studies which indicated a hippocampal origin for the
ERP old/new effect (eg. Heit et al. 1990; Puce et al. 1991; Smith and Halgren 1989;
Smith et al. 1986). It is possible, however, that differences in experimental conditions
may account for these apparently contradictory findings (Johnson 1995). Firstly, it may
be that the recognition memory processes that are employed in a continuous recognition
task may differ from those employed in a study-test task. Secondly, the lag times, with 5
intervening items between repeated items, totaled 19s in Rugg et al.’s (1991) study.
These were considerably shorter than the 4-5 minutes usually used in a study-test
design. An insufficient delay, coupled with the much lower memory loads used by Rugg
et al., could explain their failure to find a decrement in performance associated with
their reduced ERP old/new effects.
In the second of these continuous recognition-based studies (described in section
2.2.3.1), intravenously administered scopolamine, known for its detrimental effects on
explicit memory, was used to mimic the effects of left temporal lobectomy in normal
subjects (Potter et al. 1992). Although the authors had predicted that the scopolamine
would reduce ERP old/new effects, they were in fact preserved, leaving the authors to
conclude that ERP old/new effects are not dependent on explicit memory processes.
This conclusion is contrary to those drawn in nearly all other studies that have used a
study-test task design. Johnson (1995) argued, however, that once again, factors such as
low memory loads and predictable short delays between repeated items might have
contributed to Potter et al.’s findings. Because increased reaction times were also
54

Neural correlates of object recognition memory
observed, Johnson also concluded that the scopolamine had affected attention as well as
memory processes.
In summary, the study-test appears a better task design than the continuous recognition
design for the study of neural processes involved in episodic retrieval. Additional
effects more specifically associated with episodic memory retrieval have also been
revealed using the study-test design. For example, context-based study-test designs have
shown that compared with the presentation of new words, correctly recognized old
words are associated with a prolonged scalp positivity that is most prominent over left
parietal regions between 400 and 1000ms post stimulus onset, and over right frontal
regions from 400ms to beyond 1400ms. The parietal ERP old/new effect has
consistently been associated with episodic memory retrieval processes. The magnitude
of the parietal ERP old/new effect appears to be larger when the amount or quality of
information retrieved increases, or when items are better remembered or recollected.
While the left parietal ERP old/new effect has always been seen in tests requiring
recollection, the right frontal ERP old/new effect, thought to reflect right frontal
activity, is dependent on the nature of the test and is not always observed. In tests
involving contextual retrieval, the frontal effect is usually present, although because of
its prolonged duration, this is thought to represent post-retrieval operations such as
verification and monitoring, rather than processes specifically related to retrieval
success.
Clinical studies have provided information about the origin of ERP effects based on
memory retrieval. Current consensus appears to be that the scalp ERP old/new effect
over parietal regions reflects the old/new effect observed with depth recordings within
the medial temporal lobes. Furthermore, it appears that intact medial temporal lobes are
required for the generation of the ERP old/new effect, lending additional support to the
view that these ERP changes reflect explicit memory processes. Parietal and frontal
regions have also been implicated in the modulation of the ERP old/new effect.
Evidence for this comes from two main sources: the reported diffuse scalp ERP old/new
effect over parietal and frontal areas, and depth recordings of the old/new effect within
these regions. It has also been proposed that the hippocampal formation, in conjunction
with parietal and frontal regions, forms a distributed, yet highly interconnected, network
that modulates ERP activity based on memory retrieval. A few studies have also
55

Neural correlates of object recognition memory
suggested that both left and right frontal regions are involved in retrieval strategy and
retrieval effort processes. In addition, it appears that both left and right prefrontal
regions are engaged when task demands increase and when specific attributes of the
stimuli are retrieved from memory. With respect to the ERP correlates of object
recognition memory, there are, as yet, very few picture-based ERP studies. Furthermore,
findings from these studies are confused by the use of either verbalizable or non-
verbalizable pictures and a mixture of scalp-derived and intracerebral data. Picture-
based scalp ERP old/new effects have been reported over frontal and left parietal
regions. Picture-based intracerebral ERP old/new effects have indicated the involvement
of extensive regions within the left and right medial temporal lobes and within parietal
and frontal regions. The most consistent finding overall appears to be the involvement
of frontal regions, and that compared with words, pictures elicit larger responses, which
has been attributed mainly to the greater the richness of the material.
2.2.3.3 Differentiation between transient item-related and sustained task-related
ERP correlates of episodic memory retrieval
Sections 2.2.3.1 and 2.2.3.2 were concerned with transient item-related ERPs
associated, in particular, with retrieval success and post retrieval monitoring processes.
These transient ERPs occur in response to the presentation of individual items. They last
only while items are being processed, and reflect specific properties of these items and
the operations that they engage. Sustained task-related effects, however, are largely
unaffected by individual items. They are initiated by task instructions and are sustained
for the duration of the task. They reflect the ‘mode’ or ‘state’ established by the task
requirements. In the case of episodic memory retrieval, the term ‘retrieval mode’
(Tulving 1983) has been used to describe the task-related neural activity that is
sustained throughout the retrieval task. Task-related activations are usually investigated
in PET and fMRI studies, in which task blocks rather than individual items are
compared (see section 2.2.4). However, a specific ERP technique known as direct-
coupled recording (DC) allows monitoring of sustained task-related activity (Rockstroh
et al. 1989). DC ERPs are obtained by setting a higher time constant for the high-pass
filter of the recording instrumentation, such that slower changes are passed rather than
filtered-out (Cabeza 1999). The low-pass filter is unchanged, however, thereby also
56

Neural correlates of object recognition memory
allowing the recording of standard transient perturbations. Thus, the DC recording
technique can be used to monitor sustained task-related activity similar to that
monitored using PET and fMRI methods, whilst concurrently monitoring faster, item-
related changes.
Duzel and his colleagues (Duzel et al. 1999; Duzel et al. 2001) are the only group so far
to utilize this technique in the study of episodic memory retrieval. Duzel et al. (1999)
applied both PET (see section 2.1.3) and ERP techniques to identify brain regions that
mediate task and item-related activity associated with episodic memory retrieval. All
subjects were tested using both techniques, although in separate sessions. A study-test
experimental design was used. During the study phase, words were encoded by making
pleasant/unpleasant judgements. During the retrieval phase, both episodic retrieval and
retrieval from semantic memory were examined in separate, randomly presented,
blocks. For the episodic retrieval blocks, subjects were required to make old versus new
judgements, and for the semantic retrieval blocks, subjects were required to make living
versus non-living judgements. Prior to each retrieval block, subjects were oriented to the
task requirements by instructions informing them of the type of judgements to be made.
PET data showed activation within the right prefrontal cortex (BA10) and right
posterior cingulate cortex (BA23) for the episodic retrieval task relative to the semantic
retrieval task. Comparison of the semantic retrieval task with the episodic retrieval task
yielded left prefrontal cortex (BA45/47) and left temporal lobe (BA21) activations.
Significant sustained mode-related ERPs were also observed. Compared with the
semantic retrieval task, the episodic retrieval task showed a sustained positive shift over
right frontopolar regions. It is not unreasonable to suppose that this is an
electrophysiological reflection of the BA10 activation observed using PET. A similar
correspondence with the left prefrontal cortex PET activation was not seen in the ERP
data; neither the semantic nor the episodic retrieval tasks showed sustained differences
in this region.
Duzel et al noted that these PET and ERP findings are consistent with the HERA
(Hemispheric Encoding/Retrieval Asymmetry) model (Nyberg et al. 1996a; Nyberg et
al. 1998a; Tulving et al. 1994a), in which left prefrontal regions are involved in
encoding and semantic retrieval, and right prefrontal regions in episodic retrieval. They
57

Neural correlates of object recognition memory
point out that their converging PET and ERP data further substantiates the model, and
furthermore note that the asymmetry of the HERA model is based on sustained task-
related rather than transient item-related activity.
Duzel et al. drew further parallels between their ERP data and other PET data (Buckner
et al. 1998), suggesting that the slightly sluggish time-course of the DC signal may
reflect the sluggish haemodynamic response. This sluggish response was attributed to a
task-related ‘neurocognitive inertia’ that reflects a strategic, intentional, orientation by
the subject from the present to the past. The authors suggested that the converging PET
and ERP data reflect task-related operations that, in part, characterize the episodic
retrieval mode, a state in which subjects consciously think back to the encoding episode.
The PET old minus new comparison showed activations within the left medial temporal
lobe. It was suggested that these indicated regions contributing to successful retrieval
processes. The ERP old minus new comparison was characterized by the widely
reported late positivity of the parietal ERP old/new difference, peaking at around 600ms
post onset, over left parietal regions. Source analysis indicated a medial temporal lobe
origin for the left parietal positivity. Duzel et al. therefore concluded that the medial
temporal lobe is involved in transient item-specific episodic retrieval processes.
2.3 Summary of neural correlates of object recognition memory
Converging evidence obtained using a wide range of experimental methods indicates
that an integrated network of neural regions is involved in object recognition memory
(ORM). Neuropathological and neuropsychological studies in humans and non-human
primates have shown that the medial temporal lobes are important in ORM. The
perirhinal cortex appears to play a particularly crucial role, while neural linkages from
medial temporal regions to frontal and other cortical regions are critical for normal
memory function.
Object recognition memory includes recognition based both on familiarity, and on the
actual recollection of the experience of seeing the object, or episodic memory. In
58

Neural correlates of object recognition memory
humans, ORM has been studied mainly by testing episodic memory using tests in which
objects are studied and identified some time later.
Specific neural regions, which are dependent on the nature and context of the stimuli,
are engaged for the different aspects of an episodic retrieval process. One aspect is the
maintenance of the mental state adopted for the performance of a memory retrieval task,
or retrieval mode. This is thought to involve sustained mental activity directing the
mental focus to the past. More transient neural processes are involved during the actual
moments the event of interest is remembered. Episodic retrieval also involves retrieval
set processes, such as retrieval strategy and retrieval effort. Retrieval strategy refers to
the way that items are recollected, and is based on the way that they were initially
encoded. Retrieval effort refers to the difficulty associated with recalling specific events
from memory.
Evidence from PET, fMRI, and electrophysiological DC-ERP studies suggests that a
mental state or ‘mode’ of processing is established and maintained for the duration of a
retrieval task. Activations within the right pre-frontal cortex and adjacent regions have
consistently been reported when episodic retrieval tasks are compared with non-episodic
retrieval tasks, indicating the involvement of these regions in episodic retrieval.
Electrophysiological techniques have been used to examine more transient processes
involved in episodic retrieval. These have been used to identify neural regions involved
at the actual time an episode is recalled. This recollection is referred to as retrieval
success. Retrieval success has been extensively investigated by examining differences
in the EEG or ERPs associated with the identification of previously studied old and
novel new items. A significant EEG finding is that theta activity is more synchronous
during the identification of old items than new items. This is thought to reflect
hippocampal activity transmitted to the cortex via hippocampo-cortical feedback loops.
The ERP literature has consistently reported old/new differences over left parietal
regions, and also, depending on the nature of the retrieval task, over right frontal
regions. Parietal old/new differences occur regardless of whether recognition is based
on familiarity or on recollection. On the other hand, right frontal differences occur more
specifically with recollection processes. Depth electrode findings suggest that the
parietal scalp ERPs originate within the medial temporal lobes. Similarly, the scalp
59

Neural correlates of object recognition memory
ERPs over right frontal regions are thought to reflect activity in the underlying right
frontal cortex. It is thought that interaction between the medial temporal lobes and
frontal regions enables the recognition of the object to be linked to the context of
experiencing it.
The majority of both EEG and ERP studies have utilized verbal stimuli. Most studies
using pictorial stimuli have used pictures of common objects, and have reported larger
ERP responses than those obtained using word stimuli. This is generally attributed to
the greater richness of pictorial stimuli. This conclusion should, however, be
approached with caution given the paucity of studies using pictorial stimuli.
Other aspects of episodic retrieval, such as effort and strategy, termed retrieval set
effects, have also been investigated, revealing, in particular, frontal lobe involvement in
these functions. With respect to strategy, transient item-related retrieval effects are
influenced by the tonically maintained mental set adopted for a particular strategy. The
strategy adopted for retrieval may depend on the encoding strategy that had been used.
As yet, however, investigations into retrieval strategy are preliminary, and findings are
inconclusive. There has been lack of agreement over the frontal lateralization of activity
associated with retrieval effort. It has been suggested that distinct neural structures may
not be devoted to retrieval effort processes. Instead, ‘the neural correlates of increasing
effort will be manifest as increased activity of whatever brain regions are engaged by
the retrieval task in question’ (Rugg and Wilding 2000, p.114).
In conclusion therefore, lesions studies in humans and non-human primates have
identified the medial temporal and frontal lobes as the gross neural regions important in
ORM. In normally functioning humans, ORM has been investigated using tests of
episodic memory retrieval. To date, retrieval mode and retrieval success processes have,
with only one exception (Duzel et al. 1999), been investigated independently using
different techniques. The simultaneous investigation of these processes should help
further clarify the issue of whether sustained task and transient item-related frontal
activity is generated in the same neural regions. Furthermore, retrieval mode and
retrieval success findings have, in the main, been derived from studies that have used
verbal stimuli. Due to the paucity of studies that have used object stimuli, it is not clear
whether the observed patterns of neural activity are specific for verbal stimuli, or
60

Neural correlates of object recognition memory
whether these patterns are characteristic of retrieval memory in general. The present
study has attempted to add to the understanding of these issues.
61

Chapter 3 Steady-State Probe Topography
The following chapter will explain how the Steady-State Probe Topography (SSPT)
technique can be used to investigate neural activity associated with cognitive processes.
The main features of this technique will be explained, and its advantages and limitations
in relation to other neural imaging methods such as fMRI, PET and electrophysiological
techniques will be discussed. Firstly, a definition of steady-state evoked potentials and
an overview of early attempts to use steady-state evoked potentials in the study of
cognitive processes are presented. This overview is followed by a description of the
SSPT technique, first described by Silberstein et al. (1990b). Findings from a number of
studies utilizing the SSPT technique will then be summarized to provide the reader with
an appreciation of the potential of this technique. Following this synopsis of the
literature, hypotheses specific to this study dealing with object recognition memory will
be formulated.
3.1 Steady-state evoked potentials compared with transient evoked potentials
To compare and contrast transient and steady-state evoked potentials, Regan (1977a)
used an electrical engineering analogy where the brain is compared to a circuit which
receives, for example, a stepped or pulsed electrical input. The system gives an initial
response to the input, which dies away quickly, the so-called transient response. This
transient response can be compared to the traditional ERP, which is a transient electrical
potential generated by the brain in response to some stimulus (see Chapter 2).
Alternatively, the same system can be driven by a long train of rapidly repeating stimuli.
Once the system has settled, the observed response contains the same sequence of
waveforms as the input. Steady-state potentials are unlike transient evoked potentials in
that the response to one stimulus has not died away before the next stimulus is
delivered.
Transient evoked potentials, where individual peaks are well defined, are usually
described in the time domain. Conversely, the repetitive nature of a steady-state evoked

Steady-State Probe Topography
potential is more easily represented in the frequency domain, where it can be
characterized as a number of discrete frequency ‘bands’ superimposed on the
background EEG frequency spectrum. Fourier techniques are usually applied to
calculate the amplitude and phase components of a steady-state evoked potential. These
are usually calculated for the frequency of stimulation or for multiples of this frequency
(see Regan 1977b).
In a linear system, the information contained in a transient evoked potential and a
steady-state evoked potential is exactly equivalent (Regan 1977b). However, in non-
linear systems, such as the human central nervous system, steady-state evoked
potentials can provide information to complement information derived from transient
evoked potentials. One example of the clinical utility of SSVEPs is in the diagnosis of
multiple sclerosis (Regan et al. 1977). Regan et al. (1977) reported that in multiple
sclerosis, potentials evoked by a medium-frequency (13-25Hz) flicker were delayed,
whereas high-frequency (40-60Hz) evoked potentials were not. Furthermore, SSVEPs
for patterned and unpatterned stimuli were shown to be sensitive to retrobulbar neuritis
(Regan et al. 1977), whereas visually evoked potentials (VEPs) in response to a
transient flash gave no reliable indication of the disorder (Halliday 1977).
Regan (1989) described a number of advantages in using steady-state evoked potentials
over transient evoked potentials. One advantage is that it is easier to quantify steady-
state evoked potentials as these can be defined by a small number of discrete frequency
components, each consisting of two numbers, the amplitude and the phase. This reduced
set of information means that recording times can be significantly shorter than those
required for transient ERPs. This can be particularly advantageous in clinical situations
(Regan 1976). Moreover, the quantification of transient evoked potentials can be
difficult, especially when the waveform is atypical. For example, experimenters may
adopt different techniques to resolve overlapping peaks and to identify baseline levels.
In contrast, the quantification of steady-state evoked potentials ‘has a firm mathematical
basis; the quantification can be repeated in any evoked potential laboratory, and close
agreement between different experimenters is to be expected’ (Regan 1989). Regan
(1989) also noted that the recording of steady-state evoked potentials is easier as mains
interference is separated in the frequency domain, whereas in the time domain, it
overlaps with the transient evoked potential.
63

Steady-State Probe Topography
3.2 Steady-state evoked potentials in the study of cognitive processes
In contrast to the well-documented ERP changes associated with cognition, Regan
failed to show evidence for cognition-related changes in steady-state evoked potentials.
Regan indicated that ‘steady-state VEPs do not seem much affected by attention, though
this is a well documented effect for transient VEPs’ (Regan 1977a). Furthermore he
suggested that ‘if you wish to study, eg., attention or the orienting response, then steady-
state evoked potentials are most likely completely useless, so that you must use transient
EPs’ (Regan 1977b).
The first study to show a link between steady-state evoked potentials and human
cognitive activity was performed by Wilson and O’Donnell (1986). They found a
statistically significant correlation between the behavioural scores in a Sternberg
memory scanning task and the apparent latency of the SSVEP. SSVEPs generated by
stimulation in the medium frequency range (15-23Hz) were correlated with the slope of
the graph of response speed versus apparent latency. Those generated in the high
frequency range (40-59Hz) were related to the response speed intercept for zero items.
These correlations indicated that the SSVEP latency was related to the speed of
cognitive processing. In a subsequent study by this group (Wilson and O'Donnell 1988),
the relationship between cognitive processes and the SSVEP was investigated by
examining changes in the SSVEP during the performance of a mental workload task.
However, no significant correlation was found between these variables.
Steady-state evoked amplitude and phase components have been assumed to be stable
over time, and have traditionally been evaluated over long time intervals (Regan 1977a).
Regan (1989) explained that, in principle, a steady-state evoked potential is a repetitive
evoked potential ‘whose constituent discrete frequency components remain constant in
amplitude and phase over an infinitely long time period.’ In one study, SSVEP
amplitude and phase components for each stimulus frequency were evaluated over a 1
minute period (Regan 1976), and in an earlier study, fluctuations in the SSVEP were
evaluated over a 7 minute time period, with the filter time constant set to 7s (Regan
1977b). The author reported that over the 7 minute period, the SSVEP latency varied by
1 percent and the SSVEP amplitude varied by 7 percent. When a 1 minute evaluation
period was used, much more variability was observed. This was considered the result of
64

Steady-State Probe Topography
the wider bandwidth associated with the shorter evaluation period allowing more
background EEG noise through.
Galambos and Makeig (1985) suggested that the variability in steady-state evoked
potentials, which Regan attributed to background EEG noise, might be related to
cognitive processes. They attempted to find a relationship between ‘shifts in arousal’
and the steady-state auditory evoked potential (Galambos and Makeig 1988) but were
unsuccessful. Linden et al. (1987), who examined the effect of selective attention on the
auditory steady-state evoked potential, were similarly unsuccessful in demonstrating a
relationship between the steady-state evoked potential and cognitive processes despite
observing significant changes in the late components of the transient ERP during
selective attention.
Silberstein et al. (1990b) suggested factors that may have accounted for the apparent
insensitivity of steady-state evoked potentials to cognitive processes in the study of
Wilson and O’Donnell (1986) and other early studies. They suggested that cognitive
effects might have been reflected in the SSVEP amplitude rather than in the apparent
latency of the SSVEP. Furthermore, only central, occipital and parietal sites had been
investigated, limiting the possibility of observing cognition-related changes at other
sites. Using a technique subsequently termed Steady-State Probe Topography (SSPT),
Silberstein et al. (1990b) demonstrated, for the first time, a correlation between the
amplitude of the SSVEP and visual vigilance. The SSPT technique, together with other
more recent findings of SSVEP correlates of human cognitive processes, will be
outlined in the next section.
3.3 Steady-State Probe Topography (SSPT)
The SSPT technique described by Silberstein et al. (1990b) incorporated three main
features designed to maximize the possibility of observing cognition-related changes in
the SSVEP. These were (i) the Probe-ERP approach, in which an irrelevant steady-state
visual stimulus is presented in conjunction with the presentation of the cognitive task
under investigation, (ii) a high density of scalp recording sites, which should reveal
localized SSVEP topographic changes, and (iii) an analysis technique whereby the
65

Steady-State Probe Topography
SSVEP could be calculated in as little as 1s, allowing cognitive processes to be
continuously monitored over time. Each of these three features, as explained by
Silberstein et al. (1990b), will be described in turn.
‘The Probe-ERP technique was employed wherein the ERP stimulus was presented in a
manner as to be distinct from and irrelevant to the cognitive task undertaken by the
subjects.’ (Silberstein et al. 1990b, p. 338)
This technique was based on the ‘probe paradigm,’ first described by Galin and Ellis
(1975) and reviewed comprehensively by Papanicolaou and Johnstone (1984). Galin
and Ellis (1975) presented task-irrelevant visual stimuli at intervals of 3s while subjects
performed block design and writing tasks. They observed a probe-ERP asymmetry that
was task dependent, and was described as being similar to the task-related asymmetry of
EEG alpha power distribution. Shucard et al. (1977) later used auditory evoked
potentials (AEPs) as a probe response to investigate the possible lateralization of brain
function during the performance of various cognitive tasks. They found a hemispheric
asymmetry in the amplitude of the AEP that was related to the subject’s mode of
cognitive processing.
The premise underlying the Probe-ERP technique is that increases in regional cortical
activity associated with cognitive processing of the task will result in smaller potentials
evoked by an irrelevant probe stimulus (Papanicolaou and Johnstone 1984; Silberstein
et al. 1990b). The Probe-ERP premise is supported by evidence of an increase in
regional cerebral bloodflow accompanied by a reduction in the Probe-ERP amplitude
(Papanicolaou et al. 1987; Papanicolaou and Johnstone 1984). Papanicolaou and
Johnstone (1984) proposed a number of models which might explain the mechanism by
which the probe ERP is attenuated. They suggested that a ‘limited resource’ model best
accounted for the observed changes. This model assumes that cortical regions have a
limited capacity to process multiple inputs, and therefore are less responsive to the
probe stimulus when simultaneously engaged in the task of interest.
More recently, however, Nield et al. (1998) questioned the ‘limited resource model’
when reductions in the 13Hz SSVEP amplitude were found to occur simultaneously
66

Steady-State Probe Topography
with increases in the evoked 40Hz SSVEP amplitude during a heightened attentional
state.
Studies using the Probe-ERP technique in a variety of experimental situations have been
reviewed extensively by Papanicolaou and Johnstone (1984) and by Silberstein et al.
(1990b). Experimental findings include: left hemisphere attenuation of an auditory
probe ERP in a covert articulation task (Papanicolaou et al. 1983), right parietal
attenuation of a visual probe ERP in a visuo-spatial task requiring mental rotation of
geometrical figures (Johnstone et al. 1984) and differentiation between dyslexic and
control children on the basis of the amplitude of the visual probe ERP when the
demands in a reading task were increased (Johnstone et al. 1984).
Silberstein and colleagues (Silberstein et al. 1995a; Silberstein et al. 1990b) employed a
continuous visual flicker as their probe stimulus. The SSVEP produced by this stimulus
was characterized by reductions in amplitude during a period of heightened visual
vigilance (see section 3.5.1). Silberstein et al. (1995a) attributed reductions in the
amplitude of the SSVEP to increased regional brain activity. They proposed that the
amplitude reductions observed were akin to the phenomenon of event-related
desynchronisation (ERD) described by Pfurtscheller and Aranibar (1977a) and
Pfurtscheller and Klimesch (1990), in which regional increases in cortical activity are
indexed by transient reductions in alpha amplitude.
More recently, however, Silberstein (1998) reported SSVEP amplitude increases
occurring with certain types of cognitive activity. Transient increases in the amplitude
of the 13Hz SSVEP were found during the performance of a spatial working memory
task. It was proposed that with certain types of cognitive activity, specific linkages
between brain regions and between cortical layers are established. In the case of a
spatial working memory task, information is ‘reticulated between reciprocally related
neocortical regions forming reentrant loops’ (p. 34) resulting in an increased SSVEP
amplitude. Silberstein (1998) speculated that this may be the mechanism for holding
information actively or ‘on-line.’
The aforementioned interpretations of SSVEP amplitude reductions and increases were
reinforced in a recent study. Silberstein et al. (2001) reported that the ‘intake’ and ‘hold’
67

Steady-State Probe Topography
phases in an object working memory task were associated with SSVEP amplitude
decreases and increases respectively. During the ‘intake,’ or encoding phase of the task,
SSVEP amplitude reductions were observed at a number of sites, including the right
occipito-parietal region. The authors noted that reductions during the intake, or
perceptual, phase of the task are consistent with previous studies by this group in which
amplitude reductions were observed over occipito-parietal regions during a visual
vigilance task (Silberstein et al. 1990a). During the ‘hold’ phase of the task, subjects
held the encoded representation of the object(s) in working memory. Increases in
SSVEP amplitude associated with the ‘hold’ phase were likened to those reported in the
aforementioned study (Silberstein 1998) involving spatial working memory. That is,
linkages between brain regions were established, and information was reticulated
around re-entrant loops.
The phase of the SSVEP for the task of interest may be in advance of, or lag behind, the
phase for some other task, generally a reference or control task. Phase advances or lags
are also referred to as latency reductions or increases respectively. Silberstein et al.
(1996) observed that faster response times in the performance of a continuous
performance task were associated with larger frontal SSVEP latency reductions. It was
proposed that latency reductions are a manifestation of transient increases in neural
information processing speed reflecting increased coupling strength between neural
populations. More recently, Silberstein et al (2000) speculated that SSVEP latency
reductions might index excitatory processes. This suggestion was based on the fact that,
unlike normal controls, schizophrenic patients did not show prefrontal latency
reductions during a continuous performance task.
‘Brain electrical activity was recorded from 64 scalp sites within the area defined by
the International 10-20 system.’ (Silberstein et al. 1990b, p. 338)
The use of 64 scalp recording sites achieves a spatial resolution adequate for observing
effects over gross brain regions. Silberstein et al. (1990b) quote a number of studies
where the recommended electrode separation appears to be in the order of a few
centimetres. The SSPT technique is concordant with this recommendation, using the
traditional 10-20 International recording sites with additional electrodes between them
(see section 4.5 for more details).
68

Steady-State Probe Topography
‘A Fourier analyser with a 10 second integration period was used to determine SSVEP
magnitude.’ (Silberstein et al. 1990b, p. 338)
In this situation, variations in the SSVEP amplitude over time were shown to be related
to cognitive changes, rather than to background EEG noise as suggested by Regan
(1977b).
Silberstein et al. (1990b) used a temporal resolution of 10s, although in a more recent
study it was suggested ‘that it is possible to estimate the amplitude and phase of the
SSVEP using as little as 1 to 5s of recorded activity’ (Silberstein et al. 1995a). In
theory, using a 13Hz probe stimulus, the SSVEP can be calculated using as little as
1/13s (77ms) of data; however, the level of background noise would be too high to
allow any meaningful interpretation of the result. As with traditional transient ERP
methods, averaging techniques can be applied to overcome this signal-to-noise problem.
(Silberstein et al. 1995a). In the present study, a temporal resolution of 0.87s was used.
Reliable results were obtained at this resolution by averaging data for only 5 stimulus
items. Extrapolating this result to 20 or more averages could potentially yield reliable
results with a greater than 4-fold improvement in temporal resolution (ie. <200ms). In a
very recent study by this group, reliable results were in fact obtained using a temporal
resolution of 180ms (Harris et al. 2001).
The ability to tailor the evaluation period of the SSVEP allows one to focus on the
different cognitive processes that occur during a particular cognitive activity, for
example, visual vigilance, whether occurring over a period of a few hundred
milliseconds, seconds, minutes, or even hours. Silberstein et al. (1995a) argued that, in
contrast to techniques such as positron emission tomography (PET) which have a poor
temporal resolution (typically 60s), SSPT possesses the ‘temporal resolution as well as
the temporal continuity’ required to monitor relatively rapid cognitive processes.
Although not possessing the temporal resolution of ERP techniques, SSPT allows
investigation of cognitive processes over a range of time scales. For example, a short
evaluation period allows the monitoring of the more rapidly changing patterns of neural
activity associated with the processing of individual stimuli. Conversely, an evaluation
69

Steady-State Probe Topography
period the length of the task being investigated allows more sustained, state related,
neural activity to be evaluated.
3.4 SSPT and recording artifacts
Steady-state evoked potentials are, in general, more resilient to artifact than are transient
evoked potentials (see Regan 1989). If an adequate number of stimulus cycles is
averaged, non-probe related signal, such as background EEG and recording artifact, can
be greatly reduced. A greater number of averages effectually reduces bandwidth, and
therefore increases the signal to noise ratio. Regan (1977b) assessed the SSVEP over
relatively long time periods of several minutes, and thereby achieved a high signal to
noise ratio. Silberstein et al. (1990b), however, were interested in cognitive changes
occurring over a much shorter time period, and therefore far fewer stimulus cycles were
used when processing the SSVEP. Nevertheless, Silberstein et al. (1993) found that with
an integration period of only a few seconds, artifact such as EMG, EOG, eye-blinks, and
50Hz mains interference (see Figure 3.1) had very little effect on the SSVEP. Varying
amounts of known artifact were added to the EEG before the SSVEP was calculated.
Where the variances of the artifact and the EEG signal were equal, giving a much
poorer signal to noise ratio than normally experienced in practice, the resultant SSVEP
was very similar to the SSVEP with no added artifact. The source of artifact with the
greatest effect on the SSVEP, although still small, appeared to be EMG. The 50Hz
mains artifact had virtually no effect. This is understandable given that mains
interference is concentrated in a very narrow frequency band far removed from the
13Hz SSVEP.
3.5 Overview of investigations utilizing the SSPT technique
To date, the SSPT technique has been used to investigate various facets of cognition,
such as visual vigilance, attention, learning, planning and working memory. The
technique has also been employed in clinical contexts, for example, with children
diagnosed with Attention Deficit Hyperactivity Disorder (ADHD). A selection of
70
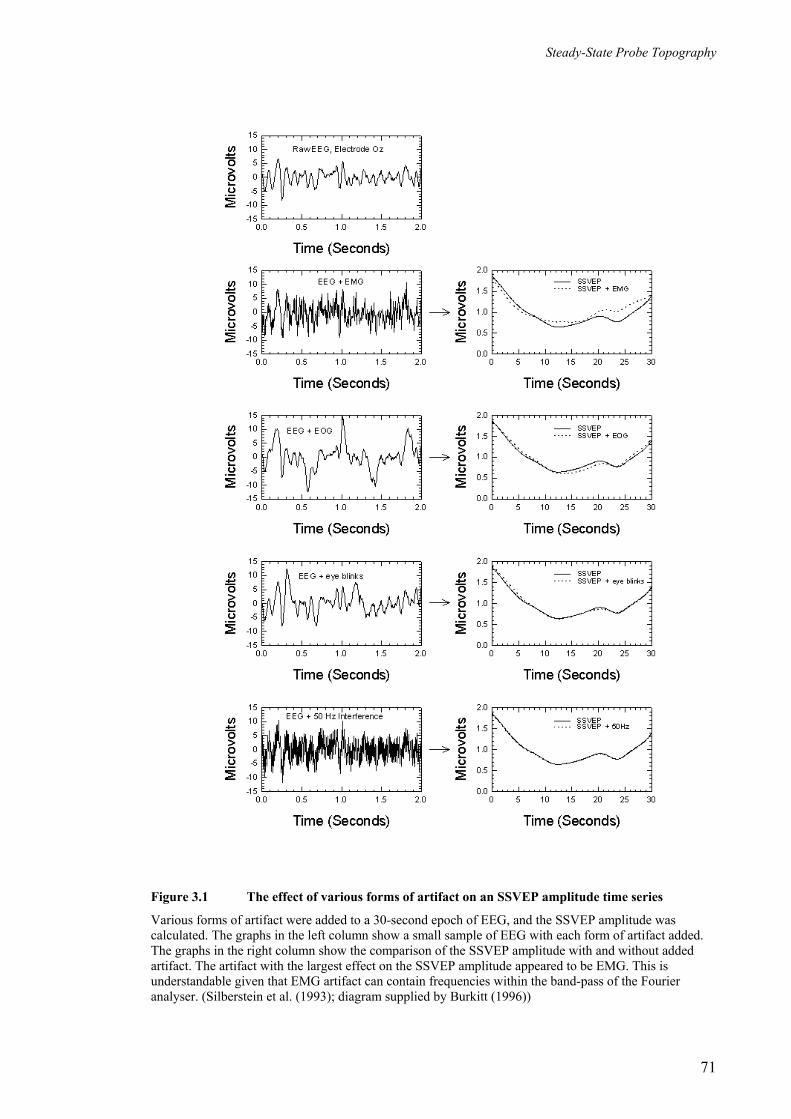
Steady-State Probe Topography
Figure 3.1 The effect of various forms of artifact on an SSVEP amplitude time series
Various forms of artifact were added to a 30-second epoch of EEG, and the SSVEP amplitude was calculated. The graphs in the left column show a small sample of EEG with each form of artifact added. The graphs in the right column show the comparison of the SSVEP amplitude with and without added artifact. The artifact with the largest effect on the SSVEP amplitude appeared to be EMG. This is understandable given that EMG artifact can contain frequencies within the band-pass of the Fourier analyser. (Silberstein et al. (1993); diagram supplied by Burkitt (1996))
71

Steady-State Probe Topography
studies will be briefly reviewed to help provide insight into the use of the technique and
to indicate the potential of the technique in the area of object recognition memory.
3.5.1 Visual vigilance
In Silberstein et al.’s (1990b) study, where a correlation between visual vigilance and
the amplitude of the SSVEP was observed, different phases of the task elicited different
SSVEP response patterns. Fourteen right-handed subjects were instructed to fixate on
the centre of a monitor and observe a series of shapes that would appear there. The total
task was divided into three parts. Each part, lasting 3 minutes, consisted of watching a
number of sequentially presented squares for 1 minute, followed by a number of
sequentially presented circles for 1 minute, and then sequentially presented squares
again for 1 minute. This sequence of shapes was presented twice, and subjects had
simply to watch throughout. Prior to commencement of the third part, subjects were told
that one of the circles had been modified and were required to identify the modification.
Subjects were naïve as to the nature of the modification and where in the sequence of
circles it would occur. A small monetary reward was offered for the correct
identification of the modification. This reward was intended to serve two purposes:
firstly, to encourage optimum performance of this part of the task, and secondly and
more importantly, to increase the level of attention during this third part in relation to
the second part. The first part served as a practice task to eliminate any novelty involved
with the recording environment and the performance of the task.
A temporal resolution of 10s was selected for the calculation of the SSVEP amplitude
and phase, as the authors were interested in monitoring slow attentional shifts associated
with changes in visual vigilance during performance of the task. The SSVEP elicited
during the presentation of the circles in the third part and the second part were
compared.
Following the presentation of the first circle, the cue for the possible appearance of the
modification, an attenuation of the SSVEP amplitude occurred at centro-parietal sites.
At this point, subjects would have been anticipating the appearance of the modified
circle. The identification of the modification, which was made to the last circle in the 1
72

Steady-State Probe Topography
minute period, was associated with an SSVEP amplitude attenuation over the occipito-
parietal and prefrontal regions.
According to the Probe paradigm, these localized reductions in SSVEP amplitude could
have reflected increases in regional cortical activity. Silberstein et al. noted that this
interpretation is consistent with neuropsychological findings, cerebral blood flow and
metabolism findings, findings from lesion studies, and electrophysiological findings.
This demonstration of a relationship between the SSVEP amplitude and cognitive
processes indicated that ‘SSPT warrants further investigation as an indicator of regional
brain activity associated with cognitive processes.’
3.5.2 Planning - Wisconsin Card Sorting Task
Using a much shorter temporal resolution of only 0.77s with the SSPT technique,
Silberstein et al. (1995a) observed more transient effects which occurred during the
performance of the Wisconsin Card Sorting (WCS) task. The WCS task is a well-
documented neuropsychological test thought to engage the prefrontal lobes (Milner
1963). Subjects are required to determine the criterion for sorting cards into four
categories. Once they have determined the sort criterion, subjects perform successful
sorts according to that criterion. After 6 to 10 successful sorts, the criterion is changed
and the process is repeated. At the point following the change in sort criterion, subjects
must dispense with the previously determined method of sorting and determine the new
one, a process which is completed in a few seconds. This period of ‘planning’ the next
move is known to place demands on the prefrontal cortex (Milner 1963; Milner 1964).
Silberstein et al. (1995a) observed a transient SSVEP amplitude reduction and a
simultaneous phase advance over prefrontal and right temporal regions at the time
subjects had to determine the new criterion for sorting. These SSVEP changes were
attributed to neural activation. As noted by the authors, previous techniques used to
examine prefrontal involvement during performance of the WCS task had lacked the
temporal resolution and sensitivity to detect cognitive processes occurring during this
short time period.
73

Steady-State Probe Topography
3.5.3 Attention - Continuous Performance Task
Transient SSVEP effects have also been observed during the performance of a
continuous performance task (CPT) (Silberstein et al. 1996). In this study, young adult
subjects viewed a series of letters on a monitor, presented at a rate of one every 2s. Prior
to the commencement of the task, subjects were instructed to identify and respond to the
appearance of the letter ‘X’ only if preceded by the letter ‘A’. The time between the
letter ‘A’ and ‘X’ is the point of greatest visual attention, and it was hypothesized that
this interval would be associated with reductions in SSVEP amplitude at scalp sites
involved in visual attention.
Again using the SSPT technique with a temporal resolution of 0.77s, SSVEP amplitude
attenuation and decreased latency were observed at parietal sites during the ‘A-X’
interval. These changes were absent during an equivalent interval where an ‘X’ was
preceded by a letter other than an ‘A.’ Another point of interest in the task was the
correct identification of the target ‘X.’ This event was associated with transient
reductions in the SSVEP amplitude and latency, interpreted as reflecting excitatory
processes, over central and frontal areas. The authors noted that this interpretation is in
agreement with other studies suggesting that these regions are involved with response
selection and execution. It was also noted that faster response times to targets were
associated with increased latency reductions frontally, suggesting that SSVEP latency
reductions index the speed of information processing, and therefore the extent of
coupling between cortical regions.
3.5.4 Clinical application of SSPT - ADHD
Several studies suggest that the SSPT technique may be applied to clinical situations.
For example, when children diagnosed with ADHD performed the continuous
performance ‘AX’ task described above, there was no attenuation in SSVEP amplitude
in the interval between the ‘A’ and the ‘X’ (Farrow et al. 1996). In contrast, young
control subjects showed a sustained parietal SSVEP amplitude attenuation similar to
that found in the young adults. The authors suggested that the lack of SSVEP
74

Steady-State Probe Topography
attenuation in the ADHD group was due to an inability to sustain attention during the
‘A-X’ interval.
Latency effects were examined by Silberstein et al. (1998), who employed a similar task
with boys diagnosed with ADHD and normal controls. In this study, only correct trials
were considered. Transient latency reductions were observed over prefrontal regions in
the normal controls following the presentation of the letters ‘A’ and ‘X’. However, the
boys with ADHD failed to show these latency reductions. It was thus concluded that the
absence of a latency reduction in ADHD subjects could have been due to inadequate
excitation of prefrontal networks.
3.5.5 Spatial working memory
In contrast to the amplitude reductions associated with heightened attention described in
previous studies, Silberstein et al. (1998) found amplitude increases associated with
cognitive effort in a spatial working memory task. They proposed that it is the nature of
the mental activity that determines whether the SSVEP amplitude increases or
decreases. In this study, subjects were instructed to hold in memory the position of 3
dots located on an imaginary circle. During the 3s hold period, the 13Hz SSVEP
amplitude showed a pronounced increase at frontal and parietal sites when compared
with that for a control task matched for perceptual and motor aspects. The authors
proposed that the increase in SSVEP amplitude in regions thought to be involved in the
task reflected the reticulation of information between ‘reciprocally related neocortical
regions forming re-entrant loops.’ (p. 34) This finding was consistent with a number of
studies where alpha activity was enhanced during certain types of cognitive task. Ray
and Cole (1985), for example, reported that during the performance of a ‘rejection’ task
where subjects had to attend to mental imagery, alpha activity was significantly
enhanced. A similar enhancement was also reported in a study using
magnetoencephalography (MEG) (Tesche et al. 1995) where subjects had to visualize a
set of imaginary movements. In another study that investigated the effect of short-term
memory on the EEG (Krause et al. 1996), enhanced alpha activity was observed when
75

Steady-State Probe Topography
subjects had to determine whether a vowel had been presented previously as a target to
be memorized.
3.6 Conclusions
Findings from the SSPT studies outlined above indicate that the SSPT technique
possesses both the temporal resolution and the temporal continuity necessary for the
investigation of cognitive processes. Moreover, the ability to vary the SSVEP
evaluation period allows investigation of cognitive processes occurring over different
time scales. The spatial resolution of SSPT using 64 electrodes, although not as good as
that obtained using PET and fMRI methods, appears satisfactory for investigating neural
activity associated with gross brain regions. Furthermore, the use of SSPT in the study
of attentional and memory processes has produced results that show the value of this
technique in monitoring neural activity associated with cognitive processes, and thus
warrants further investigation.
3.7 Formulation of hypotheses for the present investigation
The main aim of this study is to use SSPT to investigate both sustained, task-related,
and transient, item-related, neural activity associated with episodic retrieval memory.
The first hypothesis is concerned with sustained task-related effects associated with the
maintenance of a retrieval mode. The second hypothesis relates to transient effects
associated with successful retrieval from memory.
The right prefrontal region has been implicated in the establishment and maintenance of
the mental set for episodic retrieval, also referred to as retrieval mode. As explained in
Chapter 2, PET and fMRI data have consistently indicated that the prefrontal cortex
(BA10) and adjacent areas are involved in maintaining a retrieval mode. In a study
utilizing both DC-ERP recordings and PET techniques, a sustained ERP positive shift
was observed over right frontopolar regions during an episodic retrieval task, but not
during a semantic retrieval task (Duzel et al. 1999). PET data from the same study
revealed activation of the right anterior prefrontal cortex (BA10) for the episodic task
76

Steady-State Probe Topography
only. The authors suggested that the DC-ERP data represents the electrophysiological
correlate of the PET activations, and therefore concluded that episodic retrieval mode is
manifest as a tonically maintained, item-independent state. Thus, converging evidence
obtained using different techniques has indicated the involvement of right frontal
regions in the maintenance of a retrieval mode. One of the main aims of this study was
to investigate the topography of sustained task-related SSVEP effects associated with
episodic memory retrieval. This was done by comparing the average of the SSVEP
amplitude and latency values obtained over the entire duration of an episodic retrieval
task with those for a non-episodic retrieval baseline task.
It is hypothesized that retrieval mode processes engaged during the performance of an
episodic retrieval task will be reflected in sustained SSVEP amplitude and latency
effects over right frontal regions.
Transient neural activity associated with successful retrieval processes has been
investigated using EEG and ERP electrophysiological techniques (see Chapter 2), albeit
with a relatively low spatial resolution. ERP studies, in particular, have consistently
shown that ERPs associated with the recognition of previously studied items, compared
with those associated with the correct identification of unstudied items (ERP old/new
effect), show a transient positivity over parietal regions between 400 and 1000ms after
item presentation. This positivity is generally most prominent over left parietal regions.
However, nearly all such studies have utilized verbal stimuli, and so this apparent
asymmetry could be due to the nature of the stimuli. Transient ERP old/new effects
have also been observed over right frontal regions between approximately 400 and
1400ms post stimulus onset. While the parietal ERP old/new effect appears to be
observed consistently during successful retrieval, the right frontal ERP old/new effect
appears to depend on the nature of the task and is not always observed. Right frontal
ERP old/new effects have been associated with post-retrieval monitoring processes, and
are mainly observed when recognition involves recollection of the study context.
The investigation of transient effects in this study will focus on the topography of
SSVEP amplitude and latency changes associated with the retrieval from memory of
abstract, two-dimensional objects, and how these spatio-temporal patterns vary with
increases in memory load. It is anticipated that increases in memory load will result in
77

Steady-State Probe Topography
increased utilization of those neural regions involved in retrieval processes leading to
recognition of previously studied objects.
It is hypothesized that increases in memory load during successful retrieval will be
associated with transient graded SSVEP amplitude and latency changes over bilateral
parietal and right frontal regions.
78

Chapter 4 Methods
4.1 Introduction
Neural activity associated with episodic memory retrieval can be looked at from a
number of different viewpoints. For example, sustained activity associated with the
maintenance of a mental state during the course of a retrieval task has been discussed in
terms of retrieval mode (Tulving 1983). However, the performance of a retrieval task is
also associated with rapidly changing patterns of neural activity associated with more
transient processes, for example, the initial perception of a stimulus, recognition of the
stimulus, deciding whether the stimulus is a target, a motor response, and also post-
retrieval processes associated with monitoring and recollection. Steady-State Probe
Topography (SSPT) appears to be an ideal technique to study episodic memory retrieval
as the length of the evaluation period over which the SSVEP components are calculated
can be chosen to suit the time-course of cognitive changes being investigated. A long
evaluation period is appropriate for the investigation of modal effects, while a short
evaluation period is appropriate for the investigation of transient effects that occur much
more rapidly.
This chapter describes the experimental methods employed to test the hypotheses
presented in Chapter 3. Section 4.2 deals with the experimental design used to
investigate object recognition memory. It will include a description of the basic study-
test task structure and the full battery of cognitive tasks designed to examine the effect
of memory load on object recognition memory. Section 4.3 describes the procedure
used to present the battery of cognitive tasks to each subject and the software used to
control task presentation. Subjects who participated in the study are described in Section
4.4. The steady-state visual flicker employed as the probe stimulus is discussed in
Section 4.5, and recording procedures are described in Section 4.6. Section 4.7 explains
how behavioural data was analysed. Finally, section 4.8 explains the analysis of

Methods
electrophysiological data obtained using both long and short evaluation periods to
investigate modal and transient neural activity respectively.
4.2 Cognitive task design
The basic structure of each task is shown in Figure 4.1. Each task consisted of a study
phase and a test phase. During the study phase, subjects memorized 1, 3 or 5
sequentially presented, abstract, two-dimensional objects. Each object remained visible
for a relatively extended period (3.5s) to facilitate consolidation into longer-term
memory. During the test phase of the task, in the ‘task interval,’ subjects viewed
sequentially a larger number of objects that included memorized objects (targets) and
new objects (non-targets). Subjects were required to identify targets by pressing a
button with their right hand ‘as quickly and as accurately as possible.’ The ratio of non-
target to target objects was 3:1, and in each test phase, 15 non-target objects and 5 target
objects were presented (see Figure 4.2). Non-target objects were designed to look
similar to target objects, thereby making discrimination more difficult. Because a
detailed match had to be made with objects in memory, it was expected that the use of
explicit memory processes would be maximized. With more objects to remember, and a
corresponding increase in the number of distractor objects with similar appearance, the
task becomes more difficult and places a greater load on retrieval processes.
memorize objects
40s
(20 trials)(1,3, or 5)
object object
1400ms 600ms
blank
(1 trial)
40s 40s
study phase test phase
numbers interval numbers intervaltask interval(20 trials) (20 trials)
Figure 4.1 Study-test experimental design
80

Methods
In the test phase, the task interval was both preceded and followed by a 40s ‘numbers
interval.’ Subjects had been instructed to relax and press the hand-held button slowly
whenever the number ‘4’ appeared in these numbers intervals that would precede and
follow the task interval. The purpose of the first numbers interval was threefold: (i) to
ensure that remembered objects had been transferred to longer-term memory stores, (ii)
to distract subjects so that memorized items would not be rehearsed, thus limiting
working-memory or short-term memory processes, and (iii) to orient subjects to a
computerized task prior to performing the object retrieval operations of the task interval.
The final numbers interval was included so that differences in the SSVEP due to
possible differences in attention levels between the beginning and end of the task could
be compared. This analysis, however, is not included in this study.
* response required
* * * * *
11
11
11
11
11
22
22
22
22
22
33
33
33
33
33
44
44
44
44
44
* * * * *
* * * *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
* *
*
*
* *
*
*
task block (test phase)
* * * * * * * * * *12341234123412341234 12341234123412341234
study phase(memorize)
bas1bas2
m1p
m3p
m5p
m1
m3
m5
task interval
Figure 4.2 Experimental task design
Experimental task design for order 1 showing both study and test phases. For order 2, the m1p/m1 and m3p/m3 task block orders were reversed.
The complete experimental task design is illustrated in Figure 4.2. First, two baseline
task blocks were presented to all subjects. In these blocks, the requirements of the task
interval were identical to those of the numbers intervals. In other words, subjects were
required to press the button when the number ‘4’ occurred for the full two minutes.
However, only the middle 40s comprised the actual baseline task. During the first of
81

Methods
these baseline task blocks (bas1), the steady-state visual flicker (probe stimulus) was
turned-on half way through, that is, after one minute. This was done so that any
spontaneous 13Hz signal in the background EEG (first half of the task) could be
compared with the driven 13Hz SSVEP response (second half of the task). Although not
important to the current investigation, this was standard practice at the Brain Sciences
Institute as it was a straightforward method of checking that the recording system was
working correctly. The result of this analysis is provided in Appendix D. The visual
flicker remained on throughout the second baseline task block (bas2). The bas2 task
interval was used a reference task for the memory load tasks so that task-related effects
could be investigated. The baseline tasks also served to further reduce any effects of
novelty and anxiety associated with an unfamiliar environment.
Number and object stimuli were visible for 1400ms and were followed by a blank
interval of 600ms, yielding a total of 2000ms per stimulus item. Each task block
consisted of a 40s numbers interval, followed by a 40s task interval, followed by
another 40s numbers interval, giving a total duration of 120s. In other words, the
transition from numbers to task in the practice and main memory tasks occurred after 20
stimuli (40s) and from task back to numbers after another 40 stimuli (80s). The baseline
interval for analysis comprised the middle 40s of the bas2 task block.
On completion of the two baseline task blocks, six task blocks of the structure
illustrated in Figure 4.1 were presented. Three different memory load tasks were used in
which 1, 3 or 5 objects were memorized. There were two task blocks for each memory
load. The first incorporated a practice task interval (m1p, m3p or m5p, in Figure 4.2)
and was included both to familiarize subjects with the memory load, and to facilitate
consolidation of studied objects into a longer-term memory. The second task block
incorporated the task interval (m1, m3 or m5, in Figure 4.2), the SSVEP data from
which was used in the analysis. Memorized objects were identical in both practice and
main memory tasks. Thus, for each memory load (m1, m3 and m5), by the time subjects
performed the main memory task, each studied item had been viewed at least 3 times,
that is, twice during each study phase and at least once during the practice test phase.
Furthermore, the overall lag time from the first study phase to the second test phase was
approximately 4 to 5 minutes. Thus, in addition to being well encoded, the likelihood of
82

Methods
using of longer-term memory processes to retrieve targets from memory was
maximized.
Practice task blocks served primarily to reduce any possible novelty effects. In addition,
tasks were partially counterbalanced to reduce order effects. Half of the subjects were
given ‘order 1’ (shown in Figure 4.2) and the remaining subjects were given ‘order 2,’
where the m1p/m1 and m3p/m3 task orders were reversed.
As seen in Figure 4.2, targets were inserted into the same position in the sequence in the
main memory task for all three memory loads. This was done so that the preceding item
history would be comparable for targets and non-targets across each of the different
memory load tasks. However, because targets appeared in the same position for each
task interval, it was thought that subjects might have been able to predict their
appearance. Nevertheless, when asked about this after completion of the recording
session, all subjects indicated that the presentation of targets appeared to be random.
The fact that different target locations were used in the practice tasks, acting to distract
subjects from learning the sequence (see Figure 4.2) probably contributed to this.
Furthermore, behavioural and electrophysiological data (see Chapter 5) are consistent
with non-prediction of target position.
Task instructions given to subjects are provided in Appendix A.
4.3 Task presentation and stimulus parameters
Each of the stimuli shown in the ‘numbers’ and ‘task’ intervals were presented centrally
on a computer monitor. They subtended equal horizontal and vertical angles of 1.3
degrees when viewed at a distance of 1.3 metres. The stimuli had an average luminance
of approximately 15.0 Cd/m2 against the monitor background of 1.2 Cd/m2, measured
using a Tektronix J16 narrow angle Digital Photometer.
All stimuli were presented on the monitor of the ‘task computer.’ The timing of
stimulus presentation was controlled using an external millisecond timer that was
83

Methods
monitored through the games port of the task computer. Subjects’ responses were also
monitored through the games port. The computer program that controlled stimulus
presentation, timing and response was written by the author using the Microsoft Quick
C Editor and was compiled using the Microsoft C compiler, version 5.1. Each stimulus
item was ‘drawn’ in real-time to a virtual memory location of the graphics card in the
task computer. The program then waited for the next ‘screen refresh’ pulse before
displaying the stimulus (Pipingas and Maruff 1991). The games port was then
simultaneously inspected for the subject’s response and the elapsed time of stimulus
presentation. Once the presentation time had elapsed, a ‘blank’ screen was displayed. If
the response button was pressed while the stimulus item was on the screen, the response
time was stored in memory. On completion of the task, all presentation and response
times were written to a file on the hard disk for off-line analysis. Presentation and
response times were determined to an accuracy of 1ms.
As explained in section 4.2, the sequences of stimuli were pre-determined and were
stored in ‘script files’ constructed for each task. Script files were also constructed for
the presentation of target objects in the study phases. A feature of the software was that
the next task was automatically loaded once the preceding task had been completed.
Each task was ‘triggered’ by the EEG data acquisition computer via a separate line
through the games port of the task computer.
4.4 Subjects
Fifty-one male subjects participated in the study. Their ages ranged from 17 to 48 years,
with a mean of 22.50 and standard deviation of 6.28 years, and all were right handed as
assessed using the Edinburgh Handedness Inventory (Oldfield 1971). At the time of
testing, most subjects were completing undergraduate courses within the university and
had a science or engineering background. The others were tutors and lecturers in similar
disciplines. Subjects were asked to set aside approximately ninety minutes for the study,
and most sessions were completed in approximately 70 minutes. A session included
completion of a number of forms, preparation, recording and debriefing. All subjects
gave their informed consent to participate in the study and were asked to withdraw if
84

Methods
they had a history of epilepsy. The study was approved by the Swinburne University
Human Research Ethics Committee.
Prior to arrival, subjects were assigned to one of two groups, order 1, with a memory
load task order of 1, 3 then 5 items, or order 2, with a memory load task order of 3, 1
then 5 items. Subjects were naïve as to the nature of the task design and the group to
which they were assigned. Twenty-seven subjects completed order 1, and 24 subjects
completed order 2. In a subsequent off-line analysis that was used to assess the integrity
of the EEG, 11 subjects were eliminated from subsequent analysis, leaving 19 in the
order 1 group and 21 in the order 2 group (total 40).
4.5 Probe stimulus
The probe stimulus used to evoke the SSVEP consisted of a 13Hz sinusoidal flicker
subtending a horizontal angle of 160° and a vertical angle of 90°. This was presented to
subjects using specially designed glasses. Two half-mirrored strips reflected the flicker
into the eyes whilst simultaneously allowing the subject to view the task computer. The
flicker was generated by a 4 x 4 array of red light emitting diodes (LEDs) housed in a
small Faraday cage to prevent electrical contamination of the EEG. When viewed
against the background, the maximum luminance reflected at the peak of the stimulus
waveform was 3.2 Cd/m2, and the minimum was 1.2 Cd/m2. The calculated modulation
depth was 45% (see Figure 4.3). The optimum luminance was contingent on both the
amplitude of the SSVEP response and subject comfort. Results of a separate experiment
used to determine the optimum luminance for the present study is provided in Appendix
B.
85

Methods
Time (ms)
0 50 100 150 200 250
Lum
inan
ce (C
d/m
2 )
0
1
2
3
4
α
β
x
y
Calculation of Modulation Depth
45.04.40.2
2.12.32.12.3
==+−
=+−
≡=yxyx
βαDepthModulation
Figure 4.3 Calculation of modulation depth
Modulation Depth is the ratio of the mean-to-peak to the mean amplitude of the stimulus intensity waveform. This ratio can also be expressed in terms of the measured peak and background luminances x and y.
4.6 Recording
Signals at the scalp were detected using an array of electrodes mounted in a specially
designed helmet. The signals were amplified and filtered in two stages, and were
digitized and stored on computer. The 13Hz probe stimulus was synchronized to data
acquisition enabling comparison of the SSVEP phase with the phase of the stimulus
waveform. The EEG helmet and electrodes are described in section 4.6.1. Section 4.6.2
contains a description of the recording instrumentation and the method used to generate
the 13Hz probe stimulus.
86

Methods
4.6.1 Electrodes and recording setup
The electrode locations used in this study included all the scalp positions in the
International 10-20 System, with additional electrodes located between these sites to
give a total of 64 electrodes (see Figure 4.4). This montage gives an average separation
between electrodes of 3.2cm, which is adequate for observing effects localized to gross
cortical regions (Silberstein et al. 1990b). The electrodes were mounted in a helmet
designed at the Swinburne Centre for Applied Neurosciences (SCAN), now the Brain
Sciences Institute (Ciorciari et al. 1987). The electrodes were spring-loaded within the
helmet, and retracted prior to placement on the subject’s head. Once the helmet was
positioned correctly, the electrodes were gently lowered onto the scalp, and conductive
gel was introduced through each to facilitate contact between the electrode tip and the
scalp. The electrode tip consisted of a silver disc with a layer of silver chloride applied
just prior to recording. Electrode impedances varied from a minimum of 5kΩ to a
maximum of 35kΩ when measured at 40Hz and with an amplifier input impedance of
200MΩ (Ferree et al. 2001). All electrodes were referenced to a balanced non-cephalic
montage (Stephenson and Gibbs 1951) with the nose serving as the ground. Silver/silver
chloride surface electrodes were also used for these. The electrocardiogram (ECG) was
essentially eliminated using a canceling procedure described by Stephenson and Gibbs
(1951).
0
5
12
13 14 15 16 17 1820
21
22 23 24 25
27
28 29 30 31 32 33 34 35 36
37
3839 40 41
42
43
44
4547
52
59
60 61 62
63
5354
55 5657
58
67 8 9
1011
1 2 3
4
Fp1 Fp2
Fz
Cz
Pz
Oz O2O1
T5
P3 P4
T6
T3 T4C3 C4
F7F3 F4
F8
19
26
48 49 5051
46
Figure 4.4 Sixty-four scalp recording locations
Sixty-four recording locations included all International 10-20 sites with additional electrodes located between these sites.
87

Methods
4.6.2 Recording instrumentation and phase locked data acquisition
The instrumentation used to amplify, filter, digitize and store the EEG is depicted in
Figure 4.5.
The pre-amplifier stage provided initial amplification close to the subject. Each
electrode was wired into one input of a differential amplifier with a fixed gain of 4000,
an input impedance of 200MΩ, and a common mode rejection ratio greater than 100dB.
The other input of each differential amplifier was common, and was connected to the
output of the non-cephalic reference.
A second stage of instrumentation provided further amplification, filtering and sampling
of the EEG. Noise was reduced using a programmable active Butterworth filter
amplifier with a high frequency cut-off of 26Hz and a low frequency cut-off set to
0.5Hz. The gain could be further increased at this stage. A rotary switch was used
allowing five pre-set levels of amplification to cater for inter-subject variability in the
EEG amplitude. Sixty-four buffered sample-and-hold circuits were used to
simultaneously sample the outputs of the filter amplifiers.
Figure 4.5 Experimental recording arrangement
88

Methods
A plug-in card manufactured by Data Translation Corporation, together with a 486,
66MHz, IBM compatible computer, was used to sample and digitize the 64 channels of
EEG data. The EEG signals were digitized to an accuracy of 12 bits by an analogue to
digital converter (ADC) that was incorporated into the plug-in card. The 4096 digital
levels corresponded to a full-scale range of –5 to +5 volts. Equation 4.1 was used for the
conversion of digital levels back to microvolts representing the EEG amplitude. Data
was sampled to computer hard disk at a rate of 208Hz per channel. All instrumentation
except the plug-in card and computer interface were designed and constructed at the
Brain Sciences Institute (Simpson 1997).
VoltsGainGain
FSRFSRAmp
Ampset
ueanadigital
digitalactual µ 10
var
6
log
=
VoltsGain
AmpAmp digital
actual µ 6104.0
var
=
Equation 4.1 Calculation of unamplified EEG amplitude
Calculation of EEG amplitude where Ampactual is the EEG amplitude in microvolts, Ampdigital is the equivalent digital amplitude, FSRdigital (digital full-scale-range) equals 4096, FSRanalogue (analogue full-scale-range) equals 10, Gainset (initial amplification) equals 4000, Gainvar is the variable gain of the filter amplifiers. The factor of106 converts volts to microvolts.
Software for data acquisition was written in the DAOS1 language (Schier 1994). This
software was designed to acquire a pre-determined number of samples of data
equivalent to length of the task. Software was also written to set the high and low-pass
levels of the programmable filter amplifier, as well as the frequency of the sinusoidal
probe stimulus.
The instrumentation was designed to synchronize data acquisition with the sinusoidal
probe stimulus. Data acquisition was locked to the 13Hz stimulating frequency so that
acquisition always commenced on the positive zero crossing of the stimulus cycle.
Sixteen data points were sampled for all stimulus cycles, and the samples were acquired
at the same point in each stimulus cycle. The advantage of synchronizing data
1 Data Analysis Operating System (DAOS) by Laboratory Software Associates Pty. Ltd., Melbourne.
89

Methods
acquisition with stimulation is that the phase of the SSVEP can compared precisely with
that of the stimulus waveform. Coherent demodulation of the SSVEP, utilizing Fourier
techniques (see Regan 1989), can subsequently be performed off-line to calculate the
SSVEP amplitude and phase for every stimulus cycle. Furthermore, the same coherent
demodulation algorithm can be used for any frequency of stimulation, given that the
number of samples per stimulus cycle remains the same.
4.7 Analysis of behavioural data
Response times and error scores for both practice and main memory tasks were
calculated for all forty subjects. There were five targets within each ‘task interval,’ as
discussed in section 4.2. Response times for correct responses only were averaged. The
number of errors of omission (missed responses) and commission (responses to non-
targets) were noted for each individual. Individual averaged response times and error
scores were then averaged across all subjects. The Student’s t test was applied to the
response time data to determine whether differences between memory loads were
statistically significant. Memory loads of 3 and 1, 5 and 1, and 5 and 3 were compared
for both practice and main memory tasks.
4.8 Offline signal processing
The EEG data set obtained for each individual was analysed on an IBM 486 compatible
computer using a set of programs written in DAOS and in Microsoft C 5.1. The
programs extracted the 13Hz SSVEP from the EEG signal, detected and replaced
artifact-contaminated EEG data, averaged the SSVEP over the appropriate period of
time to obtain the desired temporal resolution, and averaged the data across all subjects.
4.8.1 Extraction of the 13Hz SSVEP from the EEG signal
The SSVEP Fourier coefficients were obtained for each stimulus cycle using equation
4.2. The 16 EEG data points locked to each stimulus cycle were multiplied by the
90

Methods
corresponding 16 points in the reference waveform (generated in software) and then
integrated over the 16 points to give the sine Fourier coefficient. The cosine Fourier
coefficient was obtained in a similar way, however, the reference waveform was shifted
by 90 degrees prior to multiplication with the EEG data points. All EEG data points for
every electrode and task were analysed in a similar way and stored in a file on hard disk
as a series of sine/cosine pairs. The SSVEP amplitude and phase can be calculated from
these coefficients using equation 4.3 to form a new series of phase/amplitude pairs. This
is essentially a conversion from a cartesian to a polar representation of the data. This
last calculation was performed after the EEG and Fourier coefficients were examined
and corrected for artifacts. Regan (1989) describes methods for recording SSVEPs, as
well as the technique of sine/cosine multiplication for calculating the Fourier
coefficients.
( )
∆+∆+
∆= ∑
−
=
τπττ
inTT
inTfS
aS
in
2cos)(1 1
0
( )
∆+∆+
∆= ∑
−
=
τπττ
inTT
inTfS
bS
in
2sin)(1 1
0
Equation 4.2 Calculation of single cycle Fourier coefficients
Calculation of SSVEP Fourier components, where an and bn are the cosine and sine Fourier coefficients respectively, n represents the nth stimulus cycle, S is the number of samples per stimulus cycle (16), ∆τ is the time interval between samples, T is the period of one cycle, and f(nT+i∆τ) is the EEG signal.
( )22nnamplitude baSSVEP +=
=
n
nphase
ab
SSVEP tana
Equation 4.3 Calculation of SSVEP amplitude and phase
Calculation of SSVEP amplitude and phase where an and bn are the cosine and sine Fourier coefficients respectively. Amplitude and phase components can be calculated using either single cycle Fourier coefficients or coefficients that have been calculated by integrating across multiple cycles.
91

Methods
4.8.2 Automatic detection of artifact in the EEG signal
As already explained in section 3.4, the SSVEP is relatively insensitive to artifact
because extracting narrow-band phase-locked 13Hz signal eliminates a very substantial
proportion of artifact. Nevertheless artifact cannot be ignored altogether. For example,
there may be excessive clipping at the input of the ADC due to intermittent contact with
the scalp. Also high levels of eye and muscle movement and 50Hz mains artifact can be
problematic. Muscle movement (EMG), in particular, can contain significant 13Hz
activity. However all of these can be detected and minimized using a suite of programs
developed at SCAN (Schier 1994).
Artifact can be detected using a two-stage process. The first stage makes use of the fact
that EEG has a Gaussian amplitude distribution (McEwan and Anderson 1975). To
eliminate electrodes with an unacceptable signal, amplitude histograms of EEG data for
each electrode are calculated and correlated with a Gaussian function to assess the
integrity of the EEG recording. This technique is very good for detecting data that
exceeds the input range of the ADC. When this occurs, the power spectra show maxima
at the positive and negative extremes of the amplitude histogram rather than at the mean
level of the EEG2. Using this technique, electrodes with a signal with a correlation
coefficient of less than 0.75 were classified as unacceptable.
The second stage makes use of the fact that the EEG from adjacent sites is highly
correlated. This is because the scalp signal is spatially ‘smeared’ after it has passed
through the layers of CSF, skull and scalp (Nunez 1981). This stage of detection is
particularly useful for forms of artifact, such as EMG and 50Hz mains, that display a
Gaussian-like amplitude distribution, and can therefore pass through the first stage of
detection. For this analysis, the data was divided into a series of 16 cycle sweeps. For
each sweep, the mean of the Fourier coefficients at each electrode site were correlated
with the weighted mean3 of the coefficients at the four nearest neighbouring electrode
sites. For each electrode, a look-up table that contains a record of that electrode’s nine
2 In an AC-coupled system (0.5Hz) the mean level of the EEG is zero volts. 3 The closer a neighbouring electrode is to the electrode being analysed, the more ‘weight’ it is given in
the mean.
92

Methods
nearest neighbouring electrodes was used. Extra electrodes are necessary because an
adjacent electrode may have already been flagged as unacceptable, thus requiring the
next one in the list to be used. Here, an electrode was considered unacceptable if the
correlation coefficient was less than 0.6. The SSVEP values for each electrode
identified as unacceptable were replaced with the weighted means of the SSVEP values
for that electrode’s four nearest acceptable neighbours. A subject was eliminated from
further analysis if eight or more electrodes were unacceptable in any one task.
4.8.3 Calculations for modal and transient effects
Each pair of Fourier coefficients was calculated over one stimulus cycle, a time interval
of 1/13s or 77ms. Such a short integration period gives a low signal to noise ratio.
However, as with traditional evoked potential averaging where individual ERPs can be
averaged, successive Fourier coefficients can also be averaged to give a higher signal-
to-noise ratio. It must be remembered, however, that the longer the integration period,
the lower the temporal resolution. Therefore, a trade-off situation exists where
consideration must be given to both the signal-to-noise ratio and the temporal
resolution.
In this study, Fourier coefficients were calculated using both long and short averaging
periods to investigate different aspects of cognition during performance of the tasks. A
long averaging period (ie. 40s) was used to investigate more sustained modal effects
reflecting the mental state of subjects while performing the task. In this situation,
transient SSVEP changes associated with specific events within each task cannot be
observed, rather each ‘task interval’ is considered as a whole, essentially indicating the
subject’s mental state, or processing ‘mode,’ during this time. A short 20-cycle (1.54s)
averaging period was used to investigate the dynamics of each task; that is, the transient
changes that occurred as individual stimuli were presented. With the cosine averaging
window and the computer setup used, this gave an effective temporal resolution of
870ms. Although this achieves a relatively high temporal resolution, the signal-to-noise
ratio is relatively poor. However, signal-to-noise ratio can be improved by the averaging
of data, as is the case with traditional ERP averaging. For example, data for all target
93

Methods
objects can be averaged together, as can data for non-target objects. The methods used
for both the long and short averaging periods will be described below.
4.8.3.1 Long averaging period (sustained effects)
To examine sustained or modal effects using a long averaging period of 40s, the
following procedure was followed for each electrode for each subject. For each ‘task
interval’ of the memory and bas2 task blocks, all Fourier coefficients were averaged. In
other words, forty seconds of sine and cosine Fourier coefficients were collapsed to one
pair for each task. The three resultant pairs of Fourier coefficients for each of the three
memory load conditions were further averaged to give one pair of overall averaged
Fourier coefficients. This overall average pair was compared with the corresponding
pair of Fourier coefficients for the baseline condition to examine sustained SSVEP
amplitude and latency patterns associated with retrieval mode processes. The separate
individual pairs of Fourier coefficients, averaged over each of the three ‘task intervals’
(ie. m1, m3 and m5), were compared to investigate the effect of memory load on
retrieval mode processes.
4.8.3.2 Short averaging period (transient changes)
To examine more transient effects occurring during the processing of an individual
stimulus item, Fourier coefficients were calculated using a moving cosine (Hanning)
window with a width of 20 stimulus cycles. For a stimulus frequency of 13Hz, this
would, in theory, correspond to an effective temporal resolution of 769ms. However,
computational limitations resulted in an effective temporal resolution of 870ms. At the
start of the averaging procedure, the cosine window was positioned over the first 20
Fourier coefficients of the task block. Averaging commenced at this point, and the result
was stored in a secondary file as a new Fourier pair. The window was then advanced by
one pair of Fourier coefficients. Averaging was again performed, and the result was
stored in the next position of the secondary file. This process was repeated until all data
in the initial file had been averaged. This procedure was followed separately for each
94

Methods
electrode site, for each task, and for each subject. Figure 4.6 shows graphically the
result of this procedure. It can be seen that the curves are considerably smoother for the
20-cycle averaged data than for the single cycle data. A temporal resolution of 870ms
(20-cycle averaged data) was chosen to achieve a satisfactory signal-to-noise ratio in
conjunction with a temporal resolution smaller that the interval between the appearance
of each object (2000ms), so that SSVEP changes occurring during the course of
stimulus processing could be investigated. The following section describes the
procedure for averaging data across all target and non-target presentations associated
with a correct response.
SSVEP amplitude and phase20 unit window
SS
VE
P a
mpl
itude
(mic
rovo
lt)
02468
101214
Time (s)
0 10 20 30 40 50 60 70 80 90 100 110
SS
VE
P p
hase
(radi
an)
-4-3-2-101234
Recorded EEG with superimposed SSVEP
Time (s)53 54 55 56 57
EE
G a
mpl
itude
(mic
rovo
lt)
0
50
100
150
200
250
Extracted single cycle SSVEP amplitude and phase
SS
VE
P a
mpl
itude
(mic
rovo
lt)
05
101520253035
Time (s)
0 10 20 30 40 50 60 70 80 90 100 110
SS
VE
P p
hase
(radi
an)
-4-3-2-101234
stim
ulus
on
Figure 4.6 Single cycle and averaged Fourier coefficients
SSVEP amplitude and phase data calculated from single cycle Fourier coefficients and averaged using a 20 unit window (temporal resolution = 870ms). (subject BT; electrode 61 (Oz); first baseline task(bas1) - stim-off/stim-on)
95

Methods
4.8.3.2.1 Event averaging
Analyses of data for target and non-target objects were conducted separately. Because
task presentation and data acquisition were synchronized (see section 4.3), the exact pair
of Fourier coefficients corresponding to the appearance of each object could be
identified. The following averaging procedure was observed for each electrode, task and
subject. A five-second epoch of data, which was centred on the appearance of each
correctly recognized target object, was extracted. The data for these corresponding
epochs was then averaged and, as for the long averaging period analysis, sine and cosine
coefficients were calculated separately. Data for non-target objects was then averaged in
the same way as for target objects.
Because the five-second epochs of 20-cycle averaged data contains transient SSVEP
phase changes superimposed on slow changes, when looking for transient patterns, slow
changes are a confounding factor. To eliminate these slow changes, an additional step
was included between the extraction of each epoch of data and the subsequent averaging
of data for corresponding epochs. The data in each epoch was converted to its polar
equivalent using equation 4.3. The mean phase was calculated from the series of phase
coefficients. This mean phase was then subtracted from every phase coefficient in the
data epoch. This process, in effect, eliminated any sustained, or DC, phase levels. The
mean phase for each epoch was therefore zero radians, and phase changes were related
to the transient processing of each item rather than to the sustained phase value
differences that may have existed between memory loads. Amplitude and phase
coefficients were converted back to sine and cosine Fourier coefficients prior to
averaging the data for corresponding epochs.
4.8.4 Group averaging
For both long and short averaging period analyses, only the main memory tasks for the
three memory load conditions are discussed in this section. Long averaging period
analyses for the practice tasks are shown in Appendix E.
96

Methods
So far, the 13Hz SSVEP Fourier coefficients have been extracted for each task,
averaged, and in the case of the short averaging period, averaged across target and non-
target epochs. These analyses were carried-out separately for each task, at each
electrode site, and for all subjects. Finally the data was averaged across all forty subjects
for both long and short averaging period analyses.
4.8.4.1 Group averaging: long averaging period (sustained effects)
The following steps were taken to obtain group averaged data that would show
sustained or mental state effects on the SSVEP. Prior to averaging across subjects, the
SSVEP amplitude values for each subject had to be normalized so that those individuals
showing large amplitudes did not dominate the group average. For each subject, a
representative value, or normalization factor, was obtained by calculating the mean
amplitude for the baseline condition across all 64 electrodes. All SSVEP amplitude
values for each electrode, for all three individual memory load conditions, and for the
overall average of the three memory load conditions were then divided by the unique
normalization factor for that individual. The baseline task was similarly amplitude
normalized.
The SSVEP phase value was also adjusted prior to averaging. This adjustment was
necessary because the phase values for each subject are arbitrary, and therefore
averaging across subjects could result in phase cancellation. Each individual’s phase
values for each of the three memory load conditions, and for the overall average of the
three memory load conditions, were calculated relative to the average phase for the
baseline condition. This also allowed deviations in phase from the baseline to be
examined easily. This adjustment was performed by subtracting the 64 baseline phase
values (one for each electrode) from the 64 phase values for each memory load
condition. In other words, the phase at every electrode site for the baseline condition
took the value zero, and the phase values for the memory load conditions, that is, the
three separate and the overall averaged memory load conditions, represented deviations
from this zero phase. This adjustment reduces cancellation effects when averaging
across subjects. It is also much easier to see whether there was a phase advance or phase
97

Methods
lag (ie. latency decrease or increase) relative to the baseline condition at each electrode
site.
Once the long averaging interval data had been amplitude normalized and phase
adjusted for each individual, group averaging was performed. This was done by first
converting the polar data points (amplitude and phase) back to cartesian coordinates
(sine and cosine coefficients) and then averaging the sine and cosine coefficients
separately across all subjects. The resulting cartesian average was then converted back
into polar form, allowing topographic mapping of amplitude and phase values for each
task.
SSVEP amplitude and phase differences between the average of the three memory load
conditions and the baseline condition were calculated using the averaged group data.
SSVEP amplitude and phase differences between memory loads of 3 and 1, 5 and 1, and
5 and 3, were also calculated. These differences addressed more directly the issue of
how increases in memory load affect the SSVEP response associated with retrieval
mode.
4.8.4.2 Group averaging: short averaging period (transient changes)
The short averaging period analysis used the data for each individual that had been
averaged using a 20-cycle moving window to give a temporal resolution of 870ms. The
windowed Fourier coefficients for target and non-target stimuli were event averaged
separately so that the SSVEP amplitude and phase changes engendered by the
appearance of target and non-target stimuli could be compared. As already discussed,
event averaging of SSVEP data involved adjustment of the phase prior to averaging
across successive five-second epochs of data. Thus, phase adjustment prior to
calculating the group average was not necessary as the reference phase value, in this
case the mean value over the five-second epoch for targets and non-targets for each of
the three memory load conditions, was already zero.
98

Methods
As already explained, for the long averaging period analysis, amplitude normalization
had to be conducted prior to group averaging so that individuals showing large
amplitudes did not dominate the group average. This was also required for the short
averaging period analysis and was carried-out as follows. Firstly, for each electrode, the
Fourier coefficients for the five-second epoch for non-targets with a memory load of 1
were averaged separately to give a single pair of coefficients for this epoch. The SSVEP
amplitude at each electrode site was then calculated from these averaged coefficients
and further averaged across the sixty-four sites to produce the normalization factor. A
normalization factor was calculated for each subject. Each subject’s normalization
factor was then divided into all SSVEP amplitude values within that subject’s five-
second epochs for target and non-target averages for each electrode site for all three
memory load conditions.
Once amplitude normalization had been completed, the SSVEP values were converted
to cartesian form for group averaging. For each point in the five-second epoch, sine and
cosine coefficients were averaged separately across all subjects, resulting in a five-
second epoch of group averaged coefficients. This process was repeated for each
electrode and for each experimental condition, that is, for targets and non-targets for
each of the 3 memory load conditions. All pairs of Fourier coefficients were then
converted back to polar form.
This group-averaged data was used to investigate the effect of memory load on transient
SSVEP changes by comparing SSVEP values for the different memory loads.
Differences in both amplitude and phase between memory loads were then calculated
using the group averaged data. Differences in both amplitude and phase between values
obtained for memory loads of 3 and 1, 5 and 1, and 5 and 3, were calculated for each
electrode site. Target and non-target differences were calculated separately. This
procedure showed how differences in memory load affected transient patterns in the
SSVEP response during the five second epoch centred on the appearance of an object.
For target analyses, topographic differences between high and low memory loads reflect
the effect of increased difficulty in recognizing targets on the SSVEP response.
99

Methods
Differences between SSVEP values for target and non-target objects were also
calculated. This analysis, although included for completeness, was problematic given
that targets prompted a motor response and non-targets did not. Furthermore, targets
occurred less frequently than non-targets, therefore SSVEP differences may have
reflected probability effects.
4.8.5 Topographic mapping of SSVEP data
Two-dimensional topographic maps were constructed using the SSVEP amplitude and
phase data from both the long and short averaging period analyses. Inter-electrode
values were calculated using a spherical spline interpolation procedure (Cadusch et al.
1992). Maps were displayed in a 640 x 480, 256 colour mode. Different scale ranges
and conventions were used depending on whether individual subject data, group data or
group difference data was being mapped. Specific details for each type of data will be
discussed in turn.
Amplitude and phase data for individual subjects, calculated using the long averaging
period analysis, was produced mainly for illustrative purposes (see Figures 5.3 and E-i).
The amplitude scale represents the ‘raw’ amplitude, in microvolts, of the SSVEP
elicited in response to the 13Hz visual flicker. Warmer colours represent a larger
response to the visual flicker. Topographic maps showing the SSVEP phase were
produced for individual subjects using a continuous colour scale such that the same
colour was used for both +π (+3.1) radian and –π (-3.1) radian. This is done because
phase variations for individual subjects can be large and not necessarily centred about
zero radians, resulting in phase transitions at +π and -π. In this situation, if different
colours are assigned to +π and -π, sharp discontinuities in colour can occur, making
interpretation of topographic variations more difficult. A transition to a more positive
colour is interpreted as a phase advance, and to a more negative colour, a phase lag.
Group SSVEP amplitude and phase data was mapped somewhat differently (see Figures
5.4 and E-ii). Firstly, since amplitude data was normalized prior to group averaging (see
sections 4.8.4.1 and 4.8.4.2), amplitude values are smaller. The scale range therefore
100

Methods
represents a smaller value for group data than for individual subject data. Again,
warmer colours represent a larger response to the visual flicker. Variations in phase
were much smaller for group data (±0.5 radian) than for individual subject data, and
centred on zero radians due to the phase adjustment performed prior to group averaging
(see sections 4.8.4.1 and 4.8.4.2). A continuous colour scale, like the one used for
individual subject data, was therefore not required. The baseline phase was zero at all
sites (yellow/green colour) as a result of the phase adjustment. Warmer colours
represent a phase advance, and cooler colours a lag, relative to the mean phase for the
baseline condition.
In early studies, amplitude attenuation and phase advance were interpreted as reflections
of increased neural activity. Therefore, the colour convention adopted was warmer
colours for amplitude attenuation and phase advance, and cooler colours for amplitude
increase and phase lag. While the picture has now become more complicated (see
Chapter 3) this colour convention has remained in use and is used here in the depiction
of differences between task conditions. Amplitude differences were therefore calculated
such that an SSVEP amplitude attenuation observed for higher relative to lower memory
load conditions is represented by warmer colours. Similarly, SSVEP phase differences
were calculated such that SSVEP phase advances for higher relative to lower memory
load conditions is also represented by warmer colours. Phase values can also be
expressed in terms of latency, where 1 radian is equivalent to a latency of 12.2ms. A
phase advance corresponds to a latency decrease and a phase lag to a latency increase,
therefore warmer colours represent latency decreases and cooler colours increases.
Amplitude and latency group difference data are shown in Figures 5.5, 5.6, 5.11, 5.12
and 5.13.
4.8.6 Statistical analysis and Significance Probability Mapping
Because the SSVEP consists of two components, amplitude and phase, the bivariate
Hotelling’s T2 test was used to determine the statistical significance of the SSVEP
response to differences between task conditions. The square root of the Hotelling’s T2
parameter (Hotelling’s T) was mapped rather than the T2 parameter itself because small
101

Methods
areas could contain very large T2 values which would dominate the scale. Mapping the
T values allowed for much smoother contours. This type of mapping is called
significance probability mapping (SPM) (eg. Duffy et al. 1981; Hassainia et al. 1994)
and more recently has been termed statistical parametric mapping (SPM) (eg. Acton and
Friston 1998). On the SPM maps, iso-T contours of 2.02, 2.70, 2.97 and 3.55,
corresponding to p values for a single comparison of 0.05, 0.01, 0.005 and 0.001, were
used.
A p value of 0.05, or 5%, is normally considered the threshold for statistical
significance. Long and short averaging period analyses however were not based on
single comparisons as data was analysed from 64 recording sites. A Bonferroni
correction is normally applied to p values to allow for these multiple independent
measures (Abt 1983). If all electrodes were independent of each other, 0.05 would be
divided by 64. However, because brain electrical activity from neighbouring scalp
recording sites is highly correlated, dividing the p value by 64 is incorrect (Silberstein et
al. 1995a). Rather, a value of 5 derived through spatial principal components analysis
(eg. Silberstein and Cadusch 1992) represents more accurately the degree of
independence for 64 separate, but correlated, recording sites. In this instance, therefore,
a p value of 0.01 (1%) on the Hotelling’s T topographic map was taken as the threshold
for statistical significance. It should also be noted, that where multiple time points are
sampled in a temporal sequence, a p value of 0.05/(5 x no. of time point samples)
should be used as the threshold for statistical significance (Silberstein et al. 1995a).
102

Chapter 5 Results
Response time and accuracy data were analysed in order to investigate whether an
increase in memory load resulted in an increase in task difficulty. It was anticipated that
response time would increase with memory load, and accuracy might decrease. While
response time did, in fact, increase with memory load, memory load had no significant
effect on accuracy.
Sustained task and transient item-related SSVEP responses produced during the
performance of an episodic retrieval task were examined to investigate neural activity
associated with sustained retrieval mode and transient retrieval success processes.
To investigate neural regions involved in the maintenance of a retrieval mode, the
SSVEP amplitude and latency values were averaged across the entire duration of an
episodic retrieval task and compared with the averaged SSVEP amplitude and latency
values obtained during a non-episodic retrieval baseline task. Sustained retrieval mode
processes were associated with SSVEP amplitude and latency reductions at most
recording sites. While amplitude reductions were relatively uniform across all recording
sites, latency reductions were most prominent at right fronto-temporal sites.
To investigate neural regions associated with successful retrieval from memory, the
SSVEP amplitude and latency were calculated using a short averaging period, and
values for different experimental conditions were compared at certain time points
before, during, and after the appearance of the stimulus object. Transient load-
dependent SSVEP amplitude and latency reductions occurred during successful retrieval
at right inferior frontal, bilateral parietal and right occipito-parieto-temporal sites. Both
amplitude and latency reductions at these sites were larger over the right hemisphere,
particularly for the highest memory load condition.
Behavioural results are presented in section 5.1. This section is concerned primarily
with the relationship between response time, errors and memory load. Results pertaining

Results
to sustained and transient electrophysiological effects are presented in sections 5.2.1 and
5.2.2 respectively.
5.1 Behavioural data
Data averaged across all forty subjects was used in the analysis of behavioural data. In
addition, for illustrative purposes, the data for one individual subject was also analysed.
Response times were calculated for each memory load condition, and the statistical
significance of response time differences between the conditions was calculated using
the Student’s t test.
5.1.1 Individual subject
Figure 5.1 shows mean response times and standard deviations for all correctly
recognized target objects for an individual subject (DK), for each of the 3 memory load
conditions. This subject had no errors of omission (missed responses) and no errors of
commission (responses to non-targets). The mean response time to target objects was
505ms for a memory load of 1, 600ms for a memory load of 3, and 623ms for a memory
load of 5. Mean response times were significantly different for memory loads of 1 and 3
(t=-6.65; p<0.02), as were those for memory loads of 1 and 5 (t=-6.76; p<0.02).
Appendix C (Figure C-i) shows the equivalent chart for the practice memory tasks.
104

Results
Memory Loadm1 m3 m5
Mea
n R
espo
nse
Tim
e (m
sec)
0100200300400500600700800
m1 vs m3: t=-6.65; p<0.02m1 vs m5: t=-6.76; p<0.02m3 vs m5: NS
Figure 5.1 Mean response times for target objects for each memory load for an individual subject
Mean response time and standard deviation for target objects for each of the 3 memory load conditions. Data were taken from an individual subject (DK) performing task order 2 (ie. in the order m3, m1, m5). Results of unpaired t-tests for memory load comparisons are also shown. (NS = not significant)
5.1.2 Group data
From the mean response times for correctly recognized target objects for individual
subjects, the group (n=40) mean response times and standard deviations for each of the
3 memory conditions were calculated. Similarly, the mean numbers of errors of
omission and commission, with standard deviations, were calculated. These data are
shown in Figure 5.2. Practice memory tasks were similarly analysed, and results are
shown in Appendix C (Figure C.2). The group mean response time for target objects
was 480ms for a memory load of 1, 590ms for a memory load of 3, and 620ms for a
memory load of 5. Differences between group mean response times were statistically
significant for m1 vs m3 (t=-8.08; p<0.0001), m1 vs m5 (t=-9.41; p<0.0001), and m3 vs
m5 (t=-2.83; p<0.02). Group mean errors of omission and commission were generally
small and were not significantly different between memory load conditions.
105

Results
Mea
n R
espo
nse
Tim
e (m
sec)
0100200300400500600700800
Mea
n Fr
eque
ncy
of E
rror
s of
Om
issi
on
0.0
0.2
0.4
0.6
0.8
m1 vs m3: t=-8.08; p<0.0001m1 vs m5: t=-9.41; p<0.0001m3 vs m5: t=-2.83; p<0.02
Memory Loadm1 m3 m5
Mea
n Fr
eque
ncy
of E
rror
s of
Com
mis
sion
0.0
0.2
0.4
0.6
0.8
m1 vs m3: NSm1 vs m5: NSm3 vs m5: NS
m1 vs m3: NSm1 vs m5: NSm3 vs m5: NS
Figure 5.2 Mean response time and mean number of errors for target objects versus memory load for 40 subjects
Mean response time and standard deviation for target objects for each of the 3 memory load conditions (top). Charts are shown for numbers of errors of omission (middle) and commission (bottom). Results of paired t-tests for memory load comparisons are also displayed. (NS = not significant)
106

Results
5.2 Electrophysiological data
As explained in section 4.2, during the practice baseline task block, the 13Hz probe
flicker was switched-on half way through the block in order to determine whether the
driven 13Hz response was significantly different from the background or spontaneous
13Hz activity. As this issue is not central to this thesis, results are not provided here, but
are shown in Appendix D.
To examine the sustained modal effects associated with retrieval mode processes, an
averaging period of 40s was used. This is discussed in section 5.2.1. More transient
SSVEP amplitude and latency changes that occur with the rapidly changing patterns of
neural activity associated with object retrieval were investigated using an averaging
period of 1538ms (20 stimulus cycles). This gave an effective temporal resolution of
870ms. This analysis is explained in section 5.2.2.
5.2.1 Sustained effects: long averaging period
The following results were calculated using a long averaging period by averaging
Fourier coefficients across the entire 40s ‘task interval’ of the task blocks for the
baseline and each of the three memory load conditions. The resultant SSVEP
amplitudes and latencies were compared to investigate neural changes associated with
retrieval mode processes.
5.2.1.1 Individual subjects
Topographic maps of amplitude and phase were produced for all individual subjects.
There appeared to be a large inter-subject variation in both amplitude and phase for all
task conditions. However, averaging across subjects revealed statistically significant
effects. Data for an individual subject is provided in Figure 5.3 for illustrative purposes
(and see Appendix E, Figure E.1, for the practice task equivalent). On the topographic
maps for amplitude, warmer colours represent larger SSVEP amplitudes. On the
107

Results
topographic maps for phase, warmer colours represent a phase advance relative to the
13Hz probe stimulus. Conversely, cooler colours represent smaller SSVEP amplitudes
and a phase lag relative to the probe stimulus.
5.0 +3.1
0.0 Vυ -3.1 radian
baseline
load 1
load 3
load 5
amplitude phase
Figure 5.3 Sustained effects: Mean SSVEP amplitude and phase topography for the baseline task and each of the memory load conditions calculated with a long (40s) averaging period for an individual subject
Mean SSVEP amplitude and phase topography for the baseline condition and each of the memory load conditions, calculated across the entire 40s ‘task interval.’ Warmer colours represent larger SSVEP amplitudes and a phase advance relative to the 13Hz probe stimulus. Cooler colours represent smaller SSVEP amplitudes and a phase lag relative to the probe stimulus. Data for individual subject (CL).
108

Results
5.2.1.2 Group data
Amplitude and phase data for the 40 subjects were averaged as explained in section
4.8.4. The averaged results for the long averaging period analysis are shown for
illustrative purposes in Figure 5.4 (see Appendix E, Figure E.2 for the equivalent
practice tasks). Again, warmer colours in the topographic maps represent larger SSVEP
amplitudes and a phase advance relative to the baseline phase.
To examine neural changes associated with retrieval mode processes, topographic maps
depicting SSVEP amplitude and latency differences, and the associated Hotelling’s T
statistical topographic maps, were calculated for the averaged results. To investigate
retrieval mode processes, the overall average amplitude and latency values for the 3
memory load conditions were compared with those for the baseline condition (Figure
5.5). To examine the effect of memory load on retrieval mode processes, the SSVEP
values for the separate memory load conditions were compared with each other (Figure
5.6). In contrast to the colour conventions used in Figures 5.3 and 5.4, an alternative
convention has been used for the difference maps. For the amplitude difference maps,
warmer colours represent a reduced SSVEP amplitude for the average of the 3 memory
load conditions relative to the baseline condition (Figure 5.5), and for the higher
memory load condition relative to the lower (Figure 5.6). The topographic phase maps
show phase difference in terms of latency in milliseconds. Warmer colours indicate a
reduced latency for the average of the 3 memory load conditions relative to the baseline
condition (Figure 5.5), and for the higher memory load condition relative to the lower
(Figure 5.6). The Hotelling’s T statistic indicates the consistency of SSVEP differences
across the 40 subjects investigated. Warmer colours indicate a higher level of
significance. Contour lines depict T values that represent p values of 0.05, 0.01, 0.005
and 0.001 for a single comparison.
109

Results
1.5 +0.5
0.0 -0.5 radian
baseline
load 1
load 3
load 5
amplitude phase
Figure 5.4 Sustained effects: Mean SSVEP amplitude and phase topography for the baseline condition and each of the memory load conditions averaged across 40 subjects
Mean SSVEP amplitude and phase topography for the baseline condition and each of the memory load conditions, calculated across the entire ‘task interval.’ Data taken from averaged results across 40 subjects. Warmer colours represent larger SSVEP amplitudes and a phase advance relative to the phase of the baseline condition. Cooler colours represent smaller SSVEP amplitudes and a phase lag relative to the phase of the baseline condition. The effect of the phase adjustment results in a zero phase for the baseline task (see top).
110

Results
As suggested by Silberstein et al. (1995a), a correction factor of 5 was used to allow for
64 separate, but correlated, recording sites. Therefore, p=0.01 (ie. p=0.05/5=0.01) was
taken as the level for statistical significance for the difference between the average of
the 3 memory load conditions and the baseline task. For the 3 memory load
comparisons shown in Figure 5.6, p=0.001 (ie. p=0.05/(5x3)=0.0033) was used as a
conservative value for statistical significance.
The topographic map depicting the SSVEP amplitude differences for the average of the
3 memory load conditions with respect to the average for the baseline condition (Figure
5.5) indicates that the SSVEP amplitude was diffusely attenuated (warmer colours) at
all recording sites. The topographic map showing how the latency averaged over the 3
memory load conditions compared with the average latency for the baseline condition
indicates an SSVEP latency reduction (warmer colours) at most recording sites, and
particularly prominent reductions at right fronto-temporal sites. The Hotelling’s T
topographic map indicates that SSVEP differences were statistically significant at most
recording sites. Statistical significance appears greatest at right fronto-temporal sites.
AmplitudeDifference
LatencyDifference
Hotelling's T
0.0010.0050.01
0.05
+0.3 -6.1
+6.1
6.0
0.0
m(1
,3,5
) - b
ase
-0.3
Figure 5.5 Sustained effects: Topographic differences in SSVEP amplitude and latency between the average of the 3 memory load conditions and the baseline task averaged across 40 subjects
Topographic differences for SSVEP amplitude (normalized units) and latency (ms) between the average of the 3 memory load conditions and the average for the baseline task. Warmer colours represent reduced SSVEP amplitudes and latencies for the average of the 3 memory load conditions relative to the baseline task. Topographic maps for the Hotelling’s T statistic are shown representing the statistical strength of these differences. Four contours are shown on the scale representing T values for p=0.05, 0.01, 0.005 and 0.001 for a single comparison.
111

Results
Memory load conditions were also compared directly to investigate the effect of
increased memory load, and therefore task difficulty, on retrieval mode processes.
Topographic maps depicting amplitude and latency differences between the m3 and m1,
the m5 and m1, and the m5 and m3 conditions are shown in Figure 5.6. It can be seen
that with increases in memory load, SSVEP amplitude is attenuated, as indicated by
warmer colours. The topographic map for the m5 – m1 comparison, where the memory
load difference was greatest, shows an overall amplitude attenuation that is most
AmplitudeDifference
LatencyDifference
Hotelling's T
0.001
0.0050.01
0.05
0.001
0.0050.01
0.05
0.001
0.0050.01
0.05
+0.3
+0.3
+0.3
4.0
0.0
4.0
4.0
0.0
0.0
m3-
m1
m5-
m1
m5-
m3
+6.1
+6.1
+6.1
-6.1
-6.1
-6.1
-0.3
-0.3
-0.3
Figure 5.6 Sustained effects: Topographic differences in SSVEP amplitude and latency between memory load conditions averaged across 40 subjects
Topographic differences for SSVEP amplitude (normalized units) and latency (ms) between memory load conditions. Warmer colours represent reduced SSVEP amplitudes and latencies for high relative to low memory load conditions. Topographic maps for the Hotelling’s T statistic indicate the statistical strength of these differences. Four contours are shown on the scale representing T values of p=0.05, 0.01, 0.005 and 0.001 for a single comparison.
112

Results
prominent at occipital and right occipito-temporal sites. Topographic maps depicting
latency differences between the m3 and m1, and m5 and m1 conditions indicate an
increased latency (blue colour) over frontal regions for the higher memory load
condition relative to the lower. This pattern is reversed over posterior regions where the
higher relative to the lower memory load has produced a decreased latency (pink
colour). The topographic map for the m5 – m1 comparison, where the memory load
difference was greatest, shows a particularly prominent latency reduction at right
occipital sites. As indicated by the Hotelling's T statistical maps, SSVEP amplitude and
latency reductions at right occipital regions approached, but did not reach, statistical
significance (puncorrected=0.001). Similarly, differences at left central and right temporal
sites between the SSVEP response to the m5 and m3 conditions approached, but did not
reach, statistical significance (puncorrected=0.001).
5.2.2 Transient changes: short averaging period
To observe transient changes in SSVEP amplitude and phase associated with the
occurrence of specific events within the task, a high temporal resolution analysis was
performed. The following results were calculated by firstly extracting the SSVEP
Fourier coefficients from the EEG using a single cycle integration period. Signal-to-
noise ratio was increased by using a 20-cycle moving cosine, or Hanning, averaging
window. This gave an effective temporal resolution of 870ms. Five-second epochs of
20-cycle averaged Fourier coefficients, centred on the appearance of correctly identified
target and non-target objects, were then averaged for each condition. Data for the forty
individual subjects was then averaged to give a group averaged time series for
amplitude and phase data for each electrode site and for each memory load. The SSVEP
time averaged data for different memory loads, centred on the appearance of target
objects, can be compared to highlight neural regions involved in processes associated
with successful retrieval.
113

Results
5.2.2.1 Changes with memory load
The parietal ERP old/new effect has been noted most consistently in
electrophysiological studies into episodic retrieval processes. It was anticipated that in
this study, SSVEP load-dependent changes would also be apparent over parietal
regions. Figure 5.7 shows a five-second epoch of SSVEP amplitude data, averaged over
20 cycles, for a right parieto-temporal site (electrode 42). The data for each of the 3
memory load conditions is centred on the appearance of both non-target (upper graph)
and target (lower graph) objects. As can be seen in the upper graph, the SSVEP
amplitude underwent transient changes associated with the appearance of non-target
objects. The SSVEP amplitude was lowest just prior to the appearance of an object, and
then increased, peaking 1s after the object’s appearance. The 20-cycle time-averaged
data for non-target objects shows no apparent correlation between the SSVEP amplitude
and memory load. When a five-second epoch of 20-cycle time-averaged SSVEP
amplitude data is centred on the appearance of target objects, quite a different pattern is
seen. The transient changes in amplitude appear relatively similar across all 3 memory
load conditions for times up to 500ms before target presentation and beyond 1500ms
after target presentation. Around the time of target presentation, however, the
differences between the SSVEP amplitudes associated with each of the 3 memory load
conditions markedly increase. During target presentation, at this right hemisphere
parieto-temporal electrode site, an increase in memory load was associated with a
decrease in SSVEP amplitude.
Figure 5.8 shows a five-second epoch of 20-cycle time-averaged SSVEP phase data
recorded at the same right hemisphere parieto-temporal electrode. This is also centred
on the appearance of either non-target (upper graph) or target (lower graph) objects for
each of the 3 memory load conditions. For non-target objects, variations similar to the
amplitude variations were evident. The SSVEP phase value was lowest, indicating a
phase lag, just prior to the appearance of the non-target and peaked, indicating a phase
advance, 1000ms later. The SSVEP phase data for non-target objects appeared to be
similar for each of the 3 memory load conditions. The equivalent phase data for target
objects (lower graph) also shows a rhythmic pattern over the 5s epoch. Before the
appearance of a target object, the pattern is similar to that observed for non-target
114

Results
objects. Differences between the phase values for each of the memory loads are
relatively small. However, following the appearance of a target, differences markedly
increased. Here, the higher the memory load, the more advanced the phase. Phase
values peaked at about 800ms after target appearance, and were approximately 0 radian
for a memory load of 1, 0.1 radian for a memory load of 3, and 0.2 radian, for a memory
load of 5.
Results obtained at the equivalent left hemisphere parieto-temporal site (electrode 38)
are shown in Figures 5.9 and 5.10. Left parieto-temporal variations in amplitude and
phase associated with the presentation of non-target objects are similar to those on the
right. However, variations associated with the presentation of target objects recorded at
the left hemisphere site differ from those recorded at the right, in that amplitude appears
less strongly correlated with memory load, and phase appears to be uncorrelated.
115

Results
Time (seconds)-2 -1 0 1 2
Nor
mal
ised
Am
plitu
de
0.6
0.8
1.0
1.2
Time (seconds)-2 -1 0 1 2
Nor
mal
ised
Am
plitu
de
0.6
0.8
1.0
1.2
SSVEP Amplitude - Non-Target
(Non
-)Ta
rget
Bla
nk
Non
-Targe
t
Bla
nk
(Non
-)Ta
rget
0.4
sec
post
non
-targ
et
m1m3m5
Res
pons
e
SSVEP Amplitude - Target
Non
-Tar
get
Bla
nk
Target
Bla
nk
Non
-Tar
get
0.4
sec
post
targ
et
m1m3m5
Figure 5.7 Normalized SSVEP amplitude time series for each of the memory load conditions calculated across 40 subjects
Normalized SSVEP amplitude time series with a temporal resolution of 870ms, across 40 subjects, for each of the memory load conditions. Time series data were taken from a right parieto-temporal site (electrode 42). Time zero represents the presentation of either a target or non-target object. For target presentations, the variation in the range of mean response times for the subjects is shown in grey, and the point selected for topographic mapping of differences (Figures 5.11 and 5.12) is shown by the vertical dotted line (between presentation and response).
116

Results
Time (seconds)-2 -1 0 1 2
Pha
se (r
adia
ns)
-0.4
-0.2
0.0
0.2
0.4
Time (seconds)-2 -1 0 1 2
Phas
e (r
adia
ns)
-0.4
-0.2
0.0
0.2
0.4
SSVEP Phase - Non-Target
(Non
-)Ta
rget
Blan
k
Non
-Targe
t
Bla
nk
(Non
-)Ta
rget
0.4
sec
post
non
-targ
et
m1m3m5
Res
pons
e
SSVEP Phase - Target
Non
-Tar
get
Bla
nk
Targ
et
Bla
nk
Non
-Tar
get
0.4
sec
post
targ
et
m1m3m5
Figure 5.8 SSVEP phase time series for each of the memory load conditions calculated across 40 subjects
SSVEP phase time series with a temporal resolution of 870ms, across 40 subjects, for each of the memory load conditions. Time series data were taken from a right parieto-temporal site (electrode 42). Time zero represents the presentation of either a target or non-target object. For target presentations, the variation in the range of mean response times for the subjects is shown in grey, and the point selected for topographic mapping of differences (Figures 5.11 and 5.12) is shown by the vertical dotted line (between presentation and response).
117

Results
Time (seconds)-2 -1 0 1 2
Nor
mal
ised
Am
plitu
de
0.6
0.8
1.0
1.2
Time (seconds)-2 -1 0 1 2
Nor
mal
ised
Am
plitu
de
0.6
0.8
1.0
1.2
SSVEP Amplitude - Non-Target
(Non
-)Ta
rget
Bla
nk
Non
-Targe
t
Bla
nk
(Non
-)Ta
rget
0.4
sec
post
non
-targ
et
m1m3m5
Res
pons
e
SSVEP Amplitude - Target
Non
-Tar
get
Bla
nk
Target
Bla
nk
Non
-Tar
get
0.4
sec
post
targ
et
m1m3m5
Figure 5.9 Normalized SSVEP amplitude time series for each of the memory load conditions calculated across 40 subjects
Normalized SSVEP amplitude time series with a temporal resolution of 870ms, across 40 subjects, for each of the memory load conditions. Time series data were taken from a left parieto-temporal site (electrode 38). Time zero represents the presentation of either a target or non-target object. For target presentations, the variation in the range of mean response times for the subjects is shown in grey, and the point selected for topographic mapping of differences (Figures 5.11 and 5.12) is shown by the vertical dotted line (between presentation and response).
118

Results
Time (seconds)-2 -1 0 1 2
Phas
e (r
adia
ns)
-0.4
-0.2
0.0
0.2
0.4
Time (seconds)-2 -1 0 1 2
Phas
e (r
adia
ns)
-0.4
-0.2
0.0
0.2
0.4
SSVEP Phase - Non-Target
(Non
-)Ta
rget
Bla
nk
Non
-Targe
t
Blan
k
(Non
-)Ta
rget
0.4
sec
post
non
-targ
et
m1m3m5
Res
pons
eSSVEP Phase - Target
Non
-Tar
get
Bla
nk
Target
Blan
k
Non
-Tar
get
0.4
sec
post
targ
et
m1m3m5
Figure 5.10 SSVEP phase time series for each of the memory load conditions calculated across 40 subjects
SSVEP phase time series with a temporal resolution of 870ms, across 40 subjects, for each of the memory load conditions. Time series data were taken from a left parieto-temporal site (electrode 38). Time zero represents the presentation of either a target or non-target object. For target presentations, the variation in the range of mean response times for the subjects is shown in grey, and the point selected for topographic mapping of differences (Figures 5.11 and 5.12) is shown by the vertical dotted line (between presentation and response).
119

Results
To observe the topography of the SSVEP amplitude and phase changes, differences
between memory loads were calculated for each of the 64 electrode sites, and
topographic maps were produced. It is possible to produce a topographic map for every
time point in the time-averaged data. However, it had been anticipated that the
recognition stage of object retrieval would have occurred within the interval between
object presentation and response, and therefore a time point within this interval was
examined. The precise location of this time point is not critical as the temporal
resolution used to calculate the SSVEP Fourier coefficients was 870ms, and therefore
the speed of topographic changes was of this order of magnitude. The average response
time for a memory load of 1 was 480ms, and a time point approximately 400ms after
either target or non-target appearance was selected and mapped. A time of 400ms after
target appearance was selected because 400ms is shorter than the average response time
for a memory load of 1, and therefore, SSVEP differences between memory loads were
less likely to have resulted from subject response. The same time point was also used
for mapping SSVEP differences for non-target objects.
Figure 5.11 shows SSVEP amplitude and phase differences between memory load
conditions m3 and m1, m5 and m1, and m5 and m3 at 400ms after target appearance.
Differences for non-target objects are shown in Figure 5.12. Warmer colours in the
amplitude difference maps indicate SSVEP amplitude attenuation for the higher relative
to the lower memory load condition. Conversely, cooler colours indicate SSVEP
amplitude increases for the higher relative to the lower memory load condition. Phase
differences in radians were converted to latency differences in milliseconds. Warmer
colours in the latency difference maps indicate a reduced SSVEP latency for the higher
relative to the lower memory load condition. Conversely, cooler colours indicate an
increase in SSVEP latency for the higher relative to the lower memory load condition.
The Hotelling’s T parameter for these differences was also calculated. This indicated
the consistency of SSVEP differences between memory load conditions across the 40
subjects investigated. Warmer colours indicate a higher level of significance. Contour
lines depict T values that represent p values of 0.05, 0.01, 0.005 and 0.001 for a single
comparison. Using the correction factor of 5 (for 64 separate but correlated electrodes)
suggested by Silberstein et al. (1995a), and 3 for the number of conditions, p=0.001 (ie.
120

Results
p=0.05/(5x3)=0.0033), was used as a conservative value for statistical significance (see
Methods chapter, section 4.8.6).
For target objects (Figure 5.11), SSVEP differences between memory load conditions
were apparent for both amplitude and latency. Again, warmer colours indicate lower
amplitudes and shorter latencies, and cooler colours indicate higher amplitudes and
longer latencies. The amplitude maps indicate an amplitude reduction for the higher
memory load conditions relative to the lower at most electrode sites. Amplitude
reductions for the m3 relative to m1 condition, and m5 relative to m1 condition, were
largest over the junction of the right occipital, parietal, and temporal regions, and
extended into right and left parietal, and left temporal regions. These reductions
appeared to be more extensive for the m5 relative to the m1 condition than for the m3
relative to the m1 condition. Latency maps indicate a latency reduction for the higher
memory load relative to the lower memory load conditions at most electrode sites.
Latency reductions were largest for the m3 relative to the m1 condition over parieto-
occipital regions, and for the m5 relative to m1 condition over parieto-occipital and
frontal regions. For the comparison of the m3 with the m1 condition, the Hotelling’s T
parameter indicates that SSVEP amplitude and latency differences are statistically
significant over right occipito-temporal and bilateral parietal regions. Differences
between the m5 and m1 conditions are statistically significant over additional bilateral
parietal, right parieto-temporal, and right inferior frontal regions, and represent a
reduced SSVEP amplitude and latency for the higher memory load condition. The m5
versus m3 comparison for target objects showed the same trend for amplitude and
latency differences, that is, attenuation in amplitude and reduction in latency with
increased memory load, although these differences did not reach statistical significance.
Compared with target objects, the SSVEP differences between memory load conditions
for non-target objects (Figure 5.11) were generally much smaller for both amplitude and
latency. There did not appear to be a correlation between amplitude or latency
reductions and increases in memory load. Hotelling’s T topographic maps indicated that
SSVEP differences were not statistically significant.
121

Results
AmplitudeDifference
LatencyDifference
Hotelling's T
0.001
0.0050.01
0.05
0.001
0.0050.01
0.05
0.001
0.0050.01
0.05
+0.4
+0.4
+0.4
4.0
0.0
4.0
4.0
0.0
0.0
m3-
m1
m5-
m1
m5-
m3
+4.9
+4.9
+4.9
-4.9
-4.9
-4.9
-0.4
-0.4
-0.4
Figure 5.11 Topographic differences in SSVEP amplitude and latency between memory load conditions for target objects and Hotelling's T statistic (averaged across 40 subjects)
Topographic differences in SSVEP amplitude (normalized units) and latency (ms) between memory load conditions at 400ms after the appearance of target objects. Warmer colours represent reduced SSVEP amplitudes and latencies for high relative to low memory load conditions. Topographic maps of the Hotelling’s T parameter are also shown indicating the statistical strength of these differences. Four contours are shown on the scale representing T values of p=0.05, 0.01, 0.005 and 0.001 for a single comparison.
122

Results
AmplitudeDifference
LatencyDifference
Hotelling's T
0.001
0.0050.01
0.05
0.001
0.0050.01
0.05
0.001
0.0050.01
0.05
+0.4
+0.4
+0.4
4.0
0.0
4.0
4.0
0.0
0.0
m3-
m1
m5-
m1
m5-
m3
+4.9
+4.9
+4.9
-4.9
-4.9
-4.9
-0.4
-0.4
-0.4
Figure 5.12 Topographic differences in SSVEP amplitude and latency between memory load conditions for non-target objects and Hotelling's T statistic (averaged across 40 subjects)
Topographic differences in SSVEP amplitude (normalized units) and latency (ms) between memory loads at 400ms after the appearance of non-target objects. Warmer colours represent reduced SSVEP amplitudes and latencies for high relative to low memory load conditions. Topographic maps of the Hotelling’s T parameter are also shown indicating the statistical strength of these differences. Four contours are shown on the scale representing T values of p=0.05, 0.01, 0.005 and 0.001 for a single comparison.
123

Results
5.2.2.2 Targets versus non-targets
In section 5.2.2.1, transient patterns of neural activity associated with successful
retrieval processes were investigated by varying the memory load in an episodic
memory retrieval task. With this approach, it was anticipated that the increased
difficulty associated with the retrieval of more targets would have an effect on the
SSVEP response that would help reveal the neural regions involved in successful
retrieval processes.
Neural regions involved in successful retrieval were also investigated by directly
comparing the SSVEP response to target and non-target objects. This type of direct
comparison has been used extensively in electrophysiological studies (see Chapter 2). In
this study, however, a comparison of the SSVEP response to target and non-target
identification is more problematic because targets prompted a motor response, whereas
non-targets did not. Therefore, preparation for the motor response could have been
responsible for differences in the SSVEP patterns observed for targets and non-targets.
On the other hand, it is possible for a number of reasons, that SSVEP changes prior to
the motor response were not contaminated by response processes, and therefore are
reflections of neural activity associated with successful recognition. These and other
issues relating to the comparison of target and non-target objects will be discussed in
more detail in Chapter 6.
Notwithstanding possible confounding factors, SSVEP differences between target and
non-target objects were calculated. Differences were calculated separately for each of
the 3 memory load conditions. Topographic maps in Figure 5.13 indicate amplitude and
latency differences between the SSVEP response to correctly identified target and non-
target objects at 450ms and 700ms after object appearance. The 450ms time point,
which occurs between object appearance and the mean response time, was selected
because it was anticipated that the recognition stage of object retrieval processes would
occur within this interval. Differences calculated at the 700ms time point showed that
for a memory load of 5, significant frontal differences between the SSVEP response to
targets and non-targets developed after the mean motor response time.
124

Results
AmplitudeDifference
LatencyDifference
Hotelling's T
0.001
0.0050.01
0.05
0.001
0.0050.01
0.05
0.001
0.0050.01
0.05
0.001
0.0050.01
0.05
0.001
0.0050.01
0.05
0.001
0.0050.01
0.05
700 ms
700 ms
700 ms
450 ms
450 ms
450 ms +0.4
+0.4
+0.4
+0.4
+0.4
+0.4
4.0
0.0
4.0
4.0
4.0
4.0
4.0
0.0
0.0
0.0
0.0
0.0
m1t
- m
1nt
m5t
- m
5nt
m3t
- m
3nt
+4.9
+4.9
+4.9
+4.9
+4.9
-4.9
-4.9
-4.9
-4.9
-4.9
-4.9
-0.4
-0.4
-0.4
-0.4
-0.4
-0.4 +4.9
Figure 5.13 Topographic differences in SSVEP amplitude and latency between correctly identified target and non-target objects, averaged separately for each memory load condition: pooled across 40 subjects
Topographic differences in SSVEP amplitude (normalized units) and latency (ms) between target and non-target objects at times 450ms and 700ms following the presentation of objects. Warmer colours represent reduced SSVEP amplitudes and latencies for target relative to non-target objects. Conversely, cooler colours represent increased SSVEP amplitudes and latencies for target relative to non-target conditions. Topographic maps of the Hotelling’s T statistic are also shown representing the statistical strength of these differences. Four contours are shown on the scale representing T values of p=0.05, 0.01, 0.005 and 0.001 for a single comparison.
125

Results
Warmer colours indicate both amplitude and latency reductions for target relative to
non-target objects. Conversely, cooler colours indicate both amplitude and latency
increases for target relative to non-target objects. The Hotelling’s T value, calculated for
both time points, reflects the consistency of SSVEP differences between target and non-
target data across the 40 subjects investigated. Warmer colours indicate higher levels of
statistical significance. Contour lines depict T values representing p values of 0.05,
0.01, 0.005 and 0.001 for a single comparison. Using the correction suggested by
Silberstein et al. (1995a), a factor of 5 for spatial dimensionality and 6 for the number of
conditions (ie. 0.05/(5x6)=0.0017), p=0.001 was used as a conservative level for
statistical significance (see Methods chapter, section 4.8.6).
For a memory load of 1, the topographic maps show that differences between target and
non-target amplitude and latency values were similar at both 450ms and 700ms after the
appearance of the object. The cooler colours of the amplitude maps indicate that the
SSVEP amplitude was greater for target objects than for non-target objects. Latency
differences between the target and non-target conditions were, however, very small, as
indicated by a prevalence of yellow/green colour. The Hotelling’s T parameter
approaches, but does not reach, significance (puncorrected=0.001) over occipito-temporal
regions.
For a memory load of 3, the pattern of amplitude and latency differences is quite
different. A predominance of warmer colours on topographic maps of amplitude
difference indicates an attenuation of the SSVEP amplitude for target objects relative to
non-target objects at most sites. This attenuation is greatest at parieto-temporal sites
bilaterally. The predominance of warmer colours on the difference maps for latency
indicates that latencies were generally reduced for targets relative to non-targets. The
most prominent latency reductions are apparent over left parietal regions, particularly
for the earlier 450ms time point. Hotelling’s T values are highest over parietal regions,
once again approaching, but not reaching, significance (puncorrected=0.001).
Amplitude and latency differences for a memory load of 5 were somewhat similar to
those for a memory load of 3. Again, there were SSVEP amplitude and latency
reductions, indicated by warmer colours, for target relative to non-target objects. The
126

Results
amplitude reductions, however, appeared to be more frontal and were more marked at
the later (700ms) time point. Hotelling’s T scores over pre-frontal regions indicate that
SSVEP amplitude and latency differences in these regions are statistically significant
(p<0.001) at the 700ms time point.
127

Chapter 6 Discussion
The discussion of findings for the present study is divided into five main sections.
Section 6.1 contains a brief account of behavioural results. Behavioural data were
analysed in order to determine whether increases in memory load led to corresponding
increases in task difficulty. Response time data was also required so that time points
indicative of the mental processes under investigation could be selected for data
analysis. Section 6.2 focuses on sustained SSVEP effects over frontal and right
hemisphere regions when memory load conditions are compared with the baseline
condition. These results are discussed in relation to PET, fMRI and DC-ERP findings,
namely, that right frontal regions have been associated with the establishment and
maintenance of a retrieval mode. Section 6.3 is concerned with transient effects, in
particular, load-dependent SSVEP changes associated with successful memory retrieval.
The observed SSVEP effects over parietal and right frontal regions are discussed
primarily in relation to findings from studies into the ERP old/new effect. Section 6.4
also deals with transient SSVEP changes; however here, the direct comparison of the
response to target and non-target objects is examined. Findings of frontal effects will
again be discussed in relation to the ERP old/new findings that were extensively
reviewed in Chapter 2. Finally, in section 6.5, major conclusions relating to this study
will be presented.
6.1 Behavioural results: the effect of increasing memory load on retrieval
accuracy and response time
In the present study, subjects were required to memorize 1, 3 or 5, two-dimensional
abstract objects during the study phase for the corresponding memory load condition.
These objects had to be retrieved from memory in the subsequent retrieval phase.
Recognition accuracy was high for all three memory load conditions, as evidenced by
the very low error scores across the 40 subjects. In addition, performance appeared to
improve with practice (compare Figure 5.2 with Figure C.2).

Discussion
The group mean response times increased with increases in memory load. The mean
response times for 1, 3 and 5 memory load conditions were approximately 480, 590 and
620ms respectively. Furthermore, each of the 40 subjects showed an increased response
time with increased memory load. There was considerable variability in response times
because some subjects responded slowly to all tasks, while others responded more
quickly overall. However, Student’s t tests showed that the differences in response times
associated with the different memory load conditions were highly significant.
In a post-recording interview, all subjects reported that increases in memory load led to
noticeable increases in task difficulty. That is, the task with 3 objects to be recognized
was more difficult than the task with 1 object, and the task with 5 objects was more
difficult than the task with 3. In the light of the aforementioned behavioural findings
and these personal accounts, it seems reasonable to assume that an increase in memory
load resulted in increased retrieval effort. Retrieval effort is a term that has been used to
refer to ‘the mobilization of processing resources in service of a retrieval attempt’
(Rugg and Wilding 2000, p. 110). While task difficulty is a more general term used to
describe the overall complexity of the task, retrieval effort involves transient processes
that are specifically engaged whilst determining whether an object is new or old.
The study-test task was specifically designed so that subjects would experience
increases in difficulty with increases in memory load. This was especially important
since one of the main aims of the study was to identify neural regions associated with
successful retrieval by increasing the demand on these regions. In other words, to
remember whether a presented item was studied or not should require more effort with
higher memory loads. As explained in the Methods section (section 4.2), tasks were also
designed to facilitate the encoding of objects into a more permanent store by
incorporating both multiple presentations of the studied items and relatively long
presentation times. Therefore, a high retrieval accuracy for all three memory load
conditions was not unexpected. Increased response times with increased load were also
not unexpected in the light of subjects’ personal accounts and the fact that higher
memory load tasks required the scanning of more items in memory. Sternberg (1969)
first demonstrated that response time increases linearly with the number of items stored
in short-term memory. It also appears that memory search processes are reflected in the
129

Discussion
EEG. For example, Stuss et al. (1986) showed that both the N400 amplitude and the
response time increase as the number of pictures to be named increases, and Noldy-
Cullum and Stelmack (1987) suggested that the ERP old/new effect reflects an
‘exhaustive search’ of all items in the memory set to determine whether a presented
item is old or new.
In the present experimental design, as greater numbers of target objects had to be
remembered, they also had to be differentiated from a greater number of distractor
objects of similar appearance. That is, subjects had to make a judgement based on
perceptual detail. In a similar way, Ranganath and Paller (2000) required their subjects
to make specific perceptual judgements when they had to recall whether line drawings
of objects were the same size as those studied. The authors argued that such memory
monitoring processes are an essential component of episodic memory, and furthermore,
may also be important in conscious recollection processes. It is suggested that, in the
present study, increases in response time with increasing memory load were due, in part,
to a reliance on perceptual detail for the identification of targets and non-targets.
In summary, behavioural results, particularly the response time data, served as
confirmation that increases in memory load led to increases in task difficulty.
Furthermore, given also the personal accounts that difficulty increased with memory
load, it seems reasonable to assume that retrieval effort also increased with memory
load.
6.2 Sustained SSVEP amplitude and latency changes and retrieval mode
Retrieval mode is a term specifically used in relation to episodic retrieval, and has been
defined by Tulving (1983) as a tonically maintained mental state that is necessary for
the successful retrieval of items from episodic memory. Studies using PET, fMRI and
DC-ERP techniques to investigate retrieval mode have compared patterns of activity
during the performance of episodic and non-episodic retrieval tasks. In the present
study, to identify neural regions involved in sustained retrieval mode processes, the
amplitude and latency of the SSVEP were averaged over a 40s period, during which,
130

Discussion
subjects were required to recognize previously studied items. The sustained SSVEP
effects engendered during the episodic retrieval tasks were compared with those
engendered during a low vigilance baseline task. In addition, this study attempts to
determine effects of task difficulty on retrieval mode processes by incorporating
retrieval tasks with different memory loads.
In section 6.2.1, the topography of sustained SSVEP amplitude and latency values will
be discussed, particularly in relation to the first hypothesis of this study (see section
3.7), namely, that sustained SSVEP amplitude and latency changes reflecting retrieval
mode will be most prominent over right frontal regions. While the topography of
SSVEP effects is central to this study, the direction of SSVEP effects, that is, whether
the SSVEP amplitude and SSVEP latency increase or decrease relative to a baseline
task, is also an important issue. Thus, the interpretation of SSVEP amplitude and
latency changes will be discussed in section 6.2.2.
6.2.1 The topography of sustained SSVEP changes
The first hypothesis of this study states that sustained SSVEP changes, reflecting
retrieval mode processes, will be most prominent over right frontal regions. When the
sustained SSVEP response to the episodic memory retrieval condition was compared
with the sustained response to the baseline condition, the most prominent and most
statistically significant SSVEP amplitude and latency reductions were observed over
right frontal regions. While sustained SSVEP amplitude reductions were fairly uniform
over most regions, sustained SSVEP latency reductions were clearly asymmetric and
largest over right fronto-temporal regions. The topography of these reductions is
consistent with findings obtained using other functional imaging techniques where
episodic and non-episodic memory tasks have been compared. As indicated in Chapter
2, a majority of PET and fMRI studies have reported prominent activations within right
prefrontal regions (Cabeza and Nyberg 2000). Moreover, most of these studies reported
greater right frontal than left frontal activation. This is in accordance with the HERA
model (Nyberg et al. 1996a; Tulving et al. 1994a) in which episodic memory encoding
and semantic retrieval is lateralized to left prefrontal regions, whereas episodic memory
131

Discussion
retrieval is lateralized to right prefrontal regions. In addition, Duzel et al. (1999), using
PET methods, showed that the right prefrontal cortex, BA10, was activated for an
episodic retrieval task relative to a semantic retrieval task (see Chapter 2). In the same
study, using DC-ERP techniques, sustained positive-going ERP activity was observed
over right frontal regions. Using source localization techniques, the authors found that
this activity was generated in the right frontopolar region, that is, in a region that
overlaps with BA10. In the present study, while the precise source of SSVEP amplitude
and latency reductions was not investigated, the observed patterns are suggestive of
underlying right fronto-temporal activity. The use of source localization techniques,
such as those used by Duzel et al. (1999), could help identify the neural origin(s) of
these sustained SSVEP changes.
While PET and fMRI studies have consistently yielded evidence of right prefrontal
activations with retrieval mode processes, other neural regions have also been
implicated. Activations within medial temporal, medial parieto-occipital, lateral parietal,
anterior cingulate, occipital, and cerebellar regions have also been reported in PET and
fMRI studies (Cabeza and Nyberg 2000). In the present study, when the SSVEP values
were averaged over the entire ‘task interval,’ the episodic retrieval condition relative to
the baseline condition showed significant SSVEP amplitude and latency reductions over
frontal and right hemisphere regions. The involvement of right parieto-temporal regions,
in particular, may reflect spatial and perceptual processes involved in recognizing target
objects presented amongst a larger number of distractor objects. Moreover, it is
suggested that the bias towards right hemisphere sites in this study is consistent with the
relatively abstract pictorial nature of the stimulus objects used. While the largest
sustained SSVEP amplitude and latency reductions occurred over right frontal regions,
left frontal SSVEP amplitude and latency reductions were also highly statistically
significant. As indicated in the electrophysiological literature, the involvement of left
frontal sites has been associated with task difficulty and effort in performing an episodic
retrieval task (Rugg et al. 2000). Nolde et al. (1998) suggested that while the right
prefrontal cortex is activated while subjects remember events, left prefrontal regions are
activated when more complex and reflective retrieval is required.
The investigation of neural regions involved in retrieval mode processes, as discussed
132

Discussion
above, was conducted by comparing the overall average of the SSVEP amplitude and
latency values across all three memory load conditions with the average SSVEP
amplitude and latency values for the baseline condition. A secondary analysis, also
utilizing SSVEP amplitude and latency values averaged over the 40s ‘task interval,’ was
performed to investigate the effect of task difficulty on sustained retrieval mode
processes. This was done by directly comparing the SSVEP amplitude and latency data
for the three individual memory load conditions, that is, for m3 with m1, m5 with m1,
and m5 with m3. SSVEP amplitude and latency differences for these comparisons were
statistically significant at right occipital sites for all comparisons, and at left central and
right temporal sites for the comparison of the m5 with the m3 condition. The largest
differences occurred between the m5 and m1 conditions. They were located over right
occipital regions, and were characterized by SSVEP amplitude and latency reductions
for the m5 relative to the m1 condition. In contrast to the differences between the
sustained SSVEP responses associated with the episodic memory and baseline
conditions, sustained right frontal SSVEP differences between memory load conditions
were not statistically significant. Given the suggested role of right frontal regions in
sustained retrieval mode processes, the absence of load-dependent differences over right
frontal regions suggests that retrieval mode processes are not dependent on memory
load, and therefore on task difficulty. Load-dependent amplitude and latency reductions
over occipital regions are consistent with previous findings indicating such reductions
are associated with visual vigilance (Silberstein et al. 1990b), heightened visual
attention (Silberstein et al. 1996) and object encoding (Silberstein et al. 2001). In the
present study, it seems reasonable to suppose that a more difficult task would have
promoted higher levels of attention. It is therefore suggested that the sustained load-
dependent SSVEP amplitude and latency reductions over occipital regions reflect
heightened visual attention and are not unique to retrieval mode processes.
The effect of memory load on retrieval processes can also be discussed in terms of
retrieval effort. As discussed in section 6.1, all subjects reported that the larger the
memory load, the more difficult was the retrieval task. Subjects also stated that they felt
that it took longer to respond to targets as the memory load increased, and such personal
accounts were supported by the response time data. It therefore seems reasonable to
suppose that retrieval effort also increased with increasing memory load. Rugg and
133

Discussion
Wilding (2000) suggested that retrieval effort is an item-specific process, defined as ‘the
mobilization of processing resources in service of a retrieval attempt’ (p. 110).
However, PET findings have suggested that retrieval effort is manifest as a sustained
task-related effect. A number of PET studies have reported frontal activations,
particularly on the left side, for more effortful relative to less effortful tasks (eg. Cabeza
and Nyberg 2000). Left frontal activation has also been described as being more
common in older adults than young adults, and has therefore been interpreted as a
compensatory effect (Cabeza et al. 1997a). These findings, however, do not rule-out the
possibility that left frontal activations reflect the summed activity of transient item-
related effects. In the present study, frontal differences in the SSVEP response to
different memory load conditions, reflecting differences between more effortful and less
effortful tasks, were not statistically significant. It is therefore suggested that retrieval
effort is not manifest as a sustained task-related process. It is possible, however, that
task-related SSVEP amplitude and latency differences between different memory loads,
and therefore different retrieval effort requirements, may have been too small to detect
using the SSPT technique. This is in contrast to the larger, statistically significant,
differences that were observed for the overall average of these memory load conditions
relative to the baseline condition. However, the explanation that the task-related SSVEP
differences were too small seems unlikely in light of the significant load-dependent
transient differences that were found (see section 6.3). These transient SSVEP
amplitude and latency differences were calculated using a much smaller number of
Fourier coefficients, therefore giving a much smaller signal to noise ratio, yet
differences between memory loads were significant.
6.2.2 Interpretation of sustained SSVEP amplitude and latency reductions
Thus far, SSVEP effects have been discussed in terms of topography, or scalp location.
Another important aspect of the data is the nature of the effects, that is, whether a
comparison between particular conditions produces amplitude and latency increases or
reductions. In this study, to examine sustained retrieval mode processes, the SSVEP
response to the retrieval condition was compared to the response to the baseline
condition. This comparison yielded amplitude and latency reductions for the retrieval
134

Discussion
condition relative to the baseline condition. To examine the effect of memory load on
retrieval processes, the SSVEP response to a higher memory load was compared
directly with the response to a lower memory load. Again, amplitude and latency
reductions were obtained. The interpretation of these SSVEP amplitude and latency
reductions will be discussed below.
As already mentioned, an attenuation of the 13Hz SSVEP amplitude has been
associated with a state of heightened attention in a number of experimental situations.
These include the heightened attention phases during the performance of a visual
vigiliance task (Silberstein et al. 1990b) and a continuous performance task (Silberstein
et al. 1996), and the ‘intake’ period of an object working memory task (Silberstein et al.
2001). Pipingas et al. (2000) showed graded SSVEP amplitude changes during a visual
attention task. Subjects who responded faster to the appearance of target letters in a
continuous performance task also showed a greater attenuation in SSVEP amplitude
prior to target presentation than did subjects who responded more slowly. The authors
equated a greater attenuation of the SSVEP amplitude with a heightened attention prior
to target presentation that resulted in faster responses. Silberstein et al. (1995b) has
suggested that the attenuation of the 13 Hz SSVEP amplitude associated with increased
regional brain activity may be related to the alpha event-related desynchronization
(ERD) that has been shown to index regional cortical activity in various cognitive and
motor tasks (Pfurtscheller and Aranibar 1977b; Pfurtscheller and Klimesch 1990).
More recently, however, SSVEP amplitude increases have been observed during periods
of mental effort occurring at certain phases of a cognitive task. For example, an
amplitude increase was observed during the ‘hold’ period of a spatial working memory
task (Silberstein 1998). A similar amplitude increase was also observed during the
‘hold’ period of an object working memory task (Silberstein et al. 2001). However,
during the ‘intake’ or encoding period of the same task, an amplitude decrease was
observed and was attributed to perceptual processes.
In the light of these apparently inconsistent effects of mental effort on the SSVEP
amplitude, it was posited that the type of SSVEP amplitude response depends on the
exact nature of the cognitive activity being undertaken (Silberstein et al. 2001). There
135

Discussion
appear to be similarities between the amplitude of the SSVEP response and the
amplitude of the alpha response noted by Ray and Cole (1985), namely, that attending
to visual targets engendered a decrease in alpha amplitude, whereas attending to mental
imagery engendered an increase.
In the present study, the SSVEP amplitude shows a sustained reduction for the overall
average of the three memory load conditions relative to the average of the baseline
condition. This amplitude reduction, in conjunction with a pattern of more localized
latency changes, was considered a reflection of sustained retrieval mode processes.
Because the amplitude reductions were diffuse and non-localized compared with the
latency reductions, these may not have reflected retrieval mode processes. Instead, the
amplitude reductions may have reflected non-specific aspects of the task, such as
attention or those associated with task difficulty. When the SSVEP responses to the
various individual memory load conditions were compared directly to examine the
effect of task difficulty on retrieval mode processes, the largest differences occurred at
right occipital sites and were characterized by SSVEP amplitude and latency reductions
for a higher relative to a lower memory load. Thus the observed occipital amplitude
reductions may well be attentional effects. This would be consistent with the previous
SSVEP findings showing amplitude reductions associated with perceptual attention.
The observed latency effects may specifically reflect retrieval mode processes, given the
correspondence of the location of SSVEP latency reductions over right frontal regions
to findings from neuroimaging and electrophysiological studies. Silberstein et al. (1996)
observed that faster response times in a continuous performance task were associated
with greater transient decreases in prefrontal SSVEP latency, and suggested that latency
reflects speed of information processing. The transient latency reductions were
considered to reflect transient increases in neural information processing speed resulting
from increased coupling strength between neural populations. In the present study,
sustained latency reductions occurred over right frontal regions, and in-line with the
proposal of Silberstein et al (1996), it is suggested that these are a reflection of
increased coupling between those regions required in maintaining a retrieval mode.
In summary, the sustained SSVEP effects for the retrieval task relative to the baseline
136

Discussion
task were characterized by diffuse non-localized amplitude reductions, and latency
reductions that were more prominent and were localized over right fronto-temporal
regions. While the source of these reductions was not investigated in the present study,
they occurred in the vicinity of those underlying cortical regions believed to be involved
in retrieval mode processes that have been more precisely localized in PET, fMRI, and
DC-ERP studies using source localization techniques. It is therefore proposed that
SSVEP amplitude and latency reductions at right fronto-temporal regions reflect
sustained retrieval mode processes. Furthermore, increases in task difficulty did not
appear to significantly alter this pattern of latency reduction. It is therefore also
proposed that retrieval mode processes are independent of task difficulty or retrieval
effort.
6.3 Transient SSVEP amplitude and latency changes: the effect of memory load
on successful memory retrieval
In the previous section, sustained SSVEP amplitude and latency effects were discussed
in relation to the maintenance of a retrieval mode. The sustained SSVEP effects were
obtained using a long (40s) averaging period so that neural activity maintained for the
duration of the task could be investigated. This approach essentially averages SSVEP
amplitude and latency across all time points during the task, and includes all target and
non-target presentations. However, more transient effects that reflect more rapid neural
processes, or that may differ for target and non-target objects, are masked using this
approach. In a separate analysis, SSVEP amplitude and latency data were calculated
using a short averaging period so that more transient patterns of neural activity could be
examined. This section discusses the transient SSVEP amplitude and latency changes
that occurred at a point in time when subjects were successfully retrieving target objects
from memory.
One of the main aims of this study was to investigate the SSVEP amplitude and latency
changes associated with the retrieval from memory of abstract, two-dimensional,
objects. Retrieval processes were examined by varying the memory load so that
retrieval became more difficult with increasing memory load. It was anticipated that
137

Discussion
increases in memory load would be associated with increased utilization of those neural
regions involved in successful retrieval processes. Electrophysiological findings led to
the second hypothesis: that successful retrieval would be associated with transient
SSVEP amplitude and latency changes over parietal and right frontal regions.
Furthermore, because abstract pictorial stimuli were used, it was further hypothesized
that the SSVEP changes would show a pattern with a more bilateral distribution than
that of the widely reported left parietal ERP old/new effect observed with verbal stimuli.
Transient load-dependent reductions in both SSVEP amplitude and latency that
followed the appearance of target objects were observed. More specifically, transient
amplitude reductions for the m3 relative to m1 comparison were most prominent at
occipital and bilateral parieto-temporal sites. For the m5 relative to the m1 comparison,
amplitude reductions were larger and more extensive, particularly at right hemisphere
sites. Latency reductions were more diffuse than amplitude reductions, although they
also appeared to be larger and more right lateralized as the memory load increased.
These SSVEP differences were statistically significant for the m3 relative to the m1
comparison at parietal and right occipital sites. The SSVEP differences for the m5
relative to the m1 comparison were statistically significant mainly over right
hemisphere regions, including occipital, parietal, parieto-temporal and right inferior
frontal sites. SSVEP differences for the m5 relative to the m3 comparison, while also
characterized by SSVEP amplitude and latency reductions were not statistically
significant. It is noteworthy that, compared with the load-dependent amplitude and
latency differences obtained for target objects, load-dependent differences for non-target
objects were much smaller and diffuse, and were not statistically significant at any
electrode site or for any time point.
While the topographies of transient SSVEP amplitude and latency differences were
calculated for the time point 400ms after the appearance of a target, when subjects were
likely to be engaged in memory retrieval, transient SSVEP amplitude and latency
reductions were evident throughout the entire 1400ms period that the target object was
visible. These reductions were statistically significant in the interval from
approximately 200 to 900ms after target appearance, and were located over regions
similar to those at 400ms.
138

Discussion
The second hypothesis predicted that transient SSVEP amplitude and latency load-
dependent changes would be observed at parietal and right frontal regions, regions
consistently noted in the electrophysiological literature as being involved in successful
retrieval processes. Furthermore, as the stimulus shapes were not easy to verbalize, it
was hypothesized that parietal changes would be bilateral rather than exclusively left-
sided. It appears that these hypotheses were supported. Relative to lower memory loads,
higher memory loads produced transient SSVEP amplitude and latency reductions over
bilateral parieto-temporal and right inferior frontal regions. A detailed discussion of the
transient SSVEP response over parietal and right frontal regions will follow in sections
6.3.1 and 6.3.2 respectively. The neural regions considered responsible for the observed
effects will be discussed mainly in relation to previous studies that have associated ERP
old/new differences with either familiarity or recollection-based memory processes.
Several studies have indicated that frontal changes are related to aspects of episodic
retrieval such as retrieval strategy and retrieval effort. Transient frontal SSVEP effects
will also be discussed in relation to retrieval effort in section 6.3.3. Finally, in section
6.3.4, the interpretation of transient SSVEP amplitude and latency reductions is
discussed.
6.3.1 Transient parietal effects
The parietal ERP old/new effect, where previously studied old items give rise to more
positive ERPs than novel new items, has frequently been reported in episodic memory
retrieval studies (see Chapter 2). In the present study, transient load-dependent effects
were most prominent over bilateral parietal and adjacent scalp regions, as predicted in
the second hypothesis of this study. Unlike the parietal ERP old/new effect, however,
the SSVEP differences were load-dependent, and therefore more specifically associated
with those regions more extensively utilized when subjects are making a greater effort
to retrieve studied objects from memory.
Clinical evidence, reviewed in section 2.1.2, has indicated involvement of the medial
temporal lobes in retrieval processes. It has been suggested that the parietal ERP
139

Discussion
old/new effect, which has been used to investigate neural activity associated with
successful retrieval, is a reflection of medial temporal lobe activity (Rugg et al. 1996).
Evidence supporting this proposal comes from patient studies where ERP old/new
differences, similar to those recorded over scalp areas, were obtained using depth
electrodes within the medial temporal lobes (eg. Guillem et al. 1995a; Guillem et al.
1995b; Guillem et al. 1996; Heit et al. 1990; Puce et al. 1991; Smith et al. 1986), and
where left temporal lobectomy patients failed to show a late ERP old/new effect (eg.
Smith and Halgren 1989). Guillem and colleagues (Guillem et al. 1995a; Guillem et al.
1995b) recorded directly from a number of intracerebral areas and concluded that the
ERP old/new effect is modulated by distributed, yet highly interconnected, brain
regions, including hippocampal, parietal and frontal regions. While volume conduction
of parietal and frontal cortical potentials to the scalp appears plausible, transmission
from deeper medial temporal lobe structures seems less likely. Klimesch et al. (1996),
however, suggested that theta activity, which appears to index episodic retrieval
processes, might be transmitted to the cortex via hippocampo-cortical feedback loops.
Findings from the present study do not provide further insights into the sources of the
ERP old/new effect. However, spatio-temporal similarities between transient load-
dependent SSVEP effects and ERP old/new effects suggest common origins. In future
studies, the use of source derivation techniques similar to those used by Duzel et al.
(1999) should provide valuable information regarding the sources of transient SSVEP
amplitude and latency changes. Furthermore, the use of electrophysiological techniques
with their high temporal resolution, in combination with PET and fMRI techniques with
their high spatial resolution, should also help answer this question.
Atlhough they lack the temporal resolution of electrophysiological techniques, PET and
fMRI functional neuroimaging techniques, with their superior spatial resolution, have
helped us identify neural regions involved in retrieval processes. Because these
techniques do not permit a high temporal resolution, studies using a block design have
been popular. For example, within a study, different blocks may contain different
proportions of studied to unstudied items (Cabeza and Nyberg 2000). In this way
regions showing greater activation with a greater proportion of studied items can be
identified. Such studies have indicated that the medial temporal lobes are involved in
140

Discussion
successful memory retrieval. Nyberg et al. (1996b) showed that medial temporal lobe
activation was correlated with the number of correctly retrieved old words (Nyberg et
al. 1996b).
Activation of medial temporal lobes during successful retrieval has generally been
found to occur bilaterally, regardless of the nature of the test items used (Cabeza and
Nyberg 2000). In the present study, significant transient load-dependent SSVEP effects
were found bilaterally at parietal and adjacent scalp sites. However, to suggest that these
bilateral SSVEP effects are in some way related to bilateral activity within the medial
temporal lobes would be highly speculative. The increased right lateralization of SSVEP
effects that occurred with increasing memory load is, however, discussed below.
A number of additional cortical regions have been associated with retrieval processes, in
particular, the medial parieto-occipital region, the occipital region, and lateral parietal
regions (Cabeza and Nyberg 2000). However, activation of these areas has been
attributed more specifically to processes associated with discerning the perceptual and
spatial detail of items rather than with processes specifically related to retrieval success.
These regions of cortex also lie below recording sites that showed significant load-
dependent SSVEP amplitude and latency effects and may also be involved in
discriminating between the perceptual or spatial details that determine whether items are
targets or non-targets. The spatial resolution of the SSPT technique used in the present
study did not allow a more precise localization of the specific neural regions involved.
Source localization and multimodal imaging approaches may help to address this
problem in future studies.
In addition to predicting that transient load-dependent SSVEP changes would occur
over parietal and right frontal regions, the second hypothesis also refers to the
lateralization of transient parietal effects. Because of the non-verbal nature of the
stimuli used, a relatively bilateral pattern of parietal effects was predicted rather than the
widely reported left-lateralized pattern obtained using verbal stimuli. For the m3 relative
to the m1 comparison, SSVEP amplitude and latency reductions over parietal regions
were indeed located bilaterally. For the m5 relative to the m1 comparison, both
amplitude and latency reductions were larger and right lateralized. Two possible
141

Discussion
explanations are suggested for this increasing right lateralization of amplitude and
latency reductions with increasing memory load. Firstly, with increasing memory load,
regions specifically involved in the retrieval of abstract objects become more apparent,
and these regions are located in the right hemisphere. Secondly, the right hemisphere is
engaged in a more general way when the task becomes more difficult. While both of
these explanations appear plausible, the fact that load-dependent effects were not
observed for non-target objects makes the second explanation less likely. However, the
identification of non-targets may have occurred more quickly and less laboriously than
the identification of target objects, thereby reducing differences between memory load
conditions. This, however, could not be determined because subjects were only required
to respond to target objects.
Johnson (1995) suggested that the absence of ERP old/new effects in some studies
resulted from the use of low memory loads or insufficient delay between first and
second presentations. He argued that only with sufficient memory load do retrieval
related differences become apparent. Similarly, the present findings indicate that when
sufficient memory load is applied, the right hemisphere is shown to be involved in the
retrieval of abstract objects from memory.
Schloerscheidt and Rugg (1997) hypothesized that the retrieval of objects from memory
would be associated with a relatively bilateral pattern for parietal ERP old/new
differences, while retrieval of words would show more prominent effects over left
parietal regions. This hypothesis was based on observations of verbal deficits in
individuals with damage to left medial temporal lobe structures, and deficits in retrieval
from pictorial memory in individuals with damage to right medial temporal lobe
regions. However, Schloerscheidt and Rugg (1997) observed a similar left-lateralized
parietal ERP old/new effect for both objects and words. They thus concluded that the
medial temporal lobes are not involved in the generation of the ERP old/new effect.
This conclusion appears at odds with a large body of literature suggesting that medial
temporal lobe structures are indeed associated with explicit memory processes and are
involved in the generation and modulation of the ERP old/new effect.
Schloerscheidt and Rugg may have obtained left parietal lateralization of ERP old/new
142

Discussion
effects with pictorial stimuli because the stimuli used were pictures of common objects
which may have been encoded and subsequently retrieved using verbal processes. The
abstract stimuli used in the present study were not as easily verbalized, and this could
explain the observed right lateralization of effects. An alternative explanation for the
left-sided ERP old/new effects observed by Schloerscheidt and Rugg, in line with the
proposal of Johnson (1995), is that the retrieval task contained insufficient memory
load.
It is also possible that right hemisphere load-dependent effects may reflect processes
involved more specifically in distinguishing target objects from similar-looking non-
target objects, rather than reflecting processes related to episodic memory retrieval per
se. This view is consistent with observations of activations within right lateral parietal
regions during the processing of spatial information in an episodic retrieval task.
(Moscovitch et al. 1995). Lateral parietal activations have also been associated with
perceptual aspects of recognition memory processes (Cabeza et al. 1997b), and occipital
activations have been reported for non-verbal retrieval and memory-related imagery
operations (Cabeza and Nyberg 2000). Although such processes may be engaged while
processing target stimuli in the present study, it is expected that they would also be
engaged during the processing of non-target stimuli to enable a decision to be made as
to whether a test stimulus is a target or a non-target. Given that SSVEP amplitude and
latency differences between memory loads were not statistically significant following
the presentation of non-target stimuli, it is reasonable to suppose that target SSVEP
differences arise mainly from retrieval effort, and to a lesser extent from other processes
that are necessary for performance of the task.
6.3.2 Transient right frontal effects
In addition to occurring over parietal scalp regions, the ERP old/new effect has also
been observed over right frontal regions. Moreover, the investigation of the combination
of parietal and frontal ERP old/new effects may provide information about the type of
recognition occurring. That is, it may indicate whether recognition is based on
recollection, reflecting episodic memory retrieval processes, or is based on familiarity,
143

Discussion
possibly involving implicit memory retrieval processes.
As discussed in Chapter 2, right frontal ERP old/new effects have been observed when
the context of studied items is remembered (eg. Wilding and Rugg 1996), when studied
associations are correctly retrieved (eg. Donaldson and Rugg 1998), or when specific
perceptual or spatial details of studied pictures are correctly recalled (eg. Ranganath and
Paller 2000). Wilding and Rugg (1996) required subjects to remember whether old
words had been spoken by a male or female voice. Correct contextual judgements
produced left parietal and right frontal ERP old/new effects. These effects were
prolonged, particularly at frontal regions, lasting well beyond the motor response. Given
that these effects were associated with retrieval of the specific study context, the authors
argued that they index recollection-based recognition involving episodic memory
processes. They also argued that the prolonged frontal effects were associated with the
monitoring processes that occur during recollection-based retrieval.
ERP old/new effects for incorrect contextual judgements were observed over the same
regions, but these were much smaller and less prolonged. These differences were
attributed to familiarity-based retrieval since the specific study context was not correctly
identified. However, because the ERP old/new effects for both conditions were
qualitatively similar, the authors proposed that their observations were consistent with a
model in which both recollection- and familiarity-based retrieval utilize the same
explicit memory system (Moscovitch 1992; Moscovitch 1994; Squire 1994). In this
model, familiarity-based recognition can be achieved solely with hippocampal output.
Retrieval of contextual information, or recollection, is achieved through further
integration with the prefrontal cortex.
Other researchers, however, claim that familiarity-based recognition utilizes the implicit
memory system (Squire 1992; Tulving and Schacter 1990). This view has recently been
supported by a study in which familiarity-based processes appeared to involve the
perirhinal cortex and the medial dorsal nucleus of the thalamus, and not the
hippocampus (Appleton and Brown 1999).
The location and timing of transient load-dependent effects in the present study are
144

Discussion
consistent with the findings of Wilding and Rugg (1996). Statistically significant
transient SSVEP amplitude and latency reductions over right inferior frontal regions
were observed with both the m3 relative to m1, and the m5 relative to m1, comparisons.
As already mentioned in section 6.3.1, retrieving targets from memory produced
prolonged parieto-temporal amplitude and latency reductions for higher relative to
lower memory load conditions. Amplitude and latency reductions for higher relative to
lower memory load conditions over right frontal regions were also prolonged, lasting
more than one second. They were statistically significant in the interval between 400
and 800ms after the appearance of the target. Right frontal effects described by Wilding
and Rugg (1996) occurred over a large area. In the present study, SSVEP frontal effects
were localized to right inferior frontal sites. The less diffuse nature of the effect in the
present study can possibly be explained by the fact that responses to targets under
different memory load conditions were compared. Wilding and Rugg (1996), however,
compared ERPs for targets relative to non-targets. Present findings may, in fact, have
reflected activity in underlying neural regions involved when retrieval becomes more
demanding.
As already explained, right frontal effects have been observed in studies that used the
ERP old/new effect to examine episodic memory retrieval. In the present study,
transient load-dependent effects were also observed over right frontal regions and may
also reflect episodic memory retrieval processes. Certain aspects of the observed
SSVEP amplitude and latency reductions support this proposal. Firstly, both the
location and the timing of the parieto-temporal and right frontal SSVEP amplitude and
latency reductions were similar to the spatio-temporal effects described in studies
designed to investigate context-based retrieval, and in studies where specific detail
relating to the studied events was retrieved. Secondly, transient frontal and parieto-
temporal SSVEP reductions increased with increases in memory load. These greater
reductions, in conjunction with increased response times and perceived effort, indicate
that that retrieval becomes more demanding as memory load increases. If recognition
resulted purely from familiarity, involving more automatic and less effortful processes,
load-dependent amplitude and latency reductions would not be expected. Furthermore,
while the actual cognitive operations used to retrieve objects from memory have not
been fully determined, they are likely to involve memory scanning and/or some form of
145

Discussion
perceptual or spatial discrimination in order to determine whether an object is a target or
a distractor. It seems plausible that such processes would also be affected in some way
by increases in memory load. Familiarity-based retrieval is faster (Yonelinas and Jacoby
1995) and is therefore unlikely to involve these more elaborate processes. The similarity
in the appearance of target and non-target objects in the present study meant that
subjects had to retrieve specific features of the stimuli, thus increasing the likelihood of
context-based episodic memory retrieval rather than familiarity-based retrieval
occurring. Thirdly, because encoding was intentional rather than incidental, and was
also reinforced in a practice task, targets would have been relatively well encoded, and
therefore subjects would have been confident that their judgements were correct. All of
these factors would have increased the likelihood that recognition was based on explicit
or declarative memory rather than implicit or familiarity-based memory. However, the
use of familiarity-based processes cannot be totally discounted.
As was discussed earlier, separate neural systems may be involved in familiarity-based
and recollection-based recognition (Appleton and Brown 1999). That is, familiarity-
based recognition may involve perirhinal regions, whereas recollection-based
recognition involves medial temporal regions. Given the spatial resolution of the SSPT
technique used, the neural regions responsible for the observed scalp SSVEP effects
cannot be localized more precisely. Source localization techniques or multimodal
imaging could prove useful in localizing more precisely the neural regions utilized,
therefore possibly indicating whether familiarity-based or recollection-based
recognition is being accessed.
While the time-course of the observed SSVEP load-dependent changes is not dissimilar
to findings from context-based memory retrieval studies, it must be remembered that
SSVEP amplitude and latency values were derived from Fourier coefficients averaged
over a moving window of 20 cycles, giving an effective temporal resolution of 870ms.
As a result, faster neural processes may have been reflected in the analysed data by
slower and more prolonged SSVEP amplitude and latency changes. It is therefore
possible that the conclusion drawn, namely, that recognition was more likely to have
been based on recollection rather than familiarity, is incorrect. However, the fact that
transient amplitude and latency reductions were maximum 200 to 400ms after the motor
146

Discussion
response, rather than prior to the response, indicates that these reductions reflected
slower processes more consistent with recollection-based memory retrieval.
6.3.3 Transient effects and retrieval effort
Another aspect of retrieval processing that has been discussed in the
electrophysiological literature is the effort required to retrieve items from memory (eg.
Rugg and Wilding 2000). Retrieval effort has been investigated by comparing the ERP
response to retrieval tasks with varying levels of difficulty. For such comparisons,
significant differences between ERP components over frontal regions are commonly
reported (eg. Ranganath and Paller 1999; Ranganath and Paller 2000; Rugg et al. 2000).
For example, a greater positivity was elicited over left frontal regions by the more
difficult of two retrieval tasks (Ranganath and Paller 1999). In contrast, right frontal
ERP changes were associated with the retrieval of words that had been encoded
shallowly rather than with words that had been encoded more deeply (Rugg et al. 2000).
The lack of agreement in the ERP literature over frontal lateralization of transient
retrieval effort effects also exists in the PET and fMRI literature (Rugg and Wilding
2000).
In section 6.1 it was argued on the basis of response time data and subjects’ personal
accounts that increased memory load results in both increased retrieval difficulty and
increased retrieval effort. During the successful retrieval of target objects, for higher
relative to lower memory load conditions, significant transient amplitude and latency
reductions over bilateral parieto-temporal and occipital regions, and right inferior
frontal regions were observed. These reductions may therefore reflect the increased
difficulty in retrieving objects when the memory load is increased. As discussed in
section 6.3.1, the source of the right hemisphere load-dependent SSVEP amplitude and
latency reductions may be either neural regions involved in the retrieval of abstract
objects, or neural regions recruited when the task becomes more difficult. Rugg and
Wilding (2000) suggest that these two possibilities could be reflections of the same
neural processes. That is, retrieval effort may not have a distinct neural signature;
instead, ‘the neural correlates of increasing effort will be manifest as increased activity
147

Discussion
of whatever brain regions are engaged by the retrieval task in question’ (Rugg and
Wilding 2000, p.114). This concept is of particular relevance to this study as it
underpins the use of different memory loads to identify the neural regions involved in
successful memory retrieval. The present author suggests that the employment of tasks
with different memory loads warrants further investigation as this may help to identify
more precisely neural regions associated with successful retrieval processes than do the
more traditional methods of comparing the responses to old and new items.
In order to achieve a clearer picture of the relationship between memory load and the
SSVEP response, a more recent study performed by the present author incorporated five
different memory load levels. While the experimental conditions differed considerably
from those of the present study, preliminary results indicate a correlation between the
SSVEP response and memory load (see Appendix F for poster presentation).
6.3.4 Interpretation of transient SSVEP amplitude and latency reductions
Thus far, transient SSVEP effects have been discussed in terms of scalp location, but the
nature of these effects, that is, whether they involved amplitude or latency, and whether
these were increased or reduced is also of significance. In section 6.2.2 it was explained
that amplitude reductions are thought to reflect increased neural activity, and latency
reductions are thought to reflect increased coupling between neural regions leading to
increased processing speed. It was suggested in section 6.2.2 that amplitude and latency
reductions reflect sustained retrieval mode processes, and given the topography of
latency reductions in particular, it was suggested that latency reductions might index the
coupling of neural regions during the maintenance of a retrieval mode. Differences that
exist in the interpretation of sustained and transient SSVEP amplitude and latency
reductions lie primarily in the cognitive processes that they may be indexing. Transient
SSVEP amplitude and latency reductions occur much faster than the observed sustained
reductions, and are more likely to reflect neural activity associated with more specific
memory retrieval processes. Another difference in the interpretation of sustained and
transient reductions lies in the fact that the sustained reductions were obtained for a
memory retrieval task relative to a low demand control task, whereas the transient
148

Discussion
changes were load-dependent and appear to be graded. The brief discussion below
extends the previous interpretation of sustained effects to include an interpretation of
faster load-dependent amplitude and latency reductions.
As indictated in Chapter 3, SSVEP amplitude reductions demonstrated during a visual
vigilance task (Silberstein et al. 1990b) and during the performance of the Wisconsin
Card Sorting Task (Silberstein et al. 1995b) were likened to the phenomenon of event
related desynchronisation (ERD) (eg. Pfurtscheller and Aranibar 1977a; Pfurtscheller
and Klimesch 1990) where transient alpha amplitude reductions were considered to
index regional increases in cortical activity associated with the performance of cognitive
and motor tasks. Furthermore, the extent of alpha attenuation has been associated more
specifically with the difficulty and/or relevance of a task (eg. Klimesch et al. 1990a;
Klimesch et al. 1993). Similarly, in relation to the present findings, it is suggested that
transient SSVEP amplitude reductions index the extent of utilization of those neural
regions associated with successful retrieval processes. Because SSVEP amplitude
reductions were observed for higher relative to lower memory load conditions, it is
more specifically suggested that SSVEP amplitude reductions index increased difficulty
or effort associated with performance of a higher memory load condition.
Comparisons between the m3 relative to the m1 condition, the m5 relative to the m1
condition, and the m5 relative to the m3 condition, all show SSVEP amplitude and
latency reductions over parietal and right frontal regions during the time that subjects
were considered to be actually retrieving target objects from memory. The amplitude
and latency reductions for the higher relative to the lower memory load conditions were
statistically significant for the m5 relative to the m1, and the m3 relative to the m1
comparisons, but did not reach significance for the m5 relative to the m3 comparison.
The Hotelling’s T score is a bivariate statistic that combines each pair of amplitude and
phase values into a single vector, thereby clouding the independent contribution of each
of these components. Separate statistical analyses of amplitude and latency are planned
for future studies.
Klimesch and colleagues (eg. Doppelmayr et al. 1998; Klimesch et al. 1997; Klimesch
et al. 1994) demonstrated that episodic retrieval processes are associated with specific
149

Discussion
EEG theta band changes. Transient theta increases, or theta synchronization, during a
time interval when subjects are involved in successfully recognizing previously studied
items, was reported in a number of studies by this group. It has been suggested that
these theta increases reflect hippocampal activity that is transferred to the scalp via
longitudinal hippocampal-cortical pathways (Klimesch et al. 1994). Similarly, it appears
that load-dependent 13Hz SSVEP amplitude reductions reflect certain memory retrieval
processes. An important distinction exists between the theta synchronization approach
described in the literature and the SSPT technique used in the present study. Theta
activity is a component of the brain’s intrinsic EEG, and has been observed to vary in
response to mental activity. In contrast, the 13Hz SSVEP is generated by an externally
produced visual flicker, and is therefore a driven response. This driven response appears
to be modified by neural processes associated with mental activity. It has been
suggested that SSVEP amplitude changes may in fact reflect neural processes associated
with the alpha band resonant system (Silberstein et al. 2001). In the light of the reported
relationship between theta activity and episodic memory processes, an extension to the
present study would be to investigate SSVEP amplitude and latency variations using a
probe visual flicker with a frequency in the theta range.
As mentioned above, the effects of retrieving objects from memory on the SSVEP
amplitude were load-dependent, with increases in memory load resulting in transient
reductions in amplitude. Similarly, the effects on latency were also load dependent.
Increases in memory load resulted in transient decreases in latency over parietal and
right frontal regions. In section 6.2.2, latency reductions were discussed in relation to
retrieval mode processes. It was considered that latency reductions over right frontal
regions reflect processes involved in the maintenance of an episodic retrieval mode.
Silberstein et al. (1996) have suggested that latency reductions are a manifestation of
transient increases in coupling strength between neural populations leading to increases
in information processing speed. In line with this proposal, it is speculated that transient
load-dependent latency reductions over parieto-temporal and right inferior frontal
regions reflect increased coupling strength between neural regions involved in
successful retrieval processes. It is further speculated that as memory load is increased,
connections between these regions are strengthened to enable the performance of a more
demanding retrieval task.
150

Discussion
In summary, transient load-dependent SSVEP amplitude and latency reductions
occurred while subjects were considered to be engaged in the process of retrieving
previously studied abstract objects from memory. These reductions occurred over
parietal and right frontal regions as predicted by the second hypothesis of this study.
While bilateral parietal effects were predicted, the increasing right lateralization of
amplitude and latency reductions with increasing memory load is consistent with the
fact that the abstract objects used were difficult to encode verbally. The experimental
task was specifically designed to enhance the likelihood of stimuli being stored in a
longer-term form of memory. This, in conjunction with the observation that the parietal
and right inferior frontal latency reductions were prolonged, lasting until well after the
motor response, suggests that the observed reductions were more likely to have been
associated with episodic memory retrieval processes rather than with retrieval processes
based on familiarity. An association between load-dependent SSVEP amplitude and
latency reductions and memory retrieval processes is reinforced by the fact that there
were no statistically significant findings for non-target objects.
6.4 Transient SSVEP amplitude and latency changes: target versus non-target
objects
Although problematic, for reasons that will be discussed, differences between target and
non-target objects are presented here for completeness, given that this comparison most
closely parallels the old versus new comparison of the ERP old/new effect discussed in
Chapter 2.
Transient SSVEP amplitude and latency differences between target and non-target
objects were calculated separately for each memory load condition. Only differences for
a memory load of 5 were statistically significant. Significant differences occurred at
prefrontal and right inferior frontal sites at around 700ms after the appearance of an
object, and were characterized by SSVEP amplitude and latency reductions for targets
relative to non-targets. It is suggested that amplitude reductions reflect increased
utilization of underlying neural regions for target objects, and latency reductions
151

Discussion
represent more efficient coupling between neural regions, enabling faster information
processing (see sections 6.2.2 and 6.3.4). Although response times to non-targets were
not recorded, subjects indicated that non-target objects were more rapidly and easily
identified than target objects. It is considered that memory scanning or some form of
perceptual or spatial discrimination was used to identify targets. Thus, SSVEP
amplitude and latency reductions, for target compared with non-target objects, are
consistent with more effortful and extended processing for these items. Johnson (1995)
suggested that the magnitude of the memory load is important in determining whether
ERP old/new effects are observed. The results of the present study are consistent with
this viewpoint since significant differences were only observed for a memory load of 5.
Very few studies have examined ERP old/new differences using pictorial stimuli. Those
that have can not be directly compared because their experimental conditions differ in,
for example, experimental design (continuous recognition versus study-test) and/or type
of stimulus (verbalizable versus non-verbalizable). In contrast to ERP old/new effects
observed with verbal studies, parietal differences are not always observed with pictorial
studies (eg. Friedman 1990a). It appears that the only common finding with pictorial
studies is that the ERP old/new effects involve frontal regions. In one study, ERP
old/new differences were found to be much larger for picture than for word stimuli
(Schloerscheidt and Rugg 1997). Moreover, while both pictures and words showed
early ERP old/new effects at left parietal sites, only pictures elicited significant right
frontal effects. The authors concluded that these right frontal differences are consistent
with the suggestion that pictures are more richly encoded than words, and are therefore
more likely to engage post-retrieval processes. It is difficult to determine whether the
SSVEP amplitude and latency reductions observed in the present study support the
interpretation of ERP old/new findings because of the paucity of studies and the variety
of experimental conditions used. The timing and topography of frontal SSVEP
reductions, however, are comparable to those of frontal ERP old/new effects.
The interpretation of SSVEP effects for targets relative to non-targets should be
approached with caution because a number of variables were not well controlled.
Firstly, targets required a motor response whereas non-targets did not. The observed
frontal differences may therefore be attributable to the motor response, although this
152

Discussion
seems unlikely as significant differences were largest well after the motor response.
Furthermore, differences were statistically significant only for a memory load of five;
they were not significant for loads of three and one. If differences resulted from the
motor response, then similar differences for all three memory load conditions might be
expected. Secondly, targets appeared with a probability of 0.25 while non-targets
appeared more frequently with a probability of 0.75. It could therefore be argued that
target/non-target differences reflect probability effects. If this were the case, however, a
diffuse P300-like response would be expected over parietal regions at around 300ms
after object appearance (eg. Picton 1992). In the present study, however, differences
were more frontal and occurred much later. Thirdly, there was a mismatch between the
number of targets and non-targets presented; far fewer targets were averaged resulting
in a lower signal to noise ratio. This may have reduced observed differences, but the
effect would have been the same for all memory load conditions. However, significant
differences occurred only for a memory load of five. It therefore appears unlikely that
the aforementioned factors contributed significantly to the transient amplitude and
latency differences observed. Consequently, it is tentatively suggested that the observed
transient SSVEP amplitude and latency differences reflect processes associated with
retrieval success in the same way that ERP old/new effects reflect these processes.
Furthermore, as the frontal SSVEP amplitude and latency reductions for targets
occurred after the motor response, it is further speculated that these reductions more
specifically reflect monitoring processes associated with episodic memory retrieval.
Despite the assumption that differences between the SSVEP response to targets and
non-targets were probably unaffected by the aforementioned confounding factors, the
conclusions drawn should be re-examined when such factors have been eliminated. To
address all three issues, the experiments could be repeated with an increased number of
stimulus items, equal numbers of targets and non-targets, and responses to both target
and non-target objects. Problems with interpretation due to the motor response could be
reduced in two ways. Firstly, a larger number of stimulus items would permit a higher
temporal resolution. If effects associated with memory retrieval and the motor response
occurred at different times, a higher temporal resolution would help to differentiate
these processes. Secondly, a motor response to both target and non-target objects should
lead to its effects subtracting out when comparing one condition with the other.
153

Discussion
Probability effects should be eliminated simply by the use of equal numbers of target
and non-target stimuli. For a given temporal resolution, increased numbers of target and
non-target stimuli would also improve the signal to noise ratio, increasing the likelihood
of significant effects being observed. Additional experiments employing both verbal
and pictorial material could help determine whether frontal memory retrieval effects
depend on the nature of the stimulus used. This could be extended to investigate other
types of visual stimuli such as numbers or colours, or stimuli encoded and tested in
different sensory modalities.
6.5 Conclusions and future directions
The SSPT technique was used to investigate neural correlates of object recognition
memory. Both sustained and transient SSVEP effects were analysed to examine
retrieval mode and retrieval success processes respectively during the performance of an
episodic memory retrieval task.
The first hypothesis of this study predicted that retrieval mode processes would be
reflected in sustained SSVEP effects over right frontal regions. Sustained amplitude and
latency reductions were found for the memory retrieval condition relative to the baseline
condition, and these sustained reductions were most prominent and statistically
significant over right frontal regions. While the amplitude reductions were diffuse,
latency reductions were most prominent over right fronto-temporal regions. It is
suggested that sustained SSVEP latency reductions over right fronto-temporal regions
reflect the maintenance of a retrieval mode during the performance of the task. While
this finding is consistent with the first hypothesis, and therefore with the current
literature, further investigation is required to confirm whether the source of these effects
is the same as those reported in fMRI, PET and DC-ERP studies. A secondary finding
of the present study is that sustained SSVEP amplitude and latency values do not
change significantly when the memory load is increased. It is suggested, therefore, that
retrieval mode processes are independent of the level of effort applied when performing
a memory retrieval task.
154

Discussion
The second hypothesis of this study predicted that successful retrieval of abstract
objects would be associated with transient load-dependent SSVEP changes over
bilateral parietal and right frontal regions. Load-dependent SSVEP amplitude and
latency reductions were found over these regions during a time interval when it was
considered that subjects were involved in retrieving these objects from memory. With a
small increment in memory load, load-dependent differences were observed bilaterally
over parietal regions. This finding is therefore consistent with the second hypothesis.
However, a larger increment in memory load produced larger and more right lateralized
effects. It is suggested that this pattern of right lateralized changes is consistent with the
use of stimuli that are not easily verbalized, and was probably not observed in previous
picture-based studies because of the use of lower memory loads in these studies. This
explanation is in line with Johnson’s (1995) proposal that episodic retrieval effects
become apparent when the memory loads applied, and the delays between studied and
tested items, are sufficient.
It is proposed that these transient load-dependent effects reflect episodic retrieval
processes rather than processes based on familiarity. This conclusion is drawn from a
number of observations. Firstly, the spatio-temporal patterns of both the parietal and
right frontal effects are similar to the spatio-temporal patterns of the ERP old/new effect
observed in context-based retrieval studies that have specifically investigated episodic
memory retrieval processes. Secondly, changes were load-dependent, meaning that
retrieval was effortful and not automatic. Thirdly, the experimental design was
structured to promote recollection of the perceptual details of items to be retrieved.
It is also proposed that the method of varying the memory load to investigate successful
retrieval processes may give more valid indications of retrieval processes than other
methods which have compared activity in response to old and new items, as used in
studies into the ERP old/new effect. The advantage of the method used in the present
study is that activity in response to retrieved, or old, items is not contaminated by
activity generated by a comparison task requiring, for example, the identification of new
items. While it is possible that the load-dependent transient effects of the present study
may reflect processes specific to task effort, rather than to memory retrieval, it is
suggested that this scenario is unlikely in view of the spatio-temporal similarities
155

Discussion
between transient load-dependent SSVEP effects and the ERP old/new effect. Rugg and
Wilding (2000, p. 114) have, in fact, suggested that retrieval effort doesn’t have a
unique neural signature. They suggest, instead, that activity associated with retrieval
effort may be observed in whatever neural regions are involved with retrieval success.
The present results support Rugg and Wilding’s suggestion, given the similarity
between the spatio-temporal patterns of load-dependent SSVEP effects and ERP
old/new effects. It is therefore suggested that the method of varying the memory load to
investigate retrieval processes warrants further investigation.
There are a number of important issues that remain unresolved and further questions
that arise from the present findings. Firstly, what are the neural regions that generate
these sustained and transient load-dependent effects, and are these effects generated by
the same neural regions that generate the ERP old/new effects? Secondly, while the
present findings indicate that right frontal regions may be involved in both sustained
and transient retrieval processes, it is not clear whether the same or different underlying
neural regions are involved. Thirdly, are the findings of sustained and transient effects
stimulus dependent? That is, would the use of stimuli such as words, numbers, or even
sounds result in the same patterns of activity observed in the present study with abstract
objects?
The combination of SSPT, which allows investigation of cognitive processes that occur
over different time scales, with the method of varying memory load to help identify
neural regions involved in retrieval processes, appears to be a potentially useful
approach for the investigation of these unresolved issues. Furthermore, a combination of
SSPT with source localization techniques and multi-modal imaging approaches may
also prove valuable in identifying the source of the sustained and transient effects
observed in the present study.
156

Appendix A. Task instructions

Appendix A. Task instructions
TASK INSTRUCTIONS
<read to subject>
Baseline
In the centre of the screen in front of you, a series of blue numbers, 1, 2, 3 and 4 will be
presented. The numbers will be shown to you over and over again for about 2 minutes.
Your task during the presentation of these numbers is very simple; just R-E-L-A-X,
watch the numbers and press the hand-held button S-L-O-W-L-Y whenever you see the
number 4 appear <show button press>.
Memory
In the centre of the screen in front of you, a series of numbers and shapes will be
presented one at a time. Your task is to respond to certain shapes by pressing the hand-
held button. The shapes that you must respond to will be shown to you before the task
begins.
The task will begin with the presentation of a series of BLUE numbers, 1, 2, 3 and 4.
The numbers will be shown to you over and over again for about 40 seconds. Your task
during the BLUE numbers is very simple; just R-E-L-A-X, watch the numbers and press
the button S-L-O-W-L-Y whenever you see the number 4 appear.
Following the BLUE numbers, a series of RED shapes will appear. Your task during
this segment is to C-O-N-C-E-N-T-R-A-T-E and press the button as Q-U-I-C-K-L-Y
and A-C-C-U-R-A-T-E-L-Y as you can whenever you see a TARGET shape.
Remember the target shapes will be shown to you at the beginning of each task. This
section of the task will last for about 40 seconds.
After the RED shapes segment, once again, a series of BLUE numbers will be shown to
you. As before, just RELAX and press the button SLOWLY whenever you see the
158

Appendix A. Task instructions
number 4 appear. This section will also last for about 40 seconds.
Each task, consisting of BLUE numbers, RED shapes, BLUE numbers will take about 2
minutes to complete.
<Demonstrate slow and quick button presses>
<Practice Run>
<read only the first time>
Before each task the INSTRUCTIONS will be repeated to you. If the instructions are
not clear please do not hesitate to ask.
<for investigator>
<Order 1: bas1, bas2, m1p, m1, m3p, m3, m5p, m5>
<OR>
<Order 2: bas1, bas2, m3p, m3, m1p, m1, m5p, m5
159

Appendix B. Determination of optimum luminance of
light-emitting diode (LED) arrays

Appendix B. Determination of optimum luminance of LED arrays
The maximum luminance of the light emitting diode (LED) arrays that produce the
probe stimulus was determined in a separate experiment. The voltage driving the LED
arrays was varied in a pseudo-random fashion so that the intensity of the flicker ranged
from very dim to very bright. A fixation point on the task computer was viewed through
the flicker, while the EEG was recorded for 50s at the occipital electrode, Oz. Two
subjects were used in this experiment. After every 50s recording period, each subject
gave a subjective comfort rating for the flicker: 10 represented ‘very comfortable’ and 1
represented ‘very uncomfortable.’ For each 50s period, the average SSVEP amplitude
across subjects was determined off-line and plotted against voltage. As can be seen in
Figure B.1), initially the SSVEP amplitude increased almost linearly and then reached a
plateau, further increases in voltage having little effect on the SSVEP amplitude. It is
important to use a luminance in the plateau region so that the SSVEP amplitude is not
affected by small changes in modulation depth. Regan (1972) suggested that the
modulation depth should exceed 30%. A voltage of 0.4 volts was chosen as it gives an
SSVEP amplitude in the plateau region, a modulation depth of 45%, and was not
uncomfortable for either subject.
Peak-to-peak voltage into LED goggle driver (volt)
0.0 0.2 0.4 0.6 0.8 1.0 1.2
Com
fort
(0-1
0)
0
5
10
Ave
rage
SS
VE
P a
mpl
itude
(Nor
mal
ized
uni
ts)
10
100
ComfortSSVEP amplitude
Mod
ulat
ion
dept
h =
45%
Determination of optimum luminanceof LED arrays
Figure B.1 Determination of optimum luminance of LED arrays
The peak LED-driver input voltage was determined by assessing the combination of both the SSVEP amplitude response at Oz and the subject comfort rating. A peak-to-peak voltage of 0.4 was selected giving a Modulation Depth of 45% (see also Figure 4.3).
161

Appendix C. Behavioural results for practice tasks

Appendix C: Behavioural results for practice tasks
Memory Loadm1 m3 m5
Mea
n R
espo
nse
Tim
e (m
sec)
0100200300400500600700800
m1 vs m3: NSm1 vs m5: NSm3 vs m5: NS
Figure C.1 Practice tasks: Mean response times for target objects versus memory load for an individual subject
Mean response time and standard deviation for target objects for each of the 3 practice task intervals. Data were taken from an individual subject (DK) performing task order 2 (ie. in the order m3, m1, m5). Results of unpaired t-tests for memory load comparisons are also shown. (NS = not significant)
163

Appendix C: Behavioural results for practice tasks
Mea
n R
espo
nse
Tim
e (m
sec)
0100200300400500600700800
Mea
n Fr
eque
ncy
of E
rror
s of
Om
issi
on
0.0
0.2
0.4
0.6
0.8
1.0
m1 vs m3: t=-3.84; p<0.001m1 vs m5: t=-6.27; p<0.0001m3 vs m5: NS
Memory Loadm1 m3 m5
Mea
n Fr
eque
ncy
of E
rror
s of
Com
mis
sion
0.0
0.2
0.4
0.6
0.8
1.0
m1 vs m3: NSm1 vs m5: NSm3 vs m5: NS
m1 vs m3: NSm1 vs m5: t=-2.1; p<0.005m3 vs m5: NS
Figure C.2 Practice tasks: Mean response time and errors for target objects versus memory load for 40 subjects
Mean response time and standard deviation for target objects for each of the 3 practice task intervals. Similar graphs are shown for errors of omission (missed targets) and errors of commission (responses to non-targets). Results of paired t-tests for memory load comparisons are also displayed. (NS = not significant)
164

Appendix D. The amplitude of the SSVEP
produced by turning on the probe stimulus

Appendix D: The amplitude of the SSVEP produced by turning on the probe stimulus
The 13Hz probe flicker was switched-on half way through the baseline practice task to
determine whether the driven 13Hz response (stimulus-on) was significantly different
from the background (spontaneous) 13Hz activity contained in the EEG. The SSVEP
amplitude and phase were calculated for each subject with both a high temporal
resolution (870ms) and a low temporal resolution (60s) averaging period. The high
temporal resolution analysis was performed so that the transition from stimulus-on to
stimulus-off could be visualized for each recording site. The low temporal resolution
analysis was performed by averaging all Fourier coefficients during the 60s stimulus-off
period and all Fourier coefficients during the 60s stimulus-on period. Hotelling’s T
values were used to determine the consistency of the difference between the SSVEP
response at 13Hz (stimulus-on) and the spontaneous 13Hz activity (stimulus-off) across
all 40 subjects.
The SSVEP amplitude and phase time series for the high temporal resolution averaging
period, calculated for an individual subject (BO) are shown in Figure D.1). Following
the switching on of the probe stimulus sixty seconds after commencement of the task,
the SSVEP amplitude increased and the phase appeared to stabilize. The appearance of
the SSVEP remained similar throughout the 60s stimulus on period. The topographic
maps in Figure D.2) show the mean SSVEP amplitude for the stimulus-on and stimulus-
off conditions, calculated over the whole 60s interval for each. The increase in SSVEP
amplitude is most apparent at occipito-parietal sites, although there were also smaller
increases at other recording sites.
166

Appendix D: The amplitude of the SSVEP produced by turning on the probe stimulus
SSVE
P am
plitu
de(m
icro
volt)
0123456
Time (sec)
0 10 20 30 40 50 60 70 80 90 100 110
SSVE
P ph
ase
(rad
ian)
-4-3-2-101234
stim
ulus
on
electrode - Oz
Figure D.1 SSVEP amplitude and phase during the stimulus-off and stimulus-on intervals for an individual subject
Steady-state visually evoked amplitude and phase changes occurring with the switching-on of the probe stimulus, recorded from the occipital scalp location Oz. Temporal resolution of 870ms. Data taken from individual subject (BO).
stimulus off stimulus on3.0
0.0 Vυ
Figure D.2 Mean SSVEP amplitude during the stimulus-off and stimulus-on intervals for an individual subject
Mean SSVEP amplitude calculated across the entire stimulus-off (left) and stimulus-on (right) intervals (60s of data) Data taken from individual subject (BO).
167
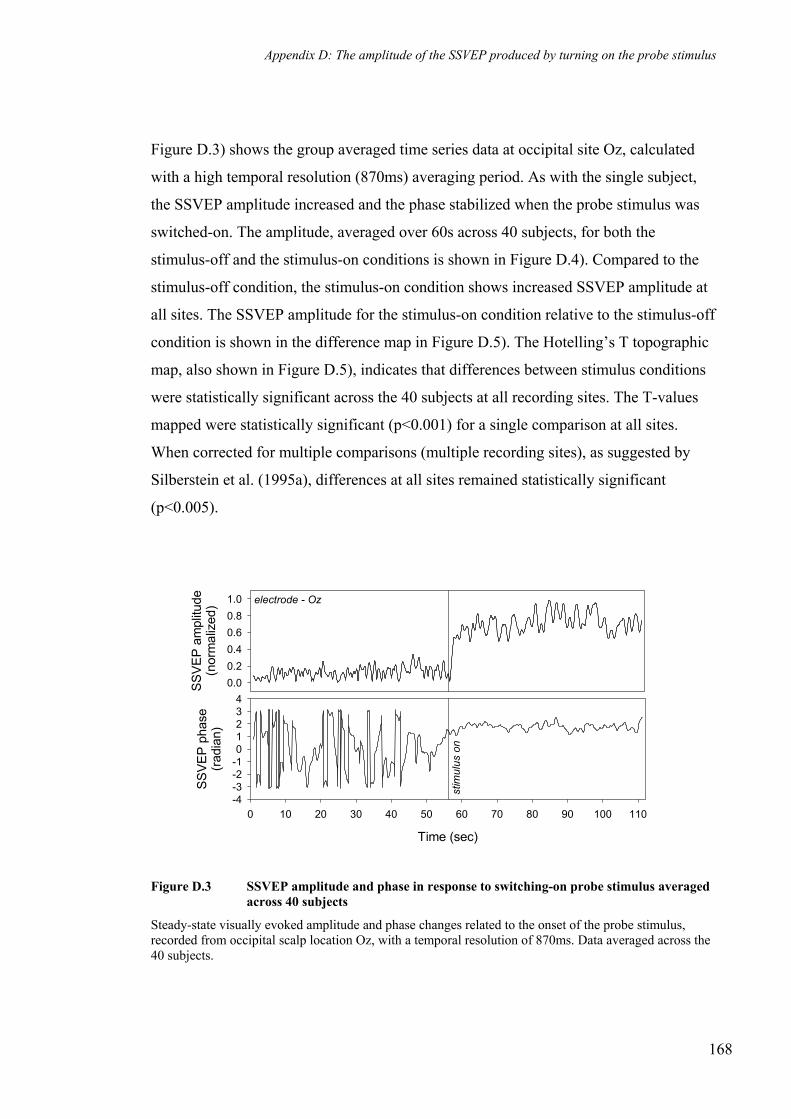
Appendix D: The amplitude of the SSVEP produced by turning on the probe stimulus
Figure D.3) shows the group averaged time series data at occipital site Oz, calculated
with a high temporal resolution (870ms) averaging period. As with the single subject,
the SSVEP amplitude increased and the phase stabilized when the probe stimulus was
switched-on. The amplitude, averaged over 60s across 40 subjects, for both the
stimulus-off and the stimulus-on conditions is shown in Figure D.4). Compared to the
stimulus-off condition, the stimulus-on condition shows increased SSVEP amplitude at
all sites. The SSVEP amplitude for the stimulus-on condition relative to the stimulus-off
condition is shown in the difference map in Figure D.5). The Hotelling’s T topographic
map, also shown in Figure D.5), indicates that differences between stimulus conditions
were statistically significant across the 40 subjects at all recording sites. The T-values
mapped were statistically significant (p<0.001) for a single comparison at all sites.
When corrected for multiple comparisons (multiple recording sites), as suggested by
Silberstein et al. (1995a), differences at all sites remained statistically significant
(p<0.005).
SSVE
P am
plitu
de(n
orm
aliz
ed)
0.00.20.40.60.81.0
Time (sec)
0 10 20 30 40 50 60 70 80 90 100 110
SSVE
P ph
ase
(rad
ian)
-4-3-2-101234
stim
ulus
on
electrode - Oz
Figure D.3 SSVEP amplitude and phase in response to switching-on probe stimulus averaged across 40 subjects
Steady-state visually evoked amplitude and phase changes related to the onset of the probe stimulus, recorded from occipital scalp location Oz, with a temporal resolution of 870ms. Data averaged across the 40 subjects.
168

Appendix D: The amplitude of the SSVEP produced by turning on the probe stimulus
stimulus off stimulus on0.8
0.0
Figure D.4 Mean SSVEP amplitude topography during the stimulus-off and stimulus-on intervals averaged across 40 subjects
Mean SSVEP amplitude calculated across the entire probe stimulus-off interval (60s off data) and probe stimulus-on interval across the 40 subjects.
Hotelling’s Tstimulus on - stimulus off+1.0
-1.0
12.0
0.0
0.0010.005
0.050.01
Figure D.5 SSVEP amplitude difference topography and Hotelling’s T values for stimulus-on relative to stimulus-off conditions averaged across 40 subjects
Topography of SSVEP amplitude differences between the mean amplitude during stimulus-on and the mean amplitude during the stimulus-off intervals. The Hotelling’s T values indicate the statistical strength of this difference. N.B. No contours are shown on the Hotelling’s T topographic map as p values for differences at all electrode locations are less than p=0.001
169

Appendix E. Sustained SSVEP effects for each
memory load condition relative to the baseline task:
practice tasks

Appendix E: Sustained SSVEP effects for each memory load condition
relative to the baseline task: practice tasks
baseline
load 1(practice)
load 3(practice)
load 5(practice)
5.0 +3.1
0.0 Vυ -3.1 radianamplitude phase
Figure E.1 Mean SSVEP amplitude and phase topography for the baseline task and each of the practice memory load tasks for an individual subject
Mean SSVEP amplitude and phase topography for each of the practice memory load tasks, calculated across the entire 40s ‘task interval.’ The mean SSVEP amplitude and phase for the baseline task was calculated over the equivalent 40s period. Data taken from individual subject (CL).
171

Appendix E: Sustained SSVEP effects for each memory load condition
relative to the baseline task: practice tasks
baseline
load 1(practice)
load 3(practice)
load 5(practice)
1.5 +0.5
0.0 -0.5 radianamplitude phase
Figure E.2 Mean SSVEP amplitude and phase topography for the baseline task and each of the practice memory load tasks averaged across 40 subjects
Mean SSVEP amplitude and phase topography for each of the practice memory load tasks calculated across the entire 40s ‘task interval.’ The mean SSVEP amplitude and phase for the baseline task was calculated over the equivalent 40s period across 40 subjects.
172

Appendix F. Retrieval of everyday objects
HeWolff
Text Box
HeWolff
Rectangle

Appendix F: Retrieval of everyday objects
Introduction Method: data acquisition
Episodic memory retrieval refers to the process of accessing personally experienced past events (1).
Recent electrophysiological and neuroimaging studies suggest that episodic retrieval is associated with both state(tonic) and transient neural activity (2-4).
Tulving (1) suggested that for successful retrieval, an individual must be in a ‘tonically’ maintained cognitive state, termed retrieval mode.
Silberstein et al. (5) demonstrated attenuation of the amplitude of the s
In this study we investigate how the amplitude of a SSVEP is affected firstly by the retrieval mode and secondly by the memory load during the retrieval period.
teady-state visually evoked potential (SSVEP) during a visual vigilance task. This attenuation is considered to be indicative of increased regional cortical activity and appears akin to the regional reductions in alpha activity associated with cognitive tasks.
Brain electrical activity was recorded from 64 scalp sites that were referenced to the nose, with the chin serving as ground. Brain electrical activity was amplified and bandpass filtered (3dB down at 0.1 Hz and 80 Hz) prior to digitization to 16 bit accuracy at a sampling rate of 500 Hz (Fig. 2).
A continuous 13Hz irrelevant sinusoidal flicker, subtending angles of 165º horizontally and 95º vertically was superimposed on the task display (Fig. 2).
For each of the 64 electrodes, the SSVEP amplitude of the response to the irrelevant flicker was extracted from the background EEG using Fourier techniques (5). The average SSVEP amplitude was calculated for the control task and for the retrieval periods of each of the 5 retrieval tasks. The resulting 6 sets of 64 SSVEP amplitudes were subsequently averaged across all subjects.
To investigate the affect of retrieval mode on the SSVEP amplitude, the data from the 5 retrieval tasks were averaged and compared with the control task data.
To investigate the effect of memory load on the SSVEP amplitude, the 5 separate sets of retrieval data were compared.
Increases in memory load were associated with an increase in the mean response time (r=0.92; p<0.01, 1-tailed) (Fig. 3B). However, no relation between memory load and accuracy, determined by the number of correctly identified old objects was apparent (r=-0.109; NS, 1-tailed) (Fig. 3A).
Episodic memory retrieval is associated with sustained SSVEP amplitude reductions, predominantly in left parieto-temporal regions. Increasing memory load is associated with reductions in the SSVEP amplitude in right parieto-temporal regions.
1. Tulving (1983). 2. Donaldson., Petersen, S. E., Ollinger, J. M., & Buckner, R. L. (2001).
(1), 129-142.3. Duzel, E., Cabeza, R., Picton, T. W., Yonelinas, A. P., Scheich, H.,
Heinze, H. J., & Tulving, E. (1999). (4), 1794-9.
4. Rugg, M. D., & Wilding, E. L. (2000). (3), 108-115.5. Silberstein, R., Ciorciari, J., & Pipingas, A. (1995).
, 24-35.
Elements of Episodic Memory.
Neuroimage, 13
Proc Natl Acad Sci U S A, 96
Trends Cogn Sci, 4
Electroencephalography and Clinical Neurophysiology, 96
Episodic memory retrieval was associated with sustained SSVEP amplitude reductions relative to the control task over parieto-temporal, parietal, temporal and frontal regions. The largest reductions were at left parieto-temporal regions (Fig. 4A).
The consistency of these differences across subjects was examined using the Student’s t score. The highest scores were observed at left parieto-temporal sites (Fig. 4B).
A negative correlation between SSVEP amplitude and memory load was found at parieto-temporal sites, particularly in the right hemisphere, indicating that with increasing memory load the SSVEP amplitude at these sites was reduced (Fig. 5).
Fig. 2. Recording set-up with 13 Hz irrelevant flicker
Fig. 4. A. Topographic map of the SSVEP amplitude of the combined retrieval tasks relative to the control task. Warmer colours represent a relative SSVEP amplitude attenuation during the retrieval task. B. Topography of Student’s t score.
Fig. 5. Topographic distribution of Pearson’s correlation coefficients. Cooler colours represent negative correlations between SSVEP amplitude during retrieval and memory load.
Fig. 1. Cognitive task design. Example images shown for a memory load of 2 and for the control task.
Method: subjects and cognitive task
Twenty-eight right-handed male subjects aged from 18 to 30 years (mean=22.5) participated in the study.
All subjects performed 5 retrieval tasks and 1 control task (see Fig. 1). The order of task presentation was counterbalanced across subjects. Each of the 5 retrieval tasks commenced with an encoding interval during which a picture of 1 everyday object or sets of 2,3,4 or 5 everyday objects were presented sequentially. Each object was displayed five times for 2 seconds. Subjects were instructed to pay careful attention as recognition of these images would be tested in the ensuing retrieval interval.
In the retrieval interval of each task, new and previously displayed (old) objects were presented for 2 seconds. The probability of an old object being presented was 0.25. Subjects were required to respond using a forced choice button press, yes or no, corresponding to old/new objects respectively. For the control task subjects were merely required to press alternately yes and no buttons in response to pictures of everyday objects similar to those in the retrieval task.
-0.5
R
B. Paired -test tA. Difference
Retrieval Mode and SSVEP amplitude
Effect of memory load on SSVEP amplitude
-2.8+0.5 +2.8
p=0.05 0.01
0.95
1 2 3 4 5
1.00
1.05
1.10
1.15
Pearson’s r
-1.0
p=0.01 0.05
+1.0
Memory load
r p=-0.967; =0.007
SSV
EP a
mpl
itude
Fig. 3. Behavioural data: Number of correctly identified old objects(accuracy) (A) and time to respond to these objects (B) versus memory load.
Memory Load1 2 3 4 5
Acc
urac
y(%
)
88
90
92
94
96
98
Memory Load1 2 3 4 5
Res
pons
etim
e(m
sec.
)
540
560
580
600
620
640
660
680
700
Res
pons
e Ti
me
(mse
c.)
Memory Load
680
640
600
560
5401 2 3 4 5
700
660
620
580
Memory Load1 2 3 4 5
88
90
92
94
96
98
Acc
urac
y (%
)
yes
yesno no no no no noyes
yes
yesno no no noyes yes
Accuracy and Response Timeversus Memory Load
A B
Figure F.1 Retrieval of everyday objects
Poster presented at the Seventh Annual Meeting of the Orgainsation for Human Brain Mapping in Brighton, UK, 10-14 June 2001
174
HeWolff
Rectangle
HeWolff
Text Box
Image not available - see printed version

Publications by the author
Journal Articles
Farrow, M., Silberstein, R.B., Levy, F., Pipingas, A., Wood, K., Hay, D.A. and Jarman,
F.C. (1996). “Prefrontal and parietal deficits in ADHD suggested by brain
electrical activity mapping during children's performance of the AX CPT.”
Educational and Developmental Psychologist (The Australian), 13, (1), 59-68.
Harris, P. G., Silberstein, R. B., Nield, G. and Pipingas, A. (2001). “Frontal lobe
contributions to perception of rhythmic group structure.” Annals of The New
York Academy of Sciences: The Biological Foundations of Music, 930, 414-417.
Patterson, J., Owen, C.M., Silberstein, R.B., Simpson, D.G., Pipingas, A. and Nield, G.
(1998). “Steady state visual evoked potential (SSVEP) changes in response to
olfactory stimulation.” Annals of the New York Academy of Sciences, 855, 625-
7.
Silberstein, R. B., Ciorciari, J. and Pipingas, A. (1993). “Rapid changes in steady-state
visually evoked potential topography associated with the Wisconsin card sort.”
Biological Psychology, 37, 43-71.
Silberstein, R. B., Ciorciari, J., and Pipingas, A. (1995). ‘Steady-state visually evoked
potential topography during the Wisconsin card sorting test.’
Electroencephalography and Clinical Neurophysiology, 96, (1), 24-35.
Silberstein, R.B., Farrow, M., Levy, F., Pipingas, A., Hay, D.A. and Jarman, F.C.
(1998). “Functional brain electrical activity mapping in boys with attention-
deficit/hyperactivity disorder.” Archives of General Psychiatry, 55, (12), 1105-
1112.
Silberstein, R. B., Harris, P. G., Nield, G. E. and Pipingas, A. (2000). “Frontal steady-
state potential changes predict long-term recognition memory performance.”
International Journal of Psychophysiology, 39, (1), 79-85.
Silberstein, R. B., Line, P., Pipingas, A., Copolov, D., and Harris, P. (2000). “Steady-
state visually evoked potential topography during the continuous performance
175

task in normal controls and schizophrenia.” Clinical Neurophysiology, 111, (5),
850-7.
Silberstein, R. B., Nunez, P. L., Pipingas, A., Harris, P. and Danieli, F. (2001). “Steady
state visually evoked potential (SSVEP) topography in a graded working
memory task.” International Journal of Psychophysiology, 42, 219-232.
Silberstein, R.B., Schier, M.A., Pipingas, A., Ciorciari, J., Wood, S.R. and Simpson,
D.G. (1990). “Steady-state visually evoked potential topography associated
with a visual vigilance task.” Brain Topography, 3, (2), 337-347.
Van Rooy, C., Stough, C., Pipingas, A., Hocking, C. and Silberstein, R. B. (2001).
“Spatial working memory and intelligence: Biological correlates.” Intelligence,
29, (4), 275-292.
Wallace, J. G., Silberstein, R. B., Bluff, K. and Pipingas, A. (1994). “Semantic
transparency, brain monitoring and evaluation of hybrid cognitive architectures.”
Connection Science, 6, (1), 43-58.
Wheaton, K. J., Pipingas, A., Silberstein, R. B. and Puce, A. (2001). “Human neural
responses elicited to observing the actions of others.” Visual Neuroscience, 18,
(3), 401-406.
Book Chapters
Harris, P. G., Silberstein, R. B., Pipingas, A. and Pressing, J. (1999). “Perceptual
grouping of pitch sequences in the steady-state visually evoked potential (SSVEP)
responses of musically trained subjects.” Music, Mind and Science, S. W. Yi, ed.,
Seoul National University Press, Seoul, Korea, 144-165.
Conference Papers
Buchan, R.J., Nagata, K., Silberstein, R.B., Nield, G., Shinohara, T., Sato, M., Pipingas,
A., Simpson, D. and Hirata, Y. (1998). “Steady state probe topography and
PET during a japanese continuous performance task.” Brain Topography Today
- Proceedings of the III Pan-Pacific Conference on Brain Topography (BTOPPS
III), 183-188. Paper presented at the III Pan-Pacific Conference on Brain
176

Topography (BTOPPS III) in Tokyo Bay, Japan, 1-4 April, 1997.
Harris, P. G., Silberstein, R. B., Pipingas, A. and Pressing, J. (1998). “Steady-state
visually evoked potential (SSVEP) responses to changes of note duration in
pitch sequences.” Proceedings of the Fifth International Conference on Music
Perception and Cognition, 115-121. Paper presented at the Fifth International
Conference on Music Perception and Cognition in Seoul, Korea, 26-30 August,
1998.
Nield, G. E., Silberstein, R. B., Pipingas, A., Simpson, D. G. and Burkitt, G. (1998).
“Effects of Visual Vigilance Task on Gamma and Alpha Frequence Range
Steady-State Potential (SSVEP) Topography.” Brain Topography Today -
Proceedings of the III Pan-Pacific Conference on Brain Topography (BTOPPS
III), 189-194. Paper presented at the III Pan-Pacific Conference on Brain
Topography (BTOPPS III) in Tokyo Bay, Japan, 1-4 April, 1997.
Pipingas, A. and Silberstein, R. B. (1995). “SSVEP changes with memory load in a
visual recognition task.” Recent Advances in Event-Related Brain Potential
Research. Proceedings of the 11th International Conference on Event-Related
Potentials (EPIC)., Paper presented at the 11th International Conference on
Event-Related Potentials (EPIC). in Okinawa, Japan, 25-30 June 1995.
Senova, M., Nagata, K., Buchan, R., Silberstein, R.B., Matsuoka, S., Nield, G.,
Pipingas, A., Simpson, D.G. and Yaguchi, K. (1998). “Effects of smoking on
brain dynamics during the continuous performance task: A pilot study.” Brain
Topography Today: Proceedings of the III Pan-Pacific Conference on Brain
Topography (BTOPPS III), 822-4. Paper presented at the III Pan-Pacific
Conference on Brain Topography (BTOPPS III) in Tokyo Bay, Japan, 1-4 April
1997.
Silberstein, R. B., Cadusch, P. J., Nield, G., Pipingas, A. and Simpson, D. G. (1996).
“Dynamic changes in the topography of the Steady State Visually Evoked
Potential associated with cognition.” Recent Advances in Event-Related Brain
Potential Research. Proceedings of the 11th International Conference on Event-
Related Potentials (EPIC), Paper presented at the Proceedings of the 11th
International Conference on Event-Related Potentials (EPIC) in Okinawa, Japan,
25-30 June, 1995.
Silberstein, R. B., Ciorciari, J., Schier, M. A., Pipingas, A. and Wood, S. (1991). “The
177

steady-state visually evoked potential topography and vigilance.” Brain
impairment: Advances in applied research, 267-274.
Silberstein, R. B., Pipingas, A., Ciorciari, J., Schier, M. A. and Wood, S. R. (1991).
“Dynamic changes in brain evoked potential laterality in a visual vigilance task.”
Brain Impairment: Advances in applied research., 267-274.
Conference Abstracts
Aranda, G., Silberstein, R. B., Nield, G. and Pipingas, A. (2000). “ Steady state visually
evoked potential topography of unfamiliar facial processing.” International
Journal of Psychophysiology, 35, (1), 31. Paper presented at the 10th World
Congress of the International Organization of Psychophysiology in Sydney
Australia, 8-13 February 2000.
Balog, O., Silberstein, R. B. and Pipingas, A. (1994). “Steady-state visually evoked
potential topography in visual and auditory variants of the continuous
performance task (CPT).” Fourth Australasian Psychophysiology Conference
Abstracts, Paper presented at the Fourth Australasian Psychophysiology
Conference in Melbourne, Australia., December 1994.
Buchan, R., Nagata, K., Silberstein, R.B., Nield, G., Shinohara, T., Satoh, M., Pipingas,
A., Simpson, D.G. and Hirata, Y. (1998). “Simultaneous steady state probe
topography (SSPT) and positron emission tomography (PET) during a Japanese
visual vigilance task.” Proceedings of the 3rd Australian Symposium on
Functional Brain Mapping, 25. Paper presented at the 3rd Australian
Symposium on Functional Brain Mapping in Howard Florey Institute of
Experimental Physiology and Medicine, University of Melbourne, Melbourne,
Australia, 16-17 April 1998.
Buchan, R., Nagata, K., Silberstein, R.B., Nield, G., Shinohara, T., Satoh, M., Pipingas,
A., Simpson, D.G. and Hirata, Y. (1997). “Simultaneous Steady State Probe
Topography (SSPT) and PET Cerebral Blood Flow Measurements During a
Japanese Visual Vigilance Task.” Proceedings of the Seventh Australasian
Psychophysiology Conference, Paper presented at the Seventh Australasian
Psychophysiology Conference in Swinburne University of Technology,
Melbourne, Australia, 6-8 December, 1997.
178

Carter, J.D., Farrow, M., Silberstein, R.B., Tucker, A., Stough, C. and Pipingas, A.
(2000). “Functional brain mapping in ADHD and control children during
performance of the Stop-Signal Task.” Program of the 10th Annual Conference
of the Australasian Society for Psychophysiology (ASP), 18. Paper presented at
the 10th Annual Conference of the Australasian Society for Psychophysiology
(ASP) in Adelaide, Australia, 3-5 December 2000.
Carter, J. D., Farrow, M., Silberstein, R. B., Tucker, A., Stough, C. and Pipingas, A.
(2000). “The stop-signal task: a comparison of auditory and visual stop-signals
set proportional to mean reaction time.” International Journal of
Psychophysiology, 35, 42. Poster presented at the 10th World Congress of the
International Organization of Psychophysiology in Sydney, Australia, 8-13
February 2000.
Chua, P., Egan, G. F., Morris, P. L. P., Saling, M., Pipingas, A., Berlangieri, S. U., Fitt,
G., Schweitzer, I. and Burrows, G. D. (1995). “A Positron Emission
Tomography (PET) activation study of an orthographic lexical retrieval task.”
The Australian Society for Psychiatric Research Annual Scientific Meeting
Abstract, Paper presented at the The Australian Society for Psychiatric Research
Annual Scientific Meeting November, 1995.
Farrow, M., Silberstein, R.B., Pipingas, A., Hay, D.A., Levy, F. and Jarman, F.C.
(1997). “Brain electrical activity mapping during the continuous performance
task in attention deficit hyperactivity disorder.” Proceedings of the Seventh
Australasian Psychophysiology Conference, Paper presented at the Seventh
Australasian Psychophysiology Conference in Melbourne, Australia, 6-8
December, 1997.
Farrow, M., Silberstein, R. B., Pipingas, A., Hay, D. A., Levy, F. and Jarman, F. C.
(1997). “Brain electrical activity mapping in attention deficit hyperactivity
disorder.” Brain Sciences Institute Symposium Program and Abstracts, Paper
presented at the Brain Sciences Institute Symposium in Melbourne, Australia, 4-
5 December, 1997.
Farrow, M., Silberstein, R. B., Pipingas, A., Levy, F., Jarman, F. C. and Hay, D. A.
(1998). “Functional brain electrical activity mapping in attention deficit
hyperactivity disorder.” The Faculty of Child and Adolescent Psychiatry, The
Royal Australian and New Zealand College of Psychiatrists Abstracts, 11th
179

Annual Conference, Paper presented at the The Faculty of Child and Adolescent
Psychiatry, The Royal Australian and New Zealand College of Psychiatrists,
11th Annual Conference in Sydney, Australia, 24 October 1998.
Farrow, M. A., Silberstein, R. B., Sergejew, A. A., Hay, D. A., Levy, F., Pipingas, A.
and Jarman, F. C. (1996). “High temporal resolution functional brain electrical
activity mapping in attention deficit hyperactivity disorder.” European
Neuropsychopharmacology, 6, (Suppl. 3), 203. Paper presented at the XXth
Collegium Internationale Neuro-psychopharmacologicum Congress in
Melbourne, Australia, 23-27 June 1996.
Farrow, M. A., Silberstein, R. B., Sergejew, A. A., Hay, D. A., Pipingas, A., Wood, K.,
Levy, F. and Jarman, F. C. (1995). “Dynamics of brain electrical activity
topography in normal and ADHD children.” The Australasian Society for
Psychiatric Research - Annual Scientific Meeting Abstracts, 62. Paper presented
at the The Australasian Society for Psychiatric Research - Annual Scientific
Meeting in Melbourne, Australia, 30 November - 1 December 1995.
Harris, P. G., Silberstein, R. B. and Pipingas, A. (1998). “Steady-state visually evoked
potential (SSVEP) responses correlate with musically trained subjects' working
memory task performance.” Australian Journal of Psychology, 50 (supplement),
90. Paper presented at the 33rd Annual Conference of the Australian
Psychological Society in Melbourne, Australia, 30 September - 4 October 1998.
Harris, P. G., Silberstein, R. B., Pipingas, A. and Pressing, J. (1997). “Steady-state
visually evoked potential responses to changes in the note duration in pitch
sequence.” Proceedings of the Seventh Australasian Psychophysiology
Conference, Paper presented at the Seventh Australasian Psychophysiology
Conference in Swinburne University of Technology, Melbourne, Australia, 6-8
December, 1997.
Line, P., Silberstein, R. B. and Pipingas, A. (1992). “Steady-state visually evoked
potential topography and mental rotation.” 2nd Australasian Psychophysiology
Conference, Paper presented at the 2nd Australasian Psychophysiology
Conference in Nelson Bay, Australia, 1992.
Owen, C., Patterson, J., Silberstein, R. B., Simpson, D. G., Nield, G. E. and Pipingas, A.
(1997). “Respiratory Monitoring and Stimulus Delivery Apparatus for use with
Brain Electrical Activity Recording.” Chemical Senses. Proceeding of the
180

International Symposium on Olfaction and Taste., 22, 765. Paper presented at
the International Symposium on Olfaction and Taste. in San Diego, California,
July, 1997.
Pantelis, C., Egan, G., Maruff, P., Pipingas, A., O'Keefe, G., Velakoulis, D., Collinson,
S. and Chua, P. (1995). “Practice dependent alterations in activation of the
anterior cingulate cortex during the Stroop task: a positron emission tomography
study.” Functional Brain Mapping Symposium Abstracts, Paper presented at the
Functional Brain Mapping Symposium in Melbourne, Australia, November,
1995.
Pantelis, C., Egan, G. F., Maruff, P., Velakoulis, D., Pipingas, A., Tharan, A. S.,
Tochon-Danguy, H. J. and Stuart, G. (1998). “Functional neuroanatomy of
stroop performance in schizophrenia: Inhibition and facilitation with practice.”
Proceedings of the 3rd Australian Symposium on Functional Brain Mapping,
21. Poster presented at the 3rd Australian Symposium on Functional Brain
Mapping in Howard Florey Institute of Experimental Physiology and Medicine,
University of Melbourne, Melbourne, Australia, 16-17 April 1998.
Pantelis, C., Egan, G. F., Maruff, P., Velakoulis, D., Pipingas, A., Tochon-Danguy, H.
J. and McKay, W. J. (1997). “Functional neuroanatomy of attentional
abnormalities in chronic schizophrenia.” Second Functional Brain Mapping
Symposium Abstracts., Paper presented at the Second Functional Brain Mapping
Symposium in Newcastle, Australia, 2 February, 1997.
Pipingas, A. (1991). “Topography variations in the steady-state visually evoked
potential during a recognition memory task.” International OSET Congress
Abstracts, Paper presented at the International OSET in Melbourne, Australia,
1991.
Pipingas, A. and Maruff, P. T. (1991). “Sources of timing errors in IBM PC controlled
experiments.” International OSET Congress Abstracts, Paper presented at the
International OSET Congress in Melbourne, Australia, 1991.
Pipingas, A., Silberstein, R., Van Rooy, C. and Aranda, G. (2001). “State dependent
changes in the steady-state visually evoked potential amplitude associated with
retrieval of everyday objects.” Neuroimage, 13, (6 (part 2)), S726. Poster
presented at the Seventh Annual Meeting of the Orgainsation for Human Brain
Mapping in Brighton, UK, 10-14 June 2001.
181

Pipingas, A. and Silberstein, R. B. (1993). “SSVEP in a memory scanning task.”
Australasian conference on physical science and engineering in medicine and
the biomedical engineering conference abstracts, Paper presented at the
Australasian conference on physical science and engineering in medicine and the
biomedical engineering conference. in Melbourne, Australia, 1993.
Pipingas, A. amd Silberstein, R. B. (1994). “Steady-state probe topography in a
memory scanning task.” Pan Pacific Conference on Brain Electric Topography
Abstracts, Paper presented at the Pan Pacific Conference on Brain Electric
Topography in Sydney, Australia, 1994.
Pipingas, A., Silberstein, R. B., Maruff, P., Pantelis, C., Egan, G. F. and Velakoulis, D.
(1997). “Topographic variations in the steady state visually evoked potential
during the Stroop task.” Second Functional Brain Mapping Symposium
Abstracts., Paper presented at the Second Functional Brain Mapping Symposium
in Newcastle, Australia, 2 February, 1997.
Pipingas, A., Silberstein, R. B. and Nield, G. E. (2000). “Correlation between pre-
target 13Hz SSVEP amplitude and response time in a visual vigilance task.”
Brain Topography , 12, (4), 314. Poster presented at the 10th World Congress
of the International Society for Brain Electromagnetic Topography 1999
(ISBET99) in Adelaide, Australia, 9-13 October, 1999.
Pipingas, A., Silberstein, R. B. and Nield, G. E. (1998). “Correlation between response
time and the 13Hz SSVEP amplitude preceding target presentation in a visual
vigilance task.” 8th Australasian Psychophysiology Conference Abstracts, 40.
Paper presented at the The 8th Australasian Psychophysiology Conference and
Annual Meeting of the Australasian Society for Psychophysiology in University
of Queensland, Brisbane, Australia, 4-6 December 1998.
Schier, M. A., Silberstein, R. B., Cadusch, P. J., Pipingas, A. and Wood, S. R. (1990).
“Spatial deconvolution of the steady state visually evoked potential topography.”
Fourth International Evoked Potential Symposium Abstracts., 161. Paper
presented at the Fourth International Evoked Potential Symposium in Toronto,
Canada, 1990.
Schier, M. A., Silberstein, R. B., Pipingas, A. and Ciorciari, J. (1993). “Steady-state
visually evoked potentials during a continuous performance task.” Brain
Topography, 5, 447.
182

Silberstein, R. B., Aranda, G., Pipingas, A. and Nield, G. (1998). “Changes in steady
state visually evoked potential associated with human face perception.”
Australasian Society for Psychiatric Research Program and Abstracts, P12.
Poster presented at the Australasian Society for Psychiatric Research in
University of Queensland, Brisbane, Australia, 1998.
Silberstein, R. B., Aranda, G., Pipingas, A., Nield, G. and Simpson, D. G. (1997).
“Dynamic Changes in the Steady-State Visually Evoked Potential Topography
Associated with Human Face Perception.” Neuroimage, S98. Poster presented
at the Third International Conference on Functional Mapping of the Human
Brain. in Copenhagen, Denmark, 19-23 May, 1997.
Silberstein, R. B., Burkitt, G. R., Line, P., Nield, G. E. and Pipingas, A. (1995).
“Steady-state visually evoked potential topography during the continuous
performance task (CPT): performance effects.” Fifth Australasian
Psychophysiology Conference Abstracts, Paper presented at the Fifth
Australasian Psychophysiology Conference in University of Wollongong,
Australia, 1995.
Silberstein, R. B., Ciorciari, J., Pipingas, A., Schier, M. A. and Ma, S. (1993). “Effects
of the Wisconsin Card Sort Test on the topography of the steady-state visually
evoked potential.” Brain Topography, 5, 447.
Silberstein, R. B. et al. (1998). Effects of Stimulant Medication on the Steady State
Visually Evoked Potential Latency Topography in Attention Deficit
Hyperactivity Disorder (ADHD). 3rd Australian Symposium on Functional
Brain Mapping Abstracts, 11. Paper presented at the 3rd Australian Symposium
on Functional Brain Mapping in University of Melbourne, Melbourne, Australia,
16-17 April , 1998.
Silberstein, R. B., Farrow, M. A., Levy, F., Pipingas, A., Jarman, F., and Hay, D. A.
(1997). “Steady state visually evoked potential latency topography in attention
deficit hyperactivity disorder.” Second Functional Brain Mapping Symposium
Abstracts., Paper presented at the Second Functional Brain Mapping Symposium
in Newcastle, Australia, 2 February, 1997.
Silberstein, R. B., Harris, P. G., Nield, G. E. and Pipingas, A. (2000). “Prefrontal
steady-state visually evoked potential (SSVEP) latency changes predict
recognition memory performance after 7 days.” International Journal of
183

Psychophysiology, 35, (1), 57. Paper presented at the 10th World Congress of
the International Organization of Psychophysiology (IOP) in Sydney, Australia,
8-13 February 2000.
Silberstein, R. B., Line, P., Nield, G. and Pipingas, A. (1994). “Steady State visually
evoked potential topography changes during the continuous performance task.”
Fourth Australasian Psychophysiology Conference Abstracts, Paper presented at
the Fourth Australasian Psychophysiology Conference in Melbourne, Australia.,
December 1994.
Silberstein, R. B., Nield, G., Pipingas, A. and Simpson, D. (1998). “Dynamic changes
in gamma and alpha frequency range steady state visually evoked potential
(SSVEP) in a visual vigilance task.” The XIIth International Conference on
Event-Related Potentials of the Brain (epic xii) Abstract Book, S01-02. Paper
presented at the The XIIth International Conference on Event-Related Potentials
of the Brain (epic xii) in Cambridge, Massachusetts, USA, 19-23 July, 1998.
Silberstein, R. B., Nield, G. E., Pipingas, A. and Simpson, D. G. (1997). “Changes in
the Alpha and Gamma Frequency Range Steady-State Visually Evoked Potential
(SSVEP) Topography during a Visual Vigilance Task.” Proceedings of the
Seventh Australasian Psychophysiology Conference, Paper presented at the
Seventh Australasian Psychophysiology Conference in Melbourne, Australia, 6-
8 December, 1997.
Silberstein, R. B., Pipingas, A., Ciorciari, J., Schier, M. and Ma, S. (1990). “Steady-
state visually evoked scalp topography in a visual vigilance task: effects of eye
movements.” Second International Congress on Brain Electromagnetic
Topography Abstracts, Paper presented at the Second International Congress on
Brain Electromagnetic Topography in Toronto, Canada, 1990.
Silberstein, R. B., Pipingas, A., Copolov, D., Line, P. and Harris, P. (2000). “Steady
state visually evoked potential topography during the continuous performance
task in normal controls and schizophrenia.” International Journal of
Psychophysiology, 35, (1), 21. Paper presented at the 10th World Congress of
the International Organization of Psychophysiology (IOP) in Sydney, Australia,
8-13 February 2000.
Silberstein, R. B., Pipingas, A., Harris, P. G., Nield, G., Saling, M., and O'Sullivan, B.
O. (1999). “Steady state visually evoked potential topography in a spatial
184

working memory task.” Society for Neuroscience Abstracts, 1, (25), 1142.
Poster presented at the Society for Neuroscience in Miami Beach, Florida, 23-28
October, 1999.
Silberstein, R. B., Pipingas, A., Saling, M. and O'Sullivan, B. (1998). “Steady state
visually evoked potential topography in a spatial working memory task.” The
8th Australasian Psychophysiology Conference Abstracts, 43. Paper presented
at the 8th Australasian Psychophysiology Conference and Annual Meeting of the
Australasian Society for Psychophysiology in University of Queensland,
Brisbane, Australia, 4-6 December, 1998.
Silberstein, R. B., Robb, D., Stanley, R., Burrows, G. and Pipingas, A. (1998). “Word
emotional valence influences steady state visually evoked potential (SSVEP)
amplitude and latency topography.” Brain Topography, 11, 71-2. Paper
presented at the The 9th World Congress of the International Society for Brain
Electromagnetic Topography 6-9 October, 1998.
Silberstein, R. B., Schier, M. A., Pipingas, A. and Ciorciari, J. (1991). “Topography of
phase and amplitude variations in the steady state visually evoked potential
during a vigilance task.” Abstracts of the Second International Congress on
Brain Electromagnetic Topography, Paper presented at the Second International
Congress on Brain Electromagnetic Topography in Toronto, Canada, 1991.
Silberstein, R. B., Schier, M. A., Pipingas, A., Ciorciari, J. and Simpson, D. G. (1990).
“Steady state probe topography, a new technique for measuring brain electrical
activity.” Proceedings of the 1990 Australian Society for Psychiatric Research
Scientific Meeting, Paper presented at the Australian Society for Psychiatric
Research Scientific Meeting in Melbourne, Australia, 1991.
Silberstein, R. B., Schier, M. A., Pipingas, A., Ciorciari, J., Wood, S. R. and Cadusch,
P. J. (1990). “Topography of the steady state visually evoked potential
associated with visual vigilance.” Fourth International Evoked Potential
Symposium Abstracts., 59. Paper presented at the Fourth International Evoked
Potential Symposium in Toronto, Canada, 1990.
Silberstein, R. B., Stough, C., Pipingas, A., Dennison, S. and Celi, E. (1996). “The
relationship between steady-state evoked potentials (SSVEPs) and cognitive
processes.” Australian Journal of Psychology, 48, 141. Paper presented at the
31st Annual Conference of the Australian Psychological Society September
185

1996.
Silberstein, R. B., Stough, C., Pipingas, A., Line, P., Celi, E. and Dennison, S. (1996).
“Steady state visually evoked potential topography in a mental rotation task:
Performance effects.” Paper presented at the Australian Psychophysiology
Conference December 1996.
Silberstein, R. B., Stough, C. K., Pipingas, A., Line, P., Celi, E. and Dennison, S.
(1997). “Dynamics of Steady-State Visually Evoked Potential Topography
Latency in a Mental Rotation Task: Performance Effects.” Brain Topography,
10, (1), 50. Paper presented at the 8th World Congress of the International
Society for Brain Electromagnetic Topography (ISBET) The Key Foundation
Symposium in Zurich, Switzerland, 6-8 March, 1997.
Silberstein, R. B., Wallace, I. G., Pipingas, A. and Bluff, K. (1993). “Steady-state
visually evoked potential topography in a counting task.” Third Australasian
Psychophysiology Conference Abstracts, Paper presented at the Third
Australasian Psychophysiology Conference
Silberstein, R. B., Wallace, J. G., Pipingas, A. and Bluff, K. (1992). “Steady-State
visually evoked topography and task automatization.” The Third International
Congress on Brain Electromagnetic Topography Abstracts, Poster presented at
the Third International Congress on Brain Electromagnetic Topography in
Amsterdam, Netherlands, 9-12 June, 1992.
Stough, C., Silberstein, R. B. S., Pipingas, A., Dennison, S., Celi, E. and Gillespie, N.
(1997). “The relationship between SSVEP and psychometric intelligence.”
Program and Abstracts of the Biennial Conference of the International Society
for the Study of Individual Differences , Poster presented at the Biennial
Conference of the International Society for the Study of Individual Differences
in Aarhus, Denmark, July 1997.
Stough, C. K., Silberstein, R. B., Celi, E., Pipingas, A. and Dennison, S. (1997). “The
relationship between steady-state visual evoked potentials (SSVEPs) and
intelligence.” Fifth European Congress of Psychology Abstracts, 32. Paper
presented at the Fifth European Congress of Psychology in Dublin, Ireland, 6-11
July, 1997.
Thompson, J., Tzambazis, K., Stough, C., Pipingas, A. and Silberstein, R. B. (2000).
“Changes in prefrontal steady state visually evoked potential in a visual
186

vigilance task: effects of nicotine.” International Journal of Psychophysiology,
35, (1), 33-34. Paper presented at the 10th World Congress of the International
Organization of Psychophysiology (IOP) in Sydney, Australia, 8-13 February
2000.
Thompson, J. C., Tzambazis, K., Stough, C., Pipingas, A., Silberstein, R. B. and
Nathan, P. J. (1999). “Changes to prefrontal electrophysiological activity
during enhanced attention following nicotine.” European
Neuropsychopharmacology, 9, (Suppl. 5), S356 (P.6.029). Poster presented at
the European College of Neuropsychopharmacology (ECNP) in London, UK,
21-25 Septemer, 1999.
Thompson, J. C., Tzambazis, K., Stough, C., Pipingas, A., Silberstein, R. B. and
Nathan, P. J. (1998). “Changes to prefrontal electrophysiological activity
during attention following nicotine.” The Australasian Society of Psychiatric
Research (ASPR) Conference Abstracts, Paper presented at the Australasian
Society of Psychiatric Research (ASPR) Conference in Brisbane, 1998.
Van Rooy, C., Stough, C. K., Pipingas, A. and Silberstein, R. B. (1997). “Spatial
Working Memory and Intelligence.” Proceedings of the Seventh Australasian
Psychophysiology Conference, Paper presented at the Seventh Australasian
Psychophysiology Conference in Melbourne, Australia, 6-8 December, 1997.
Wallace, J. G., Silberstein, R. B., Bluff, K. and Pipingas, A. (1993). “Learning in a
hybrid cognitive architecture: a brain monitoring study.” World Conference on
Artificial Intelligence in Education, Paper presented at the World Conference on
Artificial Intelligence in Education in Edinburgh, Scotland., 1993.
Wallace, J. G., Silberstein, R. B., Bluff, K. and Pipingas, A. (1992). “Semantic
tranparency, brain monitoring and the definition of hybrid systems.”
International Conference on Artificial Neural Networks, Paper presented at the
International Conference on Artificial Neural Networks 1992.
Wallace, J. G., Silberstein, R. B., Bluff, K. and Pipingas, A. (1992). “Semantic
transparency, brain monitoring and the integration of neural and symbolic
processes.” Tenth National Conference on Artificial Intelligence, Paper
presented at the Tenth National Conference on Artificial Intelligence in San
Hose., July, 1992.
Wheaton, K. J., Pipingas, A., Silberstein, R. B. and Puce, A. (2000). “ERPs elicited to
187

observing the actions of others.” International Journal of Psychophysiology, 35,
(1), 59-60. Paper presented at the 10th World Congress of the International
Organization of Psychophysiology (IOP) in Sydney, Australia, 8-13 February
2000.
188

Bibliography
Abt, K. (1983). “Significance testing of many variables - problems and solutions.”
Neuropsychobiology, 9, 47-51.
Acton, P. D., and Friston, K. J. (1998). “Statistical parametric mapping in functional
neuroimaging: beyond PET and fMRI activation studies.” European Journal of
Nuclear Medicine, 25(7), 663-667.
Adrian, E. D., and Matthews, B. H. C. (1934). “The berger thythm: Potential changes
from the occipital lobes in man.” Brain, 57, 355-385.
Appleton, J. P., and Brown, M. W. (1999). “Episodic memory, amnesia and the
hippocampal anterior thalamic axis.” Behavioural Brain Sciences, 22, 452-489.
Arnolds, D. E., Lopes da Silva, F. H., Aitink, J. W., Kamp, A., and Boeijinga, P. (1980).
“The spectral properties of hippocampal EEG related to behaviour in man.”
Electroencephalography and Clinical Neurophysioliology, 50(3-4), 324-8.
Baddeley, A. D. (1999). Essentials of human memory, Psychology Press, U.K.
Berger, H. (1929). “Uber das elektrenkephalogramm des menschen.” Arch Psychiat
Nervenkr 1929;87:527-570. English translation by Gloor P. Hans Berger on the
electroencephalogram of man. Electroencephalography and Clinical
Neurophysiology 1969;(Suppl. 28):37-73.
Besson, M., Kutas, M., and Van Petten, C. (1992). “An event-related potential (ERP)
analysis of semantic congruity and repetition effects in sentences.” Journal of
Cognitive Neuroscience, 4, 132-149.
Buckner, R. L., Koutstaal, W., Schacter, D. L., Dale, A. M., Rotte, M., and Rosen, B. R.
(1998). “Functional-anatomic study of episodic retrieval. II. Selective averaging
of event-related fMRI trials to test the retrieval success hypothesis.”
Neuroimage, 7(3), 163-175.
Buckner, R. L., and Tulving, E. (1995). “Neuroimaging studies of memory: theory and
recent PET results.” Handbook of Neuropsychology, J. C. Baron, ed., Elsevier,
New York, 439-466.
Buffalo, E. A., Reber, P. J., and Squire, L. R. (1998). “The human perirhinal cortex and
recognition memory.” Hippocampus, 8(4), 330-339.
189

Burkitt, G. (1996). “Steady-state visual evoked responses and travelling waves,” PhD
thesis, Swinburne University of Technology, Melbourne.
Cabeza, R. (1999). “Functional neuroimaging of episodic memory retrieval.” Memory,
consciousness and the brain: the Tallinn conference, E. Tulving, ed., Psychology
Press, Philadelphia, 76-90.
Cabeza, R., Grady, C. L., Nyberg, L., McIntosh, A. R., Tulving, E., Kapur, S., Jennings,
J. M., Houle, S., and Craik, F. I. (1997a). “Age-related differences in neural
activity during memory encoding and retrieval: a positron emission tomography
study.” Journal of Neuroscience, 17(1), 391-400.
Cabeza, R., Kapur, S., Craik , F., and McIntosh, A. (1997b). “Functional neuroanatomy
of recall and recognition: A PET study of episodic memory.” Journal of
Cognitive Neuroscience, 9(2), 254-265.
Cabeza, R., and Nyberg, L. (2000). “Imaging cognition II: An empirical review of 275
PET and fMRI studies.” Journal of Cognitive Neuroscience, 12(1), 1-47.
Cadusch, P. J., Breckon, W., and Silberstein, R. B. (1992). “Spherical splines and the
interpolation, deblurring and transformation of topographic EEG data.” Brain
Topography, 5.
Ciorciari, J., Silberstein, R. B., Simpson, D. G., and Schier, M. S. “The multichannel
electrode helmet.” Conference on Engineering and Physical Sciences in
Medicine, Melbourne, Australia.
Courtney, S. M., Ungerleider, L. G., Keil, K., and Haxby, J. V. (1996). “Object and
spatial visual working memory activate separate neural systems in human
cortex.” Cerebral Cortex, 6(1), 39-49.
Dawson, G. D. (1947). “Cerebral responses to electrical stimulation of peripheral nerve
in man.” Journal of Neurology, Neurosurgery, and Psychiatry, 10, 134-140.
Desimone, R., Albright, T. D., Gross, C. G., and Bruce, C. (1984). “Stimulus-selective
properties of inferior temporal neurons in the macaque.” Journal of
Neuroscience, 4(8), 2051-2062.
Desimone, R., and Duncan, J. (1995). “Neural mechanisms of selective visual
attention.” Annual Review Neuroscience, 18, 193-222.
Donaldson, D. I., and Rugg, M. D. (1998). “Recognition memory for new associations:
electrophysiological evidence for the role of recollection.” Neuropsychologia,
36(5), 377-395.
190

Donchin, E., Kutas, M., and McCarthy, G. (1977). “Electrocortical indices of
hemispheric utilization.” Lateralization in the nervous system, S. Harnad, R. W.
Doty, L. Godstein, J. Jaynes, and G. Krauthamer, eds., Academic Press, New
York, 339-384.
Doppelmayr, M., Klimesch, W., Schwaiger, J., Auinger, P., and Winkler, T. (1998).
“Theta synchronization in the human EEG and episodic retrieval.” Neuroscience
Letters, 257(1), 41-44.
Doppelmayr, M., Klimesch, W., Schwaiger, J., Stadler, W., and Rohm, D. (2000). “The
time locked theta response reflects interindividual differences in human memory
performance.” Neuroscience Letters, 278(3), 141-144.
Duffy, F. H., Bartels, P. H., and Burchfiel, J. L. (1981). “Significance probability
mapping: An aid in the topographic analysis of brain electrical activity.”
Electroencephalography and Clinical Neurophysiology, 51, 455-462.
Duffy, F. H., and McAnulty, G. (1988). “Electrophysiological studies.” Geriatric
neuropsychology, M. S. Albert and M. B. Moss, eds., The Guildford Press, New
York, 262-289.
Dujardin, K., Derambure, P., Bourriez, J. L., Jacquesson, J. M., and Guieu, J. M.
(1993). “P300 component of the event-related potentials (ERP) during an
attention task: effects of age, stimulus modality and event probability.”
International Journal of Psychophysiology, 14, 255-267.
Duzel, E., Cabeza, R., Picton, T. W., Yonelinas, A. P., Scheich, H., Heinze, H. J., and
Tulving, E. (1999). “Task-related and item-related brain processes of memory
retrieval.” Proceedings National Academy Sciences U S A, 96(4), 1794-1749.
Duzel, E., Picton, T. W., Cabeza, R., Yonelinas, A. P., Scheich, H., Heinze, H. J., and
Tulving, E. (2001). “Comparative electrophysiological and hemodynamic
measures of neural activation during memory-retrieval.” Human Brain Mapping,
13(2), 104-123.
Duzel, E., Yonelinas, A. P., Mangun, G. R., Heinze, H. J., and Tulving, E. (1997).
“Event-related brain potential correlates of two states of conscious awareness in
memory.” Proceedings National Academy Sciences U S A, 94(11), 5973-5978.
Ebbinghaus, H. (1964). Memory, Reprinted by Dover (originally published in 1885),
New York.
Fabiani, M., Karis, D., and Donchin, E. (1990). “Effects of mnemonic strategy
191

manipulation in a Von Restorff paradigm.” Electroencephalography and
Clinical Neurophysioliogy, 75(2), 22-35.
Farrow, M., Silberstein, R. B., Levy, F., Pipingas, A., Wood, K., Hay, D. A., and
Jarman, F. J. (1996). “Prefrontal and parietal deficits in ADHD suggested by
brain electrical activity mapping during children's performance of the.”
Educational and Developmental Psychologist (The Australian), 13(1), 59-68.
Felleman, D. J., and Van Essen, D. C. (1991). “Distributed hierarchical processing in
the primate cerebral cortex.” Cerebral Cortex, 1(1), 1-47.
Ferree, T. C., Luu, P., Russell, G. S., and Tucker, D. M. (2001). “Scalp electrode
impedance, infection risk, and EEG data quality.” Clinical Neurophysiology,
112(3), 536-544.
Friedman, D. (1990a). “Cognitive event-related potential components during continuous
recognition memory for pictures.” Psychophysiology, 27(2), 136-148.
Friedman, D. (1990b). “ERPs during continuous recognition memory for words.”
Biological Psychology, 30(1), 61-87.
Fujita, I., Tanaka, K., Ito, M., and Cheng, K. (1992). “Columns for visual features of
objects in monkey inferotemporal cortex [see comments].” Nature, 360(6402),
343-346.
Fuster, J. M., and Jervey, J. P. (1982). “Neuronal firing in the inferotemporal cortex of
the monkey in a visual memory task.” Journal of Neuroscience, 2(3), 361-375.
Galambos, R., and Makeig, S. “Studies on the auditory high-rates responses: Methods
and major findings.” Abstract in Proceedings of International conference on
dynamics of sensory and cognitive processing in the brain.
Galambos, R., and Makeig, S. (1988). “Dynamic changes in steady-state responses.”
Springer Series in Brain Dynamics 1, E. Basar, ed., Springer-Verlag, Berlin,
103-122.
Galin, D., and Ellis, R. (1975). “Asymmetry in evoked potentials as an index of
lateralized cognitive processes: Relation to EEG alpha asymmetry.”
Neuropsychologia, 13, 45-50.
Gardiner, J. M. (1988). “Functional aspects of recollective experience.” Memory
Cognition, 16, 309-313.
Gawne, T. J., and Richmond, B. J. (1993). “How independent are the messages carried
by adjacent inferior temporal cortical neurons?” Journal of Neuroscience, 13(7),
192

2758-2771.
Gloor, P. (1994). “Berger lecture. Is Berger's dream coming true?”
Electroencephalography and Clinical Neurophysiology, 90, 253-266.
Guillem, F., N'Kaoua, B., Rougier, A., and Claverie, B. (1995a). “Effects of temporal
versus temporal plus extra-temporal lobe epilepsies on hippocampal ERPs:
physiopathological implications for recognition memory studies in humans.”
Brain Research Cognitive Brain Research, 2(3), 147-153.
Guillem, F., N'Kaoua, B., Rougier, A., and Claverie, B. (1995b). “Intracranial
topography of event-related potentials (N400/P600) elicited during a continuous
recognition memory task.” Psychophysiology, 32(4), 382-392.
Guillem, F., N'Kaoua, B., Rougier, A., and Claverie, B. (1996). “Differential
involvement of the human temporal lobe structures in short- and long-term
memory processes assessed by intracranial ERPs.” Psychophysiology, 33(6),
720-730.
Halgren, E., Clarke, J. M., and Herve, N. (1992). “Roles des enregistrements profonds
dans la localisation, l'agencement sequential, et la caracterisation des etapes de
traitement de l'information dans le cerveau human.” Psychologie Francaise, 37,
149-166.
Halgren, E., and Smith, M. E. (1987). “Cognitive evoked potentials as modulatory
processes in human memory formation and retrieval.” Human Neurobiology,
6(2), 129-139.
Halgren, E., Squires, N. K., Wilson, C. L., Rohrbaugh, J. W., Babb, T. L., and Crandall,
P. H. (1980). “Endogenous potentials generated in the human hippocampal
formation and amygdala by infrequent events.” Science, 210(4471), 803-5.
Halliday. (1977). “Clinical applications of transient, flash visually evoked potentials.”
Visual evoked potentials in man: New developments, J. E. Desmedt, ed.,
Clarendon Press, Oxford.
Halliday, A. M., McDonald, W. I., and Mushin, J. (1972). “Delayed visual evoked
response in optic neuritis.” Lancet, 1, 982-985.
Harris, P. G., Silberstein, R. B., Nield, G. E., and Pipingas, A. (2001). “Frontal lobe
contributions to perception of rhythmic group structure. An EEG investigation.”
Annals of the New York Academy of Sciences, 930, 414-417.
Hassainia, F., Petit, D., and Montplaisir, J. (1994). “Significance probability mapping:
193

the final touch in t-statistic mapping.” Brain Topography, 7(1), 3-8.
Haxby, J. V., Grady, C. L., Horwitz, B., Ungerleider, L. G., Mishkin, M., Carson, R. E.,
Herscovitch, P., Schapiro, M. B., and Rapoport, S. I. (1991). “Dissociation of
object and spatial visual processing pathways in human extrastriate cortex.”
Proceedings National Academy Sciences U S A, 88(5), 1621-1625.
Heit, G., Smith, M. E., and Halgren, E. (1990). “Neuronal activity in the human medial
temporal lobe during recognition memory.” Brain, 113, 1093-1112.
Horel, J. A., Pytko-Joiner, D. E., Voytko, M. L., and Salsbury, K. (1987). “The
performance of visual tasks while segments of the inferotemporal cortex are
suppressed by cold.” Behavioural Brain Research, 23(1), 29-42.
Ibatoullina, A. A., Vardaris, R. M., and Thompson, L. (1994). “Genetic and
environmental influences on the coherence of background and orienting
response EEG in children.” Intelligence, 19, 65-78.
Iragui, V. J., Kutas, M., Mitchiner, M. R., and Hillyard, S. A. (1993). “Effects of aging
on event-related brain potentials and reaction times in an auditory oddball task.”
Psychophysiology, 30, 10-22.
Iwai, E. (1978). “The visual learning area in the inferotemporal cortex of monkeys.”
Integrative control functions of the brain, M. Ito, ed., Kodansha, Tokyo, 419-
427.
Iwai, E. (1981). “Visual mechanisms in the temporal and prestriate association cortices
of the monkey.” Advances Physiological Sciences, 17, 279-286.
Iwai, E. (1985). “Neuropsychological basis of pattern vision in macaque monkeys.”
Vision Research, 25(3), 425-439.
Iwai, E., and Mishkin, M. (1969). “Further evidence on the locus of the visual area in
the temporal lobe of the monkey.” Exp Neurol, 25(4), 585-94.
Jacoby, L. L. (1991). “A process dissociation framework: separating automatic from
intentional uses of memory.” Journal Memory Language, 30, 513-541.
Johnson, M. K., Hashtroudi, S., and Lindsay, D. S. (1993). “Source monitoring
[Review].” Psychology Bulletin, 114, 3-28.
Johnson, R. (1995). “Event-related potential insights into the neurobiology of memory
systems.” Handbook of Neuropsychology, J. C. Baron, ed., Elsevier, New York,
135-163.
Johnson, R., Jr., Pfefferbaum, A., and Kopell, B. S. (1985). “P300 and long-term
194

memory: latency predicts recognition performance.” Psychophysiology, 22(5),
497-507.
Johnstone, J., Galin, D., Fein, C., Yingling, C., Herron, J., and Marcus, M. (1984).
“Regional brain activity in dyslexic and control children during reading tasks:
visual probe event-related potentials.” Brain and Language, 21, 233-254.
Kandel, E. R., Schwartz, J. H., and Jessell, T. M. (2000). Principles of neural science,
McGraw-Hill, New York.
Kinsbourne, M., and Hiscock, M. (1983). “The normal and deviant development of
functional lateralization of the brain.” Carmichael's manual of child psychology,
P. H. Mussen, ed., John Wiley, New York, 157-208.
Klimesch, W. (1996). “Memory processes, brain oscillations and EEG
synchronization.” International Journal Psychophysiology, 24(1-2), 61-100.
Klimesch, W., Doppelmayr, M., Schimke, H., and Ripper, B. (1997). “Theta
synchronization and alpha desynchronization in a memory task.”
Psychophysiology, 34(2), 169-176.
Klimesch, W., Doppelmayr, M., Schwaiger, J., Winkler, T., and Gruber, W. (2000).
“Theta oscillations and the ERP old/new effect: independent phenomena?”
Clinical Neurophysiology, 111(5), 781-793.
Klimesch, W., Doppelmayr, M., Stadler, W., Pollhuber, D., Sauseng, P., and Rohm, D.
(2001a). “Episodic retrieval is reflected by a process specific increase in human
electroencephalographic theta activity.” Neuroscience Letters, 302(1), 49-52.
Klimesch, W., Doppelmayr, M., Yonelinas, A., Kroll, N. E., Lazzara, M., Rohm, D.,
and Gruber, W. (2001b). “Theta synchronization during episodic retrieval:
neural correlates of conscious awareness.” Brain Research Cognitive Brain
Research, 12(1), 33-38.
Klimesch, W., Pfurtscheller, G., Mohl, W., and Schimke, H. (1990a). “Event-related
desynchronization, ERD-mapping and hemispheric differences for words and
numbers.” International Journal Psychophysiology, 8(3), 297-308.
Klimesch, W., Pfurtscheller, G., and Schimke, H. (1992). “Pre- and post stimulus
processes in category judgement tasks as measured by event related
desynchronization (ERD).” Journal Psychophysiology, 6, 188-203.
Klimesch, W., Schimke, H., Ladurner, G., and Pfurtscheller, G. (1990b). “Alpha
frequency and memory performance.” Journal of Psychophysiology, 4, 381-390.
195

Klimesch, W., Schimke, H., and Pfurtscheller, G. (1993). “Alpha frequency, cognitive
load and memory performance.” Brain Topography, 5(3), 241-251.
Klimesch, W., Schimke, H., and Schwaiger, J. (1994). “Episodic and semantic memory:
an analysis in the EEG theta and alpha band.” Electroencephalography Clinical
Neurophysiology, 91(6), 428-441.
Kohler, S., Kapur, S., Moscovitch, M., Winocur, G., and Houle, S. (1995).
“Dissociation of pathways for object and spatial vision: a PET study in humans.”
Neuroreport, 6(14), 1865-1868.
Kolb, B., and Whishaw, I. (1996). “The occipital lobes.” Fundamentals of
Neuropsychology, W.H. Freeman and Co., New York, 243-264.
Krause, C. M., Lang, A. H., Laine, M., Kuusisto, M., and Porn, B. (1996). “Event-
related desynchronization and synchronization during an auditory memory task.”
Electroencephalography and Clinical Neurophysiology, 98, 319-326.
Kroll, N. E. A., and Klimesch, W. (1992). “Semantic memory. Complexity and
connectivity?” Mem.Cogn., 20, 192-210.
Kutas, M., and Dale, A. (1997). “Electrical and magnetic readings of mental functions.”
Cognitive Neuroscience, M. D. Rugg, ed., Psychology Press, London, 169-242.
Kutas, M., and Hillyard, S. A. (1980). “Reading senseless sentences: brain potentials
reflect semantic incongruity.” Science, 207(4427), 203-205.
Leonard, B. W., Amaral, D. G., Squire, L. R., and Zola-Morgan, S. (1995). “Transient
memory impairment in monkeys with bilateral lesions of the entorhinal cortex.”
Journal of Neuroscience, 15(8), 5637-5659.
Linden, R. D., Picton, T. W., Hamel, G., and Campbell, K. B. (1987). “Human auditory
evoked potentials during selective attention.” Electroencephalography and
Clinical Neurophysiology, 66, 145-159.
Logothetis, N. K., and Sheinberg, D. L. (1996). “Visual Object Recognition.” Annual
Review Neuroscience, 19, 577-621.
Mandler, G. (1981). “Recognizing: the judgment of prior occurrence.” Psychology
Review, 87, 252-271.
Martinson. (1939). “A study of brain potentials during mental blocking.” Journal of
Experimental Psychology, 21(1), 101-105.
McEwan, J. A., and Anderson, G. B. (1975). “Modeling the stationarity and gaussianity
of spontaneous electroencephalographic activity.” IEEE Transactions and
196

Biomedical Engineering, 22(5), 361-369.
Menon, R. S., and Kim, S. G. (1999). “Spatial and temporal limits in cognitive
neuroimaging with fMRI.” Trends in Cognitive Sciences, 3, 207-215.
Meunier, M., Bachevalier, J., Mishkin, M., and Murray, E. A. (1993). “Effects on visual
recognition of combined and separate ablations of the entorhinal and perirhinal
cortex in rhesus monkeys.” Journal of Neuroscience, 13(12), 5418-5432.
Milner, B. (1958). “Psychological defects produced by temporal lobe excision.” Recent
publications of the association for research in nervous and mental disease, 38,
244-257.
Milner, B. (1963). “Effects of different brain lesions on card sorting.” Archives of
Neurology, 9, 90-100.
Milner, B. (1964). “Some effects of frontal lobectomy in man.” The frontal granular
cortex and behaviour, J. M. Warren and K. Akert, eds., McGraw-Hill, New
York, 313-334.
Milner, B. (1968). “Visual recognition and recall after right temporal lobe excision in
man.” Neuropsychologia, 6, 191-209.
Milner, B. (1970). “Memory and the medial temporal regions of the brain.” Biological
Bases of Memory, K. H. Pribham and D. E. Broadbent, eds., Academic Press,
New York.
Mishkin, M. (1982). “A memory system in the monkey.” Philos Trans R Soc Lond B
Biol Sci, 298(1089), 83-95.
Moscovitch, C., Kapur, S., Kohler, S., and Houle, S. (1995). “Distinct neural correlates
of visual long-term memory for spatial location and object identity: a positron
emission tomography study in humans.” Proceedings National Academy
Sciences U S A, 92(9), 3721-3725.
Moscovitch, M. (1992). “Memory and working-with-memory: a component process
model based on modules and central systems.” Journal Cognitive Neuroscience,
4, 257-267.
Moscovitch, M. (1994). “Models of consciousness and memory.” The cognitive
neurosciences, M. S. Gazzaniga, ed., MIT Press, Cambridge, Mass.
Nagy, M. E., and Rugg, M. D. (1989). “Modulation of event-related potentials by word
repetition: the effects of inter-item lag.” Psychophysiology, 26(4), 431-436.
Neville, H. J., Kutas, M., Chesney, G., and Schmidt, A. L. (1986). “Event-related brain
197

potentials during initial encoding and recognition memory of congruous and
incongruous words.” Journal of Memory and Language, 25, 75-92.
Nield, G. E., Silberstein, R. B., Pipingas, A., Simpson, D. G., and Burkitt, G. R. (1998).
“The effects of a visual vigilance task on gamma and alpha frequency range
steady-state visually evoked potential (SSVEP) topography.” Brain Topography
Today, Y. Koga and K. Nagata, eds., Elsevier Science, Amsterdam, 189-194.
Nolde, S. F., Johnson, M. K., and Raye, C. L. (1998). “The role of prefrontal cortex
during tests of episodic memory.” Trends in cognitive sciences, 2(10), 399-406.
Noldy-Cullum, N. E., and Stelmack, R. M. (1987). “Recognition memory for pictures
and words: The effect of incidental and intentional learning on N400.” Current
research in event-related potentials. Electroencephalography and Clinical
Neurophysiology, Supplement 40, R. Johnson, J. W. Rohrbaugh, and R.
Parasuraman, eds., Elsevier, Amsterdam, 350-354.
Nunez, P. L. (1981). Electric fileds of the brain: the neurophysics of EEG, Oxford
University Press, New York.
Nyberg, L., Cabeza, R., and Tulving, E. (1996a). “PET studies of encoding and
retrieval: The HERA model.” Psychonomic Bulletin & Review, 3(2), 135-148.
Nyberg, L., Cabeza, R., and Tulving, E. (1998a). “Asymmetric frontal activation during
episodic memory: what kind of specificity?” Trends in Cognitive Sciences,
2(11), 419-420.
Nyberg, L., McIntosh, A. R., Houle, S., Nilsson, L. G., and Tulving, E. (1996b).
“Activation of medial temporal structures during episodic memory retrieval [see
comments].” Nature, 380(6576), 715-717.
Nyberg, L., McIntosh, A. R., and Tulving, E. (1998b). “Functional brain imaging of
episodic and semantic memory with positron emission tomography.” J Mol Med,
76(1), 48-53.
Oldfield, R. C. (1971). “The assessment and analysis of handedness: The Edinburgh
Inventory.” Neuropsychologia, 9, 97-113.
Osaka, M. (1984). “Peak alpha frequency of EEG during a mental task: task difficulty
and hemispheric differences.” Psychophysiology, 21(1), 101-105.
Paller, K. A., Bozic, V. S., Ranganath, C., Grabowecky, M., and Yamada, S. (1999).
“Brain waves following remembered faces index conscious recollection.” Brain
Research Cognitive Brain Research, 7(4), 519-531.
198

Paller, K. A., Gonsalves, B., Grabowecky, M., Bozic, V. S., and Yamada, S. (2000).
“Electrophysiological correlates of recollecting faces of known and unknown
individuals.” Neuroimage, 11(2), 98-110.
Paller, K. A., and Kutas, M. (1992). “Brain potentials during retrieval provide
neurophysiological support for the distinction between consious recollection and
priming.” Journal Cognitive Neuroscience, 4, 375-391.
Papanicolaou, A. C., Deutsch, G., Bourbon, W. T., Will, K. W., Loring, D. W., and
Eisenberg, H. M. (1987). “Convergent evoked potential and cerebral blood flow
evidence of task-specific hemispheric differences.” Electroencephalography and
Clinical Neurophysiology, 66, 515-520.
Papanicolaou, A. C., Eisenberg, H. M., and Levy, R. S. (1983). “Evoked potential
correlates of left hemisphere dominance in covert articulation.” International
Journal of Neuroscience, 20, 289-294.
Papanicolaou, A. C., and Johnstone, J. (1984). “Probe evoked potentials: theory,
method, and applications.” International Journal of Neuroscience, 24, 107-131.
Parker, A., and Gaffan, D. (1998). “Interaction of frontal and perirhinal cortices in
visual object recognition memory in monkeys [In Process Citation].” European
Journal of Neuroscience, 10(10), 3044-3057.
Petri, H. L., and Mishkin, M. (1994). “Behaviorism, cognitivism and the
neuropsychology of memory.” American Scientist, 82, 30-37.
Pfurtscheller, G., and Aranibar, A. (1977a). “Event-related cortical desynchronization
detected by power measurements of scalp EEG.” Electroencephalography and
Clinical Neurophysiology, 42, 138-146.
Pfurtscheller, G., and Aranibar, A. (1977b). “Event-related cortical desynchronization
detected by power measurements of scalp EEG.” Electroencephalography and
Clinical Neurophysiology, 42(6), 817-826.
Pfurtscheller, G., and Klimesch, W. (1990). “Topographic display and interpretation of
event-related desynchronization during a visual-verbal task.” Brain Topography,
3, 85-93.
Pfurtscheller, G., and Klimesch, W. (1991). “Event-related desynchronization during
motor behaviour and visual information procesing.” Event-related brain
research, C. H. M. Brunia, G. Mulder, and M. N. Verbaten, eds., Elsevier,
Amsterdam, 58-65.
199

Picton, T. W. (1992). “The P300 wave of the human event-related potential.” Journal of
Clinical Neurophysiology, 9(4), 456-479.
Picton, T. W., Lins, O. G., and Scherg, M. (1995). “The recording and analysis of event-
related potentials.” Handbook of neuropsychology, F. Boller and J. Grafman,
eds., Elsevier Science B.V., Amsterdam, 3-73.
Pipingas, A., and Maruff, P. T. “Sources of timing errors in IBM PC controlled
experiments.” International OSET Congress, Melbourne, Australia.
Pipingas, A., Silberstein, R. B., and Nield, G. E. (2000). “Correlation between pre-target
SSVEP amplitude and response time in a visual vigilance task.” Brain
Topography, 12(4), 314; Poster presented at the 10th World Congress of the
International Society for Brain Electromagnetic Topography (ISBET), in
Adelaide, Australia, 9-13 October, 1999.
Potter, D. D., Pickles, C. D., Roberts, R. C., and Rugg, M. D. (1992). “The effects of
Scopolamine in event-related potentials in a continuous recognition memory
task.” Psychophysiology, 29(1), 29-37.
Pritchard, W. S. (1981). “Psychophysiology of P300.” Psychol. Bull., 90, 506-540.
Puce, A., Andrewes, D. G., Berkovic, S. F., and Bladin, P. F. (1991). “Visual
recognition memory. Neurophysiological evidence for the role of temporal white
matter in man.” Brain, 114(Pt 4), 1647-1666.
Ranganath, C., and Paller, K. A. (1999). “Frontal brain activity during episodic and
semantic retrieval: insights from event-related potentials.” J Cogn Neurosci,
11(6), 598-609.
Ranganath, C., and Paller, K. A. (2000). “Neural correlates of memory retrieval and
evaluation.” Brain Research Cognitive Brain Research, 9(2), 209-222.
Ray, W. J., and Cole, H. W. (1985). “EEG alpha activity reflects attentional demands,
and beta activity reflects emotional and cognitive processes.” Science,
228(4700), 750-2.
Regan, D. (1972). Evoked potentials in psychology, sensory physiology and clinical
medicine, Chapman and Hall, London.
Regan, D. (1976). “Latencies of evoked potentials to flicker and to pattern speedily
estimated by simultaneous stimulation method.” Electroencephalography and
Clinical Neurophysiology, 40, 654-660.
Regan, D. (1977a). “Fourier analysis of evoked potentials; some methods based on
200

Fourier analysis.” Visual evoked potentials in man: New developments, J. E.
Desmedt, ed., Clarendon Press, Oxford, 110-117.
Regan, D. (1977b). “Steady-state evoked potentials.” J. Opt. Soc. Am., 67(11), 1475-
1488.
Regan, D. (1989). Human Electrophysiology, Elsevier, New York.
Regan, D., Milner, B. A., and Heron, J. R. (1977). “Slowing of visual signals in
multiple sclerosis, measured psychophysically and by steady-state evoked
potentials.” Visual evoked potentials in man: New developments, J. E. Desmedt,
ed., Clarendon Press, Oxford, 461-469.
Reid, R. (1999). “Vision.” Fundamental Neuroscience, F. Bloom, S. Landis, J. Roberts,
and L. Squire, eds., Academic Press, San Diego, 821-851.
Rockstroh, B., Elbert, T., Canavan, A., Lutzenberger, W., and Birbaumer, N. (1989).
Slow cortical potentials and behaviour, Urban and Schwarzenberg, Baltimore
M.D.
Rolls, E. T. (2000). “Functions of the primate temporal lobe cortical visual areas in
invariant visual object and face recognition.” Neuron, 27(2), 205-218.
Rugg, M. D. (1987). “Dissociation of semantic priming, word and non-word repetition
effects by event-related potentials.” Q. Journal Experimental Psychology, 39,
123-148.
Rugg, M. D. (1990). “Event-related brain potentials dissociate repetition effects of high
and low frequency words.” Memory and Cognition, 18, 367-379.
Rugg, M. D. (1995). “Cognitive event-related potentials: intracranial and lesion
studies.” Handbook of neuropsychology, F. Boller and J. Grafman, eds., Elsevier
Science B.V., Amsterdam, 165-185.
Rugg, M. D., Allan, K., and Birch, C. S. (2000). “Electrophysiological evidence for the
modulation of retrieval orientation by depth of study processing.” Journal
Cognitive Neuroscience, 12(4), 664-678.
Rugg, M. D., Cox, C. J., Doyle, M. C., and Wells, T. (1995). “Event-related potentials
and the recollection of low and high frequency words.” Neuropsychologia,
33(4), 471-484.
Rugg, M. D., and Doyle, M. C. (1992). “Event-related potentials and recognition
memory for low- and high-frequency words.” Journal Cognitive Neuroscience,
4, 69-79.
201

Rugg, M. D., and Nagy, M. E. (1989). “Event-related potentials and recognition
memory for words.” Electroencephalography and Clinical Neurophysiology,
72(5), 395-406.
Rugg, M. D., Roberts, R. C., Potter, D. D., Pickles, C. D., and Nagy, M. E. (1991).
“Event-related potentials related to recognition memory. Effects of unilateral
temporal lobectomy and temporal lobe epilepsy.” Brain, 114(Pt 5), 2313-2332.
Rugg, M. D., Schloerscheidt, A. M., Doyle, M. C., Cox, C. J., and Patching, G. R.
(1996). “Event-related potentials and the recollection of associative
information.” Brain Res Cogn Brain Res, 4(4), 297-304.
Rugg, M. D., and Wilding, E. L. (2000). “Retrieval processing and episodic memory.”
Trends in Cognitive Sciences, 4(3), 108-115.
Saletu, B., and Grunberger, J. (1985). “Memory dysfunction and vigilance:
neurophysiological and psychopharmacological aspects.” Annals New York
Academy Sciences, 444, 406-427.
Sanquist, T. F., Rohrbaugh, J. W., Syndulko, K., and Lindsley, D. B. (1980).
“Electrocortical signs of levels of processing: perceptual analysis and
recognition memory.” Psychophysiology, 17(6), 568-576.
Schacter, D. L. (1987). “Memory, amnesia, and frontal lobe dysfunction.”
Psychobiology, 25, 21-36.
Schacter, D. L. (1992). “Implicit knowledge: New perspectives on unconscious
processes.” Proceedings National Academy Sciences U S A, 89(23), 11113-
11117.
Schacter, D. L., and Crovitz, H. F. (1977). “Memory function after closed head injury:
A review of quantitative research.” Cortex, 13, 150-176.
Schacter, D. L., Kaszniak, A. W., Kihlstrom, J. F., and Valdiserri, M. (1991). “The
relation between source memory and aging.” Psychology Aging, 6(4), 559-568.
Scherg, M. (1990). “Fundamentals of dipole source potential analysis.” Auditory
evoked magnetic fields and potentials. Anvances in audiology, F. Grandori and
G. L. Romani, eds., Basel: Karger, 40-69.
Schier, M. A. (1994). “Human steady-state visually evoked potential topography and
attention,” PhD, University of Melbourne, Melbourne.
Schloerscheidt, A. M., and Rugg, M. D. (1997). “Recognition memory for words and
pictures: an event-related potential study.” Neuroreport, 8(15), 3281-3285.
202

Schmitt, F., Stehling, M. K., and Turner, R. (1998). “Echo-planar imaging. Theory,
technique and application.” , Springer-Verlag, Berlin, 662.
Scoville, W. B., and Milner, B. (1957). “Loss of recent memory after bilateral
hippocampal lesions.” Journal of Neurology, Neurosurgery and Psychiatry, 20,
11-21.
Sereno, M. I., Dale, A. M., Reppas, J. B., Kwong, K. K., Belliveau, J. W., Brady, T. J.,
Rosen, B. R., and Tootell, R. B. H. (1995). “Borders of multiple visual areas in
humans revealed by functional magnetic resonance imaging.” Science, 268, 889-
893.
Shucard, D. W., Shucard, J. L., and Thomas, D. G. (1977). “Auditory evoked potentials
as probes of hemispheric differences in cognitive processing.” Science, 197,
1295-1298.
Silberstein, R. B. (1998). “Neocortical resonances and holding memory.” Brain
Topography Today, Y. Koga, K. Nagata, and K. Hirata, eds., Elsevier Science,
Amsterdam, 34-38.
Silberstein, R. B., Burkitt, G. R., and Wood, S. R. (1993). “Artifact sensitivity of
Fourier-based analysis of steady-state visually evoked potentials.” Biological
Psychology, 37, 43-71.
Silberstein, R. B., and Cadusch, P. J. (1992). “Measurement processes and spatial
principal components analysis.” Brain Topography, 4, 267-276.
Silberstein, R. B., Cadusch, P. J., Nield, G. E., Pipingas, A., and Simpson, D. G. (1996).
“Steady-state visually evoked potential topography dynamics and cognition.”
Recent advances in event-related brain potential research, C. Ogura, Y. Koga,
and M. Shimokochi, eds., Elsevier, Amsterdam, 379-385.
Silberstein, R. B., Ciorciari, J., and Pipingas, A. (1995a). “Steady-state visually evoked
potential topography during the Wisconsin card sorting test.”
Electroencephalography and Clinical Neurophysiology, 96, 24-35.
Silberstein, R. B., Ciorciari, J., and Pipingas, A. (1995b). “Steady-state visually evoked
potential topography during the Wisconsin card sorting test.”
Electroencephalography and Clinical Neurophysiology, 96(1), 24-35.
Silberstein, R. B., Farrow, M., Levy, F., Pipingas, A., Hay, D. A., and Jarman, F. C.
(1998). “Functional brain electrical activity mapping in boys with attention-
deficit/hyperactivity disorder.” Archives of General Psychiatry, 55(12), 1105-
203

1112.
Silberstein, R. B., Line, P., Pipingas, A., Copolov, D., and Harris, P. (2000). “Steady-
state visually evoked potential topography during the continuous performance
task in normal controls and schizophrenia.” Clinical Neurophysiology, 111(5),
850-857.
Silberstein, R. B., Nunez, P. L., Pipingas, A., Harris, P., and Danieli, F. (2001). “Steady
state visually evoked potential (SSVEP) topography in a graded working
memory task.” International Journal of Psychophysiology, 42(2), 125-138.
Silberstein, R. B., Schier, M. A., Pipingas, A., Ciorciari, J., Wood, S. R., and Cadusch,
P. J. (1990a). “Topography of the steady state visually evoked potential
associated with visual vigilance.” Fourth International Evoked Potential
Symposium. Book of abstracts. Toronto, 1990. p. 59.
Silberstein, R. B., Schier, M. A., Pipingas, A., Ciorciari, J., Wood, S. R., and Simpson,
D. G. (1990b). “Steady-state visually evoked potential topography associated
with a visual vigilance task.” Brain Topography, 3(2), 337-347.
Simpson, D. G. (1997). “Instrumentation for high spatial resolution steady-state visually
evoked potentials,” Masters thesis, Swinburne University of Technology,
Melbourne.
Smith, M. E. (1993). “Neurophysiological manifestations of recollective experience
during recognition memory judgements.” Journal Cognitive Neuroscience, 5, 1-
13.
Smith, M. E., and Guster, K. (1993). “Decomposition of recognition memory event-
related potentials yields target, repetition, and retrieval effects.”
Electroencephalography and Clinical Neurophysiology, 86(5), 335-343.
Smith, M. E., and Halgren, E. (1988). “Attenuation of a sustained visual processing
negativity after lesions that include the inferotemporal cortex.”
Electroencephalography and Clinical Neurophysiology, 70(4), 366-369.
Smith, M. E., and Halgren, E. (1989). “Dissociation of recognition memory components
following temporal lobe lesions.” Journal of Experimental Psychology:
Learning Memory and Cognition, 15(1), 50-60.
Smith, M. E., Stapleton, J. M., and Halgren, E. (1986). “Human medial temporal lobe
potentials evoked in memory and language tasks.” Electroencephalography and
Clinical Neurophysiology, 63(2), 145-159.
204

Smith, M. L. (1989). “Memory disorderss associated with temporal-lobe lesions.”
Handbook of neuropsychology, F. Boller and J. Grafman, eds., Elsevier,
Amsterdam, 91-106.
Squire, L. R. (1992). “Declarative and non-declarative memory: multiple brain systems
supporting learning and memory.” Journal of Cognitive Neuroscience, 2, 232-
243.
Squire, L. R. (1994). “Declarative and nondeclarative memory: multiple brain systems
supporting learning and memory.” Memory systems, D. L. Schacter and E.
Tulving, eds., MIT Press, Cambridge (MA), 203-231.
Squire, L. R., and Zola, S. M. (1996). “Structure and function of declarative and
nondeclarative memory systems.” Proceedings of the National Academy of
Sciences U S A, 93(24), 13515-13522.
Squire, L. R., and Zola-Morgan, S. (1991). “The medial temporal lobe memory system.”
Science, 253, 1380-1386.
Stephenson, W. A., and Gibbs, M. D. (1951). “A balanced non-cephalic reference
electrode.” Electroencephalography and Clinical Neurophysiology, 3, 237-240.
Sternberg, S. (1969). “The Discovery of processing stages: extensions of Donder's
Method.” Attention and Performance II (Acta Psychol., 30), W. G. Koster, ed.,
North-Holland Publ., Amsterdam, 276-315.
Stuss, D. T., Picton, T. W., and Cerri, A. M. (1986). “Searching for the names of
pictures: An event-related potential study.” Psychophysiology, 23, 215-223.
Sutton, S., Braren, M., Zubin, J., and John, E. R. (1965). “Evoked potential correlates of
stimulus uncertainty.” Science, 150, 1187-1188.
Suzuki, W. A., and Amaral, D. G. (1994). “Perirhinal and parahippocampal cortices of
the macaque monkey: cortical afferents.” Journal of Comparative Neurology,
350(4), 497-533.
Tanaka, K. (1993). “Neuronal mechanisms of object recognition.” Science, 262(5134),
685-688.
Tendolkar, I., Doyle, M. C., and Rugg, M. D. (1997). “An event-related potential study
of retroactive interference in memory.” Neuroreport, 8(2), 501-506.
Tesche, C. D., Uusitalo, M. A., Ilmoniemi, R. J., and Kajola, M. (1995). “Characterizing
the local oscillatory content of spontaneous cortical activity during mental
imagery.” Cog Brain Res, 2, 243-249.
205

Tootell, R. B., Dale, A. M., Sereno, M. I., and Malach, R. (1996). “New images from
human visual cortex.” Trends in Neurosciences, 19(11), 481-489.
Tootell, R. B., Mendola, J. D., Hadjikhani, N. K., Ledden, P. J., Liu, A. K., Reppas, J.
B., Sereno, M. I., and Dale, A. M. (1997). “Functional analysis of V3A and
related areas in human visual cortex.” Journal of Neuroscience, 17(18), 7060-
7078.
Tulving, E. (1983). Elements of episodic memory, Oxford University Press, New York.
Tulving, E. (1993). “What is episodic memory?” Current Directions in Psychological
Sciences, 2, 67-70.
Tulving, E., Kapur, S., Craik, F. I. M., Moscovitch, M., and Houle, S. (1994a).
“Hemispheric encoding/retrieval asymmetry in episodic memory: positron
emission tomography findings.” Proceedings of the National Academy of
Sciences U S A, 91(6), 2016-2020.
Tulving, E., Kapur, S., Markowitsch, H. J., Craik, F. I., Habib, R., and Houle, S.
(1994b). “Neuroanatomical correlates of retrieval in episodic memory: auditory
sentence recognition.” Proceedings of the National Academy of Sciences U S A,
91(6), 2012-2015.
Tulving, E., and Schacter, D. L. (1990). “Priming and human memory systems.”
Science, 247, 301-306.
Ungerleider, L. G., and Haxby, J. V. (1994). “'What' and 'where' in the human brain.”
Current Opinion in Neurobiology, 4(2), 157-165.
Ungerleider, L. G., and Mishkin, M. (1982). “Two cortical visual systems.” Analysis of
visual behavior, D. J. Ingle, M. A. Goodale, and R. J. W. Mansfield, eds., MIT
Press, Cambridge, Mass.
Van Essen, D. C. (2002). “Organization of visual areas in macaque and human cerebral
cortex.” Visual Neurosciences, L. M. Chalupa and J. S. Werner, eds., MIT Press,
Cambridge MA.
Von Bonin, G., and Bailey, P. (1947). The Neocortex of Macaca Mulatta, Univ. Ill.
Press, IL.
Von Bonin, G., and Bailey, P. (1950). The Neocortex of the Chimpanzee, Univ. Ill.
Press, IL.
Waugh, N. C., and Norman, D. A. (1965). “Primary memory.” Psychological Review,
72, 89-104.
206

Webster, M. J., Bachevalier, J., and Ungerleider, L. G. (1993). “Subcortical connections
of inferior temporal areas TE and TEO in macaque monkeys.” Journal of
Comparative Neurology, 335(1), 73-91.
Webster, M. J., Ungerleider, L. G., and Bachevalier, J. (1991). “Connections of inferior
temporal areas TE and TEO with medial temporal- lobe structures in infant and
adult monkeys.” Journal of Neuroscience, 11(4), 1095-1116.
Wilding, E. L. (1999). “Separating retrieval strategies from retrieval success: an event-
related potential study of source memory.” Neuropsychologia, 37(4), 441-454.
Wilding, E. L. (2000). “In what way does the parietal ERP old/new effect index
recollection?” International Journal of Psychophysiology, 35(1), 81-87.
Wilding, E. L., Doyle, M. C., and Rugg, M. D. (1995). “Recognition memory with and
without retrieval of context: an event-related potential study.”
Neuropsychologia, 33(6), 743-767.
Wilding, E. L., and Rugg, M. D. (1996). “An event-related potential study of
recognition memory with and without retrieval of source [published erratum
appears in Brain 1996 Aug;119(Pt 4):1416].” Brain, 119(Pt 3), 889-905.
Wilson, G. F., and O'Donnell, R. D. (1986). “Steady state evoked responses:
correlations with human cognition.” Psychophysiology, 23(1), 57-61.
Wilson, G. F., and O'Donnell, R. D. (1988). “Measurement of operator workload with
the neuropsychological workload test battery.” Human mental workload, P. A.
Hancock and N. Meshkati, eds., Elsevier Science Publishers B. V., North
Holland, 63-100.
Yonelinas, A. P., and Jacoby, L. L. (1995). “The relation between remembering and
knowing as the basis for recognition: effects of size congruency.” Journal of
Experimental Memory and Language, 34, 622-643.
Young, M. P. (1993). “Visual cortex: modules for pattern recognition.” Current
Biology, 3, 44-46.
Young, M. P., and Rugg, M. D. (1992). “Word frequency and multiple repetition as
determinants of the modulation of event-related potentials in a semantic
classification task.” Psychophysiology, 29, 664-676.
Zola-Morgan, S., Squire, L. R., and Amaral, D. G. (1989). “Lesions of the amygdala
that spare adjacent cortical regions do not impair memory or exacerbate the
impairment following lesions of the hippocampal formation.” Journal of
207

Neuroscience, 9(6), 1922-1936.
Zola-Morgan, S., Squire, L. R., Rempel, N. L., Clower, R. P., and Amaral, D. G.
(1992). “Enduring memory impairment in monkeys after ischemic damage to the
hippocampus.” Journal of Neuroscience, 12(7), 2582-2596.
208

![United States Patent [19] 5,938,688 · Hinman et al., “Depth Evoked Potential and Single Unit Correlates of Vertex Midlatency Auditory Evoked Responses,” Brain Research, 264:57—67](https://static.fdocuments.net/doc/165x107/5f74697eb013c00c2b73061c/united-states-patent-19-5938688-hinman-et-al-aoedepth-evoked-potential-and.jpg)














![[DL輪読会]Neural Episodic Control/Model-Free Episodic Control](https://static.fdocuments.net/doc/165x107/5a64790d7f8b9a57568b463b/dlneural-episodic-controlmodel-free-episodic-control.jpg)

