
Self-maintained escort cells form a germline stem cell...
11
5087 DEVELOPMENT AND STEM CELLS RESEARCH ARTICLE INTRODUCTION Adult tissues are often maintained by a population of tissue-specific stem cells, which have the capacity to self-renew and generate differentiated cells throughout the organism’s lifetime. Many fast turnover tissues, such as blood, skin, intestine and testis, are known to be maintained by stem cells (Li and Xie, 2005; Morrison and Spradling, 2008). Some slow turnover tissues are either maintained by stem cells as in the brain or by self-duplication as for insulin- secreting cells in the pancreas (Dor et al., 2004; Zhao et al., 2008). These adult stem cells are maintained in the niche for long- term self-renewal (Li and Xie, 2005; Morrison and Spradling, 2008). However, it remains unclear whether stem cell lineage differentiation is also controlled by a niche mechanism. In the Drosophila ovary, two or three germline stem cells (GSCs) are anchored physically to their niche, which is composed of five to seven cap cells, through E-cadherin-mediated cell adhesion at the tip of the germarium (Song et al., 2002). Recently, a population of escort stem cells (ESCs), which directly contacts GSCs and cap cells, has been proposed to generate the differentiated escort cells (ECs) that accompany differentiated germ cells to the middle region of the germarium where ECs undergo apoptosis (Decotto and Spradling, 2005). The germ cells released from ECs are subsequently surrounded by follicle cells, which are produced by two follicular stem cells (FSCs), to form individual egg chambers (Margolis and Spradling, 1995; Song and Xie, 2002). Therefore, ESCs have been proposed to behave similarly to cyst progenitor cells in the Drosophila testis, which produce differentiated somatic cells that wrap around differentiated germ cells to support their differentiation (Gonczy et al., 1992; Kiger et al., 2000; Tran et al., 2000; Schultz et al., 2002; Decotto and Spradling, 2005). A previous study has shown that stem cell tumor (stet) encodes a germ cell-specific protein related to Rhomboid, which is required for the formation or maintenance of the long EC cellular processes and germ cell differentiation (Schultz et al., 2002). The Rhomboid family of transmembrane proteases is involved in processing EGF (epidermal growth factor) ligands (Urban et al., 2001). Indeed, another recent study showed that three EGF ligands function in germ cells to activate EGF receptor (EGFR) signaling in ECs, which in turn represses Dally expression (Liu et al., 2010). Dally is a proteoglycan protein that binds bone morphogenetic protein (BMP) and facilitates its signaling (Jackson et al., 1997; Belenkaya et al., 2004; Akiyama et al., 2008). In addition, Dally is also important for restricting BMPs inside the GSC niche in the Drosophila ovary (Guo and Wang, 2009; Hayashi et al., 2009). These findings suggest a model in which differentiated germ cells activate EGFR signaling in ECs, which prevents BMP diffusion from the GSC niche to ECs and thus promotes germ cell differentiation. However, it remains unclear how EC cellular process-mediated interactions between ECs and germ cells are regulated. In this study, we show that ECs are maintained by self-duplication instead of by ESCs. Furthermore, EC cellular processes are dependent on differentiated germ cells, and the physical interactions between ECs and germ cells are essential for germ cell differentiation. Therefore, we propose that self-maintained ECs form a niche that controls germ cell differentiation. Development 138, 5087-5097 (2011) doi:10.1242/dev.067850 © 2011. Published by The Company of Biologists Ltd Stowers Institute for Medical Research, 1000 East 50th Street, Kansas City, MO 64110, USA and Department of Cell Biology and Anatomy, University of Kansas Medical Center, 3901 Rainbow Boulevard, Kansas City, KS 66160, USA. *These authors contributed equally to this work ‡ Present address: Temasek Life Sciences Laboratory, 1 Research Link, National University of Singapore, Singapore 117604 § Author for correspondence ([email protected]) Accepted 30 September 2011 SUMMARY Stem cell self-renewal is controlled by concerted actions of niche signals and intrinsic factors in a variety of systems. In the Drosophila ovary, germline stem cells (GSCs) in the niche continuously self-renew and generate differentiated germ cells that interact physically with escort cells (ECs). It has been proposed that escort stem cells (ESCs), which directly contact GSCs, generate differentiated ECs to maintain the EC population. However, it remains unclear whether the differentiation status of germ cells affects EC behavior and how the interaction between ECs and germ cells is regulated. In this study, we have found that ECs can undergo slow cell turnover regardless of their positions, and the lost cells are replenished by their neighboring ECs via self- duplication rather than via stem cells. ECs extend elaborate cellular processes that exhibit extensive interactions with differentiated germ cells. Interestingly, long cellular processes of ECs are absent when GSC progeny fail to differentiate, suggesting that differentiated germ cells are required for the formation or maintenance of EC cellular processes. Disruption of Rho functions leads to the disruption of long EC cellular processes and the accumulation of ill-differentiated single germ cells by increasing BMP signaling activity outside the GSC niche, and also causes gradual EC loss. Therefore, our findings indicate that ECs interact extensively with differentiated germ cells through their elaborate cellular processes and control proper germ cell differentiation. Here, we propose that ECs form a niche that controls GSC lineage differentiation and is maintained by a non- stem cell mechanism. KEY WORDS: Drosophila, Germline stem cell, Niche, Differentiation, Escort cell Self-maintained escort cells form a germline stem cell differentiation niche Daniel Kirilly* ,‡ , Su Wang* and Ting Xie § DEVELOPMENT
Transcript of Self-maintained escort cells form a germline stem cell...

5087DEVELOPMENT AND STEM CELLS RESEARCH ARTICLE
INTRODUCTIONAdult tissues are often maintained by a population of tissue-specificstem cells, which have the capacity to self-renew and generatedifferentiated cells throughout the organism’s lifetime. Many fastturnover tissues, such as blood, skin, intestine and testis, are knownto be maintained by stem cells (Li and Xie, 2005; Morrison andSpradling, 2008). Some slow turnover tissues are either maintainedby stem cells as in the brain or by self-duplication as for insulin-secreting cells in the pancreas (Dor et al., 2004; Zhao et al.,2008). These adult stem cells are maintained in the niche for long-term self-renewal (Li and Xie, 2005; Morrison and Spradling,2008). However, it remains unclear whether stem cell lineagedifferentiation is also controlled by a niche mechanism.
In the Drosophila ovary, two or three germline stem cells(GSCs) are anchored physically to their niche, which is composedof five to seven cap cells, through E-cadherin-mediated celladhesion at the tip of the germarium (Song et al., 2002). Recently,a population of escort stem cells (ESCs), which directly contactsGSCs and cap cells, has been proposed to generate thedifferentiated escort cells (ECs) that accompany differentiated germcells to the middle region of the germarium where ECs undergoapoptosis (Decotto and Spradling, 2005). The germ cells releasedfrom ECs are subsequently surrounded by follicle cells, which are
produced by two follicular stem cells (FSCs), to form individualegg chambers (Margolis and Spradling, 1995; Song and Xie, 2002).Therefore, ESCs have been proposed to behave similarly to cystprogenitor cells in the Drosophila testis, which producedifferentiated somatic cells that wrap around differentiated germcells to support their differentiation (Gonczy et al., 1992; Kiger etal., 2000; Tran et al., 2000; Schultz et al., 2002; Decotto andSpradling, 2005). A previous study has shown that stem cell tumor(stet) encodes a germ cell-specific protein related to Rhomboid,which is required for the formation or maintenance of the long ECcellular processes and germ cell differentiation (Schultz et al.,2002). The Rhomboid family of transmembrane proteases isinvolved in processing EGF (epidermal growth factor) ligands(Urban et al., 2001). Indeed, another recent study showed that threeEGF ligands function in germ cells to activate EGF receptor(EGFR) signaling in ECs, which in turn represses Dally expression(Liu et al., 2010). Dally is a proteoglycan protein that binds bonemorphogenetic protein (BMP) and facilitates its signaling (Jacksonet al., 1997; Belenkaya et al., 2004; Akiyama et al., 2008). Inaddition, Dally is also important for restricting BMPs inside theGSC niche in the Drosophila ovary (Guo and Wang, 2009; Hayashiet al., 2009). These findings suggest a model in whichdifferentiated germ cells activate EGFR signaling in ECs, whichprevents BMP diffusion from the GSC niche to ECs and thuspromotes germ cell differentiation. However, it remains unclearhow EC cellular process-mediated interactions between ECs andgerm cells are regulated. In this study, we show that ECs aremaintained by self-duplication instead of by ESCs. Furthermore,EC cellular processes are dependent on differentiated germ cells,and the physical interactions between ECs and germ cells areessential for germ cell differentiation. Therefore, we propose thatself-maintained ECs form a niche that controls germ celldifferentiation.
Development 138, 5087-5097 (2011) doi:10.1242/dev.067850© 2011. Published by The Company of Biologists Ltd
Stowers Institute for Medical Research, 1000 East 50th Street, Kansas City, MO64110, USA and Department of Cell Biology and Anatomy, University of KansasMedical Center, 3901 Rainbow Boulevard, Kansas City, KS 66160, USA.
*These authors contributed equally to this work‡Present address: Temasek Life Sciences Laboratory, 1 Research Link, NationalUniversity of Singapore, Singapore 117604§Author for correspondence ([email protected])
Accepted 30 September 2011
SUMMARYStem cell self-renewal is controlled by concerted actions of niche signals and intrinsic factors in a variety of systems. In theDrosophila ovary, germline stem cells (GSCs) in the niche continuously self-renew and generate differentiated germ cells thatinteract physically with escort cells (ECs). It has been proposed that escort stem cells (ESCs), which directly contact GSCs, generatedifferentiated ECs to maintain the EC population. However, it remains unclear whether the differentiation status of germ cellsaffects EC behavior and how the interaction between ECs and germ cells is regulated. In this study, we have found that ECs canundergo slow cell turnover regardless of their positions, and the lost cells are replenished by their neighboring ECs via self-duplication rather than via stem cells. ECs extend elaborate cellular processes that exhibit extensive interactions withdifferentiated germ cells. Interestingly, long cellular processes of ECs are absent when GSC progeny fail to differentiate,suggesting that differentiated germ cells are required for the formation or maintenance of EC cellular processes. Disruption ofRho functions leads to the disruption of long EC cellular processes and the accumulation of ill-differentiated single germ cells byincreasing BMP signaling activity outside the GSC niche, and also causes gradual EC loss. Therefore, our findings indicate that ECsinteract extensively with differentiated germ cells through their elaborate cellular processes and control proper germ celldifferentiation. Here, we propose that ECs form a niche that controls GSC lineage differentiation and is maintained by a non-stem cell mechanism.
KEY WORDS: Drosophila, Germline stem cell, Niche, Differentiation, Escort cell
Self-maintained escort cells form a germline stem celldifferentiation nicheDaniel Kirilly*,‡, Su Wang* and Ting Xie§
DEVELO
PMENT

5088
MATERIALS AND METHODSDrosophila strains and cultureInformation about the Drosophila stocks used in this study is eitheravailable from Flybase (http://www.flybase.org) or specified here: X-15-29(Harrison and Perrimon, 1993), X-15-33 (Harrison and Perrimon, 1993),UAS-mEGFP (a membrane-tethered GFP generated by a fusion of the Srcmembrane signal peptide with GFP) (Kirilly et al., 2005), FRT52B (Kirillyet al., 2005), UAS-EGFP, FRT19A, tubulin-gal80 (Lee et al., 2000), tubulin-gal4 (Lee et al., 2000), c587 (Song et al., 2004), hsFLP, UAS-RhoDN (adominant-negative Rho) (Strutt et al., 1997), PZ1444 (Margolis andSpradling, 1995), UAS-dsRed, UAS-dcr2, UAS-dallyRNAi lines (onchromosome 2 and 3; kindly provided by Dr Xinhua Lin, CincinnatiChildren’s Hospital Medical Center, Cincinnati, USA), UAS-dppRNAi lines(TR00047A: 47A; TR00047R: 47R; kindly provided by Dr NorbertPerrimon, Harvard Medical School, Boston, USA), UAS-capuRNAi(VDRC#110404), dpphr4 and dpphr56. Flies were maintained and crossed atroom temperature on standard cornmeal/molasses/agar media unlessspecified. For maximizing the effect of RNAi-mediated knockdown orgene overexpression, newly eclosed flies were shifted to 29°C for a weekbefore the analysis of ovarian phenotypes.
Generating positively labeled EC clonesThree different positive marking systems were used for labeling ECs in theDrosophila ovary: the lacZ-based positive labeling system (Harrison andPerrimon, 1993), the UAS-GAL4-based positively marked mosaic lineage(PMML) system (Kirilly et al., 2005) and the GAL80-based mosaicanalysis with a repressible cell marker (MARCM) system (Lee et al.,2000). We generated mitotic clones according to our previously publishedprocedures for labeling the FSC lineage (Kirilly et al., 2005). One- to three-day-old females with genotypes hs-FLP; X-15-33/X-15-29 and hs-FLPUAS-mEGFP; FRT52B/FRT52B UAS-EGFP and hs-FLP FRT19A gal80/FRT19A; tub-gal4 UAS-GFP were subjected to incubation in a water bathat 37°C for 30 or 60 minutes to induce FLP expression and FRT-mediatedmitotic recombination. Flies were transferred daily to the fresh yeast-containing food, and the marked EC clones were detected one week, twoweeks and three weeks after the heatshock treatment.
BrdU labeling and retention assaysTwo different types of BrdU incorporation assays were utilized toinvestigate the proliferation patterns of ECs: two-hour BrdU labeling andBrdU retention. For the two-hour BrdU labeling, the ovaries wereincubated in Grace’s medium containing 75 mM BrdU for 2 hours at 25°C.For the BrdU retention assay, female flies were fed on food with yeastpaste containing BrdU (10 mg/ml) for three consecutive days, and then onfood with BrdU-free yeast paste for 14 days. The ovaries from these twotypes of assays were then fixed and processed for BrdU label detectionalong with other protein markers according to our previously publishedprocedures (Xie and Spradling, 1998).
ImmunohistochemistryAntibody staining was performed according to our previously publishedprocedures (Xie and Spradling, 1998; Song and Xie, 2002). The followingantibodies were used in this study: rabbit polyclonal anti--galactosidaseantibody (1:100, Cappel), rabbit polyclonal anti-GFP antibody (1:200,Molecular Probes), mouse monoclonal anti-Fasciclin3 antibody [1:3,Developmental Studies Hybridoma Bank (DSHB)], mouse monoclonalanti-Hts antibody (1:3, DSHB), rat monoclonal anti-Vasa antibody (1:3,DSHB) and monoclonal anti-BrdU antibody (1:20, Oncogene). All imageswere taken with either a Leica TCS SP2 or a Leica TCS SP5 confocalmicroscope.
RESULTSECs in different regions of the germariumundergo slow cell turnover and proliferationThe ESC model predicts that only ESCs at the tip of the germariumare mitotically active and that ECs undergo apoptosis at the 2a/2bjunction (Decotto and Spradling, 2005). To investigate whetherECs die only at the 2a/2b junction, we used Apoptag labeling to
detect apoptotic ECs. c587 is a GAL4 line that drives UAS-GFPexpression in ESCs, ECs and early follicle cell progenitors (Songet al., 2004) (Fig. 1A). PZ1444 is a lacZ enhancer trap line that isexpressed in cap cells and ECs (Margolis and Spradling, 1995; Xieand Spradling, 2000) (Fig. 1B). lacZ-positive ECs can be easilyidentified in the germarium in which germ cells are labeled withvasa-GFP (Sano et al., 2002) (Fig. 1B). Here, we used PZ1444,c587-driven UAS-dsRed and vasa-GFP to assist in theidentification of ECs. One or more dying EC is present in the 2a/2bjunction area of 14.7% of the 8-day-old germaria (n116), whereasonly 4.3% of those harbor one or more dying ECs in region 1 or 2a(Fig. 1C-F). These results indicate that ECs at different positionscan undergo slow turnover but with the highest turnover rate forthe most posterior ECs.
To investigate whether those lost ECs in regions 1 and 2a arerepopulated by their neighboring ECs, we performed two-hourBrdU labeling of PZ1444 ovaries in vitro to detect proliferativeECs. Following two hours of BrdU incorporation, 9.6% of thegermaria contain at least one BrdU-positive EC at the 2a/2bboundary, and only 0.8% of them contain a BrdU-positive EC in theregion 1 or 2a (n125) (Fig. 1G). This result indicates that ECs areslow cycling cells and that the ones at the 2a/2b boundary proliferatefaster than those in more anterior regions. To investigate further theslow cycling property, we also performed the BrdU retention assayby feeding PZ1444 or vasa-GFP flies for 3 days with BrdU-containing food and then 15 days with BrdU-free food. In the vasa-GFP germarium, ECs are reliably identified in the periphery of theanterior half of the germarium by absence of GFP expression,whereas ECs in the PZ1444 germarium are readily identified bylacZ-expression (Fig. 1B). After 3 days of BrdU feeding, 40% theGSCs in the PZ1444 germaria (n155 germaria) and 48.9% of theGSCs in the vasa-GFP germaria become BrdU-positive because oftheir fast proliferative nature (Fig. 1H,I). For ECs, only a smallnumber of them in either a PZ1444 germarium or a vasa-GFPgermarium are BrdU-positive, and most of those BrdU-positive ECsare localized in the 2a/2b junction (Fig. 1H,I). After 15 days ofchase, no GSCs in either PZ1444 (n212 germaria) or vasa-GFP(n154 germaria) germaria retain the BrdU label, indicating that fastcycling GSCs and FSCs do not retain BrdU label (Fig. 1J-K). Thisresult is consistent with the notion that not all adult stem cells retainBrdU label as is proposed for some adult stem cell types inmammalian systems (Morrison and Spradling, 2008). For thePZ1444 germaria, 34.3% of them harbor one or more BrdU-labeledECs in the 2a/2b junction area, and only 13.8% of them have one ormore BrdU-positive ECs in region 1 or 2a (n137; Fig. 1J). Thevasa-GFP germaria exhibit a similar BrdU labeling pattern amongthe ECs (Fig. 1K,L). The previously defined ESCs are rarely BrdU-positive (Fig. 1L), suggesting that ESCs are unlikely to beresponsible for generating ECs. These results show that ECsproliferate infrequently owing to slow cell turnover and most of theproliferative cells are restricted to the 2a/2b junction area, which isconsistent with the EC turnover pattern.
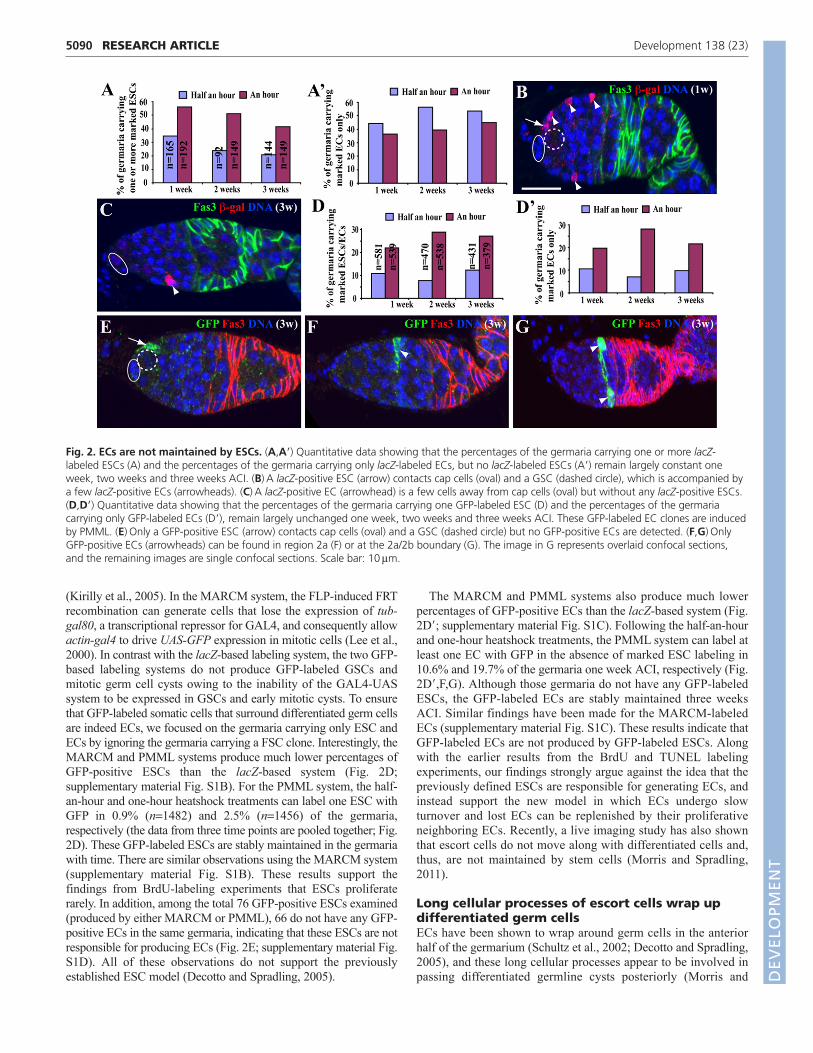
ECs are not maintained by ESCsIn the previous study, the lacZ-based positive labeling system wasused to show that the most anterior ECs, which directly contact capcells and GSCs, were proposed to function as ESCs to generatedifferentiated ECs (Decotto and Spradling, 2005). In this system,the FLP-induced FRT mitotic recombination leads to the fusionbetween the -galactosidase (lacZ) gene and the tubulin (tub)promoter and, thereby, the reconstitution of tub-lacZ in mitoticcells, and tub-lacZ expression further positively and permanently
RESEARCH ARTICLE Development 138 (23)
DEVELO
PMENT

marks mitotic cells and their progeny (Harrison and Perrimon,1993). This system has been used to label GSCs, FSCs, ESCs andtheir progeny (Margolis and Spradling, 1995; Decotto andSpradling, 2005). In the current study, to ensure that lacZ-labeledsomatic cells that surround differentiated germ cells are indeedECs, we focused on the germaria only carrying lacZ-labeled ESCsor ECs and disregarded other germaria carrying lacZ-positive FSCor GSC clones. To verify whether ESCs are indeed responsible forproducing ECs, we used the same system to repeat the lacZ-basedEC lineage tracing experiments using half-an-hour and one-hourheatshock treatments. These germaria are also labeled for Fasciclin3 (Fas3), which is expressed in follicle cell progenitors but not inECs (Zhang and Kalderon, 2001). As the previous study reported(Decotto and Spradling, 2005), both heatshock regimes canefficiently induce lacZ-labeled ESCs and ECs although the one-hour treatment generates higher lacZ-labeling efficiencies than thehalf-an-hour treatment (Fig. 2A,A�). For example, one week afterclone induction (ACI), 34.5% and 56.2% of the germaria carry atleast one lacZ-marked ESC after the half-an-hour and one-hourheatshock treatments, respectively (Fig. 2A). These lacZ-labeledESCs are relatively stable three weeks ACI, which is consistentwith the prediction of the ESC model. In addition, 82.4% of thegermaria carrying one or more lacZ-marked ESCs also have one ormore lacZ-positive ECs for the half-an-hour heatshock treatment,and 99.1% of the germaria carrying one or more lacZ-markedESCs have one or more lacZ-positive ECs for the one-hourheatshock treatment (Fig. 2B). These observations could explain
why the previous study concludes that ECs are produced by ESCs(Decotto and Spradling, 2005) (Fig. 2A,B). However, carefulanalyses of the lacZ-marked ECs have revealed the serious inherentproblem of the lacZ labeling system. First, the percentages ofgermaria carrying only marked ECs but not ESCs, which areinduced by either of the heatshock treatments, remain constant oneweek, two weeks and three weeks ACI (Fig. 2A�,C). This resultindicates that lacZ-labeled ECs can be stably maintained in theabsence of lacZ-labeled ESCs. Second, some germaria carrying onelacZ-labeled ESC do not contain any lacZ-positive ECs(supplementary material Fig. S1A), suggesting that they might notbe responsible for EC production. Third, high lacZ labeling ratesfor ESCs and ECs are not supported by our BrdU labeling results.This system has recently been shown to generate lacZ-positive cellseven in mitotically inactive Drosophila polyploid intestinal cells(Fox and Spradling, 2009). Our results also suggest that this systemcan reconstitute the tub-lacZ gene in an FLP-dependent butdivision-independent manner in ECs, and, therefore, is not asuitable system for determining the presence of ESCs.
To investigate further whether ESCs are responsible for producingECs, we used two other FLP-FRT-mediated positive lineage labelingsystems, positively marked mosaic lineage (PMML) and mosaicanalysis with a repressible cell marker (MARCM), to generatepositive GFP-marked EC clones and study their behavior. In thePMML system, FLP-induced FRT recombination can randomlygenerate the fusion gene between the tubulin promoter and the yeastgal4 gene, which can drive UAS-GFP expression in mitotic cells
5089RESEARCH ARTICLERoles of escort cells
Fig. 1. ECs at different positions can undergo apoptosis and cell proliferation. (A)c587 drives UAS-GFP expression in ESCs and ECs inregions 1 and 2a, and also in early follicle cells (FCs) in region 2b. GSCs (dashed lines) and differentiated germ cells, including 16-cell cysts, can beidentified by presence of a spectrosome (SS) and branched fusome (FS), respectively. (B)PZ1444 can also label ESCs (arrow), ECs (arrowheads) andcap cells (oval) in the germarium in which germ cells are labeled by Vasa-GFP. (C-F)ApopTag-positive ECs are found in region 2a (C,D, arrows) andthe 2a/2b boundary (E,F, arrows). (G)An EC at the 2a/2b boundary is positive for BrdU after two hours of BrdU incorporation. (H,I)GSCs (whitearrowheads), mitotic cysts and follicle cells in the germarium are positive for BrdU (green) one day after three days of BrdU feeding. Most of the ECs(black arrowheads) are negative for BrdU, but some ECs (arrows) are BrdU-positive. (J-L)After two weeks of chase following three days of BrdUfeeding, all the GSCs and mitotic cysts are BrdU-negative, but some ECs (arrows) remain positive for BrdU. ESCs are normally negative for BrdU(black arrowhead, K), but are rarely found to be positive (black arrowhead, L). Scale bars: 10mm in B-L.
DEVELO
PMENT
5090
(Kirilly et al., 2005). In the MARCM system, the FLP-induced FRTrecombination can generate cells that lose the expression of tub-gal80, a transcriptional repressor for GAL4, and consequently allowactin-gal4 to drive UAS-GFP expression in mitotic cells (Lee et al.,2000). In contrast with the lacZ-based labeling system, the two GFP-based labeling systems do not produce GFP-labeled GSCs andmitotic germ cell cysts owing to the inability of the GAL4-UASsystem to be expressed in GSCs and early mitotic cysts. To ensurethat GFP-labeled somatic cells that surround differentiated germ cellsare indeed ECs, we focused on the germaria carrying only ESC andECs by ignoring the germaria carrying a FSC clone. Interestingly, theMARCM and PMML systems produce much lower percentages ofGFP-positive ESCs than the lacZ-based system (Fig. 2D;supplementary material Fig. S1B). For the PMML system, the half-an-hour and one-hour heatshock treatments can label one ESC withGFP in 0.9% (n1482) and 2.5% (n1456) of the germaria,respectively (the data from three time points are pooled together; Fig.2D). These GFP-labeled ESCs are stably maintained in the germariawith time. There are similar observations using the MARCM system(supplementary material Fig. S1B). These results support thefindings from BrdU-labeling experiments that ESCs proliferaterarely. In addition, among the total 76 GFP-positive ESCs examined(produced by either MARCM or PMML), 66 do not have any GFP-positive ECs in the same germaria, indicating that these ESCs are notresponsible for producing ECs (Fig. 2E; supplementary material Fig.S1D). All of these observations do not support the previouslyestablished ESC model (Decotto and Spradling, 2005).
The MARCM and PMML systems also produce much lowerpercentages of GFP-positive ECs than the lacZ-based system (Fig.2D�; supplementary material Fig. S1C). Following the half-an-hourand one-hour heatshock treatments, the PMML system can label atleast one EC with GFP in the absence of marked ESC labeling in10.6% and 19.7% of the germaria one week ACI, respectively (Fig.2D�,F,G). Although those germaria do not have any GFP-labeledESCs, the GFP-labeled ECs are stably maintained three weeksACI. Similar findings have been made for the MARCM-labeledECs (supplementary material Fig. S1C). These results indicate thatGFP-labeled ECs are not produced by GFP-labeled ESCs. Alongwith the earlier results from the BrdU and TUNEL labelingexperiments, our findings strongly argue against the idea that thepreviously defined ESCs are responsible for generating ECs, andinstead support the new model in which ECs undergo slowturnover and lost ECs can be replenished by their proliferativeneighboring ECs. Recently, a live imaging study has also shownthat escort cells do not move along with differentiated cells and,thus, are not maintained by stem cells (Morris and Spradling,2011).
Long cellular processes of escort cells wrap updifferentiated germ cellsECs have been shown to wrap around germ cells in the anteriorhalf of the germarium (Schultz et al., 2002; Decotto and Spradling,2005), and these long cellular processes appear to be involved inpassing differentiated germline cysts posteriorly (Morris and
RESEARCH ARTICLE Development 138 (23)
Fig. 2. ECs are not maintained by ESCs. (A,A�) Quantitative data showing that the percentages of the germaria carrying one or more lacZ-labeled ESCs (A) and the percentages of the germaria carrying only lacZ-labeled ECs, but no lacZ-labeled ESCs (A�) remain largely constant oneweek, two weeks and three weeks ACI. (B)A lacZ-positive ESC (arrow) contacts cap cells (oval) and a GSC (dashed circle), which is accompanied bya few lacZ-positive ECs (arrowheads). (C)A lacZ-positive EC (arrowhead) is a few cells away from cap cells (oval) but without any lacZ-positive ESCs.(D,D�) Quantitative data showing that the percentages of the germaria carrying one GFP-labeled ESC (D) and the percentages of the germariacarrying only GFP-labeled ECs (D�), remain largely unchanged one week, two weeks and three weeks ACI. These GFP-labeled EC clones are inducedby PMML. (E)Only a GFP-positive ESC (arrow) contacts cap cells (oval) and a GSC (dashed circle) but no GFP-positive ECs are detected. (F,G)OnlyGFP-positive ECs (arrowheads) can be found in region 2a (F) or at the 2a/2b boundary (G). The image in G represents overlaid confocal sections,and the remaining images are single confocal sections. Scale bar: 10mm.
DEVELO
PMENT

Spradling, 2011). To investigate further to what extent germ cellsinteract with ECs, we used c587-driven UAS-GFP expression tolabel ECs with GFP, which allows the visualization of EC cellularprocesses. In the whole-mount germaria immunostained for GFPand Hts, the long cellular processes of ECs wrap arounddifferentiated germ cells, which are identified by the presence ofbranched fusomes labeled by Hts protein (Fig. 3A). Hts protein isrich in spherical spectrosomes in GSCs and cystoblasts (CBs) andin branched fusomes in differentiated germ cells (Lin et al., 1994).In cryosections of c587-gal4;UAS-GFP germaria labeled for GFPand Vasa, EC long cellular processes are shown to separateindividual germ cells from one another (Fig. 3B). Vasa is acommonly used germ cell-specific marker (Hay et al., 1988; Laskoand Ashburner, 1988). Our results further support the previousfinding that individual early differentiated germline cysts interactextensively with long EC cellular processes.
Previous studies were unable to determine the morphology ofindividual ECs using nuclear lacZ-labeled ECs and c587-driven orpatch-gal4-driven UAS-GFP expression (Margolis and Spradling,1995; Schultz et al., 2002; Song et al., 2004; Decotto andSpradling, 2005). To investigate the EC morphology of individualECs, we used the PMML system described above to label one or afew ECs with tub-gal4-driven expression of UAS-mGFP (amembrane-targeted GFP; the fusion between the Src membranelocalization signal and GFP). In the ovariole that lacks a GFP-labeled FSC clone, the GFP-positive ECs in regions 1, 2a and
2a/2b can be readily identified (Fig. 3C-I). The most anterior GFP-labeled ECs, which correspond to the previously identified ESCs,directly contact GSCs and wrap around GSCs with their cellularprocesses as previously described (Decotto and Spradling, 2005)(Fig. 3C). The ECs in region 1 that interact with CBs and mitoticgerm cysts have short cellular processes (Fig. 3D,E; supplementarymaterial Movie 1). In region 2a, where 16-cell cysts occupy halfthe diameter of the germaria, individual ECs have slightly longercellular processes to encase individual 16-cell cysts (Fig. 3F,G;supplementary material Movie 2). In the 2a/2b junction areas,where germline cysts start to adopt a lens shape crossing thegermarium, the ECs extend their cellular processes across thewhole width of the germarium to wrap around individual 16-cellcysts (Fig. 3H,I; supplementary material Movie 3). These resultsshow that ECs at different positions adopt distinct morphologiesbased on the size and morphology of underlying differentiatedgerm cell cysts, further suggesting that EC cellular processes arelikely to be regulated by the underlying differentiated germ cellcysts.
EC cellular processes might be regulated byunderlying differentiated germ cellsAlthough EC long cellular processes are only associated withdifferentiated germ cells, it remains unclear whether differentiatedgerm cells are required for the formation of long cellular processesin ECs. bag of marbles (bam) is required for germ cell
5091RESEARCH ARTICLERoles of escort cells
Fig. 3. Differentiated germ cells are wrapped by EC cellular processes. (A)Differentiated germ cell cysts (dashed lines) are encased by GFP-positive EC cellular processes. (B)A cryosectioned germarium showing that germ cell cysts (dashed lines) are separated from one another by GFP-positive EC cellular processes. (C-I)Germaria labeled for GFP (green) and -galactosidase (PZ1444, red, C), Hts (red, D) or Fas3 (red, differentiatedfollicle cells, E-I) containing a GFP-positive EC (arrow) at the ESC position (C), region 1 (D,E), region 2a (F,G) or the 2a/2b boundary (H,I). The ECs inD and E have short cellular processes encasing the cystoblast (CB) and mitotic cyst, respectively, whereas those in F-I have longer cellular processeswrapping around 16-cell cysts. The images in C, E, F and H are one confocal section, whereas images in D, G and I are overlaid confocal sections.Scale bar: 10mm.
DEVELO
PMENT

5092
differentiation, and its mutant ovaries only contain undifferentiatedcystoblast-like single germ cells (McKearin and Spradling, 1990;McKearin and Ohlstein, 1995). bamD86 (a bam deletion allele)mutant germaria are filled with undifferentiated germ cells, whichare identified by spherical spectrosomes (Fig. 4A). Interestingly, inbam mutant germaria in which ECs are labeled by c587-drivenUAS-GFP expression, the long cellular processes of most ECs failto form or to penetrate inside the germ cells (Fig. 4A). To rule outthe possibility of poor antibody penetration into tightly packedgerm cells, the c857; UAS-GFP; bamD86/ bamD86 germarialcryosections are labeled for GFP and Vasa to detect EC cellularprocesses and germ cells, respectively. Consistently, long ECcellular processes are not observed, although some ECs have smallEC protrusions (Fig. 4B, arrows). These results suggest thatdifferentiated germ cells might be required for the formation ormaintenance of long EC cellular processes.
To determine further whether the formation of long EC cellularprocesses is dependent on differentiated germ cells or Bamfunction in germ cells, we examined the formation of long ECcellular processes in germaria carrying Dpp overexpression-induced GSC-like cells. c587-driven UAS-dpp results in theformation of ovaries filled with GSC-like cells (Fig. 4C), aspreviously reported (Song et al., 2004). In the dpp-inducedtumorous germaria, EC cellular processes, which are visualized byc587-driven UAS-GFP expression, fail to penetrate into individualGSC-like aggregates and to wrap around individual single germcells (Fig. 4C). As c587-driven dpp overexpression cannotcompletely inhibit GSC differentiation owing to mosaic expression,some differentiated cysts form but some GFP-positive cellularprocesses can still be detected to wrap up the differentiated germcells (Fig. 4D). Along with the bam mutant results, these resultsfurther support the idea that differentiated germ cells might supportthe formation of EC cellular processes.
Rho functions in ECs to maintain cellularprocesses and promote germ cell differentiationThe small GTPase Rho has been extensively studied in differentorganisms for its role in the regulation of cell protrusion formationand cell-cell adhesion (Jaffe and Hall, 2005). To investigate furtherwhether EC cellular process-mediated interactions with germ cellsare important for germ cell differentiation, we overexpressed adominant negative form of Rho (RhoDN) in ECs using c587-drivenUAS-RhoDN expression. UAS-RhoDN has previously been used todisrupt specifically Rho function in different Drosophila cell types
(Strutt et al., 1997; Hacker and Perrimon, 1998; Prokopenko et al.,1999; Billuart et al., 2001; Bloor and Kiehart, 2002; Magie et al.,2002). Interestingly, in the germaria in which RhoDN is expressedin ECs, differentiated germ cells are not wrapped up by EC cellularprocesses, suggesting that Rho might regulate EC cellular processformation or maintenance (Fig. 5A-C). In those germaria, sphericalspectrosome-containing single germ cells often accumulate (Fig.5A,C). In contrast with the wild-type germaria, which contain ~5.5spectrosome-containing single germ cells including both GSCs andcystoblasts (n91), the RhoDN-expressing germaria contain anaverage of 8.9 spectrosome-containing germ cells (n69). Also, thedisruption severity of EC cellular processes is related to theincrease in the number of spectrosome-containing single germcells, suggesting that long EC cellular processes might regulategerm cell differentiation.
One of the potential mechanisms for regulation by Rho of ECcellular processes is by modulation of physical interactionsbetween ECs and germ cells through regulating actin cytoskeletonbecause Rho is known to regulate actin dynamics and cell-cellinteraction in different Drosophila tissues (Hacker and Perrimon,1998; Prokopenko et al., 1999; Bloor and Kiehart, 2002; Magie etal., 2002). To determine whether actin regulation is important forEC function and germ cell differentiation, we sought to inactivatethe function of the Formin-like actin regulator cappuccino (capu)in ECs using c587-driven UAS-capuRNAi expression. Capu isknown to regulate actin nucleation and the formation of actinmeshwork (Dahlgaard et al., 2007; Quinlan et al., 2007). Followingcapu knockdown in ECs, the ECs located in the anterior half of thegermaria generally lack cellular processes, and the anterior part ofthe germaria accumulate more spectrosome-containing single germcells, suggesting that actin regulation is crucial for maintaining ECcellular process and, thus, CB differentiation (supplementarymaterial Fig. S2A,B). However, all of the germaria appear to benormal in size, indicating that disruption of actin dynamics doesnot affect EC survival. These results further support the idea thatEC cellular processes regulated by actin dynamics are importantfor germ cell differentiation.
Because some germaria expressing RhoDN in ECs are small incomparison with wild-type germaria (Fig. 5A,B), we used thePZ1444 EC marker to examine whether the number of ECsfollowing RhoDN expression is indeed reduced. The control week-old PZ1444 germaria contain an average of 42 ECs (n31). Onlyvery few of the week-old RhoDN-expressing germaria have thenormal or close to normal number of ECs (Fig. 5D); most of them
RESEARCH ARTICLE Development 138 (23)
Fig. 4. Differentiated germ cells might maintain ECcellular processes. (A)c587;UAS-GFP; bamD86/bamD86
germarium showing that all the ECs (one by arrowhead) failto form cellular processes penetrating inside tumorous germcells. (B)c587;UAS-GFP; bamD86/bamD86 cryosectionshowing that ECs can form some protrusions (arrowhead)that fail to encase germ cells. (C)c587;UAS-dpp/UAS-GFPgermarium showing that ECs fail to form long cellularprocesses wrapping single germ cells (dashed lines).(D)c587;UAS-dpp/UAS-GFP germarium showing that ECcellular processes wrap round a differentiated cyst (arrow)but fail to penetrate inside the clusters (dashed lines)containing single germ cells. Scale bar: 10mm.
DEVELO
PMENT

have fewer ECs (Fig. 5E,F). The week-old RhoDN-expressinggermaria have an average of 21 ECs (n50), significantly fewerthan those in control germaria (Fig. 5G). Those germaria withfewer ECs tend to have more spectrosome-containing single germcells than the germaria with more ECs, suggesting that ECs areimportant for germ cell differentiation. The RhoDN-expressinggermarium in Fig. 5D has a close to normal number of ECs, butcontains excess spectrosome-containing single germ cells lyingmore posteriorly to GSCs, suggesting that disruption of EC cellularprocesses could also contribute to the delay in germ celldifferentiation. Taken together, these results suggest that Rho isrequired for EC-germ cell interactions and EC maintenance, andthat ECs are important for normal germ cell differentiation. Theexact role of long EC cellular processes in the regulation of germcell differentiation needs to be investigated further by identifyingmolecule(s) involved in the EC-cyst interaction.
The reduction of EC number in the RhoDN-expressinggermarium could be due to EC proliferation or survival. Todetermine whether Rho is required for EC proliferation or survival,we used BrdU incorporation and ApopTag-labeling to quantifyproliferating and dying ECs in control and RhoDN-expressingPZ1444 germaria, respectively. Following two hours of BrdUincorporation, 0.97% and 1.0% of control (n5145) and RhoDN-
expressing (n2213) ECs are positive for BrdU, respectively,indicating that disruption of Rho function does not change ECproliferation. By contrast, 1.8% and 4.2% of control (n4000) andRhoDN-expressing (n2795) ECs are positive for ApopTag labeling,respectively, indicating that disruption of Rho function increasesEC apoptosis (Fig. 5H,I). These results indicate that Rho isrequired for EC survival but not for EC proliferation.
Defective Rho in ECs causes the germ celldifferentiation defect by upregulating BMPsignaling activityTo characterize further the identity of accumulated spectrosome-containing single germ cells in the RhoDN-overexpressing germaria,we examined expression of bam-GFP and Dad-lacZ in these extrasingle germ cells. In the wild-type germarium, bam-GFP is areporter construct for monitoring bam transcription and is mainlyexpressed in differentiated germ cells but is repressed in GSCs byBMP signaling (Chen and McKearin, 2003), whereas Dad-lacZ isa BMP signaling activity reporter and is activated in GSCs andrepressed in differentiated germ cells (Kai and Spradling, 2003;Casanueva and Ferguson, 2004; Song et al., 2004). As expected,GSCs close to cap cells have low bam-GFP and high Dad-lacZexpression in control germaria (Fig. 6A,D). Most of the single
5093RESEARCH ARTICLERoles of escort cells
Fig. 5. Defective Rho signaling in ECs disrupts EC-germ cell interactions and germ cell differentiation. The c587;UAS-RhoDN/UAS-GFPgermaria (single confocal section) in A-C are labeled for GFP, Hts and DNA, whereas c587;UAS-RhoDN/PZ1444 germaria (overlaid confocal sections)in D-F and H-I are labeled for -galactosidase (PZ1444), Hts and DNA, and ApopTag, -galactosidase (PZ1444) and DNA, respectively. (A)The GFP-positive cellular processes of RhoDN-expressing ECs fail to wrap around a differentiated cyst (dashed lines). (B)RhoDN expression in ECs leads to areduction of regions 1 and 2a of the germarium and a defect in wrapping a differentiated cyst (dashed lines) by EC cellular processes.(C)Spectrosome-containing single germ cells and a 16-cell cyst (dashed lines) are clustered together and are not individually wrapped by RhoDN-expressing EC cellular processes. Arrowheads indicate spectrosomes. (D)Germarium contains a normal number of ECs but excess single germ cells(arrowheads) located anteriorly to differentiated cysts (arrows). (E,F)Germaria show a moderate (E) or severe (F) reduction of ECs and excessspectrosome-containing single germ cells (arrowheads). (G)RhoDN-expressing germaria have significantly fewer ECs than do control germaria. P-value is indicated. (H)A control germarium contains no apoptotic ECs. (I)A RhoDN-expressing germarium contains an apoptotic EC (arrowhead).Scale bar: 10mm.
DEVELO
PMENT

5094
germ cells express high levels of bam-GFP and low levels Dad-lacZ, suggesting that those single germ cells resemble CBs insteadof GSCs (Fig. 6B,C,E,F). However, some spectrosome-containingsingle germ cells that are located several cells away from cap cellsexpress Dad-lacZ and repress bam-GFP expression (Fig. 6C,F).This observation indicates that BMP signaling activity hasexpanded outside the GSC niche and thereby interferes with germcell differentiation. Therefore, these results demonstrate that Rhofunction is required in ECs to control CB differentiation possiblyby restricting BMP signaling to the GSC niche.
Rho in ECs helps restrict BMP signaling activity tothe GSC nicheActivation of EGFR signaling leads to phosphorylation of ERK(pERK) in ECs, and defective EGFR signaling leads to the absenceof long EC cellular processes and defective germ cell differentiation(Schultz et al., 2002). EGFR signaling in ECs promotes germ celldifferentiation by repressing the expression of dally, encoding aprotein important for Dpp diffusion and function, in ECs (Liu et al.,2010). To investigate whether the disruption of Rho function affectsEGFR signaling in ECs, we examined the expression of pERK in theECs of RhoDN-expressing germaria. Our quantitative result indicatesthat RhoDN-expressing ECs (n160) have significantly, though notdramatically, less pERK expression than control ECs (n160) (Fig.
6G-I). This finding demonstrates that defective Rho functionmoderately decreases EGFR signaling in ECs.
To determine whether the germ cell differentiation defect inRhoDN-expressing germaria is caused by upregulation of BMPsignaling outside the GSC niche, we tested whether the removal ofa copy of the dpp gene could suppress the differentiation defect. Incontrast with the results that 65% of the RhoDN-expressing germariaaccumulate excessive spectrosome-containing single germ cellsoutside the niche (Fig. 7A,B), only 15% and 20% of the RhoDN-expressing dpphr56 and dpphr4 heterozygous germaria, respectively,have extra single germ cells (Fig. 7C,D). These results indicate thatupregulation of Dpp/BMP signaling in the RhoDN-expressinggermarium contributes to the germ cell differentiation defect. Toknock down specifically dally function in ECs, two independentdally RNAi lines were used to be expressed in ECs using c587.Interestingly, EC-specific knockdown of dally in the RhoDN-expressing germaria can also slightly suppress the differentiationdefect based on the accumulation of single germ cells, indicating thatdownregulation of EGFR signaling makes a slight contribution to thegerm cell differentiation defect (Fig. 7C,E). Although EC-specificknockdown of dpp mRNAs in ECs using two independent RNAilines produces no discernible phenotype in wild-type germaria, itleads to strong suppression of the germ cell differentiation defect inthe RhoDN-expressing germaria, suggesting that RhoDN-expressing
RESEARCH ARTICLE Development 138 (23)
Fig. 6. Defective Rho signaling in ECs delays cystoblast differentiation. The germaria in A-C are labeled for GFP, Hts and DNA, those in D-Fare labeled for -galactosidase (Dad), Hts and DNA, and those in G and H are labeled for pERK, Hts and DNA. (A)Two control GSCs (solid circles)expressing low bam-GFP and differentiated cysts expressing high bam-GFP. (B,C)c587;UAS-RhoDN/bam-GFP germarium carrying many ofspectrosome-containing single germ cells, most of which express bam-GFP (cystoblast-like; dashed circles) and the remaining ones show low bam-GFP expression (GSC-like; solid circle). (D)Two control GSCs (solid circles) expressing high levels of Dad-lacZ. (E,F)c587; UAS-RhoDN/Dad-lacZgermaria carrying spectrosome-containing single germ cells, of which those further away from the niche (dashed circle) express lower Dad-lacZ thando endogenous GSCs (solid circles) contacting the niche. Some single cells away from the niche (solid circle, F) express high levels of Dad-lacZ.(G)Wild-type ECs (two indicated by arrowheads) are positive for pERK staining. (H)RhoDN-expressing ECs away from (some indicated byarrowheads) or close to (dashed circle) single germ cells remain positive for pERK staining. (I)RhoDN-expressing ECs have significantly less pERK thancontrol ECs. Error bars represent s.d. Number above horizontal line represents P-value. Scale bar: 10mm.
DEVELO
PMENT

ECs might also upregulate dpp mRNAs via transcription orstabilization (Fig. 7C,F). Taken together, Rho is required in ECs tocontrol CB differentiation by restricting BMP signaling activity tothe GSC niche.
DISCUSSIONIn the Drosophila ovary, ECs are thought to be produced by apopulation of ESCs and to move along with differentiated germcells and commit apoptosis after differentiated germ cell cysts aresurrounded by follicle cells (Decotto and Spradling, 2005). In thisstudy, we have used three positive labeling systems and BrdUlabeling experiments to demonstrate that ECs are maintained byself-duplication but not by the previously identified ESCs (Fig.7G). Furthermore, ECs exhibit extensive interactions withdifferentiated germ cells through their long cellular processes, anddifferentiated germ cells are likely to be responsible formaintaining long cellular processes. Interestingly, EC cellularprocess-mediated interactions are required for germ celldifferentiation by restricting BMP signaling to the GSC niche.Therefore, we propose that ECs form a germ cell differentiationniche by preventing BMP signaling from interfering with CBdifferentiation (Fig. 7G).
In this study, we have provided three pieces of experimentalevidence demonstrating that ECs are maintained by self-duplicationbut not by stem cells. First, contrary to one of the predictions bythe ESC model that ECs can only undergo apoptosis at the junctionbetween ECs and follicle cell progenitors, we show that ECs in anyposition can undergo apoptosis although those at the junction dierelatively more frequently. Second, contrary to another predictionof the ESC model that only ESCs can proliferate, we show thatECs in any position can proliferate slowly although those at theposterior divide relatively more frequently. Third, we have usedthree independent lineage-labeling methods to demonstrate that thepreviously defined ESCs do not produce new ECs (Fig. 7G).Therefore, the ECs in the Drosophila ovary behave similarly to thedifferentiated cells in the pancreas that are maintained by self-duplication (Dor et al., 2004). Possibly, in other slow turnovermammalian tissues, stem cells are not required for maintainingtissue homeostasis.
Long EC cellular processes might help pass on a differentiatedgerm cell cluster from one EC to other (Morris and Spradling,2011). In this study, we propose that long cellular process-mediatedintimate interactions between ECs and differentiation are requiredfor proper germ cell differentiation probably by restricting BMP
5095RESEARCH ARTICLERoles of escort cells
Fig. 7. Rho contributes to the restriction ofBMP signaling to the GSC niche. The germariain A-F are labeled for Hts and DNA. (A,B)RhoDN
expression in ECs leads to the accumulation ofexcessive spectrosome-containing single germcells. (C)The percentages of excessive single germcell-containing germaria in ovaries in which RhoDN
is specifically expressed in the ECs and which arewild type, heterozygous for dpp (dpphr4 anddpphr56), or co-expressed with RNAi constructs fordally (one on chromosome 2 and one onchromosome 3) and dpp (47A and 47R). Inaddition to endogenous GSCs, three moreadditional single cells outside the GSC niche areconsidered to be excessive single germ cellsbecause a wild-type germarium contains one totwo CB-like single germ cells. (D-F)The germ celldifferentiation defect induced by RhoDN expressioncan be dramatically suppressed by a heterozygousdpphr4 mutation (D), by co-expression of dallyRNAi (E) or by co-expression of dpp RNAi (F).(G)Working model for EC maintenance and Rhofunction in ECs. When an EC dies, its neighbor ECdivides to generate a new EC to replace the lostone. Rho works in ECs to restrict BMP signalingwithin the GSC niche by preventing Dpp diffusionvia regulation of EGFR-Dally signaling andrepressing dpp mRNA accumulation. In addition,Rho signaling is important for EC survival.
DEVELO
PMENT

5096
signaling activity to the GSC niche (Fig. 7G). EC cellular processesextensively wrap around differentiated germ cells, and are modeledaccording to the size, shape and differentiated status of theunderlying differentiated germ cells. Forced expression of adominant-negative Rho or knocking down expression of the actinregulator capu causes defects in EC cellular extension and germcell differentiation. This is consistent with the finding in theprevious study that defective EGFR signaling causes loss of ECcellular processes and germ cell differentiation defects (Schultz etal., 2002). Our genetic results have revealed two possible ways forRho in ECs to control germ cell differentiation by restricting BMPsignaling within the GSC niche. One of them is to repress dallyexpression in ECs via EGFR signaling and thereby prevent BMPdiffusion to outside the GSC niche because EGFR signaling-mediated repression of dally expression has been shown to beessential for preventing BMP diffusion and germ celldifferentiation (Guo and Wang, 2009; Hayashi et al., 2009; Liu etal., 2010). The other is to prevent BMP expression in ECs byrepressing dpp transcription or degrading dpp mRNAs throughunknown mechanisms. In addition to the regulation of EC-germcell interaction (Ridley, 2006), this study also shows that Rho isalso required for promoting EC survival. Therefore, we proposethat ECs form a niche for promoting germ cell differentiation, andthat Rho functions in ECs to promote germ cell differentiation atleast partly by preventing BMP signaling in the differentiationniche (Fig. 7G).
The differentiation niche is likely to be a conserved feature fordifferent adult stem cell systems. In the Drosophila testis, somaticcyst cells, which are produced by cyst stem cells, encasedifferentiated germ cells and move together to accompany thedifferentiated germ cells (Gonczy and DiNardo, 1996). EGFRsignaling is required in somatic cyst cells, as it is in ECs, to controlgerm cell differentiation because defective EGFR signaling in theDrosophila testis causes germ cell differentiation defects (Kiger etal., 2000; Tran et al., 2000). Although, unlike ECs, somatic cystcells are not stationary, they function like ECs to control germ celldifferentiation. Although equivalent cells for ECs in mammalianadult stem cell systems have not been defined, they are likely toexist and to play important roles in the regulation of lineage-specific differentiation because cultured adult stem cells oftenrequire a cocktail of growth factors for their proper differentiation.As easily defined stem cell niches in Drosophila contribute to abetter understanding of mammalian stem cell regulation (Li andXie, 2005; Morrison and Spradling, 2008), we anticipate that whatwe will learn from studies on the Drosophila GSC differentiationniche will be equally important for defining differentiation nichesand studying their functions in lineage differentiation inmammalian systems.
AcknowledgementsWe would like to thank L. Cai, X. Lin, N. Perrimon, D. McKearin,Developmental Studies Hybridoma Bank and Bloomington Drosophila StockCenter for reagents; the Xie laboratory members for stimulating discussions;and S. Chen, M. Lewallen and C. Flournoy for critical comments.
FundingThis work is supported by the National Institutes of Health [R01GM64428 toT.X.]; and the Stowers Institute for Medical Research [T.X.]. Deposited in PMCfor release after 12 months.
Competing interests statementThe authors declare no competing financial interests.
Supplementary materialSupplementary material available online athttp://dev.biologists.org/lookup/suppl/doi:10.1242/dev.067850/-/DC1
ReferencesAkiyama, T., Kamimura, K., Firkus, C., Takeo, S., Shimmi, O. and Nakato, H.
(2008). Dally regulates Dpp morphogen gradient formation by stabilizing Dppon the cell surface. Dev. Biol. 313, 408-419.
Belenkaya, T. Y., Han, C., Yan, D., Opoka, R. J., Khodoun, M., Liu, H. and Lin,X. (2004). Drosophila Dpp morphogen movement is independent of dynamin-mediated endocytosis but regulated by the glypican members of heparan sulfateproteoglycans. Cell 119, 231-244.
Billuart, P., Winter, C. G., Maresh, A., Zhao, X. and Luo, L. (2001). Regulatingaxon branch stability: the role of p190 RhoGAP in repressing a retractionsignaling pathway. Cell 107, 195-207.
Bloor, J. W. and Kiehart, D. P. (2002). Drosophila RhoA regulates thecytoskeleton and cell-cell adhesion in the developing epidermis. Development129, 3173-3183.
Casanueva, M. O. and Ferguson, E. L. (2004). Germline stem cell number in theDrosophila ovary is regulated by redundant mechanisms that control Dppsignaling. Development 131, 1881-1890.
Chen, D. and McKearin, D. M. (2003). A discrete transcriptional silencer in thebam gene determines asymmetric division of the Drosophila germline stem cell.Development 130, 1159-1170.
Dahlgaard, K., Raposo, A. A., Niccoli, T. and St Johnston, D. (2007). Capu andSpire assemble a cytoplasmic actin mesh that maintains microtubuleorganization in the Drosophila oocyte. Dev. Cell 13, 539-553.
Decotto, E. and Spradling, A. C. (2005). The Drosophila ovarian and testis stemcell niches: similar somatic stem cells and signals. Dev. Cell 9, 501-510.
Dor, Y., Brown, J., Martinez, O. I. and Melton, D. A. (2004). Adult pancreaticbeta-cells are formed by self-duplication rather than stem-cell differentiation.Nature 429, 41-46.
Fox, D. T. and Spradling, A. C. (2009). The Drosophila hindgut lacksconstitutively active adult stem cells but proliferates in response to tissuedamage. Cell Stem Cell 5, 290-297.
Gonczy, P. and DiNardo, S. (1996). The germ line regulates somatic cyst cellproliferation and fate during Drosophila spermatogenesis. Development 122,2437-2447.
Gonczy, P., Viswanathan, S. and DiNardo, S. (1992). Probing spermatogenesisin Drosophila with P-element enhancer detectors. Development 114, 89-98.
Guo, Z. and Wang, Z. (2009). The glypican Dally is required in the niche for themaintenance of germline stem cells and short-range BMP signaling in theDrosophila ovary. Development 136, 3627-3635.
Hacker, U. and Perrimon, N. (1998). DRhoGEF2 encodes a member of the Dblfamily of oncogenes and controls cell shape changes during gastrulation inDrosophila. Genes Dev. 12, 274-284.
Harrison, D. A. and Perrimon, N. (1993). Simple and efficient generation ofmarked clones in Drosophila. Curr. Biol. 3, 424-433.
Hay, B., Jan, L. Y. and Jan, Y. N. (1988). A protein component of Drosophilapolar granules is encoded by vasa and has extensive sequence similarity to ATP-dependent helicases. Cell 55, 577-587.
Hayashi, Y., Kobayashi, S. and Nakato, H. (2009). Drosophila glypicans regulatethe germline stem cell niche. J. Cell Biol. 187, 473-480.
Jackson, S. M., Nakato, H., Sugiura, M., Jannuzi, A., Oakes, R., Kaluza, V.,Golden, C. and Selleck, S. B. (1997). dally, a Drosophila glypican, controlscellular responses to the TGF-beta-related morphogen, Dpp. Development 124,4113-4120.
Jaffe, A. B. and Hall, A. (2005). Rho GTPases: biochemistry and biology. Annu.Rev. Cell Dev. Biol. 21, 247-269.
Kai, T. and Spradling, A. (2003). An empty Drosophila stem cell niche reactivatesthe proliferation of ectopic cells. Proc. Natl. Acad. Sci. USA 100, 4633-4638.
Kiger, A. A., White-Cooper, H. and Fuller, M. T. (2000). Somatic support cellsrestrict germline stem cell self-renewal and promote differentiation. Nature 407,750-754.
Kirilly, D., Spana, E. P., Perrimon, N., Padgett, R. W. and Xie, T. (2005). BMPsignaling is required for controlling somatic stem cell self-renewal in theDrosophila ovary. Dev. Cell 9, 651-662.
Lasko, P. F. and Ashburner, M. (1988). The product of the Drosophila gene vasais very similar to eukaryotic initiation factor-4A. Nature 335, 611-617.
Lee, T., Winter, C., Marticke, S. S., Lee, A. and Luo, L. (2000). Essential roles ofDrosophila RhoA in the regulation of neuroblast proliferation and dendritic butnot axonal morphogenesis. Neuron 25, 307-316.
Li, L. and Xie, T. (2005). Stem cell niche: structure and function. Annu. Rev. CellDev. Biol. 21, 605-631.
Lin, H., Yue, L. and Spradling, A. C. (1994). The Drosophila fusome, a germline-specific organelle, contains membrane skeletal proteins and functions in cystformation. Development 120, 947-956.
Liu, M., Lim, T. M. and Cai, Y. (2010). The Drosophila female germline stem celllineage acts to spatially restrict DPP function within the niche. Sci. Signal 3, ra57.
RESEARCH ARTICLE Development 138 (23)
DEVELO
PMENT

Magie, C. R., Pinto-Santini, D. and Parkhurst, S. M. (2002). Rho1 interacts withp120ctn and alpha-catenin, and regulates cadherin-based adherens junctioncomponents in Drosophila. Development 129, 3771-3782.
Margolis, J. and Spradling, A. (1995). Identification and behavior of epithelialstem cells in the Drosophila ovary. Development 121, 3797-3807.
McKearin, D. M. and Spradling, A. C. (1990). bag-of-marbles: a Drosophilagene required to initiate both male and female gametogenesis. Genes Dev. 4,2242-2251.
McKearin, D. and Ohlstein, B. (1995). A role for the Drosophila bag-of-marblesprotein in the differentiation of cystoblasts from germline stem cells.Development 121, 2937-2947.
Morris, L. X. and Spradling, A. C. (2011). Long-term live imaging provides newinsight into stem cell regulation and germline-soma coordination in theDrosophila ovary. Development 138, 2207-2215.
Morrison, S. J. and Spradling, A. C. (2008). Stem cells and niches: mechanismsthat promote stem cell maintenance throughout life. Cell 132, 598-611.
Prokopenko, S. N., Brumby, A., O’Keefe, L., Prior, L., He, Y., Saint, R. andBellen, H. J. (1999). A putative exchange factor for Rho1 GTPase is required forinitiation of cytokinesis in Drosophila. Genes Dev. 13, 2301-2314.
Quinlan, M. E., Hilgert, S., Bedrossian, A., Mullins, R. D. and Kerkhoff, E.(2007). Regulatory interactions between two actin nucleators, Spire andCappuccino. J. Cell Biol. 179, 117-128.
Ridley, A. J. (2006). Rho GTPases and actin dynamics in membrane protrusionsand vesicle trafficking. Trends Cell Biol. 16, 522-529.
Sano, H., Nakamura, A. and Kobayashi, S. (2002). Identification of atranscriptional regulatory region for germline-specific expression of vasa gene inDrosophila melanogaster. Mech. Dev. 112, 129-139.
Schultz, C., Wood, C. G., Jones, D. L., Tazuke, S. I. and Fuller, M. T. (2002).Signaling from germ cells mediated by the rhomboid homolog stet organizesencapsulation by somatic support cells. Development 129, 4523-4534.
Song, X. and Xie, T. (2002). DE-cadherin-mediated cell adhesion is essential formaintaining somatic stem cells in the Drosophila ovary. Proc. Natl. Acad. Sci.USA 99, 14813-14818.
Song, X., Zhu, C. H., Doan, C. and Xie, T. (2002). Germline stem cells anchoredby adherens junctions in the Drosophila ovary niches. Science 296, 1855-1857.
Song, X., Wong, M. D., Kawase, E., Xi, R., Ding, B. C., McCarthy, J. J. andXie, T. (2004). Bmp signals from niche cells directly repress transcription of adifferentiation-promoting gene, bag of marbles, in germline stem cells in theDrosophila ovary. Development 131, 1353-1364.
Strutt, D. I., Weber, U. and Mlodzik, M. (1997). The role of RhoA in tissuepolarity and Frizzled signalling. Nature 387, 292-295.
Tran, J., Brenner, T. J. and DiNardo, S. (2000). Somatic control over the germlinestem cell lineage during Drosophila spermatogenesis. Nature 407, 754-757.
Urban, S., Lee, J. R. and Freeman, M. (2001). Drosophila rhomboid-1 defines afamily of putative intramembrane serine proteases. Cell 107, 173-182.
Xie, T. and Spradling, A. C. (1998). decapentaplegic is essential for themaintenance and division of germline stem cells in the Drosophila ovary. Cell 94,251-260.
Xie, T. and Spradling, A. C. (2000). A niche maintaining germ line stem cells inthe Drosophila ovary. Science 290, 328-330.
Zhang, Y. and Kalderon, D. (2001). Hedgehog acts as a somatic stem cell factorin the Drosophila ovary. Nature 410, 599-604.
Zhao, C., Deng, W. and Gage, F. H. (2008). Mechanisms and functionalimplications of adult neurogenesis. Cell 132, 645-660.
5097RESEARCH ARTICLERoles of escort cells
DEVELO
PMENT


















