
Sebenta de Fisiologia
375
Fisiologia Fisiologia Geral Homeostasia - estado de equilíbrio que tende a manter constantes as variáveis do meio interno, sendo este um pr ocesso di nâmico. Ou seja, é a constância do meio interno, varia em limites estreitos. Todos os órgãos e tecidos do corpo exercem funções que ajudam a manter essas condições constantes: • Os pulmões fornecem oxigénio para o líquido extracelular para se repor o que é consumido pelas células; • Os rins mantêm constantes as concentrações iónicas; • O sistema gastrointestinal fornece nutrientes. Para que a homeostasia seja mantida recorre-se a: Sistemas de Controlo Homestático. “Steady-State” ou estado de equilíbrio inicial, em que a estabilidade do meio é conseguida através do balanço entre inputs e outputs, que determinam a set-point de cada variável, não é a sua magnitude que interessa mas sim a sua existência. Os sistemas de controlo homeostático não conseguem manter a constância do meio de for ma per manente, existindo um interv alo de nor malidade que, embora estreito, é fundamental para os mecanismos de regulação. “Feedback regulation”: -Feedback negativo – um aumento ou diminuição na variável que está a ser regulada desencadeia respostas que tendem a mover a variável no sentido oposto à alteração; -Feedback positivo – é o menos comum, já que tende a acelerar o processo, podendo levar a uma “explosão do sistema”. “Feedforward regulation”: antecipa al terações numa variável regulada, como por exemplo a temperatura corporal, melhorando as velocidade de res pos ta e diminuindo flutuações nessa mesma variável, isto é, reduzindo o desvio do set-point. Componentes do Sistema de Controlo Homestático: -Reflexo – resposta imediata, involuntária e específica a um determinado estímulo. 1
Transcript of Sebenta de Fisiologia

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 1/374
Fisiologia
Fisiologia Geral
Homeostasia- estado de equilíbrio que tende a manter constantes asvariáveis do meio interno, sendo este um processo dinâmico. Ou seja, é aconstância do meio interno, varia em limites estreitos.
Todos os órgãos e tecidos do corpo exercem funções que ajudam a manteressas condições constantes:
• Os pulmões fornecem oxigénio para o líquido extracelular para se reporo que é consumido pelas células;
• Os rins mantêm constantes as concentrações iónicas;
• O sistema gastrointestinal fornece nutrientes.
Para que a homeostasia seja mantida recorre-se a: Sistemas de ControloHomestático.
“Steady-State” ou estado de equilíbrio inicial, em que a estabilidade domeio é conseguida através do balanço entre inputs e outputs, que determinam aset-point de cada variável, não é a sua magnitude que interessa mas sim a suaexistência.
Os sistemas de controlo homeostático não conseguem manter a constânciado meio de forma permanente, existindo um intervalo de normalidade que,embora estreito, é fundamental para os mecanismos de regulação.
“Feedback regulation”:-Feedback negativo – um aumento ou
diminuição na variável que está a ser reguladadesencadeia respostas que tendem a mover avariável no sentido oposto à alteração;
-Feedback positivo – é o menos comum, jáque tende a acelerar o processo, podendo levar a
uma “explosão do sistema”.
“Feedforward regulation”: antecipaalterações numa variável regulada, como porexemplo a temperatura corporal, melhorando asvelocidade de resposta e diminuindo flutuaçõesnessa mesma variável, isto é, reduzindo o desviodo set-point.
Componentes do Sistema de Controlo Homestático:-Reflexo – resposta imediata, involuntária e específica a um determinado
estímulo.1

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 2/374
A via que medeia um reflexo denomina-se arco-reflexo e os seuscomponentes são os seguintes:
O centro de integração tem a função de interpretar o estímulo e determinara resposta.
Respostas Homeostáticas Locais – têm início através de um estímulo queinduz uma alteração da actividade celular que vai tentar contrariar o estímulo. No
entanto, ao contrário do reflexo, a resposta ocorre localmente, abrangendoapenas a zona lesada/estimulada.Ocorrem através de:
• Agentes autócrinas – actuam no próprio local onde são produzidas
• Agentes parácrinas – são produzidas numa dada célula à periferia dacélula onde vão actuar.
Processos relacionados com a Homeostasia:- Adaptação e Aclimatação: capacidade que um individuo tem de se
adaptar a uma alteração ambiental. A adaptação mais “estudada” é aAclimatação que se refere à capacidade de adaptação fisiológica face a umaexposição prolongada a uma alteração significativa na temperatura.
- Ritmos Circadianos: ritmos de duração cíclica (24h de duração) quepermitem que os mecanismos homeostáticos sejam utilizados imediata eautomaticamente sem que seja necessária uma activação já que ocorre porprevisão. O sono é um exemplo, assim como o “jet-lag”.
2

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 3/374
Teoria Geral dos ReceptoresReceptores - proteínas específicas, no interior da membrana ou da célula-
alvo, com a qual o mensageiro químico se combina, 2invocando uma resposta
biologicamente relevante.Propriedades dos receptores:
• Especificidade – capacidade do receptor ligar apenas um tipo ou umnúmero limitado de mensageiros químicos relacionados;
• Saturação – grau de ocupação do receptor pelos mensageiros;
• Afinidade – força de ligação do mensageiro ao receptor;
• Competição – capacidade de diferentes moléculas mensageiras,estruturalmente parecidas entre si, competirem entre si pela ligação aomesmo receptor;
o Antagonista – substância farmacologicamente activa que tem acapacidade de se ligar aos receptores e impedir a resposta celular;
o Agonista - substância farmacologicamente activa que, ao ligar-seao receptor, provoca uma resposta semelhante à que omensageiro provocaria.
Regulação dos Receptores:- Down-regulation: quando a concentração extracelular alta de mensageiro
é mantida por algum tempo, o número de receptores para o mensageiro diminui(Mecanismo de Feedback Negativo Local);
- Up-regulation: quando células expostas a uma pequena quantidade demensageiro podem possuir mais receptores para esse mensageiro – desenvolvemmaior sensibilidade – super-sensibilidade
3

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 4/374
Nota: Estas regulações só são possíveis na medida em que há degradação esíntese contínuas de receptores.
Tipos de receptores:- Intracelulares: (Mensageiros Lipossoluveís) funcionam no núcleo como
factores/supressores da transcrição num grupo específico de genes.- De Membrana: (Mensageiros Lipo-insoluvéis ou hidrossoluveís) podemfuncionar como:
• Canais-iónicos – são activados por mensageiros que induzem a aberturados canais (difusão de iões/alterações no potencial de membrana esinais eléctricos)
• Enzimas
• Proteínas jAK Quinases – têm a capacidade de fosforilar uma proteínaatravés da transferência de um grupo de fosfato do ATP para esta,
alterando a sua actividade. Ocorre por activação da parte enzimática doreceptor
• Activadores de proteína G – sendo que as proteínas G vão activarproteínas efectoras da membrana plasmática. Após activação dá-se aalteração conformacional da proteína G, a transdução de sinal, quepromove a produção de sinais citoplasmáticos de maior amplitude.Estes, por sua vez, conduzem ao aparecimento de segundosmensageiros. O complexo mensageiro-receptor liga-se à proteína G queprovoca a dissociação das suas sub-unidades e a ligação da sub-unidade
alfa ao receptor.
Mecanismo de transdução do sinal
Ligação do mensageiro ao receptorActivação do receptor Transdução do
sinalResposta celular
Nota: os mecanismos de transdução de sinal variam para os mensageirosmembranares e intracelulares.
Efectores proteicos:
• Adenilciclase – catalisa a conversão de algumas moléculas de ATP nocitosol em AMP cíclico que vai actuar como segundo mensageiro eactivar a proteína quinase A que ao fosforilar outras proteínas provoca aresposta celular
• Guanilciclase – catalisa a conversão de GTP a GMP cíclico, no citosol, quevai actuar como segundo mensageiro activando a proteína quinase G
• Fosfolipase C – catalisa a quebra de um fosfolípido da membrana
(fosfotidilnositol bifosfato-PIP2) para originar diacilglicerol (DAG) einositol trifosfato (IP3) que funcionam como segundos mensageiros
4

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 5/374
• Ca2+ - funciona também como segundo mensageiro, difundindo-se para acitosol através de um canal iónico, ou do RER
Segundos Mensageiros:
• Cálcio – activa a calmodulina e outras proteínas ligadoras de Ca2+ queactivam proteínas quinases
• AMPc – activa proteínas quinases AMPc dependentes• GMPc – activa proteínas quinases GMPc dependentes
• DAG – activa proteínas quinase c
• Eucosanóides – gerado a partir do ácido araquidónico na membrana, temefeitos autócrinas e parácrinas (relaxamento do músculo liso)
• IP3 – liberta Ca2+ do reticulo endoplasmático rugoso.
Sebenta
A generalidade dos sistemas homeostáticos necessita de assegurar umacomunicação intercelular de natureza química, sendo que o primeiro passo daacção do mensageiro químico intercelular na célula alvo consiste na sua ligação amoléculas proteicas específicas dessa célula.
Estas moléculas são conhecidas como receptores, os quais não são mais doque proteínas localizadas na membrana plasmática ou dentro da própria célula.
Os receptores de membrana são as entidades mais comuns, enquantooutros servem como "vias preferenciais" de hormonas lipossolúveis com acapacidade de atravessar a membrana plasmática e ligar-se aos receptoresespecíficos no interior da célula.
A existência de receptores explica uma importante característica da
comunicação intercelular: a especificidade. Desta forma, um mensageiro químico(uma hormona, um neurotransmissor ou um agente parácrino) pode contactarcom tipos de células muito diferentes, influenciando somente algumas delas bem
5

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 6/374
determinadas: aquelas que possuem os receptores específicos do mensageiroquímico.
É importante desde já esclarecer que, em muitos casos, onde diferentes decélulas possuem o mesmo tipo de receptor para um mensageiro particular, asrespostas das várias células ao mensageiro podem ser diferentes. É distoexemplo o neurotransmissor noradrenalina (ou norepinefrina) que provoca a
contracção do músculo liso de um vaso sanguíneo e, pela via do mesmo receptor,estimula as células do pâncreas a produzirem menos insulina. Na sua essência, oreceptor funciona como um interruptor molecular o qual, "ligado" pelomensageiro, provoca uma resposta celular. Um só tipo de receptor pode serusado para produzir diferentes respostas em diferentes tipos de células.
Existe ainda uma outra característica fundamental: uma célula pode terdiferentes tipos de receptores para um só mensageiro. A combinação deste comum dos tipos de receptor provocará uma resposta celular diferente daquela queseria provocada se o mesmo mensageiro se ligasse a outro tipo de receptor dacélula. A ligação do mensageiro aos diferentes tipos de receptores estárelacionada com a afinidade destes para o primeiro. Uma célula possui, assim,um número elevado de diferentes tipos de receptores para diferentes tipos demensageiros.
Outras características importantes da interacção entre os mensageiros e osreceptores são os fenómenos de saturação e competição:
Saturação - na maioria dos sistemas a resposta de uma célula a um mensageirocresce à medida que a sua concentração extracelular aumenta, uma vez quetambém se eleva o número de receptores ocupados por esse mensageiro. No
entanto, a resposta celular está limitada superiormente, dado que existe umnúmero finito de receptores disponíveis que ficam saturados por umadeterminada concentração extracelular de mensageiro.
Competição - a competição pelos receptores resulta da capacidade de diferentesmoléculas mensageiras, estruturalmente q1uito parecidas, competirem entre sipor um receptor. É neste contexto que surgem os conceitos de:
• Antagonista - substância farmacologicamente activa que, por serestruturalmente parecida com o mensageiro fisiológico, tem a capacidadede se ligar aos mesmos receptores bloqueando-os, não permitindo assim
que haja uma resposta celular.• Agonista - substância farmacologicamente activa que, por ser
estruturalmente parecida com o mensageiro fisiológico, tem a capacidadede se ligar aos mesmos receptores activando-os, provocando assim umaresposta celular igual àquela que seria produzida por aquele mensageiro.
Regulação dos receptores
Os receptores estão sujeitos a uma regulação fisiológica. O número destesnuma célula e a sua afinidade para um mensageiro específico podem
efectivamente ser aumentados (aumentando a sensibilidade da célula ao agente- hormonal ou outro e, portanto, aumentando a expressão de actividade desse
6

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 7/374
agente) ou diminuídos (diminuindo a sensibilidade da célula ao agente e a suaactividade fisiológica).
Um exemplo importante desta regulação é o fenómeno de regulaçãonegativa (down regulation), significando que quando uma concentraçãoextracelular alta de mensageiro é mantida por algum tempo, o número total dereceptores para o mensageiro pode diminuir. Verifica-se assim que a regulaçãonegativa tem a capacidade de diminuir a resposta da célula-alvo a uma
estimulação frequente e intensa pelo mensageiro, representando desta forma ummecanismo de feedback negativo local. Podem também ocorrer alterações nosentido oposto - regulação positiva (up regulation). As células expostas a umaconcentração muito baixa de mensageiro durante um longo período podemapresentar um número superior de receptores para esse mensageiro,desenvolvendo assim uma maior sensibilidade este.
É importante não esquecer que a regulação positiva e negativa só sãopossíveis porque há uma degradação e síntese contínuas de receptores.
Mecanismos de transdução de sinal para os receptores de membrana plasmática
A ligação de um dado mensageiro (primeiro mensageiro) a um determinadoreceptor específico provoca uma alteração na conformação deste último. Estefenómeno, conhecido por activação do receptor, é sempre o passo inicial de umaresposta celular posterior à ligação desse mensageiro, a qual pode ser umaalteração na permeabilidade, nas propriedades de transporte ou no potencialeléctrico da membrana plasmática da célula; uma alteração no metabolismo dacélula; uma alteração na actividade secretória da célula ou ainda uma alteraçãona actividade contráctil se for uma célula muscular.
Quando um determinado ligando se liga ao domínio extracelular doreceptor induz uma alteração da estrutura tridimensional deste, activando-o.Ocorre assim a transdução de sinal que irá permitir a amplificação do sinaloriginal, isto porque o receptor activado estimula uma actividade catalítica queproduz sinais citoplasmáticos de maior amplitude que o sinal original. Estes sinaisconduzem muitas vezes ao aparecimento de moléculas no interior da célula(segundos mensageiros) que serão responsáveis pela transdução do sinal inicial.
As sequências de acontecimentos entre a activação do receptor e aresposta celular podem, no entanto, ser mais complexas, sendo designadas pormecanismos de transdução de sinal. O sinal é a activação do receptor e atransdução é o processo pelo qual um estímulo é transformado numa resposta.
Os mecanismos de transdução de sinal diferem consoante se trata demensageiros lipossolúveis ou mensageiros hidrossolúveis, uma vez que oscorrespondentes receptores, como anteriormente foi referido, apresentamlocalizações distintas.
Os receptores para mensageiros lipossolúveis são intracelulares e, quandoactivados, actuam no núcleo como moduladores da transcrição genética. Osreceptores para mensageiros hidrossolúveis localizam-se na membranaplasmática, sendo classificados de acordo com os mecanismos de transdução desinal em:
1. Receptores que eles próprios funcionam como canais iónicos2. Receptores que eles próprios funcionam como proteínas quinases,especificamente tirosina quinases
7

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 8/374
3. Receptores que activam proteínas G que, por sua vez, activam proteínasefectoras na membrana plasmática, podendo estas ser:
• Adenilciclase que catalisaa formação de AMP cíclico• Guanilciclase que catalisaa formação de GMP cíclico• Fosfolipase C que catalisa a formação de diacilglicerol e inositol trifosfato• Canais iónicosOs receptores que funcionamcomo canaisiónicos são activadospor
mensageiros que induzem a abertura dos canais,da qual resultaum aumento dadifusão de iões através da membrana plasmática,estando assim associadaaalteraçõesno potencial de membrana e geraçãode sinaiseléctricos.
Os receptores que funcionam como proteínas quinases (enzimas) têm acapacidade de fosforilar uma proteína através da transferência de um grupofosfato do ATP para esta, o que origina uma alteração da actividade da proteínafosforilada que é muitas vezes também uma enzima. Desta forma, ummensageiro liga-se à face extracelular do receptor originando, uma alteração daconformação deste, de tal modo que a parte enzimática do receptor (localizadana face intracelular) é activada, fosforilando então proteínas específicas do
citosol e da membrana plasmática, incluindo ela própria. A fosforilação destasproteínas origina então alterações das suas actividades, das quais resultam asmais variadas respostas celulares. Os receptores que funcionam especificamentecomo tiro sina quinases têm a capacidade de fosforilar a porção tirosina dasproteínas, estando os mensageiros destes receptores muitas vezes envolvidos nocrescimento e desenvolvimento.
Os receptores que activam as proteínas G são maioritários, incluindoaproximadamente 100 formas distintas. Estes receptores, após activação,interagem com uma ou mais proteínas de membrana pertencentes à farm1ia dasproteínas heterotriméricas designadas por proteínas G, nome derivado deelevada afinidade que estas proteínas têm de ligar nucleótidos guanosina (GDP e
GTP). Na forma inactiva, a proteína G é formada por duas subunidadesassociadas, Gj3Y e Gsu, ligadas ao GDP (Gj3y-Gsu-GDP). O complexo mensageiro-receptor liga-se à proteína G, o que provoca a substituição do GDP pelo GTP e adissociação da subunidade u-GDP da subunidade Gj3Y.
Embora haja excepções, na maioria dos casos a subunidade u-GTP liga-seao efector proteico que pode ser um canal ou uma enzima o que, neste últimocaso, pode activar a síntese do 2.° mensageiro.
Enquanto a proteína Gs se mantiver ligada ao GTP, o efector mantém-seactivo, mas ao dar-se a hidrólise do GTP a GDP ocorre a associação dassubunidades u-GDP e Gj3Y,inactivando-se o efector.
Se existem proteínas G estimuladoras, não é menos verdade a existênciade Proteínas G inibidoras que são formadas por uma subunidade Giu que se podeligar ao GTP ou GDP por uma subunidade Gj3Y.
Os efectores proteicos podem então ser:• A adenilciclase que, ao ser activada pela proteína Gs, catalisa a conversão dealgumas moléculas de ATP no citosol em 3', 5'- adenosina monofosfato cíclica ouAMP cíclico (AMPc) que vai actuar como segundo mensageiro. O AMPc difunde-sedepois na célula para activar uma enzima conhecida como proteína quinas eAMPc-dependente (ou proteína quinase A), enzima essa que vai fosforilar outrasproteínas, originando assim a resposta celular.• A guanilcic1ase que, ao ser activada pela proteína Gs, catalisa a conversão de
algumas moléculas de GTP no citosol em 3', 5'- guanosina monofosfato cíclica ouGMP cíclico (GMPc) que vai actuar como segundo mensageiro. O GMPc difunde-sedepois na célula para activar uma enzima conhecida como proteína quinas e
8

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 9/374
GMPc-dependente (ou proteína quinase G), enzima essa que vai fosforilar outrasproteínas, originando assim a resposta celular.• A fosfolipase C que, ao ser estimulada pela proteína Gs, catalisa a quebra deum fosfolípido da membrana plamática 'denominado de fosfatidilinositol bifosfato(PIP;) para originar diacilglicerol (DAG) e inositol trifosfato (IP3). Quer o DAG, quero IP3 funcionam como segundos mensageiros, mas em diferentes vias. O DAGactiva uma proteína quinase denominada de proteína quinase C (C de cálcio,
porque o Ca2+ é necessário na formação do DAG), a, qual por sua vez fosforilaum largo número de outras proteínas, originando a resposta celular. O IP3difunde-se no citosol actuando no retículo endoplasmático (denominado deretículo sarcoplasmático no músculo), onde altera a permeabilidade damembrana deste ao ião cálcio. A concentração do Ca2+ é muito maior no interiordeste organito do que no citosol e, assim, o ião difunde-se do retículo para ocitosol, o que faz aumentar a sua concentração neste compartimento da célula. Ocálcio irá então funcionar também como segundo mensageiro, induzindo aresposta celular através de vários mecanismos.• Um canal iónico, cuja abertura e fecho estão dependentes da interacção com aproteína G.
Inicialmente a glicose está presente no compartimento 1 com umaconcentração de 20mmol/L e não há glicose no compartimento 2. Os movimentosaleatórios das moléculas de glicose no compartimento 1 levam parte dela para ocompartimento 2. A quantidade de material que atravessa uma superfície numaunidade de tempo denomina-se de fluxo. Este fluxo de glicose de sentido únicodo compartimento 1 para o 2 depende da [glicose] no 1.º compartimento. Se onúmero de moléculas num determinado volume duplicar, o fluxo de moléculasatravés de cada superfície da unidade também será duplicado, visto que semovem o dobro das moléculas em qualquer sentido num determinado tempo.
Após um curto período de tempo, parte das moléculas de glicose queentraram no compartimento 2 mover-se-ão aleatoriamente para o compartimento1 (tempo B). A magnitude do fluxo de glicose do compartimento 2 para o 1depende da [glicose] no compartimento 2 em qualquer momento.
O fluxo resultante de glicose entre os dois compartimentos em qualquermomento é a diferença entre os dois fluxos de sentido único. É o fluxo resultanteque determina o ganho final de moléculas pelo compartimento 2 e a perda finaldo compartimento 1.
Finalmente, as [glicose] nos dois compartimentos tornam-se iguais
(10mmol/L). Os dois fluxos de sentido único têm a mesma magnitude massentido oposto, sendo o fluxo resultante da glicose zero (tempo C). O sistemaalcança o equilíbrio de difusão. Nenhuma alteração na [glicose] nos dois
9

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 10/374
compartimentos ocorrerá uma vez que o número de moléculas a difundir-se nosdois sentidos é igual.
.
Movimento de solutos na membrana
10

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 11/374
Co transporte e Contratransporte
11

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 12/374
OSMOSE – difusão resultante de água através da membrana
12

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 13/374
Fenómenos Bioeléctricos de MembranasExcitáveis
Em condições fisiológicas normais umacélula apresenta:
• Meio Interno com maior quantidade deK +
• Meio Externo com maior quantidade deNa+ e também Cl-
Assim há um excesso de cargas negativas
no interior face às cargas positivas no exteriorque faz com que estas se atraiam fortemente aolongo da membrana – Potencial da Membrana oude Repouso. Esta diferença entre fluidos intra eextracelular é regularizada através da Bomba deSódio-Potássio.
13

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 14/374
A bomba expulsa para o interstício Na+ e capta K + para o citoplasma,enquanto o movimento inverso ocorre por difusão. (A bomba vai repolarizar)Assim, o potencial de equivalência do K + é de -90mV (aprox) enquanto o do Na+ éde +60mV.
Experimentalmente (através de leituras por voltímetros) verificou-se que opotencial de repouso é de -70mV, o que torna a membrana mais permeável ao K +
por estar mais perto do seu potencial de equivalência.O potencial de repouso é então gerado devido aos movimentos de K + parafora da célula, através de canais, fazendo com que o interior se torne maisnegativo.
Alterações transitórias no potencial da membrana, a partir do potencial derepouso, produzem sinais eléctricos podem estes ocorrer de duas formas:
• Potenciais GradativosSão sinais de curta distância, na medida em que resultam de alterações nopotencial de membrana confinadas a uma pequena área da mesma. Estes
potenciais chamam-se gradativos porque a magnitude da alteração depotencial pode variar e é proporcional à magnitude do estímulo. Se acorrente for negativa ocorre uma hiperpolarização da membrana, enquantose a corrente for positiva ocorre uma despolarização. Os potenciaisgradativos são os únicos meios de comunicação usados pelos neurónios,desempenhando um papel muito importante na iniciação e integração desinais por neurónios e outras células, sendo transmitidos passivamentedependendo da quantidade de carga que se desloca, da resistência local damembrana e do citoplasma, sendo por issotransmitidos decrementalmente, isto é,decresce com a distância à origem daalteração. Para além disto, estes potenciaissão somáveis, podendo ocorrer somaçõesse houver repetição do estímulo antes deocorrer repolarização da membrana.
• Potenciais de Acção:Ocorrem se a quantidade de cargadespolarizante for suficiente paraultrapassar a corrente local. Estescorrespondem a alterações rápidas nopotencial de membrana, podendo esteapresentar variações na ordem dos 100mV.As membranas capazes de produzirpotenciais de acção são chamadas demembranas excitáveis e encontram-se emcélulas nervosas, musculares, imunitárias,endócrinas e reprodutoras. A excitabilidadepode ser diminuída por diminuições do K +
intracelular, por hipercaliémia ou por inibiçãodos canais de Na+. O potencial de acção, que
14

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 15/374
ocorre sem alteração dos gradientes de concentração membranares maspor alteração das permeabilidades membranares ao sódio e ao potássio,ocorre em duas fases:
o Fase de despolarização – há abertura de canais de sódio voltagemdependentes; a concentração de iões sódio no interior aumenta (onúmero de iões sódio que entra supera o número de iões que sai
através da bomba sódio potássio) fazendo com que o potencial damembrana se aproxime do potencialde equilíbrio do sódio. Ocorre assimuma polarização positiva em que ointerior da célula fica mais positivoque o exterior.
o Fase de repolarização – dá-se oencerramento dos canais de sódiovoltagem-dependentes e a abertura
dos canais potássio. Como a difusãode potássio para fora da célula émuito superior à difusão de sódiopara dentro desta, permitindo que opotencial de membrana volte aonormal (-70mV).
Nota: 1 em cada 100000 iões de sódio entram na célula durante o processo deacção e o mesmo número de iões potássio difundem-se para o exterior da célula,
15

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 16/374
fazendo o potencial de membrana regressar aos níveis de repouso. Como omovimento de iões é tão pequeno produz somente infinitésimas alterações nas []iónicas logo o potencial de acção não está associado às alterações de [] iónicas. Afase de despolarização é exemplo de feedback positivo.
Propriedades bioeléctricas das membranas excitáveis:
• Lei do Tudo ou Nada – o potencial de acção só ocorre quando o estímulo
é capaz de atingir (em intensidade e duração) o limiar de excitação –valor de voltagem a partir do qual se dá o potencial de acção – da célula,que responde segundo a lei do tudo ou nada:o só há resposta se o estimulo atingir o limiar de excitação;o quando se atinge o limiar de excitação a resposta da célula tem
sempre a mesma magnitude, independentemente de os estímulosserem ou não supra-limiares (de maior intensidade e duração que osestímulos normais).
• Refractariedade da Membrana: os períodos refractários são o espaço de
tempo no qual a resposta da membrana se encontra alterada por umestímulo prévio.o Período Refractário Absoluto: por mais forte que seja o estímulo, não
é capaz de provocar potencial de acção, pois quase a totalidade doscanais sódio voltagem dependentes estão inactivos.
o Período Refractário Relativo: pode haver novo potencial de acção se oestímulo for supra-limiar, pois os canais de sódio já se encontramparcialmente fechados e são capazes de responder a um novoestímulo.
Isto é, depois de um potencial de acção é necessário um determinadotempo para que os canais sensíveis à voltagem voltem à suaconformação de repouso.
• Reobase, Tempo de Utilização e Cronaxia: os tecidos excitáveis sãocaracterizáveis através da determinação da intensidade mínima deestimulação e do tempo mínimo necessário para obter uma respostapropagável.o Reobase: intensidade mínima de estímulo que desencadeia o
potencial de acção.o Tempo de utilização: tempo mínimo necessário para atingir o limiar
de excitação quando aplicado um estímulo com intensidade igual àreobase.
16

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 17/374
o Cronaxia: tempo mínimo necessário para atingir o limiar de excitaçãoquando aplicado um estímulo com intensidade dupla da reobase.
• Condução dos potenciais de acção: a velocidade com que se dá apropagação de um potencial de acção é directamente proporcional aodiâmetro da fibra e depende da mielinização, sendo mais rápida nosNodos de Ranvier, onde a concentração de canais de sódio é elevada e
há uma interrupção da mielinização das fibras – condução saltatória denodo em nodo. Nos nodos de Ranvier apenas existe uma camada decélulas de Shwann chamada de neurilema. Quanto mais grossa a fibramais rápida é a propagação, pois oferecem menor resistência àscorrentes locais pelo que as zonas adjacentes atingem o limiar deexcitação mais rápido. Neste caso, e ao contrário dos potenciaisgradativos, a propagação não é decremental, o estímulo provoca umpotencial de acção que chega ao final da membrana igual ao inicial. Acondução pode ocorrer através de um ciclo de feedback positivo, ou por
um potencial gradativo (Somação) originado por um input sinápitco noneurónio, ou através de um potencial pacemaker (alteração espontâneano potencial de membrana do neurónio). Este último está implicado emcomportamentos rítmicos como a respiração, batimentos cardíacos emovimentos na parede do estômago e intestinos.
Nota: a mielina é um isolador que torna mais difícil a difusão de cargasentre os 2 lados da membrana. As fibras com mielina têm um diâmetro
superior a 1m conduzindo o impulso nervoso mais rapidamente que as
amielínicas. A condução saltatória é mais rápida do que a propagaçãonuma fibra desmielinizada dado que 1 menos número de cargas saem pelaszonas mielinizadas, traduz-se ainda por uma poupança de volume(mielinização em detrimento do aumento do diâmetro da fibra) e de energia(menor transferência de cargas e portanto menor necessidade debombagem). Potencial Receptor: nos neurónios aferentes a despolarizaçãoinicial até ao limiar de excitação é conseguida pelo potencial gradativooriginado por receptores das terminações nervosas periféricas dosneurónios.
17

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 18/374
• Somação e Acomodação: quanto mais longo for o intervalo de tempoentre os 2 estímulos, maior terá que ser a intensidade do 2.º estímulo demodo a provocar resposta. É possível provocar-se uma resposta a partirda soma de 2 estímulos abaixo do limiar de excitação (a duração do 2.ºestímulo é de 1ms).
Os Estímulos podem ser:
• Químicos: substância provoca abertura do canal;
• Mecânicos: vibração/oscilação que provoca a entrada de Na+;
• Eléctricos: modificação da carga eléctrica da célula.
Demonstrações sequenciais:
18

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 19/374
• Modelo de Hodgkin-Huxley: a membrana nervosa é representada comotendo três canais através dos quais se movimentam livremente o cloro,sódio e potássio. os iões individuais são mais forçados a movimentar-sena membrana devido aos seus gradientes electroquímicos. Estes valoresda força electromotriz são dados pela equação de Nernst.
• Somação Temporal – potencial de membrana em que duas ou mais
aferências que estão ocorrendo em diferentes momentos sãoadicionadas conjuntamente; a alteração potencial é maior que a causadapor qualquer uma das aferências isoladamente.
• Somação Espacial – adição de efeitos de entradas simultâneas paradiferentes lugares em um neurónio para produzir alteração potencialmaior que aquela causada por uma única entrada.
Sebenta
Potenciais GradativosOs potenciais gradativos são importantes em sinais de curta distância,
enquanto os potenciais de acção são sinais de longa distância de célulasnervosas e membranas das células musculares. Os potenciais gradativoscorrespondem, assim, a alterações no potencial da membrana confinados a umaregião muito curta da membrana plasmática (desaparecem ao fim de 1 a 2 mmdo sítio de origem). Podem ser alterações das quais resultará umahiperpolarização ou uma despolarização. São chamados de potenciais gradativosporque a magnitude do potencial é variável e proporcional à magnitude doestímulo.
Quando ocorre um potencial gradativo as cargas eléctricas movem-se entre
o sítio de origem do potencial e as regiões adjacentes da membrana plasmáticaque estão no potencial de repouso. Quanto maior for a alteração do potencialmaiores são as correntes. Estas correntes (movimentação de cargas) provocamdespolarização das regiões adjacentes da membrana. As correntes locais sãooriginadas pelas movimentações dos iões como K+,Na+, Cl- e HC03-.
A magnitude da corrente é decremental com o aumento da distância aosítio de origem da alteração do potencial, ou seja, a alteração do potencial demembrana é assim também decremental com o aumento da distância.
Como os potenciais gradativos decrescem com a distância só podem serusados como sinais para curtas distâncias (poucos mm). No entanto, ospotenciais gradativos são os únicos meios de comunicação usados por neurónios
e desempenham um papel muito importante na iniciação e integração de sinaispor neurónios e outras células.
Potenciais de AcçãoSão alterações rápidas do potencial de membrana que duram cerca de 1
ms, durante o qual o potencial de membrana pode apresentar variações deamplitude da ordem dos 100 mV (de -70 mV a +30 mV) e depois regressarnovamente até ao potencial de repouso. As células nervosas e musculares, bemcomo algumas células endócrinas, imunológicas e reprodutoras têm membranasplasmáticas capazes de produzir potenciais de acção. Estas membranas são
chamadas de membranas excitáveis e a sua capacidade para originar potenciaisde acção é conhecida como excitabilidade.
19

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 20/374
A magnitude do potencial da membrana em repouso depende dosgradientes de concentração e das permeabilidades para diferentes iões,particularmente o sódio e o potássio. Por seu lado, o potencial de acção resultade uma alteração transitória na permeabilidade da membrana, ao passo que osgradientes de concentração se mantêm praticamente inalteráveis.
No potencial de repouso a grande maioria dos canais de sódio estãofechados e assim este está muito mais próximo do potencial de equilíbrio do
potássio (-89 mV) do que do de sódio (+60 mV), porque a membrana é maispermeável ao K+ do que ao Na+. No entanto, durante o potencial de acção aspermeabilidades ao sódio e ao potássio vão ser alteradas. Com efeito, o potencialde acção é fundamentalmente caracterizado pela presença de duas fases, asquais traduzem as profundas alterações iónicas que se observam através damembrana celular.
A fase de despolarização do potencial de acção é devida à abertura doscanais de sódio voltagem dependentes que aumentam assim bastante apermeabilidade aos iões sódio da membrana, permitindo que os iões sódioentrem para dentro da célula. Durante este período, o n.º de iões sódio que entrapara dentro da célula supera o n.o de iões de sódio que sai de dentro da célulaatravés da bomba sódio-potássio ATPase, ocorrendo assim a despolarização emesmo uma polarização positiva (o interior da célula fica mais positivo que oexterior). É neste período que o potencial de membrana se aproxima mais dopotencial de equilíbrio do sódio.
Após a despolarização a membrana sofre uma fase de repolarização,fazendo com que o potencial de membrana volte aos níveis de repouso. Estarepolarização é devida ao encerramento dos canais de sódio (abertos durante adespolarização) e à abertura dos canais de potássio voltagem dependentes, oque faz com que a difusão de potássio para fora da célula seja muito superior à
difusão de sódio para dentro desta, permitindo assim que o potencial demembrana volte ao nível de repouso. Todo este processo pode levar-nos a pensar que seria necessário a
movimentação de uma enorme quantidade de iões para garantir este processo.Contudo, somente 1 em cada 100.000 iões de sódio entram na célula
durante o potencial de acção e aproximadamente o mesmo n.º de iões potássiodifundem-se para fora da célula, fazendo o potencial de membrana regressar aosníveis de repouso. Estes movimentos de iões são tão pequenos que produzemsomente infinitésimas alterações nas concentrações iónicas intracelulares e é porisso que se diz que o potencial de acção não está associado a alteraçõesobserváveis de concentrações iónicas, como anteriormente referido. No entanto,
se estse poucos iões que atravessam a membrana com o potencial de acção nãopudessem ser devolvidos através da bomba sódio-potássio, os gradientes deconcentração do sódio e do potássio iriam gradualmente desaparecendo e ospotenciais de acção não poderiam mais ser originados.
Resulta assim claro que, no potencial de acção, a 1.ª parte dadespolarização é primariamente devida a correntes locais que promovem aabertura dos canais de sódio. Desta forma, a despolarização faz abrir os canaisde sódio para que aumente a permeabilidade a este ião e, devido ao aumentodesta permeabilidade, os iões positivos de sódio difundem-se mais facilmentepara o interior da célula, permitindo assim a despolarização da membrana - esteé mais um exemplo de feedback positivo.
20

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 21/374
Estes períodos refractários reflectem assim o facto de, após um potencialde acção, ser necessário um determinado tempo para os canais de sódiovoltagem dependentes passarem do estado de inactivação, adquirido no fim dafase de despolarização, para o estado de fechados que corresponde à suaconformação de repouso capaz de responder a um novo estímulo.
A propagação de um potencial de acção dá-se quando uma corrente local ésuficientemente intensa para servir de estímulo que despolarizará as regiões
adjacentes da membrana até ao limiar de excitação, atingido quando a correntede Na+ para o interior da célula supera a corrente de K+ para o exterior. O ciclode feedback positivo de sódio iniciar-se-á, ocorrendo um novo potencial de acção,o qual produz então correntes locais dele próprio que, por sua vez, irãodespolarizar as regiões adjacentes, produzindo um novo potencial de acção numnovo sítio, e assim por diante, causando a sua propagação através de todo ocomprimento da membrana plasmática.
Dado que cada potencial de acção depende do ciclo de feedback positivode sódio no local da membrana onde ocorre, o potencial de acção que chega aofinal da membrana é igual ao inicial, ou seja, a condução dos potenciais de acçãonão é decremental como os potenciais gradativos.
Condução dos Potenciais de Acção
A velocidade com que se dá a propagação de um potencial de acção éproporcional ao diâmetro da fibra que o conduz e depende da fibra ser ou nãomielinizada. Quanto mais grossa for a fibra mais rápida é a propagação, dado queas fibras mais grossas oferecem menor resistência às correntes locais e, assim,as regiões adjacentes da membrana atingem mais rapidamente o limiar deexcitação.
A mielina é um isolador que torna mais difícil a difusão de cargas entre osdois lados da membrana. Assim, esta difusão ocorre nos Nçdos de Ranvier, ondea cobertura de mielina está interrompida e onde a concentração em canais desódio é elevada. Os potenciais de acção vão "saltar", desta forma, de Nodo emNodo, constituindo a denominada condução saltatória. A propagação viacondução saltatória é mais rápida que a propagação numa fibra desmielinizadade igual espessura, dado que um menor n.Ode cargas saem através das secçõesmielinizadas da membrana. Deste modo, mais cargas chegam ao Nodo adjacenteactivo e um potencial de acção é aí originado mais cedo do que se a mielina nãoestivesse presente. Para além de ser mais rápida, a condução saltatória traduz-setambém por uma poupança de volume (mielinização em detrimento do aumento
do diâmetro da fibra) e de energia (menor transferência de cargas e portantomenor necessidade de bombagem).
Nos neurónios aferentes a despolarização inicial até ao limiar é conseguidaatravés de um potencial gradativo originado nos receptores das terminaçõesnervosas periféricas dos neurónios (potencial Receptor). Em todos os outrosneurónios a despolarização até ao limiar é devida, quer a um potencial gradativooriginado por um input sináptico no neurónio, quer por uma alteração espontâneano potencial de membrana do neurónio, conhecida como Potencial Pacemaker.Este último tipo de potencial está implicado em muitos comportamentos rítmicos,como a respiração, os batimentos cardíacos e movimentos das paredes doestômago e intestinos.
Somação e Acomodação
21

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 22/374
A aplicação de um estímulo abaixo do limiar de excitação provoca um efeitoresidual no neurónio, embora o impulso não seja explícito. Este efeito é reveladopela aplicação de um segundo estímulo, também abaixo do limiar de excitação,com a duração de 1 ms, o qual provoca uma resposta explícita.
Quanto mais longo for o intervalo entre os dois estímulos maior terá queser a intensidade do segundo para provocar uma resposta. Podemos então dizer
que dois estímulos abaixo do limiar de excitação conseguem somar os seusefeitos na membrana nervosa de modo a provocar uma resposta propagávelPotencial de Acção. O primeiro estímulo sub-limiar produz uma alteração nopotencial da membrana que dura 1 ms ou mais, alteração essa que aumenta oefeito do segundo estímulo.
Condução Saltatória
Após deixarem a substância cinzenta, alguns axónios adquirem umrevestimento de uma substância lipídica chamada mielina. A mielina encontrasenormalmente associada aos neurónios somáticos de grande diâmetro,distinguindo-se assim dois tipos de fibras nervosas: as que contêm essa camadade mielina (mielínicas) e os que não a possuem (amielínicas). Normalmente asfibras com mielina têm um diâmetro superior a 1 ml-L,conduzindo o impulsonervoso mais rapidamente que as amielínicas.
Nas fibras mielínicas, o revestimento de mielina é regularmenteinterrompido em locais que se designam por Nodos de Ranvier. É devido àexistência destes locais desprovidos de mielina que se dá esta forma decondução nervosa especial nestas fibras, designada por "condução saltatória"para ilustrar a ocorrência de potenciais de acção apenas e só nos Nodos deRanvier.
Comparando com a camada mielinizada, os nodos de Ranvier contêmapenas uma camada de Células de Shwann chamada de neurilema que emcondições propícias é muito mais permeável aos iões sódio e potássio, fazendo-seassim a condução nervosa através de "saltos" entre os nodos, em vez de ser umprocesso contínuo.
Sinapse e Transmissão Sináptica
Sinapse: junção anatómica especializada, entre dois neurónios, no qual aactividade eléctrica de um neurónio (pré-sináptico) vai influenciar a actividadeeléctrica/metabólica de um segundo neurónio ou célula muscular (pós-sináptico).Anatomicamente, a sinapse para além dos neurónios pré e pós-sináptico incluitambém o espaço extracelular entre os dois neurónios a fenda sináptica.
As sinapses são estruturas fundamentais na transmissão de informaçãoproveniente do (ou dirigida ao) Sistema Nervoso Central. Trata-se de junçõesanatomicamente especializadas entre duas células efectoras, as quais sedestinam a assegurar que a actividade eléctrica da célula pré-sináptica(neurónio) seja transmitida à célula pós-sináptica (neurónio ou célula muscular).
Quando estão activas, as sinapses podem aumentar ou diminuir aactividade no neurónio pós-sináptico, produzindo neste um breve potencialgradativo. O potencial de membrana de um neurónio pós-sináptico aproxima-se
22

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 23/374
do limiar de excitação numa sinapse excitatória e afasta-se deste limiar numasinapse inibitória.
O nível de excitabilidade, a qualquer momento, de uma célula pós-sináptica, ou seja, a proximidade do potencial de membrana do limiar deexcitação, depende do número de sinapses activas e da sua qualidade excitatóriaou inibitória. Se a membrana do neurónio pós-sináptico atingir o limiar deexcitação ocorrerá um potencial de acção que se irá propagar pelo axónio até às
suas terminações, divergindo para influenciar a excitabilidade de muitos outrosneurónios.
As sinapses podem ser de 2 tipos, quanto à acção:
• Excitatórias: quando provocam um potencial de acção, ou seja, adespolarização do neurónio pós-sináptico;
• Inibitórias: quando provocam a hiperpolarização ou estabilização do nívelde potencial no neurónio pós-sináptico.
Para além disto, podemos ainda descrever 2 tipos de sinapses quanto à suaconvergência:
• Sinapses convergentes: milhares de células pré-sinápticas afectam apenasuma célula pós-sináptica;
• Sinapses divergentes: em que uma única célula pré-sináptica enviaprojecções que afectam mais que uma célula pós-sináptica.
Anatomia Funcional das Sinapses
Sinapses eléctricas: as membranas plasmáticas dos neurónios pré e pós-sináptico estão unidas por junções GAP, que permitem que correntes locaisresultantes de potenciais de acção no neurónio pré-sináptico, fluam directamenteatravés da junção, por canais, para a célula pós-sináptica, despolarizando amembrana até ao limiar de excitação originando assim um potencial de acçãonesta última célula. As membranas encontram-se separadas cerca de 2nm eencontram-se ligadas por canais compostos por 6 sub-unidades (conexinas).Através destes canais passam os iões transmitindo-se assim o impulso nervoso.
São as sinapses mais raras do sistema nervoso.
Sinapses Químicas: As sinapses químicas, mais frequentes que as eléctricas, caracterizam-se
pela presença de um mediador químico: o neurotransmissor, que assegura atransmissão do impulso. Quando são libertados mais de 2 neurotransmissoresdenomina-se cotransmissor. (As células encontram-se separadas por 20 nm)
Na sinapse química o neurónio pré-sináptico termina com uma estruturaligeiramente achatada, enquanto a membrana do neurónio pós-sináptico égeralmente pouco espessa, estando estas células separadas pela fenda sináptica,
um espaço extracelular que assim previne a propagação directa de correntes dacélula pré para a célula pós-sináptica.
23

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 24/374
Os neurotransmissores são armazenados em vesículas localizadas noterminal do axónio na célula pré-sináptica. Cada vesícula contém entre 10 4 e5x104 moléculas. Estes são libertados na fenda sináptica quando um potencial deacção da célula pré-sináptica atinge o terminal do axónio e despolariza amembrana, com abertura de canais de cálcio voltagem-dependentes, permitindoassim que o cálcio se difunda do fluido extracelular para o terminal do axónio. Alibertação do neurotransmissor ocorre por fusão das vesículas sinápticas com a
membrana plasmática da célula pré-sináptica onde se dá uma exocitose para afenda sináptica, e está dependente da permeabilidade ao cálcio por parte dacélula pré-sináptica. A acostagem das vesículas é mediada por um conjunto deproteínas auxiliares, entre as quais as proteínas SNARE, encontradas namembrana vesicular e pré-sináptica. Na fusão das vesículas parece estar tambémenvolvida uma outra proteína auxiliar designada por Sinaptotagmina, a qual seassocia às proteínas SNARE e se liga ao cálcio.
Após a exocitose, a membrana da vesícula mantém-se unida com amembrana do terminal, aumentando a superfície desta. As vesículas invaginam-se para o interior e formam novas vesículas. Estas são assim reutilizadas até
serem substituídas por novas vesículas formadas no soma e transportadas portransporte axonal até ao terminal do axónio.Casos há em que são libertados mais do que um neurotransmissor
simultaneamente, sendo o neurotransmissor adicionado chamado decotransmissor.
Uma vez libertado, o neurotransmissor difunde-se na fenda sináptica, indouma pequena parte dele ligar-se aos receptores específicos da membranaplasmática do neurónio pós-sináptico, receptores esses que podem eles própriosser canais iónicos ou podem estar associados a proteínas G que activam estescanais iónicos. Em qualquer dos casos, o resultado da ligação doneurotransmissor aos receptores é a abertura ou fecho de canais iónicosespecíficos na membrana plasmática do neurónio pós-sináptico, alterando assimo seu potencial.
Estas sinapses podem ser axodendriticas, axoaxonais ou axosomáticas.
24

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 25/374
Atraso Sináptico:atraso entre a chegada do potencial de acção ao terminal do axónio da célulapré-sináptica e a alteração do potencial da membrana da célula pós-sináptica(1ms) Este fenómeno é devido ao tempo de entrada de cálcio no terminal doaxónio e à fusão das vesículas sinápticas com a libertação do neurotransmissor.
Os canais iónicos da membrana da célula pós-sináptica regressam ao repousoquando o neurotransmissor deixa de estar ligado ao seu receptor específicopodendo transformar-se em substância ineficiente. Os neurotransmissores podemdifundir-se afastando-se dos seus receptores, ou podem ser transportadosactivamente para o axónio terminal (recaptação) ou para as células da glia +próximas.
Mecanismo de Actuação das Sinapses:
As sinapses químicas podem ser excitatórias ou inibitórias diferenciando-seassim pelos efeitos que o neurotransmissor provoca na célula pós-sináptica.
• Excitatória – da activação do receptor na membrana pós-sináptica resulta aabertura de canais de sódio, potássio, cálcio e outros pequenos iõescarregados positivamente, os quais ficam livres de se mover através damembrana de acordo com os gradientes eléctrico e químico, provocando
desta forma uma alteração do potencial de membrana – potencial pós-sináptico excitatório (EPSP). Este potencial não é mais do que um potencial
25

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 26/374
gradativo propaga-se decrementalmente para longe da sinapse paraaproximar o potencial da membrana do limiar de excitação. Durante estepotencial a excitabilidade do nervo a outros estímulos é aumentada. Oneurotransmissor deste tipo de sinapses é a Acetilcolina (Ach), e estassinapses podem sofrer somação e facilitação. Numa sinapse excitatória, aresposta pós-sináptica ao neurotransmissor é a despolarização, o que faz
com que o potencial de membrana se aproxime do limiar de excitação.• Inibitória – As alterações são sempre no sentido de causar hiperpolarização,
por potencial gradativo, designando-se estas alterações por potenciais pós-sinápticos inibitórios (IPSP). Durante este potencial a excitabilidade donervo a outros estímulos é francamente diminuído, e para gerar um IPSP osreceptores activos na membrana pós-sináptica provocam a abertura decanais de cloro e, algumas vezes, de canais de potássio. Nas células quetransportam activamente os iões cloro para fora, o potencial de equilíbriopara o cloro (-80mV) é mais negativo do que o potencial de repouso. À
medida que os canais de cloro se abrem, o cloro entra dentro da célula,produzindo assim uma hiperpolarização.
Cabe aqui ainda referir que o potencial de membrana na célula pós-sináptica éo resultado da somação temporal e espacial dos EPSP e IPSP em muitas sinapsesactivas excitatórias e inibitórias na célula.
Vesículassinápticas são de 3 tipos:
1. Pequenas e transparentes, contendo Ach, Gly, G ABA ou Glutamato;2. Pequenas e de core denso, contendo catecolaminas (naradrenalina eadrenalina);
3. Grandes e de core denso, contendo neuropéptidos (NPs – não sãonecessariamente neurotransmissores). Enquanto as pequenas vesículasse localizam junto da fenda sináptica e libertam o seu conteúdo na zonaactiva (zonas da membrana mais espessadas), as grandes vesículasestão ao longo dos terminais pré-sinápticos e libertam o seu conteúdopor exocitose ao longo de todo o terminal.
26

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 27/374
Neurotransmissores vs Neuromoduladores
Os neuromoduladores são mensageiros que produzem respostas complexasque não podem ser apenas descritas como EPSP ou IPSPs. A este tipo derespostas é dado o nome de modulação.
De facto, certos moduladores são sintetizados pelos neurónios pré-sinápticos e libertados conjuntamente com os neurotransmissores, para além deque determinadas hormonas, agentes parácrinos e certos mensageiros usadospelo sistema imunitário podem servir também como neuromoduladores, o quereflecte a complexidade do fenómeno.
Os neuromoduladores alteram frequentemente a reposta pós-sináptica adeterminados neurotransmissores, amplificando ou diminuindo a actividadesináptica, enquanto na célula pré-sináptica podem alterar a síntese, reabsorção emetabolismo de um determinado neurotransmissor.
De uma forma geral, os receptores para os neurotransmissores influenciamcanais iónicos que podem fazer parte do próprio receptor ou através dasproteínas G, actuando estes mecanismos em milissegundos. Os receptores paraos moduladores, por seu lado, provocam a maioria das vezes alteraçõesbioquímicas nos neurónios, usando para tal os mecanismos de transdução desinal via proteínas G acopladas a segundos mensageiros. Estas alterações quepodem demorar de alguns minutos a alguns dias, incluem modificações naactividade enzimática ou modificações na síntese de proteínas através dealterações na transcrição do ADN para ARN. Assim se compreende que os
neurotransmissores estão envolvidos nas comunicações rápidas, enquanto osneuromoduladores estão associados a fenómenos mais lentos como a
27

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 28/374
aprendizagem, o desenvolvimento, os estados motivacionais e até algumasactividades sensoriais ou motoras.
Eficácia Sináptica
A efectividade ou a força de uma sinapse pode ser influenciada tanto pormecanismos pré como pós-sinápticos.
Uma sinapse é afectada de forma pré-sináptica pela concentração decálcio, pelo número de receptores activados e pela natureza doneurotransmissor.
De forma pós-sináptica, as sinapses são condicionadas pelos receptoresexistentes para o neurotransmissor, o número em que os receptores estãopresentes (por haver up e down regulation) e também o tipo deneurotransmissor.
Para além disto a eficácia de uma sinapse também pode ser condicionadapor Drogas.
Os neurotransmissores/neuromoduladores mais conhecidos são:
• Acetilcolina (Ach) – é o principal neurotransmissor do sistema nervosoperiférico e no cérebro onde desempenha um papel importante nosprocessos de aprendizagem e memorização. Após ser libertada econsequentemente ter activado os receptores específicos nas células pós-sinápticas, a Ach é degradada pela enzima acetilcolinesterase que libertacolina; esta vai ser transportada activamente para os axónios-terminaispara ser usada novamente na síntese de Ach (colina+acetilenzima A) no
citoplasma dos terminais sinápticos e aramzenada em vesículas. Algunsdos receptores colinérgicos respondem à nicotina (receptor nicotínico, porcanal iónico) e à muscarina um veneno dos cogumelos (Amanita muscaria)
(receptor muscarinico), por proteínas G (Efeito da AcH no tecido pace-maker cardíaco- observa-se a dissociação das subunidades da proteína, oque abre o canal de potássio, a difusão deste para o exterior reduz afrequência cardíaca). O seu antagonista é o curare.
• Aminas Biogénicas: são sintetizadas a partir de aminoácidos. As
catecolaminas têm todas um anel catecol e um grupo amina, (dopamina,norepinefrina, epinefrina). Outros exemplos são a serotonina e histamina.As catecolaminas são metabolizadas pelos MAO (enzima monoaminooxidose) e pela catecol-o-metiltransferase (enzima COMT). Ascatecolaminas norepinefrina (ou noradrenalina) e epinefrina vão actuar no
SNP e SNC, sendo que para ambas podem existir receptores -adrenégicos
(formação de AMPc) ou -adrenégicos (activação dos 2.º mensegeiros da
fosfolipase C ou AMPc e activação directa dos canais de cálcio e potássio)(A ligação da noradrenalina ao seu receptor provoca a dissociação dasproteínas G, a ligação da subunidade alfa da proteína G à adenilciclase
28

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 29/374
activa o enzima dando origem à produção de AMPc o qual activa a proteína-kinase abrindo os canais e provocando os efeitos específicos dasubstância). A epinefrina (ou adrenalina) não é um neurotransmissorcomum no sistema nervoso, sendo no entanto a hormona que émaioritariamente segregada pela medula adrenal, uma glândula que éactivada pelo sistema nervoso periférico
Serotonina (S-hidroxitriptamina ou S-HPT): apesar de não ser umacatecolamina, é também uma importante amina biogénica, sendoproduzida a partir do triptofano (um aminoácido essencial) e metabolizadapela MAO. De uma forma geral, a serotonina desempenha um papelexcitatório nas vias que estão envolvidas no controlo muscular e um papelinibitório nas vias que regulam as sensações.
• Aminoácidos:o Excitatórios: glutamato e aspartato, são usados como
neurotransmissores pela grande maioria de sinapses excitatórias noSNC, funcionando os receptores destas substâncias como canais depotássio ou sódio, ou através da activação do sistema de segundomensageiro dependente da fosfolipase C.
o Inibitórios: GABA (gama-aminobutiríco - + abundante mediadorquímico inibidor no cérebro, formado pela descarboxilação doglutamato e libertado em mais de 20% das sinapses do SNC. Também presente na retina e é responsável pela inibição pré-sináptica. Os receptores são de 2 classes: Metabotrópicos ou GABAB
(activam proteínas G aumentando a condutância nos canais depotássio inibem a adenil-ciclase e diminuem o influxo de cálcio) ouIonotrópicos ou GABAA (compostos por subunidades que constituemos canais de cloro)) e Glicina (antagonizada pela estricnina pelo que oquadro da sua inibição é semelhante à da intoxicação por estricnina(convulsões, hiper-actividade muscular)), em que os receptoresfuncionam como canais de cloro ou abrem canais de sódio viaproteína G. (Nota: a glicina tem um efeito excitatório no cérebro (poracção sobre os receptores NMDA- N-metil-D-aspartato) é secretadapelos interneurónios na inibição directa na medula e tronco cerebral,
através do aumento da condutância de Cl-; os anestésicos geraisincluindo os gases voláteis, parecem apresentar um efeitosistemático nos canais GABAB e nos canais Cl- da glicina).Resumindo: Os seus receptores são inibitórios porque ou funcionameles próprios como canais de Cl ou abrem estes canais via proteínasG. O resultado final é uma hiperpolarização da membrana.
• Neuropeptidos: são formados por dois ou mais aminoácidos ligados entre sipor ligações peptídicas, sendo sintetizados no tecido nervoso. Estes
incluem opióides endógenos (endorfinas, dinorfinas e encefalinas) que sãopoderosos analgésicos e a Substância P.
29

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 30/374
• Gases: como o óxido nítrico (NO) que serve de mensageiro intercelularentre neurónios e entre neurónios e células efectoras, sendo transportadopor· difusão simples a partir das células que o sintetizaram. A formaçãodeste gás é catalizada pela NO sintetase a partir do aminoácido Larginina,aumentando a concentração do GMPc na célula receptora através da
activação da guanilciclase. Pensa-se que também o monóxido decarbono (CO) serve de neurotransmissor, desempenhando um papelimportante na aprendizagem por estimulação do aumento de GMPc dentrodos neurónios.
Estes dois últimos, presentes no cérebro e no trato GI apresentam elevadaafinidade à morfina. Incluem os derivados da Proencefalina, da Pro-
opimelanocortina ( -endorfina) e da Prodinorfina (dinorfinas e neoendorfinas)
e encefalinas.
Mecanismo de Libertação do Neurotransmissor
O processo que as vesículas utilizam para se movimentar até ao terminal paraque ocorra exocitose do neurotransmissor envolve um complexo conjunto deproteínas: proteína SNARE. Estas estão na membrana das vesículas, no terminalpré-sináptico e na membrana plasmática.
A iniciação da exocitose está relacionada com a presença de cálcio. Esta liga-se a uma proteína (sinaptogamina) que medeia a fusão das vesículas com a
membrana plasmática.Ocorre então a libertação do neurotransmissor na fenda sináptica que se vão
posteriormente ligar aos receptores do terminal pós-sináptico.Neurotransmissores que não estejam ligados a receptores são removidos da
fenda sináptica quando:
• São activamente transportados de volta ao terminal do axónio pré-sináptico(recaptura)
• Difundem do local dos receptores
• São enzimaticamente transformados por substâncias inactivas e são
transportadas de volta ao neurónio pré-sináptico para reutilização.
30

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 31/374
Fisiologia Neuronal e do Sistema Nervoso
Neurónio – unidade básica do sistema nervoso e funciona através de sinaiseléctricos que eles próprios geram e que depois se propagam ao longo dascélulas. É constituído por 4 partes:
• Corpo celular: contém a informação genética e as estruturas necessáriasà síntese proteica
• Dendrites: são projecções do corpo celular e que aumentam a área docorpo celular, logo a área der recepção de sinais e a capacidade decomunicação das células neurais
• Axónio: extensão singular do corpo celular onde os impulsos eléctricossão gerados e cuja parte mais próxima do corpo celular é designadasegmento inicial
• Terminais do axónio: responsáveis pela libertação de mensageirosquímicos
31

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 32/374
32

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 33/374
Tipos de Neurónios:
• Aferentes (pseudopolar) – veiculam a informação dos tecidos e órgãosdo corpo para o SNC. As terminações nervosas são formadas porreceptores que respondem a várias alterações físicas e químicas domeio que os envolve através da produção de sinais eléctricos. Osreceptores podem ser terminações especializadas dos neurónios ou
células separadas, intimamente associadas a estes. Não possuemdendrites e estão normalmente fora do SNC.
• Interneurónios – conectam os neurónios aferentes e eferentes com oSNC. Funcionam como integradores e modificadores de sinal,constituindo 99% do total de neurónios.
• Eferentes (multipolares ou bipolares) – transmitem sinais eléctricos doSNC para as células efectoras. Os corpos e dendrites destes neurónioslocalizam-se no SNC enquanto os axónios se estendem para o SNP,constituindo conjuntamente com a maioria dos axónios dos neurónios
aferentes, os nervos do SNP.
33

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 34/374
Células da Glia
• células de Schwann mielinizam os axónios periféricos;
• células satélite suportam os corpos celulares dos neurónios nos gânglios
do SNP;• oligodendrócitos mielinizam os axónios do SNC;
• microglia, defesa do SNC através da fagocitose;
• astrócitos, manutenção metabólica dos neurónios;
• células ependimárias revestem os ventrículos cerebrais e canal medular.
Sistema Nervoso
Visão Geral do Sistema Nervoso
Principais funções:1. Coordena as funções vitais2. Recebe e armazena dados3. Permite o contacto com o meio envolvente4. É a sede da actividade intelectual e emocional
Funções específicas do SN• Informação sensorial: Os receptores sensoriais monitorizam os estímulos
externos e internos como a temperatura, o tacto, o paladar, o olfacto, osom, a pressão arterial, o pH do meio interno e a posição das diferentespartes do corpo;
• Integração: O encéfalo e a espinal medula processam a informaçãosensorial e iniciam as respostas. A informação pode ter resposta imediata,ser armazenada ou ignorada;
• Homeostase: As actividades reguladoras e coordenadoras do sistemanervoso são decisivas na manutenção da hemostase (ex.: as célulascardíacas têm que se contrair a um ritmo que assegure o abastecimentoadequado de sangue);
• Actividade mental: O encéfalo é a sede de todas as actividades mentais,incluindo a consciência, o pensamento, a memória e as emoções;
• Controlo dos músculos e glândulas: Controla os principais movimentos docorpo pelo controlo da contracção do músculo esquelético; Intervém emconjunto com o sistema endócrino no controlo do músculo liso e cardíaco;Controla as secreções de muitas glândulas, como as sudoríperas, salivarese do tubo digestivo.
Glossário
Sistema Nervoso Central (SNC) - Encéfalo e Espinal Medula
34

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 35/374
Sistema Nervoso Periférico (SNP) - Nervos, Gânglios e Plexos Nervosos (fora doSNC)Neurónio de Associação (Interneurónio) - Neurónio multipolar localizado só no
SNCNeurónio Sensitivo (Neurónio Aferente) - Neurónio que transmite impulsos de umreceptor sensitivo para o SNCNeurónio Motor (Neurónio Eferente) - Neurónio que transmite impulsos do SNC
para um órgão efector (por ex. músculo)Nervo - Feixe de axónios, podendo conter fibras sensitivas e motorasNervo Motor Somático - Nervo que estimula a contracção de músculosesqueléticosNervo Motor Autónomo - Nervo que estimula ou inibe a contracção do músculoliso e cardíaco e que estimula a secreção glandularGânglio - Agrupamento de corpos celulares de neurónios localizados fora do SNCNúcleo - Agrupamento de corpos celulares de neurónios dentro do SNC Tracto (via nervosa) - Agrupamento de fibras nervosas que interconectam regiõesdo SNC
Classificação estrutural dos neurónios
Especialização funcional das diferentespartes do neurónio
35

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 36/374
Neuroglia do SN
36

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 37/374
Os astrocitos têm prolongamentos que terminam sobre os capilares e osneurónios, retirando glucose dos vasos e usando-a para produzir substratosenergéticos para estes. Estas células também retiram o neurotransmissorglutamato da sinapse e transformam-no em glutamina que é então reciclada paraos neurónios.
Sinapse Eléctrica
Nas sinapses eléctricas as membranas plasmáticas das células pré e pós-sinápticas estão unidas por junções gap, ( ou junções comunicantes ) as quaisconsistem num arranjo hexagonal de 6 conexinas ou conexónios (proteínas decanal), de cada uma das células, que formam um canal aquoso por onde passamos iões e pequenas moléculas (AMPc, etc.) que permitem a transmissão doimpulso eléctrico (PA) directamente de uma célula para a outra.
Este tipo de sinapse assegura comunicação bioeléctrica muito rápida e estámaioritariamente presente no músculo liso e cardíaco, centro respiratório dobulbo raquidiano, etc. (respostas rítmicas), onde permite a excitação rítmica degrandes massas musculares, mas são relativamente raras no sistema nervosodos mamíferos (massivamente presentes nas enguias eléctricas).
Sinapse Química
Um potencial de acção chega ao terminal pré-sináptico, provocando aabertura de canais de cálcio voltagem dependentes que permitem a passagemdeste ião para o interior do neurónio;
Os iões cálcio promovem a fusão das vesículas com a membrana pré-sinápticas com auxílio das proteínas SNARE/Sinaptotagmina e a libertação dosneurotransmissores (Ach) para a fenda sináptica;
Os neurotransmissores difundem na fenda sináptica e ligam-se aos seusreceptores na membrana da célula pós-sináptica, aumentando a permeabilidadedos canais iónicos aí presentes.
O aumento da permeabilidade aos iões origina a despolarização (ou
hiperpolarização) da membrana da célula pós-sináptica, daí resultando umpotencial de acção pós-sináptico se for ultrapassado o limiar de excitação.
37

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 38/374
Sistema Nervoso• Sistema Nervoso Central• Sistema Nervoso Periférico
o Divisão Aferente Sensitivos Somáticos Sensitivos Viscerais Sensitivos Especiais
o Divisão Eferente Sistema Nervoso Somático Sistema Nervoso Autónomo
• Simpático• Parassimpático• Entérico
Sistema Nervoso Central•
Encéfaloo Anterior – Córtex cerebral (dividido em lóbulos) e Diencéfaloo Posterior – Cerebelo e Tronco Cerebral
• Medula Espinalo Substância cinzenta: corpos celulares dos neurónios eferentes e
interneurónioso Substância branca: axónios ascendentes e descendentes
Funções dos Lóbulos Cerebrais:
• Frontal: controlo voluntário dos músculos esqueléticos e funçõesintelectuais (concentração, planeamento, expressão verbal);
• Parietal: área somatosensorial, compreensão e formulação das palavras;
• Temporal: sensações auditivas (armazenamento de experiências visuais eauditivas);
• Occipital: córtex visual, correlação das sensações visuais com outrosestímulos sensoriais.
Área de Wernica (WA) (parietal/temporal): área de interpretação geral da
linguagemÁrea de Broca (temporal): área motora do discurso
O encéfalo e a medula espinal estão protegidos pelos ossos do crânio e dacoluna vertebral, pelas meninges e pelo líquido cefalorraquidiano (LCR)Funções do Diencéfalo e Encéfalo Posterior:
• Tálamo: integração sensorial e motora; actua como estação intermédia deretransmissão dos impulsos sensitivos para o córtex;
• Hipotálamo: regulação das funções vitais (metabolismo da água, sede,apetite e temperatura); íntimo contacto e regulação da hipófise (sistema
hormonal);
38

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 39/374
• Tronco Cerebral (formação reticular): controlo do grau de actividade doSNC; centros de integração (respiratório, cardiovascular, vómito edeglutição);
• Cerebelo: postura, equilíbrio e aprendizagem motora
Cérebro: (hemisférios cerebrais) contém o córtex cerebral, que participa na
percepção, na geração de movimentos finos e delicados, razão, aprendizagem ememória; contém os núcleos subcorticais, incluindo os que participam nacoordenação da actividade do músculo esquelético; contém as vias nervosas deinterconexão (comissuras/corpo caloso);
Tronco Cerebral: contém todas as fibras nervosas que ligam a espinal medula,o encéfalo anterior e o cerebelo; contém a formação reticular e os seus várioscentros integradores, incluindo o cardiovascular, respiratório, vómito edeglutição; contém núcleos para os nervos cranianos do III ao XII.
Cerebelo: coordena os movimentos, incluindo os da postura e equilíbrio;
participa em algumas formas de aprendizagem, incluindo a de tarefas motoras.
Tálamo: funciona como estação de re-transmissão sináptica para as viassensitivas no seu percurso para o córtex cerebral; participa no controle ecoordenação do músculo esquelético; joga um papel importante na atenção evigilância.
Hipotálamo: regula a hipófise anterior, o equilíbrio hídrico, o sistemareprodutor, a temperatura corporal, o comportamento alimentar; desencadeia eregula os ritmos circadianos; participa na regulação do Sistema Nervoso
Autónomo e no desencadeamento de emoções e de comportamentos emocionais.Sistema Límbico
Trata-se de um sistema funcional que consiste na interconexão de váriasestruturas do encéfalo, incluindo porções do lobo frontal, temporal, tálamo ehipotálamo, assim como respectivas vias de interconexão. As estruturas dosistema límbico estão associadas com a aprendizagem, memória, experiências ecomportamentos emocionais e a uma grande variedade de funções viscerais eendócrinas. O hipotálamo coordena muito do output do sistema límbico referenteao comportamento e respostas endócrinas.
Estruturas principais:
- Amígdala- Núcleos olfatórios- Hipocampus
Funções principais:
- Emoções
- Comportamento emocional- Aprendizagem e memória- Inteligência Emocional??
39

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 40/374
Medula Espinal
A medula espinal (ME) é de extrema importância para o funcionamentoglobal do sistema nervoso. Constitui o elo de ligação entre o encéfalo e o sistemanervoso periférico, integrando a informação que recebe e produzindo respostasatravés de mecanismos reflexos.
Substância cinzenta central organizada nos cornos dorsais, ventrais elaterais (SNA), sendo constituída por corpos celulares neuronais, dendrites eaxónios não mielinizados.
Substância branca periférica, constituída por axónios mielinizados eorganizada nos cordões ventral, lateral e posterior, os quais estão subdivididosem feixes nervosos ou vias ascendentes ou descendentes que transportam omesmo tipo de informação para e do encéfalo.
Arco-reflexo
O AR é a unidade funcional básica do SN,
contendo, na sua expressão mais simples, capazde receber um estímulo e produzir uma resposta:um receptor sensorial, um neurónio aferente ousensitivo, um neurónio de associação ouinterneurónio, um neurónio eferente ou motor eum órgão efector. A resposta produzida chama-sereflexo, tratando-se de uma resposta automática aum estímulo sem que ocorra pensamentoconsciente. Muitos dos reflexos são integrados naME e outros no encéfalo. Os reflexos que envolvem
neurónios excitatórios resultam numa resposta(ex. contracção de um músculo); os que envolvemneurónicos inibitórios resultam na inibição de umaresposta (relaxamento de um músculo). Os centrosencefálicos superiores influenciam os reflexossuprimindo-os ou exagerando-os. Os principaisreflexos medulares são o reflexo de extensão, o reflexo dos órgãos tendinosos deGolgi, o reflexo de retirada e o reflexo extensor contralateral.
As Vias Neuronais Ascendentes
As vias neuronais ascendentes transportam informação sensitiva de e paradiferentes níveis do SNC, sendo constituídas por neurónios de 1.ª, 2.ª e 3.ªordem. A sua designação conjuga o local de partida e de chegada.
Assinale-se que a maior parte das vias apresentam cruzamento oudecussão contralateral, pelo a informação sensitiva dos estímulos à direita éprocessada no hemisfério esquerdo e vice-versa
40

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 41/374
Vias neuronais descendentes corticoespinais ou piramidais
Vias motoras descendentes corticoespinais ou piramidais – passam pelaspirâmides do bulbo; têm poucas ou nenhuma sinapse intermédia; conduçãorápida de informação motora
Assinale-se que a maior parte das vias apresentam cruzamento ou
decussão contralateral
41

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 42/374
Vias neuronais descendentes extrapiramidais
Vias multisinápticas que transportam informação envolvida no controlomotor neural superior dos músculos esqueléticos.
As vias piramidais (corticoespinais) são mostradas a cor-de-rosa e asextrapiramidais a preto.
42

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 43/374
Fornecimento de sangue ao encéfalo, barreira hemato-encefálica e LCR
- A suspensão do fornecimento de glucose ao encéfalo (represena 2% do peso docorpo, mas recebe 15% do sangue), que é praticamente o único substratoenergético para formar ATP por fosforilação oxidativa, ou de O2 por mais do que,respectivamente, 15 ou 5 mn produz lesões irreversíveis;- As tight junctions entre as células endoteliais dos capilares do encéfalo e certos
mecanismos de transporte específico constituem a barreira hemato-encefálica,que controla o tipo e velocidade de transferências a partir do sangue para otecido nervoso e vice-versa; ( ex. da heroína (CH3) versus morfina (OH), L-dopaversus dopamina, barbitúricos, nicotina, álcool e cafeína);- O LCR (soro sem proteínas), amortece os choques do tecido nervoso com ocrânio e coluna vertebral; é segregado pelos plexos coróides para o interior dosventrículos, passa ao tronco cerebral e canal raquidiano, circula no espaçosubaracnoideu e sai para o sangue pelos vilos aracnóides da superfície superiordo encéfalo.A obstrução do seu fluxo origina hidrocefalia, lesões e retardamento mental nascrianças, assim como lesões encefálicas nos adultos por aumento da pressãointracraniana.
Sistema Nervoso Periférico
As divisões aferente e eferente do SNP têm esta denominação porqueenquanto a divisão aferente é composta por neurónios que conduzeminformações a partir dos receptores sensoriais até ao sistema nervoso central(neurónios aferentes), a divisão eferente é constituída por neurónios quetransportam a informação a partir do SNC para os músculos e glândulas.
Dentro da divisão eferente encontramos o sistema nervoso autónomo e osistema nervoso somático, cujas diferenças residem não só nos órgãos enervadosmas nos tipos de neurónios.
Sistema Nervoso Autónomo
O SNA é um sistema motor relacionado com aregulação do músculo liso, do músculo cardíaco e dasglândulas. Não sendo directamente acessível aocontrole voluntário baseia-se em reflexos autónomos(resposta a um estímulo sem intervenção do SistemaNervoso Central (SNC) e no controle central, tendocomo uma das principais funções a homeostasia e a
regulação do tonos muscular liso.O SNA é assim, por definição, um sistema de
controlo da actividade fisiológica, essencial, em43

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 44/374
conjunto com o Sistema Endócrino, para a manutenção da homeostasia. Oambiente interno do organismo tem que ser constantemente monitorizado, sendoos sinais resultantes dessa monitorização transmitidos a um centro controlador.Neste é comparada a diferença entre o estado ideal e o estado real, sendo amagnitude da resposta condicionada pela discrepância entre os dois estados.
Observa-se assim que, se o aumento deuma característica não estiver devidamente
regulada e se a velocidade com que ofeedback aferente é recebido pelo centro decontrolo não se encontrar ajustado, o sistemacomeça a oscilar, dando origem a umaresposta (eferente) adequada a cadasituação.
O Sistema Nervoso Autónomo divide-seno Sistema Nervoso Simpático, SistemaNervoso Parassimpático e Sistema NervosoEntérico. É nos tecidos alvo (a generalidadedos órgãos, com excepção do músculoesquelético) que as divisões Simpática e Parassimpática actuam, querindependentemente, quer através do Sistema Nervoso Entérico.
É constituído por 2 neurónios numa séria que liga o SNC e as célulasefectoras, sendo que o 1.º neurónio tem o corpo celular no SNC (medula espinalou tronco cerebral). A sinapse entre os 2 neurónios é fora do SNC, no gânglioautónomo, e os neurónios pós-ganglionares têm aqui localizados os seus corposcelulares.
A via final do Sistema Nervoso Simpático e Parassimpático é formada por
uma sequência de dois tipos de neurónios:• Neurónios pré-ganglionares, cujos corpos celulares estão localizados na medulaespinal ou no tronco cerebral• Neurónios pós-ganglionares, cujos corpos celulares estão localizados nosgânglios autónomos periféricos.
A esta divisão do SNP eferente são atribuídas as funções de regulação dahomeostasia e do tónus muscular liso, funcionando como sistema de controlo daactividade fisiológica.
Sistema Nervoso Parassimpático:
O Sistema Nervoso Parassimpático possui uma distribuição menos dispersaque o sistema simpático. Os neurónios pré-ganglionares parassimpáticosencontram-se distribuídos pelas divisões cranial e sagrada do Sistema NervosoCentral.
Os corpos celulares dos neurónios pré-ganglionares parassimpáticos dadivisão cranial distribuem-se pelo tronco cerebral (ponte de varóleo e medulaalongada), emergindo destas áreas quatro nervos (pares) cranianos:
• Nervo Oculomotor (III par craniano)o Fibras pré-ganglionares com origem craniana
o Gânglio terminal ciliaro O olho (corpo ciliar e músculo liso da íris) é o órgão efector
44

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 45/374
• Nervo facial (VII par craniano)o Fibras pré-ganglionares com origem cranianao Gânglios terminais submandibular e pterigopalatinoo Glândulas salivares, lacrimais e mucosas são os órgãos efectores
• Nervo Glossofaringeo (IX par craniano)o Fibras pré-ganglionares de origem cranianao Gânglio óptico terminalo Glândula parótida é o órgão efector
• Nervo dorsal motor do vago (X par craniano)o Fibras pré-ganglionares de origem cranianao Gânglio terminal perto ou no órgãoo Órgãos efectores: coração, pulmões, trato gastrointestinal, fígado e
pâncreas
A divisão sagrada do sistema parassimpático tem os corpos celulares pré-ganglionares na matéria cinzenta da medula sagrada (entre S2, S3 e S4),"viajando" estas fibras através dos nervos pélvicos de modo a formarem sinapsesnos gânglios pélvicos.
• Nervos pélvicos espinhaso Fibras pré-ganglionares com origem sacral (S2 a S4)o Gânglios terminais perto dos órgãoso Órgãos efectores: metade inferior do intestino grosso, recto, bexiga e
órgãos reprodutores.
Os gânglios parassimpáticos encontram-se geralmente mais afastados doSistema Nervoso Central do que os gânglios do sistema simpático, localizando-se,nalguns casos, mesmo dentro dos órgãos enervados (gânglios intramurais).
Sistema Nervoso Simpático:
O Sistema Nervoso Simpático é um sistema com ampla distribuição. Inervaos órgãos contidos nas cavidades corporais, a pele e os músculos (lisos). Oscorpos celulares dos neurónios motores pré-ganglionares do sistema simpático
localizam-se ao nível dos segmentos lombares e torácicos da medula espinal(entre T1 e L2), concentrando-se nas zonas de matéria cinzenta denominadascolunas intermédio-lateral e intermédio-medial. Os axónios motores dosneurónios pré-ganglionares simpáticos deixam a medula espinal em associaçãocom os axónios motores somáticos, divergindo então dos nervos espinais comoramos comunicantes brancos para uma de três seguintes localizações:1. Estabelecem sinapses nesse gânglio da cadeia ganglionar simpáticaparavertebral;2. Seguem a direcção rostral (ascendente) ou caudal (descendente) pela cadeiasimpática, de modo a estabelecerem sinapses num gânglio paravertebral, noutronível;
3. Prosseguem através de um nervo esplénico de modo a estabelecerem sinapsesem gânglios pré-vertebrais (ou colaterais).
45

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 46/374
Tipo de Nervos
• Nervos Raquidianoso Axónios pós-ganglionares não são mielinizadoso Axónios seguem pelo nervo parar os órgãos que enervam: glândulas
sudoríperas, músculo liso de vasos sanguíneos esqueléticos ecutâneos e músculo liso erector do pêlo
• Nervos Simpáticoso Na região cervical os nervos formam um feixe em torno da carótida,
seguindo a partir daí o seu percurso, enervando glândulassudoríperas, salivares e músculo liso dos vasos sanguíneos, olho eerector do pêlo
o Enervam também órgãos torácicos (coração, esófago e pulmões)
• Nervos Esplâncnicoso Axónios pré-ganglionares constituem nervos esplénicos para gânglios
pré-viscerais onde sinapsamo OS órgãos alvo são: órgãos abdominopélvicos, pâncreas, fígado e
próstata
Tipos de neurónios simpáticos:
• Fásicos – alta frequência de disparo, predominantes nos gânglios para-vertebrais lombares;
• Tónicos – descargas lentas e rítmicas nos gânglios pélvicos e mesentérioinferior;
• LAH – descarregam normalmente, nos gânglios mesentérico superior e
celíaco.
Nota: Os axónios dos neurónios pré-ganglionares simpáticos têm pequenodiâmetro e são mielinizados.
Caso especial: Inervação da Glândula Supra-Renal
A Inervação do nervo esplâncnico para as supra-renais baseia-se emneurónios pré-ganglionares.
A estimulação das células da medula supre-renal pelos axónios provoca alibertação de adrenalina e noradrenalina para o sistema circulatório.
46

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 47/374
Distribuição dos nervos cranianos eespinais:
O SNP tem 43 pares de nervos:• 12 cranianos• 31 espinais:
o 8 cervicaiso 12 torácicoso 5 lombareso 5 sacraiso 1cocgígio
47

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 48/374
Divisão
Simpática (Toraco-Lombar)
Divisão Parassimpática (crânio-sacral)
48

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 49/374
49

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 50/374
Situação de stress/descarga massiva do simpático (fight-or-flight )
50

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 51/374
Relação do sistema nervoso simpático com o parassimpático
Activação unilateral: a activação do SNA considera-se unilateral quando oórgão inervado recebe apenas o SNS, como, por exemplo, a maior parte daárvore vascular. Assim, na inervação simpática da árvore vascular existe umaactividade neurológica progressiva denominada tonos autonómico.Aumentando ou reduzindo este tonos, pode efectivamente estimular-se ou inibiro alvo inervado. Existem, no entanto, algumas diferenças entre a inervação da
pele e dos órgãos viscerais e músculos. Como resposta a um estímulo térmico(ex: frio) a vasculatura periférica irá provocar vasoconstrição de modo aconservar o calor. Seria pouco lógico que essa vasoconstrição provocassetambém uma vasoconstrição sistémica.
Activação dual: a activação do SNA considera-se dual quando o órgão inervadorecebe inervação simultânea da divisão simpática e paras simpática, como é ocaso de muitos órgãos viscerais. Neste caso, os efeitos destas duas divisões dosSNA podem ser:1. Activação antagonista - Quando o Sistema Nervoso Simpático é activadoprovoca a inactivação do Sistema Nervoso Parassimpático e vice-versa. A nívelterminal a norepinefrina e a acetilcolina podem inibir a libertação doneurotransmissor oposto. Este tipo de efeitos verifica-se, por exemplo, na região
51

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 52/374
pacemaker do coração (inervação da mesma célula), sendo o simpático activador(receptor b1) e o parassimpático (receptor muscarínico) o inibidor; no tratodigestivo, sendo o passimpático activador e o simpático inibidor e no controlo daabertura pupilar, em que a estimulação simpática provoca é activadora (músculodilatador da íris) e a parassimpática inibidora (músculo constritor da íris). Músculocircular da íris com enervação parassimpática, receptores muscarínicos (miose) emúsculo radial com enervação simpática e receptores a1 (midriase)
2. Activação complementar - Embora os efeitos duais do simpático eparassimpático sejam habitualmente antagonistas, em alguns casos eles podemser complementares ou cooperativos. São complementares quando a estimulaçãosimpática e parassimpática produz efeitos similares, como por exemplo nasglândulas salivares, em que a secreção de saliva aquosa é estimulada peloparassimpático, enquanto o simpático estimula a constrição dos vasossanguíneos no trato digestivo. O consequente decréscimo do fluxo de sangue nasglândulas salivares conduz à produção de saliva espessa e mais viscosa.3. Activação cooperativa - Considera-se que existe este tipo de activaçãoquando o simpático e o parassimpático cooperam no sentido do bomdesempenho da função. Os efeitos da estimulação simpática e parassimpátca dossistemas reprodutivo e urinário são cooperativos. A erecção do pénis, porexemplo, é devida à vasodilatação resultante da estimulação dos nervosparassimpáticos; a ejaculação é devida a estimulação dos nervos simpáticos. Asduas divisões do SNA cooperam para o bom desempenho da função da funçãosexual masculina. De modo idêntico, a erecção clitoriana e as secreções vaginaissão estimuladas pelos nervos parassimpáticos, enquanto o orgasmo é umaresposta simpática, tal como no homem.
Sistema Nervoso Entérico
É constituído por neurónios e plexos ganglionares situados no tractogastrointestinal, desde o esófago até ao ânus, vesícula biliar e pâncreas,
52

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 53/374
participando no controle reflexo da motilidade (reflexo mientérico), secreção(submucoso) e fluxo sanguíneos do intestino. Rede de plexos nervosos na parededo tubo digestivo. Estes plexos têm contributos de: (1) neurónios sensitivos queligam o tubo digestivo ao SNC; (2) neurónios motores do SNA que ligam o SNC aotubo digestivo e (3) neurónios entéricos que são confinados nos plexos entéricos.Participa no controlo da motilidade, secreção e fluxo sanguíneo do intestino.
Descobriu-se recentemente que algumas células imunitárias, entre elas osmastócitos, podem ser directamente inervadas por nervos entéricos. Contémneurónios adrenérgicos e colinérgicos.
Embora possa funcionar independentemente, é regulada através de umainervação simpática e parassimpática extrínseca. O SNC monitoriza o o tubodigestivo através dos neurónios sensitivos e controla os seus músculos eglândulas através dos neurónios motores do SNA. Os neurónios entéricossensitivos detectam alterações na composição do conteúdo dos intestinos ou doseu estiramento; os neurónios motores estimulam ou inibem a contração domúsculo liso e da secreção glandular; os interneurónios ligam os dois anteriores.Embora possa funcionar de forma independente é regulado pelo SNS e SNP que
através de sinapses nos plexos que o relacionam com o SNC.Existem 3 tipos de reflexos importantes para o controle gastrointestinal que
são suportados através de uma organização hierárquica do Sistema NervosoEntérico:• Os reflexos que ocorrem apenas dentro dos plexos da parede intestinal e quesão responsáveis pelo controle da secreção, peristaltismo e contracções mistas• Um segundo grupos de reflexos envolvem os gânglios pré-vertebrais, sendoresponsáveis pela comunicação entre diferentes áreas do intestino (reflexosgastrocólico e enterogástrico)• O terceiro nível hierárquico envolvendo reflexos com extensas relações entre a
medula e o tronco cerebral (ansa longa: envolvem a medula espinal, o troncocerebral e o córtex – comando da dor e defecação).
Os Aferentes AutónomosSabe-se que quase todos os órgãos que recebem uma inervação autónoma
eferente apresentam também uma inervação aferente.Quanto às funções das fibras autónomas aferentes:
1 - São sensores do meio interno2 - Conduzem estímulos nervosos quer perceptíveis quer imperceptíveis.
Estas fibras aferentes têm os seus corpos celulares nos gânglios das raízes
dorsais e apresentam. Muitas características em comum. Esta classe de células,conhecidas como -aferentes, são muitas vezes ricas em péptidos, são
sensíveis à neurotoxina capsina e são também sensíveis ao factor decrescimento do nervo (NGF) durante a fase de desenvolvimento. Enquanto asfibras aferentes autónomas, fundamentalmente viscerais, são em menor númeroque as aferentes somáticas, inervam uma área substancialmente maior. Devidoao escasso número de células aferentes, existe uma grande divergência noimpulso aferente central que contribui para a fraca localização das sensaçõesviscerais.
Transmissão e integração ganglionar
53

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 54/374
Os gânglios autónomos representam locais capazes de processar umaquantidade considerável de informação. Esta capacidade não se deve acomplexas inter-relações anatómicas ou à presença de múltiplas populações deinterneurónios. Os neurónios dos gânglios autónomos são unipolares, nãoapresentando árvore dendrítica, formando-se as sinapses directamente no corpocelular, aumentando o processamento de informação tornando estes gânglios na
preparação ideal para o estudo da Electrofisiologia. Estes estudos revelam que,em muitas espécies, a transferência de informação desde a entrada nas fibraspré-ganglionares até à saída pelos neurónios pós-ganglionares depende dainteracção de transmissores numa diversa população de receptores existentes nacélula pós-ganglionar.
A estimulação pré-sináptica desencadeia uma complexa resposta pós-sináptica mediada pela acetilcolina e por péptidos.
A acetilcolina é mediada por 3 tipos de reposta sináptica:
• Resposta nicotínica: PPSE (potencial pós-sináptico excitatório) rápido,
responsável pela transmissão através do gânglio com a duração de 30 a 40ms;
• Resposta muscarínica: PPSE lento, demorando cerca de 1 minuto;
• Resposta muscarínica inibitória: PPSI (potencial pós-sináptico inibitório) coma duração de 1 a 2 s.
Nota: A resposta PPSE rápida parece ser a responsável pela transmissão atravésdo gânglio, uma vez que se utilizarmos drogas que bloqueiam selectivamente osreceptores nicotínicos e acetilcolinérgicos, a transmissão ganglionar é totalmentebloqueada. Os potenciais lentos (PPSE lentos e os PPSI), medeiam uma
estimulação contínua, quer aumentando, quer diminuindo a probabilidade comque os neurónios continuam a gerar potenciais de acção após um PPSE rápido.Uma modulação adicional da transmissão ganglionar ocorre através da libertaçãode péptidos, normalmente como resposta a um forte estímulo aferente. Umexemplo deste tipo de mediação é o facto de um péptido semelhante à LHRH(Hormona hipotalâmica libertadora das gonadotrofinas hipofisárias LH e FSH) tersido identificado como transmissor, mediando um PPSE peptidérgico de váriosminutos. Este péptido tem também a característica de se difundir através dogânglio, indo despolarizar uma vasta população de neurónios.
Receptores para a acetilcolina e antagonistas:• Receptor nicotínico: Antagonista curare/Hexametónio
o No músculo esquelético provoca despolarização, produzindopotenciais de acção e contracção muscular por abertura dos canaiscatiónicos no receptor.
• Receptor muscarínico: Antagonista atropina/ Bustapolaminao Músculo liso e glândulas (M3 e M4) provoca despolarização e
contracção do músculo liso e secreção de glândulas por activação deproteína G ligada ao receptor, abrindo canais cálcio. No coração (M2)
provoca hiperpolarização, diminuindo a taxa de despolarizaçãoespontânea – Ach activa o receptor. Ligado à proteína G abrindocanais potássio.
54

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 55/374
Tipos de Activação:
• Concorrente: quando é necessária activação simultânea de simpático eparassimpático para que um órgão determine a sua função. Por exemplo,glândula salivar em que: Simpático – (norepinefrina) amilase salivar;parassimpático – (acetilcolina) muco.
• Unilateral: quando não existe dupla inervação, podendo o tónus (actividadeprogressiva neurológica do simpático) ser aumentado para excitação oudiminuindo para inibição do órgão.
O Controlo Autónomo CentralO conceito de homeostasia necessita, não apenas de um conjunto de inputs
e outputs aferentes, mas também de um centro controlador. Algumas áreas doSistema Nervoso Central interagem de modo a fornecerem os mecanismos decontrolo que regem os reflexos autónomos.
O hipotálamo desempenha aqui um papel fundamental como integrador econtrolador, não sendo apenas uma estrutura de passagem de respostas.
Junção neuroefectoraDurante muitos anos pensou-se que a transmissão entre as fibras
autónomas e os seus órgãos alvo ocorria através da libertação de transmissoresque se difundiam pelo espaço extracelular indo interagir com receptorescolocados em zonas distantes da zona de libertação. Actualmente, aceita-se quea transmissão autónoma está intimamente associada à transmissão central, ondeos transmissores activam grupos de receptores restritos localizados na área delibertação.
Os neurónios autónomos podem ser classificados em grupos funcionaisatravés das suas características biofísicas. Existem 3 subclasses de neuróniossimpáticos definidas com base nos tipos de voltagem e nos canais de potássiodependentes do cálcio que são expressos na membrana do corpo celular doneurónio. Estes tipos definem a maneira como o potencial de acção é iniciadodurante a despolarização.1) Neurónios fásicos, que descarregam a uma alta frequência de disparo no iníciode uma despolarização. São predominantes nos gânglios paravertebraislombares.2) Neurónios tónicos, que descarregam lenta e ritmicamente enquanto semantém a despolarização. São predominantes nos gânglios pélvicos emesentérico inferior.
3) Neurónios LAH (long after-hyperpolarization), que descarregam normalmenteuma vez no início da despolarização, pois a excitabilidade celular após umpotencial de acção está limitada por um aumento prolongado da condutância dopotássio. São predominantes no gânglio mesentérico superior e celíaco.
Co-transmissãoÉ normalmente aceite que os neurónios sintetizam e segregam apenas um
tipo de transmissores. Este facto é sugerido pela evidência de que uma vezconhecido o transmissor podemos sempre bloquear a transmissão utilizando o
antagonista apropriado. No entanto, é frequente existirem células capazes desintetizarem três ou mais transmissores. Devido às suas reduzidas concentrações
55

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 56/374
e ao facto de se encontrarem sempre em quantidades muito inferiores a umaoutra substância, estes péptidos são de difícil detecção.
Estes péptidos são extremamente potentes e mesmo em pequenasconcentrações tem-se demonstrado que podem actuar como moduladores daacção do 1.º transmissor, ou seja, o co-transmissor interage no local pós-sinápticode modo a amplificar ou reduzir a acção do 1.0 transmissor. Temos comoexemplo no gânglio simpático um potencial pós-sináptico excitatório lento
mediado por um péptido semelhante à LHRH que estabelece uma despolarizaçãolonga que aumenta a probabilidade da célula disparar potenciais de acção comoresposta a um PPSE nicotínico rápido.
Regulação da Temperatura Corporal
Na maior parte dos mamíferos atemperatura corporal é regulada para os 37°C,sendo protegida contra aumentos e diminuiçõessignificativas.
O mecanismo responsável pelamanutenção deste set-point não é aindaconhecido, sabendo-se porém que a temperaturapode apresentar ligeiras alterações quer a curtoquer a longo prazo.• Existe um ritmo circadiano que mantém atemperatura do organismo cerca de 0,5 - O,7°Cmais elevada durante o dia (fase activa do ciclo),sendo este um exemplo de uma alteraçãofisiológica a curto prazo• As mulheres além desta variação diária
apresentam também uma variação mensal quecoincide com o ciclo menstrual, variação essaque é maior cerca de 0,5 °C durante a ovulação.
Uma das alterações mais importantes datemperatura corporal é o aparecimento de febre,quando, como resposta a um estímulopirogénico, a temperatura aumenta 1 a 2°C. Apósterminar o estímulo pirogénico, ou como respostaa um antipirético, o setpoint volta ao seu valornormal e os mecanismos de arrefecimentocorporal são reajustados de modo a
restabelecerem a temperatura normal.A manutenção da temperatura corporal
assenta num sistema de terminações nervosassensíveis à temperatura na pele e em neuróniosdo sistema nervoso central e dos órgãos visceraissensíveis à temperatura, que informam ohipotálamo sobre a temperatura existente nesta zona.
O hipotálamo integra esta informação com a sua própria informação localfornecendo uma resposta a diferentes mecanismos de modo a estabelecer atemperatura. Assim, a regulação faz-se principalmente através da vasoconstrição
periférica para conservação de calor, e na vasodilatação para a sua dissipação.Nos seres humanos os principais mecanismos de resposta que mantém a
temperatura em níveis confortáveis são mecanismos comportamentais. Nestas56

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 57/374
condições a regulação da temperatura faz-se principalmente através davasoconstrição periférica para conservação de calor e vasodilatação paradissipação de calor.
Como resposta a um estímulo térmico forte (frio) os mecanismos deprodução de calor são também activados sendo de dois tipos:1 - Produção de calor através da contracção involuntária do músculo esquelético(arrepio)
1 - Produção de calor através de uma resposta metabólica controlada pelosistema simpático, que ocorre pela activação adrenérgica do tecido adiposocastanho (caracterizado pelo elevado conteúdo em mitocôndrias).
Controle do olhoO Sistema Nervoso Autónomo desempenha um papel importante na função
normal do olho. Inerva os músculos lisos da Íris e do Cristalino de modo aoptimizar o diâmetro da pupila e a acomodação. Através da regulação do fluxosanguíneo do olho controla também o metabolismo e pode alterar a pressãoocular.
Os reflexos pupilares à luz envolvem a constrição e a dilatação do músculoda íris de modo a regular a quantidade de luz que atinge a retina. Os músculospupilares constritor e dilatador são controlados pelas divisões simpática eparassimpática do Sistema Nervoso Autónomo. O músculo dilatador é controladopelo sistema simpático, através de fibras pré-ganglionares que emergem dossegmentos medulares das colunas simpáticas paravertebrais.
Estas fibras saem da raiz ventral e são projectadas para o gânglio superiorcervical que produz as fibras pós-ganglionares simpáticas, as quais ascendemadjacentes à carótida, juntando-se ao ramo oftálmico do nervo facial que enervaa órbita. Devido a este circuito sinuoso, uma lesão na medula cervical ou torácicasuperior leva à perda do controlo do olho, produzindo assim o síndroma de
Horner.A inervação parassimpática colinérgica do esfíncter muscular da íris éregulada pela via pré-ganglionar através dos neurónios motores oculares que sesituam no núcleo de Edinger-Westphal. Os axónios destes neurónios projectam-senos gânglios ciliares que formam a inervação pós-ganglionar parassimpática domúsculo dilatador. O circuito que envolve o reflexo pupilar à luz apresenta umasequência na retina que consiste nos fotoreceptores, células bipolares e célulasgangliónicas.
As células gangliónicas fornecem as fibras do tracto óptico que seprojectam no protectum do cérebro médio que, por sua vez, proporciona osimpulsos para o núcleo de Edinger-Westphal. O circuito é finalizado por uma
transmissão pelas fibras pré e pós-ganglionares simpáticas. A incidência de luz naretina utiliza uma via de seis neurónios de modo a causar a constrição da pupila.
A acomodação é um termo que descreve as alterações que ocorrem no olhode modo a focar uma imagem visual na retina. Requer alterações na contracturado músculo ciliar de modo a alterar a curvatura do cristalino, a constrição pupilarpelos músculos do esfíncter da íris e a modificação da posição dos olhos pela viasimpática através da inervação dos neurónios motores somáticos dos músculosmediais rectos.
Os neurónios oculares parassimpáticos estão organizados de modosemelhante aos que enervam o reflexo pupilar a um estímulo luminoso. Porém, avia aferente, que controla estes neurónios parece envolver o córtex visual ealgumas partes da região média temporal. Esta última recebe impulsos do córtex
57

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 58/374
visual primário, influenciando os neurónios motores oculares através de viasmultissinápticas.
Fisiologia Muscular58

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 59/374
Características funcionais do sistema muscular:
• Contractilidade: capacidade de geração de força;
• Excitabilidade: resposta do músculo à estimulação do SN e SistemaEndócrino, que regulam a sua actividade;
• Extensibilidade: podem distender para além do seu comprimento de
repouso;• Elasticidade: após distensão têm capacidade para regressar ao seu
comprimento de repouso.
A manutenção da homeostasia do organismo necessita de um tecido capazde transformar a energia química em energia mecânica. Temos três tiposdiferentes de músculo, cada um com propriedades e funções distintas:
• Esquelético: responsável pela locomoção, expressão facial, postura,
movimentos respiratórios e outros; a sua contracção permite amovimentação de um local do organismo em relação a outro local, comopor exemplo, na flexão do antebraço. A contracção coordenada damusculatura esquelética permite a movimentação do organismo no meioambiente; as suas funções dependem do controlo voluntário ou conscientepelo Sistema Nervoso Central.
• Liso: encontra-se nas paredes dos órgãos ocos e tubos, músculos internosdo olho, paredes dos vasos sanguíneos e outros; a sua contracção provocaa alteração do diâmetro destes órgãos, permitindo a regulação da
passagem de substâncias pelo tracto gastrointestinal, o controlo do fluxo eda pressão sanguínea e a regulação do fluxo de ar nas vias aéreas; é decontrolo involuntário/inconsciente através do SNP.
• Cardíaco: é o músculo do coração e a sua função é promover ascontracções que permitem mover o sangue, permitindo disponibilizarnutrientes a todo o organismo. O seu controlo é ritmado einvoluntário/inconsciente.
Músculo esquelético: cerca de 40% do peso do corpo;responsável pela locomoção, expressão facial,fonação, postura, movimentos respiratórios e muitosoutros movimentos. As suas funções estão em largamedida dependentes de controlo voluntário ouconsciente pelo SN;
Músculo liso: paredes dos órgãos ocos e tubos,músculos internos do olho, paredes dos vasossanguíneos e outras áreas. Realiza uma grandevariedade de funções, incluindo a propulsão da urina
através do tracto urinário, mistura e mobilização dos
59

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 60/374
alimentos no tubo digestivo, dilatação e constrição das pupilas e regulação dofluxo de sangue nos vasos sanguíneos;
Músculo cardíaco: encontra-se no coração. As suas contracções produzem a forçapara mover o sangue através do sistema circulatório.
Existem três tipos de fibras musculares:
Músculo Esquelético
As fibras musculares esqueléticas que, após o nascimento forem destruídasnão podem ser substituídas pela divisão de fibras musculares já existentes.Novas fibras podem formar-se através de células indiferenciadas chamadascélulas satélite.
O músculo esquelético humano consiste em centenas de células individuaiscilíndricas (denominadas fibras musculares) ligadas entre si por tecido conjuntivo.A sua contracção é iniciada pela despolarização dos neurónios motoressomáticos, que propagam os potenciais de acção desde o cérebro, ou medulaespinal, até às fibras musculares esqueléticas. Ao chegar ao músculo a fibranervosa ramifica-se e vai enervar várias fibras musculares individuais. Apesar deum neurónio motor individual poder inervar várias fibras musculares cada fibramuscular é inervada apenas por um neurónio motor. A associação de um úniconeurónio motor, com todas as fibras musculares por ele controladas denomina-seunidade motora.
Quando o neurónio motor somático é activado, todas as fibras muscularesque são inervadas por ele respondem a esses impulsos nervosos, gerando osseus próprios sinais eléctricos que, por sua vez, levam à contracção das fibrasmusculares.
Fisiologicamente, o grau de contracção da musculatura esquelética écontrolada por:- Activação de um número necessário de unidades motoras- Controlo da frequência dos potenciais de acção do neurónio motor em cadaunidade motora.
Quando é necessário um aumento da força de contracção do músculo pararealizar uma determinada tarefa, o SNC aumenta o número de unidades motorasactivadas. Este processo designa-se por recrutamento das unidades motoras.
60

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 61/374
Em repouso, o músculo esquelético apresenta um fenómeno designado portónus muscular, um estado constante de ligeira tensão que mantém o músculonum estado de constante prontidão. O tónus muscular deve-se a uma activaçãoalternada de um pequeno número de unidades motoras dentro do músculo porcentros motores localizados no cérebro e medula espinal. O controlo demovimentos suaves no organismo (andar, nadar, etc.) é produzido porcontracções graduais da musculatura esquelética. Estas contracções graduais são
alterações da força de contracção ou de encurtamento do músculoproporcionalmente à carga colocada sobre esse mesmo músculo. O músculoesquelético é pois capaz de reagir a diferentes cargas.
Quando uma unidade motora é activada as fibras musculares geram econduzem os seus próprios potenciais de acção. Apesar do impulso nervosogerado em cada fibra ser muito fraco (cerca de 100 μV), a activação simultâneade várias fibras produz diferenças de potencial na pele circundante que sãosuficientemente elevadas para poderem ser detectadas por eléctrodos desuperfície. A detecção, amplificação e registo de alterações eléctricas da peleprovocadas pela contracção muscular subjacente denomina-se electromiografia(EMG).
O padrão estriado resulta do arranjo de filamentos finos e espessos nocitoplasma, constituindo miofibrilhas. Os filamentos espessos são basicamenteconstituídos por miosina, uma proteína contráctil; enquanto os filamentos finossão compostos por actina, troponina e tropomiosina.
Contracção muscularesquelética (sebenta antiga)
61

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 62/374
A contracção muscular esquelética processa-se, resumidamente, daseguinte forma:1- Um potencial de acção propaga-se ao longo de uma fibra nervosa motora atéàs suas terminações nas fibras musculares2- Em cada terminação, o neurónio liberta uma pequena quantidade deacetilcolina3- A acetilcolina actua sobre uma zona local da membrana da fibra muscular para
abrir múltiplos canais com receptores colinérgicos nicotínicos4- A abertura dos canais de acetilcolina permite que grandes quantidades de Na+fluam para o interior da membrana. Este estímulo é o início do potencial de acçãomuscular5- Este potencial de acção propaga-se ao longo da fibra muscular da mesmaforma que os potenciais de acção se propagam ao longo da fibra nervosa6- O potencial de acção despolariza a membrana da fibra muscular, propagando-se também em profundidade através dos túbulos T, o que provoca a libertaçãode grandes quantidades de iões cálcio a partir do retículo sarcoplasmático para osarcoplasma7- Os iões cálcio ligam-se à troponina que ao ser activada promove odeslocamento da tropomiosina, com a consequente exposição dos locais deligação da miosina na actina, o que conduz à contracção e ao eventualencurtamento do músculo, dependendo da carga8- Após uma fracção de segundo, os iões cálcio começam a ser bombeados denovo para o interior do retículo, onde permanecerão até à propagação de umnovo potencial de acção9- O abaixamento da concentração de Ca2+ no citosol faz com que este ião sedesligue da troponina, iniciando-se, por um processo inverso, o relaxamentomuscular.
Componentes estruturaisO músculo é composto de
fascículos que podem ser observados àvista desarmada; estes fascículos sãocompostos de feixes de fibrasmusculares (células musculares).
Cada célula muscular esqueléticaestá envolvida pela lâmina externacomposta de fibras reticulares quequando observada ao microscópio
óptico não se distingue da membranada fibra muscular sarcolema (do gregosarco=carne);
O endomisium (do gregomys=músculo) é uma rede de tecido conjuntivo laxo com numerosas fibrasreticulares que envolve cada fibra muscular por fora da lâmina externa;
Um feixe (fascículo) de fibrasmusculares é envolvido por outra camadade tecido conjuntivo mais denso chamadoperimisium;
O músculo é formado por umconjunto de fascículos, sendo esteconjunto envolvido pelo epimísium
62

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 63/374
(continuação da fáscia sobre o músculo), formado por tecido conjuntivo denso,fibroso e colagénico e que é contínuo com os tendões, também ligados aoendomísium, perimísium e osso.
O sarcómero é a subunidade funcional básica de contracção do músculo,consistindo numa estrutura organizada entre duas linhas Z, composta,principalmente de filamentos de actina e miosina.
Organização dos Miofilamentos
63

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 64/374
O miofilamento fino
Actina: filamento composto por duas cadeias poliméricas de actina actina F emdupla hélice formadas por monómeros de actina G, cada uma com um local
específico para ligação da miosina durante a contracção muscular e uma série demoléculas de troponina.
Tropomiosina: molécula proteica alongada que se insinua na fenda da duplahélice da actina F e cobre 7 locais activos de actina G.
Troponina: complexo de 3 proteínas constituído por Troponina I inibidora daligação cruzada à actina, Troponina T que se liga à tropomiosina e troponina Cque liga cálcio.
O
miofilamento espesso é composto por muitas moléculas de miosina, cada umaformada por 2 cadeias pesadas em dupla hélice e por duas cabeças globularesque se estendem lateralmente. 4 cadeias leves ligam-se duas a duas às cabeças
das cadeias pesadas. Cerca de 150 cabeças projectam-se para cada extremidadedo miofilamennto.As cabeças de miosina têm 3 propriedades importantes: (1) podem ligar-se
aos sítios activos das moléculas de actina formando as pontes cruzadas, (2)ligam-se à porção cilíndrica por uma zona encurvada que se pode dobrar durantea contracção (charneira) e (3) as cabeças têm actividade cinásica para fosforilar acadeia leve da miosina e ATPasica para desdobrar ATP e libertar energia paradobrar a porção em charneira da molécula de miosina durante a contracção.
64

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 65/374
Uma unidade motora consiste numneurónio motor e todas as fibrasmusculares do mesmo tipo com as quais
ele comunica através dos seus terminais.Habitualmente cada fibra tem apenasuma junção neuromuscular.
As fibras da mesma unidade motoranão são adjacentes umas das outras,encontrando-se dispersas no músculomisturadas com as fibras das outrasunidades motoras.
Um músculo é composto por váriasunidades motoras com número ediâmetro de fibra diferentes umas das
outras.Quando um PA ocorre num neurónio motor, todas as fibras musculares da
sua unidade motora são estimuladas a contraírem-se.65

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 66/374
O tamanho das unidades motoras de um músculo esquelético édeterminado pela sua função e localização no organismo. Quanto mais pequeno otamanho das unidades motoras, maior o número de neurónios necessários paracontrolar esse músculo e maior o grau de controlo do Sistema Nervoso Central(SNC) sobre a contracção. Por exemplo, os músculos que movem os dedos têmunidades motoras muito pequenas permitindo um controlo muito preciso dosmovimentos do dedo.
Junção neuro-muscular
A conexão entre o neurónio motor e a fibra muscular designa-se por junçãoneuro-muscular. Neste ponto a membrana da fibra muscular assume uma formaespecializada designada por placa motora terminal, encontrando-se o sarcolemaextensivamente invaginado.
Uma unidade motora consiste num neurónio motor e todas as fibrasmusculares que com ele comunicam, sendo que habitualmente cada fibramuscular tem apenas uma junção neuromuscular.
Cada fibra muscular esquelética está ligada a um axónio do neuróniomotor, com origem no encéfalo ou na medula espinal, só se contraindo quandoestimulada através do neurónio motor. A conexão entre o neurónio motor e afibra muscular designa-se por junção neuromuscular. Neste ponto a membranada fibra muscular assume uma forma especializada designada placa motoraterminal e nesta região da fibra muscular predominam as mitocôndrias.
O terminal do axónio motor ramifica-se penetrando na membrana da fibramuscular, no i8nterior do sacrolema. O citoplasma nas extremidades desteneurónio motor também é rico em mitocôndrias e contém vesículas sinápticas.
Junção neuromuscular (sebenta antiga)
As fibras musculares esqueléticas são inervadas por fibras nervosasgrandes e fortemente mielinizadas que se originam nos motoneurónios desde ointerior da medula. A terminação nervosa estabelece uma união denominada junção neuromuscular com a fibra muscular. Os ramos terminais da fibra nervosaformam um complexo de terminais nervosos ramificados que se invaginam nointerior da fibra muscular, mas que ficam fora da membrana plasmática destafibra. Todo este conjunto é revestido por uma ou várias células de Schwann. Aeste conjunto chama-se placa motora terminal. Quando um impulso nervosoalcança a união neuromuscular libertam-se aproximadamente 125 vesículas deacetilcolina desde os terminais para a fenda sináptica.
66

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 67/374
A chegada do potencial de acção ao terminal da fibra nervosa tambémprovoca a entrada de cálcio para a célula através da abertura de canais de cálciodependentes da voltagem. Este cálcio associa-se às vesículas de acetilcolina eestas fundem-se com a membrana libertando a acetilcolina na fenda sináptica.
Na membrana da fibra muscular existem receptores da acetilcolina(nicotínicos) que ao serem activados por esta substância, entretanto libertada nafenda sináptica, provocam a abertura dos canais de sódio associados. A
simulação desta aula assenta numa preparação simples que serve para visualizara transmissão neuromuscular e a acção de certos fármacos na sua inibição. Paratal, utiliza-se uma parte do hemidiafragma de rato. O hemidiafragma é ummúsculo respiratório esquelético grande, inervado focalmente, composto porfibras musculares rápidas. A neurotransmissão é mediada por receptoresnicotínicos. A estimulação eléctrica do nervo frénico provoca contracções rápidasneste músculo passíveis de serem registadas.
Funcionamento da junção neuro-muscular
Um potencial de acção propaga-se aoterminal pré-sináptico, provocando aabertura de canais de cálcio voltagemdependentes que permitem a passagemdeste ião para o interior do neurónio;
Os iões cálcio ligam-se àsinaptotagmina, desencadeando a
alteração conformacional do complexoSNARE (N-ethylmaleimide-sensitive fusionprotein attachment protein receptors) oque promove a acostagem e a fusão dasvesículas e a libertação daAch para a fendasináptica po exocitose;
A Ach difunde na fenda sináptica eliga-se aos receptores nicotínicos nosarcolema, aumentando a permeabilidadedos canais de sódio.
O aumento da permeabilidade aos iões sódio resulta na despolarização do
sarcolema, daí resultando sempre um potencial de acção pós-sináptico na fibramuscular.
Mecanismo de Acção (resumo)1. Um PA chega ao Terminal Pré-sináptico, provoca a abertura de canais de
cálcio voltagem dependentes, permitindo a entrada deste ião para ointerior do neurónio;
2. Os iões cálcio promovem a fusão de vesículas com a membrana pré-sináptica e libertação de Ach para a fenda sináptica;
3. O Ach difunde na fenda sináptica e liga-se aos seus receptores damembrana da fibra muscular, aumentando a permeabilidade dos canais desódio;
67

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 68/374
4. O aumento da permeabilidade aos iões sódio resulta na despolarização damembrana da fibra muscular, resultando num potencial de acção pós-sináptico se ultrapassar o limiar de excitação.
Cada potencial de acção num neurónio motor produz um potencial de acçãoem cada fibra muscular da sua unidade motora.
Em músculos esqueléticos não ocorrem potenciais inibitórios.A superfície da placa motora terminal contém acetilcolinestrase, quedegrada a Ach. A colina (resultante da degradação) é então transportadas devolta aoo axónio terminal onde é reutilizada na síntese de Ach.
Acoplamento excitação-contracção: proteínas pedunculares
Relação dos túbulos T como RS - Proteínas Pedunculares
O PA nos túbulos T altera acarga eléctrica do receptor de
voltagem dihidropiridínico (R-DHP ) inserido na parede dotúbulo.
Após a estimulação o R-DHP muda a conformação doreceptor raionidinico formado porum pedúnculo de ligação e umcanal de cálcio no RSP que abrepor acção da voltagem,libertando o cálcio para o
sarcoplasma.Do aumento do cálcio nosarcoplásma resulta a sualigação à troponina e obombeamento para o RSP o, quena ausência de estímulo, conduz ao relaxamento muscular.
68

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 69/374
Pulso contráctil, espasmo ou abalo muscular
Embora a função muscular normal sejamais complexa, a compreensão de um pulsocontráctil facilita a percepção dofuncionamento do músculo.
Um pulso contráctil ou abalo muscularé a contracção de uma fibra muscular emresposta a um único potencial de acção.
Há uma curta fase de latência após aaplicação do estímulo correspondente àexcitação, seguida de uma fase de aumentoda tensão correspondente à contracção e de
uma fase de relaxamento.
Contracção isométrica (comprimentoconstante): desenvolvimento de tensão interna sem encurtamento. Ocorre noinício da contracção (absorção de tensão pelos componentes elásticos dosarcómero) ou quando a força exercida por um objecto sobre o músculo emcontracção é superior à tensão desenvolvida pelo músculo, mantendose a cargaimóvel.
Contracção isotónica ou concêntrica: encurtamento do músculo com tensãoconstante. Inicia-se quando a tensão interna ultrapassa a força exercida pelacarga, movendo-a, mantendo-se a partir daí a tensão constante. É típica dolevantamento de pesos ou do acto de pedalar.
69

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 70/374
Contracção isotónica de alongamento ou excêntrica: alongamento do músculomantendo-se a tensão constante por a carga ser muito maior que a tensãogerada pelas pontes cruzadas. É típica do abaixamento de pesos, mas podeconduzir à lesão do músculo por rotura de ligamento ou tendão (exemplo datortura de estiramento por roldana - inquisição).
Relação tensão isométrica desenvolvida versus comprimento de repousodo sarcómero
A tensão isométrica máxima é produzida para o comprimento óptimo dosarcómero, I0.
O estiramento ou o encurtamento da fibra muscular além ou aquém de I0diminui a tensão gerada, devido à sobreposição inadequada das proteínas
contrácteis.A sobreposição de filamentos é máximo em <I0, permitindo a ligação domáximo número de pontes cruzadas aos filamentos de actina, provocando assima tensão máxima.
Quando todos os músculos esqueléticos do corpo estão relaxados, ocomprimento da maioria das fibras está perto de I0 e por isso próximo docomprimento óptimo para a geração de força.
70
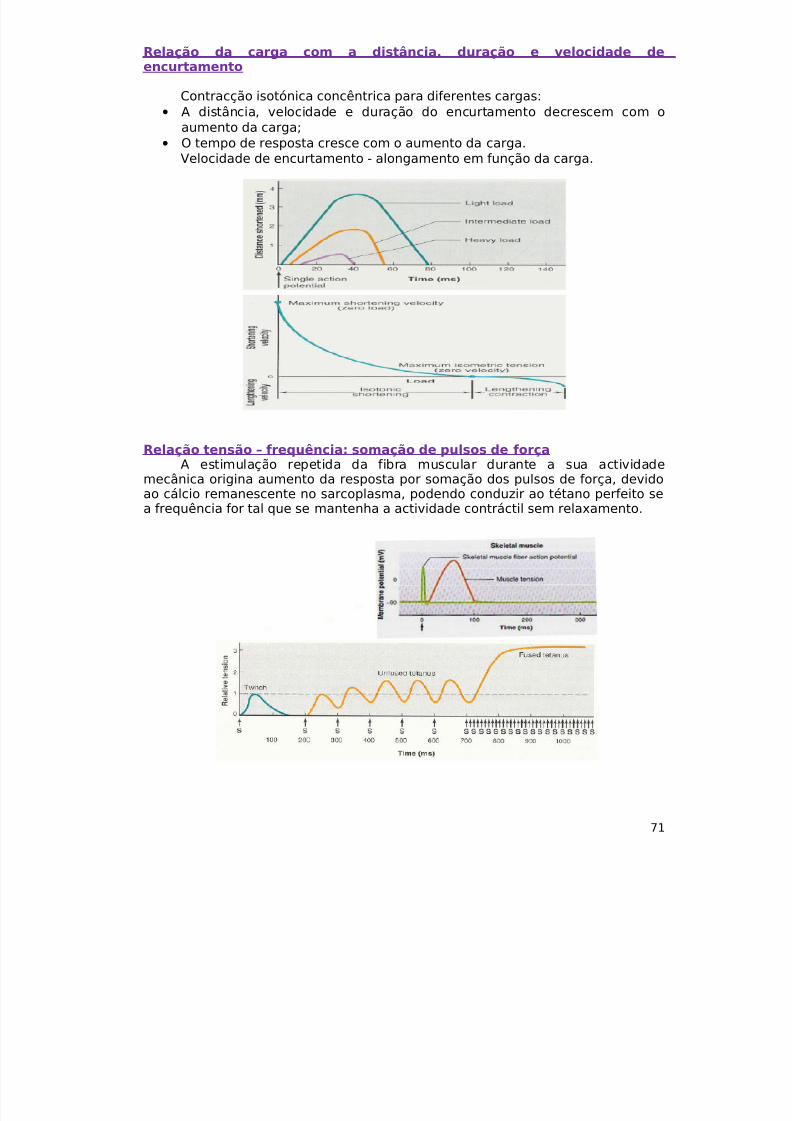
7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 71/374
Relação da carga com a distância, duração e velocidade deencurtamento
Contracção isotónica concêntrica para diferentes cargas:• A distância, velocidade e duração do encurtamento decrescem com o
aumento da carga;• O tempo de resposta cresce com o aumento da carga.
Velocidade de encurtamento - alongamento em função da carga.
Relação tensão – frequência: somação de pulsos de força
A estimulação repetida da fibra muscular durante a sua actividademecânica origina aumento da resposta por somação dos pulsos de força, devidoao cálcio remanescente no sarcoplasma, podendo conduzir ao tétano perfeito sea frequência for tal que se mantenha a actividade contráctil sem relaxamento.
71

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 72/374
Somação e Tétano
Cerca de 40% da massa total do corpo humano é músculo esquelético. Omúsculo esquelético está intimamente associado com o sistema esquelético ecombinados, estes músculos e ossos, são responsáveis pelo suporte e movimentodo corpo. Embora todas as fibras do músculo esquelético tenham sarcómeros e amesma aparência estriada, os diferentes músculos podem funcionar de
diferentes maneiras. Por exemplo, alguns são relativamente fracos e resistentesà fadiga, enquanto outros são fortes, mas cansam-se facilmente. Estascaracterísticas podem ser explicadas em termos de propriedades bioquímicas.
As fibras musculares encontradas na maior parte dos músculosesqueléticos dos mamíferos são de contracção rápida ou lenta. Cada tipo de fibracontém isozimas com diferentes actividade miosina ATPase e velocidades deligação das pontes transversas. Dentro do grupo de fibras de contracção rápida,existem as fibras que usam ambas a glicólise e a fosforilação oxidativa. Tambémexistem fibras de contracção rápida que só usam a glicólise.
Este grupo, tende a necessitar de menos oxigénio e é muito mais forte doque as fibras que também usam fosforilação oxidativa. No entanto, estas fibras
glicolíticas fortes quebram a glucose de forma muito ineficiente. Um aumentorepentino da actividade contráctil utiliza a maior parte da glucose disponível,causa a acumulação de ácido láctico e leva à fadiga mais rapidamente.
Apesar de um neurónio motor formar sinapses com muitas fibrasmusculares, todas as fibras musculares ligadas a esse neurónio motor são domesmo tipo. Assim, o estímulo de um neurónio motor em particular irá causar acontracção de apenas um tipo de fibra muscular. Os neurónios motores quefornecem as fibras oxidativas fracas e lentas, têm o limiar de excitação maisbaixo. Aqueles que inervam as fibras oxidativas rápidas de força intermédia têmo limiar de excitação mais alto e os que inervam fibras fortes e rápidas, apenasglicolíticas, têm o limiar de excitação mais elevado. Ao aumentar a quantidade deactividade que vem do encéfalo são progressivamente activados mais neurónios
72

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 73/374
motores e mais das fibras musculares fortes são incorporadas na resposta. Tudoisto ocorre através de uma única sinapse em cada fibra.
Quando um músculo tenta deslocar uma carga, primeiro o músculo geratensão interna para vencer a resistência dos vários elementos elásticos, incluindodos tendões que mantêm o músculo ligado aos ossos. O tempo dedesenvolvimento desta tensão interna antes de ocorrer movimento correspondeao período de latência e é directamente proporcional á carga deslocada pelo
músculo. Quando a tensão do músculo excede a carga, dá-se o seu encurtamentodo e o movimento da carga. O tempo que o músculo está no seu estado activo(contracção) é finito, por isso, os músculos têm menos tempo para encurtarquando estão a mover objectos mais pesados.
Nesta próxima experiência, o objectivo é demonstrar o efeito do aumentoda intensidade do estímulo na força de contracção de um músculo, o efeito doaumento de peso na amplitude de contracção e no trabalho de um músculo jácarregado e o efeito do aumento da frequência de estímulo na contracçãomuscular e na fadiga do músculo.
Esta experiência envolve a aplicação de pulsos de estímulo à pele, na parteposterior da mão, de forma a produzir uma contracção nos músculos que movemo dedo. Os pulsos de estímulo usados são originados por um isolador de estímulode alta voltagem SI-200.
Enquanto a sensação de contracção muscular provocada pode inicialmenteparecer peculiar, o estímulo dos músculos da mão é indolor e seguro. As pessoascom função cardíaca fraca, pacemakers ou qualquer outra condição que possaser agravada por estimulação eléctrica, não devem efectuar esta experiência.
Contracção do músculo como um todo: recrutamento de unidadesmotoras
Músculos que produzem movimentos delicados (dedos) têm poucas fibraspor unidades motores; ao contrário dos músculos posturais (costas e pernas).As unidades motoras rápidas glicolíticas têm fibras de grande diâmetro e
em grande número (braços).O controlo da tensão muscular é efectuado pelo aumento progressivo do
número de unidades motoras activas – recrutamento (1º LO nas contracçõesfracas, 2º RO e 3º RG para contracções progressivamente mais fortes).
O recrutamento aumenta a velocidade a que é deslocada a carga.
Fontes de ATP para a contracção muscularAs fibras musculares formam ATP por: (i) transferência de Pi do fosfato de
creatinina (CP) para o ADP, por acção da fosfoquinase; (ii) por transferência de Pide ADP para outro ADP por acção da miokinase e (iii) por fosforilação de ADP namitocôndria; (iv) por fosforilação de ADP na glicólise.
No início é consumido o glicogénio muscular, depois é a glucose e os AGsanguíneos; quando a intensidade do exercício atinge 70% do máximo a glicólisetorna-se a mais importante na produção de ATP, dado que, embora esta viaproduza menos energia é mais rápida a faze-lo.
Mecanismo de Filamentos Deslizantes
73

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 74/374
Quando há contracção muscular ocorre o encurtamento do sarcómero oque ocorresemalteraçãodotamanho
dosfilamentosde actina emiosina.Duranteesteencurtamento cada ponte cruzada da miosina ligada ao filamento de actinamovimenta-se em arco.
Ou seja: este mecanismo, que ocorre aquando da contracção muscular está
dependente da existência de pontes cruzadas da miosina ligada à actina,forçando o movimento dos filamentos de actina entre os filamentos de miosina.(actina-móvel; miosina-móvel)
Componentes do Sarcómero:
• Actina: proteína globular formada por duas cadeias helicoidais;
• Miosina: proteína composta por 2 cadeias pesadas e 4 cadeias leves. Estespolipéptidos (as cadeias) combinam-se para formar uma molécula com
duas cabeças globulares (contendo cadeias pesadas e leves) e uma cauda(longa que contém duas cadeias pesadas). Cada cabeça globular possuidois locais de ligação: 1 para a actina e outro para o ATP.
• Titina: “mola” que permite o deslizamento da actina sobre a miosina.
Ciclo das Pontes Cruzadas:Dá-se o nome de Ciclo das Pontes Cruzadas ao conjunto de eventos que
ocorre no momento em que uma ponte cruzada se liga a um filamento fino,move-se e, em seguida, é ajustada para repetir o processo.
Cada ciclo consiste em 4 etapas: 1 – fixação da ponte cruzada a umfilamento fino (de actina); 2 – movimento da ponte cruzada, produzindo tensão
74

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 75/374
no filamento de actina; 3 – desacoplamento da ponte cruzada e do filamento deactina; 4 – energização da ponte cruzada de modo a que esta possa fixar-senovamente a um filamento fino e repetir o ciclo.
As pontes cruzadas realizam o seu ciclo independentemente umas dasoutras pontes, num qualquer instante durante a contracção apenas uma porçãodas pontes cruzadas é fixada aos filamentos de actina e produz tensão, enquanto
outras estão noutra fase do seu ciclo.
Numafibra muscularem repouso a[Ca2+] é baixa e aspontes cruzadasde miosina
não podemligar-se à actina.O ciclo das
pontes cruzadasinicia-se com aentrada de cálciono
citoplasma, promovendo a ligação de uma ponte cruzada de miosina energizada
a uma molécula de actina do filamento fino:
A.+M.ADP.P. (miosina energética) A.M.ADP.P. (ligação da actina)Sendo: A (actina) e M (miosina)
A ligação da miosina energética à actina desencadeia a libertação daconformação forçada da ponte energética que produz o movimento da pontecruzada ligada, e a libertação de Pi e ADP.
A.M.ADP.P. A.M. + ADP + Pi (movimento das pontes cruzadas)
75

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 76/374
Durante o movimento das pontes cruzadas a ligação da miosina à actinadeve ser quebrada de modo a permitir que a miosina, ou melhor, a sua pontecruzada seja reenergizada para poder repetir-se o ciclo. A ligação de uma novamolécula de ATP à miosina quebra a ligação da actina à miosina.
A.M. + ATP A. + M.ATP (dissociação das pontes cruzadas de actina)
Após a dissociação da actina e miosina, o ATP ligado à miosina é separado,formando-se novamente o estádio energético da miosina.
A.+M.AMP A. + M.ADP.Pi (hidrólise de ATP)
Funções do ATP no ciclo:1. A energia libertada pela hidrólise do ATP permite que haja energia para o
movimento da ponte.
2. A ligação do ATP à miosina quebra a ligação entre este e a actina,permitindo a ocorrência de um novo ciclo.
Papel da Tropomiosina, Troponina e Cálcio na Contracção
Tropomiosina: composta pelo enrolamento de 2 polipéptidos. As cadeias detropomiosina cobrem parcialmente os locais de ligação da miosina em cadamolécula de actina. É mantida em posição pela Troponina.
Troponina: mais pequena que a tropomiosina, é globular e responsável pelamanutenção da posição da tropomiosina.
Cálcio: liga-se a sítios específicos da troponina, provocando a sua alteração,deslocando a tropomiosina e deixando a descoberto o local de ligação das pontescruzadas. Assim a [Ca2+] citosólica determina o número de locais de troponinaocupados pelo Ca2+, que por sua vez determina o número de locais de actinadisponíveis para ligação com pontes cruzadas.
Musculo Esquelético – Túbulos T e reticulo sarcoplasmáticoO sarcolema tem ao longo da sua superfície muitas invaginações tubulares
designadas por túbulos transversos ou túbulos T que se projectam para o interiordas fibras musculares junto às regiões dos sarcómeros onde os miofilamentos daactina e miosina se sobrepõem.
O lúmen de cada túbulo Testá cheio com fluidoextracelular, sendo as suasparedes contínuas com oexterior da fibra muscular. Entreos túbulos T encontra-se oretículo sarcoplasmático
(retículo endoplasmático lisoespecializado) que formadilatações, próximas dos túbulos
76

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 77/374
T, chamadas cisternas terminais, designando-se o conjunto de um túbulo T e deduas cisternas adjacentes por tríada.
A membrana do retículo sarcoplasmático transporta activamente iões cálciodo sarcoplasma para o seu lúmen, onde os iões cálcio são armazenados emelevada concentração comparativamente com o sarcoplasma.
Fases e Eventos do Pulso Contráctil
• Fase de Latência: Um potencial de acção (PA) é propagado até ao terminalpré-sináptico do neurónio motor do sistema nervoso somático aumentandoa permeabilidade da membrana ao ião cálcio que difunde para o interior doterminal, provocando, com o concurso das proteínas SNARE e dasinaptotagmina, a fusão das vesículas e a exocitose de acetilcolina (Ach)para a fenda sináptica;
A ligação da Ach ao receptor nicotínico no sarcolema causa a
abertura dos canais de sódio ligando-dependentes, o que torna amembrana pós-sináptica mais permeável a este ião que assim difunde parao interior da fibra muscular causando a despolarização local que excede olimiar de excitação produzindo PA;
A Ach é rapidamente degradada na fenda sináptica numareacção catalisada pela acetilcolinestrase, originando ácido acético e colina,o que limita o tempo de ligação da Ach ao seu receptor; Assim, o PA pré-sináptico produz um PA pós-sináptico na fibra muscular;
O PA gerado no centro da fibra muscular é propagado nosarcolema em direcção a ambos os extremos e também para o interior
através dos túbulos Transversos (T);Em reposta ao PA, há despolarização nos túbulos T que altera a
carga eléctrica do receptor de voltagem dihidropiridínico (DHP) o quemodifica a conformação do receptor de raionidina (alcalóide vegetal), quepor sua vez induz a abertura do canal de cálcio do RS que lhe estáassociado. Os iões cálcio difundem do RS para o sarcoplasma e ligam-se àtroponina, daí resultando que o complexo troponina-tropomiosina altera asua posição e expõe o local activo dos filamentos de actina aos quais seligam as pontes cruzadas da miosina.
• Fase de Contracção: As pontes cruzadas entre as moléculas de actina e de
miosina formam-se, movem-se, libertam-se e voltam a formar-se muitasvezes, gerando tensão e provocando o eventual encurtamento dosarcómero. A energia armazenada nas cabeças das moléculas de miosinapermite a ligação das pontes cruzadas e o movimento. Após a ocorrênciado movimento das pontes cruzadas, o ATP liga-se às cabeças de miosina,provocando o seu desacoplamento da actina, após o que é hidrolisado aADP+Pi, sendo alguma da energia armazenada na cabeça de miosina usadapara a ligação da próxima ponte cruzada e para o seu movimento. A maiorparte da energia é também libertada sob a forma de calor.
•
Fase de Relaxamento: Na ausência de novo estímulo, o contínuo transporteactivo dos iões cálcio do sarcoplasma para o retículo sarcoplasmáticosupera a sua libertação no sentido inverso, fazendo com que este ião se
77

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 78/374
desligue da troponina e o complexo desta com a tropomiosina bloqueia aligação de pontes cruzadas, levando assim ao relaxamento da fibramuscular.
Metabolismo EnergéticoA fibra muscular pode formar ATP de 3 formas:
1. Fosforilação do ADP pela creatina fosfato, que é muito rápida e em que aquantidade de ATP está limitado pela [] de creatina fosfato;2. Fosforilação oxidativa do ADP na mitocôndria;3. Fosforilação do ADP pela via da glicólise no citosol, a qual produz pequenas
quantidades de ATP por cada molécula de glucose metabolizada e podeocorrer na ausência de oxigénio.
Fadiga Muscular
Quando uma fibramuscular esquelética érepetidamente estimulada, atensão desenvolvida pela fibradiminui embora continue aestimulação. Há diminuição davelocidade de encurtamento erelaxamento também maislento.
A fadiga muscular écausada não por falta de ATP,mas por vários factores taiscomo alterações na acidez,variações na disponibilidade deoxigénio e falha nosmecanismos acoplamento excitação-contracção.
A força e susceptibilidade à fadiga de um músculo pode ser alterada peloexercício:
• Exercícios de baixa intensidade, mas de longa duração (endurance)
aumentam a capacidade da fibra para produzir ATP por via oxidativaatravés do aumento do número de mitocôndrias e vasos sanguíneos;• Exercícios de curta duração, mas de elevada intensidade (levantamento de
pesos) aumentam o diâmetro das fibras como resultado do aumento dasíntese de actina e miosina, o que resulta no aumento de força.
Factores determinantes da tensão muscularI.
• Tensão desenvolvida por cada fibra individual• Frequência do potencial de acção (relação tensão-frequência)•
Comprimento do sarcómero (relação tensão-comprimento)• Diâmetro da fibra• Fadiga
78

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 79/374
II.• Número de fibras activas• Número de fibras por unidade motora• Número de unidades motoras activas
Tipos de fibras musculares esqueléticas: actividade da miosina quinase versusorigem do ATP
i) Lentas oxidativas (baixa actividade ATPásica/fosforilação oxidadtiva)ii) Rápidas oxidativas (elevada actividade ATPásica/fosforilação oxidadtiva)iii) Rápidas glicolíticas (elevada actividade ATPásica/glicólise)
Os três tipos de fibras diferem na resistência á fadiga, com as rápidasglicolíticas a fatigarem-se mais rapidamente e as lentas oxidativas a resistiremmais tempo.
Classificam-se com base na velocidade máxima de encurtamento e na viapredominante da formação de ATP (fosforilação oxidativa ou glicólise), sendo queas diferenças na velocidade de encurtamento são devidas às diferentes isozimasda miosina quinase com baixa ou elevada actividade ATPásica, originando fibraslentas ou rápidas.
As fibras rápidas glicolíticas têm um diâmetro maior que as oxidativas, peloque precisam de mais energia para serem activadas, geram maior tensão, mastambém fatigam mais rapidamente.
Todas as fibras musculares de uma unidade motora pertencem ao mesmo tipoe a maior parte dos músculos contêm os três tipos de fibras, embora emdiferentes proporções.
79

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 80/374
Controlo da Tensão Muscular
A tensão total que pode ser desenvolvida por um músculo depende de:1. Quantidade de tensão desenvolvida por cada fibra;2. Número de fibras que contraem ao mesmo tempo:
o N.º de fibras em cada unidade motora;o N.º de unidades motoras activas.
A força produzida por uma fibra singular depende em parte do diâmetro dafibra: maior diâmetro maior força.
O processo que permite aumentar o n.º de unidades motoras activas num
músculo denomina-se recrutamento. Este processo torna-se possível pelaactivação de inputs sinápticos excitatórios (maior n.º de neurónios motoresactivos maior recrutamento e maior tensão).
Como a quantidade de sódio que entra numa célula numa sinapse é igualquer se trate de um neurónio motor pequeno ou grande, os neurónios pequenossão os primeiros a ser recrutados.
O recrutamento ocorre por esta ordem: 1.º unidades motoras oxidativaslentas; 2.º oxidativas rápidas; 3.º glicolíticas rápidas.
Músculo Liso
O músculo liso está largamente distribuído através do organismo e é maisvariável na sua função que os outros tipos de músculo. As suas células são maispequenas que as do músculo esquelético, apresentando-se em forma de fusocom um único núcleo localizado a meio da célula. Para além disto, os nervos comeles relacionados derivam da divisão autónoma do sistema nervoso e não dadivisão somática, quer isto dizer que em situações normais, o músculo liso nãoestá sob controlo voluntário.
Cada fibra muscular lisa é uma célula fusiforme, uninucleada, e comcapacidade de se dividir, quando estimulado por agentes parácrinos ou por lesão.
No citoplasma das fibras musculares lisas estão presentes 2 tipos de
filamentos: filamentos grossos de miosina e filamentos finos de actina. Estesúltimos são ancorados à membrana ou às estruturas denominadas corposdensos, que se assemelham às linhas Z do músculo esquelético.
80
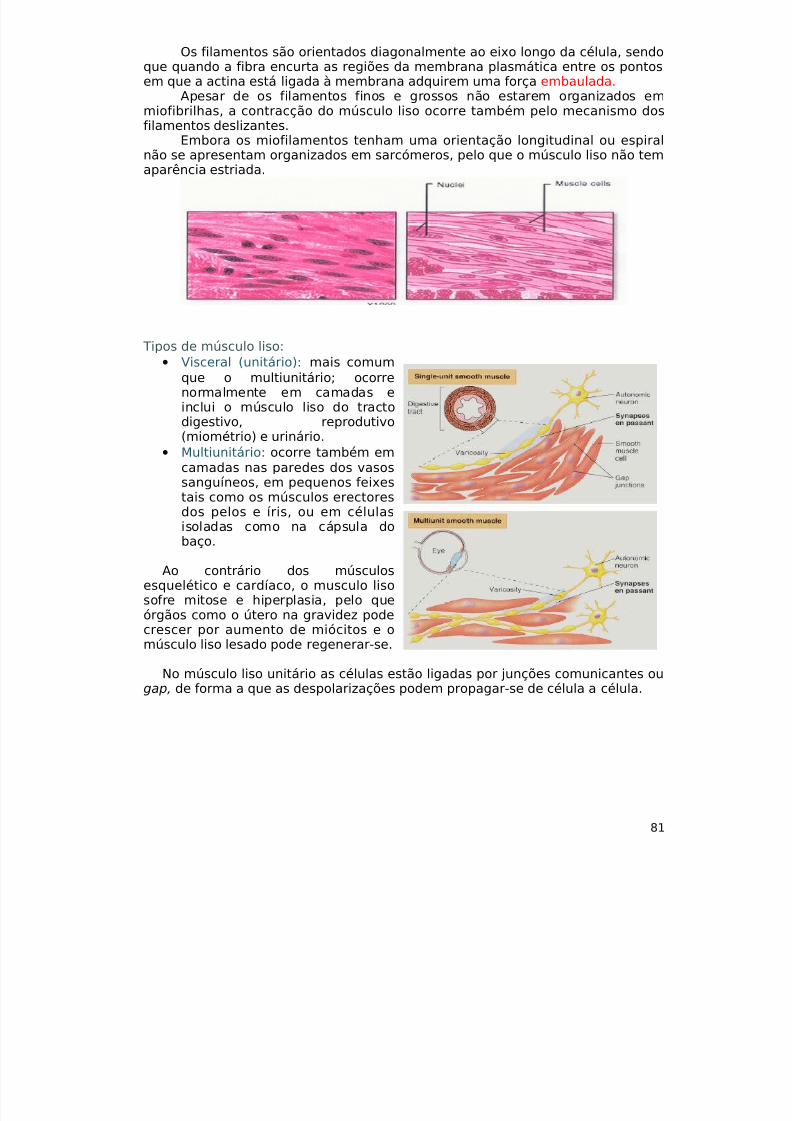
7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 81/374
Os filamentos são orientados diagonalmente ao eixo longo da célula, sendoque quando a fibra encurta as regiões da membrana plasmática entre os pontosem que a actina está ligada à membrana adquirem uma força embaulada.
Apesar de os filamentos finos e grossos não estarem organizados emmiofibrilhas, a contracção do músculo liso ocorre também pelo mecanismo dosfilamentos deslizantes.
Embora os miofilamentos tenham uma orientação longitudinal ou espiral
não se apresentam organizados em sarcómeros, pelo que o músculo liso não temaparência estriada.
Tipos de músculo liso:• Visceral (unitário): mais comum
que o multiunitário; ocorrenormalmente em camadas einclui o músculo liso do tractodigestivo, reprodutivo(miométrio) e urinário.
• Multiunitário: ocorre também em
camadas nas paredes dos vasossanguíneos, em pequenos feixestais como os músculos erectoresdos pelos e íris, ou em célulasisoladas como na cápsula dobaço.
Ao contrário dos músculosesquelético e cardíaco, o musculo lisosofre mitose e hiperplasia, pelo queórgãos como o útero na gravidez podecrescer por aumento de miócitos e omúsculo liso lesado pode regenerar-se.
No músculo liso unitário as células estão ligadas por junções comunicantes ougap, de forma a que as despolarizações podem propagar-se de célula a célula.
81
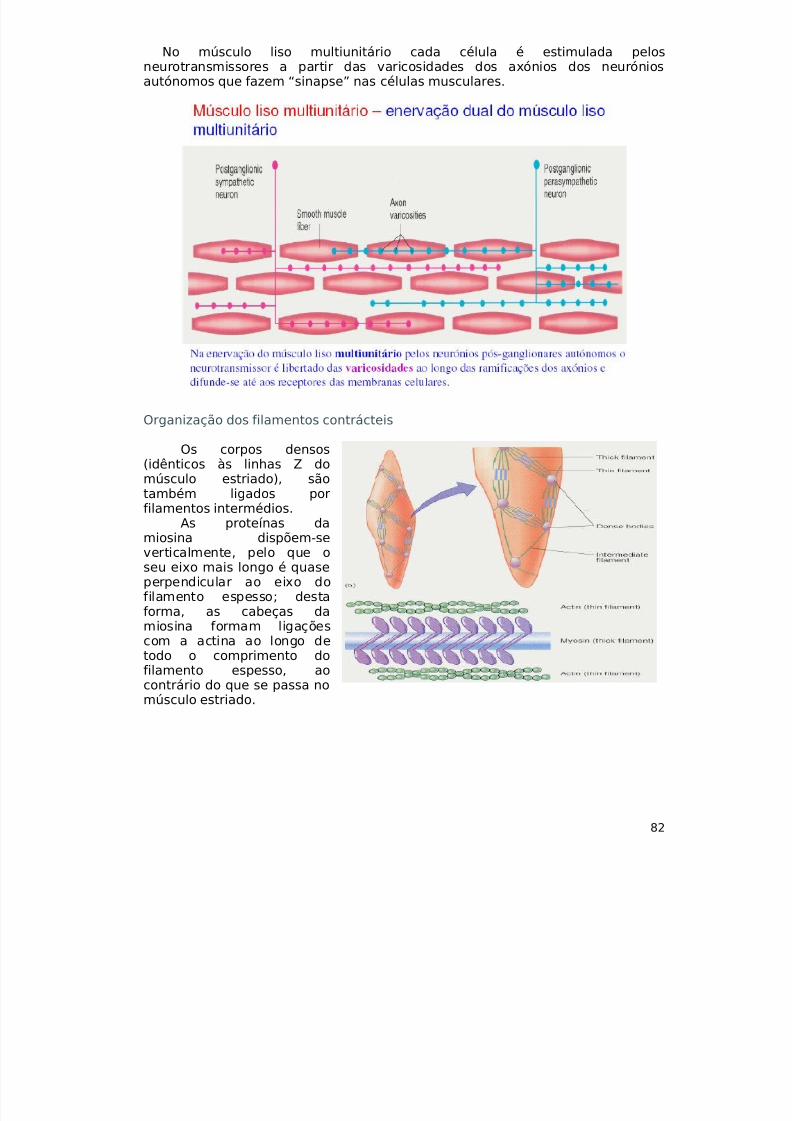
7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 82/374
No músculo liso multiunitário cada célula é estimulada pelosneurotransmissores a partir das varicosidades dos axónios dos neuróniosautónomos que fazem “sinapse” nas células musculares.
Organização dos filamentos contrácteis
Os corpos densos
(idênticos às linhas Z domúsculo estriado), sãotambém ligados porfilamentos intermédios.
As proteínas damiosina dispõem-severticalmente, pelo que oseu eixo mais longo é quaseperpendicular ao eixo dofilamento espesso; destaforma, as cabeças da
miosina formam ligaçõescom a actina ao longo detodo o comprimento dofilamento espesso, aocontrário do que se passa nomúsculo estriado.
82

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 83/374
Acoplamento excitação-contracção
O estímulo inicial para desencadear a resposta muscular é o aumento dosiões cálcio intracelulares provenientes do retículo sarcoplasmático e
principalmente do meio extracelular. Este aumento pode ser causado por umaestimulação nervosa das fibras lisas, por estimulação hormonal, ou por distensãoda fibra.
No caso do músculo liso, o ciclo das pontes cruzadas é controlado por umaenzima (calmodulina), reguladora de cálcio, que fosforila a miosina, já que estaapenas se liga à actina para dar inicio ao ciclo se estiver na forma fosforilada.Assim:
1. O Ca2+ passa através dos canais voltagem dependentes da membranasarcoplasmática (ou, em menor quantidade, pelos canais dependentesdo IP3 do RS, de hormonas ou ainda de canais mecanodependentes queabrem em resposta à distenção da célula), entra no sarcoplasma e liga-se à calmodulina, formando o complexo Ca2+ - calmodulina;
2. Este complexo activa a miosina quinase de cadeia leve (MQCL) porremoção de um grupo fosfato;
3. A MQCL fosforila as cadeias leves da miosina (CLM) a partir do ATP,activando assim as pontes cruzadas e conduzindo á contracção;
4. A contracção termina quando, por ausência de estímulo, os canais decálcio fecham, a concentração citosólica deste ião se reduz porbombeamento, o Ca2+ - calmodulina se desliga da MQCL, inactivando-ae o Pi ligado à CLM é removido pela miosina fosfatase activadadesacoplando-se assim as pontes cruzadas e dando-se o relaxamento.
83

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 84/374
Musculo liso vs músculo esquelético
Embora a maior parte dosmúsculos esqueléticos se contraiam e relaxem rapidamente, a maior parte dacontracção muscular lisa é uma contracção prolongada tónica, que por vezesdura horas ou mesmo dias. É, portanto, de esperar que as características físicas equímicas da contracção muscular lisa sejam diferentes da esquelética.
Ciclo lento das pontes transversas: A velocidade do ciclo de pontestransversas é muito menor no músculo liso do que no músculo esquelético; noentanto, pensa-se que a fracção de tempo que as pontes transversaspermanecem unidas aos filamentos de actina, e que é o principal factor que
determina a força de contracção, é muito maior no músculo liso.Atraso do começo da contracção e do relaxamento do músculo liso: Um
músculo liso começa a contrair-se 50 a 100 milissegundos após ter sido excitado,alcança a contracção máxima 0,5 segundos mais tarde e a sua força decontracção diminui após 1 a 2 segundos. Este processo completo demora cercade 1 a 3 segundos, sendo este tempo cerca de 30 vezes superior ao tempo decontracção do músculo esquelético, o que está directamente relacionado com alentidão no estabelecimento e rotura das pontes transversas.
Fontes de Cálcio citosólico:
• Retículo Sarcoplasmático: que no caso do músculo liso é uma reservainferior à do músculo esquelético, e não está arranjado de uma forma
84

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 85/374
(padrão) específico. Para além disto, no músculo liso não existem túbulos Tconectados com a membrana.
• Cálcio extracelular que entra na célula por canais extramembranares: dadoque a [] extracelular de cálcio é muito superior à intracelular, e comoexistem canais de cálcio voltagem-dependentes e controlados pormensageiros químicos, há um fluxo de cálcio do exterior para o interior dacélula. Como a célula é pequena a entrada de cálcio ocorre com rápida
ligação deste aos locais apropriados.A remoção cálcio do citosol para que ocorra relaxamento é conseguida pelotransporte activo de cálcio para o exterior, sendo que esta remoção é muitomais lenta que no caso do músculo esquelético.Apenas uma das pontes cruzadas é activada como resposta ao estímulo,
por isso a tensão gerada por uma fibra muscular lisa pode ser gradual, através de[Ca2+], quanto maior, maior o n.º de pontes cruzadas activadas e maior a tensão.
Em certos músculos lisos a [] citosólica de cálcio é suficiente para manterum baixo tónus muscular, ou seja, um baixo n.º de pontes cruzadas activas naausência de estímulo externo.
Noutros casos em que a estimulação é persistente e a [Ca 2+] citosólicacontinua elevada, a velocidade de degradação do ATP pelas pontes cruzadasdiminui mesmo que a tensão se mantenha, designando-se isto por “Laten State”,o que ocorre quando a ponte cruzada fosforilada se desfosforila enquanto aindaestá ligada à actina.
Activação Membranar: ocorre através de inputs que influenciam a actividadecontráctil do músculo liso:
1. Actividade eléctrica espontânea na membrana da fibra muscular;2. Neurotransmissores provenientes de neurónios autónomos;3. Alterações induzidas e locais, na composição química do fluído extracelular
que rodeia a fibra;4. Hormonas;5. Distenção.
1. Ocorre em certos tipos de fibras musculares lisas que geramespontaneamente potenciais de acção na ausência de inputs neurais ouhormonais; sendo que a alteração do potencial de membrana que ocorredurante a despolarização até ao limiar de excitabilidade é designadopotencial pacemaker.
2. Influenciam a resposta já que esta depende do mensageiro químico, dosreceptores e dos mecanismos de transdução de sinal intracelulares.
Músculo Cardíaco
O músculo cardíaco é estriado como o músculo esquelético, mas cadacélula contém habitualmente um único núcleo localizado perto do centro. Ascélulas são ramificadas e ligadas topo a topo por ligações especializadas célula acélula chamadas discos intercalates com desmossomas e junções comunicantesou do iato (sinapses eléctricas), permitindo que os PA passem de célula paracélula e que o miocárdio se comporte como uma unidade funcional.
Algumas células do músculo cardíaco são autorrítmicas e uma parte docoração actua normalmente como pacemaker ou marca passo. A força de
85

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 86/374
contracção das células miocárdicas pode ser aumentada pelaepinefirina/norepinefrina e pelo estiramento das cavidades cardíacas.
Apesar dos PA se originarem normalmente em células pacemaker, a taxade despolarização espontânea e a regulação da frequência cardíaca é controladapor enervação autónoma, tendo uma duração e um período refractário muitomais longos.
Miócitos e discos intercalares com desmossomas e junções comunicantes ou dohiato
86

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 87/374
87

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 88/374
88

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 89/374
89

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 90/374
Fisiologia do sistema sensorial
Principais funções do sistema sensorial:
Forma como o encéfalo recebe informação sobre o meio ambiente e sobre o corpo
• Permitir o contacto com o meio envolvente;• Receber e armazenar os dados úteis para as restantes funções, isto é, processar
os dados (internos e externos).
Sistema sensorial: parte do sistema nervoso que consiste em:
- células receptoras sensoriais que recebem estímulos do ambiente;
- vias neurais que conduzem a informação dos receptores ao SNC;
- partes do encéfalo que processam principalmente as informações.
• Informação sensorial: quando a informação se torna consciente;• Sensação: quando a informação não se torna consciente;• Percepção: nome dado ao entendimento da informação.
• Transforma a informação em sensação quando esta passa a ser consciente
• A sensação passa a percepção quando é integrada no córtex associativo e
compreendida
O início do processamento sensorial consiste na transformação da energia doestímulo em potencial gradativo e depois em potenciais de acção nas fibras nervosas.(Estimulo -> Potencial gradativo -> Potenciais de acção nas fibras nervosas)
Receptores Sensoriais
Estes receptores podem ser terminações especializadas de neurónios aferentes oucélulas separadas que sinalizam os neurónios aferentes pela libertação de mensageirosquímicos.
A energia que os activa é o estimulo e o processo pelo qual o estimulo étransformado numa resposta eléctrica e a transdução de sinal. Há 5 tipos que receptoressensoriais:
• Mecanoreceptores: detectam o frio e o calor (deformações mecânicas);• Quimioreceptores: detectam cheiro, sabor e presença de O2 e CO2 no
sangue;
90

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 91/374
• Electromagnéticos: que respondem a um determinado comprimento deonda, portando à luz;
• Nocirreceptores: que respondem a estímulos dolorosos, por lesões nostecidos.
O processo de transdução de sinal em todos os receptores sensoriais envolve a
abertura ou fecho de canais iónicos que recebem directamente, ou via 2º mensageiro,informações acerca do ambiente externo ou interno.
(Abetura dos canais -> alteração do fluxo iónico -> alteração do potencial de acção ->potencial gradativo = potencial receptor -> receptor membranar gera correntes locais ->Abertura de canais iónicos voltagem-dependentes -> Potencial de acção)
Factores que controlam a magnitude do potencial receptor:
• Força do estímulo;• Velocidade de alteração da força;• Somação temporal;• Adaptação (diminuição da sensibilidade do receptor por diminuição da
frequência dos potenciais).
91

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 92/374
Rápida - Receptores fásicos (ex: corpúsculos de Vater-Pacini)
Lenta - Receptores tónicos (ex: receptor articular)
Constituição da unidade sensorial
A unidade sensorial é composta por um único neurónio aferente com todas as suasterminações receptoras.
A porção do corpo que quando estimulada leva à actividade do neurónio aferenteé chamada de campo receptivo ou sensorial.
92

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 93/374
A precisão ou acuidade com que um estímulo pode ser localizado e diferenciadode um estímulo adjacente depende da quantidade de convergência do input neural.
(Aumento convergência do input neural -> diminuição de precisão -> aumento tamanho)
Como as terminações dos receptores de diferentes neurónios se sobrepõem, umestímulo acciona mais que uma unidade sensorial.
Inibição lateral: a informação de neurónios aferentes cujos receptores estão naextremidade do estímulo é mais inibida que a informação que está no centro doestimulo.
Aspectos do estímulo que o sistema sensorial é capaz de reconhecer:
1) Tipo de estímulo ou modalidade (especificidade do receptor), unidadessensoriais adjacentes podem ser sensíveis a diferentes tipos de estímulo
2) Intensidade (frequência dos potenciais, somação espacial)3) Localização4) Duração do estímulo (capacidade de adaptação do receptor)
Caminhos Neurais do Sistema Sensorial
Os neurónios aferentes sensoriais formam uma cadeia que consiste em 3 ou maisneurónios ligados através de sinapses. Um conjunto de 3 neurónios paralelos forma umavia sensorial, que são também conhecidas como vias de ascenção pois levam ainformação ao cérebro.
Vias de ascenção especificas : levam informação de um tipo singular de estímulo; oneurónio final vai para as áreas sensoriais específicas do córtex cerebral; olfacto ->sistema límbico; somático -> córtex somatosensorial; olhos -> córtex visual ; ouvidos ->córtex auditivo; sabor -> não tem representação no córtex cerebral. As vias específicasvão para o lado do SNC oposto ao da localização dos receptores sensoriais.
93

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 94/374
Vias de ascenção não específicas: geram informação geral na medida em que sãoactivadas por unidades sensoriais de diferentes tipos; sendo os neurónios pertencentes aesta via denominados de neurónios polimodais.
Enquanto as específicas levam informação de um estímulo específico, singular; asnão específicas são activadas por unidades sensoriais diferentes e constituídas porneurónios polimodais.
Tipos de sensações
Somáticas Especiais
94
Tácteis
Cinestésicas
Térmicas
Visão
Audição
Olfacto

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 95/374
Sensações somáticas
São as sensações da pele, músculos, ossos, tendões e articulações, e são iniciadaspor receptores somáticos especializados. Os receptores são específicos para cadaestimulo; assim cada?! receptores para calor, frio, tacto, pressão, posição; movimento edor, sendo todos eles distintos.
*Tacto (pressão): mecanorreceptores de activação rápida (tacto, movimento e vibração)ou lenta (pressão)
*Postura e movimento: receptores de estiramento do fuso muscularem e respondem àmagnitude e à intensidade do estiramento.
*Temperatura: são os termorreceptores, sendo os de calor para temperaturas entre 30ºe 43º C enquanto os de frio respondem a pequenos decréscimos de temperatura.
*Dor: os nocireceptores respondem a calor excessivo, deformação mecânica intensa eagentes químicos possíveis de causar lesão tecidular, como por exemplo histamina.
95

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 96/374
Vias neurais do sistema somatosensorial:
Anterolateral: ou espinotalãmica. 1ª sinapse – neurónio receptor e neurónio localizadona substancia cinzenta da medula espinhal. 2ª sinapse – neurónio da massa cinzenta
com neurónio cortical. Esta via processa informação de dor e temperatura.
Coluna-dorsal: neurónios receptores projectam-se ate ao tronco cerebral, onde ocorre a1ª sinapse. A 2ª sinapse ocorre no tálamo onde as projecções são enviadas para o córtexsomatosensorial.
96

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 97/374
97

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 98/374
98

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 99/374
SENSAÇÕES ESPECIAIS
Visão
O olho é composto por uma parte óptica que foca a imagem visual nas célulasreceptoras e um componente neural que transforma a imagem visual num padrão de
potenciais gradativos de acção.
Um objecto é focado ao nívelda retina, uma fina camada de tecidonervoso que possui receptores,sensíveis à luz, e neurónios. Ocristalino e a córnea constituem aestrutura óptica que foca a luz e atransforma numa imagem na retina.A imagem é focada numa área
especializada – fovea central – que éa área da retina com maior claridadevisual. Todos os ajustamentos para adistância são feitos por alterações notamanho do cristalino – acomodação.O tamanho do cristalino é controladopor músculos circulares lisos, sendoestes controlados por nervos parassimpáticos. À medida que o músculo contrai, liberta atensão nas fibras zonulares, e á medida que relaxa, a tensão das fibras zonularesaumenta.
Para focar objectos ao longe, os músculos ciliares relaxam e o cristalino adquireuma forma oval. Para objectos ao perto, o músculo ciliar contrai e o cristalino adquire aforma circular. O cristalino divide o olho em duas partes: humor-aquoso (anterior) ehumor-vitreo (posterior) sendo estes 2 fluidos incolores responsáveis pela transmissãode luz da frente do olho à retina.
O humor aquoso é composto por tecido vascular especializado que cobre omúsculo ciliar, e produzido pelo corpo ciliar que o drena através do canal de Schlemm. A
quantidade de luz que entra no olho é controlada por músculos em forma de anel, detecido pigmentoso designado íris. O buraco no centro da íris, através do qual a luz entra,é a pupila. A íris é constituída por dois anéis de músculo liso com enervação autónoma. Aestimulação de nervos simpáticos na íris aumenta a pupila, levando à contracção dasfibras musculares rodais, enquanto que a estimulação parassimpática da íris causa adiminuição da pupila, relaxa o músculo. A luz brilhante causa uma diminuição dodiâmetro da pupila.
Células fotoreceptoras
99

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 100/374
Os fotoreceptores da retina possuem um segmento exterior, o disco, que mantéma substancia química que irá responder à luz.
Existem 2 tipos de fotoreceptores, os cones e os bastonetes, diferindo no tipo dedisco. A porção sensível à luz, do fotoreceptor, encontra-se afastada da luz que entra epor isso a luz tem que passar através da camada celular da retina, antes de chegar aofotoreceptor para o estimular. A coróide, camada pigmentar por trás da retina, absorve a
luz e previne a sua reflexão de volta aos cones e bastonetes.
Os bastonetes são extremamente sensíveis e respondem a níveis muito baixos deiluminação enquanto que os cones são menos sensíveis e respondem apenas a luz maisintensa.
Os fotoreceptores contêm moléculas designadas fotopigmentos, que têm comofunção absorver luz. O fotoreceptor é único pois é o único tipo de célula que estádespolarizada quando inactivada e hiperpolarizada em resposta à activação do estímulo.
Na ausência de luz, os canais Na+ da camada exterior do segmento são mantidos
abertos pela presença de elevada concentração de GMPc. A elevação do potencial demembrana abre canais de Ca2+; este causa a libertação de neurotransmissores(glutamato), que neste caso ocorre no escuro.
Vias neurais da visão
Começam nos cones e bastonetes -> Sinapsam uns com os outros e comneurónios de 2ª ordem -> Sinapsam com os neurónios que passam a informação de umlado para o outro da retina e células ganflionares -> As células ganglionares reagemdiferentemente à cor, intensidade e movimentos da imagem.
As células ganglionares são as 1as a responder à activação através da produçãode potenciais de acção. Os axónios destas células formam o out put da retina – nervoóptico. Os 2 nervos ópticos encontram-se na base do cérebro para formar o quiasmaóptico.
Estimulo luminoso
Cones e bastonetes
Células bipolares
Gânglios
100
Retina

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 101/374
Nervo óptico
tálamo
Tronco cerebral e cerebelo Núcleo supraquiásmico
Córtex visual Córtex associativo
101

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 102/374
102

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 103/374
103

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 104/374
104

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 105/374
Audição
Transmissão do som
1ª etapa: entrada das ondas sonoras para o canal auricular externo, no fundo doqual existe uma membran: membrana timpânica. Esta passagem permite a orientaçãodo som, aquecimento do ar e filtração de partículas. Esta membrana separa o canalauricular externo do ouvido médio, que consite numa cavidade no osso temporal repletade ar.
2ª etapa: transmissão de energia sonora da membrana timpânica através doouvido médio ate ao ouvido interno (cóclea). Esta passagem é em forma de espiral cheiade fluido, também no temporal. O líquido é mais difícil de mover que o ar, a pressãosonora tem que ser amplificada, o que vai ser conseguido pelo martelo, bigorna eestribo. Estes ossiculos recebem as vibrações e diferenciam-nas para a janela oval (quesepara o ouvido médio do interno).
No ducto coclear, que divide a códea, encontram-se os receptores sensoriais dosistema auricular; sendo que de cada lado do ducto estão compartimentos cheios de
105

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 106/374
liquido. A membrana basilar, onde se situa o órgão de Carti, contém as célulasreceptoras sensitivas. Estas, ciliadas, são mecanoreceptoes. Quando os cílios se dobram,canais de Ca2+ abrem-se, despolarizando a membrana e libertando neurotransmissores(glutamato) -> gera-se assim um potencial de acção ao nível dos neurónios.
Ouvido Humano
Orgão que contém os receptores do som
Não é capaz de detectar todas as frequências detecta apenas frequências entre 20 e20.000 Hz embora detecte melhor entre os 1.000 e 4.000 Hz
Campo Tonal: Frequências que o ouvido humano detecta (audiómetro)
O ouvido humano detecta também o timbre dos diferentes instrumentos e conseguedistinguir ± 400.000 sons diferentes
A audição biauricular permite também distinguir de onde partiu o som, porque o ouvidoé capaz de medir o tempo de chegada deste
FUNÇÕES DO OUVIDO EXTERNO
(PAVILHÃOAURICULAR E CANAL AUDITIVO)
-AMPLIFICAR E DIRECCIONAR O SOM
-AQUECER O AR
-PROTEGER DO PÓ, PÓLENS, INSECTOS (através do serumen e dos pêlos)
FUNÇÕES DO OUVIDO MÉDIO
(TÍMPANO E OSSÍCULOS)
- AMPLIFICAR O SOM (1,3 vezes)
- TRANSMITIR O SOM AO OUVIDO INTERNO
- PROTEGER O OUVIDO INTERNO DOS SONS MUITO INTENSOS (ar na caixa do tímpano econtracção reflexa dos músculos do ouvido médio - tensor do tímpano e estapédio
106

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 107/374
Equilíbrio
As alterações no movimento e posição da cabeça são detectadas pelas célulasciliadas no vestíbulo do ouvido interno. Este consiste em 3 canais semicirculares,membranas e 2 projecções em forma de sáculo.
As células receptoras dos canais semicirculares possuem cílios. Estes estãoenvolvidos pela cúpula, que os mantêm juntos, estendendo-se au longo do lúmen decada canal semicircular, na ampola.
Cada célula receptora tem uma direcção de libertação máxima deneurotransmissores e quando os cílios estão dobrados nesta direcção, a céluladespolariza.
107

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 108/374
108

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 109/374
Vias Nervosas do Equilíbrio
A informação proveniente do Sistema vestibular é conduzida pelo nervo auditivoao tronco cerebral e depois dirige-se a uma região vizinha do Cortex Somatosensorial nolóbulo parietal onde é integrada com a informação proveniente das articulações, tendõese pele dando lugar à sensação de postura e movimento
IMPORTÂNCIA DA FUNÇÃO VESTIBULAR
É usada para:
- Controlar os músculos do olho
- Regular os mecanismos reflexos que mantêm a postura erecta
- A locomoção e orientação da cabeça no espaço
-Tomar consciência da posição e aceleração do corpo
Gosto
Os quimioreceptores da boca respondem a alterações químicas do exterior,estando agrupados em papilas gustativas. As sensações de gosto estão divididas emquatro; doce, amargo, salgado e ácido. Para cada uma destas sensações de gosto háuma zona da língua correspondente.
SENTIDOS QUÍMICOS
Os receptores sensíveis aos químicos - Quimioreceptores
Existem quimioreceptores nos vasos sanguíneos sensiveis às PO2; CO2 e H+
Existem quimioreceptores sensiveis a agentes químicos externos – Sabor e olfacto
Função: Aumentar a salivação e a actividade do tracto gastrointestinal, aumentaro apetite, proteger das substâncias químicas perigosas (a estricnina e arsénio sãosubstâncias amargas)
109

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 110/374
A sensação do gosto está representada no Lobo Parietal perto da região querepresenta a boca no córtex somatosensorial.
Não se sabe como é possível distinguir tantos sabores com tão poucos tipos dereceptores
Existe uma ligação estreita entre o sentido do gosto e o do olfacto.
Quando estamos constipados perdemos o gosto
Olfacto
Os neurónios receptores do olfacto encontram-se no epitélio nasal, na partesuperior da cavidade nasal. Estes receptores sobrevivem apenas dois meses, sendoconstantemente renovados, por substituição, por novas células. Estes neurónios têmuma única dendrite que se estende até à superfície do epitélio.
Os axónios das células receptoras olfactivas sinapsam nas estruturas do cérebroconhecidas como bolbos olfactórios. A informação passa dos bolbos olfactórios para ocórtex olfactório e outras partes do sistema límbico.
110

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 111/374
O sangue e a hemostase
Funções do Sangue:
• Transporte de gases, nutrientes e metabolitos;• Transmissão de sinais por hormonas e citocinas;• Funcionar como tampão dos líquidos corporais, participando no equilíbrio
ácido-base;• Participar na defesa do organismo;• Promover a hemostase.
Constituintes:
Volume de sangue – 7/8 % do peso corporal +/- 5,5 l.
64% nas veias;
13% nas artérias;
7% nos capilares;
7% no coração;
9% na circulação pulmonar.
111

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 112/374
Uma parte está armazenada no baço, fígado, pulmão e tecido sub-cutâneo.
Plasma → 55%
Eritrócitos (hematócrito) → 45%
Plasma - composto por:
- Proteínas plasmáticas, que favorecem a absorção de fluído
extracelular para dentro dos capilares sendo elas albuminas
(as mais abundantes e são sintetizadas pelo fígado), globulinas
e fibrinogénio. Estas não são absorvidas pelas células, já que
as mesmas utilizam aminoácidos para sintetizar as suas
próprias proteínas.
- Electrólitos minerais: Na+; Ca2+; Cl-; HCO3-; H+; Mg2+; etc…
- Gases: CO2; O2; N2.
- Produtos do catabolismo celular: ureia, creatinina, ácido
úrico, bilirrubina, etc…
- Nutrientes: carbohidratos, lípidos, vitaminas e aminoácidos.
- Enzimas: factores de coagulação, factores do complemento.
- Água.
- Hormonas.
• Funções das proteínas plasmáticas:
I ) Transporte. ( ex.: transferrina ou lipoproteínas);
II ) Imunidade, imunoglobulinas. ( ex.: Igm e Iga);
III ) Manutenção da pressão osmótica, albumina;
IV ) Enzimas, renina, factores do complemento ou de coagulação;
V ) Tampão, todas as proteínas;
VI ) Inibidores das proproteares, λ1 – antitripsina.
Células sanguíneas
112

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 113/374
Eritrócitos: Têm como função principal o transporte de O2, que é recebido
nos pulmões e de CO2, produzido a nível celular. Possuem
hemoglobina à qual os átomos de ferro permitem a ligação
de O2, para ser transportado.
hemoglobina
A sua membrana plasmática possui polissacáridas específicas e proteínas quediferem de pessoa para pessoa.
A eficiência no transporte de O2 e CO2, está relacionada com a elevada relaçãosuperfície/volume que estas células possuem. São produzidas na medula óssea,diferenciando-se; já que os seus precursores (reticulócitos), produzem a hemoglobinaperdendo o núcleo e organelos.
O tempo médio de vida de um eritrócito ronda os 120 dias, sendo finalmentedestruídos no fígado e baço.
A produção de eritrócitos requer os nutrientes normais para a produção dequalquer célula assim como: Ferro, Ácido Fólico, Vitamina B 12 e alguns factores de
crescimento.
O produto da degradação da hemoglobina é a bilirrubina que é de novo posto emcirculação. É responsável pela cor amarela do plasma.
Ferro
É o elemento ao qual se liga o O2 na hemoglobina.
Pequenas quantidades são perdidas na urina, suor, fezes e células escamadas dapele.
Um desequilíbrio significativo nas [ ]’ s de Fe, leva a uma produção desadequadade hemoglobina ou a um excesso que pode ser tóxico.
O controlo Homeostático do Fe reside essencialmente no epitélio intestinal, que oabsorve directamente dos alimentos.
( ↓ Fe no organismo absorção intestinal de Fe )
113
+Fe + O2

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 114/374
O organismo possui uma reserva de Fe no fígado, no qual está ligado à ferritina.
Ferro + Ferritina (no Fígado).
A reciclagem do Ferro consiste na recuperação do ferro proveniente de eritrócitosem processo de destruição, sendo depois libertados para o plasma onde se vai ligar atransferrina, uma proteína de transporte, que vai levar de novo ao local de produção deeritrócitos (medula óssea).
Ácido Fólico e vitamina B 12
O Ácido Fólico é uma vitamina requerida para a síntese da Timina (basenúcleofílica). Quando não está presente nas quantidades adequadas ocorre divisãocelular descontrolada, especialmente nas células precursoras de eritrócitos.
A vitamina B 12 também é necessária para a produção de eritrócitos, na medidaem que a actuação do ácido fólico está dependente desta.
A absorção de vitamina B 12 ocorre no tracto gastrointestinal e está dependenteda existência de: factor intrínseco (secretada pelo estômago). Sendo assim:↓ factorintrínseco → ↓vitamina B 12 → ↓ eritrócitos Anemia
Regulação da produção de Eritrócitos
O controlo directo da produção de Eritrócitos (entropoiese), é iniciada por umahormona (eritropoietina), secretada nos rins. Esta actua na medula óssea para estimulara proliferação de células precursoras de eritrócitos e a sua diferenciação em eritrócitosmaduros.
A eritropoietina é normalmente secretada em pequenas quantidades. A suasecreção é diminuída quando ocorre decréscimo do oportede O2 aos rins. Isto aconteceem situações de anemia, altitude elevada, bombeamento insuficiente de sangue para ocoração e doença pulmonar. Como resultado do aumento da secreção de eritropoitina, asua [ ] plasmática aumenta e a capacidade de transporte de O2 também.
114

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 115/374
Leucócitos: são classificados de acordo com a sua estrutura e afinidade.
São todos produzidos na medula óssea.
Monócitos Linfócitos Granulócitos polimorfonucleares
(1) (2) (3)
Neutrófilos Basófios Eosinófilos
(1) São um pouco maiores que os granulócitos, têm núcleo oval ou em forma deferradura e poucos grânulos citoplasmáticos.
(2) Têm pouco citoplasma e núcleos grandes.
(3) Têm núcleos multilobados e abundantes grânulos citoplasmáticos. Diferemquanto ao tipo de corante capaz de os corar. (pH ≠)
Plaquetas: São incolores e muito mais pequenas que os eritrócitos.
São produzidas a partir dos megacarióticos e têm como função acuagulação sanguínea.
Regulação da produção de células sanguíneas
Todas as células sanguíneas são produzidas a partir de células pluripotentes,sendo estas indifrenciadas e capazes de originar percursores de qualquer célulasanguínea.
115

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 116/374
Quando uma célula pluripotente se divide pode originar duas células filhastambém pluripotentes, ou restringi-las a uma via de desenvolvimento em particular.Assim, as células originadas podem ser células percursoras de linfócitos ou célulasmielóides, podendo estas últimas originar todas as outras células.
A proliferação está dependente de um grande número de proteínas hormonais eagentes parácrinos designados Factores de Crescimento Hematopoiético.
= Hemastase = Prevenção da perda de sangue
Quando um vaso sanguíneo é lesionado a sua resposta imediata é a constrição,diminuindo o fluxo sanguíneo na área afectada.
O estancamento da perda de sangue depende de dois outros processos que sãoindependentes e ocorrem numa sucessão rápida:
1) Formação do rolhão plaquetário;
2) Coagulação.
116

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 117/374
1) Rolhão plaquetário: Bloqueador Primário
O envolvimento de plaquetas na hemostase requer a sua adesão a uma superfície.A lesão num vaso, rompe o endotélio e expõe o tecido conjuntivo de colagénio que estáem baixo. As plaquetas aderem ao colagénio através de um intermediário – Factor deVan Willebrand – uma proteína plasmática secretada por células endoteliais e plaquetas,
que se liga ao colagénio exposto, altera a sua conformação e torna-se favorável para aligação das plaquetas.
A adesão das plaquetas ao colagénio acciona a libertação do conteúdo dasvesículas secretoras das plaquetas que vão induzir inúmeras alterações na morfologia emetabolismo das plaquetas ------ Activação das Plaquetas
Algumas destas alterações causam a aderência de novas plaquetas às maisantigas, um feedback positivo (+) designado de Agregação de Plaqueta, querapidamente cria uma camada de plaquetas dentro do vaso.
A adesão das plaquetas induz a síntese de trombaxina Az na membranaplasmática das plaquetas. Esta é libertada no fluído extracelular e actua localmente paraposteriormente estimular a agregação das plaquetas e a libertação do conteúdo dasvesículas secretadas. Estas vesículas possuem uma variedade de agentes químicos entreeles: ADP e serotonina, que contribuem para a activação plaquetária e tambémagregação.
O fibrirogénio tem também uma função importante na agregação das plaquetas jáque é responsável pela formação de pontes entre os agregados de plaquetas. Osreceptores para esta proteína tornam-se activos durante a activação das plaquetas.
A eficácia do rolhão plaquetário é aumentada pela capacidade de contracção dasplaquetas, isto porque as plaquetas quando se encontram em agregado são estimuladaspara que a sua aumente a sua concentração em actina e miosina, resultando isto numacompressão do rolhão.
Agregação [actina e missiva] compressão eficácia do rolhão
Enquanto o rolhão se forma há vasoconstrição, fazendo com que o fluxo sanguíneoseja menor. Este processo é mediado pela tromboxina A2 em resultado há contracção domusculo liso do vaso.
O rolhão não se estende ao longo de todo o endotélio já que as células endoteliaissintetizam prostaciclina, que é um inibidor da adesão plaquetária, da sua activação eagregação.
Assim, as plaquetas produzem tromboxina A2 a partir do ácido aroquidónico, e ascélulas endoteliais convertemos intermediários formados em prostaciclina.
117

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 118/374
As células endoteliais também libertam óxido nídrico, que para além de servasodilatador, inibe a activação e adesão das plaquetas.
2) Coagulação Sanguínea: Hemostase Secundária
Coagulação – transformação do sangue num gel sólido designado de coágulo outrombo, constituído essencialmente por fibrina. O processo de coagulação ocorre à voltado rolhão plaquetário e é a defesa homeostática dominante.
Tem como funções suportar e reforçar o rolhão para solidificar o sangue que semantém no canal lesado.
Os eventos que levam à coagulação são iniciados quando a lesão num vaso rompeo endotélio e permite ao sangue contactar com o tecido adjacente. Este contacto iniciauma sequência “em cascata” de activação química.
Trombina – Estabelece um feedback positivo na sua própria formação: a formaçãode trombina promove a formação de mais.
Neste processo muitos eritrócitos e outras células são aprisionadas na rede defibrina, sendo este o elemento fundamental do coágulo, já que este se forma mesmo naausência de elementos celulares (excepto plaquetas – já que as reacções da cascataocorrem sobre as plaquetas activadas, logo em plaquetas não há coagulação).
O Ca2+ é muito importante na cascata de coagulação, sendo que [Ca2+] no
plasma nunca pode ser muito baixa pois ocorreria morte por paralisia muscular.
118

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 119/374
A cascata de coagulação inicia-se na lesão tecidular e ocorre até à conversãoprotrombina trombina. Existem duas vias que conduzem a esta reacção:
I) Intrínseca
II) Extrínseca (que é a mais usual para iniciar a coagulação)
Em condições fisiológicas normais, as duas vias ocorrem sequencialmente.
Teoricamente, a coagulação pode ser iniciada pela activação do factor XII, ou pelaformação do complexo factor de tecido + factor VII a. As duas vias convergem no factorX a, que cataliza a formação de trombina, a partir da protrombina, que leva à formaçãode fibrina.
A trombina também contribui para a activação dos factores VIII, V a e X, além deactivar plaquetas.
A via extrínseca, através da formação de pequenas quantidades de trombina,providencia a activação da via intrínseca, além de facilitar o processoprotrombina trombina pela activação do factor V e activação das plaquetas.
NOTA: O fígado éum órgão importantepara a cuagulação namedida em que produzvitamina K, e esta énecessária para produzirprotrombina e outrosfactores de coagulaçãoplasmáticos.
119

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 120/374
*Sistemas de Anticoagulação *
A libertação de prostaciclina e óxido nítrico por células endoteliais inibe aagregação de plaquetas. Para além destes, o corpo tem outros mecanismospara limitar a formação de coágulos e para dissolver o coágulo após a suaformação.
Há três mecanismos que se opõem à formação de coágulos:
1º Mecanismo: Actua durante o início da fase de coagulação e utiliza a proteínaplasmática “inibidor da via do factor de tecido” (TFPI), que é secretadoprincipalmente pelas células endoteliais. O TFPI liga-se ao complexo factor detecido – factor VII a, e inibe a capacidade destes para gerar factor X a.
2º Mecanismo: É accionado pela trombina, que se liga a um receptor dascélulas endoteliais (trombomodulina) eliminando todos os efeitos de produçãode coágulos da trombina. Além disto causa a ligação da trombina à vitamina C,que inactiva os factores VIII a e V a.
3º Mecanismo: A antitrombina III, que inactiva a trombina e outros factores decoagulação, é activada através da sua ligação à heporina. Previne que ocoágulo se espalhe através da inactivação de factores de coagulação que sãolevados para longe do local lesado.
Sistema Fibrinolítico ou (trombolítico)
É o principal responsável pela remoção do coágulo, sendo constituído por umaenzima plasmática, o plasminogénio, que pode ser activada a plasmina através dasproteínas activadoras de plasminogénio. A plasmina digere a fibrina, dissolvendo o
coágulo.
120

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 121/374
Este sistema é constituído por diferentes tipos de activadores deplasminogénio e vias para os gerar( assim como para os inibir).
O mais importante é o activador de plasminogénio t-PA, que é secretado porcélulas endoteliais. A ligação do t-PA é crucial já que esta é uma enzima muito fraca naausência da fibrina.
121

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 122/374
Fisiologia do Sistema Cardiovascular
O músculo cardíaco
As células musculares do miocárdio estão arranjadas em camadas firmementeligadas entre si e rodeadas por espaços cheios de sangue. Este músculo é umacombinação entre o músculo esquelético e o liso, na medida em que as células sãoestriadas, como resultado de um arranjo dos filamentos grossos de miosina e finos deactina semelhantes aos do músculo esquelético. No entanto, as fibras cardíacas sãoconsideravelmente mais curtas que as esqueléticas, embora sofram ramificações.
As células adjacentes estão unidas pelas extremidades, por estruturasdenominadas discos intercalares. Adjacentes aos discos intercalares estão junções GAP,semelhantes às da maioria do músculo liso.
Cerca de 1% das células cardíacas não funciona na contracção, mas têmcaracterísticas especializadas essenciais para a excitação cardíaca normal, constituindoo sistema de condução do coração. Este sistema é responsável por iniciar o batimentocardíaco e disseminar rapidamente o impulso por todo o corpo.
Relativamente à inervação do músculo cardíaco, este é inervado por fibrassimpáticas e parassimpáticas, sendo estas últimas as fibras do nervo vago. As fibras pós-ganglionares simpáticas libertam norepinefrina enquanto as parassimpáticas libertamacetilcolina. Os receptores de norepinefrina no coração são principalmente beta-adrenérgicos, enquanto os receptores para a acetilcolina são muscarínicos.
Para além disto, é importante dizer que o coração secreta uma hormona peptidica(o péptico natriuretico) sendo secretado nas células atrios.
Por último, o suprimento sanguíneo ao coração faz-se através da circulaçãocoronária, na medida em que o sangue que está a ser bombeado não troca nutrientesnem produtos metabólicos com o coração.
Propriedades Globais do Coração:
-Automatocidade
- Ritmicidade- Excitabilidade
122

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 123/374
- Condutibilidade
- Contractilidade
Propriedades da Função Cardíaca
- Bomba
Propriedades mecânicas
Avaliação funcional
- Músculo
Propriedades eléctricas
ECG
Noções de pressão, circulação e resistência
1) O sangue circula sempre de uma região com pressão maior para outra de pressãomenor
2) A pressão exercida por um fluido é designada pressão hidrostática3) A resistência é a medida de fricção que impede a circulação
4) A circulação é directamente proporcional à diferença de pressão entre dois pontose inversamente proporcional à resistência
123

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 124/374
Noções de anatomia:
1) Entre o pericárdio e o coração existe um fluido que serve de lubrificante para osmovimentos do coração.
2) A válvula auricular direita é a tricúspide e a esquerda é a bicúspide, sendo que oprocesso de abertura/fecho das válvulas é passiva e resulta das diferenças depressão (o mesmo para as semi-lunares)
3) Quando a pressão intra-auricular é superior à intraventricular a válvula abre,enquanto se for inferior, fecha
4) As válvulas estão ligadas a músculos papilares das paredes ventriculares porcordas tendinosas
Coordenação do batimento cardíaco
O coração é uma bomba dupla em que os lados esquerdo e direito bombeiamsangue separadamente, mas no entanto, simultaneamente nos circuitos sistémico epulmonar.
A contracção do músculo cardíaca é iniciada pela despolarização da membrana. As junções GAP, que conectam as células miocardíacas permitem que o potencial de acçãose propague de uma célula para a outra.
A despolarização começa num pequeno grupo de células condutoras denominadaspor Nódo Sinoauricular (SA), localizado na aurícula direita, perto da entrada da veia cava.
O PA estende-se do nodo SA através da aurícula e depois dos ventrículos.
124

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 125/374
Sequência de Excitação
O Nodo SA é um normal pacemaker para todo o coração. A sua despolarizaçãogera o PA que leva á despolarização de todas as outras células cardíacas, determinandoo número de vezes que o coração contrai por minuto.
A condução através das células musculares auriculares é suficientemente rápidapara que as duas aurículas sejam despolarizadas e contraiam ao mesmo tempo.
O alastramento do PA para os ventrículos envolve uma porção de sistemacondutor, a nodo ariculoventricular (AV) que está localizado na base da aurícula direita.
A progressão do PA através das células da aurícula direita causa a despolarizaçãodo nodo AV. A propagação do PA neste nodo é relativamente lento resultando numatraso que permite que a aurícula contraia completamente antes da excitaçãoventricular ocorrer.
Depois de deixar o nodo AV, o impulso entra no sangue interventricular que possuifibras condutoras designadas de Feixe de His. As aurículas estão completamenteseparadas dos ventrículos por uma camada de tecido conjuntivo não condutor, excepto onodo AV e o feixe de His que constituem a ligação eléctrica.
O feixe de His divide-se em feixe esquerdo e direito, os quais penetram a paredeventricular e entram em contacto com as fibras de purkinge, células condutoras querapidamente distribuem o impulso através dos ventrículos. As fibras de Purkinge entramdepois em contacto com as células miocardicas.
125

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 126/374
Potenciais de acção cardíacos e excitação do nodo SA
A membrana em repouso é mais permeável ao K+ do que ao Na+. A fase dedespolarização do PA é devido principalmente à abertura dos canais voltagemdependentes do Na+. A entrada de Na+ despolariza a célula e provoca a abertura demais canais de Na+ (feedback +), a permeabilidade do K+ diminui à medida que oscanais de K+ fecham o que também contribui para a despolarização da membrana. Noentanto, a redução da permeabilidade do Na+ não é acompanhado pela repolarização damembrana. Esta mantém-se despolarizada no nível 0mv, isto porque:
1) Os canais K+ mantêm-se fechados
126

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 127/374
2) Há um aumento na permeabilidade do Ca2+
Nas células miocardiacas, a despolarização da membrana causa a abertura decanais Ca2+, o que resulta num fluxo de Ca2+ para dentro da célula a favor dogradiente electroquímico. Estes canais abrem mais lentamente que os canais Na+ emantêm-se abertos por mais tempo.
O fluxo de Ca2+ para dentro da célula contra-balança o fluxo de iões K+ para forada célula e mantém a membrana despolarizada.
A repolarização da célula ocorre com as permeabilidades voltam ao seu estadonormal.
Os potenciais de acção para as células musculares auriculares são idênticos, sóque a fase de despolarização é menor.
Em contraste existem diferenças entre os PA das células musculares do miocárdio
e as do sistema condutor.
As células do nodo SA não têm um potencial de repouso estável e são submetidasa uma despolarização lenta: potencial pacemaker.
Há 3 mecanismos de canais iónicos que contribuem para o potencial pacemaker:
1) Redução progressiva da permeabilidade do K+: os canais de K+ que estão abertosdurante o PA anterior fecham gradualmente.
2) As células pacemaker têm um leque de canais iónicos que abrem quando está avalores negativos. Estes canais conduzem para dentro correntes despolarizantesde Na+
3) Os canais pacemaker são tipo de canais de cálcio que abrem durante poucotempo mas que contribuem para o influxo de Ca2+ (canais tipo T)
O potencial pacemaker providencia ao nodo SA automaticidade: capacidade de auto-excitação espontânea e rítmica.
Diversas porções do sistema condutor são capazes de gerar potenciais pacemakermas são lentos: pacemakers ectópicos.
Electrocardiograma (ECG)
Os potenciais de acção em músculos cardíacos podem ser vistos como bateriasque causam o movimento da carga através dos fluidos corporais. As cargas emmovimento são causadas por todos os PA que ocorrem simultaneamente em célulasmiocardicas e podem ser detectadas ao colocar eléctrodos na superfície da pele.
127

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 128/374
A 1ª variação, onda P, corresponde ao fluxo de corrente durante a despolarizaçãoauricular. A 2ª variação, complexo QRS, é o resultado da repolarização ventricular.
A forma e o tamanho das ondas varia com a localização dos eléctrodos.
O ECG é uma medida das correntes geradas no fluido extracelular pelas alteraçõesocorridas simultaneamente em muitas células cardíacas. Este apenas providenciainformações relativas à actividade eléctrica do coração.
Acoplamento excitação-contracção no músculo cardíaco
128

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 129/374
O mecanismo que associa a excitaçãocom a contracção do coração é um aumentona concentração citosólica de Ca2+, devido àlibertação deste no reticulo sarcoplasmático.
O Ca2+ liga-se com a troponina eforma-se pontes cruzadas entre a actina e amiosina:
1) O PA no tubulo T abre canais de voltagemsensíveis ao Ca2+ na própria membrana dotúbulo (canais tipo2), o Ca2+ difunde dofluido extracelular através dos canais paradentro, causando um pequeno aumento na
concentração citosólica de Ca2+ que estáimediatamente adjacente ao reticulosarcoplasmático.
2) O Ca2+ liga-se a receptores de Ca2+ namembrana reticular
3) A activação dos receptores abre os canaisde Ca2+ intrínsecos e ocorre uma grandedifusão de Ca2+ +ara o citosol
A contracção termina quando a concentração citosólica de Ca2+ é restaurada portransporte activo de Ca2+ para dentro do reticulo (bomba Ca2+-ATPase). A pequenaquantidade de Ca2+ que tinha entrado aquando da excitação é activamentetransportada para fora da célula, para que a concentração de Ca2+ total se mantenhaconstante.
A quantidade citosólica de Ca2+ que aumenta durante a excitação é a maiordeterminante da força de contracção.
129

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 130/374
Período refractário do coração
O músculo ventricular é incapaz de atingir qualquer grau de somação decontracções. A incapacidade que o coração tem de gerar contracções tetânicas é oresultado de longos períodos refractários.
O período refractário absoluto dura quase toda a contracção e o músculo não podeser re-excitado.
Eventos mecânicos do ciclo cardíaco
130

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 131/374
--Sístole: período de contracção ventricular e ejecção de sangue
-Diástole: relaxamento ventricular e enchimento de sangue
Durante a primeira parte da sístole, os ventrículos estão em contracção, mas asválvulas estão fechadas – contracção ventricular isovolumétrica, já que o volume
ventricular é constante e as fibras musculares não encurtam.
Quando a pressão nos ventrículos exceder a pressão na aorta e tronco pulmonar,as válvulas abrem e a ejecção ventricular ocorre.
Durante a primeira parte da diástole, os ventrículos relaxam e as válvulas fechammantendo-se o volume ventricular – período de relaxamento ventricular isovolumétrico.
As válvulas auriculo-ventricular abrem e o enchimento dos ventrículos começa. Acontracção auricular ocorre no fim da diástole depois de quase todo o sangue terpassado.
131

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 132/374
Diástole Média e Diástole Tardia
Perto do fim da diástole, o nodo SA recebe o impulso e a aurícula despolariza econtrai.
A quantidade de sangue no ventrículo no fim da diástole é designada de volume
diastólico final (EDV).
Sístole
O ventrículo não esvazia completamente. A quantidade de sangue que permaneceapós a ejecção é designada de volume sistólico final (ESV).
Output Cardíaco – Débito Cardíaco
O volume de sangue bombeado por cada ventrículo, por minuto, é designado deoutput cardíaco. É também o volume de sangue que passa quer no circuito sistémicoquer no pulmonar, por minuto.
132

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 133/374
Controlo do Ritmo Cardíaco
O batimento rítmico docoração ocorre na ausência total dequalquer influência nervosa ouhormonal sobre o nodo SA.
Neste nodo, terminam fibrassimpáticas e parassimpáticas. Aactividade do parassimpático causaum decréscimo do ritmo cardíacoenquanto a actividade do simpáticoaumenta. Assim, é lógico que emrepouso haja mais actividadeparassimpática no coração.
A estimulação simpática aumenta o potencial pacemaker, aumentando ascorrentes de Na+ e de Ca2+ tipo T, fazendo com que as células do nodo atinjam o limiar
mais rapidamente.
133

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 134/374
A estimulação do parassimpático (medida por receptores muscarínicos do tipo M2)tem o efeito oposto, hiperpolarizando a membrana das células do nodo SA através doaumento da permeabilidade ao K+ (por diminuição do AMPc celular o que atrasa acorrente de Ca2+).
A epinefrina aumenta o ritmo cardíaco, actuando nos mesmo receptores beta-adrenergicos que a norepinefrina, ao aumentar a entrada de Ca2+.
Controlo do Volume Bombeado
As alterações na força de contracção podem ser produzidas por 3 factoresdominantes.
1) Alteração no volume do fim da diástole (pré-carga)
2) Alterações na magnitude dos inputs do SNSimpático aos ventrículos
3) Pressões arteriais contra os quais os ventrículos bombeiam (pós-carga)
O volume bombeado aumenta à medida que o volume de fim de diástole aumenta:Mecanismo de Frank-Starling (envolve o recrutamento desigual de sarcómeros, forçandoprincipalmente e progressivamente os sarcómeros curtos à medida que aumenta ovolume de enchimento.
134

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 135/374
Fisiologia Vascular
Compliance – termo utilizado para quantificar a facilidade com que uma estrutura éesticada, ou seja, quanto maior a compliance de uma estrutura mais facilmente ela podeser esticada.
Resistência – razão pela qual a pressão sanguínea não é igual ao longo de todos os vasos(vasos+pequenos -> maior resistência à passagem do sangue -> menor pressão)
Sistema Vascular
Componente comum: células endoteliais (junções activas)
- barreira permeável permite a troca de nutrientes
- sintetiza hormonas activas a partir de percursores inactivos
- mediador de angiogénese
- contribui para a formação e manutenção da matriz-extracelular
Pressão Arterial (características)
1) Nunca chega a zero (o sangue não sai todo das artérias durante a sístole, apenas1/3)
2) Pressão arterial máximo = Pressão sistólica (SP) -> Ejecção ventricular maior3) Pressão arterial mínima = Pressão diastólica (DP) depois da ejecção4) Normotensão: SP/DP = 125/75 mmHg5) Pressão de pulso (SP-DP=PP) (125-75=50 mmHg) influenciada por:
5.1) Volume ejectado
5.2) Velocidade de ejecção
135

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 136/374
5.3) Compliance arterial (variação de volume/variação de pressão)
6) Pressão arterial média (MAP=DP+1/3(SP-DP))
6.1) Não é influenciada pelo compliance
6.2) É a que leva o sangue a todos os tecidos e órgãos
7) Á medida que o sangue deixa as artérias, o volume arterial e a pressão arterialbaixam, ocorrendo a próxima contracção ventricular quando existe sangue suficiente nasartérias para os distender parcialmente.
Artérias – responsáveis pela condução do sangue para os diferentes órgãos; tubos debaixa resistência; reservatórios de pressão pois mantêm o fluxo através dos tecidosdurante a diástole
Arteríolas – vasos de menor diâmetro e maior resistência; responsáveis peladeterminação do fluxo (F) de sangue em órgãos individuais (F=MAP/resistência do órgão)
NOTA: O padrão de distribuição do fluxo sanguíneo depende do grau de contracção domúsculo liso da arteríola de cada órgão ou tecido
136

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 137/374
Alterações na Resistência
O músculo liso das arteríolas possui um tonus intrínseco ou basal que gera acontracção independente. No entanto, este tonus pode ser aumentado ou diminuidoatravés de neurotransmissores, que induzem uma alteração na concentração citosólicade Ca2+ das células musculares.
Aumento da força contráctil -> Tonus Intrínseco -> Vasoconstrição
Diminuição da força contráctil -> Tonus Intrínseco -> Vasodilatação
Mecanismos de controlo da vasoconstrição/vasodilatação
1) Controlo local
2) Controlo extrinseco (reflexo)
1) Controlo local da vasocontracção e vasodilatação:
- mecanismo independente de nervos e hormonas; inclui alterações causadas poragentes autócrinos e/ou parácrinos;
- quatro fenómenos: Hiperemia Activa, Autoregulação do fluxo, Resposta local a lesões eHiperemia Reactiva.
Hiperemia Activa – resultado directo da dilatação arteriolar nos órgãos e tecidos maisactivos, causada por alterações químicas no fluido extracelular que envolve as arteríolas,resultantes da intensa actividade metabólica das células.
Aumento da actividade metabólica -> diminuição da concentração de 0xigénio ->aumento dos metabolitos -> dilatação da arteríola -> aumento da entrada de sangue noórgão
137

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 138/374
Alterações químicas do Fluído Extracelular:
- aumento da concentração de oxigénio, usado na produção de ATP
- aumento de CO2, produto final do metabolismo oxidativo
- diminuição do pH
- aumento da concentração de adenosina, resultante da quebra de ATP
- iões K+ acumulados, resultantes de potenciais de acção repolarizantes acumulados
-> São estas alterações no fluido extracelular que vão actuar no músculo liso arteriolarcausando relaxamento.
Esta Hiperémia Activa ocorre:
Musculo esquelético; musculo cardiaco; glândulas e tecidos com maior actividademetabólica
Autoregulação do fluxo
- alterações locais mediadas na resistência arteriolar também podem ocorrer quando umtecido ou um órgão sofre alteração no seu fornecimento de sangue, resultante de umaalteração na pressão sanguinea
138

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 139/374
Aumento da pressão arterial do órgão -> diminuição de sangue para o órgão ->diminuição da concentração de oxigénio -> aumento de metabolitos -> diminuição daaderência ao órgão pelas veias -> dilatação arteriolar no órgão
-> Estas alterações visam manter o fluxo sanguíneo constante perante alterações depressão.
Os mecanismos de autoregulação são os mesmo que para a hiperémia activa.Para além disto, verifica-se também a existência de respostas miogénicas (respostasdirectas do musculo liso arteriolar se tornar mais ou menos esticado) e devem-se aalterações no movimento de Ca2+ para dentro das células musculares.
Hiperemia Reactiva
- deve-se ao facto de um órgão ou tecido ter tido o seu fornecimento de sanguecompletamente ocluso, sendo que quando a oclusão é aberta ocorre um aumentoprofundo do fluxo sanguíneo.
-> Durante o período em que não existe fluxo sanguíneo as arteriolas dilatam (no órgãoafectado)
Resposta à lesão
- a lesão de um tecido causa a libertação de várias substâncias a partir das células. Estassubstâncias fazem o músculo liso arteriolar relaxar e causam a vasodilatação na árealesada.
2) Controlo Extrínseco
Nervos simpáticos
– libertam principalmente norepinefrina que se liga a receptores alfa-adrenergicos nomusculo liso vascular de modo a causar a vasoconstrição. Também podem provocarvasodilatação na medida em que descarregam a uma velocidade intermédia que variade órgão para órgão, causando sempre alguma constrição tónica, sendo a dilatação
conseguida quando a actividade simpática desce abaixo do nível basal. Além disto, osnervos simpáticos afectam também a força e frequência de contracção por actuação aonível dos receptores beta-1.
139

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 140/374
As funções primárias destes nervos não estão relacionadas com a coordenação denecessidades metabólicas locais e fluxo sanguíneo, mas com reflexos que regulam apressão sanguínea arterial.
Nervos autónomos não-colinérgicos e não-adrenérgicos
- estes (pós glanglionares) libertam óxido-nitrico (vasodilatador). Têm um papelimportante no controlo dos vasos sanguíneos gastrointestinais. Estes neurónios irrigamtambém arteriolas do pénis, onde são responsáveis pela erecção
Hormonas
Epinefria:
- receptor alfa-adrenérgico – vasoconstrição
- receptor 2-beta-adrenérgico – vasodilatação (apenas no musculo esquelético)
Angiotensina II -> vasoconstrição
Vasopressina -> vasoconsrição porque aumenta a absorção de água; é libertado pela
hipófise posterior
Peptido-auriculo-natriurético -> vasodilatador porque diminui a pressão e aumenta aexcreção de Na+ na urina; é secretado pela aurícula cardíaca
Células endoteliais e músculo liso vascular
Certas substâncias actuam directamente no músculo liso arteriolar enquanto
outras actuam indirectamente via células endoteliais adjacentes ao músculo. As célulasendoteliais em resposta a estas substâncias secretam diversos agente parácrinos quedifundem para o musculo liso vascular adjacente e induzem tanto o relaxamento como acontracção.
- Óxido Nítrico: libertado pelas células endoteliais. Também é conhecido como factorrelaxante derivado do endotélio (EDRF). É libertado continuamente para a vasodilataçãoarteriolar basal
140

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 141/374
- Eicosanóide prostacidina (PGI2): é vasodilatador, com pouca secreção, sendo estaaumentada em resposta a inputs
- Endotelina I (ET1) – é vasoconstritor actuando como agente parácrino. No entanto, emalgumas circunstâncias, pode chegar a concentrações suficientemente altas no sanguepara funcionar como hormona
-> A resposta exercida na superfície interna da parede arterial (nas células endoteliais)pelo fluxo de sangue é designado por “shear stress”; esta aumenta à medida que o fluxode sangue nos vasos aumenta. Como resposta a este “shear stress” intenso, o endotélioarterial liberta PGI2, óxido nítrico e menos ET-1; relaxamento do músculo liso ->dilatação
Capilares
Angiogénese: desenvolvimento e crescimento dos capilares sanguíneos. Neste processo,as células endoteliais desempenham um importante papel na construção de uma novarede de capilares através da locomoção celular e divisão. São estimuladas por factoresangiogénicos secretados localmente por diversas células do tecido e pelas própriascélulas endoteliais.
Anatomia da rede de capilares
141

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 142/374
As células achatadas que constituem o tubo endotelial não estão ligadasfirmemente umas às outras, mas estão separadas por pequenos espaços cheios de água,designados de fendas intercelulares.
As células endoteliais geralmente possuem um grande número de vesículasendocíticas e exocíticas que se podem fundir para formar canais de vesículas.
A vasodilatação das arteríolas que fornecem os capilares causa um aumento nofluxo dos capilares enquanto a vasoconstrição arteriolar reduz o fluxo capilar.
Metarteriolas conectam as arteríolas com vénulas, não passando o sanguedirectamente para os capilares. Possuem células musculares lisas. O sítio em que oscapilares deixam as metarteriolas é envolvido por um anel de músculo liso (esfíncter pré-capilar) que contrai e nela em resposta a factores metabólicos locais.
Velocidade do fluxo sanguíneo nos capilares:
O fluxo lento através dos capilares, maximiza o tempo disponível para a troca desubstâncias ente o sangue e o fluido intersticial.
A velocidade do fluxo não depende da proximidade do coração, mas sim da áreade seccionamento do vaso.
Como os capilares estão muito próximos oferecem uma resistência considerável
ao fluxo, mas o elevado número de capilares providencia uma área de seccionamentotão grande que a resistência total de todos os capilares é muito menor do que a dasarteríolas.
Difusão através da parede capilar (troca de nutrientes e produtos finais do metabolismo)
Há 3 mecanismos básicos para que as substâncias que se movem através dasparedes dos capilares entrem e saiam do fluido intersticial:
1. Difusão: principal2. Transporte vesicular3. Fluxo de volume
Nos capilares do cérebro há um 4.º mecanismo: Transporte mediado, porque oscapilares do cérebro não têm fendas intercelulares (apenas tight junctions).
Nota: Se um tecido necessita de aumentar o seu metabolismo tem de obter maisnutrientes do sangue e eliminar mais produtos do metabolismo: Hiperemia activa eAumento do gradiente de difusão entre o plasma e o tecido (aumento da utilização de O2
142

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 143/374
e nutrientes e diminuição da [] de O2 e nutrientes no tecido, aumento da produção deCO2 e outros metabolitos, aumento das diferenças de [] transcapilares e aumento dedifusão).
Fluxo de Volume de Plasma: Distribuição do fluido extracelular
Normalmente existe 3 vezes mais fluido intersticial do que plasma e funcionacomo um reservatório que pode fornecer ou receber fluido do plasma.
O fluxo de volume de plasma livre de proteínas ocorre simultaneamente à troca denutrientes, O2 e produtos metabólicos por difusão. Este fluxo de volume não tem comoobjectivo na troca de nutrientes mas sim na distribuição do fluido extracelular, sendoeste composto pelo plasma e fluido intersticial.
A parede capilar é altamente permeável à água e a quase todos os solutosplasmáticos. Por isso, na presença de diferenças de pressão hidrostática, a parede docapilar comporta-se como um filtro poroso através do qual o plasma livre de proteínas semove através de um fluxo de volume de plasma capilar para o fluido intersticial atravésdos canais cheios de H2O.
A magnitude do fluxo de volume é determinado pela diferença entre a pressãosanguínea no capilar (aumenta) e pressão hidrostática do fluido intersticial (diminui).
O plasma não filtra todo para o espaço intersticial porque a força osmótica se opõeà filtração. A diferença na concentração proteica entre o plasma e o fluido intersticialsignifica que a [] de H2O no plasma é menor, induzindo o fluxo osmótico docompartimento intersticial para dentro dos capilares.
Plasma capilar e fluido intersticial no exterior possuem grandes quantidades desolutos penetrantes de baixo peso molecular: cristalóides, Na+, Cl- e glucose, sendo a []destes semelhante nos 2 lados já que os capilares são porosos, nem causam alterações
significativas na [] de H2O.
Em contraste, proteínas plasmáticas não penetrantes têm uma [] baixa no fluidointersticial; assim a diferença na [] proteica entre o plasma e o fluido intersticial significaque a [] de H2O é menor no plasma que no fluido intersticial.
A pressão de filtração de rede depende da soma de 4 variações:
- Pressão hidrostática capilar, Pc, movimento do fluido para fora dos capilares;
- Pressão hidrostática intersticial, PI, movimento de fluido para dentro dos capilares;143

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 144/374
- Força osmótica devido à [] das proteínas, πp, favorece o movimento do fluido paradentro do capilar;
- Força osmótica devido à [] proteica do fluido intersticial, πI, favorecendo o movimentodo fluido para fora do capilar.
PFR= (PC-PI)-(πP-πI)
Os 4 factores que determinam a pressão de filtração de rede são designados deforças de Starling.
A dilatação das arteríolas aumenta a pressão hidrostática capilar e a filtração éaumentada. A constrição arteriolar produz o oposto.
A filtração capilar e a absorção não têm papéis significantes na troca de nutrientes
e produtos metabólicos entre capilares e tecidos. Isto porque a quantidade de substânciaque se move para dentro ou para fora do capilar como resultado do fluxo de volume éextremamente pequena em comparação com as quantidades movidas pela difusão.
Regulação da Pressão Arterial Sistémica:
1. Uma variável interna ambiental que seja mantida constante;2. Receptores sensíveis a alterações nesta variável;
3. Vias aferentes dos receptores;4. Centro integrador que receba e integre inputs aferentes;5. Vias aferentes do centro integrador;6. Efectores direccionados pelas vias eferentes para alterar as suas actividades.
A maior variável cardiovascular, que é regulada é a pressão arterial média nacirculação sistémica.
MAP = output cardíaco x resistência periférica total
Estes 2 factores determinam o volume médio de sangue nas artérias sistémicasdurante o tempo e é esse volume de sangue que causa pressão. O output cardíaco é ovolume ejectado para as artérias por unidade de tempo.
Resistência Arterial Total
Esta resistência influencia a pressão sanguínea arterial sistémica.
144

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 145/374
Durante o exercício, o músculo esquelético arteriolar dilata o que leva a umadiminuição da resistência.
Se o output cardíaco e os diâmetros arteriolares de toda a rede vascular não sealterassem, o fluxo elevado através do músculo esquelético das arteriolas iria causar umdecréscimo na pressão arterial sistémica. Nessas situações, a resistência total dacirculação sistémica pode ser mantida por contracção das arteriolas noutros órgãos (rins,
trato GI, pele, etc).
No entanto, as arteríolas do cérebro não se alteram, assegurando o constantefornecimento do sangue.
A pressão arterial pulmonar média é mais baixa que a pressão arterial sistémicamédia. O output cardíaco das artérias pulmonares e sistémicas é o mesmo, logo aspressões apenas podem ser diferentes se as resistências forem também diferentes.Deste modo conclui-se que as arteriolas pulmonares oferecem menos resistência aofluxo que as sistémicas.
Qualquer desvio na pressão arterial vai desencadear reflexos homeostáticos paraque o output cardíaco ou a resistência periférica total sejam alterados na direcção deminimizar a alteração inicial. Estes reflexos homeostáticos designam-se reflexosbarorreceptores.
Os reflexos barorreceptores utilizam alterações na actividade de nervosautónomos que irrigam o coração e vasos sanguíneos, tal como alterações na secreçãohormonal.
Reflexos Barorreceptores
São os reflexos que regulam a pressão arterial em 1.º lugar.
As artérias carótidas comuns dividem-se em 2 artérias menores, sendo que nestadivisão a parte da artéria é mais fina que o normal e possui um grande número deramificações com muitas terminações nervosas. Esta porção da carótida toma o nome desinus carótida.
Como estas terminações nervosas são altamente sensíveis à distensão oucontracção directamente relacionadas com a pressão na artéria, o sinus carótidafunciona como barorreceptor.
O arco da aorta também tem uma zona funcional semelhante e constitui obarorreceptor do arco da aorta.
Neurónios aferentes que partem destes barorreceptores dirigem-se para o troncocerebral e providenciam inputs para os neurónios dos centros de controlo cardiovascular.
Centro cardiovascular medular
145

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 146/374
O centro integrador primário para os reflexos barorreceptores é uma rede difusade neurónios altamente interconectados, designados de centro medular cardiovascular,localizado no tronco cerebral – medula oblongata.
Quando os barorreceptores arteriais aumentam a sua velocidade de descarga, oresultado é um decréscimo no outflow simpático para o coração, arteriolas e veias e umaumento no outflow parassimpático para o coração.
Funcionamento dos reflexos barorreceptores arteriais
Os barorreceptores funcionam como reguladores de termos curtos da pressãoarterial sanguínea.
No entanto, se a pressão sanguínea se desviar do seu ponto de operação normalpor mais de alguns dias, os barorreceptores arteriais adaptam-se a esta nova pressão.
Outros barorreceptores
As grandes veias sistémicas, vasos pulmonares, e paredes do coração tambémpossuem barorreceptores que funcionam de maneira análoga.
Estes providenciam um maior grau de sensibilidade regulatória contribuindo paraum controlo da pressão arterial através do mecanismo de feedforward.
Volume de sangue e regulação da pressão arterial:
O volume de sangue o factor principal para a regulaçºão da pressão arterial. Esteinfluencia a pressão venosa, retomo venoso, volume diastólico final, volume de ejecção eoutput cardíaco.
Um elevado volume de sangue (elevada volémia) aumenta a pressão arterial, euma elevada pressão arterial diminui o volume de sangue.
146

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 147/374
147

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 148/374
148

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 149/374
149

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 150/374
150

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 151/374
151

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 152/374
152

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 153/374
153

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 154/374
154

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 155/374
155

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 156/374
156

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 157/374
157

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 158/374
158

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 159/374
159

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 160/374
160

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 161/374
161

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 162/374
162

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 163/374
163

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 164/374
164

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 165/374
165

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 166/374
166

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 167/374
167

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 168/374
168

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 169/374
Fisiologia da Respiração
Organização do Sistema Respiratório
Funções principais do aparelho respiratório
• Trocas gasosasPermite que o oxigénio contido no ar passe para o sangue e que o dióxido de
carbono do sangue passe para o ar. O aparelho circulatório transporta o oxigénio dospulmões para as células e o dióxido de carbono das células para os pulmões. Assim, osaparelhos cardiovascular e respiratório complementam-se para fornecer oxigénio e
remover dióxido de carbono de todas as células;
• Controlo do pH sanguíneoO aparelho respiratório pode fazer variar o pH sanguíneo ao alterar a concentração
de dióxido de carbono no sangue;
• FonaçãoO ar que passa através das cordas vocais é essencial para a produção de sons e da
fala;
• OlfactoA sensação de cheiro verifica-se quando as moléculas em suspensão no ar
atravessam as fossas nasais e se ligam aí a receptores específicos;
• Protecção
169

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 170/374
Protege o corpo contra agentes microbianos e químicos através dos cílios, muco efagocitose, dificultando a sua entrada e expulsando-os da superfície das vias aéreas;aquece e humidifica o ar inspirado.
Conceitos básicos
O aparelho respiratório é basicamente constituído por:
- Vias aéreas condutoras;
- Sistema trocador de gases: os pulmões;
- Bomba de ventilação: constituída, essencialmente, por:
- Parede torácica e músculos respiratórios;
- Áreas do SNC que controlam os músculos respiratóros;
- Nervos e tratos que ligam o SNC aos músculos.
Relações entre as vias respiratórias e os vasos sanguíneos
A ampliação destina-se a ilustrar a continuação das vias aéreas pelos “clusters”alveolares nos seus terminais; todo o pulmão é constituído por estes “clusters”.
170

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 171/374
Lei dos gases perfeitos
V
nR P
Lei de Dalton
O volume ocupado por um determinado número de moléculas de gás a umadeterminada temperatura e pressão é idealmente o mesmo, independentemente da
composição do gás. Portanto, a pressão exercida por um gás numa mistura de gases(pressão parcial) é igual ao produto da pressão total pela fracção desse gás na mistura.Ex.: 0,21 de oxigénio x760=160mmHg de oxigénio
Lei de Boile (/Mariot)
Num recipiente fechado a temperatura constante a pressão de um gás éinversamente proporcional ao seu volume (P = 1/V)
Lei de Henry
A concentração de um gás no líquido com o qual está em equilíbrio é determinadapela pressão parcial desse gás na fase gasosa e pela sua solubilidade na fase líquida:
Concentração do gás dissolvido = Pressão parcial do gás na fase gasosa x Coeficiente desolubilidade na fase líquida
Pressões parciais dos gases respiratórios ao nível do mar
171

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 172/374
Os volumes de gás dependem da pressão e da temperatura, pelo que os gasesrespirados precisam de ser corrigidos para as condições de determinação.
Diagrama dos elementos que compõem a parede da via aérea inferior nas suas distintasregiões
Diagrama dos elementos que compõem a parede dos vasos pulmonares nas suasdistintas regiões
172
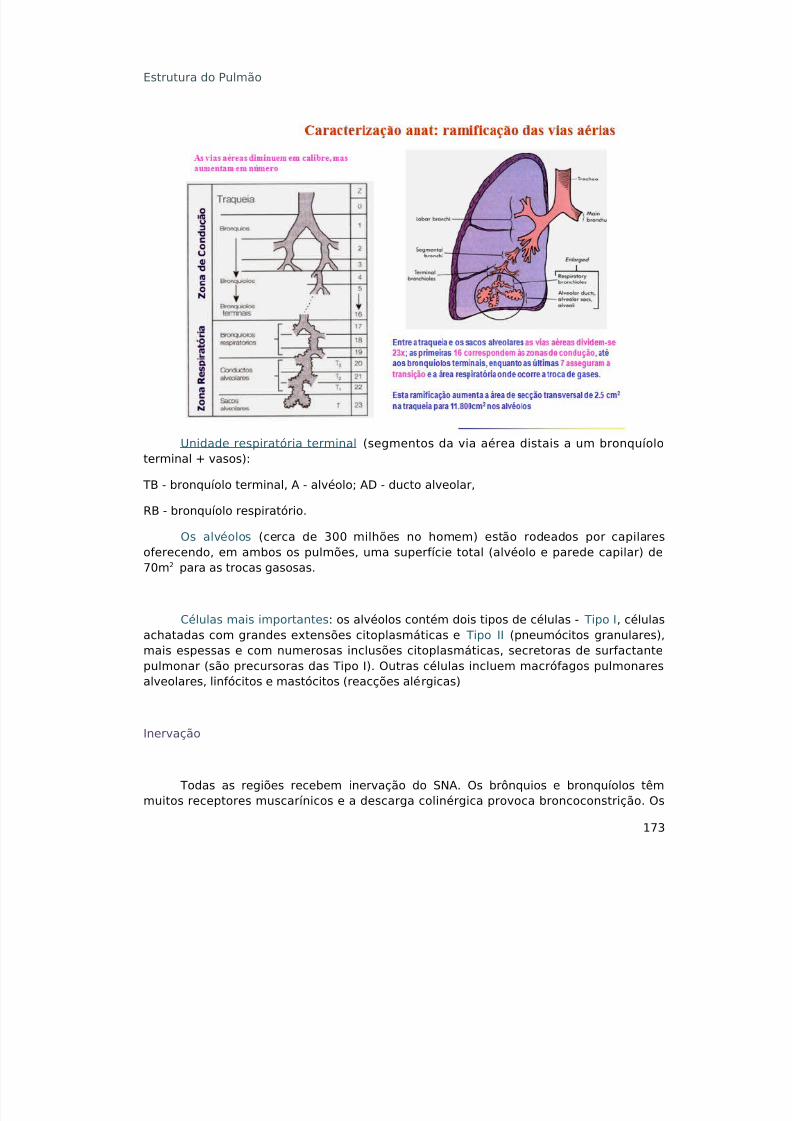
7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 173/374
Estrutura do Pulmão
Unidade respiratória terminal (segmentos da via aérea distais a um bronquíoloterminal + vasos):
TB - bronquíolo terminal, A - alvéolo; AD - ducto alveolar,
RB - bronquíolo respiratório.
Os alvéolos (cerca de 300 milhões no homem) estão rodeados por capilaresoferecendo, em ambos os pulmões, uma superfície total (alvéolo e parede capilar) de70m2 para as trocas gasosas.
Células mais importantes: os alvéolos contém dois tipos de células - Tipo I, célulasachatadas com grandes extensões citoplasmáticas e Tipo II (pneumócitos granulares),mais espessas e com numerosas inclusões citoplasmáticas, secretoras de surfactantepulmonar (são precursoras das Tipo I). Outras células incluem macrófagos pulmonaresalveolares, linfócitos e mastócitos (reacções alérgicas)
Inervação
Todas as regiões recebem inervação do SNA. Os brônquios e bronquíolos têmmuitos receptores muscarínicos e a descarga colinérgica provoca broncoconstrição. Os
173

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 174/374
receptores b2 adrenérgicos medeiam a broncodilatação estando presentes no epitéliobrônquico e no músculo liso. Nos bronquíolos existe ainda inervação não-colinérgica enão - adrenérgica que produz broncodilatação, parecendo que o VIP é o mediadorimplicado.
Vias aéreas são todos os tubos através dos quais o ar flui entre o ambiente internoe externo.
Vias aéreas:
• Superiores:o Narizo Faringeo Laringe
• Inferiores:o Zona condutora: traqueia, brônquios, bronquíolos e brônquios
terminaiso Zona respiratória: bronquíolos respiratórios, ductos alveolares e
alvéolos
As superfícies epiteliais das vais aéreas até ao final dos bronquíolos respiratóriospossuem cílios que batem constantemente na direcção da extremidade superior, alémde células epiteliais individuais que secretam muco. Este epitélio secreta também umfluído aquoso no qual o muco pode mover-se livremente.
Mecanismos Protectores: constrição dos bronquíolos em resposta à irritação emacrófagos presentes nas vias aéreas e alvéolos.
Nota: Os vasos sanguíneos pulmonares geralmente acompanham as vias aéreas etambém são submetidos a ramificações sucessivas.
Ciclo Respiratório é o nome dado ao conjunto de uma inspiração e uma expiração.
Inspiração: movimento do ar do ambiente externo através das vias aéreas para osalvéolos.
Expiração: movimento oposto à inspiração, ou seja, movimento de ar do meiointerno para o externo.
174

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 175/374
Durante o ciclo respiratório o ventrículo direito bombeia o sangue através dasartérias pulmonares e artérias para os capilares que envolvem os alvéolos.
A circulação pulmonar tem uma resistência muito pequena e por isso as pressõesnos vasos sanguíneos são determinados principalmente pelas forças gravitacionais eforças físicas passivas dentro dos pulmões.
Local de Troca Gasosa – Alvéolo
Os alvéolos são pequenos sacos abertos na extremidade, contínuos com o lúmendas vias aéreas. A parede alveolar possui uma camada contínua e simples de célulasdesignadas pneumócitos tipo I. Espalhadas entre estas células existem outras, maisespessas, designadas pneumócitos tipo II, responsáveis pela produção de surfactante.
As paredes alveolares possuem capilares e um espaço intersticial muito pequenoque consiste em fluido intersticial e tecido conjuntivo. O sangue na parede do capilaralveolar é separado do ar que está nos alvéolos por uma barreira extremamente fina.
Em certas paredes alveolares existem poros que permitem o fluxo de ar de unsalvéolos para outros, sendo estes poros muito úteis quando estamos perante alvéolosoclusos.
A superfície total dos alvéolos em contacto com os capilares é muito grande. Aextensa área e a diminuta espessura da parede dos alvéolos permite a rapidez e
eficiência das trocas gasosas entre sangue e pulmões: para o sangue é transferidagrande quantidade de O2 enquanto se liberta o CO2.
Relação entre pulmões e parede torácica
Tórax: compartimento fechado ligado no pescoço por músculos e tecido conjuntivoe completamente separado do abdómen pelo diafragma. A sua parede contém tecido
conjuntivo com propriedades elásticas.
Cada pulmão é envolvido por um saco completamente fechado (saco pleural)constituído por uma fina camada de células (pleura).
A pleura visceral está firmemente ligada ao coração por tecido conjuntivo, estandopor sua vez a pleura parietal ligada à parede torácica e diafragma.
Alterações da pressão hidrostática do fluido intrapleural causam o movimentopara dentro e para fora dos pulmões e parede torácica.
175

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 176/374
Ventilação e Mecanismo Pulmonar
A respiração engloba:
• Respiração externa (absorção de O2 e remoção de CO2 de todo o corpo);
•
Respiração interna (utilização do O2 e produção de CO2 pelas células etrocas gasosas realizadas entre elas e o meio).
A ventilação é a troca de ar entre a atmosfera e os alvéolos.
O ar move-se pelo impedimento de fluxo de uma região de elevada pressão paraoutra de pressão inferior.
(Fluxo = diferença de pressão entre dois pontos/resistência) Sendo
Para o ar fluir para fora ou p+ara dentro dos pulmões importam a pressãoatmosférica e alveolar. Sendo as pressões alveolares causadas por alterações da paredetorácica e pulmões.
Não existem músculos ligados à superfície pulmonar de modo a induzir a suaalteração de volume. Os pulmões são estruturas passivas elásticas cujo volume dependede:
1. Diferença de pressão interior e exterior dos pulmões – pressãotranspulmonar;
2. Distensibilidade dos pulmões.A pressão dentro dos pulmões é a pressão alveolar enquanto a pressão exterior
aos pulmões corresponde à pressão do fluido intrapleural (Pip) PH0 (pressãointrapulmonar) = Palv-Pip.
À medida que a parede torácica expande (contracção dos músculos da paredetorácica e diafragma), a Pip diminui, aumentando o volume pulmonar. Isto baixa a Palv etorna-a negativa relativamente a Patm. O aumento de (Patm-Palv) causa o fluxo de arpara dentro dos pulmões.
A Normopneia é de 12 - 15 movimentos /minuto, contendo cerca de 500 ml (6 a8L/min); nessas condições a troca essencial de gases por minuto é da ordem de 250 mlde O2 entrados, por cerca de 200 mL de CO2 excretados
176

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 177/374
Ventilação: fluxo de ar, gradiente de pressão e resistência.
O gás difunde de áreas de maior pressão para as de menor pressão, de acordocom o gradiente de concentração e a natureza da barreira que separa as duas zonas.
Na inspiração, a expansão da caixa torácica devido à contração dos músculosinspiratórios arrasta o folheto parietal da pleura, baixando a pressão intrapleural dosvalores pré-inspiratórios negativos (-3 mmHg) para valores pós-inspiratórios ainda maisnegativos (-6 mmHg), o que provoca o aumento do volume intrapulmonar e oabaixamento da pressão intra-alveolar, maximizando assim a pressão transpulmonar. Aexpiração reverte este processo.
A pressão transpulmonar (Pt) define-se como a diferença entre a pressão alveolar(Pa) e a pressão intrapleural (Pi):
Pt = Pa - (-Pi) = Pa + Pi
Condições necessárias para a ventilação: Quando a pressão alveolar Palv é inferior àpressão atmosférica Patm o ar entra nos pulmões. O Fluxo F é directamente proporcional àdiferença de pressões e inversamente proporcional à resistência das vias aéreas (R).
177

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 178/374
Mecânica ventilatória: variação de pressão e volume
A elasticidade dos pulmões e da parede torácica é adjuvada pela presença defluido pleural o qual facilita o deslizamento, impedindo, ao mesmo tempo, a separação. Apressão no espaço intrapleural é sub-atmosférica. Na expiração normal a tendência para“recuar” é compensada pela tendência oposta da parede torácica (a abertura da paredeprovocaria o colapso dos pulmões).
A inspiração é um processo activo - a contracção dos m. inspiratórios aumenta ovolume intratorácico enquanto que a pressão na base diminui até -6 mmHg (no esforçoinspiratório a pressão intrapleural pode atingir os 30mmHg). Os pulmões são puxadospara uma posição mais expandida, a pressão nas vias aéreas fica ligeiramente negativae o ar flui para os pulmões. No fim da inspiração o “recuo” dos pulmões impele a paredetorácica para a posição expiratória onde as pressões da parede e dos pulmões seequilibram. A pressão nas vias aéreas fica ligeiramente positiva e o ar flui para fora dospulmões.
A expiração é um processo passivo no sentido em que não existem músculos a
diminuir, através da sua contracção, o volume intratorácico (na primeira porção daexpiração existe um grau reduzido de contracção inspiratória, o que ajuda a controlar orecuo pulmonar e a abrandar a expiração)
178

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 179/374
Variação das pressões intrapleural (intratorácica) e intrapulmonar em relação à pressãoatmosférica na inspiração e expiração
Na ausência de resistência (das vias aéreas e dos tecidos) a pressão intrapleural
seguiria a linha tracejada; por essa razão a curva é desviada à esqdª
Mecânica ventilatória – músculos respiratórios
Inspiração
O movimento do diafragma é responsável em mais de 75% pelas variações dovolume intra-torácico na inspiração (pode percorrer de 1.5 a 7 cm na vertical nainspiração profunda) Os músculos intercostais externos, preenchendo obliquamente oespaço entre duas costelas contíguas, são também muito importantes na inspiração; aocontraírem empurram o esterno para a frente, aumentando principalmente o diâmetroantero-posterior. Estes 2 tipos de músculos principais são suficientes para manter aventilação no repouso. Os escalenos e os esternocleidomastoideus são músculosinspiratórios acessórios, auxiliando na elevação da caixa torácica.
179

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 180/374
Expiração
Trata-se de um processo essencialmente passivo, determinado por relaxamentodos músculos inspiratórios; é auxiliado na expiração forçada pela contracção dosmúsculos expiratórios que auxiliam na redução do volume intratorácico. Os músculosintercostais internos são especialmente importantes, pela sua disposição oblíqua, parabaixo e posterior, puxando a caixa torácica nessa direcção. Os músculos da paredeabdominal anterior auxiliam ainda a expiração, aumentando a pressão intra-abdominal, oque contribui para que o diafragma retome a sua conformação inicial.
180

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 181/374
Mecânica ventilatória: volumes e capacidadespulmonares
Volume de Tidal ou Volume Corrente (+ 500cc) - a quantidade de ar que se move paradentro e fora dos pulmões quando um indivíduo em repouso respira normalmente;
Volume de Reserva Inspiratório (+ 3100cc) - todo o volume de ar inspirado por umesforço activo, para além do volume de Tidal;
181

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 182/374
Volume de Reserva expiratório (+ 1400cc) - todo o volume de ar expelido por um esforçoexpiratório activo após uma expiração passiva;
Capacidade Vital (+ 5000cc) - volume máximo de ar que pode ser expirado após umainspiração máxima; índice clínico de Função Pulmonar, fornecendo, entre outros,informação sobre a força desenvolvida pelos músculos inspiratórios.
FEV 1 - fracção da capacidade vital expirada durante o 1ºs após a capacidade vital; é
também um indicador muito útil, sobre a resistência nas vias aéreas (caso da asma emque a CV pode ser normal mas a FEV1 é reduzida).
Volume de Reserva ou volume residual (+ 1000cc) - quantidade de ar que permanecedentro das estruturas intrapulmonares após uma expiração forçada; essencial paraevitar o “colapso”.
Volumes e Capacidades Pulmonares
O volume de ar que entra nos pulmões durante a inspiração é, normalmente,aproximadamente igual ao volume que sai dos pulmões na expiração consequente. Estevolume de ar designa-se Volume Corrente (VC) ou Volume Tidal (VT). O volume tidaldurante a respiração normal é de aproximadamente 500mL. A quantidade máxima de arque se pode aumentar sobre este volume durante uma inspiração mais prolongada édesignado de volume de reserva inspiratório (sendo cerca de 3000mL).
Após a expiração os pulmões ainda possuem uma grande quantidade de ar – é oponto de repouso dos pulmões e da parede torácica – capacidade residual funcional(CRF).
182

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 183/374
Através de uma contracção máxima activa dos músculos expiratórios é possívelexpirar muito mais do ar que fica após o volume de repouso tidal ter sido expirado. Aeste volume é dado o nome de volume de reserva expiratório (cerca de 1500mL).
Mesmo após uma expiração máxima activa, mantém-se nos pulmõesaproximadamente 1000mL de ar – volume residual.
A capacidade vital corresponde ao volume máximo de ar que uma pessoa podeexpirar após uma expiração máxima. É a soma do volume de repouso tidal, do volumede reserva inspirado e do volume de reserva expirado.
O volume expiratório forçado no 1º Segundo (FEV) é quando uma pessoa tomauma inspiração máxima e depois expira o máximo e o mais rapidamente possível.
Ventilação Alveolar
A ventilação total por minuto (VM) é:
[(ml/min)=(ml/resp)(resp/min)]
Devido ao espaço morto, nem todo o ar está disponível para a troca com o sangue.
183

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 184/374
Mecânica ventilatória: espirometria
A espirometria é uma técnica usada no diagnóstico de doenças pulmonares.Com base nos testes espirométricos de função pulmonar as doenças respiratórias podemser classificadas como restritivas ou obstrutivas. Nas doenças restritivas, como a fibrosepulmonar, a capacidade vital está reduzida abaixo do valor normal (< 4600 ml).Contudo, a velocidade com que o ar pode ser expirado de forma forçada é normal. Porcontraste, em doenças exclusivamente obstrutivas, como a asma a capacidade vital éhabitualmente normal, mas a expiração é mais difícil e demora mais tempo, porque abroncoconstrição aumenta a resistência ao fluxo de ar.
As doenças obstrutivas são assim diagnosticadas por testes que medem avelocidade de expiração, tais como o volume expiratório forçado (FEV), no qual se medea percentagem de capacidade vital que pode ser expirada no primeiro segundo (FEV1).Valores significativamente abaixo de 80% para este parâmetro sugerem a presença dedoença obstrutiva.
Equilíbrio estável entre respirações
Entre respirações os músculos respiratórios estão relaxados e não existecirculação de ar.
A pressão transpulmonar é a força que actua para expandir os pulmões,contrabalançando com o recolhimento elástico (tendência de uma estrutura elástica seopor à distensão).
Entre respirações o volume de ar que permanece nos pulmões é considerável.184

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 185/374
A diferença de pressão na parede torácica (Patm-Pip) equilibra a tendência daparede torácica, parcialmente elástica, comprimida se mover para fora para que aparede torácica e os pulmões estejam estáveis na ausência de qualquer contracçãomuscular respiratória.
A Pip é o facto essencial que mantém os pulmões parcialmente distendidos e aparede torácica parcialmente comprimida entre respirações.
Inspiração
É iniciada por uma contracção induzida pela via neural do diafragma e osmúsculos inspiradores intercostais, sendo o diafragma responsável por cerca de 75% davariação de volume intratorácico.
O diafragma ao contrair-se, move-se para baixo, aumentando o tórax. Osmúsculos intercostais ao contraírem movem-se para fora e para cima, aumentandotambém o tórax.
À medida que o tórax aumenta, a parede torácica afasta-se do pulmão e Pipdiminui enquanto Ptp aumenta ( Ptp=Palv-Pip).
No fim da inspiração o equilíbrio é de novo estabelecido, uma vez que os pulmõesmais distendidos exercem um recolhimento elástico maior que iguala Ptp.
Em situações de exercício físico, por exemplo, a expiração de volumes maiores éconseguido pela contracção de um conjunto diferente de músculos intercostais e
abdominais. Aumentam a pressão intra-abdominal e forçam o diafragma relaxado asubir.
Compliance Pulmonar
O grau de expansão pulmonar é proporcional a Ptp
sendo CP a compliance pulmonar, VP o volume pulmonar
Determinantes da CP:
i) Distensibilidade (aumento da espessura da membrana alveolardiminuição da CP)
ii) Tensão superficial nas interfaces ar - H2O nos alvéolosQuanto maior a CP, mais fácil se torna a expansão dos pulmões a uma alteração naPtp.
185

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 186/374
Se a Cp diminuir a Pip decresce bastante para atingir a expansão pulmonar, odiafragma contrai mais o que provoca um aumento de energia.
A expansão do pulmão requerenergia não só para distender o tecidoconjuntivo, mas também parar superar a tensão superficial da camada de H2O quedelimita o alvéolo.
É muito importante que os pneumócitos II secretem surfactante, que reduz as forçascoesivas entre moléculas de H2O. Por isto, o surfactante diminui a tensão superficial oque aumenta a compliance pulmonar e torna mais fácil a expansão.
Surfactante: complexo de lípidos e proteínas que evita o colapso dos alvéolos quandoestes se encontram mais pequenos, durante a expiração. A quantidade de surfactantetende a aumentar com a respiração profunda, já que esta distende os pneumócitos II eestimula-os a produzir surfactante.
186
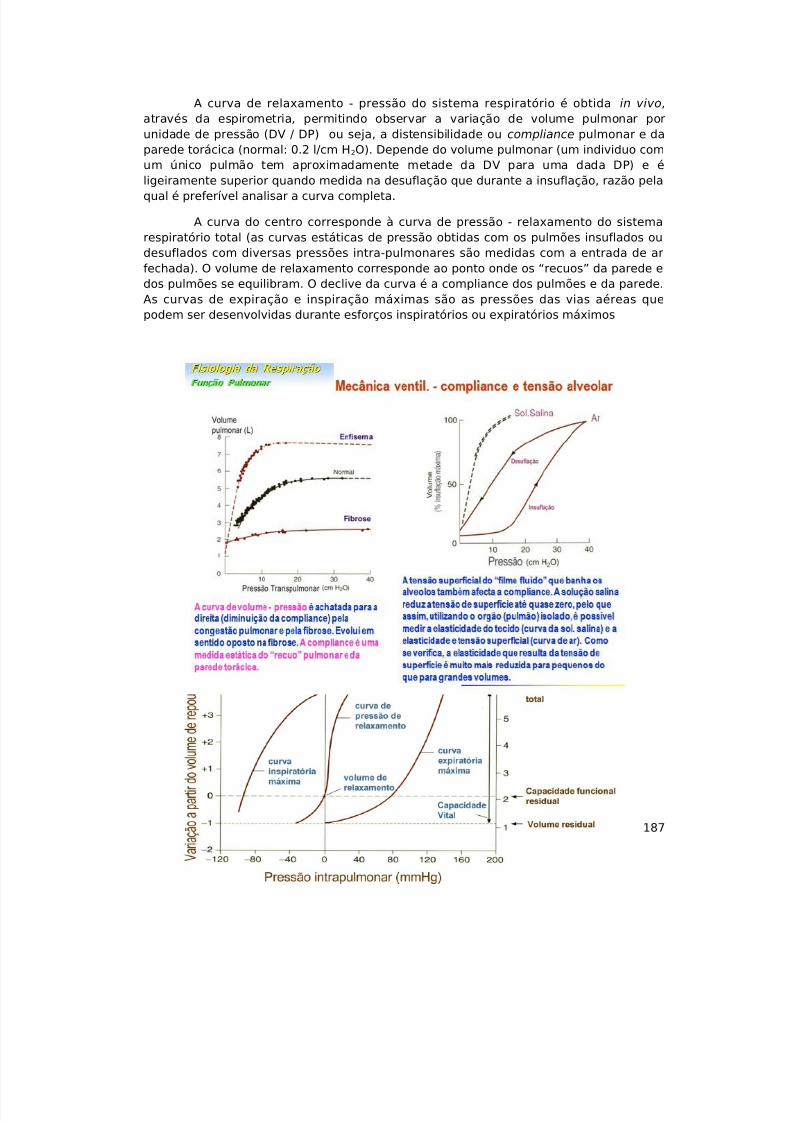
7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 187/374
A curva de relaxamento - pressão do sistema respiratório é obtida in vivo,através da espirometria, permitindo observar a variação de volume pulmonar porunidade de pressão (DV / DP) ou seja, a distensibilidade ou compliance pulmonar e daparede torácica (normal: 0.2 l/cm H
2O). Depende do volume pulmonar (um individuo com
um único pulmão tem aproximadamente metade da DV para uma dada DP) e éligeiramente superior quando medida na desuflação que durante a insuflação, razão pelaqual é preferível analisar a curva completa.
A curva do centro corresponde à curva de pressão - relaxamento do sistemarespiratório total (as curvas estáticas de pressão obtidas com os pulmões insuflados oudesuflados com diversas pressões intra-pulmonares são medidas com a entrada de arfechada). O volume de relaxamento corresponde ao ponto onde os “recuos” da parede edos pulmões se equilibram. O declive da curva é a compliance dos pulmões e da parede.As curvas de expiração e inspiração máximas são as pressões das vias aéreas que
podem ser desenvolvidas durante esforços inspiratórios ou expiratórios máximos
187

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 188/374
188

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 189/374
Resistência das Vias Aéreas
Os factores que determinam a resistência das vias aéreas são:
• O comprimento do tubo;• O raio do tubo;
• A interacção entre moléculas que se movem.
A resistência das vias aéreas ao fluxo de ar é normalmente tão pequena quediferenças de pressão muito pequenas produzem elevados volumes de fluxo de ar.
O raio das vias aéreas é afectado por factores:i) Físicos:
o A pressão transpulmonar exerce uma força de distensão nas viasaéreas e previne que as mais pequenas colapsem.
o As fibras elásticas que ligam o exterior das vias ao tecido alveolar sãopuxadas à medida que os pulmões expandem e ajudam a puxar asvias para as abrir mais do que entre respirações (tracção lateral).
Tanto a pressão transpulmonar como a tracção lateral actuam na mesma direcção,reduzindo a resistência das vias aéreas durante a inspiração.
ii) Neural:
189

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 190/374
o A epinefrina relaxa o músculo liso das vias aéreas (recepotor -
adrenérgico), o que resulta na menor resistência das vias aéreas.o Leucotrienos (eicosanóides) produzidos nos pulmões durante o
processo inflamatório, contraem o músculo.
Espaço Morto
Como as vias não permitem trocas de gás com o sangue, o espaço no seu interioré designado por espaço morto anatómico (VD).
O volume de ar renovado que entra nos alvéolos durante cada inspiração igual ovolume tidal menos o volume de ar do espaço morto.
O volume total de ar renovado que entra nos alvéolos por minuto designa-seventilação alveolar (VA):
A respiração profunda é muito mais eficaz na elevação da ventilação alveolar doque um aumento equivalente da velocidade de respiração.
Algum ar inspirado não é utilizado para a troca gasosa com o sangue apesar de
chagar aos alvéolos, porque certos alvéolos têm pouco ou nenhum suprimentosanguíneo. Este volume de ar é o espaço morto alveolar.
Espaço morto fisiológico = Espaço morto anatómico + Espaço morto alveolar
Respiração: ventilação desigual e espaço morto
A troca de gases ocorre apenas nos terminais alveolares. O restante gás queocupa o sistema respiratório não está disponível para trocas com o sangue capilar -espaço morto, + igual ao peso corporal em libras (homem com 150 lbs. + 68Kg, 150mL
<> ao espaço morto). Para um volume corrente de 500mL apenas os primeiros 350 semisturam com o ar alveolar na inspiração, enquanto que na expiração, os primeiros150mL correspondem ao espaço morto.
Distinguem-se o espaço morto anatómico correspondente ao volume total do
sistema respiratório fora dos alvéolos, e o espaço morto fisiológico ou total, relativo ao
volume de gás que não se equilibra com o sangue (ventilação desperdiçada). Emboracoincidentes no indivíduo saudável, diversas patologias alteram a dimensão e funçãodestes espaços. Para o medir utiliza-se a análise de azoto (single-breath N2 curve)
190

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 191/374
O espaço morto fisiológico ou total pode ser calculado a partir da PCO2 do arexpirado (PECO2), do PCO2 arterial (PaCO2) e do volume corrente (tidal ou V T) de acordocom a Equação de Bohr :
V D corresponde ao volume do espaço morto total e PICO2 a pressão de CO2 no ar
inspirado. Porque PICO2 x V D é muito pequeno, pode ser desprezado e a equação
resolvida em ordem a V D .
O espaço morto anatómico pode ser calculado substituindo PaCO2 pela PCO2
alveolar, isto é PACO2 . Esta corresponde à media do gás proveniente de diversos alvéolosmas proporcional à respectiva ventilação, independentemente do facto de seremperfundidos ou não.
Trocas de Gases nos Alvéolos e Tecidos
Em steady-state, o volume de O2 que deixa os capilares dos tecidos e éconsumido pelas células por unidade de tempo é igual ao volume de O2 adicionado aocorpo nos pulmões durante o mesmo período de tempo. Passa-se o mesmo com o CO2.
As quantidades de O2 consumidos pelas células e CO2 produzido não sãonecessariamente idênticas, dependendo dos nutrientes utilizados para energia.
191

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 192/374
Quociente Respiratório=CO2produzido/CO2 consumido
Pressão Parcial de Gases:
A pressão que um gás exerce é proporcional a:
i) Temperaturaii) Concentração do gás
A difusão do gás irá ocorrer de uma região onde a sua pressão parcial é elevada parauma região onde esta pressão é baixa.
Difusão de Gases em Líquidos:
No equilíbrio, a pressão parcial das moléculas de gás no líquido e fase gasosa têmde ser iguais.
Pressão de Gás Alveolar
A PO2 alveolar é menor que a PO2 atmosférica porque algum do O2 no ar queentra nos alvéolos deixa-os para entrar nos capilares pulmonares. A PO2 alveolar ésuperior à PCO2 atmosférica porque CO2 entra no alvéolo dos capilares pulmonares.
Os factores que determinam os valores precisos de PO2 alveolar são:
i) PO2 atmosféricoii) Velocidade de ventilação alveolar
192

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 193/374
iii) Velocidade de consumo de O2 total no corpo.
Há 2 termos que classificam a ventilação e são referentes somente ao CO2:
• Hipoventilação: quando ocorre um aumento da razão de produção de CO2para a ventilação alveolar. A PCO2 acima dos 40 mmHg normais.
• Hiperventilação: quando ocorre uma diminuição da razão de produção deCO2 para a ventilação alveolar. A PCO2 diminui abaixo do normal.
Troca de Gás entre Alvéolos e Sangue
A difusão dos gases termina quando as pressões parciais capilares se tornamiguais às dos alvéolos.
O sangue que deixa os capilares pulmonares e volta ao coração temessencialmente as mesmas PO2 e PCO2 que o ar alveolar.
Assim, a pressão gasosa alveolar determina a pressão gasosa arterial sistémica.
Quanto mais capilares participarem no processo de difusão mais O2 e CO2 irão sertrocados. Muitos capilares pulmonares no topo do pulmão estão normalmente fecvhados
193

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 194/374
em repouso. Durante o exercício, estes capilares abrem e recebem sangue, permitindo atroca gasosa.
Correspondência entre Ventilação e Fluxo de Sangue nos alvéolos
Para ser eficiente, a correcta proporção do fluxo de ar alveolar (ventilação) e fluxosanguíneo capilar (perfusão) devia estar disponível para cada alvéolo. Quando não está édesignado de desequilíbrio ventilação-perfusão, que tem como principal efeito adiminuição de PO2 no sangue arterial sistémico.
Estados de doença alterações regionais na compliance pulmonar, resistência dasvias aéreas e vasculares, podem causar este desentendimento.
• Podem existir alvéolos ventilados sem fornecimento de sangue;
• Pode existir fluxo de sangue em áreas não ventiladas (shunt).Para contrariar isto, desenvolvem-se respostas homeostáticas para alterar a
distribuição do fluxo sanguíneo.
Trocas gasosas entre tecido e sangue
O fornecimento de novo O2 aos alvéolos e o consumo de O2 nas células criamgradientes de PO2 que produzem difusão de O2 dos alvéolos para o sangue nos pulmões
e do sangue para as células do resto do corpo.
Transporte de Oxigénio no Sangue
O O2 está presente em duas formas:
i) Dissolvido no plasma e eritrócitos
ii) Combinado com moléculas de hemoglobina nos eritrócitos.
Cada molécula de hemoglobina é uma proteína com 4 subunidades ligadas entre si.Cada subunidade consiste num grupo heme e num polipéptido. Os 4 polipéptidos de umamolécula de hemoglobina são designadas de globina. Cada grupo heme contém umátomo de ferro ao qual se liga o oxigénio: assim uma única molécula de hemoglobinapode ligar 4 moléculas de O2.
A fracção de toda a hemoglobina na forma de oxihemoglobina é expressa comopercentagem de saturação da hemoglobina.
194

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 195/374
Saturação (%) = (O2 ligado a Hb/capacidade máxima da Hb se ligar ao O2)x100.
A quantidade total de O2 que é carregada pela hemoglobina no sangue depende nãosó da saturação de hemoglobina mas também da quantidade de Hb que existe por litrode sangue.
Efeito de PO2 na saturação da Hb
As unidades de globina da deoxihemoglobina estão firmemente ligadaselectrostáticamente numa conformação em que a afinidade para o O2 é baixa. A ligaçãodo O2 a uma molécula heme quebra algumas das ligações entre as unidades de globina,
levando a uma alteração na conformação de tal forma que os locais de ligação ao O2ficam mais expostos.
Muitas situações, incluindo elevadas altitudes e doenças pulmonares sãocaracterizadas por uma moderada redução da pressão alveolar e por isso, da PO2.Mesmo que a PO2 desça dos valores normais, a quantidade total de O2 carregado pela
Hb diminui apenas 10% factor de segurança.
O oxigénio ligado à hemoglobina não contribui para PO2 do sangue, só o O2
dissolvido contribui. A presença da hemoglobina tem um papel importante nadeterminação da quantidade total de O2 que irá difundir.
A vasta quantidade de oxigénio que difunde para o sangue dos alvéolos nãopermanece dissolvida mas combina com a hemoglobina. Por isso, a PO2sanguíneapermanece menor que a PO2 alveolar até que a hemoglobina esteja virtualmentesaturada em 100%.
Nos tecidos, o oxigénio está continuamente a difundir para dentro das célulasfazendo com que a PO2 do fluido intersticial seja menor do que a PO2 do sangue,ocorrendo a difusão de O2 do plasma dos capilares para o fluido intersticial.
A PO2 plasmática toma-se menor do que PO2 eritrocitária e o oxigénio difunde doseritrócitos para o plasma.
195

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 196/374
Monóxido de carbono e carga de Oxigénio
O monóxido de carbono tem uma elevada afinidade para os locais de ligação dooxigénio na hemoglobina.
Assim, o CO reduz a quantidade de O2 que combina com a Hb nos capilarespulmonares e altera a molécula de Hb por alterar a curva de dissociação oxigénio-Hb
para a esquerda, diminuindo a dissociação do O2 da Hb nos tecidos.
Efeitos da PCO2 sanguínea, [H+], temperatura e [DPG] sobre a saturação dahemoglobina
Transporte de CO2 no sangue
• 10% encontra-se dissolvido no plasma e eritrócitos;
• 30% reage com os grupos amina da Hb para formar carboamina Hb, sendo
esta reacção facilitada porque a deuxiHb tem maior afinidade para o CO2do que a oxiHb.
• 60% é convertido em bicarbonato:
(anidrase carbónica)
- bicarbonato
A anidrase carbónica só está presente nos eritrócitos e por isso esta reacção
ocorre preferencialmente aqui. O bicarbonato move-se para o plasma através de umtransportador que o troca por um Cl-.
196

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 197/374
Transporte de H+ entre Tecidos e Pulmões
Apenas uma pequena quantidade de H+ se mantém livre o que explica que aacidez do sangue venoso é só ligeiramente superior à do sangue arteiral.
197

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 198/374
Trocas gasosas nos pulmões - principais condicionantes
Difusão
A troca O2 - CO2 efectuada entre oalvéolo e o capilar pulmonar, ocorre de modopassivo, por difusão, sendo regulada pela Leide Fick : VGAS = [área / espessura ] x [ Pa - Pc] xD, sendo que o fluxo ou quantidade de gás (V)inversamente proporcional à espessura e
directamente proporcional à área dasuperfície de troca , à diferença de pressãoentre o alvéolo P1 e o capilar P2 e (3) áconstante de difusão D
A transferência de O2 (ou outros gases com apetência para a Hb, incluindo oCO) efectuada a partir do lumen alveolar para o glóbulo vermelho situado no capilarpulmonar, é determinada pela existência de um gradiente de pressão - maior no alvéolo(PA) que no plasma (PPL), sendo mais uma vez regulada pela Lei de Fick , permitindo o
cálculo da Capacidade de Difusão de qualquer gás :
198

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 199/374
onde DM corresponde à capacidade de difusão da membrana alveolo-capilar, VGAS corresponde à captação de O2 e PPL à sua pressão parcial no plasma. Acapacidade de transferência do O2 através do pulmão está assim dependente de doiselementos principais:a difusão da membrana alveolo-capilar p.dita, e o tempo requeridopara a
combinação do gás com a Hb.
Esquema que ilustra a travessia de um GV num capilar pulmonar e o processo deequilíbrio do oxigénio.
199

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 200/374
O gráfico inferior mostra a variação da pressão parcial de oxigénio do terminalarterial para o terminal venoso do capilar. O equilíbrio do oxigénio entre o alvéolo e ocapilar pulmonares ocorre apenas no 1º terço do capilar, em condições de repouso(curva A) e apenas um pouco adiante quando o débito cardíaco é aumentado (curva B).O oxigénio não consegue o mesmo equilíbrio em condições patológicas que envolvam oaumento da barreira de difusão (curva C)
Controlo da Respiração
Geração Neural da Respiração Rítmica
A respiração depende inteiramente da excitação de músculos respiratórios cíclicosdo diafragma e músculos intercostais, pelos seus nervos motores.
A inspiração é iniciada por potenciais de acção nos nervos dos músculos
inspiradores. Depois dos PA cessarem os músculos inspiradores relaxam e a expiraçãoocorre.
O controlo da actividade neural está nos neurónios da medula do longata-geradorrítmico respiratório.
Os ciclos alternados de descarga e paragem desta actividade nos neuróniosinspiratórios medulares são gerados por uma interacção cooperativo entre inputssinápticos de outros neurónios medulares e potenciais pacemaker intrínsecos dosneurónios inspiradores.
O input modula o output dos neurónios e pode ajudar a terminar a inspiração. Amaior fonte deste output é o centro apneustico enquanto a modulação da actividadedeste centro é da responsabilidade do centro pneumotaxico.
200

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 201/374
Outro sinal de quebra para a inspiração chega de receptores de estiramentopulmonar que se situam na camada de músculo luso das vias aéreas e são activadas poruma grande inflamação pulmonar. Os PA nas fibras nervosas aferentes dos receptores deestiramento viajam para o cérebro e inibem os neurónios inspiratórios medulares(reflexo de Hering-Brewer).
Este reflexo também é importante no ritmo respiratório sob condições de grande
volume tidal.
Controlo da Ventilação pela PO2, PCO2 e [H+]
Os inputs para os neurónios inspiradores mais importantes são os dosquimiorreceptores periféricos e dos quimiorreceptores centrais.
Os quimiorreceptores periféricos, localizados na parte superior do pescoço nabifurcação das artérias carótida comum e no tórax no arco da aorta, são designadoscorpos carótidos e corpos aórticos.
Nas duas localizações estão bastante perto dos barorreceptores arteriais e estãoem contacto íntimo com o sangue arterial.
Os corpos carótidos estão estrategicamente localizados para monitorizar osuplemente de O2 para o cérebro.
Os quimiorreceptores periféricos são compostos por células receptorasespecializadas que são estimuladas principalmente por uma diminuição da PO2 arterial eum aumento de [H+] arterial.
Estas células comunicam sinapticamente com terminais neurais através dos quaisas fibras nervosas aferentes passam para o tronco cerebral; providenciam inputssinápticos excitatórios para os neurónios inspiratórios.
Os inputs dos corpos carótidos são os dominantes.
Os quimiorreceptores centrais estão localizados na medula dolongata e são
estimulados por uma elevação de [H+] do fluido extracelular cerebral.
201

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 202/374
202

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 203/374
203

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 204/374
Controlo da Ventilação durante o Exercício:
204

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 205/374
Durante o exercício a ventilação alveolar pode aumentar até 20 vezes a ventilaçãonormal.
• PCO2: esta não aumenta no sangue arterial, como seria de esperar, emboraaumente no sangue venoso. Isto acontece porque, durante o exercíciomoderado, a ventilação alveolar aumenta na proporção exacta ao aumentoda produção de dióxido de carbono e, portanto, PCO2 alveolar, econsequentemente, PCO2 arterial não se alteram.
• PO2: embora o PO2 venosa sistémica diminua durante o exercício devido aum aumento do consumo de oxigénio nos tecidos, a PO2 alveolar econsequentemente a PO2 arterial geralmente permanecem inalteradas;devido ao aumento do consumo de oxigénio celular e ao aumento daventilação alveolar, numa proporção exacta.
• H+: visto PCO2 arterial não se alterar durante o exercício moderado ediminui em exercício intenso, não se verifica acumulação de CO2. Noentanto, durante o exercício verifica-se um aumento de [H+] arterial nãodevido à PCO2 mas à produção de ácido láctico no músculo e sua posteriorlibertação no sangue. Esta alteração na [H+] é responsável pelahiperventilação no exercício intenso.
• Outros factores: aferência reflexa dos mecanoreceptores das articulações emúsculos; aumento da temperatura corporal; aumento da concentraçãoplasmática de epinefrina; aumento da concentração plasmática de K+ (porsaída do músculo em exercício).
205

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 206/374
206

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 207/374
Pleura
207

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 208/374
Os pulmões estão contidos pelas membranas pleurais (parietal e visceral),separadas entre si por uma fina camada de liquido (liquido pleural). A pressãointrapleural é inferior á pressão atmosférica, permitindo a expansão pulmonar passiva ea entrada de ar dentro dos álveolos durante a inspiração. A pressão transpulmonardefine-se como a diferença entre a pressão alveolar e a pressão pleural, com valoresnormais que variam entre os –5 e 5cm H2O.
O líquido pleural é normalmente incolor, contendo proteína na ordem dos1.5g/dL. Calcula-se que exista na quantidade aproximada de 0.1-0.2mL/Kg peso e que oespaço pleural tenha cerca de 10 a 27 mm de largura.
A formação e reabsorção do fluido pleuralestá regulado pelas Forças de Starling, sendoimportante atender a que a pleura parietal éirrigada pela circulação sistémica e que a pressãohidrostática no espaço pleural é sub-atmosférica (-5cm H2O) e inferior à da pleura parietal (+30 cmH2O). A composição de proteínas do líquido pleuralnormal é idêntico às do interstício normal (idênticapressão oncótica).
A reabsorção de água, proteínas e outro material nol.pleural é feita através dos linfáticos da pleuraparietal
208

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 209/374
Respiração: ventilação e perfusão
O trabalho pulmonar é de natureza elástica e não-elástica; engloba o trabalhodos músculos respiratórios ao estirar os tecidos da parede respiratória e dos pulmões -trabalho elástico (+ 65%) e ao mover elementos viscosos (+ 7%) e vencer a resistêncianas vias aéreas (+ 28%) - trabalho não elástico .
Porque a pressão intrapleural é mais negativa no apex que na base do pulmão,os alvéolos da base estão menos expandidos, antes da inspiração, que os do apex erecebem, por isso, maior fracção do ar inspirado. A base do pulmão recebe tambémmaior fluxo de sangue porque a pressão transmural (para fora) ao longo dos vasospulmonares é maior, aumentando o seu diâmetro e diminuindo a sua resistência.Contudo, estes gradientes não se equilibram e a relação ventilação / perfusão (V/Q)
diminui do apex para a base.
209

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 210/374
A resistência vascular Pulmonar (RVp) expressa a relação entre a pressão e o fluxosanguíneo e é condicionada por diversos factores :
(1) Gravidade
(2) Efeito da Insuflação pulmonar sobre os vasos extra-alveolares, aumentando o seu
diâmetro (a insuflação diminui o diâmetro dos vasos intra-alveolares); o oposto ocorrecom a diminuição do volume pulmonar;
(3) Efeito das variações de fluxo sanguíneo pulmonar - a circulação pulmonar é capaz deacomodar grandes aumentos de fluxo com apenas ligeiros incrementos de pressão(arterial); este efeito é sobretudo conseguido à custa do recrutamento de vasos nãoperfundidos, em especial a nível dos vértices, assegurando também a sobre-distensãodos vasos, em especial a nível das bases
(4) Efeito das variações da composição do ar alveolar - a hipóxia alveolar e, em menorgrau, a hipercapnia produzem vasoconstrição; este fenómeno é proporcional ao grau dehipóxia e limita-se à região afectada (nos restantes locais, a hipóxia e a hipercapnia são
estímulos vasodilatadores locais)
Efeito das pressões intrapleurais (em posição vertical) sobre a ventilação. Porque apressão intra-pulmonar é atmosférica, a pressão intrapleural mais negatica no apéx fazcom que o pulmão esteja aí mais expandido no início da inspiração.
Factores Intrapulmonares
Shunt
A relação ventilação (V) perfusão (Q), designada pelo quociente V / Q é umindicador fundamental da função pulmonar. O fornecimento de ar a segmentos nãoperfundidos é desnecessário. Assim, o pulmão ventilado mas não perfundido tem umquociente V/Q infinito contribuindo para o espaço morto. O fluxo pulmonar arterial quenão é ventilado tem um V/Q igual a zero, correspondendo ao sangue venoso que passadirectamente do coração direito ao esquerdo, ultrapassando os pulmões. Face a umadeterminada patologia, a alteração das relações V/Q processa-se lenta eprogressivamente, acabando por comprometer a ventilação, a perfusão ou ambas asfunções.
O gradiente alveolo-arterial de PO2 que existe normalmente deve-se em grandeparte aos chamados shunts pós-pulmonares, os quais permitem que uma determinadaquantidade de sangue venoso aflua directamente ao compartimento arterial sem quetenha sido oxigenado. Este sangue procede sobretudo de algumas veias bronquiais queacabam nas v. pulmonares (sangue arterial) e da circulação venosa coronária (veias de
Tebésio do V.esqdº). Em qualquer caso, estas comunicações venoarteriais pós-pulmonares contribuem em condições normais, para a redução da PO2 arterial.
210

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 211/374
NOTA: Quando a PO2 arterial está reduzida por shunt , administrar oxigénio a 100% tempouco efeito prático, mesmo neste caso, o oxigénio arterial pode mesmo continuar adescer pois a Hb é rapidamente saturada e o sangue não oxigenado continua a diminuiros valores da PO2 arterial.
Efeito da variação dos rácios V/Q nas PO2 e PCO2 alveolares. As estruturas desenhadas(acima da curva) mostram as zonas de bloqueio.
Quocientes Ventilação / Perfusão
O rácio da ventilação - circulação pulmonar para o orgão inteiro, no repouso, é de0.8 (4.2L/min ar / 5.5 L/min fluxo sanguíneo). Contudo, para diversas zonas do pulmão
211

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 212/374
normal, existem marcadas diferenças para este quociente, determinadas sobretudo peloefeito da gravidade (na doença, as variações locais deste quociente são comuns). Se aventilação num alvéolo é reduzida, face à sua perfusão, a PO2 alveolar diminui enquantoque a PCO2 aumenta porque este gás é menos expirado (Esq.) Inversamente se a perfusãoé reduzida, face à sua ventilação, a PCO2 alveolar diminui porque menos CO2 é entregue ea PO2 aumenta por deficiência de entrada de O2 no sangue(Dta.) .
Factores Extrapulmonares
Ventilação
Em cada movimento respiratório são deslocados cerca de 500 mL de ar (volumecorrente) . Os cerca de 150 mL expulsos em cada expiração são essencialmenteprovenientes dos alvéolos (ar alveolar), embora ocorra sempre alguma mistura com ogás presente no espaço morto. A pressão parcial de O 2 alveolar (PAO2) pode ser calculadaatravés da
Equação do Gás Alveolar:
Sendo,
FIO2 a fracção de moléculas de O2 no gás sêco e
PIO2 o PO2 inspirado e
R o rácio da troca, ou seja, o fluxo de moléculas de CO2 / min ao longo damembrana alveolar a dividir pelo fluxo de moléculas de O 2/ min ao longo da membrana..
212

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 213/374
Factores intra e extrapulmonares determinantes das pressões parciais de O2 e de CO2
A ventilação e a perfusão diminuem, de forma linear, em pé, da base para o ápexpulmonar, pelo que os rácios V/Q são mais elevados nas porções mais elevadas
EXEMPLOS:
Emcima: áreasde
reduzido quociente V/Q (A/B) são equivalentes a unidades pulmonares onde uma
pequena fracção do débito cardíaco representa o sangue “desviado” (shunted) (C);
213

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 214/374
Em baixo: áreas de elevado quociente V/Q (A/B) são equivalentes a unidadespulmonares onde uma pequena fracção dos alvéolos representam o espaço morto (C);(por esta razão, o ápex é a zona preferencial de desenvolvimento da bactéria datuberculose, devido à elevada PO2 local)
Transporte de gases entre o pulmão e os tecidos: oxigénio
A estabilização dos gradientes de pressão parcial de O2 e de CO2 exibe um típicocomportamento de “ decaimento” constante, do ar para os tecidos, para o O2 e, dostecidos para os alvéolos, para o CO2 . Contudo, este comportamento apenas é explicadopela especificidade dos mecanismos envolvidos no transporte do oxigénio, através daHemoglobina (Hb) e da capacidade de dissolução do dióxido de carbono, no sangue :
O sistema de distribuição corporal de O2, tendo por elementos centrais o pulmão e osistema cardio-vascular, depende :
1. da quantidade de O2 que entra nos pulmões
2. de uma adequada troca de gases
3. de uma adequada perfusão do tecido
4. de um adequado transporte de O2 no sangue
A quantidade de O2 no sangue depende da quantidade de O2 dissolvido, daquantidade de Hb no sangue e da afinidade da Hb para o O2 .
214

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 215/374
O oxigénio liga-se ao átomo de Ferro enquanto o Heme se liga à cadeiapolipeptídica, através de um átomo de N para formar uma subunidade de Hb. Quatrodestas subunidades formam uma molécula de Hemoglobina.
A Hemoglobina e o transporte de O2 no sangue
Efeito da adição de Hb na distribuição de O2 em dois compartimentos comdiferentes concentrações do gás, separados por uma membrana semi-permeável. Nonovo equilíbrio os valores de PO2 são iguais, mas inferiores aos de origem, antes daadição de Hb; contudo, o total (dissolvido mais o combinado com a Hb) é agora muitomaior no lado direito da membrana.
A molécula de Hemoglobina (Hb) contém 4 átomos de Fe no estado ferroso (Fe2+)capazes de ligar, rápidamente (menos de 0.01 s) e reversivelmente o O2 - oxigenação:
215

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 216/374
a desoxigenação da Hb4O8 é também muito rápida.
A afinidade da molécula de Hemoglobina (Hb) para o O2 depende da sua estruturaquaternária. Quando a Hb captura O2 as cadeias b aproximam-se, enquanto na cedência(de O2) se afastam. Este movimento está relacionado com a mobilidade dos gruposHeme, podendo assumir uma posição relaxada - estado R que favorece a ligação de ouum estado T (tenso) que dificulta essa ligação. O estado R é sempre facilitado pela
ligação de O2 ao grupo heme. A transição entre os dois estados ocorre num máximo de108 vezes na vida da hemácia.
A Curva de dissociação Oxig - Hemoglobina
A morfologia sigmóide da curva permite que a Hb se sature quase a 100% aindaque a PO2 alveolar seja inferior ao normal (cerca de 90% de saturação são obtidos parauma PO2 alveolar de 60%) - a estes níveis a curva encontra-se na parte superior,
horizontal (zona de carga) . Por outro lado, pequenas variações da PO2 dos tecidosmodificam dramaticamente a libertação de O2 - aqui os valores de PO2 coincidem com a
porção inferior da curva (zona de descarga), na qual pequenas variações de PO2
provocam grandes variações de saturação.
O grau de variação da afinidade da Hb é indicado pela P50 ou pressão de O2
necessária para obter 50% de saturação da Hb.
O desvio à esquerda da curva de dissociação implica uma diminuição da P 50 e,portanto, maior afinidade da Hb pelo O2, maior fixação da molécula e menor libertação
de O2 à periferia (o desvio à direita terá efeito contrário)
216

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 217/374
Quando o sangue está equilibrado com 100% de O2 (PO2 de 760mmHg) ahemoglobina normal fica 100% saturada, significando que contém 1.34 mL de O2 (umpouco abaixo do valor esperado devido à existência em circulação de derivados deformas inactivas de Hb).
Curva de dissociação da Hb estabelecida para um pH de 7.4, a 38 ºC
A concentração normal de Hb é de cerca de 15g/dL, pelo que 1dL de sanguecontém 20.1 mL de O2 ligado à Hb quando esta se encontra 100% saturada.
In vivo nos capilares pulmonares a Hb está cerca de 97.5% saturada de O2 (PO2 =97mmHg) enquanto no sangue arterial periférico a saturação não ultrapassa os 97%(shunts fisiológicos). Assim:
- o sangue arterial contém cerca de 19.8 mL de O2 / dL (0.29 mL dissolvido e 19.5 mLligados à Hb);
- o sangue venoso em repouso, contém Hb apenas 75% saturada e o conteúdo total nãoultrapassa os 15.2 mL/dL.
217

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 218/374
Factores que determinam a afinidade da Hemoglobina para o Oxigénio
Os principais factores que afectam a afinidade da hemoglobina para o oxigéniosão o pH, a temperatura e a concentração de 2,3-difosfoglicerato (2,3 DPG).
O aumento da temperatura ou a queda do pH deslocam a curva para a direita, o
que quer dizer que maior PO2 será necessária para que a Hb capture uma determinadaquantidade de O2 (ou que a Hb cede o O2 com maior dificuldade para tecidos mais frios
ou mais alcalinos).
A diminuição da afinidade da Hb para o oxigénio por efeito da acidificação do meio(queda do pH) é designada por Efeito de Bohr, estando relacionada com o facto de a Hbdesoxigenada ligar H+ de forma mais eficaz que a hemoglobina oxigenada. O pH desce àmedida que o CO2 aumenta.
218

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 219/374
Transporte do Dióxido de Carbono
Destino do CO2 no sangue:
No Plasma
1. Dissolvido
2. Formação de compostos Carbamino com componentes do plasma
3. Hidratação, Tamponamento do H+ , aumento do HCO3- no plasma
Nos Glóbulos Vermelhos
1. Dissolvido
2. Formação de carbamino-Hb
3. Hidratação, Tamponamento do H+ , 70% do HCO3- entra no plasma
4. “shift” de Cloreto; aumento da P osmótica celular.
No Sangue Arterial (cerca de 49mL de CO2 / dL de sangue)
• 2.6 mL (5.3%) está dissolvido• 2.6 mL (5.3%) sob a forma de compostos carbamino•
43.8mL (89.4%) sob a forma de bicarbonatoNos Tecidos (são adicionados cerca de 3.7mL de CO2 / dL de sangue)
219

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 220/374
• 0.4 mL mantém-se em solução• 0.8 mL formam compostos carbamino• 2.5 mL formam bicarbonato
Curvas de Dissociação do CO2
Os pontos arterial (a) e venoso (v) pretendem indicar o conteúdo total de CO 2 nosangue venoso e arterial do adulto normal, em repouso.
Outras Substâncias libertadas no pulmão
Substâncias biologicamente activas metabolizadas nos pulmões220

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 221/374
Sintetizadas e utilizadas no pulmão
Surfactante pulmonar
Sintetizadas ou armazenadas e libertadas
Prostaglandinas
Histamina
Calicreína
Parcialmente removidas do sangue
Prostaglandinas
Bradicinina
Nucleótidos da adenina
Serotonina
Norepinefrina
Acetilcolina
Activação Pulmonar
Angiotensina I
Angiotensina II
Componentes do S. R. que intervêm no controlo da respiração
A respiração espontânea é resultado da descarga rítmica dependente de impulsosnervosos cerebrais transmitidos através de neurónios motores que enervam os músculosrespiratórios.
221

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 222/374
A descarga rítmica é regulada por influências químicas (alterações da PO 2, PCO2 epela concentração de H+ ) e suplementada por outras influências não - químicas.
Receptores
Destinados a receber a informação e a enviá-la aos controladores
Controladores ou Centros Respiratórios
São os responsáveis pela geração do ritmo respiratório basal e processam ainformação proveniente dos sensores em função da qual modificam a sua actividade.
Transmitem ainda aos efectores a informação necessária à manutenção (respiratória) dahomeostasia.
Efectores
Destinados a executar as ordens recebidas por parte dos centros respiratórios,incluem os músculos inspiratórios (incluindo o diafragma e os intercostais externos) e osmúsculos das vias aéreas superiores
Controlo Neural da respiração
A descarga rítmica dos neurónios da medula alongada e da pretuberância notronco cerebral produz a respiração automática:
Secção 4: a transecção medular abaixo do bulbo pára a respiração.
Secção 1: a secção dos vagos em condições normais ou no nível 1 torna o ritmo mais
lento devido à supressão das aferências vagais inibitórias do estiramento pulmonar.
222

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 223/374
Secção 2: a secção vagal neste nível mostra que o centro apneústico é periodicamenteinibido pelas aferências vagais e do c.pneumotáxico .
Secção 3: a vagotomia após inactivação do centro apneústico a este nível apenas tornao ritmo mais lento.
Centro Pneumotáxico: modula os centros respiratórios bulbares, desactivando ainspiração, regulando o volume inspiratório e também a frequência respiratória.
Centro Apneústico: parece constituir o local de integração de diversas aferências quepodem finalizar a inspiração (IO-S “inspiratory-off switch”). A estimulação vagal(aumento do volume pulmonar) bem como a estimulação do centro pneumotáxicoactivam os neurónios IO-S e terminam a inspiração.
Face dorsal do tronco cerebral. Os padrões respiratórios de A a E ilustram aconsequência da transecção a diversos níveis; (A) respiração normal, (B) respiração lentae profunda, (C) respiração apneustica, (D) respiração irregular e, (E) apneia.
O centro respiratório bulbar (Apnêustico) é constituído por 2 grupos respiratóriosdorsais e 2 ventrais.
Cada grupo dorsal é constituído por um conjunto de neurónios activos durante a
inspiração, permanecendo alguns também activos durante a expiração. Os gruposdorsais são responsáveis, principalmente, pela estimulação da contracção do diafragma,recebem aferências de outras regiões do cérebro e dos receptores periféricos,permitindo a modificação do padrão respiratório.
Cada grupo respiratório ventral é constituído por um conjunto de neurónios queestão activos durante a inspiração e a expiração, estimulando, principalmente, osmúsculos intercostais externos e internos e os abdominais.
O grupo respiratório protuberancial (Pneumotáxico) contém neurónios que só têmactividade durante a inspiração, outros durante a expiração e outros ainda em ambas.
Embora a função específica destes neurónios não esteja bem estabelecida, sabe-se quetêm ligações com o centro respiratório bulbar e parece desempenharem um papel de
223

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 224/374
alternância entre a inspiração e a expiração, não sendo contudo considerados essenciaispara desencadear a ventilação.
A respiração consciente envolve o controlo directo pelo cortex cerebral atravésdas vias corticoespinais.
Centros Bulbares: são o local de disparo rítmico essencial, determinado por doisaglomerados de neurónios: o grupo respiratório dorsal (GRD) formado pelo nTs,contendo neurónios inspiratórios de diversos tipos, e o grupo respiratório ventral (GRV)de distribuição mais difusa, formado por 3 regiões essenciais - a parte caudal, ou núcleoretroambíguo (nRA), a parte intermédia ou núcleo paraambíguo (nPA) e, a parte maisrostral formando o complexo de Boetzinger. Esta região parece ser essencial na génesedo ritmo respiratório
Cima: representação da protuberância e bulbo, mostrando a localização dosprincipais agregados de neurónios respiratórios.
Baixo: descarga rítmica dos neurónios da área do complexo de pré-Boetzinger(rato recém-nascido).
224

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 225/374
Controlo químico da função respiratória
A respiração é afectada por quimioreceptores sensíveis à PCO2, pH e PO2 dosangue:
• A PCO2 e as consequentes alterações no pH são de maior importância na regulação
respiratória que a PO2; • Os quimioreceptores centrais do bulbo raquidiano são sensíveis a alterações da
PCO2 devido às modificações resultantes no pH do LCR;• Os quimioreceptores dos corpos aórticos e caratídeos são sensíveis
indirectamente às alterações na PCO2 devido às consequentes modificações no pHsanguíneo.
Input sensorial a partir dos corpos aórticos e carotídeos
Controle Químico da resp.: sensores do sistema
225

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 226/374
Quimiorreceptores Periféricos
A elevação da PCO2 ou de H+ no sangue arterial ou a queda da PO2 aumentam aactividade neuronal respiratória na medula (alterações no sentido inverso determinamum efeito inibitório).
Os efeitos da variação química do sangue são mediados pelos Quimiorreceptores,caracteristicamente representados pelos corpos aórtico e carotídeo (glomus), os quaistêm uma localização relativamente precisa, ao contrário de muitas célulasquimiorreceptoras espalhadas por diversos territórios, iniciando impulsos que estimulamo centro respiratório.
Estes corpos são constituídos por 2 tipos de células, as do Tipo I (glomus) contêmgrandes quantidades de catecolaminas (semelhantes ás cél. cromafins da medulasuprarrenal) que são libertadas na hipóxia ou por efeito do cianeto. As células do Tipo IIsão do tipo glial servindo provavelmente de sustentação às primeiras. As células do Tipo
I contém canais de K+ sensíveis ao O2, pelo que, a hipóxia reduz o efluxo de K+,aumentando o influxo de Ca2+ via canais de Ca2+ tipo L, o que gera um potencial deacção e um impulso aferente.
Os aferentes dos corpos carotídeos atingem a medula através dos nervos do seio
carotídeo e glossofaríngeo, enquanto fibras provenientes dos corpos aórticos sobem peloVago.
Localização dos Corpos Aórtico e Carotídeo.
Note-se que o Corpo carotídeo, mostrado em maior detalhe, se situa muito próximo do
Seio Carotídeo o mais importante barorreceptor arterial.
226

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 227/374
Quimiorreceptores Centrais
Situam-se na face ventral do bulbo raquidiano e são os responsáveis pela respostahiperventilatória principal em situações de hipercápnia e/ou acidose, monitorizadocontinuamente a concentração de H+ no LCR, incluindo o fluido intersticial cerebral (oCO2 atravessa rapidamente as membranas originando ác. Carbónico.
Áreas químio-sensíveis rostral (R) e Caudal (C) na superfície ventral da medula ebulbo raquideano.
Controlo químico da função respiratória
• Decréscimos na PO2 estimulam directamente a respiração apenas quando a PO2 émais baixa que 50mmHg;
• Uma queda na PO2 também estimula indirectamente a respiração, tornando osquimioreceptores mais sensíveis a alterações de PCO2 e pH;
• A volumes tidal iguais ou superiores a 1 litro a inspiração é inibida por receptoresde estiramento dos pulmões pelo reflexo de Hering-Breuer. Um reflexo similar
pode actuar para inibir a expiração;• A barreira hematoencefálica (caixa laranja da figura), permite a passagem do CO2
para o fluído cefalorraquidiano (fluido cerebro-espinal), mas não do protão.
227

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 228/374
Efeitos da PO2, PCO2 e acidez na ventilação
a) Efeito sobre a ventilação de respirar diferentes misturas de oxigénio, mantendo aPaCO2 constante a 40mmHg. Notas:
(1) Minute ventilation = CV . Rr
(2) Alveolar ventilation = (CV-DS).Rr
228

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 229/374
b) Efeito sobre a ventilação de PaCO2 arteriais crescentes obtidas com adição deCO2 ao ar inspirado;
c) Alterações na ventilação em resposta ao aumento da acidez plasmática produzidapela adição de ácido láctico.
Influências não-químicas sobre a respiração
229

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 230/374
Os receptores das vias aéreas e pulmões são enervados por fibras vagaisamielínicas (fibras C) e mielínicas. Estas dividem-se em Receptores de Adaptação Lentae Receptores de Adaptação Rápida na base dos quais a estimulação mantida dá origemà descarga (transitória ou duradoura) das suas fibras aferentes. Outro grupo dereceptores é representado pelos terminais das fibras C e, dependendo da sua
localização, são classificados em bronquiais e pulmonares. Porque estão frequentementepróximos dos vasos pulmonares, são também chamados de Receptores J(justacapilares) . Os receptores J são estimulados pela hiperinsuflação do pulmãoproduzindo apneia reflexa seguida por taquipneia, bradicardia e hipotensão -quimioreflexo pulmonar (semelhante ao coronário; não se conhece o seu significado
fisiológico, mas pode ocorrer no embolismo ou na congestão pulmonar).
Hipóxia (deficiência de O2 ao nível dos tecidos)
Categorias principais
Hipoxia hipóxica ou hipoxémia: PO2 arterial reduzidoHipóxia química: PO2 alveolar normal, mas com conteúdo de O2 do sangue reduzido pornúmero baixo de GV, Hb deficiente ou anormal (anemia) ou competição com CO(carboxémia);
Hipóxia esquémica ou hipóxia de hipoperfusão: fluxo de sangue para os tecidosdemasiado baixo (esquémia)
Hipóxia histotóxica: A quantidade de O2 que atinge os tecidos é normal, mas as célulassão incapazes de o utilizar por interferência de um agente tóxico nos mecanismos
metabólicos da célula (cianeto)
230

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 231/374
Causas da hipoxémia
O decréscimo da PO2 arterial pode ser devida a:
Hipoventilação: (i) defeitos nas vias ou centros de controlo (morfínicos) ou nos músculosrespiratórios, (ii) anomalias na caixa torácica (cifose) e (iii) obstrução das viasrespiratórias (DPOC/ asma, enfisema). A hipoxémia da hipoventilação é sempreacompanhada pela subida da PaCO2;
Perfusão reduzida: resulta do aumento da espessura da membrana alveolar ou dodecréscimo da sua superfície (enfisema, edema pulmonar), dificultando o equilíbrio entrea PO2 do sangue com a PO2 alveolar, muitas vezes é aparente apenas durante oexercício. A PaCO2 é normal, dado que o CO2 é mais difusível que o O2, ou reduzida, se
há estimulação reflexa da ventilação;
Shunt: corresponde ao sangue que passa nos capilares pulmonares sem sercompletamente oxigenado. Pode ser devido a: (i) anormalidade anatómica do sistemacardiovascular que causa mistura de sangue venoso com sangue arterial entre o coraçãodireito e o coração esquerdo (Tetralogia de Fallot) ou (ii) defeito intrapulmonar em quesangue venoso perfunde alvéolos deficientemente ventilados (V/Q=0). A PCO2 arterialhabitualmente não sobe porque a ventilação é estimulada reflexamente pela hipoxémia;
Desigualdade ventilação-perfusão: é a principal causa da hipoxémia, ocorrendo,nomeadamente, nas doenças pulmonares crónicas obstrutivas. A PCO2 arterial pode sernormal ou aumentar, dependendo do grau de estimulação reflexa da ventilação.
Aclimatização à hipóxia de altitude
• Os quimioreceptores periféricos estimulam a ventilação;
• A eritropoetina estimula a eritropoiese;
• O DPG desvia a curva de dissociação oxigénio-hemoglobina para a direita,aumentando assim a descarga do O2 nos tecidos;
• A hipóxia induz a expressão de genes que codificam para factoresangiogénicos, levando à proliferação de novos capilares;
• Os quimioreceptores periféricos estimulam as perdas de sódio e água naurina, levando assim ao aumento dos eritrócitos e da hemoglobina.
231

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 232/374
232

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 233/374
233

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 234/374
234

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 235/374
Fisiologia Renal
Funções do Rim:
Os rins, através da filtração da porção plasmática do sangue e consequenteremoção e adição de substâncias, desempenham as seguintes funções:
i) Regulação do Equílibro Hídrico e Electrolítico;ii) Remoção de produtos finais do metabolismo (ex. fármacos);iii) Remoção de substâncias exógenas (pe fármacos) e a sua excreção;iv) Neo-glucogénese;v) Secreção hormonal de Renina, Eritropoietina e 1,25 DHC
(dihidroxicolecalciferol).
Estrutura dos Rins e Sistema Urinário
Os rins situam-se na parte posterior da parte abdominal. São órgãosretroperitoneais (por trás do peritoneu).
A urina flúi dos rins através dos ureteres para a bexiga, da qual é eliminada
através da uretra. Cada rim contém aproximadamente 1 milhão de nefrónios, sendocada nefrónio constituído por:
• Corpúsculo renal, componente inicial filtrante;• Túbulo que se estende para fora do corpúsculo.O corpúsculo renal forma um filtrado do sangue que é livre de células e de proteínas.
O filtrado deixa o corpúsculo e entra no túbulo, onde são adicionadas e removidassubstâncias. Este filtrado passa então aos ductos colectores onde deixa o rim comourina.
Anatomia dos Corpúsculos Renais
Cada corpúsculo possui um conjunto de capilares designados glomérulo oucapilares glomerulares. Cada glomérulo é irrigado por uma arteríola aferente.
235

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 236/374
O conjunto de um glomérulo com a respectiva cápsula de Bowman (que estárepleta de líquido), designa-se corpúsculo renal.
À medida que o sangue flui através do glomérulo, cerca de 20% do plasma filtrapara a cápsula de Bowman. O sangue remanescente deixa o glomérulo pela arteríolaeferente.
Entre a cápsula de Bowman e os capilares glomerulares existe um espaço cheio defluido, o espaço de Bowman, e é para este espaço que se filtra o fluido livre de proteínas.
O sangue do glomérulo está separado do fluido do espaço de Bowman por 3camadas:
i) Endotélio Simplesii) Lâmina basal proteicaiii) Epitélio Simples constituído por podócitos
236

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 237/374
As células do epitélio são designadas por podócitos e projectam um grandenúmero de extensões. Os fluidos filtrados, primeiro atravessam as células endoteliais,depois a membrana basal e finalmente as extensões dos podócitos.
Existem também células mesangiais que são células musculares lisas modificadasque envolvem os capilares glomerulares mas que não fazem parte da via de filtração.
O túbulo renal é constituído com a cápsula de Bowman. É um cilindro muitoestreito constituído por uma única camada de células epiteliais.
As células epiteliais diferem em estrutura e funções ao longo do túbulo, que édividido em porções:
i) Túbulo proximal- segmento que drena a cápsula de Bowman, inclui o túbulocontornado proximal e o túbulo recto proximal.
ii) Ansa de Henle, com porção ascendente e porção descendente.iii) Túbulo contornado distal, composto por um sistema de ductos colectores:
ducto colector cortical e ducto colector medular
Cada nefrónio é completamente diferenciado. Esta separação termina quando osductos colectores corticais convergem nos ductos colectores medulares que drenampara a pélvis renal, que é contínua com o uréter.
A porção exterior do rim, o córtex renal, contém todos os corpúsculos renais; asansas estendem-se do córtex para a parte interna do rim (a medula), assim como osductos colectores.
Cada túbulo é envolvido por capilares peritubulares que resultam da ramificaçãoda arteríola eferente. Estes capilares juntam-se para formar as veias através das quais osangue deixa os rins.
A porção ascendente da ansa passa entre as arteríolas aferente e eferente. Nesteponto existe um aglomerado de células na parede da ansa densa. Nesta zona, também aparede da arteríola aferente possui células secretoras denominadas células
justaflomerulares. A combinação da mácula densa com as células justaglomerularesdenomina-se aparelho justaglomerular e é responsável pela produção de renina.
237

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 238/374
Processos Renais Básicos
• Filtração Glomerular: filtração do plasma dos capilares glomerulares para o espaçode Bowman. O filtrado glomerular não tem células e contém todas as substânciasdo plasma, excepto proteínas, nas mesmas concentrações.
• Reabsorção Tubular: direcção do movimento do lúmen tubular para os capilaresperitubulares.
• Secreção Tubular: movimento do plasma peritubular para o lúmen tubular.
Assim, uma substância pode ter acesso ao túbulo e ser excretada na urina porfiltração glomerular e/ou secreção tubular. Uma vez no túbulo a substância não énecessariamente excretada, podendo ser reabsorvida.
Quantidade excretada = Quantidade filtrada + Quantidade excretada – Quantidadereabsorvida
238

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 239/374
Metabolismo pelas células tubulares:
As células tubulares renais removem substâncias do filtrado glomerular ou do sanguee metabolizam-nas.
As células produzem substâncias que adicionam ao sangue ou ao filtrado,
nomeadamente amónia, iões H+ e bicarbonato.
Filtração Glomerular
A filtração glomerular é um processo de fluxo de volume em que H2O esubstâncias de baixo peso molecular. Outra razão é o facto das membranascorpusculares estarem carregadas negativamente, a maioria das proteínas plasmáticassão carregadas negativamente.
Forças Envolvidas na Filtração
A filtração através doscapilares é determinada por forçasopostas: a diferença de pressãohidrostática através da parede capilarfavorece a filtração, quanto adiferença de concentração deproteínas através da parede cria umaforça osmótica que se opões àfiltração.
Isto também se aplica aoscapilares glomerulares como se vê nafigura seguinte:
239

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 240/374
Normalmente a pressão de filtração final é sempre positiva porque a pressãohidrostática capilar glomerular (PCG) é maior que a soma da pressão hidrostática no
espaço de Bowman (PEB) e a força osmótica que se opões à filtração (piCG). A pressãode filtração glomerular final inicia a formação da urina forçando o filtrado do plasma aprogredir para os glomérulos e para dentro do espaço de Boeman, e depois através dotúbulo para a pelve renal.
Taxa de Filtração Glomerular
O volume de fluido filtrado do glomérulo para o espaço de Bowman por unidade detempo é designado de Taxa de Filtração Glomerular (TFG).
A TFG é determianda pela pressão de filtração da rede, pela permeabilidade dasmembranas corpusculares e pela área de superfície disponível para a filtração. Quer istodizer que para qualquer pressão de filtração resultante determinada, o TFG édirectamente proporcional à permeabilidade e à área de superfície da membrana.
A capacidade de processar volumes tão elevados de plasma permite que os rinsregulem os constituintes do ambiente interno rapidamente e excretar grandesquantidades de produtos metabólicos.
A TFG não é um valor fixo, isto porque está sujeito a regulação fisiológica,conseguida por inputs neurais e hormonais para as arteríolas aferentes e eferentes.
• Constrição das arteríolas aferentes: a pressão capilar diminui devido à elevadaresistência arteriolar, que causa uma maior perda de pressão entre as arteríolas eos capilares. (diminui PCG)
• Constrição da arteríola eferente: aumenta a pressão, já que o sangue tende arefluir para os capilares glomerulares aumentando a PCG
• Dilatação da arteríola aferente: aumenta PCG e TFG• Dilatação da arteríola eferente: diminui PCG e TFG
• Constrição/Dilatação simultânea: das arteríolas tende a deixar a PCG inalterada.
240

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 241/374
A contracção das células mesangiais reduz a área de superfície dos capilarescausando uma diminuição na TFG a qualquer pressão de filtração.
É possível medir a quantidade total de qualquer substância não proteica filtrada para
o espaço de Bowman através do produto da TFG pela [plasmática da substânica] - cargafiltrada.
Reabsorção Tubular
Pela observação do quadro em cima podemos concluir:
i) As cargas filtradas são enormes, geralmente superiores à quantidade da
substância no corpo;ii) A reabsorção de produtos metabólicos é relativamente incompleta de modo
que grandes quantidades das cargas filtradas são excretadas na urina;
241

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 242/374
iii) A reabsorção de componentes plasmáticos necessários é relativamentecompleta.
Deve-se distinguir-se entre os processos reabsortivos que podem ser controladosfisiologicamente e os que não podem. Assim, as taxas de reabsorção da maioria dosnutrientes orgânicos são sempre muito elevadas e não são reguladas fisiologicamente,
com nenhum aparecimento na urina. Deste modo, os rins não regulam as [] plasmáticasdestes nutrientes orgânicos.
As taxas de reabsorção para a água e muitos iões, apesar de muito elevadas estãosob o controlo fisiológico.
A reabsorção tubular não ocorre por fluxo de volume. Estão envolvidos 2 processos:
i) A reabsorção de algumas substâncias ocorre por difusão, normalmente atravésde tight junctions.
ii) A reabsorção das restantes substâncias envolve o transporte mediado, comproteínas transportadoras.
O movimento do fluido intersticial para os capilares peritubulares ocorre por umacombinação de difusão e fluxo de volume (conduzidas pelas forças de starling capilares).
Reabsorção por difusão: ocorre para uma variedade de substâncias orgânicas lipo-solúveis.
Reabsorção por Transporte mediado: a substância reabsorvida por transporte mediadotem que primeiro atravessar a membrana luminal que separa o lúmen tubular do interiorda célula; depois difunde através do citosol da célula e finalmente atravessa amembrana basolateral que começa nas tight junctions e constitui a membranaplasmática dos lados e base da célula – Transporte epitelial trancelular.
No entanto, não é necessário que as duas membranas atravessadas sejam a
luminal e basolateral para ser considerado transporte activo.
242

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 243/374
Muitos dos sistemas reabsortivos por transporte mediado no túbulo renal têm um
limite na quantidade de material que podem transportar por unidade de tempo.
Transporte Máximo (TM)
Isto acontece porque os locais de ligação nas proteínas transmembranares ficamsaturados.
Secreção Tubular
A secreção move substâncias de capilares peritubulares para o lúmen tubular epode ocorrer também por difusão ou por transporte mediado.
As substâncias mais importantes secretadas são iões H+ e K+, mas também sesecreta colina e creatinina ou substâncias estranhas.
Como na reabsorção, a secreção de certas substâncias está acoplada à reabsorçãodo sódio.
Difusão dos processos nos túbulos
O papel principal dos túbulos proximais é reabsorver a maioria da água filtrada esolutos.
A ansa de Henle também reabsorve grandes quantidades de iões mas uma menorquantidade de água.
A maioria dos controlos homeostáticos são exercidos nos segmentos mais distaisdo túbulo.
Clearence Renal
A clearence renal de qualquer substância é o volume de plasma do qual asubstância é completamente removida pelos rins por unidade de tempo.
Clearence de S= m de S secretada por unidade de tempo/[S] plasmática
243

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 244/374
A clearence de uma substância é uma medida do volume de plasmacompletamente limpo da substância.
Clearence de S= [S] na urina x volume de urina/ [S] plasmática
A clearence de qualquer substância manuseada pelos rins, filtrada mas nãoreabsorvida, secretada ou metabolizada iria igualar a TFG. No entanto não existemsubstâncias assim.
Quando a clearence de uma substância é superior à TFG, essa substância foisubmetida à secreção tubular.
Quando a clearence de uma substância filtrável é menor que a TFG, essasubstância é sujeita a reabsorção.
Micção
A bexiga é um compartimento semelhante a um balão com paredes de músculoliso designado de músculo detrusor, que ao contrariar apertar a urina para dentro dolúmen da bexiga para produzir a micção.
A parte do músculo detrusor na base da bexiga funciona como esfíncter internouretral. Por baixo deste esfíncter existe outro esfíncter externo uretral de músculoesquelético.
Factores que influenciam
i) O músculo detrusor é enervado por neurónios parassimpáticos que causam acontracção muscular. Quando o músculo está relaxado o esfíncter interno estáfechado.
ii) O esfíncter interno recebe inervação simpática que causa a sua contracção.iii) O esfíncter externo é inervado por neurónios somáticos motores que causamcontracção.
Enquanto a bexiga está a encher existe um pequeno input parassimpático(músculo detrusor) mas fortes inputs simpáticos (esfíncter interno) e somatomotores(esfíncter externo) – o músculo está relaxado e os esfíncteres fechados.
Durante a micção
À medida que a bexiga enche a pressão no seu interior aumenta estimulando osreceptores de estiramento na parede da bexiga. As fibras aferentes destes receptores
244

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 245/374
entram na corda espinal e estimulam os neurónios parassimpáticos – contração domúsculo detrusor.
Esta contracção facilita a abertura do esfíncter interno. Simultaneamente o inputaferente dos receptores de estiramento inibe os neurónios simpáticos e os neuróniossomáticos.
Até aqui a micção foi descrita como reflexo espinal local mas as vias descendentesdoo cérebro também podem influenciar, determinando a capacidade de micçãovoluntária.
Regulação do Na+, H2O e K+
Equilíbrio corporal total do Na+ e H2O
Há duas formas de obter H2O:
• produção pela oxidação de nutrientes orgânicos;
• ingestão em líquidos e alimentos.
No entanto, há 5 formas de a perder:
• perda na pele; (contínuo e insensível)• perda pelas vias respiratórias; (contínuo e insensível)• tracto gastrointestinal (muito pequena);• tracto urinário;• fluxo menstrual (nas mulheres).
Sob condições normais as perdas de NaCl e H2O igualam a sua entrada noorganismo e não ocorrem alterações no corpo.
245

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 246/374
Processos renais básicos para Na+ e H2O
Tanto Na+ como H2O filtram livremente dos capilares glomerulares para o espaçode Bowman e circulam no plasma de forma livre. Ambos são submetidos a umareabsorção considerável (+ de 99%) e não são secretados.
Cerca de 2/3 do volume de H2O e Na+ são reabsorvidos no túbulo proximal, mas omaior controlo hormonal da reabsorção ocorre nos túbulos distais e ductos colectores.
Mecanismos de reabsorção de H2O e Na+
• Na+: processo activo que ocorre nos segmentos tubulares excepto na porçãodescendente da ansa.
•
H2O: difusão dependente da reabsorção de Na+.
Reabsorção de Na+
O transporte activo de Na+ para fora da célula da parede tubular através dabomba de Na+/K+ mantém a [] intracelular da Na+ baixa comparada com o lúmentubular e assim o Na+ do lúmen move-se para as células epiteliais tubulares.
O mecanismo do movimento de Na+ através da membrana luminal para as células
varia de segmento para segmento do túbulo, depende dos canais e proteínastransportadoras que estão presentes nas suas membranas luminais.
246

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 247/374
Acoplamento da reabsorção de H2O com a reabsorção de Na+
i) O Na+ é transportado do lúmen tubular para o fluído intersticial através dascélulas epiteliais.
ii) A remoção de soluto baixa a osmolaridade local do fluido tubular (elevação da[H2O]). Simultaneamente também há a elevação da osmolaridade local do fluidointersticial (baixa [H2O)].
iii) A diferença da [H2O] entre o lúmen e o
fluído intersticial causa a difusão de H2Odo lúmen através da membrana plasmáticadas células tubulares para o fluidointersticial.
iv) H2O. Na+ e tudo o resto dissolvido nofluido intersticial move-se por fluxo devolume para os capilares peritubulares.
O movimento de H2O só pode ocorrer se oepitélio for permeável a H2O. Apermeabilidade à H2O dependegrandemente da presença de canais paraH2O: aquaporinas.
A permeabilidade da H2O no tubulo proximal é sempre elevada sendo que nestazona Na+ e H2O são reabsorvidas na mesma proporção.
Nos ductos colectores corticais e medulares, a permeabilidade da H2O é sujeita acontrolo fisiológico. Este controlo é feito por uma hormona secretada pela hipófise
posterior, a vasopressina ou hormona antidiurética. A vasopressina estimula a inserçãona membrana luminal de um particular grupo de aquaporinas feitas pelas células dosductos colectores.
O aumento da urina resultante de baixa vasopressina chama-se diurese aquosa(elevado volume de urina embora sem aumento da excreção de solutos). A diureseosmótica é o resultado de um aumento na excreção de solutos.
Concentração da urina: O sistema Contra-corrente
A capacidade que os rins têm para produzir urina hiperosmótica é o maiordeterminante para a sobrevivência de uma pessoa perante quantidades limitadas deH2O.
O rim humano pode produzir uma concentração urinária máxima de1400mOsmol/L (5xsuperior à osmolaridade do plasma). A excreção diária de solutos naurina atinge cerca de 600mOsmol. Assim o volume mínimo de água na urina na qual estaquantidade de solutos pode ser dissolvida é: 600 (mOsmol/dia)/1400
(mOsmol/L)=0.444L/dia
perda de água obrigatória diariamente.
247

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 248/374
A perda deste volume mínimo de urina contribui para a desidratação quando umapessoa está privada do consumo de água.
O fluido intersticial que envolve os ductos medulares é hiperosmótico e napresença da vasopressina a H2O difunde para fora dos ductos.
Como é que o fluido intersticial medular se torna hiperosmótico?
Como o túbulo proximal reabsorve H2O e Na+ nas mesmas proporções o fluidoque entra na ansa tem a mesma osmolaridade que o plasma.
Ao longo de toda a porção ascendente da ansa, Na+ e Cl- são reabsorvidos dolúmen para o fluído intersticial medular. A porção ascendente é relativamenteimpermeável à H2O. O resultado é que o fluido intersticial medular se tornahiperosmótico quando comparado com o lúmen da porção ascendente.
A porção descendente da ansa não reabsorve NaCl e é altamente permeável à
H2O, ocorrendo uma difusão desta para fora da ansa para o fluido intersticial maisconcentrado, até que a osmolaridade dentro da porção descendente e no fluidointersticial sejam iguais. A hiperosmolaridade intersticial é mantida porque a porçãoascendente continua a reabsorver NaCl.
O multiplicador de contracorrente da ansa torna o fluido intersticial da medulaconcentrado, e é esta hiperosmolaridade que leva H2O para fora dos ductos colectores.
O sistema de contra-corrente concentra o fluido da porção descendente mas baixaa osmolaridade na porção ascendentes de modo a que o fluido que entra no túbulo distalestá mais diluído. O fluido torna-se ainda mais hiposmótico no tubo distal visto que estetambém transporta activamente NaCl e é relativamente impermeável à H2O.
A vasopressina aumenta a permeabilidade tubular à H2O nos ductos colectoresmedulares e corticais.
Na presença de grandesquantidades desta hormona, areabsorção de H2O ocorre por difusãodo fluido hiposmótico dentrodo ducto colector cortical até que o
fluido se torne isosmótico. A H2Oentra nos capilares medulares e élevada para fora dos rins pelosangue venoso. A urina final éhiperosmótica.
248

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 249/374
Circulação Medular
Os vasos sanguíneos na medula (vasa recta) formam ansas que são paralelas àansa de Henle e ductos colectores medulares.
O sangue entra no topo da ansa do vaso e à medida que flui para baixo, cada vezmais profundo na medula, Na+ e Cl- difundem para dentro dos vasos e H2O para fora-Contudo quando o sangue flui na porção ascendente o processo é completamenteinvertido.
A estrutura da ansa dos vasarecta minimiza a perda excessivade soluto do interstício pordifusão.
249

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 250/374
Regulação Renal do Sódio
Em indivíduos normais, a excreção urinária de sódio aumenta quando há umexcesso de sódio e diminui quando há défice de sódio no organismo. Estas respostas
homeostáticas são tão precisas que o sódio corporal total nunca varia demasiado.
Sódio excretado = Sódio filtrado - Sódio reabsorvido
Não existem receptores capazes de detectar a quantidade total de Na+ no corpo.As respostas que regulam a excreção urinária de Na+ são iniciadas principalmente porvários barorreceptores, como o sinus carótida.
Os barorreceptores respondem a alterações na pressão no sistema cardiovasculare iniciam reflexos que rapidamente regulam as pressões, actuando no coração, arteríolase veias.
Como o Na+ é o maior soluto extracelular, alterações no Na+ corporal totalresultam em alterações similares no volume extracelular. Como este compreende ovolume de plasma e volume de fluido intersticial, o volume de plasma também estárelacionado com Na+.
Na+ corporal baixo diminuição volume de plasma diminuição pressões
cardiovasculares Barorreceptores reflexos que influenciam arteríolas renais e
túbulos Baixar TFG aumenta a reabsorção de Na+
Aldosterona e sistema renina-angiotensiva
O córtex adrenal produz uma hormona, aldosterona, que estimula a reabsorção deNa+ pelo túbulo distal e ducto colector cortical.
Quando a aldosterona está completamente ausente, aproximadamente 2% do
Na+ filtrado não é reabsorvido. Quando a [aldosterona] é elevada, todo o Na+ que chegaao túbulo distal e colector cortical é absorvido.
250

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 251/374
A aldosterona actua induzindo a síntese de proteínas nas células alvo, proteínasestas que participam no transporte de Na+.
O que controla a secreção da aldosterona é a hormona angiotensina II que actuadirectamente no córtex adrenal.
A renina é uma enzima secretada pelas células justaglomerulares. Uma vez nacorrente sanguínea, a renina separa a angiotensina I de uma grande proteínaplasmática, o angiotensinogénio, produzido pelo fígado. A angiotensina I é submetida aquebras sucessivas para formar angiotensina II. A conversão é mediada pela enzimaconversora de angiotensina (ECA), que é encontrada na superfície luminal das célulasendoteliais dos capilares.
A angiotensina II estimula a secreção de aldosterona e a constrição das arteríolas.
O angiotensionogénio e a ECA estão normalmente presentes em excesso e porisso o factor limitante é a [] plasmática de renina.
251

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 252/374
252

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 253/374
Mecanismos pelos quais a falta de Na+ causa uma elevação na secreção de renina:
i) Nervos simpáticos renais: inervam directamente as células justaglomerulares eum aumento na actividade destes nervos estimula a secreção de renina. Estesnervos são reflexamente activados vir barorreceptores sempre que umaredução na quantidade de Na+ faz baixar a pressão cardiovascular.
ii) As células justaglomerulares estão localizadas nas paredes das arteríolasaferentes e são sensíveis à pressão funcionando como barorreceptoresintrarrenais. Quando a pressão sanguínea e o volume de plasma diminuem,estas células são menos esticadas, secretando mais renina.
iii) Mácula densa: sente a [Na+] no fluido tubular. Uma concentração de sal baixacausa um aumento na secreção da renina.
Este sistema é muito importante porque os vários mecanismos podem funcionar aomesmo tempo.
Outros factores:
• ANP (Péptido natriurético auricular) sintetizado e secretado por células auricularescardíacas, actua nos túbulos para inibir a reabsorção de Na+. Também podeactuar nos vasos sanguíneos renais para aumentar a TFG, que contribui paraaumentar a excreção de Na+. O ANP inibe directamente a secreção dealdosterona, aumentando a excreção de Na+. A secreção de ANP aumenta devidoao aumento do volume de plasma, que acompanha o aumento de Na+ corporal. Oestímulo específico do aumento na secreção de ANP é o aumento na distensãoauricular.
253

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 254/374
• Pressão arterial sanguínea: um aumento desta inibe a reabsorção de Na+,aumentando assim a sua excreção natriurese pressórica – porque:
o reduz a actividade do sistema renina – angiotensiva aldosterona;o actua localmente nos túbulos.
254

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 255/374
Regulação Renal da Água
Controlo Barorreceptor da secreção de Vasopressina
O baixo volume extracelular acciona também a secreção de vasopressina, queaumenta a permeabilidade à água dos ductos colectores. Maior quantidade de água éretida para estabilizar o volume extracelular.
Os barorreceptores diminuem a sua taxa de descarga quando as pressõescardiovasculares diminuem. Por isso, menos impulsos são transmitidos dosbarorreceptores e o resultado é uma elevada secreção de vasopressina. Esta causa aconstrição arterial, que ajuda a restaurar a pressão sanguínea arteriolar.
255

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 256/374
Controlo Osmorreceptor da secreção da vasopressina
Alterações apenas na água têm relativamente poucos efeitos no volumeextracelular. A razão é que a água distribui-se por todos os compartimentos do corpo,cerca de 2/3 entra no compartimento intracelular em vez de ficar no extracelular comofaz o Na+.
No entanto, grandes alterações na quantidade de água implicam uma alteração naosmolaridade dos fluidos corporais. Nestas condições os receptores que iniciam osreflexos que controlam a secreção de vasopressina são osmorreceptores no hipotálamo.
256

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 257/374
257

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 258/374
258

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 259/374
Regulação Renal do Potássio
O K+ é filtrável no corpúsculo renal sendo a maioria reabsorvido. Contudo, osductos colectores corticais podem secretar K+, sendo que as alterações na excreção deK+ se devem a esta secreção.
Nos ductos colectores corticais o K+ é bombeado para dentro das células atravésda membrana basal pela Na+/K+ ATP-ase e difundem para o lúmen tubular através decanais K+ na membrana luminal. A secreção de K+ está associada como reabsorção deNa+.
Factores que influenciam a secreção de K+
Uma dieta rica em K+ aumenta a [] plasmática deste, o que leva a uma elevadarecaptura através da Na+/K+ ATPase, secretando-se K+.
A aldosterona eleva a secreção de K+. As células secretoras de aldosterona docórtex adrenal são sensíveis à [] de K+ do fluido extracelular.
259

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 260/374
Regulação Renal do Cálcio
A homeostasia do cálcio depende deuma inter-relação dos ossos com os rins e avia gastrointestinal.
O intercâmbio de cálcio entre olíquido extracelular e osso não altera obalanço corporal total do cálcio mas apenasa sua distribuição corporal.
Quanto aos rins, cerca de 60% docálcio plasmático é filtrado no corpúsculorenal (o restante está ligado a proteínasplasmáticas) sendo que a maior parte destecálcio filtrado é reabsorvida.
Como não há secreção tubular de cálcio, a excreção urinária deste é-nos dada peladiferença entre a quantidade filtrada e a quantidade reabsorvida.
260

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 261/374
Assim, o controle ocorre essencialmente a nível da reabsorção, ou seja, areabsorção de cálcio aumenta quando [Ca2+] plasmática diminui e vice-versa.
261

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 262/374
Regulação Renal de Iões H+
As reacções metabólicas são altamente sensíveis à [H+] do fluido em queocorrem. Esta sensibilidade reflecte-se no funcionamento enzimático visto que o H+altera a conformação das proteínas.
Quando a perda de H+ excede o ganho, a [H+] no plasma arterial desce, o pH
sobe acima de 7.4 estamos perante uma alcalose.
Por sua vez, quando o ganho de H+ supera as perdas a [H+] plasmática sobe, apH desce e estamos perante uma acidose.
262

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 263/374
Fontes de H+
• Ganhos:o Formação de H+ pelo CO2;o Produção de ácidos não voláteis do metabolismo de proteínas ou de outras
moléculas orgânicas;o Perda de bicarbonato na urina ou diarrei.
• Perdas:o Utilização do H+ no metabolismo de aniões orgânicos;o Perda no vómito ou na urina:o Hiperventilação.
Uma grande quantidade de CO2 é gerado diariamente devido ao metabolismooxidativo. Estas moléculas de CO2 participam na formação de iões H+ durante apassagem do sangue pelos tecidos periféricos.
Esta fonte não constitui um ganho de H+ pois estes voltam a ser incorporados naágua na passagem pelos pulmões.
A retenção de CO2 ocorre na hiperventilação ou doença respiratória e causaganho de H+.
O corpo também produz ácidos orgânicos ou não, de fontes que não o CO2 (ácidosnão voláteis) como por exemplo o ácido fosfórico ou sulfúrico, que são gerados pelocatabolismo das proteínas.
A dissociação destes ácidos origina anões e H+. Contudo, o metabolismo de umavariedade de aniões orgânicos utiliza H+ para produzir bicarbonato.
263

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 264/374
Quando iões bicarbonato são perdidos é equivalente ao ganho de iões H+. Omotivo é que a perda de bicarbonato causa a reacção 1 da figura anterior.
Para além disto os rins também podem adicionar ou remover iões H+ ao plasma.
Ligações de iões H+ - Sistema Tampão
Qualquer substância que possa ligar reversivelmente H+ é designada tampão.
H+ + Tampão =HTampão
HTampão é um ácido fraco.
Quando [H+] aumento, a reacção é favorecida no sentido directo, e assim esta []não aumenta tanto.
Os tampões estabilizam a [H+] contra alterações em qualquer direcção doequilíbrio.
O tamponamento não elimina ou adiciona H+ do corpo, apenas os mantém presosaté que o equilíbrio seja restabelecido.
Integração dos Controlos Homeostáticos
264

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 265/374
Os rins normalmente excretam o excesso de H+ dos ácidos não-voláteis, ou iõesH+ resultantes de mau funcionamento respiratório ou hipoventilação, ou devido à perdade secreções gastrointestinais básicas. Se existe uma perda de iões, os rins asseguram orestauro do equilíbrio.
Os rins são os últimos equilibradores de H+.
Quando o desequilíbrio de iões de H+ é devido a causas não respiratórias, aventilação é reflexamente alterada de modo a compensar o equilíbrio.
Uma elevada [H+] estimula a ventilação e esta hiperventilação causa a reduçãoda PCO2, reduzindo [H+].
O sistema respiratório e os rins trabalham juntos. A resposta respiratória a [H+]plasmáticas alteradas é muito rápida e mantém esta [] nos mesmos valores até que aresposta mais lenta dos rins consegue eliminar a alteração.
Mecanismos Renais
Os rins eliminam ou compensam o H+ corporal através da alteração da []plasmática de bicarbonato.
Quando existe um abaixamento das [] de H+ no plasma (alcalose) a resposta dosrins é excretar uma grande quantidade de bicarbonato. Em resposta à acidose os rins
não excretam bicarbonato e as células tubulares produzem novo bicarbonato eadicionam-no ao plasma.
Manuseamento do Bicarbonato
O bicarbonato é completamente filtrável no corpúsculo renal e é submetido a
reabsorção tubular no túbulo proximal, porção ascendente da ansa e ductos colectorescorticais, onde também pode ser secretado.
A reabsorção de bicarbonato é um processo activo dependente da secreçãotubular de iões H+ que se combinam no lúmen com o bicarbonato filtrado.
Adição de novo bicarbonato ao plasma
Enquanto ainda existem quantidades significativas de bicarbonato filtrado nolúmen, quase todos os iões H+ irão combinar-se com eles.
265

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 266/374
Os iões H+, secretados a mais, combinam-se no lúmen com tampões não-bicarbonato filtrados, normalmente H2PO4-. Os iões H+ são então excretados na urinacomo parte de H2PO4-.
Quando um H+ secretado se combina com um tampão não-bicarbontao, o efeitonão é apenas a conservação do bicarbonato mas também a adição ao plasma de umbicarbonato novo (gerado na célula tubular pela anidrase carbónica). Isto aumenta a []
plasmática de bicarbonato e alcaliniza-o.
Existe um segundo mecanismo pelo qual os túbulos contribuem com um novobicarbonato para o plasma, que passa através da produção e secreção de NH4+.
Células tubulares, principalmente as do túbulos proximal, capturam glutaminatanto do filtrado glomerular como do plasma peritubular e metabolizam-no. Tanto oNH4+ como o bicarbonato são formados dentro da célula. O NH4+ é activamentesecretado para dentro do lúmen e excretado enquanto o bicarbonato se move paradentro dos capilares peritubulares e constitui o novo bicarbonato plasmático.
A contribuição renal de bicarbonato novo ao plasma é o mesmo, quer tenha sidoconseguido através de:
• Secreção de H+ e excreção em tampões que não sejam bicarbonato• Metabolismo da glutamina com excreção de NH4+A quantidade de H+ excretada na urina nestas duas formas é uma medida da
quantidade do novo bicarbonato adicionado ao plasma pelos rins.
OS rins contribuem com bicarbonato novo suficiente para compensar os iões ácidosnão-voláteis gerados no corpo.
266

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 267/374
Classificação da acidose e da alcalose
A acidose respiratória resulta darespiração alternada o ocorre quando osistema respiratório falha na eliminação deCO2 com a rapidez com que é produzido.
Por sua vez, a alcalose respiratóriaocorre quando o sistema respiratórioelimina CO2 mais rapidamente do que éproduzido.
O ponto principal da acidoserespiratória é uma elevação da PCO2arterial e da [H+]. Para a alcaloserespiratória ocorre uma redução de ambas.
Algumas causas comuns para aacidose matabólica são a produçãoexcessiva de ácido láctico ou a perdaexcessiva de bicarbonato, nomeadamentepor diarreias.
Uma causa frequente para a alcalosemetabólica é o vómito persistente, que está associado à perda de H+.
267

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 268/374
Uma elevada [H+] associada à acidose metabólica, estimulam reflexamente aventilação e baixam a PCO2 arterial.
Alterações na PCO2 plasmática na acidose/alcalose metabólica não são a causa daalcalose/acidose mas são o resultado de respostas compensatórias reflexivas aanomalias não respiratórias.
268

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 269/374
269

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 270/374
270

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 271/374
Fisiologia do Sistema Endócrino
O sistema Endócrino é, a par com o sistema nervoso, um dos principais sistemasde comunicação corporal.
Comparativamente com o sistema nervoso, o endócrino é um sistema decomunicação mais lenta, na medida em que os sinais enviados podem demorar muito
mais a chegar ao receptor e actuar durante um maior espaço de tempo.
O sistema Endócrino consiste em todas as glândulas (endócrinas) que secretam ashormonas. Estas, são diferentes das glândulas exócrinas por não possuírem ductos.
Resumo das principais Glândulas Endócrinas e suas hormonas:
• Células do Tecido Adiposo:o Leptina – taxa metabólica, reprodução
• Supra-renal/Adrenal:o Córtex
Cortisol – controlo do metabolismo orgânico, resposta ao stress,imunidade
Androgénios – impulso sexual na mulher
Aldosterona – excreção renal de Na+ e K+o Medula: (metabolismo, função cardiovascular, resposta ao stress)
Epinefrina
Norepinefrina
• Vias Gastro-intestinais: (actuam no pâncreas, fígado, vesícula biliar e viasGI)
o Gastrinao Secretinao Motilinao Colecistocininao Gip
• Gónadas:o Ovários:
Estrogénio: sistema reprodutor, mamas, crescimento edesenvolvimento dos folículos ovóricos
Progesterona: desenvolvimento, desenvolvimento dos folículos
Inibina: secreção de FSH
Relaxina: relaxamento do cervix e ligamentos púbicoso Testículos:
Testosterona: crescimento e desenvolvimento, sistemareprodutor, impulso sexual, gâmetas
Inibina: secreção de FSH Inibição de Mullerian: regressão dos ductos de mullerian.
271

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 272/374
• Coração:o ANF = Factor natriurético auricular: excreção renal de Na+, pressão
arterial
• Hipotálamo:o Hormonas hipofisiotróficas: estimulam a secreção pela hipófise
anterioro Hormona libertadora da corticotrofina (RH): estimulação da secreção
da hormona adrenocorticotróficao Hormona libertadora de tireotrofina (TRH): estimulação da secreção
da hormona tireo.estimulanteo Hormona libertadora da GHRH (hormona do crescimento):
estimulação da secreção de GHRHo Somatostatina: inibição da secreção da GHRHo Hormona libertadora da gonadotrofina (GnRH): estimulação da
secreção de hormona luteínizante e folículo-estimulanteo Dopamina: inibição da secreção da prolactina
• Rins:o Renina: secreção de aldosterona, regulaçãoda pressão arterialo Eritropoietina: produção de hemáciaso 1,25 di-hidroxivitamina D: absorção de cálcio na via GI
• Fígado:o Factores de crescimento (IGF I e II) semelhantes à insulina:
crescimento e divisão celulares
• Pâncreas: (metabolismo orgânico, glicose, aminoácidos e ácidos gordosplasmáticos)
o Insulinao Glucagono Somatostatina
• Paratiróide:o Paratormona (PTH): cálcio e fosfato plasmáticos
• Pineal:o Melatonina: maturidade sexual e ritmos corporais
• Hipófise:o Anterior
Hormona do crescimento (GH/Somatotrofina): crescimento
Hormona tireoestimulante (TSH): glândula tiróide
Hormona adrenocorticotrófica (ACTH): córtex supra-renal
Prolactina: estimulação da síntese de leite e crescimento dasmamas
Hormonas Gonadotróficas: (secreção de hormonas sexuais egâmetas (homens)
• FSH (hormona folículo-estimulante)• LH (hormona luteínizante)
272

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 273/374
o Posterior
Ocitocina: motilidade uterina e descida do leite para a aréolamamária
ADH (vasopressina): excreção dee água pelos rins, pressãoarterial
• Placenta:o Gonadotrofina coriónica humana: secreção pelo corpo lúteoo Estrogénios: ovárioso Progesterona: ovárioso Lactogénio placental: desenvolvimento das mamas, metabolismo
• Timo:o Timopoietina: função dos linfócitos T
• Tiróide:o T4 e T3: taxa metabólica, crescimento, funções e desenvolvimento
cerebraiso Calcitonina: cálcio plasmático
273

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 274/374
274

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 275/374
Estrutura e Síntese de Hormonas:
As hormonas enquadram-se em 3 classes químicas:
1. Aminas2. Péptidos e proteínas3. Esteróides
1. As hormonas aminadas são todas as que derivam do aminoácido Tirosina.Incluem as hormonas da tiróide, as catecolaminas (epinefrina enorepinefrina) e a dopamina.
2. Estas são inicialmente sintetizadas nos ribossomas das células endócrinasinicialmente como pré-hormonas. Estas são clivadas em pró-hormonas porenzimas proteolíticas no RER. A pró-hormona é então incorporada no Golgionde é novamente clivada para originar a hormona. É importante referirque muitos péptidos servem de hormonas e neurotransmissores ouneromoduladores.
3. As hormonas esteróides são lípidos, principalmente produzidas pelo córtex-
supra-renal e gónadas. O precursor destas hormonas é o colesterol.
275

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 276/374
Resumindo:
• Aminas:o Catecolaminas (epinefrina e norepinefrina) – hidrossolúveiso Dopamina – hidrossolúvelo T3 e T4, Calcitonina – não hidrossolúveis
• Péptidos: a maioria são neurotransmissores
• Esteróides: (Lipossolúveis)o Gónadas
Testosterona
Progesterona
Estrogénio
Inibinao Córtex-adrenal
Androgénio
Aldosterona
Cortisol
Transporte de Hormonas no sangue
A maioria dos péptidos e catecolaminas são hormonas hidrossoluveis, sendo
transportadas no plasma sanguíneo.
Os estertóides e hormonas da Tiróide não são solúveis em água, circulando nacorrente sanguínea ligadas a proteínas do plasma.
Quando a hormona actua no seu tecido alvo, a [] dessa no sangue tem de serreastabelecida.
A [] hormonal no plasma depende:
• Velocidade de secreção
• Velocidade de remoção do sangue
O fígado e os rins são órgãos muito importantes na excreção ou metabolização dehormonas. A hormona pode também ser metabolizada pela célula onde actua.
A resposta a uma hormona é altamente específica, envolvendo apenas as células alvopara essa hormona que possuem receptores específicos na membrana ou dentro dacélula.
Enquanto os receptores para péptidos e catecolamidas são proteínas localizadas na
membrana plasmática, os receptores para esteróides e hormonas da tiróide estãolocalizados no interior da célula.
276

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 277/374
Quando activados pela ligação à membrana, os receptores influenciam:
• Canais iónicos
• Actividade enzimática
• Actividade de Kinases
• Proteínas G
Tipos de Efeitos da Acção Hormonal
• Endócrinos: ocorrem à distância, via corrente sanguínea, atingindo muitosreceptores de forma difusa
• Neurócrinos: à distância mas para um local bem definido
• Parácrinos: afectam, localmente, as células vizinhas
•
Autócrinos: quando é a própria célula que envia a mensagem a responder àmesma
Inputs que controlam a secreção hormonal:
1. Alterações na [] de iões ou nutrientes no plasma2. Neurotransmissores libertados3. Outra hormona que actue na célula endócrina
1. A função principal da hormona é regular, por feedback negativo, aconcentração plasmática do ião ou nutriente que controla a sua secreção.Por exemplo, a secreção da insulina é estimulada pelo aumento da []plasmática de glucose. A insulina secretada vai então provocar umadiminuição da [] plasmática de glucose.
Aferências que controlam a secreção
hormonal
277

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 278/374
Hipotálamo e Hipófise
A Hipófise situa-se na sela turca doo esfenóide, na base do crânio, por baixo dohipotálamo, estando ligada a este através do infundíbulo. É composta por 2 lobos:anterior ou adenohipófise e posterior ou neurohipófise. A neurohipófise é formada portecidos neurais, com conexões com o hipotálamo.
Hormonas da neurohipófise
As hormonas são sintetizadas nos corpos celulares dos neurónios do hipotálamo emovimentam-se em vesículas através dos axónios neurais, acumulando-se nos neuróniosda neurohipófise.
As duas hormonas principais são a ocitocina e a vasopressina.
A ocitocina actua nos músculos lisos uterinos para, ao contraírem-se, provocar asaída do bebé no parto; actua também na mama para estimular a secreção do leite.
A vasopressina actua nas células do músculo liso à volta dos vasos sanguíneos demodo a causar contrcção, aumentando a pressão sanguínea.
278

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 279/374
Hormonas da Adenohipófise e Hipotálamo
Neurónios do hipotálamo secretam hormonas que controlam a secreção dehormonas da adenohipófise. Esta secreta FSH, LH, GH, prolactina e hormonasadrenocorcicotróficas.
As hormonas do hipotálamo (hipofisiotróficas), entram nos capilares e são levadospar o sistema porta hipotalâmico-hipofisário para a adenohipófise.
Se uma célula possui um receptor apropriado para uma dada hormonahipofisiotrófica, essa célula responde através de um aumento ou diminuição da secreçãohormonal.
279

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 280/374
Hormonas hipofisiotróficas
280

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 281/374
281

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 282/374
Tiróide
A glândula tiróide produz 2 moléculas iodadas de grande importância: T3 (tri-iodotironina-3 iodos) e T4 (tiroxina- 4 iodos). A T4 é convertida em T3 nas células alvo.Considera-se a T3 a hormona mais abundante na tiróide, apesar de no total a [T4] sersuperior no sangue.
Dentro da glândula encontram-se numerosos folículos compostos por célulasespecializadas que formam uma esfera fechada, rodeando o lúmen rico em proteínas.
As células foliculares são estimuladas pela TSH (hormona tireo-estimulante), quepor sua vez é estimulada pela TRH. O mecanismo de controlo básico da produção de TSHé por feedback negativo da TG da adenohipófise e hipotálamo.
Nota: TH = hormonas da tiróide
TSH
• Produção de T3 e T4
• Síntese proteica em células foliculares
• Replicação do DNA
• Divisão celular
• Aumento na quantidade de retículo
Acções metabólicas das hormonas da tiróide
As TH´s estimulam a absorção de hidratos de carbono e aumentam a libertação de
ácidos gordos e adipócitos. Estas hormonas regulam também os receptores -
adrenérgicos (efeito permissivo com as catecolaminas) especialmente no coração esistema nervoso.
Os sintomas de excesso de TH’s são idênticos aos sintomas de excesso deepinefrina e norepinefrina.
Crescimento e Desenvolvimento
As TH´s são indispensáveis para uma produção normal da GH (hormona docrescimento).
282

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 283/374
Durante o período letal, TH têm muitos efeitos no desenvolvimento do sistemanervoso, incluindo a formação de terminações de neurónios e a produção de sinapses;produção e crescimento de dendrites e formação de mielina.
A falta de TH resulta numa forma de retardamento mental ao qual é dado o nomede oretinismo.
Glândulas Supra-renais
O termo stress tem o significado de qualquer evento que aumenta a secreção decortisol. A actividade do sistema nervoso simpático, incluindo a libertação da hormonaepinefrina na medula adrenal, é também aumentada em situação de stress.
A elevada secreção de cortisol é mediada principalmente pelo sistema hipófiseanterior-hipotálamo. Um input neural chega ao hipotálamo, vindo de porções do sistema
nervoso que respondem a um stress particular, e induz a secreção de CRH. Estahormona é levada pelos vasos porta para o hipotálamo-hipófise para a hipófise anterior eestimula a secreção de ACTH. Esta, por sua vez, circula para o córtex adrenal e estimulaa libertação de cortisol.
A secreção de ACTH é estimulada também pela vasopressina, epinefrina ecitocinas (que são secretadas por células que pertencem ao sistema imunitário).
Funções Fisiológicas do Cortisol
i) Tem acções permissivas na reacção à epinefrina e norepinefrina decélulas musculares que circundam os vasos sanguíneos – é necessáriouma [] basal de cortisol para manter a pressão sanguínea nanormalidade.
ii) É necessário para manter as [] celulares de determinados enzimasenvolvidos na homeostasia metabólica. Estas enzimas estão localizadasno fígado e actuam para aumentar a produção de glucose hepática entrerefeições.
iii) Tem funções anti-inflamatórias já que inibe a formação de leucotrienos eprostaglandinas, ambos envolvidos na inflamação.
iv) Pode também ser considerado um travão no sistema imunitário, quetenderia a sobreagir a infecções mínimas na ausência de cortisol.
v) Importante na vida fetal e neofetal para o desenvolvimento,diferenciação de tecidos e glândulas, incluindo partes do cérebro,medula adrenal, intestino e pulmões.
Funções do cortisol no stress
1. Efeitos no metabolismo orgânico:283

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 284/374
• Estimulação do catabolismo proteico no osso, músculo e linfa.
• Estimulação da captura de aminoácidos e conversão em glucose dosmesmos no fígado.
• Inibição da captura de glucose e oxidação pelas células sanguíneas,excepto no cérebro.
• Estimulação do catabolismo dos triglicéridos em tecido adiposo com
libertação de glicerol e ácidos gordos, no sangue.2. Elevada reactividade vascular: elevada habilidade para manter
vasoconstrição em resposta à norepinefrina e outros estímulos.3. Efeitos protectores contra influências lesivas do stress.4. Efeitos na inflamação e respostas imunes específicas (inibição).5. Inibição de funções como reprodução e crescimento.
Stress
O stress é uma situação em que existe uma ameaça (real ou potencial) ahomeostasia. Assim, é importante manter a pressão sanguínea de modo aprovidenciar fontes de combustível adicionais no sangue e para desligartemporariamente funções não essenciais.
O cortisol aumenta a reactividade vascular, cataboliza proteínas e lípidos paraprovidenciar fontes de combustível/energia e inibe a habilidade da actuação da
insulina.
O cérebro e o músculo esquelético não necessitam de insulina para a sua acção.Ao inibir as acções da insulina, o cortisol permite a ocorrência de um mecanismo quegarante que, durante tempos de stress, os músculos e o cérebro obtenham aquantidade necessária de glucose.
As outras células do corpo não estão desprovidas de combustível, pois o cortisolprovidencia ácidos gordos para quebra de triglicéridos, que podem ser usados pelascélulas do corpo como alternativa energética.
O cortisol é altamente catabólico e com isso, células do sistema imunitário,músculos, pele e glândulas submetem-se a catabolismo para providenciar substratospara a gluconeogénese.
284

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 285/374
Funções do sistema nervoso simpático no stress
A activação do sistema nervoso simpático durante o stress é designada porresposta “luta ou foge”.
1. Elevada glicogenólise hepática e muscular providencia rápida fonte de
glucose.2. Elevada quebra de triglicéridos do tecido adiposo providencia um
suplemento de glicerol para a gluconeogénese e de ácidos gordos para aoxidação.
3. Diminuição da fadiga muscular esquelética.4. Elevada função cardíaca.5. Elevada ventilação pulmonar por estimulação de centros respiratórios
cerebrais e dilatação das vias aéreas.
Outras hormonas libertadas durante o stress:
o Reter H2O e sódio perante perda por transpiração ou hemorragia, demodo a evitar desidratação:
• Aldosterona
• Vasopressina (estimula secreção de ACTH)o Mobilizar substâncias energéticas
• GH
• Glucagon
• Insulina (diminuida)
Controlo endócrino do crescimento
285

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 286/374
As hormonas mais importantes para o crescimento são a GH, factores decrescimento (insulina I e II), hormonas da tiróide, insulina, testosterona e estrogénio.
Existe também um grupo de factores de crescimento peptidicos que actuam nasua maioria como agentes parácrinos ou autócrinos para estimular a diferenciação e/oudivisão celular de determinados tipos de células.
Existem também factores inibidores do crescimento peptidicos que modulam ocrescimento através da inibição da divisão em tecidos específicos.
As diversas hormonas e factores de crescimento não o estimulam nos mesmosperíodos da vida.
Hormona do crescimento e factores de crescimento
A hormona do crescimento secretada pela hipófise anterior tem muito pouco efeitono crescimento fetal mas é a hormona mais importante para o crescimento pós-natal:estimulação da divisão celular.
A hormona do crescimento exerce o seu efeito mitogénico indirectamente nascélulas através da mediação de um mitogénio. Este é designado de factor decrescimento (insulina I ou somatomedinas).
Sob influência da hormona do crescimento a IGF I é secretada pelo fígado, entrano sangue. A IGF I também pode ser secretada por outros tipos de células, incluindoossos e nestes casos actua como agente autócrino e parácrino.
A hormona do crescimento estimula as células percursoras dos condrócitos nacamada proliferativa a diferenciarem-se.
Durante esta diferenciação as células secretam IGF I e ficam activas.
A IGF I actua como agente autócrino e parácrino para estimular a divisão celulardos condrócitos.
A secreção e actividade de IGF I pode ser influenciada pelo estado nutricional doindivíduo e por muitas hormonas que não sejam do crescimento.
A secreção de GH é estimulada pelo GHRH e inibida pela somatostatina. Durante amaior parte do dia, existe pouca ou nenhuma GH secretada.
286

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 287/374
As hormonas sexuais, insulina e hormonas da tiróide influenciam também asecreção de GH. O resultado global da secreção da GH é maior durante a adolescência emenor nos adultos.
A baixa secreção de GH associada à idade é responsável pelo decréscimo damassa corporal, expansão do tecido adiposo e fragilização da pele.
A IGF I é requerida para o crescimento fetal do corpo normal do feto eespecificamente para maturação normal do seu sistema nervoso. No entanto, osestímulos para a secreção de IGF I durante a vida pré-natal são desconhecidos.
A IGF II, cuja secreção é independente de GH é também um mitogénio crucialdurante o período pré-natal. Apesar de continuar a ser secretado ao longo da vida,desconhece-se a sua função após o nascimento.
287

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 288/374
Cortisol: pode ter efeitos anti-crescimento, porque em elevadas [] inibe a síntese de DNAe estimula o catabolismo proteico inibindo o crescimento ósseo e a secreção de GH.
288

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 289/374
289

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 290/374
290

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 291/374
291

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 292/374
292

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 293/374
293

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 294/374
294

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 295/374
295

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 296/374
296

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 297/374
Fisiologia da Reprodução
Funções dos Gónadas
1. Gametogénese: produção de células reprodutoras (Gâmetas); espermatozóides eóvulos.
2. Secreção de hormonas esteróides: testosterona e estradiol e progesterona.
A testosterona pertence a um grupo de hormonas esteróides que têm acçõesmasculinizantes e que, colectivamente, são designadas de androgénios. Nos homens, amaioria da testosterona circulante é sintetizada nos testículos e os outros androgénioscirculantes são produzidos pelo córtex adrenal, mas estes são muito menos eficazes esão incapazes de manter a função reprodutora sozinhos.
O estradiol é secretado em grandes quantidades pelos ovários e é geralmentedesignado de estrogénios.
Os estrogénios e os androgénios não são exclusivos das mulheres e dos homens,respectivamente.
O estrogénio está presente nos homens pois é libertado em pequenas quantidadespelos testículos e é convertido a partir dos androgénios em alguns tecidos não gónadas.Nas mulheres, pequenas quantidades de androgénios são secretadas pelos ovários emaiores quantidades pelo córtex adrenal.
Todas as hormonas esteróides actuam da mesma forma: ligam-se a receptoresintracelulares, que se ligam posteriormente ao DNA para alterar a taxa de formação deum mRNA particular.
A função reprodutiva está controlada por uma cadeia de hormonas. A primeirahormona da cadeia é a GnRH hipofisiotrópica.
A secreção de GnRH é accionada por potenciais de acção nas células neuroendócrinashipotalâmicas produtoras de GnRH. O padrão pulsátil da secreção de GnRH é importanteporque as células da hipófise anterior que secretam as gonadotropinas, não respondemà GnRH se a concentração desta hormona se mantiver constantemente elevada.
Na hipófise anterior, a GnRH estimula a libertação das gonadotropinas hipofisárias:FSH e LH, e a sua estrutura é a mesma em ambos os sexos. Estas duas hormonas têmcomo resultado:
1. Maturação de espermatozóides e óvulos;2. Estimulação da secreção das hormonas sexuais.
297

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 298/374
298

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 299/374
Princípios gerais da Gametogénese
Em qualquer ponto dagametogénese, os gâmetas emdesenvolvimento são designados decélulas germinativas.
1.ª etapa
Proliferação das células germinativas pormitose. A mitose de células germinativasem que cada uma possui 46cromossomas providencia umsuplemento de células germinativas idênticas para as próximas etapas.
2.ª etapa
Meiose, em que cada gâmeta resultante recebe 23 cromossomas da células germinativaque lhe deu origem.
299

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 300/374
300

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 301/374
Fisiologia Reprodutiva Masculina
O sistema reprodutor masculino inclui os testículos, sistema de ductos quearmazenam e transportam espermatozóides, as glândulas que drenam para esses ductos
e o pénis.
A formação normal dos espermatozóides requer uma temperatura cerca de 2ºCabaixo da temperatura normal do corpo.
Os locais para a espermatogénese são os tubos seminiferos, que têm o lúmencheio de fluido que contém os espermatozóides. A parte tubular é composta por célulasgerminativas em desenvolvimento e células de Sertoli.
As células de Leydig situam-se nos pequenos espaços de tecido conjuntivo entreos túbulos, são responsáveis pela síntese e libertação de testosterona.
Os tubulos seminíferos convergem para formar uma rede de tubosinterconectados - rede testicular (rete testis). Os ductos eferentes deixam a rede edrenam para o epidídimo. Este drena para cada testículo para os ductos deferentes.
Os ductos das vesículas seminais juntam-se aos ductos deferentes para formar 2ductos ejaculadores que entram na glândula prostática e juntam-se à uretra, provenienteda bexiga.
O par de glândulas bulbouretrais drenam para a uretra após esta deixar apróstata.
301

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 302/374
302

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 303/374
Espermatogénese
- As espermatogónias começam a divisão durante a puberdade;
- O suplemento de espermatogónias indeferenciadas não diminui porque existe semprealgumas que após a mitose regridem;
- Cada espermatócito primário aumenta marcadamente de tamanho;
- A diferenciação dos espermatideos em espermatozóides envolve uma remodelaçãocelular extensiva: alongamento, a ponta do núcleo é coberta pelo acrossoma;
- A espermatogénese leva cerca de 64 dias.
Cada uma das células de Sertoli estende-se desde a membrana basal até aolúmen e está ligada à célula de sertoli adjacente de tight junctions. Estas tight junctionsdividem o túbulo em 2 compartimentos: compartimento basal (entre a membrana basale a tight junction) e o compartimento central (começa nas tight junctions e inclui olúmen). A barreira celular de Sertoli previne o movimento de qualquer químico dosangue para o lúmen e ajuda a reter o fluido luminal. Este arranjo celular tambémpermite que as etapas da espermatogénese ocorram em compartimentos diferentes.
As células de Sertoli servem de rota para os nutrientes atingirem as células em
desenvolvimento e secretam a maioria do fluido encontrado no lúmen tubular.
303

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 304/374
As células de Sertoli, em resposta à FSH e à Testosterona secretam uma variedadede mensageiros químicos que funcionam como agentes parácrinos para estimular aproliferação e diferenciação das células germinativas.
As células de Sertoli secretam a hormona proteica inibina e agentes parácrinosque afectam o funcionamento das células de Leydig.
Transporte de Espermatozóides
Os ductos deferentes e a porção do epidídimo mais perto destes servem paraarmazenar os espermatozóides até à ejaculação.
Durante a passagem pelo epidídimo a concentração de espermatozóides aumentapela absorção de fluido pelo lúmen do epidídimo; assim no epidídimo são movidos porcontracções peristálticas de músculo liso.
304

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 305/374
Erecção
Ejaculação
1. O músculo liso doepidídimo, ductosdeferentes eejaculatórios, prostáta evesícula seminalcontraem como resultadode uma estimulação de
nervos simpáticos.2. O sémen é expelido da
uretrapor umasérie de contracçõesrápidas do músculo lisouretral e do músculo
esquelético dabase do pénis.
305

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 306/374
Controlo Hormonal
• Controlo dos Testículoso As células
neuroendócrinassecretoras de GnRHaccionam umpequeno conjunto
de potenciais de acçãoa cada 90 min, secretando
GnRH.
306

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 307/374
• Testosteronao Necessária para o inicio e manutenção da espermatogénese (via células de
Sertoli);o Diminui a secreção de GnRH através de uma acção directa no hipotálamo;o Inibe a secreção de LH através de uma acção directa na hipófise anterior;o Diferenciação nos órgãos reprodutores acessórios;o Induz características sexuais secundárias, opõe-se à acção do estrogénio no
crescimento do peito;o Estimula o anabolismo proteico e crescimento ósseo;o Estimula a secreção de eritropoietina pelos rins.
307

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 308/374
A testosterona tem de ser submetida a transformações nas suas células alvo demaneira a tornar-se efectiva. Após a sua entrada no citoplasma é convertida a
dihidrostestosterona (DHT) pela 5-α-redutase, noutros tecidos é transformada emestradiol pela aromatase.
Fisiologia Reprodutiva Feminina
308

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 309/374
Oogénese
Crescimento folicular
Os folículos começam como folículos primordiais, que são um oócito primáriocircundado por uma camada de células denominadas células granulosas.
Ocorre um aumento do oócito, proliferação das células granulosas em camadas eseparação do oócito das células granulosas na zona pelúcida.
As células granulosas secretam estrogénio, inibina e pequenas quantidades deprogesterona antes da ovulação.
O oócito continua associado às células granulosas por GAP junctions, que lhepermitem receber os factores que as células granulosas secretam e que mantêm osoócitos primários em retenção da meiose.
309

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 310/374
A teca corresponde às células de tecido conjuntivo que circundam as célulasgranulosas e se diferenciam formando camadas. Esta tem um importante papel nasecreção de estrogénio pelas células granulosas.
Formação do Corpo Lúteo e Ovulação
Após o folículo maduro descarregar o ovo, ele colapsa em redor do antro e sofreuma transformação.
As células granulosas aumentam muito de tamanho e toda a estrutura formadadesigna-se corpo lúteo, que secreta estrogénio, progesterona e inibina.
É a perda da função do corpo lúteo, que leva à menstruação e ao início de umnovo ciclo menstrual.
Em termos de função ovárica, o ciclo menstrual pode ser dividido em 2 fasesaproximadamente iguais em duração e separadas pela ovulação.
310

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 311/374
Controle da Função Ovárica
O ciclo menstrual resulta de uma inter-relação entre as hormonas secretadaspelos ovários, adenohipófise e hipotálamo.
311
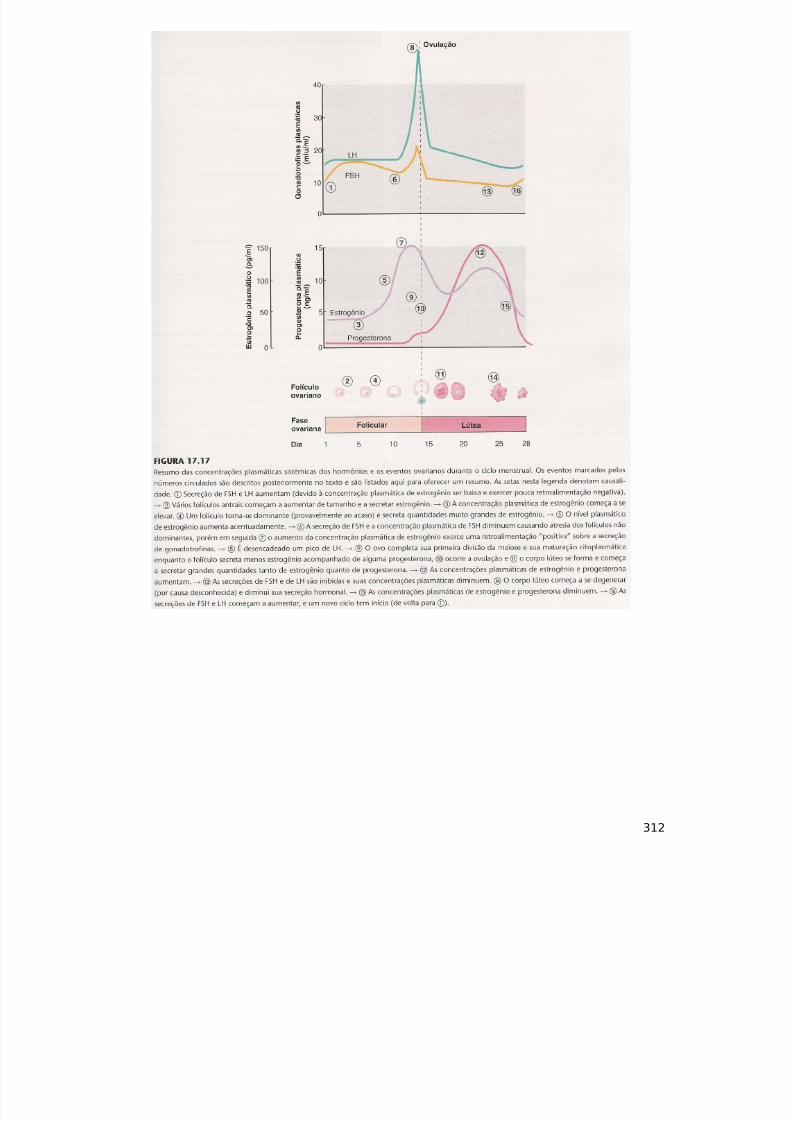
7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 312/374
312

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 313/374
Durante as fases folicular inicial e média, o FSH estimula a proliferação das célulasgranulosas e a secreção de estrogénio por estas; enquanto o LH estimula as células dateca produzirem os androgénios que as células granulosas usam para sintetizar oestrogénio.
Neste período o estrogénio exerce um feedback negativo sobre a adenohipófise,inibindo a secreção de gonadotropinas, inibindo também a secreção de GnRH pelohipotálamo. Além disto a inibina inibe preferencialmente a secreção de FSH.
Durante a fase folicular tardia, o estrogénio plasmático aumenta para desencadearum pico de LH, que então causa através das células granulosas o fim da primeira divisãoda meiose do ovo e a sua maturaçãocitoplasmática, ovulação e formaçãodo corpo lúteo.
Durante a fase lútea, sob a influênciade pequenas quantidades de LH, ocorpo lúteo secreta progesterona eestrogénio. A regressão do corpolúteo resulta da paragem da secreçãode GnRH e gonadotropinas éinibida pela combinação deprogesterona + inibina + estrogénio.
313

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 314/374
314

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 315/374
Alterações Uterinas noCiclo Menstrual
315

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 316/374
A fase folicular ovárica é equivalente às fases menstrual e proliferativa uterinas,sendo o 1.º dia correspondente ao 1.º dia do ciclo. A fase lútea ovárica é equivalente àfase secretora uterina.
A menstruação ocorre quando os níveis de estrogénio e progesterona no plasmadiminuem como resultado da regressão do corpo lúteo.
Durante a fase proliferativa, o estrogénio estimula o crescimento do endométrio edo miométrio e faz com que o muco cervical seja facilmente penetrável peloespermatozóide.
Durante a fase secretora, a progesterona converte o endométrio, que está sobacção do estrogénio, num tecido secretor e torna o muco cervical relativamenteimpenetrável ao espermo. Inibe também as contracções uterinas.
Efeitos dos Esteróides Sexuais Femininos
316

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 317/374
317

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 318/374
318

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 319/374
319

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 320/374
320

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 321/374
321

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 322/374
322

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 323/374
323

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 324/374
324

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 325/374
325

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 326/374
326

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 327/374
327

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 328/374
328

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 329/374
329

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 330/374
330

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 331/374
331

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 332/374
332

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 333/374
Fisiologia Gastrointestinal
Digestão e Absorção dos Alimentos
Função dos órgãos gastrointestinais
• Saliva:
o Mistura e lubrifica as partículas dos alimentos antes destas seremengolidas;
o Contém amilase que digere parcialmente polissacáridos complexos;o Dissolve algumas moléculas dos alimentos, pois apenas no estado
dissolvido estas moléculas podem reagir com quimioreceptores na boca,levando à sensação de sabor;
o Tem propriedades antibacterianas.
• Estômago:o As glândulas que se situam na parede do estômago secretam ácido
clorídrico e pepsina;o A função primária do ácido clorídrico é dissolver as partículas dos alimentos.
O ambiente ácido no lúmen gástrico altera a ionização de moléculaspolares, especialmente proteínas;
o
As proteínas e polissacáridos libertos pela acção do ácido clorídrico sãoparcialmente digeridos no estômago pela pepsina e pela amilase. Os lípidosnão se dissolvem por acção do ácido;
333

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 334/374
o O ácido clorídrico também destrói a maioria das bactérias que entram juntamente com os alimentos.
Quimo – fragmentos moleculares de proteínas e polissacaridos, gotículaslipídicas, sal e água.
Excepto a água, nenhuma destas moléculas consegue atravessar o epitéliogástrico. A maioria do quimo que chega ao intestino delgado é digerido eabsorvido no duodeno e no jejuno.
• Pâncreas:
o A porção exócrina do pâncreas secreta:
Enzimas digestivas;
Fluido rico em iões bicarbonato.A elevada acidez do quimo iria inactivar enzimas pancreáticas se o ácido não fosse
neutralizado pelo bicarbonato.
• Fígado:
334

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 335/374
A bílis contém iões bicarbonato, colesterol, fosfolípidos, pigmentos biliares,produtos metabólicos orgânicos e sais biliares (que solubilizam lípidos).
A bílis é secretada pelo fígado para pequenas ductos que se juntam para formar oducto hepático comum.
Entre refeições, a bílis secretada é armazenada na vesícula biliar, que concentraas moléculas orgânicas da bílis por absorção de sais e H2O.
Durante a refeição, o músculo liso da vesícula contrai, causando a ejecção da bílispara o duodeno através do ducto biliar comum.
• Intestino delgado:Aqui, monossacáridos e aminoácidos, são absorvidos por processos específicosmediados por transportadores de membrana. Os ácidos gordos passam pordifusão.
A maioria dos iões minerais são activamente transportados e H2O difundepassivamente através de gradientes osmóticos.A mobilidade do intestino delgado:i) Mistura o conteúdo luminal com as diversas secreções intestinais,
pancreáticas e bílis.ii) Promove o contacto entre a superfície e o conteúdo.iii) Movimento do conteúdo luminal em direcção ao intestino grossoiv) Completar a digestão.v) Absorver os nutrientes.vi) Impelir o que resta dos alimentos para o cólon.
VILOSIDADE INTESTINAL
Nas criptas de Lieberkuhn forma-se continuamente novas células da orlaintestinal, que migram em 3 a 5 dias para o topo da vilosidade carregadas de enzimas eproteinas de transporte nas suas membranas.
Jejuno e Ileo
• No intestino à medida que se progridediminui a espessura da parede e onúmero de pregas e vilosidades
• Na mucosa e submucosa do ileoexistem numerosos nódulos linfáticos, asplacas de Peyer
335

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 336/374
REFLEXOS INTESTINAIS
• Lei do intestino: A presença de alimento no intestino provoca a contracção da suaparede atrás deste e o relaxamento à frente
• Reflexo intestino-intestinal: A distensão de uma porção do intestino provoca orelaxamento do músculo liso intestinal na sua totalidade
• Reflexo ileogástrico: A secreção e motilidade gástricas aumentam a motilidade doíleon terminal e aceleram a passagem do conteúdo deste para o cego
Estrutura da Parede do Trato GastroIntestinal
336
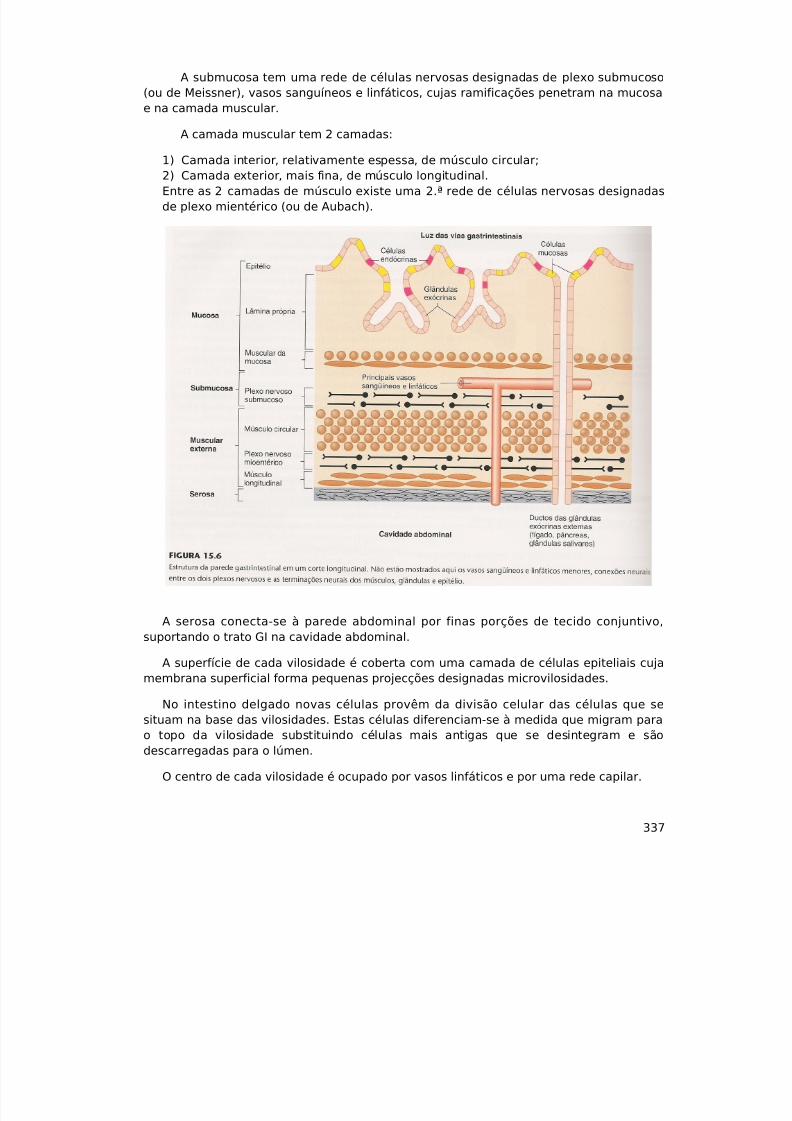
7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 337/374
A submucosa tem uma rede de células nervosas designadas de plexo submucoso(ou de Meissner), vasos sanguíneos e linfáticos, cujas ramificações penetram na mucosae na camada muscular.
A camada muscular tem 2 camadas:
1) Camada interior, relativamente espessa, de músculo circular;2) Camada exterior, mais fina, de músculo longitudinal.Entre as 2 camadas de músculo existe uma 2.ª rede de células nervosas designadasde plexo mientérico (ou de Aubach).
A serosa conecta-se à parede abdominal por finas porções de tecido conjuntivo,suportando o trato GI na cavidade abdominal.
A superfície de cada vilosidade é coberta com uma camada de células epiteliais cujamembrana superficial forma pequenas projecções designadas microvilosidades.
No intestino delgado novas células provêm da divisão celular das células que sesituam na base das vilosidades. Estas células diferenciam-se à medida que migram parao topo da vilosidade substituindo células mais antigas que se desintegram e sãodescarregadas para o lúmen.
O centro de cada vilosidade é ocupado por vasos linfáticos e por uma rede capilar.
337

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 338/374
As veias drenam do intestino delgado, grosso, pâncreas e estômago para a veia portahepática no fígado, sendo o material absorvido nos capilares intestinais processadoantes de entrar na circulação.
O fígado possui enzimas que podem metabolizar compostos tóxicos e lipofílicos quepossam ter sido ingeridos, prevenindo a sua entrada em circulação.
Regulação dos Processos GI
338

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 339/374
Os mecanismos de controlo são regulados não pelo estado nutricional do corpo,mas pelo volume e composição do conteúdo luminal.
Princípios básicos: os reflexos GI são iniciados por:
• Distensão da parede pelo volume do conteúdo luminal;
• Osmoralidade do quimo ([soluto] total)
• Acidez do quimo• [] de produtos específicos (monossacáridos, ácidos gordos, aminoácidos)
Estes estímulos actuam com receptores localizados na parede do trato GI.
Regulação Neural
O Sistema Nervoso Entérico tem 2 redes nervosas: plexo mientérico e plexosubmucoso. A actividade neural num plexo influencia a actividade no outro, e aestimulação num ponto do plexo pode levar a impulsos que são conduzidos tanto paracima como para baixo do trato.
São possíveis 2 tipos de arcos reflexos neurais:
1) Reflexos curtos de receptores através dos plexos nervosos para células efectoras;2) Reflexos longos de receptores no tracto GI para o SNC através de nervos aferentes
e de volta para os plexos nervosos e as células efectoras através de fibras
nervosas autónomas.
A fome, a visão ou o cheiro, ou ainda o estado emocional de um indivíduo pode terefeitos significativos no tracto GI – efeitos mediados pelo SNC através de neuróniosautónomos.
339

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 340/374
Regulação Hormonal
As hormonas que controlam o sistema GI são secretadas principalmente porcélulas endócrinas dispostas no epitélio do estômago e do intestino delgado.
A superfície de cada célula endócrina está exposta ao lúmen do tracto GI, sendo
estimuladas por substâncias químicas do quimo.
Cada hormona participa num sistema de controlo de feedback que regula aspectosdo ambiente luminal. Além disto, cada hormona afecta mais que um tipo de célula alvo.
Nota: CCK – colecistoquinina e GiP – péptido dependente de glucose insulinotrópico
340

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 341/374
Estas hormonas também existem no SNC e nos neurónios do plexo GI, ondefuncionam como neurotransmissores ou neuromoduladores.
Uma única célula efectora tem receptores para mais de uma hormona,neurotransmissores ou agentes parácrinos: potenciação.
A potenciação pode ser exemplificada pela interacção entre a secretina e a CCK. Asecretina estimula a secreção de bicarbonato pancreático e a CCK também, mas oestimulo é mais fraco. A soma dos estímulos faz com que a secreção seja mais intensado que o esperado, isto porque CCK amplifica a acção da secretina.
As hormonas GI têm efeitos trópicos (promotoras do crescimento) em diversostecidos: mucosa gástrica e intestinal, e porções exócrinas do pâncreas.
Fases do Controlo GastroIntestinal
O controlo neural e hormonal do sistema GI é divisível em 3 fases: cefálica,gástrica e intestinal.
I) Fase Cefálica: iniciada quando receptores na cabeça são estimuladaspela visão, sabor, odor e mastigação. É também possível de ser iniciadapor diversos estados emocionais. As vias aferentes para estes reflexossão mediadas por fibras parassimpáticas (nervo vago) que activamneurónios dos plexos nervosos GI, afectando a actividade secretora econtráctil.
II) Fase Gástrica: iniciada por 4 tipos de estímulos que iniciam os reflexos:distensão, acidez, aminoácidos e péptidos. As respostas destesestímulos são mediadas por reflexos neurais curtos e longos e pelalibertação de gastrina.
III) Fase Intestinal: é iniciada pela estimulação do tracto intestinal:distensão, acidez, osmoralidade e diversos produtos digestivos. Émediada por ambos os reflexos neurais (longos e curtos) e pelashormonas GI.
341

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 342/374
342
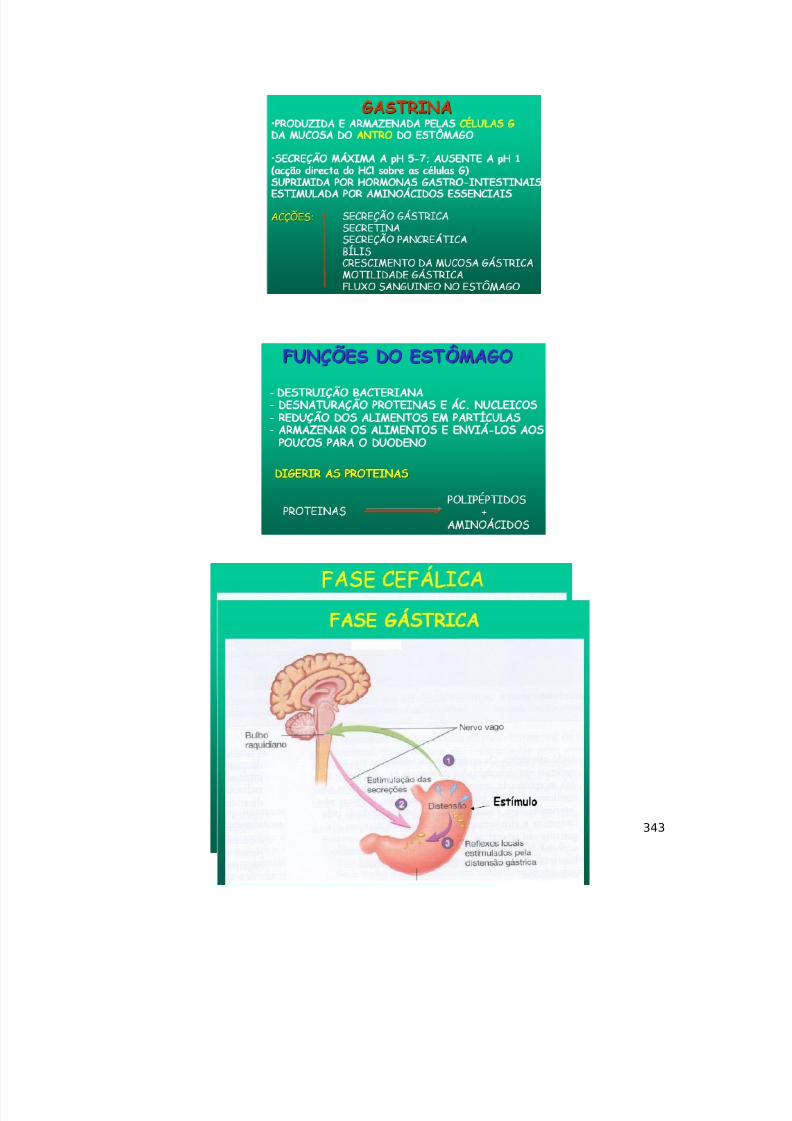
7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 343/374
343

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 344/374
344

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 345/374
Cada uma destas fases é nomeada pelo sítio no qual os estímulos iniciam os reflexose não pelos locais de actividade efectora.
Processos Contrácteis e Secretores:
1. Mastigação (Boca) – é controlada pelos nervos somáticos nos músculosesqueléticos da boca. Também são activados por reflexo, movimentosrítmicos devido à pressão da comida nas gengivas, palato e língua. Apesarde a mastigação prolongar a sensação de sabor, não altera a
2. taxa a que o alimento vai ser digerido e absorvido.3. Saliva (Boca) – a secreção da saliva é controlada tanto por neurónios
simpáticos como parassimpáticos, sendo que ambas estimulam a suasecreção. Na secreção salivar não existe regulação hormonal, sendo que naausência de alimento a taxe de secreção diminui mantendo a boca húmida.No entanto, a secreção salivar pode ser induzida, na fase cefálica, pelocheiro ou visão. A secreção salivar aumenta em resposta ao alimento,induzida por uma resposta reflexiva iniciada por quimioreceptores ereceptores de pressão na boca e língua. O volume de saliva secretada por
grama de tecido é a maior de qualquer glândula do corpo.4. Deglutição (Boca/Faringe/Esófago) – a deglutição é um reflexo complexo
iniciado quando receptores de pressão nas paredes da faringe sãoestimulados por comida ou bebida forçada para o interior da boca pelalíngua. Os receptores enviam impulsos para o centro de deglutição notronco cerebral e este possibilita a deglutição. À medida que o material semove para dentro da faringe, o palato mole é elevado e acomoda-se contraa parede posterior da faringe para prevenir a entrada de comida para acavidade nasal. Os impulsos do centro de deglutição inibem a respiração,
elevam a laringe e fecham a glote, evitando que o alimento se mova paradentro da traqueia. Devido às diferenças de pressão que existem noesófago, o ar e conteúdo gástrico tendem a ir par o esófago. Isto não
345

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 346/374
acontece porque as duas terminações do esófago estão normalmentefechadas por esfíncteres musculares. Músculo esquelético envolve oesófago mesmo abaixo da faringe formando o esfíncter esofágico superior,na última porção do esófago, músculo liso forma o esfíncter esofágicoinferior. A deglutição pode ocorrer em qualquer posição da pessoa pois éatravés de movimentos peristálticos que o alimento se move para o
estômago. A deglutição é um exemplo de um reflexo em que respostasmúltiplas ocorrem numa sequência determinada por um padrão deconexões sinápticas entre neurónios num centro coordenador cerebral.Uma vez que tanto o músculo esquelético como o liso estão envolvidos ocentro da deglutição tem de dirigir actividade eferente nos nervossomáticos e nos nervos autónomos. Simultaneamente fibras aferentes dereceptores na parede esofágica mandam informação para o centro dedeglutição que pode alterar a actividade eferente. A habilidade do esfíncteresofágico inferior para manter a barreira entre o estômago é ajudada pelo
facto da última porção do esófago estar abaixo do diafragma e ser sujeitoàs mesmas pressões abdominais que o estômago.5. Estômago - a camada epitelial que cobre o estômago invagina para a
mucosa formando glândulas tubulares. As glândulas na porção superior doestômago, corpo e fundo, secretam muco, HCl e pepsinogénio. A porçãoinferior do estômago, porção pilórica, tem uma camada muito mais espessade músculo liso. As glândulas nesta zona secretam pouco ácido, mas ascélulas endócrinas secretam gastrina. A delimitar a parede das glândulasestão células parietais que secretam ácido e factor intrínseco, e células quesecretam pepsinogénio. As células enterocromafins libertam agentesparácrinos como a histamina e as células que secretam a somatotastinaestão inseridas ao longo da glândula tubular.
6. Secreção de HCl (estômago) – o estômago secreta cerca de 2L de HCL pordia e a [H+] pode atingir os 150 mM. ATPases Na+/K+ primárias namembrana luminal das células parietais bombeiam iões H+ para o lúmendo estômago. O K+ que é bombeado para dentro da célula volta para olúmen por canais K+. À medida que os iões H+ são secretados para olúmen, iões bicarbonato são secretados ao lado oposto da célula paradentro do sangue em troca de iões Cl-. A elevada secreção ácida é o
resultado da transferência de proteínas ATPases Na+/K+ das membranasdas vesículas intracelulares para a membrana plasmática, por fusão,aumentando o n.º de bombas.
346

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 347/374
Quatro mensageiros químicos regulam a inserção das ATPases H+/K+ namembrana plasmática e por isso a secreção de ácido: gastrina, acetilcolina,histamina e somatóstatina, sendo que as membranas das células parietaispossuem receptores para os 4 agentes. A somatrostatina inibe a secreçãoácida. A histamina potencia a resposta à gastrina e à Ach.
Durante a refeição, a taxa de secreção de ácido aumenta marcadamente à
medida que estímulos provenientes das fases cefálica, gástrica e intestinalalteram a libertação dos 4 mensageiros químicos. Durante a fase cefálica, a
347

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 348/374
elevada actividade dos nervos parassimpáticos resulta na libertação de Achdos neurónios do plexo, gastrina e histamina.
Oestímulo da fase gástrica, distensão, presença de péptidos e aminoácidoslibertados pela digestão de proteínas, produz um aumento ainda maior dasecreção ácida. Estes estímulos utilizam as mesmas vias neurais usadasdurante a fase cefálica. Em adição, péptidos e aminoácidos podem actuardirectamente nas células endócrinas libertadoras de gastrina parapromover a sua secreção. Os H+ estimulam a libertação de somatostatinade células endócrinas na parede gástrica. A somatostatina actua nascélulas parietais para inibir a secreção de ácido, inibe também a libertaçãode gastrina e histamina. O resultado global é um feedback negativo quecontrola a secreção de ácido. Elevar o conteúdo proteico de uma
alimentação aumenta a secreção de ácido: 1.º aumenta a [] de péptidos nolúmen do estômago, o que estimula a secreção de ácido; 2.º as proteínasdos alimentos são um excelente tampão, à medida que as proteínas entramno estômago a []de H+ diminui à medida que H+ se liga às proteínas. Aelevada acidez no duodeno acciona reflexos que inibem a secreção gástricae ácida. Este reflexo limita a produção de ácido gástrico quando os H+ noduodeno aumentam a sua devido à entrada do quimo do estômago. Asecreção ácida durante a fase intestinal varia dependendo do volume ecomposição dos conteúdos intestinais. A inibição da secreção de ácido
gástrico durante a fase intestinal é mediada por reflexos longos e curtos epor hormonas que inibem secreção ácida.
348

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 349/374
7. Secreção de pepsina (estômago) – Apepsina é secretada na forma depercursor inactivo, designadopepsinogénio. A exposição a baixopH no lúmen do estômago causa aconversão de pepsinogénio em
pepsina por quebra de ligações(pH<2). A pepsina pode actuar nopepsinogénio de modo a promovera produção de mais pepsina. Apepsina está activa apenas napresença elevada [H+]. Éirreversivelmente inactivadaquando entra no intestino delgadoonde os H+ são neutralizados pelo
bicarbonato. Durante a fasecefálica, gástrica e intestinal, amaioria dos factores que estimulamou inibem a secreção de ácido, exercem o mesmo efeito na secreção depepsinogénio; são secreções paralelas. A pepsina não é essencial para adigestão proteica, uma vez que as proteínas podem ser completamentedigeridas por enzimas no intestino delgado. A pepsina acelera a digestãode proteínas e é importante na digestão do colagénio.
Motilidade Gástrica
O estômago vazio tem um volume de cerca de 50 mL.
Os músculos lisos do fundo e corpo relaxam antes da chegada da comida
permitindo que o volume aumente até 1,5L.
349

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 350/374
Isto é designado de relaxamento receptivo e é mediado pelos nervosparassimpáticos, coordenados pelo centro de deglutição no cérebro. O óxido nítrico eserotonina libertados por neurónios entéricos medeiam este relaxamento.
Os músculos do esfíncter pilórico contraem na chegada de uma onda peristáltica.Como consequência do fecho do esfíncter, apenas uma pequena quantidade dequimo é expelida para o duodeno em cada onda, e a maioria do conteúdo é forçado
de volta para o corpo do estômago, contribuindo para a actividade de mistura.
Quem produz as ondas peristálticas?
As ondas peristálticas são produzidas pela camada de músculo liso longitudinal.
Estas células são submetidas a ciclos de despolarização-repolarizaçãoespontâneos (ondas lentas) designadas de ritmo básico eléctrico.
Na ausência de inputs neurais ou hormonais, estas despolarizações são demasiadopequenas para causar contracções significativas.
Enquanto a frequência da contracção é determinada pelo ritmo básico eléctricointrínseco e mantém-se constante, a força de contracção e por isso a quantidade quese esvazia de cada vez é determinada reflexamente por inputs neurais e hormonais
para o músculo liso.
350

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 351/374
Todos os factores que regulam a secreção ácida também podem alterar amobilidade gástrica.
O esvaziamento gástrico é inibido por distensões do duodeno, presença degorduras, elevada acidez ou soluções hipertónicas no lúmen do duodeno.
As fibras autónomas para o estômago podem ser activadas pelo SNCindependentemente dos reflexos do estômago e do duodeno. Um aumento daactividade parassimpática aumenta a mobilidade gástrica e a actividade simpáticatem o efeito oposto.
Secreções Pancreáticas
A porção exócrina do pâncreas secreta iões bicarbonato e um número de enzimasdigestivas para ductos que convergem no ducto pancreático que se junta no ducto
biliar comum mesmo antes de entrar no duodeno.
As enzimas são secretadas por células glandulares no final do sistema de ductos eos iões bicarbonato são secretadas por células epiteliais dos ductos.
Iões H+ derivados da reacção entre CO2 e H2O catalizada pela anidrase carbónicasão activamente transportados para fora das células dos ductos e libertos no sangu,enquanto os bicarbonatos são secretados para o lúmen dos ductos.
As enzimas proteolíticas são secretadas na forma inactiva (zimogénios) sendodepois activadas. Esta activação é mediada pela enteroquinase, que está inserida na
membrana plasmática luminal das células epiteliais intestinais. É uma enzimaproteolitica que divide o tripsinogénio pancreático formando a enzima activa tripsina.
A tripsina é uma enzima proteolítica que, uma vez activada, activa os outroszimogénios pancreáticos pela quebra de fragmentos peptidicos. A tripsina tambémdigere proteínas ingeridas.
351

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 352/374
As enzimas não proteoliticas, amilase e lipase, são libertadas na forma activa.
352

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 353/374
As secreções pancreáticas aumentam durante a refeição devido à secretina (queestimula a secreção de bicarbonato) e à CCK (que estimula a secreção de enzimas).
353

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 354/374
A secreção pancreática também pode ser estimulada pelo sabor da comida e peladistensão do estômago.
Secreção de Bílis
A bílis possui 6 componentes principais:
• Sintetizados no fígado, ajudam a solubilizar lípidos1. Sais biliares2. Lecitina• Neutralizam o ácido3. Bicarbonato
• Substâncias extraídas no sangue pelo fígado e excretadas através da bílis
4. Colesterol5. Pigmentos Biliares6. Vestígios de metais
Os sais e lectina são sintetizados nofígado e ajudam a solubilizar lipidos; os iõesbicarbonato neutralizam o ácido e os 3últimos representam substâncias extraídasno sangue pelo fígado e excretadas através
da bílis.
A maioria dos sais biliares que entram notracto intestinal através da bílis sãoabsorvidos por transportadores específicosde acoplamento de Na+ no ileo. Os saisbiliares absorvidos retornam ao fígadoatravés da veia porta, e são activamentetransportados para os hepatócitos-circulaçãoenterohepática.
354

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 355/374
Para além da síntese de sais biliares a partir do colesterol, o fígado também secretacolesterol extraído do sangue para a bílis, que é removido nas fezes. O colesterol éinsolúvel em H2O, mas solúvel na bílis através da sua incorporação em micelas.
Os pigmentos biliares são formados a partir da porção heme da hemoglobina quandoos eritrócitos são destruídos no baço ou no fígado. O pigmento biliar predominante é abilirrubina, extraída do sangue, de cor amarela. Após entrar no trato intestinal, alguma
da bilirrubina é modificada por enzimas bacterianas para formar os pigmentos castanhosque dão cor às fezes.
Os hepatócitos secretam sais biliares, colesterol, lecitina e pigmentos biliares,enquanto as soluções de sais ricos em bicarbonatos são secretados por células epiteliaisdos ductos biliares estimulados pela secretina. A secreção de sais biliares é controladapela sua concentração no sangue.
Existe sempre alguma bílis a ser secretada pelo fígado. A envolver o ducto biliarcomum, no ponto em que entra no duodeno, existe um anel de músculo liso: esfíncter deOddi. Quando este esfíncter está fechado, a bílis diluída secretada pelo fígado é dirigida
para a vesícula biliar.
Intestino Delgado
355

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 356/374
Secreção
Cerca de 1500 mL de fluído é secretado pelas paredes do intestino delgado dosangue para o lúmen. O epitélio intestinal na base das vilosidades secreta Na+, Cl- e
bicarbonato para o lúmen e água por osmose. Estas secreções com muco, lubrificam asuperfície do trato intestinal e ajudam a proteger as células epiteliais de lesõesexcessivas pelas enzimas digestivas no lúmen.
O movimento de H2O para o lúmen também ocorre quando o quimo é hipertónico.
Absorção
Virtualmente todo o fluido secretado pelo intestino delgado é absorvido de novo parao sangue. Em adição, secreções salivares, gástricas, hepáticas, pancreáticas e água sãoabsorvidas.
A absorção de H2O é conseguida pelo transporte de iões, principalmente Na+, dolúmen intestinal para o sangue, seguidos pela H2O por osmose.
Mobilidade
O movimento mais comum é a contracção e relaxamento estacionário de segmentosintestinais com pouco movimento em direcção ao intestino grosso.
Cada segmento contractor tem apenas uns cm de comprimento e a contracção durauns segundos. A contracção rítmica e relaxamento, segmentação, produz uma contínuadivisão e subdivisão do conteúdo intestinal, misturando o quimo e levando-o ao contactocom a parede intestinal. Estes movimentos segmentares são iniciados por actividadeeléctrica gerada por células pacemaker que se associam com a camada muscularcircular.
A frequência da segmentação é determinada pela frequência do ritmo básicoeléctrico intestinal, que varia ao longo do comprimento do intestino em que cada regiãosucessiva tem uma frequência ligeiramente menor do que a região anterior.
A intensidade da segmentação pode ser alterada por hormonas, sistema nervosoentérico, e nervos autónomos. Os inputs produzem alterações na força de contracção domúsculo liso mas não alteram significativamente as frequências dos ritmos eléctricosbásicos.
Após a absorção, as contracções segmentares cessam e são substituídas por umpadrão de actividade peristáltica designado “complexo de migração mioeléctrico”
356

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 357/374
(MMC), começando na última porção do estômago. Ondas repetidas de actividadeperistáltica viajam uma curta distância ao longo do intestino delgado para depoisdesaparecerem; o próximo MMC começa um pouco mais a baixo.
O MMC movimenta qualquer material não digerido que ainda esteja no intestinodelgado, na direcção do intestino grosso e previne que as bactérias fiquem no intestinodelgado.
Pensa-se que a motilidade estimula o MC através dos sistemas nervosos entérico eautónomo.
• Reflexo gastroileal: a intensidade de segmentação no ileo aumenta duranteperíodos de esvaziamento gástrico.
• Reflexo intestino-intestinal: distensões, lesões ou bactérias levam a uma completaparagem da mobilidade.
Intestino grosso
A primeira porção forma um saco de fundo cego-ceco, no qual está o apêndice.
O cólon consiste em 3 segmentos: ascendente, transverso e descendente. A porçãoterminal do cólon descendente tem a forma de S, cólon sigmoide, que se esvazia norecto que termina no ânus.
O intestino grosso tem metade do comprimento do intestino delgado. A sua superfície
é invaginada e a mucosa não tem tantas vilosidades.
As suas secreções são principalmente muco e fluido com bicarbonato e K+.
O quimo entra no ceco através do esfíncter ileocecal. Este está normalmente fechadoe abre aquando da contracção da porção terminal do ileo.
A distensão do intestino grosso, produz uma contracção reflexiva do esfíncterprevenindo o refluxo.
O material que entra no intestino grosso deriva principalmente das secreções da
porção final do intestino delgado, uma vez que a maioria do alimento ingerido já foiabsorvido.
O processo absortivo primário é o transporte activo de Na+ do lúmen para o sanguecom o acompanhamento da água por osmose.
Existe um movimento de K+ do sangue para o intestino, provavelmente devido a ummecanismo activo estimulado pelo AMPc. Existe também um movimento de bicarbonatopara o lúmen, acoplado à absorção de Cl-.
As fibras são metabolizadas para ácidos gordos de cadeias curtas por bactérias e
absorvidas por difusão. O bicarbonato secretado ajuda a neutralizar a elevada acidez queresulta da formação dos ácidos gordos.
357

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 358/374
Mobilidade e Defecação
As contracções do músculo liso circular produzem a segmentação, menor no intestinogrosso que no intestino delgado.
Entre 3 a 4 vezes por dia, uma onda de contracção intensa, movimento de massa,alastra-se no segmento transverso em direcção ao recto. O músculo liso permanececontraído durante algum tempo após um movimento de massa.
A repeitda distensão da parede do recto, produzida pelo movimento de massa domaterial fecal para o seu interior, inicia o reflexo da defecação por mediação neural.
Eventos dos Estados Absortivos e Pós-absortivos
Estado Absortivo: estado durante o qual nutrientes ingeridos estão a entrar no sanguedo trato GI.
Estado Pós-absortivo: estado durante o qual o trato GI está vazio de nutrientes e aenergia tem que ser obtida a partir das reservas corporais.
Estado Absortivo
O sangue deixa o trato GI para se dirigir directamente para o fígado através da veiaporta hepática. Isto permite ao fígado alterar a composição nutricional do sangue antesde regressar ao coração a ser bombeado para o resto do corpo.
Os lípidos são absorvidos para a linfa como triglicéridos. A linfa drena para o sistemanervoso e o fígado não tem a possibilidade de modificar os lípidos absorvidos.
Hidratos de Carbono absorvidos
A glucose é a fonte corporal de maior energia durante este estado.
Os músculos esqueléticos não só catabolizam a glucose durante a fase absortiva mas
também convertem alguma da glucose a glicogénio que é armazenado.
358

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 359/374
Nos adipócitos a glucose é transformada em triglicéridos. A glucose é percursora tantodo fosfato α-glicerol como dos ácidos gordos e estas moléculas são ligadas para formartriglicéridos.
Durante este período, o fígado recolhe glucose que pode ser armazenada comoglicogénio ou transformada em fosfato α-glicerol e ácidos gordos dando origem atriglicéridos.
Alguns dos lipidos sintetizados no fígado a partir da glucose são aí armazenados masa maioria é empacotada com proteínas específicas formando as lipoproteínas dedensidade muito baixo (VLDL), pois possuem mais lípidos que proteínas.
Na corrente sanguínea os trigliceridos das VLDL são hidrolizados a monoglicéridos eácidos gordos pela enzima lipoproteína lipase. Esta enzima está localizada na superfíciedas células epiteliais dos capilares, especialmente no tecido adiposo, onde os ácidosgordos gerados difundem para os adipócitos. Aí, os ácidos gordos combinam com ofosfato α-glicerol para formar triglicéridos.
Triglicéridos Absorvidos
Os ácidos gordos são libertados principalmente em capilares do tecido adiposo pelaacção da lipoproteína lipase. Os ácidos gordos entram nos adipócitos e combinam com ofosfato α-glicerol sintetizado nos adipócitos a partir de metabolitos da glicose paraformar triglicéridos.
Aminoácidos Absorvidos
Alguns aa absorvidos entram nas células do fígado e são utilizados para sintetizarproteínas ou são convertidas em α-cetoácidos pela remoção do grupo amina que éutilizado para sintetizar ureia.
Os α-cetoácidos podem entrar no ciclo de Krebs do ácido tricarboxílico e podem sercatabolizados para providenciar energia. Podem também ser convertidos em ácidosgordos, participando na síntese de lípidos.
Durante este período ocorre uma síntese de proteínas mas apenas para restabeleceras proteínas catabolizadas durante o período pós-absortivo.
359

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 360/374
Estado Pós-Absortivo
À medida que o estado absortivotermina, a síntese de glicogénio,lípidos e proteínas cessa e ocatabolismo de todas estassubstâncias inicia-se.
Os eventos que mantêm aconcentração plasmática deglucose são:
1) Reacções que providenciamglucose no sangue;
2) Utilização celular de lípidospara energia, poupando a glucose.
360

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 361/374
Fontes de Glucose no sangue:
1. Glicogénese – a hidrólise de glicogénio armazenado ocorre no fígado e músculoesquelético. A glicogénese hepática é a primeira linha de defesa e começa em
segundos perante estímulos apropriados-activação do SNS. O músculo não tem aenzima necessária para formar glucose a partir da glucose-6-fosfato formadadurante a glicogénese. Por isso, a glucose-6-fosfato é submetida a glicólise paraoriginar piruvato e lactase, que entram no sangue e são convertidos em glucose nofígado.
2. Catabolismo de triglicéridos, em tecidos adiposos, leva à formação de glicerol eácidos gordos – lipólise. O glicerol e ácidos gordos entram no sangue e o 1.º éconvertido em glucose no fígado.
3. Proteínas – as dos músculos e outros tecidos podem ser catabolizadas sem maufuncionamento celular. A síntese de glucose a partir do piruvato, lactato, glicerol e
aa designase-se gluconeogénese.
361

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 362/374
Resumo do Metabolismo dos Nutrientes durante o Período Absortivo
1) A energia é fornecida principalmente pelos hidratos de carbono ingeridos numarefeição.
2) Há captação final de glicose pelo fígado;3) Parte dos hidratos de carbono é armazenada sob a forma de glicogénio no fígado
e no músculo, porém a maioria dos hidratos de carbono e a gordura em excessousada para energia são armazenadas principalmente como gordura no tecidoadiposo;
4) Há alguma síntese de proteínas corporais, porém parte dos aa das proteínas dadieta é usada como energia ou convertida em gordura.
Resumo do Metabolismo de Nutrientes durante o Período Pós-Absortivo
362

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 363/374
1) A síntese de glicogénio, gorduras e proteínas é interrompida e ocorre degradaçãofinal;
2) A glucose é formada no fígado tanto a partir de glicogénio aqui armazenado comotambém por gliconeogénese a partir do lactato, piruvato, glicerol e aatransportados pelo sangue. Os rins também realizam gluconeogénese durante o
jejum prolongado.3) A glucose produzida no fígado (e rins) é libertada na corrente sanguínea, porém a
sua utilização como energia é muito reduzida nos músculos e tecidos não neurais.4) A lipólise liberta ácidos gordos do tecido adiposo na corrente sanguínea, e a
oxidação destes ácidos pela maioria das células e das cetonas produzidas a partirdestes pelo fígado fornece a maior parte do suprimento energético ao corpo.
5) O cérebro continua a usar glucose, mas também usa cetonas à medida que estasse acumulam no sangue.
Controlo Neural e Endócrino dos estados
1. O que controla o anabolismo dasproteínas, glicogénio e triglicéridos nafase absortiva e o catabolismo na fasepós-absortiva?
2. O que induz a utilização primária daglucose pelas células durante a fase
absortiva e a utilização de lípidos nafase pós-absortiva?
363

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 364/374
3. O que conduz ao aprisionamento de glucose pelo fígado na fase absortiva e a sualibertação na fase pós-absortiva?
A insulina e o glucagon são hormonas peptidicas secretadas pelos ilhéus deLangerhans. As células β são as produtoras de insulina e as α as de glucagon.
Insulina
A sua secreção aumenta durante o estado absortivo e diminui no estado pós-absortivo. Os seus efeitos metabólicos são exercidos principalmente nas célulasmusculares, células do tecido adiposo e do fígado. A elevada [insulina] no plasma é amaior causa dos eventos do estado absortivo.
Como todas as hormonas péptidicas, a insulina induz os seus efeitos através da
ligação a receptores específicos na membrana plasmática, das suas células alvo. Estaligação acciona vias de transdução de sinal que influenciam as proteínas de transporteda membrana e enzimas intracelulares.
Nas células musculares, ou nos adipócitos, a insulina estimula vesículascitoplasmáticas que possuem transportador da glucose, Glut-4, na sua membrana afundir com a membrana plasmática.
364

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 365/374
Controlo da Secreção de insulina
O maior factor controlador é a [] plasmática de glucose. No entanto, a elevada []de aa também estimula a secreção de insulina.
A hormona péptido insulinotrópico dependente da glucose (GIP) secretada porcélulas endócrinas no trato GI em reposta à alimentação, também estimula a libertação
de insulina.Os neurónios autónomos também influenciam a secreção de insulina. A activação
dos neurónios parassimpáticos, que ocorre durante a ingestão de uma refeição, estimulaa secreção de insulina.
Em contraste, a activação de neurónios simpáticos nos ilhéus ou um aumento da[] plasmática de epinefrina inibe a secreção de insulina.
365

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 366/374
Principais controlos da secreção de insulina
Glucagon
Os366

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 367/374
maiores efeitos fisiológicos do glucagon são no fígado e são os opostos dos efeitos dainsulina.
1. Aumenta a quebra de glicogénio;2. Aumenta a gluconeogénese;3. Aumenta a síntese de cetonas.
O maior estímulo para a secreção de glucagon é a hipoglicémia. Uma diminuição da []plasmática de glucose induz uma elevada libertação de glucagon, que pelos seus efeitosno metabolismo, serve para restaurar a [] normal de glucose sanguínea pela glicogéneseou glicogenólise, ao mesmo tempo que suplementa ácidos gordos e cetonas.
Epinefrina e Nervos Simpáticos para o Fígado e Tecido Adiposo
A Epinefrina estimula:
1. Glicogenólise no fígado e músculo esquelético;2. Gluconeogénese no fígado;3. Lipólise nos adipócitos.A activação dos nervos simpáticos no fígado e tecido adiposo leva às mesmas
respostas por estes órgãos que a circulação de epinefrina.
Nos adipócitos a epinefrina estimula a actividade da enzima hormona sensível àlipase (HSL), que causa a quebra de triglicéridos a ácidos gordos livres e glicerol. Ambos
367

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 368/374
são libertados para o sangue onde servem de combustível ou de percursores para agluconeogénese.
Quando a [] plasmática de glucose diminui, os receptores para o glucose no SNCiniciam reflexos que levam a elevada actividade nas vias simpáticas para a medulaadrenal; fígado e tecido adiposo.
Cortisol
O cortisol tem um papel permissivo; osníveis de cortisol plasmático não necessitam deaumentar durante o jejum mas a presença decortisol no sangue mantém as [] ideais dasenzimas do fígado e tecido adiposo requeridapara a gluconeogénese e lipólise.
368

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 369/374
Por isso, em resposta ao jejum pessoas com deficiência em cortisol desenvolvemhipoglicémia de tal forma que pode interferir com o funcionamento cerebral.
Pessoas com níveis muito elevados de cortisol no plasma podem desenvolversintomas idênticos aos das pessoas cujas células não respondem correctamente àinsulina.
Hormona do Crescimento
1. Torna os adipócitos mais activos a estímulos lipolítico;2. Aumenta a gluconeogénese no fígado;3. Reduz a habilidade da insulina para a captura de glucose por músculos e tecido
adiposo.
369

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 370/374
Slides
370

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 371/374
FUNÇÕES DO FÍGADO
• Metabolismo dos carbohidratos
• Metabolismo dos lípidos
• Metabolismo das proteinas
• Armazenamento
• Síntese da bílis (600 a 1000 ml/dia)
• Destoxicação e protecção
DESTOXICAÇÃO E PROTECÇÃO
• Metabolização de hormonas esteroides e mediad. químicos
• Depuração sanguínea de bactérias e outros antigénios da circulação portal (cél.Kupffer)
• Metabolização de fármacos pelas oxidases microssomais e através de sistemas deconjugação
371

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 372/374
• Síntese e export. de glutatião (principal antioxidante celular)
• Depuração do sangue de células e proteinas danificadas e dos factores de coagulaçãoactivados
• Metabolização da amónia através do ciclo da ureia
Absorção de Água
372

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 373/374
Fisiologia Especial
Fisiologia da Resposta ao Stress
Stress – qualquer alteração do ambiente interno ou externo que provoque umdesequilíbrio à homeostasia.
Quer se trate de choque, exposição prolongada ao frio, medo ou dor a respostaintegrada ao stress processa-se sempre de forma igual.
Cortisol
Perante uma situação de stress, há um aumento da secreção de cortisol. Esteaumento é mediado pelo eixo hipotálamo-hipófise.
Assim há um estímulo hipotalâmico que leva a secreção de CRH. Esta hormona étransportada pelo sistema porta até à hipófise anterior, onde a vai estimular a secretarACTH. Por sua vez, a ACTH circula no sangue até ao córtex supra-renal onde estimula alibertação de cortisol, tendo este diversas funções numa situação de stress.
O cortisol terá então efeitos sobre o metabolismo orgânico, onde estimula ocatabolismo proteico, a gluconeogénese no fígado, inibição da função da insulina e a
estimulação de catabolismo dos triglicéridos no tecido adiposo. Além destes efeitos, ocortisol aumenta a capacidade de manter vasoconstrição em resposta à norepinefrina(aumenta reactividade vascular); inibe a inflamação e respostas imunitárias específicas,e por fim inibe funções não essenciais, como por exemplo: função reprodutora,crescimento, etc.
Funções do SNS
O SNS estimula a medula supra-renal a libertar epinefrina. Desta forma, o SNS:
- Aumenta a função cardíaca;
- Diminui a fadiga do músculo esquelético;
- Promove a vasoconstrição nas vísceras desviando o sangue para o músculoesquelético;
- Promove o aumento da glicogenólise hepática e muscular e o aumento da degradaçãode triglicéridos no tecido adiposo;
- Aumenta a ventilação pulmonar através do estímulo dos centros respiratórios cerebraise dilatação das vias respiratórias.
373

7/23/2019 Sebenta de Fisiologia
http://slidepdf.com/reader/full/sebenta-de-fisiologia 374/374
Outras hormonas
Para além do CRH, ACTH e cortisol, várias hormonas são libertadas durante o
stress. A -endorfina é co-libertada com ACTH para reduzir a dor. A vasopressinaestimula a secreção de ACTH que também actua para aumentar a retenção de água.
São também aumentadas as secreções de aldosterona, que regula a excreção de Na+ eK+ pelos rins, a hormona do crescimento e o glucagon, relacionada com o metabolismoda glucose.
Durante o stress é diminuída a secreção de insulina.
Manifestações Físicas:
• Aumenta o açúcar no sangue e o sistema digestivo atrasa-se;
• Aumento da contracção e dos batimentos cardíacos;
• Dilatação das vias respiratórias;
• Contracção muscular;
• Aumento da capacidade de focagem;
• Aumento da pressão sanguínea;
• Contracção dos vasos superficiais da pele;
•
Aumento da facilidade de coagulação sanguínea;• Contracção do músculo erector do pelo.
Regulação da Temperatura Corporal
A temperatura corporal central apresenta um ritmo circadiano, sendo mais altadurante o dia e mais baixa durante a noite.
As trocas de calor corporal com o meio externo ocorrem por radiação (em que háemissão de calor pelo corpo), condução (quando a perda de calor se dá por contactofísico com substancias mais frias, ou ganho se o contacto for com substâncias maisquentes) e convecção (perda ou ganho de calor por movimento do ar ou água próximosdo corpo). Para além disto, pode ocorrer também perda de calor por evaporação da águada pele e membranas que revestem as vias respiratórias.
O hipotálamo e outras áreas cerebrais contêm os centros de integração para osreflexos reguladores da temperatura corporal, sendo que tanto os receptores periféricos


















