
Research Article Toxicity of Superparamagnetic Iron Oxide Nanoparticles...
12
Hindawi Publishing Corporation BioMed Research International Volume 2013, Article ID 647974, 11 pages http://dx.doi.org/10.1155/2013/647974 Research Article Toxicity of Superparamagnetic Iron Oxide Nanoparticles on Green Alga Chlorella vulgaris Lotfi Barhoumi 1,2 and David Dewez 1 1 D´ epartement de Chimie, Universit´ e du Qu´ ebec ` a Montr´ eal, CP 8888, Succursale Centre-Ville, Montr´ eal, QC, Canada H3C 3P8 2 Laboratoire de Physiologie Int´ egr´ ee, Facult´ e des Sciences de Bizerte, Universit´ e de Carthage, Zarzouna 7021, Tunisia Correspondence should be addressed to David Dewez; [email protected] Received 31 July 2013; Revised 29 October 2013; Accepted 4 November 2013 Academic Editor: Aiyagari Ramesh Copyright © 2013 L. Barhoumi and D. Dewez. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Toxicity of superparamagnetic iron oxide nanoparticles (SPION) was investigated on Chlorella vulgaris cells exposed during 72 hours to Fe 3 O 4 (SPION-1), Co 0.2 Zn 0.8 Fe 2 O 4 (SPION-2), or Co 0.5 Zn 0.5 Fe 2 O 4 (SPION-3) to a range of concentrations from 12.5 to 400 g mL −1 . Under these treatments, toxicity impact was indicated by the deterioration of photochemical activities of photosynthesis, the induction of oxidative stress, and the inhibition of cell division rate. In comparison to SPION-2 and - 3, exposure to SPION-1 caused the highest toxic effects on cellular division due to a stronger production of reactive oxygen species and deterioration of photochemical activity of Photosystem II. is study showed the potential source of toxicity for three SPION suspensions, having different chemical compositions, estimated by the change of different biomarkers. In this toxicological investigation, algal model C. vulgaris demonstrated to be a valuable bioindicator of SPION toxicity. 1. Introduction Due to their physicochemical features, superparamagnetic iron oxide nanoparticles (SPION) are widely used in medical applications such as contrast agents for magnetic resonance imaging and heating mediators for cancer therapy [1, 2]. A recent review of SPION-induced toxicity studies at cellular level in animal and human cells indicated that SPION can penetrate the cellular system by both passive diffusion and endocytosis, causing several toxic effects through the alteration of genes expressions and the generation of oxida- tive radicals [3]. However, their production and extensive use may contaminate aquatic environments via wastewater input, representing a risk of toxicity for different freshwater organisms. Besides endocrine disruptor and pharmaceu- ticals, metallic engineered nanoparticles represent one of the most important hazardous materials altering freshwater qualities. Many studies showed that the toxicity of metallic engineered nanoparticles was directly related to their sur- face chemistry, hydrodynamic size, chemical composition, and solubility in aqueous solution [4, 5]. A recent study demonstrated that SPION have antibacterial properties with the ability to alter metabolic functions at a higher efficiency than antibiotics or metals salts [6]. erefore, it is of high importance to determine the toxicity potential at cellular level of hazardous metallic nanomaterials in relation to their uptake by aquatic microorganisms. In aquatic environments, green algae represent the main source of biomass production essential for animals of higher ecological trophic levels. As being able to bioaccumulate metallic contaminants, algae can be used as a bioindicator of aquatic ecosystem health. However, bioaccumulation effects of SPION on algae have been poorly examined, and such toxicological studies were mostly done on terrestrial plant species grown hydroponi- cally [7, 8]. Currently, there is only one study concerning the toxic effects of Fe 3 O 4 nanoparticles (35 nm) on the green alga Chlorella vulgaris treated 72 h to a nominal concentration range from 200 to 1600 g mL −1 . In this study, authors showed an induction of oxidative stress and an alteration of photosynthetic activity based on absorbed CO 2 fixation [9]. erefore, a more in-depth toxicological investigation needs to be performed to better characterise the toxicity of SPION on the cell physiology of green algae.
Transcript of Research Article Toxicity of Superparamagnetic Iron Oxide Nanoparticles...

Hindawi Publishing CorporationBioMed Research InternationalVolume 2013 Article ID 647974 11 pageshttpdxdoiorg1011552013647974
Research ArticleToxicity of Superparamagnetic Iron Oxide Nanoparticles onGreen Alga Chlorella vulgaris
Lotfi Barhoumi12 and David Dewez1
1 Departement de Chimie Universite du Quebec a Montreal CP 8888 Succursale Centre-Ville Montreal QC Canada H3C 3P82 Laboratoire de Physiologie Integree Faculte des Sciences de Bizerte Universite de Carthage Zarzouna 7021 Tunisia
Correspondence should be addressed to David Dewez dewezdaviduqamca
Received 31 July 2013 Revised 29 October 2013 Accepted 4 November 2013
Academic Editor Aiyagari Ramesh
Copyright copy 2013 L Barhoumi and D Dewez This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Toxicity of superparamagnetic iron oxide nanoparticles (SPION) was investigated on Chlorella vulgaris cells exposed during72 hours to Fe
3O4(SPION-1) Co
02Zn08Fe2O4(SPION-2) or Co
05Zn05Fe2O4(SPION-3) to a range of concentrations from
125 to 400 120583gmLminus1 Under these treatments toxicity impact was indicated by the deterioration of photochemical activitiesof photosynthesis the induction of oxidative stress and the inhibition of cell division rate In comparison to SPION-2 and -3 exposure to SPION-1 caused the highest toxic effects on cellular division due to a stronger production of reactive oxygenspecies and deterioration of photochemical activity of Photosystem II This study showed the potential source of toxicity for threeSPION suspensions having different chemical compositions estimated by the change of different biomarkers In this toxicologicalinvestigation algal model C vulgaris demonstrated to be a valuable bioindicator of SPION toxicity
1 Introduction
Due to their physicochemical features superparamagneticiron oxide nanoparticles (SPION) are widely used in medicalapplications such as contrast agents for magnetic resonanceimaging and heating mediators for cancer therapy [1 2] Arecent review of SPION-induced toxicity studies at cellularlevel in animal and human cells indicated that SPIONcan penetrate the cellular system by both passive diffusionand endocytosis causing several toxic effects through thealteration of genes expressions and the generation of oxida-tive radicals [3] However their production and extensiveuse may contaminate aquatic environments via wastewaterinput representing a risk of toxicity for different freshwaterorganisms Besides endocrine disruptor and pharmaceu-ticals metallic engineered nanoparticles represent one ofthe most important hazardous materials altering freshwaterqualities Many studies showed that the toxicity of metallicengineered nanoparticles was directly related to their sur-face chemistry hydrodynamic size chemical compositionand solubility in aqueous solution [4 5] A recent studydemonstrated that SPION have antibacterial properties with
the ability to alter metabolic functions at a higher efficiencythan antibiotics or metals salts [6] Therefore it is of highimportance to determine the toxicity potential at cellularlevel of hazardous metallic nanomaterials in relation to theiruptake by aquatic microorganisms In aquatic environmentsgreen algae represent the main source of biomass productionessential for animals of higher ecological trophic levels Asbeing able to bioaccumulate metallic contaminants algaecan be used as a bioindicator of aquatic ecosystem healthHowever bioaccumulation effects of SPION on algae havebeen poorly examined and such toxicological studies weremostly done on terrestrial plant species grown hydroponi-cally [7 8] Currently there is only one study concerningthe toxic effects of Fe
3O4nanoparticles (35 nm) on the green
algaChlorella vulgaris treated 72 h to a nominal concentrationrange from 200 to 1600 120583gmLminus1 In this study authorsshowed an induction of oxidative stress and an alteration ofphotosynthetic activity based on absorbed CO
2fixation [9]
Therefore a more in-depth toxicological investigation needsto be performed to better characterise the toxicity of SPIONon the cell physiology of green algae
2 BioMed Research International
In the present study the green alga Chlorella vulgariswas used as a unicellular plant model organism for the tox-icity characterisation of Fe
3O4(SPION-1) Co
02Zn08Fe2O4
(SPION-2) and Co05Zn05Fe2O4(SPION-3) Algal cells were
exposed during 24 48 and 72 hours in order to evaluate theevolution of SPION toxicity impact on the entire cellular sys-tem by evaluating the change in photochemical reactions ofphotosynthesis cell division and the induction of oxidativestress This work permitted determining the risk of SPIONtoxicity on the viability of green algae and therefore thepotential use of this algal species in a bioassay of SPIONtoxicity
2 Material and Methods
21 Biological Material The freshwater green alga Chlorellavulgariswas obtained from theCanadianPhycological CultureCentre (CPCC University of Waterloo ON Canada) Algalculture was prepared in sterile BG-11 liquid growth medium(pH = 7 ionic strength = 00201) having the following finalconcentrations of salts composition [10] 15 g Lminus1 of NaNO
3
004 g Lminus1 of K2HPO4sdot3H2O 0075 g Lminus1 of MgSO
4sdot7H2O
0036 g Lminus1 of CaCl2sdot2H2O 6 times 10minus3 g Lminus1 of C
6H8O7(citric
acid) 6 times 10minus3 g Lminus1 of C6H5FeO7(ferric citrate) 10minus3 g Lminus1
of Na2EDTAsdot2H
2O 002 g Lminus1 of Na
2CO3 286mg Lminus1
of H3BO3 181mg Lminus1 of MnCl
2sdot4H2O 0222mg Lminus1 of
ZnSO4sdot7H2O 0390mg Lminus1 ofNa
2MoO4sdot2H2O 0079mg Lminus1
of CuSO4sdot5H2O and 00494mg Lminus1 of Co(NO
3)2sdot6H2O
Algal cells were grown under continuous light intensity of100 120583molmminus2 sminus1 (SYLVANIA GRO-LUX Wide Spectrumlight F40GRQAQWS) at 24∘C plusmn 1 Aliquots of algalsamples were used for experiments when algal culturereached the exponential growth phase The change of celldensity was determined by monitoring the optical densityat 750 nm and the calculation was based on a standardcorrelation with the cell density measured using a multisizerZ3 (Beckman Coulter Inc USA)
22 Synthesis of SPION Superparamagnetic iron oxidenanoparticles (SPION) as Fe
3O4(SPION-1) Co
02Zn08Fe2O4
(SPION-2) and Co05Zn05Fe2O4(SPION-3) were produced
according to the procedure reported in [11] by usingthe polyol process starting from Co(CH
3CO2)2∙4H2O Zn
(CH3CO2)2∙2H2O and Fe(CH
3CO2)2as precursor salts and
diethylene glycol as a solvent
23 Stock Solution and SPION Characterization In thepreparation of stock solutions SPION were suspended inculture medium at a concentration of 1 g Lminus1 and homoge-nized by ultrasonication during 30min at 4∘C to break upagglomerates After sonication stock solutions weremix witha vortex for 1min and various concentrations of SPIONwereprepared in culture medium for experiments Size distribu-tionwas determined by dynamic light scattering (DLS) with aZetaPlus particle sizer (Brookhaven InstrumentCorporationUSA) using 90Plus Particles Sizing Software (Ver 420)Zeta potential of SPION suspensions in culture medium was
0
20
40
60
80
100
120
0 200 400 600
SPION-1SPION-2 SPION-3
Part
icle
s fra
ctio
ns
Diameter (nm)
Figure 1 Particle size distribution of SPION-1 (Fe3O4) SPION-2
(Co02Zn08Fe2O4) and SPION-3 (Co
05Zn05Fe2O4) suspensions in
the culture medium
0
2
4
6
8
10
12
14
16
0 100 200 300 400
24h48h72h
Solu
ble [
Fe] (120583
g mLminus
1)
[SPION-1] (120583g mLminus1)
Figure 2 Soluble fraction of free Fe released from SPION-1 (Fe3O4)
suspension in culture medium at 24 48 and 72 h of exposure
evaluated by the electrophoretic mobility method with theZetaPlus system
24 Experimental Treatments In each treatment conditionthe initial density of algal cells was of 106 cells mLminus1 in a finalvolume of 50mL Algal samples were exposed during 24 48and 72 h to 125 25 50 100 200 and 400mg Lminus1 of SPION-1 SPION-2 or SPION-3 under the same illumination andtemperature condition used for growing stock cultures Forthe control sample the samemedia compositionwas used butwithout any trace of SPION
BioMed Research International 3
0 100 200 300 4000
5
10
15Fe
Solu
ble [
Fe] (120583
g mLminus
1)
[SPION-2] (120583g mLminus1)
(a)
0
2
4
6
8
10Co
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Solu
ble
[Co]
(120583g m
Lminus1)
(b)
0
2
4
6
8
10
0 100 200 300 400
Zn
[SPION-2] (120583g mLminus1)24h48h72h
Solu
ble [
Zn] (120583
g mLminus
1)
(c)
Figure 3 Soluble fraction of free Fe Co and Zn released from SPION-2 (Co02Zn08Fe2O4) suspension in culture medium at 24 48 and 72 h
of exposure
25 Growth Inhibition Test The relative cell division rate(RCDR) was evaluated for 72 h according to [12] as RCDR =(lnW
72 hminus lnW0)72 h whereW72 h represents cell density at72 h and W
0the initial cell density
26 Bioaccumulation of Fe Co and Zn To separate SPIONfrom cells a sucrose gradient prepared in BG-11 media wasdone directly in a Beckman centrifuge tube inclined at a 30∘angle in order to get 6 layers of different sucrose densities(5mL of each sucrose solution 120 100 80 60 40 and20) Algal cells of control and SPION-treated samples werecollected by centrifugation Their pellets were slowly placedon top of sucrose gradient tubes which were centrifuged at1000 rpm during 30min in a swinging-bucket 5810R cen-trifuge (Eppendorf Germany) It resulted in the formationof a pellet of SPION at the bottom of the tube The algal cellslayer was recuperated with a glass Pasteur pipette and filtered
on a 045 120583m filter previously dried and weighted To removeSPIONweakly bound to the cell surface or the filter 3times 10mLof 10mM ethylenediaminetetraacetic acid in BG-11 mediumwas slowly passed through the filter Filters were dried at 95∘Cfor 24 h and weighted to calculate algal dry weight Filterswere then placed in acid-washed glass tubes in which 4mL ofHNO
3and 500 120583L H
2O2were added Samples were digested
during 48 h at room temperature before being diluted to 20HNO
3in Milli-Q purified water for the quantification of Fe
Co and Zn using atomic absorption spectrometry analysis(Varian SpectrAA 220 FS USA) Obtained Fe Co and Znconcentrations were normalized to the dry weight
27 Soluble Fraction of Fe Co and Zn Solubility of free FeCo and Zn released from SPION suspensions were deter-mined in culturemedium SPION suspensions was incubatedduring 24 48 and 72 h in the same condition as described
4 BioMed Research International
0
1
2
3
4Fe
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Solu
ble [
Fe] (120583
g mLminus
1)
(a)
0
1
2
3
4Co
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Solu
ble
[Co]
(120583g m
Lminus1)
(b)
0
1
2
3
4
0 100 200 300 400
Zn
[SPION-3] (120583g mLminus1)24h48h72h
Solu
ble [
Zn] (120583
g mLminus
1)
(c)
Figure 4 Soluble fraction of Fe Co and Zn released from SPION-3 (Co05Zn05Fe2O4) suspension in culture medium at 24 48 and 72 h of
exposure
above for treatment After incubation NPs were removed bycentrifugation at 12000 g for 30min and the supernatant wascollected for analysis Quantification of free Fe Co and Znin solution wasmeasured by atomic absorption spectrometry(Varian SpectrAA 220 FS USA)
28 Production of ROS The fluorescent dye 2101584071015840-dichlorodi-hydrofluorescein diacetate ((H
2DCFDA) Invitrogen Molec-
ular Probe USA) was used as an indicator of ROS accordingto [13] Cellular esterases hydrolyze this probe into the non-fluorescent compound 2101584071015840-dichlorodihydrofluorescein(H2DCF) which is better retained within cells In the
presence of ROS and cellular esterases H2DCF is trans-
formed into the highly fluorescent compound 2101584071015840-dichlo-rofluorescein (DCF) H
2DCFDA stock solution (10mM) was
prepared in ethanol in the dark After 72 h of treatment 1mLof algal samples was exposed during 15min to 02mM of
H2DCFDA in the dark The ROS level was determined by
measuring the fluorescence emission at 530 nm with a flowcytometer (FACScan system Becton Dickinson InstrumentsUSA) Cytometry results were analysed using the WinMDI28 software Algal cells were separated from noncellularparticles by using a relationship between particle size and redfluorescence level originating from chlorophyll fluorescenceemission A positive control sample using methyl viologenwas done to verify the assay (data not shown)
29 Cellular Viability Viability of algal cells was estimatedusing the fluorescein diacetate ((FDA) Invitrogen MolecularProbe USA) method according to [14] FDA is a nonpolarester compoundwhich passes through cell membranes Onceinside the cell FDA is hydrolyzed by esterases (enzymespresent in viable cells) to produce fluorescein accumulatingin cell wall and emitting fluorescence under UV illumination
BioMed Research International 5
After 72 h of treatment 1mL of algal samples was exposedduring 15min to 02mM of FDA in the dark Cell viabilitywas determined by measuring the fluorescence emissionat 530 nm with a flow cytometer (FACScan System Bec-ton Dickinson Instruments USA) Cytometry results wereanalysed using the WinMDI 28 software Algal cells wereseparated from noncellular particles by using a relationshipbetween particle size and red fluorescence level originatingfrom chlorophyll fluorescence emission A positive controlsample using methyl viologen was done to verify the assay(data not shown)
210 Chl 119886 Fluorescence Emission Photosynthetic electrontransport was monitored from the change in the rapid riseof Chl 119886 fluorescence emission using a ldquoPlant EfficiencyAnalyserrdquo fluorometer (Handy-PEA Hansatech Ltd UK)Total chlorophyll (Chl) content (119886 + 119887) was extracted in 100methanol at 65∘C and quantified with a spectrophotometer(Lambda 40 Perkin-Elmer USA) according to the formulaindicated in [15] Total Chl (120583gmLminus1) = (2493 times A
6524+
144 times A6652
)Prior to fluorescence measurements algal samples were
transferred into clean sterile 2mL glass vials and dark-adapted for 30min An aliquot of 5 120583g of total chlorophyllwas gently filtered using low pressure filtration and algal cellswere uniformly placed on a 13mm glass fibre filter (MilliporeUSA) The fluorescence induction was triggered using a 1 ssaturating flash of 3500120583molmminus2 sminus1The fluorescence inten-sity at 20120583s was considered as the 119874 value (119865
119874) fluorescence
intensities for 119870 119869 and 119868 transients were determined at300 120583s (119865
119870) 2ms (119865
119869) and 30ms (119865
119868) respectively the max-
imum fluorescence yield reached maximal value of fluores-cence intensity (119865
119872) under saturating illumination Different
photosynthetic-based fluorescence parameters related to thefunctional state of Photosystem II were calculated [16 17]the maximum efficiency of PSII electron transport 119865
119881119865119872=
(119865119872minus 119865119874)119865119872 the absorption of photons by light harvesting
antenna complexes (ABS) per active reaction center (RC)ABSRC = ((119865
119870minus 119865119874)025) times (1(119865
119869minus 119865119874)) times (119865
119872(119865119872minus
119865119874)) the relative variable fluorescence yield at 119869 transient
estimating the fraction of 119876119860
in its reduced state 119881119869=
(119865119869minus 119865119874)(119865119872minus 119865119874) and the performance index of PSII
photosynthetic activity PI = RCABS times ((119865119872minus 119865119874)119865119874) times
((1 minus 119881119869)119881119869)
211 Statistical Analysis All treatments were performed intriplicate Means and standard deviations were calculatedfor each treatment Significant differences between controland treated plants were determined by one-way analysisof variance (ANOVA) followed by a Dunnettrsquos multiplecomparison (DMC) test where 119875 value less than 005 wasconsidered significant
3 Results
31 Characterisation of SPION When SPION were sus-pended in the media nanoparticles formed agglomeratesduring the first minutes as indicated by the distribution
0
2
4
6
8
10
12
0 100 200 300 400
Bioa
ccum
ulat
ed F
e120583g
mg
of D
W
[SPION-1] (120583g mLminus1)
Figure 5 Bioaccumulated content of Fe in algal cells of Chlorellavulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square) and 72 h(998771 triangle) to different concentrations of SPION-1 (Fe
3O4)
of hydrodynamic particles size diameter which was causedby the content of salts in the media Analysis by dynamiclight scattering showed SPION-1 SPION-2 and SPION-3suspensions in culture media to have an average diameterof particle size distribution of 1959 1765 and 3472 nmrespectively (Figure 1) These distributions of hydrodynamicsize of SPION were found to be stable in the culture mediumduring the entire experimental exposure Furthermore mea-surements of zeta potential (mV) indicated that SPION werenegatively charged in themedia with values ofminus2568 (plusmn138)minus2914 (plusmn385) and minus2806 (plusmn119) respectively for SPION-1SPION-2 and SPION-3
32 Solubility and Bioaccumulation of SPION The solublefraction of free Fe Co and Zn released from SPION-1SPION-2 and SPION-3 suspensions in culture medium wasdetermined at 24 48 and 72 hours (Figures 2 3 and 4)The quantity of soluble Fe Co and Zn in the medium wasdependent on the time of exposure the SPION compositionand their concentration When C vulgaris was exposed to125 120583gmLminus1 of SPION-1 (Fe
3O4) during 72 h the proportion
of the soluble fraction of free Fe was of 27 compared tothe nominal concentration of SPION-1 and it decreased to3 for 400 120583gmLminus1 of SPION-1 (Figure 2) However thesolubility of SPION-2 (Co
02Zn08Fe2O4) was dependent on
the metal species When algal cells were exposed during72 h to 125 120583gmLminus1 of SPION-2 proportions of free FeCo and Zn in the soluble fraction were respectively of27 12 and 9 compared to the nominal concentrationof SPION-2 (Figure 3) For the exposure concentration of400 120583gmLminus1 the quantity of soluble Fe Co andZn decreasedto 3 2 and 1 respectively Concerning the solubility ofSPION-3 (Co
05Zn05Fe2O4) concentrations of Fe Co and
Zn in the soluble fraction at 72 h were of 9 each whencompared to the nominal concentration of 125120583gmLminus1 Forthe exposure concentration of 400 120583gmLminus1 concentrationsof Fe Co and Zn in the soluble fraction were less than
6 BioMed Research International
Fe
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed F
e120583
gm
g of
DW
(a)
Co
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed C
o120583
gm
g of
DW
(b)
Zn
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed Z
n120583
gm
g of
DW
(c)
Figure 6 Bioaccumulated content of Fe Co and Zn in algal cells of Chlorella vulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square)and 72 h (998771 triangle) to different concentrations of SPION-2 (Co
02Zn08Fe2O4)
05 in comparison to the nominal concentration of SPION-3 Indeed we observed that the agglomeration of SPIONincreased their precipitationsedimentation at the bottomof the experimental flask which was directly related to theincreasing concentration tested and time of exposure Thiseffect may cause the reduction of the surface contact ofnanoparticles with the medium explaining the decrease inSPION solubilisation related to their concentration and timeof exposure
The bioaccumulation of total Fe Co and Zn was alsoquantified in algal biomass of C vulgaris exposed during 2448 and 72 h to SPION (Figures 5 6 and 7) For SPION-1the accumulated contents of Fe in algal biomass increasedin dependence on the concentration of SPION and the timeof exposure (Figure 5) The bioaccumulated content of Feincreased by 35-fold for algal cells treated during 72 h to400 120583gmLminus1 of SPION-1 compared to control Under thistreatment condition Fe bioaccumulation increased by 281-fold from 24 to 72 h Concerning SPION-2 the accumulatedcontents of Fe Co and Zn increased in algal biomassin relation to the concentration of SPION (Figure 6) Thecontent of Fe reached its maximum value at 72 h whichincreased by 104-fold for 400120583gmLminus1 of SPION compared
to control On the other hand the contents of Co andZn attained their maximal values at 48 h which increasedrespectively by 537-and 628-fold for 400 120583gmLminus1 of SPIONcompared to control When C vulgaris was exposed toSPION-3 the bioaccumulation of Fe Co and Zn increasedin relation to the concentration of SPION (Figure 7) At 72 hof exposure values of Fe Co and Zn contents increasedrespectively by 442- 329- and 547-fold for 400 120583gmLminus1 ofSPION compared to control
33 Inhibition of the Relative Cell Division Rate When algalcells ofC vulgariswere exposed during 72 h to SPION at con-centrations varying from 125 to 400 120583gmLminus1 the inhibitionof the relative cell division rate based on the change of celldensity was dependent on the tested SPION concentration(Table 1) Under concentration exposure of 400120583gmLminus1 therelative cell division rate decreased significantly compared tothe control by 478 218 and 158 for SPION-1 SPION-2and SPION-3 respectively
34 Inhibition of Photosynthetic Electron Transport Thechange of Chl 119886 fluorescence emission was used to monitorthe photosynthetic electron transport when algal cells of
BioMed Research International 7
Fe
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed F
e120583
gm
g of
DW
(a)
Co
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed C
o120583
gm
g of
DW
(b)
Zn
0
1
2
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed Z
n120583
gm
g of
DW
(c)
Figure 7 Bioaccumulated content of Fe Co and Zn in algal cells of Chlorella vulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square)and 72 h (998771 triangle) to different concentrations of SPION-3 (Co
05Zn05Fe2O4)
Table 1 Change in the relative cell division rate (RCDR) when Cvulgaris was exposed to SPION during 72 h
[SPION]120583gmL
RCDR (10minus6)SPION-1 SPION-2 SPION-3
0 0335 plusmn 001 0335 plusmn 001 0335 plusmn 001
125 0305 plusmn 005 0307 plusmn 005 0317 plusmn 0004
25 0308 plusmn 0008 0298 plusmn 0006 0318 plusmn 0007
50 0307 plusmn 002 0297 plusmn 0018 0309 plusmn 0004
100 0266 plusmn 005 0291 plusmn 0007 0306 plusmn 0025
200 0220 plusmn 0008 0264 plusmn 0002 0286 plusmn 0005
400 0175 plusmn 0123 0262 plusmn 00033 0282 plusmn 0004
C vulgaris were exposed during 72 h to SPION toxicityThe kinetics of Chl 119886 fluorescence indicated a decrease offluorescence yields at 119869 119868 and119872 levels without any changein their time of appearance which was related to SPIONconcentration (See Supplementary Figures available onlineat httpdxdoiorg1011552013647974) Furthermore thechange of photosynthetic-fluorescence parameters estimated
from the fluorescence kinetic was used as biomarkersof SPION effect on PSII photochemistry (Figure 8) Thestrongest effect on photosynthetic electron transport wasobserved under 400 120583gmLminus1 of SPION treatment Underthis condition the value of ABSRC parameter increasedsignificantly compared to the control by 43 29 and 81 forSPION-1 SPION-2 and SPION-3 respectively This changeindicated a decrease of functional PSII reaction centers able toperform photochemical reactions Also119881
119869values indicating
the level of QAminusQA(total) increased significantly compared
to the control by 32 23 and 12 respectively for SPION-1SPION-2 and SPION-3 due to the inhibition of PSII electrontransport flow toward to plastoquinone pool However themaximal quantum yield of PSII (119865
119881119865119872) decreased by 14 12
and 35 respectively for SPION-1 SPION-2 and SPION-3 Moreover the PSII performance index PI was used as aglobal parameter integrating all PSII photochemical reactionsfrom light harvesting energy transfer to electron transportWhen algal cells of C vulgaris were exposed during 72 hto 400 120583gmLminus1 of SPION PI values decreased significantlycompared to the control by 78 65 and 85 for SPION-1SPION-2 and SPION-3 respectively
8 BioMed Research International
ABSRC
0 100 200 300 4000
50
100
150
200Pa
ram
eter
s val
ue (
)
[SPION] (120583g mLminus1)
(a)
0 100 200 300 4000
50
100
150
200
Para
met
ers v
alue
()
VJ
[SPION] (120583g mLminus1)
(b)
0
20
40
60
80
100
120
SPION-1SPION-2SPION-3
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)
FVFM
(c)
PI
0
20
40
60
80
100
120
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)SPION-1SPION-2SPION-3
(d)
Figure 8 Change of different photosynthetic-based fluorescence parameters for algal cells of Chlorella vulgaris exposed during 72 h todifferent concentrations of SPION-1 (Fe
3O4) SPION-2 (Co
02Zn08Fe2O4) and SPION-3 (Co
05Zn05Fe2O4) Maximal PSII quantum yield
119865119881119865119872 ratio between the number of active PSII reaction centers and light harvesting Chl antenna size ABSRC relative variable fluorescence
yield at 119869 transient 119881119869 performance index of PSII photochemical activity PI
35 Production of Reactive Oxygen Species Related to Cell Via-bility The formation of ROS per viable cells was determinedfor C vulgaris exposed during 72 h to SPION (Figure 9)Under these conditions cell viability decreased significantlycompared to control from 50 120583gmLminus1 of SPION indicatingan induction of cellular oxidative stress The productionof ROS per viable cells increased significantly comparedto control which was dependent on SPION species andconcentration
4 Discussion
41 Toxicity of SPION-1 SPION-2 and SPION-3 In thisstudy toxic effects of SPION-1 -2 and -3 were investigatedon algal cells of C vulgaris which were caused by thedeterioration of photochemical activities of photosynthesisthe induction of oxidative stress and the inhibition of cell
division rate This complex cellular alteration was dependenton SPION chemical composition and its concentration insolution Based on our results it is evident that the bioac-cumulation of free Fe Co and Zn from the soluble fractionwas contributing to the toxicity impact in algal cells It waspreviously proposed that the release of free metal ions frommetallic nanoparticle suspensions represented amajor sourceof toxicity for the growth rate of aquatic microorganisms[18 19] However it is difficult to determine if the release offree metal ions from nanoparticles is the only contributionto the toxicity impact in the algal cellular system Indeedsolubilisation of nanoparticles can take place either in themedia or inside the cell which possess an acidic pH environ-ment favorable for particle solubility [20] Therefore anotherhypothesis is that SPION may also contribute directly to thetoxicity impact Recently it was suggested for direct toxicitymechanisms of nanoparticles to induce a direct alteration of
BioMed Research International 9
0
50
100Cell viability
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(a)
ROS
0
50
100
150
200
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(b)
0
100
200
300
400
500
600
700
0 100 200 300 400
ROScell viability
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(c)
Figure 9 Change in the production of reactive oxygen species (ROS) cellular viability and the ratio ROSviable cells for Chlorella vulgarisexposed during 72 h to different concentrations of SPION-1 (⧫ diamond) SPION-2 (◼ square) and SPION-3 (998771 triangle)
cellular exchanges with the media due to particles bindingon cell membrane [21] It was also shown for bioaccumulatedFe3O4nanoparticles to cause direct toxic effects inside plant
cells [8]Moreover the increase of production of reactive oxygen
species per cell viabilitywas related to the increasing exposureconcentration of SPION indicating that the induction ofcellular oxidative stress was caused by the bioaccumulationof Fe Co and Zn Indeed it was shown in previous stud-ies that metallic nanoparticles can cause the formation ofROS via the Fenton reaction or the disruption of majorphysiological processes [22 23] Also the charged metallicsurface of nanoparticles can trigger the formation of ROS bycatalytic reduction of oxygen into superoxide anion leadingto oxidative damage into proteins lipids nucleic acids andpigments [24]
When algal cells of C vulgaris were exposed during 72 hto different concentrations of SPION suspensions the relativecellular division rate was strongly inhibited for SPION-1compared to SPION-2 and SPION-3 Under these treatmentconditions the production of ROS per viable cells was thehighest for SPION-1 Indeed Fe is well known to generate
in cell free radical oxidations causing lipid peroxidation[25] Furthermore the change of fluorescence parameter PIindicated a stronger inhibition of photosynthesis for SPION-1Therefore according to the change of these biomarkers thetoxicity impact in algal cells of C vulgaris was the strongestfor SPION-1 suspensions
42 Significance of Biochemical Biomarkers In this study twodifferent biochemical biomarkers were used to characterizethe toxicity of SPION the production of ROS per viablecells and the photosynthetic-based fluorescence parametersThe change of parameter ROScell viability permitted deter-mining the potential source of cellular toxicity causing theinhibition of cell division rate On the other hand fluores-cence parameters related to PSII photochemical reactionshave been shown to be sensitive biomarkers of SPION toxicityto C vulgaris cells Indeed the Chl 119886 fluorescence emissionis demonstrated to be related to water-splitting system func-tions at PSII reaction center and to oxidoreduction statesof electron transport carriers [26 27] Therefore our resultsshowed evidence for SPION to cause inhibitory effects on thePSII water-splitting system and the photoactivation of PSII
10 BioMed Research International
reaction centers In comparison to other fluorescence indica-tors the performance index of PSII activity representing anintegrative indicator of all PSII photochemical reactions [17]was the most sensitive biomarker of the deterioration of PSIIfunctions caused by SPION toxicity
5 Conclusion
Nowadays a large quantity of metallic nanoparticles isproduced and their toxic potential as hazardous contami-nants in aquatic environment needs to be investigated inorder to develop specific bioassays for nanoparticles toxicityassessment In this regard microalgae represent sensitiveorganisms to be used in bioassay toxicity testing for theassessment of hazardous materials In this toxicologicalinvestigation we clearly showed the potential source of tox-icity of three SPION suspensions having different chemicalcompositions The algal model C vulgaris demonstrated tobe a valuable bioindicator of SPION cellular toxicity whichwas indicated by the deterioration of photochemical activitiesof photosynthesis the induction of oxidative stress and theinhibition of cell division rateTherefore this work permittedcharacterising the cellular toxicity impact of these SPIONwith different biomarkers A good understanding of thesetoxicological interactions will permit better understandingthe risk of SPION toxicity for aquatic organisms
Abbreviations
Chl ChlorophyllNP NanoparticlesPEA Plant Efficiency AnalyzerPSII Photosystem IIROS Reactive oxygen speciesSPION Superparamagnetic iron oxide nanoparticlesTEM Transmission electron microscope
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was performed at the Department of Chem-istry (UQAM) The authors acknowledge the financial sup-port provided by both the Faculty of Sciences and theDepartment of Chemistry (UQAM) The authors wish tothank L Ben Taherc L Samia Smiric and H Abdelmelekbfor providing nanopowder of Fe
3O4 Co02Zn08Fe2O4 and
Co05Zn05Fe2O4
References
[1] A Ito M Shinkai H Honda and T Kobayashi ldquoMedicalapplication of functionalized magnetic nanoparticlesrdquo Journalof Bioscience and Bioengineering vol 100 no 1 pp 1ndash11 2005
[2] D L Huber ldquoSynthesis properties and applications of ironnanoparticlesrdquo Small vol 1 no 5 pp 482ndash501 2005
[3] N Singh G J S Jenkins R Asadi and S H Doak ldquoPoten-tial toxicity of superparamagnetic iron oxide nanoparticles(SPION)rdquo Nano Reviews vol 1 Article ID 5358 2010
[4] I Bhatt and B N Tripathi ldquoInteraction of engineered nanopar-ticles with various components of the environment and possiblestrategies for their risk assessmentrdquo Chemosphere vol 82 no 3pp 308ndash317 2011
[5] MDelay and FH Frimmel ldquoNanoparticles in aquatic systemsrdquoAnalytical and Bioanalytical Chemistry vol 402 no 2 pp 583ndash592 2012
[6] E N Taylor K M Kummer N G Durmus K Leuba K MTarquinio and T J Webster ldquoSuperparamagnetic iron oxidenanoparticles (SPION) for the treatment of antibiotic-resistantbiofilmsrdquo Small vol 8 pp 3016ndash3027 2012
[7] H Zhu J Han J Q Xiao and Y Jin ldquoUptake translocationand accumulation of manufactured iron oxide nanoparticles bypumpkin plantsrdquo Journal of Environmental Monitoring vol 10no 6 pp 713ndash717 2008
[8] Y KMushtaq ldquoEffect of nanoscale Fe3O4 TiO2and carbon par-
ticles on cucumber seed germinationrdquo Journal of EnvironmentalScience and Health A vol 46 no 14 pp 1732ndash1735 2011
[9] X Chen X Zhu R Li H Yao Z Lu and X Yang ldquoPhoto-synthetic toxicity and oxidative damage induced by nano-Fe
3O4
on Chlorella vulgaris in aquatic environmentrdquo Open Journal ofEcology vol 2 pp 21ndash28 2012
[10] R Rippka J Deruelles and J B Waterbury ldquoGeneric assign-ments strain histories and properties of pure cultures ofcyanobacteriardquo Journal of General Microbiology vol 111 no 1pp 1ndash61 1979
[11] H Basti L Ben Tahar L S Smiri et al ldquoCatechol derivatives-coated Fe
3O4and 120574-Fe
2O3nanoparticles as potential MRI
contrast agentsrdquo Journal of Colloid and Interface Science vol 341no 2 pp 248ndash254 2010
[12] W A Hoffmann and H Poorter ldquoAvoiding bias in calculationsof relative growth raterdquo Annals of Botany vol 90 no 1 pp 37ndash42 2002
[13] I B Gerber and I A Dubery ldquoFluorescence microplate assayfor the detection of oxidative burst products in tobacco cellsuspensions using 2101584071015840-dichlorofluoresceinrdquo Methods in CellScience vol 25 no 3-4 pp 115ndash122 2004
[14] P Mayer R Cuhel and N Nyholm ldquoA simple in vitro fluo-rescence method for biomass measurements in algal growthinhibition testsrdquo Water Research vol 31 no 10 pp 2525ndash25311997
[15] A R Wellburn ldquoThe spectral determination of chlorophylls aand b as well as total carotenoids using various solvents withspectrophotometers of different resolutionrdquo Journal of PlantPhysiology vol 144 no 3 pp 307ndash313 1994
[16] L Force C Critchley and J J S Van Rensen ldquoNew fluorescenceparameters for monitoring photosynthesis in plants 1 the effectof illumination on the fluorescence parameters of the JIP-testrdquoPhotosynthesis Research vol 78 no 1 pp 17ndash33 2003
[17] R J Strasser A Srivastava and M Tsimilli-Michael ldquoAnalysisof the chlorophyll a fluorescence transientrdquo in Chlorophyllfluorescence A signature of Photosynthesis Advances in Photo-synthesis and respiration G G Papageorgiou Ed pp 321ndash362Kluwer Academic Dodrecht The Netherlands 2004
[18] N M Franklin N J Rogers S C Apte G E Batley G EGadd and P S Casey ldquoComparative toxicity of nanopartic-ulate ZnO bulk ZnO and ZnCl
2to a freshwater microalga
(Pseudokirchneriella subcapitata) the importance of particle
BioMed Research International 11
solubilityrdquoEnvironmental Science andTechnology vol 41 no 24pp 8484ndash8490 2007
[19] V Aruoja H-C Dubourguier K Kasemets and A KahruldquoToxicity of nanoparticles of CuO ZnO and TiO
2tomicroalgae
Pseudokirchneriella subcapitatardquo Science of the Total Environ-ment vol 407 no 4 pp 1461ndash1468 2009
[20] A M Studer L K Limbach L Van Duc et al ldquoNanoparticlecytotoxicity depends on intracellular solubility comparison ofstabilized copper metal and degradable copper oxide nanopar-ticlesrdquo Toxicology Letters vol 197 no 3 pp 169ndash174 2010
[21] S Lin P Bhattacharya N C Rajapakse D E Brune and P CKe ldquoEffects of quantum dots adsorption on algal photosynthe-sisrdquo Journal of Physical Chemistry C vol 113 no 25 pp 10962ndash10966 2009
[22] T Xia M Kovochich J Brant et al ldquoComparison of theabilities of ambient and manufactured nanoparticles to inducecellular toxicity according to an oxidative stress paradigmrdquoNano Letters vol 6 no 8 pp 1794ndash1807 2006
[23] X Hu S Cook PWang andH-M Hwang ldquoIn vitro evaluationof cytotoxicity of engineeredmetal oxide nanoparticlesrdquo Scienceof the Total Environment vol 407 no 8 pp 3070ndash3072 2009
[24] W J Stark ldquoNanoparticles in biological systemsrdquo AngewandteChemiemdashInternational Edition vol 50 no 6 pp 1242ndash12582011
[25] F Q Schafer S Y Qian and G R Buettner ldquoIron and freeradical oxidations in cell membranesrdquo Cellular and MolecularBiology vol 46 no 3 pp 657ndash662 2000
[26] D Dewez N A Ali F Perreault and R Popovic ldquoRapidchlorophyll a fluorescence transient of Lemna gibba leaf as anindication of light and hydroxylamine effect on photosystem IIactivityrdquo Photochemical and Photobiological Sciences vol 6 no5 pp 532ndash538 2007
[27] D Dewez N Boucher F Bellemare and R Popovic ldquoUse ofdifferent fluorometric systems in the determination of fluo-rescence parameters from spinach thylakoid membranes beingexposed to atrazine and copperrdquo Toxicological and Environmen-tal Chemistry vol 89 no 4 pp 655ndash664 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 BioMed Research International
In the present study the green alga Chlorella vulgariswas used as a unicellular plant model organism for the tox-icity characterisation of Fe
3O4(SPION-1) Co
02Zn08Fe2O4
(SPION-2) and Co05Zn05Fe2O4(SPION-3) Algal cells were
exposed during 24 48 and 72 hours in order to evaluate theevolution of SPION toxicity impact on the entire cellular sys-tem by evaluating the change in photochemical reactions ofphotosynthesis cell division and the induction of oxidativestress This work permitted determining the risk of SPIONtoxicity on the viability of green algae and therefore thepotential use of this algal species in a bioassay of SPIONtoxicity
2 Material and Methods
21 Biological Material The freshwater green alga Chlorellavulgariswas obtained from theCanadianPhycological CultureCentre (CPCC University of Waterloo ON Canada) Algalculture was prepared in sterile BG-11 liquid growth medium(pH = 7 ionic strength = 00201) having the following finalconcentrations of salts composition [10] 15 g Lminus1 of NaNO
3
004 g Lminus1 of K2HPO4sdot3H2O 0075 g Lminus1 of MgSO
4sdot7H2O
0036 g Lminus1 of CaCl2sdot2H2O 6 times 10minus3 g Lminus1 of C
6H8O7(citric
acid) 6 times 10minus3 g Lminus1 of C6H5FeO7(ferric citrate) 10minus3 g Lminus1
of Na2EDTAsdot2H
2O 002 g Lminus1 of Na
2CO3 286mg Lminus1
of H3BO3 181mg Lminus1 of MnCl
2sdot4H2O 0222mg Lminus1 of
ZnSO4sdot7H2O 0390mg Lminus1 ofNa
2MoO4sdot2H2O 0079mg Lminus1
of CuSO4sdot5H2O and 00494mg Lminus1 of Co(NO
3)2sdot6H2O
Algal cells were grown under continuous light intensity of100 120583molmminus2 sminus1 (SYLVANIA GRO-LUX Wide Spectrumlight F40GRQAQWS) at 24∘C plusmn 1 Aliquots of algalsamples were used for experiments when algal culturereached the exponential growth phase The change of celldensity was determined by monitoring the optical densityat 750 nm and the calculation was based on a standardcorrelation with the cell density measured using a multisizerZ3 (Beckman Coulter Inc USA)
22 Synthesis of SPION Superparamagnetic iron oxidenanoparticles (SPION) as Fe
3O4(SPION-1) Co
02Zn08Fe2O4
(SPION-2) and Co05Zn05Fe2O4(SPION-3) were produced
according to the procedure reported in [11] by usingthe polyol process starting from Co(CH
3CO2)2∙4H2O Zn
(CH3CO2)2∙2H2O and Fe(CH
3CO2)2as precursor salts and
diethylene glycol as a solvent
23 Stock Solution and SPION Characterization In thepreparation of stock solutions SPION were suspended inculture medium at a concentration of 1 g Lminus1 and homoge-nized by ultrasonication during 30min at 4∘C to break upagglomerates After sonication stock solutions weremix witha vortex for 1min and various concentrations of SPIONwereprepared in culture medium for experiments Size distribu-tionwas determined by dynamic light scattering (DLS) with aZetaPlus particle sizer (Brookhaven InstrumentCorporationUSA) using 90Plus Particles Sizing Software (Ver 420)Zeta potential of SPION suspensions in culture medium was
0
20
40
60
80
100
120
0 200 400 600
SPION-1SPION-2 SPION-3
Part
icle
s fra
ctio
ns
Diameter (nm)
Figure 1 Particle size distribution of SPION-1 (Fe3O4) SPION-2
(Co02Zn08Fe2O4) and SPION-3 (Co
05Zn05Fe2O4) suspensions in
the culture medium
0
2
4
6
8
10
12
14
16
0 100 200 300 400
24h48h72h
Solu
ble [
Fe] (120583
g mLminus
1)
[SPION-1] (120583g mLminus1)
Figure 2 Soluble fraction of free Fe released from SPION-1 (Fe3O4)
suspension in culture medium at 24 48 and 72 h of exposure
evaluated by the electrophoretic mobility method with theZetaPlus system
24 Experimental Treatments In each treatment conditionthe initial density of algal cells was of 106 cells mLminus1 in a finalvolume of 50mL Algal samples were exposed during 24 48and 72 h to 125 25 50 100 200 and 400mg Lminus1 of SPION-1 SPION-2 or SPION-3 under the same illumination andtemperature condition used for growing stock cultures Forthe control sample the samemedia compositionwas used butwithout any trace of SPION
BioMed Research International 3
0 100 200 300 4000
5
10
15Fe
Solu
ble [
Fe] (120583
g mLminus
1)
[SPION-2] (120583g mLminus1)
(a)
0
2
4
6
8
10Co
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Solu
ble
[Co]
(120583g m
Lminus1)
(b)
0
2
4
6
8
10
0 100 200 300 400
Zn
[SPION-2] (120583g mLminus1)24h48h72h
Solu
ble [
Zn] (120583
g mLminus
1)
(c)
Figure 3 Soluble fraction of free Fe Co and Zn released from SPION-2 (Co02Zn08Fe2O4) suspension in culture medium at 24 48 and 72 h
of exposure
25 Growth Inhibition Test The relative cell division rate(RCDR) was evaluated for 72 h according to [12] as RCDR =(lnW
72 hminus lnW0)72 h whereW72 h represents cell density at72 h and W
0the initial cell density
26 Bioaccumulation of Fe Co and Zn To separate SPIONfrom cells a sucrose gradient prepared in BG-11 media wasdone directly in a Beckman centrifuge tube inclined at a 30∘angle in order to get 6 layers of different sucrose densities(5mL of each sucrose solution 120 100 80 60 40 and20) Algal cells of control and SPION-treated samples werecollected by centrifugation Their pellets were slowly placedon top of sucrose gradient tubes which were centrifuged at1000 rpm during 30min in a swinging-bucket 5810R cen-trifuge (Eppendorf Germany) It resulted in the formationof a pellet of SPION at the bottom of the tube The algal cellslayer was recuperated with a glass Pasteur pipette and filtered
on a 045 120583m filter previously dried and weighted To removeSPIONweakly bound to the cell surface or the filter 3times 10mLof 10mM ethylenediaminetetraacetic acid in BG-11 mediumwas slowly passed through the filter Filters were dried at 95∘Cfor 24 h and weighted to calculate algal dry weight Filterswere then placed in acid-washed glass tubes in which 4mL ofHNO
3and 500 120583L H
2O2were added Samples were digested
during 48 h at room temperature before being diluted to 20HNO
3in Milli-Q purified water for the quantification of Fe
Co and Zn using atomic absorption spectrometry analysis(Varian SpectrAA 220 FS USA) Obtained Fe Co and Znconcentrations were normalized to the dry weight
27 Soluble Fraction of Fe Co and Zn Solubility of free FeCo and Zn released from SPION suspensions were deter-mined in culturemedium SPION suspensions was incubatedduring 24 48 and 72 h in the same condition as described
4 BioMed Research International
0
1
2
3
4Fe
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Solu
ble [
Fe] (120583
g mLminus
1)
(a)
0
1
2
3
4Co
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Solu
ble
[Co]
(120583g m
Lminus1)
(b)
0
1
2
3
4
0 100 200 300 400
Zn
[SPION-3] (120583g mLminus1)24h48h72h
Solu
ble [
Zn] (120583
g mLminus
1)
(c)
Figure 4 Soluble fraction of Fe Co and Zn released from SPION-3 (Co05Zn05Fe2O4) suspension in culture medium at 24 48 and 72 h of
exposure
above for treatment After incubation NPs were removed bycentrifugation at 12000 g for 30min and the supernatant wascollected for analysis Quantification of free Fe Co and Znin solution wasmeasured by atomic absorption spectrometry(Varian SpectrAA 220 FS USA)
28 Production of ROS The fluorescent dye 2101584071015840-dichlorodi-hydrofluorescein diacetate ((H
2DCFDA) Invitrogen Molec-
ular Probe USA) was used as an indicator of ROS accordingto [13] Cellular esterases hydrolyze this probe into the non-fluorescent compound 2101584071015840-dichlorodihydrofluorescein(H2DCF) which is better retained within cells In the
presence of ROS and cellular esterases H2DCF is trans-
formed into the highly fluorescent compound 2101584071015840-dichlo-rofluorescein (DCF) H
2DCFDA stock solution (10mM) was
prepared in ethanol in the dark After 72 h of treatment 1mLof algal samples was exposed during 15min to 02mM of
H2DCFDA in the dark The ROS level was determined by
measuring the fluorescence emission at 530 nm with a flowcytometer (FACScan system Becton Dickinson InstrumentsUSA) Cytometry results were analysed using the WinMDI28 software Algal cells were separated from noncellularparticles by using a relationship between particle size and redfluorescence level originating from chlorophyll fluorescenceemission A positive control sample using methyl viologenwas done to verify the assay (data not shown)
29 Cellular Viability Viability of algal cells was estimatedusing the fluorescein diacetate ((FDA) Invitrogen MolecularProbe USA) method according to [14] FDA is a nonpolarester compoundwhich passes through cell membranes Onceinside the cell FDA is hydrolyzed by esterases (enzymespresent in viable cells) to produce fluorescein accumulatingin cell wall and emitting fluorescence under UV illumination
BioMed Research International 5
After 72 h of treatment 1mL of algal samples was exposedduring 15min to 02mM of FDA in the dark Cell viabilitywas determined by measuring the fluorescence emissionat 530 nm with a flow cytometer (FACScan System Bec-ton Dickinson Instruments USA) Cytometry results wereanalysed using the WinMDI 28 software Algal cells wereseparated from noncellular particles by using a relationshipbetween particle size and red fluorescence level originatingfrom chlorophyll fluorescence emission A positive controlsample using methyl viologen was done to verify the assay(data not shown)
210 Chl 119886 Fluorescence Emission Photosynthetic electrontransport was monitored from the change in the rapid riseof Chl 119886 fluorescence emission using a ldquoPlant EfficiencyAnalyserrdquo fluorometer (Handy-PEA Hansatech Ltd UK)Total chlorophyll (Chl) content (119886 + 119887) was extracted in 100methanol at 65∘C and quantified with a spectrophotometer(Lambda 40 Perkin-Elmer USA) according to the formulaindicated in [15] Total Chl (120583gmLminus1) = (2493 times A
6524+
144 times A6652
)Prior to fluorescence measurements algal samples were
transferred into clean sterile 2mL glass vials and dark-adapted for 30min An aliquot of 5 120583g of total chlorophyllwas gently filtered using low pressure filtration and algal cellswere uniformly placed on a 13mm glass fibre filter (MilliporeUSA) The fluorescence induction was triggered using a 1 ssaturating flash of 3500120583molmminus2 sminus1The fluorescence inten-sity at 20120583s was considered as the 119874 value (119865
119874) fluorescence
intensities for 119870 119869 and 119868 transients were determined at300 120583s (119865
119870) 2ms (119865
119869) and 30ms (119865
119868) respectively the max-
imum fluorescence yield reached maximal value of fluores-cence intensity (119865
119872) under saturating illumination Different
photosynthetic-based fluorescence parameters related to thefunctional state of Photosystem II were calculated [16 17]the maximum efficiency of PSII electron transport 119865
119881119865119872=
(119865119872minus 119865119874)119865119872 the absorption of photons by light harvesting
antenna complexes (ABS) per active reaction center (RC)ABSRC = ((119865
119870minus 119865119874)025) times (1(119865
119869minus 119865119874)) times (119865
119872(119865119872minus
119865119874)) the relative variable fluorescence yield at 119869 transient
estimating the fraction of 119876119860
in its reduced state 119881119869=
(119865119869minus 119865119874)(119865119872minus 119865119874) and the performance index of PSII
photosynthetic activity PI = RCABS times ((119865119872minus 119865119874)119865119874) times
((1 minus 119881119869)119881119869)
211 Statistical Analysis All treatments were performed intriplicate Means and standard deviations were calculatedfor each treatment Significant differences between controland treated plants were determined by one-way analysisof variance (ANOVA) followed by a Dunnettrsquos multiplecomparison (DMC) test where 119875 value less than 005 wasconsidered significant
3 Results
31 Characterisation of SPION When SPION were sus-pended in the media nanoparticles formed agglomeratesduring the first minutes as indicated by the distribution
0
2
4
6
8
10
12
0 100 200 300 400
Bioa
ccum
ulat
ed F
e120583g
mg
of D
W
[SPION-1] (120583g mLminus1)
Figure 5 Bioaccumulated content of Fe in algal cells of Chlorellavulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square) and 72 h(998771 triangle) to different concentrations of SPION-1 (Fe
3O4)
of hydrodynamic particles size diameter which was causedby the content of salts in the media Analysis by dynamiclight scattering showed SPION-1 SPION-2 and SPION-3suspensions in culture media to have an average diameterof particle size distribution of 1959 1765 and 3472 nmrespectively (Figure 1) These distributions of hydrodynamicsize of SPION were found to be stable in the culture mediumduring the entire experimental exposure Furthermore mea-surements of zeta potential (mV) indicated that SPION werenegatively charged in themedia with values ofminus2568 (plusmn138)minus2914 (plusmn385) and minus2806 (plusmn119) respectively for SPION-1SPION-2 and SPION-3
32 Solubility and Bioaccumulation of SPION The solublefraction of free Fe Co and Zn released from SPION-1SPION-2 and SPION-3 suspensions in culture medium wasdetermined at 24 48 and 72 hours (Figures 2 3 and 4)The quantity of soluble Fe Co and Zn in the medium wasdependent on the time of exposure the SPION compositionand their concentration When C vulgaris was exposed to125 120583gmLminus1 of SPION-1 (Fe
3O4) during 72 h the proportion
of the soluble fraction of free Fe was of 27 compared tothe nominal concentration of SPION-1 and it decreased to3 for 400 120583gmLminus1 of SPION-1 (Figure 2) However thesolubility of SPION-2 (Co
02Zn08Fe2O4) was dependent on
the metal species When algal cells were exposed during72 h to 125 120583gmLminus1 of SPION-2 proportions of free FeCo and Zn in the soluble fraction were respectively of27 12 and 9 compared to the nominal concentrationof SPION-2 (Figure 3) For the exposure concentration of400 120583gmLminus1 the quantity of soluble Fe Co andZn decreasedto 3 2 and 1 respectively Concerning the solubility ofSPION-3 (Co
05Zn05Fe2O4) concentrations of Fe Co and
Zn in the soluble fraction at 72 h were of 9 each whencompared to the nominal concentration of 125120583gmLminus1 Forthe exposure concentration of 400 120583gmLminus1 concentrationsof Fe Co and Zn in the soluble fraction were less than
6 BioMed Research International
Fe
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed F
e120583
gm
g of
DW
(a)
Co
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed C
o120583
gm
g of
DW
(b)
Zn
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed Z
n120583
gm
g of
DW
(c)
Figure 6 Bioaccumulated content of Fe Co and Zn in algal cells of Chlorella vulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square)and 72 h (998771 triangle) to different concentrations of SPION-2 (Co
02Zn08Fe2O4)
05 in comparison to the nominal concentration of SPION-3 Indeed we observed that the agglomeration of SPIONincreased their precipitationsedimentation at the bottomof the experimental flask which was directly related to theincreasing concentration tested and time of exposure Thiseffect may cause the reduction of the surface contact ofnanoparticles with the medium explaining the decrease inSPION solubilisation related to their concentration and timeof exposure
The bioaccumulation of total Fe Co and Zn was alsoquantified in algal biomass of C vulgaris exposed during 2448 and 72 h to SPION (Figures 5 6 and 7) For SPION-1the accumulated contents of Fe in algal biomass increasedin dependence on the concentration of SPION and the timeof exposure (Figure 5) The bioaccumulated content of Feincreased by 35-fold for algal cells treated during 72 h to400 120583gmLminus1 of SPION-1 compared to control Under thistreatment condition Fe bioaccumulation increased by 281-fold from 24 to 72 h Concerning SPION-2 the accumulatedcontents of Fe Co and Zn increased in algal biomassin relation to the concentration of SPION (Figure 6) Thecontent of Fe reached its maximum value at 72 h whichincreased by 104-fold for 400120583gmLminus1 of SPION compared
to control On the other hand the contents of Co andZn attained their maximal values at 48 h which increasedrespectively by 537-and 628-fold for 400 120583gmLminus1 of SPIONcompared to control When C vulgaris was exposed toSPION-3 the bioaccumulation of Fe Co and Zn increasedin relation to the concentration of SPION (Figure 7) At 72 hof exposure values of Fe Co and Zn contents increasedrespectively by 442- 329- and 547-fold for 400 120583gmLminus1 ofSPION compared to control
33 Inhibition of the Relative Cell Division Rate When algalcells ofC vulgariswere exposed during 72 h to SPION at con-centrations varying from 125 to 400 120583gmLminus1 the inhibitionof the relative cell division rate based on the change of celldensity was dependent on the tested SPION concentration(Table 1) Under concentration exposure of 400120583gmLminus1 therelative cell division rate decreased significantly compared tothe control by 478 218 and 158 for SPION-1 SPION-2and SPION-3 respectively
34 Inhibition of Photosynthetic Electron Transport Thechange of Chl 119886 fluorescence emission was used to monitorthe photosynthetic electron transport when algal cells of
BioMed Research International 7
Fe
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed F
e120583
gm
g of
DW
(a)
Co
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed C
o120583
gm
g of
DW
(b)
Zn
0
1
2
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed Z
n120583
gm
g of
DW
(c)
Figure 7 Bioaccumulated content of Fe Co and Zn in algal cells of Chlorella vulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square)and 72 h (998771 triangle) to different concentrations of SPION-3 (Co
05Zn05Fe2O4)
Table 1 Change in the relative cell division rate (RCDR) when Cvulgaris was exposed to SPION during 72 h
[SPION]120583gmL
RCDR (10minus6)SPION-1 SPION-2 SPION-3
0 0335 plusmn 001 0335 plusmn 001 0335 plusmn 001
125 0305 plusmn 005 0307 plusmn 005 0317 plusmn 0004
25 0308 plusmn 0008 0298 plusmn 0006 0318 plusmn 0007
50 0307 plusmn 002 0297 plusmn 0018 0309 plusmn 0004
100 0266 plusmn 005 0291 plusmn 0007 0306 plusmn 0025
200 0220 plusmn 0008 0264 plusmn 0002 0286 plusmn 0005
400 0175 plusmn 0123 0262 plusmn 00033 0282 plusmn 0004
C vulgaris were exposed during 72 h to SPION toxicityThe kinetics of Chl 119886 fluorescence indicated a decrease offluorescence yields at 119869 119868 and119872 levels without any changein their time of appearance which was related to SPIONconcentration (See Supplementary Figures available onlineat httpdxdoiorg1011552013647974) Furthermore thechange of photosynthetic-fluorescence parameters estimated
from the fluorescence kinetic was used as biomarkersof SPION effect on PSII photochemistry (Figure 8) Thestrongest effect on photosynthetic electron transport wasobserved under 400 120583gmLminus1 of SPION treatment Underthis condition the value of ABSRC parameter increasedsignificantly compared to the control by 43 29 and 81 forSPION-1 SPION-2 and SPION-3 respectively This changeindicated a decrease of functional PSII reaction centers able toperform photochemical reactions Also119881
119869values indicating
the level of QAminusQA(total) increased significantly compared
to the control by 32 23 and 12 respectively for SPION-1SPION-2 and SPION-3 due to the inhibition of PSII electrontransport flow toward to plastoquinone pool However themaximal quantum yield of PSII (119865
119881119865119872) decreased by 14 12
and 35 respectively for SPION-1 SPION-2 and SPION-3 Moreover the PSII performance index PI was used as aglobal parameter integrating all PSII photochemical reactionsfrom light harvesting energy transfer to electron transportWhen algal cells of C vulgaris were exposed during 72 hto 400 120583gmLminus1 of SPION PI values decreased significantlycompared to the control by 78 65 and 85 for SPION-1SPION-2 and SPION-3 respectively
8 BioMed Research International
ABSRC
0 100 200 300 4000
50
100
150
200Pa
ram
eter
s val
ue (
)
[SPION] (120583g mLminus1)
(a)
0 100 200 300 4000
50
100
150
200
Para
met
ers v
alue
()
VJ
[SPION] (120583g mLminus1)
(b)
0
20
40
60
80
100
120
SPION-1SPION-2SPION-3
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)
FVFM
(c)
PI
0
20
40
60
80
100
120
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)SPION-1SPION-2SPION-3
(d)
Figure 8 Change of different photosynthetic-based fluorescence parameters for algal cells of Chlorella vulgaris exposed during 72 h todifferent concentrations of SPION-1 (Fe
3O4) SPION-2 (Co
02Zn08Fe2O4) and SPION-3 (Co
05Zn05Fe2O4) Maximal PSII quantum yield
119865119881119865119872 ratio between the number of active PSII reaction centers and light harvesting Chl antenna size ABSRC relative variable fluorescence
yield at 119869 transient 119881119869 performance index of PSII photochemical activity PI
35 Production of Reactive Oxygen Species Related to Cell Via-bility The formation of ROS per viable cells was determinedfor C vulgaris exposed during 72 h to SPION (Figure 9)Under these conditions cell viability decreased significantlycompared to control from 50 120583gmLminus1 of SPION indicatingan induction of cellular oxidative stress The productionof ROS per viable cells increased significantly comparedto control which was dependent on SPION species andconcentration
4 Discussion
41 Toxicity of SPION-1 SPION-2 and SPION-3 In thisstudy toxic effects of SPION-1 -2 and -3 were investigatedon algal cells of C vulgaris which were caused by thedeterioration of photochemical activities of photosynthesisthe induction of oxidative stress and the inhibition of cell
division rate This complex cellular alteration was dependenton SPION chemical composition and its concentration insolution Based on our results it is evident that the bioac-cumulation of free Fe Co and Zn from the soluble fractionwas contributing to the toxicity impact in algal cells It waspreviously proposed that the release of free metal ions frommetallic nanoparticle suspensions represented amajor sourceof toxicity for the growth rate of aquatic microorganisms[18 19] However it is difficult to determine if the release offree metal ions from nanoparticles is the only contributionto the toxicity impact in the algal cellular system Indeedsolubilisation of nanoparticles can take place either in themedia or inside the cell which possess an acidic pH environ-ment favorable for particle solubility [20] Therefore anotherhypothesis is that SPION may also contribute directly to thetoxicity impact Recently it was suggested for direct toxicitymechanisms of nanoparticles to induce a direct alteration of
BioMed Research International 9
0
50
100Cell viability
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(a)
ROS
0
50
100
150
200
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(b)
0
100
200
300
400
500
600
700
0 100 200 300 400
ROScell viability
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(c)
Figure 9 Change in the production of reactive oxygen species (ROS) cellular viability and the ratio ROSviable cells for Chlorella vulgarisexposed during 72 h to different concentrations of SPION-1 (⧫ diamond) SPION-2 (◼ square) and SPION-3 (998771 triangle)
cellular exchanges with the media due to particles bindingon cell membrane [21] It was also shown for bioaccumulatedFe3O4nanoparticles to cause direct toxic effects inside plant
cells [8]Moreover the increase of production of reactive oxygen
species per cell viabilitywas related to the increasing exposureconcentration of SPION indicating that the induction ofcellular oxidative stress was caused by the bioaccumulationof Fe Co and Zn Indeed it was shown in previous stud-ies that metallic nanoparticles can cause the formation ofROS via the Fenton reaction or the disruption of majorphysiological processes [22 23] Also the charged metallicsurface of nanoparticles can trigger the formation of ROS bycatalytic reduction of oxygen into superoxide anion leadingto oxidative damage into proteins lipids nucleic acids andpigments [24]
When algal cells of C vulgaris were exposed during 72 hto different concentrations of SPION suspensions the relativecellular division rate was strongly inhibited for SPION-1compared to SPION-2 and SPION-3 Under these treatmentconditions the production of ROS per viable cells was thehighest for SPION-1 Indeed Fe is well known to generate
in cell free radical oxidations causing lipid peroxidation[25] Furthermore the change of fluorescence parameter PIindicated a stronger inhibition of photosynthesis for SPION-1Therefore according to the change of these biomarkers thetoxicity impact in algal cells of C vulgaris was the strongestfor SPION-1 suspensions
42 Significance of Biochemical Biomarkers In this study twodifferent biochemical biomarkers were used to characterizethe toxicity of SPION the production of ROS per viablecells and the photosynthetic-based fluorescence parametersThe change of parameter ROScell viability permitted deter-mining the potential source of cellular toxicity causing theinhibition of cell division rate On the other hand fluores-cence parameters related to PSII photochemical reactionshave been shown to be sensitive biomarkers of SPION toxicityto C vulgaris cells Indeed the Chl 119886 fluorescence emissionis demonstrated to be related to water-splitting system func-tions at PSII reaction center and to oxidoreduction statesof electron transport carriers [26 27] Therefore our resultsshowed evidence for SPION to cause inhibitory effects on thePSII water-splitting system and the photoactivation of PSII
10 BioMed Research International
reaction centers In comparison to other fluorescence indica-tors the performance index of PSII activity representing anintegrative indicator of all PSII photochemical reactions [17]was the most sensitive biomarker of the deterioration of PSIIfunctions caused by SPION toxicity
5 Conclusion
Nowadays a large quantity of metallic nanoparticles isproduced and their toxic potential as hazardous contami-nants in aquatic environment needs to be investigated inorder to develop specific bioassays for nanoparticles toxicityassessment In this regard microalgae represent sensitiveorganisms to be used in bioassay toxicity testing for theassessment of hazardous materials In this toxicologicalinvestigation we clearly showed the potential source of tox-icity of three SPION suspensions having different chemicalcompositions The algal model C vulgaris demonstrated tobe a valuable bioindicator of SPION cellular toxicity whichwas indicated by the deterioration of photochemical activitiesof photosynthesis the induction of oxidative stress and theinhibition of cell division rateTherefore this work permittedcharacterising the cellular toxicity impact of these SPIONwith different biomarkers A good understanding of thesetoxicological interactions will permit better understandingthe risk of SPION toxicity for aquatic organisms
Abbreviations
Chl ChlorophyllNP NanoparticlesPEA Plant Efficiency AnalyzerPSII Photosystem IIROS Reactive oxygen speciesSPION Superparamagnetic iron oxide nanoparticlesTEM Transmission electron microscope
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was performed at the Department of Chem-istry (UQAM) The authors acknowledge the financial sup-port provided by both the Faculty of Sciences and theDepartment of Chemistry (UQAM) The authors wish tothank L Ben Taherc L Samia Smiric and H Abdelmelekbfor providing nanopowder of Fe
3O4 Co02Zn08Fe2O4 and
Co05Zn05Fe2O4
References
[1] A Ito M Shinkai H Honda and T Kobayashi ldquoMedicalapplication of functionalized magnetic nanoparticlesrdquo Journalof Bioscience and Bioengineering vol 100 no 1 pp 1ndash11 2005
[2] D L Huber ldquoSynthesis properties and applications of ironnanoparticlesrdquo Small vol 1 no 5 pp 482ndash501 2005
[3] N Singh G J S Jenkins R Asadi and S H Doak ldquoPoten-tial toxicity of superparamagnetic iron oxide nanoparticles(SPION)rdquo Nano Reviews vol 1 Article ID 5358 2010
[4] I Bhatt and B N Tripathi ldquoInteraction of engineered nanopar-ticles with various components of the environment and possiblestrategies for their risk assessmentrdquo Chemosphere vol 82 no 3pp 308ndash317 2011
[5] MDelay and FH Frimmel ldquoNanoparticles in aquatic systemsrdquoAnalytical and Bioanalytical Chemistry vol 402 no 2 pp 583ndash592 2012
[6] E N Taylor K M Kummer N G Durmus K Leuba K MTarquinio and T J Webster ldquoSuperparamagnetic iron oxidenanoparticles (SPION) for the treatment of antibiotic-resistantbiofilmsrdquo Small vol 8 pp 3016ndash3027 2012
[7] H Zhu J Han J Q Xiao and Y Jin ldquoUptake translocationand accumulation of manufactured iron oxide nanoparticles bypumpkin plantsrdquo Journal of Environmental Monitoring vol 10no 6 pp 713ndash717 2008
[8] Y KMushtaq ldquoEffect of nanoscale Fe3O4 TiO2and carbon par-
ticles on cucumber seed germinationrdquo Journal of EnvironmentalScience and Health A vol 46 no 14 pp 1732ndash1735 2011
[9] X Chen X Zhu R Li H Yao Z Lu and X Yang ldquoPhoto-synthetic toxicity and oxidative damage induced by nano-Fe
3O4
on Chlorella vulgaris in aquatic environmentrdquo Open Journal ofEcology vol 2 pp 21ndash28 2012
[10] R Rippka J Deruelles and J B Waterbury ldquoGeneric assign-ments strain histories and properties of pure cultures ofcyanobacteriardquo Journal of General Microbiology vol 111 no 1pp 1ndash61 1979
[11] H Basti L Ben Tahar L S Smiri et al ldquoCatechol derivatives-coated Fe
3O4and 120574-Fe
2O3nanoparticles as potential MRI
contrast agentsrdquo Journal of Colloid and Interface Science vol 341no 2 pp 248ndash254 2010
[12] W A Hoffmann and H Poorter ldquoAvoiding bias in calculationsof relative growth raterdquo Annals of Botany vol 90 no 1 pp 37ndash42 2002
[13] I B Gerber and I A Dubery ldquoFluorescence microplate assayfor the detection of oxidative burst products in tobacco cellsuspensions using 2101584071015840-dichlorofluoresceinrdquo Methods in CellScience vol 25 no 3-4 pp 115ndash122 2004
[14] P Mayer R Cuhel and N Nyholm ldquoA simple in vitro fluo-rescence method for biomass measurements in algal growthinhibition testsrdquo Water Research vol 31 no 10 pp 2525ndash25311997
[15] A R Wellburn ldquoThe spectral determination of chlorophylls aand b as well as total carotenoids using various solvents withspectrophotometers of different resolutionrdquo Journal of PlantPhysiology vol 144 no 3 pp 307ndash313 1994
[16] L Force C Critchley and J J S Van Rensen ldquoNew fluorescenceparameters for monitoring photosynthesis in plants 1 the effectof illumination on the fluorescence parameters of the JIP-testrdquoPhotosynthesis Research vol 78 no 1 pp 17ndash33 2003
[17] R J Strasser A Srivastava and M Tsimilli-Michael ldquoAnalysisof the chlorophyll a fluorescence transientrdquo in Chlorophyllfluorescence A signature of Photosynthesis Advances in Photo-synthesis and respiration G G Papageorgiou Ed pp 321ndash362Kluwer Academic Dodrecht The Netherlands 2004
[18] N M Franklin N J Rogers S C Apte G E Batley G EGadd and P S Casey ldquoComparative toxicity of nanopartic-ulate ZnO bulk ZnO and ZnCl
2to a freshwater microalga
(Pseudokirchneriella subcapitata) the importance of particle
BioMed Research International 11
solubilityrdquoEnvironmental Science andTechnology vol 41 no 24pp 8484ndash8490 2007
[19] V Aruoja H-C Dubourguier K Kasemets and A KahruldquoToxicity of nanoparticles of CuO ZnO and TiO
2tomicroalgae
Pseudokirchneriella subcapitatardquo Science of the Total Environ-ment vol 407 no 4 pp 1461ndash1468 2009
[20] A M Studer L K Limbach L Van Duc et al ldquoNanoparticlecytotoxicity depends on intracellular solubility comparison ofstabilized copper metal and degradable copper oxide nanopar-ticlesrdquo Toxicology Letters vol 197 no 3 pp 169ndash174 2010
[21] S Lin P Bhattacharya N C Rajapakse D E Brune and P CKe ldquoEffects of quantum dots adsorption on algal photosynthe-sisrdquo Journal of Physical Chemistry C vol 113 no 25 pp 10962ndash10966 2009
[22] T Xia M Kovochich J Brant et al ldquoComparison of theabilities of ambient and manufactured nanoparticles to inducecellular toxicity according to an oxidative stress paradigmrdquoNano Letters vol 6 no 8 pp 1794ndash1807 2006
[23] X Hu S Cook PWang andH-M Hwang ldquoIn vitro evaluationof cytotoxicity of engineeredmetal oxide nanoparticlesrdquo Scienceof the Total Environment vol 407 no 8 pp 3070ndash3072 2009
[24] W J Stark ldquoNanoparticles in biological systemsrdquo AngewandteChemiemdashInternational Edition vol 50 no 6 pp 1242ndash12582011
[25] F Q Schafer S Y Qian and G R Buettner ldquoIron and freeradical oxidations in cell membranesrdquo Cellular and MolecularBiology vol 46 no 3 pp 657ndash662 2000
[26] D Dewez N A Ali F Perreault and R Popovic ldquoRapidchlorophyll a fluorescence transient of Lemna gibba leaf as anindication of light and hydroxylamine effect on photosystem IIactivityrdquo Photochemical and Photobiological Sciences vol 6 no5 pp 532ndash538 2007
[27] D Dewez N Boucher F Bellemare and R Popovic ldquoUse ofdifferent fluorometric systems in the determination of fluo-rescence parameters from spinach thylakoid membranes beingexposed to atrazine and copperrdquo Toxicological and Environmen-tal Chemistry vol 89 no 4 pp 655ndash664 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 3
0 100 200 300 4000
5
10
15Fe
Solu
ble [
Fe] (120583
g mLminus
1)
[SPION-2] (120583g mLminus1)
(a)
0
2
4
6
8
10Co
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Solu
ble
[Co]
(120583g m
Lminus1)
(b)
0
2
4
6
8
10
0 100 200 300 400
Zn
[SPION-2] (120583g mLminus1)24h48h72h
Solu
ble [
Zn] (120583
g mLminus
1)
(c)
Figure 3 Soluble fraction of free Fe Co and Zn released from SPION-2 (Co02Zn08Fe2O4) suspension in culture medium at 24 48 and 72 h
of exposure
25 Growth Inhibition Test The relative cell division rate(RCDR) was evaluated for 72 h according to [12] as RCDR =(lnW
72 hminus lnW0)72 h whereW72 h represents cell density at72 h and W
0the initial cell density
26 Bioaccumulation of Fe Co and Zn To separate SPIONfrom cells a sucrose gradient prepared in BG-11 media wasdone directly in a Beckman centrifuge tube inclined at a 30∘angle in order to get 6 layers of different sucrose densities(5mL of each sucrose solution 120 100 80 60 40 and20) Algal cells of control and SPION-treated samples werecollected by centrifugation Their pellets were slowly placedon top of sucrose gradient tubes which were centrifuged at1000 rpm during 30min in a swinging-bucket 5810R cen-trifuge (Eppendorf Germany) It resulted in the formationof a pellet of SPION at the bottom of the tube The algal cellslayer was recuperated with a glass Pasteur pipette and filtered
on a 045 120583m filter previously dried and weighted To removeSPIONweakly bound to the cell surface or the filter 3times 10mLof 10mM ethylenediaminetetraacetic acid in BG-11 mediumwas slowly passed through the filter Filters were dried at 95∘Cfor 24 h and weighted to calculate algal dry weight Filterswere then placed in acid-washed glass tubes in which 4mL ofHNO
3and 500 120583L H
2O2were added Samples were digested
during 48 h at room temperature before being diluted to 20HNO
3in Milli-Q purified water for the quantification of Fe
Co and Zn using atomic absorption spectrometry analysis(Varian SpectrAA 220 FS USA) Obtained Fe Co and Znconcentrations were normalized to the dry weight
27 Soluble Fraction of Fe Co and Zn Solubility of free FeCo and Zn released from SPION suspensions were deter-mined in culturemedium SPION suspensions was incubatedduring 24 48 and 72 h in the same condition as described
4 BioMed Research International
0
1
2
3
4Fe
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Solu
ble [
Fe] (120583
g mLminus
1)
(a)
0
1
2
3
4Co
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Solu
ble
[Co]
(120583g m
Lminus1)
(b)
0
1
2
3
4
0 100 200 300 400
Zn
[SPION-3] (120583g mLminus1)24h48h72h
Solu
ble [
Zn] (120583
g mLminus
1)
(c)
Figure 4 Soluble fraction of Fe Co and Zn released from SPION-3 (Co05Zn05Fe2O4) suspension in culture medium at 24 48 and 72 h of
exposure
above for treatment After incubation NPs were removed bycentrifugation at 12000 g for 30min and the supernatant wascollected for analysis Quantification of free Fe Co and Znin solution wasmeasured by atomic absorption spectrometry(Varian SpectrAA 220 FS USA)
28 Production of ROS The fluorescent dye 2101584071015840-dichlorodi-hydrofluorescein diacetate ((H
2DCFDA) Invitrogen Molec-
ular Probe USA) was used as an indicator of ROS accordingto [13] Cellular esterases hydrolyze this probe into the non-fluorescent compound 2101584071015840-dichlorodihydrofluorescein(H2DCF) which is better retained within cells In the
presence of ROS and cellular esterases H2DCF is trans-
formed into the highly fluorescent compound 2101584071015840-dichlo-rofluorescein (DCF) H
2DCFDA stock solution (10mM) was
prepared in ethanol in the dark After 72 h of treatment 1mLof algal samples was exposed during 15min to 02mM of
H2DCFDA in the dark The ROS level was determined by
measuring the fluorescence emission at 530 nm with a flowcytometer (FACScan system Becton Dickinson InstrumentsUSA) Cytometry results were analysed using the WinMDI28 software Algal cells were separated from noncellularparticles by using a relationship between particle size and redfluorescence level originating from chlorophyll fluorescenceemission A positive control sample using methyl viologenwas done to verify the assay (data not shown)
29 Cellular Viability Viability of algal cells was estimatedusing the fluorescein diacetate ((FDA) Invitrogen MolecularProbe USA) method according to [14] FDA is a nonpolarester compoundwhich passes through cell membranes Onceinside the cell FDA is hydrolyzed by esterases (enzymespresent in viable cells) to produce fluorescein accumulatingin cell wall and emitting fluorescence under UV illumination
BioMed Research International 5
After 72 h of treatment 1mL of algal samples was exposedduring 15min to 02mM of FDA in the dark Cell viabilitywas determined by measuring the fluorescence emissionat 530 nm with a flow cytometer (FACScan System Bec-ton Dickinson Instruments USA) Cytometry results wereanalysed using the WinMDI 28 software Algal cells wereseparated from noncellular particles by using a relationshipbetween particle size and red fluorescence level originatingfrom chlorophyll fluorescence emission A positive controlsample using methyl viologen was done to verify the assay(data not shown)
210 Chl 119886 Fluorescence Emission Photosynthetic electrontransport was monitored from the change in the rapid riseof Chl 119886 fluorescence emission using a ldquoPlant EfficiencyAnalyserrdquo fluorometer (Handy-PEA Hansatech Ltd UK)Total chlorophyll (Chl) content (119886 + 119887) was extracted in 100methanol at 65∘C and quantified with a spectrophotometer(Lambda 40 Perkin-Elmer USA) according to the formulaindicated in [15] Total Chl (120583gmLminus1) = (2493 times A
6524+
144 times A6652
)Prior to fluorescence measurements algal samples were
transferred into clean sterile 2mL glass vials and dark-adapted for 30min An aliquot of 5 120583g of total chlorophyllwas gently filtered using low pressure filtration and algal cellswere uniformly placed on a 13mm glass fibre filter (MilliporeUSA) The fluorescence induction was triggered using a 1 ssaturating flash of 3500120583molmminus2 sminus1The fluorescence inten-sity at 20120583s was considered as the 119874 value (119865
119874) fluorescence
intensities for 119870 119869 and 119868 transients were determined at300 120583s (119865
119870) 2ms (119865
119869) and 30ms (119865
119868) respectively the max-
imum fluorescence yield reached maximal value of fluores-cence intensity (119865
119872) under saturating illumination Different
photosynthetic-based fluorescence parameters related to thefunctional state of Photosystem II were calculated [16 17]the maximum efficiency of PSII electron transport 119865
119881119865119872=
(119865119872minus 119865119874)119865119872 the absorption of photons by light harvesting
antenna complexes (ABS) per active reaction center (RC)ABSRC = ((119865
119870minus 119865119874)025) times (1(119865
119869minus 119865119874)) times (119865
119872(119865119872minus
119865119874)) the relative variable fluorescence yield at 119869 transient
estimating the fraction of 119876119860
in its reduced state 119881119869=
(119865119869minus 119865119874)(119865119872minus 119865119874) and the performance index of PSII
photosynthetic activity PI = RCABS times ((119865119872minus 119865119874)119865119874) times
((1 minus 119881119869)119881119869)
211 Statistical Analysis All treatments were performed intriplicate Means and standard deviations were calculatedfor each treatment Significant differences between controland treated plants were determined by one-way analysisof variance (ANOVA) followed by a Dunnettrsquos multiplecomparison (DMC) test where 119875 value less than 005 wasconsidered significant
3 Results
31 Characterisation of SPION When SPION were sus-pended in the media nanoparticles formed agglomeratesduring the first minutes as indicated by the distribution
0
2
4
6
8
10
12
0 100 200 300 400
Bioa
ccum
ulat
ed F
e120583g
mg
of D
W
[SPION-1] (120583g mLminus1)
Figure 5 Bioaccumulated content of Fe in algal cells of Chlorellavulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square) and 72 h(998771 triangle) to different concentrations of SPION-1 (Fe
3O4)
of hydrodynamic particles size diameter which was causedby the content of salts in the media Analysis by dynamiclight scattering showed SPION-1 SPION-2 and SPION-3suspensions in culture media to have an average diameterof particle size distribution of 1959 1765 and 3472 nmrespectively (Figure 1) These distributions of hydrodynamicsize of SPION were found to be stable in the culture mediumduring the entire experimental exposure Furthermore mea-surements of zeta potential (mV) indicated that SPION werenegatively charged in themedia with values ofminus2568 (plusmn138)minus2914 (plusmn385) and minus2806 (plusmn119) respectively for SPION-1SPION-2 and SPION-3
32 Solubility and Bioaccumulation of SPION The solublefraction of free Fe Co and Zn released from SPION-1SPION-2 and SPION-3 suspensions in culture medium wasdetermined at 24 48 and 72 hours (Figures 2 3 and 4)The quantity of soluble Fe Co and Zn in the medium wasdependent on the time of exposure the SPION compositionand their concentration When C vulgaris was exposed to125 120583gmLminus1 of SPION-1 (Fe
3O4) during 72 h the proportion
of the soluble fraction of free Fe was of 27 compared tothe nominal concentration of SPION-1 and it decreased to3 for 400 120583gmLminus1 of SPION-1 (Figure 2) However thesolubility of SPION-2 (Co
02Zn08Fe2O4) was dependent on
the metal species When algal cells were exposed during72 h to 125 120583gmLminus1 of SPION-2 proportions of free FeCo and Zn in the soluble fraction were respectively of27 12 and 9 compared to the nominal concentrationof SPION-2 (Figure 3) For the exposure concentration of400 120583gmLminus1 the quantity of soluble Fe Co andZn decreasedto 3 2 and 1 respectively Concerning the solubility ofSPION-3 (Co
05Zn05Fe2O4) concentrations of Fe Co and
Zn in the soluble fraction at 72 h were of 9 each whencompared to the nominal concentration of 125120583gmLminus1 Forthe exposure concentration of 400 120583gmLminus1 concentrationsof Fe Co and Zn in the soluble fraction were less than
6 BioMed Research International
Fe
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed F
e120583
gm
g of
DW
(a)
Co
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed C
o120583
gm
g of
DW
(b)
Zn
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed Z
n120583
gm
g of
DW
(c)
Figure 6 Bioaccumulated content of Fe Co and Zn in algal cells of Chlorella vulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square)and 72 h (998771 triangle) to different concentrations of SPION-2 (Co
02Zn08Fe2O4)
05 in comparison to the nominal concentration of SPION-3 Indeed we observed that the agglomeration of SPIONincreased their precipitationsedimentation at the bottomof the experimental flask which was directly related to theincreasing concentration tested and time of exposure Thiseffect may cause the reduction of the surface contact ofnanoparticles with the medium explaining the decrease inSPION solubilisation related to their concentration and timeof exposure
The bioaccumulation of total Fe Co and Zn was alsoquantified in algal biomass of C vulgaris exposed during 2448 and 72 h to SPION (Figures 5 6 and 7) For SPION-1the accumulated contents of Fe in algal biomass increasedin dependence on the concentration of SPION and the timeof exposure (Figure 5) The bioaccumulated content of Feincreased by 35-fold for algal cells treated during 72 h to400 120583gmLminus1 of SPION-1 compared to control Under thistreatment condition Fe bioaccumulation increased by 281-fold from 24 to 72 h Concerning SPION-2 the accumulatedcontents of Fe Co and Zn increased in algal biomassin relation to the concentration of SPION (Figure 6) Thecontent of Fe reached its maximum value at 72 h whichincreased by 104-fold for 400120583gmLminus1 of SPION compared
to control On the other hand the contents of Co andZn attained their maximal values at 48 h which increasedrespectively by 537-and 628-fold for 400 120583gmLminus1 of SPIONcompared to control When C vulgaris was exposed toSPION-3 the bioaccumulation of Fe Co and Zn increasedin relation to the concentration of SPION (Figure 7) At 72 hof exposure values of Fe Co and Zn contents increasedrespectively by 442- 329- and 547-fold for 400 120583gmLminus1 ofSPION compared to control
33 Inhibition of the Relative Cell Division Rate When algalcells ofC vulgariswere exposed during 72 h to SPION at con-centrations varying from 125 to 400 120583gmLminus1 the inhibitionof the relative cell division rate based on the change of celldensity was dependent on the tested SPION concentration(Table 1) Under concentration exposure of 400120583gmLminus1 therelative cell division rate decreased significantly compared tothe control by 478 218 and 158 for SPION-1 SPION-2and SPION-3 respectively
34 Inhibition of Photosynthetic Electron Transport Thechange of Chl 119886 fluorescence emission was used to monitorthe photosynthetic electron transport when algal cells of
BioMed Research International 7
Fe
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed F
e120583
gm
g of
DW
(a)
Co
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed C
o120583
gm
g of
DW
(b)
Zn
0
1
2
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed Z
n120583
gm
g of
DW
(c)
Figure 7 Bioaccumulated content of Fe Co and Zn in algal cells of Chlorella vulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square)and 72 h (998771 triangle) to different concentrations of SPION-3 (Co
05Zn05Fe2O4)
Table 1 Change in the relative cell division rate (RCDR) when Cvulgaris was exposed to SPION during 72 h
[SPION]120583gmL
RCDR (10minus6)SPION-1 SPION-2 SPION-3
0 0335 plusmn 001 0335 plusmn 001 0335 plusmn 001
125 0305 plusmn 005 0307 plusmn 005 0317 plusmn 0004
25 0308 plusmn 0008 0298 plusmn 0006 0318 plusmn 0007
50 0307 plusmn 002 0297 plusmn 0018 0309 plusmn 0004
100 0266 plusmn 005 0291 plusmn 0007 0306 plusmn 0025
200 0220 plusmn 0008 0264 plusmn 0002 0286 plusmn 0005
400 0175 plusmn 0123 0262 plusmn 00033 0282 plusmn 0004
C vulgaris were exposed during 72 h to SPION toxicityThe kinetics of Chl 119886 fluorescence indicated a decrease offluorescence yields at 119869 119868 and119872 levels without any changein their time of appearance which was related to SPIONconcentration (See Supplementary Figures available onlineat httpdxdoiorg1011552013647974) Furthermore thechange of photosynthetic-fluorescence parameters estimated
from the fluorescence kinetic was used as biomarkersof SPION effect on PSII photochemistry (Figure 8) Thestrongest effect on photosynthetic electron transport wasobserved under 400 120583gmLminus1 of SPION treatment Underthis condition the value of ABSRC parameter increasedsignificantly compared to the control by 43 29 and 81 forSPION-1 SPION-2 and SPION-3 respectively This changeindicated a decrease of functional PSII reaction centers able toperform photochemical reactions Also119881
119869values indicating
the level of QAminusQA(total) increased significantly compared
to the control by 32 23 and 12 respectively for SPION-1SPION-2 and SPION-3 due to the inhibition of PSII electrontransport flow toward to plastoquinone pool However themaximal quantum yield of PSII (119865
119881119865119872) decreased by 14 12
and 35 respectively for SPION-1 SPION-2 and SPION-3 Moreover the PSII performance index PI was used as aglobal parameter integrating all PSII photochemical reactionsfrom light harvesting energy transfer to electron transportWhen algal cells of C vulgaris were exposed during 72 hto 400 120583gmLminus1 of SPION PI values decreased significantlycompared to the control by 78 65 and 85 for SPION-1SPION-2 and SPION-3 respectively
8 BioMed Research International
ABSRC
0 100 200 300 4000
50
100
150
200Pa
ram
eter
s val
ue (
)
[SPION] (120583g mLminus1)
(a)
0 100 200 300 4000
50
100
150
200
Para
met
ers v
alue
()
VJ
[SPION] (120583g mLminus1)
(b)
0
20
40
60
80
100
120
SPION-1SPION-2SPION-3
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)
FVFM
(c)
PI
0
20
40
60
80
100
120
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)SPION-1SPION-2SPION-3
(d)
Figure 8 Change of different photosynthetic-based fluorescence parameters for algal cells of Chlorella vulgaris exposed during 72 h todifferent concentrations of SPION-1 (Fe
3O4) SPION-2 (Co
02Zn08Fe2O4) and SPION-3 (Co
05Zn05Fe2O4) Maximal PSII quantum yield
119865119881119865119872 ratio between the number of active PSII reaction centers and light harvesting Chl antenna size ABSRC relative variable fluorescence
yield at 119869 transient 119881119869 performance index of PSII photochemical activity PI
35 Production of Reactive Oxygen Species Related to Cell Via-bility The formation of ROS per viable cells was determinedfor C vulgaris exposed during 72 h to SPION (Figure 9)Under these conditions cell viability decreased significantlycompared to control from 50 120583gmLminus1 of SPION indicatingan induction of cellular oxidative stress The productionof ROS per viable cells increased significantly comparedto control which was dependent on SPION species andconcentration
4 Discussion
41 Toxicity of SPION-1 SPION-2 and SPION-3 In thisstudy toxic effects of SPION-1 -2 and -3 were investigatedon algal cells of C vulgaris which were caused by thedeterioration of photochemical activities of photosynthesisthe induction of oxidative stress and the inhibition of cell
division rate This complex cellular alteration was dependenton SPION chemical composition and its concentration insolution Based on our results it is evident that the bioac-cumulation of free Fe Co and Zn from the soluble fractionwas contributing to the toxicity impact in algal cells It waspreviously proposed that the release of free metal ions frommetallic nanoparticle suspensions represented amajor sourceof toxicity for the growth rate of aquatic microorganisms[18 19] However it is difficult to determine if the release offree metal ions from nanoparticles is the only contributionto the toxicity impact in the algal cellular system Indeedsolubilisation of nanoparticles can take place either in themedia or inside the cell which possess an acidic pH environ-ment favorable for particle solubility [20] Therefore anotherhypothesis is that SPION may also contribute directly to thetoxicity impact Recently it was suggested for direct toxicitymechanisms of nanoparticles to induce a direct alteration of
BioMed Research International 9
0
50
100Cell viability
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(a)
ROS
0
50
100
150
200
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(b)
0
100
200
300
400
500
600
700
0 100 200 300 400
ROScell viability
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(c)
Figure 9 Change in the production of reactive oxygen species (ROS) cellular viability and the ratio ROSviable cells for Chlorella vulgarisexposed during 72 h to different concentrations of SPION-1 (⧫ diamond) SPION-2 (◼ square) and SPION-3 (998771 triangle)
cellular exchanges with the media due to particles bindingon cell membrane [21] It was also shown for bioaccumulatedFe3O4nanoparticles to cause direct toxic effects inside plant
cells [8]Moreover the increase of production of reactive oxygen
species per cell viabilitywas related to the increasing exposureconcentration of SPION indicating that the induction ofcellular oxidative stress was caused by the bioaccumulationof Fe Co and Zn Indeed it was shown in previous stud-ies that metallic nanoparticles can cause the formation ofROS via the Fenton reaction or the disruption of majorphysiological processes [22 23] Also the charged metallicsurface of nanoparticles can trigger the formation of ROS bycatalytic reduction of oxygen into superoxide anion leadingto oxidative damage into proteins lipids nucleic acids andpigments [24]
When algal cells of C vulgaris were exposed during 72 hto different concentrations of SPION suspensions the relativecellular division rate was strongly inhibited for SPION-1compared to SPION-2 and SPION-3 Under these treatmentconditions the production of ROS per viable cells was thehighest for SPION-1 Indeed Fe is well known to generate
in cell free radical oxidations causing lipid peroxidation[25] Furthermore the change of fluorescence parameter PIindicated a stronger inhibition of photosynthesis for SPION-1Therefore according to the change of these biomarkers thetoxicity impact in algal cells of C vulgaris was the strongestfor SPION-1 suspensions
42 Significance of Biochemical Biomarkers In this study twodifferent biochemical biomarkers were used to characterizethe toxicity of SPION the production of ROS per viablecells and the photosynthetic-based fluorescence parametersThe change of parameter ROScell viability permitted deter-mining the potential source of cellular toxicity causing theinhibition of cell division rate On the other hand fluores-cence parameters related to PSII photochemical reactionshave been shown to be sensitive biomarkers of SPION toxicityto C vulgaris cells Indeed the Chl 119886 fluorescence emissionis demonstrated to be related to water-splitting system func-tions at PSII reaction center and to oxidoreduction statesof electron transport carriers [26 27] Therefore our resultsshowed evidence for SPION to cause inhibitory effects on thePSII water-splitting system and the photoactivation of PSII
10 BioMed Research International
reaction centers In comparison to other fluorescence indica-tors the performance index of PSII activity representing anintegrative indicator of all PSII photochemical reactions [17]was the most sensitive biomarker of the deterioration of PSIIfunctions caused by SPION toxicity
5 Conclusion
Nowadays a large quantity of metallic nanoparticles isproduced and their toxic potential as hazardous contami-nants in aquatic environment needs to be investigated inorder to develop specific bioassays for nanoparticles toxicityassessment In this regard microalgae represent sensitiveorganisms to be used in bioassay toxicity testing for theassessment of hazardous materials In this toxicologicalinvestigation we clearly showed the potential source of tox-icity of three SPION suspensions having different chemicalcompositions The algal model C vulgaris demonstrated tobe a valuable bioindicator of SPION cellular toxicity whichwas indicated by the deterioration of photochemical activitiesof photosynthesis the induction of oxidative stress and theinhibition of cell division rateTherefore this work permittedcharacterising the cellular toxicity impact of these SPIONwith different biomarkers A good understanding of thesetoxicological interactions will permit better understandingthe risk of SPION toxicity for aquatic organisms
Abbreviations
Chl ChlorophyllNP NanoparticlesPEA Plant Efficiency AnalyzerPSII Photosystem IIROS Reactive oxygen speciesSPION Superparamagnetic iron oxide nanoparticlesTEM Transmission electron microscope
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was performed at the Department of Chem-istry (UQAM) The authors acknowledge the financial sup-port provided by both the Faculty of Sciences and theDepartment of Chemistry (UQAM) The authors wish tothank L Ben Taherc L Samia Smiric and H Abdelmelekbfor providing nanopowder of Fe
3O4 Co02Zn08Fe2O4 and
Co05Zn05Fe2O4
References
[1] A Ito M Shinkai H Honda and T Kobayashi ldquoMedicalapplication of functionalized magnetic nanoparticlesrdquo Journalof Bioscience and Bioengineering vol 100 no 1 pp 1ndash11 2005
[2] D L Huber ldquoSynthesis properties and applications of ironnanoparticlesrdquo Small vol 1 no 5 pp 482ndash501 2005
[3] N Singh G J S Jenkins R Asadi and S H Doak ldquoPoten-tial toxicity of superparamagnetic iron oxide nanoparticles(SPION)rdquo Nano Reviews vol 1 Article ID 5358 2010
[4] I Bhatt and B N Tripathi ldquoInteraction of engineered nanopar-ticles with various components of the environment and possiblestrategies for their risk assessmentrdquo Chemosphere vol 82 no 3pp 308ndash317 2011
[5] MDelay and FH Frimmel ldquoNanoparticles in aquatic systemsrdquoAnalytical and Bioanalytical Chemistry vol 402 no 2 pp 583ndash592 2012
[6] E N Taylor K M Kummer N G Durmus K Leuba K MTarquinio and T J Webster ldquoSuperparamagnetic iron oxidenanoparticles (SPION) for the treatment of antibiotic-resistantbiofilmsrdquo Small vol 8 pp 3016ndash3027 2012
[7] H Zhu J Han J Q Xiao and Y Jin ldquoUptake translocationand accumulation of manufactured iron oxide nanoparticles bypumpkin plantsrdquo Journal of Environmental Monitoring vol 10no 6 pp 713ndash717 2008
[8] Y KMushtaq ldquoEffect of nanoscale Fe3O4 TiO2and carbon par-
ticles on cucumber seed germinationrdquo Journal of EnvironmentalScience and Health A vol 46 no 14 pp 1732ndash1735 2011
[9] X Chen X Zhu R Li H Yao Z Lu and X Yang ldquoPhoto-synthetic toxicity and oxidative damage induced by nano-Fe
3O4
on Chlorella vulgaris in aquatic environmentrdquo Open Journal ofEcology vol 2 pp 21ndash28 2012
[10] R Rippka J Deruelles and J B Waterbury ldquoGeneric assign-ments strain histories and properties of pure cultures ofcyanobacteriardquo Journal of General Microbiology vol 111 no 1pp 1ndash61 1979
[11] H Basti L Ben Tahar L S Smiri et al ldquoCatechol derivatives-coated Fe
3O4and 120574-Fe
2O3nanoparticles as potential MRI
contrast agentsrdquo Journal of Colloid and Interface Science vol 341no 2 pp 248ndash254 2010
[12] W A Hoffmann and H Poorter ldquoAvoiding bias in calculationsof relative growth raterdquo Annals of Botany vol 90 no 1 pp 37ndash42 2002
[13] I B Gerber and I A Dubery ldquoFluorescence microplate assayfor the detection of oxidative burst products in tobacco cellsuspensions using 2101584071015840-dichlorofluoresceinrdquo Methods in CellScience vol 25 no 3-4 pp 115ndash122 2004
[14] P Mayer R Cuhel and N Nyholm ldquoA simple in vitro fluo-rescence method for biomass measurements in algal growthinhibition testsrdquo Water Research vol 31 no 10 pp 2525ndash25311997
[15] A R Wellburn ldquoThe spectral determination of chlorophylls aand b as well as total carotenoids using various solvents withspectrophotometers of different resolutionrdquo Journal of PlantPhysiology vol 144 no 3 pp 307ndash313 1994
[16] L Force C Critchley and J J S Van Rensen ldquoNew fluorescenceparameters for monitoring photosynthesis in plants 1 the effectof illumination on the fluorescence parameters of the JIP-testrdquoPhotosynthesis Research vol 78 no 1 pp 17ndash33 2003
[17] R J Strasser A Srivastava and M Tsimilli-Michael ldquoAnalysisof the chlorophyll a fluorescence transientrdquo in Chlorophyllfluorescence A signature of Photosynthesis Advances in Photo-synthesis and respiration G G Papageorgiou Ed pp 321ndash362Kluwer Academic Dodrecht The Netherlands 2004
[18] N M Franklin N J Rogers S C Apte G E Batley G EGadd and P S Casey ldquoComparative toxicity of nanopartic-ulate ZnO bulk ZnO and ZnCl
2to a freshwater microalga
(Pseudokirchneriella subcapitata) the importance of particle
BioMed Research International 11
solubilityrdquoEnvironmental Science andTechnology vol 41 no 24pp 8484ndash8490 2007
[19] V Aruoja H-C Dubourguier K Kasemets and A KahruldquoToxicity of nanoparticles of CuO ZnO and TiO
2tomicroalgae
Pseudokirchneriella subcapitatardquo Science of the Total Environ-ment vol 407 no 4 pp 1461ndash1468 2009
[20] A M Studer L K Limbach L Van Duc et al ldquoNanoparticlecytotoxicity depends on intracellular solubility comparison ofstabilized copper metal and degradable copper oxide nanopar-ticlesrdquo Toxicology Letters vol 197 no 3 pp 169ndash174 2010
[21] S Lin P Bhattacharya N C Rajapakse D E Brune and P CKe ldquoEffects of quantum dots adsorption on algal photosynthe-sisrdquo Journal of Physical Chemistry C vol 113 no 25 pp 10962ndash10966 2009
[22] T Xia M Kovochich J Brant et al ldquoComparison of theabilities of ambient and manufactured nanoparticles to inducecellular toxicity according to an oxidative stress paradigmrdquoNano Letters vol 6 no 8 pp 1794ndash1807 2006
[23] X Hu S Cook PWang andH-M Hwang ldquoIn vitro evaluationof cytotoxicity of engineeredmetal oxide nanoparticlesrdquo Scienceof the Total Environment vol 407 no 8 pp 3070ndash3072 2009
[24] W J Stark ldquoNanoparticles in biological systemsrdquo AngewandteChemiemdashInternational Edition vol 50 no 6 pp 1242ndash12582011
[25] F Q Schafer S Y Qian and G R Buettner ldquoIron and freeradical oxidations in cell membranesrdquo Cellular and MolecularBiology vol 46 no 3 pp 657ndash662 2000
[26] D Dewez N A Ali F Perreault and R Popovic ldquoRapidchlorophyll a fluorescence transient of Lemna gibba leaf as anindication of light and hydroxylamine effect on photosystem IIactivityrdquo Photochemical and Photobiological Sciences vol 6 no5 pp 532ndash538 2007
[27] D Dewez N Boucher F Bellemare and R Popovic ldquoUse ofdifferent fluorometric systems in the determination of fluo-rescence parameters from spinach thylakoid membranes beingexposed to atrazine and copperrdquo Toxicological and Environmen-tal Chemistry vol 89 no 4 pp 655ndash664 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 BioMed Research International
0
1
2
3
4Fe
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Solu
ble [
Fe] (120583
g mLminus
1)
(a)
0
1
2
3
4Co
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Solu
ble
[Co]
(120583g m
Lminus1)
(b)
0
1
2
3
4
0 100 200 300 400
Zn
[SPION-3] (120583g mLminus1)24h48h72h
Solu
ble [
Zn] (120583
g mLminus
1)
(c)
Figure 4 Soluble fraction of Fe Co and Zn released from SPION-3 (Co05Zn05Fe2O4) suspension in culture medium at 24 48 and 72 h of
exposure
above for treatment After incubation NPs were removed bycentrifugation at 12000 g for 30min and the supernatant wascollected for analysis Quantification of free Fe Co and Znin solution wasmeasured by atomic absorption spectrometry(Varian SpectrAA 220 FS USA)
28 Production of ROS The fluorescent dye 2101584071015840-dichlorodi-hydrofluorescein diacetate ((H
2DCFDA) Invitrogen Molec-
ular Probe USA) was used as an indicator of ROS accordingto [13] Cellular esterases hydrolyze this probe into the non-fluorescent compound 2101584071015840-dichlorodihydrofluorescein(H2DCF) which is better retained within cells In the
presence of ROS and cellular esterases H2DCF is trans-
formed into the highly fluorescent compound 2101584071015840-dichlo-rofluorescein (DCF) H
2DCFDA stock solution (10mM) was
prepared in ethanol in the dark After 72 h of treatment 1mLof algal samples was exposed during 15min to 02mM of
H2DCFDA in the dark The ROS level was determined by
measuring the fluorescence emission at 530 nm with a flowcytometer (FACScan system Becton Dickinson InstrumentsUSA) Cytometry results were analysed using the WinMDI28 software Algal cells were separated from noncellularparticles by using a relationship between particle size and redfluorescence level originating from chlorophyll fluorescenceemission A positive control sample using methyl viologenwas done to verify the assay (data not shown)
29 Cellular Viability Viability of algal cells was estimatedusing the fluorescein diacetate ((FDA) Invitrogen MolecularProbe USA) method according to [14] FDA is a nonpolarester compoundwhich passes through cell membranes Onceinside the cell FDA is hydrolyzed by esterases (enzymespresent in viable cells) to produce fluorescein accumulatingin cell wall and emitting fluorescence under UV illumination
BioMed Research International 5
After 72 h of treatment 1mL of algal samples was exposedduring 15min to 02mM of FDA in the dark Cell viabilitywas determined by measuring the fluorescence emissionat 530 nm with a flow cytometer (FACScan System Bec-ton Dickinson Instruments USA) Cytometry results wereanalysed using the WinMDI 28 software Algal cells wereseparated from noncellular particles by using a relationshipbetween particle size and red fluorescence level originatingfrom chlorophyll fluorescence emission A positive controlsample using methyl viologen was done to verify the assay(data not shown)
210 Chl 119886 Fluorescence Emission Photosynthetic electrontransport was monitored from the change in the rapid riseof Chl 119886 fluorescence emission using a ldquoPlant EfficiencyAnalyserrdquo fluorometer (Handy-PEA Hansatech Ltd UK)Total chlorophyll (Chl) content (119886 + 119887) was extracted in 100methanol at 65∘C and quantified with a spectrophotometer(Lambda 40 Perkin-Elmer USA) according to the formulaindicated in [15] Total Chl (120583gmLminus1) = (2493 times A
6524+
144 times A6652
)Prior to fluorescence measurements algal samples were
transferred into clean sterile 2mL glass vials and dark-adapted for 30min An aliquot of 5 120583g of total chlorophyllwas gently filtered using low pressure filtration and algal cellswere uniformly placed on a 13mm glass fibre filter (MilliporeUSA) The fluorescence induction was triggered using a 1 ssaturating flash of 3500120583molmminus2 sminus1The fluorescence inten-sity at 20120583s was considered as the 119874 value (119865
119874) fluorescence
intensities for 119870 119869 and 119868 transients were determined at300 120583s (119865
119870) 2ms (119865
119869) and 30ms (119865
119868) respectively the max-
imum fluorescence yield reached maximal value of fluores-cence intensity (119865
119872) under saturating illumination Different
photosynthetic-based fluorescence parameters related to thefunctional state of Photosystem II were calculated [16 17]the maximum efficiency of PSII electron transport 119865
119881119865119872=
(119865119872minus 119865119874)119865119872 the absorption of photons by light harvesting
antenna complexes (ABS) per active reaction center (RC)ABSRC = ((119865
119870minus 119865119874)025) times (1(119865
119869minus 119865119874)) times (119865
119872(119865119872minus
119865119874)) the relative variable fluorescence yield at 119869 transient
estimating the fraction of 119876119860
in its reduced state 119881119869=
(119865119869minus 119865119874)(119865119872minus 119865119874) and the performance index of PSII
photosynthetic activity PI = RCABS times ((119865119872minus 119865119874)119865119874) times
((1 minus 119881119869)119881119869)
211 Statistical Analysis All treatments were performed intriplicate Means and standard deviations were calculatedfor each treatment Significant differences between controland treated plants were determined by one-way analysisof variance (ANOVA) followed by a Dunnettrsquos multiplecomparison (DMC) test where 119875 value less than 005 wasconsidered significant
3 Results
31 Characterisation of SPION When SPION were sus-pended in the media nanoparticles formed agglomeratesduring the first minutes as indicated by the distribution
0
2
4
6
8
10
12
0 100 200 300 400
Bioa
ccum
ulat
ed F
e120583g
mg
of D
W
[SPION-1] (120583g mLminus1)
Figure 5 Bioaccumulated content of Fe in algal cells of Chlorellavulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square) and 72 h(998771 triangle) to different concentrations of SPION-1 (Fe
3O4)
of hydrodynamic particles size diameter which was causedby the content of salts in the media Analysis by dynamiclight scattering showed SPION-1 SPION-2 and SPION-3suspensions in culture media to have an average diameterof particle size distribution of 1959 1765 and 3472 nmrespectively (Figure 1) These distributions of hydrodynamicsize of SPION were found to be stable in the culture mediumduring the entire experimental exposure Furthermore mea-surements of zeta potential (mV) indicated that SPION werenegatively charged in themedia with values ofminus2568 (plusmn138)minus2914 (plusmn385) and minus2806 (plusmn119) respectively for SPION-1SPION-2 and SPION-3
32 Solubility and Bioaccumulation of SPION The solublefraction of free Fe Co and Zn released from SPION-1SPION-2 and SPION-3 suspensions in culture medium wasdetermined at 24 48 and 72 hours (Figures 2 3 and 4)The quantity of soluble Fe Co and Zn in the medium wasdependent on the time of exposure the SPION compositionand their concentration When C vulgaris was exposed to125 120583gmLminus1 of SPION-1 (Fe
3O4) during 72 h the proportion
of the soluble fraction of free Fe was of 27 compared tothe nominal concentration of SPION-1 and it decreased to3 for 400 120583gmLminus1 of SPION-1 (Figure 2) However thesolubility of SPION-2 (Co
02Zn08Fe2O4) was dependent on
the metal species When algal cells were exposed during72 h to 125 120583gmLminus1 of SPION-2 proportions of free FeCo and Zn in the soluble fraction were respectively of27 12 and 9 compared to the nominal concentrationof SPION-2 (Figure 3) For the exposure concentration of400 120583gmLminus1 the quantity of soluble Fe Co andZn decreasedto 3 2 and 1 respectively Concerning the solubility ofSPION-3 (Co
05Zn05Fe2O4) concentrations of Fe Co and
Zn in the soluble fraction at 72 h were of 9 each whencompared to the nominal concentration of 125120583gmLminus1 Forthe exposure concentration of 400 120583gmLminus1 concentrationsof Fe Co and Zn in the soluble fraction were less than
6 BioMed Research International
Fe
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed F
e120583
gm
g of
DW
(a)
Co
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed C
o120583
gm
g of
DW
(b)
Zn
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed Z
n120583
gm
g of
DW
(c)
Figure 6 Bioaccumulated content of Fe Co and Zn in algal cells of Chlorella vulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square)and 72 h (998771 triangle) to different concentrations of SPION-2 (Co
02Zn08Fe2O4)
05 in comparison to the nominal concentration of SPION-3 Indeed we observed that the agglomeration of SPIONincreased their precipitationsedimentation at the bottomof the experimental flask which was directly related to theincreasing concentration tested and time of exposure Thiseffect may cause the reduction of the surface contact ofnanoparticles with the medium explaining the decrease inSPION solubilisation related to their concentration and timeof exposure
The bioaccumulation of total Fe Co and Zn was alsoquantified in algal biomass of C vulgaris exposed during 2448 and 72 h to SPION (Figures 5 6 and 7) For SPION-1the accumulated contents of Fe in algal biomass increasedin dependence on the concentration of SPION and the timeof exposure (Figure 5) The bioaccumulated content of Feincreased by 35-fold for algal cells treated during 72 h to400 120583gmLminus1 of SPION-1 compared to control Under thistreatment condition Fe bioaccumulation increased by 281-fold from 24 to 72 h Concerning SPION-2 the accumulatedcontents of Fe Co and Zn increased in algal biomassin relation to the concentration of SPION (Figure 6) Thecontent of Fe reached its maximum value at 72 h whichincreased by 104-fold for 400120583gmLminus1 of SPION compared
to control On the other hand the contents of Co andZn attained their maximal values at 48 h which increasedrespectively by 537-and 628-fold for 400 120583gmLminus1 of SPIONcompared to control When C vulgaris was exposed toSPION-3 the bioaccumulation of Fe Co and Zn increasedin relation to the concentration of SPION (Figure 7) At 72 hof exposure values of Fe Co and Zn contents increasedrespectively by 442- 329- and 547-fold for 400 120583gmLminus1 ofSPION compared to control
33 Inhibition of the Relative Cell Division Rate When algalcells ofC vulgariswere exposed during 72 h to SPION at con-centrations varying from 125 to 400 120583gmLminus1 the inhibitionof the relative cell division rate based on the change of celldensity was dependent on the tested SPION concentration(Table 1) Under concentration exposure of 400120583gmLminus1 therelative cell division rate decreased significantly compared tothe control by 478 218 and 158 for SPION-1 SPION-2and SPION-3 respectively
34 Inhibition of Photosynthetic Electron Transport Thechange of Chl 119886 fluorescence emission was used to monitorthe photosynthetic electron transport when algal cells of
BioMed Research International 7
Fe
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed F
e120583
gm
g of
DW
(a)
Co
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed C
o120583
gm
g of
DW
(b)
Zn
0
1
2
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed Z
n120583
gm
g of
DW
(c)
Figure 7 Bioaccumulated content of Fe Co and Zn in algal cells of Chlorella vulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square)and 72 h (998771 triangle) to different concentrations of SPION-3 (Co
05Zn05Fe2O4)
Table 1 Change in the relative cell division rate (RCDR) when Cvulgaris was exposed to SPION during 72 h
[SPION]120583gmL
RCDR (10minus6)SPION-1 SPION-2 SPION-3
0 0335 plusmn 001 0335 plusmn 001 0335 plusmn 001
125 0305 plusmn 005 0307 plusmn 005 0317 plusmn 0004
25 0308 plusmn 0008 0298 plusmn 0006 0318 plusmn 0007
50 0307 plusmn 002 0297 plusmn 0018 0309 plusmn 0004
100 0266 plusmn 005 0291 plusmn 0007 0306 plusmn 0025
200 0220 plusmn 0008 0264 plusmn 0002 0286 plusmn 0005
400 0175 plusmn 0123 0262 plusmn 00033 0282 plusmn 0004
C vulgaris were exposed during 72 h to SPION toxicityThe kinetics of Chl 119886 fluorescence indicated a decrease offluorescence yields at 119869 119868 and119872 levels without any changein their time of appearance which was related to SPIONconcentration (See Supplementary Figures available onlineat httpdxdoiorg1011552013647974) Furthermore thechange of photosynthetic-fluorescence parameters estimated
from the fluorescence kinetic was used as biomarkersof SPION effect on PSII photochemistry (Figure 8) Thestrongest effect on photosynthetic electron transport wasobserved under 400 120583gmLminus1 of SPION treatment Underthis condition the value of ABSRC parameter increasedsignificantly compared to the control by 43 29 and 81 forSPION-1 SPION-2 and SPION-3 respectively This changeindicated a decrease of functional PSII reaction centers able toperform photochemical reactions Also119881
119869values indicating
the level of QAminusQA(total) increased significantly compared
to the control by 32 23 and 12 respectively for SPION-1SPION-2 and SPION-3 due to the inhibition of PSII electrontransport flow toward to plastoquinone pool However themaximal quantum yield of PSII (119865
119881119865119872) decreased by 14 12
and 35 respectively for SPION-1 SPION-2 and SPION-3 Moreover the PSII performance index PI was used as aglobal parameter integrating all PSII photochemical reactionsfrom light harvesting energy transfer to electron transportWhen algal cells of C vulgaris were exposed during 72 hto 400 120583gmLminus1 of SPION PI values decreased significantlycompared to the control by 78 65 and 85 for SPION-1SPION-2 and SPION-3 respectively
8 BioMed Research International
ABSRC
0 100 200 300 4000
50
100
150
200Pa
ram
eter
s val
ue (
)
[SPION] (120583g mLminus1)
(a)
0 100 200 300 4000
50
100
150
200
Para
met
ers v
alue
()
VJ
[SPION] (120583g mLminus1)
(b)
0
20
40
60
80
100
120
SPION-1SPION-2SPION-3
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)
FVFM
(c)
PI
0
20
40
60
80
100
120
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)SPION-1SPION-2SPION-3
(d)
Figure 8 Change of different photosynthetic-based fluorescence parameters for algal cells of Chlorella vulgaris exposed during 72 h todifferent concentrations of SPION-1 (Fe
3O4) SPION-2 (Co
02Zn08Fe2O4) and SPION-3 (Co
05Zn05Fe2O4) Maximal PSII quantum yield
119865119881119865119872 ratio between the number of active PSII reaction centers and light harvesting Chl antenna size ABSRC relative variable fluorescence
yield at 119869 transient 119881119869 performance index of PSII photochemical activity PI
35 Production of Reactive Oxygen Species Related to Cell Via-bility The formation of ROS per viable cells was determinedfor C vulgaris exposed during 72 h to SPION (Figure 9)Under these conditions cell viability decreased significantlycompared to control from 50 120583gmLminus1 of SPION indicatingan induction of cellular oxidative stress The productionof ROS per viable cells increased significantly comparedto control which was dependent on SPION species andconcentration
4 Discussion
41 Toxicity of SPION-1 SPION-2 and SPION-3 In thisstudy toxic effects of SPION-1 -2 and -3 were investigatedon algal cells of C vulgaris which were caused by thedeterioration of photochemical activities of photosynthesisthe induction of oxidative stress and the inhibition of cell
division rate This complex cellular alteration was dependenton SPION chemical composition and its concentration insolution Based on our results it is evident that the bioac-cumulation of free Fe Co and Zn from the soluble fractionwas contributing to the toxicity impact in algal cells It waspreviously proposed that the release of free metal ions frommetallic nanoparticle suspensions represented amajor sourceof toxicity for the growth rate of aquatic microorganisms[18 19] However it is difficult to determine if the release offree metal ions from nanoparticles is the only contributionto the toxicity impact in the algal cellular system Indeedsolubilisation of nanoparticles can take place either in themedia or inside the cell which possess an acidic pH environ-ment favorable for particle solubility [20] Therefore anotherhypothesis is that SPION may also contribute directly to thetoxicity impact Recently it was suggested for direct toxicitymechanisms of nanoparticles to induce a direct alteration of
BioMed Research International 9
0
50
100Cell viability
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(a)
ROS
0
50
100
150
200
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(b)
0
100
200
300
400
500
600
700
0 100 200 300 400
ROScell viability
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(c)
Figure 9 Change in the production of reactive oxygen species (ROS) cellular viability and the ratio ROSviable cells for Chlorella vulgarisexposed during 72 h to different concentrations of SPION-1 (⧫ diamond) SPION-2 (◼ square) and SPION-3 (998771 triangle)
cellular exchanges with the media due to particles bindingon cell membrane [21] It was also shown for bioaccumulatedFe3O4nanoparticles to cause direct toxic effects inside plant
cells [8]Moreover the increase of production of reactive oxygen
species per cell viabilitywas related to the increasing exposureconcentration of SPION indicating that the induction ofcellular oxidative stress was caused by the bioaccumulationof Fe Co and Zn Indeed it was shown in previous stud-ies that metallic nanoparticles can cause the formation ofROS via the Fenton reaction or the disruption of majorphysiological processes [22 23] Also the charged metallicsurface of nanoparticles can trigger the formation of ROS bycatalytic reduction of oxygen into superoxide anion leadingto oxidative damage into proteins lipids nucleic acids andpigments [24]
When algal cells of C vulgaris were exposed during 72 hto different concentrations of SPION suspensions the relativecellular division rate was strongly inhibited for SPION-1compared to SPION-2 and SPION-3 Under these treatmentconditions the production of ROS per viable cells was thehighest for SPION-1 Indeed Fe is well known to generate
in cell free radical oxidations causing lipid peroxidation[25] Furthermore the change of fluorescence parameter PIindicated a stronger inhibition of photosynthesis for SPION-1Therefore according to the change of these biomarkers thetoxicity impact in algal cells of C vulgaris was the strongestfor SPION-1 suspensions
42 Significance of Biochemical Biomarkers In this study twodifferent biochemical biomarkers were used to characterizethe toxicity of SPION the production of ROS per viablecells and the photosynthetic-based fluorescence parametersThe change of parameter ROScell viability permitted deter-mining the potential source of cellular toxicity causing theinhibition of cell division rate On the other hand fluores-cence parameters related to PSII photochemical reactionshave been shown to be sensitive biomarkers of SPION toxicityto C vulgaris cells Indeed the Chl 119886 fluorescence emissionis demonstrated to be related to water-splitting system func-tions at PSII reaction center and to oxidoreduction statesof electron transport carriers [26 27] Therefore our resultsshowed evidence for SPION to cause inhibitory effects on thePSII water-splitting system and the photoactivation of PSII
10 BioMed Research International
reaction centers In comparison to other fluorescence indica-tors the performance index of PSII activity representing anintegrative indicator of all PSII photochemical reactions [17]was the most sensitive biomarker of the deterioration of PSIIfunctions caused by SPION toxicity
5 Conclusion
Nowadays a large quantity of metallic nanoparticles isproduced and their toxic potential as hazardous contami-nants in aquatic environment needs to be investigated inorder to develop specific bioassays for nanoparticles toxicityassessment In this regard microalgae represent sensitiveorganisms to be used in bioassay toxicity testing for theassessment of hazardous materials In this toxicologicalinvestigation we clearly showed the potential source of tox-icity of three SPION suspensions having different chemicalcompositions The algal model C vulgaris demonstrated tobe a valuable bioindicator of SPION cellular toxicity whichwas indicated by the deterioration of photochemical activitiesof photosynthesis the induction of oxidative stress and theinhibition of cell division rateTherefore this work permittedcharacterising the cellular toxicity impact of these SPIONwith different biomarkers A good understanding of thesetoxicological interactions will permit better understandingthe risk of SPION toxicity for aquatic organisms
Abbreviations
Chl ChlorophyllNP NanoparticlesPEA Plant Efficiency AnalyzerPSII Photosystem IIROS Reactive oxygen speciesSPION Superparamagnetic iron oxide nanoparticlesTEM Transmission electron microscope
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was performed at the Department of Chem-istry (UQAM) The authors acknowledge the financial sup-port provided by both the Faculty of Sciences and theDepartment of Chemistry (UQAM) The authors wish tothank L Ben Taherc L Samia Smiric and H Abdelmelekbfor providing nanopowder of Fe
3O4 Co02Zn08Fe2O4 and
Co05Zn05Fe2O4
References
[1] A Ito M Shinkai H Honda and T Kobayashi ldquoMedicalapplication of functionalized magnetic nanoparticlesrdquo Journalof Bioscience and Bioengineering vol 100 no 1 pp 1ndash11 2005
[2] D L Huber ldquoSynthesis properties and applications of ironnanoparticlesrdquo Small vol 1 no 5 pp 482ndash501 2005
[3] N Singh G J S Jenkins R Asadi and S H Doak ldquoPoten-tial toxicity of superparamagnetic iron oxide nanoparticles(SPION)rdquo Nano Reviews vol 1 Article ID 5358 2010
[4] I Bhatt and B N Tripathi ldquoInteraction of engineered nanopar-ticles with various components of the environment and possiblestrategies for their risk assessmentrdquo Chemosphere vol 82 no 3pp 308ndash317 2011
[5] MDelay and FH Frimmel ldquoNanoparticles in aquatic systemsrdquoAnalytical and Bioanalytical Chemistry vol 402 no 2 pp 583ndash592 2012
[6] E N Taylor K M Kummer N G Durmus K Leuba K MTarquinio and T J Webster ldquoSuperparamagnetic iron oxidenanoparticles (SPION) for the treatment of antibiotic-resistantbiofilmsrdquo Small vol 8 pp 3016ndash3027 2012
[7] H Zhu J Han J Q Xiao and Y Jin ldquoUptake translocationand accumulation of manufactured iron oxide nanoparticles bypumpkin plantsrdquo Journal of Environmental Monitoring vol 10no 6 pp 713ndash717 2008
[8] Y KMushtaq ldquoEffect of nanoscale Fe3O4 TiO2and carbon par-
ticles on cucumber seed germinationrdquo Journal of EnvironmentalScience and Health A vol 46 no 14 pp 1732ndash1735 2011
[9] X Chen X Zhu R Li H Yao Z Lu and X Yang ldquoPhoto-synthetic toxicity and oxidative damage induced by nano-Fe
3O4
on Chlorella vulgaris in aquatic environmentrdquo Open Journal ofEcology vol 2 pp 21ndash28 2012
[10] R Rippka J Deruelles and J B Waterbury ldquoGeneric assign-ments strain histories and properties of pure cultures ofcyanobacteriardquo Journal of General Microbiology vol 111 no 1pp 1ndash61 1979
[11] H Basti L Ben Tahar L S Smiri et al ldquoCatechol derivatives-coated Fe
3O4and 120574-Fe
2O3nanoparticles as potential MRI
contrast agentsrdquo Journal of Colloid and Interface Science vol 341no 2 pp 248ndash254 2010
[12] W A Hoffmann and H Poorter ldquoAvoiding bias in calculationsof relative growth raterdquo Annals of Botany vol 90 no 1 pp 37ndash42 2002
[13] I B Gerber and I A Dubery ldquoFluorescence microplate assayfor the detection of oxidative burst products in tobacco cellsuspensions using 2101584071015840-dichlorofluoresceinrdquo Methods in CellScience vol 25 no 3-4 pp 115ndash122 2004
[14] P Mayer R Cuhel and N Nyholm ldquoA simple in vitro fluo-rescence method for biomass measurements in algal growthinhibition testsrdquo Water Research vol 31 no 10 pp 2525ndash25311997
[15] A R Wellburn ldquoThe spectral determination of chlorophylls aand b as well as total carotenoids using various solvents withspectrophotometers of different resolutionrdquo Journal of PlantPhysiology vol 144 no 3 pp 307ndash313 1994
[16] L Force C Critchley and J J S Van Rensen ldquoNew fluorescenceparameters for monitoring photosynthesis in plants 1 the effectof illumination on the fluorescence parameters of the JIP-testrdquoPhotosynthesis Research vol 78 no 1 pp 17ndash33 2003
[17] R J Strasser A Srivastava and M Tsimilli-Michael ldquoAnalysisof the chlorophyll a fluorescence transientrdquo in Chlorophyllfluorescence A signature of Photosynthesis Advances in Photo-synthesis and respiration G G Papageorgiou Ed pp 321ndash362Kluwer Academic Dodrecht The Netherlands 2004
[18] N M Franklin N J Rogers S C Apte G E Batley G EGadd and P S Casey ldquoComparative toxicity of nanopartic-ulate ZnO bulk ZnO and ZnCl
2to a freshwater microalga
(Pseudokirchneriella subcapitata) the importance of particle
BioMed Research International 11
solubilityrdquoEnvironmental Science andTechnology vol 41 no 24pp 8484ndash8490 2007
[19] V Aruoja H-C Dubourguier K Kasemets and A KahruldquoToxicity of nanoparticles of CuO ZnO and TiO
2tomicroalgae
Pseudokirchneriella subcapitatardquo Science of the Total Environ-ment vol 407 no 4 pp 1461ndash1468 2009
[20] A M Studer L K Limbach L Van Duc et al ldquoNanoparticlecytotoxicity depends on intracellular solubility comparison ofstabilized copper metal and degradable copper oxide nanopar-ticlesrdquo Toxicology Letters vol 197 no 3 pp 169ndash174 2010
[21] S Lin P Bhattacharya N C Rajapakse D E Brune and P CKe ldquoEffects of quantum dots adsorption on algal photosynthe-sisrdquo Journal of Physical Chemistry C vol 113 no 25 pp 10962ndash10966 2009
[22] T Xia M Kovochich J Brant et al ldquoComparison of theabilities of ambient and manufactured nanoparticles to inducecellular toxicity according to an oxidative stress paradigmrdquoNano Letters vol 6 no 8 pp 1794ndash1807 2006
[23] X Hu S Cook PWang andH-M Hwang ldquoIn vitro evaluationof cytotoxicity of engineeredmetal oxide nanoparticlesrdquo Scienceof the Total Environment vol 407 no 8 pp 3070ndash3072 2009
[24] W J Stark ldquoNanoparticles in biological systemsrdquo AngewandteChemiemdashInternational Edition vol 50 no 6 pp 1242ndash12582011
[25] F Q Schafer S Y Qian and G R Buettner ldquoIron and freeradical oxidations in cell membranesrdquo Cellular and MolecularBiology vol 46 no 3 pp 657ndash662 2000
[26] D Dewez N A Ali F Perreault and R Popovic ldquoRapidchlorophyll a fluorescence transient of Lemna gibba leaf as anindication of light and hydroxylamine effect on photosystem IIactivityrdquo Photochemical and Photobiological Sciences vol 6 no5 pp 532ndash538 2007
[27] D Dewez N Boucher F Bellemare and R Popovic ldquoUse ofdifferent fluorometric systems in the determination of fluo-rescence parameters from spinach thylakoid membranes beingexposed to atrazine and copperrdquo Toxicological and Environmen-tal Chemistry vol 89 no 4 pp 655ndash664 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 5
After 72 h of treatment 1mL of algal samples was exposedduring 15min to 02mM of FDA in the dark Cell viabilitywas determined by measuring the fluorescence emissionat 530 nm with a flow cytometer (FACScan System Bec-ton Dickinson Instruments USA) Cytometry results wereanalysed using the WinMDI 28 software Algal cells wereseparated from noncellular particles by using a relationshipbetween particle size and red fluorescence level originatingfrom chlorophyll fluorescence emission A positive controlsample using methyl viologen was done to verify the assay(data not shown)
210 Chl 119886 Fluorescence Emission Photosynthetic electrontransport was monitored from the change in the rapid riseof Chl 119886 fluorescence emission using a ldquoPlant EfficiencyAnalyserrdquo fluorometer (Handy-PEA Hansatech Ltd UK)Total chlorophyll (Chl) content (119886 + 119887) was extracted in 100methanol at 65∘C and quantified with a spectrophotometer(Lambda 40 Perkin-Elmer USA) according to the formulaindicated in [15] Total Chl (120583gmLminus1) = (2493 times A
6524+
144 times A6652
)Prior to fluorescence measurements algal samples were
transferred into clean sterile 2mL glass vials and dark-adapted for 30min An aliquot of 5 120583g of total chlorophyllwas gently filtered using low pressure filtration and algal cellswere uniformly placed on a 13mm glass fibre filter (MilliporeUSA) The fluorescence induction was triggered using a 1 ssaturating flash of 3500120583molmminus2 sminus1The fluorescence inten-sity at 20120583s was considered as the 119874 value (119865
119874) fluorescence
intensities for 119870 119869 and 119868 transients were determined at300 120583s (119865
119870) 2ms (119865
119869) and 30ms (119865
119868) respectively the max-
imum fluorescence yield reached maximal value of fluores-cence intensity (119865
119872) under saturating illumination Different
photosynthetic-based fluorescence parameters related to thefunctional state of Photosystem II were calculated [16 17]the maximum efficiency of PSII electron transport 119865
119881119865119872=
(119865119872minus 119865119874)119865119872 the absorption of photons by light harvesting
antenna complexes (ABS) per active reaction center (RC)ABSRC = ((119865
119870minus 119865119874)025) times (1(119865
119869minus 119865119874)) times (119865
119872(119865119872minus
119865119874)) the relative variable fluorescence yield at 119869 transient
estimating the fraction of 119876119860
in its reduced state 119881119869=
(119865119869minus 119865119874)(119865119872minus 119865119874) and the performance index of PSII
photosynthetic activity PI = RCABS times ((119865119872minus 119865119874)119865119874) times
((1 minus 119881119869)119881119869)
211 Statistical Analysis All treatments were performed intriplicate Means and standard deviations were calculatedfor each treatment Significant differences between controland treated plants were determined by one-way analysisof variance (ANOVA) followed by a Dunnettrsquos multiplecomparison (DMC) test where 119875 value less than 005 wasconsidered significant
3 Results
31 Characterisation of SPION When SPION were sus-pended in the media nanoparticles formed agglomeratesduring the first minutes as indicated by the distribution
0
2
4
6
8
10
12
0 100 200 300 400
Bioa
ccum
ulat
ed F
e120583g
mg
of D
W
[SPION-1] (120583g mLminus1)
Figure 5 Bioaccumulated content of Fe in algal cells of Chlorellavulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square) and 72 h(998771 triangle) to different concentrations of SPION-1 (Fe
3O4)
of hydrodynamic particles size diameter which was causedby the content of salts in the media Analysis by dynamiclight scattering showed SPION-1 SPION-2 and SPION-3suspensions in culture media to have an average diameterof particle size distribution of 1959 1765 and 3472 nmrespectively (Figure 1) These distributions of hydrodynamicsize of SPION were found to be stable in the culture mediumduring the entire experimental exposure Furthermore mea-surements of zeta potential (mV) indicated that SPION werenegatively charged in themedia with values ofminus2568 (plusmn138)minus2914 (plusmn385) and minus2806 (plusmn119) respectively for SPION-1SPION-2 and SPION-3
32 Solubility and Bioaccumulation of SPION The solublefraction of free Fe Co and Zn released from SPION-1SPION-2 and SPION-3 suspensions in culture medium wasdetermined at 24 48 and 72 hours (Figures 2 3 and 4)The quantity of soluble Fe Co and Zn in the medium wasdependent on the time of exposure the SPION compositionand their concentration When C vulgaris was exposed to125 120583gmLminus1 of SPION-1 (Fe
3O4) during 72 h the proportion
of the soluble fraction of free Fe was of 27 compared tothe nominal concentration of SPION-1 and it decreased to3 for 400 120583gmLminus1 of SPION-1 (Figure 2) However thesolubility of SPION-2 (Co
02Zn08Fe2O4) was dependent on
the metal species When algal cells were exposed during72 h to 125 120583gmLminus1 of SPION-2 proportions of free FeCo and Zn in the soluble fraction were respectively of27 12 and 9 compared to the nominal concentrationof SPION-2 (Figure 3) For the exposure concentration of400 120583gmLminus1 the quantity of soluble Fe Co andZn decreasedto 3 2 and 1 respectively Concerning the solubility ofSPION-3 (Co
05Zn05Fe2O4) concentrations of Fe Co and
Zn in the soluble fraction at 72 h were of 9 each whencompared to the nominal concentration of 125120583gmLminus1 Forthe exposure concentration of 400 120583gmLminus1 concentrationsof Fe Co and Zn in the soluble fraction were less than
6 BioMed Research International
Fe
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed F
e120583
gm
g of
DW
(a)
Co
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed C
o120583
gm
g of
DW
(b)
Zn
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed Z
n120583
gm
g of
DW
(c)
Figure 6 Bioaccumulated content of Fe Co and Zn in algal cells of Chlorella vulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square)and 72 h (998771 triangle) to different concentrations of SPION-2 (Co
02Zn08Fe2O4)
05 in comparison to the nominal concentration of SPION-3 Indeed we observed that the agglomeration of SPIONincreased their precipitationsedimentation at the bottomof the experimental flask which was directly related to theincreasing concentration tested and time of exposure Thiseffect may cause the reduction of the surface contact ofnanoparticles with the medium explaining the decrease inSPION solubilisation related to their concentration and timeof exposure
The bioaccumulation of total Fe Co and Zn was alsoquantified in algal biomass of C vulgaris exposed during 2448 and 72 h to SPION (Figures 5 6 and 7) For SPION-1the accumulated contents of Fe in algal biomass increasedin dependence on the concentration of SPION and the timeof exposure (Figure 5) The bioaccumulated content of Feincreased by 35-fold for algal cells treated during 72 h to400 120583gmLminus1 of SPION-1 compared to control Under thistreatment condition Fe bioaccumulation increased by 281-fold from 24 to 72 h Concerning SPION-2 the accumulatedcontents of Fe Co and Zn increased in algal biomassin relation to the concentration of SPION (Figure 6) Thecontent of Fe reached its maximum value at 72 h whichincreased by 104-fold for 400120583gmLminus1 of SPION compared
to control On the other hand the contents of Co andZn attained their maximal values at 48 h which increasedrespectively by 537-and 628-fold for 400 120583gmLminus1 of SPIONcompared to control When C vulgaris was exposed toSPION-3 the bioaccumulation of Fe Co and Zn increasedin relation to the concentration of SPION (Figure 7) At 72 hof exposure values of Fe Co and Zn contents increasedrespectively by 442- 329- and 547-fold for 400 120583gmLminus1 ofSPION compared to control
33 Inhibition of the Relative Cell Division Rate When algalcells ofC vulgariswere exposed during 72 h to SPION at con-centrations varying from 125 to 400 120583gmLminus1 the inhibitionof the relative cell division rate based on the change of celldensity was dependent on the tested SPION concentration(Table 1) Under concentration exposure of 400120583gmLminus1 therelative cell division rate decreased significantly compared tothe control by 478 218 and 158 for SPION-1 SPION-2and SPION-3 respectively
34 Inhibition of Photosynthetic Electron Transport Thechange of Chl 119886 fluorescence emission was used to monitorthe photosynthetic electron transport when algal cells of
BioMed Research International 7
Fe
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed F
e120583
gm
g of
DW
(a)
Co
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed C
o120583
gm
g of
DW
(b)
Zn
0
1
2
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed Z
n120583
gm
g of
DW
(c)
Figure 7 Bioaccumulated content of Fe Co and Zn in algal cells of Chlorella vulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square)and 72 h (998771 triangle) to different concentrations of SPION-3 (Co
05Zn05Fe2O4)
Table 1 Change in the relative cell division rate (RCDR) when Cvulgaris was exposed to SPION during 72 h
[SPION]120583gmL
RCDR (10minus6)SPION-1 SPION-2 SPION-3
0 0335 plusmn 001 0335 plusmn 001 0335 plusmn 001
125 0305 plusmn 005 0307 plusmn 005 0317 plusmn 0004
25 0308 plusmn 0008 0298 plusmn 0006 0318 plusmn 0007
50 0307 plusmn 002 0297 plusmn 0018 0309 plusmn 0004
100 0266 plusmn 005 0291 plusmn 0007 0306 plusmn 0025
200 0220 plusmn 0008 0264 plusmn 0002 0286 plusmn 0005
400 0175 plusmn 0123 0262 plusmn 00033 0282 plusmn 0004
C vulgaris were exposed during 72 h to SPION toxicityThe kinetics of Chl 119886 fluorescence indicated a decrease offluorescence yields at 119869 119868 and119872 levels without any changein their time of appearance which was related to SPIONconcentration (See Supplementary Figures available onlineat httpdxdoiorg1011552013647974) Furthermore thechange of photosynthetic-fluorescence parameters estimated
from the fluorescence kinetic was used as biomarkersof SPION effect on PSII photochemistry (Figure 8) Thestrongest effect on photosynthetic electron transport wasobserved under 400 120583gmLminus1 of SPION treatment Underthis condition the value of ABSRC parameter increasedsignificantly compared to the control by 43 29 and 81 forSPION-1 SPION-2 and SPION-3 respectively This changeindicated a decrease of functional PSII reaction centers able toperform photochemical reactions Also119881
119869values indicating
the level of QAminusQA(total) increased significantly compared
to the control by 32 23 and 12 respectively for SPION-1SPION-2 and SPION-3 due to the inhibition of PSII electrontransport flow toward to plastoquinone pool However themaximal quantum yield of PSII (119865
119881119865119872) decreased by 14 12
and 35 respectively for SPION-1 SPION-2 and SPION-3 Moreover the PSII performance index PI was used as aglobal parameter integrating all PSII photochemical reactionsfrom light harvesting energy transfer to electron transportWhen algal cells of C vulgaris were exposed during 72 hto 400 120583gmLminus1 of SPION PI values decreased significantlycompared to the control by 78 65 and 85 for SPION-1SPION-2 and SPION-3 respectively
8 BioMed Research International
ABSRC
0 100 200 300 4000
50
100
150
200Pa
ram
eter
s val
ue (
)
[SPION] (120583g mLminus1)
(a)
0 100 200 300 4000
50
100
150
200
Para
met
ers v
alue
()
VJ
[SPION] (120583g mLminus1)
(b)
0
20
40
60
80
100
120
SPION-1SPION-2SPION-3
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)
FVFM
(c)
PI
0
20
40
60
80
100
120
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)SPION-1SPION-2SPION-3
(d)
Figure 8 Change of different photosynthetic-based fluorescence parameters for algal cells of Chlorella vulgaris exposed during 72 h todifferent concentrations of SPION-1 (Fe
3O4) SPION-2 (Co
02Zn08Fe2O4) and SPION-3 (Co
05Zn05Fe2O4) Maximal PSII quantum yield
119865119881119865119872 ratio between the number of active PSII reaction centers and light harvesting Chl antenna size ABSRC relative variable fluorescence
yield at 119869 transient 119881119869 performance index of PSII photochemical activity PI
35 Production of Reactive Oxygen Species Related to Cell Via-bility The formation of ROS per viable cells was determinedfor C vulgaris exposed during 72 h to SPION (Figure 9)Under these conditions cell viability decreased significantlycompared to control from 50 120583gmLminus1 of SPION indicatingan induction of cellular oxidative stress The productionof ROS per viable cells increased significantly comparedto control which was dependent on SPION species andconcentration
4 Discussion
41 Toxicity of SPION-1 SPION-2 and SPION-3 In thisstudy toxic effects of SPION-1 -2 and -3 were investigatedon algal cells of C vulgaris which were caused by thedeterioration of photochemical activities of photosynthesisthe induction of oxidative stress and the inhibition of cell
division rate This complex cellular alteration was dependenton SPION chemical composition and its concentration insolution Based on our results it is evident that the bioac-cumulation of free Fe Co and Zn from the soluble fractionwas contributing to the toxicity impact in algal cells It waspreviously proposed that the release of free metal ions frommetallic nanoparticle suspensions represented amajor sourceof toxicity for the growth rate of aquatic microorganisms[18 19] However it is difficult to determine if the release offree metal ions from nanoparticles is the only contributionto the toxicity impact in the algal cellular system Indeedsolubilisation of nanoparticles can take place either in themedia or inside the cell which possess an acidic pH environ-ment favorable for particle solubility [20] Therefore anotherhypothesis is that SPION may also contribute directly to thetoxicity impact Recently it was suggested for direct toxicitymechanisms of nanoparticles to induce a direct alteration of
BioMed Research International 9
0
50
100Cell viability
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(a)
ROS
0
50
100
150
200
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(b)
0
100
200
300
400
500
600
700
0 100 200 300 400
ROScell viability
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(c)
Figure 9 Change in the production of reactive oxygen species (ROS) cellular viability and the ratio ROSviable cells for Chlorella vulgarisexposed during 72 h to different concentrations of SPION-1 (⧫ diamond) SPION-2 (◼ square) and SPION-3 (998771 triangle)
cellular exchanges with the media due to particles bindingon cell membrane [21] It was also shown for bioaccumulatedFe3O4nanoparticles to cause direct toxic effects inside plant
cells [8]Moreover the increase of production of reactive oxygen
species per cell viabilitywas related to the increasing exposureconcentration of SPION indicating that the induction ofcellular oxidative stress was caused by the bioaccumulationof Fe Co and Zn Indeed it was shown in previous stud-ies that metallic nanoparticles can cause the formation ofROS via the Fenton reaction or the disruption of majorphysiological processes [22 23] Also the charged metallicsurface of nanoparticles can trigger the formation of ROS bycatalytic reduction of oxygen into superoxide anion leadingto oxidative damage into proteins lipids nucleic acids andpigments [24]
When algal cells of C vulgaris were exposed during 72 hto different concentrations of SPION suspensions the relativecellular division rate was strongly inhibited for SPION-1compared to SPION-2 and SPION-3 Under these treatmentconditions the production of ROS per viable cells was thehighest for SPION-1 Indeed Fe is well known to generate
in cell free radical oxidations causing lipid peroxidation[25] Furthermore the change of fluorescence parameter PIindicated a stronger inhibition of photosynthesis for SPION-1Therefore according to the change of these biomarkers thetoxicity impact in algal cells of C vulgaris was the strongestfor SPION-1 suspensions
42 Significance of Biochemical Biomarkers In this study twodifferent biochemical biomarkers were used to characterizethe toxicity of SPION the production of ROS per viablecells and the photosynthetic-based fluorescence parametersThe change of parameter ROScell viability permitted deter-mining the potential source of cellular toxicity causing theinhibition of cell division rate On the other hand fluores-cence parameters related to PSII photochemical reactionshave been shown to be sensitive biomarkers of SPION toxicityto C vulgaris cells Indeed the Chl 119886 fluorescence emissionis demonstrated to be related to water-splitting system func-tions at PSII reaction center and to oxidoreduction statesof electron transport carriers [26 27] Therefore our resultsshowed evidence for SPION to cause inhibitory effects on thePSII water-splitting system and the photoactivation of PSII
10 BioMed Research International
reaction centers In comparison to other fluorescence indica-tors the performance index of PSII activity representing anintegrative indicator of all PSII photochemical reactions [17]was the most sensitive biomarker of the deterioration of PSIIfunctions caused by SPION toxicity
5 Conclusion
Nowadays a large quantity of metallic nanoparticles isproduced and their toxic potential as hazardous contami-nants in aquatic environment needs to be investigated inorder to develop specific bioassays for nanoparticles toxicityassessment In this regard microalgae represent sensitiveorganisms to be used in bioassay toxicity testing for theassessment of hazardous materials In this toxicologicalinvestigation we clearly showed the potential source of tox-icity of three SPION suspensions having different chemicalcompositions The algal model C vulgaris demonstrated tobe a valuable bioindicator of SPION cellular toxicity whichwas indicated by the deterioration of photochemical activitiesof photosynthesis the induction of oxidative stress and theinhibition of cell division rateTherefore this work permittedcharacterising the cellular toxicity impact of these SPIONwith different biomarkers A good understanding of thesetoxicological interactions will permit better understandingthe risk of SPION toxicity for aquatic organisms
Abbreviations
Chl ChlorophyllNP NanoparticlesPEA Plant Efficiency AnalyzerPSII Photosystem IIROS Reactive oxygen speciesSPION Superparamagnetic iron oxide nanoparticlesTEM Transmission electron microscope
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was performed at the Department of Chem-istry (UQAM) The authors acknowledge the financial sup-port provided by both the Faculty of Sciences and theDepartment of Chemistry (UQAM) The authors wish tothank L Ben Taherc L Samia Smiric and H Abdelmelekbfor providing nanopowder of Fe
3O4 Co02Zn08Fe2O4 and
Co05Zn05Fe2O4
References
[1] A Ito M Shinkai H Honda and T Kobayashi ldquoMedicalapplication of functionalized magnetic nanoparticlesrdquo Journalof Bioscience and Bioengineering vol 100 no 1 pp 1ndash11 2005
[2] D L Huber ldquoSynthesis properties and applications of ironnanoparticlesrdquo Small vol 1 no 5 pp 482ndash501 2005
[3] N Singh G J S Jenkins R Asadi and S H Doak ldquoPoten-tial toxicity of superparamagnetic iron oxide nanoparticles(SPION)rdquo Nano Reviews vol 1 Article ID 5358 2010
[4] I Bhatt and B N Tripathi ldquoInteraction of engineered nanopar-ticles with various components of the environment and possiblestrategies for their risk assessmentrdquo Chemosphere vol 82 no 3pp 308ndash317 2011
[5] MDelay and FH Frimmel ldquoNanoparticles in aquatic systemsrdquoAnalytical and Bioanalytical Chemistry vol 402 no 2 pp 583ndash592 2012
[6] E N Taylor K M Kummer N G Durmus K Leuba K MTarquinio and T J Webster ldquoSuperparamagnetic iron oxidenanoparticles (SPION) for the treatment of antibiotic-resistantbiofilmsrdquo Small vol 8 pp 3016ndash3027 2012
[7] H Zhu J Han J Q Xiao and Y Jin ldquoUptake translocationand accumulation of manufactured iron oxide nanoparticles bypumpkin plantsrdquo Journal of Environmental Monitoring vol 10no 6 pp 713ndash717 2008
[8] Y KMushtaq ldquoEffect of nanoscale Fe3O4 TiO2and carbon par-
ticles on cucumber seed germinationrdquo Journal of EnvironmentalScience and Health A vol 46 no 14 pp 1732ndash1735 2011
[9] X Chen X Zhu R Li H Yao Z Lu and X Yang ldquoPhoto-synthetic toxicity and oxidative damage induced by nano-Fe
3O4
on Chlorella vulgaris in aquatic environmentrdquo Open Journal ofEcology vol 2 pp 21ndash28 2012
[10] R Rippka J Deruelles and J B Waterbury ldquoGeneric assign-ments strain histories and properties of pure cultures ofcyanobacteriardquo Journal of General Microbiology vol 111 no 1pp 1ndash61 1979
[11] H Basti L Ben Tahar L S Smiri et al ldquoCatechol derivatives-coated Fe
3O4and 120574-Fe
2O3nanoparticles as potential MRI
contrast agentsrdquo Journal of Colloid and Interface Science vol 341no 2 pp 248ndash254 2010
[12] W A Hoffmann and H Poorter ldquoAvoiding bias in calculationsof relative growth raterdquo Annals of Botany vol 90 no 1 pp 37ndash42 2002
[13] I B Gerber and I A Dubery ldquoFluorescence microplate assayfor the detection of oxidative burst products in tobacco cellsuspensions using 2101584071015840-dichlorofluoresceinrdquo Methods in CellScience vol 25 no 3-4 pp 115ndash122 2004
[14] P Mayer R Cuhel and N Nyholm ldquoA simple in vitro fluo-rescence method for biomass measurements in algal growthinhibition testsrdquo Water Research vol 31 no 10 pp 2525ndash25311997
[15] A R Wellburn ldquoThe spectral determination of chlorophylls aand b as well as total carotenoids using various solvents withspectrophotometers of different resolutionrdquo Journal of PlantPhysiology vol 144 no 3 pp 307ndash313 1994
[16] L Force C Critchley and J J S Van Rensen ldquoNew fluorescenceparameters for monitoring photosynthesis in plants 1 the effectof illumination on the fluorescence parameters of the JIP-testrdquoPhotosynthesis Research vol 78 no 1 pp 17ndash33 2003
[17] R J Strasser A Srivastava and M Tsimilli-Michael ldquoAnalysisof the chlorophyll a fluorescence transientrdquo in Chlorophyllfluorescence A signature of Photosynthesis Advances in Photo-synthesis and respiration G G Papageorgiou Ed pp 321ndash362Kluwer Academic Dodrecht The Netherlands 2004
[18] N M Franklin N J Rogers S C Apte G E Batley G EGadd and P S Casey ldquoComparative toxicity of nanopartic-ulate ZnO bulk ZnO and ZnCl
2to a freshwater microalga
(Pseudokirchneriella subcapitata) the importance of particle
BioMed Research International 11
solubilityrdquoEnvironmental Science andTechnology vol 41 no 24pp 8484ndash8490 2007
[19] V Aruoja H-C Dubourguier K Kasemets and A KahruldquoToxicity of nanoparticles of CuO ZnO and TiO
2tomicroalgae
Pseudokirchneriella subcapitatardquo Science of the Total Environ-ment vol 407 no 4 pp 1461ndash1468 2009
[20] A M Studer L K Limbach L Van Duc et al ldquoNanoparticlecytotoxicity depends on intracellular solubility comparison ofstabilized copper metal and degradable copper oxide nanopar-ticlesrdquo Toxicology Letters vol 197 no 3 pp 169ndash174 2010
[21] S Lin P Bhattacharya N C Rajapakse D E Brune and P CKe ldquoEffects of quantum dots adsorption on algal photosynthe-sisrdquo Journal of Physical Chemistry C vol 113 no 25 pp 10962ndash10966 2009
[22] T Xia M Kovochich J Brant et al ldquoComparison of theabilities of ambient and manufactured nanoparticles to inducecellular toxicity according to an oxidative stress paradigmrdquoNano Letters vol 6 no 8 pp 1794ndash1807 2006
[23] X Hu S Cook PWang andH-M Hwang ldquoIn vitro evaluationof cytotoxicity of engineeredmetal oxide nanoparticlesrdquo Scienceof the Total Environment vol 407 no 8 pp 3070ndash3072 2009
[24] W J Stark ldquoNanoparticles in biological systemsrdquo AngewandteChemiemdashInternational Edition vol 50 no 6 pp 1242ndash12582011
[25] F Q Schafer S Y Qian and G R Buettner ldquoIron and freeradical oxidations in cell membranesrdquo Cellular and MolecularBiology vol 46 no 3 pp 657ndash662 2000
[26] D Dewez N A Ali F Perreault and R Popovic ldquoRapidchlorophyll a fluorescence transient of Lemna gibba leaf as anindication of light and hydroxylamine effect on photosystem IIactivityrdquo Photochemical and Photobiological Sciences vol 6 no5 pp 532ndash538 2007
[27] D Dewez N Boucher F Bellemare and R Popovic ldquoUse ofdifferent fluorometric systems in the determination of fluo-rescence parameters from spinach thylakoid membranes beingexposed to atrazine and copperrdquo Toxicological and Environmen-tal Chemistry vol 89 no 4 pp 655ndash664 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 BioMed Research International
Fe
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed F
e120583
gm
g of
DW
(a)
Co
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed C
o120583
gm
g of
DW
(b)
Zn
0
2
4
6
8
0 100 200 300 400
[SPION-2] (120583g mLminus1)
Bioa
ccum
ulat
ed Z
n120583
gm
g of
DW
(c)
Figure 6 Bioaccumulated content of Fe Co and Zn in algal cells of Chlorella vulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square)and 72 h (998771 triangle) to different concentrations of SPION-2 (Co
02Zn08Fe2O4)
05 in comparison to the nominal concentration of SPION-3 Indeed we observed that the agglomeration of SPIONincreased their precipitationsedimentation at the bottomof the experimental flask which was directly related to theincreasing concentration tested and time of exposure Thiseffect may cause the reduction of the surface contact ofnanoparticles with the medium explaining the decrease inSPION solubilisation related to their concentration and timeof exposure
The bioaccumulation of total Fe Co and Zn was alsoquantified in algal biomass of C vulgaris exposed during 2448 and 72 h to SPION (Figures 5 6 and 7) For SPION-1the accumulated contents of Fe in algal biomass increasedin dependence on the concentration of SPION and the timeof exposure (Figure 5) The bioaccumulated content of Feincreased by 35-fold for algal cells treated during 72 h to400 120583gmLminus1 of SPION-1 compared to control Under thistreatment condition Fe bioaccumulation increased by 281-fold from 24 to 72 h Concerning SPION-2 the accumulatedcontents of Fe Co and Zn increased in algal biomassin relation to the concentration of SPION (Figure 6) Thecontent of Fe reached its maximum value at 72 h whichincreased by 104-fold for 400120583gmLminus1 of SPION compared
to control On the other hand the contents of Co andZn attained their maximal values at 48 h which increasedrespectively by 537-and 628-fold for 400 120583gmLminus1 of SPIONcompared to control When C vulgaris was exposed toSPION-3 the bioaccumulation of Fe Co and Zn increasedin relation to the concentration of SPION (Figure 7) At 72 hof exposure values of Fe Co and Zn contents increasedrespectively by 442- 329- and 547-fold for 400 120583gmLminus1 ofSPION compared to control
33 Inhibition of the Relative Cell Division Rate When algalcells ofC vulgariswere exposed during 72 h to SPION at con-centrations varying from 125 to 400 120583gmLminus1 the inhibitionof the relative cell division rate based on the change of celldensity was dependent on the tested SPION concentration(Table 1) Under concentration exposure of 400120583gmLminus1 therelative cell division rate decreased significantly compared tothe control by 478 218 and 158 for SPION-1 SPION-2and SPION-3 respectively
34 Inhibition of Photosynthetic Electron Transport Thechange of Chl 119886 fluorescence emission was used to monitorthe photosynthetic electron transport when algal cells of
BioMed Research International 7
Fe
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed F
e120583
gm
g of
DW
(a)
Co
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed C
o120583
gm
g of
DW
(b)
Zn
0
1
2
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed Z
n120583
gm
g of
DW
(c)
Figure 7 Bioaccumulated content of Fe Co and Zn in algal cells of Chlorella vulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square)and 72 h (998771 triangle) to different concentrations of SPION-3 (Co
05Zn05Fe2O4)
Table 1 Change in the relative cell division rate (RCDR) when Cvulgaris was exposed to SPION during 72 h
[SPION]120583gmL
RCDR (10minus6)SPION-1 SPION-2 SPION-3
0 0335 plusmn 001 0335 plusmn 001 0335 plusmn 001
125 0305 plusmn 005 0307 plusmn 005 0317 plusmn 0004
25 0308 plusmn 0008 0298 plusmn 0006 0318 plusmn 0007
50 0307 plusmn 002 0297 plusmn 0018 0309 plusmn 0004
100 0266 plusmn 005 0291 plusmn 0007 0306 plusmn 0025
200 0220 plusmn 0008 0264 plusmn 0002 0286 plusmn 0005
400 0175 plusmn 0123 0262 plusmn 00033 0282 plusmn 0004
C vulgaris were exposed during 72 h to SPION toxicityThe kinetics of Chl 119886 fluorescence indicated a decrease offluorescence yields at 119869 119868 and119872 levels without any changein their time of appearance which was related to SPIONconcentration (See Supplementary Figures available onlineat httpdxdoiorg1011552013647974) Furthermore thechange of photosynthetic-fluorescence parameters estimated
from the fluorescence kinetic was used as biomarkersof SPION effect on PSII photochemistry (Figure 8) Thestrongest effect on photosynthetic electron transport wasobserved under 400 120583gmLminus1 of SPION treatment Underthis condition the value of ABSRC parameter increasedsignificantly compared to the control by 43 29 and 81 forSPION-1 SPION-2 and SPION-3 respectively This changeindicated a decrease of functional PSII reaction centers able toperform photochemical reactions Also119881
119869values indicating
the level of QAminusQA(total) increased significantly compared
to the control by 32 23 and 12 respectively for SPION-1SPION-2 and SPION-3 due to the inhibition of PSII electrontransport flow toward to plastoquinone pool However themaximal quantum yield of PSII (119865
119881119865119872) decreased by 14 12
and 35 respectively for SPION-1 SPION-2 and SPION-3 Moreover the PSII performance index PI was used as aglobal parameter integrating all PSII photochemical reactionsfrom light harvesting energy transfer to electron transportWhen algal cells of C vulgaris were exposed during 72 hto 400 120583gmLminus1 of SPION PI values decreased significantlycompared to the control by 78 65 and 85 for SPION-1SPION-2 and SPION-3 respectively
8 BioMed Research International
ABSRC
0 100 200 300 4000
50
100
150
200Pa
ram
eter
s val
ue (
)
[SPION] (120583g mLminus1)
(a)
0 100 200 300 4000
50
100
150
200
Para
met
ers v
alue
()
VJ
[SPION] (120583g mLminus1)
(b)
0
20
40
60
80
100
120
SPION-1SPION-2SPION-3
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)
FVFM
(c)
PI
0
20
40
60
80
100
120
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)SPION-1SPION-2SPION-3
(d)
Figure 8 Change of different photosynthetic-based fluorescence parameters for algal cells of Chlorella vulgaris exposed during 72 h todifferent concentrations of SPION-1 (Fe
3O4) SPION-2 (Co
02Zn08Fe2O4) and SPION-3 (Co
05Zn05Fe2O4) Maximal PSII quantum yield
119865119881119865119872 ratio between the number of active PSII reaction centers and light harvesting Chl antenna size ABSRC relative variable fluorescence
yield at 119869 transient 119881119869 performance index of PSII photochemical activity PI
35 Production of Reactive Oxygen Species Related to Cell Via-bility The formation of ROS per viable cells was determinedfor C vulgaris exposed during 72 h to SPION (Figure 9)Under these conditions cell viability decreased significantlycompared to control from 50 120583gmLminus1 of SPION indicatingan induction of cellular oxidative stress The productionof ROS per viable cells increased significantly comparedto control which was dependent on SPION species andconcentration
4 Discussion
41 Toxicity of SPION-1 SPION-2 and SPION-3 In thisstudy toxic effects of SPION-1 -2 and -3 were investigatedon algal cells of C vulgaris which were caused by thedeterioration of photochemical activities of photosynthesisthe induction of oxidative stress and the inhibition of cell
division rate This complex cellular alteration was dependenton SPION chemical composition and its concentration insolution Based on our results it is evident that the bioac-cumulation of free Fe Co and Zn from the soluble fractionwas contributing to the toxicity impact in algal cells It waspreviously proposed that the release of free metal ions frommetallic nanoparticle suspensions represented amajor sourceof toxicity for the growth rate of aquatic microorganisms[18 19] However it is difficult to determine if the release offree metal ions from nanoparticles is the only contributionto the toxicity impact in the algal cellular system Indeedsolubilisation of nanoparticles can take place either in themedia or inside the cell which possess an acidic pH environ-ment favorable for particle solubility [20] Therefore anotherhypothesis is that SPION may also contribute directly to thetoxicity impact Recently it was suggested for direct toxicitymechanisms of nanoparticles to induce a direct alteration of
BioMed Research International 9
0
50
100Cell viability
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(a)
ROS
0
50
100
150
200
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(b)
0
100
200
300
400
500
600
700
0 100 200 300 400
ROScell viability
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(c)
Figure 9 Change in the production of reactive oxygen species (ROS) cellular viability and the ratio ROSviable cells for Chlorella vulgarisexposed during 72 h to different concentrations of SPION-1 (⧫ diamond) SPION-2 (◼ square) and SPION-3 (998771 triangle)
cellular exchanges with the media due to particles bindingon cell membrane [21] It was also shown for bioaccumulatedFe3O4nanoparticles to cause direct toxic effects inside plant
cells [8]Moreover the increase of production of reactive oxygen
species per cell viabilitywas related to the increasing exposureconcentration of SPION indicating that the induction ofcellular oxidative stress was caused by the bioaccumulationof Fe Co and Zn Indeed it was shown in previous stud-ies that metallic nanoparticles can cause the formation ofROS via the Fenton reaction or the disruption of majorphysiological processes [22 23] Also the charged metallicsurface of nanoparticles can trigger the formation of ROS bycatalytic reduction of oxygen into superoxide anion leadingto oxidative damage into proteins lipids nucleic acids andpigments [24]
When algal cells of C vulgaris were exposed during 72 hto different concentrations of SPION suspensions the relativecellular division rate was strongly inhibited for SPION-1compared to SPION-2 and SPION-3 Under these treatmentconditions the production of ROS per viable cells was thehighest for SPION-1 Indeed Fe is well known to generate
in cell free radical oxidations causing lipid peroxidation[25] Furthermore the change of fluorescence parameter PIindicated a stronger inhibition of photosynthesis for SPION-1Therefore according to the change of these biomarkers thetoxicity impact in algal cells of C vulgaris was the strongestfor SPION-1 suspensions
42 Significance of Biochemical Biomarkers In this study twodifferent biochemical biomarkers were used to characterizethe toxicity of SPION the production of ROS per viablecells and the photosynthetic-based fluorescence parametersThe change of parameter ROScell viability permitted deter-mining the potential source of cellular toxicity causing theinhibition of cell division rate On the other hand fluores-cence parameters related to PSII photochemical reactionshave been shown to be sensitive biomarkers of SPION toxicityto C vulgaris cells Indeed the Chl 119886 fluorescence emissionis demonstrated to be related to water-splitting system func-tions at PSII reaction center and to oxidoreduction statesof electron transport carriers [26 27] Therefore our resultsshowed evidence for SPION to cause inhibitory effects on thePSII water-splitting system and the photoactivation of PSII
10 BioMed Research International
reaction centers In comparison to other fluorescence indica-tors the performance index of PSII activity representing anintegrative indicator of all PSII photochemical reactions [17]was the most sensitive biomarker of the deterioration of PSIIfunctions caused by SPION toxicity
5 Conclusion
Nowadays a large quantity of metallic nanoparticles isproduced and their toxic potential as hazardous contami-nants in aquatic environment needs to be investigated inorder to develop specific bioassays for nanoparticles toxicityassessment In this regard microalgae represent sensitiveorganisms to be used in bioassay toxicity testing for theassessment of hazardous materials In this toxicologicalinvestigation we clearly showed the potential source of tox-icity of three SPION suspensions having different chemicalcompositions The algal model C vulgaris demonstrated tobe a valuable bioindicator of SPION cellular toxicity whichwas indicated by the deterioration of photochemical activitiesof photosynthesis the induction of oxidative stress and theinhibition of cell division rateTherefore this work permittedcharacterising the cellular toxicity impact of these SPIONwith different biomarkers A good understanding of thesetoxicological interactions will permit better understandingthe risk of SPION toxicity for aquatic organisms
Abbreviations
Chl ChlorophyllNP NanoparticlesPEA Plant Efficiency AnalyzerPSII Photosystem IIROS Reactive oxygen speciesSPION Superparamagnetic iron oxide nanoparticlesTEM Transmission electron microscope
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was performed at the Department of Chem-istry (UQAM) The authors acknowledge the financial sup-port provided by both the Faculty of Sciences and theDepartment of Chemistry (UQAM) The authors wish tothank L Ben Taherc L Samia Smiric and H Abdelmelekbfor providing nanopowder of Fe
3O4 Co02Zn08Fe2O4 and
Co05Zn05Fe2O4
References
[1] A Ito M Shinkai H Honda and T Kobayashi ldquoMedicalapplication of functionalized magnetic nanoparticlesrdquo Journalof Bioscience and Bioengineering vol 100 no 1 pp 1ndash11 2005
[2] D L Huber ldquoSynthesis properties and applications of ironnanoparticlesrdquo Small vol 1 no 5 pp 482ndash501 2005
[3] N Singh G J S Jenkins R Asadi and S H Doak ldquoPoten-tial toxicity of superparamagnetic iron oxide nanoparticles(SPION)rdquo Nano Reviews vol 1 Article ID 5358 2010
[4] I Bhatt and B N Tripathi ldquoInteraction of engineered nanopar-ticles with various components of the environment and possiblestrategies for their risk assessmentrdquo Chemosphere vol 82 no 3pp 308ndash317 2011
[5] MDelay and FH Frimmel ldquoNanoparticles in aquatic systemsrdquoAnalytical and Bioanalytical Chemistry vol 402 no 2 pp 583ndash592 2012
[6] E N Taylor K M Kummer N G Durmus K Leuba K MTarquinio and T J Webster ldquoSuperparamagnetic iron oxidenanoparticles (SPION) for the treatment of antibiotic-resistantbiofilmsrdquo Small vol 8 pp 3016ndash3027 2012
[7] H Zhu J Han J Q Xiao and Y Jin ldquoUptake translocationand accumulation of manufactured iron oxide nanoparticles bypumpkin plantsrdquo Journal of Environmental Monitoring vol 10no 6 pp 713ndash717 2008
[8] Y KMushtaq ldquoEffect of nanoscale Fe3O4 TiO2and carbon par-
ticles on cucumber seed germinationrdquo Journal of EnvironmentalScience and Health A vol 46 no 14 pp 1732ndash1735 2011
[9] X Chen X Zhu R Li H Yao Z Lu and X Yang ldquoPhoto-synthetic toxicity and oxidative damage induced by nano-Fe
3O4
on Chlorella vulgaris in aquatic environmentrdquo Open Journal ofEcology vol 2 pp 21ndash28 2012
[10] R Rippka J Deruelles and J B Waterbury ldquoGeneric assign-ments strain histories and properties of pure cultures ofcyanobacteriardquo Journal of General Microbiology vol 111 no 1pp 1ndash61 1979
[11] H Basti L Ben Tahar L S Smiri et al ldquoCatechol derivatives-coated Fe
3O4and 120574-Fe
2O3nanoparticles as potential MRI
contrast agentsrdquo Journal of Colloid and Interface Science vol 341no 2 pp 248ndash254 2010
[12] W A Hoffmann and H Poorter ldquoAvoiding bias in calculationsof relative growth raterdquo Annals of Botany vol 90 no 1 pp 37ndash42 2002
[13] I B Gerber and I A Dubery ldquoFluorescence microplate assayfor the detection of oxidative burst products in tobacco cellsuspensions using 2101584071015840-dichlorofluoresceinrdquo Methods in CellScience vol 25 no 3-4 pp 115ndash122 2004
[14] P Mayer R Cuhel and N Nyholm ldquoA simple in vitro fluo-rescence method for biomass measurements in algal growthinhibition testsrdquo Water Research vol 31 no 10 pp 2525ndash25311997
[15] A R Wellburn ldquoThe spectral determination of chlorophylls aand b as well as total carotenoids using various solvents withspectrophotometers of different resolutionrdquo Journal of PlantPhysiology vol 144 no 3 pp 307ndash313 1994
[16] L Force C Critchley and J J S Van Rensen ldquoNew fluorescenceparameters for monitoring photosynthesis in plants 1 the effectof illumination on the fluorescence parameters of the JIP-testrdquoPhotosynthesis Research vol 78 no 1 pp 17ndash33 2003
[17] R J Strasser A Srivastava and M Tsimilli-Michael ldquoAnalysisof the chlorophyll a fluorescence transientrdquo in Chlorophyllfluorescence A signature of Photosynthesis Advances in Photo-synthesis and respiration G G Papageorgiou Ed pp 321ndash362Kluwer Academic Dodrecht The Netherlands 2004
[18] N M Franklin N J Rogers S C Apte G E Batley G EGadd and P S Casey ldquoComparative toxicity of nanopartic-ulate ZnO bulk ZnO and ZnCl
2to a freshwater microalga
(Pseudokirchneriella subcapitata) the importance of particle
BioMed Research International 11
solubilityrdquoEnvironmental Science andTechnology vol 41 no 24pp 8484ndash8490 2007
[19] V Aruoja H-C Dubourguier K Kasemets and A KahruldquoToxicity of nanoparticles of CuO ZnO and TiO
2tomicroalgae
Pseudokirchneriella subcapitatardquo Science of the Total Environ-ment vol 407 no 4 pp 1461ndash1468 2009
[20] A M Studer L K Limbach L Van Duc et al ldquoNanoparticlecytotoxicity depends on intracellular solubility comparison ofstabilized copper metal and degradable copper oxide nanopar-ticlesrdquo Toxicology Letters vol 197 no 3 pp 169ndash174 2010
[21] S Lin P Bhattacharya N C Rajapakse D E Brune and P CKe ldquoEffects of quantum dots adsorption on algal photosynthe-sisrdquo Journal of Physical Chemistry C vol 113 no 25 pp 10962ndash10966 2009
[22] T Xia M Kovochich J Brant et al ldquoComparison of theabilities of ambient and manufactured nanoparticles to inducecellular toxicity according to an oxidative stress paradigmrdquoNano Letters vol 6 no 8 pp 1794ndash1807 2006
[23] X Hu S Cook PWang andH-M Hwang ldquoIn vitro evaluationof cytotoxicity of engineeredmetal oxide nanoparticlesrdquo Scienceof the Total Environment vol 407 no 8 pp 3070ndash3072 2009
[24] W J Stark ldquoNanoparticles in biological systemsrdquo AngewandteChemiemdashInternational Edition vol 50 no 6 pp 1242ndash12582011
[25] F Q Schafer S Y Qian and G R Buettner ldquoIron and freeradical oxidations in cell membranesrdquo Cellular and MolecularBiology vol 46 no 3 pp 657ndash662 2000
[26] D Dewez N A Ali F Perreault and R Popovic ldquoRapidchlorophyll a fluorescence transient of Lemna gibba leaf as anindication of light and hydroxylamine effect on photosystem IIactivityrdquo Photochemical and Photobiological Sciences vol 6 no5 pp 532ndash538 2007
[27] D Dewez N Boucher F Bellemare and R Popovic ldquoUse ofdifferent fluorometric systems in the determination of fluo-rescence parameters from spinach thylakoid membranes beingexposed to atrazine and copperrdquo Toxicological and Environmen-tal Chemistry vol 89 no 4 pp 655ndash664 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
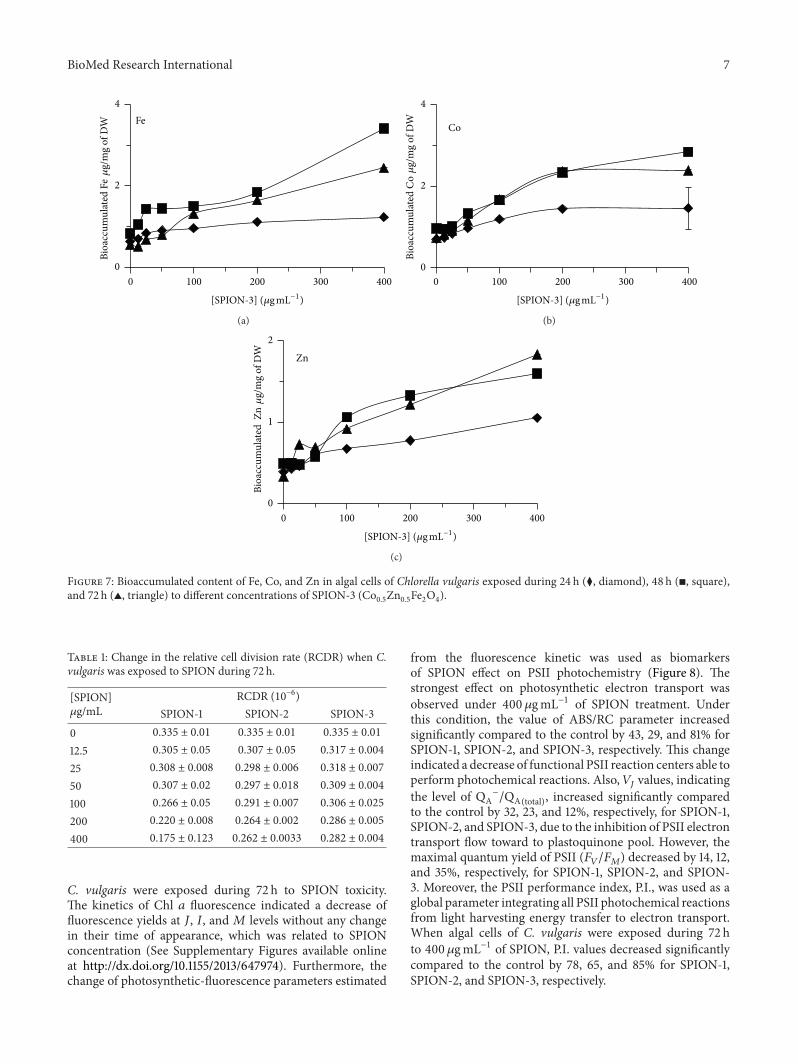
BioMed Research International 7
Fe
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed F
e120583
gm
g of
DW
(a)
Co
0
2
4
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed C
o120583
gm
g of
DW
(b)
Zn
0
1
2
0 100 200 300 400
[SPION-3] (120583g mLminus1)
Bioa
ccum
ulat
ed Z
n120583
gm
g of
DW
(c)
Figure 7 Bioaccumulated content of Fe Co and Zn in algal cells of Chlorella vulgaris exposed during 24 h (⧫ diamond) 48 h (◼ square)and 72 h (998771 triangle) to different concentrations of SPION-3 (Co
05Zn05Fe2O4)
Table 1 Change in the relative cell division rate (RCDR) when Cvulgaris was exposed to SPION during 72 h
[SPION]120583gmL
RCDR (10minus6)SPION-1 SPION-2 SPION-3
0 0335 plusmn 001 0335 plusmn 001 0335 plusmn 001
125 0305 plusmn 005 0307 plusmn 005 0317 plusmn 0004
25 0308 plusmn 0008 0298 plusmn 0006 0318 plusmn 0007
50 0307 plusmn 002 0297 plusmn 0018 0309 plusmn 0004
100 0266 plusmn 005 0291 plusmn 0007 0306 plusmn 0025
200 0220 plusmn 0008 0264 plusmn 0002 0286 plusmn 0005
400 0175 plusmn 0123 0262 plusmn 00033 0282 plusmn 0004
C vulgaris were exposed during 72 h to SPION toxicityThe kinetics of Chl 119886 fluorescence indicated a decrease offluorescence yields at 119869 119868 and119872 levels without any changein their time of appearance which was related to SPIONconcentration (See Supplementary Figures available onlineat httpdxdoiorg1011552013647974) Furthermore thechange of photosynthetic-fluorescence parameters estimated
from the fluorescence kinetic was used as biomarkersof SPION effect on PSII photochemistry (Figure 8) Thestrongest effect on photosynthetic electron transport wasobserved under 400 120583gmLminus1 of SPION treatment Underthis condition the value of ABSRC parameter increasedsignificantly compared to the control by 43 29 and 81 forSPION-1 SPION-2 and SPION-3 respectively This changeindicated a decrease of functional PSII reaction centers able toperform photochemical reactions Also119881
119869values indicating
the level of QAminusQA(total) increased significantly compared
to the control by 32 23 and 12 respectively for SPION-1SPION-2 and SPION-3 due to the inhibition of PSII electrontransport flow toward to plastoquinone pool However themaximal quantum yield of PSII (119865
119881119865119872) decreased by 14 12
and 35 respectively for SPION-1 SPION-2 and SPION-3 Moreover the PSII performance index PI was used as aglobal parameter integrating all PSII photochemical reactionsfrom light harvesting energy transfer to electron transportWhen algal cells of C vulgaris were exposed during 72 hto 400 120583gmLminus1 of SPION PI values decreased significantlycompared to the control by 78 65 and 85 for SPION-1SPION-2 and SPION-3 respectively
8 BioMed Research International
ABSRC
0 100 200 300 4000
50
100
150
200Pa
ram
eter
s val
ue (
)
[SPION] (120583g mLminus1)
(a)
0 100 200 300 4000
50
100
150
200
Para
met
ers v
alue
()
VJ
[SPION] (120583g mLminus1)
(b)
0
20
40
60
80
100
120
SPION-1SPION-2SPION-3
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)
FVFM
(c)
PI
0
20
40
60
80
100
120
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)SPION-1SPION-2SPION-3
(d)
Figure 8 Change of different photosynthetic-based fluorescence parameters for algal cells of Chlorella vulgaris exposed during 72 h todifferent concentrations of SPION-1 (Fe
3O4) SPION-2 (Co
02Zn08Fe2O4) and SPION-3 (Co
05Zn05Fe2O4) Maximal PSII quantum yield
119865119881119865119872 ratio between the number of active PSII reaction centers and light harvesting Chl antenna size ABSRC relative variable fluorescence
yield at 119869 transient 119881119869 performance index of PSII photochemical activity PI
35 Production of Reactive Oxygen Species Related to Cell Via-bility The formation of ROS per viable cells was determinedfor C vulgaris exposed during 72 h to SPION (Figure 9)Under these conditions cell viability decreased significantlycompared to control from 50 120583gmLminus1 of SPION indicatingan induction of cellular oxidative stress The productionof ROS per viable cells increased significantly comparedto control which was dependent on SPION species andconcentration
4 Discussion
41 Toxicity of SPION-1 SPION-2 and SPION-3 In thisstudy toxic effects of SPION-1 -2 and -3 were investigatedon algal cells of C vulgaris which were caused by thedeterioration of photochemical activities of photosynthesisthe induction of oxidative stress and the inhibition of cell
division rate This complex cellular alteration was dependenton SPION chemical composition and its concentration insolution Based on our results it is evident that the bioac-cumulation of free Fe Co and Zn from the soluble fractionwas contributing to the toxicity impact in algal cells It waspreviously proposed that the release of free metal ions frommetallic nanoparticle suspensions represented amajor sourceof toxicity for the growth rate of aquatic microorganisms[18 19] However it is difficult to determine if the release offree metal ions from nanoparticles is the only contributionto the toxicity impact in the algal cellular system Indeedsolubilisation of nanoparticles can take place either in themedia or inside the cell which possess an acidic pH environ-ment favorable for particle solubility [20] Therefore anotherhypothesis is that SPION may also contribute directly to thetoxicity impact Recently it was suggested for direct toxicitymechanisms of nanoparticles to induce a direct alteration of
BioMed Research International 9
0
50
100Cell viability
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(a)
ROS
0
50
100
150
200
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(b)
0
100
200
300
400
500
600
700
0 100 200 300 400
ROScell viability
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(c)
Figure 9 Change in the production of reactive oxygen species (ROS) cellular viability and the ratio ROSviable cells for Chlorella vulgarisexposed during 72 h to different concentrations of SPION-1 (⧫ diamond) SPION-2 (◼ square) and SPION-3 (998771 triangle)
cellular exchanges with the media due to particles bindingon cell membrane [21] It was also shown for bioaccumulatedFe3O4nanoparticles to cause direct toxic effects inside plant
cells [8]Moreover the increase of production of reactive oxygen
species per cell viabilitywas related to the increasing exposureconcentration of SPION indicating that the induction ofcellular oxidative stress was caused by the bioaccumulationof Fe Co and Zn Indeed it was shown in previous stud-ies that metallic nanoparticles can cause the formation ofROS via the Fenton reaction or the disruption of majorphysiological processes [22 23] Also the charged metallicsurface of nanoparticles can trigger the formation of ROS bycatalytic reduction of oxygen into superoxide anion leadingto oxidative damage into proteins lipids nucleic acids andpigments [24]
When algal cells of C vulgaris were exposed during 72 hto different concentrations of SPION suspensions the relativecellular division rate was strongly inhibited for SPION-1compared to SPION-2 and SPION-3 Under these treatmentconditions the production of ROS per viable cells was thehighest for SPION-1 Indeed Fe is well known to generate
in cell free radical oxidations causing lipid peroxidation[25] Furthermore the change of fluorescence parameter PIindicated a stronger inhibition of photosynthesis for SPION-1Therefore according to the change of these biomarkers thetoxicity impact in algal cells of C vulgaris was the strongestfor SPION-1 suspensions
42 Significance of Biochemical Biomarkers In this study twodifferent biochemical biomarkers were used to characterizethe toxicity of SPION the production of ROS per viablecells and the photosynthetic-based fluorescence parametersThe change of parameter ROScell viability permitted deter-mining the potential source of cellular toxicity causing theinhibition of cell division rate On the other hand fluores-cence parameters related to PSII photochemical reactionshave been shown to be sensitive biomarkers of SPION toxicityto C vulgaris cells Indeed the Chl 119886 fluorescence emissionis demonstrated to be related to water-splitting system func-tions at PSII reaction center and to oxidoreduction statesof electron transport carriers [26 27] Therefore our resultsshowed evidence for SPION to cause inhibitory effects on thePSII water-splitting system and the photoactivation of PSII
10 BioMed Research International
reaction centers In comparison to other fluorescence indica-tors the performance index of PSII activity representing anintegrative indicator of all PSII photochemical reactions [17]was the most sensitive biomarker of the deterioration of PSIIfunctions caused by SPION toxicity
5 Conclusion
Nowadays a large quantity of metallic nanoparticles isproduced and their toxic potential as hazardous contami-nants in aquatic environment needs to be investigated inorder to develop specific bioassays for nanoparticles toxicityassessment In this regard microalgae represent sensitiveorganisms to be used in bioassay toxicity testing for theassessment of hazardous materials In this toxicologicalinvestigation we clearly showed the potential source of tox-icity of three SPION suspensions having different chemicalcompositions The algal model C vulgaris demonstrated tobe a valuable bioindicator of SPION cellular toxicity whichwas indicated by the deterioration of photochemical activitiesof photosynthesis the induction of oxidative stress and theinhibition of cell division rateTherefore this work permittedcharacterising the cellular toxicity impact of these SPIONwith different biomarkers A good understanding of thesetoxicological interactions will permit better understandingthe risk of SPION toxicity for aquatic organisms
Abbreviations
Chl ChlorophyllNP NanoparticlesPEA Plant Efficiency AnalyzerPSII Photosystem IIROS Reactive oxygen speciesSPION Superparamagnetic iron oxide nanoparticlesTEM Transmission electron microscope
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was performed at the Department of Chem-istry (UQAM) The authors acknowledge the financial sup-port provided by both the Faculty of Sciences and theDepartment of Chemistry (UQAM) The authors wish tothank L Ben Taherc L Samia Smiric and H Abdelmelekbfor providing nanopowder of Fe
3O4 Co02Zn08Fe2O4 and
Co05Zn05Fe2O4
References
[1] A Ito M Shinkai H Honda and T Kobayashi ldquoMedicalapplication of functionalized magnetic nanoparticlesrdquo Journalof Bioscience and Bioengineering vol 100 no 1 pp 1ndash11 2005
[2] D L Huber ldquoSynthesis properties and applications of ironnanoparticlesrdquo Small vol 1 no 5 pp 482ndash501 2005
[3] N Singh G J S Jenkins R Asadi and S H Doak ldquoPoten-tial toxicity of superparamagnetic iron oxide nanoparticles(SPION)rdquo Nano Reviews vol 1 Article ID 5358 2010
[4] I Bhatt and B N Tripathi ldquoInteraction of engineered nanopar-ticles with various components of the environment and possiblestrategies for their risk assessmentrdquo Chemosphere vol 82 no 3pp 308ndash317 2011
[5] MDelay and FH Frimmel ldquoNanoparticles in aquatic systemsrdquoAnalytical and Bioanalytical Chemistry vol 402 no 2 pp 583ndash592 2012
[6] E N Taylor K M Kummer N G Durmus K Leuba K MTarquinio and T J Webster ldquoSuperparamagnetic iron oxidenanoparticles (SPION) for the treatment of antibiotic-resistantbiofilmsrdquo Small vol 8 pp 3016ndash3027 2012
[7] H Zhu J Han J Q Xiao and Y Jin ldquoUptake translocationand accumulation of manufactured iron oxide nanoparticles bypumpkin plantsrdquo Journal of Environmental Monitoring vol 10no 6 pp 713ndash717 2008
[8] Y KMushtaq ldquoEffect of nanoscale Fe3O4 TiO2and carbon par-
ticles on cucumber seed germinationrdquo Journal of EnvironmentalScience and Health A vol 46 no 14 pp 1732ndash1735 2011
[9] X Chen X Zhu R Li H Yao Z Lu and X Yang ldquoPhoto-synthetic toxicity and oxidative damage induced by nano-Fe
3O4
on Chlorella vulgaris in aquatic environmentrdquo Open Journal ofEcology vol 2 pp 21ndash28 2012
[10] R Rippka J Deruelles and J B Waterbury ldquoGeneric assign-ments strain histories and properties of pure cultures ofcyanobacteriardquo Journal of General Microbiology vol 111 no 1pp 1ndash61 1979
[11] H Basti L Ben Tahar L S Smiri et al ldquoCatechol derivatives-coated Fe
3O4and 120574-Fe
2O3nanoparticles as potential MRI
contrast agentsrdquo Journal of Colloid and Interface Science vol 341no 2 pp 248ndash254 2010
[12] W A Hoffmann and H Poorter ldquoAvoiding bias in calculationsof relative growth raterdquo Annals of Botany vol 90 no 1 pp 37ndash42 2002
[13] I B Gerber and I A Dubery ldquoFluorescence microplate assayfor the detection of oxidative burst products in tobacco cellsuspensions using 2101584071015840-dichlorofluoresceinrdquo Methods in CellScience vol 25 no 3-4 pp 115ndash122 2004
[14] P Mayer R Cuhel and N Nyholm ldquoA simple in vitro fluo-rescence method for biomass measurements in algal growthinhibition testsrdquo Water Research vol 31 no 10 pp 2525ndash25311997
[15] A R Wellburn ldquoThe spectral determination of chlorophylls aand b as well as total carotenoids using various solvents withspectrophotometers of different resolutionrdquo Journal of PlantPhysiology vol 144 no 3 pp 307ndash313 1994
[16] L Force C Critchley and J J S Van Rensen ldquoNew fluorescenceparameters for monitoring photosynthesis in plants 1 the effectof illumination on the fluorescence parameters of the JIP-testrdquoPhotosynthesis Research vol 78 no 1 pp 17ndash33 2003
[17] R J Strasser A Srivastava and M Tsimilli-Michael ldquoAnalysisof the chlorophyll a fluorescence transientrdquo in Chlorophyllfluorescence A signature of Photosynthesis Advances in Photo-synthesis and respiration G G Papageorgiou Ed pp 321ndash362Kluwer Academic Dodrecht The Netherlands 2004
[18] N M Franklin N J Rogers S C Apte G E Batley G EGadd and P S Casey ldquoComparative toxicity of nanopartic-ulate ZnO bulk ZnO and ZnCl
2to a freshwater microalga
(Pseudokirchneriella subcapitata) the importance of particle
BioMed Research International 11
solubilityrdquoEnvironmental Science andTechnology vol 41 no 24pp 8484ndash8490 2007
[19] V Aruoja H-C Dubourguier K Kasemets and A KahruldquoToxicity of nanoparticles of CuO ZnO and TiO
2tomicroalgae
Pseudokirchneriella subcapitatardquo Science of the Total Environ-ment vol 407 no 4 pp 1461ndash1468 2009
[20] A M Studer L K Limbach L Van Duc et al ldquoNanoparticlecytotoxicity depends on intracellular solubility comparison ofstabilized copper metal and degradable copper oxide nanopar-ticlesrdquo Toxicology Letters vol 197 no 3 pp 169ndash174 2010
[21] S Lin P Bhattacharya N C Rajapakse D E Brune and P CKe ldquoEffects of quantum dots adsorption on algal photosynthe-sisrdquo Journal of Physical Chemistry C vol 113 no 25 pp 10962ndash10966 2009
[22] T Xia M Kovochich J Brant et al ldquoComparison of theabilities of ambient and manufactured nanoparticles to inducecellular toxicity according to an oxidative stress paradigmrdquoNano Letters vol 6 no 8 pp 1794ndash1807 2006
[23] X Hu S Cook PWang andH-M Hwang ldquoIn vitro evaluationof cytotoxicity of engineeredmetal oxide nanoparticlesrdquo Scienceof the Total Environment vol 407 no 8 pp 3070ndash3072 2009
[24] W J Stark ldquoNanoparticles in biological systemsrdquo AngewandteChemiemdashInternational Edition vol 50 no 6 pp 1242ndash12582011
[25] F Q Schafer S Y Qian and G R Buettner ldquoIron and freeradical oxidations in cell membranesrdquo Cellular and MolecularBiology vol 46 no 3 pp 657ndash662 2000
[26] D Dewez N A Ali F Perreault and R Popovic ldquoRapidchlorophyll a fluorescence transient of Lemna gibba leaf as anindication of light and hydroxylamine effect on photosystem IIactivityrdquo Photochemical and Photobiological Sciences vol 6 no5 pp 532ndash538 2007
[27] D Dewez N Boucher F Bellemare and R Popovic ldquoUse ofdifferent fluorometric systems in the determination of fluo-rescence parameters from spinach thylakoid membranes beingexposed to atrazine and copperrdquo Toxicological and Environmen-tal Chemistry vol 89 no 4 pp 655ndash664 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 BioMed Research International
ABSRC
0 100 200 300 4000
50
100
150
200Pa
ram
eter
s val
ue (
)
[SPION] (120583g mLminus1)
(a)
0 100 200 300 4000
50
100
150
200
Para
met
ers v
alue
()
VJ
[SPION] (120583g mLminus1)
(b)
0
20
40
60
80
100
120
SPION-1SPION-2SPION-3
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)
FVFM
(c)
PI
0
20
40
60
80
100
120
Para
met
ers v
alue
()
0 100 200 300 400
[SPION] (120583g mLminus1)SPION-1SPION-2SPION-3
(d)
Figure 8 Change of different photosynthetic-based fluorescence parameters for algal cells of Chlorella vulgaris exposed during 72 h todifferent concentrations of SPION-1 (Fe
3O4) SPION-2 (Co
02Zn08Fe2O4) and SPION-3 (Co
05Zn05Fe2O4) Maximal PSII quantum yield
119865119881119865119872 ratio between the number of active PSII reaction centers and light harvesting Chl antenna size ABSRC relative variable fluorescence
yield at 119869 transient 119881119869 performance index of PSII photochemical activity PI
35 Production of Reactive Oxygen Species Related to Cell Via-bility The formation of ROS per viable cells was determinedfor C vulgaris exposed during 72 h to SPION (Figure 9)Under these conditions cell viability decreased significantlycompared to control from 50 120583gmLminus1 of SPION indicatingan induction of cellular oxidative stress The productionof ROS per viable cells increased significantly comparedto control which was dependent on SPION species andconcentration
4 Discussion
41 Toxicity of SPION-1 SPION-2 and SPION-3 In thisstudy toxic effects of SPION-1 -2 and -3 were investigatedon algal cells of C vulgaris which were caused by thedeterioration of photochemical activities of photosynthesisthe induction of oxidative stress and the inhibition of cell
division rate This complex cellular alteration was dependenton SPION chemical composition and its concentration insolution Based on our results it is evident that the bioac-cumulation of free Fe Co and Zn from the soluble fractionwas contributing to the toxicity impact in algal cells It waspreviously proposed that the release of free metal ions frommetallic nanoparticle suspensions represented amajor sourceof toxicity for the growth rate of aquatic microorganisms[18 19] However it is difficult to determine if the release offree metal ions from nanoparticles is the only contributionto the toxicity impact in the algal cellular system Indeedsolubilisation of nanoparticles can take place either in themedia or inside the cell which possess an acidic pH environ-ment favorable for particle solubility [20] Therefore anotherhypothesis is that SPION may also contribute directly to thetoxicity impact Recently it was suggested for direct toxicitymechanisms of nanoparticles to induce a direct alteration of
BioMed Research International 9
0
50
100Cell viability
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(a)
ROS
0
50
100
150
200
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(b)
0
100
200
300
400
500
600
700
0 100 200 300 400
ROScell viability
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(c)
Figure 9 Change in the production of reactive oxygen species (ROS) cellular viability and the ratio ROSviable cells for Chlorella vulgarisexposed during 72 h to different concentrations of SPION-1 (⧫ diamond) SPION-2 (◼ square) and SPION-3 (998771 triangle)
cellular exchanges with the media due to particles bindingon cell membrane [21] It was also shown for bioaccumulatedFe3O4nanoparticles to cause direct toxic effects inside plant
cells [8]Moreover the increase of production of reactive oxygen
species per cell viabilitywas related to the increasing exposureconcentration of SPION indicating that the induction ofcellular oxidative stress was caused by the bioaccumulationof Fe Co and Zn Indeed it was shown in previous stud-ies that metallic nanoparticles can cause the formation ofROS via the Fenton reaction or the disruption of majorphysiological processes [22 23] Also the charged metallicsurface of nanoparticles can trigger the formation of ROS bycatalytic reduction of oxygen into superoxide anion leadingto oxidative damage into proteins lipids nucleic acids andpigments [24]
When algal cells of C vulgaris were exposed during 72 hto different concentrations of SPION suspensions the relativecellular division rate was strongly inhibited for SPION-1compared to SPION-2 and SPION-3 Under these treatmentconditions the production of ROS per viable cells was thehighest for SPION-1 Indeed Fe is well known to generate
in cell free radical oxidations causing lipid peroxidation[25] Furthermore the change of fluorescence parameter PIindicated a stronger inhibition of photosynthesis for SPION-1Therefore according to the change of these biomarkers thetoxicity impact in algal cells of C vulgaris was the strongestfor SPION-1 suspensions
42 Significance of Biochemical Biomarkers In this study twodifferent biochemical biomarkers were used to characterizethe toxicity of SPION the production of ROS per viablecells and the photosynthetic-based fluorescence parametersThe change of parameter ROScell viability permitted deter-mining the potential source of cellular toxicity causing theinhibition of cell division rate On the other hand fluores-cence parameters related to PSII photochemical reactionshave been shown to be sensitive biomarkers of SPION toxicityto C vulgaris cells Indeed the Chl 119886 fluorescence emissionis demonstrated to be related to water-splitting system func-tions at PSII reaction center and to oxidoreduction statesof electron transport carriers [26 27] Therefore our resultsshowed evidence for SPION to cause inhibitory effects on thePSII water-splitting system and the photoactivation of PSII
10 BioMed Research International
reaction centers In comparison to other fluorescence indica-tors the performance index of PSII activity representing anintegrative indicator of all PSII photochemical reactions [17]was the most sensitive biomarker of the deterioration of PSIIfunctions caused by SPION toxicity
5 Conclusion
Nowadays a large quantity of metallic nanoparticles isproduced and their toxic potential as hazardous contami-nants in aquatic environment needs to be investigated inorder to develop specific bioassays for nanoparticles toxicityassessment In this regard microalgae represent sensitiveorganisms to be used in bioassay toxicity testing for theassessment of hazardous materials In this toxicologicalinvestigation we clearly showed the potential source of tox-icity of three SPION suspensions having different chemicalcompositions The algal model C vulgaris demonstrated tobe a valuable bioindicator of SPION cellular toxicity whichwas indicated by the deterioration of photochemical activitiesof photosynthesis the induction of oxidative stress and theinhibition of cell division rateTherefore this work permittedcharacterising the cellular toxicity impact of these SPIONwith different biomarkers A good understanding of thesetoxicological interactions will permit better understandingthe risk of SPION toxicity for aquatic organisms
Abbreviations
Chl ChlorophyllNP NanoparticlesPEA Plant Efficiency AnalyzerPSII Photosystem IIROS Reactive oxygen speciesSPION Superparamagnetic iron oxide nanoparticlesTEM Transmission electron microscope
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was performed at the Department of Chem-istry (UQAM) The authors acknowledge the financial sup-port provided by both the Faculty of Sciences and theDepartment of Chemistry (UQAM) The authors wish tothank L Ben Taherc L Samia Smiric and H Abdelmelekbfor providing nanopowder of Fe
3O4 Co02Zn08Fe2O4 and
Co05Zn05Fe2O4
References
[1] A Ito M Shinkai H Honda and T Kobayashi ldquoMedicalapplication of functionalized magnetic nanoparticlesrdquo Journalof Bioscience and Bioengineering vol 100 no 1 pp 1ndash11 2005
[2] D L Huber ldquoSynthesis properties and applications of ironnanoparticlesrdquo Small vol 1 no 5 pp 482ndash501 2005
[3] N Singh G J S Jenkins R Asadi and S H Doak ldquoPoten-tial toxicity of superparamagnetic iron oxide nanoparticles(SPION)rdquo Nano Reviews vol 1 Article ID 5358 2010
[4] I Bhatt and B N Tripathi ldquoInteraction of engineered nanopar-ticles with various components of the environment and possiblestrategies for their risk assessmentrdquo Chemosphere vol 82 no 3pp 308ndash317 2011
[5] MDelay and FH Frimmel ldquoNanoparticles in aquatic systemsrdquoAnalytical and Bioanalytical Chemistry vol 402 no 2 pp 583ndash592 2012
[6] E N Taylor K M Kummer N G Durmus K Leuba K MTarquinio and T J Webster ldquoSuperparamagnetic iron oxidenanoparticles (SPION) for the treatment of antibiotic-resistantbiofilmsrdquo Small vol 8 pp 3016ndash3027 2012
[7] H Zhu J Han J Q Xiao and Y Jin ldquoUptake translocationand accumulation of manufactured iron oxide nanoparticles bypumpkin plantsrdquo Journal of Environmental Monitoring vol 10no 6 pp 713ndash717 2008
[8] Y KMushtaq ldquoEffect of nanoscale Fe3O4 TiO2and carbon par-
ticles on cucumber seed germinationrdquo Journal of EnvironmentalScience and Health A vol 46 no 14 pp 1732ndash1735 2011
[9] X Chen X Zhu R Li H Yao Z Lu and X Yang ldquoPhoto-synthetic toxicity and oxidative damage induced by nano-Fe
3O4
on Chlorella vulgaris in aquatic environmentrdquo Open Journal ofEcology vol 2 pp 21ndash28 2012
[10] R Rippka J Deruelles and J B Waterbury ldquoGeneric assign-ments strain histories and properties of pure cultures ofcyanobacteriardquo Journal of General Microbiology vol 111 no 1pp 1ndash61 1979
[11] H Basti L Ben Tahar L S Smiri et al ldquoCatechol derivatives-coated Fe
3O4and 120574-Fe
2O3nanoparticles as potential MRI
contrast agentsrdquo Journal of Colloid and Interface Science vol 341no 2 pp 248ndash254 2010
[12] W A Hoffmann and H Poorter ldquoAvoiding bias in calculationsof relative growth raterdquo Annals of Botany vol 90 no 1 pp 37ndash42 2002
[13] I B Gerber and I A Dubery ldquoFluorescence microplate assayfor the detection of oxidative burst products in tobacco cellsuspensions using 2101584071015840-dichlorofluoresceinrdquo Methods in CellScience vol 25 no 3-4 pp 115ndash122 2004
[14] P Mayer R Cuhel and N Nyholm ldquoA simple in vitro fluo-rescence method for biomass measurements in algal growthinhibition testsrdquo Water Research vol 31 no 10 pp 2525ndash25311997
[15] A R Wellburn ldquoThe spectral determination of chlorophylls aand b as well as total carotenoids using various solvents withspectrophotometers of different resolutionrdquo Journal of PlantPhysiology vol 144 no 3 pp 307ndash313 1994
[16] L Force C Critchley and J J S Van Rensen ldquoNew fluorescenceparameters for monitoring photosynthesis in plants 1 the effectof illumination on the fluorescence parameters of the JIP-testrdquoPhotosynthesis Research vol 78 no 1 pp 17ndash33 2003
[17] R J Strasser A Srivastava and M Tsimilli-Michael ldquoAnalysisof the chlorophyll a fluorescence transientrdquo in Chlorophyllfluorescence A signature of Photosynthesis Advances in Photo-synthesis and respiration G G Papageorgiou Ed pp 321ndash362Kluwer Academic Dodrecht The Netherlands 2004
[18] N M Franklin N J Rogers S C Apte G E Batley G EGadd and P S Casey ldquoComparative toxicity of nanopartic-ulate ZnO bulk ZnO and ZnCl
2to a freshwater microalga
(Pseudokirchneriella subcapitata) the importance of particle
BioMed Research International 11
solubilityrdquoEnvironmental Science andTechnology vol 41 no 24pp 8484ndash8490 2007
[19] V Aruoja H-C Dubourguier K Kasemets and A KahruldquoToxicity of nanoparticles of CuO ZnO and TiO
2tomicroalgae
Pseudokirchneriella subcapitatardquo Science of the Total Environ-ment vol 407 no 4 pp 1461ndash1468 2009
[20] A M Studer L K Limbach L Van Duc et al ldquoNanoparticlecytotoxicity depends on intracellular solubility comparison ofstabilized copper metal and degradable copper oxide nanopar-ticlesrdquo Toxicology Letters vol 197 no 3 pp 169ndash174 2010
[21] S Lin P Bhattacharya N C Rajapakse D E Brune and P CKe ldquoEffects of quantum dots adsorption on algal photosynthe-sisrdquo Journal of Physical Chemistry C vol 113 no 25 pp 10962ndash10966 2009
[22] T Xia M Kovochich J Brant et al ldquoComparison of theabilities of ambient and manufactured nanoparticles to inducecellular toxicity according to an oxidative stress paradigmrdquoNano Letters vol 6 no 8 pp 1794ndash1807 2006
[23] X Hu S Cook PWang andH-M Hwang ldquoIn vitro evaluationof cytotoxicity of engineeredmetal oxide nanoparticlesrdquo Scienceof the Total Environment vol 407 no 8 pp 3070ndash3072 2009
[24] W J Stark ldquoNanoparticles in biological systemsrdquo AngewandteChemiemdashInternational Edition vol 50 no 6 pp 1242ndash12582011
[25] F Q Schafer S Y Qian and G R Buettner ldquoIron and freeradical oxidations in cell membranesrdquo Cellular and MolecularBiology vol 46 no 3 pp 657ndash662 2000
[26] D Dewez N A Ali F Perreault and R Popovic ldquoRapidchlorophyll a fluorescence transient of Lemna gibba leaf as anindication of light and hydroxylamine effect on photosystem IIactivityrdquo Photochemical and Photobiological Sciences vol 6 no5 pp 532ndash538 2007
[27] D Dewez N Boucher F Bellemare and R Popovic ldquoUse ofdifferent fluorometric systems in the determination of fluo-rescence parameters from spinach thylakoid membranes beingexposed to atrazine and copperrdquo Toxicological and Environmen-tal Chemistry vol 89 no 4 pp 655ndash664 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 9
0
50
100Cell viability
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(a)
ROS
0
50
100
150
200
0 100 200 300 400
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(b)
0
100
200
300
400
500
600
700
0 100 200 300 400
ROScell viability
[SPION] (120583g mLminus1)
Para
met
ers v
alue
()
(c)
Figure 9 Change in the production of reactive oxygen species (ROS) cellular viability and the ratio ROSviable cells for Chlorella vulgarisexposed during 72 h to different concentrations of SPION-1 (⧫ diamond) SPION-2 (◼ square) and SPION-3 (998771 triangle)
cellular exchanges with the media due to particles bindingon cell membrane [21] It was also shown for bioaccumulatedFe3O4nanoparticles to cause direct toxic effects inside plant
cells [8]Moreover the increase of production of reactive oxygen
species per cell viabilitywas related to the increasing exposureconcentration of SPION indicating that the induction ofcellular oxidative stress was caused by the bioaccumulationof Fe Co and Zn Indeed it was shown in previous stud-ies that metallic nanoparticles can cause the formation ofROS via the Fenton reaction or the disruption of majorphysiological processes [22 23] Also the charged metallicsurface of nanoparticles can trigger the formation of ROS bycatalytic reduction of oxygen into superoxide anion leadingto oxidative damage into proteins lipids nucleic acids andpigments [24]
When algal cells of C vulgaris were exposed during 72 hto different concentrations of SPION suspensions the relativecellular division rate was strongly inhibited for SPION-1compared to SPION-2 and SPION-3 Under these treatmentconditions the production of ROS per viable cells was thehighest for SPION-1 Indeed Fe is well known to generate
in cell free radical oxidations causing lipid peroxidation[25] Furthermore the change of fluorescence parameter PIindicated a stronger inhibition of photosynthesis for SPION-1Therefore according to the change of these biomarkers thetoxicity impact in algal cells of C vulgaris was the strongestfor SPION-1 suspensions
42 Significance of Biochemical Biomarkers In this study twodifferent biochemical biomarkers were used to characterizethe toxicity of SPION the production of ROS per viablecells and the photosynthetic-based fluorescence parametersThe change of parameter ROScell viability permitted deter-mining the potential source of cellular toxicity causing theinhibition of cell division rate On the other hand fluores-cence parameters related to PSII photochemical reactionshave been shown to be sensitive biomarkers of SPION toxicityto C vulgaris cells Indeed the Chl 119886 fluorescence emissionis demonstrated to be related to water-splitting system func-tions at PSII reaction center and to oxidoreduction statesof electron transport carriers [26 27] Therefore our resultsshowed evidence for SPION to cause inhibitory effects on thePSII water-splitting system and the photoactivation of PSII
10 BioMed Research International
reaction centers In comparison to other fluorescence indica-tors the performance index of PSII activity representing anintegrative indicator of all PSII photochemical reactions [17]was the most sensitive biomarker of the deterioration of PSIIfunctions caused by SPION toxicity
5 Conclusion
Nowadays a large quantity of metallic nanoparticles isproduced and their toxic potential as hazardous contami-nants in aquatic environment needs to be investigated inorder to develop specific bioassays for nanoparticles toxicityassessment In this regard microalgae represent sensitiveorganisms to be used in bioassay toxicity testing for theassessment of hazardous materials In this toxicologicalinvestigation we clearly showed the potential source of tox-icity of three SPION suspensions having different chemicalcompositions The algal model C vulgaris demonstrated tobe a valuable bioindicator of SPION cellular toxicity whichwas indicated by the deterioration of photochemical activitiesof photosynthesis the induction of oxidative stress and theinhibition of cell division rateTherefore this work permittedcharacterising the cellular toxicity impact of these SPIONwith different biomarkers A good understanding of thesetoxicological interactions will permit better understandingthe risk of SPION toxicity for aquatic organisms
Abbreviations
Chl ChlorophyllNP NanoparticlesPEA Plant Efficiency AnalyzerPSII Photosystem IIROS Reactive oxygen speciesSPION Superparamagnetic iron oxide nanoparticlesTEM Transmission electron microscope
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was performed at the Department of Chem-istry (UQAM) The authors acknowledge the financial sup-port provided by both the Faculty of Sciences and theDepartment of Chemistry (UQAM) The authors wish tothank L Ben Taherc L Samia Smiric and H Abdelmelekbfor providing nanopowder of Fe
3O4 Co02Zn08Fe2O4 and
Co05Zn05Fe2O4
References
[1] A Ito M Shinkai H Honda and T Kobayashi ldquoMedicalapplication of functionalized magnetic nanoparticlesrdquo Journalof Bioscience and Bioengineering vol 100 no 1 pp 1ndash11 2005
[2] D L Huber ldquoSynthesis properties and applications of ironnanoparticlesrdquo Small vol 1 no 5 pp 482ndash501 2005
[3] N Singh G J S Jenkins R Asadi and S H Doak ldquoPoten-tial toxicity of superparamagnetic iron oxide nanoparticles(SPION)rdquo Nano Reviews vol 1 Article ID 5358 2010
[4] I Bhatt and B N Tripathi ldquoInteraction of engineered nanopar-ticles with various components of the environment and possiblestrategies for their risk assessmentrdquo Chemosphere vol 82 no 3pp 308ndash317 2011
[5] MDelay and FH Frimmel ldquoNanoparticles in aquatic systemsrdquoAnalytical and Bioanalytical Chemistry vol 402 no 2 pp 583ndash592 2012
[6] E N Taylor K M Kummer N G Durmus K Leuba K MTarquinio and T J Webster ldquoSuperparamagnetic iron oxidenanoparticles (SPION) for the treatment of antibiotic-resistantbiofilmsrdquo Small vol 8 pp 3016ndash3027 2012
[7] H Zhu J Han J Q Xiao and Y Jin ldquoUptake translocationand accumulation of manufactured iron oxide nanoparticles bypumpkin plantsrdquo Journal of Environmental Monitoring vol 10no 6 pp 713ndash717 2008
[8] Y KMushtaq ldquoEffect of nanoscale Fe3O4 TiO2and carbon par-
ticles on cucumber seed germinationrdquo Journal of EnvironmentalScience and Health A vol 46 no 14 pp 1732ndash1735 2011
[9] X Chen X Zhu R Li H Yao Z Lu and X Yang ldquoPhoto-synthetic toxicity and oxidative damage induced by nano-Fe
3O4
on Chlorella vulgaris in aquatic environmentrdquo Open Journal ofEcology vol 2 pp 21ndash28 2012
[10] R Rippka J Deruelles and J B Waterbury ldquoGeneric assign-ments strain histories and properties of pure cultures ofcyanobacteriardquo Journal of General Microbiology vol 111 no 1pp 1ndash61 1979
[11] H Basti L Ben Tahar L S Smiri et al ldquoCatechol derivatives-coated Fe
3O4and 120574-Fe
2O3nanoparticles as potential MRI
contrast agentsrdquo Journal of Colloid and Interface Science vol 341no 2 pp 248ndash254 2010
[12] W A Hoffmann and H Poorter ldquoAvoiding bias in calculationsof relative growth raterdquo Annals of Botany vol 90 no 1 pp 37ndash42 2002
[13] I B Gerber and I A Dubery ldquoFluorescence microplate assayfor the detection of oxidative burst products in tobacco cellsuspensions using 2101584071015840-dichlorofluoresceinrdquo Methods in CellScience vol 25 no 3-4 pp 115ndash122 2004
[14] P Mayer R Cuhel and N Nyholm ldquoA simple in vitro fluo-rescence method for biomass measurements in algal growthinhibition testsrdquo Water Research vol 31 no 10 pp 2525ndash25311997
[15] A R Wellburn ldquoThe spectral determination of chlorophylls aand b as well as total carotenoids using various solvents withspectrophotometers of different resolutionrdquo Journal of PlantPhysiology vol 144 no 3 pp 307ndash313 1994
[16] L Force C Critchley and J J S Van Rensen ldquoNew fluorescenceparameters for monitoring photosynthesis in plants 1 the effectof illumination on the fluorescence parameters of the JIP-testrdquoPhotosynthesis Research vol 78 no 1 pp 17ndash33 2003
[17] R J Strasser A Srivastava and M Tsimilli-Michael ldquoAnalysisof the chlorophyll a fluorescence transientrdquo in Chlorophyllfluorescence A signature of Photosynthesis Advances in Photo-synthesis and respiration G G Papageorgiou Ed pp 321ndash362Kluwer Academic Dodrecht The Netherlands 2004
[18] N M Franklin N J Rogers S C Apte G E Batley G EGadd and P S Casey ldquoComparative toxicity of nanopartic-ulate ZnO bulk ZnO and ZnCl
2to a freshwater microalga
(Pseudokirchneriella subcapitata) the importance of particle
BioMed Research International 11
solubilityrdquoEnvironmental Science andTechnology vol 41 no 24pp 8484ndash8490 2007
[19] V Aruoja H-C Dubourguier K Kasemets and A KahruldquoToxicity of nanoparticles of CuO ZnO and TiO
2tomicroalgae
Pseudokirchneriella subcapitatardquo Science of the Total Environ-ment vol 407 no 4 pp 1461ndash1468 2009
[20] A M Studer L K Limbach L Van Duc et al ldquoNanoparticlecytotoxicity depends on intracellular solubility comparison ofstabilized copper metal and degradable copper oxide nanopar-ticlesrdquo Toxicology Letters vol 197 no 3 pp 169ndash174 2010
[21] S Lin P Bhattacharya N C Rajapakse D E Brune and P CKe ldquoEffects of quantum dots adsorption on algal photosynthe-sisrdquo Journal of Physical Chemistry C vol 113 no 25 pp 10962ndash10966 2009
[22] T Xia M Kovochich J Brant et al ldquoComparison of theabilities of ambient and manufactured nanoparticles to inducecellular toxicity according to an oxidative stress paradigmrdquoNano Letters vol 6 no 8 pp 1794ndash1807 2006
[23] X Hu S Cook PWang andH-M Hwang ldquoIn vitro evaluationof cytotoxicity of engineeredmetal oxide nanoparticlesrdquo Scienceof the Total Environment vol 407 no 8 pp 3070ndash3072 2009
[24] W J Stark ldquoNanoparticles in biological systemsrdquo AngewandteChemiemdashInternational Edition vol 50 no 6 pp 1242ndash12582011
[25] F Q Schafer S Y Qian and G R Buettner ldquoIron and freeradical oxidations in cell membranesrdquo Cellular and MolecularBiology vol 46 no 3 pp 657ndash662 2000
[26] D Dewez N A Ali F Perreault and R Popovic ldquoRapidchlorophyll a fluorescence transient of Lemna gibba leaf as anindication of light and hydroxylamine effect on photosystem IIactivityrdquo Photochemical and Photobiological Sciences vol 6 no5 pp 532ndash538 2007
[27] D Dewez N Boucher F Bellemare and R Popovic ldquoUse ofdifferent fluorometric systems in the determination of fluo-rescence parameters from spinach thylakoid membranes beingexposed to atrazine and copperrdquo Toxicological and Environmen-tal Chemistry vol 89 no 4 pp 655ndash664 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

10 BioMed Research International
reaction centers In comparison to other fluorescence indica-tors the performance index of PSII activity representing anintegrative indicator of all PSII photochemical reactions [17]was the most sensitive biomarker of the deterioration of PSIIfunctions caused by SPION toxicity
5 Conclusion
Nowadays a large quantity of metallic nanoparticles isproduced and their toxic potential as hazardous contami-nants in aquatic environment needs to be investigated inorder to develop specific bioassays for nanoparticles toxicityassessment In this regard microalgae represent sensitiveorganisms to be used in bioassay toxicity testing for theassessment of hazardous materials In this toxicologicalinvestigation we clearly showed the potential source of tox-icity of three SPION suspensions having different chemicalcompositions The algal model C vulgaris demonstrated tobe a valuable bioindicator of SPION cellular toxicity whichwas indicated by the deterioration of photochemical activitiesof photosynthesis the induction of oxidative stress and theinhibition of cell division rateTherefore this work permittedcharacterising the cellular toxicity impact of these SPIONwith different biomarkers A good understanding of thesetoxicological interactions will permit better understandingthe risk of SPION toxicity for aquatic organisms
Abbreviations
Chl ChlorophyllNP NanoparticlesPEA Plant Efficiency AnalyzerPSII Photosystem IIROS Reactive oxygen speciesSPION Superparamagnetic iron oxide nanoparticlesTEM Transmission electron microscope
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was performed at the Department of Chem-istry (UQAM) The authors acknowledge the financial sup-port provided by both the Faculty of Sciences and theDepartment of Chemistry (UQAM) The authors wish tothank L Ben Taherc L Samia Smiric and H Abdelmelekbfor providing nanopowder of Fe
3O4 Co02Zn08Fe2O4 and
Co05Zn05Fe2O4
References
[1] A Ito M Shinkai H Honda and T Kobayashi ldquoMedicalapplication of functionalized magnetic nanoparticlesrdquo Journalof Bioscience and Bioengineering vol 100 no 1 pp 1ndash11 2005
[2] D L Huber ldquoSynthesis properties and applications of ironnanoparticlesrdquo Small vol 1 no 5 pp 482ndash501 2005
[3] N Singh G J S Jenkins R Asadi and S H Doak ldquoPoten-tial toxicity of superparamagnetic iron oxide nanoparticles(SPION)rdquo Nano Reviews vol 1 Article ID 5358 2010
[4] I Bhatt and B N Tripathi ldquoInteraction of engineered nanopar-ticles with various components of the environment and possiblestrategies for their risk assessmentrdquo Chemosphere vol 82 no 3pp 308ndash317 2011
[5] MDelay and FH Frimmel ldquoNanoparticles in aquatic systemsrdquoAnalytical and Bioanalytical Chemistry vol 402 no 2 pp 583ndash592 2012
[6] E N Taylor K M Kummer N G Durmus K Leuba K MTarquinio and T J Webster ldquoSuperparamagnetic iron oxidenanoparticles (SPION) for the treatment of antibiotic-resistantbiofilmsrdquo Small vol 8 pp 3016ndash3027 2012
[7] H Zhu J Han J Q Xiao and Y Jin ldquoUptake translocationand accumulation of manufactured iron oxide nanoparticles bypumpkin plantsrdquo Journal of Environmental Monitoring vol 10no 6 pp 713ndash717 2008
[8] Y KMushtaq ldquoEffect of nanoscale Fe3O4 TiO2and carbon par-
ticles on cucumber seed germinationrdquo Journal of EnvironmentalScience and Health A vol 46 no 14 pp 1732ndash1735 2011
[9] X Chen X Zhu R Li H Yao Z Lu and X Yang ldquoPhoto-synthetic toxicity and oxidative damage induced by nano-Fe
3O4
on Chlorella vulgaris in aquatic environmentrdquo Open Journal ofEcology vol 2 pp 21ndash28 2012
[10] R Rippka J Deruelles and J B Waterbury ldquoGeneric assign-ments strain histories and properties of pure cultures ofcyanobacteriardquo Journal of General Microbiology vol 111 no 1pp 1ndash61 1979
[11] H Basti L Ben Tahar L S Smiri et al ldquoCatechol derivatives-coated Fe
3O4and 120574-Fe
2O3nanoparticles as potential MRI
contrast agentsrdquo Journal of Colloid and Interface Science vol 341no 2 pp 248ndash254 2010
[12] W A Hoffmann and H Poorter ldquoAvoiding bias in calculationsof relative growth raterdquo Annals of Botany vol 90 no 1 pp 37ndash42 2002
[13] I B Gerber and I A Dubery ldquoFluorescence microplate assayfor the detection of oxidative burst products in tobacco cellsuspensions using 2101584071015840-dichlorofluoresceinrdquo Methods in CellScience vol 25 no 3-4 pp 115ndash122 2004
[14] P Mayer R Cuhel and N Nyholm ldquoA simple in vitro fluo-rescence method for biomass measurements in algal growthinhibition testsrdquo Water Research vol 31 no 10 pp 2525ndash25311997
[15] A R Wellburn ldquoThe spectral determination of chlorophylls aand b as well as total carotenoids using various solvents withspectrophotometers of different resolutionrdquo Journal of PlantPhysiology vol 144 no 3 pp 307ndash313 1994
[16] L Force C Critchley and J J S Van Rensen ldquoNew fluorescenceparameters for monitoring photosynthesis in plants 1 the effectof illumination on the fluorescence parameters of the JIP-testrdquoPhotosynthesis Research vol 78 no 1 pp 17ndash33 2003
[17] R J Strasser A Srivastava and M Tsimilli-Michael ldquoAnalysisof the chlorophyll a fluorescence transientrdquo in Chlorophyllfluorescence A signature of Photosynthesis Advances in Photo-synthesis and respiration G G Papageorgiou Ed pp 321ndash362Kluwer Academic Dodrecht The Netherlands 2004
[18] N M Franklin N J Rogers S C Apte G E Batley G EGadd and P S Casey ldquoComparative toxicity of nanopartic-ulate ZnO bulk ZnO and ZnCl
2to a freshwater microalga
(Pseudokirchneriella subcapitata) the importance of particle
BioMed Research International 11
solubilityrdquoEnvironmental Science andTechnology vol 41 no 24pp 8484ndash8490 2007
[19] V Aruoja H-C Dubourguier K Kasemets and A KahruldquoToxicity of nanoparticles of CuO ZnO and TiO
2tomicroalgae
Pseudokirchneriella subcapitatardquo Science of the Total Environ-ment vol 407 no 4 pp 1461ndash1468 2009
[20] A M Studer L K Limbach L Van Duc et al ldquoNanoparticlecytotoxicity depends on intracellular solubility comparison ofstabilized copper metal and degradable copper oxide nanopar-ticlesrdquo Toxicology Letters vol 197 no 3 pp 169ndash174 2010
[21] S Lin P Bhattacharya N C Rajapakse D E Brune and P CKe ldquoEffects of quantum dots adsorption on algal photosynthe-sisrdquo Journal of Physical Chemistry C vol 113 no 25 pp 10962ndash10966 2009
[22] T Xia M Kovochich J Brant et al ldquoComparison of theabilities of ambient and manufactured nanoparticles to inducecellular toxicity according to an oxidative stress paradigmrdquoNano Letters vol 6 no 8 pp 1794ndash1807 2006
[23] X Hu S Cook PWang andH-M Hwang ldquoIn vitro evaluationof cytotoxicity of engineeredmetal oxide nanoparticlesrdquo Scienceof the Total Environment vol 407 no 8 pp 3070ndash3072 2009
[24] W J Stark ldquoNanoparticles in biological systemsrdquo AngewandteChemiemdashInternational Edition vol 50 no 6 pp 1242ndash12582011
[25] F Q Schafer S Y Qian and G R Buettner ldquoIron and freeradical oxidations in cell membranesrdquo Cellular and MolecularBiology vol 46 no 3 pp 657ndash662 2000
[26] D Dewez N A Ali F Perreault and R Popovic ldquoRapidchlorophyll a fluorescence transient of Lemna gibba leaf as anindication of light and hydroxylamine effect on photosystem IIactivityrdquo Photochemical and Photobiological Sciences vol 6 no5 pp 532ndash538 2007
[27] D Dewez N Boucher F Bellemare and R Popovic ldquoUse ofdifferent fluorometric systems in the determination of fluo-rescence parameters from spinach thylakoid membranes beingexposed to atrazine and copperrdquo Toxicological and Environmen-tal Chemistry vol 89 no 4 pp 655ndash664 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 11
solubilityrdquoEnvironmental Science andTechnology vol 41 no 24pp 8484ndash8490 2007
[19] V Aruoja H-C Dubourguier K Kasemets and A KahruldquoToxicity of nanoparticles of CuO ZnO and TiO
2tomicroalgae
Pseudokirchneriella subcapitatardquo Science of the Total Environ-ment vol 407 no 4 pp 1461ndash1468 2009
[20] A M Studer L K Limbach L Van Duc et al ldquoNanoparticlecytotoxicity depends on intracellular solubility comparison ofstabilized copper metal and degradable copper oxide nanopar-ticlesrdquo Toxicology Letters vol 197 no 3 pp 169ndash174 2010
[21] S Lin P Bhattacharya N C Rajapakse D E Brune and P CKe ldquoEffects of quantum dots adsorption on algal photosynthe-sisrdquo Journal of Physical Chemistry C vol 113 no 25 pp 10962ndash10966 2009
[22] T Xia M Kovochich J Brant et al ldquoComparison of theabilities of ambient and manufactured nanoparticles to inducecellular toxicity according to an oxidative stress paradigmrdquoNano Letters vol 6 no 8 pp 1794ndash1807 2006
[23] X Hu S Cook PWang andH-M Hwang ldquoIn vitro evaluationof cytotoxicity of engineeredmetal oxide nanoparticlesrdquo Scienceof the Total Environment vol 407 no 8 pp 3070ndash3072 2009
[24] W J Stark ldquoNanoparticles in biological systemsrdquo AngewandteChemiemdashInternational Edition vol 50 no 6 pp 1242ndash12582011
[25] F Q Schafer S Y Qian and G R Buettner ldquoIron and freeradical oxidations in cell membranesrdquo Cellular and MolecularBiology vol 46 no 3 pp 657ndash662 2000
[26] D Dewez N A Ali F Perreault and R Popovic ldquoRapidchlorophyll a fluorescence transient of Lemna gibba leaf as anindication of light and hydroxylamine effect on photosystem IIactivityrdquo Photochemical and Photobiological Sciences vol 6 no5 pp 532ndash538 2007
[27] D Dewez N Boucher F Bellemare and R Popovic ldquoUse ofdifferent fluorometric systems in the determination of fluo-rescence parameters from spinach thylakoid membranes beingexposed to atrazine and copperrdquo Toxicological and Environmen-tal Chemistry vol 89 no 4 pp 655ndash664 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology


















