
Reduction of Luteinzing Hormone Secretion Induced by Long-Term Feed Restriction in Male Rats Is...
7
Journal of Neuroendocrinology, 1999, Vol. 11, 613–619 Reduction of Luteinzing Hormone Secretion Induced by Long-Term Feed Restriction in Male Rats Is Associated with Increased Expression of GABA-Synthesizing Enzymes Without Alterations of GnRH Gene Expression S. Leonhardt, M. Shahab,* H. Luft, W. Wuttke and H. Jarr y Division of Clinical and Experimental Endocrinology, Department of Obstetrics and Gynecology, University of Go ¨ ttingen, Germany. *Department of Biological Sciences, Quaid-i-Azam University, Islamabad, Pakistan. Key words: feed restriction, luteinizing hormone, GnRH gene expression, GABA, glutamate. Abstract In rats, fasting or restriction of feed intake impairs the activity of the hypothalamic gonadotropin- releasing hormone (GnRH) pulse generator which results in reduced luteinizing hormone (LH) secretion. It is still unknown which neurotransmitters are involved in this phenomenon. However, it is known that increased GABA concentrations in the hypothalamus reduce GnRH biosynthesis and release. Therefore, we examined whether 17 days of feed restriction in male rats affected the hypothalamic gene expression of GnRH and the GABA-synthesizing enzymes glutaminase (GLS) and glutamic acid decarboxylase–which exists in two forms, GAD67 and GAD65–in the mammalian brain. Furthermore, the expression of the GnRH receptor (GnRH-R) and the GABA transporter 1 (GAT-1) were investigated. Feed restriction resulted in a 75% reduction in body weight (b.w.) compared to rats fed ad libitum. Serum concentrations of LH and testosterone in the feed restricted group were significantly reduced to #15% of that of rats fed ad libitum, while the FSH concentration remained unchanged. In the mediobasal hypothalamus (MBH) where GnRH is released into the portal vessels, mRNA levels of GAD67 and GLS were increased twofold compared to rats fed ad libitum while no changes were observed in the preoptic area of the hypothalamus (POA) where GnRH is biosynthesised. Neither the expression of preoptic GnRH mRNA nor the expression of GAD65 and of GnRH-R mRNA in both hypothalamic structures was affected by feed restriction. In the anterior pituitary, a significant reduction of the expression of GnRH-R, LH-b and the a subunit was observed in the feed restricted rats, whereas FSH-b mRNA levels remained constant. Thus, feed restriction selectively increased the expression of GABA-synthesizing enzymes in the MBH but did not modify GnRH expression in the POA. However, the reduced expression of the LH-b- and a-subunit and of the GnRH-R in the anterior pituitary indicates that pulsatile GnRH release may have been attenuated or even abolished. We suggest, that enhanced expression of GABA-synthesizing enzymes reflects increased GABAergic neurotransmission and thereby reducing GnRH release from the MBH. In many species, including humans and rats, long periods of rats, are located in the preoptic area (POA) of the anterior hypothalamus, while the axons terminate in the median under nutrition result in low circulating concentrations of luteinizing hormone ( LH ) and gonadal steroids ( 1, 2 ). Proper eminence of the mediobasal hypothalamus (MBH ) (5). A variety of intra- and extra-hypothalamic inputs appear to functioning of the pituitary-gonadal axis depends upon a neuroendocrine control system called the GnRH pulse gener- impair GnRH neuronal activity so that pituitary and gonadal function are reduced or even totally arrested (for review see ator, which causes the phasic release of GnRH and sub- sequent pulsatile gonadotropin release ( 3). The phasic activity (6, 7)). Pulsatile application of GnRH to male rats during periods of reduced feed intake can reverse the decrease of is an intrinsic property of the GnRH neurones (4) which, in Correspondence to: Professor Dr Hubertus Jarry, UFK Go ¨ ttingen Klin. & Exp. Endokrinologie, TL165 Robert-Koch-Strasse 40, 37075 Go ¨ ttingen, Germany (e-mail: [email protected]). © 1999 Blackwell Science Ltd
Transcript of Reduction of Luteinzing Hormone Secretion Induced by Long-Term Feed Restriction in Male Rats Is...

Journal of Neuroendocrinology, 1999, Vol. 11, 613–619
Reduction of Luteinzing Hormone Secretion Induced by Long-TermFeed Restriction in Male Rats Is Associated with IncreasedExpression of GABA-Synthesizing Enzymes Without Alterations ofGnRH Gene Expression
S. Leonhardt, M. Shahab,* H. Luft, W. Wuttke and H. JarryDivision of Clinical and Experimental Endocrinology, Department of Obstetrics and Gynecology, University of Gottingen, Germany.*Department of Biological Sciences, Quaid-i-Azam University, Islamabad, Pakistan.
Key words: feed restriction, luteinizing hormone, GnRH gene expression, GABA, glutamate.
Abstract
In rats, fasting or restriction of feed intake impairs the activity of the hypothalamic gonadotropin-releasing hormone (GnRH) pulse generator which results in reduced luteinizing hormone (LH)secretion. It is still unknown which neurotransmitters are involved in this phenomenon. However, it isknown that increased GABA concentrations in the hypothalamus reduce GnRH biosynthesis andrelease. Therefore, we examined whether 17 days of feed restriction in male rats affected thehypothalamic gene expression of GnRH and the GABA-synthesizing enzymes glutaminase (GLS) andglutamic acid decarboxylase–which exists in two forms, GAD67 and GAD65–in the mammalian brain.Furthermore, the expression of the GnRH receptor (GnRH-R) and the GABA transporter 1 (GAT-1)were investigated. Feed restriction resulted in a 75% reduction in body weight (b.w.) compared torats fed ad libitum. Serum concentrations of LH and testosterone in the feed restricted group weresignificantly reduced to #15% of that of rats fed ad libitum, while the FSH concentration remainedunchanged. In the mediobasal hypothalamus (MBH) where GnRH is released into the portal vessels,mRNA levels of GAD67 and GLS were increased twofold compared to rats fed ad libitum whileno changes were observed in the preoptic area of the hypothalamus (POA) where GnRH isbiosynthesised. Neither the expression of preoptic GnRH mRNA nor the expression of GAD65 and ofGnRH-R mRNA in both hypothalamic structures was affected by feed restriction. In the anteriorpituitary, a significant reduction of the expression of GnRH-R, LH-b and the a subunit was observedin the feed restricted rats, whereas FSH-b mRNA levels remained constant. Thus, feed restrictionselectively increased the expression of GABA-synthesizing enzymes in the MBH but did not modifyGnRH expression in the POA. However, the reduced expression of the LH-b- and a-subunit and ofthe GnRH-R in the anterior pituitary indicates that pulsatile GnRH release may have been attenuatedor even abolished. We suggest, that enhanced expression of GABA-synthesizing enzymes reflectsincreased GABAergic neurotransmission and thereby reducing GnRH release from the MBH.
In many species, including humans and rats, long periods of rats, are located in the preoptic area (POA) of the anteriorhypothalamus, while the axons terminate in the medianunder nutrition result in low circulating concentrations of
luteinizing hormone (LH) and gonadal steroids (1, 2). Proper eminence of the mediobasal hypothalamus (MBH) (5). Avariety of intra- and extra-hypothalamic inputs appear tofunctioning of the pituitary-gonadal axis depends upon a
neuroendocrine control system called the GnRH pulse gener- impair GnRH neuronal activity so that pituitary and gonadalfunction are reduced or even totally arrested (for review seeator, which causes the phasic release of GnRH and sub-
sequent pulsatile gonadotropin release (3). The phasic activity (6, 7)). Pulsatile application of GnRH to male rats duringperiods of reduced feed intake can reverse the decrease ofis an intrinsic property of the GnRH neurones (4) which, in
Correspondence to: Professor Dr Hubertus Jarry, UFK Gottingen Klin. & Exp. Endokrinologie, TL165 Robert-Koch-Strasse 40, 37075Gottingen, Germany (e-mail: [email protected]).
© 1999 Blackwell Science Ltd

614 Reduction of LH secretion and long-term feed restriction
LH and testosterone concentrations as compared to ad libitumfed rats, suggesting that a reproductive malfunction is due toreduced GnRH secretion (8). However, the mechanismsunderlying the alterations of GnRH release induced by fastingor feed restriction are still unclear. Among the neurotransmit-ters which can alter GnRH secretion, the amino acid GABAis of major importance, exerting an inhibitory action onGnRH neurones (9). In addition, GABA neurones maymediate the oestrogen feedback to the hypothalamus sinceoestrogen receptors were found in preoptic GABA neurones(10), but not within GnRH neurones (11, 12). A pharmacolo-gically induced increase of endogenous GABA levels in thehypothalamus, or i.c.v. application of GABA, suppresses LH Day of feed restriction
0 17 17
Bo
dy
wei
gh
t (g
) 400
200
0
2
1
0
4
3
Testicular w
eigh
t (g)
release in ovariectomized rats (13, 14). In the rat, sites ofF. 1. The effect of restricting feed for 17 days on body and testicularGnRH biosynthesis and release are spatially well separated, weight in male rats fed ad libitum (%, n=17), and those fed a restricted
so it is possible to evaluate changes in neurotransmitter diet (&, n=19). The bars show average values +SEM. *P<0.05.activity and expression at both levels. In the brain, GABA issynthesized directly from glutamate by glutamate acid decarb-
chiasm, hypothalamic grooves and mammilary bodies with a cut depth ofoxylase (GAD), and glutamate is considered to be supplied#2 mm (27).in part from glutamine hydrolysed by glutaminase (GLS)
For the isolation of total RNA from POA punches, MBH and anterior(15). It has been demonstrated that perifusion of hypothal- pituitary, the RNeasyTM Total RNA Kit (Qiagen, Hilden, Germany) was usedamic fragments in vitro with GABA resulted in diminished according to the manufacturer’s protocol. The elution of RNA was carried
out with 30 ml water for POA and with 50 ml for MBH and anterior pituitary.GnRH secretion (16), and that in vivo an i.c.v. applicationRT-PCR was conducted using SUPERSCRIPTTM for RT and SUPERMIXBof GABA reduced LH release and the expression of GnRHfor PCR (both purchased from Gibco-BRL, Eggenstein, Germany). The RTin the POA (17). Hence, a reduction in GnRH pulse generator reaction was carried out with 150 ng total RNA from MBH and anterior
activity, caused by increased endogenous GABA, may be the pituitary and with 10 ml from POA. For PCR reaction 1–3 ml cDNA and25 pM primer were added to the SUPERMIX solution. PCR amplificationresult of increased activity of the GABA-synthesizing enzymeswas carried out in a DNA thermal cycler (Biometra, Gottingen, Germany).GLS and GAD. In addition, decreased reuptake of GABATable 1 shows the primer pairs, numbers of cycles and references for theby the high affinity GABA transporter type 1 (GAT-1) mayencoding sequences used for PCR. The ribosomal protein L7 was used as an
facilitate the high GABA tone. GAD is expressed in two internal standard.forms in the brains of mammals, GAD67 and GAD65 (18) RT-PCR products were cloned using the TOPO TA CloningB Kit
(Invitrogen, Groningen, the Netherlands). DNA sequencing was performedand for both, as well as for GAT-1, alterations in mRNAon double stranded template using the dideoxy chain termination methodexpression during the oestrous cycle and under various ster-with flanking primers on the vector. The sequencing reaction was carried outoidal manipulations have been described (19–21). Using with the AutoReadTM sequencing kit and the gel was run on an automated
RT-PCR we assessed whether a reduction of LH secretion laser fluorescent DNA sequencer (A.L.F.TM, Pharmacia, Freiburg, Germany).Ten ml of the amplified PCR products were size-fractionated by electrophor-induced by long-term feed restriction results from a decrease
esis in a 1.5% agarose gel in TBE buffer and stained with ethidium bromide.of GnRH expression in the POA which may be associatedThey were visualized on a transilluminator at 254 nm (UV-Transilluminatorwith changes of hypothalamic mRNA levels of GLS,Chroma 41, Vetter GmbH, Wiesloch, Germany) and photographed with a
GAD67/65 and GAT-1 in intact male rats. In addition, we positive/negative instant sheet film (Polaroid-MP 4, Land-Camera,examined expression of the GnRH receptor (GnRH-R) and Cambridge, USA).
PCR signals on the negative film were measured with a densitometricof the gonadotropin subunits which depend on phasic GnRHscanner. The arbitrary unit of each sample was related to the correspondingrelease (22, 23).L7 signal. All data from the scanning and RIA analysis were statisticallyevaluated with an unpaired t-test using the PrismB program (GraphPadSoftware, Inc. San Diego, CA, USA). Differences were considered significant
Materials and methods if P<0.05.
Adult male rats, 80 days of age, were purchased from Winkelman GmbH(Borken, Germany) and housed under standardized conditions ( lights on
Resultsfrom 07.00 to 19.00 h, water and feed available ad libitum, 24 °C roomtemperature). After separation into single cages the rats were allowed to The effect of 50% reduced feed intake for 17 days on theadapt for 5 days. The body weights (b.w.) of all rats were obtained before
body and testicular weight of intact male rats is shown inthey were randomly assigned to restricted or ad libitum feed. For 16 dayseach rat in the restricted feed group received 8 g of rat chow/day at 8.00 am Fig. 1. The b.w. of feed-restricted rats was reduced by 35%(50% of the normal daily feed intake of the ad libitum group). All rats had (P<0.05) compared to ad libitum fed rats. Furthermore, infree access to water. On day 17 the rats were weighed and decapitated between feed restricted rats the testicular weight significantly declined10.00 and 11.00 am. Trunk blood was collected and brains and anterior
to 85% of that of ad libitum fed rats (P<0.05). Figure 2pituitaries were rapidly removed and frozen on dry ice.shows serum the concentrations of LH and testosteroneSerum LH, follicle stimulating hormone (FSH) and testosterone concentra-
tions were assayed using double antibody radioimmunoassays as described which were dramatically reduced by feed restriction to 10%previously (24). The frozen brains were sliced into serial frontal sections of those of ad libitum fed rats (P<0.01), while the secretion(600 mm in depth) on a cryostat. The POA was bilaterally punched out from of FSH was not significantly affected.frozen slices with a 1-mm-diameter stainless needle according to the method
Compared to ad libitum fed rats, the expression of LH-bof Palkovits and Brownstein (25). The stereotaxic co-ordinates were A 7.8,L 0.8, V–1.7 (26). With a scalpel, the MBH’s were dissected between optic and a subunit mRNA was significantly lower in fasted rats
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 613–619

Reduction of LH secretion and long-term feed restriction 615
T 1. Primer Pairs and References for Encoding mRNA Sequences.
Probe Primer-Sequence No. of cycles Size of PCR Ref.product
PIT POA MBH
L7 5∞ AGATGTACAGAACTGAAATTC 26 25 25 356 bp (28)3∞ TTGCTCGATCTCTTGGTAAAT
GAT-1 5∞ TTGGATGGACTGGAAAGGTG 25 24 466 bp (29)3∞ GCTATCCTCTTCTTCTCCAT
GAD65 5∞ TGTCAATTATGCACTTCTCC 29 26 627 bp (30)3∞ TTAAATGTGATGAGAGAGGG
GAD67 5∞ AGTCGTCTTGTGAGCGCCTT 25 24 340 bp (31)3∞ GATCCTGGTTGACTGTAGAG
GLS 5∞ AGGCACAGACATGGTT 25 24 471 bp (32)3∞ TGCGTACACTGGAGATG
GLAST 5∞ AATGAAGCCATCATGAGATT 25 24 480 bp (33)3∞ CAATAAGTATCACAGCCACA
GnRH 5∞ CACTATGGTCACCAGCGGG 30 375 bp (34)3∞ TCTTAGGGATCTGCGAGGAGCTCT
GnRH-R 5∞ GAAGCCCGTCCTTGGAGAAAT 30 30 30 441 bp (35)3∞ ATGGCGGCGATTAGCCTGGATCGC
LH-b 5∞ TGCAGAGAAGAATGAGTTCTGCC 24 284 bp (36)3∞ CTCAACCAATGACCTGTGAC
FSH-b 5∞ CTCCTCAGGATCTGGTGTAT 27 362 bp (37)3∞ ACCATCGGCTCCTCTATTGT
a subunit 5∞ TATGCGGCTGTCATTCTGGT 22 351 bp (38)3∞ TCCACAAGTCGTAGCTTCCA
LH FSH T
LH (
ng
/ml)
0.4
0.2
0
2
0
6
4
FSH
/T (n
g/m
l)
0.3
0.1
F. 2. The effect of restricting feed for 17 days on the serum LH, FSHLH-b FSH-b
Rat
io
4
0a-subunit
6
2
PIT
and testosterone levels of male rats fed ad libitum (%, n=17), and those F. 3. The expression of RNA encoding for the LH-b-, FSH-b-and thefed a restricted diet (&, n=19). The bars show average values +SEM. a subunit in the anterior pituitary (PIT) of male rats after 17 days of*P<0.01. restricted (&, n=19) or ad libitum (%, n=17) feeding. The arbitrary
units of each gene were related to the corresponding L7 signal and thebars show average ration values +SEM. *P<0.05.
(Fig. 3), whereas FSH-b mRNA levels remained constant.Expression of L7 RNA, the housekeeping gene used asinternal standard, was identical in both experimental groups. The mRNA levels for GAT-1, the two forms of the GABA-
synthesizing enzyme glutamic acid decarboxylase (GAD65Reduced LH concentrations after long-term feed restrictionwere not associated with diminished GnRH gene expression and GAD67) and the glutamate-forming enzyme glutaminase
(GLS) were measured in the POA and in the MBH. In thein the POA (Fig. 4). In contrast, the mRNA concentrationof the GnRH receptor (GnRH-R) was significantly reduced POA no significant difference of the mRNA concentrations
of all investigated enzymes and the GAT-1 were observedin the anterior pituitary of the FR group compared to controlrats (P<0.05); while no such changes in GnRH-R gene between feed restricted and ad libitum fed rats (Fig. 5).
In the MBH, however, the expression of GAD67 in feedexpression could be observed either in the POA or in theMBH. Again, the expression of L7 RNA in POA and MBH restricted rats was twice that determined in the ad libitum
group (Fig. 6). In addition, the expression of GLS in thewas identical in both experimental groups.
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 613–619
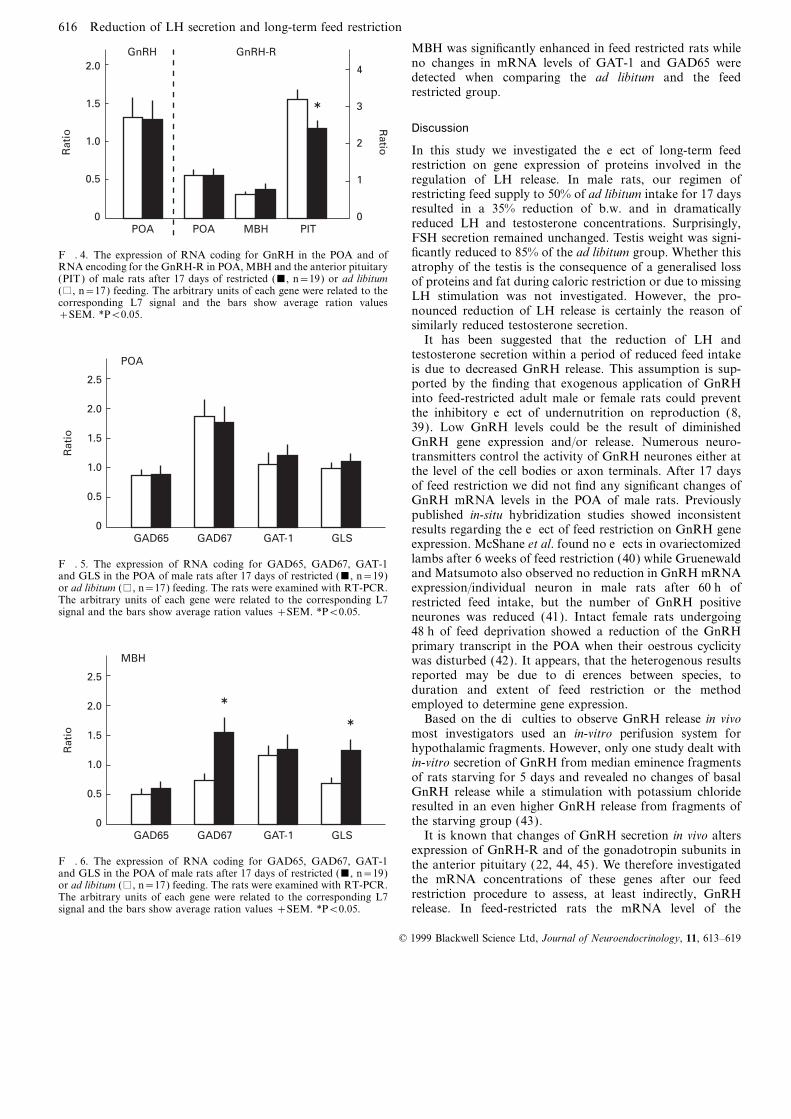
616 Reduction of LH secretion and long-term feed restriction
MBH was significantly enhanced in feed restricted rats whileno changes in mRNA levels of GAT-1 and GAD65 weredetected when comparing the ad libitum and the feedrestricted group.
Discussion
In this study we investigated the effect of long-term feedrestriction on gene expression of proteins involved in theregulation of LH release. In male rats, our regimen ofrestricting feed supply to 50% of ad libitum intake for 17 daysresulted in a 35% reduction of b.w. and in dramaticallyreduced LH and testosterone concentrations. Surprisingly,FSH secretion remained unchanged. Testis weight was signi-
POA POA PIT
Rat
io
1.5
0.5
0
2
1
0
4
3
Ratio
MBH
2.0
1.0
GnRH-RGnRH
ficantly reduced to 85% of the ad libitum group. Whether thisF. 4. The expression of RNA coding for GnRH in the POA and ofRNA encoding for the GnRH-R in POA, MBH and the anterior pituitary atrophy of the testis is the consequence of a generalised loss(PIT) of male rats after 17 days of restricted (&, n=19) or ad libitum of proteins and fat during caloric restriction or due to missing(%, n=17) feeding. The arbitrary units of each gene were related to the LH stimulation was not investigated. However, the pro-corresponding L7 signal and the bars show average ration values
nounced reduction of LH release is certainly the reason of+SEM. *P<0.05.similarly reduced testosterone secretion.
It has been suggested that the reduction of LH andtestosterone secretion within a period of reduced feed intakeis due to decreased GnRH release. This assumption is sup-ported by the finding that exogenous application of GnRHinto feed-restricted adult male or female rats could preventthe inhibitory effect of undernutrition on reproduction (8,39). Low GnRH levels could be the result of diminishedGnRH gene expression and/or release. Numerous neuro-transmitters control the activity of GnRH neurones either atthe level of the cell bodies or axon terminals. After 17 daysof feed restriction we did not find any significant changes ofGnRH mRNA levels in the POA of male rats. Previouslypublished in-situ hybridization studies showed inconsistentresults regarding the effect of feed restriction on GnRH geneexpression. McShane et al. found no effects in ovariectomizedGAD65 GAD67 GLS
Rat
io 1.5
0.5
0GAT-1
2.5
1.0
POA
2.0
lambs after 6 weeks of feed restriction (40) while GruenewaldF. 5. The expression of RNA coding for GAD65, GAD67, GAT-1and Matsumoto also observed no reduction in GnRH mRNAand GLS in the POA of male rats after 17 days of restricted (&, n=19)
or ad libitum (%, n=17) feeding. The rats were examined with RT-PCR. expression/individual neuron in male rats after 60 h ofThe arbitrary units of each gene were related to the corresponding L7 restricted feed intake, but the number of GnRH positivesignal and the bars show average ration values +SEM. *P<0.05. neurones was reduced (41). Intact female rats undergoing
48 h of feed deprivation showed a reduction of the GnRHprimary transcript in the POA when their oestrous cyclicitywas disturbed (42). It appears, that the heterogenous resultsreported may be due to differences between species, toduration and extent of feed restriction or the methodemployed to determine gene expression.
Based on the difficulties to observe GnRH release in vivomost investigators used an in-vitro perifusion system forhypothalamic fragments. However, only one study dealt within-vitro secretion of GnRH from median eminence fragmentsof rats starving for 5 days and revealed no changes of basalGnRH release while a stimulation with potassium chlorideresulted in an even higher GnRH release from fragments ofthe starving group (43).
It is known that changes of GnRH secretion in vivo altersGAD65 GAD67 GLS
Rat
io 1.5
0.5
0GAT-1
2.5
1.0
MBH
2.0
expression of GnRH-R and of the gonadotropin subunits inF. 6. The expression of RNA coding for GAD65, GAD67, GAT-1 the anterior pituitary (22, 44, 45). We therefore investigatedand GLS in the POA of male rats after 17 days of restricted (&, n=19) the mRNA concentrations of these genes after our feedor ad libitum (%, n=17) feeding. The rats were examined with RT-PCR.
restriction procedure to assess, at least indirectly, GnRHThe arbitrary units of each gene were related to the corresponding L7signal and the bars show average ration values +SEM. *P<0.05. release. In feed-restricted rats the mRNA level of the
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 613–619

Reduction of LH secretion and long-term feed restriction 617
GnRH-R in the anterior pituitary was significantly reduced. inhibitory amino acid. In the brain, GABA is synthesizedfrom glutamate by glutamate acid decarboxylase (GAD) andNappi and Rivest showed a reduction of GnRH-R mRNA
in the anterior pituitary of female rats deprived of food for glutamate is mainly formed from ketoglutarate by the actionof aminotransferases and in part from glutamine by the48 h associated with a disturbed oestrus cycle (42); and
Bergendahl et al. demonstrated a 50% reduction of anterior enzyme glutaminase (GLS) (15). GAD is expressed in twoforms GAD65 and GAD67 (30) and both mRNAs as wellpituitary GnRH-R in male rats after 6 days of fasting (46).
Regarding the gonadotropin subunit expression in the as GLS mRNA have been demonstrated in the hypothalamusof rats (54, 55). The expression of both forms of GAD in theanterior pituitary during our long-term feed restriction study
we found a 22.6% reduction of LH-b- and a 26.7% reduction hypothalamus is affected by steroids (19, 20). Furthermore,GLS mRNA is present not only in glutamate neurones, butof a subunit mRNA expression in feed restricted rats, but no
changes in the FSH-b mRNA levels. In line with this finding also in GAD-containing neurones (55). Thus, activation ofthe GABA system during a long-lasting inhibition of GnRHDalkin et al. (23) reported that in castrated/testosterone-
replaced male rats, gene expression of gonadotropin subunits release may lead to increased expression of GAD and GLS.Indeed we observed that in feed-restricted rats, the expressionwas strongly dependent on pulsatile GnRH release, and that
a lack of GnRH pulses decreased LH-b- and a-subunit of GAD67 and GLS mRNA in the MBH was significantlyincreased while no effect could be observed in the POA. ThemRNA levels while FSH-b expression remained unaffected.
Taken together, our findings of reduced GnRH-R expression expression of GAD65 was unaffected in both hypothalamicregions.in the anterior pituitary, LH secretion and expression of
gonadotropin subunits pointed to a modulated GnRH secre- Intra-hypothalamic GABA neurones are located at both,POA and MBH (56). In previous experiments of our grouption from the hypothalamus during states of malnutrition.
However, we cannot rule out as suggested by Sinclair et al. exogenous application of GABA into the POA suppressesLH secretion which results from diminished expression offor the bovine, that total reserves of LH may be directly
influenced by the animals nutritional status via yet unknown GnRH in the POA and the expression of the GnRH receptor(17, 49) in the hypothalamus and anterior pituitary ofmechanisms within the pituitary (47).
Besides the anterior pituitary, GnRH-R are expressed in ovariectomized rats (14, 57). Furthermore, prior to andduring the oestrogen induced LH surge in ovariectomizedPOA and MBH (48, 49). Little is known about the function
and regulation of these hypothalamic GnRH-R. It appears rats release of GABA in the POA but not in the MBH isdecreased. These findings prompted us to suggest that thethat they are part of an autocrine feedback loop since in the
rat i.c.v. injection of GnRH inhibts LH release (50). This POA is the major site for the action of GABA on GnRHsynthesis and release. However, there is also evidence thatautocrine control was also observed in various in-vitro experi-
ments showing a reduced GnRH secretion from hypothalamic pulsatile GnRH release is controlled in the MBH. Forexample, multiple unit activity recorded in the MBH offragments under treatment with GnRH analoga (51, 52).
Furthermore, Krsmanovic et al. demonstrated expression of ovariectomized rats strikingly correlates with LH pulses (58)and local injection of a GABA transaminase inhibitor intoGnRH-R in immortalized GnRH neurones which mediate
an autocrine regulation of GnRH release (4). Though not the arcuate nucleus reduced LH levels in orchidectomizedrats (59). In vitro, release of GnRH from hypothalamicproven, it is tempting to speculate that similar to regulation
of function and expression of GnRH-R in the pituitary, also fragments comprising only the MBH is clearly pulsatile (60)and could be modulated by activation of GABAA receptorsGnRH-R expression in the hypothalamus is dependent on
pulsatile GnRH release. Indirect evidence for this assumption (61). Thus, it appears that depending on the physiologicalconditions in the rat GnRH release may be controlled at thewas provided by Seong et al. as they observed a reduction of
GnRH-R expression in the POA, MBH and pituitary in levels of perikarya and/or terminals. Alternatively, thechanges of gene expression of GAD67 and GLS may beovariectomized rats treated i.c.v. with the GABA agonist
muscimol (49). This treatment is known to inhibit LH release related to a yet undefined physiological function like foodintake or behaviour rather than to gonadotropin secretion.via both, inhibition of GnRH release and gene expression
and therefore they suggested, that reduced GnRH-R expres- GABA is removed from the synaptic cleft through GAT-1.GAT-1 seems to be expressed only on neuronal but not onsion is the consequence of reduced intra-hypothalamic GnRH
secretion. Since the possibility exists that feed restriction glial structures (62), and co-expression with GAD67- orGABA-positive neurones has been shown (63, 64). One couldalters GnRH release, expression of GnRH-R in either POA,
MBH or pituitary may be affected also. The present data speculate that an enhanced GABA turnover could increasethe expression of GAT-1 but our results showed no effect onindicate that indeed GnRH-R expression in the anterior
pituitary is reduced while hypothalmic expression of this gene GAT-1 mRNA neither in the MBH nor in the POA. However,the concomitant increase of expression of GAD67 and GLSremains unaffected. This pointed to a selective regulation of
GnRH release, i.e. secretion into the portal vessel may be in neurones located in the MBH may be associated withenhanced GABA release in this structure. Possibly via presyn-inhibited in feed restricted rats while intrahypothalamic
release is continued. Even though this seems to be highly aptic inhibition, GABA may reduce GnRH release from themedian eminence. In summary we conclude from this studyspeculative, such a selective release and concomitant expres-
sion pattern of GnRH-R has been suggested by Roth et al. that during long-term feed restriction in the male rat GnRHrelease is reduced by a GABAergic mechanism and that thisduring ontogeny of rats (53).
A variety of neurotransmitters modulate GnRH release inhibitory action of GABA takes place in the MBH not inthe POA.(6, 7) and among these, GABA plays a major role as an
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 613–619

618 Reduction of LH secretion and long-term feed restriction
acid decarboxylase (GAD67) mRNA levels in the preoptic area: correla-Acknowledgementstion to the luteinizing hormone surge. Neuroendocrinology 1995; 62:562–570.The authors thank Martina Bremer for the excellent technical assistance.
21 Herbison AE, Augood SJ, Simonian SX, Chapman C. Regulation ofGABA transporter activity and mRNA expression by estrogen in ratAccepted 8 March 1999preoptic area. J Neurosci 1995; 15: 8302–8309.
22 Kaiser UB, Jakubowiak A, Steinberger A, Chin WW. Differential effectsof gonadotropin-releasing hormone (GnRH ) pulse frequency on gonad-Referencesotropin subunit and GnRH receptor messenger ribonucleic acid levels invitro. Endocrinology 1997; 138: 1224–1231.1 Howland BE. The influence of feed restriction and subsequent re-feeding
23 Dalkin AC, Haisenleder DJ, Ortolano GA, Ellis TR, Marshall JC. Theon gonadotrophin secretion and serum testosterone levels in male rats.frequency of gonadotropin-releasing-hormone stimulation differentiallyJ Reprod Fertil 1975; 44: 429–436.regulates gonadotropin subunit messenger ribonucleic acid expression.2 Newmark SR, Rossini AA, Naftolin FI, Todd R, Rose LI, Cahill GF JrEndocrinology 1989; 125: 917–924.Gonadotropin profiles in fed and fasted obese women. Am J Obstet
24 Leonhardt S, Jarry H, Kreipe A, Werstler K, Wuttke W. PituitaryGynecol 1979; 133: 75–80.adenylate cyclase activating polypeptide (PACAP) stimulates pituitary3 Knobil E. The electrophysiology of the GnRH pulse generator in thehormone release in male rats. Neuroendocrinol Lett 1992; 14: 319–328.rhesus monkey. J Steroid Biochem 1989; 33: 669–671.
25 Palkovits M. Isolated removal of hypothalamic or other brain nuclei of4 Krsmanovic LZ, Stojilkovic SS, Mertz LM, Tomic M, Catt KJ.the rat. Brain Res 1973; 59: 449–450.Expression of gonadotropin-releasing hormone receptors and autocrine
26 Pellegrino LJ, Pellegrino AS, Cushman AJ. A Stereotaxic Atlas of theregulation of neuropeptide release in immortalized hypothalamic neurons.Proc Natl Acad Sci USA 1993; 90: 3908–3912. Rat Brain, New York: Plenum Publishing Corporation, 1979.
5 Merchenthaler I, Setalo G, Petrusz P, Negro-Vilar A, Flerko B. 27 Arias P, Jarry H, Leonhardt S, Moguilevsky JA, Wuttke W. EstradiolIdentification of hypophysiotropic luteinizing hormone-releasing hor- modulates the LH release response to N-methyl--aspartate in adultmone (LHRH) neurons by combined retrograde labeling and immuno- female rats: studies on hypothalamic luteinizing hormone-releasing hor-cytochemistry. Exp Clin Endocrinol 1989; 94: 133–140. mone and neurotransmitter release. Neuroendocrinology 1993; 57: 710–715.
6 Kalra SP, Kalra PS, Sahu A, Allen LG, Crowley WR. The steroid- 28 Hemmerich P, von Mikecz A, Neumann F, Sozeri O, Wolff Vorbeck G,neuropeptide connection in the control of LHRH secretion. Adv Exp Zoebelein R, Krawinkel U. Structural and functional properties ofMed Biol 1987; 219: 65–83. ribosomal protein L7 from humans and rodents. Nucl Acids Res 1993;
7 Barraclough CA. Neural control of the synthesis and release of luteinizing 21: 223–231.hormone-releasing hormone. Ciba Found Symp 1992; 168: 233–246. 29 Guastella J, Nelson N, Nelson H, Czyzyk L, Keynan S, Miedel MC,
8 Bergendahl M, Perheentupa A, Huhtaniemi IT. Starvation-induced sup- Davidson N, Lester HA, Kanner BI. Cloning and expression of a ratpression of pituitary-testicular function in rats is reversed by pulsatile brain GABA transporter. Science 1990; 249: 1303–1306.gonadotropin-releasing hormone substitution. Biol Reprod 1991; 44: 30 Erlander MG, Tillakaratne NJ, Feldblum S, Patel N, Tobin AJ. Two413–419. genes encode distinct glutamate decarboxylases. Neuron 1991; 7: 91–100.
9 Jarry H, Perschl A, Wuttke W. Further evidence that preoptic anterior 31 Wyborski RJ, Bond RW, Gottlieb DI. Characterization of a cDNAhypothalamic GABAergic neurons are part of the GnRH pulse and coding for rat glutamic acid decarboxylase. Brain Res Mol Brain Ressurge generator. Acta Endocrinol (Copenh) 1988; 118: 573–579. 1990; 8: 193–198.
10 Flugge G, Oertel WH, Wuttke W. Evidence for estrogen-receptive 32 Banner C, Hwang JJ, Shapiro RA, Wenthold RJ, Nakatani Y, LampelGABAergic neurons in the preoptic/anterior hypothalamic area of the KA, Thomas JW, Huie D, Curthoys NP. Isolation of a cDNA for ratrat brain. Neuroendocrinology 1986; 43: 1–5. brain glutaminase. Brain Res 1988; 427: 247–254.
11 Shivers BD, Harlan RE, Morrell JI, Pfaff DW. Absence of oestradiol 33 Storck T, Schulte S, Hofmann K, Stoffel W. Structure, expression, andconcentration in cell nuclei of LHRH-immunoreactive neurones. Nature functional analysis of a Na (+)-dependent glutamate/aspartate trans-1983; 304: 345–347. porter from rat brain. Proc Natl Acad Sci USA 1992; 89: 10955–10959.
12 Herbison AE, Robinson JE, Skinner DC. Distribution of estrogen 34 Bond CT, Hayflick JS, Seeburg PH, Adelman JP. The rat gonadotropin-receptor-immunoreactive cells in the preoptic area of the ewe: releasing hormone: SH locus: structure and hypothalamic expression.co-localization with glutamic acid decarboxylase but not luteinizing Mol Endocrinol 1989; 3: 1257–1262.hormone-releasing hormone. Neuroendocrinology 1993; 57: 751–759. 35 Kaiser UB, Zhao D, Cardona GR, Chin WW. Isolation and characteriza-
13 Donoso AO, Banzan AM. Effects of increase of brain GABA levels on tion of cDNAs encoding the rat pituitary gonadotropin-releasing hor-the hypothalamic-pituitary-luteinizing hormone axis in rats. Acta
mone receptor. Biochem Biophys Res Commun 1992; 189: 1645–1652.Endocrinol (Copenh) 1984; 106: 298–304.
36 Chin WW, Godine JE, Klein DR, Chang AS, Tan LK, Habener JF.14 Jarry H, Leonhardt S, Wuttke W. Gamma-aminobutyric acid neurons
Nucleotide sequence of the cDNA encoding the precursor of the betain the preoptic/anterior hypothalamic area synchronize the phasic activitysubunit of rat lutropin. Proc Natl Acad Sci USA 1983; 80: 4649–4653.of the gonadotropin-releasing hormone pulse generator in ovariectomized
37 Kato Y, Ezashi T, Hirai T, Kato T. Strain difference in nucleotiderats. Neuroendocrinology 1991; 53: 261–267.sequences of rat glycoprotein hormone subunit cDNAs and gene frag-15 Kaneko T, Mizuno N. Glutamate-synthesizing enzymes in GABAergicment. Zool Sci 1990; 7: 879–887.neurons of the neocortex: a double immunofluorescence study in the rat.
38 Godine JE, Chin WW, Habener JF. alpha subunit of rat pituitaryNeuroscience 1994; 61: 839–849.glycoprotein hormones. Primary structure of the precursor determined16 McRee RC, Meyer DC. GABA control of LHRH release is dependentfrom the nucleotide sequence of cloned cDNAs. J Biol Chem 1982; 257:on the steroid milieu. Neuroscience Lett 1993; 157: 227–230.8368–8371.17 Leonhardt S, Seong JY, Kim K, Thorun Y, Wuttke W, Jarry H.
39 Goto K, Kotsuji F, Tominaga T. Divergent effects of gonadotrophin-Activation of central GABA-A- but not of GABA-B-receptors rapidlyreleasing hormone (GnRH) analogue and authentic GnRH on thereduces pituitary LH release and GnRH gene expression in the preoptic/anterior pituitary gland of rats with restricted feeding. J Endocrinol 1995;anterior hypothalamic area of ovariectomized rats. Neuroendocrinology145: 501–511.1995; 61: 655–662.
40 McShane TM, Petersen SL, McCrone S, Keisler DH. Influence of food18 Kaufman DL, Houser CR, Tobin AJ. Two forms of the gamma-restriction on neuropeptide-Y, proopiomelanocortin, and luteinizing hor-aminobutyric acid synthetic enzyme glutamate decarboxylase have dis-mone-releasing hormone gene expression in sheep hypothalami. Bioltinct intraneuronal distributions and cofactor interactions. J NeurochemReprod 1993; 49: 831–839.1991; 56: 720–723.
41 Gruenewald DA, Matsumoto AM. Reduced gonadotropin-releasing hor-19 Herbison AE, Augood S, McGowan EM. Expression of glutamic acidmone gene expression with fasting in the male rat brain. Endocrinologydecarboxylase messenger RNA in rat medial preoptic area neurones1993; 132: 480–482.during the oestrous cycle and after ovariectomy. Brain Res Mol Brain
42 Nappi RE, Rivest S. Effect of immune and metabolic challenges on theRes 1992; 14: 310–316.20 Unda R, Brann DW, Mahesh VB. Progesterone suppression of glutamic luteinizing hormone-releasing hormone neuronal system in cycling female
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 613–619

Reduction of LH secretion and long-term feed restriction 619
rats: an evaluation at the transcriptional level. Endocrinology 1997; 138: and GAD67 mRNAs suggest that the two glutamate decarboxylases playdistinctive functional roles. J Neurosci Res 1993; 34: 689–706.1374–1384.
55 Najlerahim A, Harrison PJ, Barton AJ, Heffernan J, Pearson RC.43 Warnhoff M, Dorsch G, Pirke KM. Effect of starvation on gonadotrophinDistribution of messenger RNAs encoding the enzymes glutaminase,secretion and on in vitro release of LRH from the isolated medianaspartate aminotransferase and glutamic acid decarboxylase in rat brain.eminence of the male rat. Acta Endocrinol (Copenh) 1983; 103: 293–301.Brain Res Mol Brain Res 1990; 7: 317–333.44 Kaiser UB, Jakubowiak A, Steinberger A, Chin WW. Regulation of rat
56 Tappaz ML, Brownstein MJ. Origin of glutamate decarboxylase GAD-pituitary gonadotropin-releasing hormone receptor mRNA levels in vivocontaining cells in discrete hypothalamic nuclei. Brain Res 1977; 108:and in vitro. Endocrinology 1993; 133: 931–934.371–379.45 Shupnik MA. Gonadotropin gene modulation by steroids and gonadotro-
57 Herbison AE, Chapman C, Dyer RG. Role of medial preoptic GABApin-releasing hormone. Biol Reprod 1996; 54: 279–286.neurones in regulating luteinising hormone secretion in the ovariec-46 Bergendahl M, Perheentupa A, Huhtaniemi IT. Effect of short-termtomised rat. Exp Brain Res 1991; 87: 345–352.starvation on reproductive hormone gene expression, secretion and
58 Kawakami M, Uemura T, Hayashi R. Electrophysiological correlates ofreceptor levels in male rats. J Endocrinol 1989; 121: 409–417.pulsatile gonadotropin release in rats. Neuroendocrinology 1982; 35:47 Sinclair K, Broadbent P, Hutchinson J. Naloxone evokes a nutritionally63–67.dependent LH response in post partum beef cows but not in mid-luteal
59 Donoso AO. Localized increase of GABA levels in brain areas of thephase maiden heifers. Anim Sci (Pencaitland) 1995; 61: 219–230.rat and inhibition of the plasma LH rise following orchidectomy.48 Jennes L, Conn PM. Gonadotropin-releasing hormone and its receptorsNeuroendocrinology 1988; 48: 130–137.in rat brain. Front Neuroendocrinol 1994; 15: 51–77.
60 Purnelle G, Gerard A, Czajkowski V, Bourguignon J-P. Pulsatile secre-49 Seong JY, Jarry H, Kuhnemuth S, Leonhardt S, Wuttke W, Kim K.tion of gonadotropin-releasing hormone by rat hypothalamic explants ofEffect of GABAergic compounds on gonadotropin-releasing hormoneGnRH neurons without cell bodies. Neuroendocrinology 1997; 66:receptor gene expression in the rat. Endocrinology 1995; 136: 2587–2593.305–312.50 DePaolo LV, King RA, Carrillo AJ. In vivo and in vitro examination of
61 Bourguignon J-P, Gerard A, Purnelle G, Czajkowski V, Yamanaka C,an autoregulatory mechanism for luteinizing hormone-releasing hormone. Lemaitre M, Rigo JM, Moonen G, Franchimont P. Duality of glutam-Endocrinology 1987; 120: 272–279. atergic and GABAergic control of pulsatile GnRH secretion by rat
51 Bourguignon J-P, Gerard A, Debougnoux G, Rose J, Franchimont P. hypothalamic explants: I. Effects of antisense oligodeoxynucleotides usingPulsatile release of gonadotropin-releasing hormone (GnRH) from the explants including or excluding the preoptic area. J Neuroendocrinolrat hypothalamus in vitro: calcium and glucose dependency and inhibition 1997; 9: 183–191.by superactive GnRH analogs. Endocrinology 1987; 121: 993–9. 62 Gomeza J, Gimenez C, Zafra F. Cellular distribution and regulation by
52 Feleder C, Jarry H, Leonhardt S, Moguilevsky JA, Wuttke W. Evidence cAMP of the GABA transporter (GAT-1) mRNA. Brain Res Mol Brainto suggest that gonadotropin-releasing hormone inhibits its own secretion Res 1994; 21: 150–156.by affecting hypothalamic amino acid neurotransmitter release. 63 Rattray M, Priestley JV. Differential expression of GABA transporter-1Neuroendocrinology 1996; 64: 298–304. messenger RNA in subpopulations of GABA neurones. Neurosci Lett
53 Roth C, Leonhardt S, Theiling K, Lakomek M, Jarry H, Wuttke W. 1993; 156: 163–166.Ontogeny of the GnRH-, glutaminase- and glutamate decarboxylase- 64 Borden LA, Smith KE, Vaysse PJ, Gustafson EL, Weinshank RL,gene expression in the hypothalamus of female rats. Brain Res Dev Brain Branchek TA. Re-evaluation of GABA transport in neuronal and glialRes 1998; 110: 105–114. cell cultures: correlation of pharmacology and mRNA localization.
Receptors Channels 1995; 3: 129–146.54 Feldblum S, Erlander MG, Tobin AJ. Different distributions of GAD65
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 613–619


















