Redalyc.Mecanismo de la fecundación humana
8
Revista Peruana de Ginecología y Obstetricia ISSN: 2304-5124 [email protected] Sociedad Peruana de Obstetricia y Ginecología Perú Álvarez-Diaz, Jorge Alberto Mecanismo de la fecundación humana Revista Peruana de Ginecología y Obstetricia, vol. 53, núm. 1, enero-marzo, 2007, pp. 45-51 Sociedad Peruana de Obstetricia y Ginecología San Isidro, Perú Disponible en: http://www.redalyc.org/articulo.oa?id=323428183009 Cómo citar el artículo Número completo Más información del artículo Página de la revista en redalyc.org Sistema de Información Científica Red de Revistas Científicas de América Latina, el Caribe, España y Portugal Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto
Transcript of Redalyc.Mecanismo de la fecundación humana

Revista Peruana de Ginecología y Obstetricia
ISSN: 2304-5124
Sociedad Peruana de Obstetricia y
Ginecología
Perú
Álvarez-Diaz, Jorge Alberto
Mecanismo de la fecundación humana
Revista Peruana de Ginecología y Obstetricia, vol. 53, núm. 1, enero-marzo, 2007, pp. 45-51
Sociedad Peruana de Obstetricia y Ginecología
San Isidro, Perú
Disponible en: http://www.redalyc.org/articulo.oa?id=323428183009
Cómo citar el artículo
Número completo
Más información del artículo
Página de la revista en redalyc.org
Sistema de Información Científica
Red de Revistas Científicas de América Latina, el Caribe, España y Portugal
Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto

TEMA DE REVISiÓN
MECANI SMO BE LA FECUNBACIÓNHUMANA
RESUMENSe realiza una revisión actualizada respecto alos conceptos actuales basados en la biologíadel desarrollo temprano (hoy combinando conocimientos de biología celular y genéticamolecular), deuna forma accesible y haciendo consideraciones desde fa ciencia básicapara sus implicaciones con laclínica. Eltrabajo se desarrolla dividiendo elmecanismo dela fecundaciónhumana en varias fases, con fines didácticos: la activación espermática, laincorporacióndel esperma hacia elcitoplasma del óvulo, la activación metabólica del óvulo, la formación de los pronúcleos, la migración de los pron(¡cleos, y fa iniciación de laprimera divisióncelularyeldesarrollo inicial. Así mismo, sehace algunas consideracionesespeciales en relacióncon elmecanismo de regulación epigenéfica en eldeS<irrollo tempra·no yalgunas implicacionesdel desarrollo biológico en lacuestión terminológica científicaque identifica estas etapas.
PaJabrase/ave: Mecanismos de lafecundación, gamefos humanos, espermatozoIde, óvulo.epigénesis.
JorgeAlberto Álvarez-Diaz
Rev Per Gineco! Obste/o 2oo7:5311 }:45·52
Recepción, 8de enero de 2007Aceptación: 2de lebrero de 2007
MédicoSexólogo Clínico:Magisleren BiQética:Diplomadoen Biologia Molecular yen BiomfdicinaMolecular;Doctorandoen elPrograma deCifncias Socosaníaras yHumanidadesMédicas,Univers i~ad Complutense deMadrid.CQTrespondencia a:Dr.Jorge A1~rto Áfllarez Diaz,UniversidadComplUlense deMadrid.CiudadUniverSltariafacultaddeMedicina, Plazade RamÓll yCaja ls/n,Departamento de HistOflade la Med ic ina,PabellónIV,Só1ano. DespachO 3, CP 28040, Ma OriO.EspañaTel,1/34)91394 1521.FaJ; ,(1341913 941 803.e-mas: [email protected]
ABSTRACTWedo an updatedrevisión inregards fa Ihe current concep/sofbiology-based early develop·ment(combining knowledgealcell biologyandmoleculargenelics). inan accessible wayandconsidering implicalions ofbasic science one/inicalpractice, Didaclically we divide Ihemechanism 01 human ferti liza fion In severalphases: spermactivation, incorporation ofthescemintheovumcytoplasm, me/abolicactivalion of theovum. pronuclei formation, pronucleimigra/ion, and ini/ialionofthe lirst cellular divisionandindial developmenl. Sornespecial consideralionsare also doneregarding/he mechanisms al epigenetic regulation inearly developmenl, and sorne !mpJications onbiologicaldevelopment in scien/ific namesidentifyingeese stages.
Key words: Fertilizatíon mechanisms, humangame/es, sperm, ovum, eplgenesis,
INTRODUCCIO N
Hace casi diez años que aparecióun artículo interesante sobre elmecanismo de fert ilización en laRevista Peruana de Ginecología yObstetricie'" , El propós itofundamental de este trabajo es unapuesta al día en algunos conocimientos al respecto.
Alaluzde lagenét ica ylabiologíamol ecular, se ha podidocomprender cada vez mejor el proceso dela fecundac ión. Una cuest ión importantesobre lagénesisdeunserhumanoes que lafecundación noes un evento que surja en un solo
un momento, sinounproceso largoen el t iempo ymolecula rmentecomplejo" :".
Labiología de lareproducción ylabiologíadel desarrollo han utilizadomuchasherramien tas debiología cel ular, genét ica y biologíamolecular parapodercomprendermejor lafecundac ión humana. Lafecundación podría ser definidacomo el proceso que culmina enlaunión de unnúcleo espermáticocon el núcleodel óvu lo, dentrodelcitoplasma activado del óvulo.Para que esto ocurra adecuadamente, algu nos otros eventos deben de acontecer'":
VOL 53 N°1E1'fRQ·N\ARZO 2007 -1-5

Mecanismo de lafecundaciónhumana
• Activación espermática.
• Incorporación del esperma hacia el citoplasma del óvulo, locual incluye launión yfusióndelas membranas plasmáticas delóvulo y el espermatozoide.
• Activación metabólica del óvulo y la iniciación del primer cíclocelula r.
• Formación de los pronúcleos yfusión nuclear.
• Migracióndelospronúcleos, 10quepermitelaunión genómka.
• Iniciaciónde laprimera divisióncelular y el desarrollo inicial.
Además, existen mecanismos deregulación epigenética, descubriéndose cada vez más moléculas producidasen las trompasqueson indispensables para el desarrolloulterioralafecundacióndelóvu loporelespermatozoide. Estoes, sin estas moléculas no es posible que un cigoto alcance sudesarro lloaun embrión. En mediocina reproductivaestoes bien conocido, yaquesi lamaduración enel laboratorio de un óvulo resultafallida, se recomienda hacer uncocultivo con células de la trompa, loquecrealas condiciones másparecidasalas fisiológicaspara eldesarrollo del cigoto.
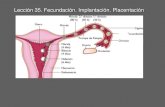
Hay que partir del conoci do hechode quetras un eyaculado de un varónfértil,entendiendoentoncesqueexistan un mínimo de 20 millonesde espermatozoides por mililitroeyaculado y un mínimo de 3 mleyaculados, no más de 200 espermatozoidesllegarán al tercio externo de latrompadeFalopio pa ra intentar interactuar con el óvulo. Porotro lado,esconocido actualmenteque el tamañodel folículo tienerelacióncon lacalidaddel óvulo,yesto
tiene repercusiones paraque se produzca ono lafecundación. ElHquido folicular tienevarios agentes quepueden inducirla quimiotaxisde losespermatozoides, entre elloslaprogesterona(que podríatener entonces una significaciónclínicasu utilizaciónen ciclos de medicina reproductiva); pero, muy probablementeno esel principal agente qulnuotáctico'",
EVENTOS ESPERMÁTICOSSi bien es muy importante unaseriede eventos espermáticos paraque se dé el proceso de la fecundación,ellos no ocurren de manera aislada, sino siempreen comunicación con elóvulo. Se sabe quelamatrizextracelular del eúmulusoóphorus (estructura única de losóvulos demamíferos superiores) esesencia l no solo paralaovulac ión,sino también paratodoel procesode la Iecuncacró n'".
l a capacitación es necesaria pa rael desarrollo de la habilidad de lareacciónacrosómlceen mamíferos.Durante lainteracción de losgametos, lareacción acrosómica Inducida por el cúmuJus oóphorus, es unprerrequisito pa ra que el espermatozoide pase la zona pelúcida, sefunda con y penetre al ÓVUIOI81• laP4 (progesterona) secretada por lascélulas del cúmulus, es un cotactorimportante para que ocurra esteevento de exocitosis. la reacciónacrosómica resultade lafusiónentrelasmembranasacrosomal externa ycelular, que llevan a laexposición de lamembrana acrosomal interna. la unión de agonistas (P4 olagl icoproteína ZP3) areceptores dela membrana plasmática activa señalesintraespermáticasyvías enzimáticasrelacionadascon lareacciónacrosómica.
Entre las molécu las descritascomo receptoresespermáticos potenciales se incluye las proteínasZP (de zonapelúcida),proteína GJGo yreceptoresde tirosinaquinasa.Los receptoresespermáticos paraP4 están pobremente caracterizados, excepto un receptorpa recidoaGABAA. la reacciónacrosómicapromovida por ZP (y P4) está mediada por un incrementoobligatorio en la concentración de calciointracelula r, que aparece primeroen el segmento ecuatorial delacrosoma y se esparcepor toda lacabeza. los canales teneos de lamembrana plasmática re lacionados con la ent rada de calcio sonoperados por una despolarizaciónde la membrana plasmát ica yfosforilaciones proteicasmediadaspor laproteinaquinasaeyuna proteína tirosinaquioasa'". Parte delincremento del calcio intracelularpuede ser debido a la liberacióndelcalcio almacenado, através decanales dependientes de IP3 ycAMPo Además, laactivacióndelaadenilato ciclasa y la fosfolipasae incremen tan el calcio intracelularyestimulanlaactividad dePlA2yla despclimerización de laactina, llevando a la fusión de lasmembranas.
En la zona pelúcida de los mamíferos se encuentran varias glicoproteínas importan tes para loseventos dereconoc imientoyadhesión celulares. Estas glicoproteínasson laZPl , ZP2YZP3, sientoestaúltima lamás estudiada'!".los residuos de ca rbohidratos delas glicoproteínas ZPtienen unrolclave en este paso tempranodelafecundación. la imagendelazonapelúcidaalamicroscopiaelectrónica ofrece unaapariencia de unaespecie de malla. ZP2 y ZP3 tie-
.t(i REVISTA PERUANA DE GiNECOLOGíA y OBSTETRIOA

'. Jorge Alberto Álvarez-Diaz
nen filamentos, ZPl entrecruzaestos filamentos como una mallatridimensional y ZP3 actúa comoun receptor del espermatozoide.La unión específica de una especie de espermatozoide a la zonapelúcida es mediada por una molécula(galactosil transferasa)en lasuperficiede lacabeza del espermatozo ide, que une el oligosacárido de unión O de la ZP3 auna proteína trrcstnaq ulnesa't-'.Una vez localizada la unión, ocurreen elespermatozoidelareacciónecrcsomat La ZP3 funcionacomomolécula de ad hes ión y comosecretagoga para la exocitosisacroscnar'" . Se haencontrado 30000 sitios de unión en laZP3. Estosugiere una unión compleja, en laque part icipan múltiples receptoresen la superficie del espermatozoide, sugiriéndose cada vez másque se trata de moléculas de la familiadel complementoexpresadasa nivel de rnembran s'!".
Lafamil ia de glicoprote ínas ZP hasidoampliamenteestudiada desdelasdécada delos 80's,en mode losderatón, yextrapolados sus resultados alosseres humanos. Sinembargo, parece ser que otras especies demamíferos (entre ellas lossereshumanos)producenunacuartaglicoproteína de estafamilia,unaZP4(l4.151,que estaría involucradajunto con la ZP3 en la activaciónespermát ica y la inducción de lareacción acrosomal(l6.17 .18l . Deaquí quenoesténclaros estos eventos aún en seres humanos, requiriéndose un mayor estudio al respecto'!". Esta serie de hechos,entre otros, ha hecho que secuestione seriamente si el modelo deratón es adecuado pa ra el estudiode las fallas en la fecundación yextrapolarlas aseres humanos?".
Al completarse lareacciónacrosómica, se liberanenzimas(como laacrosina. unaproteasa tripsinodependiente, y la neuraminidasa)que causan lisisde la zona pelúcida(21
) , teni endo importancia paraellolaactividad del proteasomaesperrn át ico?".
Antes de continuar, es importan teaclarar dos aspectos que tienenrelevancia para la clínica. Por unlado, ya se ha mencionado la bajaproporciónde espermatozoides quealcanzan el tercio externo de latrompa, aludiendo por tanto a subuenamotilidad. Sin embargo,estudiosrec ientesmuestran quemásde75%de los espermatozoides dehombresfértiles no tienecapacidadde interactuar con la zona pelúcidaI23). Esto tiene repercusiones irnportantesen entender loque significa lafertilidad olainfert ilidad, asícomo en los estudios de análisisseminal. Por otro lado, varonesot igospé rrnícos. a pesar de tenerglicoproteínasZP normales, tieneninteracciones defectuosas con lazona pel úcida, por loque cada vezse recomienda más que si acudenatratamientos de medicina reproductiva, no se pierda tiempo y dinero en inseminación y ni siquieraen una fecu ndación in vitra, sinoque se llegue directo a11CS1124l .
INCORPORACiÓN DElESPERMATOZOIDE HACIAEl CITOPLASMA DEl ÓVULOLa incorporación física del espermatozoide hacia el citoplasma delóvuloocurre simu ltáneamenteconla activación del óvulo y la reacción cortical. Al fundirseelespe rmatozoide con el oolerna (membrana del óvulo) se da origen a lasingamia, es decir, al englobamiento del espermatozoide en
el óvulo. Un grupo específico demoléculas es re lacionado con unanclajet ipointegrina entre ambosgametos, para que se dé la fusiónl251, sugi riéndose también Quepuedeestar mediada por moléculas con act ividad de metalooroteinasa?". Esto no está totalmente estudiado, teniendo lafertilina un papel importante aúnpordefinir. Los microfilamentosdeactina se ensamblanenlacortezadel óvulo, evento fundamentalpa ra la incorporacióndelespermatozoide, cuyo estudio se sugierecada vezen modelos deprim ate yno en modelos de ratón?" ,
ACTIVACiÓN METABÓLICADEl ÓVULOLos óvulos son células metabó licamente quiescentes. La unióndel espermatozoide y el óvu lo inicia una cascada de eventos quetransforman esta célulaquiescenteen una metabólicamente muyactiva,dinámica, animada:elcigoto.Aunque haytodavía opinionesmuydiversas sobre la manera precisacomo estoocurre, está claro Queentodos los sistemas de fecundaciónuna elevación delas concentraciones del calcio intra celular es elmensajero principal en lacomunicación de laseñal de activación .
Unmecanismopa ra esta elevaciónde las concentraciones de calciopodría ser laentrada de calcio extracelular, luego de interacc iónentre integrinas del óvulo conligandos para estas proteínas, enla membra na del esperma, comolatertñtnaw'. aunque podríanestar involucrados otra s moléculas,como P-selectina, receptoresFeymoléculasCD porpartedel óvulo,así como algunas otrasespecíficaspor parte del espermatozoide.

n, ,, o . Mecanismo de lafecundacl6n humana
Alternativamente,el espermatozoi de podría activar al óvulo tambiénpor lainyeccióndirecta deunaproteína derivada del esperma , laoscilina?". Sin embargo, se insisteque no sería esta proteína sola,sinoacompañada de otra seriede eventos que permitaque continúen oscilaciones de calcio, entrando haciael óvulo, que en losmamíferos puede durar hasta varias horas'> '.
Si bien la act ivac ióndel óvulose dainicialmentepor entrada de calcio,el cómo esta señal iniciael desarroIlotempranonoesclaro?" . Recientemente, se ha puesto atención alpapel de la fostorilación de proteínas tirosmacuinasasw'. ya que seconocealgunas reaccionesquímleasque llevan a la activación de unaproteínaquínasa de42 kDa, llamada ER KlMAP (demitogen activatingprotein)1331, que finalmente lleva ala destrucción de laciclinadel MPF(de M-phase promoting factor).
La síntesis inicial de proteínas estádada por el mRNA del óvulo, comomecan ismo de control, y este control espaciotemporalde laexpresióndel mRNA es esencial para un grannúmero de procesos?". Estos procesos, resultado final de la fecundación, incluyenlarecuperación delnúmero diploide de cromosomas, lavariabilidad genét ica dada por larecornbinación meiótica, ladeterminación del sexo cromosóm ico ygenéticoy el inicio de lasegmentación.
La membrana plasmática del ovocito se vue lve impermeable paraotros espermatozoides, gracias a lareacción cortica l, endonde secretagránulos que digieren proteolíticamentesitios deunión espermatica, alterando la cubierta del óvulo. Tal vez, el mejor entendido bloqueo rápidode lapolispermla es elrápidobloqueo eléctrico, endonde
laapertura de canalesiónicosresultaen una inversión del potencial demembrana, toquepreviene lafusiónadicional de otrosespermatozoides.
FORMACiÓN DE lDS PRDNÚClEDSy FUSiÓN NUCLEAREl esperma entra al óvulo con unnúcleo altamente condensado, inact ivo, y también muchos óvulosestán en rea lidad en un arresto concromosomas rneióticos condensados; lafecundaciónpermitelacondensación y remodelación de losgenomasparentales. Enelespermade mamíferos, el núcleo es modificadodurante laespermicgénesis, detal forma que las típicas proteínascrorn osórn icashistonas son reduc idas por protaminas, loque perm iteun empaquetamiento del ADN enuna conformac ión superhe licoidal?". El ADN paternose unealasntstones maternas yotras proteínasyes cubiertopor una nueva envoltura nuclear de origen materno,quetieneláminas nuclearesycompiejos de poro nuclear. Estepasoescrucial, yaquesi no se ensamblanadecuadamente los complejos deporo nuclearcon laslám inasnucleares, el transporte nucleocitcplásrniconose llevaadecuadamenteylafecundación es bloqueadal361•
La fecundación secompletacon unaseñalizaciónpor launión genó mlca.Enmamíferos, los pronúcleosno sefunden durantelainterfase: lospronúcleosadyacentes se separan porunrompimientonuclearenlaprimera mitosisy loscromosomasparentales se mezclancuando se alineanen la placa metafásica del primerhuso mitótico. Comoconsecuencia,los eventos de la fecundación demamíferos se inician durante la segunda rneiosis y no están completos hasta la primera mitosis.
MIGRACiÓN DE lDS PRDNÚClEOSy UNiÓN GENÓMICALuegode la incorporación exitosadel esperma al óvulo y la activación de éste, ycoincidentecon laformación de los pronúcleos, unnuevo sistema de motilidad seensambla dentro del óvu lo íecundado ysu organ ización es dirigidapor el centrosoma del espermatozo ide recientemente introducido(37). El áster del espermatozo ide es una estructura de microtúbu losorgan izadatridimensionalmente de forma radiada,organ izada por el centrosoma, que se encuentraadyacenteal pronúcleo. Elásterdel espermatozoide tienetresfunciones importantes descritas:
• su crecimientosubyacente a lacorteza del óvulo guía al espermatozoide hacia el cent ro delóvulo,
• cuandoel extremo distal de losmicrotúbufosdinámicos se uneal pronúcleodel óvulo, da lugara una súbitayrápida translocación haciael centro del ésterdeles permatozoide, y
. Ia elongación continua de losrnic rotúbulos del áster mueveahora alospronúcleos adyacentes hacia el centro del óvulo.
Sin embargo, el papel del ásterdurantelafecundaciónno debe sersobreenfatizado; algunas formasde fallas en la fecundación parecen ser debidas a defectos en laorganizaciónofuncionamientodelásterl381. La y-tubulina delcentrosoma del espermatozo ideatrae numerosas proteínas maternas, espec ialmente v-tubuli namaterna y proteínas del aparatomitóticonuclear:391. Esta transformación del centrosomadel espermatozoide como precursor inacti-
-18 REVISTA PERUANA DEG IM.COl OGiA y O BSTETRIOA

Jorge Alberto Alv3lez-Diaz n, o '
Q
va hacia un centroorganizador demicrotúbulos, reproduceelcentrosomadel cigoto. Se ha descritoquees crucial la integridad del espermatozoi de en orden a tener uncentrosoma patern o integro , pa raque se puedan dar posteriormen te de una forma adecuada las divisionesmitóticas poscigóticas"?' .
Siguiendo lamigración yunión delos pronúcleos, el centrosoma seduplica y separa para llegar a sera futuro los dos polos del primerhuso miótico. El ADN va a lareplicación durantela interfase, yuna nuevacascadade eventos queregulan el ciclo celular lleva a lainiciación de la primera divisiónyel desarrollo temprano.
Al completarse lameiosis 11 delóvuloque permaneció en arresto en dietioteno, se expulsa también el segundo cuerpo polar. En esta etapaocurre la integracióngenómica, esdecir, la mezcla de cromosomaspaternos y maternos. En todos losmamíferos noexisteunacariogamia{fusión de ambos pron úcíeos), porlaquenose formaun núcleodiploideverdadero conel cigoto.
MECANISMOS DEREGULACiÓN EPI GENÉTICATRAS LA FECUNDACiÓNLa blologla molecular, en años recientes, ha aportado muchos datos, con relación a susten tar elhecho biológico que un cigoto notiene toda la información necesaria para desarrollarse adecuadamente hasta un ser humano, a lolargo de todo el programa de desarrollo y desde los estadios másternpranos'" >" .
Para eldesarrollodura nte elperiodo preimplantatorio, exi sten muchos factores en las trompas que
son absolutamente necesarios eindispensables pa ra que se puedan dar de forma adecuada losestadios tempranosdel desarrol lodel producto de la fecundación .
Un ejemplo de la importancia defactoresepigenéticos que influencian el desarrollo más temprano,es el de los aminoácidos comomoduladores. Algunos de losestimuladores más potentes deldesarrollo embrionarioson la tau rinay la glicina, sugi riéndose quesu papel es mucho más amplio quesolo el de sustratos metabólicos.Una posibilidad desu mecanismoregulador podría ser a través delpH, (pH intraceluíar)'?".
Por otra parte, es muy fuerte laevidencia de que, enel desarrollopreimplantacional dentro deltracto femenino, las células generan su ATP (la molécula quealmacenayproveeenergíapor exceleneiadentrodelacélu la), apartir desustratospresentesen eloviductoyen f luidos uterinos'?".
La evidencia experimentalacumulada también indica que factoresde crecimiento de origen maternose relacionancon laregulación dedesarrollo preirnplantacional'?". Alparecer, losfactores de crecimiento estarían más relacionados conla segunda parte del desarrollopreimplantacional (formación delblastocisto ysu desarro llo)que conlaprimera parte (divisióncelular).La expresión de oncogenesespecíficos puede ser partede lasvíasde señali zación iniciadas por losfactores de crecim iento.
Tamb ién, hay evidencia que la regulación del pH" sibien está dadopor mecanismos de transporte lónico a través de la membrana delcigoto reciénformado, también es
cierto que esto depende de concentraciones de iones y sustratospresentesen el fluido del oviducto, evidenciando aquí también laparticipación mateme>".La evidencia crece cada día másalrededor de este tema. Hoyesbien conocido que latrompatieneun papel crucial al genera r un microambiente que potencia las posibilidades de fecu ndación, iniciode las primeras divisiones celularesy lamisma implantación. Tanto los gametoscomo las primerascélulas post-cigótcasexpresanhormona liberadora de gonadotropina(GnRH) y hoy ya se ha demostradotambién que la GnRH extrahipotalámicapresenteen elovicuctotiene un rol paracrinatautocrinosustancial en las primeras fases de)desarrollo mástempra no-" .
LA CUESTIÓN TERMINOLÓGICAUn término que surge inmediatamente al hablar sobre la fecundación es el de 'embrión' , por loquese hace alguna consideracionesdesde los hechos científicos.
Embrión es una palabra queetimológicamente viene del griegoembryon, de en (prefijo que significa dentro de) y bryein (hinchar,crecer, germi naro brotar)yque daaentender la idea que surge en si gloXVI Ideunente preexistente Quesolamente 'crece' o 'se desarrolla'dentro del útero de la madre. Dehecho, lapalabra como tal noexiste antes del siglo XVI I. La ra íz esprofundamente histórica. En laépoca actua l, embrión tiene otrosignificado, muy claro para la embriología, Que diceQue setratadeun "Término que se refiere al desarrollo humano en las etapas ini·ciales de la diferenciación. Por logeneralel tecnicismo noseemplea
VOLs3 N lEl'lRO·W.RZO í?007 .t9

Mecanismo de lafecundación humana
para retenral productode lafecundación sino hasta la segunda semana, despuésde formadoel disco embrionario bilarniner'"?". Deaquíquehaya aparecido el neologismo de 'pre-ernbrión', pa ra denom inar al conjunto de estadiosprevios a que apareciera el discoembrionariobilaminar. Contrarioalo que se ha expresado muchasveces, no quiere decir que al hablardepre-ernbrión se diga quenoesté vivo, que sea una cosa, queno sea humano, o que nosea merecedorde respeto por una dignidad intrínseca (que este tema yanoesde corte biológico, sino filosófico, en el marco de la otoétcacomo una ética aplicada).
Por otro lado, el significado quesepuededenominar como 't écnica' o 'científico' ofrecido por laembriología, es desconocido por lamayoría de lagente, quien no entiende precisamente eso. El diccionario, por ejemplo, recoge lossignificados que se atribuye a laspalabras por lamayoría y, en consenso, yen el caso de embrión, laRea l Academia Española lo definecomo un "servivo en lasprimeras etapasde su desarrollo, desdela fecundación hasta que el organismo adquiere las característicasmorfológicas de laespecie.//2. Enla especie humana,productode laconcepción hasta fines del tercermes de embara zc'w'.
La moderna embriología, que haincorporado datos de la biologíadel desarrollo y de la genéticamolecular,atal punto que parecieran inseparables,deja clarosalgunos datos que son relevantesparaestedebateI50) . Primero, que luego del proceso de la fecundaciónhay un cigoto, este cigoto se divide llamándose cada célula resul-
tante un blastómero, hasta el estad iodemórula, desdelos16 hasta los 64 blastómeros, aproximadamente. Así llega al útero, paraluegoformarel blastocisto, con unnúmeroaproximado de 100 células y una cavidad centra l (blastoceie), Luego deello, seformarálamasa celularexterna(trotoblastootrofoectodermo,quedará origena la placenta y otras membranasextraembri ónlcas). que rodea a lamasa celular interna (embrioblasto, ungrupode 20 a30 células que originará el embrión, yaqueel sufijo 'blasto' habladeunacélulaprecursora, inmadura). Poresto, a todos los estadios previosa la aparic ión del embrión se lesdenomina colectivamente comoun 'pre-embrión'. Vistoasí, se trataría másbien de unarealidadbiológica, más allá de una cuestiónmeramente terminológica oeufemística. Otros datos recientesapoyanlaideade que pre-embriónyembriónson dos rea lidades biológicas distintasv". Además, seacota algunas cosas, clarif icandoque un cigotocorrespondería a laentidad biológica que surge desde elcontactode los gametos hastaantes de lasingamia; asumendoque un cigotono es un embrión,el cigoto no requeriríade muchasprotecciones Que se le dan hastaahora (tal vez por ello, algunos laboratorios decen tros de medicinareproductiva han optado portomarcomo criterio para la crioprese rvaclón precisamente Que seencuentren en estado de pronúcleos, yasí evitar la consideraciónética de tener embrionescriopreservados)'?'.
Sin embargo, la ESHRE (EuropeanSociety for Human Reproductionand Embryology) cuenta con una
declara ción deética respecto delestatusdel embrión, adoptandoeltérmino de 'embriónpreimplantacional' , por motivossocioculturales más Que biológices>". Luego de exponer el cesarrollobiológicoque seda luegodeuna fertilización in vivo (no necesariamente igual al que se da invitrot, la declarac ión dicen que"Sin embargo, para evitar confusión y term inología especial, lacual puede llevar a incertidumbreen la mente pública, y con el conocimientoQuehay muchas otrasdefiniciones para la entidad queresulta de la fertilización duranteel desarrollo hacia el feto, hemosdecididoutilizarel térm ino gená r¡cade 'embrión ', elcual serefieraalas etapas de la fertilización a laformación del disco embrionario".
El debaterespectoaestacuestiónes muy amplio, ydeberíaencaminarse cada vez más a la objetividad de loshechos biológicos yalareflexión filosófica adecuada sobrelos mismos. Tal vezel futuroinmediato en este sentidovaya aencaminarse al aná lisisprofun dosobreladignidad del pre-embrión. El futuro lo dirá.
REFERENCIAS BIBLIOGRÁFICAS1. Bellido 1'. Meca nismo de fertiliz ación.
Gincco lObsrcr (Perúl. 1997;4313U 83·90.2. Edwards RG,Brody SA. Principies and prar
tke ni ds,isled human rcprodccno n. 1!>1 ed.llalllllXlfe: SJumk'r;COnlpJny.I'l 'l'i: 315·44.
3. Hcffner LO. Human rcproduction al aglcnce. NI'W Yor k: Bldckwell Scienre Ud.2001 : 45.
4. Srbarten G. Fenilizancn. En:Fauser BCJM,Rutheríord Al, Slrau» If, Van Steirteghern.Molecu lar biology in reproductive medicine. Balthnore: Ihe Parthenon PublishingCruup. 1999: 297· 310.
5. Ja;swal BS. Iur-Kcspa L Dar j, Ma, hiach S,fi,enha ch M. Hum,m sperm Chemold~ i~: 1,progcsterone a chemoau rar tam! BiolReprod.1999;60:1314.9.
6. l imo l, Kirnata K. Cumuhrs oophorus exI racc l luIMm o1l ri ~: 'h rons nucüo n and rl'gu,laeon.Cell Strurt Funct. 2001;26:1B9-96.
50 REVlSlA PEIIlIANA DE G1f'1COLOGiAy OBSTETRICIA

Jorge Alberto .Iv3rez-Di3z
7. PanatC.SerresC, lou,llUJ<'l P.Ttwacrosomereecuco in human spermerozoa. Hiel Cell.200();91:255·66.
8. D,¡rszonA, l ahMea P. Nishigaki l. h pillO'oaLlon chanu•." lsincperrnphysiolog\,. PhysiolRev. 19911;7912':481-51O.
9. Ia kasakiS, MoriE,Mori1.Snuctureofsugalehall1s includediu mammallan zona pellucida glyeuprol(·il1> and Ilwir pcictuial rolesin Ipt'rm·egg interarticn. Bkx'lurn BiophisAr1a, 1999;1473:106-15.
10. BMMC Crosby JA.Mmeno RO. Earlyslepsoíspcrm c¡:¡: inH;,laelion,during rnamrualianlertilization,Cell Biol lnt. 1996;144:33-9.
11 . Thaler CD. Caedullo RA. lile inilialmolccular intetarticn beewcen mease >permandtbeZon.l pellucida iH OIllple1\ hil'l(.!ingeven.J Biol Chern. 19% :271( 8): 23289-97.
12. OaceKS. üronson RA. Ghebrl·hiwI'IB.Sur.lace expresslon of ccnt pleme nr rece pto rgClq-R /p 33 il inr reased 011 lile plasmamernhrane01human spenuatnma añcr ca
pacítanon. Hiol Rcprod. 2002;66:823-9.13. SpargoSC, HupeRM.Evolution and remen
clature oi thc zona pelludda gene l,lmily.Biol Reprod. 2003;68:358·62.
t-t.teñecret. Conre rSj. SalpckarA,OluíowohiO, Ashlon P. P,l\"lu\'ie B. (1 al. Pour zonapellucida glycoprnll'lIl'are expressed in thelIum,lIl. Hum Reprod, !()()4;19:1580·6.
15. Yu rewiez EC Sac('(J AG. el ,11.. HelefO·oligomerizalion·dependem binding 01 pigoo·e\'le zona pellueida glycoproteins ZPBandl PCloho.Jr 'peml mcmhral1l' ",,,>ides. JHiolCllem. 1998; 273:7488·'14.
16. YOJ1ezawa N. Fukui N, KlInoM ,ShimxbM,Ga ka 5. Mitsui S, N,1k.11l0 M. Molecula rc10ningoí hovine zona pellucida glyeupro·leins lPA and lPll and ,1I1alysis lor~no·
bindingcomponenlofzona. Eurll-liochl'm.2001;268:587-94.
17.CII.tkravarty S, Surai 1< . Gupla SI<.Bacu lov ifUS-e~pr(-;""'t 1 tl'e(Mnhinamhumanzona pelludda gIY({JprUlein-B induces.t("llNl m.t1e~ocyl')"S in c.-.pacilall>f! , petmalozo.t in additiun to lOn,l pellucid,l gil"coprolein. C. Mol HUIll Reprod,2005;1 1:365·72 ,
lB.Palra t C, Auer 1. F,luque 1'. t l'and ri Rl,Jouanoe1 P, Serres C. Zon.. pelluc id.. immfenilisedhlJman ooeyh', induces ... volr'lgl'dependenlealcilJm inllu~ ,100lhe acrO'>Oll1ereadion in 'l lenO,llol o.l. bul eannol be ¡Jenetraled by sperm. BMC IJevelopm\·m,ll Hiorog\,. 2006;{,:59 dod 0.11 36114; 1-21j X.6-59 Disponihk' ('n: http://WIVW,bioowdcentral,eom/14; 1· 213X!6/59
19, Neuher t. POWI' I) RD.ls lhe mouS(> a e1 inically[('Ievanl mode!fm human fertilizalioníailu'e>/ Hum Reprod. 2000;15(1 ):171·-1,
20. PaulP,MilesDG. Penetr,llioo. adhe, iooalKJíusioninmamloalians¡Jl'f1n·egginll'raClion.Science. 2002;29(15576):2183-S,
21. Morale\ P, KOflg M. PiZM'OE ,I'a~I\'nCl'ar
licipalion 01 Ihe sperm proleasome in hu·man lerlilizalion. Hum Rl'prod. 2003;18(5);1010-7.
22. li u DY. Ganen e BakerHWG.lowproportioll' oi 'ccrm can hind 10 tbe zona pellucída oi human OOC}'ll'S. Hum Rep rod.2003;1811 1):2.182-9.
2.1.liu D'( Baker HWG. High íreqcencycfdetcctivesperm-zona ¡Jl'l lueida inleraelion inolígczoosperm¡r iníertile meno HumReprod. 2004;l '1(21:228. JJ.
24. Elesch FM, Oadella 8M. Dynamks oi lhernammalianspetrnpla,m,lruenbrane inlh(,proress of íertilization. Btorhim BiophisAd a. 2000;1469:197-235.
25. Bronson RA, FUli FM, CJlzi F, Doldi N.ferrari A,Evidence Ihal a íun(!ion,ll íerlilin.like ADAM playsa role in human spermoolemmal inlerael;ons. Mol H'lm Reprod.1999;515):433·-10.
26. Hew¡lwn l. Primatemodelsforas,i,led re-produClive ter hnofogíes. Rcprod.2004:128:293-9.
27. FinazC, H,lrtlmami-H,lIlza S.Adlll'Sion protcms exprcsvedon human gemctesurfaccs,1nd egg3Cliv,1Iion. Bio]e-n 2000:92:!35·44.
28, Parringlon J,Swann K, She\'chenko VI,SeSilyAK. la i FA. Cakium oscillation in mnmmahan egg. lriggcred bya soluble spcrm protein. Nnure. 199b;3Z:264·8.
29. NixünVL '\\cOougallA.ícres KT.Ca1+~cillalions and lhe cell cycle at íerlili,alionofmammalian and ao;cidiao eggs. BiolCell.2000;92:187-96.
30. (jappa B. ChiriS,Eggaelivalion: Up' lreamoftheÍl'nilizaliOrl cakium ,ignal. BiolCell.2000;'J2:215·JJ.
31 S~IO K. l okmakov M . Fukami Y. Ferliliza·lioo signaling aod prolein'lyrm ine kinases.Compar,llive Biochemi,try Jnd l'hy\io logy.2000;126:129·48.
J2.S hibu}·a EK. Boulton lG . Cobb MH.Ruderman IV.Aoiv,ltion oi1'42 MAPkinaseand lhe relea,e oi ooeyles írom cell cydearres!.EM80 J. 1992;11:3963·75.
n. VandelVeldcoAW.lhoma~AA. lheroleoíthe 5" unlranslated regioo oí ,In mRN.A. inlransl.11 ion regulalion duringdcvelopmenl.1m J Biochem Cell Biol. 1999;31:B7·IOb,
)4 , Hern.\nJ ez Pérez O. Balle~ le rns l M.Biologia molecular del espe,maluzoide demamiíerO'> . En: Vel,\ zljucz Mor!ezullla ).IE dJ Biologia de la reproducción. 1I.16ico,OF:Universidad AutóoomaMelropolilaoa,IZlapalapa.1Y'J8:1 9.1 ·216.
35, Rawe VY. Brugo·Olmedo S. Nodar FN.Ponlio R, 511tov~ky P, Ahnormal ass(·mblyoí annulale lamell,le.tnd nudear porecomplexC" coioeides .,.,. ¡th icttilila lioo <lIrcs! allhe pmoudear ,lage 01 human zygOlic developmenl. HIJmR('prod, 2003;18:576-82.
,lb.Sehallen G. lhe renlrosome .tnd ilS madI'of inh{'ril,lnce: lile r<'duelion 01 eentm\.ll!llCdllring ¡\arlWlo¡\\'nesis amI ¡IS reSlor,lt iondurin g ícrliliz.tlion. Dev lliol.1994;165:299-335.
37. SimerlyC, WuG. Zoran5 el al.l hel),llerllJIinl1erilancc oí the celUrosome, the eell'~
microlllhule.organizalion eenler inhlJmans
and ñte implications iar illiertillly. NanrreMro . 19lIS;IA 7·53.
38.5imelly C. Zorao SS.PavoeC. Dominko 1.5utovskyP,N,l\",uaCS,5alhhuryILSchencnG. Btparentat inheotance oí g-tubuhn duroinghuman fe" ilizalion:molecular recces..tutíon cí functionalzygctic cemroscmes íntnsermnetcdhum,lIloocyle, and inceíl-freee~1 r,lCb nucleated by human sperm. MolBiol Cell. 199'1;tO,2955---f:,Y.
39. Moo mjy M, Colorobero tr. veeck Ll.,Rosenwaks Z, Palerrno Cf). Spetm inlegrilyi~ critieal íor normal mñotíc division andcMly l·mbryonie dcc elopment. Mol HumReprod. 19Y9;519):836·44,
40. BudncCA.CCf,110 RC 'lhe zygOle: lo beoruot lo be a persono J Med Phitosophy.1989;14:64\ -5.
41.Smilh R. Brologta del embriónprcimplantacional. En: Beca lnfante IP.(COIf1¡J.l El embrión human o. Sanliago:'\Wditc'l!'\ 11(>0, 2002:! 1-49.
42. Bavlstcr Bl). MtKiernan SH. Regulauon ufharnstcr emhryo dcvclopmcnt in vuro byamino acids,En: Bavister BD. Preimplanla.tion embryo dcveloprncm. New York:SprinRet-Vcdag.1Y93;57-72-
43. tce se HI. Enelgy rncíebellsm in preimplanl,ltiOfl tkvelopmcnt. En: Il avi~er BD Pteirnplnntation embryo development. New York:SprmRer-Vellag. 1993:73-82.
44. Fischer B. GrOls1h faClor> as regulalors oim,llllma]ianpreimpl,lI1lation emhryotkvelopmcnt. En: Bavisler Bl). Pre implanlationl'mbr)"ode\'e!opmenl, Ncw York: Springer.Verlag. 1993:83.%.
46 Bal1 z lM, Bigger~ JD,lecheneClnlraeellll '1M pH R'gulJtiOfl hylhe preimplamalion em·luyo. En: Bavi.ter RO. Preimplaolalion em·hryo developmenl. New York: Springer·Verlag. 1993:97·tll ,
47Chan EM. RagaF. Bonilla·Mu,ole>f, PolanMl . Human()viduclal¡;ooadolropin·releJI·ing hormone: pú'osible impliealions in fe'·lilil ation, early emhr\,onie de \'elopmen1.and implalltalion. I Chn Endocrin Me1Jh.2000;85141:1377-81.
48 Mou'e Kl. Emb riología c1iniea 4' ed.Méxim: Itlterallwricalla. 198'1: 6,
4'1Re,ll Acadl' mia Espariol ,\, Oiecion,lriodelal (·ngua Espaiiola. 22' cd. Madrid: ElpasaCalpe. 2001:880.
50 Lacado.·na IR.Em brione\ humano,y cultivosde tejidos: rr·llexiolll'> cien l ¡íica~. é-\ ieas yjuridiCJ" Rev Der Gen H. 2000:12:1911 12.
51 B,lb,ldm G. The mara1s1 Jlu~ ohheemhryo:
the human emhryo in lhe UK HumanFett ilb,¡tion alld Em bryologyIResealCh Pur·pose» Regulalion 2001 debate. ReprodBiomed Online. 2003;171 11: 12-6.
S2 Tesarik l. Azygute is nol all emhryo:elhiealalld leg,ll considl'rations. Reprod Biom(>f!Online. 2004;19(1):13·6.
53 ESRHE la, kfOlCe 0 0 Elhin and l aw. l, lilemoral~,ltU~ ofIhe pre.implanlalion embryo.Human Reproouclion. 2001:16(5):1046·8.
VOl 53 N") ENE RO·MARZO 2007 51