
prr.hec.gov.pkprr.hec.gov.pk/jspui/bitstream/123456789/8459/1/shazia arif thesis full ...
201
i ISOLATION, CLONING AND SEQUENCING OF VITAMIN A, VITAMIN C AND FOLATE GENES (cDNA) FROM SEA BUCKTHORN ( HIPPOPHAE RHAMNOIDES L.) FRUIT BERRIES SHAZIA ARIF (Reg. No. 2001-URTB-3139) Session 2008-2011
Transcript of prr.hec.gov.pkprr.hec.gov.pk/jspui/bitstream/123456789/8459/1/shazia arif thesis full ...

i
ISOLATION, CLONING AND SEQUENCING OF VITAMIN A,
VITAMIN C AND FOLATE GENES (cDNA) FROM SEA
BUCKTHORN (HIPPOPHAE RHAMNOIDES L.) FRUIT
BERRIES
SHAZIA ARIF
(Reg. No. 2001-URTB-3139)
Session 2008-2011

ii
Department of Plant Breeding and Molecular Genetics Faculty of
Agriculture, Rawalakot University of Azad Jammu & Kashmir
ISOLATION, CLONING AND SEQUENCING OF VITAMIN
A, VITAMIN
ISOLATION, CLONING AND SEQUENCING OF VITAMIN
A,
VITAMIN C AND FOLATE GENES (cDNA) FROM SEA BUCKTHORN
(HIPPOPHAE RHAMNOIDES L.) FRUIT BERRIES
By
SHAZIA ARIF
(Reg. No. 2001-URTB-3139)
(M.Sc. (Hons.) Plant Breeding and Molecular Genetics
A thesis submitted in partial fulfillment of the requirement for the degree
of
Doctor of Philosophy
In
Plant Breeding and Molecular Genetics

iii
Session 2008-2011
Department of Plant Breeding and Molecular Genetics
FACULTY OF AGRICULTURE, RAWALAKOT THE
UNIVERSITY OF AZAD JAMMU AND KASHMIR
“In the name of ALLAHA, the most Beneficent, the
most Merciful”

iv

v

vi
DEDICATION
I dedicate this humble effort to
my affectionate and loving
Parents, my late brother, my
husband, my Brother & Sisters
TABLE OF CONTENTS
Sr. No. Page No.
I Title page ii
II Certification iii
III Declaration iv
IV Dedication v
V Table of contents v
VI List of Tables xi
VII List of Figures xii
VIII Abbreviations xxii
vii
IX Acknowledgement xxviii
X Abstract xxx
01 CHAPTER 1 01
01 GENERAL INTRODUCTION 01
1.1 Sea buckthorn (Hippophae rhamnoides) 01
1.2 Origin and History. 02
1.3 Chemical composition and Nutritional value 03
1.4 Vitamins 05
1.4.1 Ascorbic Acid 05
1.4.2 Caroteniods and Tocopherols 06
1.4.3 Flavonoids 08
1.4.4 Folate 09
1.5 Amino acids 10
1.6 Scope 11
1.7 OBJECTIVES 15
02 CHAPTER 2 16
02 ASCORBATE OXIDASE (AO) GENE (Abstract) 16
2.1 INTRODUCTION 17
2.2 MATERIALS AND METHODS 21
2.3 Designing of Primers 22
2.4 RNA isolation protocol (TRIzol® Reagent) 23
viii
2.5 Rapid Amplification of cDNA Ends (RACE-PCR) 23
2.6 RT-PCR Amplification Protocol for Hr-AO cDNA 24
2.7 Gene purification 25
2.8 Gene Cloning protocol 27
2.8.1 TA cloning Vector 27
2.8.2 Set up the ligation reaction 27
2.8.3 Host Cells 28
2.8.4 Electroporation of E. coli: 28
2.8.5 E. coli cells Preparation for Electroporation. 30
2.8.6 Colony PCR procedure 32
2.8.7 Plasmid DNA extraction 33
2.8.8 Plasmid PCR and sequencing 35
2.8.9 Gateway cloning 36
2.8.10 Gateway® Entry Clone 36
2.8.11 LR Reaction. 40
2.8.12 Transformation 41
2.9 AO gene expression analysis 42
2.9.1 Semi-quantitative RT-PCR 42
2.9.2 ATGene (Arabidopsis thaliana AO gene) expression Pattern 43
2.10 RESULTS 43

ix
2.10.1 Isolation and sequence analysis of large fragment of AO cDNA
from sea buckthorn
43
2.10.2 Gate way cloning 46
2.10.3 Differential expression of AO gene 47
2.10.4 Phylogenetic analysis 50
2. 11 DISCUSSION 54
2.12 CONCLUSIONS 58
03 CHAPTER 3 59
03 FOLATE (HPPK-DHPS) GENE (Abstract) 59
3.1 INTRODUCTION 60
3.2 MATERIALS AND METHODS 65
3.2.1 Plant Material 65
3.2.2 Designing of Primers 65
3.2.3 DNA Extraction 66
3.2.4 PCR Amplification and Molecular Cloning 67
3.2.5 Isolation of RNA and expression Analysis 67
3.2.6 ATGene (Arabidopsis thaliana gene) expression evaluation for
HPPK-DHPS
68
3.2.7 Sequence Analysis and Phylogenetic Reconstruction 69
3.2.8 Homology Modeling and Protein Structure Prediction 70

x
3.3 RESULTS 71
3.3.1 Amplification of HrHPPK-DHPS Gene from Sea buckthorn 71
3.3.2 HPPK-DHPS is differentially expressed in Sea buckthorn tissues 72
3.3.3 Sequence analysis of HPPK-DHPS identified
from sea buckthorn
75
3.3.4 Phylogenetic reconstruction of HPPK-DHPS Genes 79
3.3.5 Tertiary structure of Folate protein 81
3.4 DISCUSSION 83
3.5 CONCLUSIONS 88
04 CHAPTER 4 88
04 CAROTENIODS BIOSYNTHESIS GENES (LCY-β AND PSY)
(Abstract)
89
4.1 INTRODUCTION 90
4.2 MATERIALS AND METHODS 95
4.2.1 Plant Material 95
4.2.2 Designing of Primers 95
4.2.3 RNA extraction and PCR Amplification 96
4.2.4 Expression analysis of Genes 97
4.2.5 ATGene (Arabidopsis thaliana Gene) expression analysis of LCY
and PSY
98
4.2.6 Molecular Cloning and Sequence analysis 99
4.3 RESULTS 100

xi
4.3.1 Isolation of Hr-Lcyb and Hr-Psy Genes 100
4.3.2 Differential Gene Expression Analysis 102
4.3.3 Sequence analysis 106
4.3.4 Phylogenetic Analysis 114
4.4 DISCUSSION 117
4.5 CONCLUSIONS 122
4.6 General summary 123
4.7 Conclusions and future recommendations 127
4.8 List of published papers and books 129
05 CHAPTER 5 131
05 Literature cited 131
LIST OF TABLES
Table No. Page No
2.1 Comparative list of the Vitamin C contents in sea buckthorn berries,
seed oil and pulp.
20
2.2. Detail of Primers used in this study for H. rhamnoides AO cDNA
cloning, expression analysis (RT-PCR) and Gateway cloning.
22
xii
2.3. Overview of different AO genes, function and expression in specific
tissues with their reference source.
50
2.4 List showing organisms and accessions used in phylogenetic tree
construction.
3. 1. Detail of Primers used for Hr-HPPK-DHPS gene amplification and
expression studies.
66
4.1. Constituents of sea buckthorn Fruit 94
4.2. Detail of Primers used in this study for H. rhamnoides Lcy and Psy
cDNA cloning and expression analysis (RT-PCR)
96
LIST OF FIGURES
Figure No.
Page No.
2.1 Structure of ascorbic acid (a) ascorbic acid (reduced form), (b)
dehydroascorbic acid (oxidized form).
17
2.2. Map of Vector pTZ57R/T used for TA cloning purpose. 27
2.3. The map of pDONAR™201 vector used in gateway cloning. 38
2.4 Map of the destination vector used in Gateway cloning. 41

xiii
2.5 (a) Isolation of high quality total RNA from sea buckthorn tissues. (b)
Amplification of Hr-AO gene using RACE technique of PCR. (c) PCR
confirmation of the clones carrying recombinant plasmids with M13 and
short internal primers. M=1Kb DNA ladder.
44
2.6 Alignment of Hr-AO and S. lycopersicum nucleotide sequences with
Mac VectorTM 7.2.3. (Accerlrys Inc.)Gcg/Wisconsin pakage university
of Wisconsin). The top sequence is that of tomato AO and the bottom
HrAO-Like. The identical bases are shown against a black background.
45
2.7 (a) Amplification of Hr-AO product by standard PCR with gateway
primers. (b). Gel showing pDONAR recombinant plasmids. (c&d) The
pDEST vector pXCG-mYFP (51delta 35s) SB512 colony and plasmid
46
PCR confirmation respectively. (e) Stable integration of AO-PB clones
carrying the recombinant plasmid into destination vector. (f) pDEST AO
recombinant plasmid confirmation through PCR. M= 1 kb marker.
2.8 Hr-AO transcript amplifications in vegetative bud, shoot apex, green leaf,
green fruit and mature orange red fruit tissues of sea buckthorn through
semi quantative RT-PCR in comparisons with control Actin gene. Actin
gene is used as control, to check equal loading and PCR quality check.
47

xiv
2.9 Arabidopsis thaliana AO (AT5G21100) gene expression pattern from
global data set of microarray under different conditions: (a) in different
plant tissues, (b) hormonal conditions, (c) Abiotic stress, and (d) light
exposures respectively.
48
2.10 Phylogenetic reconstruction of ascorbate oxidase gene from different
species. Neighbor joining tree was made by means of MEGA5 software.
The Value on each node indicated bootstraps replication of 1000. This
species along with accession used in tree includes AY971876-
L.esculentum-AO, XM_002528929-R.communis-AO,
XM_003532792G.max-L-AO XM_003638399-M.truncatula-AO,
Y15295-M. truncatulaL-AO, AB457618-P.sativum-AO, AF529300-
G.max-AO, D43624-
N.tabacum-AO, XM_002312802-P.trichocarpa-AO, XM_003555611-
G.max-AO, NM_197271-O.sativa-AO, NM_147871-A.thaliana-AO,
NM_001203424-A.thaliana-AO, AY099586-A.thaliana-AO, BT003407-
A.thaliana-AO, XM_003336029-P.graminis-AO,
XM_003713188M.oryzae-AO, XM_001875289-L.bicolor-AO, AF206722-
B.juncea-AO,
53
D55677-C.maxima-AO, X55779-Pumpkin-AO, AF233594- C.melo-AO,
J04494-Cucumber-AO, FR750377-C.sativus-AO, Y10226-C.melo-AO,
AB004798-A.thaliana-AO, B1698178-I.pine-AO, FJ896040-F.ananassa-
AO, EF528482-M.domestica-AO and Gu321223-M.pumila-AO
3.1 Structure of Folate (a) and 5-methyltetrahydrofolate(b) found in sea
buckthorn.
60

xv
3.2 The systematic presentation of plant folate biosynthesis pathway. The
main pathway enzymes includes 1, GTP cyclohydrolase I; 2, nudix
hydrolase; 3, non-specific phosphatase; 4, DHN aldolase; 5, ADC
synthase; 6, ADC lyase; 7, HPPK-pyrophosphokinase; 8, dihydropteroate
synthase; 9, dihydrofolate synthase; 10, dihydrofolate reductase; 11,
folylpolyglutamate synthase. 12,DHN, dihydroneopterin; 13, ADC,
aminodeoxychorismate; 14, PABA, p-aminobenzoate, 15,-PPP
triphosphate.
62
3.3 Amplification of HrHPPK-DHPS gene. (a) Image of Genomic DNA
isolated from three different sea buckthorn samples (S1, S2, S3) run along
1kb DNA ladder. (b), PCR amplification of full length sequence of folate
gene (HPPK-DHPS) from sea buckthorn genomic
DNA. M= 1kb DNA ladder.
71
3.4 Cloning of HrHPPK-DHPS gene. (a) Positive colony PCR confirmation
with M13 primers. (b) purified recombinant plasmids HrHPPK-DHPS
gene. (c) PCR confirmation of recombinant plasmid with internal short
gene specific primers. M= 1kb DNA ladder.
72
3.5 RNA isolation from different tissues of sea buckthorn shown
against HindIII Marker. RT-PCR analysis of HPPK-DHPS gene
transc different tissues (leaf, Fruit, Bud, Seed) of sea buckthorn
against 50bp ladder. 18S rRNA was used as internal control.
λ DNA/
pts in
as shown
73
ri

xvi
3.6 AtGene Expression pattern for HPPK-DHPS gene (AT4G30000) from
global data set of microarray in: (a) different plant tissues, (b) under
different hormonal conditions, (c) abiotic stress conditions and (d)
response to different light conditions.
74
3.7 Alignment of newly isolated HrHPPK-DHPS amino acid sequence with
its ortholog from S. lycopersicum (FJ972198) using MaVectorTM7.2.3.
(Accerlrys Inc.) gcg/ Wisconsin pakage university of Wisconsin). The
similar regions are shown against a black background and differences are
exposed against white background.
76
3. 8 Alignment of newly isolated HrHPPK-DHPS nucleotide sequence with
its ortholog from S. lycopersicum (FJ972198) using MaVectorTM7.2.3.
(Accerlrys Inc.) gcg/ Wisconsin pakage university of Wisconsin). The
identical regions are shown against a black background and differences
are exposed against white background.
77
3.9 Multiple sequence alignment of newly isolated HrHPPK-DHPS amino
acid sequence with Mac VectorTM
7.2.3. (Accerlrys Inc.) gcg/ Wisconsin
pakage university of Wisconsin) from sea buckthorn and its ortholog from
S. lycopersicum, G. max, A. thaliana and T. aestivum amino acid in
blocks highlight similarity. The strictly conserved
78
regions are shown against black background.

xvii
3.10 Phylogenetic reconstruction of HrHPPK-DHPS gene. Neighbor joining
tree is constructed using MEGA 5.0 software. Values on the nodes indicate
the bootstrap replication of 1000. Phylogenetic inference of enzymes
containing the two domains HPPK and DHPS was made using 15
complete coding sequences H. rhamnoides, S. lycopersicum (GenBank
accession No. FJ972198) T. aestivum (EF208803), B. distachyon
(XM003562616), O. sativa (NM001066822), O. sativa (AK068210), P.
sativum (Y08611), M. truncatula (XM003614065),
C. arietinum (XM004490156) G. max (XM003518734), G. max
(XM003517802), A. thaliana (NM105586), A. thaliana
(NM001203939), A. lyrata (XM002888660) and A. thaliana
(BT033093) were included in the alignment. Four clusters are enclosed by
brackets. A scale is given at the bottom.
80
3.11 Conserved domain of newly isolated HrHPPK-DHPS gene obtained from
domain search data base showed hppk superfamily and
pterin_binding domain.
81
3.12 Predicted 3D structure of HrHPPK-DHPS proteins from different plant
species shown at the same scale. A, X-ray Structure of the template
Bifunctional 6-hydroxymethyl-7, 8- Dihydroxypterin Pyrophosphokinase
Dihydropteroate Synthase From Saccharomyces
Cerevisiae (Lawrence et al., 2005) B, 3D structure of sea buckthorn
HPPK-DHPS produced using MODELER 9v10, C, 3D model of S.
82

xviii
lycopersicum, D, 3D model of A. thaliana, E, 3D Model of G. max. F,
Model of T. aestivum. Models in different protein structure were colored
according to different domains. Pterin binding domain DHPS was given
Blue color, HPPK domain was shown by red color. Interdomain linker was
given grey color.
4.1 Structure of important caroteniods reported in Sea buckthorn. (a)
αcarotene, (b) Lycopene.
91
4. 2 Structures of different forms of Carotenoids reported to be present in the
sea buckthorn.
92
4.3. Systematic presentation of caroteniod biosynthetic pathway in plant. The most
important enzymes that participate in pathway includes geranylgeranyl
pyrophosphate (GGPP), phytoene synthase (PSY), phytoene desaturase
(PDS), lycopene-β-cyclase (LCYB lycopene-ε-cyclase (LCYE) β-ring
hydroxylase (HYDβ) ε-ring hydroxylase (HYDε), zeaxanthin epoxidase
(ZEP), Violaxanthin de-epoxidase (NSY) and Epoxycarotenoid
dioxygenase (NCED).
93
4.4 (a) Comparison of RNA band intensity from different tissues including
bud, root, apex, leaves, seed and fruits. (2) Amplification of full length
cDNA of Hr-Lcyb (c) Hr-Psy gene amplification through RT-PCR. M
stand for 1kb DNA ladder.
101
4.5 (A) a. Positive colony PCR of Hr-Lcyb with M13 primers. b. Purified
selected recombinant plasmid with target gene inserts. (B). a. positive
colony of Hr-PSY confirmed by PCR with M13 primers. b. Recombinant
101

xix
purified plasmid of Hr-PSY gene. c. Recombinant Hr-PSY plasmid PCR
confirmation. M stand for 1kb DNA ladder.
4.6 (a) Photograph showing different tissues of sea buckthorn a. bud, b. apex,
c. root. D. leaf, e. seed, f. fruit. (b) RT-PCR analysis of Hr-lcyb and HrPsy
gene transcripts in above mentioned tissues of sea buckthorn in
comparison with internal control actin-1. M stand for 1kb DNA ladder.
102
4.7. Relative Gene Expression pattern for Lycopene β-cyclase gene
(AT3G10230) from global data set of microarray: (a) in different plant
tissues, (b) hormonal conditions, (c) abiotic stress and (c) light exposures
respectively.
104
4. 8. Relative Gene Expression pattern for Phytoene synthase gene
(AT5G17230) from global data set of microarray: (a) in different plant
tissues, (b) hormonal conditions, (c) abiotic stress and (d) light exposures
respectively.
105
4.9 Alignment of newly isolated Hr-Lcyb nucleotide sequence with its
ortholog from S. lycopersicum using MaVectorTM7.2.3. (Accerlrys Inc.)
gcg/ Wisconsin pakage university of Wisconsin). The similar regions are
shown against a black background and differences are exposed against
white background.
108

xx
4.10 Alignment of newly isolated Hr-Lcyb amino acid sequence with its
ortholog from S. lycopersicum using MaVectorTM7.2.3. (Accerlrys Inc.)
gcg/ Wisconsin pakage university of Wisconsin). The conservations in
sequence are shown against a black background and differences are
109
exposed against white background.
4.11 Alignment of newly isolated Hr-Psy nucleotide sequence with its ortholog
from S. lycopersicum using MaVectorTM7.2.3. (Accerlrys Inc.) gcg/
Wisconsin pakage university of Wisconsin). The similar regions are
shown against a black background and differences are exposed against
white background.
110
4.12 Alignment of newly isolated Hr-Psy amino acid sequence with its ortholog
from S. lycopersicum using MaVectorTM7.2.3. (Accerlrys Inc.) gcg/
Wisconsin pakage university of Wisconsin) software. The similarities are
shown against a black background and differences are exposed against
white background.
111

xxi
4.13 Multiple sequence Alignments of newly isolated Hr-Lcyb amino acid
sequence with its ortholog from S. lycopersicum (NM_001247297), V.
vinifera (JQ319639), N. pseudonarcissus (GQ327929), M. truncatula
(XM-003624977), N. tazetta (JQ797381), C. sinensis (DQ235259) and L.
barbarum (AY906864) using MaVectorTM7.2.3. (Accerlrys Inc.) gcg/
Wisconsin pakage university of Wisconsin). The strictly conserved
regions are shown against a black background and differences are exposed
against white background.
112
4.14 Multiple Alignment of amino acid sequence of Hr-Psy gene with its
representative from other plant species. The names and accession numbers
of these members are: Lopomoea (AB499050), T. erecta
(AF251015), S. lycopersicum (EF534739), N. tabacum (JF461341) and
113
Tomato (M84744). The strictly conserved regions are shown against a
black background and differences are exposed against white background.

xxii
4.15 Phylogenetic analysis of Hr-Lcyb and other selected plant members.
Neighbor joining tree is constructed using MEGA 5.0 software. Values on
the nodes indicate the bootstrap replication of 1000. Phylogenetic
inference was made using 25 complete coding sequences H. rhamnoides,
S. lycopersicum (NM_001247297), C. maxima (AY217103), C. sinensis
(DQ496224), C. papaya (DQ415894), P. trichocarpa (XM_002308867),
R. communis (XM_002531452), E. japonica (JX089591), V. vinifera
(JQ319639), B. nivea (EU122344), C. moschata (JN559395), S. europaea
(AY789516), C. arietinum (XM_004493356), M. truncantula
(XM_003624977), G. max (XM_006576725), P. vulgaris (HQ199604),
A. palaestina (AF321534), L. barbarum (AY906864) Lpomoea_sp.
Kenyan (AB499055), N. tabacum (X81787),
Chrysanthemum_X_morifolium (AB205041) T. officinale (AB247456), N.
pseudonarcissus (GQ327929) and N. trzetta (JQ797381).
114
4.16. The detail and accession no of the plant species for the phytoene synthase
gene which were used for the phylogenetic analysis includes H.
rhamnoides, P,mume (AB253628), F.vesca (XM_004289519), G. max
(XM_003544910), C. arietinum (XR_189445), D.Kaki (FJ594485), C.
sinensis (EF545005), D. carota (DQ192187), A. deliciosa (FJ797304), M.
indica (JN001197), p. trichocarpa (XM_002327528), R.communis
(XM_002532929), M. esculenta (GU111719), C. papaya (DQ666828),
116

xxiii
P. sitchensis (EF676285), T.erecta (AF251015), C.melo (GU361622),
C.lanatus, M.charanita (AY494789), M. cochinchinensis (KF233991), S.
lycopersicum (M84744), O. fragrans (JQ699273), M. germanica
(AY986508), A. palaestina (AY661705), C. roseus (HQ438241), G.
jasminoides (HQ599860), C. canephora (DQ157164), B. oleracea
(JF920036), B. napus (AB454517), E. japonica (JX097048), B.
distachyon (XM_003579062), T. aestivum (BT009537), H. vulgare
(AK358888) and P. juncea (HM539711).
4.17 Conserved domains of Hr-Lycopene β-cyclase
showing main NAD_binding domain.
118
4.18 Conserved domains of Hr-Phytoene synthase gene consisting of Six main
regions.
119
ABBREVIATIONS
AA Ascorbic acid
AAC antioxidant activity coefficient

xxiv
ABA Abscisic acid
ADC Aminodeoxychorismate
Al Aluminum
AMV-RT Avian Myeloblastosis Virus Reverse Transcriptase
AOX antioxidant value
AO Ascorbate oxidase
AsA L-ascorbic acid
ATP Adenosine triphosphate
BAC Bacterial artificial chromosome
Bp Base pair
Ca Calcium
Cm Centimeters
CaMV Cauliflower mosaic virus
CCS Capsanthin–capsorubin synthase
cDNA complementary DNA
Cd Cadmium
CPTA 2-(4-chlorophenylthio)-triethylamine
Cu Cupper
DHA Dehydroascorbate
DMSO Dimethyl sulfoxide
DNTPs Deoxinuceotide Triphosphate
xxv
DXS Deoxyxylulose 5-phosphate synthase
3D 3-dimentational
E. coli Escherichia coli
EC Electrical conductivity
EDTA Ethylenediaminetetraacetic acid
ESTs Expressed sequence tag
EtBr Ethidium Bromide
EMSAs Electrophoretic mobility shift assays
Fe Ferric
Fe Iron
FPGS Folylpolyglutamate synthase
G Gram
GDC Glycine Decarboxylase Complex
GGPP Geranylgeranyl pyrophosphate
GGDP Geranylgeranyl diphosphate
GR Glutathione reductase
GSRP Gene-specific reverse primer
GPP/FPP Geranyl/farnesyl diphosphates
GSH Glutathione
GW Gateway cloning

xxvi
HPPK-DHPS Hydroxymethyldihydropterin pyrophosphokinase–
dihydropteroate synthase
Hr Hippophae rhamnoides
Hr-Lcyb Hippophae rhamnoides Lycopene beta cyclase
HYDβ) β-ring hydroxylase
HYDε ε-ring hydroxylase
IPTG Isopropyl- β -D-thiogalactopyranoside
IPPS Transisoprenyl diphosphate synthases
Kbp Kilo base pair
Kg Kilogram
K Potassium
L Litter
LB Luria-Bertani
LCYβ Lycopene beta cyclase
LCYE lycopene--ε-cyclase
Li Lithium
MDHA Monodehydroascorbate
MDHAR Monodehydroascorbate reductase
MEGA Molecular Evolutionary Genetics Analysis
MEP Methylerythritol 4-phosphate
Mg Magnesium
xxvii
MgCl2 Magnesium chloride
Mn Manganese
mRNA Messenger RNA
MTX Methotrexate
N Nitrogen
NADPH Nicotinamide adenine dinucleotide phosphate
NAOH Sodium Hydroxide
Na Sodium
NCED Epoxycarotenoid dioxygenase
NCBI National Center for Biotechnology Information
NIGAB National Institute for Genomics and Advanced Biotechnology
NSY Violaxanthin de-epoxidase
OD Optical density
ORR oxidation rate ratio
P Phosphorus
PABA p-aminobenzoate
PAGE Polyacrylamide gel electrophoresis
Pb Lead
PCR Polymerase Chain Reaction
PDB Protein Data Bank
PDS phytoene desaturase
xxviii
PEG Polyethylene glycol
PFRA Prairie Farm Rehabilitation Administration
PlF Phytochrome-interacting factor
PPP Triphosphate.
PSY phytoene synthase
PVP Polyvinylpyrrolidone
qPCR Quantitative real-time PCR
QTLs Quantitative trait loci
RACE-PCR Rapid amplification of cDNA ends
Rb Rubidium
RDA Recommended dietary allowance
ROS Reactive oxygen species
RT-PCR Reverse transcription Polymerase chain reaction
SBT Sea buckthorn
SDS Sodium Dodecyl Sulfate
SHMT Serine hydroxymethyltransferase
SL Solanum lycopersicum
SNM Species not Mentioned
Taq Thermus aquaticus
TBE Trisbuffer ethylene diamine tetra acetic acid
TE Tris ethylene diamine tetra acetic acid
xxix
THF TetrahydroFolate
TLC Thin layer chromatography
TILLING Targeting Induced Local Lesions In Genomes
µg Micro gram
µl Micro liter
USSR Union of Soviet Socialist Republics
UV Ultra violet
WT Wild-type
X-Gal 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside
ZEP zeaxanthin epoxidase
Zn Zinc

xxx
ACKNOWLEDGEMENTS
First and above all, I praise Allah, the almighty for providing me this opportunity and granting
me the capability to precede my PhD studies successfully. Without Allah’s will, I would not
have been able to conduct this research. My humble gratitude to the Muhammad (S.A.W) the
Last Prophet of God, who brought the humanity under the umbrella of peace and prosperity,
whose way of life has been a continuous guidance for me
I would like to thank the following friends and family members for their support throughout the
process of completing this dissertation. First of all, to my parents, I express utmost love and
appreciation for allowing me to explore my academic and artistic interests with the freedom
necessary to grow as both a student and as an individual. Their prayer for me was what sustained
me thus far.
I would like to thank my supervisor and mentor Professor Dr. Syed Dilnawaz Ahmad Gardezi,
Vice Chancellor, AJ&K University, whose support and guidance made my thesis work possible.
He has been actively interested in my work. I am very grateful for his patience, motivation,
enthusiasm, and immense knowledge taken together, makes him a great mentor.
I would like to thank my co-advisor Professor Dr. Muhammad Ramzan Khan on the spot for
providing me support and guidance, warm encouragement, thoughtful ideas, and critical
comments, made my thesis work at NIGAB, NARC Islamabad a success. Special thanks to
Professor Dr. Ghulam Muhammad Ali for his kind behavior, knowledge and provision of Lab
facilities during Molecular work at NIGAB as an external student. I am thankful to my thesis
committee members Professor Dr. Muhammad Fareed Khan Chairman PB&MG, Professor Dr.
Abdul Hamid department of Horticulture) for their support and guidance.

xxxi
I thank Dr. Ghulam Hasnain University of Florida for his advices and help in providing foreign
acceptance and his friendly assistance with various problems all the time, especially for his help
with the paperwork, and his help outside the laboratory.
I would like to thank the Higher Education Commission (HEC), Pakistan for selecting me as a
research scholar in Indigenous PhD scheme for financial aid to my university during my PhD
studies and giving me the prospect to achieve my PhD degree. I am obliged to HEC for six
month foreign exposure at the University Of Florida (USA), under International Research
Support Initiative Program (IRSIP). Moreover, thanks are due to Prof. Dr. Rathinasabapathi,
Balasubramani for accommodating me as guest researcher in his group and allowing me to use
lab facilities at Institute of Food and Agriculture Sciences, University of Florida (USA) apart
from his scholastic discussion.
I want to express my deep thanks to my esteemed to research associate Zaheer Abbas for the
trust, the insightful discussion, offering valuable advice, for his support during the whole period
of the study. I would like to express appreciation and special thanks to Assistant Professor Dr.
Asad Hussain Shah and Dr. Shahid iqbal Awan department of PB&MG for providing assistance
and sincere guidance in research. Very special thanks to my colleagues, Saira Ishaq, Nazia
Rahman, Sajeela Ahmed and Sameera Rehman for their help and cooperation during my
research work.
I would like to express appreciation to my beloved Husband for his continuous support and
encouragement to finish this work. I do not have words to acknowledge his contribution during
the evolution of this write up.
Shazia Arif

32
32
ABSTRACT
Plants are enormously resourceful biochemical units, capable of synthesizing almost all complement of
indispensable dietetic micronutrients; though, these are disproportionately scattered among special plant parts. Sea
buckthorn (Hippophae rhamnoides L.) is a fruit berry plant known for its therapeutic and nutraceautical
exceptionality. It is enriched with storming range of nutrients in its berry, seed, leaf and bark included vitamins,
essential oils and minerals. Most of our food produce are often undersupplied with most of these nutrients.
Functional candidate’s micronutrient genes from sea buckthorn were therefore chosen from the pathway enzymes
known to be involved in vitamins biosynthesis and metabolism. Specific gene primers were designed and RT-PCR
and PCR amplification was carried out to amplify desired gene fragments. The list of cDNA cloned and sequenced
included genes coding for enzymes in the metabolism and synthesis of Ascorbate, Folate and vitamin A.
Differences and similarities were found in homology and phylogenic evaluation with genes from other plant
species.
On the first part the amplification and cloning of full length cDNA of ascorbate oxidase gene was described. A full
length coding sequence containing a unique fragment of 2158 bp as compared to tomato gene sequence was isolated
and cloned. There was a difference in length of new gene sequence as compared to reference gene sequence with
87% gene homology. The relative abundance of the total RNA coding for Hr-AO was estimated using semi-
quantitative RT-PCR with maximum transcript accumulation in green leaf and young green fruit tissues.
Second part of the study was concerned with the folate (Hydroxymethyldihydropterin pyrophosphokinase–
dihydropteroate synthase) linked in the pathway of folate biosynthesis from sea buckthorn. The target gene HPPK-
DHPS was successfully isolated and cloned and its structure was compared with other plants. The sequence
analysis revealed that this novel genomic locus is 2354bp in size. The coding region is interrupted by a single large
intron. Its length is 1539bp which is similar to its ortholog in tomato. Expression profile of HrHPPK-DHPS with

33
33
semi-quantitative RT-PCR revealed the higher accumulation of transcripts in leaf and fruits tissues. In this study
an attempt was also made to compare the 3D homology modeling structure of new protein.
Third part of the research deals with carotenoids which produces yellow colors of fruits and vegetables and are
almost universally distributed in living things. Carotenoids are a chief source of vitamin A in humans nutritional.
Basic challenges were to study two important caroteniods pathway genes Lycopene β-cyclase and Phytoene
synthase from vitamin A rich plant “sea buckthorn” (Hippophae rhamnoides). The newly isolated full length
coding sequences of Hr-Lcyb and Hr-Psy showed homology with reference gene sequence. Despite the similarities
in nucleotide gene sequences several differences in structural domains were also found. Maximum transcript
accumulation is accordingly detected in a variety of plant tissues (leaf & Fruit), even though at trace level in some
tissues like seed and root. Awareness and identification about the value of these important micronutrient genes in
sea buckthorn plant will increases its long term value for genetic engineering in future.

34
34
Chapter 1
GENERAL INTRODUCTION
1.1 Sea buckthorn (Hippophae rhamnoides)
Sea buckthorn is well-known as nature’s most impartial fruit bearing plant. It is
considered as the “Holy Fruit of the Himalayas”. It has been treasured by native
Tibetans for centuries for its extensive nutritional values. This fruit is also
recognized as Sandthorn, Sandorn, and the "Wonder Berry" all over the
world. This fruit never stops to astonish with its marvelous nutritional base due to
high amount of vitamins.
Both medicinal and nutritional values of sea buckthorn, berries, leaves and seeds are
not authenticated. This medicinal plant needed to be searched of its hidden potential
for commercial crops biofortification using modern molecular biological approach.
Many important crops are lacking nutrients and most of the nutrients can be provided
by this plant. Many developments have been made in the application of molecular
biology to improve nutritients in plants.
The micronutrients deficiency is preventing about one third of world population from
reaching their physical and intellectual potential. Many varieties of crop with
increased amounts of essential vitamins and minerals have been developed due to
genetic engineering. This technique has also improved the profiles of nutraceautical
compounds. Much of the research into vitamins and minerals has been focused to
create new varieties of food crops to improve the diet of population in developing
countries. Biofortification is an economical approach to improve food qualities that
harmonize other technological and social involvement.

35
35
1.2 Origin and History
Sea buckthorn is a universal name that is particular to the winter hardy, deciduous
and dioecious spiny shrub or small tree with yellow, red or orange berry of the genus
Hippophae (Bailey and Bailey, 1978). This genus is in Elaeagnaceae family. It
consists of six species and ten subspecies, along with the most economically
important Hippophae rhamnoides L. generally identified as sea buckthorn (Rongsen,
1992). Northern areas of Pakistan have only one subspecies known as turkestanica
(Rongsen, 1996).
Sea buckthorn (Hippophae rhamnoides L.) is a versatile plant species native to
Europe and Asia. It has been used by humans for centuries that are written by
prehistoric Greek scholars such as Dioscorid. Leaves and branches of Sea buckthorn
were used for rapid weight gain and shiny coat for the horses, therefore this plant
was considered as healthy food by ancient Greece. That’s why; in Latin this plant is
given the name “Hippo”-horse, ‘phaoas’ to shine (Rongsen, 1992).
H. rhamnoides has a very wide division; it grows well on hilly areas, in valleys and
river beds, along sea coasts and islands, in small separated or large permanent pure
stands or in mixed stands with other plant species (Yao, 1994). The remaining
species in the genus have a quite restricted allocation and found only in china and
some nearest countries alongside the Himalaya Mountains (Rousi, 1971; Liu and He,
1978; Lian, 1988; Yu et al., 1989).
The genus Hippophae is dispersed between 270-690N latitude and 7ºW to 122ºE
longitude (Rousi, 1971; Pan et al., 1989; Yu et al., 1989). H. rhamnoides L. subsp.
turkestanica occur in North areas of Pakistan usually distributed all over the
karakarum ranges at altitudes of 2,000-4,200m (Gilgit, Hunza, Skardu, Nagar,

36
36
Shigar, Khaploo etc). Northern parts of earlier state of Jammu Kashmir are the centre
of origin of ssp. turkestanica (Ahamad and Kamal, 2002). Presently the reproduction
and adaptation of sea buckthorn have been tried in Euro Asia and North America
(Yao, 1994). It has been expected that there were about 3000 hectare of sea
buckthorn forest in Pakistan, with the annual production of 12002250 tones of sea
buckthorn fruit (Rongsen, 1996).
Sea buckthorn plant is a thorny, multibranched, dioecious, spinescent shrub attaining
size of 2-4 meter in tallness with solid branches shaping frequently symmetrical
round head. It has brown or black coarse bark and a thick grayish green crown
(Rousi, 1971). A tree like shape is frequently found as merely the bud on the outer
part of the plant germinates and branch. There is tremendous variability in height
starting from a little bush less than 50cm to a tree more than
20m tall in sea buckthorn plant (Rousi, 1994; Yu et al., 1989; Yao and Tigerstedt,
1994). It is considered a first woody species that habitate in open areas such as
dumped farmland, harsh environment, and rocky islands (Rousi, 1965; Yao and
Zhu, 1985; Yao, 1994).
1.3 Chemical composition and Nutritional value
Sea buckthorn berries are considered as vitamins C and E rich and nutrients rich such
as carbohydrates, organic acids, and amino acids. The fruit has more than 100 types
of bioactive substances, nutrients and more than 22 minerals. Variety of antioxidant
chemicals such as vitamins C and E, many carotenoids such as beta carotene,
flavonoid, and certain enzymes are present in Sea buckthorn berries. The berries
carry important components which are being used in therapeutic assignations. These

37
37
components are vitamins A, B1, B2, B6, C, and E, carotene, fatty acid, palmitolein
acids, palmitin, and β-sitosterol (Bernath and Foldesi, 1992).
Berries from Uzbekistan the protein content of Sinensis press juice ranged from 1.26
to 1.40% and was also reported as 9.1 to 9.4 % . Supercritical CO2 extracted press
juice gave a compactly clouded juice of significant cloud firmness (Beveridge et al.,
2004). Sea buckthorn berries are also having plenty of several amino acids. Berries
also have low levels of the sugar, alcohols, mannitol, xylitol and sorbitol. There are
several mineral elements present in sea buckthorn seeds, juice and berries. Sea
buckthorn juice contains 24 chemical elements. A plentiful amount of proteins and
free amino acids is present in Sea buckthorn. Sea buckthorn fruit contains a total of
18 amino acids (Zhang et al., 1989; Mironov, 1989).
Major elements are phosphorus, nitrogen, iron, manganese, calcium, boron, silicon
and aluminum (Tong et al., 1989; Wolf and Wegert, 1993; Zhang, et al., 1989).
Potassium is the most plentiful of all the elements found in juice or berries (Chen,
1988; Kallio et al., 1999; Tong et al., 1989; Zhang, Yan et al., 1989).
Sea buckthorn berries are acidic in nature has exotic flavor, but have an excellent
potentiality for producing different refined products like syrup, beverage, squash,
jam and jellies. Astringent quality of sea buckthorn juice or pulp can be minimized
by blending of juice or pulp with other fruits like orange, apple and papaya in
different ratios. Many sea buckthorn products are available in the market like oil,
juice, and food additives, jellies, candies, cosmetics, and shampoos (Schroeder and
Yao, 1995).

38
38
Sea buckthorn berry oil support tissue and wound healings because it contains 35%
of the rare and valuable palmitoleic acid which is known as component of skin fat.
The seed oil is has high oleic acid content (17%), omega-3 (34%) and omega-6
(31%). Omega-3 and omega-6 are present in one to one ratio. Because of one to one
ratio the two omegas is significant because they self check each other and control
thousands of metabolic functions by prostaglandin pathways. Fatty acid composition
is different in the seed and pulp oil. Pulp oil extraction contains saturated fatty acids;
mainly palmitic acid and palmitoleic acid (Kallio et al., 2002).
Sea buckthorn berries have very important components known as berries oils. In
general, the oil from the pulp/peel portion is combined due to the complexity
concerned with taking apart. High total lipid content like tocopherols, tocotrienols,
carotenoids, and omega-3 and omega-6 fatty acid have identified both in seeds and
berry pulp. Sea buckthorn seed and pulp oils composition is different according to
the subspecies, its origin, cultivating activities, harvesting time, and the extraction
techniques of berries (Yang and Kallio, 2002).
reported that sea buckthorn oil is rich in essential fatty acids, antioxidants and
vitamins contents therefore it is being widely used as an anti bacterial,
antiinflammatory, analgesic, regeneration of tissues and to maintain women health.
Residue left from squashed seed oil and pulp juice can be used in medicine and
cosmetics. Sea buckthorn berries is rich in bioactive phytochemicals, therefore it is
widely used in the industries of Pakistani northern areas, China and Russia
cosmoceutical and nutraceautical products (Zeb, 2004).

39
39
1.4 Vitamins
1.4.1 Ascorbic Acid
Sea buckthorn is amongst the most nutritive and vitamins affluent plant. Sea
buckthorn fruit has plenty of nutrients like carbohydrates, amino acids, organic acids
and vitamins. Vitamin C in fruits usually varies from 200 to 1500 mg/ 100 g, which
is 5 to 100 times more than any other specific fruit like apple. Sea buckthorn berries
are popular for their very high levels of vitamin C which varies from 360 mg/100 g
of berries for the European subspecies rhamnoides (Plekhanova, 1988;
Rousi and Aulin, 1977; Wahlberg and Jeppsson, 1990, 1992; Yao et al., 1992) to
2500 mg/100 g of berries for the Chinese subspecies sinensis (Yanget el al., 1999;
Yao and Tigerstedt, 1994; Zhao et al., 1991). The oil composition of the seeds
(7.3% w/w dry basis), pulp (1.7% w/w), and whole berry in wild sinensis berries
(2.1% w/w) was also investigated (Yang and Kallio, 2001).
Sea buckthorn fruit from Portland contain rich vitamin C substance ranging from
114 to 1550 mg/100 g with a normal substance (695 mg/100 g) which is 12 times
more than oranges, grading fruit of sea buckthorn amongst the highest in vitamin C
content. Strawberry, kiwi, orange, tomato, carrot and hawthorn have lower
concentration of vitamin C than sea buckthorn fruit (Dharmananda, 2004). Sea
buckthorn berries pulp from india have 223.2 mg/100 g of vitamin C. Roughly 75%
of the vitamin C in the Sea buckthorn berries pulp retain in the juice throughout the
process, resulting in 168.3–184.0 mg/100 g of vitamin C in the last patent juice
(Arimboor et al., 2006). The turkestanica sea buckthorn fruit has vitamin C
substance in the range of 200 to 1500 mg/100 which is 5 to 100 times greater than
any other fruit or vegetable (Ahmad & Kamal, 2002).

40
40
1.4.2 Caroteniods and Tocopherols
Carotenoids attained significance amongst biologically active agents to a great
extent, as they have numerous actions such as anti-oxidant, anti-mutagenic and anti-
tumour (Britton et al., 2009). Earlier research on sea buckthorn berries has shown
variation in constitution and content of carotenoids (Andersson et al., 2009; Bal et
al., 2011). Forty-one different carotenoids have been found in various varieties, with
zeaxanthin, b-cryptoxanthin, and b-carotene as the major ones (Andersson et al.,
2009; Raffo et al., 2004). Alpha-Carotene, c-carotene, dihydroxy-b-carotene,
lycopene, and canthaxanthin are found to be the minor ones (Yang & Kallio, 2005).
The presence of carotenoid esters has been found in sea buckthorn berries (Giuffrida
et al., 2011; Pintea et al., 2005).
The carotene substances of berries vary from 30 to 40 mg/100 g (Bernath and
Foldesi, 1992, Wolf and Wegert, 1993). The carotenoids content generally increases
during ripening (Andersson et al., 2009). Plenty of lipoproteins and pigments are
present in tissue layers and the fleshy mesocarp of Sea buckthorn fruits.
Carotenolipoprotein compound are found mainly in fruit membranes where polar
lipids might function as link compounds among the polar (protein) and nonpolar
(carotenoids) moieties (Pintea et al., 2001). Carotenoid substance is the main factor
by which sea buckthorn oil is traded commercially (Beveridge et al., 1999).
Carotenoids differ extensively depending on the oil resource; vary from 314 to 2139
mg/100 g for Chinese grown sea buckthorn (Zhang and Xu., 1989). Fleshy tissue
and fruit oils are rich reservoir of carotenoids as can be observed by their colors, at
about 900–1000 mg/100 g for Pamirs sea buckthorn (Mironov, 1989). Berries have
Vitamin E concentration up to 160 mg/100 g (Zhang et al., 1989; Ma and Cui, 1989;

41
41
Eliseev, 1989; Schapiro, 1989, Wahlberg and Jeppsson, 1990, 1992). Sea buckthorn
is also enriched with good quantity of water and fat soluble vitamins. In sea
buckthorn vitamin E is more than that of maize, wheat embryo, soybean and
safflower (Bernath & Foldesi, 1992). Sea buckthorn is also enriched in B1, B2, K
and bioflavonoids (Bekker & Glushenkova, 2001).
The seed oil has more than 95% of the retrievable tocopherols. It is present at a high
concentration (140 mg/100 ml). 1% phytosterols and small amounts of tocotrienols
are also present (Parimelazhagan et al., 2005. Sea buckthorn seeds have Radical-
scavenging proanthocyanidins (Fan et al., 2007). Sea buckthorn is considered as a
useful medicine for many diseases because it contains considerable amounts of
vitamin E and β-carotene (Ahmad & Kamal, 2002).
The vitamin E in sea buckthorn berries is 160 mg/100 g (Eliseev, 1989; Ma & Cui,
1989; Wahlberg and Jeppsson, 1990, 1992; Zhang, et al., 1989). Juice of Chinese
varieties has 162–255 mg/100 g (Zhang, et al., 1989) while Pakistani varieties pulp
has 481 mg/100 g (Zeb, 2004a). Seeds of Chinese varieties also have 40.1–103.0
mg/100 g vitamin E (Ma et al., 1989).
Many forms of Sea buckthorn leaves and Berries showed up total carotenoid
components which range between 3.5 and 4.2 mg/100 g dry weight in leaves, and 53
and 97 mg/100 g dry weight in berries. The carotenoid di-esters characterize the key
portion among berry forms include zeaxanthin di-palmitate as main compound,
while leaves have free carotenoids only like viollaxanthin, neoxanthin, lutein, and b-
carotene. Component study showed the appropriate carotenoid biomarkers feature
for the Carpathians’ sea buckthorn from Romania with contribution to their
taxonomical catogorization and validity identification (Raluca et al., 2014).

42
42
1.4.3 Flavonoids
Sea Buckthorn’s flavonoid “quercetin,” is recognized as a natural antihistamine and
anti-inflammatory, and studies suggest that it may relieve hay fever, eczema,
sinusitis, and asthma, and may help protect against heart disease and certain cancers
(Tamara et al., 2010) 11 to 22% crude protein, 3 to 6% of crude fat and some
flavonoides are present in its leaves (Wang et al., 2000; Rongsen, 1996). The
flavoniods mg/100 g in different sea buckthorn species are listed in table 1.3. It was
found that these phenolics are either quercertin derivative or isorhamnetin
derivatives. Sea buckthorn contains quecertin, gallic acid, catechin, epicatechin and
other small phenolics in its composition (Hakkinen et al., 1999). To a great extent
the quercetin compound represent as 3 o glycosides with glucose, rutinose or
rhamnose sugars making up the glycoside (Rosch et al., 2004b). It is also found that
the flavonoids content is as maximal as 1000 mg/100 g (Tian, 1985; Wang, 1987;
Xu, 1956).
Sea buckthorn fruits are rich in total flavonoid contents. Fresh fruit has highest
flavonoids according to the ex-Soviet Union researcher, which is 854 mg/100 g,
whereas, the average flavonoids contents in fresh fruit is 354 mg/100 g according to
a Chinese researcher. Lots of study has shown that in sea buckthorn flavonoids
contents from elevated level of sea were maximum (Yuzhen and Fuheng, 1997).
1.4.4 Folate
Folate is a water-soluble vitamin B which is identified as advantageous to human
health. Folate is helpful to prevent neural tube defect in babies, improve a
cardiovascular disease that is caused by high plasma homocysteine and few types of

43
43
cancer. Sea buckthorn fruit is considered as high source of folate ranging 29 μg/100
g fresh weights (Marcus, 2005; Virendra S. 2006).
The whole folate substances of sea buckthorn fruit berries and juice analyzed ranged
from 29 up to 81 μg/100 g. In contrast sea buckthorn folate berries contents with that
of other berries, these berries including 81μg/100 g were almost more rich in folates
as rose hips, which confirmed the maximum folate contents of 96 μg/100 g amongst
berries investigated 5-Methyltetrahydrofolate is a methylated
derivate of tetrahydrofolate. (Strålsjö et al., 2003; Gutzeit et al., 2008).
It was reported that folate contents varied significantly among species and also with
the origination of the plants tissues. For instance wheat germ has higher folate levels
whereas darker greens vegetable normally comprised much folate than roots such as
carrot and fresh fruits. Amongst the fruit plants, maximum concentration was
reported in orange and some other citrus fruits (Scott et al., 2000). However
divergence found in folate pathway genes between HPPKs and DHPs from different
species effectively suggested substantial variation in the genes evolution. The
association among folic acid accumulations in photosynthetic tissues is not
emphasized until now. It was found that the photo-respiratory process necessitates
both GDC and the SHMT folate dependant enzyme, which accumulated in the
mitochondria with rejuvenation. (Douce et al., 2001). It was reported in some plants
that folate is accumulated in cotyledon and embryo during the process of
germination, although higher HPPK-DHPS contents were only noticed in embryo.
The higher capability of developing embryo and meristematics tissue for
synthesizing and accumulating folate is associated with cellular metabolic process
and higher demands for nucleotide synthesis. These most important cellular

44
44
functions involved folate coenzymes with light playing important part in folate
biosynthesis in leaves (Jabrin et al., 2003).
1.5 Amino acids
In sea buckthorn fruit out of 22 known amino acids (Mironov, 1989; Zhang et al.,
1989), half of which are very important because they play a significant role in
different functions within our bodies like cells and muscles formation, production of
energy, loss of fats, and many functions of brain. Juice of sea buckthorn is enriched
in many free amino acids. Eighteen types of free amino acids in the juice of Chinese
sea buckthorn were found (Chen, 1988). Total amino acids content of Chinese sea
buckthorn which contains more apartic acid (426.6 mg/100 g) were also discovered.
Amongst these amino acids eight amino acids are very important for human body.
These amino acids are threonine, valine, methionine, leucine, lysine, trytophan,
isoleucine, and phenylalanine (Zhang et al., 1989; Chen, 1988).
1.6 Scope
An Adequate amount of micronutrients are one of the basic requirements for human
health in the daily diet (Caballero and Black, 2003). To overcome micronutrient
deficiency problem, the an ingredient is added for the purpose of enrichment and has
demonstrated to be very effective for certain micronutrients; for instance, salt
iodination or fortification of tap water and toothpaste (Darnton and Nalubola,
2002). For each of the micronutrient fortifying procedure is different; though, it will
possibly be harder for Fe owed to its rapid oxidations (Boccio and Lyengar, 2003).
Cereals fortification requires complicated techniques. A substitute method to fortify
cereals by food processing and agricultural management is aggregation of

45
45
micronutrients directly in cereals seeds using traditional breeding or genetic
engineering (Zimmermann and Hurrel, 2002).
Vitamin C is very important for heart function, immunity, connective tissue, and iron
consumption. Although most of the animals and plants can make Vitamin C, human
being is short of L-gulono-1,4-lactone oxidoreductase, which is necessary for the
final step in Vitamin C preparation. In human being fruits and vegetables are major
nutritional resource of vitamin C. Latest studies indicate that as Vitamin C uptake
is increased (from 60 to 200 mg d-1) health benefits are increased (Carr and Frei,
1999; Levine et al., 1999). Vitamin C is required in cell division method, regulation
and expansion (Smirnoff, 1996; Pignocchi et al., 2003).
Folate is considered a crucial nutrient in the human diet. Many physiologic upsets
like anemia and neural tube defects are found in neonates due to folate deficiency
(Lucock, 2000). Folate deficiency causes mental disorders such as psychiatric
syndrome and decreased congnitive functioning in old people (Calvaresi and Bryan,
2001; Hultberg et al., 2001). Folate is also considered to protect against heart
diseases and various cancer categories (Boushey et al., 1995; Brattstrom et al., 2000;
Lucock, 2000).
It was reported that of folate level can be improved in food plants through metabolic
engineering. Lack of Folate deficiency has philosophical influence on healthiness of
human beings effecting large population. This problem can be solved through
modification of crops with high contents of folate through genetic engineering.
Plants are rich in dietetic folate so this technique may help to resolve the worldwide
folate malnutrition. Presently some advance techniques are being assessed for the
improvement of micronutrients in plants with possible application of folate increase.

46
46
It was reported that vitamin B comprised of water-soluble enzyme cofactors. In
plants as well as in animals and microorganisms various metabolic processes depend
on derivatives of vitamin B where they play important role. Seven vitamins form this
complex: B1, B2, B3, B5, B6, B8 and B9 (folate). Humans lack enzymatic system
to all these seven B vitamins required in the human diet for proper nutrition. The
principle objective was to review the current information about the synthesis of
vitamin B in plants (Sanja, 2007).
The lack of nutrient caused several physical and intellectual problems. In developing
countries genetic engineering techniques like biofortification of cereals to cover
daily folate requirements is a striking approach to address malnutrition. In this study
wheat seeds were used to isolate folate genes 6-hppk/dhps, 4-amino-
4deoxychorismate synthase and folypolyglutamate synthetase and their homologues
were identified in rice genome. Gene expression studies confirmed de novo folate
production in growing wheat seeds and exclusively in transcripts of mature seeds
with potential to refill its own pool of vital glutamated folates during whole life cycle
(Shan and Robert, 2008). It was also found in literature that iron contents in cereals
and rice crops are very low. Most of these Fe contents are lost during processing of
grains. Folate deficiency is affecting large group of people around the world. The
approaches like food fortification and folate supplement intake programs are not
always productive. A substitute to solve this problem is through biofortification of
iron. Here, some direct genetic modification and some conventional breeding
techniques for rice biofortification for iron were addressed. Here the breakthrough
of new genes and QTLs related to folate biofortification is highlighted (Raul et al.,
2012).

47
47
Folate is needed for almost all living organisms as important cofactors. Among plant
produce cereals are the most extensively used but unable to add enough folate. It was
illustrated through GTP cyclohydrolase 1 mRNAs from growing tissues of wheat
seed, roots and leaves that de novo synthesis of folate is taking place all over the
wheat plant. Folate biosynthetic pathway is constantly active in developing wheat
seeds and it is important for folate production depending upon seed viability (Shane
et al., 2008).
Folate deficiency in the developing and developed countries, where it is responsible
for serious health problems among people was also studied. Currently some
progressive food fortification and folic acid supplementation were effectively used
modern approaches established. The genes and enzymes of folate synthesis are
sufficiently understood to enable metabolic engineering of the pathway and their
study consequences from engineering studies in plants are supporting (Samir et al.,
2008).
The presence of genes for different micronutrients at one place may have extra
benefit as the same promoter elements working at sea buckthorn mesocarp are active
during fruit ripening may have joint action and can help in the transport of many
genes in one go. Gene transformation of micronutrients in cereals by genetic
engineering techniques shows best results than traditional breeding techniques. Sea
buckthorn fruits are rich in micronutrients like Vitamin A, Vitamin C, E and Iron
which shows numerous genes related to the synthesis of such phytochemicals, which
are essential for the growth and health of human being. These micronutrient genes
can be used for the cereals biofortification.

48
48
1.7. Objectives
The main objectives of this doctoral dissertation are as follows:
To identify the proper stage of fruit development for gene expression via analysis of
genes involved in vitamin A, vitamin C and folate genes in different tissues of sea
buckthorn.
Isolation of mRNA from Sea buckthorn fruit berries to synthesize cDNA of these
micronutrient genes i.e. vitamin C, folate and vitamin A using gene specific primers
in RT-PCR.
Amplification, cloning and sequence analysis of these genes for their later
comparison with other genes in different species having potential for staple food
biofortification.

49
49
Chapter 2
ASCORBATE OXIDASE
Abstract
Ascorbic acid (vitamin C) is a well-known molecule for its nutritional importance.
However the major aspects of its metabolic processes and a few aspects of its
functions in plants are poorly understood. For instance, its biosynthetic pathways
were not definitely recognized although it attains milli-molar concentration in nearly
all tissues. Human beings and some animals are dependent on ascorbate contents in
diet. A Functional candidate gene known to be involved in ascorbate metabolism
was therefore chosen. A full length cDNA sequence of Ascorbate oxidase containing
a unique fragment of the 2158 bp as compared to tomato cDNA sequence (1737 bp)
has been amplified and cloned through RT-PCR based cloning of cDNA. The amino
acid residues encoded by Hr-AO was 719aa. There was a difference in length of
new cDNA sequence as compared to cDNA with 87% gene homology. Expression
analysis of this gene sequence from six different tissues including vegetative bud,
seed, shoot apex, green leaves, green fruit and mature (orange red) fruits has showen
maximum transcript accumulation in green leaf and young green fruit tissues. This
is the first report on description of relationship among the expression of Hr-AO and
fruits development in such type of bush plant. This novel gene isolated from sea

50
50
buckthorn will help to understand the regulatory role of this enzyme in ascorbic acid
metabolism. The investigation suggested that this pathway gene could merely
contribute toward Ascorbic acid function or may be specific for further genetic
engineering of crops.
2.1 INTRODUCTION
Vitamin C or ascorbic acid (ascorbate) is an indispensable nutritive element for
living organisms. Vitamin C relates to numerous vitamers having activities of
vitamin C. Ascorbate oxidase belongs to class of multi copper enzyme catalyzing
the oxidation of ascorbic acid to dehydroascorbic acid. Ascorbate oxidase is a cell
wall localized enzyme that utilizes oxygen for oxidation of ascorbate (AA) to the
unstable radical monodehydroascorbate (MDHA) which quickly disproportionate to
give dehydroascorbate (DHA) and ascorbic acid, and hence added to regulation of
the ascorbic acid redox state (Vasileios et al., 2006). The organization of both
reduced and oxidized types of ascorbic acid is shown below (Fig. 2.1).
(a) (b)
Fig. 2.1: Structure of ascorbic acid (a) ascorbic acid (reduced form) (b)
dehydroascorbic acid (oxidized form).

51
51
Vitamin C is crucial for cardiovascular functions, immune cell development,
connective tissues, and iron consumption. However plants and most animal have the
ability to synthesize ascorbic acid whereas L-gulono-1,4-lactone oxidoreductase
enzyme is absent in humans which is required for the last stage in AsA synthesis.
Because AsA cannot store in the body, the vitamin should be attained frequently
from dietary origins. Fruits and vegetables constitute the main dietary sources of
ascorbic acid in humans and current reports suggested that better
AsA consumption (from 60 to 200 mg /day) may impart health benefit (Carr, et al.,
1999; Levine et al., 1999).
The isolation and sequencing of genes encoding AO were described in many plants.
Ascorbate oxidase is an enzyme of cell wall and their mRNA coded for a principal
signaling sequences distinctive of extra-cellular protein (Esaka et al., 1990, Ohkawa
et al., 1989). The member of Cucurbitaceae family such as pumpkin, cucumber,
squash, zucchini and melon are naturally occurring richest source of AO. The
extensive biochemical and expression studies were carried out in these species
(Carvalho et al., 1981, Esak et al., 1990, 1992; Lee and Dawson,
1973; Moser and Kanellis, 1994; Nakamura et al., 1968). Most importantly vitamin
C is an antioxidant having significant role in regenerating vitamin E from oxidize
forms (Carr and Frei, 1999; Bruno et al., 2006). AO expressions are regulated
through composite transcription and translation control (Esaka et al., 1992). The
activities and expressions level of ascorbate oxidase are tightly associated to cell
development (Kato and Esaka, 2000).
Vegetables and fruits are important constituents of the daily diet which contributed
carbohydrates especially dietetic fiber, vitamin and mineral to the body. Vitamin C

52
52
is commonly observed in several fruits and vegetables (Deman, 1973). This is water-
soluble having antioxidant property well known for health and suitable functions of
the human body (Benzie, 1999; Davey et al., 2000). It controls several syndromes
like scurvy and also has a tendency of preventing several contagious diseases,
including viral and bacterial diseases. This is also essential for curing injuries, burns
and cracked bones. This vitamin is necessary for the production of all connective
tissues (Heimann, 1980). Additionally the foods ample in fresh fruits and vegetables
are defensive against chronic, degenerative diseases (Joshipura et al., 1999; Lampe,
1999; Cox et al., 2000)
Ascorbate (AA) is the richest antioxidant found in plants and contributed mainly to
cell redox state (Smirnoff, 2000). The largest part of the AA is contained in the
cytoplasm; almost 10% of the AA contents of the total leaf is transferred and localize
in the apoplastic region, where it was present in millimolar concentrations (Noctor
and Foyer, 1998). Apoplastic ascorbic acid is supposed to characterize firstly
protection against extraneous oxidants causing potential damage. These also have a
significant function to mediate reaction to stress rendering an improved oxidative
load (Barnes et al., 2002; Pignocchi and Foyer, 2003).
In plants ascorbic acid makes a central part in the production of hydroxyprolinerich
proteins (collagen development) that are constituents of the plant cell wall.
Additionally these have important role in cell growth and divisions. Currently the
ascorbic acid biosynthetic pathway explained in plants looks quite diverse from that
in animals (Wheeler et al., 1998). The comparison of vitamin C among different
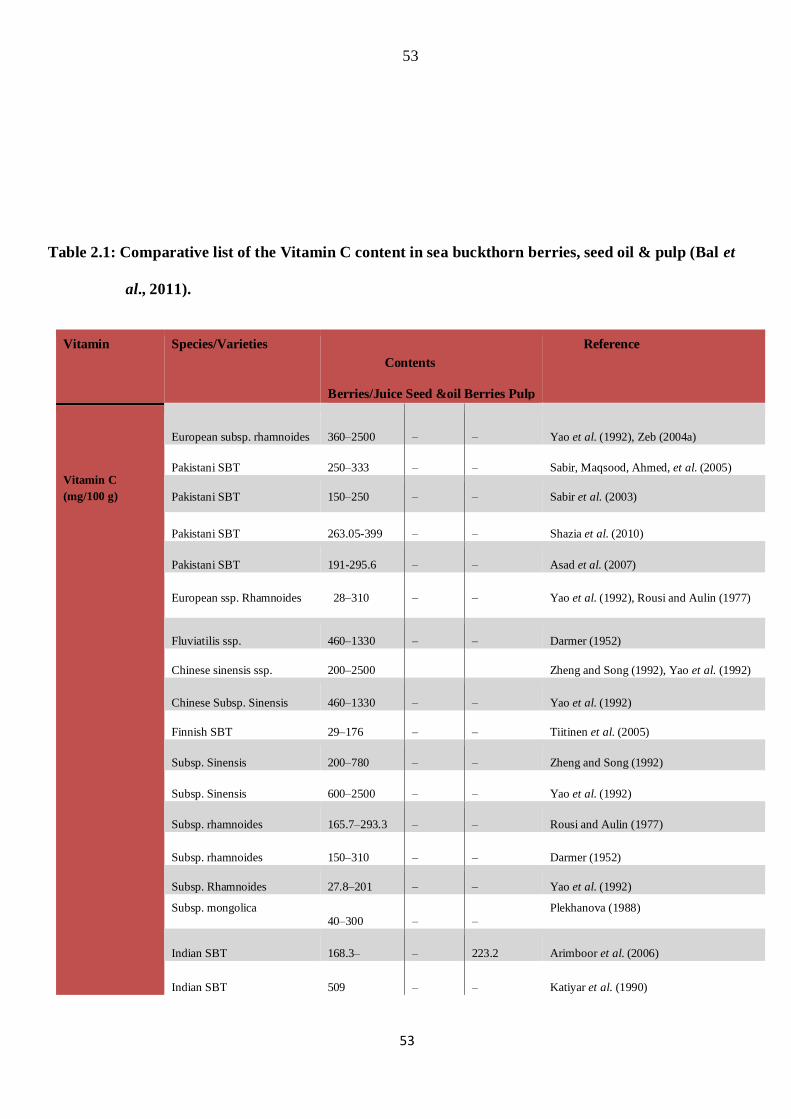
species and varieties are shown in table 2.1.
53
53
Table 2.1: Comparative list of the Vitamin C content in sea buckthorn berries, seed oil & pulp (Bal et
al., 2011).
Vitamin Species/Varieties
Contents
Berries/Juice Seed &oil Berries Pulp
Reference
Vitamin C (mg/100 g)
European subsp. rhamnoides 360–2500 – – Yao et al. (1992), Zeb (2004a)
Pakistani SBT 250–333 – – Sabir, Maqsood, Ahmed, et al. (2005)
Pakistani SBT 150–250 – – Sabir et al. (2003)
Pakistani SBT 263.05-399 – – Shazia et al. (2010)
Pakistani SBT 191-295.6 – – Asad et al. (2007)
European ssp. Rhamnoides 28–310 – – Yao et al. (1992), Rousi and Aulin (1977)
Fluviatilis ssp. 460–1330 – – Darmer (1952)
Chinese sinensis ssp. 200–2500 Zheng and Song (1992), Yao et al. (1992)
Chinese Subsp. Sinensis 460–1330 – – Yao et al. (1992)
Finnish SBT 29–176 – – Tiitinen et al. (2005)
Subsp. Sinensis 200–780 – – Zheng and Song (1992)
Subsp. Sinensis 600–2500 – – Yao et al. (1992)
Subsp. rhamnoides 165.7–293.3 – – Rousi and Aulin (1977)
Subsp. rhamnoides 150–310 – – Darmer (1952)
Subsp. Rhamnoides 27.8–201 – – Yao et al. (1992)
Subsp. mongolica 40–300 – –
Plekhanova (1988)
Indian SBT 168.3– – 223.2 Arimboor et al. (2006)
Indian SBT 509 – – Katiyar et al. (1990)
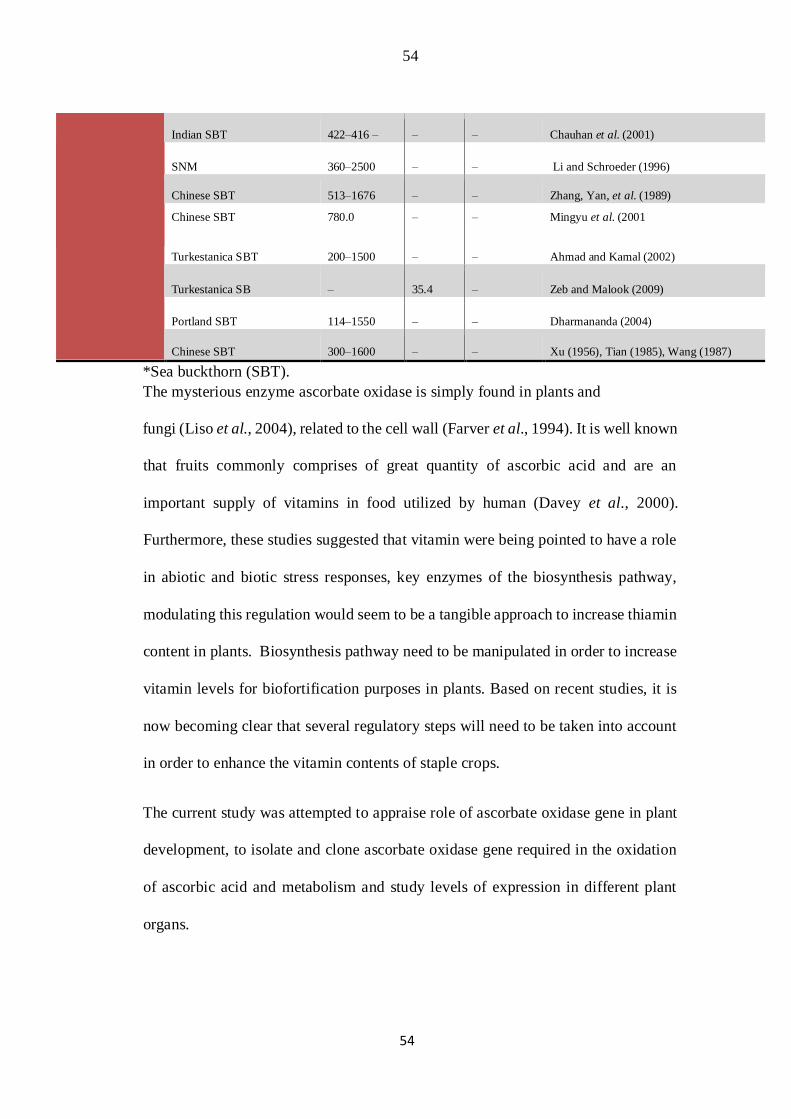
54
54
Indian SBT 422–416 – – – Chauhan et al. (2001)
SNM 360–2500 – – Li and Schroeder (1996)
Chinese SBT 513–1676 – – Zhang, Yan, et al. (1989)
Chinese SBT 780.0 – – Mingyu et al. (2001
Turkestanica SBT 200–1500 – – Ahmad and Kamal (2002)
Turkestanica SB – 35.4 – Zeb and Malook (2009)
Portland SBT 114–1550 – – Dharmananda (2004)
Chinese SBT 300–1600 – – Xu (1956), Tian (1985), Wang (1987)
*Sea buckthorn (SBT).
The mysterious enzyme ascorbate oxidase is simply found in plants and
fungi (Liso et al., 2004), related to the cell wall (Farver et al., 1994). It is well known
that fruits commonly comprises of great quantity of ascorbic acid and are an
important supply of vitamins in food utilized by human (Davey et al., 2000).
Furthermore, these studies suggested that vitamin were being pointed to have a role
in abiotic and biotic stress responses, key enzymes of the biosynthesis pathway,
modulating this regulation would seem to be a tangible approach to increase thiamin
content in plants. Biosynthesis pathway need to be manipulated in order to increase
vitamin levels for biofortification purposes in plants. Based on recent studies, it is
now becoming clear that several regulatory steps will need to be taken into account
in order to enhance the vitamin contents of staple crops.
The current study was attempted to appraise role of ascorbate oxidase gene in plant
development, to isolate and clone ascorbate oxidase gene required in the oxidation
of ascorbic acid and metabolism and study levels of expression in different plant
organs.

55
55
2.2 MATERIALS AND METHODS
Wild sea buckthorn (Hippophae rhamnoides sp. Sinensis) berries were grown at the
research station at National Institute for Genomics and Advanced Biotechnology
(NIGAB) Islamabad. For nucleic acid extraction, the tissue samples were harvested
from plants at different bud, leaf, fruit and seed developmental stages. Fruits were
harvested once or twice a week during the ripening period from sea buckthorn
nursery, frozen immediately in liquid nitrogen and placed at -80ᴼC until processed
for nucleic acid extraction and eventually expression analysis.
2.3 Designing of Primer
Nucleotide sequences of ascorbate oxidase genes were retrieved from National
Center for Biotechnology Information (NCBI) database. The gene specific primers
were designed from the conserved region of acorbate oxidase gene for the
amplification of full length coding sequence of AO cDNA. Primers for expression
analysis through RT-PCR were designed from newly isolated sequences of AO
cDNA (Table. 2.2).
Table 2.2: Detail of primers used in this study for H. rhamnoides AO cDNA cloning,
expression analysis (RT-PCR) and Gateway cloning.

56
56
2.4 RNA isolation protocol (TRIzol® Reagent)
The sea buckthorn fruits and leaves samples were homogenize in liquid Nitrogen at
room temperature. About 0.1 g of tissue was transferred in ice cold micro centrifuge
tubes. About 500 ul RNA reagent was added, vortexed. It was incubated at room
temperature for 5 min. For this period of incubation micro centrifuge tubes were kept
horizontally. The Samples were centrifuged for 2 min at 12800 rpm at room
temperature, and supernatant was transferred to the new tube. 100 ul of 5M Nacl was
added and mixed for a while, and then 300 ul of chloroform was also added and
mixed thoroughly. The samples were then centrifuged at 4oC for 10 min at 12800
rpm. The uppermost aqueous phase was transfer to new micro centrifuge tube and
equal volume of isoproponal, was added, mixed and kept at room temperature for 10
min. Centrifuged again for 10 min at 4oC at 12800 rpm. The

57
57
RNA pallet was washed with 70% ethanol. The RNA was stored at -80oC for RTPCR
experiments.
2.5 Rapid Amplification of cDNA Ends (RACE-PCR)
This good quality total RNA was utilized for the synthesis of cDNA with AMVRT
reverse transcriptase enzyme. In this case cDNA synthesis is primed using oligo (dT)
primer I, which aliquots (1 µl of a 20 µl reaction mix) of this reaction are subjected
to a PCR reaction using 5′-primer and a gene-specific reverse primer (GSRP)
designed for investigation. The reverse primer from the gene specific primer pair
AO-R 5՜ TTATAGAATTTAAGGCCTGTGGAA 3՜ was used. The next
procedure is made optimize to construct cDNA for further reactions.
Reagent Concentration
Total RNA 4 µl
Gene specific Primer(Reverse primer) 1 µl
Nuclease free water 8-9 µl 8-9 µl
These ingredients were mildly mixed and centrifuged briefly for few seconds and
incubated at 65oC for 5 minutes. Chilled with ice for 2-3 min. Then following
components were added in the specific order below.
Reagent Concentration
5x AMV RT buffer 4 µl
Ribolock TM RNAase inhibitor 0.5 ul
dNTP Mix, 10mM each 2 ul
AMV Reverse transcriptase 0.5 ul
Total volume 20 ul
58
58
It was mixed gently and centrifuged briefly. Then mixture was incubated for 60min
at 50oC. The reactions were terminated by heating up for 5 minutes at 85oC. This
cDNA was good enough for further experiments.
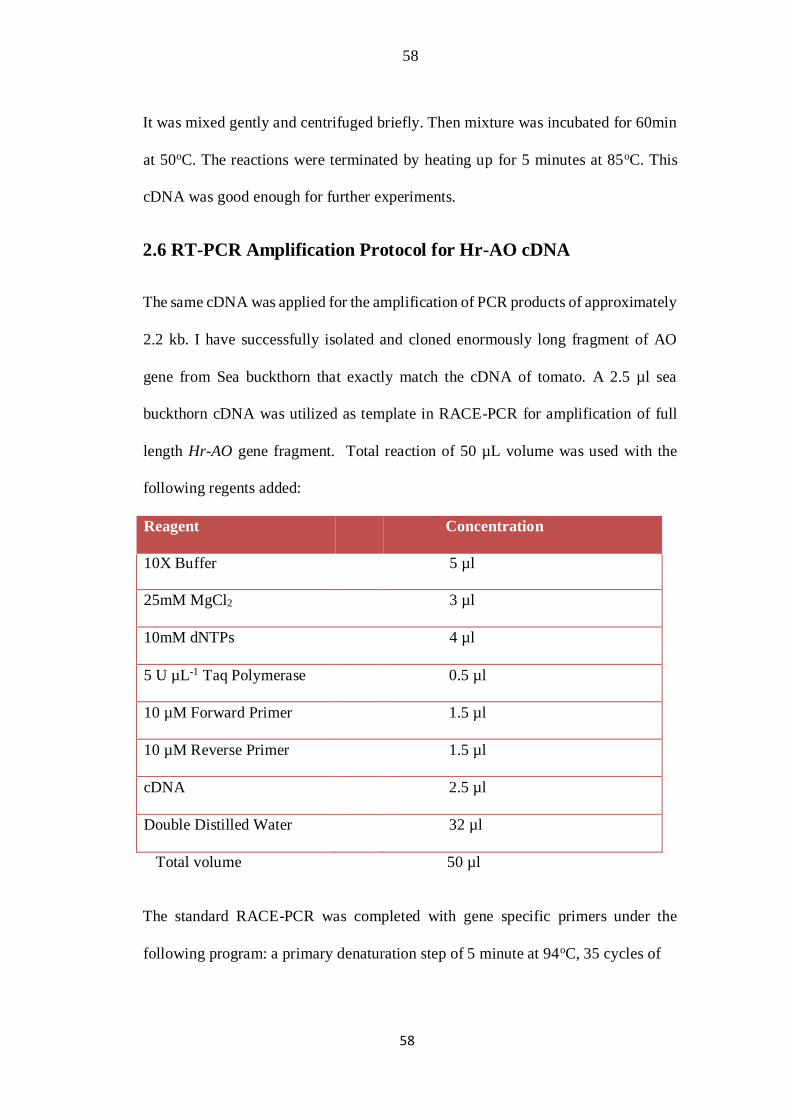
2.6 RT-PCR Amplification Protocol for Hr-AO cDNA
The same cDNA was applied for the amplification of PCR products of approximately
2.2 kb. I have successfully isolated and cloned enormously long fragment of AO
gene from Sea buckthorn that exactly match the cDNA of tomato. A 2.5 µl sea
buckthorn cDNA was utilized as template in RACE-PCR for amplification of full
length Hr-AO gene fragment. Total reaction of 50 µL volume was used with the
following regents added:
Reagent Concentration
10X Buffer 5 µl
25mM MgCl2 3 µl
10mM dNTPs 4 µl
5 U µL-1 Taq Polymerase 0.5 µl
10 µM Forward Primer 1.5 µl
10 µM Reverse Primer 1.5 µl
cDNA 2.5 µl
Double Distilled Water 32 µl
Total volume 50 µl
The standard RACE-PCR was completed with gene specific primers under the
following program: a primary denaturation step of 5 minute at 94oC, 35 cycles of

59
59
94oC for 60s, 59oC for 60s and 68oC of 165s, followed by last extension step of 68oC
for 10 min. Furthermore 1% agarose gel was used for the examination of amplicon
and photographs were taken. These amplified fragments were gels purify and
sequencing was carried out to validate the target sequence.
2.7 Gene purification
The total PCR products was first run on 2% high resolution agarose gel and then
purify by PCR GeneJET PCR Purification Kit (K0701). The following purification
protocol was used for Gel elution.
Gel pieces comprising the DNA fragments were excised with clear scalpels. To
reduce the gel volume the DNA was sliced as closer as feasible. Then slices were
placed in a pre weighted 1.5 ml centrifuge tube and weighed. The purified fragment
for cloning reactions was avoided from UV light damages. UV exposure was
minimized by putting the slices of gene fragment on a glass plate under UV exposure.
The binding buffer 1:1 was added to the gel slices. To melt the agarose gel fully the
mixture was incubated for 10 minutes at 50-60oC. These tubes were inverted and
shaken after few minute to dissolve it properly. When gel was completely dissolved
its color became yellow. Then 800 µl of the soluble gel mixture was poured out into
GeneJET™ purifications columns and centrifuge for one minute. The flow through
was disposed off and column was placed again into that collection tubes.
Additionally 100 µl of binding buffer was added to the purified gene fragments in
column used for sequencing purposes. Centrifuge it again for one minute and
discarded the flow through. The column was set back in the respective collection
tube.

60
60
It was mixed with 700 µl of wash buffer and was centrifuge again for one min. The
liquid buffer was discarded from the flow through and column was set back into the
same collection tube. The emptied column was centrifuged once more for one minute
to completely remove the remaining wash buffer. A new 1.5 ml microcentrifuge tube
was taken into which GenJET™ purifications column was shifted. Finally Elution
Buffer (About 50 µl) was transferred into the central membrane of the column. It
was Centrifuge for one min to elute DNA. The column was discarded and purified
DNA was stored at -20oC. The purification product was transported to MACROGEN
(Korea) intended for sequencing.
2.8 Gene Cloning protocol
2.8.1 TA cloning Vector
TA cloning is a commonly used lab technique for sub-cloning purposes without use
of restriction enzymes. It is much easy and faster as compared to conventional sub-
cloning methods. The following vector was chosen for cloning purpose (Fig.
2.2).
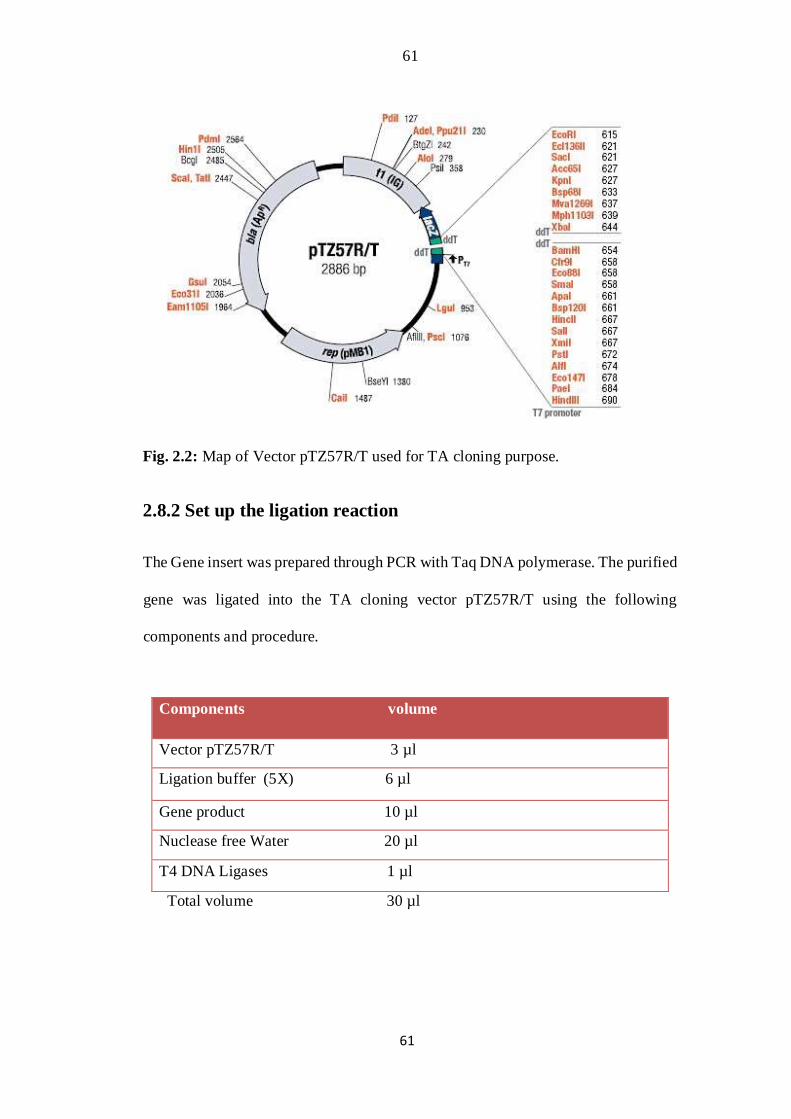
61
61
Fig. 2.2: Map of Vector pTZ57R/T used for TA cloning purpose.
2.8.2 Set up the ligation reaction
The Gene insert was prepared through PCR with Taq DNA polymerase. The purified
gene was ligated into the TA cloning vector pTZ57R/T using the following
components and procedure.
Components volume
Vector pTZ57R/T 3 µl
Ligation buffer (5X) 6 µl
Gene product 10 µl
Nuclease free Water 20 µl
T4 DNA Ligases 1 µl
Total volume 30 µl

62
62
The whole mix was vortexed in brief following centrifugation for 3-5s. These
ligations were incubated at 22oC for 60 minutes. For maximum number of
transformants it was incubated overnight at 4oC.
2.8.3 Host Cells
In our cloning experiments E. coli strain DH5α was used which is well-suited for
lacZ blue/white screening processes. It is quite easier to transform with recovery of
high-quality plasmids from transformed colonies. The following procedure was
applied to prepare electro-competent cells of DH5α strain.
2.8.4 Electroporation of E. coli:
LB-medium (1 L)
The E. coli cells were grown on LB liquid medium. For 1 L of LB media, following
ingredients were mixed in a Flask container with a stir bar until everything was
dissolved.
Components To make 1L
Tryptone 10 g
Yeast extract 10 g
NaCl 5 g
Water, nuclease-free 950 ml
Total 1000 ml
The pH of the medium was adjusted to 7.0 with 1N NaOH and volume was brought
up to one litter. The media was then autoclaved for twenty minutes at 15 pounds per

63
63
square inch. It was allowed to cool down to 55°C at room temperature. The
antibiotics like ampicillin (50 µg/mL) were poured if required. The media was stored
at room temperatures or +4°C.
Stock solution (Amp. 50 mg/ml)
Ampicillin (2.5 g) was dissolved in 50 ml deionize H2O. It was filter sterilized and
store in number of aliquot (50 μl) at -20°C for later use.
X-Gal (20 mg/ml)
X-Gal 200 mg was dissolved in 10 ml N,N-dimethylformamide and was stocked at
-20°C in darkness.
IPTG (100 mM)
About 1.2 grams of IPTG were dissolve in 50 ml of deionize water. It was filter
sterilized, aliquoted and stored at 4°C. About 50 μl per plate was used.
LB plate Preparation (Antibiotics)
Once the LB medium was prepared and autoclaved, it was cooled to
approximately 55°C. After that 1ml stock solution of ampicillin (50 mg/ml), 50 μl
of X-Gal (20 mg/ml) and 50 μl of IPTG 100 mM were added. The media was
smoothly mixed and pour in Petri-plates. Then plates were dried and opened
underneath UV light for thirty minutes at room temperature and stored at +4°C in
darkness.
2.8.5 E. coli cells Preparation for Electroporation.
Procedure
For preparation of electro-competent cells 5 ml LB media was poured into falcon
tube with 5 ul of ampicillin added. A single colony from freshly grown DH5α LB

64
64
plate was used to inoculate this media and cell were grown at 37oC overnight with
continuous shaking. The following procedure was followed:
• The overnight grown culture was diluted with 500 ml LB media in a 1L Flask
(OD˷0.2).
• The cells were grown for two to three hours at 37oC with vigorous shaking until the
cells reached to OD600˷0.2 to 0.4. The culture was chilled on ice and transferred to
centrifuge tube.
• The cell pallet was harvested by centrifuging at 8500 rpm at 4oC for 20 minutes in
pre-cooled sterile centrifuge tubes. For each experiment autoclaved bottles were
used.
• The pellet was resuspended with 350 ml ice cold H2O/500 ml. Pellet was dissolved
by continuous vortexing (gently) and pipetting until cells became resuspend.
• The cell suspension was centrifuged at 85,00 rpm for 20 min and supernatant was
removed cautiously as soon as possible.
• The cell pellet was resuspended/washed next time in 250 ml sterilized chilled water
and dissolved by the use of similar procedure as depicted before. The cell
suspensions were centrifuged at 7000 rpm for 20 minutes at 4oC.
• The cell pellet was resuspended again in 20 ml of cold water in falcon tubes and
centrifuged at 5000 rpm at 4oC to harvest the cells.
• The clean cell pellet was then resuspended in 800 ul 7% DMSO/500 ml culture or
70% glycerol was used for 50 ul of cells.

65
65
• The final volume of about 50 ul aliquots of cells was made in pre-cooled eppies.
These competent cells were either utilized directly or kept frozen in aliquot in icecold
vials at-80oC.
• Electroporation of AO gene (construct) into E. coli strain DH5α.
• Transformation of the resulting plasmid (pTZ57R/T) with gene insert into E. coli
DH5α strain was mad using electroporation method of transformation. The
following material in various volumes was used in the experiments.
Material volume
Vector 2 µl
LBmedia liquid 1 ml
Cuvette (chilled) 1 -2 ml
Tips (sterilized) 1 -1000 µl
LB solid plates 25 ml/plate
Electroporator 1800 volts
The required number of micro centrifuge tubes, LB liquid media and electroporation
cuvettes were marked and Placed on ice. The electroporation machine was also set
at 1800 volts. Using a micro pipette about 1-10 µl of ligation mixture was mix with
50 µl competent cell and carefully transferred into ice-cold cuvettes with no bubbles
formed. It was made sure that cell deposited around base of cuvette. It was
Electroporated at 1800 volts by pressing the button two times after the pulse is
delivered. Immediately 1 ml of LB media shifted into cuvette, gently mixed with
pipetting twice up and down. It was then transferred to sterilize micro-centrifuge
tubes. It was kept at 37°C for 1-2 hour with vigorous Shaking (250 rpm). These cells
were diluted appropriately and 200-500 μl of cells was spread onto a pre heated LB
antibiotic plates. These plates were placed overnight in incubator at 37°C.
Consequently over 90% of cell colonies comprised the vectors having gene inserts.

66
66
Blue/white colony selection was used to identify the recombinants (clones). The
white colonies with recombinant plasmids were selected marked for colony PCR and
plasmid isolation
2.8.6 Colony PCR procedure
This protocol was considered for quickly screening of plasmids with insert
immediately from colonies of E. coli cells. These techniques were successfully
applied to find out inserts length with M13 primers. The positive colonies were
marked and further propagated by short strike of individual colonies on ampicillin
plates. The following colony PCR reaction was used for AO gene insert validation,
and reagents were mix together resting on ice with enzymes added at the end.
Reagent Concentration
10X Buffer 5 µl
25mM MgCl2 4 µl
10mM dNTPs 1 µl
5 U µL-1 Taq Polymerase 0.5 µl
M13 Forward Primer (10µM) 1 µl
M 13 Reverse Primer (10µM) 1 µl
Double Distilled Water 37.5 µl
Total 50 µl
The cool PCR reactions in tube were properly mixed and a smaller amount of each
colony was added. The selected colonies were crushed with a pipette tip and mixed
by pipetting up and down. A short strike could be made over the ampicillin culture
plate to save the individual colonies for re-propagation. This type of adequate mixing
resulted in cell breakdown completely by good yield.
The following PCR profile was used for colony PCR reaction.
• 1 cycle of 5 minutes at 95°C –to denature DNA and break cells.
• 34 cycles of 30 second at 94°C, 30seconds at 52°C and 2 minutes at 68°C.

67
67
• 1 cycle of 12 min at 68°C -last extensions step to ensure fully sized product.
These PCR amplifications were run on agarose gel for presence of expected product
size. After PCR confirmation individual colonies with positive confirmation were
cultured for plasmid DNA purification before analysis.
2.8.7 Plasmid DNA extraction
General protocol
The recombinant plasmids were purified from overnight bacterial culture using a
reliable Favorprep™ plasmid DNA extraction Mini Kit. The following procedure
was used.
• 1-4 ml of well-grown bacteria cultures were transferred to a micro-centrifuge tube.
• The bacteria were descended by centrifuging for 2 min at maximum speed and
supernatant was discarded completely.
• 200 µl of FAPD1 buffer (RNase A added) was mixed with pellet. Then
resuspended the cells completely by pipetting. No cell pellet was left visible after
resuspension of the cells.
• About 200 µl of FAPD2 buffer was added up. The tubes were inverted gently 10
times to lyse cells. Incubate it at room temperature for two minutes. These tubes may
be inverted continuously until lysate became clear. This step was preceded with 5
minutes.
• 300 µl of FAPD3 buffer was added and the tube was inverted 10 times
immediately but gently.
• Centrifuged for 5-10 min at full speed at 14000rpm. During centrifugation placed
FAPD column in collecting tube.

68
68
• Supernatant was transported carefully into FAPD column. Then Centrifuge it for 30
seconds. The flow-through was discarded and the FAPD column was placed back
in the collection tube. In adding it was make sure that no white pellet was transferred
into the column.
• 400 µl of W1 buffer was Add to FAPD column. Centrifuge it again for 30 seconds
and discard the flow through. The FAPD columns were put again in the collecting
tubes.
• 600 µl of wash buffer was added into FAPD column. Then centrifuged for 30
seconds then discarded the flow-through and the FAPD column was place back in
the collection tube.
• Then Centrifuge it for three more minutes for drying the column. It will assure the
removal of the remaining liquids completely that may hinder successive enzyme
reaction.
• FAPD columns were place into clean 1.5ml micro-centrifuge tubes.
• 50 ul-100 ul Elution buffer were add to central membranes of FAPD column. The
column was left standing for 2 min.
• Centrifuge it again for one min for elution of plasmid DNA.
• The plasmid DNA was Store at 4oC or -20oC.
2.8.8 Plasmid PCR and sequencing
The purified plasmids were again confirmed through plasmid PCR with internal
short gene primers or M13 primers. For this purpose the plasmids were diluted (1µl
plasmid DNA & 4 µl of dd H2O) and used for PCR following the procedure as under
Reagent Concentration
10X Buffer 5 µl
25mM MgCl2 3 µl

69
69
10mM dNTPs 1 µl
5 U µL-1 Taq Polymerase 0.5 µl
M13 Forward Primer (10µM) 1 µl
M 13 Reverse Primer (10µM) 1 µl
Double Distilled Water 37.5 µl
Plasmid (Diluted) 1 µl
Total 50 µl
PCR Profile
The following PCR profile was used for plasmid PCR reaction with
• One cycle of five minutes at 95°C for initial denaturation.
• 34 cycle of 30 second at 94°C, 30 seconds at 52°C and 2 min at 68°C.
• 1 cycle of 12 min at 68°C for final extension step.
The PCR product were analyzed on gel and sent for sequencing to Macrogen Korea.
2.8.9 Gateway cloning
This technique was used for efficient transference of DNA fragment among plasmid
vectors. This wass accomplished by the use of recombinant sequence set of
"Gateway att" site and two enzyme mixes, named "LR and BP Clonase". We
successfully transferred gene fragment among various vectors used for cloning.
Initially the gene fragments were first inserted in plasmids having both adjoining
recombinant sequences att L 1” and att L 2” in order to create a Gateway Entry clone.
These Entry clones were made in following two steps.
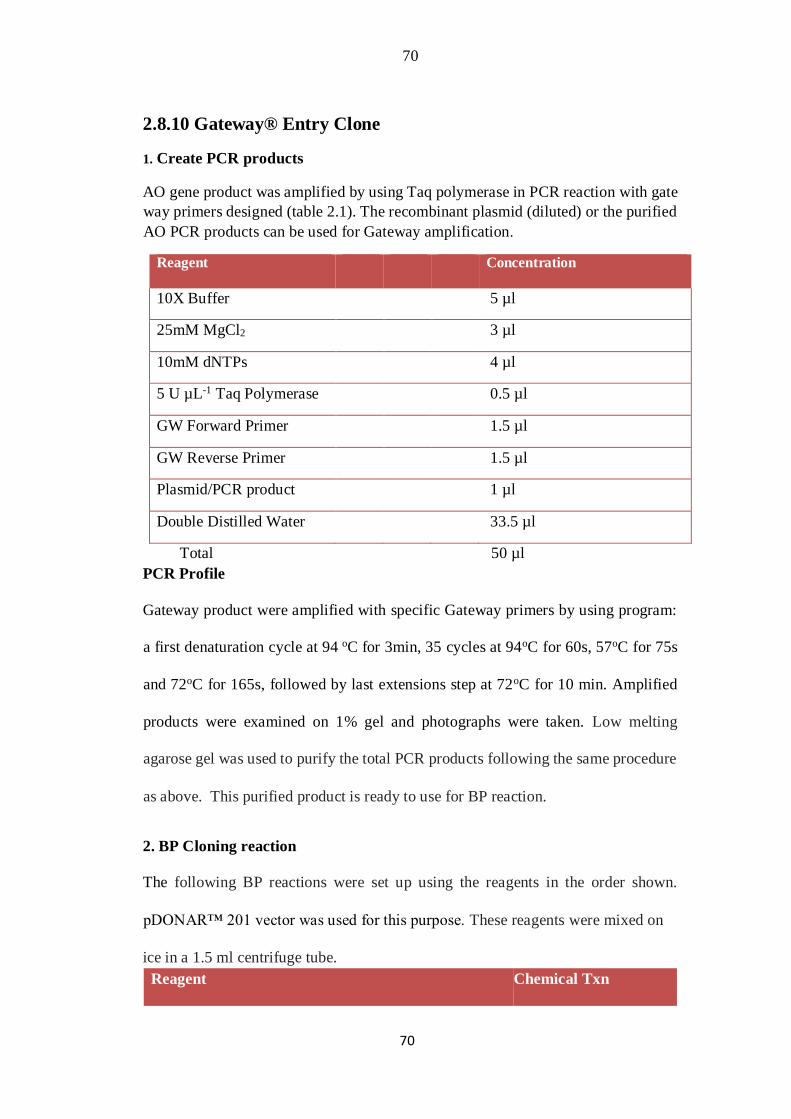
70
70
2.8.10 Gateway® Entry Clone
1. Create PCR products
AO gene product was amplified by using Taq polymerase in PCR reaction with gate
way primers designed (table 2.1). The recombinant plasmid (diluted) or the purified
AO PCR products can be used for Gateway amplification.
Reagent Concentration
10X Buffer 5 µl
25mM MgCl2 3 µl
10mM dNTPs 4 µl
5 U µL-1 Taq Polymerase 0.5 µl
GW Forward Primer 1.5 µl
GW Reverse Primer 1.5 µl
Plasmid/PCR product 1 µl
Double Distilled Water 33.5 µl
Total 50 µl
PCR Profile
Gateway product were amplified with specific Gateway primers by using program:
a first denaturation cycle at 94 oC for 3min, 35 cycles at 94oC for 60s, 57oC for 75s
and 72oC for 165s, followed by last extensions step at 72oC for 10 min. Amplified
products were examined on 1% gel and photographs were taken. Low melting
agarose gel was used to purify the total PCR products following the same procedure
as above. This purified product is ready to use for BP reaction.
2. BP Cloning reaction
The following BP reactions were set up using the reagents in the order shown.
pDONAR™ 201 vector was used for this purpose. These reagents were mixed on
ice in a 1.5 ml centrifuge tube.
Reagent Chemical Txn

71
71
Fresh PCR product (purified gateway clone) 1-7 µl
pDONAR™ 201 1 µl
TE Buffer (pH 8.0) 8 µl
These components were mixed carefully and incubated at room temperature for five
min. After thawing BP Clonase™ II enzymes by ice mixed it for about 2 min.
Then 2 µl of BP Clonase™ II enzyme was added to the reaction mixture by vortexing
briefly twice and centrifuged briefly. The reactions were then incubated for 1 hour
at 25°C. These reactions may be finished by addition of 1 µl of the Proteinase K into
all samples and vortex for a short time. These mixtures were incubated for 10 min at
37°C. These samples were placed on ice before proceeding to transformed electro-
competent E. coli cells. The map of pDONAR™201 vector was constructed in Sim
Vector 4.6 software as shown in
Fig. 2.3.
Fig. 2.3: The map of pDONAR™201 vector used in gateway cloning.

72
72
3. Transformation into electro Competent E. coli cells
Single vial of E. coli cells (DH5α) were thawed resting on ice. About 2 µl of the BP
mix was poured in vial of electro-competent E. coli (50 µl) cells and was mixed
carefully. It was incubated for few min lying on ice and transferred to pre cold
cuvette. The cells were then Electroporated at 1800 volts for transformation of
pDONAR™ 201 vector into DH5α cells with no shakiness. These tubes were
transferred into ice at once. About 500 µl LB liquid media was added, mixed gently
by pipetting and transferred to sterile micro centrifuge tube. It was incubated for one
to three hours by temperature of 37°C through continuous agitating. The bacterial
cultures (250-300 µl/plate) were spread on a pre-warmed LB agar plate which
contained kanamycin (100 µg/ml). Kanamycin was used as suitable marker for
selection of donor vectors. These plates were incubated at 37°C overnight. All
transformed colonies have produced white color competently. The well grown
colonies were selected and colony PCR was performed to confirm the transformants.
Reagent Concentration
Premix X Taq 12.5 µl
AO Forward primer 1 µl
AO Reverse primer 1 µl
dd H2O 10.5 µl
Colony Single
Total 25 µl

73
73
PCR Profile
Colony PCR was carried out using following program: an initial denaturation step at
94 for 3min, 35 cycles of 94oC for 60s, 57oC for 75s and 72oC for 165s, and last
cycle of extension at 72oC for 10 minutes. Positive colonies were selected by
analyzing the PCR product on 1% agarose gel and photographed. The positive
colonies were also grown further by striking on separate kanamycin resistant LB
plate. The colonies were used to inoculate 5 ml LB liquid media (added 5 µl
kanamycin) and were grown overnight. This culture was used to purify plasmid for
further reaction and sequencing. The BP purified plasmids were also confirmed
through PCR with gene specific primers. The primers were diluted for PCR (1 µl of
plasmid DNA & 4 µl dd H2O) and same PCR conditions were used as for colony
PCR.
Reagent Concentration
Premix X Taq 12.5µl
AO Forward primer 1 µl
AO Reverse primer 1 µl
dd H2O 9.5 µl
Plasmid DNA 1 µl
Total 25 µl
The purified plasmids were sequenced from Macrogen Korea with gene specific
primers. This entry clone was now prepared for entry into destination vector.
2.8.11. LR Reaction

74
74
LR reaction was used to transfer Hr-AO gene from Gateway entry clone into
destination vector. This was easier method and components were added following
the same order as above at room temperature.
Reagent Chemical Txn
Entry clone 1-7 µl
Destination vector 1 µl
TE Buffer (pH 8.0) 8 µl
LR Clonase ™ II enzyme was thaw on top of ice and mixed for 2 minutes. LR
Clonase ™ II enzymes were vortexes and mix it shortly two times. About 2 µl of LR
Clonase ™II enzyme mix was poured into the reaction and mixed by vortexing
briefly and then centrifuged. This reaction was incubated at 25°C for 1 hour.
Additionally 1 µl of the Proteinase K mix may be added to LR reaction mixture to
finish the reaction. The sample was incubated at 37°C for 10 min. The map of
destination vector used was shown in Fig. 2.4.

75
75
Fig. 2.4: Map of the destination vector used in Gateway cloning.
2.8.12 Transformation
The same protocol was followed as designated for BP reaction for transformation,
with the exception that we used appropriate selection markers for the LB plate
suitable for our destination vectors (typically 100 µg/ml ampicillin) instead of
kanamycin. The total LR reaction was used for transformation and plating. The
transformed cells were analyzed by colony PCR and positive colonies were marked
and grown for plasmid purification. The construct was ready to use for further
sequencing, cloning and expression process.
2.9 AO gene expression analysis
2.9.1 Semi-quantitative RT-PCR
The semi-quantitative RT-PCR amplifications were used to differentiate expression
pattern of ascorbate oxidase gene from Sea buckthorn plant. Different tissues
including vegetative buds, fresh seeds, shoot apex, green leaves, young green fruits
and mature orange red fruits were collected from the plant grown in the glass house
at the NIGAB Islamabad. Samples were instantly frozen using liquid nitrogen and
were utilized for total RNA isolation with Trizol reagent. The RNA was treated with
rDNAse to remove any DNA contamination and was quantify through thermo
Scientific’s NanoDropTM Lite spectrophotometer. 1.5% agarose gel was used to
evaluate the total RNA quality.
The AO gene’s primers were designed in NCBI primer picking program. These
primers were designed based on the sequence of new gene isolated. The cDNA

76
76
synthesized from different tissues was utilized as template in RT- PCR reaction with
the subsequent program: first cycle of denaturing at 95˚C 5 of min, 37 cycles of 94˚C
for 30sec, 58˚C for 30 sec and 68˚C for 30 sec, following last extension step of 68˚C
for 10 min. Actin-1 reference gene was utilized as control gene to check equal PCR
loading. These reactions were repeatedly carried out thrice for both biological and
technical replicates. The transcript products were examined on 1% agarose gel
stained with Ethidium Bromide (EtBr). The gel documentation system was used to
photograph the amplicon. The transcript accumulation was evaluated from band
intensity.
2.9.2 ATGene (Arabidopsis thaliana AO gene) Expression Pattern
For comparision of “Arabidopsis thaliana gene” AO expression Patterns with that
of sea buckthorn Hr-AO, the bioinformatics tool AtGenExpress Visualization
(http://jsp.weigelworld.org/expviz/expviz.jsp) was used to accomplish the analysis
of data set. For data processing, standardization, and statistical analysis of these
genes Microsoft Excel software was used. The results of the statistical analysis are
presented as a mean and ± SD.
2.10 RESULTS
2.10.1 Isolation and sequence analysis of large fragment of AO cDNA
from Sea buckthorn.
RACE-PCR techniques were utilized for amplification of full length fragment of Hr-
AO cDNA. This newely isolated sequence has full size of 2158 bp as compared to
1737 bp size of reference cDNA from tomato. The distinct high molecular weight

77
77
upper band of Hr-AO (2158bp) cDNA observed on agarose gel (Fig. 2.5) was picked,
purified and cloned. The length and purity of new cDNA was determined by the gel
electrophoresis method and sequencing. The positive clones, containing AO gene
insert, were picked out from thousands of transformed cells by blue white screening.
Plasmid extraction was carried out with Favorprep™ plasmid DNA extraction Mini
Kit. The plasmid containing 2.2 kb fragment of this cDNA sequence were confirmed
through PCR and sequencing. The new sequence of
cDNA sequence was analyzed using (NCBI) Search System
(www.ncbi.nlm.nih.gov/). The sequence similarity was searched by using BLAST
program against EMBL plant DNA sequence and swissport protein database. The
identity score of Hr-AO compared to some other plant AO sequences was commonly
found in range from 60 to 87%. Comparatively the expected Hr-AO nucleotides
sequences of sea buckthorn plant with that of tomato plant AOs indicated high
conservation in sequences as shown in Fig. 2. 6. Our sequence analysis implied that
Hippophae rhamnoides AO genes encoded enzymes homological to AOs from
different plants having similarities to other multicopper oxidases genes. Here I have
reported elaborate molecular study of ascorbate oxidase gene addressing differences
in length and amino acid residues.
ClustalW alignments from Hr-AO cDNA with reference sequence at both
nucleotide and amino acid level have been created and considerable variability was
observed in the sequence and size of this gene. Differences were found in the
nucleotide sequence of Hr-AO, and all these are the source of change in protein in
comparision with reference cDNA sequence. The deduced amino acids sequences
from the nucleotide sequence indicated that product of translation was a precursor

78
78
of about 719 amino acid residues with E value of 7.49e-82. The sequence identity to
tomato sequence was 87% at amino acid level.
Fig. 2.5: (a) Isolation of high quality total RNA from sea buckthorn tissues. (b)
Amplification of Hr-AO gene using RACE technique of PCR. (c) PCR confirmation
of the clones carrying recombinant plasmids with M13 and short internal primers. M
= 1Kb DNA ladder.

79
79
Fig. 2.6: Alignment of Hr-AO and S. lycopersicum nucleotide sequences with Mac
VectorTM 7.2.3. The top sequence is that of tomato AO and the bottom that of Hr-
AO. The identical bases are shown against a black background.

80
80
2.10.2 Gate way cloning
In addition to the molecular cloning and sequencing Gateway cloning was carried
out to transfer gene to expression vector. This is a robust scheme of creating an
extensive range of expressions construct for function in manifold host system. Gate
way cloning was used to easily shuttle our targeted gene (Hr-AO) accompanying
recombination with attB PCR products and attB expression clones. AO gene specific
primers were used to analyze entry clones (Fig. 2.7) by using PCR reactions. The
Entry clones were purified and sequence verified. After creating entry clones, our
concerned gene were transferable in great collection of expressions vector by means
of Gateway® LR reactions involving entry clone and destination vector of our
interest. This clone was ready to be used for further introduction and expression into
the system of choice.
Figure 2.7: (a) Amplification of Hr-AO product by standard PCR with gateway
primers. (b). Gel showing pDONAR recombinant plasmids. (c&d) The pDEST
vector pXCG-mYFP (51delta 35s) SB512 colony and plasmid PCR confirmation
respectively. (e) Stable integration of AO-PB clones carrying the recombinant
plasmid into destination vector. (f) pDEST AO recombinant plasmid confirmation
through PCR. M: 1 kb marker.

81
81
2.10.3 Differential expression of AO gene
In order to differentiate the expression pattern of ascorbate oxidase gene in various
tissues of Sea buckthorn total RNA from six different tissues like vegetative bud,
fresh seed, shoot apex, leaf, green fruit and mature fruit were extracted. This method
involves RT-PCR amplification of gene transcripts using mRNA reverse
transcription and a second step of semi-quantative PCR amplification (PCR) of the
cDNA synthesis. Normalization was done using constitutively expressed actin-1
gene. In our expression studies we find that transcript signals of AO are strongly
detectable in the green leaf tissues (Fig. 2.8). There was no band in case of seed
tissues. These reactions were repeatedly carried out thrice for both biological and
technical replicates.
Fig. 2.8: Hr-AO transcript amplifications in vegetative bud shoot apex, green leaf,
green fruit and mature orange red fruit tissues of Sea buckthorn through semi
quantative RT-PCR in comparisons with control Actin gene. . Actin gene is used as
control, to check equal loading and PCR quality check.
To check the gene expression pattern of ascorbate oxidase gene at different
development stages, abiotic stress, hormones and light under a set of multiple
biological conditions Arabidopsis thaliana gene expression data was used as
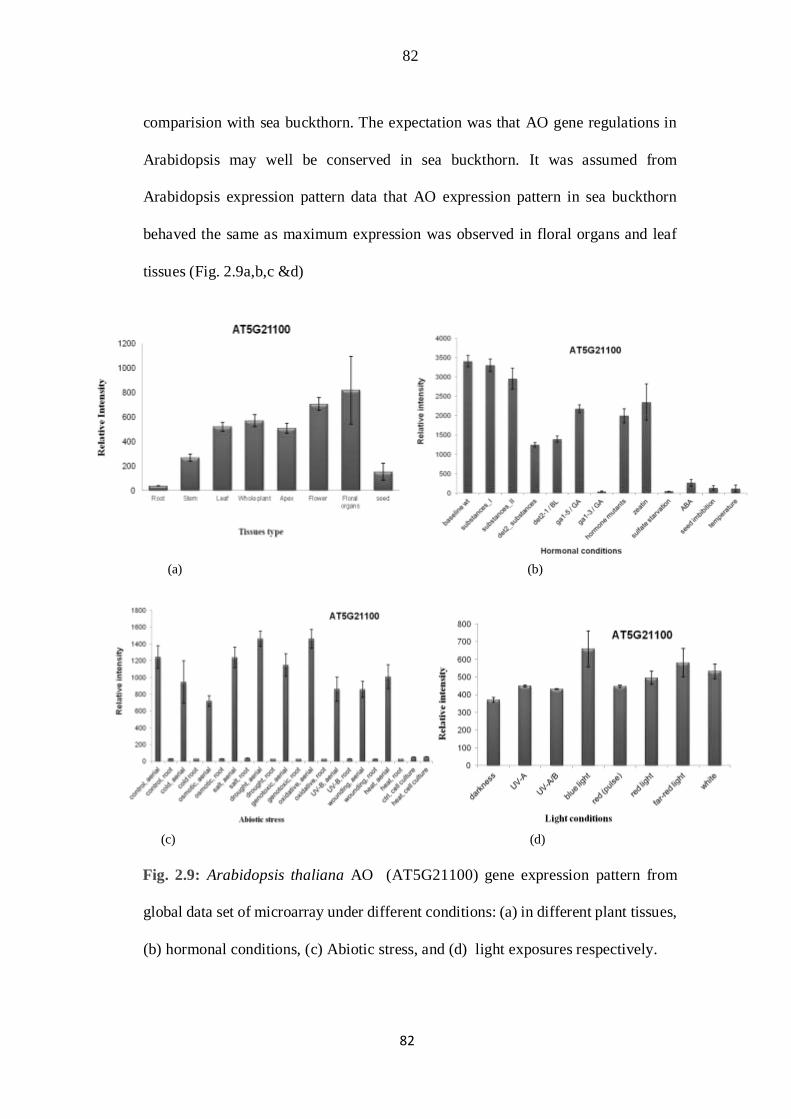
82
82
comparision with sea buckthorn. The expectation was that AO gene regulations in
Arabidopsis may well be conserved in sea buckthorn. It was assumed from
Arabidopsis expression pattern data that AO expression pattern in sea buckthorn
behaved the same as maximum expression was observed in floral organs and leaf
tissues (Fig. 2.9a,b,c &d)
(a) (b)
(c) (d)
Fig. 2.9: Arabidopsis thaliana AO (AT5G21100) gene expression pattern from
global data set of microarray under different conditions: (a) in different plant tissues,
(b) hormonal conditions, (c) Abiotic stress, and (d) light exposures respectively.

83
83
The transcript signals were also significant in case of green fruit tissues. However
weak band intensity was found in case of vegetative buds and shoot apex with
insignificant band intensity. More interestingly no amplicon were detected in case
of fresh seeds tissues.
I can imagine that ascorbic acid contents in matured fruit are less which can be due
to over-ripening of fruit with some degradation by enzyme then the activity growing
leaf and green tissues. Moreover the transcript intensity was higher in green fruit
tissues than the ripened fruit tissues. Leaf tissues showed the maximum transcript
accumulation which showed maximum ascrobate oxidase accumulation and
expression in apoplastic region. Gene expression changes are highly dynamic, and
gene expression pattern vary and often more complex. Numerous reports showed
that it is possible to improve ascorbate accumulation in plant cells via regulating the
ascorbate recycling process.
I was already reported that ascorbate oxidase transcript levels are highest in the
actively growing tissues. The ascorbate content in plants varies with different tissues
and plant species. Photosynthetically active tissues in addition to fruit and other
storage organs usually contain relatively higher concentration of ascorbate (Loewus
and Loewus, 1987).
The overview of different AO genes, function and expression in specific tissues with
their reference source are showen in table 2.3.

84
84
Table 2.3: Overview of different AO genes, function and expression in specific tissues
with their reference source.
Lf=leaf, Fl=flower, Sp=sepal, Pt=petal, St=stem, Fr=fruit, Rt=root, += expression and /= no
expression.
2.10.4 Phylogenetic analysis
Phylogenetic analysis revealed that the Hr-AO is the member of the copper ion
binding, oxidoreductase, family and it was assigned the unique name Hr-AO.
In order to invoke an evolutionary model between newly isolated sequence and those
in the database, phylogenetic reconstruction was carried out. Multiple sequence
alignments were completed with Clustal W alignments using Mega 5.0 program and
homology searches were done with BLAST programs in NCBI databases. For this

85
85
purpose the coding cDNA sequences of AO homologue were collected from
various plants species including L. esculentum, R. communis G. max, M. truncatula,
M. truncatula, P. sativum, G. max, N. tabacum, P. trichocarpa, G. max, O. sativa,
A. thaliana, A. thaliana, A. thaliana, A. thaliana, P. graminis, M. oryzae, L. bicolor,
B. juncea, C. maxima, Pumpkin, C. melo, Cucumber, C. sativus, C. melo, A.
thaliana, I. pine, F.ananassa, M. domestica, M. pumila were used. The tree was built
by majority decree and strict consensus. Terminal gaps were eliminated before
analyzing the sequences. Whereas the inner alignments gaps were left and analyzed
using scoring gap as character or as missing character. A tree was created using
neighbor-joining method as shown in figure 2.10. 1000 bootstrap replicate were used
for bootstrap analysis.
This tree displayed that sequences are evidently differentiated into different clades
with five different clusters. Higher bootstrap values are indicative of increased
reliability of the tree. Fascinatingly divergence in sequences was found with H.
rhamnoides sequence lay close to L. esculentum, then close to G. max, R. communis
and some other members of the Fabaceae family. The list of the organisms and
accession used in phylogenetic tree construction along with their evalue are shown
in table 2.4.
Table 2.4: List showing organisms and accessions used in phylogenetic tree
construction.

86
86
This is very outstanding discovery that Hr-AO along with Solanaceae member
showed close relation with Fabaceae member where AOs were strongly associated
in a cluster. The other members of the cucurbitaceae AO, brassicaceae and poaceae
were more distantly related. The members of the pine and Malus seem to be the
progenitors.

87
87
Fig 2.10: Phylogenetic reconstruction of ascorbate oxidase gene from different
species. Neighbor joining tree was made by means of MEGA5 software. The Value
on each node indicated bootstraps replication of 1000. This species along with
accession used in tree includes AY971876-L.esculentum-AO, XM_002528929-
R.communis-AO,XM_003532792-G.max-L-AO, XM_003638399M.truncatula-AO,
Y15295-M. truncatula-L-AO, AB457618-P.sativum-AO, AF529300-G.max-AO,
D43624-N.tabacum-AO, XM_002312802-P.trichocarpaAO, XM_003555611-
G.max-AO, NM_197271-O.sativa-AO, NM_147871A.thaliana-AO,
NM_001203424-A.thaliana-AO, AY099586-A.thaliana-AO,
BT003407-A.thaliana-AO, XM_003336029-P.graminis-AO, XM_003713188-
M.oryzae-AO, XM_001875289-L.bicolor-AO, AF206722-B.juncea-AO,
D55677C.maxima-AO, X55779-Pumpkin-AO, AF233594- C.melo-AO,
J04494Cucumber-AO, FR750377-C.sativus-AO, Y10226-C.melo-AO, AB004798-
A.thaliana-AO, B1698178-I.pine-AO, FJ896040-F.ananassa-AO, EF528482-
M.domestica-AO and Gu321223-M.pumila-AO

88
88
2.11 DISCUSSION
Sea-buckthorn is one of nature’s true super foods, as it packs a powerful nutritional
punch with a broad spectrum of health-promoting vitamins, minerals,
phytonutrients, and essential fatty acids. Whether you use it for a specific issue or to
promote overall health, give this little super food a try!
Vitamin C or ascorbic acid is a water-soluble vitamin most commonly found in
mammals and other animals. While humans are unable to synthesize ascorbic acid
and dependent on diet for their vitamin C contents. However the chemistry of
synthetic and food originated vitamin C is similar. The most fruit and vegetable are
abundant in several micronutrients and photochemical contents that could affect
bioavailability of vitamin C.
The fruits berry is the richest source of nutrient in diet proportional to energy
contents. Wild sea buckthorn berries are also good source of vitamin C. During our
study we successfully find coding fragment of AO ortholog from sea buckthorn
which is involved in ascorbic acid metabolism and biosynthesis. However reliable
and sensitive amplification of AO fragment (2158bp) was most important discovery
achieved through RT-PCR reaction with difference of about 421 bp between length
of the new amplicon and tomato gene. The non-synonymous differences were
randomly distributed among the whole sequence making difference with tomato
gene. Allignments at both nucleotide and amino acid levels showed conservation and
differences, which provide new tool for determining the function of proteins and
domains. In addition to examine gene functions it is essential to clone the target gene
into various types of plasmids. In case of plant biological methods the targeted gene
is commonly cloned in binary-vector for the purpose of Agrobacterium

89
89
transformations (Chakrabarty et al., 2007). The Hrascorbate oxidase was cloned
both through TA cloning and gate way cloning techniques to further study its level
of expression and function.
The capability of direct cloning and selecting recombination products in Gateway
cloning system is less time consuming non-laborious, and decreased possible
contaminant associated with handling of transformants. In our transformation system
the desired construct was firstly cloned in Entry vector (pENTR201) and was then
transfered into Destination vector pXCG-mYFP (51delta 35s) SB512 by site-specific
recombination. BP products (containing AO gene) were verified by sequencing.
Colony PCR was used to substantiate Gateway LR products, because there was no
change in sequence outside the recombination taking place at the Att site. A variety
of recombinant pDEST vectors obtained could also be utilized for the transformation
of Arabidopsis plants by floral dip system of transformation (Clough and Bent,
1998). This Hr-AO gene construct could potentially be used to simplify and improve
the efficiencies of gene cloning in A. tumefaciens for transformations studies in
plants or protein expression vectors and can possibly be adapted for high- throughput
applications.
However extensive expression and enzyme structural studies of AO gene have been
carried out but its exact function in higher plant species left unidentified. Current
investigation was centered on modifications in the Hr-AO expression pattern in
different tissues of sea buckthorn. Hence, differences in expression pattern in
different tissues are quite outstanding. The comparison of Arabidopsis thaliana AO
microarray data on gene expression pattern in different tissues and under different
conditions was also found consistent with that of sea buckthorn AO expression

90
90
profile. Ascorbate content in rapidly-growing tissues is higher than that of aging
tissues, as ascorbate is generally accumulated in the tissues with active growth such
as the meristem (Luwe, 1993). The ripening fruits in tomato accumulate more
ascorbate than immature fruits (Smirnoff, 1996).
In our work ascorbate oxidase gene transcript accumulation in sea buckthorn was
compared among different tissue, which corresponds to gene expression studies in
melon. Gene-specific expressions work in melon plant indicated that simply CmAO1
and CmAO4 genes showed transcriptional activity and differential regulation
depending upon tissues, development stages in addition to stimulus by outside.
Further they found CmAO1 gene transcript in floral and fruits tissue, while CmAO4
transcripts preferably accumulated in vegetative tissue without any expression of
CmAO genes in seeds of melon. Expression activity of CmAO4 gene was observed
during germination. Moreover regulation in activity of CmAO4 as a result of stresses
by heating, hormones and wound were also noticed (Sanmartin et al., 2007). These
results corresponded to our expression studies in sea buckthorn plant where I have
found AO expression activities in actively growing fruit and vegetative tissues.
Further reports of maximum expression of AO were found in different active
growing parts of plant with considerable increase in activities and expression level
of melon AO transcripts through the particular developmental stage of fruits (Esaka
et al. 1992; Moser and Kanellis 1994; Diallinas et al., 1997; Al-Madhoun et al.,
2003). Our results are also in coincidence with expression work in strawberry
(Fragaria×ananassa) where high expression level of FaAO was found in more
young parts (young fruits) with low expression in late maturing stages. It could be
linked to the facts of active growth in young fruit tissues with important effects of

91
91
FaAO on increasing fruits development (Yuanxiu et al., 2013).
Gene expression in sea buckthorn tissues followed divergent profile. The increase in
ascorbate level in actively growing tissues and fruits possibly will be the effect of
combine actions of oxidized and recycled enzyme. It was reported already that
during the process of earlier fruit growth there is an increase in transcript of AO
showing function of the enzyme in fruit development. Ascorbate oxidase enzymes
have also been suggested to participate in regulation of cell divisions and expansions
by control of ascorbate redox reactions (Davey et al., 2000; Potters et al., 2000;
Tabata et al., 2001; Sanmartin et al., 2007). There was a massive decrease in
transcript level on ripening of fruits. In melon fruit developmental process a
considerable gain in ascorbate oxidase contents during ripening of fruits having
significant part in metabolic process of cell wall (Moser and Kanellis, 1994). Some
results established differential regulation in expression of ascorbate oxidase gene at
the levels of transcripts increase (George et al., 1997).
However diverse biosynthesis pathways were connected with specific tissue,
development stage, or peripheral pressure and environmental conditions.
Appreciative effort about ascorbic acid metabolisms and biosynthetic processes in
fruit tissue is entailed with the purpose of regulating food plants with high ascorbic
acid contents. The scrutinization of cloning and expressions of ascorbate
biosynthetic enzyme included is useful approaches of this study. The assessment of
cumulative data from cDNA sequence, amino acid sequence, phylogenetic
reconstruction and expression pattern demonstrated that there is a considerable
variability in the gene constitution and evolutionary association of ascorbate oxidase
gene with other homolog.

92
92
2.12 CONCLUSIONS
The manifold roles of ascorbate oxidase in various plant physiological process and
ascorbic acid biosynthesis and recycling may bequeath with a more effectual
approach to improve ascorbate content of food crops. The current study exhibited
isolation, cloning and sequencing of ascorbate oxidase gene from sea buckthorn. The
expression studies showed that AO is vigorously expressed in dynamic fraction of
the plant tissues. Identification of genes controlling ascorbic acid buildup is
promising. The increasing knowledge about ascorbic acid genes should facilitate
engineering or modulating its accumulation.
Chapter 3

93
93
FOLATE (HPPK-DHPS) GENE
Abstract
Sea buckthorn (Hippophae rhamnoides L.) is a hardy, fruits growing plant. Well
recognized for its value in medicines and nutraceautical products, it is enriched with
astounding array of nutrient in its fruit, seed, leaf and bark contains vitamins,
essential oils and minerals. Most of the nutrients are lacking in our food crops. This
study was envisaged to isolate the folate (HPPK-DHPS) ortholog
from sea buckthorn. The target gene Hydroxymethyldihydropterin
pyrophosphokinase–dihydropteroate synthase linked in the folate biosynthesis
pathway was successfully isolated and cloned. The sequences analysis revealed that
this novel genomic locus is 2354bp in size. The coding region is interrupted by a
single large intron. Its length is 1539bp which is similar to its ortholog in tomato.
The newly isolated gene has prominent nucleotide differences compared to tomato
sequence, randomly distributed throughout the full sequence length. Expression
profile of HrHPPK-DHPS with semiquantitative RT-PCR revealed the higher
transcripts accumulation in leaf and fruits tissues. Phylogentic reconstruction
revealed its association with other plant species. In this study an attempt was also
made to compare the 3D homology modeling structure of new protein, though
considerable conservation in 3dimensional structure of folate proteins was observed;
however, notable differences in substrate binding pockets were also visible. This
study has future implications in utilizing the potential of wild medicinal plants for
genetic improvement of folate deficient staple food crops.

94
94
3.1 INTRODUCTION
Sea buckthorn fruit is considered as vitamins and other much important bioactive
substance repository (Xurong et al., 2001). Sinensis a subspecies of sea buckthorn
fruits have large amount of vitamins A, B2, C as compared to carrot, tomato and
orange. Sea buckthorn berries also have vitamin B1, P and K (Lu, 1992) and folate
vitamers (Gutzeit et al., 2008) to a very great extent.
It was also reported that total folate contents in sea buckthorn berries in the range of
about 39 μg/100 g and 5-methyltetrohydrofolate was illustrated as the principal
folate vitamers found in sea buckthorn berry. Tracings of tetrahydrofolate were
noticed although were distantly lower than the detecting point (Strålsjö et al.,
2003). The structure of folate and 5-Methyltetrahydrofolate are shown in fig. 3.1.
(a) Folate
(b) 5-Methyltetrahydrofolate
Fig. 3.1: Structure of Folate (a) and 5-methyltetrahydrofolate(b) found in sea
buckthorn.
Folate belongs to B-group vitamin that is important for cells function as well as cell
division. Human diet contains important nutrients such as vitamins and minerals.

95
95
One third of the world population cannot reach their intellectual and physical
potentiality due to deficiency. Many severe diseases such as blood anemia and neural
tube defects in newborns (Lucock, 2000), psychiatric syndromes in the elders and
reduce cognitive functioning (Calvaresi and Bryan, 2001; Hultberg et al., 2001) are
caused due to folate deficiency.
There are a number of genes that codes for vitamins such as folate in this
nutraceautical plant. The plant bifunctional HPPK-DHPS is an important enzyme of
the folate biosynthetic pathway. Tetrahydrofolate and its one-carbon derivative,
conjointly named folate, are important cofactors for one-carbon reactions in the
biosynthesis of amino acids such as purines, pantothenate, and thymidylate (Hanson
et al., 2000; Ravanel et al., 2004).
Folate is a molecule consisting of p-aminobenzoate condensed with pterin ring and
glutamate moieties. Their biosynthetic pathways in plant are individually separated in
three sub-cellular sections as showen in fig 3.2. The initial committing stage in folate
synthesis provided the pteridine by the catalytic function of GTP cyclohydrolase in
cytosolic region (Hossain et al., 2004; Schoedon et al., 1992; Yoneyama et al., 2001).
The synthesis of p-aminobenzoate occurred in plastid from chorismate in two steps
originated by aminodeoxychorismate synthase (Basset et al., 2004a). The
condensation of pABA and pteridine moieties take place in mitochondria
catalyzed by 6-hydroxymethyl-7,8-dihydropterin pyrophosphokinase, and
7,8-dihydropteroate synthase. These two are specific to plants and microorganisms,
permitting for de novo folate synthesis (Re´ beille´ et al., 1997).

96
96
Fig. 3.2: The systematic presentation of plant folate biosynthesis pathway redrawn
from source (Rébeillé and Douce, 1999; Rocío I et al., 2007). The main pathway
enzymes includes 1. GTP cyclohydrolases I; 2. nudix hydrolase; 3. non-specific
phosphatase; 4. DHN aldolase; 5. ADC synthase; 6. ADC lyase; 7.
HPPKpyrophosphokinase; 8. dihydropteroate synthase; 9. dihydrofolate synthase;
10. dihydrofolate reductase; 11. folylpolyglutamate synthase. 12. DHN,
dihydroneopterin; 13. ADC, aminodeoxychorismate; 14. PABA, p-aminobenzoate,
15. PPP triphosphate.

97
97
Our study was mainly focused to obtain full length gene fragment of HPPK-DHPS
from genomic DNA of sea buckthorn. It is well known that there is correlation
between number of introns found and gene expression in higher organisms. Several
new studies indicate that rapidly cycling cells constrain gene-architecture toward
short genes with a few introns, allowing efficient expression during short cell cycles.
In contrast, longer genes with long introns exhibit delayed expression, which can
serve as timing mechanisms for patterning processes (Heyn et al., 2015) Earlier
investigations on genome DNA sequencing in a wide variety of organism
demonstrated great variability in intron-exon structures of homologous genes in
various organisms (Rodríguez-Trelles et al., 2006). Recently much research on
complete genome of eukaryote has depicted the introns length and densities varied
substantially among related groups. The value and cost of spliceosom intron in
eukaryotic organisms was not demonstrated. The most documented effects of introns
splicing is to enhance the genes expression but their regulatory mechanism is still
undefined. Some earlier study revealed that intron splicing is a lengthy process. It
indicated that splicing cannot decrease the time needed in transcriptional activities
and spliced pre-mRNA processing. Alternatively it may be facilitating the afterward
cycles of transcriptions (Deng-ke and Yu-Fei, 2011).
The folate synthesis was reported to occur preferably in tissues with higher division
and photosynthetic activity during the process of cellular differentiation.
Additionally the folate contents in fruit could change during the maturing processes,
such as in tomato it was decreased with fruits ripening from mature green to red
ripening phase (Basset et al., 2004a).

98
98
The divergence detected between HPPKs and DHPSs from various species
powerfully suggested the substantial variation in genes progressions. However the
functions of these evolutions are poorly interpreted, a potential benefit of the
bifunctional property of HPPK/DHPS may possibly take both catalytic sites close;
as such actions are sequential in folate biosynthesis.
Sea buckthorn has a great nutraceautical potential for genetic improvement of
cereal crops through genetic engineering, hitherto little efforts has been made in
isolating folate gene from this plants. But before this application, it is very important
to isolate and analyze these genes and to study their expression patterns using
molecular techniques. Therefore we set out, 1) to isolate and clone HPPKDHPS
gene, 2) to analyze the sequence of this gene using bioinformatics tools, 3) to analyze
expression patterns of this genes in different tissues at various developmental stages
and, 4) Comparative homology modeling for deciphering the 3D structure of HPPK-
DHPS gene.
Based on experimental and sequences analysis I hypothesized that there existed a
novel HPPK-DHPS gene in sea buckthorn, recruited in folate synthesis pathway. No
deletion or insertions were detected but non-synonymous substitutions in
comparision with tomato sequence were found in the entire sequence.
Phylogenetic reconstruction revealed that HPPK-DHPS from sea buckthorn is
sister to tomato ortholog. Expression profile of HrHPPKDHPS with
quantitative RT-PCR indicated the higher accumulation of transcripts in leaf
and fruits tissues.

99
99
3.2 MATERIAL AND METHODS
3.2.1 Plant material
Sea buckthorn (Hippophae rhamnoides ssp. Sinensis) plants were grown in glass
house as well as in the field of National Institute for Genomics and Advanced
Biotechnology. For DNA/RNA extraction, the leaf, berries, bud and seed samples
were harvested from plants at various stage of development. Fruits harvesting was
carried out once or twice a week during the ripening period from sea buckthorn
nursery. The samples were frozen in liquid nitrogen and stored at -
80ᴼC until processed for further nucleic acid extraction and expression analysis.
3.2.2 Designing of primers
Nucleotide sequences of HPPK-DHPS genes were retrieved from
National Center for Biotechnology Information (NCBI) databases. The gene
specific primers were designed from conserved region of HPPK-DHPS gene
sequence of S. lycopersicum (accession No. FJ972198) was utilized for
amplifications of full length coding region of HrHPPK-DHPS gene. Primers for
expression analysis through RT-PCR were designed from newly isolated gene
sequences of HrHPPK-DHPS gene (table 3.1) .

100
100
Table 3.1: Primers used for HrHPPK-DHPS gene amplification and
expression. studies
3.2.3 DNA extraction
A CTAB method of DNA extraction was set out to isolate the HrHPPK-DHPS gene
from sea buckthorn plant instead of cDNA. DNA was extracted using the leaf
samples of sea buckthorn with a standard CTAB method (Doyle and Doyle, 1987).
The samples were grind with liquid nitrogen and homogenize in
1ml of 2×CTAB buffer (2% cetyltrimethylammonium bromide (CTAB); 100mM
Tris HCl with pH 8.0, 1.4M NaCl; 20mM EDTA; 1% PVP; 0.2% β-
mercaptoethanol). Incubate it at 65°C for 40 minutes. Each lysate was extracted two
times by equivalent volumes of chlorophorm: isoamyl alcohol (24:1). Then

101
101
precipitate DNA by 100% isopropanol followed by centrifugation at 13,000 rpm for
10 min). Each DNA pellet was washed using cold 70% ethanol, then dry it and was
suspended in 100 μl of TE buffer. Add 4 μl RNaseA (10 μg/ml) to the dissolved
DNA and incubated for removing traces of total RNA. The DNA was run on agarose
gel and photographed as shown in fig. 3.3a.
3.2.4 PCR amplification and molecular cloning
Diluted DNA was utilized as template in standard PCR reaction for HrHPPKDHPS
gene amplification using the Takara Ex Taq™ polymerase. PCR
amplifications were carried out in a reaction volume of total 50 μl by the use of gene
specific primers with initial denaturation at 94ᴼC for 3 min, amplification with 35
cycles at 94ᴼC for 60s, annealing at 57ᴼC for 75s, elongation at 72ᴼC for 145s,
followed by final extensions step of 72ᴼC for 10 min. The amplification products of
HPPK-DHPS gene were purified from gel by the use of “Gene JET™ gel extraction
kit” following cloning into pTZ57R/T vector for sequencing to MACROGEN
(Korea).
3.2.5 Isolation of RNA and expression analysis
Different tissues from sea buckthorn were used for total RNA extraction by means
of the TRIZOL® Reagent (Invitrogen). The quantification of RNA was
accomplished with thermo Scientific’s NanoDropTM Lite. 1.5% agarose gel was
used to evaluate the quality of total RNA extracted. Fermentas AMV-RT reverse
transcriptase enzyme was used for first strand cDNA synthesis. A 2µg sample of
total RNA with oligo (dt) primer was incubated at 65ᴼC for 5min, chilled on ice and
mixed briefly. The AMV-RT enzyme, buffer and deoxyribonucleotides were added

102
102
and incubate the reactions for 60min at 50ᴼC. Then terminate the reaction at 85ᴼC
for 5 minutes. Multiplex semi-qualitative RT-PCR was followed up to differentiate
the expressions pattern of freshly amplified HrHPPK-DHPS gene transcript in leaf,
fruits bud and seed of sea buckthorn as described in Khan et al., (2012a). First strand
cDNA synthesized using the oligo (dT) primers from various tissues were utilized
as templates in a reaction volume of total 20µl by Fermentas Taq polymerase.
The following PCR profile was used with an initial denaturation of 5 min at 95oC
following 35 cycles at 94oC for 30 sec,
58oC for 30 sec and 68oC for 30 sec, and a final extension of 68oC for 7 min. The
18SrRNA gene was applied as an internal control. The reactions were repeated three
times for each of the biological and technical replicate. The products amplified were
resolved using 2% agarose gel and photographs were taken.
3.2.6 ATGene (Arabidopsis thaliana gene) expression evaluation for
HPPK-DHPS
The Arabidopsis thaliana gene Hydroxymethyldihydropterin pyrophosphokinase
dihydropteroate (HPPK-DHPS) expression pattern under a set of multiple
biological conditions (different tissues, hormones, abiotic stress and light) was also
compared with sea buckthorn. The tool in bioinformatics such as AtGenExpress
(http://jsp.weigelworld.org/expviz/expviz.jsp) was used to accomplish the analysis
of data set from HPPK-DHPS model system. For further statistical analysis, data
normalization, and processing of these genes Microsoft Excel software was used.
The results of the statistical analysis were obtained as a mean and ± SD.

103
103
3.2.7 Sequence analysis and phylogenetic reconstruction
After the sequences were retrieved for each gene family, the sequence and
phylogenetic analysis was completed with Mac Vector version 7.2 and MEGA 5
(Kumar et al., 2008). Clustal W (Thompson et al., 1994) in Mac Vector 7.2 was used
as a multiple alignment tool for aligning the amino acids. HPPK-DHPs Nucleotide
and protein sequences were retrieved from NCBI data base from different species.
The Fasta formate of different protein sequences were aligned in Mac Vector 7.2
software to see the difference in newly isolated amino acids sequence with
previously isolated sequences.
The nucleotide sequences were translated in the Mac Vector program. The default
parameters were used. Sequences which were too diverged disturbed the alignments
so they were removed manually while performing the multiple alignment procedure.
Using the alignment file phylogenetic tree was constructed following Neighbor
Joining methods (Russo et al., 1996; Saitou and Nei, 1987) implemented in MEGA5.
The total deletions options were utilized for omitting any position which postulates
gaps in sequence. As amino acids substitution models, un-corrected proportions (p)
of amino acid variations and distances were utilized. MEGA 5.0 software was used
to construct the Phylogenetic tree. The bootstrap method (Felsenstein, 1985) was
used to test the authenticity of the generated tree topologies. The method (at 1000
pseudo replicates) gave the bootstraps probabilities of all inner branches in tree.

104
104
3.2.8 Homology modeling and protein structure prediction
The three dimensional (3D) structural modeling of Folate protein (HrHPPKDHPS)
was performed by MODELLAR 9V10 homology modeling program. The amino
acid sequence of HrHPPK-DHPS amino acid residues (512) retrieved in FASTA
format were submitted to protein-protein blast (Blastp) searches adjacent to protein
data bank (PDB) for identification of an appropriate templates structure for relative
modeling. The 2BMB_A was preferred as a proper template structure by query
sequences making 33% identities and query coverage of 89% and have E-value of
7e-70. The modeler quality was assessed based on sequence alignment by blast and
template structure used. These structures were predicted using MODELLAR having
alpha helice and beta-pleated sheet envisioned by Chimera 1.6.

105
105
3.3 RESULTS
3.3.1 Amplification of HrHPPK-DHPS gene from sea buckthorn
In order to screen for the candidate gene linked in the pathway of folate biosynthesis
gene specific primer pair HPPK-F and HPPK-R were used for amplifying HPPK-
DHPS gene from sea buckthorn. A full length 2354 bp target genomic fragment was
amplified which was interrupted by single introns. The distinct high molecular
weight band of HPPK-DHPS observed on agarose gel (Fig. 3.3b) was purified and
cloned. E. coli strain DH5α was used as host for plasmid PTZ57R/T which carries a
gene for ampicillin resistance. Four positive clones, containing HPPK-DHPS gene
insert, were picked out from thousands of transformed cells (Fig. 3.4a) Extraction of
these plasmids for nucleotide sequence analysis was carried out with Favorprep™
plasmid DNA extraction Mini Kit. The plasmid containing 2.3 kb fragment of this
gene were confirmed through PCR and sequencing (Fig. 3.4 b&c).
(a) (b)
Fig. 3.3: Amplification of HrHPPK-DHPS gene. (a) Image of Genomic DNA
isolated from three different sea buckthorn samples (S1, S2, S3) run along 1kb DNA
ladder. (b), PCR amplification of full length sequence 2354bp of folate gene (HPPK-
DHPS) from sea buckthorn genomic DNA. M stand for 1kb DNA
ledder.
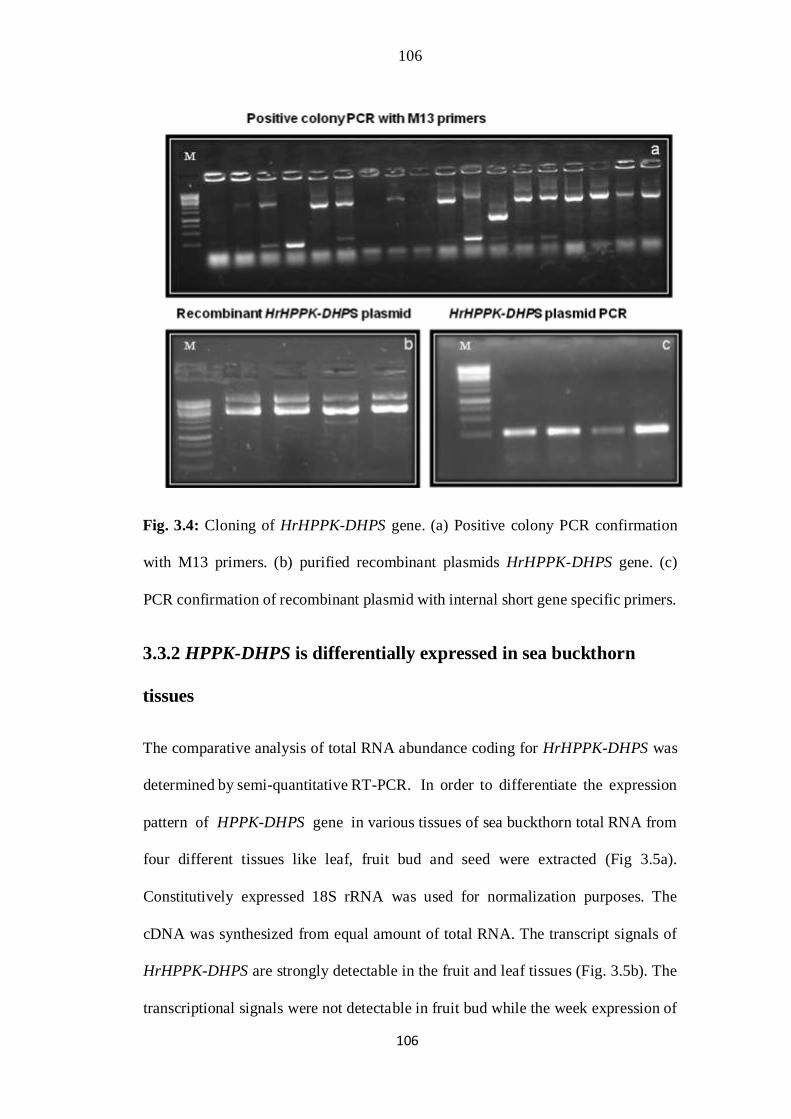
106
106
Fig. 3.4: Cloning of HrHPPK-DHPS gene. (a) Positive colony PCR confirmation
with M13 primers. (b) purified recombinant plasmids HrHPPK-DHPS gene. (c)
PCR confirmation of recombinant plasmid with internal short gene specific primers.
3.3.2 HPPK-DHPS is differentially expressed in sea buckthorn
tissues
The comparative analysis of total RNA abundance coding for HrHPPK-DHPS was
determined by semi-quantitative RT-PCR. In order to differentiate the expression
pattern of HPPK-DHPS gene in various tissues of sea buckthorn total RNA from
four different tissues like leaf, fruit bud and seed were extracted (Fig 3.5a).
Constitutively expressed 18S rRNA was used for normalization purposes. The
cDNA was synthesized from equal amount of total RNA. The transcript signals of
HrHPPK-DHPS are strongly detectable in the fruit and leaf tissues (Fig. 3.5b). The
transcriptional signals were not detectable in fruit bud while the week expression of

107
107
this gene was found in seed. Hence, differences in expression pattern in different
tissues are quite outstanding. The present study exhibited that HrHPPK-DHPS genes
are strongly expressed in active part of the sea buckthorn plant tissues comparatively.
Fig. 3.5: (a) RNA isolation from different tissues of sea buckthorn shown against λ
DNA/ HindIII Marker. (b) RT-PCR analysis of HPPK-DHPS gene transcripts (270
bp) in different tissues (leaf, Fruit, Bud, Seed) of sea buckthorn as shown against
50bp ladder. 18S rRNA was used as internal control.
The Arabidopsis thaliana gene Hydroxymethyldihydropterin pyrophosphokinase
dihydropteroate (HPPK-DHPS) expression pattern under a set of multiple
biological conditions (different tissues, hormones, abiotic stress and light) was also
compared with sea buckthorn as shown in fig 3.6a,b,c&d.
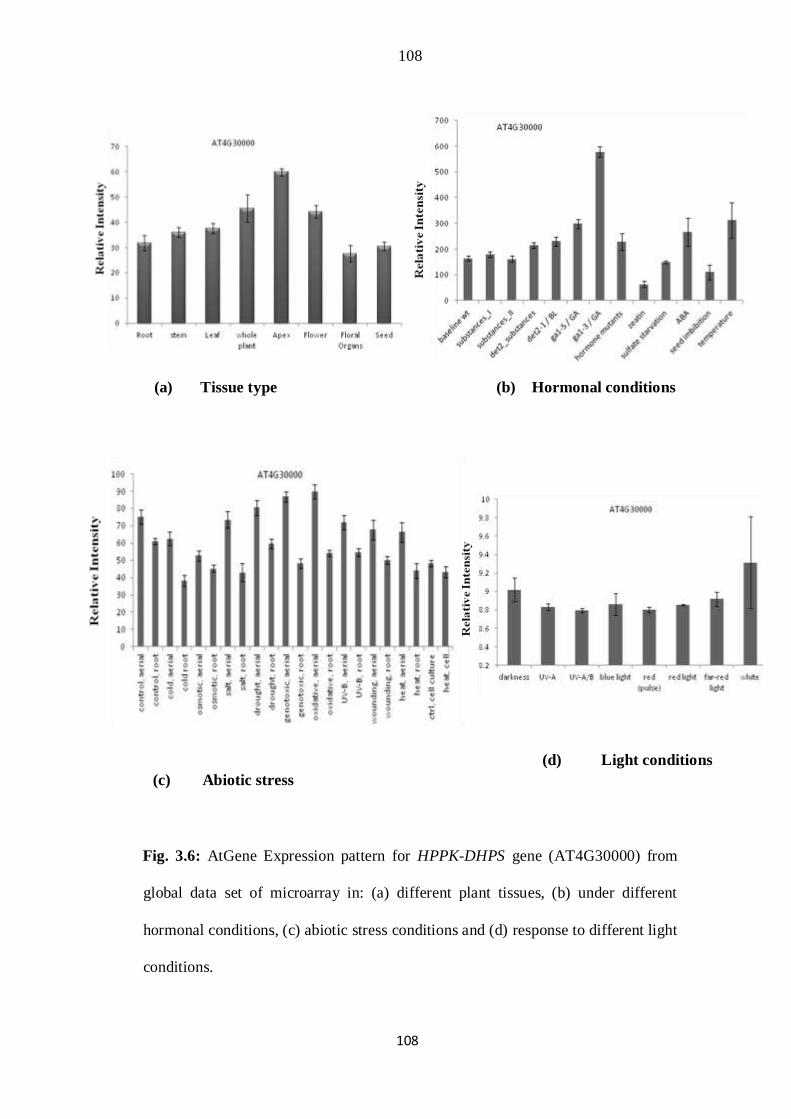
108
108
(a) Tissue type (b) Hormonal conditions
(d) Light conditions
(c) Abiotic stress
Fig. 3.6: AtGene Expression pattern for HPPK-DHPS gene (AT4G30000) from
global data set of microarray in: (a) different plant tissues, (b) under different
hormonal conditions, (c) abiotic stress conditions and (d) response to different light
conditions.

109
109
3.3.3 Sequence analysis of HPPK-DHPS identified from Sea buckthorn
The newly isolated sequence was edited using Mac Vector™ 7.2.3 software. Blasts
result presented maximum hit with HPPK-DHPS gene of S. lycopersicom followed
by P. sativum, M. truncatula, C. arietinum, G. max, A. thaliana, A. lyrata T.
aestivum, B. distachyo and O. sativa etc. thereby confirming the presence of new
sequence of HPPK-DHPS gene in sea buckthorn. All positions of codon comprising
gaps and missed data were removed. The length of sequenced clone was 2.3 kb due
to the presence of a single large intron. The 1539 bp coding sequence was extracted
and it was found to be exactly equal in length to tomato gene coding sequence. Thus,
HrHPPK-DHPS gene sequence revealed no variation in the length of the nucleotide
sequence. ClustalW alignments of HPPK-DHPS gene with the tomato gene
sequence were generated and considerable variability was observed in the sequence
of this novel gene. Alignments of newly translated amino acid with the tomato
sequence were also made and shown in fig. 3.7. A total of 35 nucleotide differences
were observed in sequence in the final dataset as shown in Fig. 3.8 as compared to
tomato ortholog sequence. These differences were simply of substitutions type, and
no deletion and insertion were detected. These were randomly distributed in the
entire sequence without specific domain and resulted in a change in the protein
structure (non-synonymous type). The amino acids sequences inferred from the
nucleotides sequences indicated that product of translation was a precursor of about
512 amino acid residues. The sequence identity was 97% at amino acid level
compared to tomato sequence.

110
110
HPPK-DHPs protein sequences were retrieved from NCBI protein data base from
different species. Multiple sequence alignment has shown the high sequence
similarity of HrHPPK-DHPS translated amino acid sequence with different proteins
from number of plant species including tomato, arabidopsis, soya bean and wheat.
However considerable differences were observed at the N-terminus and C-terminus
regions of these proteins. Sequence similarities and difference among selected
protein homologue sequences are shown in Fig. 3.9.
Figure 3.7: Alignment of newly isolated HrHPPK-DHPS amino acid sequence with
its ortholog from S. lycopersicum (FJ972198) using Mac VectorTM7.2.3. (Accerlrys
Inc.) gcg/ Wisconsin pakage university of Wisconsin). The similar regions are shown
against a black background and differences are exposed against white background.

111
111
Figure 3.8: Alignment of newly isolated HrHPPK-DHPS nucleotide sequence with
its ortholog from S. lycopersicum (FJ972198) using MaVectorTM7.2.3. (Accerlrys
Inc.) gcg/ Wisconsin pakage university of Wisconsin). The identical regions are
shown against a black background and differences are exposed against white
background.

112
112
Fig. 3.9: Multiple sequence alignment of newly isolated HrHPPK-DHPS amino acid
sequence with Mac VectorTM7.2.3. (Accerlrys Inc.) gcg/ Wisconsin pakage
university of Wisconsin) from sea buckthorn and its ortholog from S. lycopersicum,
G. max, A. thaliana and T. aestivum amino acid in blocks highlight similarity. The
strictly conserved regions are shown against black background.

113
113
3.3.4 Phylogenetic reconstruction of HPPK-DHPS gene
To search the evolutionary relationships among newly isolated sequence and those
in the database, phylogenetic reconstructions were completed. The coding sequences
of HPPK-DHPS homologue from different plant species including S .
lycopersicum, T. aestivum, B. distachyon, C. arietinum, O. sativa, A. thaliana, P.
sativum, M. truncatula, G. max, and A. lyrata were used. A neighbour-joining tree
shown in figure 3.10 demonstrated that sequences were clearly separated into various
clades. Four groups containing brassicaceous HPPK-DHPS, cereals, pea family and
tomato as a member of Solanaceae were observed. The un-rooted tree clearly
separates out HPPK-DHPS of sea buckthorn and establishes its relationship with
tomato. This indicates divergence in sequences. The monocots i.e. T. aestivum B.
distachyon, O. sativa, appeared to be the progenitor because these were present at
the base. The increase in reliability of the tree was indicated by highest bootstraps
values.
All the result established that HPPK-DHPS genes were substantially differentiated
in various clusters. The isolated new HrHPPK-DHPS sequence however
presented clustering with tomato gene, therefore divergence was subsisted. To
what extent variations in HrHPPK-DHPS contribute to functional divergence,
characterization during site directed mutagenesis and transformation is needed.
Although earlier to transformation it is preponderating to perform the expression
studies of the HrHPPK-DHPS in the different tissue of the sea buckthorn. The
amino acid composition was calculated in BioEdit software

114
114
Fig. 3.10: Phylogenetic reconstruction of HrHPPK-DHPS gene. Neighbor joining
tree is constructed using MEGA 5.0 software. Values on the nodes indicate the
bootstrap replication of 1000. Phylogenetic inferences of enzyme comprising both
Domains HPPK and DHPS were accomplished utilizing complete 15 coding
sequences of H. rhamnoides, S. lycopersicum (GenBank accession No. FJ972198)
T. aestivum (EF208803), B. distachyon
(XM003562616), O. sativa (NM001066822), O. sativa (AK068210), P. sativum
(Y08611), M. truncatula (XM003614065), C. arietinum (XM004490156) G. max
(XM003518734), G. max (XM003517802), A. thaliana (NM105586), A. thaliana
(NM001203939), A. lyrata (XM002888660) and A. thaliana (BT033093) were
included in the alignment. Four clusters are enclosed by brackets. A scale is given at
the bottom.

115
115
3.3.5 Tertiary structure of folate protein
HPPK-DHPs belong to the Pterin-binding superfamily. There have been a no of
techniques in molecular structural modeling. 3D structure of H. rhamnoides, S.
lycopersicum T. aestivum, A. thaliana and G. max HPPK-DHPS protein was
constructed and visualized using the MODELLER 9v10 and Chimera 6.1 software.
The 3D structure of the selected original saccharomyces cerevisiae gene templates
(Lawrence et al., 2005) were used as starting point for homology modeling. The
domain architecture consisting of Pterin domain from 230-498 residues and HPPK
domain from 47-172 amino acid residues was evaluated using the chimera program
for sea buckthorn protein. The HrHPPKDHPS has e-value of 5.77e-118 DHPS and
2.94e-52 for HPPK. The two consreved domains of HrHPPK-DHPS genes were searched
from NCBI data base conserve
domain search as shown in fig. 3. 11.
Fig. 3.11: Conserved domain of newly isolated HrHPPK-DHPS gene obtained from
domain search data base showed hppk superfamily and pterin_binding domain.
All the 3D structures of 5 different protein fold classes ranging in size from
512-561 amino acid residues. The ~ 250-residue pterin-binding (DHPS) domains
were found to have (beta/alpha) 8 barrel fold, which adopted shape of a distorted

116
116
cylinder. It has eight alpha-helices gathered around the outside of an inner cylinder of
parallel beta-strands. The pterin ring binds at the bottom of the (beta/alpha;) The
pterin ring was partly hidden inside the (beta/alpha) 8 barrels. The pterins bonding
residuals depicted higher conservation among five species. Modeled tertiary protein
structure of different plant species were shown in (Fig 3.12). HPPK was the upstream
and the adjoining enzyme to DHPS in the folate biosynthetic pathways. The 3D
structure of the target protein was also sought out from protein data bank (PDB) but
no 3D structure of folate protein from plant folate synthesis pathway had been
reported to date.
Fig. 3.12. Predicted 3D structure of HrHPPK-DHPS proteins from different plant
species shown at the same scale. A, X-ray structure of the template bifunctional
enzyme HPPK-DHPS from S. Cerevisiae (Lawrence et al., 2005) B, 3D structure of
sea buckthorn HPPK-DHPS produced using MODELER
9v10, C, 3D model of S. lycopersicum, D, 3D model of A. thaliana, E, 3D Model

117
117
of G. max. F, Model of T. aestivum. Models in different protein structure were
colored according to different domains. Pterin binding domain DHPS was given
Blue color, HPPK domain was shown by red color. Inter-domain linker
was given grey color.
3.4 DISCUSSION
The berries of sea buckthorn are enriched with great array of substances possessing
substantial biological activities. There is possibly an enormous accumulation of
nutritionally valuable photochemicals waiting to be discovered. Sea buckthorn fruit
comprises solid carotenoids substances, amino acid, vitamin B (Folic Acid), B1, B2,
B6, B12, B15, and K, dietary minerals, polyphenolicacids and βsitosterol. The
problems of micro-nutrients insufficiency can be solved by fortifying staple crops,
which is an established practice and found to be more effective for several
micronutrients.
The isolation and cloning of a single gene (HPPK-DHPS) encoding key enzyme
in folate biosynthesis pathway was successfully carried out for the first time from
sea buckthorn in this study. A full length fragment of HPPK-DHPS gene containing
large single intron was obtained using standard PCR amplification. The HPPK-
DHPS genes were already cloned from numerous plants included Arabidopsis, dicots
and monocots. As there were no deletions and insertions in the extracted coding
sequence, the size of newly isolated sequence remain conserved. The length of
coding sequence was restricted to 1539 bp for this gene. The amino acids sequences
derived from the nucleotides sequences indicated that product of translation was a
precursor of about 512 amino acid residues. In pea leaf the synthesis of a HPPK-
DHPS enzyme, having open reading frame of 1545 bp and inferred amino

118
118
acids sequences corresponding to polypeptides of 515 residuals was reported, which
match our results (Fabrice et al., 1997). The homology analysis using ClustalW
alignments showed that the HrHPPK-DHPS sequence was 97% identical to the
sequence of S. lycopersicum cultivar (SlHPPK-DHPS).
The HrHPPK-DHPS fragment cloned in this study contained a large single intron. It
is well established that introns results in increased stability and expression of genes
(Buchman and Berg, 1988; Chung and Perry, 1989; Okkema et al., 1993). The gene
expression in plants and in several eukaryotes can be affected importantly by introns
in different ways. An enhancer element or alternative promoters are found in few
introns whereas a large number raise mRNA accumulation by various procedures.
It is obvious from expression study that there were differences remarkably in the
accumulation patterns of HPPK-DHPS among different stages of development. The
expression was not detectable in bud while its accumulation gradually lowered in
seed but in the fruit and leaf tissues transcript expression was rather similar and was
considerably higher than other tissues (Fig. 4.8). Our results are in corroboration
with already reported expression studies of HPPK-DHPS gene in tomato,
Arabidopsis and pea in dissimilar tissues and at different developmental stages
(Fabrice et al., 1997). The expression profile of HPPK-DHPS gene in different
tissues of the sea buckthorn has shown same behavior as observed by Arabidopsis
thaliana HPPK-DHPS microarray gene data.
The proteins that synthesize folate are different from species to species. The
phylogenetic analysis differentiated the HPPK-DHPS genes from different
species into 4 different groups. HPPK-DHPS from sea buckthorn conglomerated
with S. lycopersicum gene. It can be inferred that these two proteins might have

119
119
the same properties. But functional characterization of these two proteins through
ectopic expression or knocking down is required to validate this hypothesis.
Nevertheless, the divergence among different species exists to their clustering.
It is obvious from expression study that there was significant divergence in
accumulating profile of HPPK-DHPS involving different stages of development.
The expression was not detectable in bud while its accumulation gradually lowered
in seed but in the fruit and leaf tissues transcript expression was rather similar and
was considerably higher than other tissues. Our results are in corroboration with
already reported expression studies of HPPK-DHPS gene in tomato, Arabidopsis
and pea in dissimilar tissues and at different developmental stages.
In general all these investigations effectively suggested that synthesis of folate is
under strong control. These premises also implicated differences in requirements
among tissues and are fluctuating with plant developmental processes. The entire
folate content is different from one plant to another. Folate content also varies with
the type of the organ or tissue as the folate contents shows fluctuation throughout the
developmental process of plant, suggests that folate synthesis is strongly managed
and changed as a role of the metabolic necessity (Basset et al., 2004a; Jabrin et al.,
2003). It is found that tissues like cotyledons, roots and stems with decreased
metabolic activities have fixed quantity in pea seedlings during development (jabrin
et al., 2003; R beill et al., 2006).
This investigation demonstrated that sea buckthorn has an ortholog of HPPKDHPS
gene therefore it is a different gene. This sequence was equated with the sequences
present in gene pools, which predict that this novel gene sequence, belong to HPPK-
DHPS. The HrHPPK-DHPS fragment cloned in this study

120
120
contained a large single intron. This was further cloned in expression vector for
maximum expression. It is found that there is big difference of sequences among a
specified intron and exon into newly isolated sequence. There is a single intron in
the full length DNA sequence of HPPK-DHPD gene. The exon sequence is greatly
more preserved which suggest that the factual sequences of the intron was not
extremely very significant. The intron was spliced out and coding region sequence
encodes a putative protein which is conserved among tomato pea and sea buckthorn.
Intron can considerably have an effect on genes expression in plant as well as in
number of eukaryote in different manners. It is well established that introns results
in increased stability and expression of genes (Buchman and Berg, 1988; Chung and
Perry, 1989; Okkema et al., 1993).
Our study was mainly focused to obtain full length gene fragment of HPPK-DHPS
from genomic DNA of sea buckthorn. It is well known that there is positive
correlation between number of introns found and gene expression and that introns
containing transgenes showed enhancements of gene expression both transcription
and translation than the same gene lacking introns (Brinster et al., 1988). Earlier
investigations on genome DNA sequencing in a wide variety of organism
demonstrated great variability in intron-exon structures of homologous genes in
various organisms (Rodríguez-Trelles et al., 2006).
Phylogenetic analysis differentiated the HPPK-DHPS genes from different species
into 4 different groups. HPPK-DHPS from sea buckthorn conglomerated with S.
lycopersicum gene. It can be inferred that these two proteins might have the same
properties. But functional characterization of these two proteins through ectopic
expression or knocking down is required to validate this hypothesis. However, the

121
121
divergence among different species exists due to their clustering. This might provide
the evolutionary advantage to this family for proteins.
However in our study homology modeling of 3D structure of HPPK-DHPS gene
from sea buckthorn plant in comparison with other plant species was also reported.
HPPK-DHPS proteins are found in them Mitochondria, chloroplast and cytosol of
higher plants. The evolutionary trajectory of a protein through sequence analysis is
constrained by its function. Polypeptide consists of both HPPK and DHPs structural
domains that are relevant mono-functional domains (Sandeep et al., 2012). These
domains allowed us to construct a 3D protein structure of H. rhamnoides by applying
Homology modeling approach. The rationale behind this method is the observation
that proteins with similar sequence usually share the same overall 3D folding
patterns. Therefore experimentally determined protein structures were usually used
as template for predicting the confirmation of an additional protein with identical
amino acids sequences (Havel and Snow, 1991; David, 1997).
This work also described the significant structural variation in petering binding
domain among the various orthologus proteins. We have found slight but notable
difference with respect to the substrate binding pockets opening. There might be a
clue about functional attributes regarding the 3D structure of different binding
proteins that is triggering the function. The eminence of homology modeling
completed using MODELLER and Chimera program
depends upon the excellence of the sequence alignments by Blast and protein
structure used as template. The structural analysis and figures generated with
MODELLER software were presented in Fig. 3.11. These structures provide a good
foundation for functional analysis of the gene. The comparison of data from DNA

122
122
sequence, amino acid sequence, phylogenetic reconstruction, homology modeling
and expression pattern demonstrated that there is a considerable variability in the
gene structure and evolutionary relationship of HrHPPK-DHPS with other
homologs.
3.5. CONCLUSIONS
In a nutshell, the current investigation presents lines of evidences for varying
expression level of the bifunctional HPPK-DHPS on the folate status in plants.
Based on sequence similarity, clustering and phylogenetic differentiation of genes
it is concluded that HPPK-DHPS isolated from sea buckthorn is a novel gene.
In this study we predict and compare three dimensional structure of HPPK-DHPS
protein which was not available in plant species This study has future implications
in exploring the potential of wild medicinal plants for genetic improvement of staple
food crops to meet the nutrients deficiency in human diet.
Chapter 4

123
123
CAROTENIODS BIOSYNTHESIS GENES (LCY-β AND PSY)
Abstract
Caroteniods are organic pigments that are dispersed almost everywhere in living
things. These are important for the growth of plants and photosynthesis. These are
main nutritional source of vitamin A for humans being. Basic challenges were to
study two important caroteniods pathway genes, lycopene β-cyclase and phytoene
synthase from vitamin A rich plant sea buckthorn. The newly isolated full length
cDNAs sequence of Hr-Lcyb has a complete open reading frame of 1503 bp and Hr-
Psy has an open reading of 1238 bp in length. The length of the coding sequences of
Hr-Lcyb and Hr-Psy showed homology with tomato gene sequences. RT-PCR
analysis was carried out to show maximum transcript accumulation in different
tissues of sea buckthorn plant. In case of lycopene beta cyclase maximum transcript
signals were observed in case of fruit and leaf tissues. Significant level of expression
was also found in shoot apex and newly growing bud tissues. Whereas very minute
traces were found in root and seed tissues. In case of phytoene synthase maximum
transcript abundance was found in fruit, leaf and apex. Some expression was also
found in bud and seed tissues. In contrast to Lcy gene no transcripts signals were
found in root tissues. In this assessment we recapitulated the existing knowledge of
the carotenoid transcript signals and discussed about the knowledge of their
biosynthesis regulation in bud, shoot apex, roots, seeds and fruits. This work
recommends that the study of different genes encoding caroteniods with diverse
activity will also help to meet the problem associated with vitamin A deficiency.

124
124
4.1 INTRODUCTION
Carotenoids are one of the major groups of pigments that are widely spread in plants.
They are responsible for the red, orange and yellow, coloration of tissues. They are
biologically important in many organisms from bacteria and fungi to higher plants
(Goodwin, 1980). These are concerned in photo protection, lightharvesting, and
pollination in plants (Tracewell et al., 2001; Szabo et al., 2005; Dong et al., 2007).
Forty-one different carotenoids have been found in various varieties of sea buckthorn
berries (Bekker and Glushenkova, 2001); the main carotenoid’s types are β -
cryptoxanthin, β-carotene and zeaxanthin, while canthaxanthin, alphacarotene,
dihydroxy-β-carotene, γ-carotene and lycopene are known as the minor ones (Yang
and Kallio, 2005).
As b-carotene, carotenoids, and cantaxanthin work as anti-mutagenic and
anticancerogenic and anti-swelling therefore these compounds act as safeguard from
some kinds of skin cancer (Mashkovski, 1997). Enzymes have genetic code which
is responsible to catalyze major steps in the biosynthetic pathway of carotenoid have
been replicated and their expression have been analyzed in various species (Zar et
al., 2009; Charles et al., 2009; Ha et al., 2007).
Carotenoids oil contents have found to be valuable remedies against burn, ulcers,
cryopathies, skin cancers and many gynecological disorders (Akulinin, 1958;
Chugaeva et al., 1964; Gatin, 1963; Goodwin, 1980; Mashkovski, 1997; Rahimv et
al., 1983). The structure of α-carotene and lycopene were shown in Fig. 4.1.
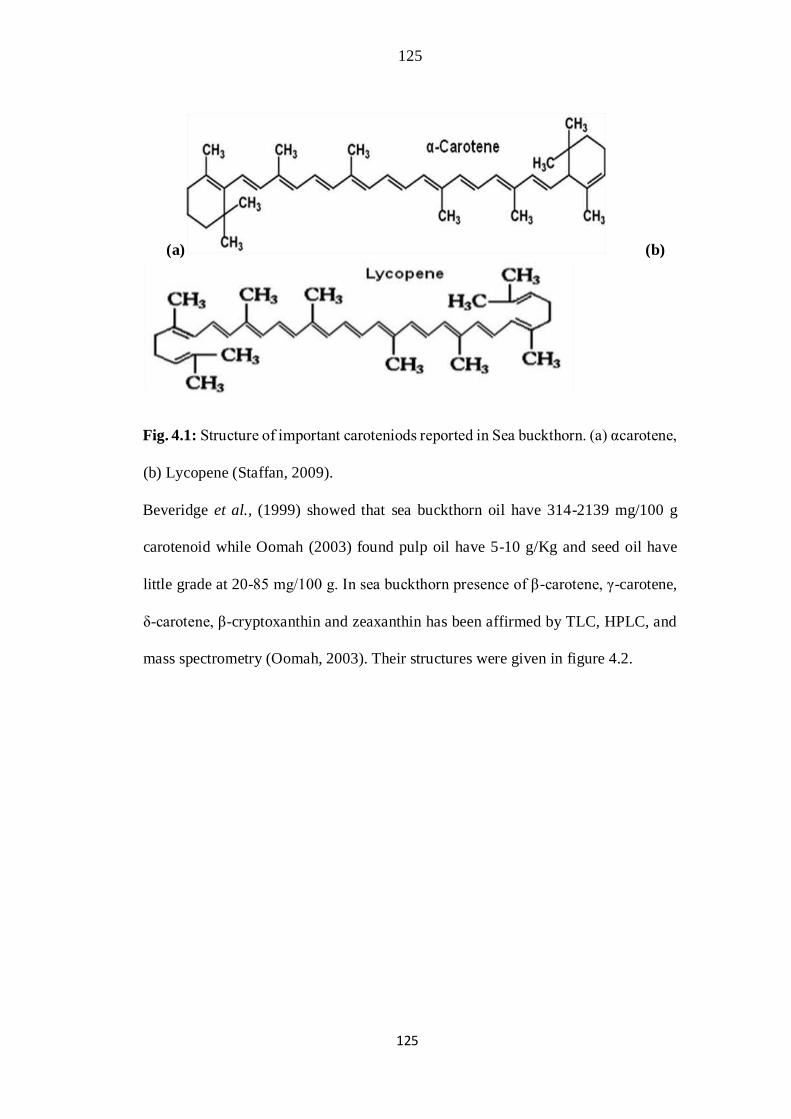
125
125
(a) (b)
Fig. 4.1: Structure of important caroteniods reported in Sea buckthorn. (a) αcarotene,
(b) Lycopene (Staffan, 2009).
Beveridge et al., (1999) showed that sea buckthorn oil have 314-2139 mg/100 g
carotenoid while Oomah (2003) found pulp oil have 5-10 g/Kg and seed oil have
little grade at 20-85 mg/100 g. In sea buckthorn presence of β-carotene, γ-carotene,
δ-carotene, β-cryptoxanthin and zeaxanthin has been affirmed by TLC, HPLC, and
mass spectrometry (Oomah, 2003). Their structures were given in figure 4.2.

126
126
Fig 4.2: Structures of different forms of carotenoids reported to be present in the sea
buckthorn (Staffan, 2009).
The carotenoid biosynthesis pathway in plants begins with the formation of phytoene
from geranylgeranyl pyrophosphate, by the enzyme phytoene synthase
“PSY” (Fraser & Bramley, 2004; Gallagher et al., 2004). With two isomerization
and four desaturation reactions phytoene is then transformed to all-translycopene
(Chen et al., 2010). Two branches, the β,ε branch and the β,β branch, in the pathway
are formed by the cyclization of lycopene. The formation of α-carotene from the β,ε
branch depend on enzymes like lycopene epsilon cyclase and lycopene beta cyclase;
the formation of β-carotene from the β,β branch depends on LCYβ (Fig. 4.3). In β-
carotene hydroxylation of the two β-ionone rings is the most powerful nutritional
source of vitamin A, (Rafael et al., 2014).

127
127
Figure 4.3: Systematic presentation of carotenoid biosynthetic pathway in plant
(Clotault et al., 2008).
High amount of phytochemicals like carotenoids, ascorbic acid, tocopherols and
phenolic compounds are present in the Sea buckthorn berries (Zadernowski et al.,
1997; Andersson et al., 2008; Zadernowski et al., 2005). Skin, mucosa,
cardiovascular and immune system disorders are being treated by sea buckthorn
berries (Yang and Kallio, 2002; Raffo et al., 2004). Earlier studies have shown that

128
128
red and orange-red fruits of sea buckthorn have more carotenoids than yellow and
orange-yellow (Lian et al., 2000). As fruit matures the carotenoid content is
increased (Zhang et al., 1989).
For nutritional and health products preparation, it is significant to have access to raw
material that has high content of carotenoids (Deli and Molnar, 2002; Maoka et al.,
2001). Earlier studies have shown that sea buckthorn berries have differences in the
contents and composition of carotenoids (Yang and Kallio, 2002; Beveridge et al.,
1999) which may be due to many reasons like environment and growing conditions,
berry parts study, genetic variation, and variation between years, storage
environment, and methods of examination. The constituents of sea buckthorn fruit
are showen in table. 4.1.
Table. 4.1: Constituents of Sea buckthorn Fruit (Alam, Dharmananda, 2004).
Sr. No. Sea buckthorn fruit constituents (Per 100 grams fresh berries)
1 The major unsaturated fatty acids are linolenic acid(omega-
3) , palmitic acid and linoleic acid (omega-6),, palmitoleic
acid (omega-7), oleic acid (omega-9),; there are also
saturated oils and sterols (mainly β-sitosterol)
6–11% (3–5% in fruit pulp, 8–18% in
seed) fatty acid composition and total oil
content vary with subspecies
2 Vitamin C 28–310 mg (typical amount: 600 mg)
3 Carotenoids, including beta carotene, lycopene, and
zeaxanthin; these contribute to the yellow-orange-red colors of the fruit
Total caroteniods
32–45 mg fatty acids (oils)
(432.4 IU 100 g-1) (Stobdan et al.,
2010). 527.8 mg/100g (Beveridge et al.,
1999; Cenkowski et al., 2006; Lian et
al., 2000).10- 1,200 mg/kg (yang et al.,
2002)
4 Vitamin E (mixed tocopherols) Up to 180 mg (equal to about 270 IU)
5 Folic acid Up to 80 mg
6 Organic acids for example, quinic acid, malic acid;
ingredients similar to those found in cranberries Quantity not determined, expressed juice
has pH of 2.7–3.3
7 Flavonoids (e.g., mainly isorhamnetin, quercetin glycosides,
and kaempferol) 50–500 mg (0.05% to 0.5%)

129
129
Despite several studies on caroteniods contents in sea buckthorn no attempt was
made to exploit important vitamin genes to alleviate micronutrient deficiencies:
micronutrient supplementation, food fortification, and biofortification. Biosynthesis
of carotenoid has been studied in plants like pepper, tomato and Arabidopsis
(Hirschberg et al., 2001).
Sea buckthorn gives nearly all nutrition contents which are necessary for human
being, because main cereals usually have poor dietary products. Basic challenges of
this study were the identification of caroteniods biosynthetic pathway genes. Here
we report molecular cloning of carotenoid biosynthesis genes (Lcy & Psy) and have
analyzed the sequence identity with same genes from different species.
4.2 MATERIAL AND METHOD
4.2.1 Plant Material
Sea buckthorn (Hippophae rhamnoides subsp. Sinensis) plants were grown both in
the green house and in the field of National Institute for Genomics and Advanced
Biotechnology. The plant samples including buds, root, shoot apex, leaves, seed and
fruit tissues were collected at different growth points. Fruits were harvested once or
twice a week during the ripening period from sea buckthorn nursery, all these
samples were put in liquid nitrogen to freeze the samples which were stored at -80ᴼC
till it is processed to extract nucleic acid.
4.2.2 Designing of Primers
Nucleotide sequence of Lcyb and Psy genes were collected from NCBI
(www.ncbi.nlm.nih.gov). Primers were utilized for both genes with the known
sequences of S. lycopersicum cDNAs. These were use to synthesize the cDNAs made

130
130
from RNAs of the sea buckthorn (Hippophae rhamnoides) fruit. The primers for
gene expression studies were designed from newly isolated cDNA sequences. The
detail and accession number and sequence of the gene specific primers used in the
study are given in Table 4.2.
Table 4.2: Detail of Primers used in this study for H. rhamnoides Lcy and Psy cDNA
cloning and expression analysis (RT-PCR).
4.2.3 RNA extraction and PCR Amplification
The roots, leaves, buds, shoot apex, seed and fruits of sea buckthorn were used to
extract RNA with the help of a reagent known as TRIZOL. To extract RNAs
instructions of “InvitrogenTM Life Technologies Corporation” were used. Extracted
RNA was evaluated qualitatively on 1.5% agarose gel. The cDNA was synthesized
using Fermentas AMV-RT reverse transcriptase (#EP0641) kit. A 2µg sample of

131
131
total extracted RNA with gene-specific primer was incubated at 65ᴼC for 5min,
chilled on ice, mix briefly. The AMV-RT enzyme, buffer and
deoxyribonucleotides were added. At 50ᴼC reaction was incubated for 60 minutes
followed by 85ᴼC for 5 minutes.
For gene amplification of Hr-Lcyb and Hr-Psy, fruit cDNA of sea buckthorn were
utilized as template in RT-PCR which was performed using Takara Ex Taq™
polymerase amplification kit under the following program with initial denaturation
step of 94ᴼC for 5 minutes, amplification with 35 cycles of 94ᴼC for 60s, annealing
at 58ᴼC for Hr-Lcyb and 59ᴼC for Hr-Psy for 60s elongation at 68ᴼC for 150s,
accompanied by final extension at 68ᴼC for 10 minutes.
4.2.4 Expression analysis of Genes
To recognize the expression forms of newly separated Hr-LCY and Hr-Psy genes in
different tissues semi quantitative RT-PCR was done. For this purpose the roots,
leaves, buds, shoot apex, seed and fruits of sea buckthorn were used to extract RNA
using TRIZOL reagent according to manufacturer instructions as previously
described in gene isolation procedure (Fig. 4.4a). As far as RNA concentration in
different tissues and yield is concerned high RNA yield was found in leaf, apex and
fruit tissues which showed maximum transcript accumulation. RNA yield from bud
root and seed tissue was low which showed very low transcript accumulation. From
newly isolated gene sequence all the short primers for RT-PCR analysis were
designed in Bio-Edit program (Table 4.2). Then, the standardized RNA from various
tissues was used as templates for cDNA synthesis with oligo (dt) primers using
AMV-RT enzyme (#EP0641). The transcripts amplification of Lcy and Psy gene was
carried out with short gene specific expression primers by RT-PCR in total reaction

132
132
volume of 20 µL using Takara Ex Taq™ polymerase kit. The following PCR profile
was run with first denaturizing cycle for 5 min at 95˚C, follow by 34 cycles at 94˚C
for 30 sec, 58˚C for 30 sec and 68˚C for 30 sec, and a last extension step of 68˚C for
7 min. Actin-1 gene which express in all eukaryotic tissues was selected as control
to check equal loading and PCR quality check. . These qRT-PCR reactions were
performed thrice involving both technical as well as biological replicate.
After that, gene expression pattern was examined electrophoretically using 2%
agarose gel with ethidium bromides staining and then photographs were taken.
Cumulative data of transcript signals were evaluated according to the band intensity,
which was translated into relative quantity and compared with the reference band.
4.2.5 AtGene (Arabidopsis thaliana Gene) expression analysis of LCY
and PSY.
AtGenExpress is a tool to analyze the transcript abundance changes of gene under
study in Arabidopsis using global microarray data collection. Here the focus was on
differential expression estimates computation of caroteniods gene (Lcyb &Psy) in
different tissues and different set of particular biological conditions. This
AtGenExpress data site (http://jsp.weigelworld.org/expviz/expviz.jsp) was used to
accomplish the analysis of data set. The Arabidopsis thaliana caroteniods gene for
lycopene β-cyclase (AT3G10230) and phytoene synthase (AT5G17230) were
therefore selected to compare its expression pattern with sea buckthorn. For data
normalization, processing and statistical analysis of these genes Microsoft Excel
software was used. The results of the statistical analysis are presented as a mean and
± SD.

133
133
4.2.6 Molecular Cloning and Sequence analysis
The targeted products of both genes were purified from agarose gel by means of the
Gene JET™ Gel extraction kits. The genes of interest were ligated into the
pTZ57R/T vector, which is then transformed into DH5α strain of E.coli cells. Blue-
white screening was used to choose colonies with desired insert (white colonies).
Recombinant plasmids comprising DNA inserts of both genes were therefore
purified from transformed cells. Plasmid purification was carried out with
Favorprep™ plasmid DNA extraction Mini Kit and sequencing was done from
MACROGEN (Korea). The BioEdit and Mac Vector™ 7.2.3 software were used to
edit the nucleotide sequences of amplified gene. In addition to nucleotide alignment
amino acids formatted alignment was also completed using Mac Vector™ 7.2.3
program. ClustalW alignments have been made to compare these newly isolated
sequences with previously isolated tomato sequences. Phylogenetic relationships
among different plant species were also analyzed. A neighbor joining methodology
was utilized for constructing the Lcyb and Psy gene sequence tree via MEGA 5.0
software. A bootstrap test based on 1,000 replicates was completed to verify the
reliability of the phylogenetic tree.

134
134
4.3 RESULTS
4.3.1 Isolation of Hr-Lcyb and Hr-Psy Genes
The Fig. 4.4 b&c showed the targeted size products of Hr-Lcyb and Hr-Psy amplicons
from sea buckthorn plant respectively. These cDNA amplifications were purified
and inserted into fast cloning vector pTZ57R/T for transformation into E. coli strains
DH5α as shown in Fig. 4.5 A&B. The recombinant plasmids were purified and
sequenced. The newly isolated full length cDNAs confirm that the Hr-Lcyb has a
complete open reading frame of 1503 bp and Hr-Psy has an open reading of 1238
bp in length. Hr-lcyb encoded proteins of 501aa with e-value of
2.69e-175 and Hr-Psy encodes 412aa amino acids residues with e-value of 2.45e90.
The length of the cDNA remains the same as compared to tomato sequence. By
contrast with tomato gene sequence only substitutions were found and no insertions
or deletions were found in the new gene sequences.
All these nucleotide differences were of non-synonymous type that does not lead to
appreciable protein change. In any part of protein, substitution of amino acid can
occur that does not affect secondary structure or function of protein in a significant
way. A replacement of an amino acid may occur by another amino acid of same
chemical property and protein still has normal functions. These non-synonymous
types are actually favoring novel expressions of the genetic material leading to the
development of more improved functions.
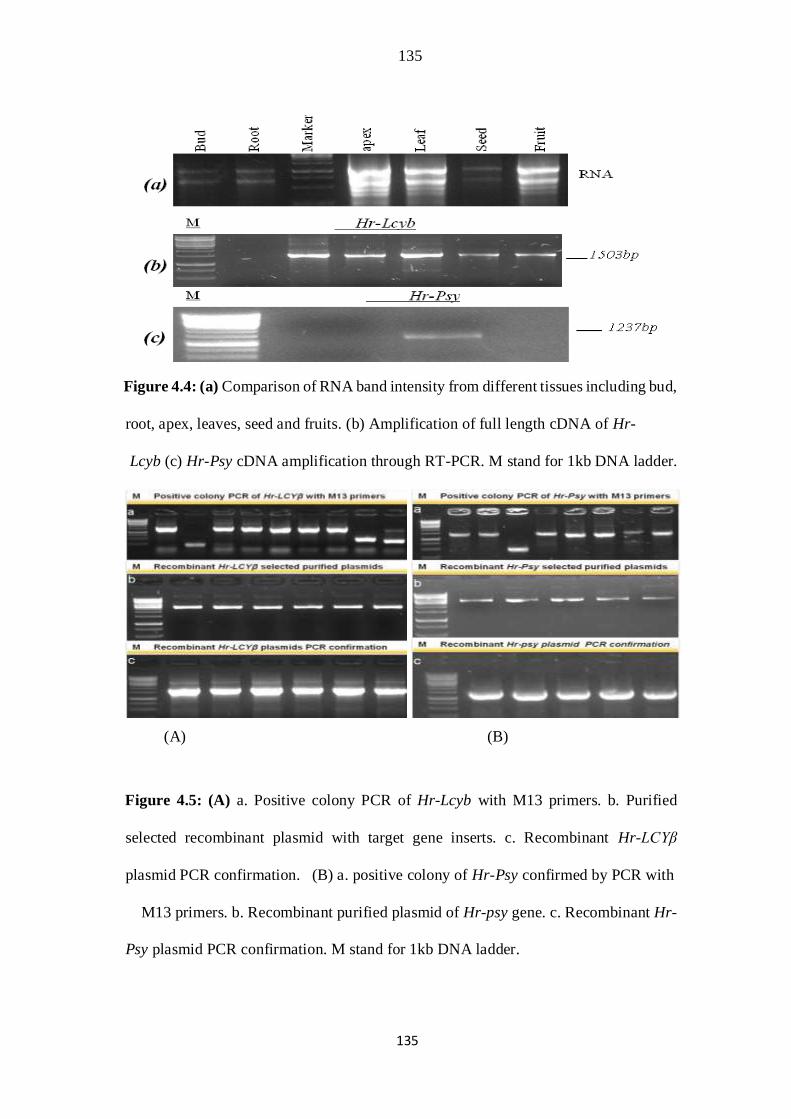
135
135
Figure 4.4: (a) Comparison of RNA band intensity from different tissues including bud,
root, apex, leaves, seed and fruits. (b) Amplification of full length cDNA of Hr-
Lcyb (c) Hr-Psy cDNA amplification through RT-PCR. M stand for 1kb DNA ladder.
(A) (B)
Figure 4.5: (A) a. Positive colony PCR of Hr-Lcyb with M13 primers. b. Purified
selected recombinant plasmid with target gene inserts. c. Recombinant Hr-LCYβ
plasmid PCR confirmation. (B) a. positive colony of Hr-Psy confirmed by PCR with
M13 primers. b. Recombinant purified plasmid of Hr-psy gene. c. Recombinant Hr-
Psy plasmid PCR confirmation. M stand for 1kb DNA ladder.

136
136
4.3.2 Differential Gene Expression Analysis
To substantiate further the quantitative nature of PCR Semi quantitative PCR was
used for Gene expression analysis. Gene expression levels varied in different plant
tissues (bud, apex, root, leaf, seed and fruit tissues) which potentially effect
caroteniods accumulation. The differential expression in different tissues was
explained in part by the presence of two active forms of caroteniod biosynthesis
genes (Lcyb & Psy) in comparison with house-keeping gene Actin-1 which is used
as control, to check equal loading and PCR quality check. Relative amplifications
were used to find out divergence in expressions of the targeted gene. Lycopene beta
cyclase showed maximum transcript signals in case of fruit and leaf tissues (Fig.4.6).
Significant level of expression was also found in shoot apex and newly growing bud
tissues. Whereas very minute traces were found in root and seed
tissues.
Figure 4.6: Photograph showing different tissues of sea buckthorn a. bud, b. apex,
c. root. d. leaf, e. seed, f. fruit. RT-PCR analysis of Hr-lcyb and Hr-Psy gene
transcripts in above mentioned tissues of sea buckthorn in comparison with internal
control actin-1. M stand for 1kb DNA ladder.

137
137
In case of phytoene synthase maximum transcript abundance was found in fruit, leaf
and apex. Some expression was also found in bud and seed tissues. In contrast to
Lcy gene no traces of transcripts were found in root tissues (Fig.4.6).
It can be recommended that these differences in gene expression level may be due
to environmental factors (growth stage, light), relatively low abundant RNAs all
factors contribute to variability. However, it was reported already that endogenous
and environmental factors, for example the type and the stage of the tissue
development, the mutations and the ultraviolet light, could also be the causes of
increased or reduced gene and protein expressions (Giuliano et al., 1993; Namitra et
al., 2011; Gady et al., 2012; Lazzeri et al., 2012).
The computational technique of AtGene express was considered to identify selective
expression using Arabidopsis thaliana microarrays data to monitor genome-wide
gene expression in sea buckthorn plant. It was assumed from Arabidopsis expression
pattern that lycopene beta cyclase gene expression in sea buckthorn would behave
the same as shown in fig. 4.7 a, b, c&d.
This was an indirect approach to estimate the protein abundance by examining the
gene expression level. Therefore, it is reasonable to explain that elevated levels of
gene expression observed in the leaf and fruit leads to the higher activity of these
enzymes in converting phytoene to carotene and lycopene in these tissues. This
observation was consistent with the high amount of carotenoid, especially Lycopene
accumulation in the petal and fruit tissues of tomato from high lycopene tomato
cultivar, Solanum lycopersicum KKU-T34003 (Krittaya and Klanrit, 2013). They
reported that Psy1 gene fragment have open reading frame of 1,239 base pair length,
which corresponds to our results (Krittaya and Klanrit, 2013). Similar sequence
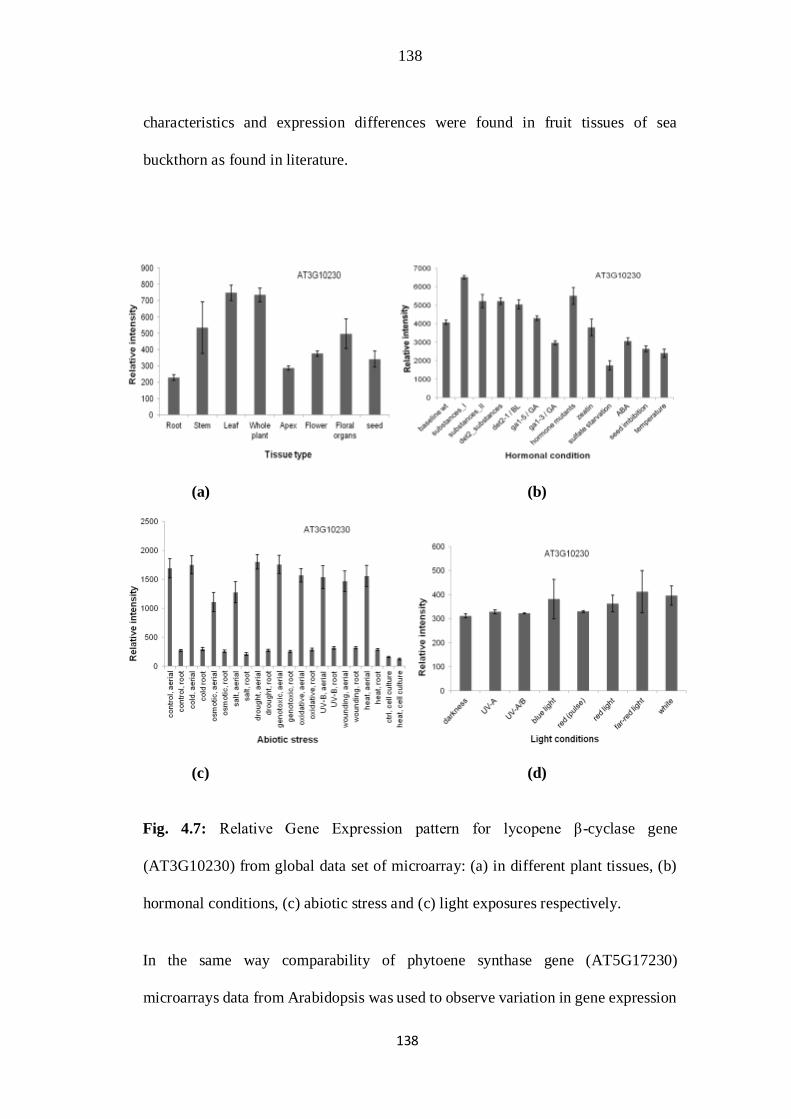
138
138
characteristics and expression differences were found in fruit tissues of sea
buckthorn as found in literature.
(a) (b)
(c) (d)
Fig. 4.7: Relative Gene Expression pattern for lycopene β-cyclase gene
(AT3G10230) from global data set of microarray: (a) in different plant tissues, (b)
hormonal conditions, (c) abiotic stress and (c) light exposures respectively.
In the same way comparability of phytoene synthase gene (AT5G17230)
microarrays data from Arabidopsis was used to observe variation in gene expression
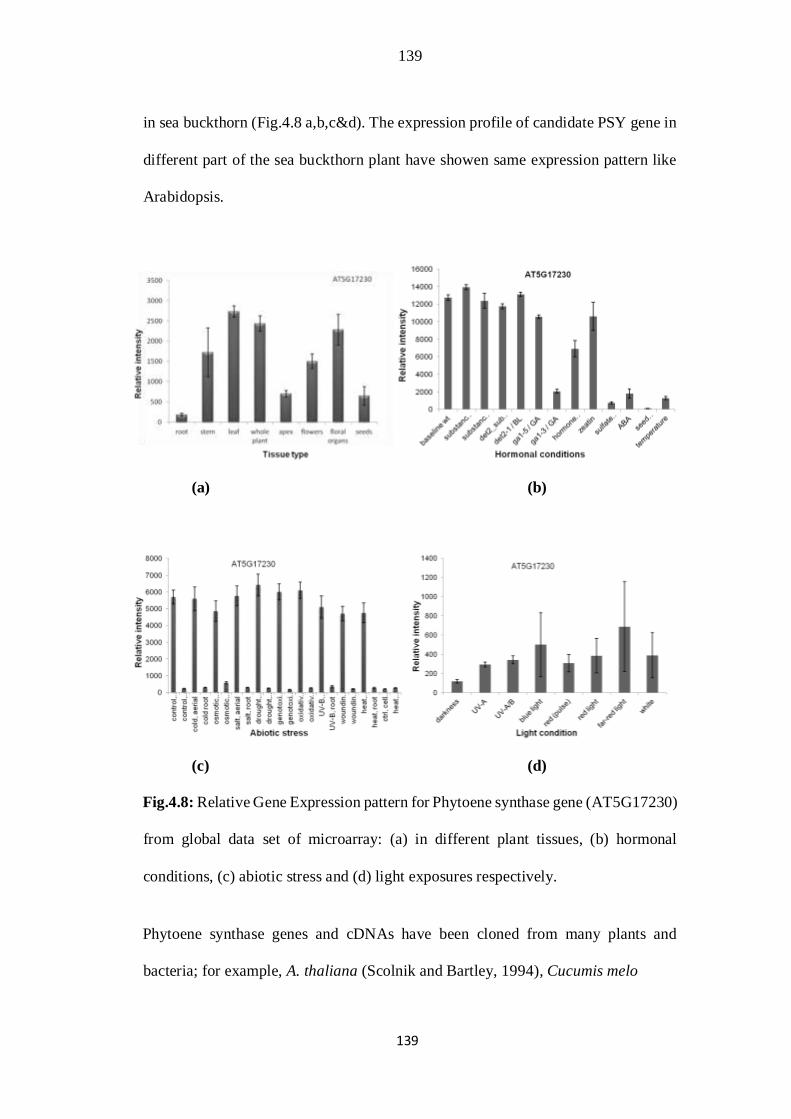
139
139
in sea buckthorn (Fig.4.8 a,b,c&d). The expression profile of candidate PSY gene in
different part of the sea buckthorn plant have showen same expression pattern like
Arabidopsis.
(a) (b)
(c) (d)
Fig.4.8: Relative Gene Expression pattern for Phytoene synthase gene (AT5G17230)
from global data set of microarray: (a) in different plant tissues, (b) hormonal
conditions, (c) abiotic stress and (d) light exposures respectively.
Phytoene synthase genes and cDNAs have been cloned from many plants and
bacteria; for example, A. thaliana (Scolnik and Bartley, 1994), Cucumis melo

140
140
(Karvouni et al., 1995), Narcissus pseudonarcissus (Schledz et al., 1996), Solanum
lycopersicum (Giuliano et al., 1993; Giorio et al., 2008) and Nicotiana tabacum
(Busch et al., 2002). Because of this rate-limiting feature, PSY has expanded much
interest to be cloned and targeted for transformation to boost carotenoid levels of food
crops.
4.3.3 Sequence analysis
The cDNA sequences of Hr-Lcyb &Hr-Psy were edited with Mac Vector™ 7.2.3
software. Both pairwise and multiple Sequence alignments tool were utilized to
identify region of similarities that may indicate functional, structural and/or
evolutionary relationships between two and multiple biological sequences. Pairwise
alignments were made on both nucleotide and amino acid levels respectively.
Pairwise alignment of new Hr-Lcyb sequence with the tomato gene sequence showed
considerable nucleotide variation randomly distributed throughout the whole
sequence as shown in fig. 4.9. The nucleotide sequences were translated in the
MacVector program. The default parameters were used. The fasta formate of new
Hr-Lcyb protein sequences were aligned in Mac-Vector 7.2 software to see the
difference in newly isolated amino acids sequence with previously isolated
sequences as shown in fig.4.10. Sequences which were too diverged disturbed the
alignments so they were removed manually while performing the multiple alignment
procedure. Similarly in case of Hr-Psy Pairwise alignments of new cDNA sequences
with the target tomato cDNA sequence show considerable variability in the new
sequence (Fig.4.11). The Hr-Psy nucleotide sequence were translated and aligned
with the tomato protein sequence to see the conservation in new sequence as shown
in fig. 4.12. Analysis of the homology of the deduced amino acid sequence of Hr-

141
141
Lcyb and Hr-Psy reveal various degrees of similarities and variations. Sequence
alignments revealed that the newly isolated sequences were not 100% identical to
other plant sequences; there are differences in the underlying coding sequences. The
conservation in regions of Hr-Lcyb were analyzed by comparing with the amino acid
sequence of S. lycopersicum,V. vinifera, N, pseudonarcissus, M. truncatula, N.taetta,
C. sinensis, L. barbarum which belongs to species that present caroteniods
accumulation. The strictly conserved amino acid residues are shown against a black
background as shown in Fig. 4.13. The possible amino acid changes found do not
alter the function of HrLcyb sequence isolated from sea buckthorn. The translated
amino acid sequence of Hr-Psy was aligned with other sequences from different
plant species including some member of family solanaceae, family convolvulaceae,
and asteraceae. The detail and accession no of the amino acid sequence uniqueness
and similarities for multiple sequence alignments of new and other phytoene
synthase enzymes, degree of sequence similarity and the locations of the more highly
conserved regions were compared and illustrated in Fig. 4.14. Differences were
found at both N-terminus and C-terminus domains of LCY and PSY proteins as
shown in case of multiple sequence alignments.

142
142
Fig. 4.9: Alignment of newly isolated Hr-Lcyb nucleotide sequence with its ortholog
from S. lycopersicum using MaVectorTM7.2.3. (Accerlrys Inc.) gcg/
Wisconsin pakage university of Wisconsin). The similar regions are shown
against a black background and differences are exposed against white
background.

143
143
Fig 4.10: Alignment of newly isolated Hr-Lcyb amino acid sequence with its ortholog
from S. lycopersicum using MaVectorTM7.2.3. (Accerlrys Inc.) gcg/ Wisconsin
pakage university of Wisconsin). The conservations in sequence are shown against
a black background and differences are exposed against white background.

144
144
Fig. 4.11: Alignment of newly isolated Hr-Psy nucleotide sequence with its ortholog
from S. lycopersicum using MaVectorTM7.2.3. (Accerlrys Inc.) gcg/ Wisconsin
pakage university of Wisconsin). The similar regions are shown against a black
background and differences are exposed against white background.

145
145
Fig. 4.12: Alignment of newly isolated Hr-Psy amino acid sequence with its ortholog from
S. lycopersicum using MaVectorTM7.2.3. (Accerlrys Inc.) gcg/ Wisconsin pakage
university of Wisconsin) software. The similarities are shown against a black
background and differences are exposed against white background.

146
146
Figure 4.13: Multiple sequence Alignment of newly isolated Hr-Lcyb amino acid sequence
with its ortholog from S. lycopersicum (NM_001247297), V. vinifera (JQ319639),
N. pseudonarcissus (GQ327929), M. truncatula (XM-003624977), N. tazetta
(JQ797381), C. sinensis (DQ235259) and L. barbarum (AY906864) using
MaVectorTM7.2.3. (Accerlrys Inc.) gcg/ Wisconsin pakage university of
Wisconsin). The strictly conserved regions are shown against a black background
and differences are exposed against white background.

147
147
Figure 4.14: Multiple Alignment of amino acid sequence of Hr-Psy gene with its
representative from other plant species. The names and accession numbers of these
members are: Lopomoea (AB499050), T. erecta (AF251015), S. lycopersicum
(EF534739), N. tabacum (JF461341) and Tomato (M84744). The strictly conserved
regions are shown against a black background and differences are exposed against
white background.

148
148
4.3.4 Phylogenetic Analysis
In order to show the inferred evolutionary relationships phylogenetic reconstruction
was carried out. The evolutionary history was inferred using the Neighbor-Joining
method (Saitou and Nei, 1987). The evolutionary distances were computed using the
Maximum Composite Likelihood method (Tamura et al., 2004). Evolutionary
analyses were conducted in MEGA5 (Tamura et al., 2011).
Figure 4.15: Phylogenetic analysis of Hr-Lcyb and other selected plant members. Neighbor
joining tree is constructed using MEGA 5.0 software. Values on the nodes indicate
the bootstrap replication of 1000. Phylogenetic inference was made using 25
complete coding sequences H. rhamnoides, S. lycopersicum (NM_001247297), C.
maxima (AY217103), C. sinensis (DQ496224), C. papaya
(DQ415894), P. trichocarpa (XM_002308867), R. communis (XM_002531452),

149
149
E. japonica (JX089591), V. vinifera (JQ319639), B. nivea (EU122344), C. moschata
(JN559395), S. europaea (AY789516), C. arietinum (XM_004493356),
M. truncantula (XM_003624977), G. max (XM_006576725), P. vulgaris
(HQ199604), A. palaestina (AF321534), L. barbarum (AY906864) Lpomoea_sp.
Kenyan (AB499055), N. tabacum (X81787), Chrysanthemum_X_morifolium
(AB205041) T. officinale (AB247456), N. pseudonarcissus (GQ327929) and N.
trzetta (JQ797381).
For this purpose the coding sequences of lycopene beta cyclase homologue from
diverse plant species H. rhamnoides, C. maxima, C. sinensis, C. papaya, P.
trichocarpa, R. communis, E. japonica, V. vinifera, B. nivea, C.moschata, S.
europaea, C. arietinum, M. truncantula, G. max, P. vulgaris, A. palaestina, L.
barbarum, Lpomoea_sp. Kenyan, N. tabacum, S. lycopersicum,
Chrysanthemum_X_morifolium, T.officinale, N. pseudonarcissus and N. trzetta were
used and tree was built as shown in Fig. 4.15
The detail of the plant species used for the Psy tree includes H. rhamnoides, P,mume,
F.vesca, G. max, C. arietinum, D.Kaki, C. sinensis, D. carota, A. deliciosa, M.
indica, p. trichocarpa, R.communis, M. esculenta, C. papaya, P. sitchensis, T.erecta,
C.melo,C.lanatus, M.charanita, M. cochinchinensis, S. lycopersicum, O. fragrans,
M. germanica, A. palaestina, C. roseus, G.jasminoides, C. canephora, B. oleracea,
B. napus, E. japonica, B. distachyon, T. aestivum, H. vulgare, P. juncea. The
sequences were edited alligned, analyze and neighbor joining tree in MEGA 5.0
software was constructed and displayed with tree view in Fig. 4.16. All positions
containing gaps and missing data were eliminated. To postulate a possible
phylogenetic relationship among this group of enzymes, we have used the maximum
likelihood method of sequence analysis.

150
150
Figure 4.16: The detail and accession no of the plant species for the phytoene synthase gene
which were used for the phylogenetic analysis includes H. rhamnoides, P,mume
(AB253628), F.vesca (XM_004289519), G. max (XM_003544910), C. arietinum
(XR_189445), D.Kaki (FJ594485), C. sinensis (EF545005), D. carota (DQ192187),
A. deliciosa (FJ797304), M. indica (JN001197), p. trichocarpa (XM_002327528),
R.communis (XM_002532929), M. esculenta (GU111719), C. papaya (DQ666828),
P. sitchensis (EF676285), T.erecta (AF251015), C.melo (GU361622), C.lanatus,
M.charanita (AY494789), M. cochinchinensis (KF233991), S. lycopersicum
(M84744), O. fragrans (JQ699273), M. germanica

151
151
(AY986508), A. palaestina (AY661705), C. roseus (HQ438241), G. jasminoides
(HQ599860), C. canephora (DQ157164), B. oleracea (JF920036), B. napus
(AB454517), E. japonica (JX097048), B. distachyon (XM_003579062), T. aestivum
(BT009537), H. vulgare (AK358888) and P. juncea (HM539711).
4.4 DISCUSSION
Sea buckthorn berries contain high levels of vitamin, carotenoid, tocopherol,
chlorophyll, flavonoid, and fatty acid. All the bioactive compounds have potentiality
of useful effects on human health (Singh, 2005; Suryakumar and Gupta, 2011).
Amongst all bioactive compounds carotenoids are the major compounds they have
several actions like anti-oxidant, anti-tumour and antimutagenic (Britton et al.,
2009). Earlier studies have shown that sea buckthorn berry has many difference in
the contents and composition of carotenoids (Andersson et al., 2009; Bal et al.,
2011). Despite of these investigation still there is no data reported on caroteniods
biosynthesis gene isolation from this wild medicinal entity.
In order to investigate the active genes that correlate with caroteniods accumulation
and biosynthesis full length coding sequences of Hr-Lcyb and HrPsy were isolated,
cloned and their sequences were analyzed by both pair wise and multiple sequence
alignments. The of Hr-Lcyb cDNAs has a complete ORF of 1503 bp and Hr-Psy
cDNA has an ORF of 1238 bp in length encoding proteins of 501aa and 412aa amino
acids residues respectively. The length of the cDNA remains the same as compared
to tomato sequence.The expected amino acid sequence of the sea buckthorn lycopene
beta cyclase (EC 5.5.1.19) is rather similar to those of the other recognized plant beta
cyclase from tomato, pepper, (Hugueney et al., 1995); and from tobacco and tomato
(Pecker et al., 1996). It also resembles the B cyclase from the V. vinifera, M.

152
152
truncatula, N. tazetta, C. sinensis, L. barbarum, and N. pseudonarcissus, as was
found in our multiple amino acid sequence alignments of these species. Multiple
sequence alignments make it possible to examine and find out relationships from an
evolutionary point of view by homological sequence. These finding implied that this
enzyme is of relatively same origin compared with the β cyclase of divergent species,
and that it may have come to pass by slight modification of a duplicated β cyclase
gene in tomato (Giuliano et al., 1993).
We have found a copy of this lycopene beta cyclase cDNA in Sea buckthorn. The
conserved domain of Hr-lycopene β-cyclase were also searched from NCBI
conserved domain search which appear to have evolved from the plant lycopene
cyclases consisting of NAD(P)-like binding domain (Fig.4.17).
Fig. 4.17: Conserved domains of Hr-Lycopene β-cyclase showing main
NAD_binding domain.
Comparison was done between predicted amino acid sequence of Hr-Psy and amino
acid sequence of different plant species. Protein alignments study revealed that the
protein encoded by Hr-Psy gene can be grouped in the class called terpene cyclase
that synthesize geranyl/farnesyl diphosphates and superfamily of transisoprenyl
diphosphate synthases. Further study of conserved domains and motifs showed that
Psy gene consist of six regions as follows, substrate binding pocket, substrate Mg2+

153
153
binding site, active site lid residue, aspartate-rich region-1 and aspartate-rich region-
2 (Fig. 4.18) which corresponds to findings of Lao et al.,
(2011). Although all of the homolog’s aligned varied considerably in their Ntermini.
Fig. 4.18: Conserved domains of Hr-Phytoene synthase gene consisting of six main
regions
The protein sequence alignments were used to identify the homologous sequences
and to employ conservation in sequence and structure which will be useful to
visualize the biochemical activities and biological function of these proteins.
Alignment at the protein level also indicated the difference of sequences among
different plant species. To look at the result in an evolutionary context we compared
the subsets of clusters representing four clusters for lycopene gene and five clusters
for phytoene synthase gene. It is demonstrated by this phylogenic tree that Hr-Lcyb
as well as Hr-Psy belongs to the correspondent subfamilies as anticipated, signifying
that our cDNA clones actually codes for the related enzyme. Phylogenetic analysis
showed considerable homology of our genes in all known carotenoid-producing
organisms.
RT-PCR was used to analyze Hr-Lcyb & Hr-Psy for their level of expression to
evaluate maximum accumulation of these gene transcripts in buds, apex, fruit, leaf,
and seed tissues of sea buckthorn. Normalization was done using constitutively

154
154
expressed actin-1 as internal control. The transcript signals of both Hr-Lcy and Hr-
Psy were strongly detectable in the fruit and leaf tissues. Although, a large variation
was noticed between different plants tissues positioned in the carotenoid
accumulation, we found that Lycopene is produced at low quantity in seed and root
tissues. It is reported in literature that ripening-specific genes are involve to activate
carotenoid pathway during ripening stage (Ronen et al., 1999; Ronen et al., 2000;
Alba et al., 2005).
Our results indicated that the comparative amount of corresponding carotenoids
biosynthesis enzymes mRNA changed considerably during different
developmental stages. It could be concluded from the Arabidopsis expression pattern
that caroteniods biosynthesis genes in different part of the sea buckthorn plant have
depicted the same expression pattern.
The accumulation of carotenoids is reported in the majority of plants tissue, such as
green shoot, flower, fruit, and seed and root tissues. Even though the content as well
as type of carotenoid in green tissues showed comparative conservation among most
of the plant species. whereas the level of carotenoid and its profiles in non green
parts, including flower, fruit and seed tissues, showed considerable variability under
the influence of main factors such as stage of development, environmental
conditions, stresses or a grouping of these factors (Howitt and Pogson, 2006).
Beta-carotene concentration usually enhanced with the stages of development. As
fruit matures, carotenoid contents were increased to larger concentration. These
caroteniods are responsible for orange red color in sea buckthorn berries that are
solely synthesized and accumulate during fruit ripening. The ripened fruit berries of
sea buckthorn were preferably chosen for our current gene study as there is much

155
155
caroteniods accumulation at this peak point which is steady with the perceptual
structure found in other plants like pepper, mango, and papaya. (Fabi et al., 2007;
Ha et al., 2007). Now it is fully recognized that transcriptional regulation of genes
that encodes phytoene synthase (PSY), is the major force that triggers the production
of carotenoids in different plant species. Phytoene synthase (PSY) is considered as
the first enzyme of the pathway that determines the rate at which the overall reaction
proceeds.
Considerable enhancement in carotenoids contents were reached through genetic
modification of carotenoids biosynthetic pathways in canola, rice, potato and maize
(Aluru et al., 2008; Shewmaker, 1999).
Wild fruits and berries formed one of the most important food sources of the people.
Gene reserve of these wild species can be used productively for genetic engineering
of food crops for better nutrition. Sea buckthorn plants are remarkable in their
capacity to synthesize a variety of macro and micro nutrients significant to human
health. The targeted expression of these important micro-nutrients gene can be
utilized to channeled metabolic flux in new pathway in grain crops with
micronutrient malnutrition. The lack of micronutrients is due to the deficiency of
vitamin and mineral elements in food. People in poor countries especially have to
face the danger of nutrient deficiency because their diet comprised chiefly on grain
and does not included most important fruits, vegetables and animal products.
Proteins obtained from plants make available amino acids essential to human health.
Staple crops can express proteins with much beneficial amino acids as a result of
modification. The valuable affects of berry and berry fraction on health of human
were widely examined and supported by investigation, which suggested a enormous

156
156
prospective for berry to maintain and promote humans health (Cheng et al., 2007;
Gao et al., 2000; Johansson et al., 2000; Rösch et al., 2003; Suleyman et al., 2002;
Shukla et al., 2006; Vijayaraghavan et al., 2006; Yang and Kallio, 2002; Yang and
Kallio, 2005).
The biosynthetic pathways can be functionally altered with the prospective for
improving the nutrition and survival properties of plants. This is of substantial value
to address food security in the face of adverse climatic changes. Our finding showed
that analyses of both Hr-Lcyb and Hr-Psy gene from sea buckthorn (Hippophae
rhamnoides) have identical structures and biological functions to the subsequent
genes from other plants species. The genetic diversity and wild gene pool of sea
buckthorn plant can be used for meeting current and future demands. The
identification and indispensable use of these berries and important molecules studied
generate baseline data for biotechnological interest in the plants. Many attempts to
improve vitamin A malnutrition were concentrated on manipulations of carotenoids
biosynthesis pathway enzymes for increasing the nutritional contents and functional
attributes of food grains. The activity of these genes and their encoded products can
be regulated and characterized in numerous ways.
4.5 CONCLUSIONS
Our study effort was to contribute to caroteniods gene identification, isolation and
analysis to provide important knowledge to caroteniods synthesis in this medicinal
plant. In this assessment I have summarized the existing knowledge of the carotenoid
transcript signals (LCYβ & PSY) and discussed about the knowledge of their
biosynthesis regulation in bud, shoot apex, roots, seeds and fruits. The importance
of these micro nutrients and protective effect conferred by sea buckthorn deserves

157
157
further investigation. This work on wild fruit berries will provide an advantage to
access all these compounds to combat micronutrient malnutrition in the developing
world, additionally one might argue that the quantities are generally higher, but use
of these is not sizable
4.6 GENERAL SUMMARY
This investigation symbolizes a novel and unique approach through an evaluation of
the nutritional use of traditional medicinal plants sea buckthorn by examining
essential micronutrients genes. Plants are well known for their biochemical
versatility, proficient with synthesis of almost all complements of dietary
micronutrient requirements; although having uneven distribution amongst various
plant portions. Sea buckthorn (Hippophae rhamnoides L.) is fruit-bearing plant
identified for therapeutic and nutraceutical potential. The biologically active
compounds found in sea buckthorn berries have fascinated major research interests
equally for their antioxidants and for central enzyme of metabolism. Its fruit, seed,
leaf and bark comprised of astounding range of nutrient elements like vitamins,
substantial oils and minerals. The majority of our regular staples crops are poor in
most of these nutritious elements.
Functional candidate’s genes from sea buckthorn were therefore chosen from the
pathway enzymes known to be involved in vitamins biosynthesis and metabolism.
Specific gene primers were designed and RT-PCR and PCR amplification was used
to amplify desired gene products. The products were sequenced to check the desired
gene amplification from sea buckthorn plant. The list of cDNA cloned and
sequenced included Ascorbate oxidase (AO), Folate (HPPK-DHPS), vitamin A
(LCYβ and PSY). Differences and similarities were found in homology and

158
158
phylogenic evaluation with other plant species. The first amplification was
concerned about full length cDNA of ascorbate oxidase (Ascorbic acid) gene.
Ascorbic acid (vitamin C) is a well-known for its dietetic significance; most
attributes of its metabolism and some attributes of its function in plants are feebly
understood similarly biosynthetic pathway has not been definitely recognized
nevertheless in the majority of tissues it attained millimolar concentration. We
carried out successful amplification of full length coding sequence of AO gene
(2158 bp) with reference to reference gene sequence with an open reading frame of
1737 bp. The total amino acid residues of Hr-AO were 719aa. Moreover differences
were found in length of new gene sequence in comparison with tomato gene
sequence shearing 87% gene homology. The relative abundance of the total RNA
coding for Hr-AO was estimated using semi-quantitative RT-PCR amplification
with maximum transcript accumulation in green leaf and young green fruit tissues.
This was the first report to differentiate the association involving the expression of
Hr-AO and fruit growth in such type of bush plant. This novel gene isolated from
sea buckthorn will help to understand the regulatory role of this enzyme in ascorbic
acid metabolism.
The second gene isolated was Folate (Hydroxymethyldihydropterin
pyrophosphokinase–dihydropteroate synthase) involved in the folate biosynthesis
pathway. The target gene HPPK-DHPS was successfully isolated and cloned from
sea buckthorn and its protein structure was compared with other plants. The
sequence analysis exposed that this novel gene was 2354 bp in size. The coding
region was interrupted by a single large intron. The length of the ORF was found to
be 1539 bp which was similar to its ortholog in tomato.

159
159
Prominent nucleotide differences found were randomly distributed throughout the
full sequence length. Semi-quantitative RT-PCR Expression analysis revealed
higher HrHPPK-DHPS transcripts accumulation in leaf and fruits tissues.
Phylogenetic reconstruction exposed gene duplication in soybean and Arabidopsis.
In this study an attempt was also made to compare the 3D homology modeling
structure of new protein, though considerable conservation in 3dimensional structure
of folate proteins was observed; however, notable differences in substrate binding
pockets were also visible. The study will have future implications in utilizing the
potential of wild medicinal plants for genetic improvement of folate deficient staple
food crops.
Third part of the research deals with carotenoids which produces yellow colors of
fruits and vegetables and are almost universally distributed in living things.
Carotenoids are well-known for its role in plant development and photosynthesis,
and are a chief source of vitamin A in humans nutritional. Among these caroteniods
lycopene is the carotene most commonly found in ripened fruit tissues. Lycopene
βcyclase catalyzed the conversion of Lycopene into β-carotene which is the major
dietary predecessor of vitamin A. The colored carotenoids are derived from
Phytoene. Phytoene synthase converts (GGDP) geranylgeranyl diphosphate to
Phytoene in the first stage of carotenoid biosynthesis pathway. Basic challenges were
to study two important caroteniods pathway genes Lycopene β-cyclase and phytoene
synthase from vitamin A rich plant “sea buckthorn” (Hippophae rhamnoides). The
newly isolated full length coding sequences of Hr-Lcyb and Hr-Psy showed
homology with original gene sequence. Despite the similarities in nucleotide gene
sequences several differences in structural domains were also found. These novel

160
160
genes sequence from wild plant sea buckthorn are closely linked with other
carotenoids biosynthetic genes and can therefore be useful in identifying novel
protein accumulation and function. Maximum transcript accumulation is accordingly
detected in a variety of plant tissues (leaf & Fruit), even though at trace level in some
tissues like seed and root. In this assessment we recapitulated the existing knowledge
of the carotenoid transcript signals and discussed what is acknowledged regarding
the regulation of their biosynthesis in bud, shoot apex, root, seed and fruit tissues.
This work recommends that the study of different genes encoding caroteniods with
diverse activity will also help to meet the problem associated with vitamin A
deficiency. The likelihood of intensifying the vitamins content of plants to recuperate
their nutritive significance is promising. However exploitation of natural usable gene
pool is worthwhile approach that should receive considerable attention in research.
Our effort added important knowledge after searching genes useful for future
improvement of food crops through biofortification.
The availability of biosynthetic genes from different species that can function
cooperatively to produce visually detectable products makes carotenoids
biosynthesis an ideal model system to explore engineering strategies. In addition to
the inherent value of carotenoids as chemicals for medicine and industry, carotenoids
biosynthesis is an excellent model system to develop and test tools for the creation
of novel multienzyme pathways in recombinant organisms using combinatorial and
directed evolution tool.Awareness and identification about the value of these
important micronutrient genes in sea buckthorn plant will increases its long term
value for genetic engineering in future. These genes will be further functionally

161
161
characterized and transformed to plants lacking such genes. Cereals are staple foods,
widely consumed and are deficient in essential micronutrients.
4.7 GENERAL CONCLUSIONS AND RECOMMENDATIONS
• Recent interests for investigating sea buckthorn plant and their fruit berries
were depended intimately on their reported health benefits and their valuable
bioactive compound to a certain extent. It necessitated an increase in the
understanding of these important contents of vitamins in the fruits, leaves and
berry raw material etc.
• The Micronutrients genes like ascorbic acid, folate and caroteniods pathway
genes rich in sea buckthorn leaves and fruits showed the presence of multiple
genes for such bio-chemicals, which have much significance for human
growth and health. These micronutrient genes can successfully be used for the
biofortification of staple food crops on large scale.
• In this thesis work the genes of vitamin C, folate and caroteniods were
investigated and compared with other plant species with remarkable
differences in sequence analysis and protein configuration. The functional
role of ascorbate oxidase has never been fully elucidated before, owing to the
complexity in discerning the existence of an enzyme specially oxidizing
ascorbate without any apparent benefit as a result of consumption of
ascorbate. The isolation and expression studies of ascorbate oxidase gene
from sea buckthorn will generate sufficient date in understanding their role.
• Futher more the identification of folate biosynthetic gene and dominant
carotenoids genes from sea buckthorn are promising for future biofortification

162
162
of staple food crops. The levels of these different biologically active
substances are relatively higher in comparision with other fruit and berries.
• The assessment of cumulative data from cDNA sequence, amino acid
sequence, phylogenetic reconstruction and expression pattern demonstrated
that there is a considerable variability in the genes constitution and
evolutionary association with other ortholog.
• The Cereals are staple foods with wider consumptions and are mostly
deficient in important micro-nutrients. The ortologues genes isolated in the
study will possibly be characterized functionally by transformation into
cereals crop (wheat, rice) to meet the problem of micronutrient malnutrition.
Ascorbic acid cannot be stored in the body; therefore this vitamin must be
acquired regularly from dietary sources.
• These studies approach entails genetic modification of plant folate
metabolism for improved bio-availability. The most evident and simple
approaches toward an increase in folate level is through engineering the folate
biosynthetic pathway genes.
• This work also recommends that the caroteniods genes with diverse activities
will also help to meet the problem associated with vitamin A deficiency. Most
efforts to alleviate vitamin A deficiency have focused on the manipulation of
carotenoids biosynthetic pathway enzymes to improve the nutritional content
or functional properties of grains.
• The identification and indispensable use of these berries and important
molecules studied generate baseline data for biotechnological interest in the
plants.

163
163
• This study has future implications in exploring the potential of wild medicinal
plant for genetic improvement of staple food crops for nutrients.
• In future investigations these genes will be cloned into expression vector
through Gateway cloning technology for functional characterization. The
activity of these genes and their encoded products can be regulated and
characterized in numerous ways.
4.8 LIST OF PUBLISHED PAPERS AND BOOKS
Asad Hussain Shah, S Dilnawaz A, Mubasher S, Shazia A, Ishtiaque K and Farhat B.
2007. Biochemical and nutritional evaluations of sea buckthorn (hippophae
rhamoides L. ssp. turkestanica from different locations of Pakistan. Pak. J. Bot.,
39 (6): 2059-2065.
Shazia arif, S Dilnawaz A, Asad H S, Lutful H, Shahid I A, Abdul H and Farhat B. 2010.
Determination of optimum harvesting time for vitamin C, Oil and mineral elements
in berries of sea buckthorn (hippophae rhamnoides) Pak. J. Bot., 42 (5): 3561-
3568.
Aurangzeb Rao, S Dilnawaz A, Mubashar S, Shahid I A, Asad H S, M. Fareed K, Sardar
A K, Saima S, Shazia A, Rizwan A and Maria G. 2013.. Antioxidant activity and
lipid peroxidation of selected wheat cultivars under salt stress.
Journal of Medicinal Plants Research Vol. 7(4), pp. 155-164.
Aurangzeb Rao, S Dilnawaz A, Mubashar S, Shahid I A, Asad H S, M. Fareed K, Saima
S, Shazia A and Rizwan A. 2013. Potential biochemical indicators improve salt
tolerance in fifteen cultivars of wheat (Triticum aestivum L.) from Pakistan
International Journal of Scientific & Engineering Research. Volume 4(6): ISSN
2229-5518.

164
164
Shazia Arif, M Ramzan K, S Dilnawaz A, Zaheer A and G Muhammad A. 2014. novel
hydroxymethyldihydropterin pyrophosphokinase-dihydropteroate synthase
(HHPK-DHPS) gene from a nutraceutical plant sea buckthorn, involved in folate
pathway is predominantly expressed in fruit tissue. Journal of BMC molecular
biology (accepted).
Books
Syed Dilnawaz Ahmed and Shazia A. 2010. A Book entitled “Comparisons of few
important bio-chemicals from Sea buckthorn: Bio-chemicals from Sea buckthorn
expressed at three stages of fruit development and folate genes for
biofortification”. ISBN-13: 978-3838383378 ISBN-10: 3838383370.
Shazia Arif, Syed Dilnawaz A and Asad HS. 2012. A Book entitled “ Sea Buckthorn A
Magic plant in Azad Kashmir” subtitle Modified Methods for the Isolation of High
quality RNA from Sea buckthorn rich in Polyphenols & secondary metabolites, by
LAMBERT Academic publishing. ISBN-13: 978-3-659-27183-0
ISBN-10: 3659271837

165
165
Chapter 5
LITERATURE CITED
Ahmad, S.D. and M. Kamal. 2002. Morpho-molecular characterization of local
genotypes of Hippophaė rhamnoides L. ssp. turkestanica a multi-purpose plant
from northern areas of Pakistan. Online J. Biol Sci., 2: 351-354.
Akulinin, I.A. 1958. Using sea buckthorn oil in treating burns. Sov. Med. 11: 137-
138.
Alam, Z. 2004. Chemical and Nutritional Constituents of Sea Buckthorn Juice. Pak.
J. Nutr., 3: 99-106.
Alba, R., P. Payton, Z. Fei, R. McQuinn, P. Debbie, G.B. Martin, S.D. Tanksley and
J.J. Giovannoni. 2005. Transcriptome and selected metabolite analyses reveal
multiple points of ethylene control during tomato fruit
development. Plant Cell, 17: 2954-2965.
Al-Madhoun, A., M. Sanmartin and A.K. Kanellis. 2003. Expression of ascorbate
oxidase isoenzymes in cucurbits and during development and ripening of
melon fruit. Postharvest Biol. Technol., 27: 137-146.
Aluru, M, Y. Xu, R. Guo, Z. Wang, S. Li, W. White, K. Wang and S. Rodermel.
2008. Generation of transgenic maize with enhanced provitamin A content. J.
Exp. Bot., 59: 3551-3562.

166
166
Andersson, S.C., K. Rumpunen, E. Johansson and M.E. Olsson. 2008. Tocopherols
and tocotrienols in sea buckthorn (Hippophae rhamnoides L.) berries during
ripening. J. Agric. Food Chem., 56: 6701-6706.
Andersson, S.C., M.E. Olsson, E. Johansson and K. Rumpunen. 2009. Carotenoids
in sea buckthorn (Hippophae rhamnoides L.) berries during ripening and use
of pheophytin a as a maturity marker. J. Agric. Food Chem., 57: 250-258.
Arimboor, R., V.V. Venugopalan, K. Sarinkumar, C. Arumughan and R.C.
Sawhney. 2006. Integrated processing of fresh Indian sea buckthorn
(Hippophaė rhamnoides) berries and chemical evaluation of products. J. Sci.
Food Agr., 86: 2345-2353.
Asad, H.S., S.D. Ahmed, M. Sabir, S. Arif, I. khaliq and F. batool. 2007.
Biochemical and nutritional evaluations of sea buckthorn (Hyppophae
rhamnoides L. spp. turkestanica) from different locations of Pakistan. Pak. J.
Bot., 39: 2059-2065.
Bailey, L.H. and E.Z. Bailey. 1978. Hortus Third. A concise dictionary of plants
cultivated in the United States and Canada. MacMillan Pub. Co. Inc. 1290 pp.
Bal, L.M., V. Meda, S.N. Naik and S. Satya. 2011. Sea buckthorn berries: A potential
source of valuable nutrients for nutraceuticals and cosmoceuticals.
Food Res. Int., 44: 1718-1727.
Barbehenn, R.V., A. Jaros, L. Yip, L. Tran, A.K. Kanellis and C.P. Constabel. 2008.
Evaluating ascorbate oxidase as a plant defense against leaf-chewing insect
using transgenic Poplar. J. Chem. Ecol., 34: 1331-1340.

167
167
Barnes, J., Y. Zheng and T. Lyons. 2002. Plant resistance to ozone: the role of
ascorbate. In: Omasa K, Saji H, Youssefian S, Kondo N, eds. Air pollution
and plant biochemistry: prospects for phytomonitoring and phytoremediation.
Berlin, Heidelberg, New York: Springer, 235-252 pp.
Basset, G.J., E.P. Quinlivan, S. Ravanel, F. Rébeillé, B.P. Nichols, K. Shinozaki,
M. Seki, L.C. Adams-Phillips, J.J. Giovannoni, III. J.F. Gregory and A.D.
Hanson. 2004. Folate synthesis in plants: the p-aminobenzoate branch is
initiated by a bifunctional PabA–PabB protein that is targeted to plastids. Proc.
Natl. Acad. Sci. USA, 101: 1496-1501.
Bekker, N.P. and A.I. Glushenkova. 2001. Components of certain species of the
Elagnaceae family. Chem. Nat. Comp., 37: 97-116.
Benzie, I.F.F. 1999. Prospective functional markers for defining optimal nutritional
status: Vitamin C. Proc. Nutr. Soc., 58: 1-8.
Bernath, J. and D. Foldesi. 1992. Sea buckthorn (Hippophaė rhamnoides L.): A
promising new medicinal and food crop. J Herbs Spices Med. Plants, 1: 27-
35.
Beveridge, T., T.S.C. Li, B.D. Oomah and A. Smith. 1999. Sea buckthorn products:
Manufacture and composition. J. Agric. Food Chem., 47: 34803488.
Beveridge, T.H.J. 2002. Opalescent and cloudy fruit juices: formation and particle
stability. Crit. Rev. Food Sci. Nutr., 42: 317-37.
Beveridge, T.H.J., M. Cliff and P. Sigmind. 2004. Supercritical carbon dioxide
percolation of sea buckthorn press juice. J. Food Qual., 27: 41-54.

168
168
Birghila, S., S. Dobrinas, N. Matei, V. Magearu, V. Popescu and A. Soceanu.
2004. Distribution of Cd, Zn and ascorbic acid in different stages of tomato
(Lycopersicum esculentum Solanaceae) plant growing. Rev. Chim-
Bucharest., 55: 683-685.
Boccio, J.R. and V. Lyengar. 2003. Iron deficiency – causes, consequences, and
strategies to overcome this nutritional problem. Biol. Trace Elem. Res., 94: 1-
31.
Boushey, C.J., S.A. Beresford, G.S. Omenn and A.G. Motulsky. 1995. A quantitative
assessment of plasma homocysteine as a risk factor for vascular disease.
Probable benefits of increasing folic acid intakes. JAMA., 274:
1049-1057.
Brattstrom, L. and D.E. Wilcken. 2000. Homocysteine and cardiobascular disease:
cause or effect? Am. J. Clin. Nutr., 72: 315-323.
Bruno, R.S., S.W. Leonard, J. Atkinson, T.J. Montine, R. Ramakrishnan, T.M.
Bray and M.G. Traber. 2006. Faster plasma vitamin E disappearance in
smokers is normalized by vitamin C supplementation. Free Radic Biol. Med.,
40: 689-697.
Britton, G., S. Liaaen-Jensen and H. Pfander. 2009. Carotenoids. Volume 5:
Nutrition and Health. Basel, Switzerland: Birkhauser Verlag. 8pp.
Brinster, R.L, J.M. Allen, R.R. Behringer, R.E. Gelinas and R.D. Palmiter. 1988.
Introns increase transcriptional efficiency in transgenic mice. Proc. Natl.
Acad. Sci. USA, 85: 836-840.

169
169
Buchman, A.R. and P. Berg. 1988. Comparison of intron-dependent and
intronindependent gene expression. Mol. Cell Biol., 8: 4395-4405.
Busch, M., A. Seuter and R. Hain. 2002. Functional analysis of the early steps of
carotenoid biosynthesis in tobacco. Plant Physiol., 128: 439-453.
Caballero, B., 2003. Fortification, supplementation, and nutrient balance. Eur. J.
Clin. Nutr., 57: S76-S78.
Calvaresi, E., J. Bryan. 2001. B vitamins, cognition, and aging: a review. J. Gerontol.
B. Psychol. Sci Soc Sci., 56: 327-339.
Carr, A.C. and B. Frei. 1999. Toward a new recommended dietary allowance for
vitamin C based on antioxidant and health effects in humans. Am. J. Clin.
Nutr., 69: 1086-1107.
Carvalho, J. L.B., C.J. Lima and P.H. Medeiros. 1981. Ascorbate oxidase from
Cucurbita maxima. Phytochem., 20: 2423-2424.
Cenkowski, S., R. Yakimishen, R. Przybylski and W.E. Muir. 2006. Quality of
extracted sea buckthorn (Hippophae rhamnoides L.) seed and pulp oil.
Can. Biosyst. Eng., 48: 9-16.
Chakrabarty, R., R. Banerjee, S.M. Chung, M. Farman, V. Citovsky, S.A.
Hogenhout, T. Tzfira and M. Goodin. 2007. pSITE vectors for stable
integration or transient expression of autofluorescent protein fusions in plants:
probing Nicotiana benthamiana-virus interactions. Mol. Plant-
Microbe Interact., 20: 740-750.

170
170
Charles, A.D., T. McGhie, R. Wibisono, M. Montefiori, R.P. Hellens and A.C. Allan.
2009. The kiwifruit lycopene beta-cyclase plays a significant role in carotenoid
accumulation in fruit. J. Exp. Bot., 60: 3765-3779.
Chauhan, A.S., M.N. Rekha, R.S. Ramteke and W.E. Eipeson. 2001. Preparation and
quality evaluation of processed products from sea buckthorn (Hippophaė
rhamnoides L.) berries. Beverage Food World, 27: 31-34.
Cheng, J.Y., K. Kondo, Y. Suzuki, Y. Ikeda, X. Meng and K. Umemura. 2007.
Inhibitory effects of total flavones of Hippophaë rhamnoides L. on thrombosis
in mouse femoral artery and in vitro platelet aggregation. Life Sci., 72: 2263-
2271.
Chen, T. 1988. Studies of the biochemical composition of Hippophaė and its quality
assessment in Gansu Province. Hippophaė, 1: 19-26.
Chen, Y., F. Li and E.T. Wurtzel. 2010. Isolation and characterization of the Z-ISO
gene encoding a missing component of carotenoid biosynthesis in plants.
Plant Physiol., 153: 66-79.
Chou, W.M., T.Shigaki, C. Dammann, Y.Q. Liu and M.K. Bhattacharyya. 2004.
Inhibition of phosphoinositide-specific phospholipase C results in the
induction of pathogenesis related genes in soybean. Plant Biol., 6: 664-672.
Chugaeva, V.N., A.I. Gurvich and M.G.Ushakova. 1964. Mater. Conf. on Vitamins
from Natural Raw Mat. Kuybushev., 172-177pp
Chung, S. and R. Perry. 1989. Importance of introns for expression of mouse
ribosomal protein gene rpL32. Mol. Cell Biol., 9: 2075-2082.

171
171
Clotault, J., D. Peltier, R. Berruyer, M. Thomas, M. Briard and E. Geoffriau. 2008.
Expression of carotenoids biosynthesis genes during carrot root development.
J. Exp. Bot., 59: 3563-73.
Clough, S.J. and A.F. Bent. 1998. Floral dip: a simplified method for
Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J., 16:
735-743.
Cox, B.D., M.J. Wlichelow and A.T. Prevost. 2000. Seasonal consumption of salad
vegetables and fresh fruit in relation to the development of cardiovascular
disease and cancer. Public Health Nutr., 3: 19-29.
Cunningham, F.X., Z.J. Sun, D. Chamovitz, J. Hirschberg and E.
Gantt. 1994. Molecular structure and enzymatic function of lycopene cyclase
from the cyanobacterium Synechococcus sp. strain PCC7942. Plant Cell, 6:
1107-1121.
Darmer, G. 1952. Der Sanddornals Wild-und Kulturepflaze. Leipzig, Germany: S.
Hirzel Verlag – Publisher. 89 pp.
Darnton, H.I. and R. Nalubola. 2002. Fortification strategies to meet micronutrient
needs: successes and failures. Proc. Nutr. Soc., 61: 231-241.
Davey, M.W., M.V. Montagu, D. Inze, M. Sanmartin, A. Kanellis, N. Smirnoff,
I.F.F. Benzie, J.J. Strain, D. Favell and J. Fletcher. 2000. Plant L-ascorbic acid:
chemistry, function, metabolism, bioavailability and effects of processing. J.
Sci. Food Agr., 80: 825-860.
David, T.J. 1997. Progression in protein structure prediction. Curr. Optn Struc.

172
172
Btol., 7: 377-387.
Deli J, P.P. Molnar. 2002. Carotenoids: analysis, isolation, structure elucidation.
Curr. Org. Chem., 6: 1197-1219.
Deman, J.M. 1973. Principle of Food Chem. Oxford University Press, London. 311-
320 pp.
Deng-Ke, N. and Y. Yu-Fei. 2011. Why eukaryotic cells use introns to enhance gene
expression: Splicing reduces transcription-associated mutagenesis by
inhibiting topoisomerase I cutting activity. Biol. Direct., 6: 24.
De-Tullio, M.C., S. Ciraci, R. Liso and O. Arrigoni. 2007. Ascorbic acid oxidase is
dynamically regulated by light and oxygen. A tool for oxygen management in
plants. J. Plant Physiol., 164: 39-46.
Dharmananda, S. 2004. Sea buckthorn, Institute of Traditional Medicine, Portland,
Oregon. http://www.itmonline.org/arts/sea buckthorn.htm.
Diallinas, G., I. Pateraki, M. Sanmartin, A. Scossa, E. Stilianou, N.J. Panopoulos and
A.K. Kanellis. 1997. Melon ascorbate oxidase: cloning of a multigene family,
induction during fruit development and repression by wounding.
Plant Mol. Biol. 34: 759- 770.
Dong, H., Y. Deng, J. Mu, Q. Lu, Y. Wang, Y. Xu, C. Chu, K. Chong, C. Lu, J. Zuo.
2007. The Arabidopsis Spontaneous Cell Death1 gene, encoding a
zetacarotene desaturase essential for carotenoid biosynthesis, is involved in
chloroplast development, photoprotection and retrograde signalling. Cell Res.,
17: 458-470.

173
173
Douce, R., J. Bourguignon, M. Neuburger and F. Rébeillé. 2001. The glycine
decarboxylase system: a fascinating complex. Trends Plant Sci., 6: 167-176.
Doyle, J.J. and J.L. Doyle. 1987. A rapid DNA isolation procedure for small
quantities of fresh leaf tissue. Phytochem. Bull., 19: 11-15.
Eleonora, N.,A.D. Venere, N. Rosato, A. Rossi, A.F. Agro and G. Mei. 2006.
physico-chemical properties of molten dimer Ascobate oxidase. The FEBS J.,
273: 5194-5204.
Eliseeve, I.P. 1989. Evolutionary genetic aspects in assessment of achievements and
perspectives of sea buckthorn selection in USSR. Proc. Int. Symp. Sea
buckthorn (Hippophae rhamnoides L.), Xian, China. 184-193 pp.
Esaka, M., H. Fukui, K. Suzuki and K. Kubota. 1989. Secretion of ascorbate oxidase
by suspension-cultured pumpkin cells. Phytochem., 28: 117-119.
Esaka, M., K. Fujisawa, M. Goto and Y. Kisu. 1992. Regulation of ascorbate oxidase
expression in pumpkin by auxin and copper. Plant Physiol., 100: 231-237.
Esaka, M., T. Hattori, K. Fujisawa, S. Sakajo and T. Asahi. 1990. Molecular cloning
and nucleotide sequence of full-length cDNA for ascorbate oxidase from
cultured pumpkin cells. Eur. J. Biochem., 191: 537-541.
Fabi, J.P., B.R. Cordenunsi, G.P. de Mattos Barreto, A.Z. Mercadante, F.M. Lajolo,
J.R. Oliveira do Nascimento. 2007. Papaya fruit ripening: response to ethylene
and 1-methylcyclopropene (1-MCP). J. Agric. Food Chem., 55:
6118-6123.
Fabrice, R., D. Macherel, J.M. Mouillon, J.R. Garin and D. Roland. 1997. Folate
biosynthesis in higher plants: purification and molecular cloning of a

174
174
bifunctional 6-hydroxymethyl 7,8- dihydropterin
pyrophosphokinase/7,8dihydropteroate synthase Localized in mitochondria.
The EMBO J., 16: 947957.
Fan, J., X. Ding and W. Gu. 2007. Radical-scavenging proanthocyanidins from sea
buckthorn seed. Food Chem., 102: 168-177.
Farver, O., S. Wherland and I. Pecht. 1994. Intramolecular electron transfer in
ascorbate oxidase is enhanced in the presence of oxygen. J. Biol. Chem., 269:
22933-6.
Felsenstein, J. 1985. Confidence limits on Phylogenies: An approach using the
Bootstrap. Evolution, 39: 783-791.
Fotopoulos, V., M. Sanmartin and A.K. Kanellis. 2006. Effect of ascorbate oxidase
over-expression on ascorbate recycling gene expression in response to agents
imposing oxidative stress. J. Exp. Bot., 57: 3933-3943.
Foyer, C.H., P. Descourvieres and K.J. Kunert. 1994. Protection against oxygen
radicals: an improved defence mechanism study in transgenic plants. Plant Cell
Environm., 17: 507-523.
Fraser, P.D. and P.M. Bramley. 2004. The biosynthesis and nutritional uses of
carotenoids. Prog Lipid Res., 43: 228-265.
Gady, A.L.F., W.H. Vriezen, M.H.B.J. Van de Wa, P. Huang, A.G. Bovy and
R.G.F. Visser. 2012. Induced point mutations in the phytoene synthase 1 gene
cause differences in carotenoid content during tomato fruit ripening. Mol.
Breeding, 29: 801-812.

175
175
Gallagher, C.E., P.D. Matthews, L. Faqiang and E.T. Wurtzel. 2004. Gene
duplication in the carotenoid biosynthetic pathway preceded evolution of the
grasses. Plant Physiol., 135: 1776-1783.
Gamas, P., C. Niebel Fde, N. Lescure and J. Cullimore. 1996. Use of a subtractive
hybridization approach to identify new Medicago truncatula genes induced
during root nodule development. Mol. Plant Microbe Interact., 9: 233-42.
Gao, X.Q., M. Ohlander, N. Jeppsson, L. Bjork and V. Trajkovski. 2000. Changes
in antioxidant effects and their relationship to phytonutrients in fruits of sea
buckthorn (Hippophae rhamnoides L.) during maturation. J. Agric. Food
Chem., 48: 1485-1490.
Garcia-Pineda, E., E. Castro-Mercado and E. Lozoya-Gloria. 2004. Gene expression
and enzyme activity of pepper (Capsicum annuum L.) ascorbate oxidase during
elicitor and wounding stress. Plant Sci., 166: 237–243
Gatin, J.I. 1963. Sea buckthorn, Moscow. 159 pp.
George, D.P., M. Sanmartin, A. Scossa, E. Stilianou, N.J. Panopoulos and A.K.
Kanellis. 1997. Melon ascorbate oxidase: cloning of a multigen family,
induction during fruit development and repression by wounding. Plant Mol.
Biol., 34: 759-770.
Giuffrida, D., A. Pintea, P. Dugo, G. Torre, R.M. Pop and L. Mondello. 2011.
Determination of carotenoids and their esters in fruits of sea buckthorn (Hippophae rhamnoides
L.) by HPLC-DAD-APCI-MS. Phytochem. Anal., 23: 267-273.

176
176
Giuliano, G., G.E. Bartley and P. Scolnik. 1993. Regulation of caroteniods
biosynthesis during tomato development. Plant Cell, 5: 379-387.
Goodwin, T.W. 1980. The biochemistry of the carotenoids, 2nd edition volume 1.
Landon. Chapman and Hall 19, London, NY.
Gorbatsova, J., T. Lõugas, R. Vokk, and M. Kaljurand. 2007. Comparison of the
contents of various antioxidants of sea buckthorn berries using CE.
Electrophoresis, 28: 4136-4142.
Goyer, A., V. Illarionova, S. Roje, M. Fischer, A. Bacher and A.D. Hanson. 2004.
Folate biosynthesis in higher plants. cDNA cloning, heterologous expression,
and characterization of dihydroneopterin aldolases. Plant Physiol., 135: 103111.
Gutzeit, D., M. Sabine, J. Gerold, W. Peter and R. Michael. 2008. Folate content in
sea buckthorn berries and related products (Hippophaë rhamnoides L. ssp.
rhamnoides): LC-MS/MS determination of folate vitamer stability influenced
by processing and storage assessed by stable isotope dilution assay. Anal.
Bioanal. Chem., 391: 211-219.
Hampele, I.C., A. D'Arcy, G.E. Dale, D. Kostrewa, J. Nielsen, C. Oefner, M.G. Page,
H.J. Schönfeld , D. Stüber and R.L. Then. 1997. Structure and Function of the
dihydropteroate Synthase from Staphylococcus aureus. J. Mol. Biol.
268: 21-30.
Hanson, A.D. 2010. Developmental and feedforward control of the expression of
folate biosynthesis genes in tomato fruit. Mol. Plant., 3: 66-77.

177
177
Hagihara, T., M. Hashi, Y. Takeuchi and N. Yamaoka. 2004. Cloning of soybean
genes induced during hypersensitive cell death caused by syringolide elicitor.
Planta, 218: 606–614
Hakkinen, S.H., S.O. Karenlampi, I.M. Heinonen, H.M. Mykkanen and A.R.
Torronen. 1999. Content of the flavonols quercetin, myricetin, and kaempferol
in 25 edible berries. J. Agric. Food Chem., 47: 2274-9.
Hanson, A.D., D.A. Gage and Y. Shachar-Hill. 2000. Plant one carbon
metabolism and its engineering. Trends Plant Sci., 5: 206-213.
Hanson, A.D. and S. Roje. 2001. One-carbon metabolism in higher plants. Annu.
Rev. Plant Physiol. Plant Mol. Biol., 52: 119-137.
Havel, T.F. and M.E. Snow. 1991. A new method for building protein conformations
from sequence alignments with homologous of known
structures. J. Mol. Btol., 217: 1-7.
Ha, S.H., J.B. Kim, J.S. Park, S.W. Lee and K.J. Cho. 2007. A comparison of the
carotenoid accumulation in capsicum varieties that show different ripening
colours: deletion of the capsanthin-capsorubin synthase gene is not a
prerequisite for the formation of a yellow pepper. J. Exp. Bot., 58: 31353144.
Heimann, W. 1980. Fundamentals of Food Chemistry 1st Edn., Ellis Horwood
Limited.
Heyn, P., T.K. Alex, T. Pavel and M.N. Karla. 2015. Introns and gene expression:
Cellular constraints, transcriptional regulation, and evolutionary consequences.
BioEssays, 37: 148-154.
Hirschberg, J. 2001. Carotenoid biosynthesis in flowering plants. Curr. Opin. Plant

178
178
Biol., 4: 210-218.
Hong, Y.L., P.A. Hossler, D.H. Calhoun and S.R. Meshnick. 1995. Inhibition of
recombinant Pneumocystis carinii dihydropteroate synthetase by sulfa drugs.
J. Antimicrob. Chemother., 39: 1756-1763.
Hossain, T., I. Rosenberg, J. Selhub, G. Kishore, R. Beachy and K. Schubert. 2004.
Enhancement of folates in plants through metabolic engineering. Proc. Natl.
Acad. Sci. USA, 101: 5158-5163.
Howitt, C.A. and B.J. Pogson. 2006. Carotenoid accumulation and function in seeds
and non-green tissues. Plant Cell Environ., 29: 435-445.
Hugueney, P., A. Badillo, H.C. Chen, A. Klein, J. Hirschberg, B. Camara and M.
Kuntz. 1995. Metabolism of cyclic carotenoids: a model for the alteration of
this biosynthetic pathway in Capsicum annuum chromoplasts. Plant J., 8: 417-
424.
Hultberg, B., K.N. Isaksson and L. Gustafson. 2001. Markers for the functional
availability of obalamin/folate and their association with neuropsychiatric
symptoms in the elderly. Int. J. Geriatr Psych., 16: 873-878.
Ioannidi E., M.S. Kalamaki, C. Engineer , I. Pateraki , D. Alexandrou , I.
Mellidou, J. Giovannonni and A.K. Kanellis. 2009. Expression profiling of ascorbic acid-related
genes during tomato fruit development and ripening and in response to stress conditions. J. Exp.
Bot., 60: 663-78.
http://www.ncbi.nlm.nih.gov/pubmed/?term=Kalamaki%20MS%5BAuthor%5D&cauthor=true&cauthor_uid=19129160
http://www.ncbi.nlm.nih.gov/pubmed/?term=Kalamaki%20MS%5BAuthor%5D&cauthor=true&cauthor_uid=19129160

179
179
Jabrin, S., S. Ravanel, B. Gambonnet, R. Douce and F. Rébeillé. 2003. One-carbon
metabolism in plants. Regulation of tetrahydrofolate synthesis during
germination and seedling development. Plant Physiol., 131: 1431-1439.
Jeppsson, N. and X.Q. Gao. 2000. Changes in the contents of kaempherol, quercetin
and L-ascorbic acid in sea buckthorn berries during maturation. Agri. Food Sci.
Finland, 9: 17-22.
Johansson, A., H. Korte, B. Yang, J. Stanley and H. Kallio. 2000. Sea buckthorn
berry oil inhibits platelet aggregation. J. Nutr. Biochem., 11: 491-495.
Joshipura, K., A. Ascherio, A.E. Manson, M.J. Stampfer, E.B. Rimm, F.E Speizer,
C.H. Hennekens, D. Spiegeleman and W.C. Willett. 1999. Fruit and vegetable
intake in relation to risk of ischaemic stroke. J. Am. Med. Assoc., 282: 1233-
1239.
Kallio, H., B. Yang and P. Peippo. 2002. Effects of different origins and harvesting
time on vitamin C, tocopherols, and tocotrienols in sea buckthorn (Hippophaë
rhamnoides) berries. J. Agric. Food Chem., 50: 6136-6142.
Kallio, H., B. Yang, P. Peippo, R. Tahvonen and R. Pan. 2002. Triacylglycerols,
glycerophospholipids, tocopherols, tocotrienols in berries and seeds of two
species (ssp. sinensis and mongolica) of sea buckthorn (Hippophaė
rhaminoides). J. Agric. Food Chem., 50: 3004-3009.
Kallio, K., B.R Yang, R. Tahvonen and M. Hakala. 1999. Composition of sea
buckthorn berries of various origins. Proc. of Int. Symp. on Sea Buckthorn
(Hippophaė rhamnoids L.), Beijing, China.

180
180
Karvouni, Z., I. John, J.E. Taylor, C.F. Watson, A.J. Turner and D. Grierson. 1995.
Isolation and characterization of a melon cDNA clone encoding phytoene
synthase. Plant Mol. Biol., 27: 1153-1162.
Kato, N. and Esaka M. 2000. Expansion of transgenic tobacco protoplasts expressing
pumpkin ascorbate oxidase is more rapid than that of wild-type protoplasts.
Planta. 210: 1018-1022.
Katiyar, S.K., K. Sharma, N. Kumar and A.K. Bhatia. 1990. Composition of some
unconventional Himalayan wild fruits. J. Food Sci. Technol., 27: 309-310.
Khan, M.I. and M. Kamran. 2004. “Cost analysis of sea buckthorn berries picking
techniques and food products preparation at domestic level: A study conducted
in Ghulkin, Upper Hunza, Northern Areas of Pakistan”. WWF-
Pakistan, Jutial, Gilgit.
Khan, M.R., I. Khan and G.M. Ali. 2013. MPF2-like MADS-box genes affecting
SOC1 and MAF1 expression are implicated in flowering time control. Mol.
Biotechnol., 54: 25-36.
Krittaya, S. and P. Klanrit. 2013. cDNA cloning and expression analyses of phytoene
synthase 1, phytoene desaturase and ζ-carotene desaturase genes from
Solanum lycopersicum KKU-T34003. Songklanakarin. J. Sci. Technol.,
35: 517-527.
Kumar, S., J. Dudley, M. Nei and K. Tamura. 2008. MEGA: a biologist-centric
software for evolutionary analysis of DNA and protein sequences. Brief
Bioinform., 9: 299-306.
http://www.ncbi.nlm.nih.gov/sites/entrez?cmd=retrieve&db=pubmed&list_uids=18417537&dopt=AbstractPlus
http://www.ncbi.nlm.nih.gov/sites/entrez?cmd=retrieve&db=pubmed&list_uids=18417537&dopt=AbstractPlus

181
181
Lampe, J.W. 1999. Health effects of vegetables and fruit: assessing mechanism of
action in human experimental studies. Am. J. Clin. Nutr., 70: 475S-490S.
Lao, Y.M.1., L. Xiao, Z.W. Ye, J.G. Jiang and S.S. Zhou. 2011. In silico analysis of
phytoene synthase and its promoter reveals hints for regulation mechanisms of
carotenogenesis in Duanliella bardawil, Bioinformatics, 27:
2201-8.
Lawrence, M.C., P. Iliades, R.T. Fernley, J. Berglez, P.A. Pilling and I.G. Macreadie.
2005. The three-dimensional structure of the bifunctional 6hydroxymethyl-7,8-
dihydropterin pyrophosphokinase/dihydropteroate synthase of Saccharomyces
cerevisiae. J. Mol. Biol., 348: 655-70.
Lazzeri, V., V. Calvenzani, K. Petroni, C. Tonelli, A. Castagna and A. Ranieri. 2012.
Carotenoid profiling and biosynthetic gene expression in flesh and peel of wild
type and hp-1 tomato fruit under UV-B depletion. J. Agric. Food Chem., 60:
4960-4969.
Levine, M., S.C. Rumsey, R. Daruwala, J.B. Park and Y.H. Wang. 1999. Criteria
and recommendations for vitamin C intake, Journal of American Med. Assn.,
281: 1415-1423.
Lee, M.H. and C.R. Dawson. 1973. Ascorbate oxidase. Spectral characteristics of
the enzyme. J. Biol. Chem., 248: 6603-6609.
Lian, Lu T.S.G., S.K. Xue and X.L. Chen. 2000. Biology and chemistry of the genus
Hyppophae. Dansu Scientific and Technological publishing House, Lanzhou,
China. 88-91 pp.

182
182
Lian, Y.S., S.G. Lu, S.K. Xue and X.L. Chen. 2000. Biology and Chemistry of the
Genus Hippophae. Lanzhou Gansu Science Technology Press. 1-226 pp.
Li, M., F. Ma, D. Liang, J.Li and Y. Wang. 2010. Ascorbate biosynthesis during
early fruit development is the main reason for its accumulation in Kiwi. PLos
ONE, 5: e14281.
Lisiewska, Z., W. Kmiecik and A. Korus. 2006. Content of vitamin C, carotenoids,
chlorophylls and polyphenols in green parts of dill (Anethum graveolens L.)
depending on plant height. J. Food Compos. Anal., 19: 134-140.
Liso, R.1., M.C. D. Tullio, S. Ciraci, R. Balestrini, N. L. Rocca, L. Bruno, A.
Chiappetta, M.B. Bitonti, P. Bonfante and O. Arrigoni. 2004. Localization of
ascorbic acid, ascorbic acid oxidase, and glutathione in roots of Cucurbita
maxima L. J. Exp. Bot., 55: 2589-97.
Li, T.S.C. 1999. Sea buckthorn: New crop opportunity. In J. Janick (Ed.),
Perspectives on new crops and new uses. Alexandria, VA: ASHS Press.
335337 pp.
Liu, S.W. and T.N. He. 1978. The genus Hippophae from Qing-Zang Plateau. Acta
Phytotax. Sinic. 16: 106-108. (in Chinese).
Loewus, F.A. and M.W. Loewus. 1987. Biosynthesis and metabolism of
ascorbicacid in plants. Crit. Rev. Plant Sci., 5: 101-119.
Lucock, M. 2000. Folic acid: nutritional biochemistry, molecular biology, and role
in disease processes. Mol. Genet. Metab., 71: 121-38.
Lu, R. 1992. Sea buckthorn-A multipurpose plant species for fragile mountains,
ICIMOD Occasional Paper No. 20. Katmandu, Nepal. 62 pp.
http://www.ncbi.nlm.nih.gov/pubmed?term=Balestrini%20R%5BAuthor%5D&cauthor=true&cauthor_uid=15520029
http://www.ncbi.nlm.nih.gov/pubmed?term=Balestrini%20R%5BAuthor%5D&cauthor=true&cauthor_uid=15520029
http://www.ncbi.nlm.nih.gov/pubmed?term=La%20Rocca%20N%5BAuthor%5D&cauthor=true&cauthor_uid=15520029
http://www.ncbi.nlm.nih.gov/pubmed?term=La%20Rocca%20N%5BAuthor%5D&cauthor=true&cauthor_uid=15520029
http://www.ncbi.nlm.nih.gov/pubmed?term=La%20Rocca%20N%5BAuthor%5D&cauthor=true&cauthor_uid=15520029
http://www.ncbi.nlm.nih.gov/pubmed?term=La%20Rocca%20N%5BAuthor%5D&cauthor=true&cauthor_uid=15520029
http://www.ncbi.nlm.nih.gov/pubmed?term=Chiappetta%20A%5BAuthor%5D&cauthor=true&cauthor_uid=15520029

183
183
Luwe, M.W.F., U. Takahama and U. Heber. 1993. Role of ascorbate in detoxifying
ozone in the apoplast of spinach (Spinacia oleracea L) leaves. Plant Physiol.,
101: 969-976.
Maite, S. I. Pateraki, F. Chatzopoulou and A.K. Kanellis. 2007. Diferential
expression of the ascorbate oxidase multigene family during fruit development
and in response to stress. Planta, 225: 873-885.
Maoka, T., K. Mochida, M. Kozuka, Y. Ito, Y. Fujiwara, K. Hashimoto, F. Enjo, M.
Ogata, Y. Nobukuni, H. Tokuda and H. Nishino. 2001. Cancer
chemopreventive activity of carotenoids in the fruits of red paprika Capsicum
annuum L. Cancer Lett., 172: 103-109.
Marcus, C.C.W.E. 2005. Sea buckthorn - Hippophae rhamnoides L. A Whole Food
Answer to Better Nutrition. riyasilver.trdinfotech.com/sea buckthorn.htm
Mashkovski, A.D. 1997. Lecarstvennaye Sredstv., Moscow. 625 pp.
Ma, Z., Y. Cui and G. Feng, 1989. Studies on the fruit character and biochemical
compositions of some forms within Chinese sea buckthorn (Hippophaė
rhamnoides subsp. sinensis) in Shanxi, China. Proc. Of int. symp. on sea
buckthorn (H. rhamnoids L.), Xian, China. 106-113 pp.
Mingyu, X., S. Xiaoxuan and C. Jinhua. 2001. The medicinal research on sea
buckthorn. International workshop on sea buckthorn. 18-21 pp.
Mironov, V.A., T.N. Guseva-Denskaya, Y. Dubrovina, G.A. Osipov, E.A.
Shabanova, A.A. Nikulin, N.S. Amirov and I.G. Trubitsina. 1989. Chemical

184
184
composition and biological activity of extracts from sea buckthorn fruit
components. Khim-Farm. Zh., 23: 1357-64. (In Russian).
Moser, O. and A.K. Kanellis. 1994. Ascorbate oxidase of Cucumis melo L. var.
reticulatus: purification, characterization and antibody production. J. Exp. Bot.
45: 717–724.
Nakamura, T., N. Makino and Y. Ogura. 1968. Purification and properties of
ascorbate oxidase from cucumber. J. Biochem. 64: 189-95.
Namitra, K.K., S.N. Archana and P.S. Negi. 2011. Expression of carotenoid
biosynthetic pathway genes and changes in carotenoids during ripening in
tomato (Lycopersiconm esculentum). Food Funct., 2: 168-173.
Naohiro, K. and E. Muneharu. 1996. cDNA cloning and gene expression of ascorbate
oxidase in tobacco. Plant Mol. Biol., 30: 833-837.
Noctor, G. and C.H. Foyer 1998. Ascorbate and glutathione: keeping active oxygen
under control. Annu. Rev. Plant Physiol. Plant Mol. Biol., 49: 249-
279.
Ohkawa, J., N. Okada, A. Shinmyo and M. Takano. 1989. Primary structure of
cucumber (Cucumis sativus) ascorbate oxidase deduced from cDNA sequence:
Homology with blue copper proteins and tissue-specific expression. Proc. Natl.
Acad. Sci. USA, 86: 1239-1243.
Okkema, P.G., S.W. Harrison, V. Plunger, A. Aryana and A. Fire. 1993. Sequence
requirements for myosin gene expression and regulation in Caenorhabditis
elegans. Genetics, 135: 385-404.

185
185
Oomah, B. D. 2003. Sea buckthorn lipids. Li T. S. C., Beveridge T. (Eds). Sea
buckthorn (Hippophaė rhamnoides): Production and utilization. NRC
Research Press, Ottawa, ON. 51-68 pp.
Pan, R., Z. Zhang, Y. Ma, Z. Sun and B. Deng. 1989. The distribution characters of
sea-buckthorn (H. rhamnoides L.) and its research progress in China. Proc. Int.
Symp. Sea-buckthorn (H. rhamnoides L.) Xian, China. 1-16 pp.
Parimelazhagan, T., O.P. Chaurasia, and Z. Ahmed. 2005. Sea buckthorn: Oil with
promising medicinal value. Curr. Sci., 88: 8-9.
Pecker, I., R. Gabbay, F.X. Cunningham and J. Hirschberg. 1996. Cloning and
characterization of the cDNA for lycopene beta-cyclase from tomato reveals
decrease in its expression during fruit ripening. Plant Mol. Biol., 30: 807-819.
Pignocchi, C., J.M. Fletcher, J.E. Wilkinson, J. Barnes and C.H. Foyer. 2003. The
function of ascorbate oxidase in tobacco (Nicotiana tabacum L.). Plant
Physiol., 132: 1631-1641.
Pignocchi, C. and C.H. Foyer. 2003. Apoplastic ascorbate metabolism and its role in
the regulation of cell signalling. Curr. Opin. Plant Biol., 6: 379-389.
Pintea, A., A. Marpeau, M. Faye, C. Socaciu and M. Gleizes. 2001. Polar lipid and
fatty acid distribution in carotenolipoprotein complexes extracted from sea
buckthorn fruits. Phytochem. Anal., 12: 293-298.
Pintea, A., A. Varga, P. Stepnowski, C. Socaciu, M. Culea and H.A. Diehl. 2005.
Chromatographic analysis of carotenol fatty acid esters in Physalis alkekengi
and Hippophae rhamnoides. Phytochem. Anal., 16: 188-195.

186
186
Plekhanova, M.N. 1988. A report on Sea buckthorn in Russia. 77 pp.
Potters, G., N. Horemans, R.J. Caubergs and H. Asard. 2000. Ascorbate and
dehydroascorbate influence cell cycle progression in tobacco cell suspension.
Plant Physiol., 124: 17-20.
Raffo, A., F. Paoletti and M. Antonelli. 2004. Changes in sugar, organic acid,
flavonol and carotenoid composition during ripening of berries of three sea
buckthorn (Hippophae rhamnoides L.) cultivars. Eur. Food Res. Technol., 219:
360-368.
Rafael, da Silva M., V. Galli, S.D. Dos Anjos E Silva and C.V. Rombaldi. 2014.
Carotenoid biosynthetic and catabolic pathways: Gene expression and
carotenoid content in grains of maize landraces. Nutrients, 6: 546-563.
Rahimv, I.F., L.D. Lebedeva and N.D. Khasanshina. 1983. In: Biol. Chem. Pharm.
of Sea buckthorn Novosibirsk. 109-120 pp.
Raluca, P.M, Y. Weesepoel, C. Socaciu, A. Pintea, J.P. Vincken and H. Gruppen.
2014. Carotenoid composition of berries and leaves from six Romanian sea
buckthorn (Hippophae rhamnoides L.) varieties. Food Chem., 147: 1-9.
Ranjith, A., K.S. Kumar, V.V. Venugopalan, C. Arumughan, R.C. Sawhney and V.
Singh. 2006. Fatty acids, tocols, and carotenoids in pulp oil of three sea
buckthorn species (Hippophae rhamnoides, H. salicifolia, and H. tibetana)
grown in the Indian Himalayas. Am. Oil Chem. Soc., 83: 359-364.
Ravanel, S., H. Cherest, S. Jabrin, D. Grunwald, Y. Surdin-Keran, R. Douce and F.
Re’beille. 2001. Tetrahydrofolate biosynthesis in plants; molecular and
functional characterization of dihydrofolate synthetase and three isoforms

187
187
of folylpolyglutamate synthetase in Arabidopsis thaliana. Proc. Natl. Acad.
Sci. USA, 98: 1560-15365.
Ravanel, S., R. Douce and F. Re’beille 2004. The uniqueness of tetrahydrofolate
synthesis and one-carbon metabolism in plants. In Plant Mitochondria: From
Genome to Function, Day, D.A. Millar, H. and Whelan, J. eds. (Great Britain:
Kluwer). 277-292 pp.
R beill , F., S. Ravanel, S. Jabrin, R. Douce, S. Storozhenko and D. Van Der
Straeten. 2006. Folate in plants: Biosynthesis, distribution and enhancement.
Physiol. Plant, 126: 330-342
Re’beille, F., D. Macherel, J.M. Mouillon, J. Garin and R. Douce. 1997. Folate
biosynthesis in higher plants: purifcation and molecular cloning of a
bifunctional 6-hydroxymethyl-7,8-dihydropterin
pyrophosphokinase/7,8dihydropteroate synthase localized in mitochondria.
EMBO J., 16: 947-957.
Rébeillé, F. and R. Douce. 1999. Folate synthesis and compartmentation in higher
plants. In: Regulation of Primary Metabolic Pathways in Plants. Kruger NJ,
Hill SA, Ratcliffe RG, eds. Dordrecht, Kluwer. 53-99 pp.
Ren, J., H. Gao, J. Zhou, X. Hou and Y. Li. 2013. Molecular cloning and
characterization of ascorbate oxidase gene in non-heading Chinese cabbage.
Russ. J. Plant Physiol., 60: 756-763.
Rodríguez-Trelles F., R. Tarrío and F.J. Ayala. 2006. "Origins and evolution of
spliceosomal introns". Annu. Rev. Genet. 40: 47-76.

188
188
Ronen, G., M. Cohen, D. Zamir, J. Hirschberg. 1999. Regulation of carotenoid
biosynthesis during tomato fruit development: expression of the genes for
Lycopene-epsilon-cyclase is down-regulated during ripening and is elevated in
the mutant Delta. Plant j., 17: 341-51.
Rongsen, L. 1992. Sea buckthorn a multipurpose plant species for Fragile
Mountains. ICIMOD, Khathmando, Nepal. 20: 5-20.
Rongsen, L. 1996. Feasibility study on sea buckthorn development in Pakistan.
ICIMOD, Kathmandu, Nepal.
Ronen, G., L. Carmel-Goren, D. Zamir and J. Hirschberg. 2000. An alternative
pathway to B-formation in plant chromoplasts discovered by map-based
cloning of beta and old-gold color mutations in tomato. Proc. Natl. Acad. Sci.
USA, 97: 11102-11107.
Ronen, G., M. Cohen, D. Zamir and J. Hirschberg. 1999. Regulation of carotenoids
biosynthesis during tomato fruit development: expression gene for Lycopene
epsilon-cyclase is down-regulated during ripening and is elevated in the mutant
delta. The Plant J., 17: 341-351.
Rosch, D., C. Mugge, V. Fogliano and L.W. Kroh. 2004b. Antioxidant oligomeric
proanthocyanidins from sea buckthorn (Hippophae rhamnoides L.) Pomace. J.
Agric. Food Chem., 52: 6712-8.
Rösch, R., M. Bergmann, D. Knorr and L.W. Kroh. 2003. Structure–antioxidant
efficiency relationships of phenolic compounds and their contribution to the

189
189
antioxidant activity of sea buckthorn juice. J. Agric. Food Chem., 51:
42334239.
Rousi, A. and H. Aaulin. 1977. Ascorbic acid content in relation to ripeness in fruit
of six H. rhamnoides clones form pyharanta, SW. Finland. Ann. Agric. Fenn.,
16: 80-87.
Rousi, A. 1965. Observation on the cytology and variation of European and Asiatic
populations of Hippophae rhamnoides. Ann. Bot. Fenn. 2: 1-18.
Rousi, A. 1971. The genus Hippophae L. A taxonomic study. Annales Botanici.
Fennici., 8: 177-227.
Rupert, G.F., A. Wallace, P.D. Fraser, D. Valero, P. Hedden, P.M. Bramley and D.
Grierson. 2002. Constitutive expression of a fruit phytoene synthase gene in
transgenic tomatoes causes dwarfism by redirecting metabolites from the
gibberellin pathway. The Plant J., 8: 693-701.
Russo, C.A.M., N. Takezaki and M. Nei. 1996. Efficiencies of different genes and
tree-building methods in recovering a known vertebrate phylogeny.
Mol. Biol. Evol., 13: 525-536.
Sabir, S.M., H. Maqsood, S.D. Ahmed, A.H. Shah and M.Q. Khan. 2005. Chemical
and nutritional constituents of sea buckthorn (Hipophae rhamnoides ssp.
turkestanica) berries from Pakistan. Ital. J. Food Sci., 17: 455-462.
Sabir, S.M., H. Maqsood, I. Hayat, M.Q. Khan and A. Khaliq. 2005. Elemental and
nutritional analysis of sea buckthorn (Hippophaė rhamnoides spp.
turkestanica) berries of Pakistani origin. J. Med. Food, 8: 518-522.

190
190
Sabir, S.M., S.D. Ahmed and N. Lodhi. 2003. Morphological and biochemical
variation in Hippophaė rhammnoides ssp. turketanica, a multipurpose plant
for fragile mountains of Pakistan. S. Afr. J. Bot., 69: 587-592.
Sabot, I., E. Bergantino and G.M. Giacometti. 2005. Light and oxygenic
photosynthesis: energy dissipation as a protection mechanism against
photooxidation. EMBO Rep., 6: 629-634.
Saitou, N. and M. Nei. 1987. The neighbor-joining method: A new method for
reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
Samir, B., S. Storozhenko, P. Mehrshahi, M.J. Bennett, W. Lambert, J.F. Gregory,
K. Schubert, J. Hugenholtz, D. Van Der Straeten and A.D. Hanson. 2008.
Folate biofortification in food plants. Trends Plant Sci., 13: 28-35.
Sandeep, C., D. Olan, M.C. Brett, N. Janet, S.S. Jamie, G.M. Ian, F. Ross, S.P.
Thomas and D.S. James. 2012. Structure of S. aureus HPPK and the
Discovery of a New Substrate Site Inhibitor. PLoS ONE, 7: e29444.
Sanmartin, M., P.D. Drogoudi, T. Lyons, I. Pateraki, J. Barnes and A.K. Kanellis.
2003. Over-expression of ascorbate oxidase in the apoplast of transgenic
tobacco results in altered ascorbate and glutathione redox states and increased
sensitivity to ozone. Planta, 216: 918-928.
Sanmartin, M., I. Pateraki, F. Chatzopoulou and A.K. Kanellis. 2007. Diferential
expression of the ascorbate oxidase multigene family during fruit development
and in response to stress. Planta, 225: 873-885.
Sanja, R. 2007. Vitamin B biosynthesis in plant. Phytochemistry, 68: 1904-1921.

191
191
Schledz, M., S. al-Babili, J. von Lintig, H. Haubruck, S. Rabbani, H. Kleinig and P.
Beyer. 1996. Phytoene synthase from Narcissus pseudonarcissus: functional
expression, galactolipid requirement, topological distribution in chromoplasts
and induction during flowering. Plant J., 10: 781-792.
Schapiro, D.C. 1989. Biochemical studies on some hopeful forms and species of sea
buckthorn in USSR. In: Proceedings first International Symposium on Sea
Buckthorn, Xi’an, china. 64-66 pp.
Schroeder, W.R. and Y. Yao. 1995. Sea buckthorn: A promising multipurpose crop
for Saskatchewan; PFRA Shelterbelt Center, supplementary report. 95-102 pp.
Schroeder, W.R. 1995. Improvement of conservation trees and shrubs. PFRA
Shelterbelt Centre Supp. Rpt., 42: 95-1.
Schoedon, G., U. Redweik, G. Frank, R. Cotton and N. Blau. 1992. Allosteric
characteristics of GTP cyclohydrolase I from Escherichia coli. Eur. J.
Biochem., 210: 561-568.
Scolnik, P.A. and G.E. Bartley. 1994. Nucleotide sequence of an Arabidopsis cDNA
for phytoene synthase. Plant Physiol., 104: 1471-1472.
Scott, J.M., F. Rbeille and J. Fletcher. 2000. Folic acid and folates the feasibility of
nutritional enhancement in plant foods. J. Sci. Food Agric., 80: 795-824.
Shane, R.M. and R.J. Henry. 2008. Genes of folate biosynthesis in wheat. J. Cereal
Sci. 48: 632-638
Shane, R.M., D. Brushett and R.J. Henry. 2008. GTP cyclohydrolase 1 expression
and folate accumulation in the developing wheat seed. J. Cereal Sci. 48:
503512.

192
192
Shane, R.M. and R.J. Henry. 2008. Genes of folate biosynthesis in wheat. J. Cereal
Sci., 48: 632-638
Shazia, A., S.D. Ahmed, A.H. Shah, L. Hassan, S.I. Awan, A. Hamid and F. batool.
2010. Determination of optimum harvesting time for vitamin C, oil and mineral
elements in berries Sea buckthorn (Hippophae rhamnoides). Pak. J. Bot., 42:
3561-3568.
Shukla, S.K., P. Chaudhary, I.P. Kumar, N. Samanta, F. Afrin and M.L. Gupta. 2006.
Protection from radiation-induced mitochondrial and genomic DNA damage
by an extract of Hippophaë rhamnoides. Environ. Mol. Mutagen.,
47: 647-656.
Simrnoff, N. 1996. The function and metabolism of ascorbic acid in plants. Annales
Botanici., 78: 661-669.
Singh, V. 2005. Free radicals diseases anti-oxidants and anti-oxidant properties of
sea buckthorn (Hippophae rhamnoides L.) V. Singh (Ed.), Sea buckthorn
(Hippophae L). A Multipurpose Wonder Plant. Daya Publishing House, India.
3-69 pp.
Smirnoff, N. 1996. The function and metabolism of ascorbic acid in plants. Ann.
Bot., 78: 661-669.
Smirnoff, N. 2000. Ascorbic acid: metabolism and functions of a multi-facetted
molecule. Curr. Opin. Plant Biol., 3: 229-235.
Sperotto, R.A., F.K. Ricachenevsky, A. Waldow Vde and J.P. Fett. 2012. Iron
biofortification in rice: It’s a long way to the top. Plant Sci., 190: 24-39.

193
193
Staffan, C.A. 2009. Carotenoids, Tocochromanols and Chlorophylls in Sea
Buckthorn Berries (Hippophae rhamnoides) and Rose Hips (Rosa sp.)
(Doctoral Thesis report). ISSN: 1652-6880.
Stobdan, T, O.P. Chaurasia, G. Korekar, S. Mundra, Z. Ali, A. Yadav and S.B. Singh.
2010. Attributes of sea buckthorn (Hippophae rhamnoides L.) to meet
nutritional requirements in high altitude. Defence Sci. J., 60: 226-230.
Storozhenko, S., O. Navarrete, S. Ravanel, V. De Brouwer, P. Chaerle, G.F. Zhang,
O. Bastien, W. Lambert, F. Rébeillé and D. Van Der Straeten. 2007.
Cytosolic Hydroxymethyldihydropterin Pyrophosphokinase/Dihydropteroate Synthase from
Arabidopsis thaliana: a specific role in early development and stress response. J Biol. Chem.,
282: 10749-10761.
Stover, P.J. 2004. Physiology of folate and vitamin B12 in health and disease. Nutr.
Rev., 62: S3-12.
Stralsjo, L., H. Ahlin, C.M. Witthoft and J. Jastrebova. 2003. Folate determination
in Swedish berries by radioprotein-binding assay (RPBA) and high
performance liquid chromatography (HPLC). Eur. Food Res. Technol., 216:
264-269.
Suleyman, H., K. Gumustekin, S. Taysi, S. Keles, N. Oztasan and O. Aktas. 2002.
Beneficial effects of Hippophae rhamnoides Lon nicotine induced oxidative
stress in rat blood compared with vitamin E. Biol Pharm. Bull., 25: 11331136.
Suryakumar, G. and A. Gupta. 2011. Medicinal and therapeutic potential of Sea
buckthorn (Hippophae rhamnoides L.). J. Ethnopharmacol., 138: 268-278.

194
194
Tabata, K., K. Oba, K. Suzyki and M. Esaka. 2001. Generation and properties of
ascorbic acid-deficient transgenic tobacco cells expressing antisense RNA for
L-galactono-1,4-lactone dehydrogenase. The Plant J., 27: 139-148.
Tamura, K., M. Nei and S. Kumar. 2004. Prospects for inferring very large
phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci.
USA, 101: 11030-11035.
Tamura, K., D. Peterson, N. Peterson, G. Stecher, M. Nei and S. Kumar. 2011.
MEGA5: Molecular Evolutionary Genetics Analysis using Maximum
Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol.
Biol. Evol., 28: 2731-9.
Tamara, E. 2010. What’s in Sea Buckthorn: Flavonoids! http://www.mont-
echo.com.
Thompson, J.D.1., D.G. Higgins and T.J. Gibson. 1994. CLUSTAL W: improving
the sensitivity of progressive multiple sequence alignment through sequence
weighting, position-specific gap penalties and weight matrix choice. Nucleic
Acids Res., 22: 4673-80.
Tian, H. 1985. Sea buckthorn. Water and soil conserve. Commun., 2: 5-32.
Tong, Z.J.C., Z. Zhao, Y. Yang and K. Tian. 1989. The determination of the
physical-chemical constants and sixteen mineral elements in raw sea buckthorn
juice. Proc. of Int. Symp. Sea buckthorn (H. rhamnoides L.), Xian, China. 132-
137 pp.

195
195
Tracewell, C.A., A. Cua, D.H. Stewart, D.F. Bocian and G.W. Brudvig. 2001.
Characterization of carotenoid and chlorophyll photooxidation in photosystem
II. Biochem. J., 40: 193-203.
Tuskan, G.A., S. Difazio and S. Jansson et al. 2006. The genome of black
cottonwood, populous trichocarpa (Torr. & Gray). Science, 313: 1596-1604.
Vasileios, F., S. Maite and K.K. Angelos. 2006. Effect of ascorbate oxidase
overexpression on ascorbate recycling gene expression in response to agents
imposing oxidative stress. J. Exp. Bot., 57: 3933-3943.
Velioglu, Y.S., G. Mazza, L. Gao and B.D. Oomah. 1998. Antioxidant activity and
total phenolics in selected fruits, vegetables and grain products. J. Agric.
Food Chem., 46: 4113-4117.
Vijayaraghavan, R., A. Gautam, O. Kumar, S.C. Pant, M. Sharma and S. Singh.
2006. Protective effect of ethanolic and water extracts of sea buckthorn against
the toxic effects of mustard gas. Indian J. Exp. Biol., 44: 821-831.
Virendra, S. 2006. Sea buckthorn (Hippophae L.) a multipurpose wonder plant
Vol. II: Biochem. and Pharmacol.
Wahlberg, K. and N. Jeppsson. 1990. Development of cultivars and growing
techniques for sea-buckthorn, black chokeberry, Lonicera and Sorbus.
Sverigges Lantbruksuniversitedt Balsgård-Avdelningenför Hortikulturell
Växförädling Verksamhetsberättelse. 80-93 pp. (in Swedish, summary in
English).
Waller, J.C., T.A. Akhtar, A. Lara-Núñez, J.F. Gregory, R.P. McQuinn, J.J.

196
196
Giovannoni, A.D. Luciana, C.C., T.M. de Oliveira, A.F. Mendes, A.F.
Macedo, E.I. Floh , A.S. Gesteira, W.S. Soares-Filho and M.G. Costa. 2012.
Ectopic expression of a fruit phytoene synthase from Citrus paradisi Macf.
promotes abiotic stress tolerance in transgenic tobacco. Plant Mol. Biol. Rep.,
39: 10201-10209.
Wang, G., 1987. Comparative study of sea buckthorn oil in the Northern Zone of
China. Journal of Northwest Forest college, 2: 55-60.
Wang, Z.R., L. Wang, H.H. Yin, F.J. Yang, Y.O. Gao and Z.J. Zhang. 2000. Effect
of total flavonoids of Hippophae rhamnoides on contractile mechanics and
calcium transfer in stretched myocyte. Space Med. Med. Eng., 13: 6-9.
Wheeler, G.L., M.A. Jones, N. Smirnoff. 1998. The biosynthetic pathway of vitamin
C in higher plants. Nature, 393: 365-369.
Wolf, D. and F. Wegert. 1993. Experience gained in the cultivation, harvesting and
utilization of sea-buckthorn. In: Cultivation and Utilization of Wild Fruit
Crops. Bernhard Thalacker Verlag GmbH & Co. 23-29 pp. (in German).
Xu, Z. 1956. Studies on sea buckthorn juice. Acta Nutritioni., 1: 334-349.
Yao, Y. and C. Zhu. 1985. Investigation on natural resources of Hippophae
rhamnoides L. in West-Shanxi Plateau. Shansxi Sci. and Tech. of Forestry. 4:
105-119. (in Chinese).
Yao, Y. and P.M.A. Tigerstedt. 1993. Lsozyme studies of genetic diversity and
evolution in Hippophae. Genet Resour. Crop Ev., 40: 153-164.
Yao, Y. and P.M.A. Tigerstedt. 1994. Genetic diversity in Hippophae L. and its use
in plant breeding. Euphytica., 77: 165-169.

197
197
Yao, Y. and P.M.A. Tigerstedt. 1995. Geographic variation of growth rhythm, height
and hardiness and their relations in Hippophaė rhamnoides. J. Am.
Soc. Hortic. Sci., 120: 691-698.
Yang, B., K. Kalimo, L. Mattila, S. Kallio, J. Katajisto, O. Peltola and H. Kallio.
1999. Effect of dietary supplementation with sea buckthorn (Hippophaë rhamnoides) seed and
pulp oils on atopic dermatitis. J. Nutr. Biochem., 10: 622-630.
Yang, B. and H. Kallio. 2002a. Composition and physiological effects of sea
buckthorn (Hippophaė) lipids. Trends Food Sci. Technol., 13: 160-167.
Yang, B. and H. Kallio. 2002b. Effects of harvesting time on triacylglycerols and
glycerophospholipids of sea buckthorn (Hippophaė rhamnoides L.) berries of
different origins. J. Food Compost. Anal., 15: 143-157.
Yang, B and H. Kallio. 2005. Lipophilic components of sea buckthorn (Hippophae
rhamnoides L.) seeds and berries V. Singh (Ed.), Sea buckthorn (Hippophae
L), A Multipurpose Wonder Plant. Daya Publishing House, India. 70-97 pp.
Yang, B. and H. Kallio. 2002. Lipophilic components in seeds and berries of sea
buckthorn and physiological effects of sea buckthorn oils. Trends Food Sci.
Technol., 13: 160-167.
Yang, B. and H. Kallio. 2002. Composition and physiological effects of sea
buckthorn (Hippophae) lipids. Trends Food Sci. Technol., 13: 160-167
Yang, B., R.M. Karlsson, P.H. Oksman and H.P. Kallio. 2001. Phytosterols in sea
buckthorn (Hippophaė rhamnoides L.) berries: Identification and effects of
different origins and harvesting times. J. Agric. Food Chem., 49: 5620-5629.

198
198
Yang, B.R. 2001. Lipophilic components of sea buckthorn (Hippophaė rhamnoides)
seeds and berries and physiological effects of sea buckthorn oils. PhD
dissertation, Turku University, Finland.
Yang, B., M. Ahotupa, P. Maatta and H. Kallio. 2011. Composition and antioxidative
activities of supercritical CO2-extracted oils from seeds and soft parts of
northern berries. Food Res. Int., 44: 2009-2017.
Yang, H., S. Wang and X. Su. 1988. A study on the dynamic variation of ascorbic
acid in Hippophae. Hippophae, 4: 41-44 (in Chinese).
Yoneyama, T., L. Wilson and K. Hatakeyama. 2001. GTP cyclohydrolase I feedback
regulatory protein-dependent and -independent inhibitors of GTP
cyclohydrolase I. Arch. Biochem. Biophys., 388: 67-73.
Yuanxiu, L.X.G., H. Tang, Y. Hou, D. Yu and X. Zhang. 2013. Cloning and
Expression Analysis of an Ascorbate Oxidase (AO) Gene from Strawberry
(Fragaria×ananassa cv. Toyonaka). Journal of Agricultural Sci., 5: 14-22.
Yuzhen, Z. and W. Fuheng. 1997. Sea buckthorn flavonoids and their medicinal
value. Hippophaė., 10: 39-41.
Yu, Z., F. Ao and Y. Lian. 1989. Discussion on the problems of origin, classification,
community and resource of sea-buckthorn in China. Proc. Int. Symp. Sea-
buckthorn (H. rhamnoides L.), Xian, China. 21-30 pp.
Yuanxiu, L., X. Gu, H.Tang, Y. Hou, D. Yu and X. Zhang. 2013. Cloning and
Expression Analysis of an Ascorbate Oxidase (AO) Gene from Strawberry
(Fragaria×ananassa cv. Toyonaka). J. Agr.Sci., 5: 14-22.

199
199
Yu, L., Z. Hong, C. Wei, H. Shao-zhen and L. Qing-chang. 2013. Cloning and
Functional Analysis of Lycopene ε-Cyclase (IbLCYe) Gene from
Sweetpotato, Ipomoea batatas (L.) Lam. Journal of Integ. Agr., 12: 773-780.
Zadernowski, R., H. Nowak-Polakowska, B. Lossow and J.
Nesterowicz., 1997. Sea-buckthorn lipids. J. Food Lipids, 4: 165-172.
Zadernowski, R., M. Naczk, S. Czaplicki, M. Rubinskiene and M. Szalkiewicz.
2005. Composition of phenolic acids in sea buckthorn (Hippophae rhamnoides
L.) berries. J. Am. Oil Chem. Soc., 82: 175-179.
Zar, B.A., L. Zacaryas and M.J. Rodrigo. 2009. Molecular and functional
characterization of a novel chromoplast-specific lycopene β-cyclase from
Citrus and its relation to Lycopene accumulation. J. Exp. Bot., 60: 17831797.
Zeb, A., and I. Malook. 2009. Biochemical characterization of sea buckthorn
(Hippophaė rhamnoides L. spp. turkestanica) seed. Afr. J. Biotechnol., 8:
1625-1629.
Zeb, A. 2004a. Chemical and nutritional constituents of sea buckthorn juice. Pak. J.
Nut., 3: 99-106.
Zeb, A. 2004b. Important therapeutic uses of sea buckthorn (Hippophaė): A review.
J. Biol. Sci., 4: 687-693.
Zhang, W., J. Yan, J. Duo, B. Ren and J. Guo. 1989. Preliminary Study of
Biochemical Constituents of Berry of Sea buckthorn growing in Shanxi
Province and their Changing Trend. In: Proc. of Int. Symp. Sea buckthorn
(Hippophae rhamnoides L.), Xian, China. 96-105 pp.

200
200
Zhang, P. 1989. Anti-cancer activities of sea buckthorn seed oil and its effects on the
weight of immune organs. Sea buckthorn. 2: 31-34.
Zhang, P.X., L.M. Ding and Li. 1989. Anti tumor effects of fruit juice and seed oil
of (Hyppophae rhamnoides L.)and their influences on immune function.
Proc. Int. Symp. Sea buckthorn (H. rhamnoides L.), Xian, China. 373-381 pp.
Zhang, W., J. Yan, J. Duo, B. Ren and J. Guo. 1989. Preliminary study of
biochemical constitutions of berry of sea buckthorn growing in Shanxi
province and their changing trend. Proc. of int. symp. on sea buckthorn (H.
rhamnoides L.), Xian, China. 96-105 pp.
Zhang, Y., J. Xu and N. Yu. 1989. Research on the centrifuge technology of sea
buckthorn juice and oil. Proc. of Int. Symp. on sea buckthorn (H. rhamnoides
L.), Xian, China. 307-310 pp.
Zheng, X.W. and X.J. Song. 1992. Analysis of the fruit nutrient composition of nine
types of sea buckthorn in Liaoning, China North. Fruits of China., 3: 22-
24.
Zhou, X. and C. Chen. 1989. Research on the process technology of turbid type sea
buckthorn beverage. Proceedings of international symposium on sea buckthorn
(H. rhamnoides L.), Xian, China. 310-313 pp.
Zimmermann, M.B. and R.F. Hurrel. 2002. Improving iron, Zinc and vitamin A
nutrition through plant biotechnology. Curr. Opin. Biotechnol., 13: 142-145.

201
201
Zou, L., H. Li, B. Ouyang, J. Zhang and Z. Ye. 2006. Cloning and mapping of genes
involved in tomato ascorbic acid biosynthesis and metabolism. Plant Sci., 170:
120-127.


















