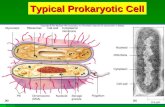
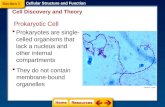
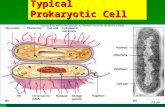
Prokaryotic eukaryoticgenome
94
Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings The Genetics of Viruses and Bacteria • Viruses called bacteriophages – Can infect and set in motion a genetic takeover of bacteria, such as Escherichia coli
-
Upload
andrew-mccaskill -
Category
Documents
-
view
134 -
download
2
description
Transcript of Prokaryotic eukaryoticgenome

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
The Genetics of Viruses and Bacteria
• Viruses called bacteriophages
– Can infect and set in motion a genetic takeover of bacteria, such as Escherichia coli

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Recall that bacteria are prokaryotes
– With cells much smaller and more simply organized than those of eukaryotes
• Viruses
– Are smaller and simpler still
Figure 18.20.25 m
Virus
Animalcell
Bacterium
Animal cell nucleus

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• A virus has a genome but can reproduce only within a host cell
• Scientists were able to detect viruses indirectly
– Long before they were actually able to see them
Figure 18.3
For example: Tobacco mosaic disease stunts the growth of tobacco plants and gives their leaves a mosaic coloration

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• In the late 1800s
– Researchers hypothesized that a particle smaller than bacteria caused tobacco mosaic disease
• In 1935, Wendell Stanley
– Confirmed this hypothesis when he crystallized the infectious particle, now known as tobacco mosaic virus (TMV)

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Structure of Viruses
• Viruses
– Are very small infectious particles consisting of nucleic acid enclosed in a protein coat and, in some cases, a membranous envelope
• Viral genomes may consist of
– Double- or single-stranded DNA
– Double- or single-stranded RNA

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Figure 18.4a, b
18 250 mm 70–90 nm (diameter)
20 nm 50 nm(a) Tobacco mosaic virus (b) Adenoviruses
RNADNACapsomere
Glycoprotein
Capsomereof capsid
Capsids and Envelopes
• A capsid
– Is the protein shell that encloses the viral genome
– Can have various structures

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Some viruses have envelopes
– Which are membranous coverings derived from the membrane of the host cell
Figure 18.4c
80–200 nm (diameter)
50 nm(c) Influenza viruses
RNA
Glycoprotein
Membranousenvelope
Capsid

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Bacteriophages, also called phages
– Have the most complex capsids found among viruses
Figure 18.4d
80 225 nm
50 nm(d) Bacteriophage T4
DNA
Head
Tail fiber
Tail sheath

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
General Features of Viral Reproductive Cycles
• Viruses are obligate intracellular parasites
– They can reproduce only within a host cell
• Each virus has a host range
– A limited number of host cells that it can infect

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Viruses use enzymes, ribosomes, and small molecules of host cells
– To synthesize progeny virusesVIRUS
Capsid proteins
mRNA
Viral DNA
HOST CELL
Viral DNA
DNACapsid
Figure 18.5
Entry into cell anduncoating of DNA
ReplicationTranscription
Self-assembly of new virus particles and their exit from cell

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Reproductive Cycles of Phages
• Phages
– Are the best understood of all viruses
– Go through two alternative reproductive mechanisms: the lytic cycle and the lysogenic cycle

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
The Lytic Cycle
• The lytic cycle
– Is a phage reproductive cycle that culminates in the death of the host
– Produces new phages and digests the host’s cell wall, releasing the progeny viruses

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• The lytic cycle of phage T4, a virulent phage
Phage assembly
Head Tails Tail fibersFigure 18.6
Attachment. The T4 phage usesits tail fibers to bind to specificreceptor sites on the outer surface of an E. coli cell.
1Entry of phage DNA and degradation of host DNA.The sheath of the tail contracts,injecting the phage DNA intothe cell and leaving an emptycapsid outside. The cell’sDNA is hydrolyzed.
2
Synthesis of viral genomes and proteins. The phage DNAdirects production of phageproteins and copies of the phagegenome by host enzymes, usingcomponents within the cell.
3Assembly. Three separate sets of proteinsself-assemble to form phage heads, tails,and tail fibers. The phage genome ispackaged inside the capsid as the head forms.
4
Release. The phage directs productionof an enzyme that damages the bacterialcell wall, allowing fluid to enter. The cellswells and finally bursts, releasing 100 to 200 phage particles.
5

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
The Lysogenic Cycle
• The lysogenic cycle
– Replicates the phage genome without destroying the host
• Temperate phages
– Are capable of using both the lytic and lysogenic cycles of reproduction

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• The lytic and lysogenic cycles of phage , a temperate phage
Many cell divisions produce a large population of bacteria infected with the prophage.
The bacterium reproducesnormally, copying the prophageand transmitting it to daughter cells.
Phage DNA integrates into the bacterial chromosome,becoming a prophage.
New phage DNA and proteins are synthesized and assembled into phages.
Occasionally, a prophage exits the bacterial chromosome, initiating a lytic cycle.
Certain factorsdetermine whether
The phage attaches to ahost cell and injects its DNA.
Phage DNAcircularizes
The cell lyses, releasing phages.Lytic cycleis induced
Lysogenic cycleis entered
Lysogenic cycleLytic cycle
or Prophage
Bacterialchromosome
Phage
PhageDNA
Figure 18.7

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Viral Envelopes
• Many animal viruses
– Have a membranous envelope
• The broadest variety of RNA genomes
– Is found among the viruses that infect animals
• Viral glycoproteins on the envelope
– Bind to specific receptor molecules on the surface of a host cell Table 18.1

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
RNA
Capsid
Envelope (withglycoproteins)
HOST CELL
Viral genome (RNA)
Template
Capsidproteins
Glyco-proteins
mRNA
Copy ofgenome (RNA)
ER
Figure 18.8
• The reproductive cycle of an enveloped RNA virus Glycoproteins on the viral envelope bind to specific receptor molecules(not shown) on the host cell, promoting viral entry into the cell.
1
Capsid and viral genomeenter cell2
The viral genome (red)functions as a template forsynthesis of complementary RNA strands (pink) by a viral enzyme.
3
New copies of viralgenome RNA are madeusing complementary RNAstrands as templates.
4
Complementary RNAstrands also function as mRNA,
which is translated into bothcapsid proteins (in the cytosol)and glycoproteins for the viral
envelope (in the ER).
5
Vesicles transportenvelope glycoproteins to
the plasma membrane.
6
A capsid assemblesaround each viral
genome molecule.
7
New virus8

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Retroviruses, such as HIV, use the enzyme reverse transcriptase
– To copy their RNA genome into DNA, which can then be integrated into the host genome as a provirus
Figure 18.9
Reversetranscriptase
Viral envelope
Capsid
Glycoprotein
RNA(two identicalstrands)

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• The reproductive cycle of HIV, a retrovirus
Figure 18.10
mRNA
RNA genomefor the nextviral generation
Viral RNA
RNA-DNAhybrid
DNA
ChromosomalDNA
NUCLEUSProvirus
HOST CELL
Reverse transcriptase
New HIV leaving a cell
HIV entering a cell
0.25 µm
HIV Membrane of white blood cell
The virus fuses with thecell’s plasma membrane.The capsid proteins areremoved, releasing the viral proteins and RNA.
1
Reverse transcriptasecatalyzes the synthesis of aDNA strand complementaryto the viral RNA.
2
Reverse transcriptasecatalyzes the synthesis ofa second DNA strandcomplementary to the first.
3
The double-stranded DNA is incorporatedas a provirus into the cell’s DNA.
4
Proviral genes are transcribed into RNA molecules, which serve as genomes for the next viral generation and as mRNAs for translation into viral proteins.
5
The viral proteins include capsid proteins and reverse transcriptase (made in the cytosol) and envelope glycoproteins (made in the ER).
6
Vesicles transport theglycoproteins from the ER tothe cell’s plasma membrane.
7 Capsids areassembled aroundviral genomes and reverse transcriptase molecules.
8
New viruses budoff from the host cell.9

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Evolution of Viruses
• Viruses do not really fit our definition of living organisms (no cellular structure)
• Viruses, viroids, and prions are formidable pathogens in animals and plants
• Diseases caused by viral infections
– Affect humans, agricultural crops, and livestock worldwide

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Viral Diseases in Animals
• Viruses may damage or kill cells
– By causing the release of hydrolytic enzymes from lysosomes
• Some viruses cause infected cells
– To produce toxins that lead to disease symptoms

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Vaccines
– Are harmless derivatives of pathogenic microbes that stimulate the immune system to mount defenses against the actual pathogen
– Can prevent certain viral illnesses
• Emerging viruses
– Are those that appear suddenly or suddenly come to the attention of medical scientists

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Severe acute respiratory syndrome (SARS)
– Recently appeared in China
(a) Young ballet students in Hong Kong wear face masks to protect themselves from the virus causing SARS.
(b) The SARS-causing agent is a coronavirus like this one (colorized TEM), so named for the “corona” of glycoprotein spikes protruding from the envelope.
Outbreaks of “new” viral diseases in humansAre usually caused by existing viruses that expand their host territory

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Viral Diseases in Plants
• More than 2,000 types of viral diseases of plants are known
• Common symptoms of viral infection include
– Spots on leaves and fruits, stunted growth, and damaged flowers or roots
Figure 18.12

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Plant viruses spread disease in two major modes
– Horizontal transmission, entering through damaged cell walls
– Vertical transmission, inheriting the virus from a parent

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Viroids and Prions: The Simplest Infectious Agents• Viroids
– Are circular RNA molecules that infect plants and disrupt their growth
• Prions
– Are slow-acting, virtually indestructible infectious proteins that cause brain diseases in mammals
– Propagate by converting normal proteins into the prion version
Figure 18.13
Prion
Normalprotein
Originalprion
Newprion
Many prions

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Rapid reproduction, mutation, and genetic recombination contribute to the genetic diversity of bacteria
• Bacteria allow researchers
– To investigate molecular genetics in the simplest true organisms

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
The Bacterial Genome and Its Replication
• The bacterial chromosome
– Is usually a circular DNA molecule with few associated proteins
• In addition to the chromosome
– Many bacteria have plasmids, smaller circular DNA molecules that can replicate independently of the bacterial chromosome

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Bacterial cells divide by binary fission
– Which is preceded by replication of the bacterial chromosome
Replicationfork
Origin of replication
Termination of replication
Figure 18.14

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Mutation and Genetic Recombination as Sources of Genetic Variation
• Since bacteria can reproduce rapidly
– New mutations can quickly increase a population’s genetic diversity

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Mechanisms of Gene Transfer and Genetic Recombination in Bacteria
• Three processes bring bacterial DNA from different individuals together
– Transformation
– Transduction
– Conjugation

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Transformation
• Transformation
– Is the alteration of a bacterial cell’s genotype and phenotype by the uptake of naked, foreign DNA from the surrounding environment

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Transduction
• In the process known as transduction
– Phages carry bacterial genes from one host cell to another
1
Figure 18.16
Donorcell
Recipientcell
A+ B+
A+
A+ B–
A– B–
A+
Recombinant cell
Crossingover
Phage infects bacterial cell that has alleles A+ and B+
Host DNA (brown) is fragmented, and phage DNA and proteins are made. This is the donor cell.
A bacterial DNA fragment (in this case a fragment withthe A+ allele) may be packaged in a phage capsid.
Phage with the A+ allele from the donor cell infects a recipient A–B– cell, and crossing over (recombination)between donor DNA (brown) and recipient DNA(green) occurs at two places (dotted lines).
The genotype of the resulting recombinant cell (A+B–) differs from the genotypes of both the donor (A+B+) and the recipient (A–B–).
2
3
4
5
Phage DNA
A+ B+

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Conjugation and Plasmids
• Conjugation
– Is the direct transfer of genetic material between bacterial cells that are temporarily joined
Figure 18.17 Sex pilus 1 m

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
The F Plasmid and Conjugation
• Cells containing the F plasmid, designated F+ cells
– Function as DNA donors during conjugation
– Transfer plasmid DNA to an F recipient cell
Figure 18.18a
A cell carrying an F plasmid(an F+ cell) can form amating bridge with an F– celland transfer its F plasmid.
A single strand of the F plasmid breaks at a specific point (tip of blue arrowhead) and begins tomove into the recipient cell. As transfer continues, the donor plasmid rotates(red arrow).
2 DNA replication occurs inboth donor and recipientcells, using the single parental strands of the F plasmid as templates to synthesize complementary strands.
3 The plasmid in the recipient cell circularizes. Transfer and replication result in a compete F plasmid in each cell. Thus, both cells are now F+.
4
F Plasmid Bacterial chromosome
Bacterial chromosome
F+ cell
F+ cell
F+ cell
Mating bridge
1
Conjugation and transfer of an F plasmid from an F+ donor to an F– recipient
(a)
F– cell

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Chromosomal genes can be transferred during conjugation
– When the donor cell’s F factor is integrated into the chromosome
• A cell with the F factor built into its chromosome
– Is called an Hfr cell
• The F factor of an Hfr cell
– Brings some chromosomal DNA along with it when it is transferred to an F– cell

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Conjugation and transfer of part of the bacterial chromosome from an Hfr donor to an F– recipient, resulting in recombination
F+ cell Hfr cell
F factorThe circular F plasmid in an F+ cellcan be integrated into the circularchromosome by a single crossoverevent (dotted line).
1The resulting cell is called an Hfr cell (for High frequency of recombination).
2
Since an Hfr cell has all the F-factor genes, it can form a mating bridge with an F– cell and transfer DNA.
3 A single strand of the F factorbreaks and begins to move through the bridge. DNA replication occurs in both donor and recipient cells, resulting in double-stranded DNA
4 The location and orientation of the F factor in the donor chromosome determine the sequence of gene transfer during conjugation. In this example, the transfer sequence for four genes is A-B-C-D.
5 The mating bridgeusually breaks well before the entire chromosome andthe rest of the F factor are transferred.
6
Two crossovers can result in the exchange of similar (homologous) genes between the transferred chromosome fragment (brown) and the recipient cell’s chromosome (green).
7 The piece of DNA ending up outside thebacterial chromosome will eventually be degraded by the cell’s enzymes. The recipient cell now contains a new combination of genes but no F factor; it is a recombinant F– cell.
8
Temporarypartialdiploid
Recombinant F–
bacterium
A+B+ C+
D+
F– cell A–B–
C–
D–
A–B–
C–
D– D–
A–
C–B–
A+
B+C+D+A+B+
D+C+
A+
A+
B+
A–B–
C–
D–
A–B+
C–
D–
A+
B+ B–
A+
Hfr cell
D–
A–
C–
B–
A+
B+C+D+
A+
B+
Conjugation and transfer of part of the bacterial chromosome from an Hfr donor to an F– recipient, resulting in recombination
(b)
Figure 18.18b

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
R plasmids and Antibiotic Resistance
• R plasmids
– Confer resistance to various antibiotics
• Transposable elements
– Can move around within a cell’s genome
– Are often called “jumping genes”
– Contribute to genetic shuffling in bacteria

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Figure 18.19a
(a) Insertion sequences, the simplest transposable elements in bacteria, contain a single gene that encodes transposase, which catalyzes movement within the genome. The inverted repeats are backward, upside-down versions of each other; only a portion is shown. The inverted repeat sequence varies from one type of insertion sequence to another.
Insertion sequence
Transposase geneInvertedrepeat
Invertedrepeat
3
5
3
5
A T C C G G T…
T A G G C C A …
A C C G G A T…
T G G C C T A …
Insertion Sequences
• An insertion sequence contains a single gene for transposase
– An enzyme that catalyzes movement of the insertion sequence from one site to another within the genome

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Transposons
• Bacterial transposons
– Also move about within the bacterial genome
– Have additional genes, such as those for antibiotic resistance
Figure 18.19b
(b) Transposons contain one or more genes in addition to the transposase gene. In the transposon shown here, a gene for resistance to an antibiotic is located between twin insertion sequences. The gene for antibiotic resistance is carried along as part of the transposon when the transposon is inserted at a new site in the genome.
Inverted repeats Transposase gene
Insertion sequence
Insertion sequence
Antibioticresistance gene
Transposon
5
3
5
3

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Individual bacteria respond to environmental change by regulating their gene expression
• E. coli, a type of bacteria that lives in the human colon
– Can tune its metabolism to the changing environment and food sources

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• This metabolic control occurs on two levels
– Adjusting the activity of metabolic enzymes already present
– Regulating the genes encoding the metabolic enzymes
Figure 18.20a, b
(a) Regulation of enzyme activity
Enzyme 1
Enzyme 2
Enzyme 3
Enzyme 4
Enzyme 5
Regulationof geneexpression
Feedbackinhibition
Tryptophan
Precursor
(b) Regulation of enzyme production
Gene 2
Gene 1
Gene 3
Gene 4
Gene 5
–
–

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Operons: The Basic Concept
• In bacteria, genes are often clustered into operons, composed of
– An operator, an “on-off” switch
– A promoter
– Genes for metabolic enzymes

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• An operon
– Is usually turned “on”
– Can be switched off by a protein called a repressor

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Video Clip – trp operon
• The trp operon: regulated synthesis of repressible enzymes
Figure 18.21a
(a) Tryptophan absent, repressor inactive, operon on. RNA polymerase attaches to the DNA at the promoter and transcribes the operon’s genes.
Genes of operon
Inactiverepressor
Protein
Operator
Polypeptides that make upenzymes for tryptophan synthesis
Promoter
Regulatorygene
RNA polymerase
Start codon Stop codon
Promoter
trp operon
5
3mRNA 5
trpDtrpE trpC trpB trpAtrpRDNA
mRNA
E D C B A

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
DNA
mRNA
Protein
Tryptophan(corepressor)
Active repressor
No RNA made
Tryptophan present, repressor active, operon off. As tryptophanaccumulates, it inhibits its own production by activating the repressor protein.
(b)
Figure 18.21b

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Repressible and Inducible Operons: Two Types of Negative Gene Regulation
• In a repressible operon
– Binding of a specific repressor protein to the operator shuts off transcription
• In an inducible operon
– Binding of an inducer to an innately inactive repressor inactivates the repressor and turns on transcription

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Video Clip – Lac Operon
• The lac operon: regulated synthesis of inducible enzymes
Figure 18.22a
DNA
mRNA
ProteinActiverepressor
RNApolymerase
NoRNAmade
lacZlacl
Regulatorygene
Operator
Promoter
Lactose absent, repressor active, operon off. The lac repressor is innately active, and inthe absence of lactose it switches off the operon by binding to the operator.
(a)
5
3

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
mRNA 5'
DNA
mRNA
Protein
Allolactose(inducer)
Inactiverepressor
lacl lacz lacY lacA
RNApolymerase
Permease Transacetylase-Galactosidase
5
3
(b) Lactose present, repressor inactive, operon on. Allolactose, an isomer of lactose, derepresses the operon by inactivating the repressor. In this way, the enzymes for lactose utilization are induced.
mRNA 5
lac operon
Figure 18.22b

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Inducible enzymes
– Usually function in catabolic pathways (break down)
• Repressible enzymes
– Usually function in anabolic pathways (build up)

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Regulation of both the trp and lac operons
– Involves the negative control of genes, because the operons are switched off by the active form of the repressor protein
Figure 18.22a
DNA
mRNA
ProteinActiverepressor
RNApolymerase
NoRNAmade
lacZlacl
Regulatorygene Operator
Promoter
Lactose absent, repressor active, operon off. (a)
5
3
DNA
mRNA
Protein
Tryptophan(corepressor)
Active repressor
No RNA made
Tryptophan present, repressor active, operon off
(b)
Figure 18.21b

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Positive Gene Regulation
• Some operons are also subject to positive control
– Via a stimulatory activator protein, such as catabolite activator protein (CAP)

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Promoter
Lactose present, glucose scarce (cAMP level high): abundant lac mRNA synthesized.If glucose is scarce, the high level of cAMP activates CAP, and the lac operon produces large amounts of mRNA for the lactose pathway.
(a)
CAP-binding site OperatorRNApolymerasecan bindand transcribe
InactiveCAP
ActiveCAPcAMP
DNA
Inactive lacrepressor
lacl lacZ
Figure 18.23a
• In E. coli, when glucose, a preferred food source, is scarce
– The lac operon is activated by the binding of a regulatory protein, catabolite activator protein (CAP)

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Video Clip
• When glucose levels in an E. coli cell increase
– CAP detaches from the lac operon, turning it off
Figure 18.23b(b)Lactose present, glucose present (cAMP level low): little lac mRNA synthesized.
When glucose is present, cAMP is scarce, and CAP is unable to stimulate transcription.
Inactive lacrepressor
InactiveCAP
DNA
RNApolymerasecan’t bind
Operator
lacl lacZ
CAP-binding site
Promoter

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Eukaryotic Genomes
• In eukaryotes, the DNA-protein complex, called chromatin
– Is ordered into higher structural levels than the DNA-protein complex in prokaryotes
Figure 19.1

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Chromatin structure is based on successive levels of DNA packing
• Eukaryotic DNA
– Is precisely combined with a large amount of protein
• Eukaryotic chromosomes
– Contain an enormous amount of DNA relative to their condensed length

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Nucleosomes, or “Beads on a String”
• Proteins called histones
– Are responsible for the first level of DNA packing in chromatin
– Bind tightly to DNA
• The association of DNA and histones
– Seems to remain intact throughout the cell cycle

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• In electron micrographs
– Unfolded chromatin has the appearance of beads on a string
• Each “bead” is a nucleosome
– The basic unit of DNA packing
Figure 19.2 a
2 nm
10 nm
DNA double helix
Histonetails
His-tones
Linker DNA(“string”)
Nucleosome(“bad”)
Histone H1
(a) Nucleosomes (10-nm fiber)

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Nucleosome
30 nm
(b) 30-nm fiber
Higher Levels of DNA Packing
• The next level of packing
– Forms the 30-nm chromatin fiber
Figure 19.2 b

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• The 30-nm fiber, in turn
– Forms looped domains, making up a 300-nm fiber
Figure 19.2 c
Protein scaffold
300 nm
(c) Looped domains (300-nm fiber)
Loops
Scaffold

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• In a mitotic chromosome
– The looped domains themselves coil and fold forming the characteristic metaphase chromosome
Figure 19.2 d
700 nm
1,400 nm
(d) Metaphase chromosome

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• In interphase cells
– Most chromatin is in the highly extended form called euchromatin

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Gene expression can be regulated at any stage, but the key step is transcription
• All organisms
– Must regulate which genes are expressed at any given time
• During development of a multicellular organism
– Its cells undergo a process of specialization in form and function called cell differentiation

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Differential Gene Expression
• Each cell of a multicellular eukaryote
– Expresses only a fraction of its genes
• In each type of differentiated cell
– A unique subset of genes is expressed
Figure 19.3
Signal
NUCLEUSChromatin
Chromatin modification:DNA unpacking involvinghistone acetylation and
DNA demethlation
Gene
DNAGene availablefor transcription
RNA Exon
Transcription
Primary transcript
RNA processing
Transport to cytoplasm
Intron
Cap mRNA in nucleus
Tail
CYTOPLASM
mRNA in cytoplasmDegradation
of mRNA
Translation
Polypetide
CleavageChemical modificationTransport to cellular
destination
Active protein
Degradation of protein
Degraded protein

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Histone Modification
• Chemical modification of histone tails
– Can affect the configuration of chromatin and thus gene expression
Figure 19.4a (a) Histone tails protrude outward from a nucleosome
Chromatin changes
Transcription
RNA processing
mRNA degradation
Translation
Protein processingand degradation
DNAdouble helix
Amino acidsavailablefor chemicalmodification
Histonetails

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Histone acetylation
– Seems to loosen chromatin structure and thereby enhance transcription
Figure 19.4 b(b) Acetylation of histone tails promotes loose chromatin structure that permits transcription
Unacetylated histones Acetylated histones

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Organization of a Typical Eukaryotic Gene
• Associated with most eukaryotic genes are multiple control elements
– Segments of noncoding DNA that help regulate transcription by binding certain proteins
Figure 19.5
Enhancer(distal control elements)
Proximalcontrol elements
DNA
UpstreamPromoter
Exon Intron Exon Intron
Poly-A signalsequence
Exon
Terminationregion
Transcription
Downstream
Poly-Asignal
ExonIntronExonIntronExonPrimary RNAtranscript(pre-mRNA)
5
Intron RNA
RNA processing:Cap and tail added;introns excised andexons spliced together
Coding segment
P P PGmRNA
5 Cap 5 UTR(untranslated
region)
Startcodon
Stopcodon
3 UTR(untranslated
region)
Poly-Atail
Chromatin changes
Transcription
RNA processing
mRNAdegradation
Translation
Protein processingand degradation
Cleared 3 endof primarytransport

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Enhancers and Specific Transcription Factors
• Proximal control elements
– Are located close to the promoter
• Distal control elements, groups of which are called enhancers
– May be far away from a gene or even in an intron

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Distal controlelement
Activators
Enhancer
PromoterGene
TATAbox General
transcriptionfactors
DNA-bendingprotein
Group ofMediator proteins
RNAPolymerase II
RNAPolymerase II
RNA synthesisTranscriptionInitiation complex
Chromatin changes
Transcription
RNA processing
mRNAdegradation
Translation
Protein processingand degradation
A DNA-bending proteinbrings the bound activators
closer to the promoter.Other transcription factors,
mediator proteins, and RNApolymerase are nearby.
2
Activator proteins bindto distal control elementsgrouped as an enhancer in the DNA. This enhancer hasthree binding sites.
1
The activators bind tocertain general transcription
factors and mediatorproteins, helping them form
an active transcriptioninitiation complex on the promoter.
3
Video Clip
• An activator
– Is a protein that binds to an enhancer and stimulates transcription of a gene
Figure 19.6

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Some specific transcription factors function as repressors
– To inhibit expression of a particular gene
• Some activators and repressors
– Act indirectly by influencing chromatin structure

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Cancer results from genetic changes that affect cell cycle control
• The gene regulation systems that go wrong during cancer
– Turn out to be the very same systems that play important roles in embryonic development

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Types of Genes Associated with Cancer
• The genes that normally regulate cell growth and division during the cell cycle
– Include genes for growth factors, their receptors, and the intracellular molecules of signaling pathways

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Oncogenes and Proto-Oncogenes
• Oncogenes
– Are cancer-causing genes
• Proto-oncogenes
– Are normal cellular genes that code for proteins that stimulate normal cell growth and division

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• A DNA change that makes a proto-oncogene excessively active
– Converts it to an oncogene, which may promote excessive cell division and cancer
Figure 19.11
Proto-oncogene
DNA
Translocation or transposition:gene moved to new locus,under new controls
Gene amplification:multiple copies of the gene
Point mutationwithin a controlelement
Point mutationwithin the gene
OncogeneOncogene
Normal growth-stimulatingprotein in excess
Hyperactive ordegradation-resistant protein
Normal growth-stimulatingprotein in excess
Normal growth-stimulatingprotein in excess
Newpromoter

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• The p53 gene encodes a tumor-suppressor protein
– That is a specific transcription factor that promotes the synthesis of cell cycle–inhibiting proteins
Figure 19.12b
UVlight
DNA
Defective ormissingtranscriptionfactor, such asp53, cannotactivatetranscription
MUTATION
Protein thatinhibitsthe cell cycle
pathway, DNA damage is an intracellularsignal that is passed via protein kinasesand leads to activation of p53. Activatedp53 promotes transcription of the gene for aprotein that inhibits the cell cycle. Theresulting suppression of cell division ensuresthat the damaged DNA is not replicated.Mutations causing deficiencies in anypathway component can contribute to thedevelopment of cancer.
(b) Cell cycle–inhibiting pathway. In this1
3
2
Protein kinases2
3 Activeformof p53
DNA damagein genome
1

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Mutations that knock out the p53 gene
– Can lead to excessive cell growth and cancer
Figure 19.12c
EFFECTS OF MUTATIONS
Proteinoverexpressed
Cell cycleoverstimulated
Increased celldivision
Cell cycle notinhibited
Protein absent
Effects of mutations. Increased cell division, possibly leading to cancer, can result if the cell cycle is overstimulated, as in (a), or not inhibited when it normally would be, as in (b).
(c)

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
The Multistep Model of Cancer Development
• Normal cells are converted to cancer cells
– By the accumulation of multiple mutations affecting proto-oncogenes and tumor-suppressor genes

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• A multistep model for the development of colorectal cancer
Figure 19.13
Colon
Colon wall
Normal colonepithelial cells
Small benigngrowth (polyp)
Larger benigngrowth (adenoma)
Malignant tumor(carcinoma)
2 Activation ofras oncogene
3 Loss oftumor-suppressorgene DCC
4 Loss oftumor-suppressorgene p53
5 Additionalmutations
1 Loss of tumor-suppressorgene APC (orother)

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Certain viruses
– Promote cancer by integration of viral DNA into a cell’s genome
• Individuals who inherit a mutant oncogene or tumor-suppressor allele
– Have an increased risk of developing certain types of cancer

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Eukaryotic genomes can have many noncoding DNA sequences in addition to genes
• The bulk of most eukaryotic genomes
– Consists of noncoding DNA sequences, often described in the past as “junk DNA”
• However, much evidence is accumulating
– That noncoding DNA plays important roles in the cell (Probably is no such thing as ‘junk DNA’)

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
The Relationship Between Genomic Composition and Organismal Complexity
• Compared with prokaryotic genomes, the genomes of eukaryotes
– Generally are larger
– Have longer genes
– Contain a much greater amount of noncoding DNA both associated with genes and between genes

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Transposable Elements and Related Sequences
• The first evidence for wandering DNA segments
– Came from geneticist Barbara McClintock’s breeding experiments with Indian corn
Figure 19.15

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
TransposonNew copy oftransposon
Transposonis copied
DNA of genome
Insertion
Mobile transposon
(a) Transposon movement (“copy-and-paste” mechanism)
RetrotransposonNew copy of
retrotransposon
DNA of genome
RNA
Reversetranscriptase
(b) Retrotransposon movement
Insertion
Movement of Transposons and Retrotransposons
• Eukaryotic transposable elements are of two types
– Transposons, which move within a genome by means of a DNA intermediate
– Retrotransposons, which move by means of an RNA intermediate
Figure 19.16a, b

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Sequences Related to Transposable Elements
• Multiple copies of transposable elements and sequences related to them
– Are scattered throughout the eukaryotic genome
• In humans
– A large portion of transposable element–related DNA consists of a family of similar sequences called Alu elements

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Other Repetitive DNA, Including Simple Sequence DNA
• Simple sequence DNA
– Contains many copies of tandemly repeated short sequences
– Is common in centromeres and telomeres, where it probably plays structural roles in the chromosome

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Genes and Multigene Families
• Most eukaryotic genes
– Are present in one copy per haploid set of chromosomes
• The rest of the genome
– Occurs in multigene families, collections of identical or very similar genes

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
DNA RNA transcripts
Non-transcribedspacer Transcription unit
DNA18S 5.8S 28S
rRNA
5.8S28S
18S
• Some multigene families
– Consist of identical DNA sequences, usually clustered tandemly, such as those that code for RNA products
Figure 19.17a Part of the ribosomal RNA gene family

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• The classic examples of multigene families of nonidentical genes
– Are two related families of genes that encode globins
Figure 19.17b The human -globin and -globin gene families
-GlobinHeme
Hemoglobin
-Globin
-Globin gene family -Globin gene family
Chromosome 16 Chromosome 11
Embryo
Fetusand adult Embryo Fetus Adult
GA
2
1
2 1

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Duplications, rearrangements, and mutations of DNA contribute to genome evolution
• The basis of change at the genomic level is mutation
– Which underlies much of genome evolution
Does the following produce new and novel information?

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Duplication of Chromosome Sets
• Accidents in cell division
– Can lead to extra copies of all or part of a genome, which may then diverge if one set accumulates sequence changes

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Duplication and Divergence of DNA Segments
• Unequal crossing over during prophase I of meiosis
– Can result in one chromosome with a deletion and another with a duplication of a particular gene
Figure 19.18
Nonsisterchromatids
Transposableelement
Gene
Incorrect pairingof two homologuesduring meiosis
Crossover
and

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Rearrangements of Parts of Genes: Exon Duplication and Exon Shuffling
• A particular exon within a gene
– Could be duplicated on one chromosome and deleted from the homologous chromosome

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• In exon shuffling
– Errors in meiotic recombination lead to the occasional mixing and matching of different exons either within a gene or between two nonallelic genes
Figure 19.20
EGF EGF EGF EGF
Epidermal growthfactor gene with multipleEGF exons (green)
F F F F
Fibronectin gene with multiple“finger” exons (orange)
Exonshuffling
Exonduplication
Exonshuffling
K
F EGF K K
Plasminogen gene with a“kfingle” exon (blue)
Portions of ancestral genes TPA gene as it exists today

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
How Transposable Elements Contribute to Genome Evolution
• Movement of transposable elements or recombination between copies of the same element
– Occasionally generates new sequence combinations that are beneficial to the organism (however, no new information was added)
• Some mechanisms
– Can alter the functions of genes or their patterns of expression and regulation