
Production of clonal planting materials from Gracilaria ... · changii and Kappaphycus alvarezii...
18
Production of clonal planting materials from Gracilaria changii and Kappaphycus alvarezii through tissue culture and culture of G. changii explants in airlift photobioreactors Hui-Yin Yeong & Siew-Moi Phang & C. R. K. Reddy & Norzulaani Khalid Received: 26 May 2013 /Revised: 15 August 2013 /Accepted: 16 August 2013 /Published online: 27 September 2013 # Springer Science+Business Media Dordrecht 2013 Abstract Global demand for seaweed resources has increased due to their emergent use as sources of biopharmaceuticals, nutraceuticals and biofuels. These high-valued products make possible the use of micropropagation techniques that may be more costly than conventional mariculture. This study reports the successful tissue culture of Kappaphycus alvarezii (Doty) Doty ex P. C. Silva and Gracilaria changii (B. Xia & Abbott) Abbott, Zhang and Xia. Callus induction of K. alvarezii was successfully developed following an explant sterilisation pro- tocol. Callus formation and regeneration of K. alvarezii was observed in solidified Provasoli’ s enriched seawater medium. Different culture conditions such as agar concentration, growth hormones, nutrients, irradiance and enrichment media were investigated to determine the suitable conditions for explant culture of G. changii . Proliferations of adventitious shoots were induced under the most suitable culture conditions. G. changii explants were successfully cultured in airlift photo- bioreactors, with no decrease in the carbohydrate content in the G. changii explants. This micropropagation technique can provide a useful alternative system for seedling production of economically important seaweeds. Keywords Tissue culture . Callus induction . Formation and regeneration . Explant culture . Plant growth regulators . Red seaweeds . Kappaphycus . Gracilaria . Micropropagation . Algae biotechnology Introduction World production of seaweed biomass has increased greatly from 1.995 million t (fresh weight) in 1970 to 19 million t (fresh weight) in 2010 (FAO 2007, 2012) to meet the demands for phycocolloid production, food and the emerging seaweed- based pharmaceutical, nutraceutical and biofuel industries. In 2010, 95.5 % of world seaweed biomass was produced through mariculture (FAO 2012). This led to the rapid devel- opment of seaweed cultivation technology aimed at the ad- vancement of industrial scale production of biomass. Efforts have been focussed on developing techniques for consistent supply of high-quality seed stock, strain improvement and efficient mass culture of high yielding commercial strains. Gracilaria and Kappaphycus are the two economically important red seaweeds in the world trade market especially for phycocolloid production. Gracilaria contributes 60–80 % of the world’ s agar (Schramm 1991; Armisen 1995; Bixler and Porse 2011). In 2009, agar production reached 12,500 t annually from the processing of 57,500 t of raw material (Bixler and Porse 2011). Approximately 71 % of the world’ s carrageenophyte resources, about 120,000 dry t.y -1 , is harvested from Kappaphycus alvarezii (Doty) Doty ex P. C. Silva (McHugh 2003) and has a market value of US $240 million annually. In Malaysia, Kappaphycus farming is found mainly in Sabah_s waters, and recently, a small-scale Kappaphycus farm (tambalang variety) was established at Pangkor Island, west coast Peninsular Malaysia (Phang 2006; Phang et al. 2010). There is a total 22 taxa of Gracilaria in Malaysian waters (Lim and Phang 2004; Phang 2006). Gracilaria changii (B. H.<Y. Yeong (*) : S.<M. Phang Institute of Ocean and Earth Sciences (IOES), University of Malaya, 50603 Kuala Lumpur, Malaysia e-mail: [email protected] S.<M. Phang : N. Khalid Institute of Biological Sciences, University of Malaya, 50603 Kuala Lumpur, Malaysia C. R. K. Reddy Discipline of Marine Biotechnology and Ecology, CSIR—Central Salt and Marine Chemicals Research Institute, Bhavnagar 364002, India J Appl Phycol (2014) 26:729–746 DOI 10.1007/s10811-013-0122-4
-
Upload
nguyendiep -
Category
Documents
-
view
217 -
download
0
Transcript of Production of clonal planting materials from Gracilaria ... · changii and Kappaphycus alvarezii...

Production of clonal planting materials from Gracilariachangii and Kappaphycus alvarezii through tissue cultureand culture of G. changii explants in airlift photobioreactors
Hui-Yin Yeong & Siew-Moi Phang & C. R. K. Reddy &
Norzulaani Khalid
Received: 26 May 2013 /Revised: 15 August 2013 /Accepted: 16 August 2013 /Published online: 27 September 2013# Springer Science+Business Media Dordrecht 2013
Abstract Global demand for seaweed resources has increaseddue to their emergent use as sources of biopharmaceuticals,nutraceuticals and biofuels. These high-valued products makepossible the use of micropropagation techniques that may bemore costly than conventional mariculture. This study reportsthe successful tissue culture of Kappaphycus alvarezii (Doty)Doty ex P. C. Silva and Gracilaria changii (B. Xia & Abbott)Abbott, Zhang and Xia. Callus induction of K. alvarezii wassuccessfully developed following an explant sterilisation pro-tocol. Callus formation and regeneration of K. alvarezii wasobserved in solidified Provasoli’s enriched seawater medium.Different culture conditions such as agar concentration, growthhormones, nutrients, irradiance and enrichment media wereinvestigated to determine the suitable conditions for explantculture of G. changii . Proliferations of adventitious shootswere induced under the most suitable culture conditions. G.changii explants were successfully cultured in airlift photo-bioreactors, with no decrease in the carbohydrate content in theG. changii explants. This micropropagation technique canprovide a useful alternative system for seedling production ofeconomically important seaweeds.
Keywords Tissue culture . Callus induction . Formation andregeneration . Explant culture . Plant growth regulators . Redseaweeds .Kappaphycus .Gracilaria . Micropropagation .
Algae biotechnology
Introduction
World production of seaweed biomass has increased greatlyfrom 1.995 million t (fresh weight) in 1970 to 19 million t(fresh weight) in 2010 (FAO 2007, 2012) to meet the demandsfor phycocolloid production, food and the emerging seaweed-based pharmaceutical, nutraceutical and biofuel industries. In2010, 95.5 % of world seaweed biomass was producedthrough mariculture (FAO 2012). This led to the rapid devel-opment of seaweed cultivation technology aimed at the ad-vancement of industrial scale production of biomass. Effortshave been focussed on developing techniques for consistentsupply of high-quality seed stock, strain improvement andefficient mass culture of high yielding commercial strains.
Gracilaria and Kappaphycus are the two economicallyimportant red seaweeds in the world trade market especiallyfor phycocolloid production. Gracilaria contributes 60–80 %of the world’s agar (Schramm 1991; Armisen 1995; Bixlerand Porse 2011). In 2009, agar production reached 12,500 tannually from the processing of 57,500 t of raw material(Bixler and Porse 2011). Approximately 71 % of the world’scarrageenophyte resources, about 120,000 dry t.y−1, is harvestedfrom Kappaphycus alvarezii (Doty) Doty ex P. C. Silva(McHugh 2003) and has a market value of US $240 millionannually. InMalaysia,Kappaphycus farming is foundmainly inSabah_s waters, and recently, a small-scale Kappaphycus farm(tambalang variety) was established at Pangkor Island, westcoast Peninsular Malaysia (Phang 2006; Phang et al. 2010).
There is a total 22 taxa of Gracilaria in Malaysian waters(Lim and Phang 2004; Phang 2006). Gracilaria changii (B.
H.<Y. Yeong (*) : S.<M. PhangInstitute of Ocean and Earth Sciences (IOES), University of Malaya,50603 Kuala Lumpur, Malaysiae-mail: [email protected]
S.<M. Phang :N. KhalidInstitute of Biological Sciences, University of Malaya,50603 Kuala Lumpur, Malaysia
C. R. K. ReddyDiscipline of Marine Biotechnology and Ecology, CSIR—CentralSalt and Marine Chemicals Research Institute,Bhavnagar 364002, India
J Appl Phycol (2014) 26:729–746DOI 10.1007/s10811-013-0122-4

Xia & Abbott) Abbott, Zhang and Xia, being one of the mostabundant agarophytes in Malaysia, contains high amounts ofagar (12 to 25 % dry weight) and agarose (13 to 16 % dryweight) with high gel strength (up to 563 g.cm−2 for agar and950 g.cm−2 for agarose) (Phang et al. 1996) and high amountsof β-carotene and essential fatty acids (20:5 ω3) (Phang et al.1996; Marinho-Soriano et al. 2001; Chu et al. 2003), bioactivecompounds (Wong et al. 1994), as well as possesses anti-inflammatory, gastroprotective and anti-ulcerogenic properties(Shu et al. 2013). Extensive research has been reported by theAlgae Research Laboratory of the University of Malaya onthis species including its genetic diversity (Sim et al. 2007;Yow et al. 2011, 2013), proteomics (Wong et al. 2006), func-tional genomics (Chan et al. 2004; Song et al. 2013),transcriptomics (Teo et al. 2007, 2009; Ho et al. 2009; Siowet al. 2012, 2013), genetic transformation (Gan et al. 2003,2006) and protoplast regeneration (Yeong et al. 2008) contrib-uting to prospective utilization of this alga for both applied andbasic research purposes. Thus, G. changii has great potentialfor commercial development in Malaysia. Furthermore, thesuccess in transformation of G. changii (Gan et al. 2003,2006) and protoplast isolation and regeneration of G. changii(Yeong et al. 2008) makes possible the commercialisation ofG. changii especially through bioprocess development for thepharmaceutical, nutraceutical and phycocolloid industries.
In general, tissue culture techniques developed for seaweedmicropropagation are basically established from tissue culturetechniques of higher plants. The first report of seaweed tissueculture (Chen and Taylor 1978) involved the red algaChondrus crispus . Since then, many species of red and brownalgae have been studied for callus induction and thallus re-generation. However, seaweed tissue culture is still in the stateof development and lags far behind that of higher plants(Reddy et al. 2008; Baweja et al. 2009). Parameters such asgrowth media, plant growth regulators and nutrients and cul-ture conditions such as temperature, irradiance and day lightcycle can influence the success of callus induction. Previousstudies showed success in the use of tissue culture techniquesfor the production of planting materials from Kappaphycus.Dawes and Koch (1991) reported the first successful tissueculture of K. alvarezii and then further established propaguleproduction for laboratory and field trials (Dawes et al. 1993,1994). Reddy et al. (2003) demonstrated better growth rates(1.5–1.8 times higher) for tissue culture plantlets compared tofield plants during the field trial (Reddy et al. 2003).Successful production of K. alvarezii plantlets from explantsculture (Hurtado and Biter 2007) and improved productionrate of explants for commercial nursery stocks (Yunque et al.2011) have been reported. However, withGracilaria spp., theproduction of planting materials through tissue culture re-mains at the laboratory stage. Successful callus inductionshave been reported for Gracilaria verrucosa (Gusev et al.1987; Kaczyna andMegnet 1993),Gracilaria textorii (Huang
and Fujita 1997), Gracilaria vermiculophylla (Yokoya et al.1999),Gracilaria chilensis (Collantes et al. 2004),Gracilariatenuistipitata , Gracilaria perplexa (Yokoya et al. 2004),Gracilaria corticata (Kumar et al. 2007) and Gracilariopsistenuifrons (Yokoya 2000), but no callus formation and regen-eration into new thalli has been reported for G. changiialthough ball-shaped friable callus-like structures were ob-served once (Tan 1999).
The production of clonal planting materials for commer-cially important seaweeds is not only to increase the seedlingstock production for cultivation but can also provide a con-stant supply of selected breeds with desired characteristics.Tissue culture techniques allow the production of disease-free(particularly virus-free) plants and rapid propagation of spe-cies which are difficult to multiply by conventional vegetativemeans. Malaysia as a Kappaphycus farming country has aneed to improve the current seaweed farming system by use ofhigh-quality planting materials to ensure the stability of pro-duction and the sustainable development. Our objective in thisstudy was to develop seaweed tissue culture techniques forproducing clonal planting materials for the two most promis-ing seaweeds, namely K. alvarezii and G. changii . The effectof six plant growth regulators, sources of nitrogen and phos-phorus, a polyamine, pH, irradiance and light–dark cycle, oncallus formation was investigated. In addition, the culture ofexplants of G. changii was investigated using two types ofairlift photobioreactors.
Materials and methods
Healthy Gracilaria changii plants were collected from man-grove areas of Morib, Selangor, west coast PeninsularMalaysia (02°45′48.5″N, 101°26′8.6″E) during low tidewhile K. alvarezii plants were collected from theKappaphycus farm at Semporna, Sabah, East Malaysia andkept cool in an ice-chest with some seawater for transport backto the laboratory. In the laboratory, the samples were cleanedof mud and epiphytes using a soft brush and filtered seawater.The samples were maintained as unicultures in tanks of fil-tered and aerated seawater in both the hatchery and laboratory(25±2 °C, 25 μmol photons m−2 s−1 irradiance, 12-h:12-hlight – dark cycle, 28±2 ppt seawater salinity) for about4 weeks before they were used in the studies.
Thalli of G. changii or K. alvarezii were thoroughlycleared of epiphytes using forceps and autoclaved seawaterunder a dissecting microscope. The thalli were then cut into5 cm pieces and rinsed several times with autoclaved artificialseawater (Instant Ocean; Atkinson and Bingman 1997)followed by sonication in autoclaved artificial seawater for3 min before treatment with 1 % w/v PVP-I for 30 sec. Thethalli segments were then immersed in autoclaved artificialseawater with an antibiotic mixture containing 0.100 g L−1
730 J Appl Phycol (2014) 26:729–746

kanamycin, 0.300 g L−1 penicillin G, 0.020 g L−1 polymyxinB sulphate, 0.001 g L−1 nalidixic acid, 0.020 g L−1 cefotaxineand 1.000 g L−1 streptomycin sulphate for 12 h (Yeong et al.2008). Sterility was tested using LB broth (Merck, Germany)and Marine Agar 2216E (Difco) medium.
Tissue culture
The sterilised thalli were cut into short explants (length~5 mm) using a sterile surgical blade and inoculated in thedesired culture media as listed in Tables 1 and 2. Differentculture media were prepared according to the experimentalrequirements, and autoclaved sterile artificial seawater(Instant Ocean; Atkinson and Bingman 1997) was used asbase medium. Five explants were inoculated in a 90-mm Petridish for solid media, or in a 200-mL culture vessel for liquidmedia (25 °C, 25 μmol photons m−2 s−1 irradiance, 12-h:12-hlight – dark cycle, unless specified otherwise). Fresh mediawere supplied weekly throughout the culture period.Observations of callus and adventitious branch induction weremade weekly up to day 56.
In order to determine the suitable tissue culture media andconditions for callus induction in the G. changii explantsculture, the effect of agar (liquid media and solid media using0.8%w/vBactoAgar), enrichment liquidmedia as Provasoli’senriched seawater (PES) media (James 1978) or Guillard’s(f/2) marine water enrichment solution (GMWES) (Guillard1975), plant growth regulators (PGRs) 2,4-dichlorophenoxyacetic acid (2,4-D) (Technology Laboratories), indole-3-aceticacid (IAA) (Sigma), α-naphthalene acetic acid (NAA)(Sigma), zeatin (Sigma), nutrient supplementation (phosphateas K2PO4, sulphate as K2SO4, nitrogen as NH4Cl and urea),polyamines, irradiance, light–dark cycle and pH was investi-gated (Table 1). For K. alvarezii explant culture, the effect ofagar (liquid media and solid media using 1.0 % w/v BactoAgar), enrichment liquid media as PES medium, PGRs (singlePGR (2,4-D) or combination of PGRs (2,4-D and kinetin) wasinvestigated (Table 2). All studies were conducted in threereplicates (minimum) and incubated at 25 °C in a growthchamber (Protech Model GC-1050).
Simultaneous studies on callus induction
Thalli of G. changii and K. alvarezii were sterilised using theoptimized sterilization protocol described above. Thesterilised thalli were cut into short explants (length ~5 mm)using a sterile surgical blade, and ten explants were inoculatedinto 1.0 % Bacto agar solidified PES medium in each 90-mmpetri dish. Both explant cultures were placed in an incubatorwith 25 μmol photons m−2 s−1 irradiance, 12-h:12-h light –dark cycle at 25 °C. The experiments were repeated threetimes using three different batches of seaweed samples, andabout 60–100 explants were tested each time for each species.
These experiments were conducted at the same time for thetwo species, and all protocols carried out were the same inorder to compare the responses of the two species to the sameconditions for callus induction.
Effect of airlift photobioreactor on explant culture of G.changii
Two types of airlift photobioreactors, namely a 5-L tubularairlift photobioreactor (working volume 3 L) and a 1-L spher-ical airlift photobioreactor (working volume 500 mL), wereused (Fig. 1). PES liquid culture media with or without plantgrowth regulators (0.1 mg L−1 2,4-D+0.1 mg L−1 kinetin)were used as the base media. Healthy axenic thalli sections ofG. changii were cut into short explants (lengths ~10 and~20 mm) using a sterile surgical blade. Approximately0.002 g mL−1 of explants of two lengths (~10 and ~20 mm)with equal weight per volume was inoculated into the 5-Lairlift photobioreactor, while only ~10-mm explants wereinoculated into the 1-L airlift photobioreactor. The airliftphotobioreactors were placed under an irradiance of25 μmol photons m−2 s−1 with 12-h:12-h light – dark cycleat 25 °C. Fifty percent of the culture medium was replacedwith fresh medium weekly. Observations of adventitiousbranch generation were made weekly up to day 72. After72 days of culture, G. changii explants were harvested forcarbohydrate extraction. Carbohydrate contained in G.changii explants were extracted in 2 N HCl and quantifiedusing the phenol–sulphuric acid method (Kochert 1978).
Results
Tissue culture of G. changii
In experiment 1, explants cultured in Guillard’s (f/2) GMWESwith 3.315 mg L−1 2,4-D showed the highest percentage ofexplants with adventitious branches (12.00±4.90 %). About8.00 % of explants cultured in 1.105, 2.210 and 4.420 mg L−1
2,4-D treatment produced adventitious branches. In the IAAand NAA treatments, adventitious branches were only ob-served in 1.105 mg L−1 IAA and 0.221 mg L−1 NAA (6.67±6.67 %), while no branches formed at all other concentra-tions of IAA and NAA tested. Explants cultured in GMWESonly and with 0.221 mg L−1 2,4-D also did not produce anybranches. Those explants without new branches were found tohave lost their pigmentation and became bleached by the endof day 56. However, Kruskal–Wallis ANOVA showed nosignificant differences on the effect of plant growth regulatorson formation of adventitious branches in the explants (p >0.05)(Fig. 2).
In experiment 2, five different concentrations of 2,4-D inboth 0.8 % w/v Bacto agar solidified GMWES medium and
J Appl Phycol (2014) 26:729–746 731
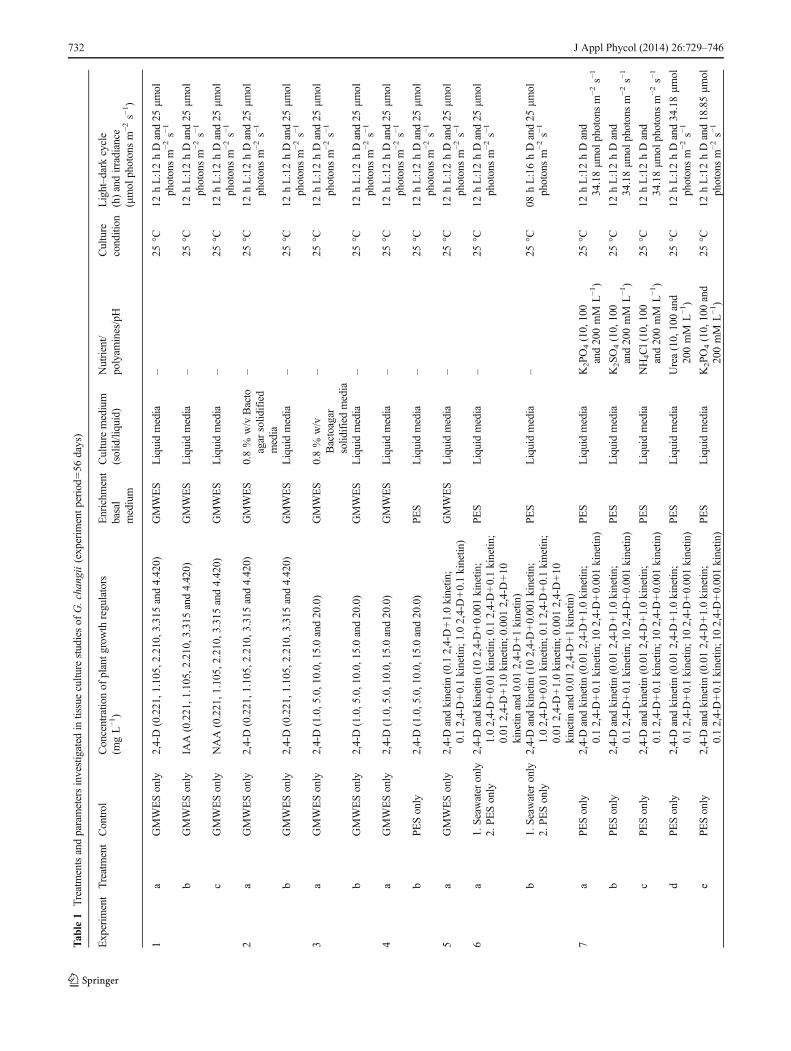
Tab
le1
Treatmentsandparametersinvestigated
intissuecultu
restudiesof
G.changii(experim
entp
eriod=56
days)
Experim
entTreatmentControl
Concentratio
nof
plantg
rowth
regulators
(mgL−1)
Enrichm
ent
basal
medium
Culture
medium
(solid/liquid)
Nutrient/
polyam
ines/pH
Culture
condition
Light–dark
cycle
(h)andirradiance
(μmol
photonsm
−2s−
1)
1a
GMWESonly
2,4-D(0.221,1.105,2.210,3.315
and4.420)
GMWES
Liquidmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
bGMWESonly
IAA(0.221,1.105,2.210,3.315
and4.420)
GMWES
Liquidmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
cGMWESonly
NAA(0.221,1.105,2.210,3.315
and4.420)
GMWES
Liquidmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
2a
GMWESonly
2,4-D(0.221,1.105,2.210,3
.315
and4.420)
GMWES
0.8%
w/v
Bacto
agar
solid
ified
media
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
bGMWESonly
2,4-D(0.221,1.105,2.210,3
.315
and4.420)
GMWES
Liquidmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
3a
GMWESonly
2,4-D(1.0,5.0,10.0,15.0and20.0)
GMWES
0.8%
w/v
Bactoagar
solid
ifiedmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
bGMWESonly
2,4-D(1.0,5.0,10.0,15.0and20.0)
GMWES
Liquidmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
4a
GMWESonly
2,4-D(1.0,5.0,10.0,15.0and20.0)
GMWES
Liquidmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
bPESonly
2,4-D(1.0,5.0,10.0,15.0and20.0)
PES
Liquidmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
5a
GMWESonly
2,4-Dandkinetin
(0.12,4-D+1.0kinetin
;0.12,4-D+0.1kinetin
;1.02,4-D+0.1kinetin
)GMWES
Liquidmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
6a
1.Seaw
ater
only
2,4-Dandkinetin
(102,4-D+0.001kinetin
;1.02,4-D+0.01
kinetin
;0.12,4-D+0.1kinetin
;0.01
2,4-D+1.0kinetin
;0.001
2,4-D+10
kinetin
and0.01
2,4-D+1kinetin
)
PES
Liquidmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
12.PE
Sonly
b1.Seawater
only
2,4-Dandkinetin
(102,4-D+0.001kinetin
;1.02,4-D+0.01
kinetin
;0.12,4-D+0.1kinetin
;0.01
2,4-D+1.0kinetin
;0.001
2,4-D+10
kinetin
and0.01
2,4-D+1kinetin
)
PES
Liquidmedia
–25
°C08
hL:16hDand25
μmol
photonsm
−2s−
12.PE
Sonly
7a
PESonly
2,4-Dandkinetin
(0.012,4-D+1.0kinetin
;0.12,4-D+0.1kinetin
;102,4-D+0.001kinetin
)PE
SLiquidmedia
K2PO4(10,100
and200mM
L−1)
25°C
12hL:12hDand
34.18μm
olphotonsm
−2s−
1
bPESonly
2,4-Dandkinetin
(0.012,4-D+1.0kinetin
;0.12,4-D+0.1kinetin
;102,4-D+0.001kinetin
)PE
SLiquidmedia
K2SO4(10,100
and200mM
L−1)
25°C
12hL:12hDand
34.18μm
olphotonsm
−2s−
1
cPESonly
2,4-Dandkinetin
(0.012,4-D+1.0kinetin
;0.12,4-D+0.1kinetin
;102,4-D+0.001kinetin
)PE
SLiquidmedia
NH4Cl(10,100
and200mM
L−1)
25°C
12hL:12hDand
34.18μm
olphotonsm
−2s−
1
dPESonly
2,4-Dandkinetin
(0.012,4-D+1.0kinetin
;0.12,4-D+0.1kinetin
;102,4-D+0.001kinetin
)PE
SLiquidmedia
Urea(10,100and
200mM
L−1)
25°C
12hL:12hDand34.18μm
olphotonsm
−2s−
1
ePESonly
2,4-Dandkinetin
(0.012,4-D+1.0kinetin
;0.12,4-D+0.1kinetin
;102,4-D+0.001kinetin
)PE
SLiquidmedia
K2PO4(10,100and
200mM
L−1)
25°C
12hL:12hDand18.85μm
olphotonsm
−2s−
1
732 J Appl Phycol (2014) 26:729–746
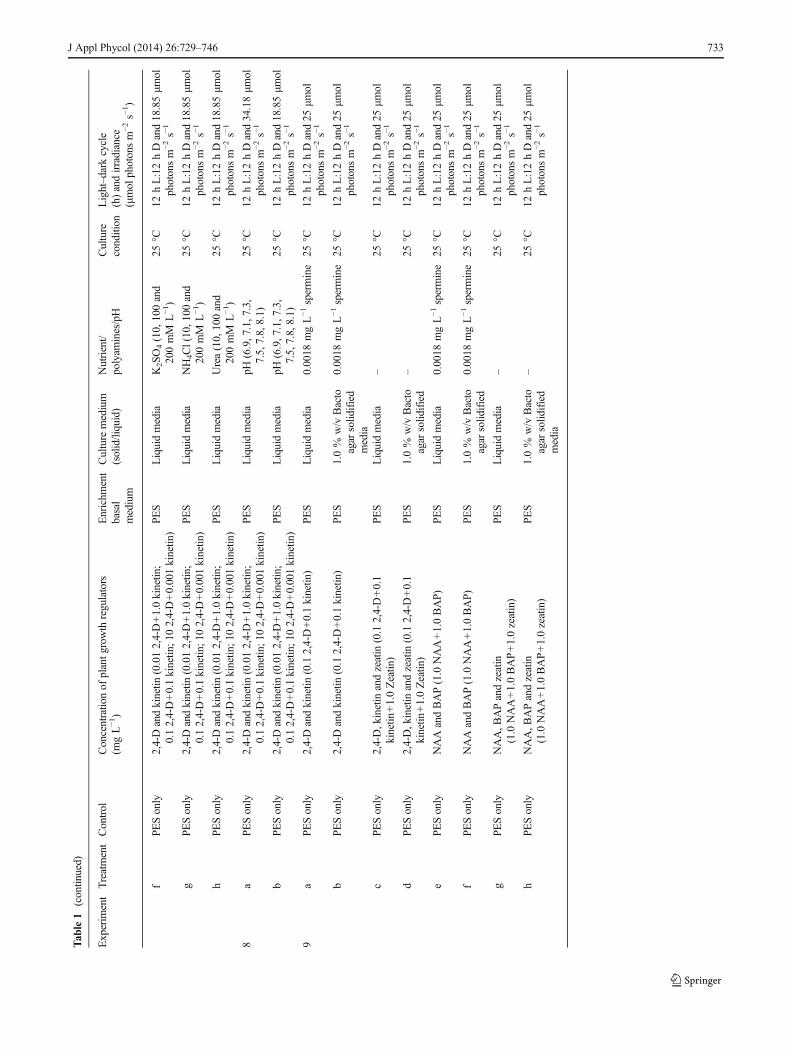
Tab
le1
(contin
ued)
Experim
entTreatmentControl
Concentratio
nof
plantg
rowth
regulators
(mgL−1)
Enrichm
ent
basal
medium
Culture
medium
(solid/liquid)
Nutrient/
polyam
ines/pH
Culture
condition
Light–dark
cycle
(h)andirradiance
(μmol
photonsm
−2s−
1)
fPESonly
2,4-Dandkinetin
(0.012,4-D+1.0kinetin
;0.12,4-D+0.1kinetin
;102,4-D+0.001kinetin
)PE
SLiquidmedia
K2SO4(10,100and
200mM
L−1)
25°C
12hL:12hDand18.85μm
olphotonsm
−2s−
1
gPESonly
2,4-Dandkinetin
(0.012,4-D+1.0kinetin
;0.12,4-D+0.1kinetin
;102,4-D+0.001kinetin
)PE
SLiquidmedia
NH4Cl(10,100
and
200mM
L−1)
25°C
12hL:12hDand18.85μm
olphotonsm
−2s−
1
hPESonly
2,4-Dandkinetin
(0.012,4-D+1.0kinetin
;0.12,4-D+0.1kinetin
;102,4-D+0.001kinetin
)PE
SLiquidmedia
Urea(10,100and
200mM
L−1)
25°C
12hL:12hDand18.85μm
olphotonsm
−2s−
1
8a
PESonly
2,4-Dandkinetin
(0.012,4-D+1.0kinetin
;0.12,4-D+0.1kinetin
;102,4-D+0.001kinetin
)PE
SLiquidmedia
pH(6.9,7.1,7.3,
7.5,7.8,8.1)
25°C
12hL:12hDand34.18μm
olphotonsm
−2s−
1
bPESonly
2,4-Dandkinetin
(0.012,4-D+1.0kinetin
;0.12,4-D+0.1kinetin
;102,4-D+0.001kinetin
)PE
SLiquidmedia
pH(6.9,7.1,7.3,
7.5,7.8,8.1)
25°C
12hL:12hDand18.85μm
olphotonsm
−2s−
1
9a
PESonly
2,4-Dandkinetin
(0.12,4-D+0.1kinetin
)PES
Liquidmedia
0.0018
mgL−1spermine
25°C
12hL:12hDand25
μmol
photonsm
−2s−
1
bPESonly
2,4-Dandkinetin
(0.12,4-D+0.1kinetin
)PES
1.0%
w/v
Bacto
agar
solid
ified
media
0.0018
mgL−1spermine
25°C
12hL:12hDand25
μmol
photonsm
−2s−
1
cPESonly
2,4-D,kinetin
andzeatin
(0.12,4-D+0.1
kinetin
+1.0Zeatin
)PE
SLiquidmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
dPESonly
2,4-D,kinetin
andzeatin
(0.12,4-D+0.1
kinetin
+1.0Zeatin
)PES
1.0%
w/v
Bacto
agar
solid
ified
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
ePESonly
NAAandBAP(1.0NAA+1.0BAP)
PES
Liquidmedia
0.0018
mgL−1spermine
25°C
12hL:12hDand25
μmol
photonsm
−2s−
1
fPESonly
NAAandBAP(1.0NAA+1.0BAP)
PES
1.0%
w/v
Bacto
agar
solid
ified
0.0018
mgL−1spermine
25°C
12hL:12hDand25
μmol
photonsm
−2s−
1
gPESonly
NAA,B
APandzeatin
(1.0NAA+1.0BAP+1.0zeatin)
PES
Liquidmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
hPESonly
NAA,B
APandzeatin
(1.0NAA+1.0BAP+1.0zeatin)
PES
1.0%
w/v
Bacto
agar
solid
ified
media
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
J Appl Phycol (2014) 26:729–746 733
liquid GMWES medium were tested. Solid media with3.315 mg L−1 2,4-D produced the highest number of explantswith new branches (32.00±12.00 %) while no branch forma-tion was observed in liquid media without 2,4-D and with0.221 mg L−1 2,4-D (Fig. 3). All solid media either with orwithout 2,4-D induced adventitious branches while in liquidmedia, adventitious branch formation was only observed intreatments with 1.105 mg L−1 2,4-D. A significant differencewas found in the effect of media on the success of adventitiousbranch formation (p <0.05) but not in the effect of differentconcentrations of plant growth regulator.
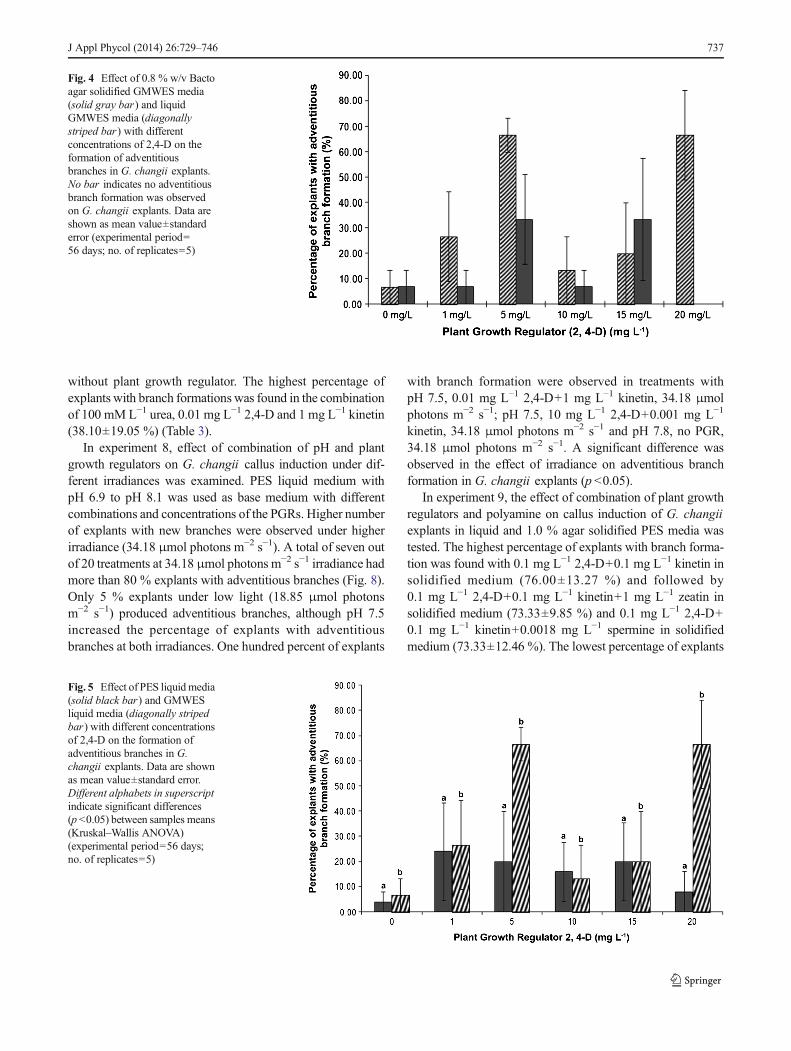
Higher concentrations of 2,4-D (1, 5, 10, 15 and 20mg L−1)in both 0.8 % w/v Bacto agar solidified GMWES and liquidGMWES media were investigated in experiment 3. A highernumber of adventitious branches were observed in liquid me-dia compared to solid media (Fig. 4). The highest percentage ofexplants with branches was 66.67 % observed in liquid mediawith either 5 mg L−1 or 20 mg L−1 2,4-D. No branch formationwas observed on solid media with 20 mg L−1 2,4-D, while thelowest percentage (6.67%) of explants with new branches wasin liquid media without 2,4-D and solid media with 0, 1 and10 mg L−1 2,4-D.
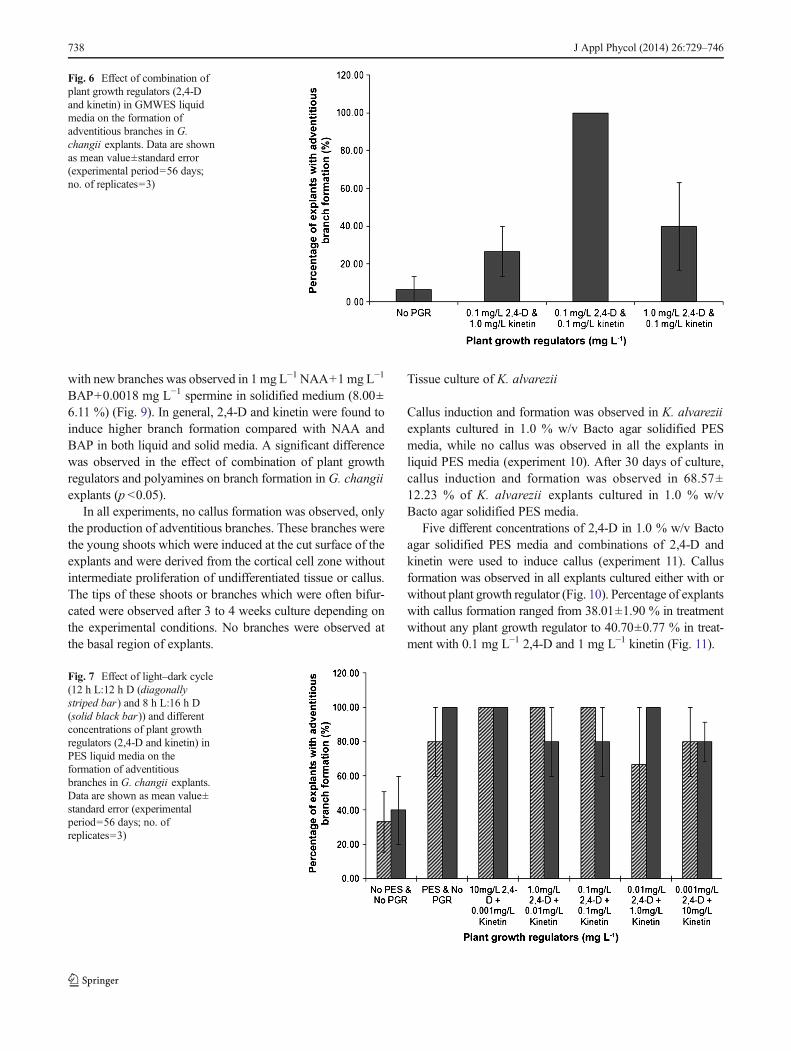
In experiment 4, both PES and GMWES media with fivedifferent concentrations of 2,4-D were used. GMWES witheither 5 or 20 mg L−1 2,4-D produced the highest percentageof explants with new branch formation (66.67±6.67 and66.67±17.64 %, respectively) followed by PES liquid mediawith 1 mg L−1 2,4-D (24.00±19.39 %). PES liquid mediumwithout 2,4-D had the lowest percentage of explants with newbranch formation (4.00±4.00 %) (Fig. 5). A significant dif-ference was found in the effect of enrichment media on thepercentage of explants with new branch formation (p <0.05),but not in the effect of different concentrations of plant growthregulators.
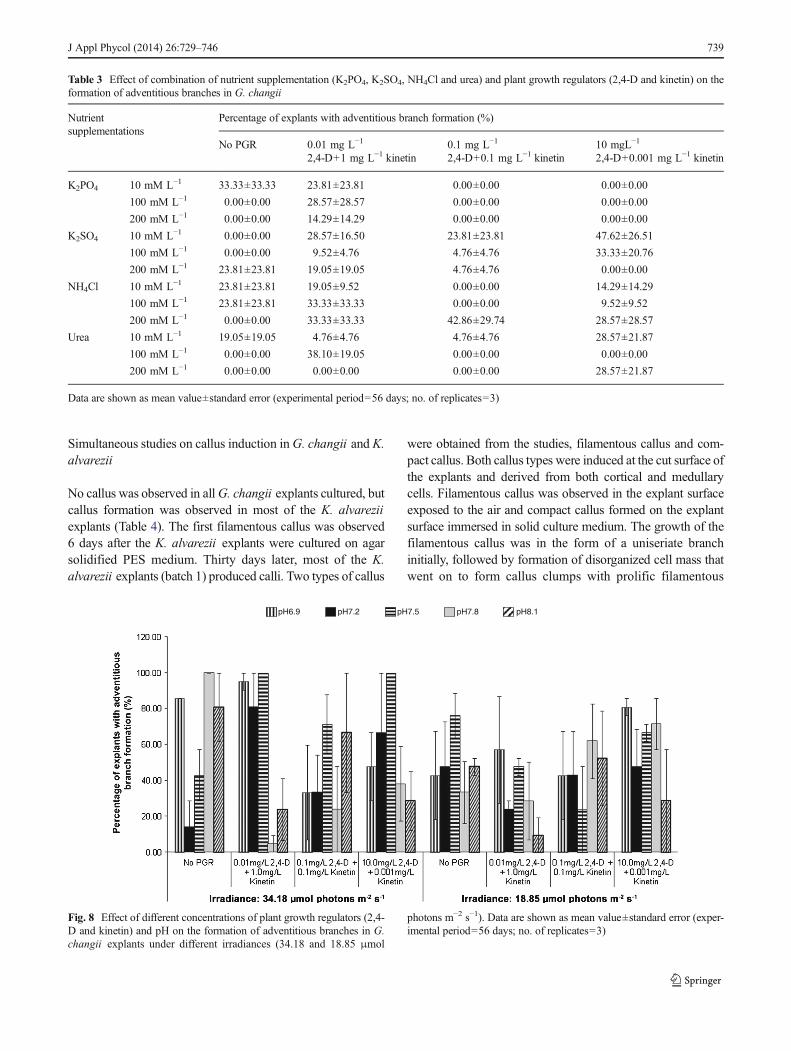
In experiment 5, three combinations of plant growth weretested with GMWES as the basal medium. A combination of0.1 mg L−1 2,4-D and 0.1 mg L−1 kinetin was the best with100% of the explant cultures producing adventitious branches(Fig. 6). This was followed by 1 mg L−1 2,4 D and 0.1 mg L−1
kinetin (40.00±23.09 %) and 0.1 mg L−1 2,4-D and 1 mg L−1
kinetin (26.67±13.33 %). Treatments without plant growthregulators (control) formed the lowest number of newbranches (6.67±6.67 %). However, there was no significantdifference in the effect of plant growth regulators on newbranch formation. Furthermore, no callus induction was ob-served in G. changii explants in all treatments.
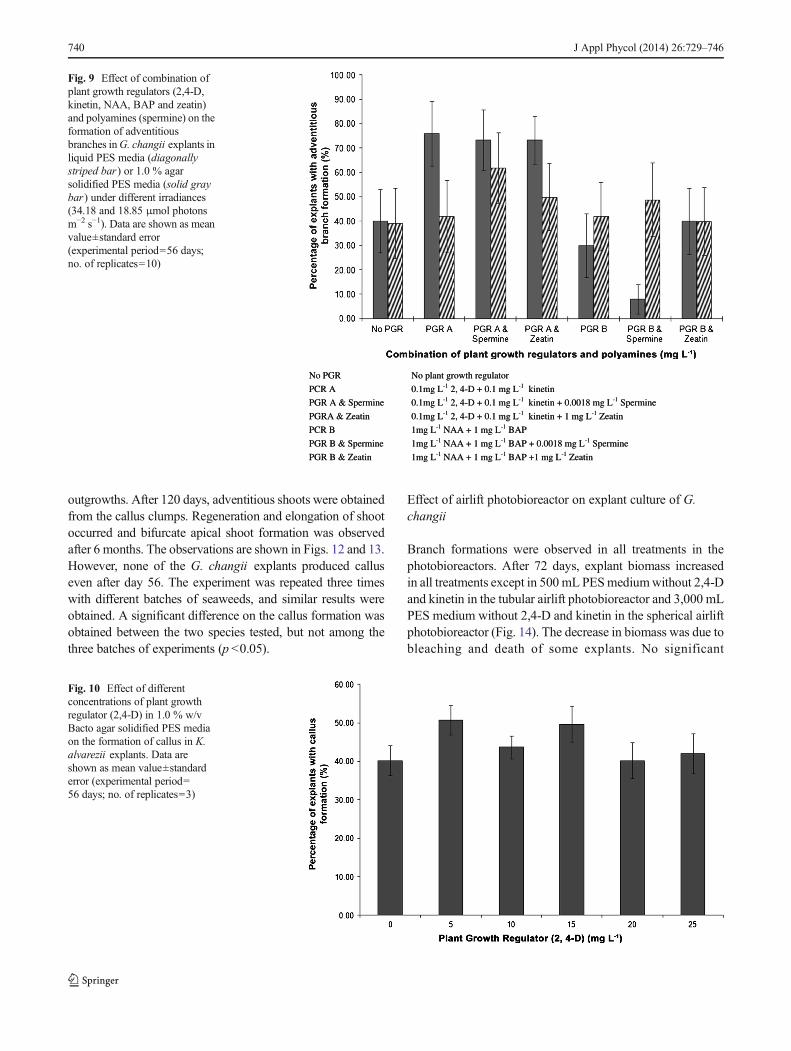
When variation in the light–dark cycle ( 12-h:12-h L:D or8-h:16-h L:D) was combined with different concentrations of2,4-D and kinetin on G. changii callus induction (experiment6), the use of PES enrichment medium as base mediumtogether with 2,4-D and kinetin increased adventitious branchformations in all treatments. In the 12 h light – 12 h dark cycle,adventitious branch formation was observed in 100 % of theT
able2
Treatmentsandparametersinvestigated
intissuecultu
restudiesof
K.alvarezii(experim
entp
eriod=56
days)
Experim
ent
Treatment
Control
Concentratio
nof
plantg
rowth
regulators(m
gL−1)
Enrichm
ent
basalm
edium
Culture
medium
(solid/liquid)
Nutrient/
polyam
ines/pH
Culture
condition
Light–dark
cycle(h)and
irradiance
(μmol
photonsm
−2s−
1)
10a
PESonly
–PES
1.0%
w/v
Bacto
agar
solid
ifiedmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
bPESonly
–PE
SLiquidmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
11a
PESonly
2,4-D(0,5,10,15,20)
PES
1.0%
w/v
Bacto
agar
solid
ifiedmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
12a
PESonly
2,4-Dandkinetin
(0.1
2,4-D+1.0kinetin
;0.1
2,4-D+0.1kinetin
;1.0
2,4-D+0.1kinetin
)PES
1.0%
w/v
Bacto
agar
solid
ifiedmedia
–25
°C12
hL:12hDand25
μmol
photonsm
−2s−
1
734 J Appl Phycol (2014) 26:729–746

explants in PES liquid media with 10 mg L−1 2,4-D+0.001 mg L−1 kinetin; 1 mg L−1 2,4-D+0.01 mg L−1 kinetinand 0.1 mg L−1 2,4-D+0.1 mg L−1 kinetin while in a 8-hlight – 16-h dark cycle, 100 % adventitious branch formationwas observed in PES liquid media with 0 mg L−1 2,4-D+0 mg L−1 kinetin; 10 mg L−1 2,4-D+0.001 mg L−1 kinetinand 0.01 mg L−1 2,4-D+1 mg L−1 kinetin (Fig. 7). Liquidmedia without PES enrichment and plant growth regula-tors had the lowest percentage of adventitious branchformation in both light–dark cycle treatments (33.33±17.64 % in 12-h:12-h L:D cycle and 40.00±20.00 % ina 8-h:16-h L:D cycle).
In experiment 7, a total of three concentrations of fournutrients, namely phosphate (K2PO4), sulphate (K2SO4), ni-trogen (NH4Cl) and urea, were tested together with three
different combinations of 2,4-D and kinetin in PES liquidmedia. Adventitious branch formations were observed in G.changii explants in selected treatments (Table 3). For thetreatments with a combination of phosphate (K2PO4) andplant growth regulators, adventitious branch formations werefound with 0.01 mg L−1 2,4-D+1 mg L−1 kinetin with K2PO4
(10, 100 and 200 mM L−1) with 23.81±23.81, 28.57±28.57and 14.29±14.29 % of explants with new branch formation,respectively. For the treatments with a combination of sul-phate (K2SO4) and plant growth regulators, adventitiousbranch formations were found in all the treatments except inthe combinations of 10 and 100 mM L−1 K2SO4 without 2,4-D and kinetin and the combination of 200 mM L−1 K2SO4
with 10 mg L−1 2,4-D+0.001 mg L−1 kinetin. The highestpercentage of explants with adventitious branch formations
c
a
b
Fig. 1 The three types of airliftphotobioreactors used in theexperiment. a 5 L spherical airliftphotobioreactor. b 5 L tubularairlift photobioreactor. c 1 Ltubular airlift photobioreactor
J Appl Phycol (2014) 26:729–746 735
was found in the treatment with the combination of10 mM L−1 K2SO4 with 10 mg L−1 2,4-D+0.001 mg L−1
kinetin (47.62±26.51 %). In the treatments on combination ofnitrogen (NH4Cl) and plant growth regulators, adventitiousbranch formations were not found in the combinations of 10and 100 mM L−1 NH4Cl with 0.1 mg L−1 2,4-D+0.1 mg L−1
kinetin and in 200 mM L−1 NH4Cl without plant growth
regulators. However, the highest percentage of explants withnew branch formations (42.86±29.74 %) was found in thetreatment with the combination 200 mM L−1 NH4Cl with0.1 mg L−1 2,4-D+0.1 mg L−1 kinetin. For the treatments oncombination of urea and plant growth regulators, no specifictrend was observed. Adventitious branch formations wereobserved in the combination of 10 mM L−1 urea with and
Fig. 2 Effect of different concentrations of plant growth regulators (2,4-D, IAA and NAA) in GMWES liquid media on the formation of adven-titious branches in G. changii explants. No bar indicates no adventitious
branch formation was observed on G. changii explants. Data are shownas mean value±standard error (experimental period=56 days; no. ofreplicates=5)
Fig. 3 Effect of 0.8 % w/v Bacto agar solidified GMWES media (solidgray bar) and liquid GMWES media (diagonally striped bar) withdifferent concentrations of 2,4-D on the formation of adventitiousbranches inG. changii explants. No bar indicates no adventitious branch
formation was observed onG. changii explants. Data are shown as meanvalue±standard error. Different alphabets in superscript indicate signif-icant differences (p <0.05) between samples means (Kruskal–WallisANOVA) (experimental period=56 days; no. of replicates=5)
736 J Appl Phycol (2014) 26:729–746
without plant growth regulator. The highest percentage ofexplants with branch formations was found in the combinationof 100 mM L−1 urea, 0.01 mg L−1 2,4-D and 1 mg L−1 kinetin(38.10±19.05 %) (Table 3).
In experiment 8, effect of combination of pH and plantgrowth regulators on G. changii callus induction under dif-ferent irradiances was examined. PES liquid medium withpH 6.9 to pH 8.1 was used as base medium with differentcombinations and concentrations of the PGRs. Higher numberof explants with new branches were observed under higherirradiance (34.18 μmol photons m−2 s−1). A total of seven outof 20 treatments at 34.18 μmol photons m−2 s−1 irradiance hadmore than 80 % explants with adventitious branches (Fig. 8).Only 5 % explants under low light (18.85 μmol photonsm−2 s−1) produced adventitious branches, although pH 7.5increased the percentage of explants with adventitiousbranches at both irradiances. One hundred percent of explants
with branch formation were observed in treatments withpH 7.5, 0.01 mg L−1 2,4-D+1 mg L−1 kinetin, 34.18 μmolphotons m−2 s−1; pH 7.5, 10 mg L−1 2,4-D+0.001 mg L−1
kinetin, 34.18 μmol photons m−2 s−1 and pH 7.8, no PGR,34.18 μmol photons m−2 s−1. A significant difference wasobserved in the effect of irradiance on adventitious branchformation in G. changii explants (p <0.05).
In experiment 9, the effect of combination of plant growthregulators and polyamine on callus induction of G. changiiexplants in liquid and 1.0 % agar solidified PES media wastested. The highest percentage of explants with branch forma-tion was found with 0.1 mg L−1 2,4-D+0.1 mg L−1 kinetin insolidified medium (76.00±13.27 %) and followed by0.1 mg L−1 2,4-D+0.1 mg L−1 kinetin+1 mg L−1 zeatin insolidified medium (73.33±9.85 %) and 0.1 mg L−1 2,4-D+0.1 mg L−1 kinetin+0.0018 mg L−1 spermine in solidifiedmedium (73.33±12.46 %). The lowest percentage of explants
Fig. 4 Effect of 0.8 % w/v Bactoagar solidified GMWES media(solid gray bar) and liquidGMWES media (diagonallystriped bar) with differentconcentrations of 2,4-D on theformation of adventitiousbranches in G. changii explants.No bar indicates no adventitiousbranch formation was observedon G. changii explants. Data areshown as mean value±standarderror (experimental period=56 days; no. of replicates=5)
Fig. 5 Effect of PES liquid media(solid black bar) and GMWESliquid media (diagonally stripedbar) with different concentrationsof 2,4-D on the formation ofadventitious branches in G.changii explants. Data are shownas mean value±standard error.Different alphabets in superscriptindicate significant differences(p <0.05) between samples means(Kruskal–Wallis ANOVA)(experimental period=56 days;no. of replicates=5)
J Appl Phycol (2014) 26:729–746 737
with new branches was observed in 1 mg L−1 NAA+1mg L−1
BAP+0.0018 mg L−1 spermine in solidified medium (8.00±6.11 %) (Fig. 9). In general, 2,4-D and kinetin were found toinduce higher branch formation compared with NAA andBAP in both liquid and solid media. A significant differencewas observed in the effect of combination of plant growthregulators and polyamines on branch formation in G. changiiexplants (p <0.05).
In all experiments, no callus formation was observed, onlythe production of adventitious branches. These branches werethe young shoots which were induced at the cut surface of theexplants and were derived from the cortical cell zone withoutintermediate proliferation of undifferentiated tissue or callus.The tips of these shoots or branches which were often bifur-cated were observed after 3 to 4 weeks culture depending onthe experimental conditions. No branches were observed atthe basal region of explants.
Tissue culture of K. alvarezii
Callus induction and formation was observed in K. alvareziiexplants cultured in 1.0 % w/v Bacto agar solidified PESmedia, while no callus was observed in all the explants inliquid PES media (experiment 10). After 30 days of culture,callus induction and formation was observed in 68.57±12.23 % of K. alvarezii explants cultured in 1.0 % w/vBacto agar solidified PES media.
Five different concentrations of 2,4-D in 1.0 % w/v Bactoagar solidified PES media and combinations of 2,4-D andkinetin were used to induce callus (experiment 11). Callusformation was observed in all explants cultured either with orwithout plant growth regulator (Fig. 10). Percentage of explantswith callus formation ranged from 38.01±1.90 % in treatmentwithout any plant growth regulator to 40.70±0.77 % in treat-ment with 0.1 mg L−1 2,4-D and 1 mg L−1 kinetin (Fig. 11).
Fig. 6 Effect of combination ofplant growth regulators (2,4-Dand kinetin) in GMWES liquidmedia on the formation ofadventitious branches in G.changii explants. Data are shownas mean value±standard error(experimental period=56 days;no. of replicates=3)
Fig. 7 Effect of light–dark cycle(12 h L:12 h D (diagonallystriped bar) and 8 h L:16 h D(solid black bar)) and differentconcentrations of plant growthregulators (2,4-D and kinetin) inPES liquid media on theformation of adventitiousbranches in G. changii explants.Data are shown as mean value±standard error (experimentalperiod=56 days; no. ofreplicates=3)
738 J Appl Phycol (2014) 26:729–746
Simultaneous studies on callus induction inG. changii and K.alvarezii
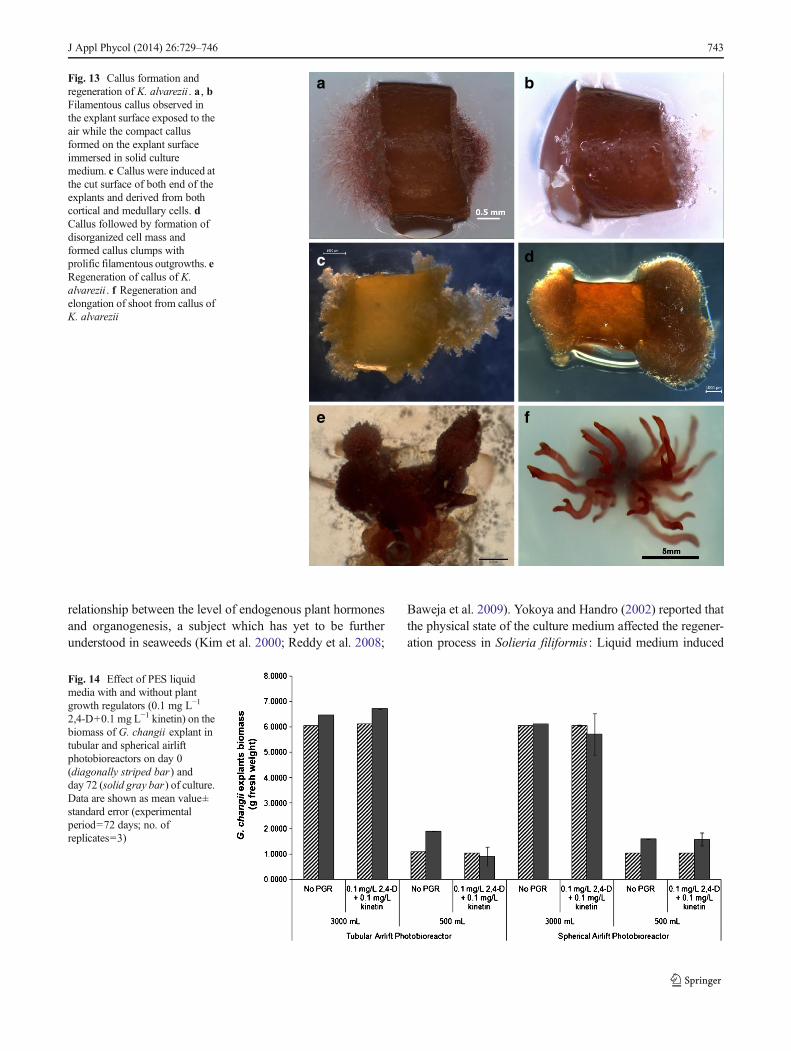
No callus was observed in allG. changii explants cultured, butcallus formation was observed in most of the K. alvareziiexplants (Table 4). The first filamentous callus was observed6 days after the K. alvarezii explants were cultured on agarsolidified PES medium. Thirty days later, most of the K.alvarezii explants (batch 1) produced calli. Two types of callus
were obtained from the studies, filamentous callus and com-pact callus. Both callus types were induced at the cut surface ofthe explants and derived from both cortical and medullarycells. Filamentous callus was observed in the explant surfaceexposed to the air and compact callus formed on the explantsurface immersed in solid culture medium. The growth of thefilamentous callus was in the form of a uniseriate branchinitially, followed by formation of disorganized cell mass thatwent on to form callus clumps with prolific filamentous
Table 3 Effect of combination of nutrient supplementation (K2PO4, K2SO4, NH4Cl and urea) and plant growth regulators (2,4-D and kinetin) on theformation of adventitious branches in G. changii
Nutrientsupplementations
Percentage of explants with adventitious branch formation (%)
No PGR 0.01 mg L−1
2,4-D+1 mg L−1 kinetin0.1 mg L−1
2,4-D+0.1 mg L−1 kinetin10 mgL−1
2,4-D+0.001 mg L−1 kinetin
K2PO4 10 mM L−1 33.33±33.33 23.81±23.81 0.00±0.00 0.00±0.00
100 mM L−1 0.00±0.00 28.57±28.57 0.00±0.00 0.00±0.00
200 mM L−1 0.00±0.00 14.29±14.29 0.00±0.00 0.00±0.00
K2SO4 10 mM L−1 0.00±0.00 28.57±16.50 23.81±23.81 47.62±26.51
100 mM L−1 0.00±0.00 9.52±4.76 4.76±4.76 33.33±20.76
200 mM L−1 23.81±23.81 19.05±19.05 4.76±4.76 0.00±0.00
NH4Cl 10 mM L−1 23.81±23.81 19.05±9.52 0.00±0.00 14.29±14.29
100 mM L−1 23.81±23.81 33.33±33.33 0.00±0.00 9.52±9.52
200 mM L−1 0.00±0.00 33.33±33.33 42.86±29.74 28.57±28.57
Urea 10 mM L−1 19.05±19.05 4.76±4.76 4.76±4.76 28.57±21.87
100 mM L−1 0.00±0.00 38.10±19.05 0.00±0.00 0.00±0.00
200 mM L−1 0.00±0.00 0.00±0.00 0.00±0.00 28.57±21.87
Data are shown as mean value±standard error (experimental period=56 days; no. of replicates=3)
pH6.9 pH7.2 pH7.5 pH7.8 pH8.1
Fig. 8 Effect of different concentrations of plant growth regulators (2,4-D and kinetin) and pH on the formation of adventitious branches in G.changii explants under different irradiances (34.18 and 18.85 μmol
photons m−2 s−1). Data are shown as mean value±standard error (exper-imental period=56 days; no. of replicates=3)
J Appl Phycol (2014) 26:729–746 739
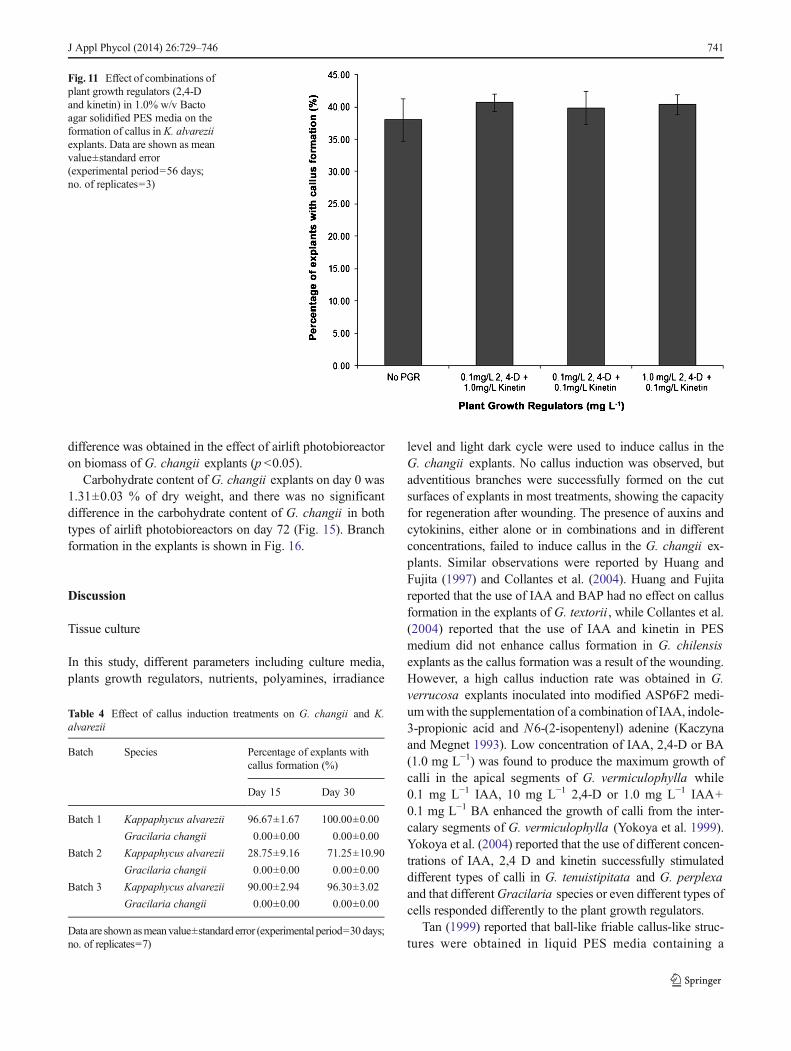
outgrowths. After 120 days, adventitious shoots were obtainedfrom the callus clumps. Regeneration and elongation of shootoccurred and bifurcate apical shoot formation was observedafter 6 months. The observations are shown in Figs. 12 and 13.However, none of the G. changii explants produced calluseven after day 56. The experiment was repeated three timeswith different batches of seaweeds, and similar results wereobtained. A significant difference on the callus formation wasobtained between the two species tested, but not among thethree batches of experiments (p <0.05).
Effect of airlift photobioreactor on explant culture of G.changii
Branch formations were observed in all treatments in thephotobioreactors. After 72 days, explant biomass increasedin all treatments except in 500mL PESmediumwithout 2,4-Dand kinetin in the tubular airlift photobioreactor and 3,000 mLPES medium without 2,4-D and kinetin in the spherical airliftphotobioreactor (Fig. 14). The decrease in biomass was due tobleaching and death of some explants. No significant
No PGR No plant growth regulator
PCR A 0.1mg L-1 2, 4-D + 0.1 mg L-1 kinetin
PGR A & Spermine 0.1mg L-1 2, 4-D + 0.1 mg L-1 kinetin + 0.0018 mg L-1 Spermine
PGRA & Zeatin 0.1mg L-1 2, 4-D + 0.1 mg L-1 kinetin + 1 mg L-1 Zeatin
PCR B 1mg L-1 NAA + 1 mg L-1 BAP
PGR B & Spermine 1mg L-1 NAA + 1 mg L-1 BAP + 0.0018 mg L-1 Spermine
PGR B & Zeatin 1mg L-1 NAA + 1 mg L-1 BAP +1 mg L-1 Zeatin
No PGR No plant growth regulator
PCR A 0.1mg L-1 2, 4-D + 0.1 mg L-1 kinetin
PGR A & Spermine 0.1mg L-1 2, 4-D + 0.1 mg L-1 kinetin + 0.0018 mg L-1 Spermine
PGRA & Zeatin 0.1mg L-1 2, 4-D + 0.1 mg L-1 kinetin + 1 mg L-1 Zeatin
PCR B 1mg L-1 NAA + 1 mg L-1 BAP
PGR B & Spermine 1mg L-1 NAA + 1 mg L-1 BAP + 0.0018 mg L-1 Spermine
PGR B & Zeatin 1mg L-1 NAA + 1 mg L-1 BAP +1 mg L-1 Zeatin
Fig. 9 Effect of combination ofplant growth regulators (2,4-D,kinetin, NAA, BAP and zeatin)and polyamines (spermine) on theformation of adventitiousbranches inG. changii explants inliquid PES media (diagonallystriped bar) or 1.0 % agarsolidified PES media (solid graybar) under different irradiances(34.18 and 18.85 μmol photonsm−2 s−1). Data are shown as meanvalue±standard error(experimental period=56 days;no. of replicates=10)
Fig. 10 Effect of differentconcentrations of plant growthregulator (2,4-D) in 1.0 % w/vBacto agar solidified PES mediaon the formation of callus in K.alvarezii explants. Data areshown as mean value±standarderror (experimental period=56 days; no. of replicates=3)
740 J Appl Phycol (2014) 26:729–746
difference was obtained in the effect of airlift photobioreactoron biomass of G. changii explants (p <0.05).
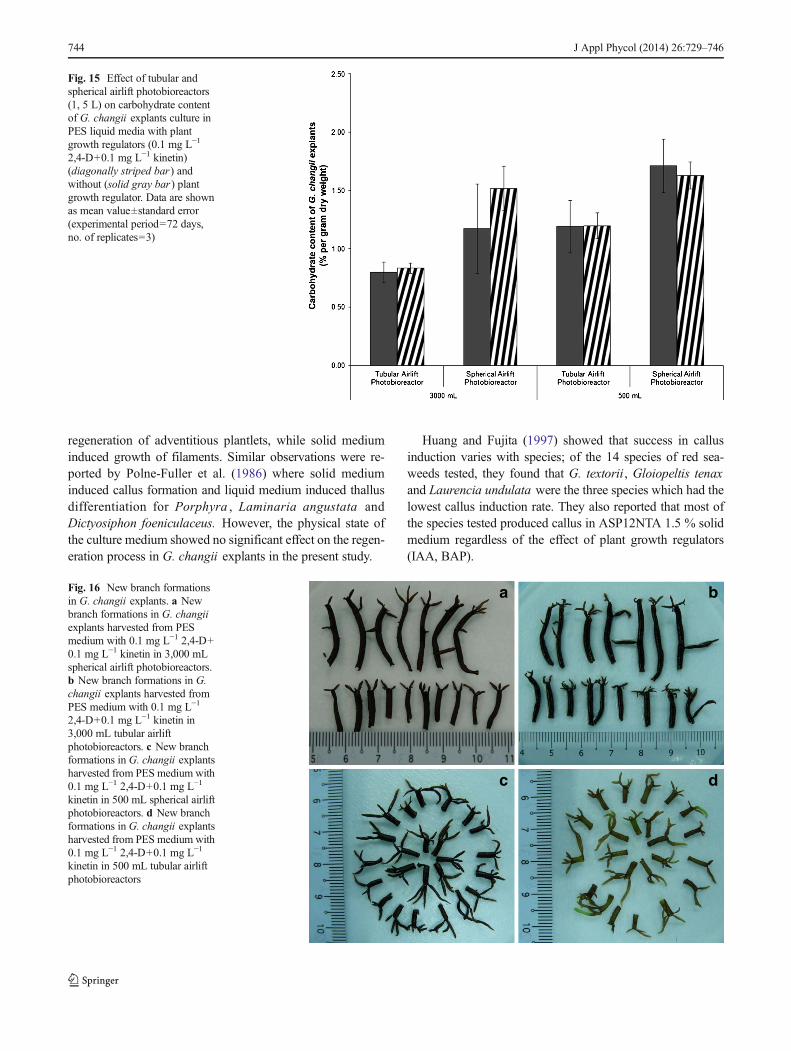
Carbohydrate content of G. changii explants on day 0 was1.31±0.03 % of dry weight, and there was no significantdifference in the carbohydrate content of G. changii in bothtypes of airlift photobioreactors on day 72 (Fig. 15). Branchformation in the explants is shown in Fig. 16.
Discussion
Tissue culture
In this study, different parameters including culture media,plants growth regulators, nutrients, polyamines, irradiance
level and light dark cycle were used to induce callus in theG. changii explants. No callus induction was observed, butadventitious branches were successfully formed on the cutsurfaces of explants in most treatments, showing the capacityfor regeneration after wounding. The presence of auxins andcytokinins, either alone or in combinations and in differentconcentrations, failed to induce callus in the G. changii ex-plants. Similar observations were reported by Huang andFujita (1997) and Collantes et al. (2004). Huang and Fujitareported that the use of IAA and BAP had no effect on callusformation in the explants of G. textorii , while Collantes et al.(2004) reported that the use of IAA and kinetin in PESmedium did not enhance callus formation in G. chilensisexplants as the callus formation was a result of the wounding.However, a high callus induction rate was obtained in G.verrucosa explants inoculated into modified ASP6F2 medi-umwith the supplementation of a combination of IAA, indole-3-propionic acid and N6-(2-isopentenyl) adenine (Kaczynaand Megnet 1993). Low concentration of IAA, 2,4-D or BA(1.0 mg L−1) was found to produce the maximum growth ofcalli in the apical segments of G. vermiculophylla while0.1 mg L−1 IAA, 10 mg L−1 2,4-D or 1.0 mg L−1 IAA+0.1 mg L−1 BA enhanced the growth of calli from the inter-calary segments of G. vermiculophylla (Yokoya et al. 1999).Yokoya et al. (2004) reported that the use of different concen-trations of IAA, 2,4 D and kinetin successfully stimulateddifferent types of calli in G. tenuistipitata and G. perplexaand that differentGracilaria species or even different types ofcells responded differently to the plant growth regulators.
Tan (1999) reported that ball-like friable callus-like struc-tures were obtained in liquid PES media containing a
Fig. 11 Effect of combinations ofplant growth regulators (2,4-Dand kinetin) in 1.0% w/v Bactoagar solidified PES media on theformation of callus in K. alvareziiexplants. Data are shown as meanvalue±standard error(experimental period=56 days;no. of replicates=3)
Table 4 Effect of callus induction treatments on G. changii and K.alvarezii
Batch Species Percentage of explants withcallus formation (%)
Day 15 Day 30
Batch 1 Kappaphycus alvarezii 96.67±1.67 100.00±0.00
Gracilaria changii 0.00±0.00 0.00±0.00
Batch 2 Kappaphycus alvarezii 28.75±9.16 71.25±10.90
Gracilaria changii 0.00±0.00 0.00±0.00
Batch 3 Kappaphycus alvarezii 90.00±2.94 96.30±3.02
Gracilaria changii 0.00±0.00 0.00±0.00
Data are shownasmeanvalue±standard error (experimental period=30days;no. of replicates=7)
J Appl Phycol (2014) 26:729–746 741

combination of 2,4-D and kinetin. Similar treatments werecarried out in the present study, but no callus formation wasobserved in G. changii explants. This may be because not allparts of seaweed thalli respond similarly to the given plantgrowth regulators as reported by Yokoya et al. (2004). Yokoyaet al. (2004) concluded that both auxins (IAA and 2,4-D) andcytokinin (kinetin) play a regulatory role in the growth andmorphogenetic processes of G. tenuistipitata and G.perplexa , and the differential sensitivity of the explants wasdue to the seaweed-specific development system. The differ-ential sensitivity of explants to plant growth regulators mightbe due to a regulatory mechanism rather than the concentra-tion of plant growth regulators (Trewavas 1982). Baweja et al.(2009) pointed out the lack of attention on the factor of cellularcompetence to plant growth regulators in seaweed tissueculture studies and that more fundamental research shouldbe done to answer the question.
However, the use of combinations of auxin and cytokininsuccessfully produced higher number of explants with adven-titious branches or shoots inG. changii compared to the use of
single hormones or their omission from the medium. Theseadventitious shoots originated from the cortical zone of the cutsurface of the explants, directly without an intervening callusphase, and were followed by shoot elongation and bifurcationat the apices of the shoots. Titlyanov et al. (2006) reported thatyoung fronds developed directly from the cut edges of themeristematic fragments in the superficial cortical layers ofPalmaria palmata thallus.
In higher plants, shoot formation can be executed directlyor indirectly with an intervening callus phase in response tothe extra-cellular signals (Hicks 1994; George et al. 2008;Elhiti and Stasolla 2012), which may be described as a com-petent phenomenon which has been indentified in three dis-tinct phases: competence acquisition, canalization and mor-phogenesis. Elhiti and Stasolla (2012) explained that in thefirst phase, cells within the explant acquire competence torespond to inductive signals, then “canalised” into a shootdevelopment program which results in “morphogenesis”.These responses may be linked to signals from the presenceof auxins and cytokinins (plant growth regulators) and also the
a b
c d
e f
Fig. 12 Callus induction andformation of K. alvarezii . aFilamentous callus was observedon day 10 in the K. alvareziiexplants cultured in agarsolidified PES medium. bFilamentous callus was observedon day 15 in the K. alvareziiexplants cultured in agarsolidified PES medium. cFilamentous callus was observedon day 30 in the K. alvareziiexplants cultured in agarsolidified PES medium. dFilamentous callus was observedon day 45 in the K. alvareziiexplants cultured in agarsolidified PES medium. eFilamentous callus was observedin the K. alvarezii explantscultured in agar solidified PESmedium. f Compact callus wasobserved in the K. alvareziiexplants cultured in agarsolidified PES medium
742 J Appl Phycol (2014) 26:729–746
relationship between the level of endogenous plant hormonesand organogenesis, a subject which has yet to be furtherunderstood in seaweeds (Kim et al. 2000; Reddy et al. 2008;
Baweja et al. 2009). Yokoya and Handro (2002) reported thatthe physical state of the culture medium affected the regener-ation process in Solieria filiformis : Liquid medium induced
a b
c d
e f
Fig. 13 Callus formation andregeneration of K. alvarezii . a , bFilamentous callus observed inthe explant surface exposed to theair while the compact callusformed on the explant surfaceimmersed in solid culturemedium. c Callus were induced atthe cut surface of both end of theexplants and derived from bothcortical and medullary cells. dCallus followed by formation ofdisorganized cell mass andformed callus clumps withprolific filamentous outgrowths. eRegeneration of callus of K.alvarezii . f Regeneration andelongation of shoot from callus ofK. alvarezii
Fig. 14 Effect of PES liquidmedia with and without plantgrowth regulators (0.1 mg L−1
2,4-D+0.1 mg L−1 kinetin) on thebiomass of G. changii explant intubular and spherical airliftphotobioreactors on day 0(diagonally striped bar) andday 72 (solid gray bar) of culture.Data are shown as mean value±standard error (experimentalperiod=72 days; no. ofreplicates=3)
J Appl Phycol (2014) 26:729–746 743
regeneration of adventitious plantlets, while solid mediuminduced growth of filaments. Similar observations were re-ported by Polne-Fuller et al. (1986) where solid mediuminduced callus formation and liquid medium induced thallusdifferentiation for Porphyra , Laminaria angustata andDictyosiphon foeniculaceus. However, the physical state ofthe culture medium showed no significant effect on the regen-eration process in G. changii explants in the present study.
Huang and Fujita (1997) showed that success in callusinduction varies with species; of the 14 species of red sea-weeds tested, they found that G. textorii , Gloiopeltis tenaxand Laurencia undulata were the three species which had thelowest callus induction rate. They also reported that most ofthe species tested produced callus in ASP12NTA 1.5 % solidmedium regardless of the effect of plant growth regulators(IAA, BAP).
Fig. 15 Effect of tubular andspherical airlift photobioreactors(1, 5 L) on carbohydrate contentof G. changii explants culture inPES liquid media with plantgrowth regulators (0.1 mg L−1
2,4-D+0.1 mg L−1 kinetin)(diagonally striped bar) andwithout (solid gray bar) plantgrowth regulator. Data are shownas mean value±standard error(experimental period=72 days,no. of replicates=3)
a b
c d
Fig. 16 New branch formationsin G. changii explants. a Newbranch formations in G. changiiexplants harvested from PESmedium with 0.1 mg L−1 2,4-D+0.1 mg L−1 kinetin in 3,000 mLspherical airlift photobioreactors.b New branch formations in G.changii explants harvested fromPES medium with 0.1 mg L−1
2,4-D+0.1 mg L−1 kinetin in3,000 mL tubular airliftphotobioreactors. c New branchformations in G. changii explantsharvested from PES medium with0.1 mg L−1 2,4-D+0.1 mg L−1
kinetin in 500 mL spherical airliftphotobioreactors. d New branchformations in G. changii explantsharvested from PES medium with0.1 mg L−1 2,4-D+0.1 mg L−1
kinetin in 500 mL tubular airliftphotobioreactors
744 J Appl Phycol (2014) 26:729–746

When the same protocols were repeated with G. changiiand K. alvarezii , it was found that, although callus was notinduced in G. changii , more than 90 % of K. alvarezii ex-plants produced callus after 30 days culture. The success ofcallus induction in K. alvarezii explants indicated that therewas probably no significant problem in the callus inductionprotocol, the personal skill and techniques as well as theculture (incubation) environment. An important factor in sea-weed callus formation is its response to injury or wounding(Kumar et al. 2004, 2007). Callus formation in G. verrucosa(Kaczyna and Megnet 1993) and G. corticata (Kumar et al.2007) has been reported as a result of wound responses. In thepresent study, callus formation was found in 68.57±32.37 %of K. alvarezii explants cultured in 1.0 % w/v Bacto agarsolidified PES medium for 30 days without any plant growthregulator indicating that the wounding effect may be respon-sible for inducing callus formation in K. alvarezii explants.However, the wounding effect (cutting) at both ends of the G.changii explants did not induce any callus formation in G.changii explants but produced adventitious shoots instead.
The failure to induce callus inG. changii may be due to themicroenvironment of its original habitat.G. changii inhabitedthe mangroves while the Kappaphycus was farmed usingvegetative propagation on long lines. The Gracilaria grewepiphytically on the roots of mangrove trees, and propagationwas probably through spores produced during the triphasiclife cycle. Perhaps microflora is present in the mangrove soilwhich produces micronutrients—essential for the growth ofthe seaweed, especially under artificial conditions. Mangrovesoils are sulphur rich as well, and the use of sulphate inthe culture medium could be an interesting study. Thepresent study can be used as guideline for further stud-ies. Kappaphycus , on the other hand, has been cultivat-ed artificially in Sabah for several decades throughcuttings and has probably developed the capacity fortotipotency.
In conclusion, callus formation and regeneration was suc-cessfully obtained for K. alvarezii explants. No callus forma-tion was observed in G. changii explants; instead, prolifera-tion of adventitious shoots was successfully obtained. The useof airlift photobioreactors not only increased the biomass ofG.changii explants but also showed that there were no signifi-cant changes in the carbohydrate content of the explantscompared to the original plants. These findings will contributeto successful production of clonal planting materials to sup-port the expanding seaweed farming in Malaysia.
Acknowledgments This research was funded by the Ministry of Sci-ence, Technology and Innovation, Malaysia (E-Science Fund: 02-01-03-SF0148 and 04-01-03-SF0664; Top-down Project: 06-02-02-003 BTK/ER/016, Grant No: 39-02-03-9002/Oracle 8301902), the MalaysianToray Science Foundation (Japan) Grant (MTSF 1050-2009A) and theUniversity of Malaya Research Grants (UMRG Grant No: RG100-10SUS, F0233/2004D, P0114/2006A).
References
Armisen R (1995) World-wide use and importance of Gracilaria. J ApplPhycol 7:231–243
Atkinson MJ, Bingman C (1997) Elemental composition of commercialseasalts. J Aquaric Aquat Sci 8:39–43
Baweja P, Sahoo D, García-Jiménez P, Robaina RR (2009) Seaweedtissue culture as applied to biotechnology: problems, achievementsand prospects. Phycol Res 57:45–58
Bixler HJ, Porse H (2011) A decade of change in the seaweed hydrocol-loids industry. J Appl Phycol 23:321–335
Chan CX, Teo SS, Ho CL, Rofima Yasmin O, Phang SM (2004)Optimisation of RNA extraction for Gracilaria changii(Gracilariales, Rhodophyta). J Appl Phycol 16:297–301
Chen LCM, Taylor RA (1978) Medullary tissue culture of the red algaChondrus crispus. Can J Bot 56:883–886
Chu WL, Norazmi M, Phang SM (2003) Fatty acid composition of someMalaysian seaweeds. Malays J Sci 22:21–27
Collantes G, Melo C, Candia A (2004) Micropropagation by explants ofGracilaria chilensis Bird, McLachlan and Oliveira Gloria. J ApplPhycol 16:203–213
Dawes CJ, Koch EW (1991) Branch, micropropagule and tissue cultureof the red algae, Eucheuma diculatum and Kappaphycus alvareziifarmed in the Philippines. J Appl Phycol 3:247–257
Dawes CJ, Trono GC Jr, Lluisma AO (1993) Clonal propagation ofEuchuema denticulatum and Kappaphycus alvarezii for Philippineseaweed farms. Hydrobiologia 260/261:379–383
Dawes CJ, Lluisma AO, Trono GC (1994) Laboratory and field growthstudies of commercial strains of Eucheuma denticulatum andKappaphycus alvarezii in the Philippines. J Appl Phycol 6:21–24
Elhiti M, Stasolla C (2012) In vitro shoot organogenesis and hormoneresponse are affected by the altered level of Brassica napus meri-stem genes. Plant Sci 190:40–51
Food and Agriculture Organization of the United Nations (FAO) (2007)FAO year book, vol. 100/2. FAO, Rome
Food and Agriculture Organization of the United Nations (FAO) (2012)World review of fisheries and aquaculture. In: The State of WorldFisheries and Aquaculture 2012. FAO, Rome, pp 3–100
Gan SY, Qin S, Rofina Yasmin YD, Phang SM (2003) Transient expres-sion of lacZ in particle bombarded Gracilaria changii(Gracilariales, Rhodophyta). J Appl Phycol 15:315–353
Gan SY, Rofina Yasmin O, Qin S, Phang SM (2006) Crop improvementin seaweed cultivation. In: Phang SM, Critchley A, Ang P (eds)Advances in seaweed cultivation and utilization in Asia.University of Malaya Maritime Research Center, Kuala Lumpur,pp 81–103
George EF, Hall MA, Klerk GD (2008) Plant propagation by tissueculture, volume 1. The background. Springer, Netherlands
Guillard RR (1975) Culture of phytoplankton for feeding marine inver-tebrates. In: Smith WL, Chanley MH (eds) Culture of marineinvertebrate animals. Plenum, New York, pp 26–60
Gusev MV, Tambiev AH, Kirikova NN, Shelyastina NN, Aslanyan RR(1987) Callus formation in seven species of agarophyte marinealgae. Mar Biol 95:593–597
Hicks G (1994) Shoot induction and organogenesis in vitro: a develop-ment perspective. In Vitro Cell Dev 30:10–15
Ho CL, Teoh S, Teo SS, Rahim RA, Phang SM (2009) Profiling thetranscriptome of Gracilaria changii (Rhodophyta) in response tolight deprivation. Mar Biotechnol 11:513–519
Huang W, Fujita Y (1997) Callus induction and thallus regeneration insome species of red algae. Phycol Res 45:105–111
Hurtado AQ, Biter AB (2007) Plantlet regeneration of Kappaphycusalvarezii var. adik-adik by tissue culture. J Appl Phycol 19:783–786
James DE (1978) Culturing algae. Carolina Biological Supply,Burlington
J Appl Phycol (2014) 26:729–746 745

Kaczyna F, Megnet WH (1993) The effects of glycerol and plant growthregulators on Gracilaria verrucosa (Gigartinales, Rhodophyceae).Hydrobiologia 268:57–64
Kim JW, Han SK, Kwon SY, Lee HS, Lim YP, Liu JR, Kwak SS (2000)High frequency shoot induction and plant regeneration from coty-ledonary hypocotyl explants of cucumber (Cucumis sativus L.)seedlings. J Plant Physiol 157:136–139
Kochert AG (1978) Carbohydrate determination by the phenol–sulphuricacid method. In: Hellbust JA, Craigie JS (eds) Handbook of phyco-logical methods: physiological and biochemical methods.Cambridge University Press, Cambridge, pp 95–97
Kumar GR, Reddy CRK, Ganesan M, Thiruppathi S, Dipakkore S,Eswaran K, Rao PYS, Jha B (2004) Tissue culture and regenerationof thallus from callus ofGelidiella acerosa (Gelidiaies, Rhodophyta).Phycologia 43:596–602
Kumar GR, Reddy CRK, Jha B (2007) Callus induction and thallusregeneration from callus of phycocolloid yielding seaweeds fromthe Indian coast. J Appl Phycol 19:15–25
Lim PE, Phang SM (2004)Gracilaria species (Gracilariales, Rhodophyta)of Malaysia including two new records. Malays J Sci 23:71–80
Marinho-Soriano E, Silva TSF, Moreira WSC (2001) Seasonal variationin the biomass and agar yield from Gracilaria cervicornis andHydropuntia cornea from Brazil. Bioresour Technol 77:115–120
McHugh DJ (2003) A guide to the seaweed industry. FAO fisheriestechnical paper no 441. FAO, Rome, p 105
Phang SM (2006) Seaweed resources in Malaysia: current status andfuture prospects. Aquat Ecosyst Health Manag 9:185–202
Phang SM, Shaharuddin S, Noraishah H, Sasekumar A (1996) Studies onGracilaria changii (Gracilariales, Rhodophyta) from Malaysianmangroves. Hydrobiologia 326/327:347–352
Phang SM, Lim PE, Yeong HY (2010) Malaysian seaweed resources inthe South China Sea and their potential economic and ecologicalapplications. J Sci Technol Trop 6:87–109
Polne-Fuller M, Saga N, Gibor A (1986) Algal cell, callus and tissuecultures and selection of algal strains. In: Barclay WR (ed) Algalbiomass technologies: an interdisciplinary perspective. Cramer,Berlin, pp 30–36
Reddy CRK, Kumar GRK, Siddhanta AK, Tewari A, Eswaran K (2003)In vitro somatic embryogenesis and regeneration of somatic embry-os from pigmented callus of Kappaphycus alvarezii (Doty) Doty(Rhodophyta, Gigartinales). J Appl Phycol 39:610–616
Reddy CRK, Jha B, Fujita Y, Ohno M (2008) Seaweed micropropagationtechniques and their potentials: an overview. JAppl Phycol 20:609–617
Schramm W (1991) Cultivation of unattached seaweeds. In: Guiry MD,Blunden G (eds) Seaweed resources in Europe, uses and potential.Wiley, New York, pp 379–408
Shu MH, Appleton D, Zandi K, AbuBakar S (2013) Anti-inflammatory,gastroprotective and antiulcerogenic effects of red algae Gracilariachangii (Gracilariales, Rhodophyta) extract. BMC ComplAlternative Med 13:61
Sim MC, Lim PE, Gan SY, Phang SM (2007) Identification of randomamplified polymorphic DNA (RAPD) markers for differentiatingmale from female and sporophyte thalli of Gracilaria changii(Rhodophyta). J Appl Phycol 19:763–769
Siow RS, Teo SS, Ho WY, Mohd. Yunus Abd. S, Phang SM, Ho CL(2012) Molecular cloning and biochemical characterization ofgalactose-1-phosphate uridylyltransferase from Gracilaria changii(Rhodophyta). J Phycol 48:155–162
Siow RS, Teoh S, Teo SS, Abd. Shukor MY, Phang SM, Ho CL (2013)Molecular cloning and characterization of GDP-mannose-3′,5′-
epimerase from Gracilaria changii . J Appl Phycol. doi:10.1007/s10811-013-9987-5
Song SL, Lim PE, Phang SM, Lee WW, Lewmanomont K, Largo DB,Abdul Han N (2013) Microsatellite markers from expressed se-quence tags (ESTs) of seaweeds in differentiating variousGracilaria species. J Appl Phycol 25:839–846
Tan EL (1999) Tissue culture and protoplast isolation of Gracilariachangii (Rhodophyta). Master’s thesis, University of Malaya,Kuala Lumpur
Teo SS, Ho CL, Teoh S, Lee WW, Tee JM, Abdul Rahim R, Phang SM(2007) Analyses of expressed sequence tags (ESTs) from anagarophyte Gracilaria changii (Gracilariales, Rhodophyta). Eur JPhycol 42:41–46
Teo SS, Ho CL, Teoh S, Rahim RA, Phang SM (2009) Transcriptomicanalysis of Gracilaria changii (Rhodophyta) in response to hyper-and hypo-osmotic stresses. J Phycol 45:1093–1099
Titlyanov EA, Titlyanova TV, Kadel P, Luning K (2006) Newmethods of obtaining plantlets and tetraspores from fragmentsand cell aggregates of meristematic and submeristematic tissueof the red alga Palmaria palmata . J Exp Mar Biol Ecol 339:55–64
Trewavas AJ (1982) Growth substance sensitivity: the limiting factor inplant development. Physiol Plant 55:60–72
Wong WH, Goh SH, Phang SM (1994) Antibacterial properties ofMalaysian seaweeds. In: Phang SM et al (eds) Algal biotechnologyin the Asia-Pacific region. University of Malaya, Kuala Lumpur, pp75–81
Wong PF, Tan LJ, Nawi H, Abu Bakar S (2006) Proteomics of the redalga, Gracilaria changii (Gracilariales, Rhodophyta). J Phycol 42:113–120
Yeong HY, Khalid N, Phang SM (2008) Protoplast isolation and regen-eration fromGracilaria changii (Gracilariales, Rhodophyta). J ApplPhycol 20:641–651
Yokoya NS (2000) Apical callus formation and plant regeneration con-trolled by plant growth regulators on axenic culture of red algaGracilariopsis tenuifrons (Gracilariales, Rhodophyta). Phycol Res48:133–142
Yokoya NS, Handro W (2002) Effects of plant growth regulators andculture medium on morphogenesis of Solieria filiformis(Rhodophyta) cultured in vitro . J Appl Phycol 14:97–102
Yokoya NS, Kakita H, Obika H, Kitamura T (1999) Effects of environ-mental factors and plant growth regulators on growth of the red algaGracilaria vermiculophylla from Shikoku Island, Japan.Hydrobiologia 398/399:339–347
Yokoya NS,West JA, Luchi AE (2004) Effects of plant growth regulatorson callus formation, growth and regeneration in axenic tissue cul-tures of Gracilaria tenuistipitata and Gracilaria perplexa(Gracilariales, Rhodophyta). Phycol Res 52:244–254
Yow YY, Lim PE, Phang SM (2011) Genetic diversity of Gracilariachangii (Gracilariaceae, Rhodophyta) from west coast, PeninsularMalaysia based on mitochondrial cox1 gene analysis. J Appl Phycol23:219–226
Yow YY, Lim PE, Phang SM (2013) Assessing the use of mitochondrialcox1 gene and cox2-3 spacer for genetic diversity study ofMalaysian Gracilaria changii (Gracilariaceae, Rhodophyta) fromPeninsular Malaysia. J Appl Phycol 25:831–838
Yunque DAT, Tibubos KR, Hurtado AQ, Critchley AT (2011)Optimization of culture conditions for tissue culture production ofyoung plantlets of carrageenophyte Kappaphycus . J Appl Phycol23:615–621
746 J Appl Phycol (2014) 26:729–746


















