
PPARγ Regulates Mouse Meibocyte Differentiation and Lipid ...
11
ALI DJALILIAN, MD, EDITOR PPARg Regulates Mouse Meibocyte Differentiation and Lipid Synthesis JAMES V. JESTER,PHD, 1 ERIC POTMA,PHD, 2 AND DONALD J. BROWN,PHD 1 ABSTRACT Purpose: Previous reports suggest that age- related meibomian gland atrophy is associated with decreased expression of the lipid-sensitive nuclear receptor, PPARg. The purpose of this study was to identify the role of PPARg in modulating meibocyte lipid synthesis. Methods: Cytoplasmic and nuclear fractions from meibomian glands of young (2M) and old (2Y) C57Bl6 mice were probed using antibodies specific for PPARg. Mouse meibocytes were cultured, immortalized using a SV40 lentiviral vector, and evaluated for lipid synthesis using LipidTox staining and CARS/Raman microspectroscopy. Lipid synthesizing clones were tested for effects of PPARg agonist, rosiglitazone, on lipid synthesis and PPARg localization, post-translational modification and induction of PPARg response genes. Re- sults: The cytoplasmic fraction in young mice contained both 50 and 72 kDa PPARg bands that were absent or reduced by 75% in older mice, respectively. Cultured mei- bocytes produced neutral lipid containing equal amounts of wax and cholesterol esters, similar to mouse meibum. Addition of rosiglitazone (10-50 mM) significantly increased lipid production (P<.05) in meibocytes, associated with SUMO1 sumoylation and cytoplasmic accumulation of the 72 kDa PPARg. Rosiglitazone also increased the localization of PPARg to the cytoplasm and up-regulated of PPARg, ADP and ADFP mRNA. Conclusions: This study confirms the loss of cytoplasmic/vesicular PPARg localization in older, atrophic mouse meibomian glands. Furthermore, PPARg stimulates lipid synthesis in mouse meibocytes, associated with PPARg sumoylation and translocation to the cytoplasm. Taken together these data suggest that lipid synthesis in older mice is down regulated by a PPARg mediated pathway. KEY WORDS eyelid, lipid, meibocytes, meibomian gland, PPARg, tear film I. INTRODUCTION M eibomian glands are lipid-excreting, holocrine glands that are embedded in the tarsal plate of the mammalian eyelid and provide lipid to the tear film. 1,2 Loss of tear film lipid is known to cause tear film instability, increase tear evaporation, and increase tear film osmolarity, causing an evaporative form of dry eye. 3-6 Evaporative dry eye is generally thought to be an age- related disorder that is a common clinical complaint of pa- tients visiting the ophthalmologist or optometrist and may comprise from 37% to 47% of the average patient popula- tion. 7 Recently, there has been increasing awareness of the association between evaporative dry eye and meibomian gland dysfunction (MGD) in the form of meibomian gland dropout and altered lipid quantity and/or quality. 8-10 While the risk of evaporative dry eye and MGD increases with age, much remains to be clarified regarding the under- lying cellular and molecular mechanisms. 11 Past clinical and animal studies have established that meibomian glands are susceptible to keratinization and that genetic alterations and chemical agents, i.e., chlorinate biphenyls and epinephrine, can lead to replacement of the meibomian gland with keratic cysts. 12-14 These observations have formed the basis for an obstructive model of MGD where hyperkeratinization of the meibomian gland duct leading to ductal occlusion and plugging is thought to cause cystic dilation and a ‘disuse at- rophy’ of the gland. 6 Androgens have also been implicated in the development of evaporative dry eye, based on findings that neutral and polar lipid profiles of meibomian glands show distinct differences related to sex and the presence of functional androgen receptors. 15,16 While keratinization and androgens are clearly important to gland function, the overall Accepted for publication August 2016. From 1 Gavin Herbert Eye Institute, University of California, Irvine, and 2 Department of Chemistry and Beckman Laser Institute, University of California, Irvine, Irvine, CA. Supported by NEI EY021510, The Skirball Program in Molecular Ophthalmology and Research to Prevent Blindness, Inc., Unrestricted Grant. The authors have no commercial or proprietary interest in any concept or product discussed in this article. Single-copy reprint requests to James V. Jester, PhD (address below). Corresponding author: James V. Jester, PhD, Gavin Herbert Eye Institute, University of California Irvine, Hewitt Hall, 843 Health Sciences Road, Building 843, 2nd Floor, Irvine, CA 92697. Tel: 949-824-8047. E-mail address: [email protected] Ó 2016 Elsevier Inc. All rights reserved. The Ocular Surface ISSN: 1542- 0124. Jester JV, Potma E, Brown DJ. Pparg regulates mouse meibocyte differentiation and lipid synthesis. 2016;14(4):484-494. Original Research 484 THE OCULAR SURFACE / OCTOBER 2016, VOL. 14 NO. 4 / www.theocularsurface.com
Transcript of PPARγ Regulates Mouse Meibocyte Differentiation and Lipid ...

Original Research
ALI DJALILIAN, MD, EDITORPPARg Regulates Mouse MeibocyteDifferentiation and Lipid Synthesis
JAMES V. JESTER, PHD, 1 ERIC POTMA, PHD, 2 AND DONALD J. BROWN, PHD1
ABSTRACT Purpose: Previous reports suggest that age-related meibomian gland atrophy is associated withdecreased expression of the lipid-sensitive nuclear receptor,PPARg. The purpose of this study was to identify the role ofPPARg in modulating meibocyte lipid synthesis. Methods:Cytoplasmic and nuclear fractions from meibomian glandsof young (2M) and old (2Y) C57Bl6 mice were probed usingantibodies specific for PPARg. Mouse meibocytes werecultured, immortalized using a SV40 lentiviral vector, andevaluated for lipid synthesis using LipidTox staining andCARS/Raman microspectroscopy. Lipid synthesizing cloneswere tested for effects of PPARg agonist, rosiglitazone, onlipid synthesis and PPARg localization, post-translationalmodification and induction of PPARg response genes. Re-sults: The cytoplasmic fraction in young mice containedboth 50 and 72 kDa PPARg bands that were absent orreduced by 75% in older mice, respectively. Cultured mei-bocytes produced neutral lipid containing equal amounts ofwax and cholesterol esters, similar to mouse meibum.Addition of rosiglitazone (10-50 mM) significantly increasedlipid production (P<.05) in meibocytes, associated withSUMO1 sumoylation and cytoplasmic accumulation of the72 kDa PPARg. Rosiglitazone also increased the localizationof PPARg to the cytoplasm and up-regulated of PPARg, ADP
Accepted for publication August 2016.
From 1Gavin Herbert Eye Institute, University of California, Irvine, and2Department of Chemistry and Beckman Laser Institute, University ofCalifornia, Irvine, Irvine, CA.
Supported by NEI EY021510, The Skirball Program in MolecularOphthalmology and Research to Prevent Blindness, Inc., UnrestrictedGrant.
The authors have no commercial or proprietary interest in any concept orproduct discussed in this article.
Single-copy reprint requests to James V. Jester, PhD (address below).
Corresponding author: James V. Jester, PhD, Gavin Herbert Eye Institute,University of California Irvine, Hewitt Hall, 843 Health Sciences Road,Building 843, 2nd Floor, Irvine, CA 92697. Tel: 949-824-8047. E-mailaddress: [email protected]
� 2016 Elsevier Inc. All rights reserved. The Ocular Surface ISSN: 1542-0124. Jester JV, Potma E, Brown DJ. Pparg regulates mouse meibocytedifferentiation and lipid synthesis. 2016;14(4):484-494.
484 THE OCULAR SURFACE / OCTOBER 2016
and ADFP mRNA. Conclusions: This study confirms the loss ofcytoplasmic/vesicular PPARg localization in older, atrophicmouse meibomian glands. Furthermore, PPARg stimulateslipid synthesis in mouse meibocytes, associated with PPARgsumoylation and translocation to the cytoplasm. Takentogether these data suggest that lipid synthesis in older miceis down regulated by a PPARg mediated pathway.
KEY WORDS eyelid, lipid, meibocytes, meibomian gland,PPARg, tear film
I. INTRODUCTION
M eibomian glands are lipid-excreting, holocrineglands that are embedded in the tarsal plate ofthe mammalian eyelid and provide lipid to the
tear film.1,2 Loss of tear film lipid is known to cause tearfilm instability, increase tear evaporation, and increase tearfilm osmolarity, causing an evaporative form of dry eye.3-6
Evaporative dry eye is generally thought to be an age-related disorder that is a common clinical complaint of pa-tients visiting the ophthalmologist or optometrist and maycomprise from 37% to 47% of the average patient popula-tion.7 Recently, there has been increasing awareness of theassociation between evaporative dry eye and meibomiangland dysfunction (MGD) in the form of meibomian glanddropout and altered lipid quantity and/or quality.8-10
While the risk of evaporative dry eye and MGD increaseswith age, much remains to be clarified regarding the under-lying cellular and molecular mechanisms.11 Past clinical andanimal studies have established that meibomian glands aresusceptible to keratinization and that genetic alterations andchemical agents, i.e., chlorinate biphenyls and epinephrine,can lead to replacement of the meibomian gland with keraticcysts.12-14 These observations have formed the basis for anobstructive model of MGD where hyperkeratinization ofthe meibomian gland duct leading to ductal occlusion andplugging is thought to cause cystic dilation and a ‘disuse at-rophy’ of the gland.6 Androgens have also been implicatedin the development of evaporative dry eye, based on findingsthat neutral and polar lipid profiles of meibomian glandsshow distinct differences related to sex and the presence offunctional androgen receptors.15,16 While keratinization andandrogens are clearly important to gland function, the overall
, VOL. 14 NO. 4 / www.theocularsurface.com

OUTLINE
I. Introduction
II. Materials and Methods
A. Animals
B. Isolation, Culture, Immortalization and Character-ization of Mouse Meibocytes
C. Immunocytochemistry
D. Assessment of Meibocyte Lipid Synthesis
E. Protein Isolation and Western Blotting
F. Immunoprecipitation
G. RNA Isolation and Analysis of Gene Expression
H. Statistical Analysis
III. Results
A. Cytoplasmic and Nuclear PPARg Expression
B. Characterization of Immortalized MouseMeibocytes
C. Lipid Synthesis and PPARg Localization in CulturedMeibocytes
D. Effects of Rosiglitazone on Lipid Synthesis andPPARg Localization
IV. Discussion
V. Conclusion
MEIBOCYTE PPARg / Jester, et al
regulation of meibomian gland differentiation and lipid syn-thesis remains unknown.
Recently, a telomerase immortalized human meibomiangland epithelial cell line has been developed that has beenused to evaluate the influence of androgens, growth factors,serum, neurotransmitters, and antibiotics onmeibomian glandepithelial proliferation and lipid synthesis.17-25 In these studies,gene expression analysis has shown that peroxisome prolifera-tive activated receptor (PPAR) signaling can be differentiallyregulated based on induction of proliferation or differentiation.PPARs are a family of lipid-sensitive nuclear receptors thatshow distinct tissue localizations and physiologic activities.One member, PPARg, has been shown to play an importantrole in regulating lipid synthesis and differentiation of adipo-cytes and sebocytes and is known to interact with other nuclearhormone receptors, including androgens and retinoic acid.26-28
While past studies have shown that PPARg is expressed indeveloping and adult mouse meibomian glands,29 the role ofPPARg signaling on meibocyte lipid synthesis is not known.
Interestingly, during aging of humans and mouse meibo-mian glands, PPARg undergoes a change in the immunocyto-chemical localization within meibocytes from a vesicular/cytoplasmic and nuclear localization in young glands to a pre-dominantly nuclear pattern in older glands. This change occursconcurrently with decreased basal acinar cell proliferation,gland atrophy and clinically identified meibomian glanddropout.30-32 Overall, these findings support the hypothesisthat aging of the meibomian gland involves altered meibocytedifferentiation and PPARg receptor signaling that is associatedwith meibomian gland dropout and altered lipid quality.
To test this hypothesis we have evaluated the subcellularlocalization of PPARg in meibomian glands of young and
THE OCULAR SURFACE / OCTOBER 2016, VO
old mice and cultured mouse meibocytes. In this paper wepresent data showing that the cytoplasm of meibocytescontain a 50/72 kDa PPARg that is decreased or absent inmeibomian glands from older mice. Furthermore, treatmentof cultured mouse meibocytes with the PPARg agonist, rosi-glitazone, induces lipid synthesis coincident with sumoyla-tion and increased localization of PPARg to the cytoplasmalong with up-regulation of PPARg response genes. Takentogether, these findings indicate that PPARg regulates lipidsynthesis in meibocytes and suggest that age-related atrophyof mouse meibomian glands involves downregulation ofPPARg-induced lipid synthesis.
II. MATERIALS AND METHODSA. Animals
C57Bl/6 mice were used in this study. All animal pro-cedures were approved by the University of California,Irvine, Institutional Animal Care and Use Committee andwere conducted in accordance with the ARVO Statementfor the Use of Animals in Ophthalmic and Vision Research.All animals were humanely sacrificed by cervical neck dislo-cation following sedation with ketamine (100 mg/Kg bodyweight) and xylazine (20 mg/Kg body weight) prior to tissuecollection.
B. Isolation, Culture, Immortalization and Character-ization of Mouse MeibocytesIsolation and Culture: The upper and lower eyelids of
twenty 6-week-old mice were removed, disinfected with70% ethanol and dissected using a Zeiss SteREO Discov-ery.V12 microscope (Carl Zeiss MicroImaging, LLC, UnitedStates). Tissues were first washed with phosphate buffer sa-line (PBS, pH 7.2) and then immediately placed in Dulbec-co’s modified Eagle’s medium (DMEM) supplemented withantibiotics (Invitrogen, Carlsbad, CA). The conjunctiva andexcess skin was removed and the tarsal plates carefullyexcised and placed in PBS containing 0.25% collagenase A(Invitrogen, Carlsbad, CA) and 0.6U/ml dispase II (Invitro-gen). Glands were then digested at 37�C overnight followedby centrifugation at 1,500 RPM for 15 minutes. The cell pel-let was then suspended in Keratinocyte Growth Medium(KGM, Lonza Walkersville, Inc., Walkersville, MD) contain-ing human epidermal growth factor (hEGF), insulin(bovine), Hydrocortisone and antibiotics (Gentamicin) sup-plemented with 10% fetal bovine serum. Cells were culturedon 6-well primaria culture plates (Becton Dickinson andCompany, Franklin Lakes, NJ) and maintained in a humid-ified 5% CO2 incubator at 37�C. In the reported studies, 2batches of mice were used to establish meibocyte culturesfor characterizing lipid synthesis and immortalization,respectively.
Immortalization of Meibocytes: Primary mouse meibo-cytes at 40-50% confluence were immortalized using aSV40 large T-antigen inserted in a lentiviral vector (AppliedBiological Materials Inc., Richmond, BC Canada). Briefly,viral supernatant containing 0.8 mg/ml of polybrene
L. 14 NO. 4 / www.theocularsurface.com 485

MEIBOCYTE PPARg / Jester, et al
transfection reagent (Millipore Corporation, Phillisburg, NJ)was added to meibocytes cultures. Cells were then culturedovernight and the viral supernatant removed and replacedby serum supplemented KGM complete growth mediumfor an additional 72 hours. Cells were then plated in 60-mm dishes at a limited dilution of 50 cells/dish. Singleclones were then isolated using a cloning disc soaked withTrypsin-EDTA and subcultured to 6-well plates. Initially,8 clones were isolated, and one clone (CN-G2) that pro-duced abundant lipid droplets was selected for further study.
Characterization of Clone CN-G2: The lipid-producing clone,CN-G2, was characterized for the production of lipid and the ef-fects of serum and PPARg agonist, rosiglitazone. Cells at passage23 and above were plated at 30% confluence in either glass cham-ber slides (BD Falcon 4-well CultureSlide, Rockville, MD) forimmunocytochemistry and lipid staining or 100-mm diameterdishes for biochemical and RNA analysis. Cells were then culturedfor 7 days (3 coverslips and 3 dishes per condition) in either 0%,1%, 2% or 10% serum containing KGM growth media to deter-mine the effects of serum on lipid synthesis and PPARg localiza-tion. To assess the effects of rosiglitazone, cells were plated at 30%confluence in 2% serum containing KGM growth media for 7 daysand then switched to media containing rosiglitazone at 10 mM,20 mM and 50 mM (Enzo Life Sciences, Plymouth Meeting, PA)with changes in the media every other day. Cells were thencollected at 1, 3, and 5 days (3 coverslips per condition) and7 days (3 coverslips and 3 dishes per condition) for analysis of lipidsynthesis or PPARg localization and gene expression.
C. ImmunocytochemistryFor analysis of meibomian glands, eyelids from 3 young
(2-month-old) and 3 old (2-year-old) mice was initially fixedovernight in 2% paraformaldehyde in PBS at 4�C, washed inPBS, embedded in Tissue Tech O.C.T. Compound (SakuraFinetek USA, Inc, Torrance, CA), frozen in liquid nitrogenand stored at -80�C and then sectioned using a LeicaCM1850 Cryotome (Leica, Wetzlar, Germany), and mountedonto glass slides. For analysis of cultured meibocytes, cellswere fixed in 2% paraformaldehyde in PBS for 2 hours.
For immunostaining, cells and tissue sections were per-meablized in PBS containing 0.5% dimethyl sulfoxide and0.5% Triton X (pH 7.2) for 5 minutes and then washed inPBS. Slides were then incubated in goat serum (1/30) for30 minutes at 37�C and then incubated with rabbit anti-PPARg (1:50, Abcam, Cambridge, MA). Slides were thenwashed with PBS, stained with FITC conjugated goat anti-rabbit IgG (1:200, Invitrogen) for 1 hour at 37�C and thencounterstained with DAPI (Invitrogen). For negative con-trols, primary antibodies were either omitted or replace bynonspecific rabbit sera. The samples were then evaluatedand imaged using a Nikon Eclipse E600 epifluorescence mi-croscope (Nikon Inc, Melville, NY).
D. Assessment of Meibocyte Lipid SynthesisTo assess whether immortalized meibocytes synthesized
lipids characteristic of the meibomian gland, CN-G2 cellswere evaluated by coherent anti-Stokes Raman scattering
486 THE OCULAR SURFACE / OCTOBER 2016
(CARS) and Raman microspectroscopy using previouslypublished techniques.33 Briefly, multimodal CARS micro-scopy was carried out using an inverted confocal microscope(Fluoview 300, Olympus) and an optical parametric oscil-lator (Levante Emerald OPO, APE, Berlin) pumped by thesecond harmonic of a Nd:vanadate picosecond mode-locked laser (PicoTrain, High-Q). CARS images were ob-tained using a 20x, 0.70 NA objective (UplanSApo,Olympus), which has a w0.5 mm lateral and w3.5 mm axialfocal volume. Raman microspectroscopy was carried out us-ing a frequency doubled Nd:vanadate laser (Verdi V5,Coherent) tuned to 532 nm. Raman microspectroscopywas coupled to CARS through the back port of the micro-scope and used the same objective lens. Switching betweenCARS and Raman detection was accomplished using acarrousel, allowing detection of lipid droplets using CARS,which were then directly analyzed by Raman.
To measure the lipid content, cells were stained withneutral lipid fluorescent probe, HCS LipidTOXTM (Invitro-gen. Carlsbad, CA). Cells were fixed in 2% paraformalde-hyde and then rinsed in PBS. Cells were then incubated inHCS LipidTox solution (dilution 1:1000) for 20 minutes atroom temperature. Coverslips were rinsed in PBS, counter-stained with DAPI nuclear stain and fluorescence imagedusing the Nikon Eclipse E600 microscope. A total of 5random images for each coverslip (3 coverslips totaling 15image per treatment) were digitized and then analyzed usingMetamorph Image Processing Software (Molecular Devices,Downington, PA, USA). All coverslips from each experi-ment were stained and evaluated on the same day to reducethe effects of random variations in staining intensity and im-aging. Regions of fluorescence were first thresholded usingthe Threshold subroutine to include all high intensity pixelsrepresenting lipid droplets within cells and generallyincluded pixel intensities from 100 to 255. This thresholdwas then applied to all subsequent images taken duringthe same experiment. Thresholded pixel area was thenmeasured using the Measure subroutine in Metamorphand the area recorded. For each image the number of cellswas also determined by counting the nuclei using theManual Count subroutine. To calculate the lipid contentper cell in each image, the thresholded lipid pixel area wasthen divided by the total number of cells in each image.
E. Protein Isolation and Western BlottingFor mouse meibomian glands, 5 young (2-month-old)
and 5 old (2-year-old) mice were sacrificed and the meibo-mian glands isolated from the upper and lower eyelids andpooled together to obtain enough protein for multiple west-ern blots. Tissues were then homogenized using a Polytron(PT-1200, Kinematic, Bohemia, NY). For extraction of pro-tein from tissue cultured cells, 100 mm dishes were firstrinsed 3 times in Dulbecco’s phosphate buffered saline,and then cells were scraped from the dish using a polyeth-ylene Cell Lifter (Corning Inc., Corning, NY). Proteinsfrom tissues and cells were fractionated using the NE-PERNuclear & Cytoplasmic Extraction Reagent kit from Thermo
, VOL. 14 NO. 4 / www.theocularsurface.com

MEIBOCYTE PPARg / Jester, et al
Scientific (Rockford, IL). All extraction buffers were supple-mented with the Calbiochem Protease Inhibitor Cocktail SetIII (EMD Chemical Inc, Gibbstown, NJ) and PhosphataseInhibitor Cocktail 2 (Sigma-Aldrich, St Louis, MO). Theprotein content of the cell and tissue extracts was thenmeasured using the RC DC Protein Assay (Bio-Rad Labora-tories, Hercules, CA) and the samples run on 12% SDSPAGE gels. Proteins were then transferred to PVDF mem-branes using iBlot Gel Transfer Device (Invitrogen, Carls-bad, CA). Membranes were then blocked using 5% non-fatdry milk in PBS-T (0.2% Tween 20) and then immuno-stained using antibodies to PPARg (Dilution 1:500). Mem-branes were then rinsed with PBS-T and then incubatedwith goat anti-rabbit IgG (HþL) HRP (Dilution 1:2500;Invitrogen), rinsed with PBS-T and incubated with Super-Signal� West Pico Chemiluminescent Substrate (ThermoScientific, Rockford, IL). Immunostained bands were thendetected using photographic film. All western blots wererepeated 3 times.
F. ImmunoprecipitationCytoplasmic extracts from cultured meibocytes were
first immunoprecipitated using goat anti-PPARg antibodies(sc-22020, Santa Cruz Biotechnology, Dallas, TX) linked toDynabeads-Protein G (Novex, Life Technologies, Oslo, Nor-way). Proteins were eluted and then western blotted usingrabbit anti-PPARg antibodies (ab27649, Abcam, Cam-bridge, MA) to identify presence of cytoplasmic 50 and72 kDa PPARg. Blots were then stripped using a mild strip-ping protocol and then reacted with rabbit monoclonal anti-body to SUMO1 (ab133352, Abcam).
G. RNA Isolation and Analysis of Gene ExpressionGene expression was evaluated by real time PCR using
previously published methods.34,35 Briefly, cells were directlylysed using RLT buffer and the RNA isolated over RNeasyspin columns as suggested by the manufacturer (Quiagen,Valencia CA). RNA yield and quality was evaluated withthe aid of a Nanodrop spectrophotometer (Thermo Scienti-fic, Wilmington, DE). 0.5 mg RNA was reverse transcribedusing oligo dT and random primers as supplied in theQuantiTect Reverese Transcription Kit (Quiagen). Realtime PCR was performed using Power Sybr Green reagents(ABI Life Technologies, Foster City, CA) and validated realtime PCR primers obtained from SA Biosciences (Quiagen).Products were evaluated by melt curve analysis and sizingon agarose gels. Relative quantization was performed usingthe delta CT method using both GAPDH and beta actin asthe normalizing housekeeper genes as previouslydescribed.34
H. Statistical AnalysisAll results are reported as mean � standard deviation.
Differences between groups were assessed by one-wayANOVA and Bonferroni multiple comparisons (SigmaStat version 3.11, Systat Software Inc, Point Richmond,CA). All experiments were repeated at least 3 times.
THE OCULAR SURFACE / OCTOBER 2016, VO
III. RESULTSA. Cytoplasmic and Nuclear PPARg Expression
As shown earlier,31 immunostaining of mouse eyelids withantibodies specific for PPARg show a cytoplasmic/vesicularand nuclear localization of PPARg within the acinar cells ofmeibomian glands in young mice (Figure 1A). This is incontrast to the predominantly nuclear localization that isdetected in acinar cells of meibomian glands in older mice(Figure 1B). Subcellular fractionation of pooled mouse meibo-mian gland tissue (5 mice/age group) into cytoplasmic and nu-clear fractions showed that the nucleus contains almostexclusively a 50 kDa PPARg in both young (2M) and old(2Y) glands that is consistent with the expression of the g1splice variant of PPARg (Figure 1C).36 Furthermore, stainingof western blots with antibodies specific for g1 showed positivestaining while antibodies to g2 failed to stain (data not shown).Evaluation of the cytoplasmic fraction showed that in additionto the 50 kDa PPARg splice variant there was a 72 kDaPPARg variant suggesting post translational modification ofPPARg. Comparison of young (2M) and old (2Y) meibomianglands indicated that there was a marked decrease in both the50 and 72 kDa PPARg forms in the cytoplasmic compartmentof the old meibomian glands, with almost complete loss of the50 kDa PPARg. Densitometry measurements based onnormalization to GAPDH staining (Figure 1D) showed a sig-nificant (P < .05) 75% decrease in the 72 kDa band and com-plete loss for the 50 kDa band. Additionally, the nuclear50 kDa PPARg was reduced by 40% based on densitometrynormalized to nuclear Histone 3 expression (H3).
B. Characterization of Immortalized MouseMeibocytesAfter immortalization and cloning, we compared the
characteristics of primary cultured, normal mouse meibo-cytes to that of the SV40 transformed and immortalizedmouse meibocyte clone, CN-G2 (Figure 2). Cultured normalmouse meibocytes showed an epithelial morphology withvery slow growth that required weeks to come to confluence.In culture many cells showed accumulation of perinuclearvacuoles (Figure 2A) that appeared to enlarge over time.By contrast, CN-G2 immortalized cells were fast growingbut also showed an epithelial morphology and the accumu-lation of perinuclear vacuoles that at times filled the cyto-plasm (Figure 2B, arrows). Staining of cells withantibodies specific to PPARg showed that both normal mei-bocytes (Figure 2C) and CN-G2 cells (Figure 2D) containedboth nuclear and cytoplasmic localized PPARg. Cytoplasmicstaining was both diffuse and filamentous with some cellsthat contained perinuclear vacuoles, presumably represent-ing lipid, showing strong diffuse PPARg staining(Figure 2D, arrow). Staining of CN-G2 cells (Figure 2E)and normal meibocytes (data not shown) with the neutrallipid probe, LipidTox (Green), showed strong fluorescentstaining of cytoplasmic vacuoles indicating the synthesis ofneutral lipids. These fluorescent images were digitized andthen thresholded using digital imaging software as shownin Figure 2F to measure lipid area.
L. 14 NO. 4 / www.theocularsurface.com 487

Figure 1. PPARg localization inmouse meibomian gland identifiedby immunostaining (A and B) andwestern blotting (C and D) (Repre-sentative of 3 mice per age forimmunocytchemistry and pooledmeibomian gland proteins from 5mice for each age). Young mice (A)showed both nuclear and cyto-plasmic PPARg staining while oldmice (B) showed predominantly nu-clear staining (green ¼ PPARg,red ¼ nuclei). Western blots (C) anddensitometry averaged from 3western blots and normalized usingGAPDH for cytoplasmic fraction andHistone 3 for nuclear fraction (D)show that the nucleus contains onlythe 50 kDa PPARg band which issignificantly (P<.05) reduced in oldmice, while the cytoplasmic fractionshows loss of the 50 kDa band and asignificantly (P<.05) reduced 72 kDaPPARg band in old glands (errorbar ¼ standard deviation).
MEIBOCYTE PPARg / Jester, et al
To verify that the cells were meibocytes and synthe-sized meibomian gland lipid, CN-G2 cells were analyzedusing CARS and Raman spectroscopy. Cells that con-tained visible lipid droplets by brightfield microscopyshowed the presence of bright spots using CARS imaging[Figure 3A]. In general, lipid droplets ranged in size from1.3-4.0 mm in diameter, which is sufficiently large toprobe with Raman spectroscopy. The spontaneous Ramanspectrum that was obtained from the lipid droplets isshown in Figure 3B;Red line ¼ CN-G2 cells. For compar-ison, the Raman spectrum obtained from mouse meibo-mian gland lipid is also shown [Figure 3B; Blackline ¼ MG Acini]. It should be noted that the twomain peaks at 2846 cm�1 and 2886 cm�1 represent therespective symmetric and asymmetric CH2 stretching vi-brations of purified wax and cholesterol esters, character-istic of the major meibomian gland lipids that have beenpreviously reported.33 Importantly, the lipids synthesizedby CN-G2 cells show similar peak vibrational componentsindicating the lipid contains high amounts of wax estersand cholesterol esters. By contrast, lipids from sebaceousgland and fat tissue, which have reduced cholesterol esters,show remarkably different Raman spectra with a reducedvibrational peak at 2886 cm�1 [Figure 3C;Black ¼ sebocyte and Blue ¼ adipocyte]. Of interestwas the finding that there is a third broad peak around2940 cm-1 [Figure 3B, arrow], which corresponds to alower peak identified in the meibomian gland acinar lipidand represents a CH3 stretch vibration associated withprotein. This peak has been shown to vary dependingon the location of lipid from the acini to the duct, andmay contain information regarding the protein contentor lipid fluidity.33
488 THE OCULAR SURFACE / OCTOBER 2016
C. Lipid Synthesis and PPARg Localization inCultured MeibocytesSince serum contains unsaturated and essential free fatty
acids, which are ligands for PPARg, cells were cultured indifferent serum concentrations to evaluate the effects ofserum on lipid synthesis. When cells were cultured for7 days and then stained with LipidTox, there was a markedreduction in the neutral lipid staining in low serum concen-trations compared to cells maintained in 10% serum(Figures 4A and 4B, respectively). Quantifying the amountof lipid present in cultures showed that there was a signifi-cant reduction (P < .05) in total lipid area/cell when cellswere cultured in media contain 0%, 1% or 2% serum withan 82%, 83% and 37% reduction in lipid synthesis, respec-tively (Figure 4C). Although 2% serum containing mediaappeared to promote lipid synthesis compared to serumfree or 1% serum, 2% serum maintained cell viability whileat the same time significantly reducing lipid synthesiscompared to 10% serum treated cultures. Reduced serumalso altered the localization of PPARg as identified by cellfractionation and western blotting (Figure 4D). While nochange was detected in the localization of nuclear PPARg,cultures grown in low serum showed a significant (P<.05)loss of the cytoplasmic 72 kDa PPARg compared to cellsgrown in 10% serum containing media. Overall, these find-ings indicate that reduced serum concentrations in the cul-ture media lead to significant loss of lipid synthesis that isassociated with loss of the 72 kDa PPARg protein.
D. Effects of Rosiglitazone on Lipid Synthesis andPPARg LocalizationCN-G2 cells were grown in media containing 2% serum for
7 days, and then treated with rosiglitazone, a member of the
, VOL. 14 NO. 4 / www.theocularsurface.com

Figure 2. Brightfield (A and B)and PPARg immunostaining (C andD) of primary cultured mousemeibocytes (A and C) and SV40immortalized CN-G2 meibocytes (Band D). Note that both cells inculture appear to contain promi-nent vacuoles (arrows) and showcyotplasmic and nuclear PPARgstaining. Cytoplasmic vacuolesshowed strong staining with fluo-rescent LipidTox (E) indicating thepresence of neutral lipids. Area ofLipidTox staining was thenthresholded (F, orange area) tomeasure lipid area. Each image isrepresentative of 3 separate cov-erslips.
MEIBOCYTE PPARg / Jester, et al
thiazolidineiones family of synthetic PPARg agonists that hasbeen shown to have important anti-type 2 diabetic actions.28 Itstimulates sebum production in diabetic patients and lipid syn-thesis in sebocyte cell cultures.37 Cells treated for 7 days withrosiglitazone showed a significant (P<.05), dose- and time-dependent, 8-10-fold increase in lipid area per cell followingexposure to 10 mM, 20 mM and 50 mM rosiglitazone whencompared tobaseline lipid levels (Figure 5). Fractionationof cells
THE OCULAR SURFACE / OCTOBER 2016, VO
andwestern blotting for the cytoplasmic fraction of PPARg after7 days exposure to rosiglitazone (Figure 6A) also showed a sig-nificant (P<.05) dose-dependent increase in cytoplasmicPPARg compared to untreated CN-G2 cells (0 mM). Interest-ingly, cytoplasmic fractions from CN-G2 cells also showedincreased amounts of both the 50 kDa and 72 kDa PPARg pro-tein with the appearance of multiple bands. These additionalbands most likely represent post translational modifications
L. 14 NO. 4 / www.theocularsurface.com 489

Figure 3. Coherent Anti-StokesRaman (CARS) image focused on2845 cm�1 (A) and Raman micro-spectroscopy (B and C) of CN-G2lipid vacuoles. CN-G2 cells (B andC, Red line) show major peaks at2846 cm�1 and 2886 cm�1, similarto that of lipid within meibomiangland acini (B) indicating theabundance of wax and cholesterolesters, respectively. Lipid in CN-G2cells was distinct from that oflipid within sebocytes and adipo-cytes (C, black and blue, respec-tively), which contain reducedcholesterol esters. Also note thepresence of an elevated peak at2940 cm�1 from the CH3 stretchregion representative of protein inCN-G2 cells (B, arrow) that is alsopresent in acinar lipid at reducedlevels that show maturationalchanges within the meibomiangland lipid.
MEIBOCYTE PPARg / Jester, et al
involving serine phosphorylation and sumoylation. To identifywhether these higher molecular weight bands were due topost-translationalmodification of PPARg, the cytoplasmic frac-tion was immunoprecipitated to isolate PPARg using a goatanti-PPARg antibody (Figure 6B). Staining of western blotswith rabbit anti-PPARg confirmed the immunoprecipitationof both the 50 kDa and72 kDaPPARgbands (Lane 1). Strippingand reprobing of western blots with antibodies against SUMO1showed that the 72 kDa PPARg band was sumolyated and posttranslationally modified (Lane 2). RNA extracted from culturestreated with 50 mM rosiglitazone also showed enhanced expres-sion of PPARg, adiponectin and ADFP (Figure 6C, numberabove bar¼ range) as measured by real-time PCR.
IV. DISCUSSIONThis report establishes that aging in the mouse meibo-
mian gland leads to a loss in the expression of the lipid-sensitive nuclear receptor, PPARg, with complete loss of
490 THE OCULAR SURFACE / OCTOBER 2016
the 50 kDa PPARg1 splice variant and substantial loss ofa 72 kDa PPARg band within the cytoplasmic compartmentof the gland. Cell culture studies using an SV40 transformedmeibocyte cell line also confirm that decreased lipid synthe-sis associated with reduced serum concentrations was asso-ciated with loss of the 72 kDa PPARg. Furthermore, thisreport establishes for the first time that the PPARg agonist,rosiglitazone, dose-dependently increases lipid synthesisconcomitant with increased PPARg cytoplasmic localizationof both the 50 kDa and 72 kDa proteins, identifies the sumo-lyation of PPARg by SUMO1 as potentially playing a role innuclear export of PPARg, and establishes the upregulationof PPARg response genes involved in lipid synthesis, i.e.adiponectin and ADFP. Taken together, these findings sug-gest that lipid synthesis in the meibomian gland is regulatedby PPARg and that during aging there is decreased PPARgreceptor activation, leading to deceased lipid synthesis andage-related atrophy of the meibomian gland.
, VOL. 14 NO. 4 / www.theocularsurface.com
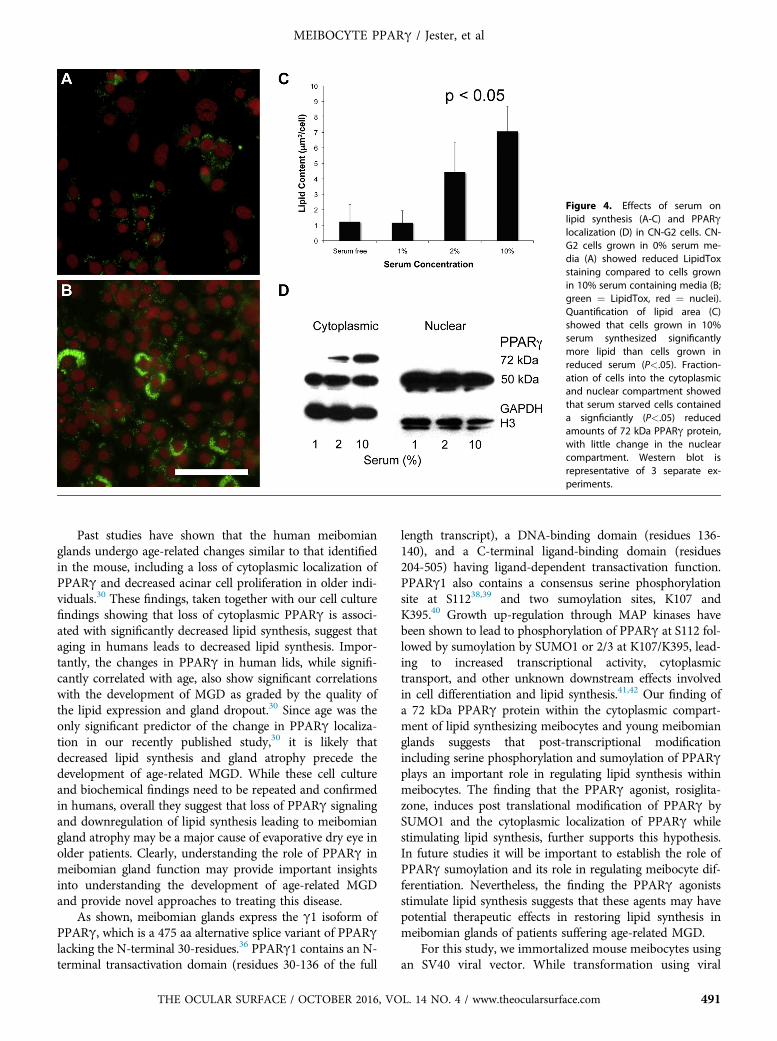
Figure 4. Effects of serum onlipid synthesis (A-C) and PPARglocalization (D) in CN-G2 cells. CN-G2 cells grown in 0% serum me-dia (A) showed reduced LipidToxstaining compared to cells grownin 10% serum containing media (B;green ¼ LipidTox, red ¼ nuclei).Quantification of lipid area (C)showed that cells grown in 10%serum synthesized significantlymore lipid than cells grown inreduced serum (P<.05). Fraction-ation of cells into the cytoplasmicand nuclear compartment showedthat serum starved cells containeda signficiantly (P<.05) reducedamounts of 72 kDa PPARg protein,with little change in the nuclearcompartment. Western blot isrepresentative of 3 separate ex-periments.
MEIBOCYTE PPARg / Jester, et al
Past studies have shown that the human meibomianglands undergo age-related changes similar to that identifiedin the mouse, including a loss of cytoplasmic localization ofPPARg and decreased acinar cell proliferation in older indi-viduals.30 These findings, taken together with our cell culturefindings showing that loss of cytoplasmic PPARg is associ-ated with significantly decreased lipid synthesis, suggest thataging in humans leads to decreased lipid synthesis. Impor-tantly, the changes in PPARg in human lids, while signifi-cantly correlated with age, also show significant correlationswith the development of MGD as graded by the quality ofthe lipid expression and gland dropout.30 Since age was theonly significant predictor of the change in PPARg localiza-tion in our recently published study,30 it is likely thatdecreased lipid synthesis and gland atrophy precede thedevelopment of age-related MGD. While these cell cultureand biochemical findings need to be repeated and confirmedin humans, overall they suggest that loss of PPARg signalingand downregulation of lipid synthesis leading to meibomiangland atrophy may be a major cause of evaporative dry eye inolder patients. Clearly, understanding the role of PPARg inmeibomian gland function may provide important insightsinto understanding the development of age-related MGDand provide novel approaches to treating this disease.
As shown, meibomian glands express the g1 isoform ofPPARg, which is a 475 aa alternative splice variant of PPARglacking the N-terminal 30-residues.36 PPARg1 contains an N-terminal transactivation domain (residues 30-136 of the full
THE OCULAR SURFACE / OCTOBER 2016, VO
length transcript), a DNA-binding domain (residues 136-140), and a C-terminal ligand-binding domain (residues204-505) having ligand-dependent transactivation function.PPARg1 also contains a consensus serine phosphorylationsite at S11238,39 and two sumoylation sites, K107 andK395.40 Growth up-regulation through MAP kinases havebeen shown to lead to phosphorylation of PPARg at S112 fol-lowed by sumoylation by SUMO1 or 2/3 at K107/K395, lead-ing to increased transcriptional activity, cytoplasmictransport, and other unknown downstream effects involvedin cell differentiation and lipid synthesis.41,42 Our finding ofa 72 kDa PPARg protein within the cytoplasmic compart-ment of lipid synthesizing meibocytes and young meibomianglands suggests that post-transcriptional modificationincluding serine phosphorylation and sumoylation of PPARgplays an important role in regulating lipid synthesis withinmeibocytes. The finding that the PPARg agonist, rosiglita-zone, induces post translational modification of PPARg bySUMO1 and the cytoplasmic localization of PPARg whilestimulating lipid synthesis, further supports this hypothesis.In future studies it will be important to establish the role ofPPARg sumoylation and its role in regulating meibocyte dif-ferentiation. Nevertheless, the finding the PPARg agonistsstimulate lipid synthesis suggests that these agents may havepotential therapeutic effects in restoring lipid synthesis inmeibomian glands of patients suffering age-related MGD.
For this study, we immortalized mouse meibocytes usingan SV40 viral vector. While transformation using viral
L. 14 NO. 4 / www.theocularsurface.com 491

Figure 5. Effects of 0, 10 mM, 20 mM, and 50 mM rosiglitazone on lipidsynthesis in CN-G2 cells. Note that rosiglitazone dose and time-dependently stimulated a significant (P<.05), 8-10 fold increase in lipidarea within CN-G2 cells. (Data is representative of 3 separateexperiments).
Figure 6. Effects of rosiglitazone on PPARg localization (A), posttranslational modification (B) and PPARg response gene expression (C).Induced lipid synthesis by rosiglitazone was associated with a significant(P<.05) increase in cytoplasmic localization of PPARg proteins showingmultiple bands consistent with post translational modification involvingserine phosphorylation and sumoylation (A). Immunoprecipitation ofPPARg using goat anti-PPARg antibodies pulled down both the 50 kDAand 72 kDa proteins (B, Lane 1). Stripping and re-probing of western blotusing a rabbit anti-SUMO1 antibody showed staining of the 72 kDaband, indicating sumoylation. Rosiglitazone also enhanced expression ofPPARg and other PPARg response genes, adiponectin and ADFP (C).
MEIBOCYTE PPARg / Jester, et al
oncogenes are known to produce cell lines showing geneticinstability and altered cell growth regulation and differenti-ation, a similar approach has been used for the sebaceousgland to establish a lipid synthesizing cell line for whichthe molecular mechanisms of sebaceous gland functionhave been evaluated.43-46 Recently, a human telomeraseimmortalized meibomian gland epithelial cell line has beengenerated that contains neutral lipids, expresses genesinvolved in lipid synthesis and responds to androgens.21
Since telomerase immortalization has been shown to main-tain normal cell differentiation,47 future studies are neededto compare our findings regarding regulation of lipid syn-thesis by PPARg in this meibomian gland epithelial cellline. Additionally, the effects of androgens, known to playan important role in the development of MGD in patients,on PPARg signaling are also needed.
Finally, Raman spectral analysis of CN-G2 lipid detectedan elevated vibrational peak around 2940 cm�1 in addition tothe two main peaks at 2846 cm�1 and 2886 cm�1 associatedwith wax and cholesterol esters. Though less prominent, thispeak was also identified in the mouse meibomian glandacinar lipid and showed progressive decreasing levels as lipidmoved from the acinus to the central meibomian gland ductsuggesting maturation of the lipid.33 The significance of this
492 THE OCULAR SURFACE / OCTOBER 2016
peak is possibly related to either increased protein associatedwith the lipid or differences in lipid fluidity. The finding thatCN-G2 cells show an enhanced vibrational peak in this re-gion is consistent with this maturation hypothesis; newly syn-thesized meibomian gland lipid would contain the highestamount of associated protein or change in fluidity. Furtherstudy is needed to determine the significance of this regionand whether lipid produced by meibocytes in culture un-dergoes similar maturation following synthesis of lipid.
V. CONCLUSIONThis study shows that lipid synthesis in the mouse is
regulated by the lipid-sensitive nuclear receptor, PPARg.
, VOL. 14 NO. 4 / www.theocularsurface.com

MEIBOCYTE PPARg / Jester, et al
Furthermore, downregulation of lipid synthesis by serumstarvation leads to decreased cytoplasmic localization ofPPARg in cultured meibocytes, similar to that observed inolder mice and human meibomian glands. Taken together,these findings suggest that there is an age-related decreasein lipid synthesis by the meibomian gland that may explain,in part, age-related MGD and the development of evapora-tive dry eye in this patient population.
ACKNOWLEDGMENTThe authors wish to recognize and thank Chyong Jy Nien, MD, Salina
Massei, Gloria Lin, Ginna Kim, and Tejas Shah who have provided assis-tance in the collection and analysis of the data presented in this paper.
REFERENCES1. Bron AJ, Tripathi RC, Tripathi BJ. Wolff’s Anatomy of the Eye and
Orbit. 8th ed. London, Chapman & Hall Medcial, 19972. Jester JV, Nicolaides N, Smith RE.Meibomian gland studies: histologic and
ultrastructural investigations. Invest Ophthalmol Vis Sci 1981;20:537-473. Mishima S, Maurice DM. The oily layer of the tear film and evaporation
from the corneal surface. Exp Eye Res 1961;1:39-454. Mathers WD, Lane JA. Meibomian gland lipids, evaporation, and tear
film stability. Adv Exp Med Biol 1998;438:349-605. Gilbard JP, Rossi SR, Heyda KG. Tear film and ocular surface changes
after closure of the meibomian gland orifices in the rabbit. Ophthal-mology 1989;96:1180-6
6. Foulks GN, Bron AJ. Meibomian gland dysfunction: a clinical scheme fordescription, diagnosis, classification, and grading. Ocul Surf 2003;1:107-26
7. Lemp MA, Nichols KK. Blepharitis in the United States 2009: a survey-based perspective on prevalence and treatment. Ocul Surf 2009;7:S1-14
8. Tong L, Chaurasia SS, Mehta JS, Beuerman RW. Screening for meibomiangland disease: its relation to dry eye subtypes and symptoms in a tertiaryreferral clinic in singapore. Invest Ophthalmol Vis Sci 2010;51:3449-54
9. Viso E, Gude F, Rodriguez-Ares MT. The association of meibomiangland dysfunction and other common ocular diseases with dry eye: apopulation-based study in Spain. Cornea 2011;30:1-6
10. Nichols KK, Foulks GN, Bron AJ, et al. The international workshop onmeibomian gland dysfunction: executive summary. Invest OphthalmolVis Sci 2011;52:1922-9
11. Knop E, Knop N, Millar T, et al. The international workshop on mei-bomian gland dysfunction: report of the subcommittee on anatomy,physiology, and pathophysiology of the meibomian gland. Invest Oph-thalmol Vis Sci 2011;52:1938-78
12. Jester JV, Nicolaides N, Kiss-Palvolgyi I, Smith RE. Meibomian glanddysfunction. II. The role of keratinization in a rabbit model of MGD.Invest Ophthalmol Vis Sci 1989;30:936-45
13. Jester JV, Rajagopalan S, Rodrigues M. Meibomian gland changes in therhino (hrrhhrrh) mouse. Invest Ophthalmol Vis Sci 1988;29:1190-4
14. Ohnishi Y, Kohno T. Polychlorinated biphenyls poisoning in monkeyeye. Invest Ophthalmol Vis Sci 1979;18:981-4
15. Sullivan BD, Evans JE, Cermak JM, et al. Complete androgen insensi-tivity syndrome: effect on human meibomian gland secretions. ArchOphthalmol 2002;120:1689-99
16. Sullivan BD, Evans JE, Dana MR, Sullivan DA. Influence of aging onthe polar and neutral lipid profiles in human meibomian gland secre-tions. Arch Ophthalmol 2006;124:1286-92
17. Ding J, Kam WR, Dieckow J, Sullivan DA. The influence of 13-cis ret-inoic acid on human meibomian gland epithelial cells. Invest Ophthal-mol Vis Sci 2013;54:4341-50
THE OCULAR SURFACE / OCTOBER 2016, VO
18. Ding J, Sullivan DA. The effects of insulin-like growth factor 1 andgrowth hormone on human meibomian gland epithelial cells. JAMAOphthalmol 2014;132:593-9
19. Kam WR, Sullivan DA. Neurotransmitter influence on human meibo-mian gland epithelial cells. Invest Ophthalmol Vis Sci 2011;52:8543-8
20. Khandelwal P, Liu S, Sullivan DA. Androgen regulation of gene expres-sion in human meibomian gland and conjunctival epithelial cells. MolVis 2012;18:1055-67
21. Liu S, Hatton MP, Khandelwal P, Sullivan DA. Culture, immortaliza-tion, and characterization of human meibomian gland epithelial cells.Invest Ophthalmol Vis Sci 2010;51:3993-4005
22. Liu S, Kam WR, Ding J, et al. Effect of growth factors on the prolifer-ation and gene expression of human meibomian gland epithelial cells.Invest Ophthalmol Vis Sci 2013;54:2541-50
23. Liu Y, Kam WR, Ding J, Sullivan DA. Effect of azithromycin on lipidaccumulation in immortalized human meibomian gland epithelial cells.JAMA Ophthalmol 2014;132:226-8
24. Liu Y, KamWR, Ding J, Sullivan DA. Can tetracycline antibiotics dupli-cate the ability of azithromycin to stimulate human meibomian glandepithelial cell differentiation? Cornea 2015;34:342-6
25. Sullivan DA, Liu Y, Kam WR, et al. Serum-induced differentiation ofhuman meibomian gland epithelial cells. Invest Ophthalmol Vis Sci2014;55:3866-77
26. Rosen ED, Spiegelman BM. PPARgamma: a nuclear regulator of meta-bolism, differentiation, and cell growth. J Biol Chem 2001;276:37731-4
27. Fan W, Yanase T, Nomura M, et al. Androgen receptor null male micedevelop late-onset obesity caused by decreased energy expenditure andlipolytic activity but show normal insulin sensitivity with high adipo-nectin secretion. Diabetes 2005;54:1000-8
28. Hammarstedt A, Andersson CX, Rotter Sopasakis V, Smith U. The ef-fect of PPARgamma ligands on the adipose tissue in insulin resistance.Prostaglandins Leukot Essent Fatty Acids 2005;73:65-75
29. Nien CJ, Massei S, Lin G, et al. The development of meibomian glandsin mice. Mol Vis 2010;16:1132-40
30. Nien CJ, Massei SR, Lin G, et al. Effects of age and dysfunction on hu-man meibomian glands. Arch Ophthalmol 2011;129:462-9
31. Nien CJ, Paugh JR, Massei S, et al. Age-related changes in the meibo-mian gland. Exp Eye Res 2009;89:1021-7
32. Jester BE, Nien CJ, Winkler M, et al. Volumetric reconstruction of themouse meibomian gland using high-resolution nonlinear optical imag-ing. Anat Rec (Hoboken) 2011;294:185-92
33. Lin CY, Suhalim JL, Nien CL, et al. Picosecond spectral coherent anti-Stokes Raman scattering imaging with principal component analysis ofmeibomian glands. J Biomed Opt 2011;16:021104
34. Brown DJ, Lin B, Holguin B. Expression of neuregulin 1, a member ofthe epidermal growth factor family, is expressed as multiple splice var-iants in the adult human cornea. Invest Ophthalmol Vis Sci 2004;45:3021-9
35. Spirin KS, Ljubimov AV, Castellon R, et al. Analysis of gene expressionin human bullous keratopathy corneas containing limiting amounts ofRNA. Invest Ophthalmol Vis Sci 1999;40:3108-15
36. Tontonoz T, Spiegelman BM. Fat and beyond: The diverse biology ofPPARgamma. Ann Rev Biochem 2008;77:289-312
37. Trivedi NR, Cong Z, Nelson AM, et al. Peroxisome proliferator-activated receptors increase human sebum production. J Invest Derma-tol 2006;126:2002-9
38. Adams M, Reginato MJ, Shao D, et al. Transcriptional activation byperoxisome proliferator-activated receptor gamma is inhibited by phos-phorylation at a consensus mitogen-activated protein kinase site. J BiolChem 1997;272:5128-32
L. 14 NO. 4 / www.theocularsurface.com 493

MEIBOCYTE PPARg / Jester, et al
39. Hu E, Kim JB, Sarraf P, Spiegelman BM. Inhibition of adipogenesisthrough MAP kinase-mediated phosphorylation of PPARgamma. Sci-ence 1996;274:21003
40. Geiss-Friedlander R, Melchior F. Concepts in sumoylation: a decade on.Nat Rev Mol Cell Biol 2007;8:947-56
41. Burgemeister I, Seger R. MAPK kinases as nucleo-cytoplasmic shuttlesfor PPARg. Cell Cycle 2007;6:1539-48
42. van Beekum O, Fleskens V, Kalkhoven E. Post translational modifica-tions of PPAR-gamma: fine-tuning the metabolic master regulator.Obesity (Silver Spring) 2009;17:213-9
43. Makrantonaki E, Zouboulis CC. Testosterone metabolism to 5a-dihy-drotestosterone and synthesis of sebaceous lipis is regulated by the
494 THE OCULAR SURFACE / OCTOBER 2016
peroxisome proliferator-activated receptor ligand linoleic acid in hu-man sebocytes. Br J Dematol 2007;156:428-32
44. Wrobel A, Seltmann H, Fimmel S, et al. Differentiation and apoptosisin human immortalized sebocytes. J Invest Dermatol 2003;120:175-81
45. Xia L, Zouboulis CC, Ju Q. Culture of human sebocytes in vitro. Der-matoendocrinol 2009;1:92-5
46. Zouboulis CC, Seltmann H, Neitzel H, Orfanos CE. Establishment andcharacterization of an immortalized human sebaceous gland cell line(SZ95). J Invest Dermatol 1999;113:1011-20
47. Robertson DM, Li L, Fisher S, et al. Characterization of growth and dif-ferentiation in a telomerase-immortalized human corneal epithelial cellline. Invest Ophthalmol Vis Sci 2005;46:470-8
, VOL. 14 NO. 4 / www.theocularsurface.com


















