
Population Response of Giant Galápagos Tortoises to Feral Goat Removal
5
RESEARCH ARTICLE Population Response of Giant Gal´ apagos Tortoises to Feral Goat Removal Cruz M´ arquez, 1 James P. Gibbs, 1,2,3 Victor Carri´ on, 4 Sixto Naranjo, 4 and Alizon Llerena 1 Abstract Efforts to eradicate nonnative mammals to restore oceanic island ecosystems have become increasingly successful but parallel tracking of response by native species for which control efforts are intended to benefit has been rare. A major campaign to eradicate nonnative goats and burros was initiated in 1995 on Alcedo Volcano in the Gal´ apagos Archipelago that ultimately removed 62,868 goats and eliminated them by 2006. Planners of the eradication pro- gram had the foresight to invest in intensive monitoring of the status of the volcano’s giant tortoise (Chelonoidis nigra ) population whose welfare was a primary motivation for the eradication effort. Monitoring revealed an increase in the proportion of juveniles among all tortoises as well as increased growth rates of individual tortoises on Alcedo Volcano from earlier to later phases of the eradication cam- paign. Over the same time frame in a control population on nearby Santa Cruz Island (where goats and donkeys were not removed) juvenile fraction and individual growth rates remained unchanged. Although goat removal coin- cided with occurrence of a rare climatic event that simul- taneously boosted forage availability for tortoises, failure to observe a comparable improvement in the control pop- ulation implies that removal of goats and burros was the primary causative factor of improving population status of tortoises on Alcedo Volcano. Key words: eradication, Gal´ apagos Islands, giant tortoise, goats, invasive species, population restoration. Introduction Introduction of mammals to islands has lead to destruction of endemic species and degradation of ecosystems around the globe (Towns et al. 2006). In response, nonnative mammal eradication efforts have become widespread (Donlan & Wilcox 2008). Integrating monitoring of the species targeted for erad- ication within an adaptive management framework underlies much of the recent and striking upswing of success of eradi- cation operations world-wide (Lavoie et al. 2007). This said, parallel tracking of population- and community-level response by the endemic species for which control efforts are often intended to benefit is rare (Courchamp et al. 2003). Lack of evaluation of population response by endemic species to non- native species control programs hinders development of sup- port for further eradication efforts to restore island ecosystems. On the Gal´ apagos Archipelago, feral goats (Capra hircus ) and donkeys (Equus asinus ) (Fig. 1a & 1f) cause major damage to the vegetation and fauna (Hamann 1993). Alcedo Volcano on Isabela Island is an example. Goats first arrived 1 Charles Darwin Foundation, Puerto Ayora, Gal´ apagos, Ecuador 2 Department of Environmental and Forest Biology, State University of New York College of Environmental Science and Forestry, Syracuse, NY 13210, U.S.A. 3 Address correspondence to J. P. Gibbs, email [email protected] 4 Program for Protection and Conservation of Terrestrial Ecosystems, Gal´ apagos National Park Service, Puerto Ayora, Gal´ apagos, Ecuador © 2012 Society for Ecological Restoration doi: 10.1111/j.1526-100X.2012.00891.x from the adjacent volcano of Sierra Negra to the south in the late 1960s and were reported in the early 1980s in such restricted areas and small numbers as to be considered not to represent a threat to the biota (Duffy 1981). By the mid 1990s, however, the goat population had increased from 80 to 100,000 animals, causing severe damage to the vegetation (Freire 1992). Heightened concern initially expressed by MacFarland (1991) and others about competition for food between goats and endemic giant tortoises (Chelonoidis nigra vandenburghi ) on Alcedo gave rise to concerted control efforts by the Gal´ apagos National Park Service under the aegis of Project Isabela in 1995, which lead to removal of 62,868 individuals at a cost of US$4.1 million and eradication by 2006 (Carrion et al. 2011). The architects of Project Isabela invested in monitoring not only progress on goat and donkey removal (Carrion et al. 2011) but also endemic species’ responses to invasive mammal control efforts (e.g. birds: Donlan et al. 2007 and invertebrates: Desender et al. 1999). Herein we describe the population response by giant tortoises on Alcedo Volcano (Fig. 1b) whose welfare was one of the primary motives for Project Isabela. Giant tortoises are an iconic animal for Gal´ apagos and also an important “ecosystem engineer” there (Gibbs et al. 2008, 2009; Blake et al. in press). Goats primarily and donkeys secondarily are considered major competitors for food with tortoises as well as causing other forms of habitat degradation, such as removing shade trees, denuding shrubs of the foliage upon which mist condenses and drips from to the ground Restoration Ecology 1
-
Upload
cruz-marquez -
Category
Documents
-
view
213 -
download
0
Transcript of Population Response of Giant Galápagos Tortoises to Feral Goat Removal

R E S E A R C H A R T I C L E
Population Response of Giant Galapagos Tortoisesto Feral Goat RemovalCruz Marquez,1 James P. Gibbs,1,2,3 Victor Carrion,4 Sixto Naranjo,4 and Alizon Llerena1
Abstract
Efforts to eradicate nonnative mammals to restore oceanicisland ecosystems have become increasingly successful butparallel tracking of response by native species for whichcontrol efforts are intended to benefit has been rare. Amajor campaign to eradicate nonnative goats and burroswas initiated in 1995 on Alcedo Volcano in the GalapagosArchipelago that ultimately removed 62,868 goats andeliminated them by 2006. Planners of the eradication pro-gram had the foresight to invest in intensive monitoringof the status of the volcano’s giant tortoise (Chelonoidisnigra) population whose welfare was a primary motivationfor the eradication effort. Monitoring revealed an increasein the proportion of juveniles among all tortoises as well
as increased growth rates of individual tortoises on AlcedoVolcano from earlier to later phases of the eradication cam-paign. Over the same time frame in a control populationon nearby Santa Cruz Island (where goats and donkeyswere not removed) juvenile fraction and individual growthrates remained unchanged. Although goat removal coin-cided with occurrence of a rare climatic event that simul-taneously boosted forage availability for tortoises, failureto observe a comparable improvement in the control pop-ulation implies that removal of goats and burros was theprimary causative factor of improving population status oftortoises on Alcedo Volcano.
Key words: eradication, Galapagos Islands, giant tortoise,goats, invasive species, population restoration.
Introduction
Introduction of mammals to islands has lead to destructionof endemic species and degradation of ecosystems around theglobe (Towns et al. 2006). In response, nonnative mammaleradication efforts have become widespread (Donlan & Wilcox2008). Integrating monitoring of the species targeted for erad-ication within an adaptive management framework underliesmuch of the recent and striking upswing of success of eradi-cation operations world-wide (Lavoie et al. 2007). This said,parallel tracking of population- and community-level responseby the endemic species for which control efforts are oftenintended to benefit is rare (Courchamp et al. 2003). Lack ofevaluation of population response by endemic species to non-native species control programs hinders development of sup-port for further eradication efforts to restore island ecosystems.
On the Galapagos Archipelago, feral goats (Capra hircus)and donkeys (Equus asinus) (Fig. 1a & 1f) cause majordamage to the vegetation and fauna (Hamann 1993). AlcedoVolcano on Isabela Island is an example. Goats first arrived
1 Charles Darwin Foundation, Puerto Ayora, Galapagos, Ecuador2 Department of Environmental and Forest Biology, State University of New YorkCollege of Environmental Science and Forestry, Syracuse, NY 13210, U.S.A.3 Address correspondence to J. P. Gibbs, email [email protected] Program for Protection and Conservation of Terrestrial Ecosystems, GalapagosNational Park Service, Puerto Ayora, Galapagos, Ecuador
© 2012 Society for Ecological Restorationdoi: 10.1111/j.1526-100X.2012.00891.x
from the adjacent volcano of Sierra Negra to the south inthe late 1960s and were reported in the early 1980s in suchrestricted areas and small numbers as to be considered not torepresent a threat to the biota (Duffy 1981). By the mid 1990s,however, the goat population had increased from 80 to 100,000animals, causing severe damage to the vegetation (Freire1992). Heightened concern initially expressed by MacFarland(1991) and others about competition for food between goatsand endemic giant tortoises (Chelonoidis nigra vandenburghi )on Alcedo gave rise to concerted control efforts by theGalapagos National Park Service under the aegis of ProjectIsabela in 1995, which lead to removal of 62,868 individualsat a cost of US$4.1 million and eradication by 2006 (Carrionet al. 2011).
The architects of Project Isabela invested in monitoring notonly progress on goat and donkey removal (Carrion et al.2011) but also endemic species’ responses to invasive mammalcontrol efforts (e.g. birds: Donlan et al. 2007 and invertebrates:Desender et al. 1999). Herein we describe the populationresponse by giant tortoises on Alcedo Volcano (Fig. 1b) whosewelfare was one of the primary motives for Project Isabela.Giant tortoises are an iconic animal for Galapagos and alsoan important “ecosystem engineer” there (Gibbs et al. 2008,2009; Blake et al. in press). Goats primarily and donkeyssecondarily are considered major competitors for food withtortoises as well as causing other forms of habitat degradation,such as removing shade trees, denuding shrubs of the foliageupon which mist condenses and drips from to the ground
Restoration Ecology 1

Giant Tortoise-Feral Goat Interactions
(a) (b)
(c) (d)
(e) (f)
Figure 1. One of the 62,868 feral goats (a) removed from Alcedo Volcano, Galapagos Islands, Ecuador, between 1995 and 2006 to benefit the volcano’sendemic species, including giant tortoises (b), whose mass was recorded (c) along with length/growth rate (d) and sex and age [(e), mature male top,young female below] to measure tortoise population response to eradication of goats and donkeys [(f), note donkey on left rolling on potential tortoisenesting site].
where tortoises can drink it, and, in the case of donkeys,trampling tortoise nests (Fowler 1983). Our intent was to testthe hypothesis that eradication of goats and donkeys benefitedgiant tortoises on Alcedo Volcano.
Methods
Alcedo Volcano (−0.424731◦, −91.118421◦) is one of the fivemassive shield volcanoes comprising Isabela Island, the largestof the Galapagos Islands, an oceanic archipelago located1,000 km west of continental Ecuador. The volcano is unin-habited by humans. On the basis of genetic criteria tortoiseson Alcedo Volcano are demographically isolated from thoseon adjacent volcanoes, likely representing an endemic species(Caccone et al. 2002). Seasonal patterns of distribution oftortoises on the volcano remain poorly known; therefore,Alcedo’s tortoise population was sampled along permanenttransects 2–8 km long established at random across an ele-vational gradient among the four ecological zones present on
the volcano: (1) the semi-dry zone at 500 m, (2) the semi-humid zone at 600–900 m, (3) the wet zone between 900and 1,128 m, and (4) the volcano’s summit at 1,128 m. Tran-sects were monitored over 11 years: twice annually between1995 and 2000 and annually between 2001 and 2005. Duringtransect surveys tortoises were identified individually using aduplicate system to minimize misidentifications: (1) applyingnumbers in succession as brands to the rear costal scutes cou-pled with (2) simultaneous insertion of a passive integratedtransponder (PIT) tag (Avid Identification Systems Inc., Norco,CA, U.S.A.) into the left rear leg of each tortoise. Tortoisemass was recorded using an electronic scale suspended froma custom-built tetrapod upon which tortoises were raised fromthe ground via a pulley system (Fig. 1c). Curved carapacelength, the standard measure of tortoise size used in Galapagos,was measured as the distance from the notch at the anteriorof the carapace over the tops of the vertebral scutes to the tipof the supracaudal scute (Fig. 1d). Sex and age were assessedbased on external morphology (Fig. 1f).
2 Restoration Ecology

Giant Tortoise-Feral Goat Interactions
We used as a “control” population tortoises on SantaCruz Island (−0.695857◦, −90.409946◦) where the tortoisepopulation was surveyed during the same time frame and usingthe same methods as on Alcedo by biologists of the CharlesDarwin Foundation and park guards of the Galapagos NationalPark Service who repeatedly walked transects connectinghighland (feeding) and lowland (nesting) areas within theTortoise Reserve. The Tortoise Reserve on Santa Cruz Islandwas deemed a suitable control because it supports tortoisesof comparable size and morphology (domed-shaped carapacetype) to those of Alcedo; moreover, its habitats span a similarecological gradient (from dry lowlands to moist highlands).Because tortoises migrate over a gradient of elevation onSanta Cruz Island annually we selected from the data availablethose collected at two, adjacent and well-delineated sites (“elchato” and “las fincas”) during the same season each year(the dry season, August to December) to avoid conflatingelevation- or season-associated changes in tortoise populationparameters with a long-term temporal change in populationstatus. Tortoises on Santa Cruz were measured and markedusing the same methods as on Alcedo but were not weighed;therefore, we were able to contrast relative changes in growthand sex and age structure but not body condition (analyzed forAlcedo only).
We contrasted population parameters for tortoises on bothAlcedo Volcano and Santa Cruz Island during the high goatdensity phase (1995–1999) versus later (2000–2005) projectsegments of Project Isabela because about 85% of the goatsultimately removed from Alcedo Volcano were removed dur-ing the initial project phase. Independence in representationof juveniles among all the tortoises sampled in relation totime (initial phase of Project Isabela: 1995–1999; final phase:2000–2005) was assessed with G-tests (Zar 1984). General-ized additive mixed models (GAMMs) were used to evaluatebody condition (relative mass per unit length) and growth ratesfor tortoises (cf., Chaloupka & Limpus 1997; Seminoff et al.2002). GAMM’s enabled controlling for the nonlinear relation-ship between body mass and body size while partitioning outthe effects of categorical covariates: status (juvenile, female,male), season (dry: July to December; wet: January to June),and phases of Project Isabela (initial phase: 1995–1999; finalphase: 2000–2005). GAMMs were also used to model the rela-tionship between somatic growth rates and body size (curvedcarapace length), status (juvenile, female, or male), and projectphase. Growth rates (cm/year) were estimated from changein curved carapace length—the standard measure of tortoiselength used in Galapagos—between subsequent measurementsof recaptured individual tortoises divided by intervening timeinterval. PIT number was initially included in GAMMs forboth body condition and growth rate as a random effect toaccount for repeated measures on individuals (Crawley 2007)but did not contribute to variation in either parameter andtherefore was excluded. Tortoises whose status (sex, age) wasclassified as “unknown” (typically those in transition betweenjuvenile and adult) were not included in the analysis. All sta-tistical differences associated with α < 0.05 were interpretedas “significant.”
Results
A total of 8,112 tortoises were captured, measured, marked,and released between 1995 and 2005 on Alcedo Volcano witha median annual sample of 324 tortoises captured per year.Age composition (frequency of juveniles among all tortoises)varied between 1995–1999 and 2000–2005: the proportionof juveniles among all tortoises captured increased from0.05 to 0.24, respectively (Gadj = 626.7, df = 1, p < 0.001).In contrast, at both “el chato” and “las fincas” on SantaCruz Island the proportion of juveniles among all tortoisesremained unchanged between 1995–1999 and 2000–2005 (“elchato”: 0.03 to 0.01, Gadj = 1.31, df = 1, p = 0.25, n = 465tortoises; “las fincas”: 0.01 to 0.02, Gadj = 1.69, df = 1,p = 0.19, n = 844 tortoises).
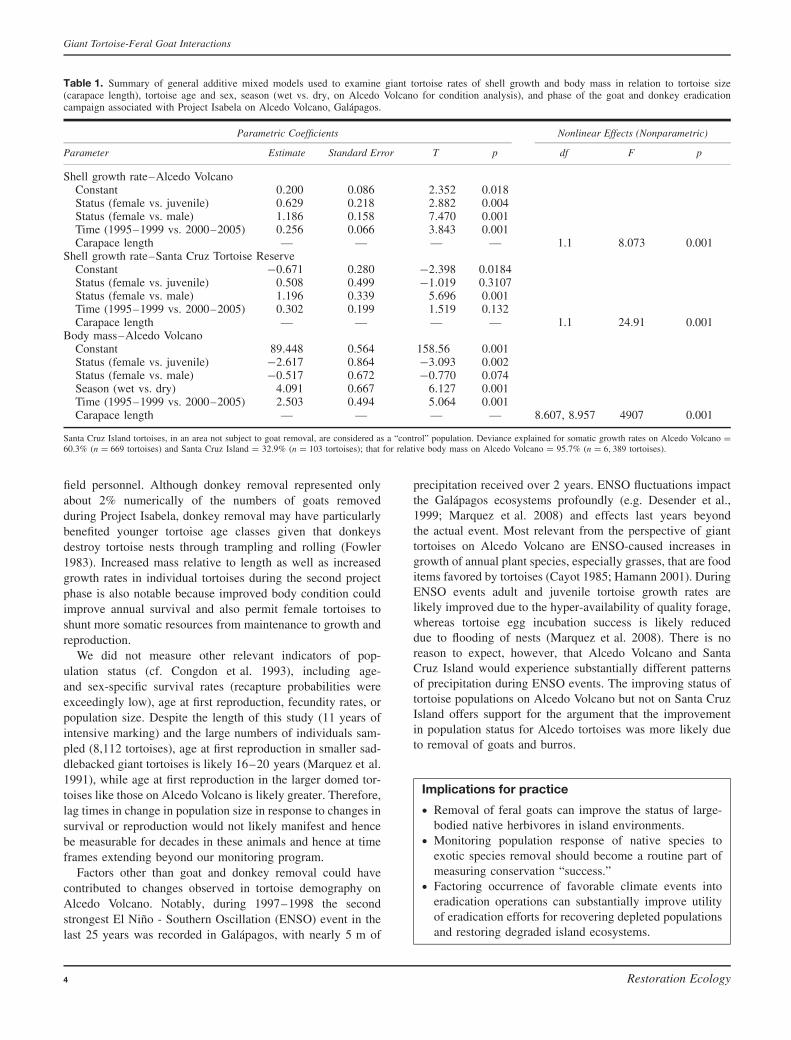
During 1995–2005, 669 PIT-tagged tortoises were recap-tured on average 2.6 years apart on Alcedo Volcano. Annualsomatic growth rates (centimeter per year in curved carapacelength) of individual, recaptured tortoises averaged 0.54 cm±1.29 cm (SD), with 60.1% of deviance explained by sexand age, carapace length, and project phase (Table 1). Juve-niles grew an average of 2.70 cm/year (±1.07 SD, n = 78),that is, 5× faster than males (0.53 cm/year ±1.15, n = 232)and 35× faster than females (0.076 cm/yr ± 0.87, n = 359).Growth rates of tortoises whose mean capture year (average ofinitial capture and recapture year) fell within the early phaseof Project Isabela (1995–1999) were lower than those withinthe later phase (2000–2005) (Table 1). On Santa Cruz Islandduring the same time frame, 103 tortoises were recaptured ineither “el chato” or “las fincas” on average 2.9 years apartand grew an average of 0.88 cm/year (±1.2SD). Only 32.9%of deviance in growth rates was explained by age and sex(Table 1). Juveniles grew much faster (2.15 cm/year ± 1.30)than males (0.96 cm/year ± 1.12), which in turn grew fasterthan females (0.27 cm/year ± 0.77). Growth rates of tortoiseson Santa Cruz were independent of phase of Project Isabela(Table 1).
Our suite of independent variables accounted for virtuallyall variation (96%) in tortoise body mass on Alcedo Volcano(Table 1); after controlling for the inherent size–mass relation-ship, females were relatively heavier than juveniles and males(marginally so: p = 0.07), and tortoises were relatively lighterduring the initial than final project phase.
Discussion
Observed changes in the demographic attributes of the gianttortoise population on Alcedo Volcano versus the controlpopulation on Santa Cruz were consistent with a positiveresponse by tortoises to goat and donkey eradication. Notably,juvenile tortoises were virtually undetected on Alcedo Volcanoduring the initial phase of Project Isabela but reappearedduring the project’s final phase to represent nearly a quarterof all tortoises encountered. This increase in representation ofjuveniles is likely underestimated given that rapid regrowthof vegetation following goat removal would likely havedecreased probability of detection of juvenile tortoises by
Restoration Ecology 3
Giant Tortoise-Feral Goat Interactions
Table 1. Summary of general additive mixed models used to examine giant tortoise rates of shell growth and body mass in relation to tortoise size(carapace length), tortoise age and sex, season (wet vs. dry, on Alcedo Volcano for condition analysis), and phase of the goat and donkey eradicationcampaign associated with Project Isabela on Alcedo Volcano, Galapagos.
Parametric Coefficients Nonlinear Effects (Nonparametric)
Parameter Estimate Standard Error T p df F p
Shell growth rate–Alcedo VolcanoConstant 0.200 0.086 2.352 0.018Status (female vs. juvenile) 0.629 0.218 2.882 0.004Status (female vs. male) 1.186 0.158 7.470 0.001Time (1995–1999 vs. 2000–2005) 0.256 0.066 3.843 0.001Carapace length — — — — 1.1 8.073 0.001
Shell growth rate–Santa Cruz Tortoise ReserveConstant −0.671 0.280 −2.398 0.0184Status (female vs. juvenile) 0.508 0.499 −1.019 0.3107Status (female vs. male) 1.196 0.339 5.696 0.001Time (1995–1999 vs. 2000–2005) 0.302 0.199 1.519 0.132Carapace length — — — — 1.1 24.91 0.001
Body mass–Alcedo VolcanoConstant 89.448 0.564 158.56 0.001Status (female vs. juvenile) −2.617 0.864 −3.093 0.002Status (female vs. male) −0.517 0.672 −0.770 0.074Season (wet vs. dry) 4.091 0.667 6.127 0.001Time (1995–1999 vs. 2000–2005) 2.503 0.494 5.064 0.001Carapace length — — — — 8.607, 8.957 4907 0.001
Santa Cruz Island tortoises, in an area not subject to goat removal, are considered as a “control” population. Deviance explained for somatic growth rates on Alcedo Volcano =60.3% (n = 669 tortoises) and Santa Cruz Island = 32.9% (n = 103 tortoises); that for relative body mass on Alcedo Volcano = 95.7% (n = 6, 389 tortoises).
field personnel. Although donkey removal represented onlyabout 2% numerically of the numbers of goats removedduring Project Isabela, donkey removal may have particularlybenefited younger tortoise age classes given that donkeysdestroy tortoise nests through trampling and rolling (Fowler1983). Increased mass relative to length as well as increasedgrowth rates in individual tortoises during the second projectphase is also notable because improved body condition couldimprove annual survival and also permit female tortoises toshunt more somatic resources from maintenance to growth andreproduction.
We did not measure other relevant indicators of pop-ulation status (cf. Congdon et al. 1993), including age-and sex-specific survival rates (recapture probabilities wereexceedingly low), age at first reproduction, fecundity rates, orpopulation size. Despite the length of this study (11 years ofintensive marking) and the large numbers of individuals sam-pled (8,112 tortoises), age at first reproduction in smaller sad-dlebacked giant tortoises is likely 16–20 years (Marquez et al.1991), while age at first reproduction in the larger domed tor-toises like those on Alcedo Volcano is likely greater. Therefore,lag times in change in population size in response to changes insurvival or reproduction would not likely manifest and hencebe measurable for decades in these animals and hence at timeframes extending beyond our monitoring program.
Factors other than goat and donkey removal could havecontributed to changes observed in tortoise demography onAlcedo Volcano. Notably, during 1997–1998 the secondstrongest El Nino - Southern Oscillation (ENSO) event in thelast 25 years was recorded in Galapagos, with nearly 5 m of
precipitation received over 2 years. ENSO fluctuations impactthe Galapagos ecosystems profoundly (e.g. Desender et al.,1999; Marquez et al. 2008) and effects last years beyondthe actual event. Most relevant from the perspective of gianttortoises on Alcedo Volcano are ENSO-caused increases ingrowth of annual plant species, especially grasses, that are fooditems favored by tortoises (Cayot 1985; Hamann 2001). DuringENSO events adult and juvenile tortoise growth rates arelikely improved due to the hyper-availability of quality forage,whereas tortoise egg incubation success is likely reduceddue to flooding of nests (Marquez et al. 2008). There is noreason to expect, however, that Alcedo Volcano and SantaCruz Island would experience substantially different patternsof precipitation during ENSO events. The improving status oftortoise populations on Alcedo Volcano but not on Santa CruzIsland offers support for the argument that the improvementin population status for Alcedo tortoises was more likely dueto removal of goats and burros.
Implications for practice
• Removal of feral goats can improve the status of large-bodied native herbivores in island environments.
• Monitoring population response of native species toexotic species removal should become a routine part ofmeasuring conservation “success.”
• Factoring occurrence of favorable climate events intoeradication operations can substantially improve utilityof eradication efforts for recovering depleted populationsand restoring degraded island ecosystems.
4 Restoration Ecology

Giant Tortoise-Feral Goat Interactions
Acknowledgments
This work was supported and funded by the GalapagosNational Park Service, Charles Darwin Foundation, Inc. (nowGalapagos Conservancy), Darwin Initiative for BiodiversityConservation (U.K. Government), European Union, FrankfurtZoological Society, and the Global Environment Facility. M.Chaloupka kindly offered statistical advice and S. Blake andtwo anonymous reviewers commented on the manuscript. Thework would not have been possible without the work anddedication of many “guardaparques” of the Galapagos NationalPark Service.
LITERATURE CITED
Blake, S., M. Wikelski, F. Cabrera, A. Guezou, M. Silva, E. Sadeghayobi, C.Yackulic, and P. Jaramillo. Seed dispersal by giant Galapagos tortoises.Journal of Biogeography (in press).
Caccone, A., G. Gentile, J. P. Gibbs, T. H. Fritts, H. L. Snell, J. Betts, andJ. R. Powell. 2002. Phylogeography and history of giant galapagostortoises. Evolution 56:2052–2066.
Carrion, V., C. J. Donlan, K. J. Campbell, C. Lavoie, and F. Cruz. 2011.Archipelago-wide island restoration in the Galapagos Islands: Reducingcosts of invasive mammal eradication programs and reinvasion risk. PLoSOne 6:e18835.
Cayot, L. J. 1985. Effects of El Nino on giant tortoises and their environment.El Nino in the Galapagos islands: the 1982–1983 event. FundacionCharles Darwin para las islas Galapagos, Quito, Ecuador.
Chaloupka, M. Y., and C. J. Limpus. 1997. Robust statistical modelling ofhawksbill sea turtle growth rates (southern Great Barrier Reef). MarineEcology Progress Series 146:1–8.
Congdon, J. D., A. E. Dunham, and R. C. Van Loben Sels. 1993. Delayedsexual maturity and demographics of blanding turtles (Emydoideablandingi i), Implications for conservation and management of long-livedorganisms. Conservation Biology 7:826–833.
Courchamp, F., J.-L. Chapuis, and M. Pascal. 2003. Mammal invaderson islands: Impact, control and control impact. Biological Reviews78:347–383.
Crawley, M. J. 2007. The R book. John Wiley & Sons, West Sussex, UnitedKingdom.
Desender, K., L. Baert, J.-P. Maelfait, and P. Verdyck. 1999. Conservation onVolcan Alcedo (Galapagos): terrestrial invertebrates and the impact ofintroduced feral goats. Biological Conservation 87:303–310.
Donlan, C., K. Campbell, W. Cabrera, C. Lavoie, V. Carrion, and F. Cruz.2007. Recovery of the Galapagos Rail (Laterallus spilonotus) fol-lowing the removal of invasive mammals. Biological Conservation138:520–524.
Duffy, D. C. 1981. Ferals that failed. Noticias de Galapagos 33:21–22.Donlan, C. J., and C. Wilcox. 2008. Integrating invasive mammal eradications
and biodiversity offsets for fisheries bycatch: conservation opportuni-ties and challenges for seabirds and sea turtle. Biological Invasions10:1053–1060.
Fowler, L. 1983. The population and feeding ecology of tortoises and feral bur-ros on Volcano Alcedo, Galapagos Islands. Ph.D. disseration, Universityof Florida. Pages 150.
Freire, M. 1992. Ecología de los chivos ferales (Capra hircus L.) en elVolcan Alcedo, Isla Isabela, Galapagos, Ecuador. “Licenciado” Thesis,Universidad Tecnica de Ambato, Ambato, Ecuador.
Gibbs, J. P., C. Marquez, and E. J. Sterling. 2008. The role of endangeredspecies reintroduction in ecosystem restoration: Tortoise-cactus interac-tions on Espanola Island, Galapagos. Restoration Ecology 16:88–93.
Gibbs, J. P., E. J. Sterling, and F. J. Zabala. 2009. Giant tortoises as ecologicalengineers: a long-term quasi-experiment in the Galapagos Islands.Biotropica 48:208–214.
Hamann, O. 1993. On vegetation recovery, goats and giant tortoises on PintaIsland, Galapagos, Ecuador. Biodiversity and Conservation 2:138–151.
Hamann, O. 2001. Demographic studies of three indigenous stand-formingplant taxa (Scalesia, Opuntia, and Bursera) in the Galapagos Islands,Ecuador. Biodiversity and Conservation 10:223–250.
Lavoie, C., C. J. Donlan, K. Campbell, F. Cruz, and G. V. Carrion. 2007. Geo-graphic tools for eradication programs of insular non-native mammals.Biological Invasions 9:139–148.
MacFarland, C. 1991. Goats on Alcedo volcano in the Galapagos Islands - helpneeded. Conservation Biology 5:6–7.
Marquez, C., G. Morillo, and L. J. Cayot. 1991. Pages 17–18. A 25-year management program pays off: repatriated tortoises on Espanolareproduce. Noticias de Galapagos.
Marquez, C., D. A. Wiedenfeld, S. Naranjo, and W. Llerena. 2008. Pages7–10. The 1997-8 El Nino and the Galapagos tortoises Geochelonevandenburghi on Alcedo volcano. Galapagos Research, Galapagos.
Seminoff, J. A., A. Resendiz, W. J. Nichols, and T. T. Jones. 2002. Growthrates of wild green turtles (Chelonia mydas) at a temperate foraging areain the Gulf of California, Mexico. Copeia 610–617.
Towns, D. R., I. A. E. Atkinson, and C. H. Daugherty. 2006. Have the harmfuleffects of introduced rats on islands been exaggerated? BiologicalInvasions 8:863–891.
Zar, J. H. 1984. Biostatistical analysis. 2nd edition. Prentice Hall, EnglewoodCliffs, New Jersey.
Restoration Ecology 5


















