
Plant Nutrition 1: Membrane energetics and transport ... · Cellular and tissue distribution of...
96
© 2014 American Society of Plant Biologists Plant Nutrition 1: Membrane energetics and transport, potassium nutrition and sodium toxicity
Transcript of Plant Nutrition 1: Membrane energetics and transport ... · Cellular and tissue distribution of...

© 2014 American Society of Plant Biologists
Plant Nutrition 1: Membrane
energetics and transport,
potassium nutrition and sodium
toxicity

© 2014 American Society of Plant Biologists
Outline
• Introduction to plant nutrition
• Overview of nutrient uptake and transport
• Energizing the membrane
• H+-ATPases and H+-PPases
• Potassium
• Uptake, transport and homeostasis
• Sodium
• Toxicity, transport and tolerance

© 2014 American Society of Plant Biologists
Plant nutrition: Introduction
44% Oxygen
42% Carbon
CO2,
photo-
synthesis
7% Hydrogen
H2O water
93% of plant
dry mass is
composed of
C, O and H
Plants are ~70 to
>90% water by
weight
7% Other,
from soil
K
Potassium
N
Nitrogen
P Phosphorus
Ca Calcium
Mg Magnesium
S Sulfur
Si Silicon
Cl Chlorine
Other
These elements are
obtained mainly from soil,
are often referred to as
mineral nutrients, and are
the subject of the topic
Plant Nutrition

© 2014 American Society of Plant Biologists
Plants assimilate mineral nutrients
from their surroundings
K+
K+
PO43-
PO43-
PO43-
NO3-
NO3-
K+ K+
K+
K+
K+
K+
PO43-
PO43-
PO43-
NO3-
NO3-
Nutrient assimilation is very
energetically demanding –
the nutrients have to be
moved against a
concentration gradient and
often a charge gradient
Algae
Bryophytes
Tracheophytes

© 2014 American Society of Plant Biologists
Nutrient uptake, assimilation and
utilization involve many processes Nutrient
acquisition
efficiency
Nutrient usage
efficiency
Root system
architecture
Root
exudates
Rhizosphere
microbiota
Symbioses
P
P
N N
NH3
Transporters
and pumps
Intercellular
transport
efficiency
X R-X
Assimilation and
remobilization
efficiency
Regulatory and
homeostatic
networks

© 2014 American Society of Plant Biologists
Nutrients removed from soils can be
replenished with fertilizers
Total nutrient requirement
Typical fertilizer application
Corn
Soy W
he
at
Co
tto
n
Ric
e
Kg
/ha
K
g/h
a
1000
800
600
400
200
0
0
200
400
Nitrogen
Phosphate
Potash
Magnesium
Sulfur
Most fertilizers
contain nitrogen
(N), phosphorus
(P) and potassium
(K). Some include
other elements
Fertilizers can be
complex waste
products or
refined blends of
nutrient salts
Plants remove nutrients from the soil
Source: USGS

© 2014 American Society of Plant Biologists
Global mineral nutrient resources
are unevenly distributed
Supply > Demand
Supply < Demand
FAO (2011) Current world fertilizer trends and outlook to 2015.
N
P2O5
K2O

© 2014 American Society of Plant Biologists
The global trade in fertilizers is
worth billions of dollars annually
IFIA
Ammonium Urea Potash Diammonium
phosphate
Monoammonium
phosphate
Phosphate
rock
Sulfur Sulfuric
acid

© 2014 American Society of Plant Biologists
How much is the right amount of
fertilizer to apply to a field?
Photo by Michael Russelle.
Species / variety
of plant: Different
plants have
different needs
Soil
characteristics:
Residual nutrients,
rate of nutrient
leaching, pH,
particle size,
presence of
microbes etc. affect
optimal application
Cultivation
practices: Is
unharvested
material removed,
or left to replenish
the soil?
Abiotic and biotic
factors: Temperature,
rain, stress and pests
or pathogens affect
nutrient needs
Developmental stage affects plant needs
Interactions between nutrients:
There are both positive and negative
interactions between various nutrients
Financial considerations:
Balancing the cost of fertilizers with
the gain reaped from their use

© 2014 American Society of Plant Biologists
Fertilizer use can cause
environmental and health problems Nitrogen fixation is
energy demanding
Phosphate and potash
mining is destructive
Image source: Lamiot; Alexandra Pugachevsky
Transport requires energy
Human and animal waste
can spread disease N O N
Nitrous oxide (N2O)
derived from fertilizer is a
major greenhouse gas
Nutrient runoff pollutes
waterways and can lead
to eutrophication
Plants need
nutrients, but their
application isn’t
always optimal or
sustainable – how
can plant science
contribute to
better practices?

© 2014 American Society of Plant Biologists
Pumps, channels
and carriers are the
molecular
mediators of these
processes
Membrane transport can
consume 1/3 of a cell’s
metabolic energy (or more)
Nutrients must be
transported across
membranes to enter
the plant
Cross membrane
into living cell in
root hair Symplastic or
transcellular pathway
Apoplastic pathway
Cross membrane
into living cell in
at endodermis
Bidirectional transport
between xylem parenchyma
cells and apoplastic
transpiration stream
Casparian strip
Nutrient uptake and transport:
Overview

© 2014 American Society of Plant Biologists
Plants assimilate mineral nutrients
mainly as cations or anions
μmol / g (dry wt)
Element Assimilated form
250 Potassium (K) K+
1000 Nitrogen (N) NO3-, NH4
+
60 Phosphorus (P)
HPO42-,
H2PO4-
30 Sulfur (S) SO42-
80 Magnesium (Mg)
Mg2+
125 Calcium (Ca) Ca2+
μmol / g (dry wt)
Element Assimilated form
2 Iron (Fe) Fe3+, Fe2+
0.002 Nickel (Ni) Ni2+
1 Manganese (Mn)
Mn2+
0.1 Copper (Cu) Cu2+
0.001 Molybdenum (Mo)
MoO42+
2 Boron (B) H3BO3
3 Chlorine (Cl) Cl-
0.3 Zinc (Zn) Zn2+
MACRONUTRIENTS MICRONUTRIENTS
See Taiz, L. and Zeiger, E. (2010) Plant Physiology. Sinauer Associates; Marschner, P. (2012) Mineral Nutrition of Higher Plants. Academic Press, London

© 2014 American Society of Plant Biologists
Nutrients are concentrated in the
plant relative to the environment
[K+]o
0.1 – 1 mM
[H2PO4-]o
[HPO42-]o
< 1 μM
[NO3-]o
<100 μM –
>1 mM
[NH4+]o
<100 μM –
>1 mM
[K+]i 50 - 100 mM
Soil abundance
(ranges or typical
values)
Cell [H2PO4
-]i [HPO4
2-]i
5 - 10 mM
[NO3-]i
10 mM
[NH4+]i
~1 mM
Energy is expended to assimilate nutrients
against a steep concentration gradient The driving force of
the nutrient’s
chemical gradient is
outwards

© 2014 American Society of Plant Biologists
Transport can be down or against an
electrochemical gradient Down an electrochemical gradient
(Diffusion or facilitated diffusion)
Through
membrane
Through
channel
Through
carrier
Against an electrochemical gradient
(Active transport)
ATP ADP + Pi
Primary active transport:
Directly coupled to ATP
hydrolysis
Secondary active
transport: Indirectly
coupled to ATP
hydrolysis
Symport Antiport
OUT
IN

© 2014 American Society of Plant Biologists
Solutes cross membranes through
different types of transporters
X
X
Pumps are often drawn
like lollipops, with a large
cytoplasmic catalytic
domain
Pumps: • Move solutes against a
chemical or charge gradient
• Couple transport to hydrolysis
of ATP or pyrophosphate
ATP ADP
ATP
ADP
The multisubunit vacuolar
proton pump VH+-ATPase
Reprinted from Schumacher, K. and Krebs, M. (2010). The V-ATPase: small cargo, large effects. Curr. Opin. Plant Biol. 13: 724-730 with permission from Elsevier.

© 2014 American Society of Plant Biologists
Solutes cross membranes through
different types of transporters
X
X X
Channels are often
drawn as two adjacent
ovals (or a cross-section
of a doughnut)
Channels: • are protein-formed holes in the membrane
• can be open or closed
• move one type of solute at a time
• do not provide an energy source for the
movement; solutes can only move down
their electrochemical gradient
Reprinted from Long, S.B., Campbell, E.B. and MacKinnon, R. (2005). Crystal structure of a mammalian voltage-dependent Shaker family K+ channel. Science. 309: 897-903 with permission from AAAS.
Side (L) and Top (R) views
of a potassium channel

© 2014 American Society of Plant Biologists
Solutes cross membranes through
different types of transporters
Carriers /
Coupled Transporters • are membrane proteins
• can be active or inactive
• can move more than one solute at a time
• The driver (usually H+ in plants) moves down
its electrochemical gradient, which provides
the energy for the co-transported solute’s
transport
X
X
Coupled transporters
are often drawn as
circles with arrows
indicating the direction
of flow for each ion
H+ H+
X
H+
Schematic domain structure
(L) and Top-down (R) views
of an HKT1 Na+ transporter
Cotsaftis, O., Plett, D., Shirley, N., Tester, M. and Hrmova, M. (2012). A two-staged model of Na+ exclusion in rice explained by 3D modeling of HKT transporters and alternative splicing. PLoS ONE. 7: e39865. Chérel,
I., Lefoulon, C., Boeglin, M. and Sentenac, H. (2014). Molecular mechanisms involved in plant adaptation to low K+ availability. J. Exp. Bot. 65: 833-848, by permission of Oxford University Press.

© 2014 American Society of Plant Biologists
Pumps, channels and carriers are
also involved in nutrient distribution
Reprinted from Ahmad, I. and Maathuis, F.J.M. (2014). Cellular and tissue distribution of potassium: Physiological relevance, mechanisms and regulation. J. Plant Physiol. 171: 708–714 with permission from Elsevier.
• Nutrient uptake is
just the first step
• The assimilated
nutrients have to be
transported to
where they are
needed, including
leaves and seeds
• The vacuole is an
important storage
compartment
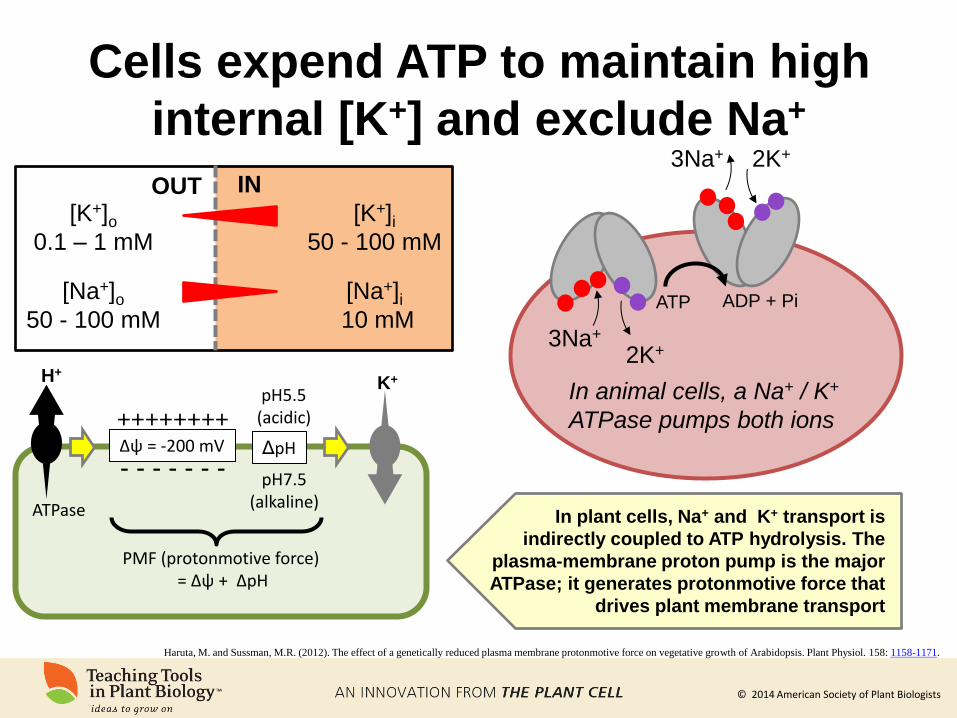
© 2014 American Society of Plant Biologists
Cells expend ATP to maintain high
internal [K+] and exclude Na+
3Na+
2K+ 3Na+
2K+
In animal cells, a Na+ / K+
ATPase pumps both ions
In plant cells, Na+ and K+ transport is
indirectly coupled to ATP hydrolysis. The
plasma-membrane proton pump is the major
ATPase; it generates protonmotive force that
drives plant membrane transport
[K+]o
0.1 – 1 mM
[K+]i 50 - 100 mM
[Na+]o
50 - 100 mM
[Na+]i 10 mM
OUT IN
ATP ADP + Pi
H+ K+
ATPase
Δψ = -200 mV ΔpH
pH5.5 (acidic)
pH7.5 (alkaline)
PMF (protonmotive force) = Δψ + ΔpH
++++++++
- - - - - - -
Haruta, M. and Sussman, M.R. (2012). The effect of a genetically reduced plasma membrane protonmotive force on vegetative growth of Arabidopsis. Plant Physiol. 158: 1158-1171.

© 2014 American Society of Plant Biologists
The membrane as a battery:
Mitchell’s big idea
Mitchell was awarded the Nobel
Prize in Chemistry in 1978
Energy used to
form gradient Energy released
by dissipating
gradient; this
energy can
perform work
Peter Mitchell recognized the
importance of membrane-
bound ATPases in
membrane energetics. In this
drawing he highlights the
reversibility of the ATPase
Reprinted from Mitchell, P. (1961). Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature 191: 144-148.

© 2014 American Society of Plant Biologists
Plant cells have a membrane
potential (Em) -100 to -200 mV
0 mV
+100 mV -100 mV
Reference
electrode Recording
electrode
+
+ +
+
+ - - - -
Voltmeter
The electrical gradient is
maintained by the outward
pumping of protons (H+),
and the accumulation of
anions such as malate in
the cytosol
H+
Malate

© 2014 American Society of Plant Biologists
The electrochemical gradient is
important for ion transport
[K+]o
0.1 – 1 mM
[H2PO4-]o
[HPO42-]o
< 1 μM
[NO3-]o
<100 μM –
>1 mM
[NH4+]o
<100 μM –
>1 mM
[K+]i 50 - 100 mM
[H2PO4-]i
[HPO42-]i
5 - 10 mM
[NO3-]i
10 mM
[NH4+]i
~1 mM
The cell’s electrical gradient
drives anions OUT and cations IN
The electrochemical gradient
defines the energetic demands for
transport, and integrates the
electrical and concentration
gradients
Em = ~ -150 mV

© 2014 American Society of Plant Biologists
Nutrient transport requires energy
and selective transporters
Plant cells pump protons
out to make proton and
charge gradients - -
-
+
+
+
K+
K+
H+
Proton (H+) gradient
Charge gradient
The electrical (charge) and
proton gradients drive the
movements of ions across
membranes through selective
transporters

© 2014 American Society of Plant Biologists
Energizing the membrane: Plant H+-
ATPases and H+-PPases
Sze, H., Li, X. and Palmgren, M.G. (1999). Energization of plant cell membranes by H+-pumping ATPases: Regulation and biosynthesis. Plant Cell. 11: 677-689.
The plasma-membrane
H+-ATPase uses energy
from ATP to pump
protons out of the cell
The vacuolar-type proton
pumps transport protons into
the lumen of endomembrane
compartments (e.g., vacuole)
The H+-PPase uses energy
stored in pyrophosphate
The VH+-ATPase
is a multimeric
protein complex

© 2014 American Society of Plant Biologists
The PM H+-ATPase is a “master
enzyme” and “powerhouse”
See Palmgren, M.G. (2001). Plant plasma membrane H+-ATPases: Powerhouses for nutrient uptake. Annu. Rev. Plant Physiol. Plant Mol.
Biol. 52: 817-845; Figure adapted from Michelet, B. and Boutry, M. (1995). The plasma membrane H+-ATPase. Plant Physiol. 108: 1-6.
ADP H+ ATP
pH ~ 5 - 6
pH ~ 7.5 - - - - -
~ -150 mV
+ + + + +
By pumping protons out
of the cell, PM H+-
ATPases produce electric
and pH gradients
Cations Anions
Channels Antiporters
H+ H+
Symporters Uniporters The electrochemical
gradient produced by the
PM H+-ATPase drives other
transport processes

© 2014 American Society of Plant Biologists
PM H+-ATPases were first
characterized in fungi
Goffeau, A. and Slayman, C.W. (1981). The proton-translocating ATPase of the fungal plasma membrane. Biochim. Biophys. Acta (BBA) 639: 197-223 by permission of Elsevier; Serrano, R., Kielland-Brandt, M.C. and
Fink, G.R. (1986). Yeast plasma membrane ATPase is essential for growth and has homology with (Na+ + K+), K+- and Ca2+-ATPases. Nature. 319: 689-693; Hager, K.M., Mandala, S.M., Davenport, J.W., Speicher,
D.W., Benz, E.J. and Slayman, C.W. (1986). Amino acid sequence of the plasma membrane ATPase of Neurospora crassa: deduction from genomic and cDNA sequences. Proc. Natl. Acad. Sci. USA. 83: 7693-7697.
Predicted structure of cloned plasma-
membrane ATPase PMA1 from yeast. White bars indicate regions of homology with
other cation pumps
PM H+-ATPases were
initially cloned from fungi
Detailed physiological studies in
Neurospora characterized the
plasma membrane proton-
translocating ATPase

© 2014 American Society of Plant Biologists
Plant and fungal PM H+-ATPases are
members of a larger family
Reprinted by permission from Macmillan Publishers Ltd from Kühlbrandt, W. (2004). Biology, structure and mechanism of P-type ATPases. Nat. Rev. Mol. Cell Biol. 5: 282-295; see also Baxter, I., Tchieu, J., Sussman,
M.R., Boutry, M., Palmgren, M.G., Gribskov, M., Harper, J.F. and Axelsen, K.B. (2003). Genomic Comparison of P-Type ATPase Ion Pumps in Arabidopsis and Rice. Plant Physiol. 132: 618-628.
Mammalian
Na+/ K+-ATPase
Plant and
fungal plasma
membrane
H+-ATPases
3Na+
2K+ 3Na+
2K+
ATP ADP + Pi
H+

© 2014 American Society of Plant Biologists
Several differentially expressed
genes encode plant PM H+-ATPases
Arango, M., Gévaudant, F., Oufattole, M. and Boutry, M. (2003). The plasma membrane proton pump ATPase: the significance of gene subfamilies. Planta. 216: 355-365.Okumura, M., Inoue, S.-i.,
Takahashi, K., Ishizaki, K., Kohchi, T. and Kinoshita, T. (2012). Characterization of the plasma membrane H+-ATPase in the liverwort Marchantia polymorpha. Plant Physiol. 159: 826-834. DeWitt, N.D.
and Sussman, M.R. (1995). Immunocytological localization of an epitope-tagged plasma membrane proton pump (H+-ATPase) in phloem companion cells. Plant Cell. 7: 2053-2067. See also Okumura, M.,
Takahashi, K., Inoue, S.-i. and Kinoshita, T. (2012). Evolutionary appearance of the plasma membrane H+-ATPase containing a penultimate threonine in the bryophyte. Plant Signal. Behav. 7: 979 - 982.
AHA3 is highly expressed in phloem companion cells
Negative
control
Antibody
stain
Phylogeny showing functional
genes in Arabidopsis (AHA)
and four in the liverwort
Marcantia polymorpha (MpHA)
O
Other PM H+-ATPase-encoding genes are
expressed in other tissues, and many are
upregulated by stress or other factors

© 2014 American Society of Plant Biologists
PM H+-ATPase activity also is
regulated post-transcriptionally
The monomer is a
single polypeptide
with 10 membrane-
spanning domains
Reprinted from Kühlbrandt, W., Zeelen, J. and Dietrich, J. (2002). Structure, mechanism, and regulation of the Neurospora plasma membrane H+-ATPase. Science. 297: 1692-1696 with permission from AAAS;
Ottmann, C., et al. (2007). Structure of a 14-3-3 coordinated hexamer of the plant plasma membrane H+-ATPase by combining X-Ray crystallography and electron cryomicroscopy. Mol. Cell. 25: 427-440.
IN
OUT
ATP ADP + Pi
H+
Post-translational regulation
includes phosphorylation,
interaction with 14-3-3 proteins
and multimer formation

© 2014 American Society of Plant Biologists
Plant PM H+-ATPases are essential
for nutrient uptake and allocation
Sondergaard, T.E., Schulz, A. and Palmgren, M.G. (2004). Energization of transport processes in plants. Roles of the plasma membrane H+-ATPase. Plant Physiol. 136: 2475-2482.
Through their combined
actions, PM H+-ATPases
contribute to the movement of
nutrients throughout the plant
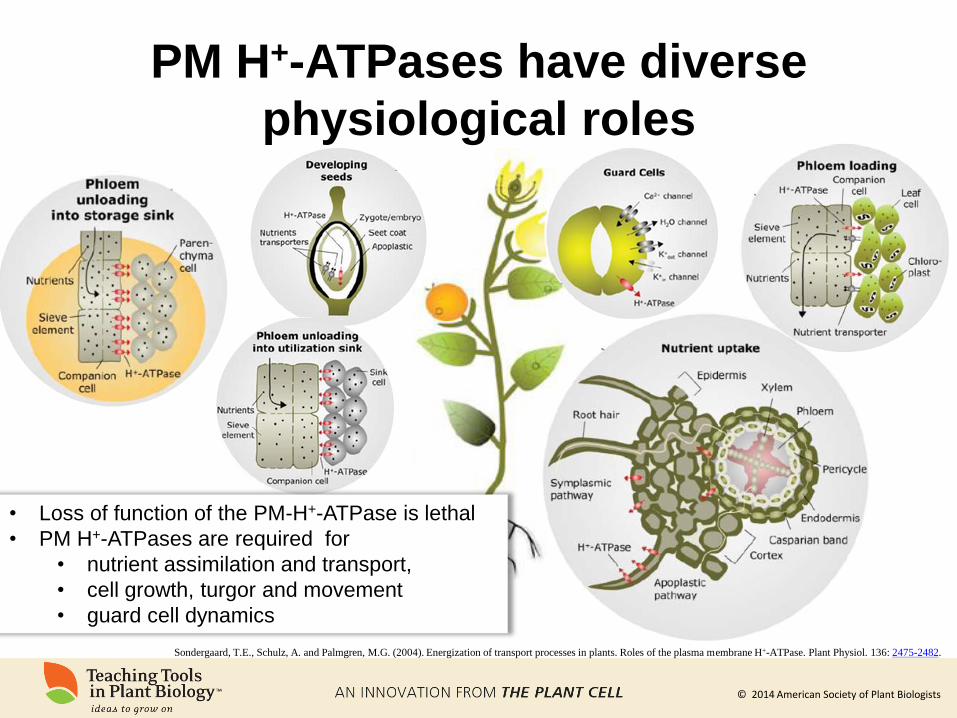
© 2014 American Society of Plant Biologists
PM H+-ATPases have diverse
physiological roles
• Loss of function of the PM-H+-ATPase is lethal
• PM H+-ATPases are required for
• nutrient assimilation and transport,
• cell growth, turgor and movement
• guard cell dynamics
Sondergaard, T.E., Schulz, A. and Palmgren, M.G. (2004). Energization of transport processes in plants. Roles of the plasma membrane H+-ATPase. Plant Physiol. 136: 2475-2482.

© 2014 American Society of Plant Biologists
Vacuolar pumps pump protons into
the vacuole and endocompartments
pH 3 - 6
pH 5 - 6
pH 7.5
Sze, H., Li, X. and Palmgren, M.G. (1999). Energization of plant cell membranes by H+-pumping ATPases: Regulation and biosynthesis. Plant Cell. 11: 677-689.
Isayenkov, S., Isner, J.C. and Maathuis, F.J.M. (2010). Vacuolar ion channels: Roles in plant nutrition and signalling. FEBS letters. 584: 1982-1988.
Em = ~ -30 mV
Protons are pumped into the
vacuole by:
• Vacuolar H+-ATPases (VH+-
ATPases) and
• Vacuolar pyrophosphatases
(H+-PPases)
H+
H+
H+
H+
PPi 2 x Pi
ADP + Pi H+
ATP

© 2014 American Society of Plant Biologists
The VH+-ATPase is a large multi-
subunit enzyme
Schumacher, K. and Krebs, M. (2010). The V-ATPase: small cargo, large effects. Curr. Opin. Plant Biol. 13: 724-730; Reprinted by permission from Macmillan Publishers Ltd from Nishi, T. and Forgac, M. (2002). The vacuolar (H+)-
ATPases — nature's most versatile proton pumps. Nat. Rev. Mol. Cell Biol. 3: 94-103; Ward, J.M. and Sze, H. (1992). Subunit composition and organization of the vacuolar H+-ATPase from oat roots. Plant Physiol. 99: 170-179.
Hydrolyzes ATP to
generate proton gradient Uses proton gradient
to synthesize ATP
VH+-ATPase F-ATPase
VH+-ATPases are related to mitochondrial F-ATPases,
even though their transport directions are reversed
The knob-like V1
domains of VH+-ATPase
are visible on a plant
membrane

© 2014 American Society of Plant Biologists
VH+-ATPases contribute to growth,
salt tolerance & ion uptake / storage
Gogarten, J.P., Fichmann, J., Braun, Y., Morgan, L., Styles, P., Taiz, S.L., DeLapp, K. and Taiz, L. (1992). The use of antisense mRNA to inhibit the tonoplast H+ ATPase
in carrot. Plant Cell. 4: 851-864. Krebs, M., Beyhl, D., Görlich, E., Al-Rasheid, K.A.S., Marten, I., Stierhof, Y.-D., Hedrich, R. and Schumacher, K. (2010). Arabidopsis V-
ATPase activity at the tonoplast is required for efficient nutrient storage but not for sodium accumulation. Proc. Natl. Acad. Sci. USA. 107: 3251-3256.
Phenotypes associated with
decreased VH+-ATPase activity:
• Decreased growth rate
• Male sterility
• Altered nutrient storage
capabilities
Decreased growth rate in carrot
plants expressing an antisense
VH+-ATPase A construct
A mutant lacking functional tonoplast
VH+-ATPase is sensitive to Zn2+ toxicity
(cannot sequester it into vacuole)
© 2014 American Society of Plant Biologists
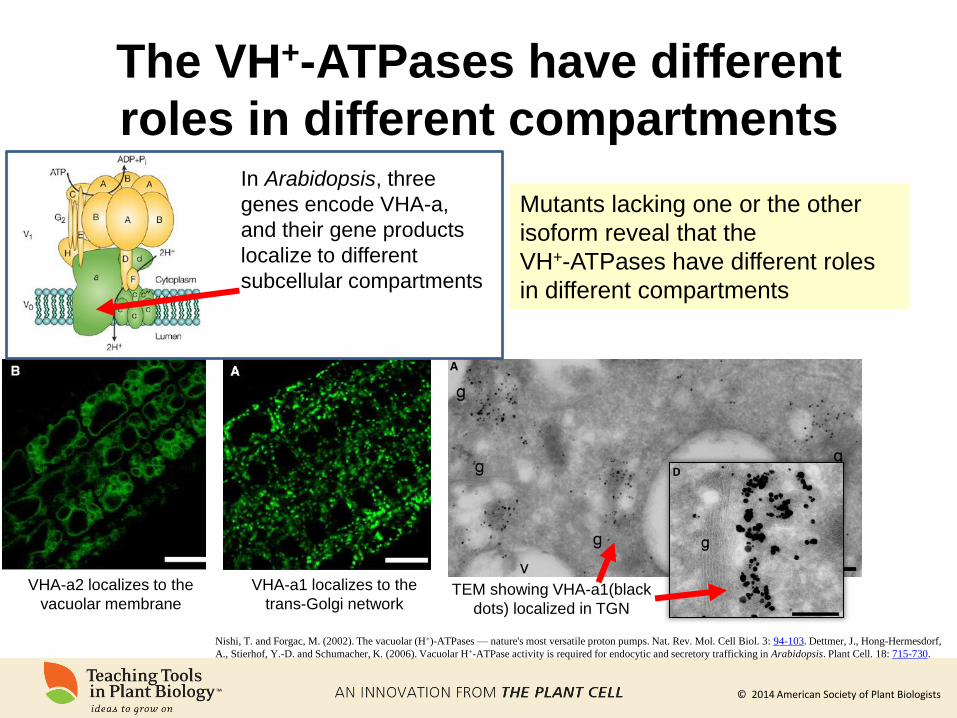
The VH+-ATPases have different
roles in different compartments
VHA-a2 localizes to the
vacuolar membrane
VHA-a1 localizes to the
trans-Golgi network
Mutants lacking one or the other
isoform reveal that the
VH+-ATPases have different roles
in different compartments
In Arabidopsis, three
genes encode VHA-a,
and their gene products
localize to different
subcellular compartments
TEM showing VHA-a1(black
dots) localized in TGN
Nishi, T. and Forgac, M. (2002). The vacuolar (H+)-ATPases — nature's most versatile proton pumps. Nat. Rev. Mol. Cell Biol. 3: 94-103. Dettmer, J., Hong-Hermesdorf,
A., Stierhof, Y.-D. and Schumacher, K. (2006). Vacuolar H+-ATPase activity is required for endocytic and secretory trafficking in Arabidopsis. Plant Cell. 18: 715-730.

© 2014 American Society of Plant Biologists
VH+-ATPases have multiple
functions in plants and animals VH+-ATPases have roles in
pH homeostasis in
endomembranes and also
in membrane trafficking
Functions in plants
• Development
• Cell expansion
• Nutrient assimilation
Functions in humans
• Development
• Kidney function
• Bone resorption
• Tumor cell metastasis
Reprinted from Schumacher, K. and Krebs, M. (2010). The V-ATPase: small cargo, large effects. Curr. Opin. Plant Biol. 13: 724-730 with permission from Elsevier.
© 2014 American Society of Plant Biologists
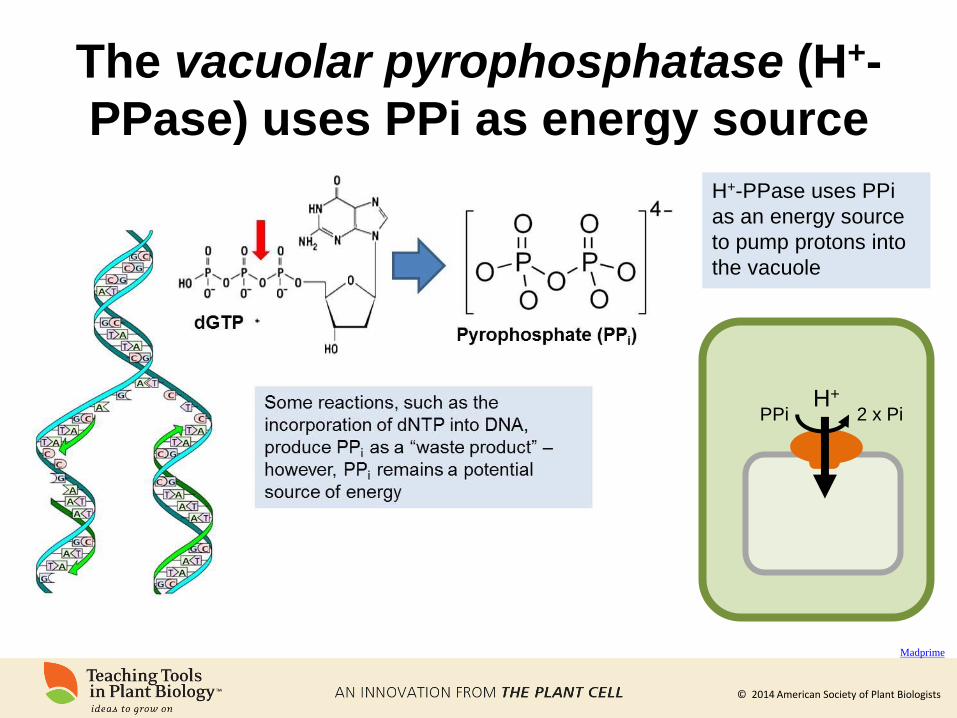
The vacuolar pyrophosphatase (H+-
PPase) uses PPi as energy source
Madprime
H+
PPi 2 x Pi
H+-PPase uses PPi
as an energy source
to pump protons into
the vacuole

© 2014 American Society of Plant Biologists
Plants have 2 types of H+-PPases
Reprinted from Drozdowicz, Y.M. and Rea, P.A. (2001). Vacuolar H+ pyrophosphatases: from the evolutionary backwaters into the mainstream. Trends Plant Sci. 6: 206-211 with permission from Elsevier; Gaxiola, R.A.,
Sanchez, C.A., Paez-Valencia, J., Ayre, B.G. and Elser, J.J. (2012). Genetic manipulation of a “vacuolar” H+-PPase: From salt tolerance to yield enhancement under phosphorus-deficient soils. Plant Physiol. 159: 3-11.
Blue indicates
eubacteria
Red indicates
Archaea
Green indicates
eukaryotes
Eukaryotes with H+-
PPases are limited to
plants and green
algae (circled in
green) and parasitic
protists (circled in
orange)
Type 2:
Golgi-localized
K+ insensitive
Strongly inhibited by Ca2+
Type 1:
Tonoplast-localized
Require K+ for activity
Moderately inhibited by Ca2+

© 2014 American Society of Plant Biologists
Each H+-PPase subunit has 16
membrane spanning domains
PPi
Reprinted by permission from Macmillan Publishers Ltd from Lin, S.-M., Tsai, J.-Y., Hsiao, C.-D., Huang, Y.-T., Chiu, C.-L., Liu, M.-H., Tung, J.-Y., Liu,
T.-H., Pan, R.-L. and Sun, Y.-J. (2012). Crystal structure of a membrane-embedded H+-translocating pyrophosphatase. Nature. 484: 399-403.
H+-PPase functions
as a homodimer
PPi
Structure of a plant H+-PPase
© 2014 American Society of Plant Biologists
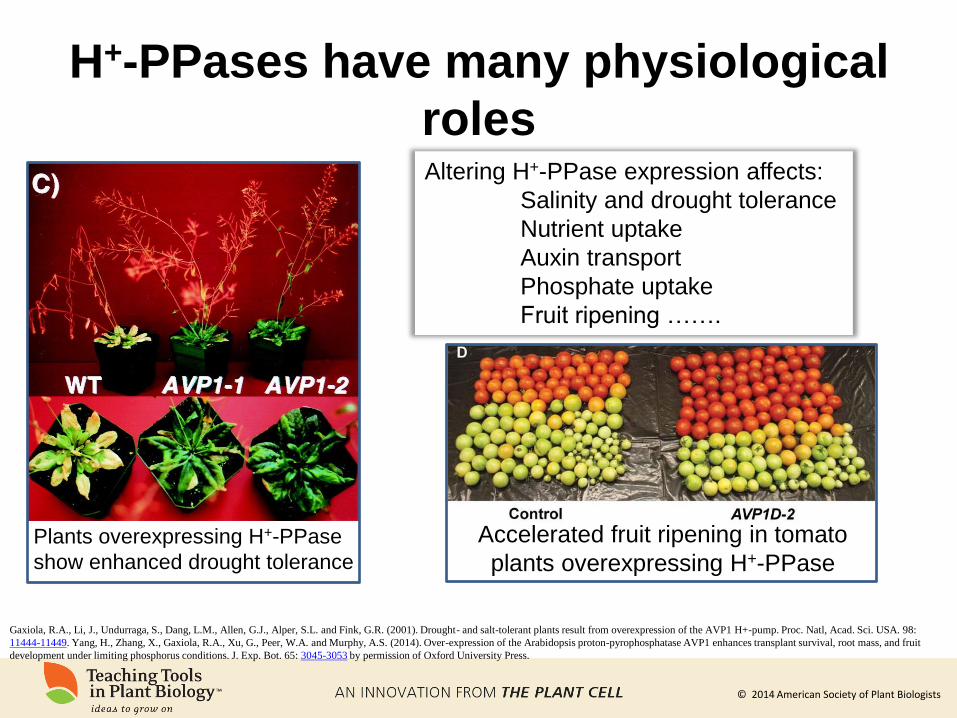
H+-PPases have many physiological
roles
Gaxiola, R.A., Li, J., Undurraga, S., Dang, L.M., Allen, G.J., Alper, S.L. and Fink, G.R. (2001). Drought- and salt-tolerant plants result from overexpression of the AVP1 H+-pump. Proc. Natl, Acad. Sci. USA. 98:
11444-11449. Yang, H., Zhang, X., Gaxiola, R.A., Xu, G., Peer, W.A. and Murphy, A.S. (2014). Over-expression of the Arabidopsis proton-pyrophosphatase AVP1 enhances transplant survival, root mass, and fruit
development under limiting phosphorus conditions. J. Exp. Bot. 65: 3045-3053 by permission of Oxford University Press.
Plants overexpressing H+-PPase
show enhanced drought tolerance
Altering H+-PPase expression affects:
Salinity and drought tolerance
Nutrient uptake
Auxin transport
Phosphate uptake
Fruit ripening …….
Accelerated fruit ripening in tomato
plants overexpressing H+-PPase

© 2014 American Society of Plant Biologists
Energizing the membrane: Summary
• Electrochemical gradients across membranes
(pH and charge) store energy that can be used
to move materials across the membrane
• Plant cells are energized by proton pumps that
reside in the plasma membrane or
endomembranes

© 2014 American Society of Plant Biologists
K+ and Na+ - “The twins”. So alike
yet so different
K
Sodium (Na) and potassium (K):
• Same column of the periodic table
• Both have a single electron in the
outer shell so form monovalent
cations
• Both are very abundant elements
Potassium
deficiency
NaCl
toxicity
And yet, potassium is an
essential nutrient, and
sodium frequently is toxic
Benito, B., Haro, R., Amtmann, A., Cuin, T.A. and Dreyer, I. (2014).The twins K+ and Na+ in plants. J. Plant Physiol. 171: 723–731. FAO

© 2014 American Society of Plant Biologists
Potassium uptake, transport and
homeostasis
Regulates
stomatal
conductance,
photosynthesis
and transpiration
Maintains turgor
and reduces wilting
Strengthens
cell walls Maintains ionic
homeostasis Stimulates
photosynthate
translocation
Enhances
fertility
Promotes stress
tolerance
See Wang, M., Zheng, Q., Shen, Q. and Guo, S. (2013). The critical role of potassium in plant stress response. Intl. J. Mol. Sci. 14: 7370-7390; Sin Chee Tham /Photo; Purdue extension; Onsemeliot.
Symptoms of
potassium deficiency
[K+] in soil = ~0.1 – 1 mM
[K+] in plant cell
cytoplasm = ~100 mM
Potassium is an essential macronutrient
Regulates
enzyme activities

© 2014 American Society of Plant Biologists
Potassium fertilizers are mined from
underground reserves as “potash”
Almost half of the world’s reserved of potash
are found in Saskatchewan, Canada
Potash is a term that encompasses
many forms of potassium:
• KCl (potassium chloride, aka sylvite)
• K2SO4 (potassium sulfate)
• K2CO3 (potassium carbonate)
• K2Ca2Mg(SO4)4·2H2O (polyhalite)
• etc.
Canada Potash; Lmbuga
KCl, sylvite
For historical reasons, potash
is measured in units of K2O
equivalents, even though it is
rarely found in the form of K2O

© 2014 American Society of Plant Biologists
Potash provides K+ for fertilizers,
which supplement natural sources
manure
decomposition
Terrestrial
cycle: Plant /
Animal / Soil Underground reserves
Water with
dissolved K+
salts returned
to surface
Water
pumped
underground
Salts
recovered by
evaporation
90 – 98%
insoluble
minerals
1 – 3%
exchangeable
salts
0.1 – 0.2% soil
solution K+
Potash
fertilizer
application
Adapted from International Potash Institute

© 2014 American Society of Plant Biologists
Potash prices can be volatile and
there are few suppliers
1.06 cm
Canada is #1
in production
(11.2 Mt) and
reserves
(4,400 Mt)
Russia is #2
in production
(7.4 Mt) and
reserves
(3,300 Mt)
Brazil
3.2 Mt
210 Mt
Chile
0.8 Mt
130 Mt
US
1.1 Mt
130 Mt
China
3.2 Mt
210 Mt
Belarus
5.5 Mt
750 Mt
World
reserves
9500 Mt
World
production
(2011)
37 Mt
Jordan
1.4 Mt
40 Mt
Israel
2.0 Mt
40 Mt
Germany
3.3 Mt
150 Mt
Spain
0.4 Mt
20 Mt
UK
0.4 Mt
22 Mt
Adapted from International Potash Institute

© 2014 American Society of Plant Biologists
Potassium is an essential plant
nutrient
Reprinted from Maathuis, F.J.M. (2009). Physiological functions of mineral macronutrients. Curr. Opin. Plant Biol. 12: 250-258 with permission from Elsevier.
K+ uptake
involves high
and low affinity
transporters
K+ is a counter ion for
negatively charged molecules
including DNA and proteins
K+ is a cofactor for
some enzymes
As the major cation in
the vacuole, K+
contributes to cell
expansion and
movement, including
that of guard cells
K+ moves in and
out of the vacuole
through specific
transporters
© 2014 American Society of Plant Biologists
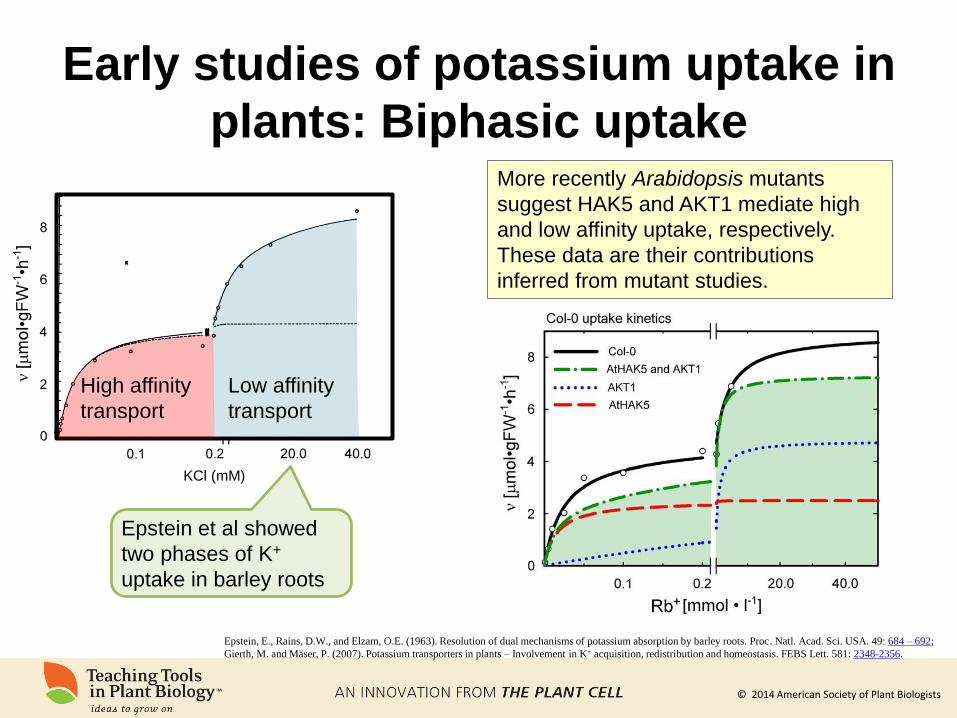
Early studies of potassium uptake in
plants: Biphasic uptake
Epstein, E., Rains, D.W., and Elzam, O.E. (1963). Resolution of dual mechanisms of potassium absorption by barley roots. Proc. Natl. Acad. Sci. USA. 49: 684 – 692;
Gierth, M. and Mäser, P. (2007). Potassium transporters in plants – Involvement in K+ acquisition, redistribution and homeostasis. FEBS Lett. 581: 2348-2356.
KCl (mM)
Low affinity
transport
High affinity
transport
Epstein et al showed
two phases of K+
uptake in barley roots
More recently Arabidopsis mutants
suggest HAK5 and AKT1 mediate high
and low affinity uptake, respectively.
These data are their contributions
inferred from mutant studies.

© 2014 American Society of Plant Biologists
More energy must be expended to
take up K+ when it is scarce
See Britto, D.T. and Kronzucker, H.J. (2008). Cellular mechanisms of potassium transport in plants. Physiol. Plant. 133: 637-650; Nieves-Cordones, M., Alemán, F.,
Martínez, V. and Rubio, F. (2014). K+ uptake in plant roots. The systems involved, their regulation and parallels in other organisms. J. Plant Physiol. 171: 688–695.
K+ uptake from low [K+]ext
requires more energy than
when [K+]ext is higher
K+ K+ H+
H+
ATP
2 x H+
2 x ATP
At very high [K+]ext, non-
selective cation channels
(NSCC) also may contribute
Low affinity
transport
High affinity
transport
© 2014 American Society of Plant Biologists
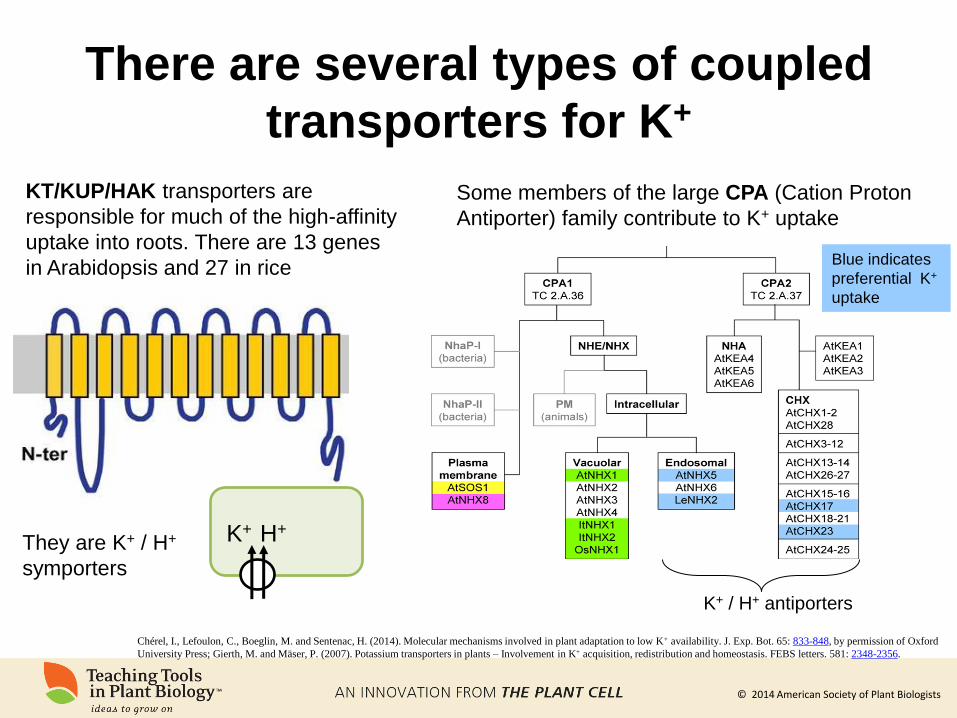
There are several types of coupled
transporters for K+
Chérel, I., Lefoulon, C., Boeglin, M. and Sentenac, H. (2014). Molecular mechanisms involved in plant adaptation to low K+ availability. J. Exp. Bot. 65: 833-848, by permission of Oxford
University Press; Gierth, M. and Mäser, P. (2007). Potassium transporters in plants – Involvement in K+ acquisition, redistribution and homeostasis. FEBS letters. 581: 2348-2356.
c
c KT/KUP/HAK transporters are
responsible for much of the high-affinity
uptake into roots. There are 13 genes
in Arabidopsis and 27 in rice
K+ H+ They are K+ / H+
symporters
Some members of the large CPA (Cation Proton
Antiporter) family contribute to K+ uptake
Blue indicates
preferential K+
uptake
K+ / H+ antiporters

© 2014 American Society of Plant Biologists
There are 15 genes encoding
potassium channels in Arabidopsis
Used with permission from Hedrich, R. (2012). Ion channels in plants. Physiol.Rev. 92: 1777-1811.
9 Shaker-
type (Kv)
(Plasma
membrane)
5 Two Pore K+
(TPK) channels
(mostly tonoplast)
1 Non-selective
two-pore cation
(TPC) channel
1 Kir-like
potassium
channel

© 2014 American Society of Plant Biologists
Shaker channels form as tetramers
that can be heteromers
Hedrich, R. (2012). Ion channels in plants. Physiol.Rev. 92: 1777-1811; Chérel, I., Lefoulon, C., Boeglin, M. and Sentenac, H. (2014). Molecular mechanisms involved in plant adaptation
to low K+ availability. J. Exp. Bot. 65: 833-848, by permission of Oxford University Press; Reprinted from Anschütz, U., Becker, D. and Shabala, S. (2014), Going beyond nutrition:
Regulation of potassium homoeostasis as a common denominator of plant adaptive responses to environment. J. Plant Physiol. 171: 670-687 with permission from Elsevier.
Each monomer has
6 transmembrane
domains
Model of a Shaker K+ channel tetramer with
each monomer shown in a different color
© 2014 American Society of Plant Biologists
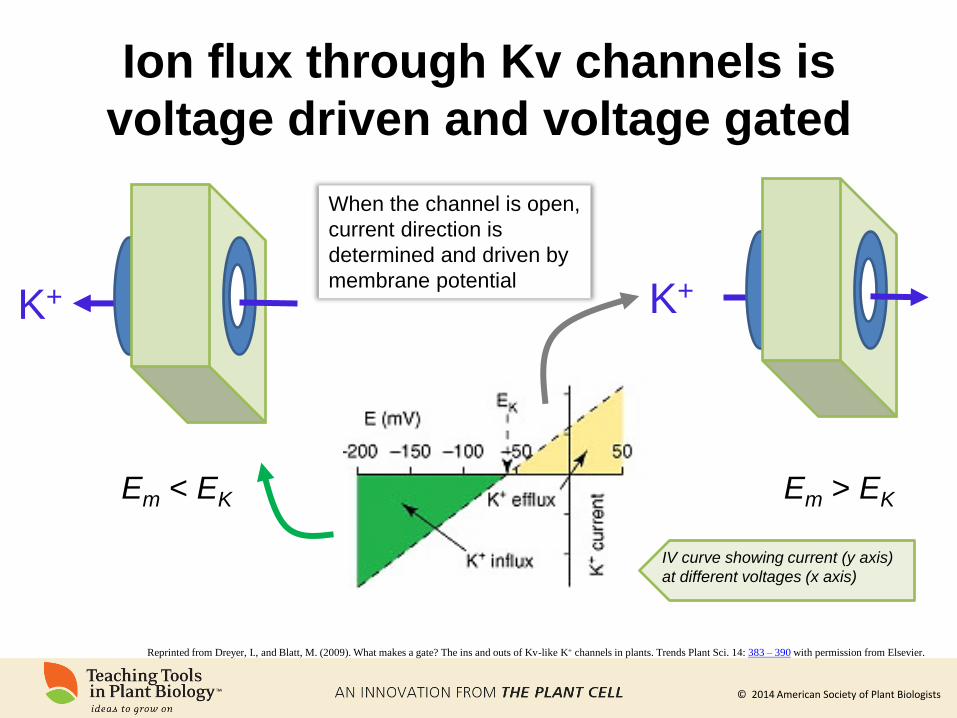
Ion flux through Kv channels is
voltage driven and voltage gated
K+ K+
Em < EK
When the channel is open,
current direction is
determined and driven by
membrane potential
Em > EK
IV curve showing current (y axis)
at different voltages (x axis)
Reprinted from Dreyer, I., and Blatt, M. (2009). What makes a gate? The ins and outs of Kv-like K+ channels in plants. Trends Plant Sci. 14: 383 – 390 with permission from Elsevier.

© 2014 American Society of Plant Biologists
Kv channels are voltage gated: Their
“openness” is voltage sensitive
Voltage-sensitive potassium
channels (Kv) are closed at
some membrane voltages
Inward rectifying channels
are open with a fixed
midpoint voltage V1/2
Notice that the relative
conductance is the relative
probability that the gate is open
V1/2, the half-maximal
activation voltage, is
the key parameter
Outward rectifying
channels are open with a
midpoint voltage that is
[K]out dependent
Reprinted from Dreyer, I., and Blatt, M. (2009). What makes a gate? The ins and outs of Kv-like K+ channels in plants. Trends Plant Sci. 14: 383 – 390 with permission from Elsevier.

© 2014 American Society of Plant Biologists
Current depends on the product of
membrane voltage and gating
Gating Current Voltage
potential x =
Inward
rectifying
channel
Outward
rectifying
channel
Reprinted from Dreyer, I., and Blatt, M. (2009). What makes a gate? The ins and outs of Kv-like K+ channels in plants. Trends Plant Sci. 14: 383 – 390 with permission from Elsevier.

© 2014 American Society of Plant Biologists
Different Shaker-type channels
respond differently to voltage
Although their primary
sequences are similar,
some are voltage-gated and
some not, and some are
inward rectifying and some
outward rectifying
Kout
Ksilent
Kweak
Kin
Reprinted from Dreyer, I., and Blatt, M. (2009). What makes a gate? The ins and outs of Kv-like K+ channels in plants. Trends Plant Sci. 14: 383 – 390 with permission from Elsevier;
Pilot, G., Pratelli, R., Gaymard, F., Meyer, Y. and Sentenac, H. (2003). Five-group distribution of the Shaker-like K+ channel family in higher plants. J. Mol. Evol. 56: 418-434.
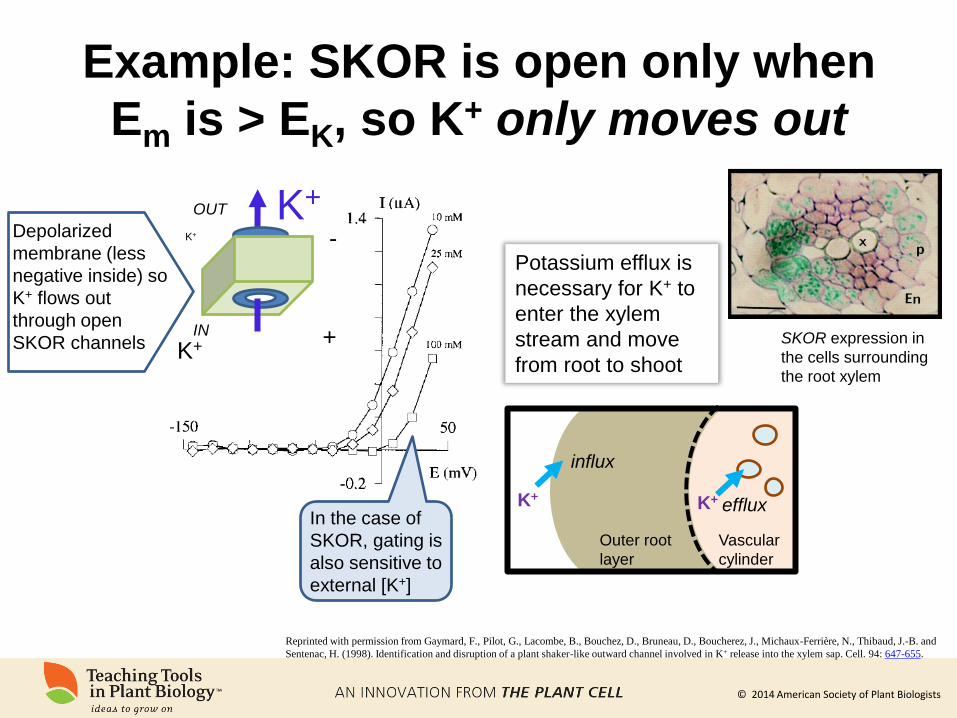
© 2014 American Society of Plant Biologists
Example: SKOR is open only when
Em is > EK, so K+ only moves out
In the case of
SKOR, gating is
also sensitive to
external [K+]
Reprinted with permission from Gaymard, F., Pilot, G., Lacombe, B., Bouchez, D., Bruneau, D., Boucherez, J., Michaux-Ferrière, N., Thibaud, J.-B. and
Sentenac, H. (1998). Identification and disruption of a plant shaker-like outward channel involved in K+ release into the xylem sap. Cell. 94: 647-655.
Outer root
layer
Vascular
cylinder
Potassium efflux is
necessary for K+ to
enter the xylem
stream and move
from root to shoot
Depolarized
membrane (less
negative inside) so
K+ flows out
through open
SKOR channels
K+
K+
-
+
OUT
IN SKOR expression in
the cells surrounding
the root xylem
influx
efflux K+ K+
K+

© 2014 American Society of Plant Biologists
Guard cells are model systems for
the study of K+ transport OPEN CLOSING
H+
H+
H+
H+
H+
H+
K+ K+
KAT
PM-H+-ATPase
V-ATPase
V-PPase
GORK
K+
K+
A-
A-
TPK
TPC
Hills, A., Chen, Z.-H., Amtmann, A., Blatt, M.R. and Lew, V.L. (2012). OnGuard, a computational platform for quantitative kinetic modeling of guard cell physiology. Plant Physiol. 159: 1026-1042 Chen, Z.-H., Hills, A.,
Bätz, U., Amtmann, A., Lew, V.L. and Blatt, M.R. (2012). Systems dynamic modeling of the stomatal guard cell predicts emergent behaviors in transport, signaling, and volume control. Plant Physiology. 159: 1235-1251.
The depth and breadth of
information available for
stomatal guard cells has
made them the premier cell
system in plants for studies
of membrane transport,
signaling, and homeostasis

© 2014 American Society of Plant Biologists
The activity of membrane pumps
and channels is coordinated ABA
K+in
channels K+
out
channels
Anion
channels
H+-PPase V-H+-ATPase
PM-H+-ATPase
V-K+in
channels
V-K+out
channels
V-Anion
channels
[Ca2+]cyt Protein
phosphorylation
pHcyt
Stomata opening Stomata closing
Plasma
membrane
Tonoplast
Adapted from Chen, Z.H., and Blatt, M.R. (2010) Membrane Transport in Guard Cells. In: Encyclopedia of Life Sciences (ELS). John Wiley & Sons, Ltd: Chichester

© 2014 American Society of Plant Biologists
Potassium homeostasis: Responses
to low K+ availability
Adapted from Chérel, I., Lefoulon, C., Boeglin, M. and Sentenac, H. (2013). Molecular mechanisms
involved in plant adaptation to low K+ availability. J. Exp. Bot. 65: 833-848.
Low K
Membrane
hyperpolarization
More efficient
uptake through
K+ channels
Transcriptional
induction of HAK5
K+ channel
Hormonal changes
(auxin, ethylene)
Calcium
signaling
K+ uptake
Enhanced root
growth and
gravitropic
responses
Direct effects
Indirect effects

© 2014 American Society of Plant Biologists
K+ mobilization is critical for K+
homeostasis
Adapted from Amtmann, A., and Leigh, R. (2010). Ion homeostasis. In Abiotic Stress Adaptation in Plants: Physiological, Molecular and Genomic
Foundation, A. Pareek, S.K. Sopory, H.J. Bohnert and Govindjee (eds) (Dordrecht, The Netherlands: Springer), pp. 245 – 262.
Cytosol
Vac.
Supraoptimal K+
can be stored in
the vacuole As K+ becomes
limiting, it becomes
preferentially allocated
to the cytosol

© 2014 American Society of Plant Biologists
K+ mobilization is critical for K+
homeostasis
Cytosol
Vac.
Prioritized
Non-
Prioritized
As K+ becomes
limiting, it becomes
preferentially allocated
to the cytosol
K+ can be remobilized
from less essential tissues
into prioritized tissues
such as growing and
photosynthetic tissues
Adapted from Amtmann, A., and Leigh, R. (2010). Ion homeostasis. In Abiotic Stress Adaptation in Plants: Physiological, Molecular and Genomic
Foundation, A. Pareek, S.K. Sopory, H.J. Bohnert and Govindjee (eds) (Dordrecht, The Netherlands: Springer), pp. 245 – 262.

© 2014 American Society of Plant Biologists
Summary: Potassium uptake,
transport and homeostasis
• Potassium is an essential macronutrient
required in large amounts
• Potassium is transported by channels and
transporters which are regulated transcriptionally
and post-transcriptionally, by membrane voltage
potential, and signals such as pH, Ca2+ and
hormones
• K+ uptake, transport and remobilization are
regulated extensively to ensure that the plant’s
critical tissues are preferentially supported

© 2014 American Society of Plant Biologists
Sodium toxicity, transport and
tolerance
Colum, P. (1918). The Adventures of Odysseus and the Tale of Troy. Project Gutenberg; USDA, USDA, Peggy Greb; FAO
To demonstrate his (fake)
madness, Odysseus plowed
salt into his field
You can’t take salt
out of soil easily;
once it is there it
stays there
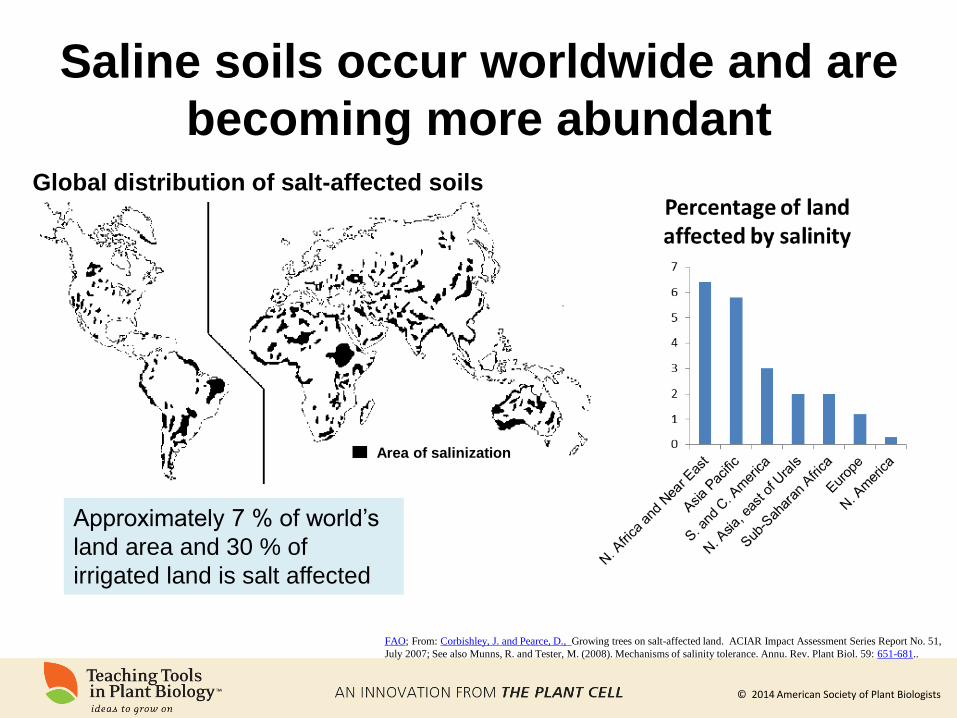
© 2014 American Society of Plant Biologists
Saline soils occur worldwide and are
becoming more abundant
FAO; From: Corbishley, J. and Pearce, D., Growing trees on salt-affected land. ACIAR Impact Assessment Series Report No. 51,
July 2007; See also Munns, R. and Tester, M. (2008). Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 59: 651-681..
Global distribution of salt-affected soils
Approximately 7 % of world’s
land area and 30 % of
irrigated land is salt affected
Area of salinization
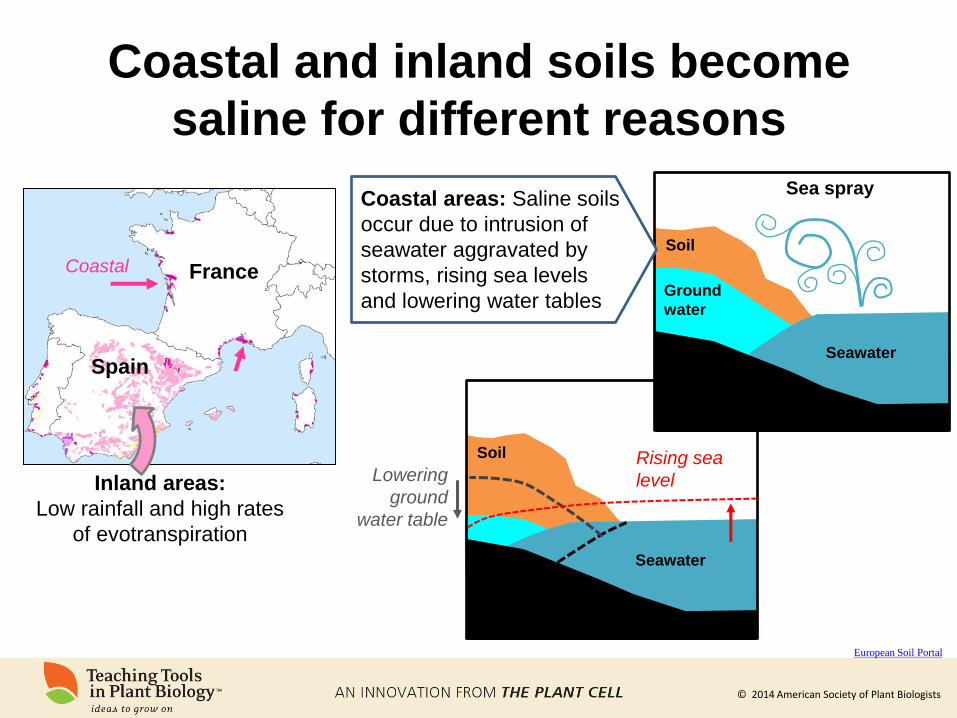
© 2014 American Society of Plant Biologists
Coastal and inland soils become
saline for different reasons
European Soil Portal
Inland areas:
Low rainfall and high rates
of evotranspiration
Seawater
Ground
water
Soil
Sea spray
Seawater
Soil Rising sea
level Lowering
ground
water table
France
Spain
Coastal
Coastal areas: Saline soils
occur due to intrusion of
seawater aggravated by
storms, rising sea levels
and lowering water tables

© 2014 American Society of Plant Biologists
Melting land ice is raising sea levels
and threatening agricultural lands
Reprinted from Nicholls, R.J. and Cazenave, A. (2010). Sea-level rise and its impact on coastal zones. Science. 328: 1517-1520 by permission of AAAS; Vermeer, M. and Rahmstorf, S. (2009). Global
sea level linked to global temperature. Proc. Natl. Acad. Sci. 106: 21527-21532.See also Cazenave, A. and Llovel, W. (2010). Contemporary sea level rise. Annu. Rev. Marine Sci. 2: 145-173. IRRI
Low-lying countries such as Bangladesh are
particularly vulnerable –here a farmer
surveys the damage caused by a cyclone
Se
a leve
l ch
an
ge
(cm
)
Sea levels are
expected to be
>1 m higher by 2100
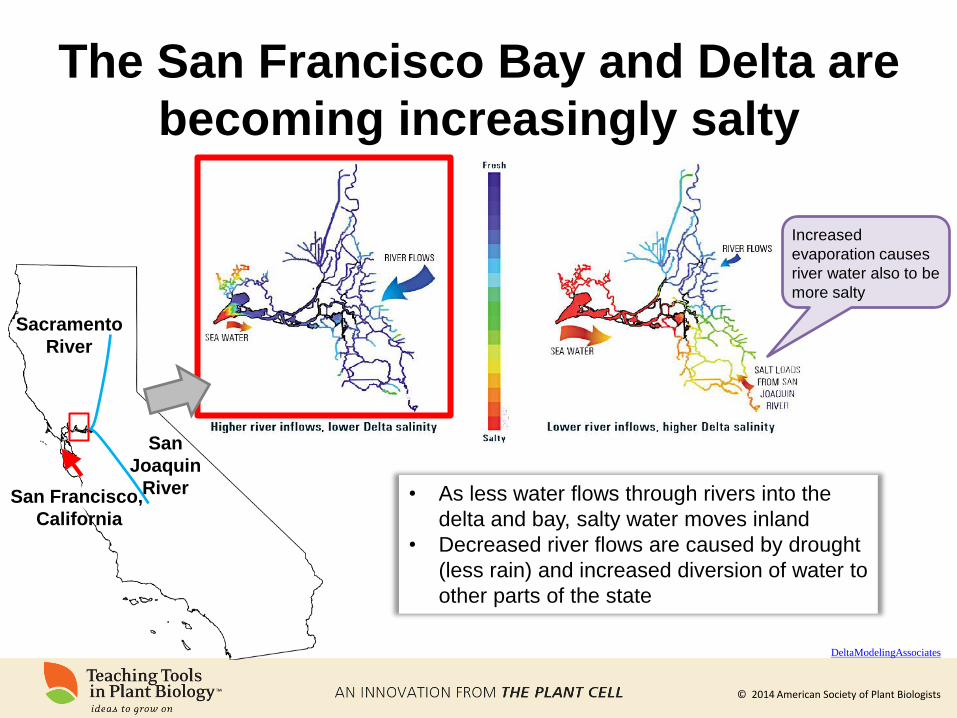
© 2014 American Society of Plant Biologists
The San Francisco Bay and Delta are
becoming increasingly salty
San Francisco,
California
Sacramento
River
San
Joaquin
River • As less water flows through rivers into the
delta and bay, salty water moves inland
• Decreased river flows are caused by drought
(less rain) and increased diversion of water to
other parts of the state
Increased
evaporation causes
river water also to be
more salty
DeltaModelingAssociates
© 2014 American Society of Plant Biologists
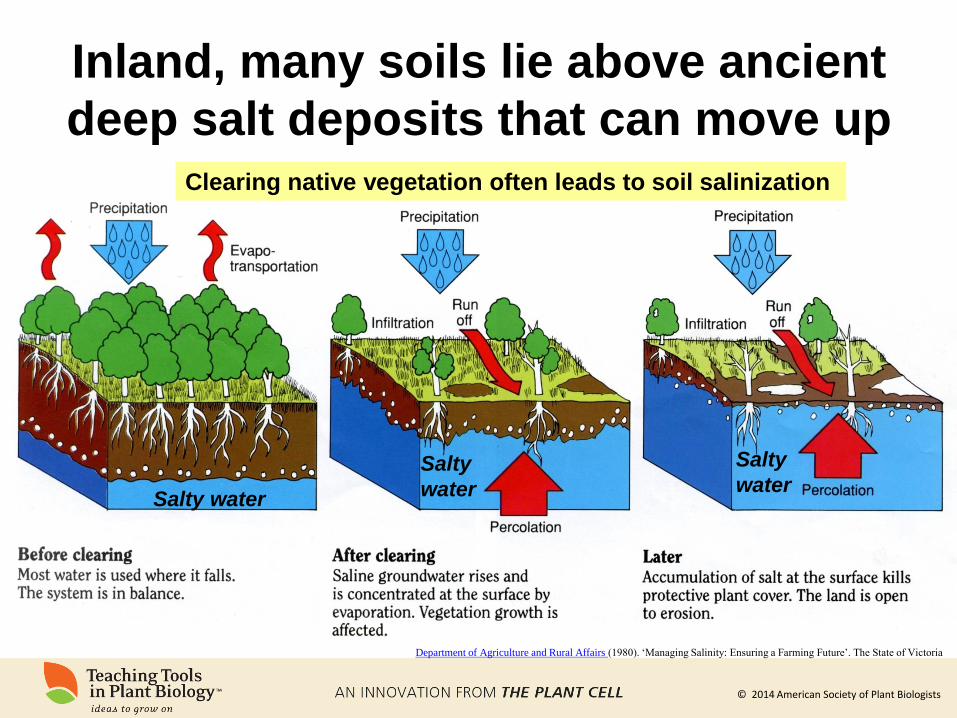
Inland, many soils lie above ancient
deep salt deposits that can move up
Salty water
Salty
water
Salty
water
Department of Agriculture and Rural Affairs (1980). ‘Managing Salinity: Ensuring a Farming Future’. The State of Victoria
Clearing native vegetation often leads to soil salinization

© 2014 American Society of Plant Biologists
Irrigation also contributes to soil
salinity by mobilizing deep salts
Without irrigation
rainwater does not
penetrate below the
rootzone
Excessive irrigation
penetrates into deeper,
salty soils, dissolves the
salts and draws them
upwards into the rootzone
Rain
Evaporation
Rain
Salt
(dissolved)

© 2014 American Society of Plant Biologists
How can we address the problems
caused by soil salinization?
Identify
responses to salt
stress in salt-
sensitive species
(glycophytes)
Introduce salinity-tolerance
traits into crop plants through
breeding and engineering
Identify halophytes
that can be used as
food or energy crops
Munns, R., James, R.A., Xu, B., Athman, A., Conn, S.J., Jordans, C.,
Byrt, C.S., Hare, R.A., Tyerman, S.D., Tester, M., Plett, D. and
Gilliham, M. (2012). Wheat grain yield on saline soils is improved by
an ancestral Na+ transporter gene. Nat Biotech. 30: 360-364.
CSIRO; The State of Victoria; Maurice Chédel; Marco Schmidt
Avoid adding to the
problem by better
management of
fragile soil systems
Areas of concern
Learn about salt
tolerance from
naturally salt-
tolerant species
(halophytes)
Arthrocnemum
macrostachyum
Study salt-tolerant relatives
of crop plants
Chenopodium
quinoa
Thinopyrum
ponticum
Salicornia europaea
Geng, Y., Wu, R., Wee, C.W., Xie, F., Wei, X., Chan, P.M.Y.,
Tham, C., Duan, L. and Dinneny, J.R. (2013). A spatio-temporal
understanding of growth regulation during the salt stress response
in Arabidopsis. Plant Cell. 25: 2132-2154.

© 2014 American Society of Plant Biologists
Plant species have a broad range of
salinity tolerances
Arabidopsis
and rice are
quite sensitive
Saltbush (Atriplex amnicola)
is a halophyte that can
tolerate very salty soil
Reprinted by permission of Annual Reviews from Munns, R. and Tester, M. (2008). Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 59: 651-681.
Q. Can we identify and
exploit the mechanistic
basis of increased
salinity tolerance?
A. YES!
© 2014 American Society of Plant Biologists
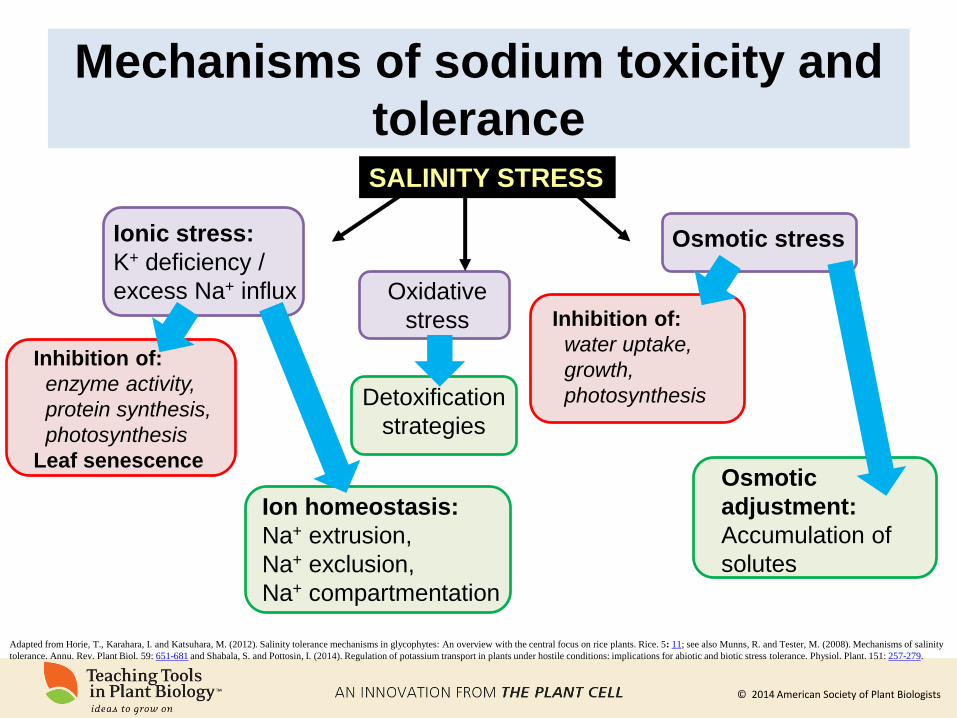
Mechanisms of sodium toxicity and
tolerance
Adapted from Horie, T., Karahara, I. and Katsuhara, M. (2012). Salinity tolerance mechanisms in glycophytes: An overview with the central focus on rice plants. Rice. 5: 11; see also Munns, R. and Tester, M. (2008). Mechanisms of salinity
tolerance. Annu. Rev. Plant Biol. 59: 651-681 and Shabala, S. and Pottosin, I. (2014). Regulation of potassium transport in plants under hostile conditions: implications for abiotic and biotic stress tolerance. Physiol. Plant. 151: 257-279.
SALINITY STRESS
Ionic stress:
K+ deficiency /
excess Na+ influx Inhibition of:
water uptake,
growth,
photosynthesis
Osmotic
adjustment:
Accumulation of
solutes
Inhibition of:
enzyme activity,
protein synthesis,
photosynthesis
Leaf senescence
Ion homeostasis:
Na+ extrusion,
Na+ exclusion,
Na+ compartmentation
Osmotic stress
Oxidative
stress
Detoxification
strategies

© 2014 American Society of Plant Biologists
General sodium tolerance strategy:
Keep sodium out of cytosol & shoot
IN
OUT
“OUT”
1. Keep Na+ from
entering plant / cells
2. Pump out any
Na+ that leaks in
3. Compartmentation
of Na+ in vacuole
4. Extrude Na+
via salt glands
6. Synthesize
compatible solutes for
osmotic balance
Na+
Na+
Na+
Na+
K+ 5. Accumulate K+
to maintain a high
ratio of K+ to Na+
Compatible
solutes
7. Prevent Na+
from moving into
the shoot and
leaves

© 2014 American Society of Plant Biologists
Na+ transport and exclusion is an
integral part of Na+ tolerance
As Na+ becomes more
prevalent, it is preferentially
sequestered into the
vacuole via transporters
Cytosol
Vac.
Na+ can be sequestered in
less essential tissues and
excluded from growing
and photosynthetic tissues
Prioritized
Non-
Prioritized
Adapted from Amtmann, A., and Leigh, R. (2010). Ion homeostasis. In Abiotic Stress Adaptation in Plants: Physiological, Molecular and Genomic
Foundation, A. Pareek, S.K. Sopory, H.J. Bohnert and Govindjee (eds) (Dordrecht, The Netherlands: Springer), pp. 245 – 262.
© 2014 American Society of Plant Biologists
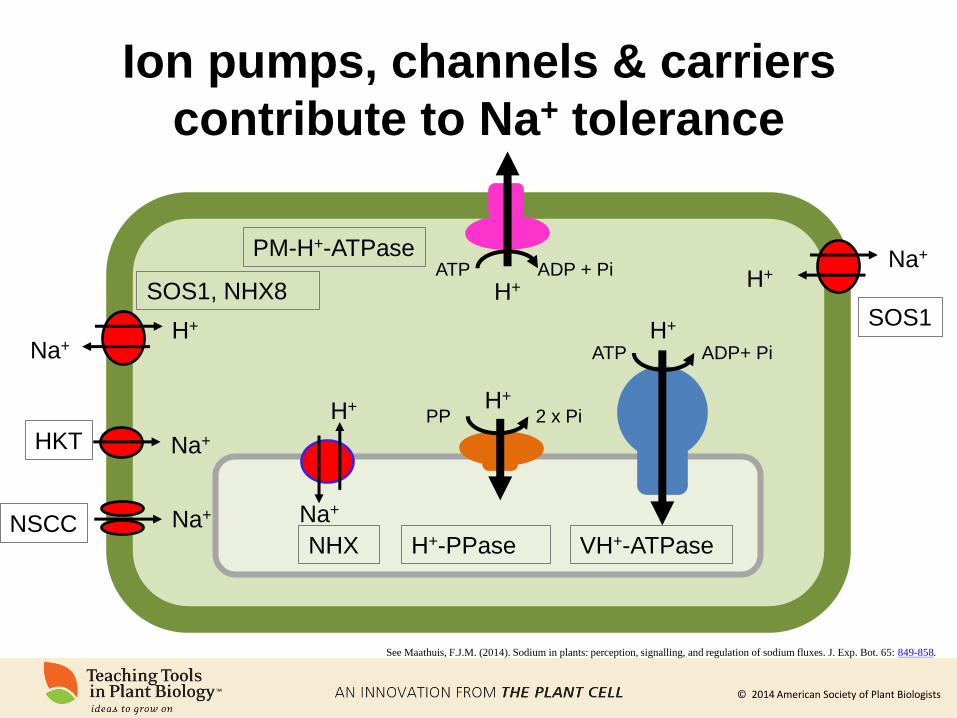
Ion pumps, channels & carriers
contribute to Na+ tolerance
H+
H+
H+ ATP ADP + Pi
ATP ADP+ Pi
PP 2 x Pi
Na+
H+
Na+ H+
H+ Na+
SOS1
VH+-ATPase H+-PPase NHX
PM-H+-ATPase
SOS1, NHX8
NSCC Na+
Na+ HKT
See Maathuis, F.J.M. (2014). Sodium in plants: perception, signalling, and regulation of sodium fluxes. J. Exp. Bot. 65: 849-858.
© 2014 American Society of Plant Biologists
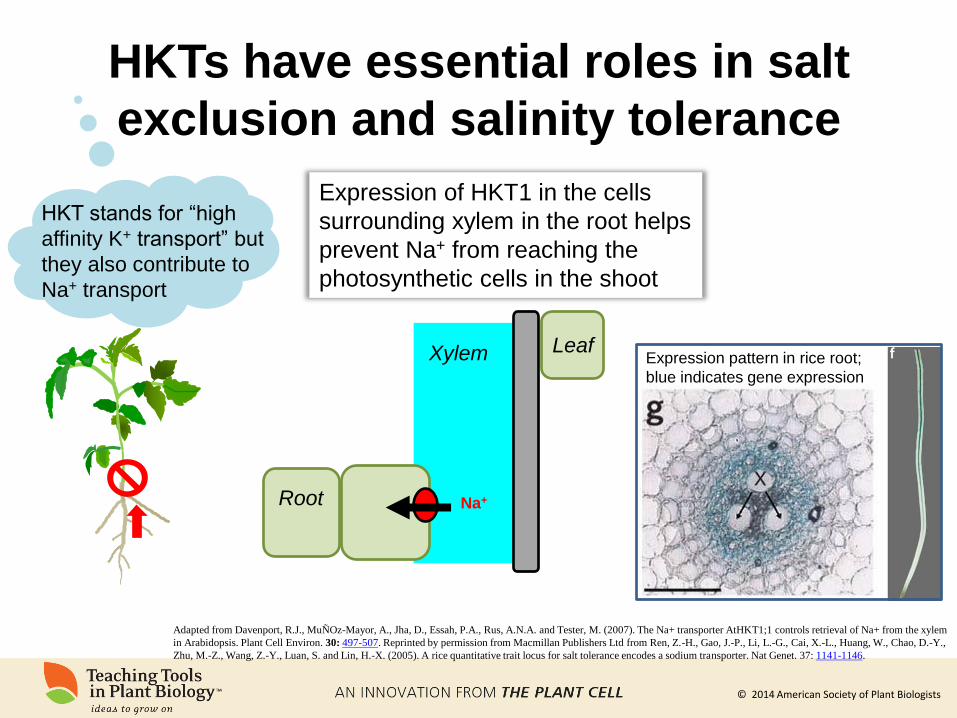
HKTs have essential roles in salt
exclusion and salinity tolerance
HKT stands for “high
affinity K+ transport” but
they also contribute to
Na+ transport
Na+ Root
Leaf Xylem
Expression of HKT1 in the cells
surrounding xylem in the root helps
prevent Na+ from reaching the
photosynthetic cells in the shoot
Adapted from Davenport, R.J., MuÑOz-Mayor, A., Jha, D., Essah, P.A., Rus, A.N.A. and Tester, M. (2007). The Na+ transporter AtHKT1;1 controls retrieval of Na+ from the xylem
in Arabidopsis. Plant Cell Environ. 30: 497-507. Reprinted by permission from Macmillan Publishers Ltd from Ren, Z.-H., Gao, J.-P., Li, L.-G., Cai, X.-L., Huang, W., Chao, D.-Y.,
Zhu, M.-Z., Wang, Z.-Y., Luan, S. and Lin, H.-X. (2005). A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat Genet. 37: 1141-1146.
Expression pattern in rice root;
blue indicates gene expression

© 2014 American Society of Plant Biologists
HKT1 expression level and activity is
correlated with Na+-tolerance
Mäser, P., Eckelman, B., Vaidyanathan, R., Horie, T., Fairbairn, D.J., Kubo, M., Yamagami, M., Yamaguchi, K., Nishimura, M., Uozumi, N., Robertson, W., Sussman, M.R. and Schroeder,
J.I. (2002). Altered shoot/root Na+ distribution and bifurcating salt sensitivity in Arabidopsis by genetic disruption of the Na+ transporter AtHKT1. FEBS letters. 531: 157-161.
Salt-tolerance in varieties of
rice, wheat and barley has
been genetically mapped to
variation in HKT activity
Loss of function =
more salt sensitive
Gain of function =
more salt tolerant
© 2014 American Society of Plant Biologists
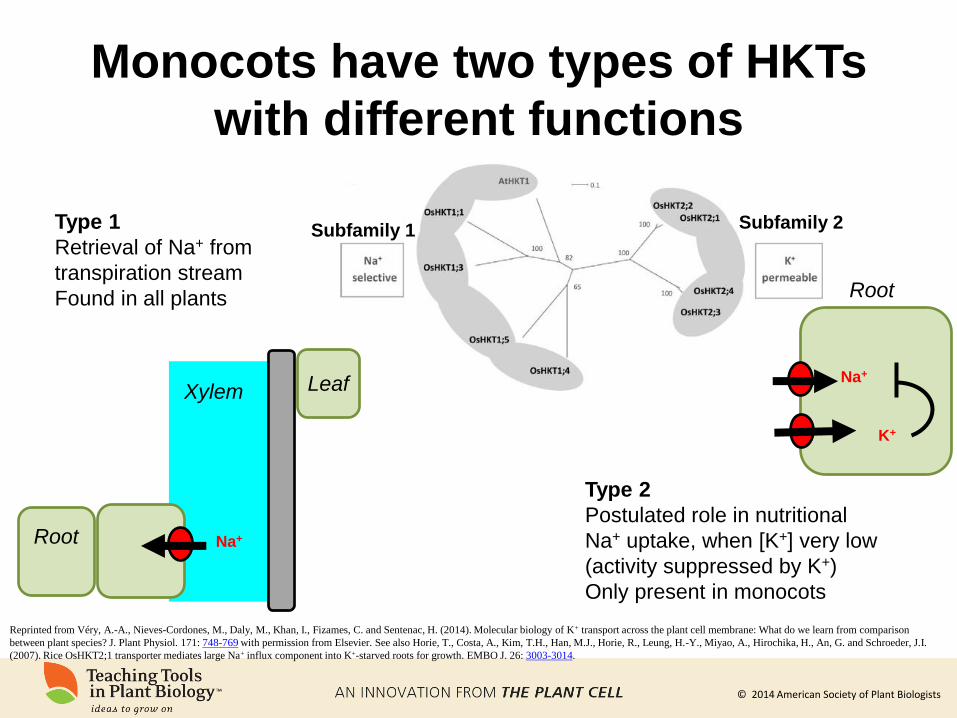
Monocots have two types of HKTs
with different functions
Subfamily 1 Subfamily 2
Reprinted from Véry, A.-A., Nieves-Cordones, M., Daly, M., Khan, I., Fizames, C. and Sentenac, H. (2014). Molecular biology of K+ transport across the plant cell membrane: What do we learn from comparison
between plant species? J. Plant Physiol. 171: 748-769 with permission from Elsevier. See also Horie, T., Costa, A., Kim, T.H., Han, M.J., Horie, R., Leung, H.-Y., Miyao, A., Hirochika, H., An, G. and Schroeder, J.I.
(2007). Rice OsHKT2;1 transporter mediates large Na+ influx component into K+-starved roots for growth. EMBO J. 26: 3003-3014.
Type 1
Retrieval of Na+ from
transpiration stream
Found in all plants
Type 2
Postulated role in nutritional
Na+ uptake, when [K+] very low
(activity suppressed by K+)
Only present in monocots
Root
Na+
K+
Na+ Root
Leaf Xylem

© 2014 American Society of Plant Biologists
NHX (Sodium / proton exchangers)
are part of the CPA family
Chérel, I., Lefoulon, C., Boeglin, M. and Sentenac, H. (2013). Molecular mechanisms involved in plant adaptation to low K+ availability. J. Exp. Bot. 65: 833-848. Gierth, M. and Mäser, P.
(2007). Potassium transporters in plants – Involvement in K+ acquisition, redistribution and homeostasis. FEBS letters. 581: 2348-2356. Chanroj, S., Wang, G., Venema, K., Zhang, M.W.,
Dalwiche, C.F., and Sze, H. (2012). Conserved and diversified gene families of monovalent cation / H+ antiporters from algae to flowering plants. Front. Plant Sci. 3: 25.
Arabidopsis: 8 NHX transporters
AtNHX 1 – 4 Vacuole
AtNHX 5 – 6 Endosome
AtNHX 7 (SOS1) – 8 PM CPA = Cation / proton antiporter
© 2014 American Society of Plant Biologists
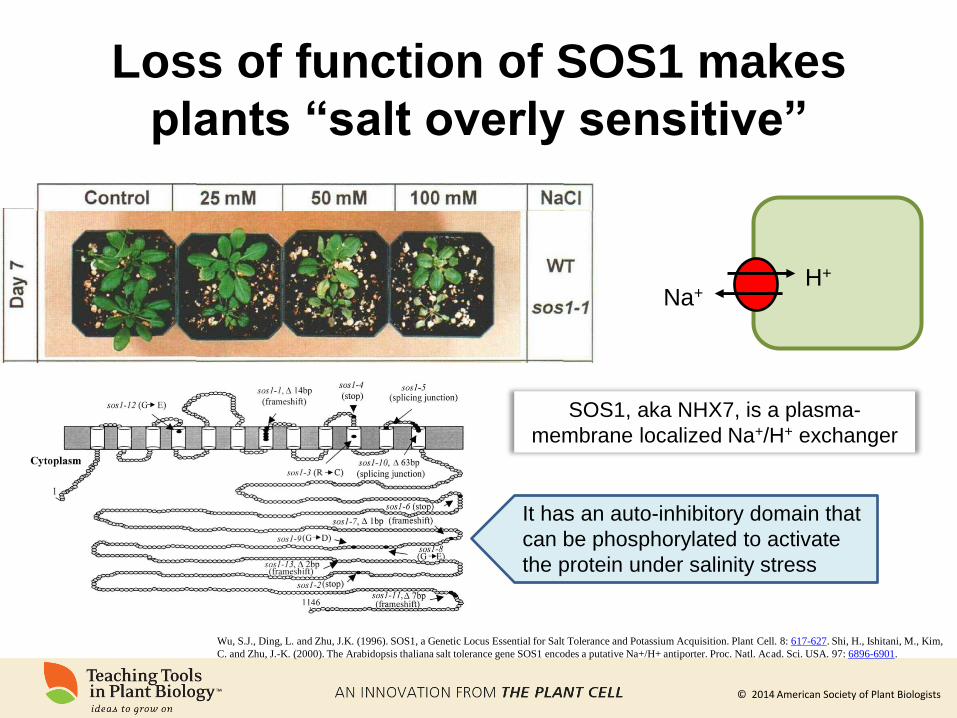
Loss of function of SOS1 makes
plants “salt overly sensitive”
Wu, S.J., Ding, L. and Zhu, J.K. (1996). SOS1, a Genetic Locus Essential for Salt Tolerance and Potassium Acquisition. Plant Cell. 8: 617-627. Shi, H., Ishitani, M., Kim,
C. and Zhu, J.-K. (2000). The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA. 97: 6896-6901.
Na+ H+
SOS1, aka NHX7, is a plasma-
membrane localized Na+/H+ exchanger
It has an auto-inhibitory domain that
can be phosphorylated to activate
the protein under salinity stress

© 2014 American Society of Plant Biologists
NHXs roles include Na+, K+ and H+
transport and homeostasis
Bassil, E., Coku, A. and Blumwald, E. (2012). Cellular ion homeostasis: emerging roles of intracellular NHX Na+/H+ antiporters in plant growth and development. J. Exp. Bot. 63: 5727-5740, by permission of Oxford University Press.
Na+ sequestration in vacuole Outward Na+ across PM
Regulation of pH of
endocytic
compartments for
proper protein
sorting and
modification

© 2014 American Society of Plant Biologists
Identification of salt tolerance in
halophytes and crop relatives
Chick pea
(Cicer arietinum)
See Flowers, T.J., Galal, H.K. and Bromham, L. (2010). Evolution of halophytes: multiple origins of salt tolerance in land plants.
Functional Plant Biology. 37: 604-612. Photo credits: Sanjay Acharya; Z. Hell; Fritz Geller-Grimm; Javier Martin, H2O
Salicornia spp.
Arthrocnemum
macrostachyum
Most sensitive Most tolerant
Glycophytes Halophytes
Salinity tolerance is a complex
trait and plants have a wide range
of salinity tolerances Halophytes are often defined as
having the ability to complete their
lifecycle on >200 mM NaCl
Wheat – intermediate sensitivity
(Triticum aestivum)

© 2014 American Society of Plant Biologists
Salt tolerance has evolved
repeatedly and independently
Flowers, T.J., Galal, H.K. and Bromham, L. (2010). Evolution of halophytes: multiple origins of salt tolerance
in land plants. Funct. Plant Biol. 37: 604-612; Bennett, T.H., Flowers, T.J. and Bromham, L. (2013). Repeated
evolution of salt-tolerance in grasses. Biol. Lett. 9: 20130029, by permission of the Royal Society.
RED indicates order that
includes some halophytes
All
vascu
lar
pla
nts
200 salt tolerant species
within the grasses, from
~76 independent events

© 2014 American Society of Plant Biologists
Halophytes can be grown on saline
soils for food and fodder
Image credits: M. Fagg, Australian National Botanic Gardens; Arizona State University. See Glenn, E.P., Anday, T., Chaturvedi, R., Martinez-Garcia, R., Pearlstein, S., Soliz, D., Nelson, S.G. and
Felger, R.S. (2013). Three halophytes for saline-water agriculture: An oilseed, a forage and a grain crop. Env. Exp. Bot. 92: 110-121;Flowers, T.J. and Colmer, T.D. (2008). Salinity tolerance in
halophytes*. New Phytol. 179: 945-963. Shabala, S. (2013). Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 112: 1209-1221.
Atriplex nummularia
(old man saltbush)
The genus Atriplex includes many
edible halophytes and is being
grown for fodder in Australia
Palmer’s grass (or nipa grass; Distichlis
palmeri) grows in tidal marshes of the Gulf
of California and was a food grain eaten by
the indigenous people of the region
Mex
USA

© 2014 American Society of Plant Biologists
Quinoa is a facultative halophyte
and a popular food grain
Bonales-Alatorre, E., Shabala, S., Chen, Z.-H. and Pottosin, I. (2013). Reduced tonoplast fast-activating and slow-activating channel
activity is essential for conferring salinity tolerance in a facultative halophyte, Quinoa. Plant Physiol. 162: 940-952. Maurice Chédel
Quinoa (Chenopodium quinoa) evolved in the
Andes and can tolerate saline soils
Young leaves (left) extrude salt into salt
bladders, older leaves (right) store it in vacuoles Quinoa is also a
useful model for
studies of salinity
tolerance
Vacuolar sodium channel activity is decreased in old
leaves (right) grown under salinity; no salinity effect is
observed in young leaves
Control
Salt
Control
Salt

© 2014 American Society of Plant Biologists
Models for salt tolerance:
Eutrema spp. (salt /saltwater cress)
Reprinted from Amtmann, A. (2009). Learning from evolution: Thellungiella generates new knowledge on essential and
critical components of abiotic stress tolerance in plants. Mol. Plant. 2: 3-12 by permission of Oxford University Press.
Arabidopsis thaliana
(Previously known as
Thellungiella halophila
or the related
Thellungiella
salsuginea)
Strategies for salt tolerance
include expansion of several gene
families (HKT, AVP) and lower
accumulation of Na+ in the shoot
as compared to Arabidopsis..
Higher selectivity
for K+ than Na+
uptake in the root
© 2014 American Society of Plant Biologists
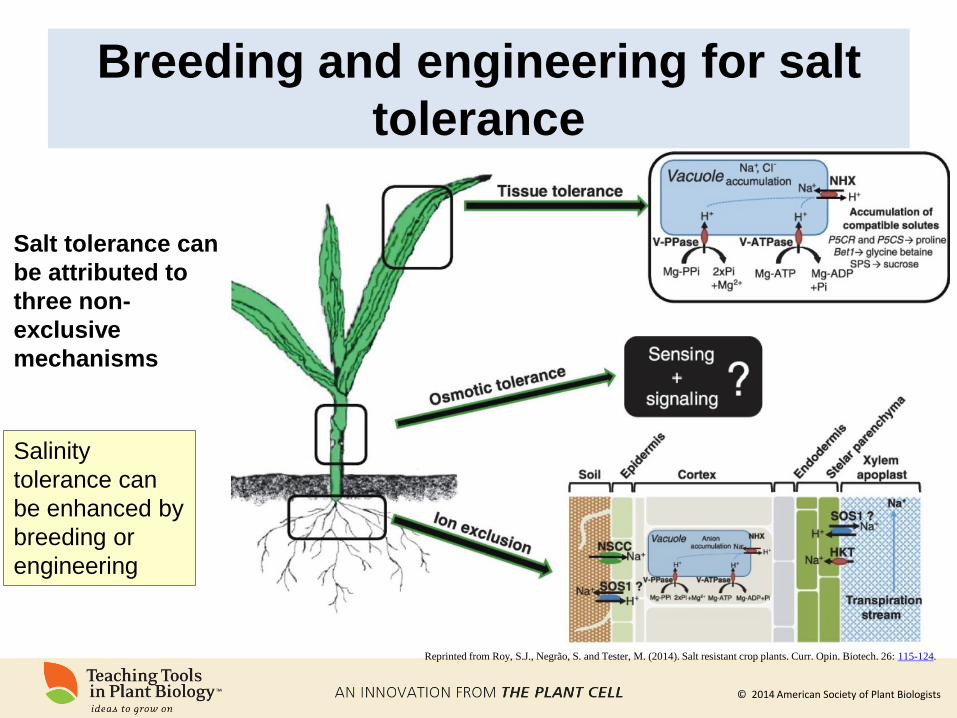
Breeding and engineering for salt
tolerance
Reprinted from Roy, S.J., Negrão, S. and Tester, M. (2014). Salt resistant crop plants. Curr. Opin. Biotech. 26: 115-124.
Salt tolerance can
be attributed to
three non-
exclusive
mechanisms
Salinity
tolerance can
be enhanced by
breeding or
engineering

© 2014 American Society of Plant Biologists
Wheat yield on saline soils improved
by an ancestral Na+ transporter gene
Huang, S., Spielmeyer, W., Lagudah, E.S. and Munns, R. (2008). Comparative mapping of HKT genes in wheat, barley, and rice, key determinants
of Na+ transport, and salt tolerance. J. Exp. Bot. 59: 927-937 by permission of Oxford University Press; Credit: Dr Richard James, CSIRO
A pair of genes derived from a
relative of wheat confers enhanced
salinity tolerance
Tetraploid
pasta wheat
Hexaploid
bread wheat
Durum wheat carrying salt-
tolerance genes
Because these species are
closely related, the genes
can be introduced into
cultivated wheat without
using GM methods

© 2014 American Society of Plant Biologists
Nax1 and Nax2 exclude Na+ from leaf
blades by removal from xylem
Reprinted by permission from Macmillan Publishers Ltd from Munns, R., et al and Gilliham, M. (2012). Wheat grain yield on saline soils is improved by an ancestral Na+ transporter gene. Nat.
Biotech. 30: 360-364. Schroeder, J.I., et al and and Sanders, D. (2013). Using membrane transporters to improve crops for sustainable food production. Nature. 497: 60-66. Huang, S., Spielmeyer,
W., Lagudah, E.S. and Munns, R. (2008). Comparative mapping of HKT genes in wheat, barley, and rice, key determinants of Na+ transport, and salt tolerance. J. Exp. Bot. 59: 927-937.
When expressed in Xenopus
oocytes, the transporters
conduct Na+ but not K+
In plants, Nax1 and Nax2
pump Na+ into the cells
surrounding the xylem so it
does not reach the leaf blade
Nax1
Nax2

© 2014 American Society of Plant Biologists
The candidate gene approach has
had some success
Transgenic plants carrying vacuolar NHXs,
vacuolar H+-PPases and plasma membrane NHXs
have demonstrated enhanced salinity tolerance
H+
H+
H+ ATP ADP + Pi
ATP ADP+ Pi
PP 2 x Pi
Na+
H+
H+ Na+
SOS1
V-H+-ATPase H+-PPase NHX
PM-H+-ATPase
NSCC Na+
Na+ HKT
Enhanced ROS
detoxification and
synthesis of
compatible solutes
is also correlated
with enhanced
salinity tolerance
See for example Roy, S.J., Negrão, S. and Tester, M. (2014). Salt resistant crop plants. Curr. Opin. Biotechnology. 26: 115-124; Gaxiola, R.A., Li, J., Undurraga, S., Dang, L.M., Allen,
G.J., Alper, S.L. and Fink, G.R. (2001). Drought- and salt-tolerant plants result from overexpression of the AVP1 H+-pump. Proc. Natl. Acad. Sci. USA 98: 11444-11449; Apse, M.P.,
Aharon, G.S., Snedden, W.A. and Blumwald, E. (1999). Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science. 285: 1256-1258.

© 2014 American Society of Plant Biologists
The intersection of potassium
nutrition and sodium toxicity
Cuin, T.A., Miller, A.J., Laurie, S.A. and Leigh, R.A. (2003). Potassium activities in cell compartments of salt‐grown barley leaves. J. Exp. Bot. 54: 657-661 with permission from Oxford University Press.
K+ext Na+
ext
K+ uptake
Na+ uptake
Na+ and K+ interfere with
each other’s uptake
When barley plants are grown
on 200 mM NaCl, they
accumulate Na+ at the
expense of K+ in their leaves

© 2014 American Society of Plant Biologists
As [Na+]ext increases and enters the
cell, K+ is driven out
H+
ATP
ADP + Pi
Non-selective
cation channel Na+
K+
Na+
Membrane
depolarizes
1. Steep concentration
gradient for Na+
2. Na+ leaks in
through NSCCs
3. Increased negative
charge within;
depolarized membrane
4. K+ driven out
through Kv channel
Some salt-tolerant plants
maintain elevated K+ by higher
activity of PM H+-ATPase
Chen, Z., Pottosin, I.I., Cuin, T.A., Fuglsang, A.T., Tester, M., Jha, D., Zepeda-Jazo, I., Zhou, M., Palmgren, M.G., Newman, I.A. and Shabala, S. (2007). Root plasma membrane transporters
controlling K+/Na+ homeostasis in salt-stressed barley. Plant Physiol. 145: 1714-1725; Shabala, S. and Cuin, T.A. (2008). Potassium transport and plant salt tolerance. Physiol. Plant. 133: 651-669.
Na+
K+
Furthermore,
Na+ directly
competes with
K+ for low- and
high-affinity
transporters

© 2014 American Society of Plant Biologists
Interaction between K+ nutrition and
Na+ toxicity
Cytosol
Vac.
Prioritized
Non-
Prioritized
K+ / Na+ ratio
K+ / Na+ ratio
Plants must
coordinate the
actions of K+ and Na+
transporters to
maintain a high ratio
of K+ to Na+ in
prioritized tissues
Adapted from Amtmann, A., and Leigh, R. (2010). Ion homeostasis. In Abiotic Stress Adaptation in Plants: Physiological, Molecular and Genomic
Foundation, A. Pareek, S.K. Sopory, H.J. Bohnert and Govindjee (eds) (Dordrecht, The Netherlands: Springer), pp. 245 – 262.

© 2014 American Society of Plant Biologists
Salinity tolerance: Summary
• Saline soils are detrimental to plants and
are widespread
• Sodium toxicity is primarily due to
interfering with K+ nutrition
• Sodium tolerance depends on exclusion,
extrusion an sequestration
• Breeding and engineering for salinity
tolerance have had mixed success so far

© 2014 American Society of Plant Biologists
Summary and ongoing research
K+
K+
PO43-
PO43-
PO43-
NO3-
NO3-
• Nutrient uptake is extremely energetically demanding
• Proton motive force generated by proton pumps is
essential for nutrient uptake
• Dozens of membrane transporters are involved in
uptake, allocation and homeostasis of mineral nutrients
• Most plants require a high cytosolic ratio of K+ to Na+
• Plants require large amounts of potassium for optimal
growth
• Sodium toxicity is a real and growing problem
• The mechanisms of sodium tolerance are being
identified and exploited for plant breeding


















