
OXIDATIVE PHOSPHORYLATIONS: PATHWAYS AND YIELD … · oxidative phosphorylations: pathways and...
15
OXIDATIVE PHOSPHORYLATIONS: PATHWAYS AND YIELD IN MITOCHONDRIAL PREPARATIONS* BY J. H. COPENHAVER, JR.,? AND HENRY A. LARDY (From the Institute jar Enzyme Research, University of Wisconsin, Madison, Wisconsin) (Received for publication, August 31, 1951) Present evidence is in general agreement that 2 or 3 moles of phosphate may be esterified per atom of oxygen consumed during the oxidation of most intermediary metabolites by animal tissue preparations (l-6). These former studies have been concerned mainly with the complete oxidation of a particular substrate. While such studies may give indication of the over-all efficiency of such oxidations, they do not permit the exact estima- tion of the efficiency of a particular oxidative reaction. Ochoa (7) studied the single step oxidation of a-ketoglutarate to succinate and Lehninger and Smith (4) have studied the phosphorylations involved in the oxidation of P-hydroxybutyrate to acetoacetate. Lehninger (8) has also studied the phosphorylations occurring during the oxidation of reduced diphospho- pyridine nucleotide by’mitochondrial preparations, and has concluded that the phosphate fixed in that series of reactions accounts for all the phosphate fixed during /3-hydroxybutyrate oxidation. This paper summarizes results of a study of the phosphorylations asso- ciated with a number of one-step and two-step oxidations in rat liver mitochondria. Such mitochondria, prepared in isotonic sucrose (9), have a minimum of phosphatase activity (10) and the oxidations which they carry out are compulsorily coupled with phosphorylation (11). In addi- tion to the use of oxygen as the ultimate electron acceptor, studies were also made of the phosphorylations associated with the reduction of arti- ficial electron acceptors. A comparison of phosphorylation efficiencies with various electron acceptors should provide some insight as to the loci of high energy phosphate generation within the electron transport chain. EXPERIMENTAL Livers were obtained from young mature male rats of the Holtzman strain. The tissue was chilled and a 10 per cent homogenate prepared with cold 0.25 M sucrose in a moderately loose Potter-Elvehjem homoge- * Supported by grants from the American Cancer Society upon recommendation of the Committee on Growth of the National Research Council and from the Life Insurance Medical Research Fund. t Postdoctorate Fellow of the National Heart Institute. Present address, De- partment of Pharmacology, Southwestern Medical School, Dallas, Texas. 225 by guest on July 16, 2018 http://www.jbc.org/ Downloaded from
Transcript of OXIDATIVE PHOSPHORYLATIONS: PATHWAYS AND YIELD … · oxidative phosphorylations: pathways and...

OXIDATIVE PHOSPHORYLATIONS: PATHWAYS AND YIELD IN MITOCHONDRIAL PREPARATIONS*
BY J. H. COPENHAVER, JR.,? AND HENRY A. LARDY
(From the Institute jar Enzyme Research, University of Wisconsin, Madison, Wisconsin)
(Received for publication, August 31, 1951)
Present evidence is in general agreement that 2 or 3 moles of phosphate may be esterified per atom of oxygen consumed during the oxidation of most intermediary metabolites by animal tissue preparations (l-6). These former studies have been concerned mainly with the complete oxidation of a particular substrate. While such studies may give indication of the over-all efficiency of such oxidations, they do not permit the exact estima- tion of the efficiency of a particular oxidative reaction. Ochoa (7) studied the single step oxidation of a-ketoglutarate to succinate and Lehninger and Smith (4) have studied the phosphorylations involved in the oxidation of P-hydroxybutyrate to acetoacetate. Lehninger (8) has also studied the phosphorylations occurring during the oxidation of reduced diphospho- pyridine nucleotide by’mitochondrial preparations, and has concluded that the phosphate fixed in that series of reactions accounts for all the phosphate fixed during /3-hydroxybutyrate oxidation.
This paper summarizes results of a study of the phosphorylations asso- ciated with a number of one-step and two-step oxidations in rat liver mitochondria. Such mitochondria, prepared in isotonic sucrose (9), have a minimum of phosphatase activity (10) and the oxidations which they carry out are compulsorily coupled with phosphorylation (11). In addi- tion to the use of oxygen as the ultimate electron acceptor, studies were also made of the phosphorylations associated with the reduction of arti- ficial electron acceptors. A comparison of phosphorylation efficiencies with various electron acceptors should provide some insight as to the loci of high energy phosphate generation within the electron transport chain.
EXPERIMENTAL
Livers were obtained from young mature male rats of the Holtzman strain. The tissue was chilled and a 10 per cent homogenate prepared with cold 0.25 M sucrose in a moderately loose Potter-Elvehjem homoge-
* Supported by grants from the American Cancer Society upon recommendation of the Committee on Growth of the National Research Council and from the Life Insurance Medical Research Fund.
t Postdoctorate Fellow of the National Heart Institute. Present address, De- partment of Pharmacology, Southwestern Medical School, Dallas, Texas.
225
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from

226 OXIDATIVE PHOSPHORYLATION
nizer. The mitochondria were separated by a modification (11) of the procedure of Schneider (9). The final mitochondrial sediment was made up to a volume equal to the original weight of tissue sample.
The basic incubation mixture in the Warburg vessels was as follows: main compartment, 0.5 ml. of mitochondrial suspension; 0.036 PM of cyto- chrome c; 22.5 PM of MgS04; 6 PM of ATP;’ 40 PM potassium phosphate buffer at pH 7.3; and 20 uM of substrate. One side arm contained 0.03 or 0.05 ml. of yeast hexokinase preparation and 50 pM of glucose. The other side arm contained 0.3 ml. of 50 per cent perchloric acid. All flask com- ponents were added as essentially isotonic solutions and the final volume, exclusive of the perchloric acid, was made up to 3.0 ml. with 0.15 M KCl. NaOH and a fluted filter paper were used in the center wells.
The hexokinase-glucose system was used in all experiments as a means of trapping the high energy phosphate by converting it to hexose phos- phate. The yeast hexokinase preparation was a suspension obtained at Step 3a according to the procedure of Berger et al. (12). The cytochrome c used in these experiments was from the Sigma Chemical Company or purified samples from other sources. Adenosinetriphosphate was the pure disodium tetrahydrate salt obtained from the Pabst Laboratories. All other reagents were reagent grade or purified in the laboratory.
The flasks were kept in an ice bath until the enzyme preparations were added and then attached to manometers. They were equilibrated in the water bath at 30” for 3 minutes, the contents of the glucose-containing side arm were then flushed into the main compartment, and the equilibra- tion allowed to proceed for 2 minutes.2 During this time the stop-cocks were closed and the manometer fluid brought to scale. The initial read- ings were taken and one or two zero time flasks were removed and the contents quickly deproteinized. All flasks were kept chilled for later chemi- cal analyses. Oxidation was allowed to proceed for 15 or 25 minutes in the remaining flasks. After the final readings, the flask contents were deproteinized.
The flask contents were usually analyzed for orthophosphate by the method of Lowry and Lopez (13). In the presence of ferricyanide, ortho- phosphate was determined in the following manner. A suitably diluted aliquot, containing about 1 PM of phosphate, was made alkaline and 0.2 ml. of 25 per cent barium acetate added. The tubes were chilled, centri- fuged in the cold, and the supernatant decanted and drained by inverting the tubes on filter paper. The precipitate was dissolved in 1 N HCl; a
1 The following abbreviations are employed: KG = a-ketoglutarate, Pyr = pyruvate, AcAc = acetoacetate, ATP = adenosinetriphosphate, DPN = diphospho- pyridine nucleotide, NP = high energy phosphate.
* Control experiments demonstrated that the 5 minute preincubation period was adequate to obtain thermal equilibrium.
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from

J. H. COPENHAVER, JR., AND H. A. LARDY 227
volume equal to the original sample was taken and the very smail volume of the precipitate neglected. An aliquot of this was taken for the deter- mination of orthophosphate by the method of King (14). Standard amounts of orthophosphate gave better than 95 per cent recovery with this method.
Acetoacetate was determined by Barkulis and Lehninger’s (15) modi- fication of the Greenberg and Lester (16) method for the determination of ketone bodies. This method was not applicable to samples containing ferricyanide. Acetoacetate production in the presence of this electron acceptor was determined by distillation of an aliquot of the filtrate in the presence of aniline hydrochloride. The acetone was trapped as the 2,4- dinitrophenylhydrazone and this was then carried through the extraction procedure outlined by Barkulis and Lehninger (15).
Pyruvate and ac-ketoglutarate were determined by a modification of the procedure of Friedemann and Haugen (17). Ferricyanide was determined by direct calorimetry, with a 420 rnp filter. Malate was measured by the fluorometric method of Hummel (18), while citrate was determined by the procedure of Natelson et al: (19).
The tabular data include every experiment which has been done under the conditions described. All experiments are numbered consecutively and comparisons may be made of the results obtained with different sub- strates within a given experiment by reference to corresponding experiment numbers in the different tables.3 In many cases zero time flasks and in all cases experimental flasks were run in duplicate.
Results
It has been the aim of the present work to study the generation of high energy phosphate coupled with a single oxidative step and to determine substrate disappearance or end-product accumulation simultaneously when- ever possible. In this manner, a balance sheet can be constructed which would indicate the occurrence of dismutations or additional oxidations. The results of such studies with three substrates are summarized in Tables I, II, and III.
The data in Table I show that the oxidation of ac-ketoglutarate to suc- cinate is accompanied by the uptake of a maximum of 4 moles of phos- phate. It is evident that malonate prevented the further oxidation of succinate since no more than 1 atom of oxygen was consumed per mole of a-ketoglutarate removed. Also, since the oxygen: ol-ketoglutarate ratios are very nearly 1, dismutations. are probably not occurring. Therefore, it seems justifiable to assume that the measured P:O ratios of 3.2 and 3.6
3 Since space does not permit publication of the data for individual experiments, the original tables have been mimeographed and are available on request.
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from

228 OXIDATIVE PHOSPHORYLATION
in the absence and presence of fluoride are a measure of the efficiency of the one-step oxidation of cr-ketoglutarate to succinate in the mitochondrial preparation. Assuming that the theoretical P:O ratio for this one-step oxidative reaction is a whole number, it appears that the ratio for this reaction is probably 4, a value which has been approximated in four ex- periments.
The ratios of orthophosphate disappearance to oxygen consumption are
TABLE I
Phosphorylation Associated with Oxidation of a-Ketoglutarate to Succinate with Oxuaen As Electron Acceptor
A o AKG P:O P:KG 0:KG
-~ ~__
M w. CM micro- atoms PM
9 0.01 2.4 17.2 4.8 5.3 3.6 (3.3-4.1) 3.3 0.9 (0.8-1.1) 12 None 2.0 16.6 4.6 4.9 3.2 (2.2-4.1) 3.2 1.0 (0.8-1.1)
The figures are averages for all experiments; ranges indicated in parentheses. 0.01 or 0.02 M malonate in all flasks.
TABLE II
Phosphurylation Associated with Oxidation of Pyruvate to Acetoacetate with Oxygen As Electron Acceptor
A 0 Afir Atic P:O P:Pyr 0:Pyr Pyr:AcAc
~__ ~~~
M w. PM zoczs- PM PM
17 0.01 1.7 11.6 4.4 4.8 2.4 2.6 (2.1-3.2) 2.5 0.9 2.2 9 None 1.8 18.0 7.2 8.6 3.6 2.6 (2.0-2.9) 2.3 1.0 2.1
The figures are averages for all experiments; ranges indicated in parentheses. 0.004 M malonate present in ail flasks.
calculated from the values obtained by the chemical determination of orthophosphate and the manometric determination of oxygen consump- tion. The accuracies of these determinations are such that P: 0 ratios calculated from duplicate flasks seldom differ by as much as 10 per cent. Thus, the two experiments in Table I resulting in P:O ratios of 4.1 do not exceed the value of 4 by a significant amount.
Lehninger (20), using washed residues of rat liver homogenate, and Recknagel and Potter (21), using whole homogenates of rat liver, showed that the oxidation of pyruvate in the absence of 4-carbon dicarboxylic acid and in the presence of malonate results in the accumulation of 1 mole of acetoacetate for every 2 moles of pyruvate disappearing. The data on
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from
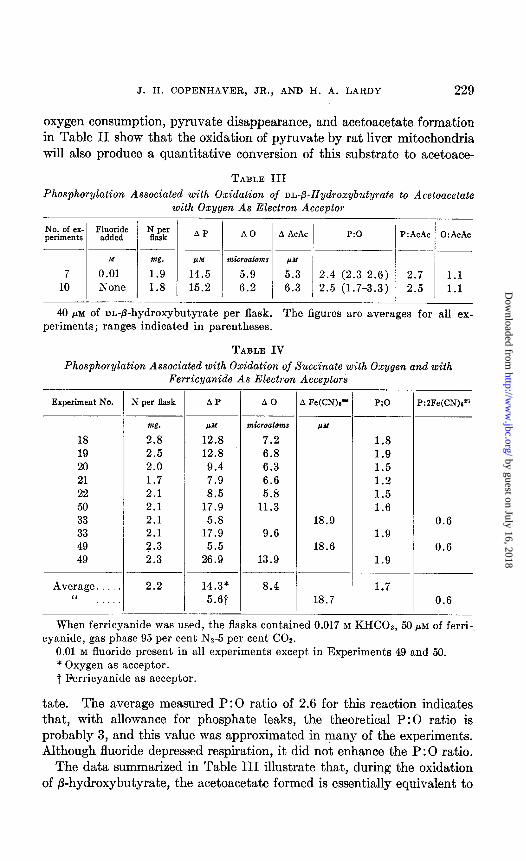
J. H. COPENHAVER, JR., AND H. A. LARDY 229
oxygen consumption, pyruvate disappearance, and acetoacetate formation in Table II show that the oxidation of pyruvate by rat liver mitochondria will also produce a quantitative conversion of this substrate to acetoace-
TABLE III
Phosphorylation Associated with Oxidation of m-P-Hydroxybutyrate to Acetoacetate with Oxygen As Electron Acceptor
No. of ex- FFw&ii periments ;,p,"k' A P A0 A AcAc P:O P:AcAc 0:AcAc
M w. w microatons p.M
7 0.01 1.9 14.5 5.9 5.3 2.4 (2.3-2.6) 2.7 1.1 10 None 1.8 15.2 6.2 6.3 2.5 (1.7-3.3) 2.5 1.1
40 PM of nn-fl-hydroxybutyrate per flask. The figures are averages for all ex- periments; ranges indicated in parentheses.
TABLE IV
Phosphorylation Associated with Oxidation of Succinate with Oxygen and with Ferricyanide As Electron Acceptors
Experiment No. N per tlask AP A0 I Fe(CN)s’ P:O
w. PM
2.8 12.8 2.5 12.8 2.0 9.4 1.7 7.9 2.1 8.5 2.1 17.9 2.1 5.8 2.1 17.9 2.3 5.5 2.3 26.9
microatoms
7.2 6.8 6.3 6.6 5.8
11.3
w
18 19 20 21 22 50 33 33 49 49
1.8 1.9 1.5 1.2 1.5 1.6
9.6 18.9
18.6 13.9
1.9
1.9
1.7 Average.. 2.2 “
14.3* 5.6t
8.4
0.6
0.6
18.7 0.6
-
I ‘:ZFe(CN)s’
When ferricyanide was used, the flasks contained 0.017 M KHCOa, 50 ,AM of ferri- cyanide, gas phase 95 per cent Nz-5 per cent COZ.
0.01 M fluoride present in all experiments except in Experiments 49 and 50. * Oxygen as acceptor. t Perricyanide as acceptor.
tate. The average measured P:O ratio of 2.6 for this reaction indicates that, with allowance for phosphate leaks, the theoretical P:O ratio is probably 3, and this value was approximated in many of the experiments. Although fluoride depressed respiration, it did not enhance the P:O ratio.
The data summarized in Table III illustrate that, during the oxidation of P-hydroxybutyrate, the acetoacetate formed is essentially equivalent to
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from

230 OXIDATIVE PHOSPHORYLATION
the oxygen consumed. Thus, it may be assumed that this is a single step oxidation. This is in accord with the concept that acetoacetate is not metabolized by rat liver preparations. The experimentally determined P:O ratio of 2.4 during a 15 minute period is somewhat greater than the values reported by Lehninger and Smith (4), who used adenylic acid as a
Phosphorylation Associated with Oxidation of Citrate to Succinate with Oxygen and with Ferricyanide As Electron Acceptors
Experiment No . P < per flask AP
41 42 43 44 44 45 45 46 46 47 47 48 48 50 50
w. PM 2.2 8.6 2.0 11.6 2.5 13.5 1.9 7.6 1.9 10.1 2.3 9.2 2.3 4.8 1.8 7.0 1.8 7.0 2.2 5.7 2.2 9.0 2.4 9.2 2.4 18.3 2.1 7.2 2.1 5.3
Average . “ . . .
. . .
. . .
2.1
A0
nicroatoms
3.3 3.9 4.4
4.2 3.4
FI A citric P:O I
2.6 3.1 3.1
12.0 2.4 2.7
3.3
4.0
5.8 3.3
8.8 8.6
11.6
14.0
2.0 2.6 3.1
2.1
2.3
2.5 3.2 2.3
10.9
4.0 2.6 11.0
‘:‘ZFe(CN)s=
1.3
1.1 1.6
1.0
1.3
1.0
1.2
When ferricyanide was used, the flasks contained 0.017 M KHCOS, 50 PM of ferri- cyanide, gas phase 95 per cent Nt-5 per cent COZ.
0.007 M malonate in all flasks except as follows: Experiment 41,0.003 M malonate; Experiment 42, 0.01 M malonate.
* Oxygen as acceptor. t Ferricyanide as acceptor.
phosphate acceptor. Differences in the lability of the accumulating phos- phate compounds or in the preparation of the mitochondria may be re- sponsible for these results.
Table IV is a compilation of data for succinate as the substrate. An average experimental P: 0 ratio of 1.7 was obtained. With this substrate, no attempt was made to block the further metabolism of fumarate; there- fore, some phosphorylation resulting from subsequent oxidations cannot be ruled out.
With intact mitochondria, it was not possible to carry out the one-step
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from

J. H. COPENHAVER, JR., AND H. A. LARDY 231
oxidation of citrate or glutamate to cr-ketoglutarate in the presence of a binding agent such as semicarbazide. As an alternative, the oxidation of these two substrates was carried out in the presence of malonate. Under such conditions, the P : 0 ratios obtained are the averages of two oxidative steps. As shown by the data of Table I, the average P: 0 ratio of the one-step oxidation of a-ketoglutarate in the absence of fluoride is 3.2. As shown by Tables V and VI, the experimental P:O ratios are 2.6 for the two-step oxidation of both citrate and glutamate. The oxidations of
TABLE VI
Phosphorylation Associated with Oxidation of Glutamate to Succinate with Oxygen and with Ferricyanide As Electron Acceptors
Experiment No. N per flask AP
41 42 43 44 44 48 48 50 50
w. wf
2.2 12.2 2.0 9.9 2.5 17.2 2.3 15.0 2.3 8.7 2.4 5.0 2.4 9.7 2.1 10.0 2.1 14.9
- - A0
microatoms 4.4 3.9 6.6 5.6
4.6
5.5
L
Average. “ . . ..I 2.3 1 1;::; 1 5.’
14.3 10.6
16.6
13.8
P:O
2.8 2.5 2.6 2.7
2.1
2.7
2.6
1 P:ZFe(CN)c,'
1.2 0.9
1.2
1.1
When ferricyanide was used, the flasks contained 0.017 M KHCO,, 50 pM of ferri- cyanide, gas phase 95 per cent Nz-5 per cent COZ.
0.007 M maionate in ail flasks except as follows: Experiment 41, 0.003 M maionate; Experiment 42, 0.01 M malonate.
* Oxygen as acceptor. t Ferricyanide as acceptor.
both substrates are relatively slow, and a 25 minute experimental period was used instead of the usual 15 minutes. With such prolonged experi- ments, it is possible that leaks from the pool of esterified phosphate would be greater than with the shorter experiments (22). In addition, the deter- mination of cy-ketoglutarate in one group of such experiments showed the accumulation of about 0.5 FM of this acid in the oxidation of 2 to 3 PM of either citrate or glutamate, indicating that more than half of the oxygen consumption was due to the single step oxidation leading to a-ketogluta- rate. Therefore, it appears justified to assign a value of 3 to the theoretical P:O ratios for the oxidation of both citrate and glutamate to cr-ketoglu- tarate.
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from

232 OXIDATIVE PHOSPHORYLATION
In an attempt to gain further information concerning the sites in the electron transport chain at which - P is generated, a study was made with various electron acceptors in place of oxygen. It has been shown (23) that certain of these oxidizing agents have a preferential, if not ob- ligatory, enzyme or coenzyme from which they accept electrons.
Methylene blue gave variable results. In the oxidation of certain sub- strates, such as a-ketoglutarate, it permitted the oxidation to proceed at a rate comparable to that obtained with oxygen as the acceptor. Under these conditions, there was significant phosphate uptake, giving P: 0 ratios approaching 2. However, with other substrates such as pyruvate or /3- hydroxybutyrate, methylene blue not only completely inhibited the up- take of phosphate but also greatly diminished the rate of oxidation. It is not known whether such effects were the result of the deleterious action of hydrogen peroxide on the enzymes concerned or the direct effect of methylene blue on labile enzymes. The addition of relatively large amounts of crystalline catalase did not affect the results obtained with methylene blue.
The use of ferricyanide as the electron acceptor proved to be more satisfactory. The results of these studies are summarized in Tables IV to IX. It is evident that there is significant phosphorylation occurring during the oxidation of each of the substrates studied. The results of Tables VII and VIII show that one-step oxidation of a-ketoglutarate and pyruvate leads to the esterification of more than 1 mole of phosphate per 2 moles of ferricyanide reduced. These results will be discussed later. In some of the pyruvate experiments, acetoacetate production was de- termined by the distillation procedure described in the experimental sec- tion and was found to be in close agreement with the theoretical yield.
The data obtained in the study of other substrates indicate that the theoretical P:2Fe(CNJZ ratio for the other one-step oxidations studied is 1. Of particular interest are the results from the oxidation of /3-hydroxy- butyrate, which are summarized in Table IX. In contrast, Lehninger (24) was unable to detect any esterification of phosphate during the oxi- dation of P-hydroxybutyrate in the presence of ferricyanide.
It has been demonstrated with some reconstructed systems (23) that ferricyanide does not accept electrons directly from dehydrogenases but rather accepts them from pyridine nucleotides. Other purified oxidative enzymes are known to react with ferricyanide in the absence of DPN (25). It is obvious that the data do not demonstrate the site at which ferri- cyanide taps into the electron transport system in the intact mitochondria. The possibility exists that, in mitochondria, the point at which ferricyanide acts may depend upon the momentary ratio of ferri- to ferrocyanide and perhaps the momentary ratio of the oxidized to the reduced forms of the various enzymes themselves.
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from

J. H. COPENHAVER, JR., AND H. A. LARDY 233
To define more precisely the site at which ferricyanide accepts electrons, use was made of the antibiotic, antimycin. Ahmad et al. (26) and Potter and Reif (27) have shown that antimycin A is a powerful inhibitor of
TABLE VII
Phosphorylation Associated with Oxidation of a-Ketoglutarate to Succinate with Ferricyanide As Electron Accepfor
No. of experi- N per flask A P malts Fe&F AKG P:ZFe(CN)o' P:KG ZFe(CN)?:KG
w. PM PM CM
10 2.0 11.7 15.5 7.2 1.5 (1.2-1.9) 1.6 1.1
The figures are averages for all experiments; ranges indicated in parentheses. The flasks contained 0.017 M KHCO,, 50 PM of potassium ferricyanide, gas phase 95 per cent Nz-5 per cent COZ. 0.017 M malonate in all flasks.
TABLE VIII
Phosphorylation Associated with Oxidation of Pyruvate to Acetoacefate with Ferricyanide As Electron Acceptor
ex$&,ts Nperflask A P F&J).$' A Pyr P:ZFe(CN)s- P:Pyr 2Fe(CN)?:Pyr
~____
w. &M @M PM
7 2.0 9.0 15.5 7.4 1.2 (0.8-1.5) 1.2 1.0
The figures are averages for all experiments; ranges indicated in parentheses. The flasks contained 0.017 M KHCOS, 50 pM of potassium ferricyanide, 0.004 M malo- nate, gas phase 95 per cent N2-5 per cent COZ.
TABLE IX
Phosphorylation Associated with Oxidation of DL-&Hydroxybutyrate to Acetoacetate with Ferricyanide As Electron Acceptor
No. of experiments N per flask AP A Fe(CN)s= P:ZFe(CN)a'
w. &M PM
7 2.1 5.4 13.0 0.8 (0.6-1.1)
The figures are averages for all experiments; ranges indicated in parentheses. The flasks contained 0.017 M KHCOI, 50 pM of potassium ferricyanide, 40 pM of DLL
&hydroxybutyrate.
succinoxidase and the oxidation of other substrates of the Krebs cycle. The evidence (27) indicates that this antibiotic combines with some com- ponent of the electron transport mechanism between the dehydrogenase and cytochrome c (or oxidase). Table X shows that antimycin A, at the level of 1 y per flask, completely inhibits the oxidation of p-hydroxybu- tyrate when oxygen is the ultimate electron acceptor. It has, however,
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from

234 OXIDATIVE PHOSPHORYLATION
no effect on either the efficiency of phosphorylation or the rate of ferri- cyanide reduction when the latter is the ultimate electron acceptor. The incubation of antimycin A with ferricyanide did not diminish the ability of the antibiotic to inhibit the oxidation of succinate under aerobic con- ditions. Thus the conclusion is warranted that the electron carrier or carriers donating electrons to ferricyanide are proximal to that factor in- hibited by antimycin A.
An attempt was made to study the single step oxidation of malate in the ferricyanide system by the addition of cyanide, which reputedly binds the oxalacetate formed. Since the amount of ferricyanide reduced was in excess of the amount of malate disappearing, it would seem that malate is oxidized more than one step, either by circumventing or breaking through the cyanide block. With 20 UM of cyanide per flask, the reduction of ferricyanide was about double that predicted from the disappearance of
TABLE X
Effect of Antimycin A on Oxidative Pho&horylation of DL-&Hydroxybutyrate
Electron acceptor AP A0 A Fe(CN)e= P:O
PM microaloms PM
Ferricyanide. 6.1 14.9 + 1 y antimycin A.. 6.2 14.6
Oxygen......................... 20.6 8.6 2.4 + 1 y antimycin A.. -1.7 0.0 0.0
0.8 0.8
malate, and the P: 2Fe(CN)b ratio was close to 2. With 40 uM of cyanide, the reduction of ferricyanide was only about 50 per cent in excess of that predicted from the disappearance of malate, but the P:2Fe(CN)r ratio was slightly less than 1. The latter value is probably closer to the actual efficiency of this particular single step oxidation, although the possible effect of larger amounts of cyanide per se on this efficiency cannot be over- looked. Slater (28) has shown that increasing amounts of cyanide de- crease the efficiency of phosphorylation coupled with the oxidation of or- ketoglutarate when cytochrome c was the ultimate electron acceptor.
The two-step oxidation of citrate and glutamate in the presence of malonate was also carried out in the anaerobic ferricyanide system. The data are summarized in Tables V and VI. Since the data in Table VII show that the actual P:2Fe(CN)r ratio obtained for the single step oxida- tion of oc-ketoglutarate is 1.5, the values in Tables V and VI would indicate that the theoretical value for the single step oxidation of citrate or glu- tamate to cr-ketoglutarate is probably 1.
A comment should be made at this point as to the effect of ferricyanide
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from

J. H. COPENHAVER, JR., AND H. A. LARDY 235
on the rate of oxidation of these various substrates. With most of the substrates studied, the rate of oxidation was approximately the same with either oxygen or ferricyanide as the electron acceptor. The simplest ex- planation of this result is that the rate-limiting reaction in these oxidations is found somewhere proximal to the site at which ferricyanide taps into the electron transport mechanism. However, a-ketoglutarate oxidation is noticeably faster with ferricyanide as the electron acceptor than when oxygen is the acceptor. This effect is apparent, to a lesser extent, in the two-step oxidation of citrate or glutamate. These results seem to indicate that the rate-limiting reaction in the oxidation of ar-ketoglutarate is distal to the site at which ferricyanide will accept electrons. Such an accelera- tion was not noticed when methylene blue was used as the electron carrier, although additional problems related to the use of methylene blue, dis- cussed previously, might negate this finding.
DISCUSSION
Since the original findings of Belitzer and Tsibakova (29) and Ochoa (30) that more than 1 mole of phosphate is esterified for every atom of
p- I~~ROXYSUTYRATE
AGETOACETATE
GLUTAMATE 1 NP
q*
&ETOGLUTARATE
ISOGITRATE i \‘\\“” I
OhLOSUCCINATE 4,
\ T\
d-KETOGLUTARATE NP
SUCGINATE
PYRUVATE
oxygen consumed, it has been postulated that some of the phosphate is fixed during the transportation of electrons over the chain of intermediary catalysts between substrate and oxygen. Experimental verification of this hypothesis has been achieved by Lehninger (8) who found the oxidation of DPNH by mitochondria to yield almost 2 moles of high energy phos- phate per atom of oxygen consumed. Assuming that the fixation of inor- ganic phosphate occurs in discrete steps, the results obtained by others and in the present studies may be summarized in the accompanying scheme.
The electron transport mechanisms associated with the oxidation of /3-hydroxybutyrate, glutamate, and isocitrate are represented as similar pathways in the scheme. Electron carriers designated S, T, and U may
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from

236 OXIDATIVE PHOSPHORYLATION
be pyridine-nucleotide, a flavoprotein and Slater’s factor (31), or cyto- chrome c respectively. Similarly, the electron carrier labeled 2, which is associated with succinate oxidation, may be cytochrome b (32, 33).
As to the oxidation of pyruvate, the earlier work of Lipmann (34) indi- cates a possibility of a “substrate phosphorylation” during the oxidation of pyruvate analogous to that which occurs during the oxidation of triose phosphate (35) or ol-ketoglutarate (36). Although both acetyl moieties must be activated by phosphorylation before they may condense to aceto- acetate (37), there is a possibility that the potential energy of the aceto- acetyl - enzyme linkage or the acetoacetyl - coenzyme linkage may be preserved as high energy phosphate. Were this the case, one might expect 1.5 moles of phosphate to be fixed per mole of pyruvate oxidized to aceto- acetate by ferricyanide. Yields approaching this value were actually ob- tained. It is quite evident from the data of Table VII that 2 moles of phosphate (1 at the substrate level (36)) are fixed per mole of cu-ketoglu- tarate oxidized in the presence of ferricyanide. By analogy between the two a-keto acids, it might be proposed that the oxidation of pyruvate to acetoacetate by ferricyanide would yield 0.5 mole of -phosphate per mole of pyruvate oxidized at the substrate level and 1 additional mole at the level of an electron carrier. Such a hypothesis would fit well the data obtained from the oxidation of pyruvate by ferricyanide.
It would not, however, fit the experimental results obtained by the oxi- dation of pyruvate with oxygen as the final electron acceptor. The hy- pothesis would require a theoretical P: 0 ratio of 3.5 for the oxidation of pyruvate to acetoacetate in the presence of oxygen. The data in Table II show that such a ratio was seldom approached on an experimental basis. Hunter and Spector (36) have shown that the oxidation of cr-ketoglutarate produces a phosphorylation which is very resistant to the uncoupling action of dinitrophenol and which they presume to be a phosphorylation at the substrate level. If there is a “substrate phosphorylation” in the oxidation of pyruvate which may be preserved, then, by analogy, this phosphoryla- tion should be resistant to dinitrophenol. The results of two experiments indicate that in the oxidation of pyruvate to acetoacetate 2 X 10db M 2,4-dinitrophenol will completely prevent the fixation of inorganic phos- phate without affecting the formation of acetoacetate or the rate of oxida- tion. Such evidence indicates that the potential energy of an aceto- acetyl-enzyme linkage or an acetoacetyl-coenzyme linkage is not pre- served as high energy phosphate. Therefore, the question must be left open whether there are one or two intermediary carriers (X and Y) which carry out phosphorylations during electron transport between pyru- vate and ferricyanide.
On the basis of present evidence, it is not possible to state whether
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from

J. H. COPENHAVER, JR., AND H. A. LARDY 237
cytochrome c or cytochrome oxidase participates directly in the fixation of inorganic phosphate during electron transport. The studies of Slater (28) have indicated that all phosphorylation accompanying the oxidation of (Y- ketoglutarate occurs prior to the oxidation of cytochrome c. In a dif- ferent approach, Friedkin and Lehninger (38) were unable to find significant fixation of radioactive orthophosphate accompanying the oxidation of re- duced cytochrome c. On the other hand, in a report which has just appeared, Judah (39) has studied the oxidation of cytochrome c reduced by ascorbic acid by rat liver mitochondria and shown significant phos- phorylation with P: 0 ratios less than 1. Results of similar studies in this laboratory are intermediate between those of Friedkin and Lehninger and Judah. The oxidation of reduced cytochrome c produced in most experiments a small uptake of phosphate with P: 0 ratios approaching 0.5. The incubation of mitochondria in the presence of all flask components except an oxidizable substrate results in a slow liberation of inorganic phosphate from the ATP and mitochondrial coenzymes. However, no such leak occurred during the oxidation of ascorbic acid. These findings may support the results of Judah.
SUMMARY
The phosphorylations associated with the oxidation of various inter- mediary metabolites by rat liver mitochondria were studied under con- ditions allowing the one- or two-step oxidation of the substrate in question. The oxidation of pyruvate to acetoacetate gave P:O ratios (inorganic phosphate removed to atoms of oxygen consumed) averaging 2.6. When ferricyanide replaced oxygen, the P:2Fe(CN)r ratios obtained averaged 1.2. The one-step oxidation of a-ketoglutarate produced P:O ratios of 3.2 and 3.6 in the absence and presence of fluoride. Replacement of oxygen by ferricyanide gave P : 2Fe(CN)r, averaging 1.5. The oxidation of /3-hydroxybutyrate to acetoacetate gave an average P:O ratio of 2.4 and an average P:2Fe(CN)T ratio of 0.8. The oxidation of succinate produced an average P:O ratio of 1.7 and a P:aFe(CN)b ratio of 0.6. The two-step oxidation of citrate or glutamate to succinate gave P: 0 ratios averaging 2.6 and P:2Fe(CN)r ratios of 1.2 and 1.1 respectively. From the data obtained for the oxidation of a-ketoglutarate to succinate, it may be inferred that the theoretical P: 0 ratio for the oxidation of iso- citrate or glutamate to a-ketoglutarate is 3, while the P:2Fe(CN)r ratio is probably 1.
The antibiotic, antimycin A, completely inhibits the oxidation of P- hydroxybutyrate in the presence of oxygen but has no effect on either oxidation or high energy phosphate generation when ferricyanide is the final electron acceptor.
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from

238 OXIDATIVE PHOSPHORYLATTON
The possible loci of high energy phosphate generation within the elec- tron transport chain are discussed in the light of the evidence presented.
BIBLIOGRAPHY
1. Ochoa, S., J. Biol. Chem., 161, 493 (1943). 2. Cross, R. J., Taggart, J. V., Covo, G. A., and Green, D. E., J. Biol. Chem., 17’7,
655 (1949). 3. Hunter, F. E., Jr., and Hixon, W. S., J. Biol. Chem., 181, 73 (1949). 4. Lehninger, A. L., and Smith, S. W., J. Biol. Chem., 181, 415 (1949). 5. Johnson, R. B., and Lardy, H. A., J. Biol. Chem., 184, 235 (1950). 6. Judah, J. D., and Williams-Ashman, H. G., Biochem. J., 48, 33 (1951). 7. Ochoa, S., J. Biol. Chem., 166, 87 (1944). 8. Lehninger, A. L., J. BioZ. Chem.; 180, 345 (1951). 9. Schneider, W. C., in Umbreit, W. W., Burris, R. H., and Stauffer, J. F., Mano-
metric techniques and tissue metabolism, Minneapolis, revised edition (1949). 10. Potter, V. R., and Recknagel, R. O., in McElroy, W. D., and Glass, B., Phosphorus
metabolism, Baltimore, 377 (1951). 11. Lardy, H. A., and Wellman, H., J. BioZ. Chem., 196, 215 (1952). 12. Berger, L., Slein, M. W., Colowick, S. P., and Cori, C. F., J. Gen. Physiol., 29,
379 (1946). 13. Lowry, 0. H., and Lopez, J. A., J. Biol. Chem., 182, 421 (1946). 14. King, E. J., Biochem. J., 28,292 (1932). 15. Barkulis, S. S., and Lehninger, A. L., J. BioZ. Chem., 190, 339 (1951). 16. Greenberg, L. A., and Lester, D., J. Biol. Chem., 164, 177 (1944). 17. Friedemann, T. E., and Haugen, G. E.: J. Biol. Chem., 147, 415 (1943). 18. Hummel, J. P., J. Biol. Chem., 180, 1225 (1949). 19. Natelson, S., Lugovoy, J. K., and Pincus, J. B., J. BioZ. Chem., 170, 597 (1947). 20. Lehninger, A. L., J. BioZ. Chem., 184, 291 (1946). 21. Recknagel, R. O., and Potter, V. R., J. BioZ. Chem., 191, 263 (1951). 22. Kielley, W. W., and Kielley, R. K., J. Biol. Chem., 191, 485 (1951). 23. Dixon, M., and Zerfas, L. G., Biochem. J., 34, 371 (1940). 24. Lehninger, A. L., J. BioZ. Chem., 178, 625 (1949). 25. Schweet, R. S., Fuld, M., Cheslock, K. C., and Paul, M. H., in McElroy, W. D.,
and Glass, B., Phosphorus metabolism, Baltimore, 246 (1951). 26. Ahmad, K., Schneider, H. G., and Strong, F. M., Arch. Biochem.,iB, 281 (1950). 27. Potter, V. R., and Reif, A. E., Federation Proc., 10, 234 (1951). 28. Slater, E. C., Nature, 166, 982 (1950). 29. Belitzer, V. A., and Tsibakova, E. T., Biokhimiya, 4, 516 (1939). 30. Ochoa, S., J. BioZ. Chem., 138, 751 (1941). 31. Slater, E. C., Biochem. J., 46, 484 (1950). 32. Pappenheimer, A. M., Jr., and Hendee, E. D., J. Biol. Chem., 180, 597 (1949). 33. Clark, H. W., Neufeld, H. A., and Stotz, E., Federation Proc., 10, 172 (1951). 34. Lipmann, F., Cold Spring Harbor Symposia Quant. Biol., 7, 248 (1939). 35. Negelein, E., and Bromel, H., Biochem. Z., 303, 132 (1939). 36. Hunter, F. E., Jr., and Spector, S., Federation Proc., 10, 201 (1951). 37. Stadtman, E. R., Doudoroff, M., and Lipmann, F., J. BioZ. Chem., 191,377 (1951). 38. Friedkin, M., and Lehninger, A. L., J. Biol. Chem., 178, 611 (1949). 39. Judah, J. D., Biochem. J., 49, 271 (1951).
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from

J. H. Copenhaver, Jr. and Henry A. LardyMITOCHONDRIAL PREPARATIONS
PATHWAYS AND YIELD IN OXIDATIVE PHOSPHORYLATIONS:
1952, 195:225-238.J. Biol. Chem.
http://www.jbc.org/content/195/1/225.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/195/1/225.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on July 16, 2018http://w
ww
.jbc.org/D
ownloaded from


















