
Oxidation of Benzaldehydes to Benzoic Acid Derivatives by Three ...
6
Vol. 56, No. 7 APPLIED AND ENVIRONMENTAL MICROBIOLOGY, JUlY 1990, p. 2228-2233 0099-2240/90/072228-06$02.00/0 Oxidation of Benzaldehydes to Benzoic Acid Derivatives by Three Desulfovibrio Strains GERHARD ZELLNER,' HELMUT KNEIFEL,2 AND JOSEF WINTER'* Department of Microbiology, University of Regensburg, Universitatsstrasse 31, D-8400 Regensburg,1 and Department of Biotechnology 3, Kernforschungsanlage Julich GmbH, D-5170 Jiilich, Federal Republic of Germany Received 17 January 1990/Accepted 4 May 1990 Desulfovibrio vulgaris Marburg, "Desulfovibrio simplex" XVI, and Desulfovibrio sp. strain MP47 used benzaldehydes such as vanillin, 3,4,5-trimethoxybenzaldehyde, protocatechualdehyde, syringaldehyde, p- anisaldehyde, p-hydroxybenzaldehyde, and 2-methoxybenzaldehyde as electron donors for sulfate reduction and carbon dioxide and/or components of yeast extract as carbon sources for cell synthesis. The aldehydes were oxidized to their corresponding benzoic acids. The three sulfate reducers oxidized up to 7 mM vanillin and up to 4 mM p-anisaldehyde. Higher concentrations of vanillin or p-anisaldehyde were toxic. In addition, pyridoxal hydrochloride and o-vanillin served as electron donors for sulfate reduction. Salicylaldehyde, pyridine-2- aldehyde, pyridine-4-aldehyde, and 4-hydroxy-3-methoxybenzylalcohol were not oxidized. No molecular hydrogen was detected in the gas phase. The oxidized aldehydes were not further degraded. Lignin is an inexpensive source of phenolic compounds (24). Several aromatic compounds, including vanillin, co- niferyl aldehyde, syringaldehyde, sinapaldehyde, p-hydrox- ybenzoic acid, vanillic acid, and syringic acid, have been extracted from hydrolyzed aspen wood (51). Sulfite pulp, a wastewater of cellulose production (43), is the main source for, e.g., vanillin, a flavoring agent for beverages, foods, confectionery, and perfumery. Microbial degradation of lignin has been investigated for more than 50 years (37) but is still not totally understood. The initial depolymerization steps were regarded as aerobic reactions, catalyzed by fungi (14) or by bacteria (15, 16) such as Xanthomonas sp. (42) and Streptomyces sp. (13). Ferulic acid, cinnamic acid, veratrylaldehyde, homocatechol, 3- methoxy-4-hydroxytoluene, syringic acid, vanillic acid, and vanillin, which could be degraded aerobically and anaerobi- cally, were found as metabolites (1, 6, 21, 22, 27, 29, 49, 58, 59). Some of these compounds transiently inhibited solvent production from xylose by Klebsiella pneumoniae, but van- illin was apparently detoxified by conversion to vanillyl alcohol (41). Anaerobic degradation of lignin monomers may proceed under sulfate-reducing and methanogenic conditions, by nitrate respiration, by anaerobic photocatabolism, and by homoacetogenic fermentation (7, 12, 19, 27, 50). However, the participating microorganisms have not been character- ized in detail. In mixed methanogenic cultures, vanillin, vanillic acid, and several other methoxylated aromatic com- pounds were degraded to CO2 and CH4 (27, 28, 30, 31). The catabolism of these compounds included demethoxylation or demethylation (18) and other tranformation reactions, which finally could lead to ring fission and degradation to CO2 (34). While Acetobacterium woodii (2), Clostridium thermoaceti- cum, Clostridium thermoautotrophicum (17), an unnamed gram-negative anaerobe (21, 22), and Pseudomonas sp. strain PN-1 (53) carried out demethoxylation of, e.g., vanil- lin, other anaerobes catalyzed demethoxylations or decar- boxylations prior to ring fission (50). While Streptomyces viridosporus could oxidize vanillin to vanillic acid (47), A. woodii (2) or Syntrophococcus sucromutans (36) trans- * Corresponding author. formed vanillic acid to protocatechuic acid and used the eliminated substituents for homoacetogenesis. Chen et al. (10) reported the oxidation of dehydrodivanillin to dehy- drodivanillic acid and the further bioconversion of dehy- drodivanillic acid to vanillic acid by anaerobic bacteria from rumen fluid. A complete degradation of certain aromatic compounds was catalyzed by pure cultures of sulfate-reducing bacteria, e.g., the degradation of benzoate was catalyzed by Desulfo- coccus multivorans, Desulfosarcina variabilis, or Desul- fonema magnum (46); that of indole was catalyzed by Desulfobacterium indolicum (3); that of phenol was cata- lyzed by Desulfobacterium phenolicum (4); and that of catechol was catalyzed by Desulfobacterium catecholicum (52). In this paper, we report on the ability of three Desul- fovibrio strains to oxidize several substituted benzaldehydes to the respective benzoic acid derivatives under sulfate- reducing conditions with CO2 as a primary carbon source. MATERIALS AND METHODS Organisms. Desulfovibrio vulgaris Marburg DSM 2119 was obtained from Hans Hippe, Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Federal Republic of Germany. "Desulfovibrio simplex" XVI DSM 4141 (60) and Desulfovibrio sp. strain MP47 were isolated from a laboratory digester for the biomethanation of whey (62, 63). Strain MP47 is presumably another strain of Desulfovibrio vulgaris (63). Cultivation of organisms. Desulfovibrio vulgaris Marburg, "Desulfovibrio simplex" XVI, and Desulfovibrio sp. strain MP47 were routinely cultured in a medium that contained the following (per liter): 4.25 g of Na2SO4, 0.255 g of K2HPO4, 0.255 g of KH2PO4, 0.255 g of (NH4)2S04, 0.45 g of NaCl, 0.10 g of MgSO4 - 7H20, 2.0 g of yeast extract (no. 3753; E. Merck AG, Darmstadt, Federal Republic of Germany), 60 mg of CaCl2, 4 mg of FeSO4, 24 mg of Ni(NH4)2(SO4)2, and 1 mg of resazurin. The medium was supplemented with 10 ml each of vitamin and mineral solutions and reduced by the addition of 0.5 g of L-cysteine hydrochloride and 0.5 g of Na2S 9H20 per liter, according to the method of Balch et al. (5). Serum bottles received 20 ml of medium and an H2-C02 (80:20%; 300 kPa) gas atmo- 2228
Transcript of Oxidation of Benzaldehydes to Benzoic Acid Derivatives by Three ...

Vol. 56, No. 7APPLIED AND ENVIRONMENTAL MICROBIOLOGY, JUlY 1990, p. 2228-22330099-2240/90/072228-06$02.00/0
Oxidation of Benzaldehydes to Benzoic Acid Derivatives by ThreeDesulfovibrio Strains
GERHARD ZELLNER,' HELMUT KNEIFEL,2 AND JOSEF WINTER'*Department of Microbiology, University of Regensburg, Universitatsstrasse 31, D-8400 Regensburg,1 and Department of
Biotechnology 3, Kernforschungsanlage Julich GmbH, D-5170 Jiilich, Federal Republic of Germany
Received 17 January 1990/Accepted 4 May 1990
Desulfovibrio vulgaris Marburg, "Desulfovibrio simplex" XVI, and Desulfovibrio sp. strain MP47 usedbenzaldehydes such as vanillin, 3,4,5-trimethoxybenzaldehyde, protocatechualdehyde, syringaldehyde, p-anisaldehyde, p-hydroxybenzaldehyde, and 2-methoxybenzaldehyde as electron donors for sulfate reductionand carbon dioxide and/or components of yeast extract as carbon sources for cell synthesis. The aldehydes wereoxidized to their corresponding benzoic acids. The three sulfate reducers oxidized up to 7 mM vanillin and upto 4 mM p-anisaldehyde. Higher concentrations of vanillin or p-anisaldehyde were toxic. In addition, pyridoxalhydrochloride and o-vanillin served as electron donors for sulfate reduction. Salicylaldehyde, pyridine-2-aldehyde, pyridine-4-aldehyde, and 4-hydroxy-3-methoxybenzylalcohol were not oxidized. No molecularhydrogen was detected in the gas phase. The oxidized aldehydes were not further degraded.
Lignin is an inexpensive source of phenolic compounds(24). Several aromatic compounds, including vanillin, co-niferyl aldehyde, syringaldehyde, sinapaldehyde, p-hydrox-ybenzoic acid, vanillic acid, and syringic acid, have beenextracted from hydrolyzed aspen wood (51). Sulfite pulp, awastewater of cellulose production (43), is the main sourcefor, e.g., vanillin, a flavoring agent for beverages, foods,confectionery, and perfumery.
Microbial degradation of lignin has been investigated formore than 50 years (37) but is still not totally understood.The initial depolymerization steps were regarded as aerobicreactions, catalyzed by fungi (14) or by bacteria (15, 16) suchas Xanthomonas sp. (42) and Streptomyces sp. (13). Ferulicacid, cinnamic acid, veratrylaldehyde, homocatechol, 3-methoxy-4-hydroxytoluene, syringic acid, vanillic acid, andvanillin, which could be degraded aerobically and anaerobi-cally, were found as metabolites (1, 6, 21, 22, 27, 29, 49, 58,59). Some of these compounds transiently inhibited solventproduction from xylose by Klebsiella pneumoniae, but van-illin was apparently detoxified by conversion to vanillylalcohol (41).Anaerobic degradation of lignin monomers may proceed
under sulfate-reducing and methanogenic conditions, bynitrate respiration, by anaerobic photocatabolism, and byhomoacetogenic fermentation (7, 12, 19, 27, 50). However,the participating microorganisms have not been character-ized in detail. In mixed methanogenic cultures, vanillin,vanillic acid, and several other methoxylated aromatic com-pounds were degraded to CO2 and CH4 (27, 28, 30, 31). Thecatabolism of these compounds included demethoxylation ordemethylation (18) and other tranformation reactions, whichfinally could lead to ring fission and degradation to CO2 (34).While Acetobacterium woodii (2), Clostridium thermoaceti-cum, Clostridium thermoautotrophicum (17), an unnamedgram-negative anaerobe (21, 22), and Pseudomonas sp.strain PN-1 (53) carried out demethoxylation of, e.g., vanil-lin, other anaerobes catalyzed demethoxylations or decar-boxylations prior to ring fission (50). While Streptomycesviridosporus could oxidize vanillin to vanillic acid (47), A.woodii (2) or Syntrophococcus sucromutans (36) trans-
* Corresponding author.
formed vanillic acid to protocatechuic acid and used theeliminated substituents for homoacetogenesis. Chen et al.(10) reported the oxidation of dehydrodivanillin to dehy-drodivanillic acid and the further bioconversion of dehy-drodivanillic acid to vanillic acid by anaerobic bacteria fromrumen fluid.A complete degradation of certain aromatic compounds
was catalyzed by pure cultures of sulfate-reducing bacteria,e.g., the degradation of benzoate was catalyzed by Desulfo-coccus multivorans, Desulfosarcina variabilis, or Desul-fonema magnum (46); that of indole was catalyzed byDesulfobacterium indolicum (3); that of phenol was cata-lyzed by Desulfobacterium phenolicum (4); and that ofcatechol was catalyzed by Desulfobacterium catecholicum(52). In this paper, we report on the ability of three Desul-fovibrio strains to oxidize several substituted benzaldehydesto the respective benzoic acid derivatives under sulfate-reducing conditions with CO2 as a primary carbon source.
MATERIALS AND METHODSOrganisms. Desulfovibrio vulgaris Marburg DSM 2119
was obtained from Hans Hippe, Deutsche Sammlung vonMikroorganismen und Zellkulturen GmbH, Braunschweig,Federal Republic of Germany. "Desulfovibrio simplex" XVIDSM 4141 (60) and Desulfovibrio sp. strain MP47 wereisolated from a laboratory digester for the biomethanation ofwhey (62, 63). Strain MP47 is presumably another strain ofDesulfovibrio vulgaris (63).
Cultivation of organisms. Desulfovibrio vulgaris Marburg,"Desulfovibrio simplex" XVI, and Desulfovibrio sp. strainMP47 were routinely cultured in a medium that containedthe following (per liter): 4.25 g of Na2SO4, 0.255 g ofK2HPO4, 0.255 g of KH2PO4, 0.255 g of (NH4)2S04, 0.45 gof NaCl, 0.10 g of MgSO4 - 7H20, 2.0 g of yeast extract(no. 3753; E. Merck AG, Darmstadt, Federal Republic ofGermany), 60 mg of CaCl2, 4 mg of FeSO4, 24 mg ofNi(NH4)2(SO4)2, and 1 mg of resazurin. The medium wassupplemented with 10 ml each of vitamin and mineralsolutions and reduced by the addition of 0.5 g of L-cysteinehydrochloride and 0.5 g of Na2S 9H20 per liter, accordingto the method of Balch et al. (5). Serum bottles received 20ml of medium and an H2-C02 (80:20%; 300 kPa) gas atmo-
2228

OXIDATION OF BENZALDEHYDES 2229
sphere. The pH of the medium was adjusted to 7. Cultureswere grown at 37°C on a rotary shaker at 100 rpm (InforsGmbH, Munich, Federal Republic of Germany). Utilizationof yeast extract components as electron donors was tested inthe same medium with an N2-CO2 (80:20%; 300 kPa) gasatmosphere.For tests of aromatic compounds as alternative hydrogen
donors because of their limited solubility, exact amounts wereweighed into serum bottles, which were then transferred intoan anaerobic chamber and filled with medium. The gas phaseof the anaerobic chamber housing the serum bottles wasreplaced by N2-CO2 (80:20%; 300 kPa) at a gassing manifold(5) before the bottles were sterilized in an autoclave.
Autooxidation of benzaldehyde to benzoic acid was ex-cluded by analysis of both uninoculated and autoclavedcontrols in parallel with cultures of the Desulfovibrio teststrains. No oxidation was measurable during the period ofincubation. In addition, no oxidation of the benzaldehydeswas observed when autoclaved cells were used.Measurement of growth. Optical density of cultures was
measured with a photometer (Stasar II; Gilford InstrumentLaboratories, Inc., Oberlin, Ohio) at a wavelength of 578nm. Samples were withdrawn from the serum bottles with asyringe, transferred into a quartz cuvette, and reduced witha grain of sodium dithionite. All growth measurements werecarried out with the same cuvette and controlled by micro-scopic counting. Optical density measurements of parallelcultures were corrected for the absorbance measured imme-diately after inoculation and for the absorbance reached incontrol cultures in the absence of an added electron donorfor sulfate reduction.
Analyses. (i) UV-visible-spectrum spectroscopy. The spec-tra of filtered samples (disposable filters, 0.2-,um pore size;Sartorius GmbH, Gottingen, Federal Republic of Germany)and of authentic compounds were recorded on a double-beam spectrophotometer (UVIKON 860; Kontron, Eching,Federal Republic of Germany). Spectra of substrates andtransformation products were compared with those of refer-ence compounds.
(ii) Separation of aromatic compounds by thin-layer chro-matography. Aromatic substances were chromatographedon thin-layer plates (silica gel 60 F254; Merck). The solventsystems used were TLC1 (1-butanol-acetic acid-water, 4:1:1[vol/vol/vol]), TLC2 (2-butanol-formic acid-water, 6:1:2[vol/vol/vol]), TLC3 (diethylether-acetic acid, 90:5 [vol/vol]), TLC4 (benzene-ethylacetate, 9:1 [vollvol]), TLC5 (2-propanol-benzene-concentrated ammonium hydroxide, 5:4:1 [vol/vol/vol]), and TLC6 (chloroform-ethanol-aceticacid, 18:1:1 [vol/vol/vol]). Compounds were detected viaquenching of fluorescence upon illumination with UV light(wavelength, 254 nm). The best separation of aldehydes andacids was achieved with solvent systems TLC5 and TLC6with the following Rf values for authentic references (Rfs ofTLC5 and of TLC6): vanillin, 0.22/0.76; vanillic acid, 0.04/0.61; p-hydroxybenzaldehyde, 0.29/0.42; p-hydroxybenzoicacid, 0.05/0.35; protocatechualdehyde, 0.10/0.18; protocate-chuic acid, 0.12/0.09; syringaldehyde, 0.12/0.70; syringicacid, 0.03/0.56; 3,4,5-trimethoxybenzaldehyde, not deter-mined/0.99; 3,4,5-trimethoxybezoic acid, not determined/0.69; and benzoic acid, 0.13/0.73.
(iii) HPLC of aromatic compounds. Aromatic compoundswere separated with a high-performance liquid chromatog-raphy (HPLC) system (Kontron) equipped with a gradientprogrammer (model 205), two pumps (model T-414), and aUVIKON 820 UV-visible-light spectrum spectrophotome-ter. Peaks were quantified with a Shimadzu C-R3A integra-
tor. Separation of compounds was achieved by using areverse-phase column (inside diameter, 0.5 cm, length, 25cm, 5-,um particles, Spherisorb 5 ODS 2; Kontron) at roomtemperature with the following elution system. Solvent Awas 5% (vol/vol) aqueous acetic acid. Solvent B was 70%(vol/vol) aqueous methanol. The flow rate was 0.75 ml/minthroughout the separation, with a gradient as follows: 0 to 1min, 100% solvent A; 2 to 15 min, linear gradient from 100%solvent A to 50% solvent A-50% solvent B; 16 to 18 min,50% solvent A-50% solvent B; and 19 to 24 min, 100%solvent A. For elution of syringaldehyde and p-anisalde-hyde, 100% solvent B instead of 50% A-50% B was used.UV absorption (wavelength, 254 nm) was measured. Somesubstances (see Table 2) were additionally analyzed with anHPLC system (Waters Millipore GmbH, Eschborn, FederalRepublic of Germany) consisting of two M6000A pumps, agradient controller (M680), an automatic injector (WISP710B), a photodiode array detector (1040; Hewlett Packard,Munich, Federal Republic of Germany), and an integrator(D-2000; Merck). Separation was achieved with a reverse-phase column (15 by 0.4 cm, 5-,im particles, ODS 1; Latek,Heidelberg, Federal Republic of Germany). Elution of cul-ture samples and reference substances was at room temper-ature with the following gradient program: 0 to 60% aceto-nitrile in 0.1 M ortho-phosphoric acid within 20 min, with aflow rate of 1 mllmin. All samples for HPLC were passedthrough membrane filters (0.2-,um-pore-size cellulose acetateor 0.45-,um-pore-size cellulose nitrate; Minisart, SartoriusGmbH) prior to injection. Double-distilled water and analyt-ical grade solvents were filtered through sintered glass witha pore size of 1.6 iLm prior to utilization. Solvents weredegassed under a vacuum in an ultrasonic water bath,regassed with oxygen-free nitrogen, and stored anaerobi-cally until use.
(iv) Gas chromatography-mass spectrometry analysis ofcompounds. Gas chromatography-mass spectrometry analy-ses were carried out according to the method of Gupta et al.(25).
(v) Analysis of fatty acids, hydrogen, hydrogen sulfide, andsulfate. Fatty acids, H2, and H2S were determined by gaschromatography as described earlier (61). Sulfate was deter-mined colorimetrically (54).
Chemicals. Vanillin, o-vanillin, vanillic acid, benzalde-hyde, benzoic acid, p-hydroxybenzaldehyde, p-hydroxyben-zoic acid, 3,4,5-trimethoxybenzaldehyde, 3,4,5-trimethoxy-benzoic acid, protocatechualdehyde, protocatechuic acid,syringaldehyde, syringic acid, 2-methoxybenzaldehyde, 2-methoxybenzoic acid, DL-mandelic acid, p-anisaldehyde,p-anisic acid, and pyridoxal hydrochloride were of analyticalgrade and were purchased from Serva (Heidelberg, FederalRepublic of Germany), Merck, or Aldrich Chemie GmbH &CoKG (Steinheim, Federal Republic of Germany). All otherchemicals were of analytical grade and were purchased fromMerck.
RESULTS
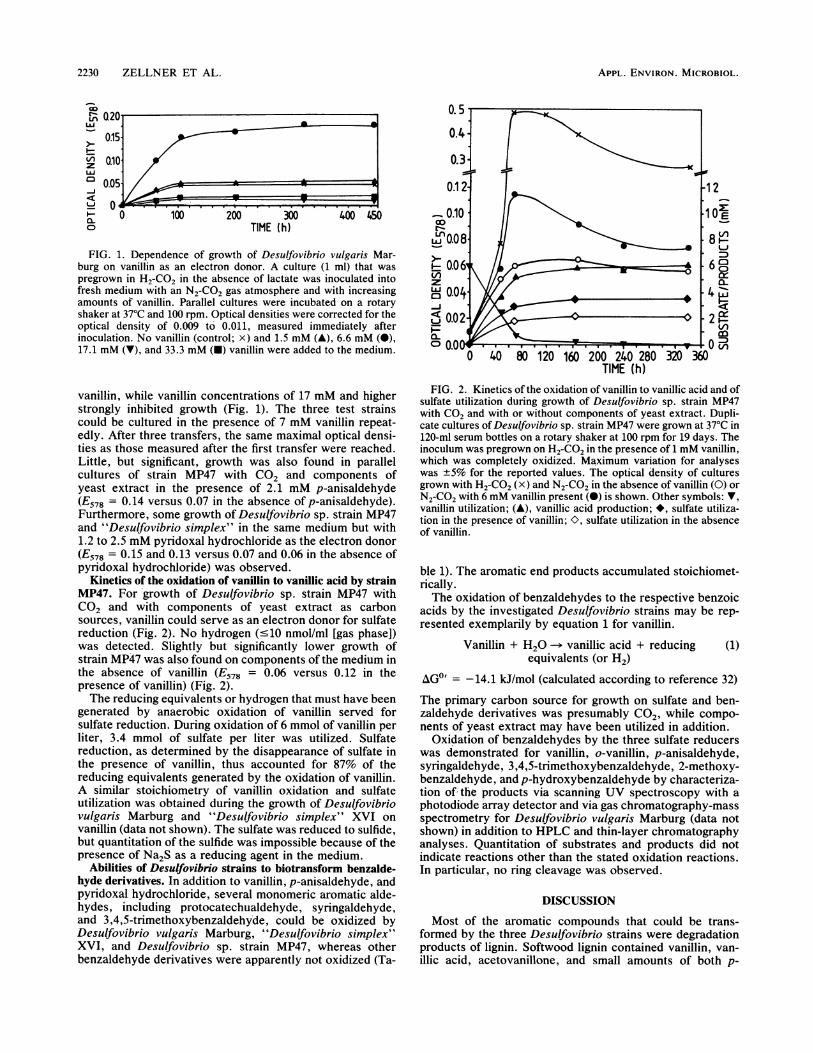
Oxidation of vanillin, p-anisaldehyde, and pyridoxal hydro-chloride by Desulfovibrio strains. Desulfovibrio vulgaris Mar-burg, "Desulfovibrio simplex" XVI, and Desulfovibrio sp.strain MP47 used vanillin as an electron donor for sulfatereduction and CO2 and/or components of yeast extract ascarbon sources. The maximal optical densities of Desul-fovibrio vulgaris Marburg (E578 = 0.17) (Fig. 1), "Desul-fovibrio simplex" (E578 = 0.1), and Desulfovibrio sp. strainMP47 (E578 = 0.25) were obtained in the presence of 7 mM
VOL. 56, 1990
2230 ZELLNER ETAL.APLENRO.MCBo.
Goi
I-05
C) TIME (h)
FIG. 1. Dependence of growth of Desulfovibrio vulgaris Mar-burg on vanillin as an electron donor. A culture (1 ml) that waspregrown in H2-C02 in the absence of lactate was inoculated intofresh medium with an N2-C02 gas'atmosphere and with increasingamounts of vanillin. Parallel cultures were incubated on a rotaryshaker at 370C and 100 rpm. Optical densities were corrected for theoptical density of 0.009 to' 0.011, measured immediately afterinoculation. No vanillin (control; x) and 1.5 mM (A), 6.6 mM (0),17.1 mM (V), and 33.3 mM (U) vanillin were added to the medium.
vanillin, while vanillin concentrations of 17 mM and higherstrongly inhibited growth (Fig. 1). The three test strainscould be cultured in the presence of 7 mM va'nillin repeat-edly. After three transfers, the same maximal optical densi-ties as those measured after the first transfer were reached.Little, but significant, growth was also found in parallelcultures of strain MP47 with CO2 and components ofyeast extract in the presence of 2.1 mM p-anisaldehyde(E578 = 0.14 versus 0.07 in the absence of p-anisaldehyde).Furthermore, some gro'wth of Desulfovibrio sp. strain MP47and "Desulfovibric simplex" in the same medium but with1.2 to 2.5 mM pyridoxal hydrochloride as the electron donor(E578 = 0.15 and 0.13 versus 0.07 and 0.06 in the absence ofpyridoxal hydrochloride) was observed.
Kinetics of the oxidation of vanillin to vanillic acid by strainMP47. For growth of Desulfovibrio sp. strain MP47 withCO2 and with components of yeast extract as carbonsources, vanillin could serve as an electron donor for sulfatereduction (Fig. 2). No hydrogen (-<10 nmol/ml [gas phase])was detected. Slightly but significantly lower growth ofstrain MP47 was also found on components of the medium inthe absence of vanillin (E578 = 0.06 versus 0.12 in thepresence of vanillin) (Fig. 2).The reducing equivalents or hydrogen that must have been
generated by anaerobic oxidation of vanillin served forsulfate reduction. During oxidation of 6 mmol of vanillin perliter, 3.4 mmol of sulfate per liter was utilized. Sulfatereduction, as determined by the disappearance of sulfate inthe presence, of vanillin, thus accounted for 87% of thereducing equivalents generated by the oxidation of vanillin.A similar stoichiometry of van'illin oxidation and sulfateutilization was obtained during the growth of Desulfovibriovulgaris Marburg and "Desulfovibrio simplex" XVI onvanillin (data not shown). The sulfate was reduced to sulfide,but quantitation of the sulfide was impossible because of thepresence of Na2S as a reducing agent in the medium.
Abilities of Desulfovibrio strains to biotransform benzalde-hyde derivatives. In addition to vanillin, p-anisaldehyde, andpyridoxal hydrochloride, several monomeric aromatic aide-hydes, including protocatechualdehyde, syringaldehyde,and 3,4,5-trimethoxybenzaldehyde, could be oxidized byDesulfovibrio vulgaris Marburg, "Desulfovibrio simplex"XVI, and Desulfovibrio sp. strain MP47, whereas otherbenzaldehyde derivatives were apparently not oxidized (Ta-
40 80 120 160 200 240 280 320
TIME (h)
Li
CDc
L/)
FIG. 2. Kinetics of the oxidation of vanillin to vanillic acid and ofsulfate utilization during growth of Desulfovibrio sp. strain MP47with CO2 and with or without components of yeast extract. Dupli-cate cultures of Desulfovibrio sp. strain MP47 were grown at 370C in120-mi serum bottles on a rotary shaker at 100 rpm for 19 days. Theinoculum was pregrown on H2-C02 in the presence of 1 mM vanillin,which was completely oxidized. Maximum variation for analyseswas ±5% for the reported values. The optical density of culturesgrown with H2-C02 (x) and N2-C02 in the absence of vanillin (0) orN2-C02 with 6 mM vanillin present (0) is shown. Other symbols: V,vanillin utilization; (A), vanillic acid production; *, sulfate utiliza-tion in the presence of vanillin; K>, sulfate utilization in the absenceof vanillin.
ble 1). The aromatic end products accumulated stoichiomet-rically.The oxidation of benzaldehydes to the respective benzoic
acids by the investigated Desulfovibrio strains may be rep-resented exemplarily by equation 1 for vanillin.
Vanillin + H20 ---* vanillic acid + reducingequivalents (or H2)
(1)
AGI' = -14.1 kJ/mol (calculated according to reference 32)The primary carbon source for growth on sulfate and ben-zaldehyde derivatives was presumably C02, while compo-nents of yeast extract may have been utilized in addition.
Oxidation of benzaldehydes by the three sulfate reducerswas demonstrated for vanillin, o-vanillin, p-anisaldehyde,syringaldehyde, 3,4,5-trimethoxybenzaldehyde, 2-methoxy-benzaldehyde, and p-hydroxybenzaldehyde by characteriza-tion of, the products via scanning UV spectroscopy with aphotodiode 'array detector and via gas chromatography-massspectrometry for Desulfovibrio vulgaris Marbur'g (data notshown) in addition to HPLC and thin-layer chromatographyanalyses. Quantitation of substrates and products did notindicate reactions other than the stated oxidation reactions.In particular, no ring cleavage was observed.
DISCUSSION
Most of the aromatic compounds that could be trans-formed by the three Desulfovibrio strains were degradationproducts of lignin. Softwood lignin contained vanillin, van-illic acid, acetovanilione, and small amounts of both p-
APPL. ENVIRON. MICROBIOL.

OXIDATION OF BENZALDEHYDES 2231
TABLE 1. Biotransformation of aromatic aldehydes bysulfate-reducing bacteria
Growth of':
Compounds Desulfovibrio Desulfovibrio Desulfovibriovulgaris sp. strain sp. strainMarburg MP47 XVI
Vanillin + + +o-Vanillin +bb _Vanillic acid - - -Protocatechualdehyde +CProtocatechuic acidSyringaldehyde + + +Syringic acid3,4,5-Trimethoxy- + + +benzaldehyde
3,4,5-Trimethoxy- - - NDbenzoic acid
p-Hydroxybenzaldehyde + + NDp-Hydroxybenzoic acid - - ND2-Methoxybenzaldehyde + + ND2-Methoxybenzoic acidp-Anisaldehyde + + +p-Anisic acidPhenolPyridoxal hydrochloride + + +
a -, No growth, compound not transformed; +, little growth (E578 = 0.05to 0.2), more than 80% of supplied compound transformed to the respectivebenzoic acid; ND, not determined. Parallel cultures in serum bottles contain-ing 20 ml of medium and 250 mg of any one of the substrates mentioned aboveper liter were incubated under N2-CO2 (80:20%, 300 kPa) at 37°C on a shaker(100 rpm) for 4 weeks. Biotransformation of the substrates was analyzed by atleast two methods, including HPLC with two detector systems, UV spectros-copy at 254 nm (293 nm for pyridoxal hydrochloride), and thin-layer chroma-tography.
b No growth; however, there was oxidation after more than 4 weeks ofincubation.
c No growth, but there was transformation to protocatechuic acid.
hydroxybenzaldehyde and syringaldehyde, while in hard-wood lignin, higher amounts of syringaldehyde were found.Grass lignin and cane lignin were sources ofp-hydroxybenz-aldehyde (24, 38, 51). A variety of mono- and dinuclearphenolic compounds, including p-hydroxybenzaldehyde, p-hydroxybenzoic acid, protocatechualdehyde, protocate-chuic acid, vanillin, vanillic acid, carboxyvanillin, carboxy-vanillic acid, syringaldehyde, and syringic acid, could beobtained through hydrolysis of lignin (26).
In the presence of oxygen, oxidation of vanillin to vanillicacid was shown for Streptomyces viridosporus (47) andSerratia marcescens (45). The oxidation of vanillin tovanillic acid as an intermediate step in the degradation oftrans-ferulic acid has been demonstrated for Pseudomonasfluorescens and Pseudomonas acidovorans (9, 56). ANAD+-dependent alcohol dehydrogenase in a Pseudomonassp. was shown to catalyze the oxidation of vanillin to vanillicacid (44). The oxidation of benzaldehyde to benzoic acid aswell as the oxidation of p-hydroxybenzaldehyde to p-hy-droxybenzoic acid was involved in the degradation of man-delic acid by Pseudomonas putida, Pseudomonas convexa,Acinetobacter calcoaceticus, and Aspergillus niger (20). Inaddition, only recently halogenated aromatic aldehydes wereshown to be oxidized to the corresponding aromatic acids bymixed cultures, but the oxidation could not be assigned to aspecific metabolic group of bacteria (40).
In this paper, the oxidation of vanillin to vanillic acid inthe absence of oxygen was demonstrated for three Desul-fovibrio strains that were able to growth with CO2 as thecarbon source (46, 60, 63). While the reduction of sulfate
with molecular hydrogen is exergonic with a AG0' of -151.9kJ/mol under standard conditions (55), the oxidation ofvanillin for the generation of hydrogen or reducing equiva-lents may add little additional energy (AG0' = -14.1 kJ/mol,calculated according to reference 32). In contrast to thepossibility of an unlimited supply of hydrogen as an electrondonor for sulfate reducers, vanillin and other benzaldehydederivatives could only be supplied at low, growth-limitingconcentrations, because of their inhibitory effects at higherconcentrations (Fig. 1). However, in nature, lignin degrada-tion proceeds only at low rates and thus the concentration ofbenzaldehyde derivatives in anaerobic ecosystems presum-ably stays well below inhibitory concentrations for sulfatereducers.The toxicity of several plant phenolic acids and aldehydes
for rumen bacteria and Klebsiella pneumoniae has beenextensively investigated (8, 11, 41, 57). Severe structuraldamage to the cell walls of Bacteroides succinogenes, Ru-minococcus flavefaciens, and Ruminococcus albus wascaused by 50 mM vanillic acid (8, 11). However, the pres-ence of up to 10 mM vanillic acid had no effect or only asmall inhibitory effect on the cellulolytic activity of thesestrains, whereas the presence of 10 mM syringic, 4-hydroxy-benzoic, or hydrocinnamic acid might even stimulate cellu-lose decomposition. For the Desulfovibrio strains tested, thetoxicity of vanillin apparently was in the same range as thatfor rumen microorganisms.
Vanillic acid, produced from vanillin by sulfate reducers,could be degraded to protocatechuic acid and acetate bypure cultures of Acetobacterium woodii (2). Therefore, inaddition to demethoxylation (2, 9, 17, 35, 39), the oxidationof the aldehyde function to a carboxy function might facili-tate ring cleavage by other organisms. For anaerobic phenoldegradation, carboxylation to benzoate has been shown asan essential step in undefined (23) and defined consortia (33).While the degradation or transformation of several aro-
matic compounds by pure cultures of sulfate-reducing bac-teria was recently shown (3, 4, 46, 52), for neither one ofthem, nor for one of the presently known sulfate-reducingbacteria (46, 48), has the biotransformation of benzaldehydederivatives to the respective benzoic acids been reported.The three Desulfovibrio strains used in this study couldbiotransform several benzaldehydes to their respective ben-zoic acids; however, none of the tested strains was able tocatalyze further tranformation or even a complete degrada-tion of the generated benzoic acid derivative. The energeticbenefit of benzaldehyde oxidation for the sulfate reducersand the ecological significance remain to be investigated indetail in future work. For further degradation of benzoic acidderivatives, other sulfate reducers, such as Desulfococcus,Desulfonema, and Desulfosarcina strains, which are knownto degrade at least unsubstituted benzoate, might be goodcandidates (46).
ACKNOWLEDGMENTS
This investigation was supported by funds of the Bundesministe-rium fur Forschung und Technologie.We gratefully acknowledge the expert recording of mass spectra
carried out by Cornelia Jebsen.
LITERATURE CITED
1. Aulin-Erdtman, G., and L. Hegbom. 1957. Spectrographic con-tributions to lignin chemistry VII. Sven. Papperstidn. 60:671-681.
2. Bache, R., and N. Pfennig. 1981. Selective isolation of Aceto-
VOL. 56, 1990

2232 ZELLNER ET AL.
bacterium woodii on methoxylated aromatic acids and determi-nation of growth yields. Arch. Microbiol. 130:255-261.
3. Bak, F., and F. Widdel. 1986. Anaerobic degradation of indoliccompounds by sulfate-reducing enrichment cultures, and de-scription of Desulfobacterium indolicum gen. nov., sp. nov.Arch. Microbiol. 146:170-176.
4. Bak, F., and F. Widdel. 1986. Anaerobic degradation of phenoland phenol derivatives by Desulfobacterium phenolicum sp.nov. Arch. Microbiol. 146:177-180.
5. Balch, W. E., G. E. Fox, L. J. Magrum, C. R. Woese, and R. S.Wolfe. 1979. Methanogens: reevaluation of a unique biologicalgroup. Microbiol. Rev. 43:260-296.
6. Benner, R., A. E. Maccubin, and R. E. Hodson. 1984. Anaerobicbiodegradation of the lignin and polysaccharide components oflignocellulose and synthetic lignin by sediment microflora.Appl. Environ. Microbiol. 47:998-1004.
7. Berry, D. F., A. J. Francis, and J.-M. Bollag. 1987. Microbialmetabolism of homocyclic and heterocyclic aromatic com-pounds under anaerobic conditions. Microbiol. Rev. 51:43-59.
8. Borneman, W. S., D. E. Akin, and W. P. vanEseltine. 1986.Effect of phenolic monomers on ruminal bacteria. Appl. Envi-ron. Microbiol. 52:1331-1339.
9. Cartwright, N. J., and A. R. W. Smith. 1967. Bacterial attack onphenolic ethers. Biochem. J. 102:826-841.
10. Chen, W., K. Ohmiya, S. Shimizu, and H. Kawakami. 1985.Degradation of dehydrodivanillin by anaerobic bacteria fromcow rumen fluid. Appl. Environ. Microbiol. 49:211-216.
11. Chesson, A., C. S. Stewart, and R. J. Wallace. 1982. Influence ofplant phenolic acids on growth and cellulolytic activity of rumenbacteria. Appl. Environ. Microbiol. 44:597-603.
12. Colberg, P. J., and L. Y. Young. 1982. Biodegradation oflignin-derived molecules under anaerobic conditions. Can. J.Microbiol. 28:886-889.
13. Crawford, D. L. 1981. Microbial conversions of lignin to usefulchemicals using a lignin-degrading Streptomyces. Biotechnol.Bioeng. Symp. 11:275-291.
14. Crawford, D. L., and R. L. Crawford. 1980. Microbial degrada-tion of lignin. Enzyme Microb. Technol. 2:11-22.
15. Crawford, R. L. 1981. Lignin biodegradation and transforma-tion, p. 154-167. J. Wiley & Sons, Inc., New York.
16. Crawford, R. L., and D. L. Crawford. 1984. Recent advances instudies of the mechanisms of microbial degradation of lignins.Enzyme Microb. Technol. 6:434-42.
17. Daniel, S. L., Z. Wu, and H. L. Drake. 1988. Growth ofthermophilic acetogenic bacteria on methoxylated aromaticacids. FEMS Microbiol. Lett. 52:25-28.
18. DeWeerd, K. A., A. Saxena, D. P. Nagle, and J. M. Suflita. 1988.Metabolism of the '8O-methoxy substituent of 3-methoxyben-zoic acid and other unlabeled methoxybenzoic acids by anaer-obic bacteria. Appl. Environ. Microbiol. 54:1237-1242.
19. Evans, W. C. 1977. Biochemistry of the bacterial catabolism ofaromatic compounds in anaerobic environments. Nature (Lon-don) 270:17-22.
20. Fewson, C. A. 1988. Microbial metabolism of mandelic acid: amicrocosm of diversity. FEMS Microbiol. Rev. 54:85-110.
21. Frazer, A. C., and L. Y. Young. 1985. A gram-negative anaer-obic bacterium that utilizes 0-methyl substituents of aromaticacids. Appl. Environ. Microbiol. 49:1345-1347.
22. Frazer, A. C., and L. Y. Young. 1986. Anaerobic C1 metabolismof the O-methyl-14C-labeled substituent of vanillate. Appl. En-viron. Microbiol. 51:84-87.
23. Genthner, B. R. S., G. T. Townsend, and P. J. Chapman. 1989.Anaerobic transformation of phenol to benzoate via para-car-boxylation. Use of fluorinated analogues to elucidate the mech-anism of transformation. Biochem. Biophys. Res. Commun.162:945-951.
24. Goldstein, I. S. 1975. Perspectives on production of phenols andphenolic acids from lignin and bark. Appl. Polymer Symp.28:259-267.
25. Gupta, J. K., C. Jebsen, and H. Kneifel. 1986. Sinapic aciddegradation by the yeast Rhodotorula glutinis. J. Gen. Micro-biol. 132:2793-2799.
26. Hanselmann, K. W. 1982. Lignochemicals. Experientia 38:176-189.
27. Healy, J. B., and L. Y. Young. 1979. Anaerobic biodegradationof eleven aromatic compounds to methane. Appl. Environ.Microbiol. 38:84-89.
28. Healy, J. B., Jr., L. Y. Young, and M. Reinhard. 1980. Meth-anogenic decomposition of ferulic acid, a model lignin deriva-tive. Appl. Environ. Microbiol. 39:436 444.
29. Higuchi, T. 1982. Biodegradation of lignin: biochemistry andpotential applications. Experientia 38:159-166.
30. Kaiser, J. P., and K. W. Hanselmann. 1982. Aromatic chemicalsthrough anaerobic microbial conversion of lignin monomers.Experientia 38:167-176.
31. Kaiser, J. P., and K. W. Hanselmann. 1982. Fermentativemetabolism of substituted monoaromatic compounds by a bac-terial community from anaerobic sediments. Arch. Microbiol.133:185-194.
32. Kienitz, H. 1961. Experimentelle theoretische GrundgroBen derThermochemie, p. 1-38. In H. R. Landolt-B6rnstein (ed.),Zahlenwerte und Funktionen aus Physik, Chemie, Astronomie,Geophysik und Technik, 6. Auflage, Bd. II. KalorischeZustandsgrofen. Springer Verlag KG, Berlin.
33. Knoll, G., and J. Winter. 1989. Degradation of phenol viacarboxylation by a defined, obligate syntrophic consortium ofanaerobic bacteria. Appl. Microbiol. Biotechnol. 30:318-324.
34. Kreikenbohm, R., and N. Pfennig. 1985. Anaerobic degradationof 3,4,5-trimethoxylbenzoate by a defined mixed culture ofAcetobacterium woodii, Pelobacter acidigallici, and Desulfo-bacter postgatei. FEMS Microbiol. Ecol. 31:29-38.
35. Krumholz, L. R., and M. P. Bryant. 1985. Clostridium pfennigiisp. nov. uses methoxyl groups of monobenzenoids and pro-duces butyrate. Int. J. Syst. Bacteriol. 35:454-456.
36. Krumholz, L. R., and M. P. Bryant. 1986. Syntrophococcussucromutans sp. nov. gen. nov. uses carbohydrates as electrondonors and formate, methoxymonobenzenoids or Methanobre-vibacter as electron acceptor systems. Arch. Microbiol. 143:313-318.
37. Levine, M., G. H. Nelson, D. Anderson, and P. B. Jacobs. 1935.Utilization of agricultural wastes. I. Lignin and microbial de-composition. Ind. Eng. Chem. 27:195-200.
38. McCarty, P. L., L. Y. Young, J. M. Gossett, 0. C. Stuckey, andJ. B. Healy. 1977. Heat treatment for increasing methane yieldfrom organic materials, 179-199. In H. G. Schlegel and J.Barnes (ed.), Microbial energy conversion. Pergamon Press,Oxford.
39. Mountfort, D. O., and W. D. Grant. 1988. Eubacterium callan-deri sp. nov. that demethoxylates 0-methoxylated aromaticacids to volatile fatty acids. Int. J. Syst. Bacteriol. 38:254-258.
40. Neilson, A. H., A.-S. Allard, P.-A. Hynning, and M. Remberger.1988. Transformation of halogenated aromatic aldehydes bymetabolically stable anaerobic enrichment cultures. Appl. En-viron. Microbiol. 54:2226-2236.
41. Nishikawa, N. K., R. Sutcliffe, and J. N. Saddler. 1988. Theinfluence of lignin degradation products on xylose fermentationby Klebsiella pneumoniae. Appl. Microbiol. Biotechnol. 27:549-552.
42. Odier, E., and B. C. R. Monties. 1978. Biodegradation of wheatlignin by Xanthomonas. Ann. Microbiol. 129A:361-377.
43. Pearl, I. A., and D. L. Beyer. 1950. Studies on lignin and relatedproducts. IV. The oxidation of fermented spent liquors withcupric oxide under pressure. TAPPI 33:544-548.
44. Pelmont, J., C. Tournesac, A. Mliki, M. Barrelle, and C. Beguin.1989. A new bacterial alcohol dehydrogenase active on de-graded lignin and several low molecular weight aromatic com-pounds. FEMS Microbiol. 57:109-114.
45. Perestelo, F., M. A. Falcon, and G. de la Fuente. 1989. Produc-tion of vanillic acid from vanillin by resting cells of Serratiamarcescens. Appl. Environ. Microbiol. 55:1660-1662.
46. Pfennig, N., F. Widdel, and H. G. Truper. 1981. The dissimila-tory sulfate-reducing bacteria, 926-940. In M. P. Starr, H.Stolp, H. G. Truper, A. Balows, and H. G. Schlegel (ed.), Theprocaryotes. Springer-Verlag, Berlin.
47. Pometto, A. L., III, and D. L. Crawford. 1983. Whole-cell
APPL. ENVIRON. MICROBIOL.

OXIDATION OF BENZALDEHYDES 2233
bioconversion of vanillin to vanillic acid by Streptomyces viri-dosporus. Appl. Environ. Microbiol. 45:1582-1585.
48. Postgate, J. R. 1979. The sulphate-reducing bacteria, 1st ed.Cambridge University Press, Cambridge.
49. Scheline, R. R. 1973. Metabolism of foreign compounds bygastrointestinal microorganisms. Pharmacol. Rev. 25:452-523.
50. Sleat, R., and J. P. Robinson. 1984. The bacteriology of anaer-obic degradation of aromatic compounds. J. Appl. Bacteriol.57:381-394.
51. Stanek, D. A. 1958. A study of the low-molecular weight phenolsformed upon hydrolysis of aspenwood. TAPPI 41:601-609.
52. Szewzyk, R., and N. Pfennig. 1987. Complete oxidation ofcatechol by the strictly anaerobic sulfate-reducing Desulfobac-terium catecholicum sp. nov. Arch. Microbiol. 147:163-168.
53. Taylor, B. F. 1983. Aerobic and anaerobic catabolism of vanillicacid and some other methoxy-aromatic compounds by Pseudo-monas sp. strain PN-1. Appl. Environ. Microbiol. 46:1286-1292.
54. Terho, T. T., and K. Hartiala. 1971. Method for determinationof the sulfate content of glycosaminoglycans. Anal. Biochem.41:471-476.
55. Thauer, R. K., K. Jungermann, and K. Decker. 1977. Energyconservation in chemotrophic anaerobic bacteria. Bacteriol.Rev. 41:100-180.
56. Toms, A., and J. M. Wood. 1970. The degradation of trans-
ferulic acid by Pseudomonas acidovorans. Biochemistry 9:337-343.
57. Varel, V. H., and H.-J. G. Jung. 1986. Influence of foragephenolics on ruminal fibrolytic bacteria and in vitro fiber degra-dation. Appl. Environ. Microbiol. 52:275-280.
58. Zeikus, J. G. 1981. Lignin metabolism and the carbon cycle.Adv. Microb. Ecol. 5:211-243.
59. Zeikus, J. G., A. L. Wellstein, and T. K. Kirk. 1982. Molecularbasis for the biodegradative recalcitrance of lignin in anaerobicenvironments. FEMS Microbiol. Lett. 15:193-197.
60. Zellner, G., P. Messner, H. Kneifel, and J. Winter. 1989.Desulfovibrio simplex spec. nov., a new sulfate-reducing bacte-rium from a sour whey digester. Arch. Microbiol. 152:329-334.
61. Zeliner, G., E. Stackebrandt, H. Kneifel, P. Messner, U. B.Sleytr, E. Conway de Macario, H.-P. Zabel, K. 0. Stetter, and J.Winter. 1989. Isolation and characterization of a thermophilic,sulfate reducing archaebacterium, Archaeoglobus fulgidusstrain Z. Syst. Appl. Microbiol. 11:151-160.
62. Zellner, G., P. Vogel, H. Kneifel, and J. Winter. 1987. Anaerobicdigestion of whey and whey permeate with suspended andimmobilized complex and defined consortia. Appl. Microbiol.Biotechnol. 27:306-314.
63. Zeliner, G., and J. Winter. 1987. Analysis of a highly efficientmethanogenic consortium producing biogas from whey. Syst.Appl. Microbiol. 9:284-292.
VOL. 56, 1990
















![One-step synthesis of pyridines and dihydropyridines in a … · 2013-09-30 · tion of benzyl alcohol oxidation [27], the esterification of benzoic acid in a microwave tubular flow](https://static.fdocuments.net/doc/165x107/5e904433c4617d5874147025/one-step-synthesis-of-pyridines-and-dihydropyridines-in-a-2013-09-30-tion-of-benzyl.jpg)

