
Mechanism ofAction of Benzoic Acid …BENZOIC ACID ANDZYGOSACCHAROMYCESBAILII 3411 withdrawnat...
5
Vol. 57, No. 12 Mechanism of Action of Benzoic Acid on Zygosaccharomyces bailii: Effects on Glycolytic Metabolite Levels, Energy Production, and Intracellular pH ALAN D. WARTHt Division of Food Processing, Commonwealth Scientific and Industrial Research Organization, P.O. Box 52, North Ryde, New South Wales 2113, Australia Received 7 March 1989/Accepted 8 September 1991 The effects of benzoic acid in the preservative-resistant yeast Zygosaccharomyces bailii were studied. At concentrations of benzoic acid up to 4 mM, fermentation was stimulated and only low levels of benzoate were accumulated. Near the MIC (10 mM), fermentation was inhibited, ATP levels declined, and benzoate was accumulated to relatively higher levels. Intracellular pH was reduced but not greatly. Changes in the levels of metabolites at different external benzoic acid levels showed that glycolysis was limited at pyruvate kinase and glyceraldehyde dehydrogenase-phosphoglycerate kinase steps. Inhibition of phosphofructokinase and several other glycolytic enzymes was not responsible for the inhibition of fermentation. Instead, the results suggest that the primary action of benzoic acid in Z. bailii is to cause a general energy loss, i.e., ATP depletion. Although benzoic acid and other weak-acid-type preser- vatives have been commonly used to preserve acidic foods, beverages, and pharmaceuticals for many years, there is uncertainty about which biochemical events are important in inhibiting cell growth under industrially relevant conditions. Inhibition of a variety of enzymes in vitro has been reported for sorbic acid (24), but this has not been clearly related to growth inhibition or cell death. There are few reports of significant inhibition of enzymes by benzoic acid or the aliphatic acids. In fact, glycolytic metabolism is stimulated by concentrations of preservative that inhibit cell growth (6, 26). Preservatives inhibit transport in a variety of cells (12). This may account for their effectiveness in preventing the growth of many sensitive bacteria and yeasts under some conditions, but it is not a likely cause of growth inhibition under conditions such as anaerobic fermentation of nutrient- rich foods (29). On the other hand, weak acids are well known to reduce cytoplasmic pH, particularly under the acidic conditions in which the weak-acid-type preservatives are used and are most effective. Krebs et al. (14) suggested that benzoic acid inhibited the growth of Saccharomyces cerevisiae by lowering the intracellular pH, which inhibited glycolysis, specifically by inhibition of phosphofructokinase. Zygosaccharomyces bailii is a yeast which tolerates high levels of preservative and causes spoilage of beverages and other acidic foods (25, 29). When grown in the presence of benzoic acid, Z. bailii in particular, as well as several other yeasts, tolerate higher levels of the acid. These cells accu- mulate less benzoate than expected from their intracellular pH, and benzoic acid at sublethal concentrations stimulates fermentation rates and reduces growth yields (26, 30). It was suggested that cells grown in the presence of benzoic acid became permeable to or transported benzoate anion, thus reducing the concentration of benzoate in the cell. Benzoic acid would then cycle in and out of the cell in undissociated and dissociated forms, respectively, and the intracellular pH t Deceased 21 June 1990. All revisions were completed by Dr. Kenneth W. Nickerson, University of Nebraska. Address reprint requests to Dr. Michael Eyles, Commonwealth Scientific and Indus- trial Research Organization. would tend to drop. The energy demand for export of protons and maintenance of intracellular pH may stimulate fermentation rates and lower the growth yields. In the present study, the effects of benzoic acid on fermentation rate, ATP level, intracellular pH, and benzoate accumulation level were determined over a range of concen- trations up to that causing full inhibition of growth to elucidate the events that may be critical in producing growth inhibition in Z. bailii. Metabolite levels were measured to determine which reaction steps in glycolysis were affected. MATERIALS AND METHODS Growth. Z. bailii FRR 1292 was grown in yeast extract medium (pH 3.5) (27) containing 5% glucose in place of the fructose. Benzoic acid (2 mM) was added (26). Cultures were stirred at 25°C in plugged flasks containing 30% headspace and harvested at the late exponential phase (0.3 to 1 mg [dry weight] per ml). Cells were washed twice in 0.1 M potassium citrate buffer (pH 3.5) and kept at 10°C for 1 to 3 h prior to incubation. MICs were determined by using microwell plates with inocula grown in the presence of benzoic acid (28). Growth rates were measured at 25°C by using shaken screw-cap tubes. Incubation. Cells were incubated at 25°C in screw-cap tubes fitted with a septum and flushed with N2. Glucose (20%) was added after 10 min, and 200 mM potassium benzoate and 0.8 equivalent of 1 M HCI were added 2 min later. Final concentrations were as follows: cells, 13 g (dry weight) per liter; glucose, 50 g/liter; potassium citrate (pH 3.5), 0.1 M; benzoic acid, 0 to 18 mM. Three independent experiments were done, and the same trends were found in each; results are plotted for one experiment only. Analyses. To determine the fermentation rate, samples (0.5 ml) were withdrawn 3 and 13 min after the addition of benzoate, diluted with cold 1% 2-propanol, frozen with dry ice, and stored at -180°C. For analysis, the samples were rapidly thawed and filtered. Ethanol was determined by gas chromatography (27), and glycerol was determined enzymat- ically (Boehringer test combination; Boehringer Mannheim Australia Ltd.). Samples for metabolite determinations were 3410 APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Dec. 1991, p. 3410-3414 0099-2240/91/123410-05$02.00/0 Copyright © 1991, American Society for Microbiology on April 17, 2018 by guest http://aem.asm.org/ Downloaded from
Transcript of Mechanism ofAction of Benzoic Acid …BENZOIC ACID ANDZYGOSACCHAROMYCESBAILII 3411 withdrawnat...

Vol. 57, No. 12
Mechanism of Action of Benzoic Acid on Zygosaccharomyces bailii:Effects on Glycolytic Metabolite Levels, Energy
Production, and Intracellular pHALAN D. WARTHt
Division ofFood Processing, Commonwealth Scientific and Industrial Research Organization,P.O. Box 52, North Ryde, New South Wales 2113, Australia
Received 7 March 1989/Accepted 8 September 1991
The effects of benzoic acid in the preservative-resistant yeast Zygosaccharomyces bailii were studied. Atconcentrations of benzoic acid up to 4 mM, fermentation was stimulated and only low levels of benzoate wereaccumulated. Near the MIC (10 mM), fermentation was inhibited, ATP levels declined, and benzoate wasaccumulated to relatively higher levels. Intracellular pH was reduced but not greatly. Changes in the levels ofmetabolites at different external benzoic acid levels showed that glycolysis was limited at pyruvate kinase andglyceraldehyde dehydrogenase-phosphoglycerate kinase steps. Inhibition of phosphofructokinase and severalother glycolytic enzymes was not responsible for the inhibition of fermentation. Instead, the results suggest thatthe primary action of benzoic acid in Z. bailii is to cause a general energy loss, i.e., ATP depletion.
Although benzoic acid and other weak-acid-type preser-vatives have been commonly used to preserve acidic foods,beverages, and pharmaceuticals for many years, there isuncertainty about which biochemical events are important ininhibiting cell growth under industrially relevant conditions.Inhibition of a variety of enzymes in vitro has been reportedfor sorbic acid (24), but this has not been clearly related togrowth inhibition or cell death. There are few reports ofsignificant inhibition of enzymes by benzoic acid or thealiphatic acids. In fact, glycolytic metabolism is stimulatedby concentrations of preservative that inhibit cell growth (6,26). Preservatives inhibit transport in a variety of cells (12).This may account for their effectiveness in preventing thegrowth of many sensitive bacteria and yeasts under someconditions, but it is not a likely cause of growth inhibitionunder conditions such as anaerobic fermentation of nutrient-rich foods (29). On the other hand, weak acids are wellknown to reduce cytoplasmic pH, particularly under theacidic conditions in which the weak-acid-type preservativesare used and are most effective. Krebs et al. (14) suggestedthat benzoic acid inhibited the growth of Saccharomycescerevisiae by lowering the intracellular pH, which inhibitedglycolysis, specifically by inhibition of phosphofructokinase.Zygosaccharomyces bailii is a yeast which tolerates high
levels of preservative and causes spoilage of beverages andother acidic foods (25, 29). When grown in the presence ofbenzoic acid, Z. bailii in particular, as well as several otheryeasts, tolerate higher levels of the acid. These cells accu-
mulate less benzoate than expected from their intracellularpH, and benzoic acid at sublethal concentrations stimulatesfermentation rates and reduces growth yields (26, 30). It wassuggested that cells grown in the presence of benzoic acidbecame permeable to or transported benzoate anion, thusreducing the concentration of benzoate in the cell. Benzoicacid would then cycle in and out of the cell in undissociatedand dissociated forms, respectively, and the intracellular pH
t Deceased 21 June 1990. All revisions were completed by Dr.Kenneth W. Nickerson, University of Nebraska. Address reprintrequests to Dr. Michael Eyles, Commonwealth Scientific and Indus-trial Research Organization.
would tend to drop. The energy demand for export ofprotons and maintenance of intracellular pH may stimulatefermentation rates and lower the growth yields.
In the present study, the effects of benzoic acid onfermentation rate, ATP level, intracellular pH, and benzoateaccumulation level were determined over a range of concen-trations up to that causing full inhibition of growth toelucidate the events that may be critical in producing growthinhibition in Z. bailii. Metabolite levels were measured todetermine which reaction steps in glycolysis were affected.
MATERIALS AND METHODS
Growth. Z. bailii FRR 1292 was grown in yeast extractmedium (pH 3.5) (27) containing 5% glucose in place of thefructose. Benzoic acid (2 mM) was added (26). Cultures werestirred at 25°C in plugged flasks containing 30% headspaceand harvested at the late exponential phase (0.3 to 1 mg [dryweight] per ml). Cells were washed twice in 0.1 M potassiumcitrate buffer (pH 3.5) and kept at 10°C for 1 to 3 h prior toincubation. MICs were determined by using microwell plateswith inocula grown in the presence of benzoic acid (28).Growth rates were measured at 25°C by using shakenscrew-cap tubes.
Incubation. Cells were incubated at 25°C in screw-captubes fitted with a septum and flushed with N2. Glucose(20%) was added after 10 min, and 200 mM potassiumbenzoate and 0.8 equivalent of 1 M HCI were added 2 minlater. Final concentrations were as follows: cells, 13 g (dryweight) per liter; glucose, 50 g/liter; potassium citrate (pH3.5), 0.1 M; benzoic acid, 0 to 18 mM. Three independentexperiments were done, and the same trends were found ineach; results are plotted for one experiment only.
Analyses. To determine the fermentation rate, samples (0.5ml) were withdrawn 3 and 13 min after the addition ofbenzoate, diluted with cold 1% 2-propanol, frozen with dryice, and stored at -180°C. For analysis, the samples wererapidly thawed and filtered. Ethanol was determined by gaschromatography (27), and glycerol was determined enzymat-ically (Boehringer test combination; Boehringer MannheimAustralia Ltd.). Samples for metabolite determinations were
3410
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Dec. 1991, p. 3410-34140099-2240/91/123410-05$02.00/0Copyright © 1991, American Society for Microbiology
on April 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from
BENZOIC ACID AND ZYGOSACCHAROMYCES BAILII 3411
withdrawn at 14 min. Adenine nucleotides were extracted byinjecting the sample rapidly into an equal volume of ethanolat 78°C. After 5 min, the samples were cooled, dilutedfourfold, and stored at -18°C. ATP was measured bybioluminescence by using a model M1070 luminometer andLumit reagent (Lumac B.V., The Netherlands). AMP +ADP + ATP and ADP + ATP were determined as ATP byusing pyruvate kinase and adenylate kinase (1). For othermetabolites, the sample was mixed with an equal volume of2 M HCl04 at 0°C for 30 min, pH was adjusted to 6.2 to 6.7with 2 M K2CO3, and the mixture was frozen with dry iceand thawed twice and centrifuged. Phosphoenolpyruvateand glycerate 2-phosphate were determined within 4 h, andother metabolites were determined after storage at -180°C.The following glycolytic metabolites were measured enzy-matically: glucose 6-phosphate and fructose 6-phosphate(19), fructose 1,6-bisphosphate and triosephosphates (20),glycerate 3-phosphate (11), glycerate 2-phosphate and phos-phoenolpyruvate (15), pyruvate (16), acetaldehyde (2), andNAD (13). Pi was estimated by the method of Lowry andLopez (17). Intracellular concentrations of metabolites werecalculated by using a value of 1.8 pil of cell water per mg (dryweight) (26). The amount of benzoic acid in cells and in themedium was determined by high-pressure liquid chromato-graphy (27) after centrifugation for 2 min at 20,000 x g. Acorrection was applied for interstitial fluid.
Intracellular pH determination. Since determination ofintracellular pH by distribution of weak acid requires theassumption of impermeability of the anion, intracellular pHwas estimated by the freeze-thaw method (4, 26, 30). Cells(250 mg [wet weight]) were rapidly filtered on Nucleporemembranes and washed with 0.5 ml of cold water, and thepellet was scraped off into a polythene cap (9 mm [innerdiameter]) and frozen in liquid N2. The caps were thenthawed in air or in hexane and immediately refrozen. Aftersix cycles, the temperature was quickly raised to 25°C andthe pH was determined by using a flat, combination elec-trode. No addition of fluid was necessary, as the pelletbecame a viscous fluid.
Calculation of mass action substrate/product ratios. Levelsof several metabolites were below the limit of detection bythe methods used. Glyceraldehyde 3-phosphate and glycer-ate 2,3-bisphosphate concentrations were calculated by as-suming equilibration of triosephosphate isomerase and phos-phoglycerate mutase, respectively, as these steps are notcommonly limiting (22). The lack of data for glyceric 1,3-bisphosphate means that glyceraldehyde-3-phosphate dehy-drogenase and phosphoglycerate kinase reactions cannot bedistinguished. Glyceraldehyde-3-phosphate dehydrogenasehas been taken as being in equilibrium, but the results mayapply to either step. The enolase reaction equilibrium re-quired that the phosphoenolpyruvate concentration in thesample with the lowest benzoic acid concentration be lessthan 0.039 mM, which is less than the limit of detection (0.1mM). This value was used at each benzoate concentration.Modeling of higher values allowed at other benzoic acidconcentrations did not significantly affect any conclusions.NAD+/NADH ratios were calculated from ethanol and ace-taldehyde concentrations, assuming equilibration by alcoholdehydrogenase.
RESULTS
Growth inhibition. The MIC of benzoic acid for Z. bailii,determined by using microwell plates, was 10 mM (28).Benzoic acid progressively reduced the growth rate, causing
I1 15
I
E 10E0-
0o' S
20n.
0
0 2 4 6 8 10 12
Benzoic acid in medium (mM)14
3
2 iE.-II-
1 <
0
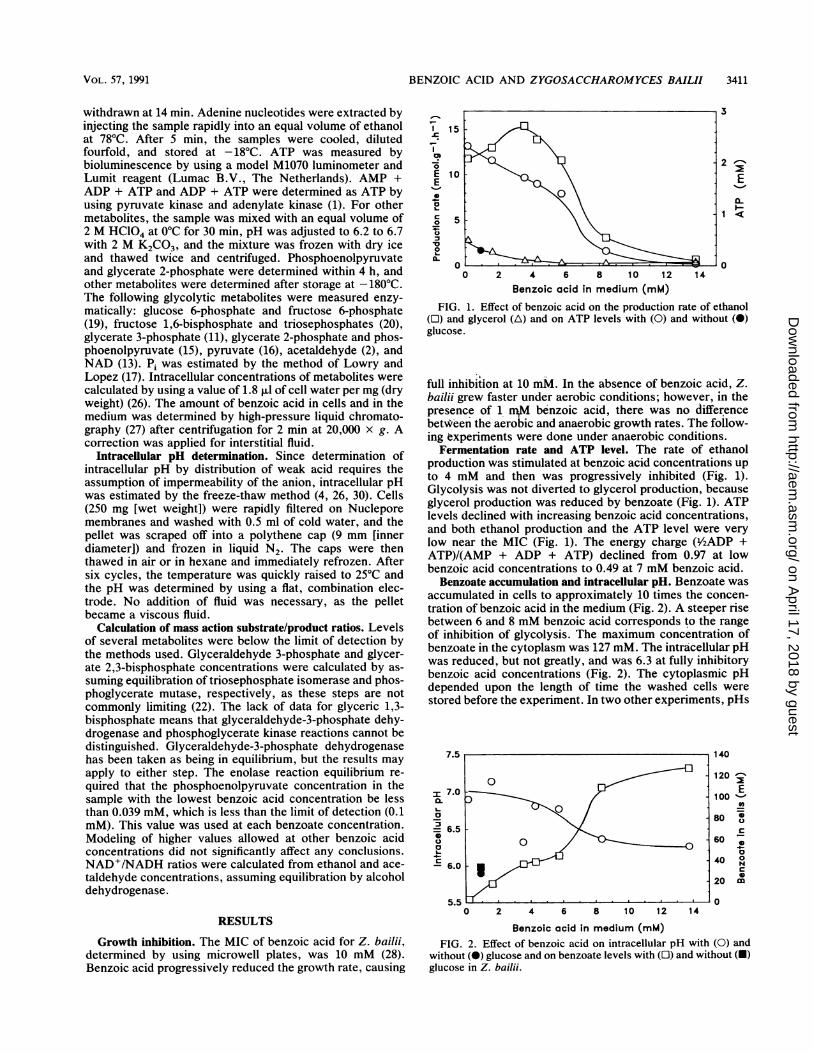
FIG. 1. Effect of benzoic acid on the production rate of ethanol(O) and glycerol (A) and on ATP levels with (0) and without (0)glucose.
full inhibition at 10 mM. In the absence of benzoic acid, Z.bailii grew faster under aerobic conditions; however, in thepresence of 1 nmM benzoic acid, there was no differencebetween the aerobic and anaerobic growth rates. The follow-ing experiments were done under anaerobic conditions.
Fermentation rate and ATP level. The rate of ethanolproduction was stimulated at benzoic acid concentrations upto 4 mM and then was progressively inhibited (Fig. 1).Glycolysis was not diverted to glycerol production, becauseglycerol production was reduced by benzoate (Fig. 1). ATPlevels declined with increasing benzoic acid concentrations,and both ethanol production and the ATP level were verylow near the MIC (Fig. 1). The energy charge (Y2ADP +ATP)/(AMP + ADP + ATP) declined from 0.97 at lowbenzoic acid concentrations to 0.49 at 7 mM benzoic acid.
Benzoate accumulation and intracellular pH. Benzoate wasaccumulated in cells to approximately 10 times the concen-tration of benzoic acid in the medium (Fig. 2). A steeper risebetween 6 and 8 mM benzoic acid corresponds to the rangeof inhibition of glycolysis. The maximum concentration ofbenzoate in the cytoplasm was 127 mM. The intracellular pHwas reduced, but not greatly, and was 6.3 at fully inhibitorybenzoic acid concentrations (Fig. 2). The cytoplasmic pHdepended upon the length of time the washed cells werestored before the experiment. In two other experiments, pHs
7.5 140
120
I7.0 100 E5.5
0 8
6.5 840 60,0~~~
40 0
6.0~~~~~~~~~~~~20 m
5.5 j00 2 4 6 8 10 12 14
Benzoic acid in medium (mM)FIG. 2. Effect of benzoic acid on intracellular pH with (0) and
without (0) glucose and on benzoate levels with (O) and without (-)glucose in Z. bailii.
VOL. 57, 1991
on April 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from
APPL. ENVIRON. MICROBIOL.
12
-z 10 LE
8
00
4.0C- 2
0
0 2 4 6 8 10 12 14
Benzoic acid in medium (mM)
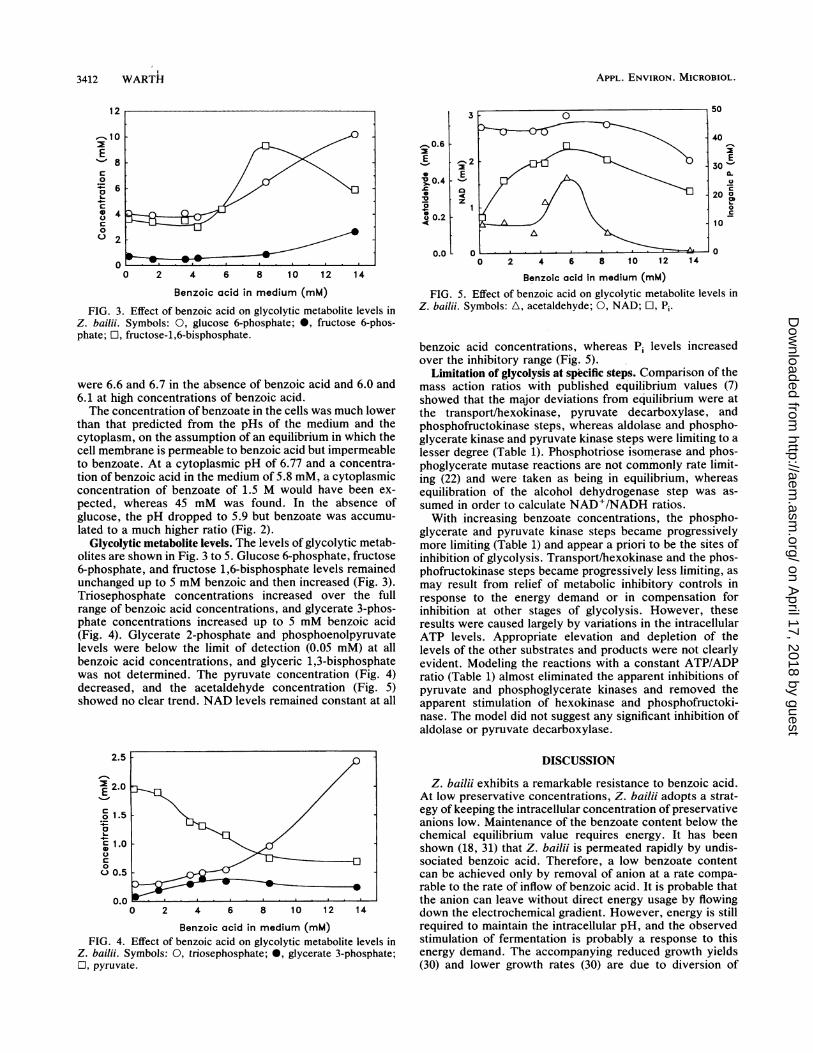
FIG. 3. Effect of benzoic acid on glycolytic metabolite levels inZ. bailii. Symbols: 0, glucose 6-phosphate; 0, fructose 6-phos-phate; 0, fructose-1,6-bisphosphate.
were 6.6 and 6.7 in the absence of benzoic acid and 6.0 and6.1 at high concentrations of benzoic acid.The concentration ofbenzoate in the cells was much lower
than that predicted from the pHs of the medium and thecytoplasm, on the assumption of an equilibrium in which thecell membrane is permeable to benzoic acid but impermeableto benzoate. At a cytoplasmic pH of 6.77 and a concentra-tion of benzoic acid in the medium of 5.8 mM, a cytoplasmicconcentration of benzoate of 1.5 M would have been ex-pected, whereas 45 mM was found. In the absence ofglucose, the pH dropped to 5.9 but benzoate was accumu-lated to a much higher ratio (Fig. 2).
Glycolytic metabolite levels. The levels of glycolytic metab-olites are shown in Fig. 3 to 5. Glucose 6-phosphate, fructose6-phosphate, and fructose 1,6-bisphosphate levels remainedunchanged up to 5 mM benzoic and then increased (Fig. 3).Triosephosphate concentrations increased over the fullrange of benzoic acid concentrations, and glycerate 3-phos-phate concentrations increased up to 5 mM benzoic acid(Fig. 4). Glycerate 2-phosphate and phosphoenolpyruvatelevels were below the limit of detection (0.05 mM) at allbenzoic acid concentrations, and glyceric 1,3-bisphosphatewas not determined. The pyruvate concentration (Fig. 4)decreased, and the acetaldehyde concentration (Fig. 5)showed no clear trend. NAD levels remained constant at all
- 0.62E
n 0.4
-0
o 0.2
0.0
E30
c.20 a
0C-
0 2 4 6 8 10 12 14
Benzoic acid in medium (mM)FIG. 5. Effect of benzoic acid on glycolytic metabolite levels in
Z. bailii. Symbols: A, acetaldehyde; 0, NAD; 0, Pi.
benzoic acid concentrations, whereas P1 levels increasedover the inhibitory range (Fig. 5).
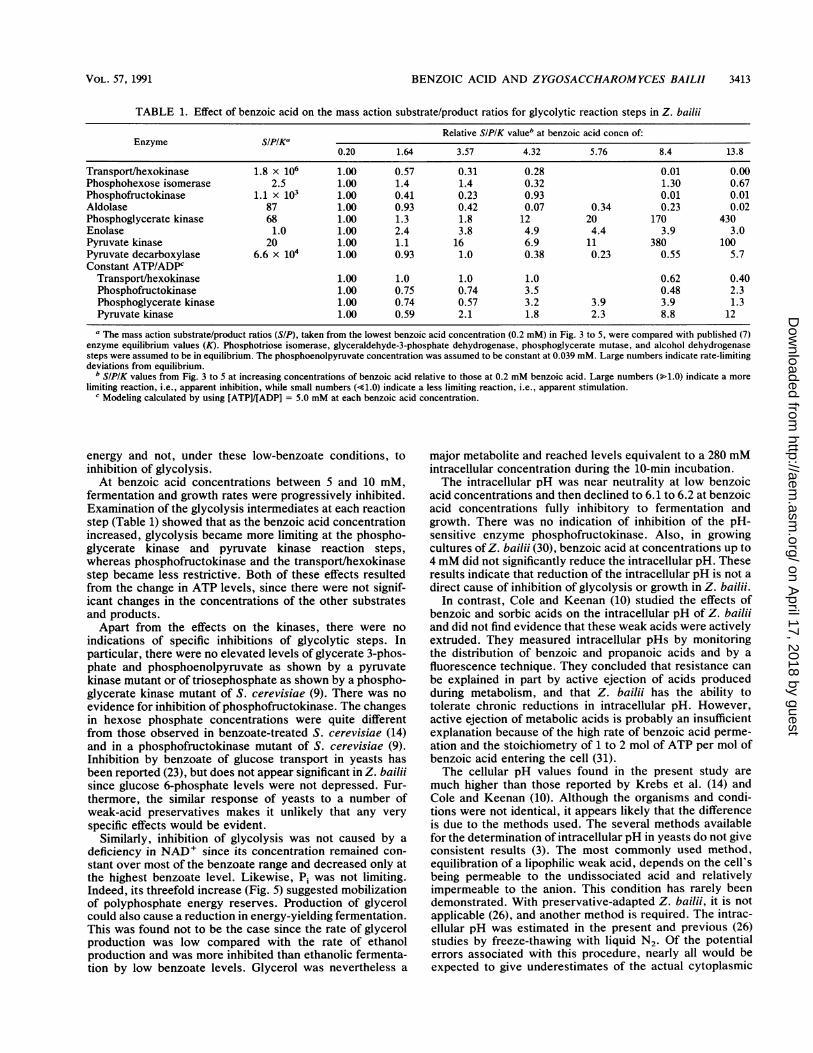
Limitation of glycolysis at specific steps. Comparison of themass action ratios with published equilibrium values (7)showed that the major deviations from equilibrium were atthe transport/hexokinase, pyruvate decarboxylase, andphosphofructokinase steps, whereas aldolase and phospho-glycerate kinase and pyruvate kinase steps were limiting to alesser degree (Table 1). Phosphotriose isomerase and phos-phoglycerate mutase reactions are not commonly rate limit-ing (22) and were taken as being in equilibrium, whereasequilibration of the alcohol dehydrogenase step was as-sumed in order to calculate NAD+/NADH ratios.With increasing benzoate concentrations, the phospho-
glycerate and pyruvate kinase steps became progressivelymore limiting (Table 1) and appear a priori to be the sites ofinhibition of glycolysis. Transport/hexokinase and the phos-phofructokinase steps became progressively less limiting, asmay result from relief of metabolic inhibitory controls inresponse to the energy demand or in compensation forinhibition at other stages of glycolysis. However, theseresults were caused largely by variations in the intracellularATP levels. Appropriate elevation and depletion of thelevels of the other substrates and products were not clearlyevident. Modeling the reactions with a constant ATP/ADPratio (Table 1) almost eliminated the apparent inhibitions ofpyruvate and phosphoglycerate kinases and removed theapparent stimulation of hexokinase and phosphofructoki-nase. The model did not suggest any significant inhibition ofaldolase or pyruvate decarboxylase.
2.5 ,> DISCUSSION
E2.0 Z. bailii exhibits a remarkable resistance to benzoic acid.At low preservative concentrations, Z. bailii adopts a strat-
o 1.5 egy of keeping the intracellular concentration of preservativeanions low. Maintenance of the benzoate content below thechemical equilibrium value requires energy. It has beenshown (18, 31) that Z. bailii is permeated rapidly by undis-sociated benzoic acid. Therefore, a low benzoate content
00 0.5 can be achieved only by removal of anion at a rate compa-
rable to the rate of inflow of benzoic acid. It is probable that0.0 the anion can leave without direct energy usage by flowing
0 2 4 6 8 10 12 14 down the electrochemical gradient. However, energy is stillBenzoic acid in medium (mM) required to maintain the intracellular pH, and the observed
FIG. 4. Effect of benzoic acid on glycolytic metabolite levels in stimulation of fermentation is probably a response to thisZ. bailii. Symbols: 0, triosephosphate; 0, glycerate 3-phosphate; energy demand. The accompanying reduced growth yields0, pyruvate. (30) and lower growth rates (30) are due to diversion of
3412 WARTii
on April 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from
BENZOIC ACID AND ZYGOSACCHAROMYCES BAILII 3413
TABLE 1. Effect of benzoic acid on the mass action substrate/product ratios for glycolytic reaction steps in Z. bailii
Relative SIPIK valueb at benzoic acid concn of:Enzyme SIPIKa
0.20 1.64 3.57 4.32 5.76 8.4 13.8
Transport/hexokinase 1.8 x 106 1.00 0.57 0.31 0.28 0.01 0.00Phosphohexose isomerase 2.5 1.00 1.4 1.4 0.32 1.30 0.67Phosphofructokinase 1.1 x 103 1.00 0.41 0.23 0.93 0.01 0.01Aldolase 87 1.00 0.93 0.42 0.07 0.34 0.23 0.02Phosphoglycerate kinase 68 1.00 1.3 1.8 12 20 170 430Enolase 1.0 1.00 2.4 3.8 4.9 4.4 3.9 3.0Pyruvate kinase 20 1.00 1.1 16 6.9 11 380 100Pyruvate decarboxylase 6.6 x 104 1.00 0.93 1.0 0.38 0.23 0.55 5.7Constant ATP/ADPC
Transport/hexokinase 1.00 1.0 1.0 1.0 0.62 0.40Phosphofructokinase 1.00 0.75 0.74 3.5 0.48 2.3Phosphoglycerate kinase 1.00 0.74 0.57 3.2 3.9 3.9 1.3Pyruvate kinase 1.00 0.59 2.1 1.8 2.3 8.8 12
a The mass action substrate/product ratios (SIP), taken from the lowest benzoic acid concentration (0.2 mM) in Fig. 3 to 5, were compared with published (7)enzyme equilibrium values (K). Phosphotriose isomerase, glyceraldehyde-3-phosphate dehydrogenase, phosphoglycerate mutase, and alcohol dehydrogenasesteps were assumed to be in equilibrium. The phosphoenolpyruvate concentration was assumed to be constant at 0.039 mM. Large numbers indicate rate-limitingdeviations from equilibrium.
b SIPIK values from Fig. 3 to 5 at increasing concentrations of benzoic acid relative to those at 0.2 mM benzoic acid. Large numbers (>1.0) indicate a morelimiting reaction, i.e., apparent inhibition, while small numbers (s1.0) indicate a less limiting reaction, i.e., apparent stimulation.
c Modeling calculated by using [ATP]/[ADP] = 5.0 mM at each benzoic acid concentration.
energy and not, under these low-benzoate conditions, toinhibition of glycolysis.At benzoic acid concentrations between 5 and 10 mM,
fermentation and growth rates were progressively inhibited.Examination of the glycolysis intermediates at each reactionstep (Table 1) showed that as the benzoic acid concentrationincreased, glycolysis became more limiting at the phospho-glycerate kinase and pyruvate kinase reaction steps,whereas phosphofructokinase and the transport/hexokinasestep became less restrictive. Both of these effects resultedfrom the change in ATP levels, since there were not signif-icant changes in the concentrations of the other substratesand products.Apart from the effects on the kinases, there were no
indications of specific inhibitions of glycolytic steps. Inparticular, there were no elevated levels of glycerate 3-phos-phate and phosphoenolpyruvate as shown by a pyruvatekinase mutant or of triosephosphate as shown by a phospho-glycerate kinase mutant of S. cerevisiae (9). There was noevidence for inhibition of phosphofructokinase. The changesin hexose phosphate concentrations were quite differentfrom those observed in benzoate-treated S. cerevisiae (14)and in a phosphofructokinase mutant of S. cerevisiae (9).Inhibition by benzoate of glucose transport in yeasts hasbeen reported (23), but does not appear significant in Z. bailiisince glucose 6-phosphate levels were not depressed. Fur-thermore, the similar response of yeasts to a number ofweak-acid preservatives makes it unlikely that any veryspecific effects would be evident.
Similarly, inhibition of glycolysis was not caused by adeficiency in NAD+ since its concentration remained con-stant over most of the benzoate range and decreased only atthe highest benzoate level. Likewise, Pi was not limiting.Indeed, its threefold increase (Fig. 5) suggested mobilizationof polyphosphate energy reserves. Production of glycerolcould also cause a reduction in energy-yielding fermentation.This was found not to be the case since the rate of glycerolproduction was low compared with the rate of ethanolproduction and was more inhibited than ethanolic fermenta-tion by low benzoate levels. Glycerol was nevertheless a
major metabolite and reached levels equivalent to a 280 mMintracellular concentration during the 10-min incubation.The intracellular pH was near neutrality at low benzoic
acid concentrations and then declined to 6.1 to 6.2 at benzoicacid concentrations fully inhibitory to fermentation andgrowth. There was no indication of inhibition of the pH-sensitive enzyme phosphofructokinase. Also, in growingcultures of Z. bailii (30), benzoic acid at concentrations up to4 mM did not significantly reduce the intracellular pH. Theseresults indicate that reduction of the intracellular pH is not adirect cause of inhibition of glycolysis or growth in Z. bailii.
In contrast, Cole and Keenan (10) studied the effects ofbenzoic and sorbic acids on the intracellular pH of Z. bailiiand did not find evidence that these weak acids were activelyextruded. They measured intracellular pHs by monitoringthe distribution of benzoic and propanoic acids and by afluorescence technique. They concluded that resistance canbe explained in part by active ejection of acids producedduring metabolism, and that Z. bailii has the ability totolerate chronic reductions in intracellular pH. However,active ejection of metabolic acids is probably an insufficientexplanation because of the high rate of benzoic acid perme-ation and the stoichiometry of 1 to 2 mol of ATP per mol ofbenzoic acid entering the cell (31).The cellular pH values found in the present study are
much higher than those reported by Krebs et al. (14) andCole and Keenan (10). Although the organisms and condi-tions were not identical, it appears likely that the differenceis due to the methods used. The several methods availablefor the determination of intracellular pH in yeasts do not giveconsistent results (3). The most commonly used method,equilibration of a lipophilic weak acid, depends on the cell'sbeing permeable to the undissociated acid and relativelyimpermeable to the anion. This condition has rarely beendemonstrated. With preservative-adapted Z. bailii, it is notapplicable (26), and another method is required. The intrac-ellular pH was estimated in the present and previous (26)studies by freeze-thawing with liquid N2. Of the potentialerrors associated with this procedure, nearly all would beexpected to give underestimates of the actual cytoplasmic
VOL. 57, 1991
on April 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
pH. Also, for yeast cells at external pHs near 3.5 with anadequate energy supply, several studies have found near-neutral internal pH values. For S. cerevisiae, nuclear mag-netic resonance studies gave a value of 7.1 (21), comparedwith 6.7 from freeze-thawing (5), and 6.4 from the weak-acidmethod (8). Lower values have frequently been obtained,but these can be attributed to an inadequate energy supply.Above the MIC for growth, cells contained high concen-
trations of benzoate. This is an appropriate response toconserve energy for long-term survival. Cells can retainviability under these conditions for many days while the highbenzoate anion concentration could well cause direct inhibi-tion of glycolysis and other cell functions. However, theintracellular pH values did not drop to levels which wouldrequire no energy to maintain, at least not within the timescale of this experiment. The residual fermentation rates,although low, may be necessary for survival.
In summary, the action of benzoic acid on Z. bailiidepends upon its concentration. At low concentrations,growth rates are reduced and fermentation rates are stimu-lated because energy is used to lower the internal concen-tration of benzoate. At higher concentrations, fermentationitself becomes inhibited, leading to a collapse of the system.The reasons for this inhibition of fermentation are not fullyclear. Internal benzoate concentrations increased in parallelwith the inhibition of fermentation, but they may not be themajor direct cause of inhibition. pH reduction also does notappear to be a prime cause of inhibition in Z. bailii, since theinternal pH was not severely depressed. Instead, the patternof glycolytic intermediate changes suggests that ATP limita-tion was important. This ATP limitation may be direct ormay result from regulatory mechanisms designed to con-serve the remaining energy.
REFERENCES1. Bali, W. J., and D. E. Atkinson. 1975. Adenylate energy charge
in Saccharomyces cerevisiae during starvation. J. Bacteriol.121:975-982.
2. Beutler, H.-O. 1984. Acetaldehyde, p. 606-613. In H. U. Berg-meyer (ed.), Methods of enzymatic analysis, vol. VI. AcademicPress, Inc., New York.
3. Borst-Pauwels, G. W. F. H. 1981. Ion transport in yeast.Biochim. Biophys. Acta 650:88-127.
4. Borst-Pauwels, G. W. F. H., and J. Dobbelmann. 1972. Deter-mination of the yeast cell pH. Acta Bot. Neerl. 21:149-154.
5. Borst-Pauwels, G. W. F. H., and P. H. J. Peters. 1977. Effect ofthe medium pH and the cell pH upon the kinetical parameters ofphosphate uptake by yeast. Biochim. Biophys. Acta 466:488-495.
6. Bosund, I. 1962. The action of benzoic and salicylic acids on themetabolism of microorganisms. Adv. Food Res. 11:331-353.
7. Burton, K. 1957. Free energy data of biological interest, p.275-285. In H. A. Krebs and H. L. Kornberg (ed.), Energytransformations in living matter: a survey. Springer Verlag KG,Berlin.
8. Cartwright, C. P., J.-G. Juroszek, M. J. Beavan, F. M. S. Ruby,S. M. F. De Morais, and A. H. Rose. 1986. Ethanol dissipates theproton-motive force across the plasma membrane of Saccharo-myces cerevisiae. J. Gen. Microbiol. 132:369-377.
9. Ceriacy, M., and I. Breitenbach. 1979. Physiological effects ofseven different blocks in glycolysis in Saccharomyces cerevi-siae. J. Bacteriol. 139:152-160.
10. Cole, M. B., and M. H. J. Keenan. 1987. Effects of weak acids
and external pH on the intracellular pH of Zygosaccharomycesbailii, and its implication in weak-acid resistance. Yeast 3:23-32.
11. Czok, G. 1984. D-Glycerate 3-phosphate, p. 537-541. In H. U.Bergmeyer (ed.), Methods of enzymatic analysis, vol. VI.Academic Press, Inc., New York.
12. Freese, E., and C. Levin. 1978. Action mechanisms of preser-vatives and antiseptics. Dev. Ind. Microbiol. 19:207-227.
13. Klingenberg, M. 1984. Nicotinamide-adenine dinucleotides anddinucleotide phosphates, p. 251-256. In H. U. Bergmeyer (ed.),Methods of enzymatic analysis, vol. VI. Academic Press, Inc.,New York.
14. Krebs, H. A., D. Wiggins, M. Stubbs, A. Sols, and F. Bedoya.1983. Studies on the antifungal action of benzoate. Biochem. J.214:657-663.
15. Lamprecht, W., and F. Heinz. 1984. Pyruvate, p. 570-577. InH. U. Bergmeyer (ed.), Methods of enzymatic analysis, vol. VI.Academic Press, Inc., New York.
16. Lamprecht, W., and F. Heinz. 1984. D-Glycerate 2-phosphateand phosphoenolpyruvate, p. 555-561. In H. U. Bergmeyer(ed.), Methods of enzymatic analysis, vol. VI. Academic Press,Inc., New York.
17. Lowry, 0. H., and J. A. Lopez. 1946. The determination ofinorganic phosphorous in the presence of labile phosphateesters. J. Biol. Chem. 162:421-428.
18. Macris, B. J. 1975. Mechanism of benzoic acid uptake bySaccharomyces cerevisiae. Appl. Microbiol. 30:503-506.
19. Michal, G. 1984. D-Glucose 6-phosphate and D-fructose 6-phos-phate, p. 191-198. In H. U. Bergmeyer (ed.), Methods ofenzymatic analysis, vol. VI. Academic Press, Inc., New York.
20. Michal, G. 1984. D-Fructose 1,6-bisphosphate, dihydroxyace-tone phosphate and D-glyceraldehyde 3-phosphate, p. 342-350.In H. U. Bergmeyer (ed.), Methods of enzymatic analysis, vol.VI. Academic Press, Inc., New York.
21. Navon, G., R. G. Shulman, Y. Yamane, T. R. Eccleshall, K. B.Lam, J. J. Baronofsky, and J. Marmur. 1979. Phosphorus-31nuclear magnetic resonance studies of wild-type and glycolyticpathway mutants of Saccharomyces cerevisiae. Biochemistry18:4487-4499.
22. Rose, I. A., and Z. B. Rose. 1969. Glycolysis: regulation andmechanisms of the enzymes, p. 93-161. In M. Florkin and E. H.Stotz (ed.), Comprehensive biochemistry, vol. 17. ElsevierBiomedical Press, Amsterdam.
23. Scharff, T. G., M. Z. Badr, and R. J. Doyle. 1982. The nature ofsalicylate inhibition of sugar transport in yeast. Fundam. Appl.Toxicol. 2:168-172.
24. Sofos, J. N., and F. F. Busta. 1981. Antimicrobial activity ofsorbate. J. Food Prot. 44:614-622.
25. Thomas, D. S., and R. R. Davenport. 1985. Zygosaccharomycesbailii-a profile of characteristics and spoilage activities. FoodMicrobiol. 2:157-169.
26. Warth, A. D. 1977. Mechanism of resistance of Saccharomycesbailii to benzoic, sorbic and other weak acids used as foodpreservatives. J. Appl. Bacteriol. 43:215-230.
27. Warth, A. D. 1985. Resistance of yeast species to benzoic andsorbic acids and to sulfur dioxide. J. Food Prot. 48:564-569.
28. Warth, A. D. 1986. Effect of nutrients and pH on the resistanceof Zygosaccharomyces baiii to benzoic acid. Int. J. FoodMicrobiol. 3:263-271.
29. Warth, A. D. 1986. Preservative resistance of Zygosaccharo-myces bailii and other yeasts. CSIRO Food Res. Q. 46:1-8.
30. Warth, A. D. 1988. Effect of benzoic acid on growth yield ofyeasts differing in their resistance to preservatives. Appl. Envi-ron. Microbiol. 54:2091-2095.
31. Warth, A. D. 1989. Transport of benzoic and propanoic acids byZygosaccharomyces bailii. J. Gen. Microbiol. 135:1383-1390.
3414 WARTH
on April 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from


















