
Omara-Opyene, A.L. and Gedamu, L. 1997
21
Molecular and Biochemical Parasitology 90 (1997) 247 – 267 Molecular cloning, characterization and overexpression of two distinct cysteine protease cDNAs from Leishmania dono6ani chagasi 1 Archangel Levi Omara-Opyene, Lashitew Gedamu * Department of Biological Sciences, Uni6ersity of Calgary, Calgary, Alta. T2N 1N4, Canada Received 20 April 1997; accepted 5 September 1997 Abstract We have cloned and characterized two distinct cysteine protease cDNAs from Leishmania dono6ani chagasi. One of the cDNAs, Ldccy2, was isolated from a cDNA library prepared from total promastigote RNA while the other cDNA, Ldccys1, was isolated from a cDNA library prepared from total amastigote RNA. Ldccys2 has an open reading frame of 471 amino acids and Ldccys1 has an open reading frame of 447 amino acids. Comparison of the predicted protein sequences of the two distinct cysteine proteases with those of cysteine proteases from Leishmania pifanoi, a member of the L. mexicana complex, showed that the cysteine proteases from the two species of Leishmania are similar in their protein sequences. Each of the two cDNAs is distinct in genomic arrangement and chromosome location. Ldccys1 belongs to a family of cysteine proteases encoded by tandemly organized genes located on chromosome 7 while Ldccys2 appears to be a single cysteine protease gene located on chromosome 10. The organization of the two families of cysteine protease genes in L. dono6ani dono6ani was also found to be similar. In this species, the Lddcys1 genes are located on chromosome 5 while the Lddcys2 gene is located on chromosome 8. The Ldccys1 genes are expressed abundantly in the amastigotes recovered from infected hamsters, but at a very low level in the promastigote stage of development. On the other hand, the Ldccys2 gene is expressed both in the promastigote and amastigote stages. We have overexpressed the two cDNAs of cysteine proteases in Leishmania cells and the over-produced cysteine proteases are biologically active and are inhibited by cysteine protease inhibitors. Furthermore, the over-produced and indigenous amastigote specific cysteine protease, Ldccys1, reacted with poly- clonal antibodies raised against this protein. © 1997 Elsevier Science B.V. Keywords: Leishmania dono6ani chagasi ; Cysteine proteases; Amastigote specific; Promastigote specific; cDNA; Overexpression Abbre6iations: PBS, phosphate buffered saline; PCR, polymerase chain reaction; SDS-PAGE, sodium dodecyl sulfate-polyacry- lamide gel electrophoresis. * Corresponding author. Tel.: +1 403 2205556; fax: +1 403 2899311; e-mail: [email protected] 1 Note: Leishmania dono6ani chagasi Ldccys1 and Ldccys2 cDNA sequences have been assigned EMBL and GenBank™ nucleotide accession numbers AF004592 and AF004593, respectively. 0166-6851/97/$17.00 © 1997 Elsevier Science B.V. All rights reserved. PII S0166-6851(97)00158-8
-
Upload
archangel-levi-omara-opyene-phd -
Category
Documents
-
view
92 -
download
0
Transcript of Omara-Opyene, A.L. and Gedamu, L. 1997

Molecular and Biochemical Parasitology 90 (1997) 247–267
Molecular cloning, characterization and overexpressionof two distinct cysteine protease cDNAs from
Leishmania dono6ani chagasi1
Archangel Levi Omara-Opyene, Lashitew Gedamu *
Department of Biological Sciences, Uni6ersity of Calgary, Calgary, Alta. T2N 1N4, Canada
Received 20 April 1997; accepted 5 September 1997
Abstract
We have cloned and characterized two distinct cysteine protease cDNAs from Leishmania dono6ani chagasi. One ofthe cDNAs, Ldccy2, was isolated from a cDNA library prepared from total promastigote RNA while the othercDNA, Ldccys1, was isolated from a cDNA library prepared from total amastigote RNA. Ldccys2 has an openreading frame of 471 amino acids and Ldccys1 has an open reading frame of 447 amino acids. Comparison of thepredicted protein sequences of the two distinct cysteine proteases with those of cysteine proteases from Leishmaniapifanoi, a member of the L. mexicana complex, showed that the cysteine proteases from the two species of Leishmaniaare similar in their protein sequences. Each of the two cDNAs is distinct in genomic arrangement and chromosomelocation. Ldccys1 belongs to a family of cysteine proteases encoded by tandemly organized genes located onchromosome 7 while Ldccys2 appears to be a single cysteine protease gene located on chromosome 10. Theorganization of the two families of cysteine protease genes in L. dono6ani dono6ani was also found to be similar. Inthis species, the Lddcys1 genes are located on chromosome 5 while the Lddcys2 gene is located on chromosome 8.The Ldccys1 genes are expressed abundantly in the amastigotes recovered from infected hamsters, but at a very lowlevel in the promastigote stage of development. On the other hand, the Ldccys2 gene is expressed both in thepromastigote and amastigote stages. We have overexpressed the two cDNAs of cysteine proteases in Leishmania cellsand the over-produced cysteine proteases are biologically active and are inhibited by cysteine protease inhibitors.Furthermore, the over-produced and indigenous amastigote specific cysteine protease, Ldccys1, reacted with poly-clonal antibodies raised against this protein. © 1997 Elsevier Science B.V.
Keywords: Leishmania dono6ani chagasi ; Cysteine proteases; Amastigote specific; Promastigote specific; cDNA;Overexpression
Abbre6iations: PBS, phosphate buffered saline; PCR, polymerase chain reaction; SDS-PAGE, sodium dodecyl sulfate-polyacry-lamide gel electrophoresis.
* Corresponding author. Tel.: +1 403 2205556; fax: +1 403 2899311; e-mail: [email protected] Note: Leishmania dono6ani chagasi Ldccys1 and Ldccys2 cDNA sequences have been assigned EMBL and GenBank™ nucleotide
accession numbers AF004592 and AF004593, respectively.
0166-6851/97/$17.00 © 1997 Elsevier Science B.V. All rights reserved.
PII S 0166 -6851 (97 )00158 -8

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267248
1. Introduction
Parasitic protozoa of the genus Leishmaniacause a wide spectrum of human cutaneous, mu-cocutaneous, and visceral leishmaniasis that afflictan estimated 12 million people and is endemic in86 countries around the world [1]. Human infec-tion begins with a bite of an infected sandflywhich inoculates promastigotes into subcutaneoustissue where they are taken up by tissuemacrophages. In the macrophages, promastigotesestablish themselves within the phagolysosomeswhere they transform into a non-flagellated formcalled amastigotes. The lysosomes fuse with thevacuole containing the parasite and release acidhydrolases but Leishmania resist destruction bythese enzymes. How the Leishmania amastigotesescape hydrolysis by lysosomal enzymes is un-known. The parasite’s success in establishing aninfectious process depends on how much it canmultiply within this hostile environment. Al-though the exact roles of cysteine proteases inLeishmania pathogenesis have yet to be defined[2], these enzymes appear to be crucially impor-tant for the survival of both the promastigote andamastigote forms of these parasites. Evidence insupport of this view first came from studies whichshow that inhibitors of cysteine proteases arrestthe growth of the amastigote within macrophages[3,4] but these reports were controversial becausethese inhibitors are also toxic to macrophages.The controversies have been resolved by evidencewhich show that two fluoromethyl ketone-deriva-tized peptide inhibitors of cysteine proteases areeffective in killing both the promastigote andamastigote forms of the parasites but not the hostcells [2]. The effectiveness of these inhibitors inkilling the parasites have led some investigators topropose that cysteine proteases are potentialchemotherapeutic targets [2,5,6]. Recently, evi-dence from the disruption of Leishmania mexicanacysteine protease gene array (lmcpb) suggest theseenzymes are virulence factors [7].
Cysteine proteases have been described frommany pathogenic organisms such as bacteria [8],viruses [9], fungi [10], and parasites [5,6,11–21]. InLeishmania, cysteine proteases have been de-scribed mainly from members of the L. mexicana
complex [12–18]. All the cysteine protease geneswhich have been cloned from both Leishmaniaand T. cruzi appear to be developmentally regu-lated [16–20]. However, prior to our work, effortsto detect cysteine proteases from the L. dono6aniand L. major using gelatin SDS-PAGE have beenunsuccessful [13,15]. As an initial step in identify-ing the possible functions of cysteine proteases invisceral leishmaniasis, we describe in this paperthe molecular cloning of two distinct cysteineprotease cDNAs from Leishmania dono6anichagasi, a member of the L. dono6ani complex.One of the genes, Ldccys2, is expressed in boththe promastigote and amastigote stages of devel-opment and the other gene, Ldccys1, is expressedpredominantly during the amastigote stage of de-velopment. While Ldccys2 appears to be repre-sented by a single gene, Ldccys1 belongs to amajor family of cysteine proteases encoded bytandemly organized genes. The two cysteineprotease cDNAs were overexpressed in Leishma-nia chagasi cells and the over-produced cysteineproteases were shown to be biologically active andinhibited by cysteine protease inhibitors.
2. Materials and methods
2.1. Growth and isolation of parasites
A Brazilian isolate of Leishmania dono6anichagasi (L. d. chagasi ) was kindly provided by R.Pearson, University of Virginia, Charlottesville,VA. Promastigotes from this isolate were culturedat 26°C in a modified minimal essential medium(HOMEM) (GIBCO) supplemented with 10%heat-inactivated fetal calf serum and hemin(GIBCO). The parasites were harvested in thelogarithmic (log) or stationary phase of growth. ASudanese strain of L. dono6ani dono6ani (L. d.dono6ani ) (LD3) was kindly provided by T. Pear-son, University of Victoria, Victoria, B.C.,Canada. Promastigotes from this species of Leish-mania were cultured at 26°C in Schneider’sDrosophila medium (GIBCO) supplemented with20% heat-inactivated fetal calf serum (GIBCO).To maintain infectivity, hamsters were infectedwith the parasites by intracardiac inoculation at a

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267 249
rate of 1×107 promastigotes per animal. Theanimals were sacrificed 4 weeks post infection andthe amastigotes recovered from the spleens asdescribed by others previously [22]. Amastigotesconvert to promastigotes when cultured at 26°Cin HOMEM medium with hemin and heat-inacti-vated foetal calf serum [22].
2.2. Southern and Northern (RNA) analysis
Genomic DNA was isolated from L. d. chagasiand L. d. dono6ani by lysing about 1010 pro-mastigotes in 10 ml of lysis buffer (10 mM Tris–HCl (pH 8.3), 50 mM EDTA (pH 8.0), and 1%SDS) and incubating with 1 mg of RNase A(Boehringer Mannheim) at 37°C for 1 h.Proteinase K (Boehringer Mannheim) was thenadded to a final concentration of 100 mg ml−1
and incubated overnight at 42°C. The DNA wasfurther purified by phenol-chloroform extractionand ethanol precipitation. The isolated DNA wasdigested with various restriction enzymes, sepa-rated on a 1% agarose gel and transferred ontonylon membrane (Hybond N+; Amersham) bycapillary blotting [23]. Total RNA was isolatedfrom the promastigote of L. d. chagasi and fromthe spleen of uninfected and infected hamsters bythe guanidinium isothiocyanate method [27]. To-tal RNA from uninfected and infected hamsterswas a gift from Dr Mary E. Wilson, University ofIowa College of Medicine, Iowa City, IA. Fifteenmicrograms of total RNA was loaded on separatewells and resolved on a 1.2% formaldehyde-agarose gel and transmitted to Hybond N+ nylonmembrane. Filters were hybridized at 60°C with32P-labelled DNA probes indicated in each figurelegend. All RNA blots were probed with 32P-la-belled constitutively expressed Leishmania chagasiribosomal protein (LcPO) probe (kindly providedby Drs S.G. Reed and Y. Slieky, Infectious Dis-ease Research Institute, Seattle, WA) to act as aloading control. Filters were autoradiographed onKodak X-Omat films with an intensifying screen.
2.3. PCR amplification of cysteine protease genes
Two oligonucleotide primers used to amplifythe cysteine protease gene fragments were based
on conserved regions from closely related organ-isms as reported by Eakin et al. [11]. The se-quences of the primers are: Sense primer (LCP-1):5% - ACAGAATTCCAGGGCCAGTGCGGCTC -GTGCTGG-3%, underline indicates EcoRI restric-tion site; antisense primer (LCP-2): 5%-CG-CAAGCTTCCACGAGTTCTTCACGATCCAGTA-3%, underline indicates HindIII restriction site.Conditions for PCR were as follows: A 100 m lreaction mixture containing 10 mM Tris–HCl(pH 8.2), 50 mM MgCl2, 200 mM each deoxynu-cleoside triphosphate, 2.5 U of Taq DNA poly-merase (Pharmacia), 100 pmol of each primer,and 100 ng of genomic DNA were subjected to 30cycles of amplification (94°C for 1 min, 42°C for2 min, 72°C for 2 min followed by an extension at72°C for 10 min) with a PHC-2 Dri-Block Cyler(Techne, Cambridge, UK). PCR products of 500bp length were digested sequentially with HindIIIand EcoRI, gel purified and subcloned inpGEMR-2 (Promega). Plasmid DNA was isolatedfrom recombinant colonies using the alkaline lysismethod [23], and sequenced using universal andSP6 primers as described below.
2.4. Screening, isolation and sequencedetermination of cysteine protease cDNAs
Two L. dono6ani chagasi cDNA libraries inl-ZAP II were kindly provided by Dr Mary E.Wilson, University of Iowa College of Medicine,Iowa City, IA. The first cDNA library screenedwas prepared from total promastigote RNA andthe second cDNA library screened was preparedfrom total amastigote RNA as described [24]. Theamastigotes used were purified from the spleens ofhamsters infected with L. d. chagasi as described[21] and are believed to represent the trueamastigotes found in clinical cases in terms ofgene expression. A total of 80 000 PFU was platedand duplicated filters were lifted. Screening of thepromastigote cDNA library was performed with a500 bp PCR product from L. dono6ani chagasigenomic DNA which was labelled with [a-32P]dCTP using PCR (the 500 bp PCR fragmentcontains Ldccys1 and Ldccys2 sequences) whilethe amastigote cDNA library was screened with a500 bp fragment containing Ldccys1 sequence.

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267250
The blots from the amastigote cDNA library werestripped and rehybridized with a 500 bp whichcontains the Ldccys2 sequence. Only plaques withstrong and duplicated signals were picked and aBluescript plasmid derivative was excised from thel-ZAP II recombinant phages in vivo with thehelper phage R-408 (Strategene protocol). Se-quence analyses were performed on several clonesusing the method of Sanger et al. [25]. Briefly,sequencing reactions were performed on double-stranded DNA plasmids with external vector-based primers and designed primers in thepresence of either T7 sequencing mixes or theDeaza T7 sequencing mixes (Pharmacia) and la-belled nucleotides ([a-35S]dATP or [a-35S]dCTP,1000 mCi/mmol; Dupont). The aminio acid align-ment was performed using the Clustal V program.
2.5. Pulsed field gel electrophoresis (PFGE)
Pulsed field gel electrophoresis was carried outas described by Liu and Sanderson [26]. Thechromosomes were separated using a Bio-RadDRII contour-clamped homogeneous electric fieldapparatus or a Bio-Rad Mapper. The pulse timesof 60–150 s were routinely used and the totalrunning time was 36–48 h. Electrophoresis wascarried out in 0.7% agarose gels containing 1.5 mgml−1 ethidium bromide and 0.5× TBE buffer(1× buffer contains 90 mM Tris, 90 mM boricacid, 2 mM EDTA (pH 8.0)). Southern blot ofpulsed field gels was performed as describedabove.
2.6. O6erexpression of cysteine protease inLeishmania cells
Two oligonucleotide primers containing BamHIrestriction sites were used in PCR to amplify thecysteine protease coding region from each of thetwo distinct cDNAs. The sequences of the fourprimers are: Sense primer (Ldccys1-GST1): 5%-TTCTCTTCCCCTCGCTGGATCCTCCGCGA-TGG-3%; antisense primer (Ldccys1-GST2): 5%-AGACGCAGGATCCACCCCTACGTG-3%;sense primer (Ldccys2-GST1): 5%-GCACGTTG-CACCAACAAGGATCCGCAAAG-3%; antisenseprimer (Ldccys2-GST2): 5%-GCACAGTGGATC-
CTGCCAGTCCTA-3%. The underline indicatesthe position of BamHI restriction sites in all thefour primers. The PCR reactions were performedas described above and the PCR products whichrepresented the coding region of each of the twocysteine protease cDNAs was digested withBamHI, gel purified and ligated into the BamHIsite of pX (kindly provided by Dr Steven Beverleyfrom Harvard Medical School, Boston, MA). Theligation reaction was used to transform the DH5a
strain of Escherichia coli by electroporation asdescribed [28]. Plasmid clones containing the cD-NAs were selected by restriction analysis andsequencing. Transfections of L. d. chagasi pro-mastigotes were performed according to protocolsdescribed by others [29,30]. Briefly, log phase L. d.chagasi promastigotes were washed twice in phos-phate buffered saline (PBS) and resuspended at arate of 1×108 ml−1 in electroporation buffer (21mM Hepes, pH 7.5, 137 mM NaCl, 5 mM KCl,0.7 mM Na2PO4, 6 mM glucose) and 50 mg ofplasmid pX-Ldccys1 or pX-Ldccys2 or pX alone.The promastigotes resuspended in electroporationbuffer with 50 mg of plasmid DNA (total volume400 m l) were placed in 0.2 cm cuvette and electro-poration was performed using Gene Pulser (BIO-RAD) delivering a pulse of 1.2 kV andcapacitance at 250 mF. Electroporation was re-peated three times at 10 min intervals with pro-mastigotes in cuvette sitting on ice in between.Transfected promastigotes were transferred todrug-free HOMEM media overnight. After selec-tion in HOMEM media containing 50 mg ml−1 ofG418 (GIBCO BRL) for about 2 weeks, resistantpromastigotes appeared. The concentration ofG418 was then increased at a weekly interval by50 mg ml−1 until it reached 400 mg ml−1.
2.7. Assays for proteolytic acti6ities of cysteineprotease
Biological activity of recombinant cysteineprotease was detected in gelatin SDS-PAGE (non-denaturing) prepared according to the recipe ofHotez et al. [31] with some modifications. Briefly,the separating gel contained 2.8 ml of 1.5 MTris–HCl (pH 8.8) and 0.4% SDS, 3.0 ml of28.4% acrylamide and 1.6% bis-acrylamide (final

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267 251
concentration was equal to 12.5%), 0.8 ml of 1%gelatin (while still hot), 0.8 ml of water, 10 m l ofTEMED (N,N,N %,N %-tetra-methyl-ethylene-di-amine), and 60 m l of 10% ammonium persulfate.The stacking gel contained, 1.25 ml of 0.5 MTris–HCl (pH 6.8), and 0.4% SDS, 0.5 ml of28.4% acrylamide and 1.6% bis-acrylamide, 0.5 mlof 1% gelatin (while still hot), 2.75 ml of water, 10m l of TEMED, and 50 m l of 10% ammoniumpersulfate. The protein samples were applied tothe gels in an SDS sample buffer (62.5 mM Tris–HCl (pH 6.8), 2% SDS, 10% glycerol and bro-mophenol blue) without heating. The procedurewas modified to include SDS in both the gel andthe running buffer so as to allow separation ofcysteine protease as a function of molecularweight. The gel was run at low voltage (approx.50 V) to prevent heating during the run. Uponcompletion of the run, the gel was washed once in2.5% Triton X-100 (Sigma) for 1 h then washed in0.1 M sodium acetate (pH 5.5) and 1 mM DDT(Sigma) for 1 h. The gel was then incubated infresh 0.1 M sodium acetate (pH 5.5)+1 mMDDT+20 mM Z-Phe-Arg-MCA (Peninsula Lab-oratories, CA) overnight at 37°C. The gels werestained the next day by placing in 50% methanol,10% acetic acid, containing 0.2% Coomassie bluefor 30 min. The gel was then destained for at least2 h in a solution containing 7% methanol, 5%acetic acid in water. Once bands of proteolysisbecame visible, gels were photographed. Toconfirm that the enzymatic activity was due tocysteine protease, inhibitors of cysteine protease(2 mM leupeptin and 2 mM iodoacetamide bothfrom Sigma) were included in all the buffers usedto incubate control gels.
2.8. Generation of antibody against cysteineprotease by DNA immunization of mice
The direct DNA injection technology [32] wasemployed to generate antibodies against Ldccys1and Ldccys2. Briefly, the coding regions of thetwo cDNAs of cysteine proteases were amplifiedby PCR using the same primers described underoverexpression and subcloned into the BamHIsite of a plasmid VR1012 containing a eukaryoticpromoter (plasmid VR1012; kindly provided by
Dr Jon Norman, Vical, San Diego, CA). Theplasmid clones having inserts in the correct orien-tation were selected by restriction analysis andconfirmed by sequencing. These constructs weredelivered to experimental animals by direct injec-tion into the hind limb quadriceps muscles. Micewere injected with 50 mg of DNA in saline and ina total volume of 100 m l. All mice were boostedafter 21 days and tested for antibody response 7days after the boost. Some mice were boosted fora second time after another 21 days from the firstboost. A titre of 1:500 was regarded as goodresponse.
2.9. SDS-PAGE and Western blot analysis ofLeishmania cells o6erexpressing cysteine protease
SDS-PAGE was performed according to themethod of Laemmli [33] using 12% polyacry-lamide gels. Transfected and untransfected pro-mastigotes were washed and resuspended in lysisbuffer (20 mM Tris pH 7.3, 40 mM NaCl, 10 mMEDTA and 20 mM phenylmethylsulphonylfluoride (PMSF)) [34] and sonicated for 5×15 son ice with a Virsonic cell disrupter at a setting of60 using a small probe. Cell lysate was treatedwith an equal volume of 2% SDS and one part of5× sample buffer (formulated as described byBIO-RAD) was added to four parts of the solubi-lized protein. The proteins were then denaturedfor 3 min in a 100°C boiling water bath andloaded into wells of a 4% stacking. After elec-trophoresis the protein was transferred to aPVDF-Plus transfer membrane (Micron Separa-tions) and probed with antibody raised againstLdccys1 through direct DNA injection technol-ogy. Reactivity was detected by using the ECLsystem (Amersham).
3. Results
3.1. PCR amplification of cysteine protease genes
In order to determine the population of cysteineprotease genes in the genome of L. d. chagasi wecarried out the PCR amplification of the cysteineprotease DNA fragment from genomic DNA us-

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267252
Fig. 1. DNA and the deduced amino acid sequences of (i) Ldccys1 and (ii) Ldccys2 cDNAs. The Ldccys1 and Ldccys2 cDNAs haveopen reading frames of 447 and 471 amino acids, respectively. In the 5% UTR of Ldccys1, an open reading frame of 23 amino acidsis shown in bold. The LCP-1 and LCP-2 primers used to amplify the 500 bp fragment are underlined.

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267 253
Fig. 1. (i) (Continued)
ing two primers (LCP1 and LCP2). A 500 bpPCR product was obtained. The PCR productwas digested sequentially with HindIII andEcoRI, gel purified and subcloned in pGEMR-2(Promega). Plasmid DNA from a large number ofclones were isolated and sequenced. Our resultsshowed the presence of two distinct types of cys-teine protease DNA sequence which we desig-nated as Ldccys1 and Ldccys2. The Ldccys1 genewas more abundant in the L. d. chagasi genomethan the Ldccys2 gene. We also found the pres-ence of a related variant of the Ldccys1 genewhich only differs with the deletion of 13 aminoacids close to the LCP2 primer and this variantwas under-represented in the genome relative tothe Ldccys1 gene (data not shown).
3.2. Screening of the cDNA libraries
In order to isolate full length cDNAs represent-ing the Ldccys1 and Ldccys2 cysteine proteasegenes, we screened promastigote and amastigotespecific L. dono6ani chagasi cDNA libraries in
l-ZAP II as described in Section 2. From eachcDNA library we sequenced several positiveclones. All the 16 clones from the promastigotespecific cDNA library corresponds to Ldccys2 asdetermined by sequencing within the 500 bp. Sim-ilarly, all the 20 clones from the amastigote spe-cific cDNA library corresponds to Ldccys1 (alsodetermined by sequencing within the 500 bp) ex-cept one clone which contains the variant form. Inaddition to the 13 amino acid deletion, this vari-ant form also differed extensively 3% to LCP2towards the carboxy-terminal end (data notshown). Furthermore, when the amastigotecDNA library blots were stripped and rehy-bridized with the Ldccys2 probe, 4 clones werepicked from positions which differed fromamastigote clones. However, we did not furtheranalyse these clones. These findings suggest thatLdccys1 is amastigote specific, while Ldccys2 ap-pears to be not stage-specific.
The sequences of Ldccys1 and Ldccys2 cDNAsare shown in Fig. 1. These cDNAs may not be fulllength since the 5% ends of both Ldccys1 and

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267254
(ii)
Fig. 1. (ii) (Continued)

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267 255
Fig. 1 (ii). (Continued)
Ldccys2 do not coincide with the published se-quence for L. dono6ani splice leader [35]. Ldccys2has an open reading frame of 471 amino acidsand Ldccys1 has an open reading frame of 447amino acids (Fig. 1). The difference in lengthbetween the two proteins is mainly due to a 21amino acid deletion from the carboxy-terminalregion of Ldccys1 (Fig. 2). It is interesting to notethat there is an open reading frame of 23 aminoacids in the 5%-untranslated region of the Ldccys1cDNA. At the amino acid level, comparativeanalysis of the cDNA sequences for Ldccys1 andLdccy2 show a similarity of 40%, suggesting thetwo cDNAs are distinct. In order to determine thedegree of similarity, we aligned the deducedamino acid sequences of the two distinct cysteineproteases with those of cysteine proteases fromLeishmania pifanoi, a member of L. mexicanacomplex (Fig. 2). The amino acid sequence ofLdccys1 when compared with that of Lpcys2
which is the corresponding gene from L. pifanoishow 75% similarity while Ldccys2 is 78% similarto the available amino acid sequence of Lpcys1which is the corresponding gene from L. pifanoi[18].
The deduced amino acid sequence from Ldc-cys1 and Ldccys2 cDNAs contain all the featurescharacteristic of the different members of the cys-teine protease family of proteins (Fig. 2). Thus, itcontains the active site cysteine at positions 149and 150 in Ldccys1 and Ldccys2, respectively.Glycine which is involved in substrate binding inpapain is also present at positions 147 (in Ldc-cys1) and 148 (in Ldccys2). In addition, otheramino acid residues important in catalysis includ-ing histidine and asparagine are present. Histidineis located at positions 286 (in Ldccys1) and 289(in Ldccys2) while asparagine is present at posi-tions 306 and 309 in Ldccys1 and Ldccys2, respec-tively. In addition, other amino acids well

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267256
Fig. 2. Comparison of the deduced amino acid sequences from Ldccys1 and Ldccy2 to the deduced amino acid sequences of Lpcys1and Lpcy2 from Leishmania pifanoi. Ldccys1 is homologous to Lpcys2 and Ldccys2 is homologous Lpcys1. For alignment of aminoacid sequences the Clustal V program was used. Amino acid residues common to the cysteine proteases are shaded and boxed.Shaded residues imply similarity/identity amino acids among the cysteine proteases. The ‘–’ symbol indicates gap introduction intothe sequences to maximize alignment.
conserved in other cysteine proteases [17,18] arepresent in Ldccys1 and Ldccys2.
From this amino acid sequence alignment (Fig.2) and sequence information on other cysteineproteases [17], the Ldccys1 and Ldccys2 proteins
are composed of pre- and pro-regions followed bya mature protease core and a C-terminal exten-sion. The N-terminal region (pre-) of both cys-teine protease sequence is hydrophobic andappears to be represented by residues 1–37 and

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267 257
Fig. 2. (Continued)
1–39 for Ldccys1 and Ldccys2, respectively. Thepro-region for Ldccys1 appears to be cleavedbetween amino acid residues 123 and 124 generat-ing an alanine as in the case of Lpcys2 and othercysteine proteases. Similarly, the pro-region ofLdccys2 also appears to have a cleavage site be-tween amino acids 127 and 128 resulting in aglycine and this position has been shown to bedifferent in other cysteine proteases (Fig. 2, [17]).The cleavage site between the mature proteasecore and the C-terminal extension region found incruzain [36], which is Val-Gly/-Gly-Pro was notfound in both L. d. chagasi cysteine proteases andthus, the cleavage site at this point is not clear. Inaddition, a stretch of amino acid sequences rich inAla, Gly, Pro, Ser, and Thr which have beendescribed downstream of the cleavage site of themature protease core and C-terminal region for L.pifanoi and L. mexicana cysteine proteases werenot present in both L. d. chagasi cysteineproteases. In the C-terminal region, Ldccys1 andLdccys2 contain ten and seven cysteine residues,respectively, which align with the cysteine residuesin other cysteine proteases (Fig. 2, [17]). In addi-tion, Ldccys2 has two major insertions betweenamino acids 30–36 and 429–449 (Fig. 2). Taken
together, these structural comparisons stronglysuggest that the cDNAs presented in this paperare cysteine proteases.
3.3. Southern blot analysis
In order to understand the organization of cys-teine protease genes in the Leishmania genome,two identical Southern blot of L. d. chagasi andL. d. dono6ani genomic DNA digested with SacI,PstI and SalI restriction enzymes were preparedas described in Section 2. One blot was hybridizedwith a 32P-labelled 500 bp fragment of Ldccys1and the other blot was hybridized with a 32P-la-belled 500 bp fragment from Ldccys2. The resultsin Fig. 3 (Panel A) show L. d. chagasi genomicDNA blot hybridized with Ldccys1 probe. SacIdigest gave a single hybridizable band of about 15kb (this was clear when the blot was underex-posed); PstI digest gave a strong band at about1.2 kb and two relatively fainter bands of 0.6 and1.7 kb. SalI gave a strong band at about 3.1 kbwith two fainter bands at 3.3 and 2.2 kb. Similarbands were detected when the L. d. dono6anigenomic DNA was cut with the same enzymesand hybridized with the Ldccys1 probe. On the
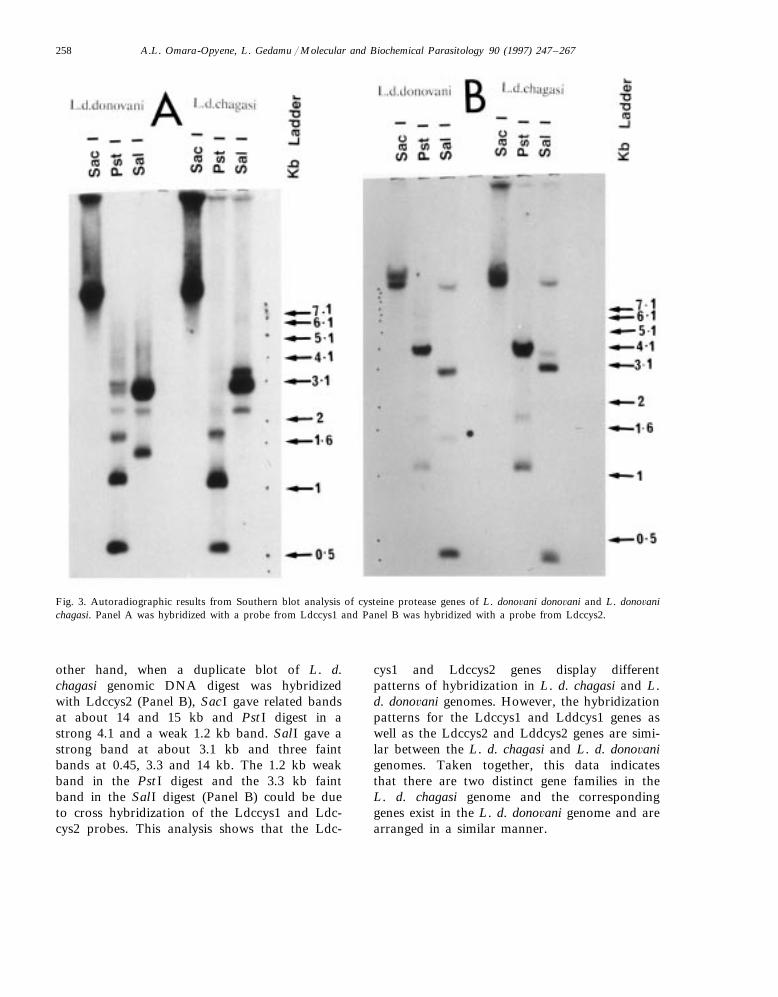
A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267258
Fig. 3. Autoradiographic results from Southern blot analysis of cysteine protease genes of L. dono6ani dono6ani and L. dono6anichagasi. Panel A was hybridized with a probe from Ldccys1 and Panel B was hybridized with a probe from Ldccys2.
other hand, when a duplicate blot of L. d.chagasi genomic DNA digest was hybridizedwith Ldccys2 (Panel B), SacI gave related bandsat about 14 and 15 kb and PstI digest in astrong 4.1 and a weak 1.2 kb band. SalI gave astrong band at about 3.1 kb and three faintbands at 0.45, 3.3 and 14 kb. The 1.2 kb weakband in the PstI digest and the 3.3 kb faintband in the SalI digest (Panel B) could be dueto cross hybridization of the Ldccys1 and Ldc-cys2 probes. This analysis shows that the Ldc-
cys1 and Ldccys2 genes display differentpatterns of hybridization in L. d. chagasi and L.d. dono6ani genomes. However, the hybridizationpatterns for the Ldccys1 and Lddcys1 genes aswell as the Ldccys2 and Lddcys2 genes are simi-lar between the L. d. chagasi and L. d. dono6anigenomes. Taken together, this data indicatesthat there are two distinct gene families in theL. d. chagasi genome and the correspondinggenes exist in the L. d. dono6ani genome and arearranged in a similar manner.

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267 259
3.4. Arrangement of cysteine protease genes inthe L. d. chagasi genome
In order to determine the arrangement of cys-teine protease genes in the genome of L. d.chagasi, we tested whether the cysteine proteasegenes we have isolated are arranged in tandem.To do this, a restriction enzyme which cuts thecDNA cloned once was first identified. SalI andStuI were found to cut the Ldccys1 and theLdccys2 cDNAs, respectively. Thus, genomicDNA from L. d. chagasi was digested with theseenzymes separately to completion and the twoidentical blots were prepared for each digest. Aduplicate blot from SalI digest was hybridizedwith a probe representing sequences 5% end to theSalI site of the Ldccys1 cDNA. The other dupli-cate blot was hybridized with a probe representingsequences 3% end to the SalI site. Similarly, hy-bridization was carried out using probes repre-senting sequences 5% and 3% end of StuI site of theLdccys2 cDNA. The results of these studies areshown in Fig. 4. As shown in Panel A, a 3.1 kbfragment hybridized when the blot was probedwith either probes. However, the 5% end probehybridized to a 3.6 kb fragment. On the otherhand, a 2.4 kb fragment hybridized faintly to the3% end probe. The 3.6 and 2.4 kb bands areprobably the 5% and 3% ends of the Ldccys1 genecluster. Similar analysis was performed on theLdccys2 gene(s). As shown in Panel B, 15 and 2.8kb hybridizable bands were obtained using 5% and3% end probes, respectively. From this data, itappears that there is at least one Ldccys2 gene inthe L. d. chagasi genome while the Ldccys1 genesare represented by multi-copy genes which arearranged in tandem.
3.5. Chromosomal localization of cysteineprotease genes
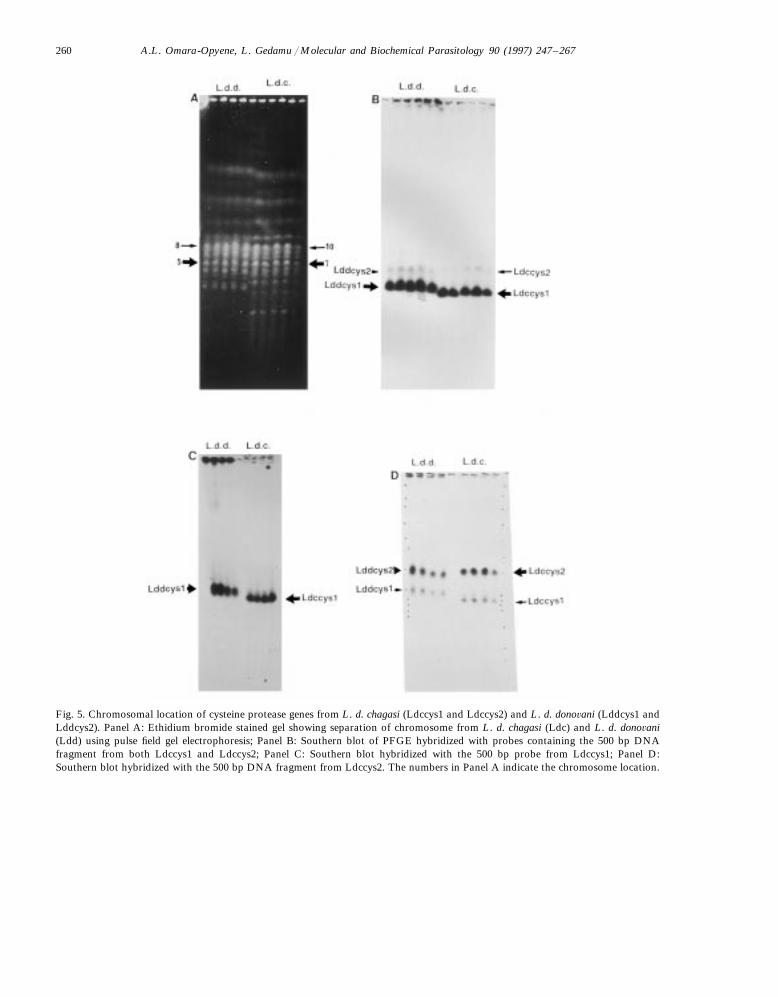
In an attempt to find out the chromosomeswhere each of the cysteine protease genes is lo-cated in the Leishmania genome, pulsed field gelelectrophoresis was used to resolve chromosomesize DNA. The resolved DNA was blotted andhybridized separately with 32P-labelled 500 bpfragment as probe containing (i) both Ldccys1
and Ldccys2, (ii) Ldccys1 and (iii) Ldccys2. Theresults of these analyses is shown in Fig. 5. Ethid-ium bromide stained gel showing the separationof L. d. chagasi (Ldc) and L. d. dono6ani (Ldd) ispresented in Fig. 5A. When probes containing amixture of Ldccys1 and Ldccys2 sequences wereused, two hybridizable bands, one very strong andone very faint, were detected in both L. d. chagasiand L. d. dono6ani (Fig. 5B). In L. d. chagasi, thestrong band is located on chromosome 7 and aweak one on chromosome 10. On the other hand,in L. d. dono6ani, the strong and the faint bandsare located on chromosomes 5 and 8, respectively.The chromosomal numbering was on the basis ofcounting from the smallest visible chromosomesize DNA.
Fig. 4. Arrangement of Ldccys1 and Ldccys2 genes. Autora-diographic results from the Southern blot analysis. Panel A:The lane labelled SalI 5% was hybridized with probe represent-ing sequences 5% to SalI site in Ldccys1. The lane labelled SalI3% was hybridized with probe representing sequences 3% to SalIsite in Ldccys1. Panel B: The lane labelled StuI 5% was hy-bridized with probe representing sequences 5% to StuI site inLdccys2. The lane labelled StuI 3% was hybridized with proberepresenting sequences 3% to StuI site in Ldccys2.
A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267260
Fig. 5. Chromosomal location of cysteine protease genes from L. d. chagasi (Ldccys1 and Ldccys2) and L. d. dono6ani (Lddcys1 andLddcys2). Panel A: Ethidium bromide stained gel showing separation of chromosome from L. d. chagasi (Ldc) and L. d. dono6ani(Ldd) using pulse field gel electrophoresis; Panel B: Southern blot of PFGE hybridized with probes containing the 500 bp DNAfragment from both Ldccys1 and Ldccys2; Panel C: Southern blot hybridized with the 500 bp probe from Ldccys1; Panel D:Southern blot hybridized with the 500 bp DNA fragment from Ldccys2. The numbers in Panel A indicate the chromosome location.

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267 261
In order to determine the chromosomal localiza-tion of each of the cysteine protease gene family,we hybridized the blot with specific probes. Whenthe blot was hybridized with the probe containingLdccys1 sequences, only the strong band wasobserved in both L. d. chagasi and L. d. dono6anigenomes (Fig. 5C). This data suggested that in L.d. chagasi, the Ldccys1 genes are located on chro-mosome 7 while the corresponding cysteine proteasegenes in L. d. dono6ani (Lddcys1) are located onchromosome 5. On the other hand, when the blotwas hybridized with the probe containing Ldccys2sequences, two hybridizable bands were obtained inboth L. d. chagasi and L. d. dono6ani genomes (Fig.5D). The stronger bands were on chromosome 10for L. d. chagasi and 8 of L. d. dono6ani genomes.The fainter bands were on chromosome 7 of L. d.chagasi and 5 of L. d. dono6ani genomes. Interest-ingly, the position of the faint hybridizable bandsis in the same chromosomes for the Ldccys1/Ldd-cys1 genes. It is not clear at present whether genesrelated to Ldccys2/Lddcys2 are located on that samechromosome. Alternatively, the probe may havecross-reacted with related sequences in the Ldccys1/Lddcys1 gene cluster. In any event, this data suggestthat the Ldccys2 and Lddcys2 genes are located onchromosome 10 of the L. d. chagasi and 8 of theL. d. dono6ani genomes, respectively. Taken to-gether, this data shows that the two distinct cysteineprotease genes are located on two separate chromo-somes in the L. d. chagasi genome and the organi-zation of the corresponding genes are similar in theL. d. dono6ani genome.
3.6. mRNA expression of Ldccys1 and Ldccys2
In order to determine the expression pattern ofthe two distinct cysteine protease genes, we exam-ined the level of mRNAs using Northern blotanalysis. For this purpose, we utilized the 500 bpfragment probe specific for each cysteine proteasegene. We isolated total RNA from log and station-ary stages of promastigote as well as from amastig-otes as described in Section 2. As shown in Fig. 6B(top panel), a 2.6 kb message (this mRNA is theexpected size from the promastigote specific cysteineprotease cDNA sequence) hybridized with theLdccys2 probe in the log (lane 1), stationary (lane
2), and amastigote (lane 3) stages of development.The levels of the message hybridized were compara-ble in all stages if we consider the amount of totalparasite RNA present in the amastigote stage (lane3) and this message is not detectable in spleens fromuninfected hamsters (lane 4). On the other hand,when the same blot was hybridized with the Ldccys1probe, a predominant 2.4 kb message (this mRNAis the expected size from the amastigote specificcysteine protease cDNA sequence) with a moder-ately abundant 4.0 kb transcript and a very low levelof transcript approximately 5.0 kb were detected inthe RNA from amastigote stage (Fig. 6A, lane 3),but not in the log and stationary stages of thepromastigote (Fig. 6A, lanes 1 and 2) and in spleensof uninfected hamsters (Fig. 6A, lane 4). It isinteresting to note that the level of Ldccys1 expres-sion is significantly higher than that of Ldccys2 inthe amastigotes considering that the same blot wasused for both hybridizations (top panels A and B)after stripping the probe from the membrane. Thelevels of loading were assessed by stripping the blotsand rehybridizing with the LcPo probe which isexpressed constitutively (Fig. 6A,B, middle panels)as well as from ethidium bromide stained gels (Fig.6A,B, lower panels). The faint hybridizable LcPobands in the amastigote stage is due to a low amountof the RNA from the amastigotes. Taken together,our data suggest that the Ldccys1 genes are ex-pressed abundantly in that stage of development butat very low level in the promastigote stage ofdevelopment. Conversely, the Ldccys2 gene is ex-pressed in both the promastigote and amastigotestages of development.
3.7. O6erexpression of cysteine protease inLeishmania dono6ani chagasi cells
In order to show that the two cDNAs code forcysteine proteases, each cDNA was cloned in thepX vector. L. d. chagasi promastigotes were trans-fected with the recombinant plasmids and selectedwith G418. The presence of the plasmids contain-ing the cDNAs were confirmed by Southern blotanalysis of BamHI digested total DNA fromtransfected cells. As shown in Fig. 7, the cellstransfected with recombinant plasmids show hy-bridizable bands at positions 1.3 (lane 2) and 1.4

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267262
Fig. 6. Northern blot analysis of Ldccys1 and Ldccys2 transcripts. Total RNA isolated from: lane 1, log phase promastigoteparasites; lane 2, stationary phase promastigote parasites; lane 3, spleens of hamsters infected with L. d. chagasi ; and lane 4, spleensof uninfected hamsters. Top panels A and B are blots hybridized with Ldccys1 and Ldccys2 probes, respectively. The blot shownin Panel A was first hybridized with a Ldccys1 probe, then it was stripped and rehybridyzed with a Ldccys2 (Panel B). The middlepanels A and B are blots hybridized with LcPO probe which is constitutively expressed in L. d. chagasi and used as an internalcontrol. All blots were exposed for 16 h except lanes 3 and 4 (top panel A) which were exposed for 48 h. The bottom panels A andB are ethidium bromide stains of RNA used as loading controls. Leishmania RNA has three bands. RNA from spleens of infectedhamsters shows 28S and 18S (strong bands). The two faint bands are due to Leishmania RNA. However, the third band co-migratewith the 18S RNA from macrophages of infected hamsters. The arrows indicate transcript sizes and the size of the RNA marker(M) is also shown.
kb (lane 3) for the Ldccys1 and Ldccys2 genes,respectively. However, such bands were absentfrom cells transfected with the pX vector alone(lanes 1 and 4). In addition, a band representingeach cysteine protease gene in the genome wasalso detected. These results demonstrate the pres-ence of the constructs in the transfected cells.
In order to show that the transfected cells areover-producing biologically active cysteineproteases, enzymatic activities of cell lysates wereanalyzed in gelatin SDS-PAGE (non-denaturing).Only cells transfected with recombinant con-structs were found to over-produce biologicallyactive cysteine proteases (Fig. 8A, lanes 1 and 3).
Enzymatic activities were seen at about 43 and 30kDa (lanes 1 and 3), the latter appears to be theprocessed form of cysteine proteases [34]. When asimilar gel is incubated in a buffer which includedspecific inhibitors to cysteine proteases, the enzy-matic activities seen at about 43 and 30 kDadisappeared (Fig. 8B), confirming that the enzy-matic activities are indeed due to the over-ex-pressed cysteine proteases. This data shows thatbiologically active cysteine proteases are over-pro-duced in L. d. chagasi transfected promastigotes.
In order to further demonstrate the overexpres-sion, protein extracts from cells transfected withthe recombinant plasmid containing the Ldccys1

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267 263
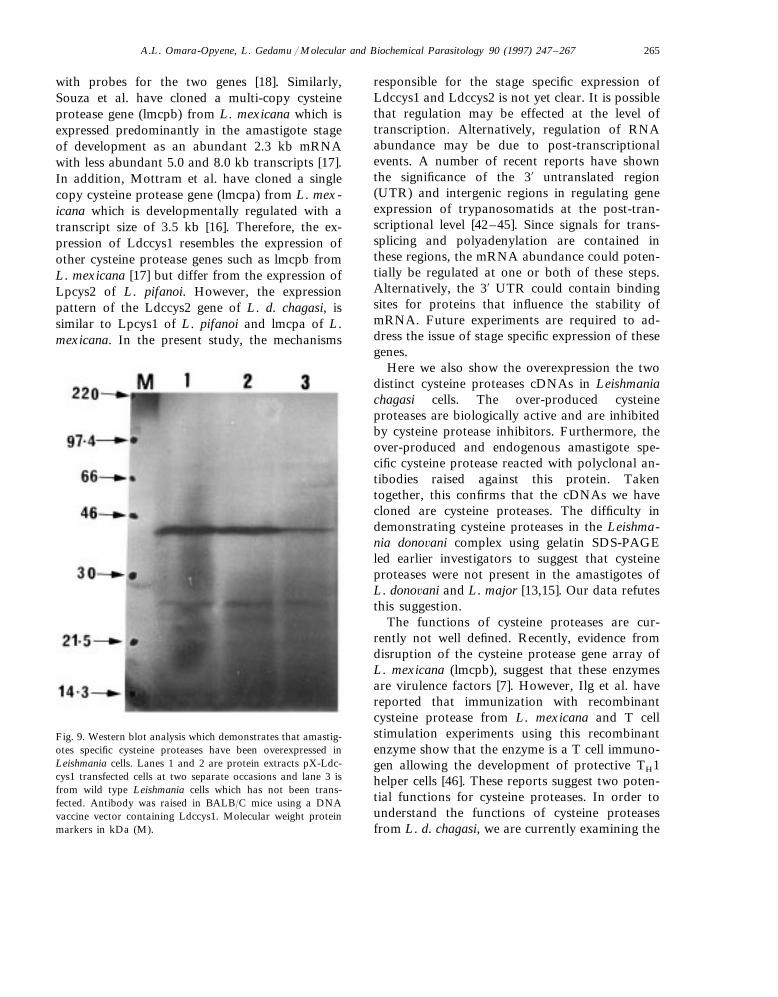
cDNA were analyzed by Western blot analysisusing a specific polyclonal antibody generated bydirect DNA injection technology as described inSection 2 as well as in the literature [32,39–41].As shown in Fig. 9, a 43 kDa band correspondingto the unprocessed form of cysteine protease wasdetected. This band was present at a higher level
in the transfectants (lanes 1 and 2) compared tothe wild type transfected with the pX vector alone(lane 3). In addition, there are minor bands atabout 26 kDa which are probably due to theprocessed forms of this enzyme. This data furtherconfirms the overexpression of amastigote specificcysteine proteases in Leishmania cells. However,the difficulty of raising polyclonal antibody toLdccys2 prevented us from carrying out similaranalysis on protein extracts from cells transfectedwith the recombinant plasmid containing Ldccys2cDNA.
4. Discussion
For the first time we show that there are twodistinct cysteine protease genes in the genome ofLeishmania don6ani chagasi, a member of theLeishmania dono6ani complex, which causes vis-ceral leishmaniasis. The two genes isolated werecharacterized and found to have structural char-acteristics of cysteine proteases which have previ-ously been isolated from other Trypanosomatids.However, the two genes differ at the levels ofamino acid sequence, genomic organization andgene expression. At the level of the amino acidsequence, there is a 40% similarity between Ldc-cys1 and Ldccys2 which is comparable to thesimilarity found between Lpcys2 and Lpcys1 of L.pifanoi [18] as well as between lmcpb and lmcpa ofL. mexicana [16,17]. On the other hand, Ldccys1is homologous to Lpcys2 of L. pifanoi becausethey are 75% similar at the level of protein se-quence. The Ldccys2 is also homologous to Lp-cys1 of L. pifanoi as they are 78% similar in theirprotein sequence. This fact indicates that the twocysteine protease genes are distinct and probablyarose by some duplicate event.
At the level of genomic organization, we showthat Ldccys1 belongs to a family of cysteineproteases encoded by tandemly organized geneswhile Ldccys2 appears to be a single cysteineprotease gene. The organization of the twofamilies of cysteine protease genes in L. d. dono-6ani was also found to be similar. In this regard,Ldccys1 is similar to other multi-copy cysteineprotease genes such as lmcpb from L. mexicana
Fig. 7. Southern blot analysis of total DNA from L. d. chagasitransfected cells. The total DNA was digested with BamHI.Lanes 1 and 4, pX transfected cells. Lanes 2 and 3, pX-Ldc-cys1 and pX-Ldccys2 transfected cells, respectively. Lanes 1and 2 were hybridized with Ldccys1 probe while lanes 3 and 4were hybridized with Ldccys2 probe. The strong bands at 1.3and 1.4 kb in lanes 2 and 3 indicate the presence of the codingregions of Ldccys1 and Ldccys2, respectively. The upper bandsin lanes 1 and 2 are due to endogenous Ldccys1 in L. d.chagasi genome present in several copies. The endogenousLdccys2 could not be detected in lanes 3 and 4 due to the shortexposure of the blots because it is less abundant in L. d.chagasi genome.

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267264
Fig. 8. Panel A: Gelatin SDS-PAGE (non-denaturing) showing the enzymatic activity of biologically active cysteine proteasesover-expressed in L. d. chagasi cells. Panel B: Shows a similar gel incubated with inhibitors of cysteine proteases (2 mM leupeptinand 2 mM iodoacetamide). The enzymatic activity of wild type gp63 at 63 kDa and above was not affected by inhibitors of cysteineproteases and confirms equal loading. Leishmania cells are transfected with pX-Ldccys1 (lane 1), pX-Ldccys2 (lane 3) and pX alone(lanes 2 and 4). The size of the proteins is represented by the marker lane (M).
[17], Lpcys2 from L. pifanoi [18], cysteine proteasefrom T. brucei [37] and cruzain from T. cruzi [38].On the other hand, Ldccys2 resembles other singlecopy cysteine protease genes such as lmcpa fromL. mexicana [17] and Lpcys1 from L. pifanoi [18].
At the level of gene expression, we show thatthe Ldccys1 genes are expressed predominantly inthe amastigote stage of development but at a verylow level at the promastigote stage of develop-ment. Thus, this gene is expressed in a stage-spe-cific manner. On the other hand, the Ldccys2 geneis expressed throughout the parasite development,albeit, at a comparable level. Furthermore, theLdccys1 gene is expressed at a significantly higherlevel than the Ldccys2 gene at the amastigotestage. Although further functional analysis is re-quired, these results suggest that the Ldccys1genes are important and may play a significant
role at the amastigote stage of development. Thetranscript size of Ldccys2 is 2.6 kb. On the otherhand, Ldccys1 has a 2.4 kb abundant transcriptwith a relatively high level transcript of 4.0 kband a weak 5.0 kb transcript. Traub-Cseko et al.found Lpcys1 which is homologous to Ldccys2and Lpcys2 which is homologous to Ldccys1 areexpressed in both promastigotes and amastigotesof L. pifanoi, although the levels of expression ishigher for both in the amastigotes [18]. The pre-dominant mRNAs for Lpcys1 are 2.6 and 4.6 kbwith weaker transcripts of approximately 7.5 and8.1 kb. However, for Lpcys2 the predominantmRNA is 2.3 kb, with a moderately abundanttranscript of 4.4 kb and very low levels of tran-scripts of approximately 7 kb [5]. Similar observa-tion was also made for amastigotes derived fromtissue infected with L. amazonensis and hybridized
A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267 265
with probes for the two genes [18]. Similarly,Souza et al. have cloned a multi-copy cysteineprotease gene (lmcpb) from L. mexicana which isexpressed predominantly in the amastigote stageof development as an abundant 2.3 kb mRNAwith less abundant 5.0 and 8.0 kb transcripts [17].In addition, Mottram et al. have cloned a singlecopy cysteine protease gene (lmcpa) from L. mex-icana which is developmentally regulated with atranscript size of 3.5 kb [16]. Therefore, the ex-pression of Ldccys1 resembles the expression ofother cysteine protease genes such as lmcpb fromL. mexicana [17] but differ from the expression ofLpcys2 of L. pifanoi. However, the expressionpattern of the Ldccys2 gene of L. d. chagasi, issimilar to Lpcys1 of L. pifanoi and lmcpa of L.mexicana. In the present study, the mechanisms
responsible for the stage specific expression ofLdccys1 and Ldccys2 is not yet clear. It is possiblethat regulation may be effected at the level oftranscription. Alternatively, regulation of RNAabundance may be due to post-transcriptionalevents. A number of recent reports have shownthe significance of the 3% untranslated region(UTR) and intergenic regions in regulating geneexpression of trypanosomatids at the post-tran-scriptional level [42–45]. Since signals for trans-splicing and polyadenylation are contained inthese regions, the mRNA abundance could poten-tially be regulated at one or both of these steps.Alternatively, the 3% UTR could contain bindingsites for proteins that influence the stability ofmRNA. Future experiments are required to ad-dress the issue of stage specific expression of thesegenes.
Here we also show the overexpression the twodistinct cysteine proteases cDNAs in Leishmaniachagasi cells. The over-produced cysteineproteases are biologically active and are inhibitedby cysteine protease inhibitors. Furthermore, theover-produced and endogenous amastigote spe-cific cysteine protease reacted with polyclonal an-tibodies raised against this protein. Takentogether, this confirms that the cDNAs we havecloned are cysteine proteases. The difficulty indemonstrating cysteine proteases in the Leishma-nia dono6ani complex using gelatin SDS-PAGEled earlier investigators to suggest that cysteineproteases were not present in the amastigotes ofL. dono6ani and L. major [13,15]. Our data refutesthis suggestion.
The functions of cysteine proteases are cur-rently not well defined. Recently, evidence fromdisruption of the cysteine protease gene array ofL. mexicana (lmcpb), suggest that these enzymesare virulence factors [7]. However, Ilg et al. havereported that immunization with recombinantcysteine protease from L. mexicana and T cellstimulation experiments using this recombinantenzyme show that the enzyme is a T cell immuno-gen allowing the development of protective TH1helper cells [46]. These reports suggest two poten-tial functions for cysteine proteases. In order tounderstand the functions of cysteine proteasesfrom L. d. chagasi, we are currently examining the
Fig. 9. Western blot analysis which demonstrates that amastig-otes specific cysteine proteases have been overexpressed inLeishmania cells. Lanes 1 and 2 are protein extracts pX-Ldc-cys1 transfected cells at two separate occasions and lane 3 isfrom wild type Leishmania cells which has not been trans-fected. Antibody was raised in BALB/C mice using a DNAvaccine vector containing Ldccys1. Molecular weight proteinmarkers in kDa (M).

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267266
impact of Leishmania overexpressing cysteineproteases on Leishmania pathogenesis. We arealso attempting to disrupt these genes for furtherfunctional analyses. In addition, we are studyingthe mechanism of regulation of these family ofgenes.
Acknowledgements
This investigation received financial supportfrom the Medical Research Council (MRC) ofCanada and the UNDP/World Bank/WHO Spe-cial Programme for Research and Training inTropical Diseases to L.G. Pulsed field gel elec-trophoresis was run for us in the laboratory of DrKen Sanderson of the Department of BiologicalSciences, University of Calgary. The nucleic andamino acid sequences in this paper were alignedfor us by Julian Wood of the Department ofBiological Sciences, University of Calgary. We arevery grateful for all this help.
References
[1] WHO Technical Report Series 793. Control of Leishma-niasis. 1990.
[2] McKerrow JH. The proteases and the pathogenicity ofparasitic protozoa. Ann Rev Microbiol 1993;47:821–53.
[3] Combs GH, Baxter J. Inhibition of Leishmania amastig-ote growth by antipain and leupeptin. Ann Trop MedParasitol 1984;78:21–4.
[4] Coombs GH, Hart DH, Capaldo J. Proteinase inhibitorsas antileishmanial agents. Trans R Soc Trop Med Hyg1982;76:660–3.
[5] Robertson CL, Coombs GH. Multiple high activity cys-teine proteases of Leishmania mexicana are encoded bythe lmcpb gene array. Microbiology 1994;140:417–24.
[6] North MJ, Mottram JC, Coombs GH. Cysteineproteinases of parasitic protozoa. Parasitol Today1990;6:270–5.
[7] Mottram JC, Souza EA, Hutchinson JE, Carter R, FrameMJ, Coombs GH. Evidence from disruption of lmcpbgene array of Leishmania mexicana that cysteineproteinases are virulence factors. Proc Natl Acad SciUSA 1996;93:6008–13.
[8] Morihara K. Comparative specificity of microbialproteinases. Adv Enzymol 1974;41:179–243.
[9] Bazan JF, Fletterick JR. Viral cysteine proteinases arehomologous to the trypsin-like family of serine proteases:Structural and functional implications. Proc Natl AcadSci USA 1988;85:7872–6.
[10] Apodaca G, McKerrow JH. Regulation of Trichophytonrubrum proteolytic activity. Infect Immun 1989;57:3081–90.
[11] Eakin AE, Bouvier J, Sakanari JA, Craik CS, McKerrowJH. Amplification and sequencing of genomic DNA frag-ments encoding cysteine proteases from protozoan para-site. Mol Biochem Parasitol 1990;39:1–8.
[12] North MJ, Coombs GH. Proteinases of Leishmania mexi-cana amastigotes and promastigotes: Analysis by gel elec-trophoresis. Mol Biochem Parasitol 1981;3:293–300.
[13] Coombs GH. Proteinases of Leishmania and other flagel-late protozoan. Parasitology 1982;84:149–55.
[14] Pupkis MF, Combs GH. Purification and characteriza-tion of proteolytic enzymes of Leishmania amastigotesand promastigotes. J Gen Microbiol 1984;130:2375–83.
[15] Pupkis MF, Tetley F, Coombs GH. Leishmania mexi-cana : amastigote hydrolases in unusual lysosomes. ExpParasitol 1986;62:29–39.
[16] Mottram JC, Robertson CD, Coombs GH, Barry JD. Adevelopmentally regulated cysteine proteinase gene ofLeishmania mexicana. Mol Microbiol 1992;6:1925–32.
[17] Souza AE, Waugh S, Coombs GH, Mottram JC. Charac-terization of a multi-copy gene for a major stage-specificcysteine proteinase of Leishmania mexicana. FEBS Lett1992;311:124–7.
[18] Traub-Cseko YM, Duboise M, Boukai LK, McMahon-Pratt M. Identification of two distinct cysteine proteinasegenes of Leishmania pifanoi axenic amastigotes usingpolymerase chain reaction. Mol Biochem Parasitol1993;57:101–16.
[19] Pamer EG, So M, Davis CE. Identification of develop-mentally regulated cysteine protease of Trypanosomacruzi. Mol Biochem Parasitol 1989;33:27–32.
[20] Campetella O, Martinez J, Cazzullo JJ. A major cysteineproteinase is developmentally regulated in Trypanosomacruzi. FEMS Microbiol Lett 1990;67:145–50.
[21] Lockwood BC, North MJ, Mallinson DJ, Coombs GH.Analysis of Leishmania proteinases reveals developmen-tally changes in species-specific forms and a common68-kDa activity. FEMS Microbiol Lett 1987;48:345–50.
[22] Pearson RD, Steigbigel RT. Mechanism of lethal effect ofhuman serum upon Leishmania dono6ani. J Immunol1980;125:2195–201.
[23] Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning.A Laboratory Manual. 2nd ed. Cold Spring Harbor, NewYork: Cold Spring Harbor Laboratory, 1989.
[24] Wilson ME, Young BM, Andersen KP, Weinstock JV,Metwali A, Ali AK, Donelson JE. A recombinant Leish-mania chagasi antigen that stimulates cellular immuneresponses in infected mice. Infect Immun 1995;63:2062–9.
[25] Sanger F, Nicklen S, Coulson AR. DNA sequencing withchain-terminating inhibitors. Proc Natl Acad Sci USA1977;74:5463–7.
[26] Liu S-L, Sanderson KE. A physical map of Salmonellatyphimurium LT2 genome made by using XbaI analysis. JBacteriol 1992;174:1662–72.

A.L. Omara-Opyene, L. Gedamu / Molecular and Biochemical Parasitology 90 (1997) 247–267 267
[27] Chomczynski P, Sacchi N. Single-step method of RNAisolation by acid guanidium thiocyanate-phenol-chloro-form extraction. Anal Biochem 1987;162:156–9.
[28] Miller EM, Nickoloff JA. Escherichia coli electrotransfor-mation. In: Nickoff JA, editor. Methods in MolecularBiology, vol. 47, Electroporation Protocols for Microor-ganisms. Totowa (NJ): Humana, 1995:105–115.
[29] Laban A, Tobin JF, Curotto de Lafaille MA, Wirth DF.Stable expression of the bacterial neor gene in Leishmaniaenriettii. Nature 1990;343:572–4.
[30] Beverley SM, Clayton CE. Transfection of Leishmaniaand Trypanosoma brucei by electroporation. MethodsMol Biol 1993;21:333–48.
[31] Hotez PJ, Narasimhan S, Haggerty J, Milstone L, Bho-pale V, Schad GA, Richards FF. Hyaluronidase frominfective Ancylostoma hookworm larvae and its possiblefunction as a virulence factor in tissue invasion and incutaneous larvae migrans. Infect Immun 1992;160:1018–23.
[32] Ulmer JB, Donnelly JJ, Parker SE, Rhodes GH, FelgnerPL, Dwarki VJ, Gromkowski SH, Deck RR, DeWittCM, Friedman A, Hawe LA, Leander KR, Martinez D,Perry HC, Shiver JW, Montogomery DL, Liu MA. Het-erologous protection against influenza by injection ofDNA encoding a viral protein. Science 1993;259:1745–9.
[33] Laemmli UK. Cleavage of structural proteins during as-sembly of the head of bacteriophage T4. Nature1970;227:680–5.
[34] Duboise SM, Vannier-Santos MA, Costa-Pinto D, RivasL, Pan AA, Traub-Cseko Y, De Sauza W, McMahon-Pratt D. The biosynthesis, processing, and immunolocal-ization of Leishmania pifanoi amastigote cysteineproteinases. Mol Biochem Parasitol 1994;68:119–32.
[35] Wilson K, Hanson S, Landfear S, Ullman B. Nucleotidesequence of the Leishmania dono6ani medRNA gene. Nu-cleic Acids Res 1991;19:5787.
[36] Eakin EE, Mills AA, Harth G, McKerrow JH, Craik CS.The sequence, organization, and expression of the majorcysteine protease (cruzain) from Trypanosoma cruzi. JBiol Chem 1992;268:6115–8.
[37] Mottram JC, North MJ, Barry JD, Coombs GH. Acysteine proteinase cDNA from Trypanosoma brucei pre-dicts an enzyme with an unusual c-terminus extension.FEBS Lett 1989;258:211–5.
[38] Campetella O, Henriksson J, Asund L, Frasch ACC,Petersson U, Cazzullo JJ. The major cysteine proteinase(cruzipain) from Trypanosoma cruzi is encoded by multi-ple polymorphic tandemly organized genes located ondifferent chromosomes. Mol Biochem Parasitol1992;50:225–34.
[39] Waine GJ, McManus DP. Nucleic acids: vaccines of thefuture. Parasitol Today 1995;11:113–6.
[40] McDonnell WM, Askari FK. DNA Vaccines. New Engl JMed 1996;334:42–5.
[41] Ulmer JB, Donnelly JJ, Liu MA. DNA vaccines promis-ing: A new approach to inducing protective immunity.Am Soc Microbiol News 1996;62(9):476–9.
[42] Jefferies D, Tebabi P, Pays E. Transient activity assays ofthe Trypanosoma brucei variant surface glycoprotein genepromoter: control of gene expression at the post-tran-scriptional level. Mol Cell Biol 1991;11:338–43.
[43] Ramamoorthy R, Donelson JE, Paetz KE, Maybodi M,Roberts SC, Wilson ME. Three distinct RNAs for thesurface protease gp63 are differentially expressed duringdevelopment of Leishmania dono6ani chagasi promastig-otes to an infectious form. J Biol Chem 1992;267:1888–95.
[44] Teixeira SM, Kirchhoff L, Danelson J. Post-transcrip-tional elements regulating expression of mRNAs from theamastin/tuzin gene cluster of Trypanosoma cruzi. J BiolChem 1995;270:22586–94.
[45] Charest H, Zhang W-W, Matlashewski G. The develop-mental expression of Leishmania dono6ani A2 amastigote-specific genes is post-transcriptionally mediated andinvolves elements located in the 3%-untranslated region. JBiol Chem 1996;271:17081–90.
[46] Ilg T, Fuchs M, Gnau V, Wolfram M, Harbecke D,Overath P. Distribution of parasite cysteine proteases inlesions of mice infected with Leishmania mexicanaamastigotes. Mol Biochem Parasitol 1994;67:193–203.
.














![Cuba · 2012. 10. 18. · 3 Omara & Chucho Omara Portuondo [voz], Chucho Valdés [piano] Montuno: 2011 Jazz Cubano Madrid: Grupo Zeta, 2000 Rumba, Mambo, Cha-cha …](https://static.fdocuments.net/doc/165x107/60b6b5d95c847d097f3b3041/cuba-2012-10-18-3-omara-chucho-omara-portuondo-voz-chucho-valds.jpg)



